
Hypophysectomy, replacement therapy, and the tolerance of the euryhaline killifish, Fundulus...
21
GENERAL AND COMl’~H.ITIVE E~DOCHINOLOGY 5, t6+180 (1965) Hypophysectomy, Replacement Therapy, and the Tolerance of the Euryhaline Killifish, Fundulus heteroclitus, to Hypotonic Media GRACE E. PICKFORD,’ EMILY E. ROBERTSOK, AND WILBIJR H. SAWYER’ Received July 26. 1964 Hypophysectomized F~dalrts heferoclit MS, w t)reviously reported, survive only a few days after transfer from salt to fresh water. Survival time is the same when fish are tested 2 weeks or 2 months after hgpophysectomy. Gradual dihxtion of the external medium produces failure at an average salinity of 0.26% (0.67-0.02%). This salinity is comparable to that causing failure aft,er abrupt transfer. Although failure is associated with hypochlorcmia, daily injections of a balanced salt solution or 0.6% NaCl shorten survival. Daily injections of telcostean neurohypophysial peptides, arginine vasotocin (225 and 0.0225 pressor mU per gram) and isotocin ( 1.32 and 0.026 oxytocic mU per gram) do not prolong survival. An extract of caudal nemosecretory tissue (urophysis) from male Z’iZapla mossambica had no beneficial action at doses of 50 fig per gram every other day. Purified ovine prolartin, 105 mU per gram on alternate days, maintained survival of 10 of 11 fish for the duration of the 28&y test period in fresh water, irrespective of the period of pretreatment in salt water. Purified bovine prolactin prolonged survival and 2 of 6 fish were maintained for 28 days. Purified bovine growth hormone was ineffective. Monkey growth hormone containing 2 U per milli- gram of prolactin activity, administered at a dose of 20 pg per gram (equivalent to 40 mU per gram prolactin acbvitg), prolonged survival. Human grolvth hormone containing 4 U per milligram of prolactin activity. at a dose of 20 /.QZ per gram (equivalent to 80 mU per gram prolartin nativity) also prolonged survival and 4 of 6 fish were maintained for 28 days. The ability of pituitary preparations to l)rotec’t hypophysectomized F. hctrroclitzts from failure in fresh water appears to he related to their prolactin activity and not to their content of nrurohypophysial peptides. Thi s suggests that an adenohypophpsial hormone resembling mammalian prolactin is essential for survival of this fish in hypotonic external media. Carp pituitary preparations do not protect hypophysec- tom&d F. helprorlifus alt,hough F. heteroclitus pituitaries are known to be cffect,ive. Burden (1956 \, confirming preliminary observations of Pickford (1953)) demon- water. Failure usually occurrccl in 6-7 days at 15°C and was accompanied by asthema strated that hypophysectomized Fundulus and a fall in serum chloride. Attempted re- heteroclitus could not survive in fresh placement’ therapy with several mammalian ’ Supported by grant G-16247 from the National pituitary hormones, thyroxine, or deoxycor- Science Foundation. ticosterone failed to promote survival, but ‘Supported in part by grant G-17660 from the the hypophyaectomized fish could btl kept National Science Foundation and grant AM-01910 alive and healthy in fresh water by injec- and a General Research Support Grant from the tions of a brei of F. heteroclitus pituitaries. National Institutes of Health, U. S. Public, Health Perch pituitaries were partially effective Service. but not polluck glands. Pickford and Phil- 160
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Hypophysectomy, replacement therapy, and the tolerance of the euryhaline killifish, Fundulus...

GENERAL AND COMl’~H.ITIVE E~DOCHINOLOGY 5, t6+180 (1965)
Hypophysectomy, Replacement Therapy, and the Tolerance of the
Euryhaline Killifish, Fundulus heteroclitus, to Hypotonic Media
GRACE E. PICKFORD,’ EMILY E. ROBERTSOK, AND WILBIJR H. SAWYER’
Received July 26. 1964
Hypophysectomized F~dalrts heferoclit MS, w t)reviously reported, survive only a few days after transfer from salt to fresh water. Survival time is the same when fish are tested 2 weeks or 2 months after hgpophysectomy. Gradual dihxtion of the external medium produces failure at an average salinity of 0.26% (0.67-0.02%). This salinity is comparable to that causing failure aft,er abrupt transfer. Although failure is associated with hypochlorcmia, daily injections of a balanced salt solution or 0.6% NaCl shorten survival.
Daily injections of telcostean neurohypophysial peptides, arginine vasotocin (225 and 0.0225 pressor mU per gram) and isotocin ( 1.32 and 0.026 oxytocic mU per gram) do not prolong survival. An extract of caudal nemosecretory tissue (urophysis) from male Z’iZapla mossambica had no beneficial action at doses of 50 fig per gram every other day. Purified ovine prolartin, 105 mU per gram on alternate days, maintained survival of 10 of 11 fish for the duration of the 28&y test period in fresh water, irrespective of the period of pretreatment in salt water. Purified bovine prolactin prolonged survival and 2 of 6 fish were maintained for 28 days. Purified bovine growth hormone was ineffective. Monkey growth hormone containing 2 U per milli- gram of prolactin activity, administered at a dose of 20 pg per gram (equivalent to 40 mU per gram prolactin acbvitg), prolonged survival. Human grolvth hormone containing 4 U per milligram of prolactin activity. at a dose of 20 /.QZ per gram (equivalent to 80 mU per gram prolartin nativity) also prolonged survival and 4 of 6 fish were maintained for 28 days.
The ability of pituitary preparations to l)rotec’t hypophysectomized F. hctrroclitzts from failure in fresh water appears to he related to their prolactin activity and not to their content of nrurohypophysial peptides. Thi s suggests that an adenohypophpsial hormone resembling mammalian prolactin is essential for survival of this fish in hypotonic external media. Carp pituitary preparations do not protect hypophysec- tom&d F. helprorlifus alt,hough F. heteroclitus pituitaries are known to be cffect,ive.
Burden (1956 \, confirming preliminary observations of Pickford (1953)) demon-
water. Failure usually occurrccl in 6-7 days at 15°C and was accompanied by asthema
strated that hypophysectomized Fundulus and a fall in serum chloride. Attempted re- heteroclitus could not survive in fresh placement’ therapy with several mammalian
’ Supported by grant G-16247 from the National pituitary hormones, thyroxine, or deoxycor-
Science Foundation. ticosterone failed to promote survival, but
‘Supported in part by grant G-17660 from the the hypophyaectomized fish could btl kept National Science Foundation and grant AM-01910 alive and healthy in fresh water by injec- and a General Research Support Grant from the tions of a brei of F. heteroclitus pituitaries. National Institutes of Health, U. S. Public, Health Perch pituitaries were partially effective Service. but not polluck glands. Pickford and Phil-
160

FRESH-WATER TOLERANCE OF FUXDULCS 161
lips (1959) found subsequently that both cortisol and aldosterone accelerated failure in fresh water. Cortisol is a naturally occurring adrenal steroid in this species (Chester Jones et al., 1959)) and the doses employed were estimated from known cir- culating levels. Cod corpuscles of Stannius and hog renin were ineffective. Parathyroid hormone, kindly donated by Dr. Howard Rasmussen, also gave negative results at a daily dose of 0.9 U per gram (Pickford, unpublished data). Partial protect.ion was obtained with a crude ext,ract of rat pitui- tary glands, and survival was maintained for 3 weeks with sheep prolactin. These findings suggest, that a pituitary hormone resembling prolactin may participate in osmoregulation by F. heteroclihts.
0ther ruryhaline teleosts also appear to require the presence of t,he pituitary for fresh-water adaptation. Ball (1962) and Ball et ul. (1963) report that. hypophysec- tomized Poecilia (fMollien,esia) latipinna and P. (1W.) fornzosa succumb in fresh water. Schreibman and Kallman (1963) observed t.he same phenomenon in Sipho- phorus maczrlatus and X. helleri. The hy- pophysectomized eel, on the other hand, can withstand transfer to fresh water (Fontaine et al., 1949; Callamand et nl., 1951). Simi- larly, the euryhaline but, more habitually fresh-water cyprinodont, Fundulus diaph- anus, can survive indefinitely in this me- dium in the absence of the pituitary (Pick- ford and Ball, unpublished data). Moreover, various stenohaline fresh-water teleosts sur- vive in fresh water after hypophysectomy. The older data are summarized in Pickford and Atz (1957). In the euryhaline plains killifish, FunMus kansae, Dr. W. R. Flcm- ing, Department of Zoology, I-niversity of Missouri ipersonal communication) finds that hypophysectomy impairs fresh-water survival but that the fish can he maintained by addition of calcium to the medium. The role of calcium has not been explored in t,hc case of F. heterocbitus, but, the calcium con- tent of Kew Haven city water is low (ca. 9 ppm Ca-- according to data supplied by the New Haven Water Company). The present report is not concerned with the mechanisms of osmoregulation but. is de-
voted primarily to the nature of the hor- mone, or hormones, that maintain fresh- water survival.
MATERIALS AKD METHODS
All esperirnents were made on hypophq-secto- mized male F. heteroclitzts, according to proce- dures described previously (Burden, 1956; Pick- ford and Phillips, 1959). The fish were maintained and tested at 15” -ir l”C, unless otherwise stated; failure is more rapid at higher temperatures.
Two series of fish were employed: Series LVII
B fish were capt.ured in late summer and autumn, 1962; hypophysectomised in mid-January, 1963; and assigned to experiments in June, July, and .%ugust, 1963. Series LVIII C fish were captured in August, 1963 ; hypophysectomized in late Septem- ber, 1963; and employed in experiments between October, 1963, and Map, 1964. Both groups were
collect,ed in the J-icinity of New Haven, Connecti- cut.
At a suitable interval after removal of the hypophysis, usually 4 weeks or longer (see below), each fish was tested for its lark of ability to live in fresh water. When signs of asthenia developed, the fish was returned to salt water and allowed to recover before it was assigned to an experiment. Recovery was judged by resumption of normal feeding, and, as a rule, this required 2-3 weeks. Seventy fish were surccssfully tested, 8 others died in fresh water or failed to recover in salt water, and 3 survived for more than a month in fresh water. These fish are presumed to have been in- completely hvpophysectomized and were therefore rejected.
Fish that completrly recovered from one cxperi- ment were frequently assigned to a second, or even a third or fourth experiment. In certain experiment.al groups, fish that began to fail in fresh water, or fish that had been successfully ma.intained in this medium, were killed. Blood samples were taken and tissues were fixed for histological examination. The gills and skin of such fish are being studied by Dr. Noel Nussbaum (Department of Biology, Bowdoin College) and will be the subject of a latrr communication. All fish that died during or after an experiment were examined for possible collateral causes of death, in particular, for occlusion of the excretory ducts by renal calculi. Autopsies werp ultimately I)rrformrd on those fish that survivrtl.
Intraperitonral injections were ntiminirtpred tlai1.v or on :dtcrnatr t1:t.v~ untlcr light anesthesia with tricaine methane sulfonate (MS 222). All preparations were given in a standard volumr of 0.6% NaCl: 0.01 ml per gram weight of fish. Each

fish was weighed to the nc:trc:st 0.05 prim, but, es-
cept for neurohyl)ol)llysinl peptides, the volumc~ Was approximated to the nearest 0.005 ml. :kgi- nine vasot.ocin and isot ocin were injec’lrcl with a Hamilton microsyringe to the nearest 0.0005 ml.
In eslieriments in which fish were injcctcd on :iltcrnate days, t,imc to failure has tieen al)proxi- rnat.ed to the nearest whole day. It is impossible to rescue every fish at exact,ly the ,same stage of tiikrss, especially when failure occurs during the night. In experiments involving daily injections t,he fish were observed more closely, and failure time is approximated to the nearest 6 hours,
In the initial, noncsperimental screening tests, the fish invariably ceased to eat on the first 01 second day in fresh water. This was also true in many of the hormonal exl>eriments. However, some treatments promoted a continuation of feed- ing, even though ultimate failure might supervene. In such instances, data on feeding have been included in the tables.
Changes of salinity, in the gradual dilution experiment (Exp. 3) were measured with an RBZ- 3341 Solu Bridge conductivity meter (Industrial Instruments, Inc.) in the range above 40 Bmhos per centimeter (0.22%). Near fresh-water levels water samples were tested with a precision hy- drometer, since our conductivity m&r did not operate in this range.
Gcy’s solution, the mammalian balanced salt solution employed in one esperiment, was pur- chased from the Hyland Laboratories. It has the following composition per 1000 ml: NaCl, 8 gm; KCI. 0.375 gm: CaCh (anhydrous), 0.275 gm; MgCh.6 H,O, 0.21 gm; NaZHPOa.2H?0, 0.15 gm; KH?PO,, 0.025 gm; dextrose (anhydrous), 2.0 gm : SaHCO,, was not added.
Argir&Le Vasotocin. A solution containing 1 my per milliliter of synthetic arginine vasotocin (Berde et al., 1962) was generously supplied by Dr. B. Berde of the Pharmacological Laboratories of Sandoz Ltd., Basel. This was diluted with 0.05M acetic acid and 0.5% chlorbutanol to a concentration of 100 gg per milliliter. The result- ing solution contained 22.5 rat vasopressor (USP) units per milliliter. Final dilutions, with 0.6% NaCl, were prepared immediately before injection.
Zsotoci,~. This was also supplied in solution by Dr. Berde. The solution used contained 20 gg per milliliter of synthetic 4-serine, 8-isoleucine oxyto- tin (Guttmann et al., 1962). It contained 2.64 rat uterus (oxytocic) units per milliliter and was diluted with 0.6% NaCl immediat.ely before injection.
I ~iYlpllys;s. Ttwllill~ll qm:tl ~r(l~ irom m:ilt~ ‘l’hpin rtt~wsr~n~bicn collcrtrtl t)y Dr. Howar(1 Bern, I)cl~artmc~nt of Zoology, Uni\-ersily of (!:ili- Eornia, I~ork(~ley, and dehytlrnted in acrtone, wcrt~ tritur:itc,d in 0.6:;. NaCl. The volume of the suspension \vas adjusted so that 50 pg dry weight was injectctl llrr gram body weighf A similar 1)reparntion of Tilnpin brain (telrnccyhalon) n’;i> used for control injections. The solutions were frozen in daily injection ctuota. Assays of thrst~ tissues for neurohypophysial-like activities have been reported elsewhere (Sawyer and Bern, 1963).
Mumnmlian Adermhypophyskl Bornmm. Puri- fied bovine growth hormone (NIH-GH-B6) and purified ovine prolact in (NIH-P-S4) were donated by the Endocrinology Study Section of the National Institutes of Health. The following prep- arations were received from Dr. -4. E. Wilhelmi, Department of Biochemistry, Emory University : bovinc prolactin (B 671 B), monkey growth hor- mone (M 655 A), and human growth hormone (NIH-GH-HS-269). Solutions were prepared in advance and frozen in daily injection quota.
Carp Pilltitclry Preprrralions. Freshly frozen carp pituitary glands were purchased from StolIcr Fisheries, Inc., Spirit Lake, Iowa. They wcr(’ shipped in dry ice to Emory University. Dr. Wil- helmi prepared whole lyophilized powders from material collected in gugust, 1961 and in June, 1962. The August sample was also used for the preparation of fractions by a variation of the alkaline ethanol method of Bates and Riddle (1935) for prolactin. Thirt,y gm of the powder was susllrnded in 1 liter of 6056 ethanol, 0.01 N in SaOH. Solvent. was added at 5°C and the ext.rar- tion was conducted at room tempemture, for 3 hours: the pH was maintained at 10.0 by addition of 4 N NaOH as required. The suspension was centrifuged for 30 minutes at 10,4OOg, and the supernatant solution was decanted through glass wool to hold back floating particles. The clear d:trk green solution, 920 ml, was adjusted to pH 5.5 with 4 N HCl, and precipitation was completed liy the addition of 2500 ml of cold 95% ethanol. .ift,er the mixture had stood overnight in the cold. the prrcipit,atr had settled well, so that all but 500 ml of the solution could he siphoned off. The remainder was centrifuged for 30 minutes at 10,400 9. One-fourth of t,he precipitate was washed once with 95% ethanol, centrifuged again, SUR- pcnded in water, and lyophilized directly (Fc7148). The remaining three-fourths of the precipitate was suspended in water, dialyzed against distilled water, and lyophilized (Fc714B). Solutions were prepared in advance and frozen in daily injection quota.

Aden
ohyp
ophy
sial
activ
ities
Stan
dard
as
says
6 O
bser
vatio
ns
on
Fund
ulm
~ Ne
uroh
ypop
hysia
l ac
tivitie
s (m
U/m
n)
Prep
arat
ion
Inte
r- hle
lano-
M
elano
- G
II Pr
olacti
n u/
m!3
TS
H
FSH
Gro
wth
Thyr
oid
Tevt
is
rena
l ge
nesis
LI
I AC
TH
cyte
u/
mg
stim
. st
im.
stirn
. st
im.
stim
. pr
olif.
Antid
iuret
ic T:
aSOp
K?SS
Or
Oxy
tocic
Gr;a
v;;e
horm
one
Mon
key
Hum
an
Prola
ctin
Ovin
e Bo
vine
Carp
pit
uitar
y Cr
ude,
Ju
ne
Crud
e,
Augu
st
Frsc
tmn
Fc71
4.4
Frac
tion
Fc71
4B
1 3
<S
: C.
40
35
0.00
4.5
0.5
!)
Urop
hysis
Ti
lapi
a,
mal
e
Neur
ohyp
ophy
sial
Argin
ine
vsso
tocin
~-
Iso
tocin
1.9
ca.
0.25
04
10
ca
. 2
negl
. ra
. 4
0.4
0.12
21
(11
.:j-3
0 24
+++
:: $q
c Nk
? ;-y
; No
ne
30
44(3
9-49
) NO
M
1 so0
19
00(1
670-
2180
) -
+++
++
110
loo@
-105
)
Gi
Ge
p)
F T
No
ne
8 No
ne
33
40(3
2-42
) -
f NO
X 21
6(20
4-22
8)
- ++
+ ~-
- i
NOW
? 11
14(1
79~1
89)
0.11
128(
116-
142)
23
00(2
10&2
600)
29
0(28
2-30
0)
<I
5 <2
.;,
1704
-20)
10
4(98
-109
) 3X
0(32
0-45
0)
:353
(33
3-38
1,
<2
u/w
-_
__
0.65
13
8(13
1-14
5)
132(
121~
144)
a Fi
gure
8 in
pare
nthe
ses
repr
eeen
t 95
v/0
conf
idenc
e lim
its
b Da
ta
supp
lied
by
the
Endo
crino
logy
Stud
y Se
ction
(N
IH),
and
Dr.
A.
E.
Wilh
elmi.
ml./
mg
unles
s ot
hcrw
iv st
ated
. c
Base
d on
th
e pr
esen
t or
ea
rlier
expe
rimen
ts.
Arbi
trary
ra
tmgn
: it
to
+ +
+.

164 PICIiFOHD, ROBERTSOX, AND SAWYER
Riologicaz A.&wys (Tahlc 1)
Assuys for Xeuroh~pophyaial ActiLli1ir.s. Anti- diuretic activity was estimated by intravenous inject.ion into rats under rthanol anesthesia. Vase- pressor activity was determined by assays on pithed rat’s. Otherwise the method was that of the United Stat,es Pharmacopeia (USP). Oxytovic ac- tivitp was measured by Munsick’s (1960) modi- fication of the Holton assay on rat uteri suspended in a solution containing no magnesium ion. B USP reference standard was used in all assays. Details of the methods apljear elsrwhcrc (Sawyrr, 1964).
Asanys for Mn~~~~dirr/~ Atlolohypophysial Ac- tildies. The bovine growth hormone, and ovine prolactin, donated by the Endocrinology Stud] Section, Nat,ional Institutes of Health (NIH), were’ acrompanictl by specifications for atieno- hypophysial activities, and thrsc data are given in Table 1. Growth potency is givc,n in terms of the USP reference standard and was assayed by the l&day body weight gain test in hypophyserto- mized fcmalr rats. Lutrinizing hormone (LH), assayed by the ovarian ascorbic acid depletion method of Parlow, is expressed in terms of NIH- LH-Sl standard. Follicle-stimulating hormone (FSH), by the HCG-augmrntation mc%hod of St.c&~lan and Poh1c.v. iz; similarly rxpresscd in trrms of NIH-FSH-Sl rtandald. For c.onvrnience, in Tablet 1. WV hare cxprc~ssctl both thrsc, activi- ties as milliunit s. Thyroid-stimulating hormone (TSH). in terms of USP thyrot,ropin *tandanl. was assayed by the chiceli thyroidal P:” uptake mrthod of Lamberg. Adrcnocorticotropir hormonr (ACTH) was estimated in terms of CSP rrfrrcnccx Standard by the Munson modific~ation of thc~ Sayers assay. Prolactin activity was determined by the pigeon croij sac wc+ght, method of Riddlr, et nl.
Thr data for bovine prolactin, monkey growth hormone, and human growth hormonr were supplied by Dr. -4. E. Wilhelmi.
(lbsereutions OWL Fundulus. These observations throw further light on rollateral activities of t,he mammalian adenohypophpsial hormones and pro- vide some informat,ion concerning hormonal activi- ties of the carp pituitary preparations. Growth- promoting activity could not h(T estimated hecause the experiments were made at 15°C. At this tem- perature, administration of bovine growth hormone elicits no growth response in hypophysectomized F. hrferoclitus (Pickford and Atz, 1957). Data on thyroid ~11 height, which reflec*ts the TSH artiv- ity of the preparation, are available for only two preparations: monkey growth hormone and carp lyophilized pituitary. Testis stimulation, shown previously to he an LH-stimulated response (Pirkford and Atz. 1957), is reflected by the dc~l-
opmcnt of nuptial coloration. This is ~~s~~rcssc~l in arbitrar.v units ranging from i- (t ingc’ ot’ y~~llow on prlvic> and anal fins) to + + + (maximum yc~llo~~- over entire vf,ntral surfacr) (Pir~kfonl. 1953). In some rxperimc,nts t,his is supplemrntcd by data on the gonosomal.ir index (GSI = weight oi trstes x lOO/weight of body). Strong testis stimulat,ion i+ reflected by the appearance of flo\+%lg milt. ran- taining mot,& spermatozoa, and is readily ob- served when the fish are handled, During each rxperiment, ohserrat.ions were also made on the degrcr of dorsal darkening and on I)ossible pro- lifer&ion of newly-formed melanoc.vtes on the belly. Previous studies have shown that restoration of melanin pigment in the faded mrlanophores of hypophyscctomizrd 8’. heterodit 1~s is elicited b> ovine prolactin, whereas the proliferation of new melanocytes requires the presence of int,ermedin (Pickford and Kosto. 1957: Kosto ci nl., 1959).
HEWl,TS
The experiments are grouped in four categories, alt,hough they were not neces- sarily performed in the sequence in which they are presented.
Group I: Collateral Problems
Experiment 1. Fresh-water survival 2 weeks after hypophysectomy was compared with that. of the same fish, retested after 2 months, and, in four instances, after recov- ery from an intervening experiment with isotocin. There was no significant, difference in survival time between the initial test and the retest (Table 2). Fish 2 was the only exception; it had failed on the two previous tests but subsequently was able to survive normally in fresh water. The head was fixed in sublimated Bouin-Hollande and sent to Dr. John N. Ball, Department of Zoology, University of Liverpool. Dr. Ball discov- ered the anticipated presence of a regener- ated hypophysis, regarding which he writes (personal communicat,ioni : “The regener-
ated adenohypophysis is composed almost entirely of rostra1 erythrosinophiles, cells which have been identified as the source of the fish prolactin-like hormone (Ball and Pickford, 19641. A full report will be pub- lished subsequently.”
The only difference between the initial test, 2 weeks after hypophysectomy, and subsequent fresh-water tests was the mag- nitude of the abnormal behavior on return
FRESH-WATER TOLERANCE OF FCNDULUS 165
TABLE 2 COMPARISON OF FRESH-WATER SURVIVAL Two
WEEKS AFTER HYPOPHYSECTOMY WITH RESVLTS OF B RETEST Two MONTHS
LATER (EXPERIMENT 1)
Days to failure in fresh wata Ser. LVIIIC; Hypophysect. Imtocin.
IFT;;4ti:3; Init~l&st, X .28.63; Retest, Later .a Exp. Zad 1.9.64 Fat@ exp.
I 6 2 5 3 5 4 6 5** 6 6 4
ti R 151) 27+ The -
6 R 15b 5 R 15b 6 R 151) 5 R 15b
a One asterisk indicates that kidney stones were found in urinary ducts at autopsy, during or at termination of experiment; two asterisks indicates that kidney stones were found after full recovery or in a later experiment.
b Key to symbols: A = Autopsy, either of fish failing in fresh water or at successful termination of experiment (indicated by symbol + after the number of days in fresh water). Dfw = Died in fresh water during test. Dsw = Failed to recover in salt water. Dl = Died subsequently, after complete recovery in salt water. R = Recovered and resumed eating in salt water. T = Survived for the number of days indicated in fresh water; healthy and eating normally unless otherwise indicated; experiment terminated by return to salt water, or by autopsy if so indicated (A).
c Residual pituitary regenerated; see text, d Approximated to nearest whole day (cf. Table
6).
to salt water. The fish showed signs of ex- treme distress for several days, swimming wildly and spinning when disturbed. Such violent behavior is rarely observed in ex- periments on fish that have been hypophy- sectomized for a longer period of time.
Experhent 2. The effects of daily injec- tions of 0.6% NaCl, 0.01 ml per gram weight, were compared with the effects of similar volumes of a balanced saline soIu- tion (Gey’s solution). The results indicat,e that both solutions accelerate failure in fresh water (Table 3). All fish had gained weight at the t,ime of failure, and the mean increase for bot.h groups taken t,ogether was 7.6%.
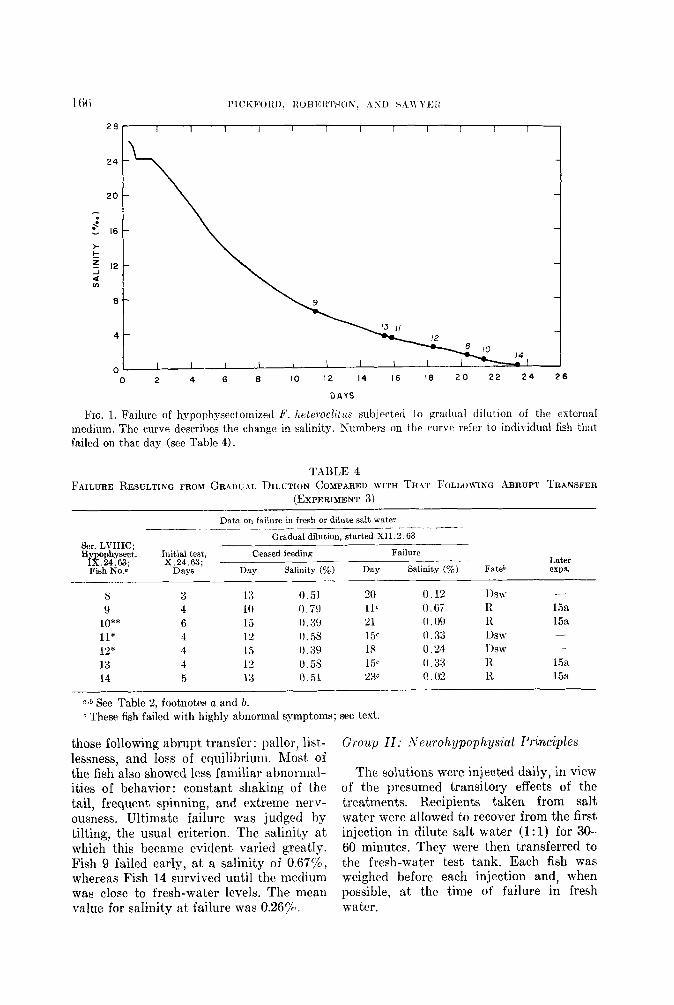
Experiment 3. Hypophysectomized fish were subjected to gradual dilution of the external medium to ascertain whether they would become adapted to fresh water, as compared with their failure following abrupt transfer. Dechlorinated city water was allowed to drip slowly into an aquarium equipped with overflow siphon. The salinity was measured twice daily and also at the time of failure of each fish.
The results show that gradual diMon did not enable any of the seven fish tested to survive indefinitely in a hypotonic me- dium (Table 4 and Fig. 1). The fish stopped eating and began to show signs of discom- fort at salinities ranging from 0.39 to 0.75pl. Signs of distress resembled, in part,
TABLE 3 COMPARISON OF THE EFFECTS OF INJECTION OF 0.6% NACL AND GEY’S SOLUTION ON FRESH WATER
SURVIVAL (EXPERIMENT 2)
Data on failure in fresh water (days survival)
ser. LVIIIC: Hypophyaeet. 1X.24-25.63;
Fish No.a
Initial teat Exp. 2. IV. 27.64
No. of Previous Date Days Treatment inj.
Weight Days change (%) Fat& CCp*.
22** X1.24.63 5 0. G%NaCl I 8 4 11.8 R Dl 10
29 X11.8.63 8 0. 6ycNaC1 2 ‘2 12.9 R 10 3o*c X11.8.63 6 0. G%NaCI 1 1 - R 10 32 X11.8.63 3 0. 6yGNaC1 1 3 4 6.7 Dsw 10
25* X1.24.63 5 33 X11.8.63 6 34 X11.8.63 6 35 X11.8.63 10
osb See footnotes a and b of Table 2. c See Table 9, footnote c.
Gey’s ‘2 1 7.1 Dfw 5 Gey ‘s ‘2 1; 8.3 Dfw 10
Gey’s ‘2 1; 3 0 Dsw 10 Gey ‘s 3 t? 3.4 Dsw 10
I (iti
24
4
0 0 2 4 6 6 IO I2 I4 I6 I6 20 22 24 26
DAYS
FIG. 1. Failure of hypophysectomized F. hetcroclitus subjcctetl to gradual dilution of the external medium. The curve describes the change in salinity. Kumbers on the curve refer to individual fish that failed on that day (see Table 4).
TABLE 4 FAILURE RESULTING FROM GRADU.\L I~ILUTION COMP.IREI) WITH T~.lrr FOLLOWING ABRUPT TRANSFER
(EXPERIMENT 3)
Data on failure in fresh or dilute salt water
Gradual dilution, started XII. 2.63 Ser. LVIIIC; nypOpbysect. Initial test, Ceased feedinn Failure
;$i;4tiE3 ; .a xzf3;
Later Day Salirlity (%) Day Salinity (%) Fate* exps.
8 3 13 0.51 20 0.12 L)s\\ - 9 4 10 0. T!J IlC 0.67 R 15a
10** 6 15 0.39 21 0, O!) R 15a 11* 4 12 0.5s 15c 0.33 IhW - 12* 4 15 0.39 1S 0.24 I>SW - 13 4 12 0.5s 15c 0.33 R 15a 14 5 1:: 0.51 ‘TJC 0.02 R 15a
n*b See Table 2, footnotes a and b. C These fish failed with highly abnormal symptoms; see text.
those following abrupt transfer: pallor, list- lessness, and loss of equilibrium. Most of the fish also showed less familiar abnormal- ities of behavior: constant shaking of the tail, frequent spinning, and extreme nerv- ousness. Ultimate failure was judged by tilting, the usual criterion. The salinit,y at which this became evident varied greatly. Fish 9 failed early, at a salinity of 0.67$, whereas Fish 14 survived until the medium was close to fresh-water levels. The mean value for salinity at failure was 0.265%.
Group II: Xeurohvpophysiat Prkiples
The solutions were injected daily, in view of the presumed transitory effects of the treatments. Recipients taken from salt water were allowed to recover from the first injection in dilute salt water (1: 1) for SO- 60 minutes. They were then transferred to the fresh-water test tank. Each fish was weighed before each injection and, when possible, at the time of failure in fresh water.
FRESH-WATER TOLERANCE OF FUNDULUS 167
TABLE 5
ARGININE VASOTOCIN (AVT) (EXPERIMENT 4)
Data on failure in fresh water
Ser. LVII B; Hypophysect.
L2&2&63; .a
Initial test (a) AV;f~::j~U/gm) (b) AVT(O.O225mU/gm)
VII.23.63
Day8 Inj. SWV. % wt. Inj. SWV. Date S”TV. days days change days days
% wt. change Fat+
21 V.22.63 8 - - - 3 3: -1.5 &
26 V.22.63 11 - 4 4$ 0.9 AC
56 VI.11.63 6 4 4; 10.0 5 51 17.6 A
58 VI.11.63 6 3 3 6.1 4 46 0 A
100 VI.10.63 5 2 ',3 - "4 3 3; 4.5 A
102 VI.10.63 5 2 2; - 3 3: 3.8 Dfw
orb See Table 2, footnotes a and b. c Used one month Dreviouslv in EXD. 14: still showed marked nuptial coloration on pelvic and anal fins _
resulting from testis stimulation.
Experiment 4. (a) Arginine vasotocin resulting from previous treatment with a (AVT) was injected daily into four fish at crude carp pituitary preparation. This made a moderate dose, 2.25 pressor mU per gram. no difference to their survival time. In the (b) After full recovery, the same four fish majority of tests a weight. increase that together with two others from Exp. 14 were averaged 5.4% was noted at the time of tested at a lower dose, 0.0225 mIJ per gram. failure,
Neither dose of AVT enabled the fish to withstand fresh water (Table 5). Failure followed the second, third, or fourth injec- tion, usually within 6-18 hours; only Fish 56 received a fifth injection. Failure was accelerated in comparison with the initial test in all instances. Two of the recipients in Exp. 4b had markedly stimulated testes
Experiment 5. (a) Isotocin, the teleostean oxytocin-like principle (TOP), was injected into four fish at a moderate dose, 1.32 oxy- tocic mU per gram. (b) Another group of four fish were tested at a lower dose, 0.0264 mU per gram.
The results resemble those with AVT. The treatment did not promote fresh-water
TABLE 6 ISOTOCIN, THE TELEOSTEAN OXYTOCIN-LIKE PRINCIPLE (TOP) (EXPERIMENT 5)
Data on failure in fresh water
Ser. LVIIIC; Hypophysect.
1X.24.63; Fish No.0
Initial test (a) TO;,W&dJ/gm)
Days Date
Inj. Days % wt. Subsequent mm”. days swv. change Fate” eXpS.
2 3 5** 6
X.7.63 x.7.63 S.i.63 X.7.63
-3.2 6.3
9.4
R 1 R 1 R 1 R 1
151, 15b 15b
Hypophysect. 1X.24-25.63
(b) TO~!O.;~;~ mu/pm)
24** X1.24.63 5 2 2$ 3.7 RDl - 25** X1.24.63 5 ‘2 aa 2.9c R 2
26 X1.24.63 5 2 '2 t 9.3 RDl - 28* X1.24.63 6 s 2+ 9.5c Dsw -
a,b See Table 2, footnotes a and b. c Weight taken at last injection, 2-3 hours before failure.

168 PICKFORD, ROBERTSON, AKD SA\VTP:R
survival but accelerated the t,ime of failure as compared with the initial test (Table 6). In most instances a weight increase averag- ing 5.6% was evident at the t,ime of failure.
Group III: The C’nwlab Xewosecretoq Organ (Uroph ysis’l
Experimen.t 6. Three fish received injec- tions of Tilapia urophysis and three re- ceived brain; the dose was 50 pg per gram. The frog bladder activity of the male urophysis was 25 mU per milligram (Saw- yer and Bern, 19631. At the 50 pg per gram dose, each fish therefore received 1.25 mU per gram. This is comparable to the frog bladder activity of the lower dose of AVT, administered in Exp. 4b, which was of the order of 2 mU per gram. As with neurohy- pophysial principles, the first injection was given to fish taken from salt water, and recovery in dilute salt water was followed by t,ranefer to fresh water. Subsequent in- jections were given on alternate days.
The rcsult,s fail to show any beneficial effects of the t,reatment, (Table 7).
Group II-: Gowth Hormone. Prolactin, nvd Carp Pituitar?y Prepawtions
Erperiment 7. Before testing primate growth hornlone preparations, it was necce- sary to confirm t.hc findings of Burden (1956) that bovine growth hormone was ineffective. Fish used as controls in an assay of growth promoting activity in fish pitui-
t,ary preparations hati rc)ccGvetl injec.tions of bovine growth hormonc~ CSIH-GH-B6 ) at 20 pg per gram thrice wc~~kly for 6 wcckh at about 20°C. The preparation contained a t,race of prolactin (Table 1 ) rquivalent. to 8 niV per gram at the dostl c~ploytd, but, no darkening was observed. At the termi- nation of the experiment, these fish show.t~d an average length increment of 4.57c and slight testi:: stimulation that was reflected in the development, of traces of nuptial coloration on the pelvic and anal fins. The fish were then acclimated for 8 days t,o the lower temperature necessary for fresh- water survival tests (ea. 15”Ci, and injec- tions were continued on alternate days. On the day aft.er the fourth injection at the lower temperature, the fish were transferred to the fresh-water test tank and injections were given on alt.ernate days until failure. At the on& of distress, blood samples and t,issues wcrc taken for subsequent study.
The results confirm that treatment with purified bovine growth hormone does not enable hy~~o~~hysertomizccl F. heternclit~rs to live in fresh water (Table 8). This is the only group of fi?h that, had not been pre- tested in fresh water. The time?; of failure (6-10 days I arc of the order usually cn- countered in screening tests of uninjectcd fish, but :I proper comparison must he made with the shortened survival time of control fish receiving saline injections (Table 3, and Burden, 1956). The fish continued to eat for
TABLE 7 UROPHYSIS ANI) BRAIN OF Tilapiu mossamhica (EXPERIMENT tin
Days to failure in fresh water
Ser. LVIIIC; Hypophyseot.
1X.24.63 Fish No.a
Tilapia preparations0 X11.4.63
(a) Urophysis (b) Brain Later
Fate” EXps. -
15 6 4 - R 13 16 4 32 - Ik3w 20 6 32 - R 13
18 4 53" R 13 19 6 42 R 13 21 4 3% R 13
a,b See Table 2, footnotes a and b. c All fkh received two injections, on the first and third days. Fish 18 survived unt.il day 5, hut was in such
poor condition that a third injection was not attempted.

FRESH-WATER TOLERANCE OF FUNDULUS 169
several days, and failure supervened despite this presumably beneficial response to the treatment.
Experiment 8. A previous investigation had shown that sheep prolactin promoted fresh-water survival although the fish did not eat normally and were not in good con- dition (Pickford and Phillips, 1959). It Tvas thought that, a combination of ovine prolac- tin and bovine growth hormone, now shown to prolong feeding, might give more favor- able results. The prolactin used in the 1959 experiment (C. II. Li) had an activity of 35 U per milligram: at the 10 pg per gram dose each fish therefore received 350 mU per gram. In the present experiment each fish was given a mixture containing ovine prolactin (10 pg or 210 mU per gram) and bovine growt,h hormone (20 ,ug per gram). Four fish were pretreated for 1 week in salt water, as in the 1959 experiment. They were transferred t.o fresh water on t,he day following the fourth injection, and treat- ment was continued on alternate days.
The results show that the treatment pro- longed survival in three of the four recipi- ents, and feeding cont.inued at least, inter-
mittently (Table 8). Fish 55 began to eat on day 15 and was autopsied after 3 weeks in fresh water; it was the only one to sur- vive. In the 1959 experiment, four fish sur- vived for 20 days when the experiment was terminated, but only one of these was eating. Combination with bovine growth hormone certainly did not improve survival. As in all experiments with ovine prolactin, the fish began to darken on the second or third day of treatment, due to restoration of melanin pigment in depigmented melano- phores. No new pigment cells appeared.
Experiment 9. Three fish were pretreated for 24 days with ovine prolactin at 5 pg per gram (105 mU per gram) daily for the first 7 days and on alternate days thereafter (a total of 15 injections). On the day following the Iast injection they were transferred to fresh water, and treatment was cont)inued on alternate days. After 28 days in fresh water the recipients were still alive and healthy (Table 9), eating normally, and in excellent condition. The experiment, was terminated by autopsy. No visible renal calculi were observed. All fish became very dark dorsally due to restoration of melanin
TABLE 8 EXPERIMENTS WITH BEEF GROWTH HORMONE (NIH-GH-B6) ALONE OR IN COMBINATION WITH SHEEP
PROLACTIN (NIH-P-S4) (EXPERIMENTS 7 AND 8)
Days eating, and failure or survival in fresh water
Ser. LVII B; Hypophysect.
1.23-25.63; Fish NO.” Date
Initial test
Eatinge D&Y8
V.13.63 Exp. 7d
Eating< DSYS Fateb
32* 2 7 A 33 5 6 A 34* 8 10 A 35* - 4 7 A
As above V.29.63 Exp. 88
31 IV.22.63 N4 10 6.NlO 12 A 42* V.29.63 0 8 5 6 Dfw 51 V.10.63 0 3 6.N8 17 Dfw 55 V.2.63 1 8 B15 22-t TA
a*b See Table 2, footnotes a and II. c Days during which fish continued to eat normally: N = Nibbled on day indicated; B = began to eat on
day indicated. d Experiment 7: GH-B6,20 pg/gm; pretreated for 7 qeeks in salt water; see text. e Experiment 8: GH-B6,20 pg/gm plus P-S4, 10 pg/gm; pretreated for 1 week in salt water.

Ii0 PICKFOIID, IWBERTSOS, AND SAWYEll
pigment without proliferation of ncw-ly- formed melanocytes. Nuptial colors wcrc lacking.
Erpetin&ent 10. Experiment 9 diffcrcd from the 1959 prolactin experiment in two respects; the lower dose (5 pg as compared with 10 pg per gram) and the longer period of pretreatment (24 days as compared with 8 days). Experiment, 10 was designed to establish whether pretreatment was in itself ‘a decisive factor. One group of four fish was injected with ovine prolactin thrice weekly for 1 month, at 5 pg per gram, in salt water; a control group received 0.67, NaCl. On the day preceding transfer to fresh water, the fish in the control group were also given prolactin. Prolactin injections were contin- ued on alternate days for the duration of the experiment.
The results show that pretreatment with prolactin is not essential for prolonged fresh-water survival (Table 9). Only one fish died during the experiment, and this was in the group that had been pretreated. Fish 35 had gill rot but survived neverthe- less. The others, pretreated or not, remained
liealt’hy nnll continued to eat during the ent,ire experiment. As usual, all fish became dark dorsally, but without proliferation of new pigrncnt cells. Fish 32 may have had slight testis stimulation since nuptial colors were evident on the pelvic and anal fins, After the injections were discontinued, fol- lowing r&urn to salt, water, the seven sur- vivors gradually became paler.
Experiment 11. A bovine prolactin prepa- ration was injected at the 5 pg per gram dose successfully employed in Exps. 9 and 10 with ovine prolactin. The crop gland stimulating activity, in view of the range of variation of report#ed assays (Table I), was essentially similar (120 mU as compared with 105 mU per gram). Six fish were pre- treated thrice weekly for 1 month in salt water and were t,ransferred to fresh water on the day following the 13th inject,ion. After that they were injected on altcrnatc days.
The results indicate that beef prolactin can promote fresh-water survival, but at the dose employed this preparation was lees effective than ovine prolactin (Table IO).
TABLE 9 FURTHER EXPERIMENTS WITH SHEEP PROLACTIN (NIH-PS-4) AT A LOWER DOSE; WITH .LNI) WITHOUT
PROLONGED PRETREATMENT (EXPERIMENTS 9 AND 10)
Days to failure or survival in fresh water
Series and
date hypophysect.
Exp. 10: Exp. 9: PS-4 5rg/gm;
Initial test PS-45fig/gm; Fish - pretreated (a) Pretreated (b) Not pretr. Later NO.5 Date Days VII.24.63 11.23.64 Fat+ l%Xp.
-- - LVII B 1.23-25.63 46 V.22.63 11 28+ - TA
59 V.10.63 9 28+ - TB
101 V.10.63 4 28+ TA - .~
LVIIIC 1X.25.63 22** X1.24.63 - - 28+ - T ‘2
29 X11.8.63 ii 28+ T ‘2
3o*c X11.8.63 6 28+ T 2
31 X11.8.63 8 9 Dfw -
32 X11.8.63 3 - 28+ T 2
33 X11.8.63 6 28+ T 2
34 X11.8.63 6 28+ T 2
35 X11.8.63 10 28+ Td 2
-1) See Table 2, footnotes a and b. C A small calculus, lodged in the urinogenital papilla, could be seen at each injection. d Did not eat well during experiment, was found to have gill rot and was treated with malachite green;
rrrovered in salt water.

FRESH-WATER TOLERANCE OF FUNDULUS 171
TABLE 10 BEEF PROLACTIN (B-671B, WILHELMI) (EXPERIMENT 11)
Days eating and survival or failure in fresh water
Ser. LvIIIc; Hypophysect.
1X.25.63; Fish No.-
Exp. 11. B671B, Bpg/gm Initial test 111.3.64~ X11.23.63
Survival Last ate Failed Last ate or failure
36 1 10 5 8 Dfw 37** 1 9 23d 28$ T 38** 1 7 4 28+ T Dl 40 2 3 34d 27 R 41 2 6 1 9 DSW 42 1 6 19d 23 Dsw
O@b See Table 2, footnotes a and b. c Pretreated in salt water for 1 month. d Feeding intermittent after first few days. e A nematode, Eustrongyloides sp., encapsulated in viscera.
Two fish failed early in the experiment, but one of these had a parasitic nematode dis- tending the viscera and its failure may be discounted (Fish 41). The remaining four fish ate intermittently; they were pale and did not appear to be healthy. One failed on day 23 and one on day 27, just before termination of the experiment,. Two were still alive after 28 days in fresh water. At the time of failure, or at termination, no fish showed any trace of testis stimulation (nuptial colors lacking) and there was no significant change in length, Surprisingly, in contrast to fish treated with ovine prolactin, there was little or no restoration of melanin pigmentation.
onset of distress each of the four long survivors was autopsied. No fish had changed significantly in length, the nuptial colors remained negative, and the gonoso- matic indices were below 0.5; these results indicate a lack of testis stimulation. Meas- urements of thyroid cell height suggest that very little thyrotropic activity was present in the preparation. All fish became dark due to increased melanin pigmentation in faded melanophores, but no proliferation of neu pigment cells was observed.
Experiment 12. Five fish were injected with monkey growth hormone on alternate days for 1 week in salt water. On the day after the fourth injection they were trans- ferred to fresh water. The dose employed, 20 pg per gram, contained relatively low crop gland activity (ca. 40 mU per gram).
Experiment 13. Human growth hormone, like monkey gr0wt.h hormone, was adminis- tered at a dose of 20 lug per gram. The pro- lactin activity of the preparation was such that each fish received 80 mU per gram per injection. Six fish, of which four had pre- viously been used in Exp. 6, were pretreated thrice weekly for 1 month in salt water; they were placed in fresh water on the day after the 13th injection, and injections were given subsequently on alternate days.
The results indicate considerable bene- The results show good survival (Table ficial action (Table 11). Four of the five 11). Fish 19 failed rather soon after trans- recipients survived for 18-22 days in fresh fer to fresh water; Fish 18 survived until water and continued to eat for the greater part of this t,ime. The survival time is all
day 24; the remaining four recipients were
the more remarkable since the temperature still alive and were eating and healthy
control system failed and the water in the after 1 month in fresh water. The experi-
test tank rose to about 20°C during the last ment was terminated by return to salt water. The testes of these fish were un-
week of the experiment. It is possible that survival would have been longer if the
doubtedly strongly stimulated, which re-
temperature had remained lower. At the flects the known contamination of the prep- aration with LH (Table 1). All except

172 PICKFOKD, IWRl!XTHOK, AND SA\VYF:R
PRIMATE GROWTH HORMONES: MONKEY (M655.1, WILHELMI) .ANU H~M.\N CSIH-(;H-HS-‘,‘G!r)
(EXPERIMENTS 12 ANI) 13)
LVII B 4** VI.24.63 6 2 (5 R 1.22-24.63 8 3 18 201 0.13 A
10 7 15 1Xf 0.41 .\ 20 6 18 pf I 0.40 .I .-
48 5 18 1Sf 0.33 An
Exp. 13, HS-269 20 pg/gm, II.3.64h
5:: Failure Nuptial Flowing
days COIOTS Jp~~lll
LVIIIC 15 X1.6.63 6 28+ 28f + 111.22.64 ‘I 6a 1X.25.63 17 7 28+ 2+ + T
18 4 10. N17” 24 + III, “0.64 Dsw 6b 19 6 4 5 - DSW 6b 20 6 2X+ 2X+ + 111.20.64 ‘I 6a 21 4 28+ 28+ + III. 22.64 T 111 6b
nrb See Table 2, footnotes a and b. c Rated from negative (-1 to maximum (+ + +). d Gonosomatic index = testis wt. X lOO/body wt. e Pretreated for 1 week in salt water. f Owing to malfunction of temperature control system, the water temperature rose from about 15°C to
aljout 20°C 15 days after start of fresh-water test and remained high for 1 week. g Fish 48 had an extensive brain tumor. h Pretreated for 1 month in salt water. i N = nibbled on day indicated.
Fish 19, which died early, showed marked tion of new pigment, cells which spread nuptial coloration, and flowing sperm was over t,he belly. evident in four instances after 7 weeks of Experiment l-6. A lyophilized whole pitui- treatment. Length changes were not signifi- tary preparation from carp collected in cant. All fish became extremely dark and June was administered at a dose of 100 pg there was a more or less ext,ensive prolifera- per gram on alternate days ; pretreatment
TABLE 12
LYOPHILIZED CARP PITUITARY; JUNE (EXPERIMENT 14)
Ser. LVII B; Hypophyseot.
1.22-25.63: Fish No.0
21 24**
26 30
107
Days eating aIld to failure in fresh water
Exp. 14.. June carp Initial test 100 cg/gm. VI .24.63c
Later Date Eating Failure Ent.ing Failure Fate” lXP8.
V.22.63 ‘2 8 3 11 R 4b V.22.63 2 7 3 4 R -
V.22.63 2 11 8 12 R 4b V 22 63 V: 13:63
2 7 .3 Dsw 1 9 3 R
a,b See Table 2, footnotes a and b. c Pret,reated for 1 week in salt water.

FRESH-WATER TOLERANCE OF FUNDULUS 173
for 1 week in salt water was followed by transfer to fresh wat,er on the day after the fourth injection.
The results fail to indicate any beneficial effect on survival time in fresh water (Table 12). The fish continued to eat, how- ever, for at least 3 days. The recipients showed t,ransitory pallor after each injec- tion, which was attributed to the teleostean melanophore-concentrating hormone. Even the comparatively short period of treatment was sufficient, to initiate testis stimulation, which was reflected in the development of nuptial coloration on the pelvic and anal fins. This stimulation persisted for almost a month after termination of the experiment and was still evident in two survivors t.hat were later assigned to Exp. 4b.
Experiment 15. Although essentially neg- at,ive results were obtained in Exp. 14, there are possible collateral causes for failure, and a further investigation m-as undertaken.
The crude lyophiliaed carp pituitary powder was prepared from material collected at a different time of year (August) ; the dose was lower (20 pg per gram). Two fractions derived from this material by the alkaline ethanol extraction procedure for prolact,in were also tested at the same dose. Fraction Fc 714 B was dialyzed; fraction Fc 714 A was not (see Materials and Methods).
Recipients were injected thrice weekly for 1 month in salt water. On the day after the 13th injection they were placed in fresh water, and subsequent injections were given on alternate days. The results are essen- tially negative (Table 13). All fish ceased eating after the first day in fresh water. Fish 4, receiving Fe 714 A, survived for 13 days but was in poor condition and was pale and quiescent. No fish showed any significant length change during treatment eit,her in salt or fresh water. Transitory pal- lor developed after each injection, but the
‘i’-1BLE 13 C:\RP PITTXTARY FRACTIONS (EXPERIMENT 15)
Data on failure in fresh water
Ser. LVIIIC; Initial test Hypophysect. -
Exp. 15, 20 pglgm, III. 17.63~
IX. 24-26.63; Failure F;Cl;e NupotiaJ Flowing Previous Fish No.a Date days Fraction SPtXlW Fat@ exps.
9 X.24.63 4 (a) Lyophilizrd pituitary (August) 8 + 32fw Dsw 3 10** X.24.63 6 2 + -R 3 13 X.24.63 4 2 * - RDl 3 14 X.24.63 5 3’ ++ 32 fw Dfw S
1 X.i.63 6 (h) Fc714.4 5/ ++ - R 1 3 X.7.63 5 2 ++ 28s~ R 1, 5a 4 X.7.63 6 13 + 28s~ R 1 5** X.7.63 6 2 ++ - R 1, 5a 6 X.7.63 4 4’ + 32fw R 1, 5a
49 1.2.64 5 2 ++ 30fw R
43** 1.2.64 4 (c) Fc714B 5 + -R - 44 1.2.64 5 3’ + 32 fw Dfw - 45 1.2.64 7 9 + 32 fw R 46 1.2.64 5 2 IL -R 47 1.2.64 6 5’ + - R 48 1.2.64 6 5’ ++ - R -
avb See Table 2, footnotes a and 6. c Pretreated for 1 month in salt water. d Rated from negative (- ) to maximum (+ + +). B Day on which flowing sperm was first observed, counted from initiation of treatment (day 0); fish put in
fresh water on day 29; symbols SW and fw indicate whether sperm wae first seen in salt or fresh water. f Edema evident previous to failure.

li4 PICKFORD, ROBERTSON, AND SA\VYER
over-all color, when not under the immedi- ate influence of the melanophore-concen- trating hormone, showed litt,le or no restora- tion of melanin pigment. However, in an earlier experiment with the lyophilized powder at a higher dose (100 pg per gram), significant melanization was observed. In the same experiment, which was at 2O”C, the crude carp material caused a slight growth increment but little or no stimula- tion of the thyroid; the testes were strongly stimulated. Strong testis stimulation was also evident in the present experiment: the nuptial coloration, originally absent., ad- vanced to a + (yellow on anal and pelvic fins) and, especially with fraction Fc 714 A, to a + or ++(yellow over entire belly). Flowing motile sperm developed in eight of t,he fish, either while the fish were still in salt water or after t,ransfer to fresh water. The time of treatment (2&32 days) rather than the salinity of the medium is probably the decisive factor. Four of the fish with flowing sperm were receiving fraction Fe 714 A, but the numbers are not sufficient to decide whether the gonadotropin content of this nondialyzed fraction is significantly greater t.han that of the dialyzed fraction Fc 714 B. Significant edema was evident in several fish prior to failure in fresh water.
DISCWSIOS
FACTORS AFFECTING FAILURE IN FRESH WATER
The experiments reported in Group I brought to light some interesting results. The time of failure in fresh wat’er remains fairly constant (Exp. 1)) and, within rea- sonable limits, repeated use of hypophysec- tomized fish is permissible. The initial test in fresh water should not be made too soon after hypophysectomy. Not only do recently operated fish react badly when returned from fresh to salt water, a problem that demands further study, but there is also the danger, observed in one instance, that a functional pituitary may be regenerated.
Burden (1956) was unable to maintain hypophysectomized F. heteroclitw in fresh water by injections of NaCl, although he
demonst,ratcd that failure> was associateti with a decrease in blood chloride. Such replacement therapy could fail owing to the administration of insufficient salt,. Burden’s results showed, however, that. the treatment itself accelerated failure. Experiment,s on intact F. heteroclitus (Slither and Pickford, unpublished data), with the l-hour leuco- penic response as an index of stress (Slichcr, 1958, 1961; Slither et al., 1962), have indi- cated that injections of 0.6% NaCl at a dose of 0.02 ml per gram are stressful. On the other hand, injections of a balanced salt solution (Gey’s solution) at a smaller vol- ume per gram weight, 0.005 ml per gram, failed to elicit l-hour leucopenia. In the present investigation, however, Gey’s solu- tion, at 0.01 ml per gram, accelerated fail- ure in fresh water in t,he same manner as 0.6% NaCl (Exp. 2).
Burden reported that preadaptation to fresh water before hypophysectomy did not delay failure. Furt,hermore, he found that hypophysectomized F. heteroclitus cannot, survive indefinitely in diluted sea water, even at a concentration essentially isos- motic with the blood. The experiment nou reported (Exp. 3) on gradual dilution sup- ports these findings. Failure occurred at a mean salinity of 0.2670. Such a medium was not tolerated in Burden’s experiments. It is t’herefore clear that hypophysectomized F. heteroclitus cannot adapt themselves to gradual dilution of salt water.
Renal lithiasis, resulting from the forma- tion of carbonate apatite calculi, is of fre- quent occurrence in hypophysectomized F. heteroclitus (Pickford and Ate, 1957). Ball and Kallman (1962) have observed the same thing in hypophysectomized P. (44.) for~zoso. The presence or absence of visible calculi in the excretory ducts was studied at autopsy and is recorded in Tables 2-13 for every experimental fish. There is no apparent correlation between failure or sur- vival in fresh water and the incidence of calculi. Calculi were present in at least some fish after prolonged treatment with effective hormones, such as sheep prolactin, and equally so after prolonged treatment with ineffective crude carp preparations.

FRESH-WATER TOLERANCE OF FLWDULUS 175
These observations exclude partial obstruc- tion of the urinary ducts as a cause of failure.
NEUROHYPOPHYSIAL PRINCIPLES
The possible participation of the neuro- hypophysis in teleostean osmoregulation has been reviewed by Maetz (1963) and Dodd and Perks (1964). Sodium influx in goldfish increases within an hour aft,er intraperi- toneal injection of oxytocin (0.4 mU per gram) (Maetz, 1963). Diuresis also occurs during the first 2 hours after injection. This response can be elicited with lower doses, 0.05 and 0.035 mU per gram, respectively, of AVT or whole pollack pituitary extract. According to these results, teleostean neuro- hypophysial preparations have a dual effect on the intact goldfish: transitory diuresis, with concomitant loss of sodium, offset by a more lasting increase in sodium uptake from the external medium.
In respect to euryhaline marine teleosts, Motais and Maetz (1964) report that high doses of oxytocin increase both outflux and influx of sodium in the flounder, Platichthys flesus, in fresh water. The first response is sodium loss and is followed by increased uptake. These authors also report that oxy- tocin increases sodium loss by flounders transferred from fresh to salt water. They conclude that neurohypophysial hormones promote sodium flux across the gills against the existing osmotic gradient, i.e., inward in the goldfish in fresh water and outward in the flounder in salt water.
The experiments of Meier and Fleming (1962) on intact Fundulus kansae are no less perplexing. Oxytocin, at high doses, seemed to have effects that would promote either fresh- or salt-water adaptation.
Other investigators suggest that oxytocin may increase sodium loss by teleosts in fresh water. Holmes and McBean (1963) report that oxytocin and vasopressin mark- edly enhance glomerular filtration rate in rainbow trout. Unfortunately they did not collect urine for sodium determinations. The increase in filtration rate observed by Maetz (1963) in goldfish injected with oxy- tocin and arginine vasotocin, and the in-
creased filtrat,ion rate in African lungfish (Sawyer, unpublished data) in response to arginine vasotocin and oxytocin, are both associated with sodium loss. Bentley and Follett (1963) also report that arginine vasotocin causes increased renal sodium loss by river lampreys. It is possible, there- fore, that neurohypophysial peptides in- jected into intact fishes act primarily to increase renal salt loss. Whether such re- sponses have any physiological meaning remains questionable (Bentley and Follett, 1963).
Our results with the teleostean neurohy- pophysial peptides, arginine vasotocin and isotocin, give no support to the hypothesis that these substances play a significant role in the maintenance of fresh-water survival in hypophysectomized F. heteroclitus. The negative results report,ed by Burden (1956) could be questioned since he employed a pharmacological dose of ox posterior lobe powder, but in our experiments the doses were low. Furthermore, there is no correla- tion between the posterior lobe activities of adenohypophysial preparations and their ability to maintain fresh-water survival (Fig. 2). Ovine prolactin, which was highly effective (Exps. 9 and lo), had little or no antidiuretic or oxytocic activit,ies; the same is true of bovine prolactin which was at least partially effective (Exp. 11). The two primate growth hormone preparations, both partially effective (Exps. 12 and 13), were more or less heavily contaminated; the simian preparation, in particular, contained such an amount that each fish received 46 mU oxytocic activity per gram. Ineffective doses of carp pituitary preparations (Exps. 14 and 15) contained neurohypophysial activities of the same order as the effective human growth hormone preparation.
Negative results could have been antici- pated. It is certain that in Fundulus, as in other species, the hypophysectomized fish are not deprived of hypothalamic neurose- cretions whose release may be mediated, as in the eel (Arvy et al., 1955), by osmotic stress. No direct comparison can be made with the work of Maetz and other investi- gators, who employed intact animals. In

176 PICKFORD, ROBERTSON. AND SA\VYIGR
such, the role of the neurohypophysial prep- ‘arations could be indirect, i.e., mediated through the release of some effective a&no- hypophysial hormone.
26 I
l sP l HGH 1
20 @BP
h4GHa i
OXYTOCIC ACTIVITY, mU/g.day
MGH
I BGH i
&OLACTIN !&TMTY, 30
mU/g.day 100
FIG. 2. Survival of hypophysectomized F. hetc,.- oclitus in fresh water and the influence of replare- ment therapy. Each point represents mean survival time after transfer to fresh water. Survival is plott’ed in the upper diagram against daily dose of neurohypophysial principles as indicated by oxy- tocic assays. In the lower diagram survival is
plotted against dose of prolactin activity detclr- mined by crop gland assays in the same prepara- tions. All trials terminated at 28 days. The frac- tions shown above the arrows indicate the number
of fish still healthy at 28 days over the total num- ber of fish tested. The vertical lines indicate the range of failure times within each treated group. Abbreviations: SP = sheep prolactin; BP = beef prolact,in; HGH = human growth hormone; MGH = monkey growth hormone; BGH = beef growth hormone; Cl and C2 = carp pituitary, June and August: FA and FB = Fc714A and B; TL- = Tilnpia urophysis; AVT = arginine vaso- torin : TOP = 4-serine, S-isoleucine osytocin.
UROPHYHIAL EXTRACT
The experiments described under Group IV can be subdivided into several categor- ies. In the first place, it was decisively proven that preparat’ions containing pigeon crop gland stimulating activity maintain fresh-water survival. Limited success, re- ported in an earlier investigation with ovine prolactin (Pickford and Phillips, 1959) can perhaps be attributed to the high dose (350 mU per gram). Almost complete suc- cess was obtained, in the present investiga- tion, with 105 mU per gram (Exps. 9 and 10). Experiment 10 also explored the possi- bility that a long period of hormonal prc- treatment might favor fresh-water survival. It was thought that prolactin might elicit a slow morphological change, such as the restoration of normal gill histology, as contrasted to a more rapid physiological action. However, the only fish that failed in fresh water was a member of the pre- treated group. A rapid cytological change, taking place w&in 24 hours, may occur.
The experiments with Tila~u urophysis, The experiments discussed above em-
reported in Group III, must be considered ployed purified ovine prolactin. An experi- in the light of controversial information ment with bovine prolactin (Exp. 11)) at a regarding the supposed osmoregulatory similar dose, was less successful although
function of the tcleostcan caudal IICUI’O- secretory storage-release center I Enami, 1956; Enami et al., 1956; Takasugi and Bern, 1962). Holmgren (1964) has reported briefly that urophysectomized F. hetero- clitus are less able to withstand transfer from salt to fresh water than the controls. Maetz et al. (1963, 1964) found that t’he injection of saline extracts of fresh goldfish urophysis into unstressed goldfish has :m
effect similar to that previously reported for oxytocin, arginine vasotocin, and fish pitui- tary preparations: an increased uptake of sodium t,hrough the gills and an increase in urine flow. However, their assays and those of Sawyer and Bern (1963) suggest that the active agent is not a known neurohy- pophysial peptidc. Since Maetz et al. (1964) obtained negative results wit,11 a boiled acetic extract of a&one-dried urophyses, the interpretation of our negative results with saline extracts of acetone-dried glands must remain in doubt,.
ADENOHTPOPHTSIAL PREPARATIONS

FRESH-XATER TOLERANCE OF FUNDULUS 177
t,he treatment was obviously beneficial. However, the bovine prolactin lacked a collateral activity which, in the case of ovine prolactin, stimulates melanin synthe- sis (Pickford and Kosto, 1957; Kosto et al., 1959). There is little reason to suspect the presence of significant amounts of known active agents in either the ovine or bovine prolactins employed in the present investi- gation (Table 1). The two prolactins are stated to be closely similar, but minor dif- ferences have been reported (Li, 1961). However, according to Dr. A. E. Wilhelmi (personal communication), the bovine pro- lactin was prepared by a different method. Furt.her investigations are under considera- tion.
Two experiments were done with primate growth hormone preparations containing prolactin activity. Some investigators be- lieve that this property is inherent in the primate growth hormone molecule (Wallace and Ferguson, 1961), but this has been disputed (Brauman and Brauman, 1963). For our purposes the cont,roversy is imma- terial. Before embarking on an experiment with primat.e growth hormone, we confirmed the findings of Burden (1956) that purified beef growth hormone is essentially ineffec- tive (Exp. 7). A trace of prolactin contained in the preparation, at the dose employed 8 mU per gram weight of fish, was seem- ingly sufficient to offset the accelerated times of failure that are regularly observed after saline injections (Table 3) or other ineffective treatments. Burden (1956) em- ployed a different beef growth hormone preparation. It had no such minor beneficial action, but its prolactin assay is unknown.
.4s a further control, we studied the effect of combining bovine growt,h hormone with ovine prolactin (Exp. 8). Prolactin was administered at a high dose similar to that known from an earlier investigation to be partially but not completely effective (Pickford and Phillips, 1959). Simultane- ous administ.ration of bovine growth hor- mone conferred no greater advantage.
With this information at our disposal, and with the knowledge that ovine prolactin at 5 pg per gram was highly effective (Exp. 91, we started an experiment with simian
growth hormone (Exp. 12). The dose was such that the fish received 40 mU per gram of prolactin activity per injection. The re- sults demonstrate, beyond reasonable doubt, that simian growth hormone tends to pro- mote fresh-water survival. Four of the five recipients survived for 18-22 days and con- tinued to cat for a greater part of this time. Ultimate failure might be attributed to various causes: the relatively low dose of prolactin, the presence of contaminating hormones, and the possibility of species specificit,y. Simian growth hormone, in con- trast to bovine growth hormone, elicits a poor growth response in F. heteroclitus (Pickford et al.. 19591. It is of interest that the fish went, dark, as with ovine prolactin, demonstrating the presence of the melano- genesis fact.or.
Human growth hormone gave strikingly favorable results (Exp. 131. Four out of six recipients were in excellent health and were still eating normally after 28 days in fresh water. In comparing these results with the somewhat less successful response to simian growth hormone, one may observe, first., that the dose of prolactin was higher (80 mU as compared with 40 mU per gram). Since the minimum effective dose for pro- lactin, from anv source, has not been estab- lished, the significance of this difference is a matter of speculation. In the second place, the human growth hormone was heavily contaminated with gonadotropin and thyro- t,ropin, but cont’ained lesser amounts of vasopressor and oxytocic act,ivities. None of these hormones have been shown to have any beneficial action, but deleterious effects of chronic treatment wit.h neurohypophysial preparations cannot, be tliscount.ed. Possible species differences between simian and human prolactine are an unknown factor, but human growth hormone, like simian growth hormone, elicits a poor growth re- sponse in hypophysectomized F. heteroclitua (Pickford et al., 1959). The human growth hormone, like the monkey preparation, con- tained the hormone of melanogenesis, but unlike the latter, it evidently contained intermedin which stimulated the multiplica- tion of new melanocyt,es. Intermedin, per se, has not been etnployed in fresh-water sur-

178 PICKFORD, IiOBE11TSOK7 AND SAW1-14:Ii
viva1 tests, but it is not present in sheep prolactin preparations that have proved beneficial: it was absent from the C. H. Li preparation employed by Pickford and Phillips (1959) and may be presumed ab- sent in NIH-P-S4 since no proliferation of pigment cells \vas observed.
At this point one might summarize the results as showing that mammalian adeno- hypophysial preparations with crop gland stimulating activity favor fresh-water sur- vival (Fig. 2). The best results, with highly purified sheep prolactin, appear to be decisive.
The final experiments, in which an at- tempt was made to demonstrate prolactin- like activity in carp pituitary preparations (Exps. 14 and 15), were disappointing. Neither whole lyophilized carp pituitary powder, at low or high doses, nor fractions prepared by a standard method for the sep- aration of mammalian prolactin, gave any indication of activity. Recipients failed to darken significantly although, in an earlier experiment (Pickford, unpublished data) some darkening had been observed after administration of lyophilized carp pituitary powder at a high dose. If the melanogenesis factor was present in the experiments rc- ported here, it must have been in such low amounts as to be completely masked by the simultaneous presence of t,he melano- phore-concentrating hormone. Survival tests in fresh water gave no evidence of favor- able, prolactin-like action. The resu1t.s are the more surprising since prolactin-like activity has been demonstrated in carp pituitary preparations by the red eft test (Grant and Pickford, 1959). The prepara- tions gave negative results for prolactin by the pigeon crop gland test (Wilhelmi, per- sonal communication) but, since it is now known that no prolactin-like activity can be demonst.rated by this method in fish pituitary extracts (Nicoll and Bern, 1964), this result is meaningless, Clearly further studies are needed with teleostcan prepara- tions that show high prolactin-like activity by the red eft test.
CONCLUSION
In reviewing the negat,ive results with neurohypophysial peptides and Tilapin
urophysi*, it wits pointed out that’ suc11 preparations might participate in osmoreg- ulation by intact fish. Our negative results On liypopliysecton?iz~~~ fish do not rule alit this possibility. In fish, as in mammals, the release of adenoliypophysial hormones i:: probably regulated by neurohypophysial- like peptides arising in the hypothalamus, or in other neurosecrctory systems. This hypothesis receives support from the work of Ball and his collaborators on the function of t’he transplanted pituitary in hypophy- sectomized Poe&a (Mollienesia) formosa. discussed below, and from the work of Roy (1963). Roy reported that the urophysis of Indian carp contains a corticotropin rcleas- ing factor, effective either on fish or labora- tory mammals. There is, of course, no rea- son to believe that stimulation of ACTH release in F. heferoclitus would favor frcsh- water adaptation. Neither ACTH nor adrenal corticoids, at presumed physio- logical doses, enabled hypophysectomized recipients to withstand fresh water (Burden, 1956; Pickford and Phillips, 1959).
Intcrcst. therefore turns to t,lic mecha- nisms regulating the storage and release of the presumed prolactin-like hormone of the teleostean hypophysis. The presence of crythrosinophile cells, resembling mam- malian luteotrophs, is now well established (Olivercau, 1963). Moreover, these cells show hypcrtrophy and hyperplasia in eury- haline species t.hat have been adapted to fresh water. Olivereau and Ball (1963) demonstrated t,his response in Poecilia (dlollienesia) latipinna, and more recently, convincing changes have been observed in the hypophysis of fresh-water adapted F. hetero4ifu.s (Ball and Pickford, 1964). To this may be added the observations of Ball on the regenerated adenohypophysis of Fish 2 in Exp. 1. Studies on hypophysectomized P. (M.) formosa have shown that the pres- ence of pituitary t,ransplants enables this species to live in fresh water (Ball and Kallman, 1962) and erythrosinophile cells can be demonstrated in the grafted gland (Olivcreau and Ball, 1963). Normal hy- pothalamic connect,ions are therefore not necessary, in this teleost, for the storage and release of the hypothetical prolactin- like hormone. We arc not in a position t,o

FRESH-WATER TOLERANCE OF FUNDULUS 179
state that the hypothalamus exerts an in- hibitory action, as in tetrapods. The com- plex problems of prolactin release in higher vertebrates are reviewed by Jorgensen and Larsen (1963). The demonstration by Masur (1962) that transplanting the red eft pituitary initiates the water drive re- sponse is of particular interest.. The im- portant point which we wish to make is that all experiments on the endocrine control of salt and water metabolism in teleosts, as in other vertebrates, must take into considera- tion that investigations on intact animals carry an inherent limitation and must be evaluated accordingly.
REFERENCES ARVY, L., FONTAINE, M., AND GAEE, M. (1955). Ac-
tions des solutions salines hypertoniques sur la neurosecretion hypothalamique chez I’anguilie hypophysectomisee. Con@. Rend. Sot. Biol. 149, 22.5228.
BALL, J. N. (1962). Brood-production after hy- pophysectomy in the viviparous teleost MoZZi- enesia latipinua Le Sucur. N&lx 194, 787.
B.~LL, J. N., AND KALLMAN, K. D. (1962). Func- tional pituitary transplants in the all-female, gynogenetic teleost, Mollienesia jormosa (Gir- ard). Am. Zool. ‘2, 389 (Abstract)
B.~LL, J. N., OLIVEREAU, M., AND KALLM.~PI’, K. D. (1963). Secretion of thyrotrophic hormone by pituitary transplants in a teleost fish. Na(/(re 199, 618-620.
BALL, J. N., AND PICKFORD, G. E. (1964). Pituitary cytology and fresh water adaptation in the killi- fish, Fund&s heteroclitus. Anat. Record 148, 358 (Abstract).
BATES, R. W.. AA-D RIDDLE, 0. (1935). The prepara- tion of prolactin. J. Pharmacol. 55, 365-371.
BENTLEY, P. J., AND FOLLETT, B. K. (1963). Kidney function in a primitive vertebrate, the cyclos- tome, Lampetra fluviatilis. J. Physiol. 169, 902- 918.
BERDE, B., HUGUESIN, R., .~ND ST~~RMER, E. (1962). The biological activities of arginine-vasotocin obtained by a new synthesis. Ezperienlia 18, 444-445.
BRAUMAN, J., AND BRAUMAN, H. (1963). Growth hormone secretion measured by immunoassay compared with lactogenic activity in the human hypophysis i?l vitro. Gen. Camp. Endoc&oZ. 3, 689 (Abstract).
BURDEN, C. E. (1956). The failure of hypophysec- tomized Fundulus heteroclitus to survive in fresh water. Biol. Bull. 110, t&28.
CALLAMAND, O., FONTAINE, M., OLIVEREAU, M., AND RAFFY, A. (1951). Hypophyse et osmorkgulation
chez les poissons. Bull. Inst. Oceataog. 984, 1-7. CHESTER JONES, I., PHILLIPS, J. G., AND HOLMES,
W. N. (1959). Comparative physiology of the adrenal cortex. Zrc “Comparative Endocrinology” (A. Gorbman, ed.), pp. 582-610. Wiley, New York.
DODD, J. M., END PERKS, A. M. (1964). Neuro- physiological function in lower vertebrates. In “The Pit,uitary” (G. Harris and B. Donovan, eds.), Vol. 3 (in press). Butterworths, London.
EX~MI, M. (1956). Studies in neurosecretion. VIII. Changes in the caudal neurosecretory system of the loach (MisgrLmzLs nng&llicar~datz~s) in re- sponse to osmotic stimuli. Proc. Japun Acutl. 32, 759764.
ENAMI, M., MIY~SHITA, S., AND IMAI, K. (1956). Studies in neurosecretion. IS. Possibility of oc- currence of a sodium-regulating hormone in the caudal neurosecretory system of teleosts. En- docrinol. Japan. 3, 28&290.
FONTAINE, M., CALLAMAND, O., AND OLIVEREAU, M. (1949). Hypophyse et euryhalinite chez l’an- guille. Compt. Rend. Aead. Sci. MS, 513-514.
GRANT, W. C., AND PICKEQRD, G. E. (1959). Pres- ence of red eft water-drive factor prolactin in the pituitaries of teleosts. Biol. Bull. 116, 42% 435.
GUTTMANN, S., BERDE, B., AND ST~RMER, E. (1962). The synthesis and some pharmacological effects of serine’-isoleucine’-oxytocin, a probable neu- rohypophysial hormone. Experientia 18, 445446.
HOLMES, W. N., AND MCLEAN, R. L. (1963). Stud- ies on the glomerular filtration rate of rainbow trout, (Salmo gnirdneri). J. Exptl. Biol. 40, 335- 341.
HOLMCREN, U. (1964). Neurosecretion in teleost fishes: The caudal neurosecretory system. Am. Zool. 4, 37-45.
JGRGENSEN, C. B., .~ND LARSEN, L. 0. (1963). Neuro-adenohypophysial relationships. Symp. Zool. Sot. (London) 9, 59-82.
KOSTO, B., PICKFORD, G. E., AND FOSTER, M. (1959). Further studies of the hormonal induction of melanogenesis in the killifish, Fund,ulus hetero- clitus. Endocrinology 65, 869881.
Lr, C. H. (1961). Biochemistry of prolactin. In “Milk: The Mammary GIand and Its Secre- tion” (S. K. Kon and A. T. Cowie, eds.), Vol. 1, pp. 205208. Academic Press, New York.
MAETZ, J. (1963). Physiological aspects of neuro- hypophysial function in fishes with some refer- ence to the Amphibia. Symp. Zool. Sot. (Lon- don) 9, 107-140.
MAETZ, J., BOURGUET, J., ASD LAHLOUH, B. (1963) Action de l’urohypophyse sur les Cchanges de sodium (&dies It l’aide du “Na) et sur I’excrC- tion urinaire du t&%steen Cnrasaius aumtus. J. Physiol. Park 55, 159-166.

180 PICKFOHD, IWBE:KTSOS, AND SAWYEN
MAETZ, J., BOURGUET, J., AND J,AHLOUH, B. (1964).
Urophyse et osmor&gulation chcz Camssins nnr,r- tus. Gen. Camp. Endocrinol. 4, 401-414.
MAETZ, J., AND JUIEN, M. (misprint for Julic,n. M.) ( 1961). Action of neurohgpophysral hormonc~s on 1,lr.e sodium fluxes of a freshwater tc~lcost. Srrt((rcs 189, 152.
M.~suR, S. K. (1962). Autotransplantation of the pituitary in t,he red eft. Am. Zool. 2, 538 (Ah- stract).
MEIER, A. H., .~ND FLEMING, W. H. (1962). Tho effects of Pitocin and Pitressin on water and sodium movements in the euryhaline killifish, Fundulus knnsne. Comp. Biochem. Physiol. 6, 215-231.
MOTAIJ, It., AND MAETZ, J. (1964). Artion des hor- mones neurohypophysaires sur les &changes dc sodium (mesur& B l’aide du radio-sodium Na”) chez un tdlkostken euryhaline: Plntichthys pescts L. Germ. Comlj. ET&ocriiLol. 4, 210-224.
MTJNSICK, R. A. (1960). Effect of magnesium ion on the response of the rat uterus to neurohg- pophysial hormones and annlogues. Enrlocritrol- ogy 66, 451457.
NICOLL, C. S., .~ND BERN, H. A. (1964). “Prolactin” and thrx pituitary glands of fishes. Gen. Camp. Eutlocrino/. 4, 457-471.
OLIVEKEAU, M. (1963). Cytophysiologie du lobe distal de l’hypophyse des agnathes et des pois- sons, a l’cxclusion de celle concernant la fonc- tion gonadotrope. Colloy. Intern. Centw Natl. Rech. Sci. (Paris) 128, 315-329.
OLIVEHEAU, M., AND BALL, J. N. (1963). Cytophysi- ologic adOnohppophysaire chez un Poisson: Mel-
lienesia spccics. Gen. Comp. Endocrinol. 3, 723 (Abstract).
PICKFORD, G. E. (1953). A study of the hypophy- sectomized male killifish, Fundulus heteroclitus (Linn.). Bull. Binghnm Oceanog. Collect. 14, (2) 541.
PICKFORD, G. E., AND ATZ, J. W. (1957). “The Physiology of t,hc Pituitary Gland of Fishes,” 613 pp. New York Zool. Sot.
PICKFORD, G. E., .~ND KOSTO, B. (1957). Hormonal induction of melanogenesis in hypophysecto-
~‘I~KF~I~. G. I+:.. .\ND PHILIDS, .I. (;. (1959). I’ro- lac*r 111. :I iac+or in promoting survival of II>.- l~ol~ll~,~rr(olrlizc~tl killifish in frrsh watc,r. S’cic,/c,c 130, 454-465.
PICKFORIJ, G. E.. WILHELMI, A. 15.. ASD N[.wB~~x.
N. (1959). Comparative studies of the responrc, of hypophyscctomized kill&h, Flc&frlus hetew- clitlbs, to growth hormone preparations. Annl. Record 134, 624-625 (Abstract).
ROY, B. B. (1963). Histological a,nd experimental observations on the caudal neurosecretory sys- t,em of some Indian fishes. Proc. Natl. Inst. Sci. In&u 28, 449-477.
SAWYER, W. H. (1964). Biological assays ior neuro- hypophysial principles in tissues and in blood. IIL “The Pituitary” (G. Harris and B. Donovan. cds.), Vol. 3 (in press). Butterworths, London.
S.~WTEH. W. H.. .\ND BERN, H. A. (1963). Examina- tion of the caudal neurosecretory -;yst,em of Il’iEapicc naossc~mbica (Teleostei Cichlitlne) fo1 the presence of neurohypophyseal-likrr :rc+iviby. Am. Zool. 3, 555556 (Abstract).
SCHREIBMAS, M. P., AND KALLi+lAN, K. D. (lQ63). Effects of hypophysectomy on Sipho/jhor/cs. Am. Zool. 3, 556 (Abstract).
SLICHER, A. M. (1958). The effect of ACTH, corti- sol and cold shock on the peripheral white cell count of fishes. Anot. Record 131, 600 (Ah-
&art). SLICHER, A. M. (1961). Endocrinological and
hematological studies in FundlJus heteroclitus (Linn.). BnZZ. Biagham Clccnnog. Collect. 17,
(3) l-55. ,SLICHER, 8. M., PICKFORD, G. E., AND BALL, J. X.
(1962). Effects of ACTH and cold shock on the white cell count of fishes. Annt. Record 142, 327 (8bstract).
T.4KASuG1, N., AND BERN, H. A. (1962). Experi- mental studies on the caudal neurosecretory sys- tem of T&pin wlossnmbica. Comp. Biochem. Phy.sioZ. 6, 28S303.
W.~LL.~CE, B. J,. C., AND FERGUSON, K. A. (1961). Preparation of human growth hormone. J. Endocrinol. 23, 285-290.




















