
Hv ZIP7 mediates zinc accumulation in barley (Hordeum vulgare)
13
HvZIP7 mediates zinc accumulation in barley (Hordeum vulgare) at moderately high zinc supply Jingwen Tiong 1 , Glenn K. McDonald 2 , Yusuf Genc 2 , Pai Pedas 3 , Julie E. Hayes 1 , John Toubia 1 , Peter Langridge 1 and Chun Y. Huang 1 1 Australian Centre for Plant Functional Genomics, The University of Adelaide, Hartley Grove, Urrbrae, SA 5064, Australia; 2 School of Agriculture, Food and Wine, University of Adelaide, Glen Osmond, SA 5064, Australia; 3 Plant and Soil Science Section, Department of Agriculture and Ecology, University of Copenhagen, Copenhagen, Denmark Author for correspondence: Chun Y. Huang Tel: +61 8 8303 7160 Email: [email protected] Received: 26 June 2013 Accepted: 29 July 2013 New Phytologist (2013) doi: 10.1111/nph.12468 Key words: accumulation, Hordeum vulgare (barley), low affinity, translocation, trans- porter, uptake, zinc (Zn), ZIP. Summary High expression of zinc (Zn)-regulated, iron-regulated transporter-like protein (ZIP) genes increases root Zn uptake in dicots, leading to high accumulation of Zn in shoots. However, none of the ZIP genes tested previously in monocots could enhance shoot Zn accumulation. In this report, barley (Hordeum vulgare) HvZIP7 was investigated for its functions in Zn trans- port. The functions of HvZIP7 in planta were studied using in situ hybridization and transient analysis of subcellular localization with a green fluorescent protein (GFP) reporter. Transgenic barley lines overexpressing HvZIP7 were also generated to further understand the functions of HvZIP7 in metal transport. HvZIP7 is strongly induced by Zn deficiency, primarily in vascular tissues of roots and leaves, and its protein was localized in the plasma membrane. These properties are similar to its closely related homologs in dicots. Overexpression of HvZIP7 in barley plants increased Zn uptake when moderately high concentrations of Zn were supplied. Significantly, there was a specific enhancement of shoot Zn accumulation, with no measurable increase in iron (Fe), manganese (Mn), copper (Cu) or cadmium (Cd). HvZIP7 displays characteristics of low-affinity Zn transport. The unique function of HvZIP7 provides new insights into the role of ZIP genes in Zn homeostasis in monocots, and offers opportunities to develop Zn biofortification strategies in cereals. Introduction Zinc (Zn) is an essential micronutrient for all organisms. It is involved in many biochemical processes (Welch, 1995; Maret, 2004). All organisms must maintain adequate Zn in their cells. To achieve sustained Zn uptake from fluctuating environments, plants and other eukaryotes are equipped with a dual-transporter system, which consists of both high-affinity and low-affinity Zn transporters (Eide, 2006). In yeast, the high-affinity transporter gene (Zrt1) is responsible for uptake of Zn in Zn-limiting media. When Zn is abundant in external media, Zrt1 is repressed and the low-affinity transporter (Zrt2) mediates Zn uptake. Zrt1 has a remarkably high affinity for Zn with an estimated apparent K m of 10 nM for free Zn 2+ ions, whereas Zrt2 has a lower affinity of apparent K m of c. 100 nM (Eide, 2006). The single-celled alga, Chara corallina, is similar to yeast, and also has a dual transporter system for Zn influx (Reid et al., 1996). The high-affinity system saturates at c. 0.1 lM, and the low-affinity system shows a linear dependence on Zn concentration from 0.5 lM up to at least 50 lM (Reid et al., 1996). A kinetic study of Zn influx in a higher plant, wheat, also shows the presence of a dual Zn uptake system (Hacisalihoglu et al., 2001). Apparent K m values of the high-affinity system are between 0.6 and 2 nM, while the K m values of the low-affinity system are between 2 and 5 lM (Hacisalihoglu et al., 2001). Soils with low available Zn occur over a significant proportion of the world’s agricultural area (Alloway, 2004), which not only affects grain yield but also the Zn concentration of the grain. Low Zn in cereal grains exacerbates Zn deficiency in humans (Cunningham-Rundles et al., 2005; Hotz et al., 2005), and con- sequently increasing grain Zn content in cereals (biofortification) is considered to be important for the alleviation of human Zn deficiency (Welch & Graham, 2004). Understanding the func- tional role of the major genes that control Zn uptake and trans- port is important for improvement of Zn nutrition in plants and enhancement of Zn concentrations in grains. The Zn-regulated, iron-regulated transporter-like protein (ZIP) family is considered to be the primary group of transporters controlling plant Zn influx at the plasma membrane (Grotz et al., 1998; Guerinot, 2000; Eide, 2006). There are 15 members of the ZIP gene family identified in the Arabidopsis thaliana genome (Grotz & Guerinot, 2006), and 16 have been identified in the rice genome Ó 2013 Australian Centre for Plant Functional Genomics New Phytologist Ó 2013 New Phytologist Trust New Phytologist (2013) 1 www.newphytologist.com Research
Transcript of Hv ZIP7 mediates zinc accumulation in barley (Hordeum vulgare)

HvZIP7 mediates zinc accumulation in barley (Hordeum vulgare)at moderately high zinc supply
Jingwen Tiong1, Glenn K. McDonald2, Yusuf Genc2, Pai Pedas3, Julie E. Hayes1, John Toubia1, Peter Langridge1
and Chun Y. Huang1
1Australian Centre for Plant Functional Genomics, The University of Adelaide, Hartley Grove, Urrbrae, SA 5064, Australia; 2School of Agriculture, Food and Wine, University of Adelaide,
Glen Osmond, SA 5064, Australia; 3Plant and Soil Science Section, Department of Agriculture and Ecology, University of Copenhagen, Copenhagen, Denmark
Author for correspondence:Chun Y. Huang
Tel: +61 8 8303 7160Email: [email protected]
Received: 26 June 2013
Accepted: 29 July 2013
New Phytologist (2013)doi: 10.1111/nph.12468
Key words: accumulation, Hordeum vulgare(barley), low affinity, translocation, trans-porter, uptake, zinc (Zn), ZIP.
Summary
� High expression of zinc (Zn)-regulated, iron-regulated transporter-like protein (ZIP) genes
increases root Zn uptake in dicots, leading to high accumulation of Zn in shoots. However,
none of the ZIP genes tested previously in monocots could enhance shoot Zn accumulation.
In this report, barley (Hordeum vulgare) HvZIP7 was investigated for its functions in Zn trans-
port.� The functions of HvZIP7 in planta were studied using in situ hybridization and transient
analysis of subcellular localization with a green fluorescent protein (GFP) reporter. Transgenic
barley lines overexpressing HvZIP7 were also generated to further understand the functions
of HvZIP7 in metal transport.� HvZIP7 is strongly induced by Zn deficiency, primarily in vascular tissues of roots and leaves,
and its protein was localized in the plasma membrane. These properties are similar to its
closely related homologs in dicots. Overexpression of HvZIP7 in barley plants increased Zn
uptake when moderately high concentrations of Zn were supplied. Significantly, there was a
specific enhancement of shoot Zn accumulation, with no measurable increase in iron (Fe),
manganese (Mn), copper (Cu) or cadmium (Cd). HvZIP7 displays characteristics of low-affinity
Zn transport.� The unique function of HvZIP7 provides new insights into the role of ZIP genes in Zn
homeostasis in monocots, and offers opportunities to develop Zn biofortification strategies in
cereals.
Introduction
Zinc (Zn) is an essential micronutrient for all organisms. It isinvolved in many biochemical processes (Welch, 1995; Maret,2004). All organisms must maintain adequate Zn in their cells.To achieve sustained Zn uptake from fluctuating environments,plants and other eukaryotes are equipped with a dual-transportersystem, which consists of both high-affinity and low-affinity Zntransporters (Eide, 2006). In yeast, the high-affinity transportergene (Zrt1) is responsible for uptake of Zn in Zn-limiting media.When Zn is abundant in external media, Zrt1 is repressed andthe low-affinity transporter (Zrt2) mediates Zn uptake. Zrt1 hasa remarkably high affinity for Zn with an estimated apparent Km
of 10 nM for free Zn2+ ions, whereas Zrt2 has a lower affinity ofapparent Km of c. 100 nM (Eide, 2006). The single-celled alga,Chara corallina, is similar to yeast, and also has a dual transportersystem for Zn influx (Reid et al., 1996). The high-affinity systemsaturates at c. 0.1 lM, and the low-affinity system shows a lineardependence on Zn concentration from 0.5 lM up to at least50 lM (Reid et al., 1996). A kinetic study of Zn influx in ahigher plant, wheat, also shows the presence of a dual Zn uptake
system (Hacisalihoglu et al., 2001). Apparent Km values of thehigh-affinity system are between 0.6 and 2 nM, while the Km
values of the low-affinity system are between 2 and 5 lM(Hacisalihoglu et al., 2001).
Soils with low available Zn occur over a significant proportionof the world’s agricultural area (Alloway, 2004), which not onlyaffects grain yield but also the Zn concentration of the grain.Low Zn in cereal grains exacerbates Zn deficiency in humans(Cunningham-Rundles et al., 2005; Hotz et al., 2005), and con-sequently increasing grain Zn content in cereals (biofortification)is considered to be important for the alleviation of human Zndeficiency (Welch & Graham, 2004). Understanding the func-tional role of the major genes that control Zn uptake and trans-port is important for improvement of Zn nutrition in plants andenhancement of Zn concentrations in grains. The Zn-regulated,iron-regulated transporter-like protein (ZIP) family is consideredto be the primary group of transporters controlling plant Zninflux at the plasma membrane (Grotz et al., 1998; Guerinot,2000; Eide, 2006). There are 15 members of the ZIP gene familyidentified in the Arabidopsis thaliana genome (Grotz & Guerinot,2006), and 16 have been identified in the rice genome
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013) 1
www.newphytologist.com
Research

(Narayanan et al., 2007; Chen et al., 2008). Only four barley ZIPgenes (HvIRT1, HvZIP3, HvZIP5 and HvZIP8; Pedas et al.,2008, 2009) and one from the tetraploid emmer wheat (TdZIP1;Durmaz et al., 2010) have been described so far in temperatecereals, which are major staple foods for humans (Graham &Welch, 2001). Therefore, a better understanding of the contribu-tion of ZIP transporters to Zn homeostasis in cereal plants isimportant for biofortification (Cakmak, 2008; Palmgren et al.,2008; Zhao & McGrath, 2009; Waters & Sankaran, 2011).
The complementation of yeast mutants defective in both thehigh-affinity transporter, Zrt1, and low-affinity transporter, Zrt2,has been a key tool in characterization of the Zn transport abilityof plant ZIP family transporters (Grotz et al., 1998; Ramesh et al.,2003; Ishimaru et al., 2005; Pedas & Husted, 2009; Pedas et al.,2009; Lee et al., 2010a; Stephens et al., 2011). Kinetic studies onthese plant ZIP transporters by expression in yeast indicate thatapparent Km values are between 2 and 18.5 lM Zn (Grotz et al.,1998; Ramesh et al., 2003; Stephens et al., 2011); these values aresimilar to the Km range determined in planta for low-affinity trans-port systems (Reid et al., 1996; Hacisalihoglu et al., 2001).Whether these plant ZIP transporters are high- or low-affinity Zntransporters remains to be elucidated in planta, although tissue-specific expression using promoter-reporters suggests that some ofthese ZIP transporters may be high-affinity (Milner et al., 2012).
Studies of metal hyperaccumulating dicotyledonous species,such as Noccaea caerulescens (previously known as Thlaspicaerulescens) and Arabidopsis halleri, show that high expression ofmembers of the ZIP family, NcZNT1, AtZIP4 and AtIRT3, con-tributes to high Zn accumulation (Hanikenne et al., 2008; Linet al., 2009; Milner et al., 2012). By contrast, overexpression ofOsZIP4, OsZIP5 and OsZIP8 in rice increased Zn concentrationin roots, but reduced Zn accumulation in shoots and grains (Ishi-maru et al., 2007; Lee et al., 2010a,b). It is not known if anymembers of the ZIP family in monocotyledonous species can leadto increased Zn accumulation in shoots and/or grains if expres-sion is enhanced.
In this report, we describe the identification of additional ZIPgenes from the barley genome, and functional characterization ofone HvZIP gene, HvZIP7, in planta. Phylogenetic analysis showsthat HvZIP7 is closely related to NcZNT1, AtZIP4 and AtIRT3.HvZIP7 is highly induced in both roots and shoots by Zn defi-ciency, and its protein is localized in the plasma membrane.Overexpression of HvZIP7 in barley increases root uptake and/orroot to shoot translocation of Zn, but only at moderately highconcentrations of Zn supply. Our results provide new insightsinto the functional role of ZIP genes in Zn homeostasis in mono-cotyledonous species.
Materials and Methods
Phylogenetic analysis
Sequences of 16 ZIP family members from rice (Oryza sativa L.)and 18 from Arabidopsis (Arabidopsis thaliana L.) were used tosearch for homologous sequences in barley (Hordeum vulgare L.),Brachypodium (Brachypodium distachyon L.) and wheat
(Triticum aestivum L.). Rice and Arabidopsis ZIP family mem-bers were retrieved from the Rice Genome Annotation ProjectDatabase release 6.1 (http://rice.plantbiology.msu.edu/) at Mich-igan State University and from A. thaliana genome annotationdatabase release 9 (http://www.arabidopsis.org/) at The Arabid-opsis Information Resource (TAIR). Similarity searches were per-formed primarily by BLASTp (Altschul et al., 1997, 2005)querying the JGI v1.0, 89 assembly of Brachypodium at Phyto-zome, version 7 (http://www.phytozome.net/), and the nonre-dundant database of proteins at NCBI (http://blast.ncbi.nlm.nih.gov/Blast.cgi) for barley and wheat. ZIP protein sequences werealigned by standalone MAFFT v6.905b using the L-INS-Imethod with associated default parameters (Katoh et al., 2009),and imported into ClustalW2 v2.1 (Larkin et al., 2007). The un-rooted tree was generated in ClustalW2 using the neighbor-join-ing method with all parameters set to default. One thousandbootstrap datasets were generated to estimate the confidence lim-its of nodes. The tree was visualized using the Molecular Evolu-tionary Genetics Analysis (MEGA) package, version 5.05(Tamura et al., 2011).
Transcript analysis
Barley (cv Lofty Nijo) seedlings used for examining effects of Zndeficiency on transcript abundance of HvZIP7 (Fig. 1a) weregrown in hydroponics with 0.005 (�Zn) or 0.5 lMZn (+Zn) for14 d. In brief, barley seeds were surface-sterilized with 70% ethanolfor 1 min, 3% hypochlorite for 5 min, rinsed with deionized waterand incubated in Petri dishes for 2 d at room temperature. Seedswith emerged radicles were put into a seedling cup which wasplaced in the lid of a black plastic container. Each container con-tained four seedling cups and was filled with 1 l of nutrient solu-tion. Each cup contained two plants. The basal nutrients were asfollows (in lM); Ca(NO3)2, 1000; KNO3, 1000; NH4H2PO4,100; MgSO4, 250; KCl, 50; H3BO3, 12.5; Fe-HEDTA, 10;MnSO4, 0.4; CuSO4, 0.1; NiSO4, 0.1; and H2MoO4, 0.1. 2-[N-morpholino] ethane-sulfonic acid-KOH of 2 mM was used tobuffer pH to 6.0. Macronutrients and micronutrients were sup-plied at half and full strength, respectively, until D9, and fullstrength of all nutrients thereafter. The nutrient solution was aer-ated continuously and replaced at D9 and D12, respectively.Plants were grown in a growth room at 20 : 15°C day : nighttemperature and with a photoperiod of 14 : 10 h day : night at300 lmol m�2 s�1 photon flux intensity at the plant level. AtD14, plants were harvested. Roots were briefly rinsed in deion-ized water, separated from shoots, and excess water was blottedonto fresh laboratory tissues. The roots and shoots were frozenimmediately in liquid nitrogen for transcript analysis.
RNA isolation and quantitative real-time reverse transcriptionpolymerase chain reaction (RT-PCR) analysis of transcripts wereconducted as described by Huang et al. (2011). The transcriptabundances of four control genes (barley a-tubulin, heat shockprotein 70, glyceraldehyde-3-phosphate dehydrogenase andcyclophilin) were determined for all cDNA samples, and themost similar three of these four genes were used as normalizationcontrols. The primers 5′-TGGAAGGCATCCTCGACTCTG-3′
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist2
and 5′- CAATCAGATGGACACAGGCACAT-3′ were used forendogenous HvZIP7, and the primers 5′- TACTCATATACATGGCGCTGGT-3′ and 5′- TTTATTGCCAAATGTTTGAACG-3′ were used for the HvZIP7 transgene. The normalizedcopies lg�1 RNA were used to represent transcript abundances.For semiquantitative RT-PCR analysis (Supporting Information,
Fig. S2b), RNA isolation and cDNA synthesis were the same asdescribed earlier, transgene-specific transcripts were amplifiedfrom cDNA using the primer pair of 5′-GCTGGTTTGTCCTCATTTTACG-3′ and 5′-ATGATAATCATCGCAAGACCG-3′, and the transcripts of the barley glyceraldehydes-3-phosphate dehydrogenase (HvGAPDH) gene were also amplifiedas a loading control using the primer pair of 5′-GTGAGGCTGGTGCTGATTACG-3′ and 5′-TGGTGCAGCTAGCATTTGACAC-3′.
In situ PCR analysis
Barley plants (cvGolden Promise) were grown in hydroponics withno Zn addition. Plant growth conditions were the same as thosedescribed for transcript analysis. Roots and leaves were collectedfrom16-d-old plants. In situPCRwas performed using themethoddescribed by Koltai & Bird (2000) with a fewmodifications. Rootsand leaves were fixed in FAA (63% ethanol, 5% acetic acid and 2%formaldehyde, v/v), embedded in 5% agarose in 19 PBS (w/v),and then sectioned using a microtome (Leica VT1200S,Germany). The sections were treated with DNase (Ambion), fol-lowed by heat inactivation. cDNA synthesis was carried out usingSuperscript III RT (Invitrogen) with a gene-specific primer(5′-CACGTTATCTGATGTATGTATG-3′). PCR reactionswere performed in a volume of 50 ll containing 19 PCR buffer(Invitrogen), 1.5 mM MgCl2, 200 lM dNTP, 0.2 nM digoxige-nin-11-dUTP (Roche), 1.5 U Taq polymerase (Invitrogen) and150 ng of both forward and reverse primers (5′-ACCAGGT-TCTACGAGACCAAG-3′ and 5′-TTAGGAGCGCACGTGTC-3′) for the variable region of theHvZIP7 coding sequence.
Subcellular localization of HvZIP7
HvZIP7 fragments containing the full-length open reading framewere amplified from barley cDNA by PCR with the primer pairof 5′- TCAGGGCATGATGATCGGTGTA-3′ and 5′-CAATCAGATGGACACAGGCACAT-3′. The HvZIP7 fragments werecloned into a pGEM®T-easy vector (Promega) and sequenced.The coding sequence of HvZIP7 without the stop codon wasamplified from pGEM®T-easyHvZIP7 plasmid using the primerpair of 5′-ATGATGATCGGTGTAGCAGGCTTC-3′ and5′-GGCCCAGACTGCAAGCAT-3′. The PCR fragment wasligated into the pCR8-GW-TOPO vector (Invitrogen) and trans-ferred into pMDC83 containing the mGFP gene (Curtis &Grossniklaus, 2003). The resulting plasmid (pMDC83HvZIP7::GFP) places HvZIP7 upstream of mGFP. Transient expression ofHvZIP7::GFP in onion epidermal cells and visualization of greenfluorescent protein (GFP) in subcellular locations were con-ducted as described by Preuss et al. (2010). The plasmolysis ofonion epidermal cells was performed by immersion in 1Msucrose for 1 min before confocal image analysis.
Generation of HvZIP7 overexpressing barley plants
The full-length open reading frame of HvZIP7 was amplifiedfrom the pGEM�T-easyHvZIP7 plasmid by PCR using the
(a)
(b) (c)
(d) (e)
(f) (g)
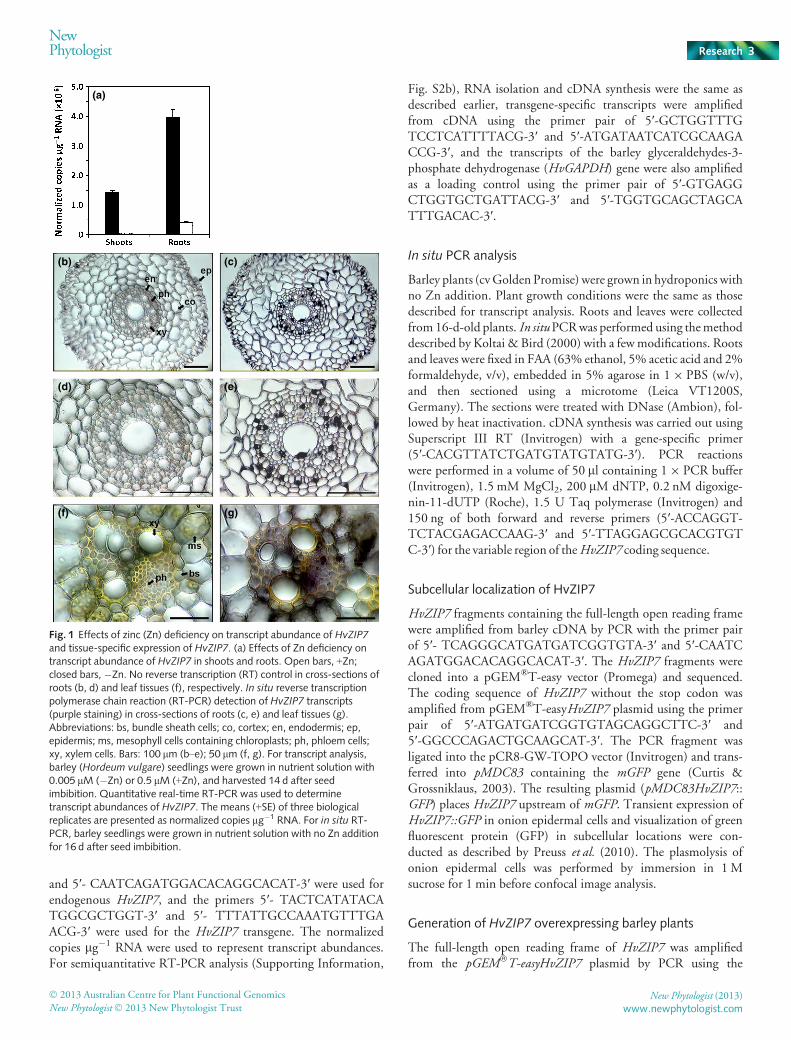
Fig. 1 Effects of zinc (Zn) deficiency on transcript abundance of HvZIP7and tissue-specific expression of HvZIP7. (a) Effects of Zn deficiency ontranscript abundance of HvZIP7 in shoots and roots. Open bars, +Zn;closed bars, �Zn. No reverse transcription (RT) control in cross-sections ofroots (b, d) and leaf tissues (f), respectively. In situ reverse transcriptionpolymerase chain reaction (RT-PCR) detection of HvZIP7 transcripts(purple staining) in cross-sections of roots (c, e) and leaf tissues (g).Abbreviations: bs, bundle sheath cells; co, cortex; en, endodermis; ep,epidermis; ms, mesophyll cells containing chloroplasts; ph, phloem cells;xy, xylem cells. Bars: 100 lm (b–e); 50 lm (f, g). For transcript analysis,barley (Hordeum vulgare) seedlings were grown in nutrient solution with0.005 lM (�Zn) or 0.5 lM (+Zn), and harvested 14 d after seedimbibition. Quantitative real-time RT-PCR was used to determinetranscript abundances of HvZIP7. The means (+SE) of three biologicalreplicates are presented as normalized copies lg�1 RNA. For in situ RT-PCR, barley seedlings were grown in nutrient solution with no Zn additionfor 16 d after seed imbibition.
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 3

primer pair of 5′-CTGGTACCATGGTGATCGGTGTAGCAG-3′ and 5′-TACTCGAGTTCAGGCCCAGACTGC-3′.The PCR product was ligated into the pCR8-GW-TOPO vectorand then transferred into the pMDC32 vector (Invitrogen) by LRreaction according to the manufacturer’s instructions. The result-ing pMDC32HvZIP7 plasmid was used for barley (cv GoldenPromise) transformation using an Agrobacterium-mediatedmethod described by Tingay et al. (1997) and Matthews et al.(2001). The constitutive expression of HvZIP7 was driven by adouble CaMV35S promoter. The number of transgene loci intransgenic plants was determined by standard Southern hybrid-ization (Fig. S2a), and transgene segregation in progenies wasanalyzed by PCR using the transgene-specific primers describedearlier for transcript analysis.
Plant growth and accumulation of Zn and Cd in soil
Zinc-deficient siliceous sand (DTPA-extractable Zn, 0.07 mgkg�1 soil) was used in soil experiments. Calcium carbonate of0.5% (w/w) was added to the sand to simulate calcareous sandysoil for maintaining high pH at 8.0 and reducing bioavailabilityof Zn (Genc et al., 2007). Basal nutrients were as described byGenc et al. (2007). Micronutrients (mg kg�1 soil) includedFeSO4.7H2O (16.8), MnSO4.H2O (3), CuSO4.5H2O (5),H3BO3 (0.3), CoSO4.7H2O (1), MoO3 (0.005), and NiSO4.6H2O(0.15). Four Zn rates (0, 1.0, 5.0, and 12.5 mg Zn kg�1 soil) asZnSO4 were applied to cover plant Zn nutrition range from defi-ciency to luxury. Two barley grains with emerged radicles wereplanted into each pot containing 1 kg soil. Plants were thinned toone per pot 9 d after imbibition (D9). Plants were grown in agrowth room at 20 : 15°C day : night temperature and a photo-period of 14 : 10 h day : night at 300 lmol m�2 s�1 photon fluxintensity at the plant level. At D28, plants were harvested. Rootswere washed free of soil, briefly rinsed in deionized water andexcess water was blotted on fresh laboratory tissues. The rootsand shoots were then oven-dried at 80°C for 48 h for nutrientanalysis.
For Cd accumulation, the experimental setup was similar tothat of the Zn accumulation experiment. Two rates of Cd (3 and10 mg Cd kg�1 soil as CdCl2) were added into the soil with basalnutrients. At D26, shoots were harvested and oven-dried at 80°Cfor 48 h for nutrient analysis.
Plant growth and Zn accumulation in nutrient solution
Plant growth conditions and basal nutrients in nutrient solutionwere the same as described earlier in transcript analysis except forplant number, harvest dates and replacement of nutrient solu-tion. Three of the four seedling cups in each container containedtwo plants for the first three harvests at 14 d after seed imbibition(D14), D15 and D16, and one plant for the last harvest at D20.All plants were grown with 0.5 lM Zn until D14. Four Zn treat-ments (0.5, 5, 10, and 20 lM Zn supplied as ZnSO4) wereapplied at D14. Macronutrients and micronutrients were sup-plied at half and full strength, respectively, until D10, and at fullstrength of all nutrients thereafter. The nutrient solution was
aerated continuously and replaced at D10, D12, D14, and D17,respectively. The pH of the solution was constant at pH 6.0 dur-ing the experiment. Light and temperature settings for plantgrowth were identical to those for soil-grown plants describedearlier. At each harvest, roots were briefly rinsed in deionizedwater and excess water was blotted onto fresh laboratory tissues.The roots and shoots were separated and then oven-dried at 80°Cfor 48 h for mineral element analysis.
Plant growth in potting mix
Plants were grown in pots (15 cm diameter 915 cm height) con-taining 1 kg of potting mix. The potting mix consisted of cocopeat (540 l), sand (60 l), hydrated lime (0.24 kg), agriculturallime (0.6 kg), dolomite lime (calcium magnesium carbonate,0.18 kg), calcium nitrate (0.45 kg), superphosphate (0.18 kg),iron sulfate (0.45 kg), iron chelate (0.03 kg), Microplus (0.18 kg;Langley Fertilizers, Perth, WA, Australia) and mini-Osmocote(1.8 kg), and the pH of the mix was 6.5. Microplus contains0.4% (w/w) of soluble Zn (equivalent to 17.8 mg Zn kg�1 pot-ting mix) as well as Fe, Mn, Mo, B and Mg. One barley seed withemerged radicle was planted in the pots. Deionized water wasused for watering during the experiment. Plants were grown in aglasshouse under c. 24 : 13°C day : night temperature and12 : 12 h day : night cycle. Shoots and grains were harvested atmaturity and oven-dried at 80°C for 48 h.
Mineral element analysis
Dry plant samples and grains were weighed to c. 0.3 g, digestedwith an 11 ml nitric acid/perchloric acid mixture (10 : 1 v/v),boiled until the volume was reduced to c. 1.0 ml, added to a finalvolume of 25 ml using deionized water, and then analyzed formineral elements by inductively coupled plasma optical emissionspectrometry at Waite Analytical Services, Urrbrae, South Australia(Wheal et al., 2011).
Statistical analysis
All experiments were set up as a completely randomized blockdesign with three to four replicates. The data were analyzed usingthe Genstat Statistical Program (version 11.1; VSN InternationalLtd, Hemel Hempstead, UK), and pairwise comparisons of themeans were made using the least significance difference (LSD)test at P = 0.05. To overcome the problem of nonhomogeneity ofvariances, log-transformed data were used for the ANOVA.
Results
Identification of members of the ZIP family from the barleygenome
Sequences of 16 ZIP family members of rice (O. sativa) and 15reported members of A. thaliana, along with three newly identi-fied members (AtIAR1, AtPutZnT and AtZTP29) (Table S1)were used to search for candidate members in barley as well as
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist4

Brachypodium (B. distachyon) and wheat (T. aestivum). Ninenew barley ZIP proteins (HvZIP1, HvZIP2, HvZIP6, HvZIP7,HvZIP10, HvZIP11, HvZIP13, HvZIP14 and HvZIP16) wereidentified in addition to the four HvZIP proteins (HvIRT1,HvZIP3, HvZIP5 and HvZIP8) reported by Pedas et al. (2008,2009). NcZNT1, a close homolog of AtZIP4, was also includedfor phylogenetic analyses of ZIP proteins, as it has been shown tobe responsible for the Zn/Cd hyperaccumulation inN. caerulescens (Lasat et al., 2000; Milner et al., 2012). Phylo-genetic analyses showed that HvZIP7 was closely related toOsZIP7, forming a unique clade with HvZIP10 and OsZIP10.Three AtZIP proteins (AtZIP4, AtIRT3, and AtZIP9) andNcZNT1 were grouped to the clade of HvZIP7 and OsZIP7(Fig. S1). HvZIP1 and HvZIP2 (249 amino acid residues),which were 82% identical to the wheat ZIP protein, TaZIP2,were clustered with OsZIP1, OsZIP2, AtZIP2 and AtZIP11(Fig. S1). HvZIP3 formed a distinct clade together with OsZIP3and OsZIP4. HvZIP5 and HvZIP8 were 86% similar in proteinsequence, and they were grouped with OsZIP9 and OsZIP5(Fig. S1). HvZIP5 and HvZIP8 were also closely related to Os-ZIP8 and HvZIP13 (Fig. S1). HvZIP6, HvZIP11, HvZIP14and HvZIP16 formed a distinct clade with OsZIP6, OsZIP11,OsZIP14 and OsZIP13/OsZIP16, respectively. Each of thesefour clades also contained one Arabidopsis homolog, AtZIP6,AtPutZnT, AtIAR1 and AtZTP29, respectively (Fig. S1).
Sixteen members of the ZIP gene family were identified in thefully sequenced genome of Brachypodium, a new model forgrasses (Brkljacic et al., 2011). Almost all distinct clades of Brac-hypodium and rice ZIP gene families contain at least one barleyhomolog (Fig. S1), indicating that barley homologs are repre-sented in the major clades of the ZIP gene family in monocotyle-donous species. At least seven wheat ZIP (TaZIP) proteins werefound in distinct homologous groups for HvZIP proteins(Fig. S1). The results indicate that the ZIP family proteins arerelatively conserved in monocotyledonous lineages. By compari-son, some members of the AtZIP family, such as AtZIP1, At-ZIP3, AtZIP5 and AtZIP12, branched separately from all othersequences in the tree with low bootstrap values (Fig. S1), suggest-ing that sequence divergence occurs in some members of the ZIPfamily between monocotyledonous and dicotyledonous species.HvZIP7 was selected for further studies because it belongs to aphylogenetic clade different from those (OsZIP4, OsZIP5 andOsZIP8) previously studied (Ishimaru et al., 2007; Lee et al.,2010a,b), and its homologs (NcZNT1, AtZIP4 and AtIRT3)have been shown to play an important role in Zn hyperaccumula-tion (Lasat et al., 2000; Hanikenne et al., 2008; Lin et al., 2009).
Expression of HvZIP7 in both roots and shoots is increasedby Zn deficiency
Transcript abundance of HvZIP7 was low in roots and shoots ofZn-adequate barley plants (Fig. 1a). Zn deficiency increased thetranscript abundance of HvZIP7 in both roots and shoots by atleast ninefold (Fig. 1a). To find out in which cell types HvZIP7 isspecifically expressed, in situ RT-PCR was performed in cross-sections of roots and leaves of Zn-deficient plants. There were no
HvZIP7 transcripts detected in the no-RT controls of either rootsor leaves (Fig. 1b,d,f). HvZIP7 transcripts in the roots weredetected mainly in epidermal cells and the cells within the vascu-lar bundle of roots (Fig. 1c,e), while in the cross-section of leavesHvZIP7 transcripts were found primarily in the cells of the vascu-lar bundle (Fig. 1g). These results suggest that HvZIP7 isinvolved in root uptake and translocation of Zn and Zn translo-cation in leaves.
HvZIP7 localizes to the plasma membrane
To determine subcellular localization of HvZIP7, an HvZIP7::GFP construct was generated for transient expression in onionepidermal cells. The green fluorescence of HvZIP7::GFPappeared in the location of the plasma membrane (PM; Fig. 2a,b).To distinguish PM from the vacuolar membrane, the onion epi-dermal cell, which expressed HvZIP7::GFP, was plasmolyzed forvisualization of the Hechtian strands connecting PM to the cellwall (Fig. 2c,d). A close-up view shows adhesion of the greenfluorescent strands of PM to the cell wall (Fig. 2e,f), confirmingthat the fluorescent signal derived from HvZIP7::GFP is associ-ated with PM.
HvZIP7 overexpression in plants results in higher Znaccumulation in shoots when Zn supply is moderatelyhigh in soil
A number of attempts to complement two yeast strains involvedin Zn homeostasis (zrt1zrt2 mutant defective in Zn uptake andzrc1cot1 mutant with a Zn sensitive phenotype) with HvZIP7were unsuccessful. Therefore, transgenic lines overexpressingHvZIP7 were generated in barley for gain-of-function analyses.Transgenic plants with a single locus insert of the HvZIP7
(a) (b)
(c) (d)
(e) (f)
Fig. 2 Subcellular localization of HvZIP7::GFP. An onion epidermal cellexpressing HvZIP7::GFP under UV (a) or bright light (b). An onionepidermal cell expressing HvZIP7::GFP after partial plasmolysis under UV(c) or bright light (d). A close-up view of Hechtian strands (Hs) from thesame cell shown in (c) under UV (e) or bright light (f). Arrows indicate theHechtian strands with green fluorescent protein (GFP) fluorescence. Bars,50 lm.
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 5

transgene were identified by Southern blot analysis (Fig. S2a) andselected for propagation. Progenies of three independent, singlelocus-insert lines (OX-2, OX-7 and OX-10) were genotyped byPCR to identify homozygous lines with or without transgene(null). As the processes of tissue culture, selection, and plantregeneration were employed for Agrobacterium-mediated barleytransformation (Tingay et al., 1997; Matthews et al., 2001), theseprocesses could potentially induce genetic alterations in theresulting transformants. If such changes are present in the trans-formants selected, a null line arising from segregation of thetransgene would be a more appropriate control because both ofthem have been exposed to the same tissue culture conditions,unlike the wildtype plant. Therefore, null lines were included as acontrol in all further molecular and physiological experiments,and the wildtype control was added whenever necessary. Semi-quantitative RT-PCR analysis showed that the HvZIP7 transgene
was expressed in all three independent transgenic lines, while notranscripts were detected in either the corresponding null line orthe wildtype (Fig. S2b).
The three independent homozygous transgenic lines were ini-tially examined for Zn accumulation in various growth condi-tions supplemented with low to moderately high Zn supply.There were no differences in plant growth and Zn accumulationobserved between transgenic and null lines or the wildtype whenZn supply was low to adequate. However, when the transgeniclines were grown with moderately high Zn supply, a significantdifference in plant Zn accumulation was observed between thetransgenic and null lines or the wildtype. To substantiate theseearly findings, one of the three transgenic lines, OX-10, wasselected to study plant Zn accumulation in both soil and nutrientsolution. When OX-10 was grown in soil supplemented with 0to 12.5 mg Zn kg�1 soil (covering the plant Zn nutrition range
(a) (b)
(c) (d)
(e) (f)
Fig. 3 Effects of a wide range of zinc (Zn)supplement rates in soil on growth, Znconcentration and Zn content of the plantsoverexpressing HvZIP7. (a, b) Dry weight(DW) of shoots (a) and roots (b). (c, d) Znconcentrations of shoots (c) and roots (d). (e,f) Zn content of shoots (e) and roots (f).Barley (Hordeum vulgare) seedlings of null (anull segregant of the OX-10 transgenic linewith no transgene; white bars) and OX-10transgenic (Tr; black bars) lines as well as thewildtype (WT; gray bars) were grown in acalcareous sandy soil supplemented with fourZn rates as ZnSO4. Plants were harvested28 d after seed imbibition. Mean (+SE) valuesof three replicates are presented. Asterisksindicate the least significant difference of thelog-transformed dataf at P < 0.05.
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist6
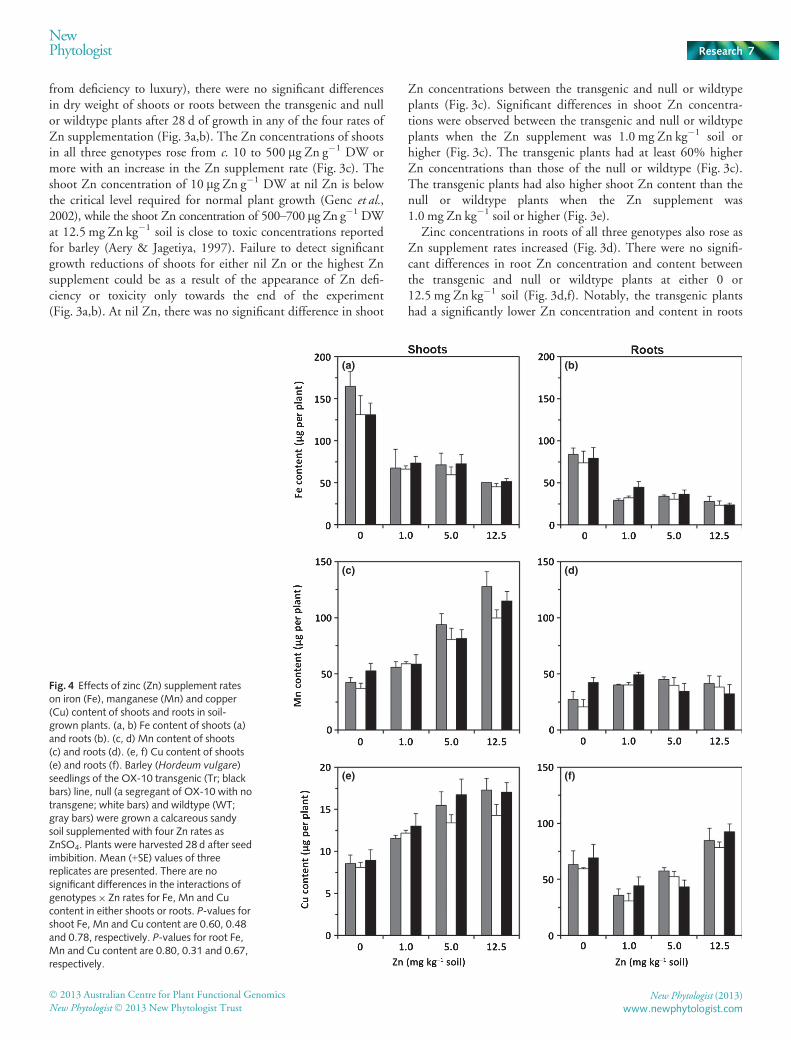
from deficiency to luxury), there were no significant differencesin dry weight of shoots or roots between the transgenic and nullor wildtype plants after 28 d of growth in any of the four rates ofZn supplementation (Fig. 3a,b). The Zn concentrations of shootsin all three genotypes rose from c. 10 to 500 lg Zn g�1 DW ormore with an increase in the Zn supplement rate (Fig. 3c). Theshoot Zn concentration of 10 lg Zn g�1 DW at nil Zn is belowthe critical level required for normal plant growth (Genc et al.,2002), while the shoot Zn concentration of 500–700 lg Zn g�1 DWat 12.5 mg Zn kg�1 soil is close to toxic concentrations reportedfor barley (Aery & Jagetiya, 1997). Failure to detect significantgrowth reductions of shoots for either nil Zn or the highest Znsupplement could be as a result of the appearance of Zn defi-ciency or toxicity only towards the end of the experiment(Fig. 3a,b). At nil Zn, there was no significant difference in shoot
Zn concentrations between the transgenic and null or wildtypeplants (Fig. 3c). Significant differences in shoot Zn concentra-tions were observed between the transgenic and null or wildtypeplants when the Zn supplement was 1.0 mg Zn kg�1 soil orhigher (Fig. 3c). The transgenic plants had at least 60% higherZn concentrations than those of the null or wildtype (Fig. 3c).The transgenic plants had also higher shoot Zn content than thenull or wildtype plants when the Zn supplement was1.0 mg Zn kg�1 soil or higher (Fig. 3e).
Zinc concentrations in roots of all three genotypes also rose asZn supplement rates increased (Fig. 3d). There were no signifi-cant differences in root Zn concentration and content betweenthe transgenic and null or wildtype plants at either 0 or12.5 mg Zn kg�1 soil (Fig. 3d,f). Notably, the transgenic plantshad a significantly lower Zn concentration and content in roots
(a) (b)
(c) (d)
(e) (f)
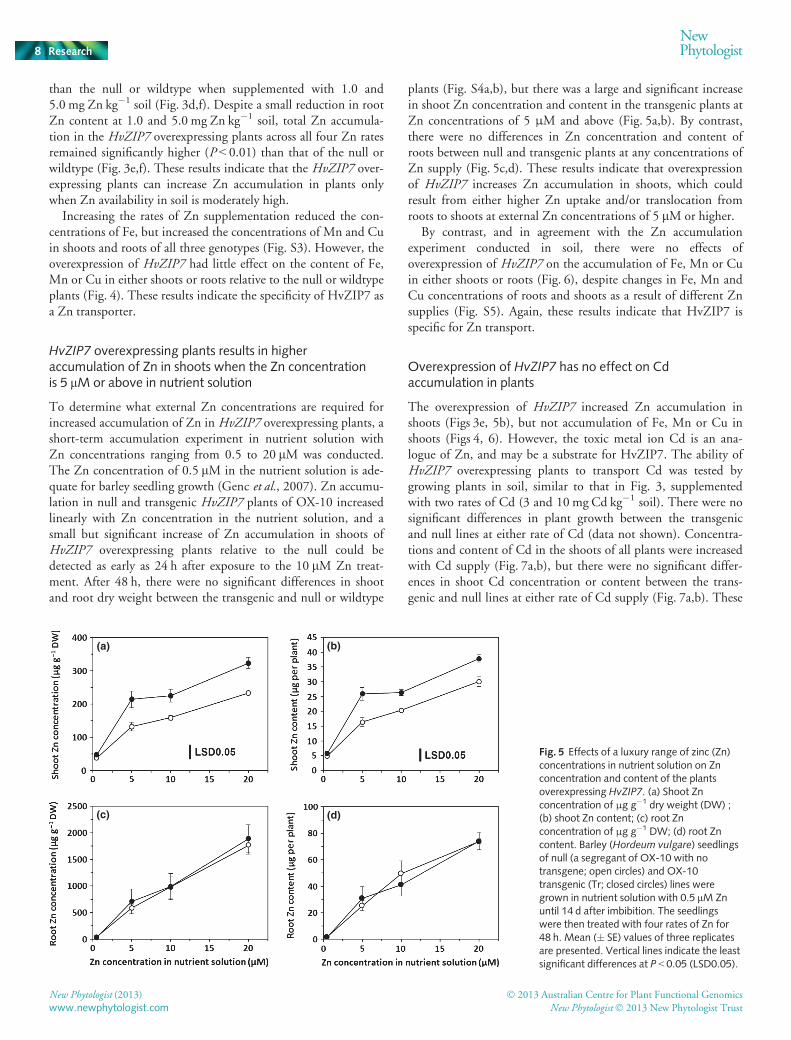
Fig. 4 Effects of zinc (Zn) supplement rateson iron (Fe), manganese (Mn) and copper(Cu) content of shoots and roots in soil-grown plants. (a, b) Fe content of shoots (a)and roots (b). (c, d) Mn content of shoots(c) and roots (d). (e, f) Cu content of shoots(e) and roots (f). Barley (Hordeum vulgare)seedlings of the OX-10 transgenic (Tr; blackbars) line, null (a segregant of OX-10 with notransgene; white bars) and wildtype (WT;gray bars) were grown a calcareous sandysoil supplemented with four Zn rates asZnSO4. Plants were harvested 28 d after seedimbibition. Mean (+SE) values of threereplicates are presented. There are nosignificant differences in the interactions ofgenotypes9 Zn rates for Fe, Mn and Cucontent in either shoots or roots. P-values forshoot Fe, Mn and Cu content are 0.60, 0.48and 0.78, respectively. P-values for root Fe,Mn and Cu content are 0.80, 0.31 and 0.67,respectively.
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 7
than the null or wildtype when supplemented with 1.0 and5.0 mg Zn kg�1 soil (Fig. 3d,f). Despite a small reduction in rootZn content at 1.0 and 5.0 mg Zn kg�1 soil, total Zn accumula-tion in the HvZIP7 overexpressing plants across all four Zn ratesremained significantly higher (P < 0.01) than that of the null orwildtype (Fig. 3e,f). These results indicate that the HvZIP7 over-expressing plants can increase Zn accumulation in plants onlywhen Zn availability in soil is moderately high.
Increasing the rates of Zn supplementation reduced the con-centrations of Fe, but increased the concentrations of Mn and Cuin shoots and roots of all three genotypes (Fig. S3). However, theoverexpression of HvZIP7 had little effect on the content of Fe,Mn or Cu in either shoots or roots relative to the null or wildtypeplants (Fig. 4). These results indicate the specificity of HvZIP7 asa Zn transporter.
HvZIP7 overexpressing plants results in higheraccumulation of Zn in shoots when the Zn concentrationis 5 lM or above in nutrient solution
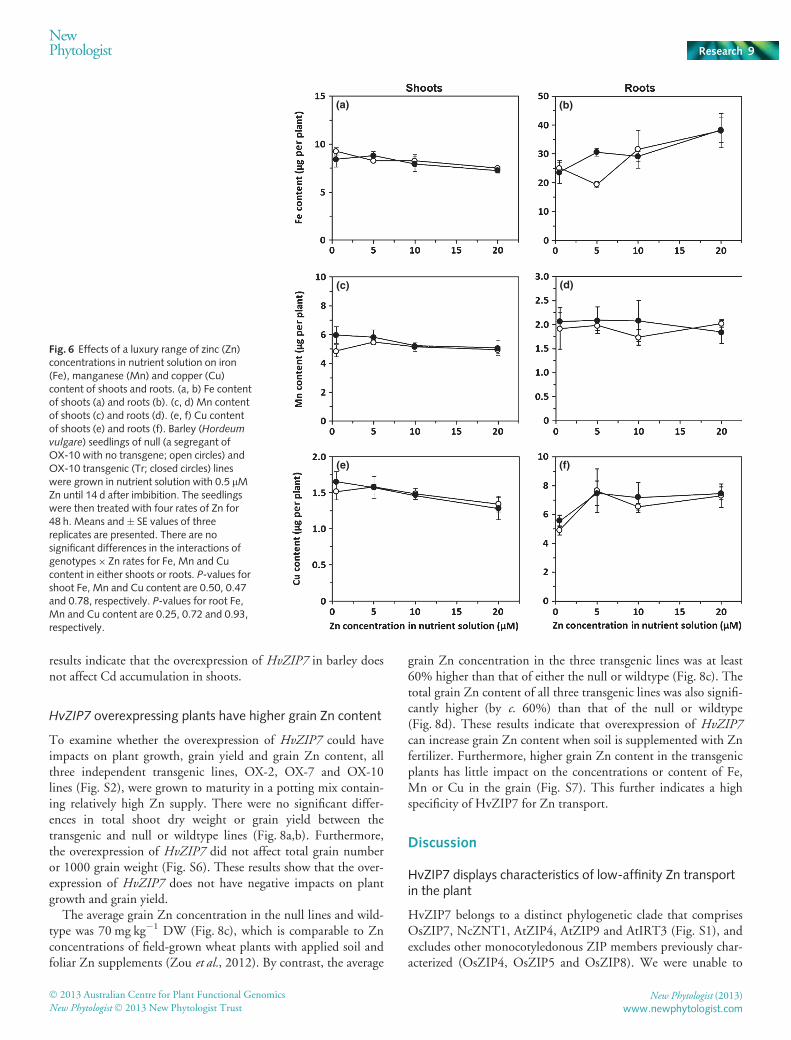
To determine what external Zn concentrations are required forincreased accumulation of Zn in HvZIP7 overexpressing plants, ashort-term accumulation experiment in nutrient solution withZn concentrations ranging from 0.5 to 20 lM was conducted.The Zn concentration of 0.5 lM in the nutrient solution is ade-quate for barley seedling growth (Genc et al., 2007). Zn accumu-lation in null and transgenic HvZIP7 plants of OX-10 increasedlinearly with Zn concentration in the nutrient solution, and asmall but significant increase of Zn accumulation in shoots ofHvZIP7 overexpressing plants relative to the null could bedetected as early as 24 h after exposure to the 10 lM Zn treat-ment. After 48 h, there were no significant differences in shootand root dry weight between the transgenic and null or wildtype
plants (Fig. S4a,b), but there was a large and significant increasein shoot Zn concentration and content in the transgenic plants atZn concentrations of 5 μM and above (Fig. 5a,b). By contrast,there were no differences in Zn concentration and content ofroots between null and transgenic plants at any concentrations ofZn supply (Fig. 5c,d). These results indicate that overexpressionof HvZIP7 increases Zn accumulation in shoots, which couldresult from either higher Zn uptake and/or translocation fromroots to shoots at external Zn concentrations of 5 lM or higher.
By contrast, and in agreement with the Zn accumulationexperiment conducted in soil, there were no effects ofoverexpression of HvZIP7 on the accumulation of Fe, Mn or Cuin either shoots or roots (Fig. 6), despite changes in Fe, Mn andCu concentrations of roots and shoots as a result of different Znsupplies (Fig. S5). Again, these results indicate that HvZIP7 isspecific for Zn transport.
Overexpression of HvZIP7 has no effect on Cdaccumulation in plants
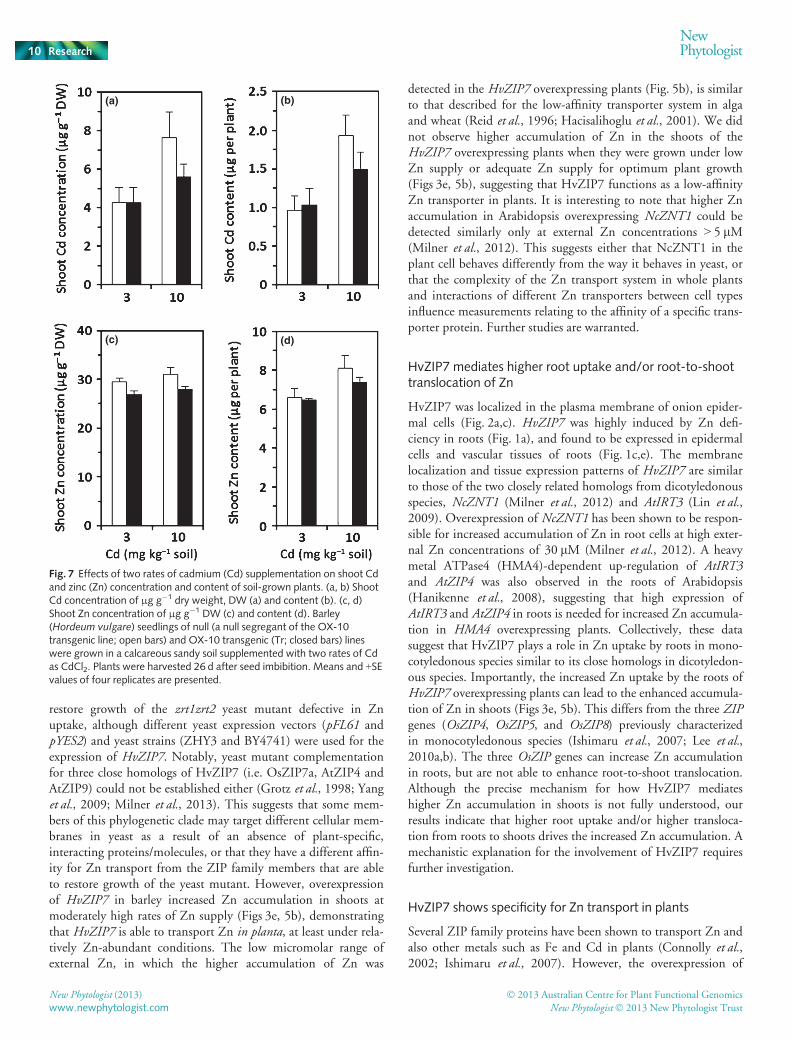
The overexpression of HvZIP7 increased Zn accumulation inshoots (Figs 3e, 5b), but not accumulation of Fe, Mn or Cu inshoots (Figs 4, 6). However, the toxic metal ion Cd is an ana-logue of Zn, and may be a substrate for HvZIP7. The ability ofHvZIP7 overexpressing plants to transport Cd was tested bygrowing plants in soil, similar to that in Fig. 3, supplementedwith two rates of Cd (3 and 10 mg Cd kg�1 soil). There were nosignificant differences in plant growth between the transgenicand null lines at either rate of Cd (data not shown). Concentra-tions and content of Cd in the shoots of all plants were increasedwith Cd supply (Fig. 7a,b), but there were no significant differ-ences in shoot Cd concentration or content between the trans-genic and null lines at either rate of Cd supply (Fig. 7a,b). These
(a) (b)
(c) (d)
Fig. 5 Effects of a luxury range of zinc (Zn)concentrations in nutrient solution on Znconcentration and content of the plantsoverexpressing HvZIP7. (a) Shoot Znconcentration of μg g�1 dry weight (DW) ;(b) shoot Zn content; (c) root Znconcentration of μg g�1 DW; (d) root Zncontent. Barley (Hordeum vulgare) seedlingsof null (a segregant of OX-10 with notransgene; open circles) and OX-10transgenic (Tr; closed circles) lines weregrown in nutrient solution with 0.5 lM Znuntil 14 d after imbibition. The seedlingswere then treated with four rates of Zn for48 h. Mean (� SE) values of three replicatesare presented. Vertical lines indicate the leastsignificant differences at P < 0.05 (LSD0.05).
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist8
results indicate that the overexpression of HvZIP7 in barley doesnot affect Cd accumulation in shoots.
HvZIP7 overexpressing plants have higher grain Zn content
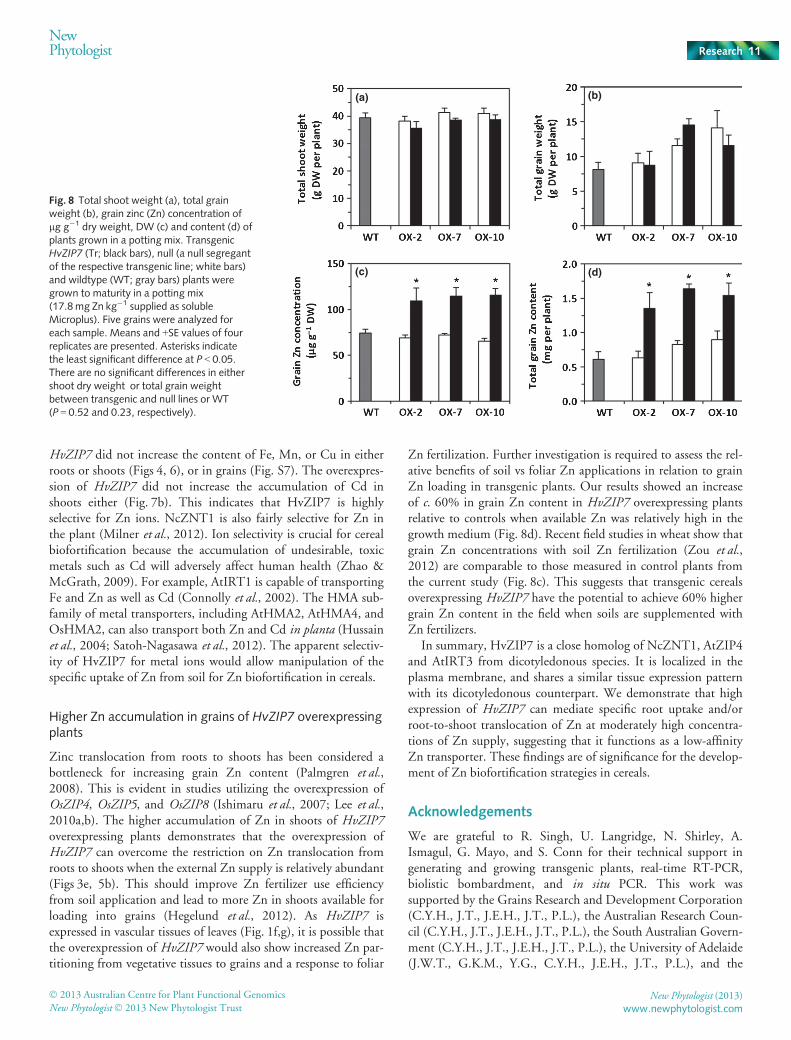
To examine whether the overexpression of HvZIP7 could haveimpacts on plant growth, grain yield and grain Zn content, allthree independent transgenic lines, OX-2, OX-7 and OX-10lines (Fig. S2), were grown to maturity in a potting mix contain-ing relatively high Zn supply. There were no significant differ-ences in total shoot dry weight or grain yield between thetransgenic and null or wildtype lines (Fig. 8a,b). Furthermore,the overexpression of HvZIP7 did not affect total grain numberor 1000 grain weight (Fig. S6). These results show that the over-expression of HvZIP7 does not have negative impacts on plantgrowth and grain yield.
The average grain Zn concentration in the null lines and wild-type was 70 mg kg�1 DW (Fig. 8c), which is comparable to Znconcentrations of field-grown wheat plants with applied soil andfoliar Zn supplements (Zou et al., 2012). By contrast, the average
grain Zn concentration in the three transgenic lines was at least60% higher than that of either the null or wildtype (Fig. 8c). Thetotal grain Zn content of all three transgenic lines was also signifi-cantly higher (by c. 60%) than that of the null or wildtype(Fig. 8d). These results indicate that overexpression of HvZIP7can increase grain Zn content when soil is supplemented with Znfertilizer. Furthermore, higher grain Zn content in the transgenicplants has little impact on the concentrations or content of Fe,Mn or Cu in the grain (Fig. S7). This further indicates a highspecificity of HvZIP7 for Zn transport.
Discussion
HvZIP7 displays characteristics of low-affinity Zn transportin the plant
HvZIP7 belongs to a distinct phylogenetic clade that comprisesOsZIP7, NcZNT1, AtZIP4, AtZIP9 and AtIRT3 (Fig. S1), andexcludes other monocotyledonous ZIP members previously char-acterized (OsZIP4, OsZIP5 and OsZIP8). We were unable to
(a) (b)
(c) (d)
(e) (f)
Fig. 6 Effects of a luxury range of zinc (Zn)concentrations in nutrient solution on iron(Fe), manganese (Mn) and copper (Cu)content of shoots and roots. (a, b) Fe contentof shoots (a) and roots (b). (c, d) Mn contentof shoots (c) and roots (d). (e, f) Cu contentof shoots (e) and roots (f). Barley (Hordeumvulgare) seedlings of null (a segregant ofOX-10 with no transgene; open circles) andOX-10 transgenic (Tr; closed circles) lineswere grown in nutrient solution with 0.5 lMZn until 14 d after imbibition. The seedlingswere then treated with four rates of Zn for48 h. Means and � SE values of threereplicates are presented. There are nosignificant differences in the interactions ofgenotypes9 Zn rates for Fe, Mn and Cucontent in either shoots or roots. P-values forshoot Fe, Mn and Cu content are 0.50, 0.47and 0.78, respectively. P-values for root Fe,Mn and Cu content are 0.25, 0.72 and 0.93,respectively.
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 9
restore growth of the zrt1zrt2 yeast mutant defective in Znuptake, although different yeast expression vectors (pFL61 andpYES2) and yeast strains (ZHY3 and BY4741) were used for theexpression of HvZIP7. Notably, yeast mutant complementationfor three close homologs of HvZIP7 (i.e. OsZIP7a, AtZIP4 andAtZIP9) could not be established either (Grotz et al., 1998; Yanget al., 2009; Milner et al., 2013). This suggests that some mem-bers of this phylogenetic clade may target different cellular mem-branes in yeast as a result of an absence of plant-specific,interacting proteins/molecules, or that they have a different affin-ity for Zn transport from the ZIP family members that are ableto restore growth of the yeast mutant. However, overexpressionof HvZIP7 in barley increased Zn accumulation in shoots atmoderately high rates of Zn supply (Figs 3e, 5b), demonstratingthat HvZIP7 is able to transport Zn in planta, at least under rela-tively Zn-abundant conditions. The low micromolar range ofexternal Zn, in which the higher accumulation of Zn was
detected in the HvZIP7 overexpressing plants (Fig. 5b), is similarto that described for the low-affinity transporter system in algaand wheat (Reid et al., 1996; Hacisalihoglu et al., 2001). We didnot observe higher accumulation of Zn in the shoots of theHvZIP7 overexpressing plants when they were grown under lowZn supply or adequate Zn supply for optimum plant growth(Figs 3e, 5b), suggesting that HvZIP7 functions as a low-affinityZn transporter in plants. It is interesting to note that higher Znaccumulation in Arabidopsis overexpressing NcZNT1 could bedetected similarly only at external Zn concentrations > 5 lM(Milner et al., 2012). This suggests either that NcZNT1 in theplant cell behaves differently from the way it behaves in yeast, orthat the complexity of the Zn transport system in whole plantsand interactions of different Zn transporters between cell typesinfluence measurements relating to the affinity of a specific trans-porter protein. Further studies are warranted.
HvZIP7 mediates higher root uptake and/or root-to-shoottranslocation of Zn
HvZIP7 was localized in the plasma membrane of onion epider-mal cells (Fig. 2a,c). HvZIP7 was highly induced by Zn defi-ciency in roots (Fig. 1a), and found to be expressed in epidermalcells and vascular tissues of roots (Fig. 1c,e). The membranelocalization and tissue expression patterns of HvZIP7 are similarto those of the two closely related homologs from dicotyledonousspecies, NcZNT1 (Milner et al., 2012) and AtIRT3 (Lin et al.,2009). Overexpression of NcZNT1 has been shown to be respon-sible for increased accumulation of Zn in root cells at high exter-nal Zn concentrations of 30 lM (Milner et al., 2012). A heavymetal ATPase4 (HMA4)-dependent up-regulation of AtIRT3and AtZIP4 was also observed in the roots of Arabidopsis(Hanikenne et al., 2008), suggesting that high expression ofAtIRT3 and AtZIP4 in roots is needed for increased Zn accumula-tion in HMA4 overexpressing plants. Collectively, these datasuggest that HvZIP7 plays a role in Zn uptake by roots in mono-cotyledonous species similar to its close homologs in dicotyledon-ous species. Importantly, the increased Zn uptake by the roots ofHvZIP7 overexpressing plants can lead to the enhanced accumula-tion of Zn in shoots (Figs 3e, 5b). This differs from the three ZIPgenes (OsZIP4, OsZIP5, and OsZIP8) previously characterizedin monocotyledonous species (Ishimaru et al., 2007; Lee et al.,2010a,b). The three OsZIP genes can increase Zn accumulationin roots, but are not able to enhance root-to-shoot translocation.Although the precise mechanism for how HvZIP7 mediateshigher Zn accumulation in shoots is not fully understood, ourresults indicate that higher root uptake and/or higher transloca-tion from roots to shoots drives the increased Zn accumulation. Amechanistic explanation for the involvement of HvZIP7 requiresfurther investigation.
HvZIP7 shows specificity for Zn transport in plants
Several ZIP family proteins have been shown to transport Zn andalso other metals such as Fe and Cd in plants (Connolly et al.,2002; Ishimaru et al., 2007). However, the overexpression of
(a) (b)
(c) (d)
Fig. 7 Effects of two rates of cadmium (Cd) supplementation on shoot Cdand zinc (Zn) concentration and content of soil-grown plants. (a, b) ShootCd concentration of μg g�1 dry weight, DW (a) and content (b). (c, d)Shoot Zn concentration of μg g�1 DW (c) and content (d). Barley(Hordeum vulgare) seedlings of null (a null segregant of the OX-10transgenic line; open bars) and OX-10 transgenic (Tr; closed bars) lineswere grown in a calcareous sandy soil supplemented with two rates of Cdas CdCl2. Plants were harvested 26 d after seed imbibition. Means and +SEvalues of four replicates are presented.
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist10
HvZIP7 did not increase the content of Fe, Mn, or Cu in eitherroots or shoots (Figs 4, 6), or in grains (Fig. S7). The overexpres-sion of HvZIP7 did not increase the accumulation of Cd inshoots either (Fig. 7b). This indicates that HvZIP7 is highlyselective for Zn ions. NcZNT1 is also fairly selective for Zn inthe plant (Milner et al., 2012). Ion selectivity is crucial for cerealbiofortification because the accumulation of undesirable, toxicmetals such as Cd will adversely affect human health (Zhao &McGrath, 2009). For example, AtIRT1 is capable of transportingFe and Zn as well as Cd (Connolly et al., 2002). The HMA sub-family of metal transporters, including AtHMA2, AtHMA4, andOsHMA2, can also transport both Zn and Cd in planta (Hussainet al., 2004; Satoh-Nagasawa et al., 2012). The apparent selectiv-ity of HvZIP7 for metal ions would allow manipulation of thespecific uptake of Zn from soil for Zn biofortification in cereals.
Higher Zn accumulation in grains of HvZIP7 overexpressingplants
Zinc translocation from roots to shoots has been considered abottleneck for increasing grain Zn content (Palmgren et al.,2008). This is evident in studies utilizing the overexpression ofOsZIP4, OsZIP5, and OsZIP8 (Ishimaru et al., 2007; Lee et al.,2010a,b). The higher accumulation of Zn in shoots of HvZIP7overexpressing plants demonstrates that the overexpression ofHvZIP7 can overcome the restriction on Zn translocation fromroots to shoots when the external Zn supply is relatively abundant(Figs 3e, 5b). This should improve Zn fertilizer use efficiencyfrom soil application and lead to more Zn in shoots available forloading into grains (Hegelund et al., 2012). As HvZIP7 isexpressed in vascular tissues of leaves (Fig. 1f,g), it is possible thatthe overexpression of HvZIP7 would also show increased Zn par-titioning from vegetative tissues to grains and a response to foliar
Zn fertilization. Further investigation is required to assess the rel-ative benefits of soil vs foliar Zn applications in relation to grainZn loading in transgenic plants. Our results showed an increaseof c. 60% in grain Zn content in HvZIP7 overexpressing plantsrelative to controls when available Zn was relatively high in thegrowth medium (Fig. 8d). Recent field studies in wheat show thatgrain Zn concentrations with soil Zn fertilization (Zou et al.,2012) are comparable to those measured in control plants fromthe current study (Fig. 8c). This suggests that transgenic cerealsoverexpressing HvZIP7 have the potential to achieve 60% highergrain Zn content in the field when soils are supplemented withZn fertilizers.
In summary, HvZIP7 is a close homolog of NcZNT1, AtZIP4and AtIRT3 from dicotyledonous species. It is localized in theplasma membrane, and shares a similar tissue expression patternwith its dicotyledonous counterpart. We demonstrate that highexpression of HvZIP7 can mediate specific root uptake and/orroot-to-shoot translocation of Zn at moderately high concentra-tions of Zn supply, suggesting that it functions as a low-affinityZn transporter. These findings are of significance for the develop-ment of Zn biofortification strategies in cereals.
Acknowledgements
We are grateful to R. Singh, U. Langridge, N. Shirley, A.Ismagul, G. Mayo, and S. Conn for their technical support ingenerating and growing transgenic plants, real-time RT-PCR,biolistic bombardment, and in situ PCR. This work wassupported by the Grains Research and Development Corporation(C.Y.H., J.T., J.E.H., J.T., P.L.), the Australian Research Coun-cil (C.Y.H., J.T., J.E.H., J.T., P.L.), the South Australian Govern-ment (C.Y.H., J.T., J.E.H., J.T., P.L.), the University of Adelaide(J.W.T., G.K.M., Y.G., C.Y.H., J.E.H., J.T., P.L.), and the
(a) (b)
(c) (d)
Fig. 8 Total shoot weight (a), total grainweight (b), grain zinc (Zn) concentration oflg g�1 dry weight, DW (c) and content (d) ofplants grown in a potting mix. TransgenicHvZIP7 (Tr; black bars), null (a null segregantof the respective transgenic line; white bars)and wildtype (WT; gray bars) plants weregrown to maturity in a potting mix(17.8mg Zn kg�1 supplied as solubleMicroplus). Five grains were analyzed foreach sample. Means and +SE values of fourreplicates are presented. Asterisks indicatethe least significant difference at P < 0.05.There are no significant differences in eithershoot dry weight or total grain weightbetween transgenic and null lines or WT(P = 0.52 and 0.23, respectively).
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 11

Danish Strategic Research Council (PP, NUTRIEFFICIENT;grant no. 10-093498).
References
Aery NC, Jagetiya BL. 1997. Relative toxicity of cadmium, lead, and zinc on
barley. Communications in Soil Science and Plant Analysis 28: 949–960.Alloway BJ. 2004. Zinc in soils and crop nutrition. Brussels, Belgium: IZA
Publications.
Altschul SF, Madden TL, Schaffer AA, Zhang JH, Zhang Z, Miller W, Lipman
DJ. 1997. Gapped BLAST and PSI-BLAST: a new generation of protein
database search programs. Nucleic Acids Research 25: 3389–3402.Altschul SF, Wootton JC, Gertz EM, Agarwala R, Morgulis A, Schaffer AA, Yu
YK. 2005. Protein database searches using compositionally adjusted
substitution matrices. FEBS Journal 272: 5101–5109.Brkljacic J, Grotewold E, Scholl R, Mockler T, Garvin D, Vain P, Brutnell T,
Sibout R, Bevan M, Budak H et al. 2011. Brachypodium as a model for the
grasses: today and the future. Plant Physiology 157: 3–13.Cakmak I. 2008. Enrichment of cereal grains with zinc: agronomic or genetic
biofortification? Plant and Soil 302: 1–17.Chen W, Feng Y, Chao Y. 2008. Genomic analysis and expression pattern of
OsZIP1, OsZIP3, and OsZIP4 in two rice (Oryza sativa L.) genotypes withdifferent zinc efficiency. Russian Journal of Plant Physiology 55: 400–409.
Connolly EL, Fett JP, Guerinot ML. 2002. Expression of the IRT1 metal
transporter is controlled by metals at the levels of transcript and protein
accumulation. Plant Cell 14: 1347–1357.Cunningham-Rundles S, McNeeley DF, Moon A. 2005.Mechanisms of
nutrient modulation of the immune response. Journal of Allergy and ClinicalImmunology 115: 1119–1128.
Curtis MD, Grossniklaus U. 2003. A gateway cloning vector set for high-
throughput functional analysis of genes in planta. Plant Physiology 133: 462–469.Durmaz E, Coruh C, Dinler G, Grusak M, Peleg Z, Saranga Y, Fahima T,
Yazici A, Ozturk L, Cakmak I et al. 2010. Expression and cellular localization
of ZIP1 transporter under zinc deficiency in wild emmer wheat. PlantMolecular Biology Reporter 29: 1–15.
Eide DJ. 2006. Zinc transporters and the cellular trafficking of zinc. Biochimica etBiophysica Acta-Molecular Cell Research 1763: 711–722.
Genc Y, Huang CY, Langridge P. 2007. A study of the role of root
morphological traits in growth of barley in zinc-deficient soil. Journal ofExperimental Botany 58: 2775–2784.
Genc Y, McDonald GK, Graham RD. 2002. Critical deficiency concentration of
zinc in barley genotypes differing in zinc efficiency and its relation to growth
responses. Journal of Plant Nutrition 25: 545–560.Graham RD, Welch RM. 2001.Micronutrient interactions in humans: setting goalsfor plant breeders and agronomists. Dordrecht, the Netherlands: Kluwer
Academic Publishers.
Grotz N, Fox T, Connolly E, Park W, Guerinot ML, Eide D. 1998.
Identification of a family of zinc transporter genes from Arabidopsis that
respond to zinc deficiency. Proceedings of the National Academy of Sciences, USA95: 7220–7224.
Grotz N, Guerinot ML. 2006. Molecular aspects of Cu, Fe and Zn
homeostasis in plants. Biochimica et Biophysica Acta–Molecular Cell Research1763: 595–608.
Guerinot ML. 2000. The ZIP family of metal transporters. Biochimica etBiophysica Acta 1465: 190–198.
Hacisalihoglu G, Hart JJ, Kochian LV. 2001.High- and low-affinity zinc
transport systems and their possible role in zinc efficiency in bread wheat. PlantPhysiology 125: 456–463.
Hanikenne M, Talke IN, Haydon MJ, Lanz C, Nolte A, Motte P, Kroymann J,
Weigel D, Kraemer U. 2008. Evolution of metal hyperaccumulation
required cis-regulatory changes and triplication of HMA4. Nature 453:391–395.
Hegelund JN, Pedas P, Husted S, Schiller M, Schjoerring JK. 2012. Zinc fluxes
into developing barley grains: use of stable Zn isotopes to separate root uptake
from remobilization in plants with contrasting Zn status. Plant and Soil 361:241–250.
Hotz C, DeHaene J, Woodhouse LR, Villalpando S, Rivera JA, King JC. 2005.
Zinc absorption from zinc oxide, zinc sulfate, zinc oxide plus EDTA, or
sodium-zinc EDTA does not differ when added as fortificants to maize tortillas.
Journal of Nutrition 135: 1102–1105.Huang CY, Shirley N, Genc Y, Shi BJ, Langridge P. 2011. Phosphate utilization
efficiency correlates with expression of low-affinity phosphate transporters and
noncoding RNA, IPS1, in barley. Plant Physiology 156: 1217–1229.Hussain D, Haydon MJ, Wang Y, Wong E, Sherson SM, Young J, Camakaris J,
Harper JF, Cobbett CS. 2004. P-type ATPase heavy metal transporters with
roles in essential zinc homeostasis in Arabidopsis. Plant Cell 16: 1327–1339.Ishimaru Y, Masuda H, Suzuki M, Bashir K, Takahashi M, Nakanishi H, Mori
S, Nishizawa NK. 2007.Overexpression of the OsZIP4 zinc transporter
confers disarrangement of zinc distribution in rice plants. Journal ofExperimental Botany 58: 2909–2915.
Ishimaru Y, Suzuki M, Kobayashi T, Takahashi M, Nakanishi H, Mori S,
Nishizawa NK. 2005.OsZIP4, a novel zinc-regulated zinc transporter in rice.
Journal of Experimental Botany 56: 3207–3214.Katoh K, Asimenos G, Toh H. 2009.Multiple alignment of DNA sequences
with MAFFT. In: Posada D, ed. Bioinformatics for DNA sequence analysis.Totowa, NJ, USA: Humana Press Inc, 39–64.
Koltai H, Bird DM. 2000.High throughput cellular localization of specific plant
mRNAs by liquid-phase in situ reverse transcription-polymerase chain reaction
of tissue sections. Plant Physiology 123: 1203–1212.Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA,
McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R et al. 2007. ClustalW and clustal X version 2.0. Bioinformatics 23: 2947–2948.
Lasat MM, Pence NS, Garvin DF, Ebbs SD, Kochian LV. 2000.Molecular
physiology of zinc transport in the Zn hyperaccumulator Thlaspi caerulescens.Journal of Experimental Botany 51: 71–79.
Lee S, Jeong HJ, Kim SA, Lee J, Guerinot ML, An G. 2010a.OsZIP5 is a
plasma membrane zinc transporter in rice. Plant Molecular Biology 73: 507–517.Lee S, Kim SA, Lee J, Guerinot ML, An G. 2010b. Zinc deficiency-inducible
OsZIP8 encodes a plasma membrane-localized zinc transporter in rice.
Molecules and Cells 29: 551–558.Lin Y-F, Liang H-M, Yang S-Y, Boch A, Clemens S, Chen C-C, Wu J-F, Huang
J-L, Yeh K-C. 2009. Arabidopsis IRT3 is a zinc-regulated and plasma
membrane localized zinc/iron transporter. New Phytologist 182: 392–404.Maret W. 2004. Zinc and sulfur: a critical biological partnership. Biochemistry43: 3301–3309.
Matthews PR, Wang MB, Waterhouse PM, Thornton S, Fieg SJ, Gubler F,
Jacobsen JV. 2001.Marker gene elimination from transgenic barley, using
co-transformation with adjacent ‘twin T-DNAs’ on a standard Agrobacterium
transformation vector.Molecular Breeding 7: 195–202.Milner MJ, Craft E, Yamaji N, Koyama E, Ma JF, Kochian LV. 2012.
Characterization of the high affinity Zn transporter from Noccaea caerulescens,NcZNT1, and dissection of its promoter for its role in Zn uptake and
hyperaccumulation. New Phytologist 195: 113–123.Milner MJ, Seamon J, Craft E, Kochian LV. 2013. Transport properties of
members of the ZIP family in plants and their role in Zn and Mn homeostasis.
Journal of Experimental Botany 64: 369–381.Narayanan NN, Vasconcelos MW, Grusak MA. 2007. Expression profiling of
Oryza sativa metal homeostasis genes in different rice cultivars using a cDNA
macroarray. Plant Physiology and Biochemistry 45: 277–286.Palmgren MG, Clemens S, Williams LE, Kraemer U, Borg S, Schjorring JK,
Sanders D. 2008. Zinc biofortification of cereals: problems and solutions.
Trends in Plant Science 13: 464–473.Pedas P, Husted S. 2009. Zinc transport mediated by barley ZIP proteins are
induced by low pH. Plant Signaling & Behavior 4: 842–845.Pedas P, Schjoerring JK, Husted S. 2009. Identification and characterization of
zinc-starvation-induced ZIP transporters from barley roots. Plant Physiologyand Biochemistry 47: 377–383.
Pedas P, Ytting CK, Fuglsang AT, Jahn TP, Schjoerring JK, Husted S. 2008.
Manganese efficiency in barley: identification and characterization of the metal
ion transporter HvIRT1. Plant Physiology 148: 455–466.Preuss CP, Huang CY, Gilliham M, Tyerman SD. 2010. Channel-like
characteristics of the low-affinity barley phosphate transporter PHT1;6 when
expressed in Xenopus oocytes. Plant Physiology 152: 1431–1441.
New Phytologist (2013) � 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist Trustwww.newphytologist.com
Research
NewPhytologist12

Ramesh SA, Shin R, Eide DJ, Schachtman DP. 2003. Differential metal
selectivity and gene expression of two zinc transporters from rice. PlantPhysiology 133: 126–134.
Reid RJ, Brookes JD, Tester MA, Smith FA. 1996. The mechanism of zinc uptake
in plants – characterisation of the low-affinity system. Planta 198: 39–45.Satoh-Nagasawa N, Mori M, Nakazawa N, Kawamoto T, Nagato Y, Sakurai K,
Takahashi H, Watanabe A, Akagi H. 2012.Mutations in rice (Oryza sativa)heavy metal ATPase 2 (OsHMA2) restrict the translocation of zinc and
cadmium. Plant and Cell Physiology 53: 213–224.Stephens BW, Cook DR, Grusak MA. 2011. Characterization of zinc transport
by divalent metal transporters of the ZIP family from the model legume
Medicago truncatula. BioMetals 24: 51–58.Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011.
MEGA5: molecular evolutionary genetics analysis using maximum likelihood,
evolutionary distance, and maximum parsimony methods.Molecular Biologyand Evolution 28: 2731–2739.
Tingay S, McElroy D, Kalla R, Fieg S, Wang MB, Thornton S, Brettell R.
1997. Agrobacterium tumefaciens-mediated barley transformation. Plant Journal11: 1369–1376.
Waters BM, Sankaran RP. 2011.Moving micronutrients from the soil to the
seeds: genes and physiological processes from a biofortification perspective.
Plant Science 180: 562–574.Welch RM. 1995.Micronutrient nutrition of plants. Critical Reviews in PlantSciences 14: 49–82.
Welch RM, Graham RD. 2004. Breeding for micronutrients in staple food
crops from a human nutrition perspective. Journal of Experimental Botany 55:353–364.
Wheal MS, Fowles TO, Palmer LT. 2011. A cost-effective acid digestion method
using closed polypropylene tubes for inductively coupled plasma optical
emission spectrometry (ICP-OES) analysis of plant essential elements.
Analytical Methods 3: 2854–2863.Yang X, Huang J, Jiang Y, Zhang H-S. 2009. Cloning and functional
identification of two members of the ZIP (Zrt, Irt-like protein) gene family in
rice (Oryza sativa L.).Molecular Biology Reports 36: 281–287.Zhao FJ, McGrath SP. 2009. Biofortification and phytoremediation. CurrentOpinion in Plant Biology 12: 373–380.
Zou CQ, Zhang YQ, Rashid A, Ram H, Savasli E, Arisoy RZ, Ortiz-Monasterio
JI, Simunji S, Wang ZH, Sohu V et al. 2012. Biofortification of wheat
with zinc through zinc fertilization in seven countries. Plant and Soil 361:119–130.
Supporting Information
Additional supporting information may be found in the onlineversion of this article.
Fig. S1 Phylogenetic tree of ZIP proteins in Arabidopsis, rice,Brachypodium, barley and wheat.
Fig. S2 Southern blot analysis and semiquantitative RT-PCRanalysis of transcript abundances of the HvZIP7 transgene.
Fig. S3 Effects of Zn supplement rates on Fe, Mn and Cu con-centrations of shoots and roots in soil-grown plants.
Fig. S4 Effects of Zn concentrations in nutrient solution on dryweight of the plants overexpressing HvZIP7.
Fig. S5 Effects of a luxury range of Zn concentrations in nutrientsolution on Fe, Mn and Cu concentrations of shoots and roots.
Fig. S6 Total grain number and 1000 grain weight of plantsgrown in a potting mix.
Fig. S7 Fe, Mn and Cu concentration and content of barleygrains.
Table S1 Identifiers and annotations for the Arabidopsis and ricegenes described in this study
Please note: Wiley-Blackwell is not responsible for the content orfunctionality of any supporting information supplied by theauthors. Any queries (other than missing material) should bedirected to the New Phytologist Central Office.
� 2013 Australian Centre for Plant Functional Genomics
New Phytologist� 2013 New Phytologist TrustNew Phytologist (2013)
www.newphytologist.com
NewPhytologist Research 13




















