
Neuropeptide Release from Dental Pulp Cells by RgpB via Proteinase-Activated Receptor-2 Signaling

BIOTECHNOLOGICALLY RELEVANT ENZYMES AND PROTEINS
Heterologous expression and characterizationof a glucose-stimulated β-glucosidase from the termiteNeotermes koshunensis in Aspergillus oryzae
Cristiane Akemi Uchima & Gaku Tokuda &
Hirofumi Watanabe & Katsuhiko Kitamoto &
Manabu Arioka
Received: 10 August 2010 /Revised: 11 October 2010 /Accepted: 12 October 2010 /Published online: 6 November 2010# Springer-Verlag 2010
Abstract Neotermes koshunensis is a lower termite thatsecretes endogenous β-glucosidase in the salivary glands.This β-glucosidase (G1NkBG) was successfully expressedin Aspergillus oryzae. G1NkBG was purified to homoge-neity from the culture supernatant through ammoniumsulfate precipitation and anion exchange, hydrophobic,and gel filtration chromatographies with a 48-fold increasein purity. The molecular mass of the native enzymeappeared as a single band at 60 kDa after gel filtrationanalysis, indicating that G1NkBG is a monomeric protein.Maximum activity was observed at 50 °C with an optimumpH at 5.0. G1NkBG retained 80% of its maximum activity attemperatures up to 45 °C and lost its activity at temperaturesabove 55 °C. The enzyme was stable from pH 5.0 to 9.0.G1NkBG was most active towards laminaribiose and p-nitrophenyl-β-D-fucopyranoside. Cellobiose, as well ascello-oligosaccharides, was also well hydrolyzed. Theenzyme activity was slightly stimulated byMn2+ and glycerol.
The Km and Vmax values were 0.77 mM and 16 U/mg,respectively, against p-nitrophenyl-β-D-glucopyranoside. Anunusual finding was that G1NkBG was stimulated by 1.3-fold when glucose was present in the reaction mixture at aconcentration of 200 mM. These characteristics, particularlythe stimulation of enzyme activity by glucose, makeG1NkBG of great interest for biotechnological applications,especially for bioethanol production.
Keywords Glucose-stimulated β-glucosidase .Neotermeskoshunensis . Heterologous expression . Aspergillus oryzae
Introduction
One of the biggest concerns of the twenty-first century isclimate change resulting from an increasing emission ofgreenhouse gasses such as CO2, N2O, and methane into theatmosphere. It is estimated that 85% of energy consumptioncomes from burning petroleum-based fuels, which contributesto a major fraction of CO2 emissions (Carroll and Somerville2009). This fact, associated with the depletion of fossil fuels,makes the utilization of bioethanol as an attractive option forcombating both global warming and consumption of crudeoil (Dashtban et al. 2009). A further increase in bioethanolproduction may come from lignocellulosic biomass, such asagricultural residues and wood. Lignocellulose in wasteproducts is abundant, inexpensive, and renewable (Balatet al. 2008).
Plant biomass is a mixture of carbohydrate polymers,like cellulose, hemicellulose, pectin, and the non-carbohydrate polymer lignin. Cellulose, which is a homo-polysaccharide composed of β-D-glucopyranose unitslinked by β-1,4 glycosidic bonds, comprises around 40–
Electronic supplementary material The online version of this article(doi:10.1007/s00253-010-2963-y) contains supplementary material,which is available to authorized users.
C. A. Uchima :K. Kitamoto :M. Arioka (*)Department of Biotechnology, University of Tokyo,1-1-1 Yayoi, Bunkyo-ku,Tokyo 113-8657, Japane-mail: [email protected]
G. TokudaCenter of Molecular Biosciences, University of the Ryukyus,Nishihara,Okinawa 903-0213, Japan
H. WatanabeNational Institute of Agrobiological Science,Owashi,Tsukuba, Ibaraki 305-8634, Japan
Appl Microbiol Biotechnol (2011) 89:1761–1771DOI 10.1007/s00253-010-2963-y

50 wt.% of dry wood and is a potential feedstock forbiomass conversion. However, for bioethanol production,cellulose must be converted into the fermentable sugar,glucose, in a complex process, which requires synergisticaction of three different enzymes known as endoglucanase(EC. 3.2.1.4), cellobiohydrolase (EC. 3.2.1.91), and β-glucosidase (EC. 3.2.1.21) (Béguin and Aubert 1994).While endoglucanase attacks randomly amorphous cellu-lose chain at internal bonds, cellobiohydrolase acts on thereducing and non-reducing ends releasing cellobiose fromcellulose fibers. Although the formation of cellobioseinhibits the action of these enzymes, β-glucosidase reducessuch inhibition catalyzing hydrolysis of cellobiose or cello-oligomers to glucose, thus playing a critical role incellulose degradation (Bhatia et al. 2002; Lynd et al.2002). Another important point that must be taken intoconsideration is that most β-glucosidases are also stronglyinhibited by the end-product glucose, which may restrictthe whole degradation process of cellulose (Bhatia et al.2002; Bothast and Saha 1997; Karnchanatat et al. 2007;Kaur et al. 2007; Lynd et al. 2002; Yang et al. 2008; Yoonet al. 2008). On the other hand, some studies have reportedthat fungal β-glucosidases are tolerant to (Günata andVallier 1999; Riou et al. 1998; Saha and Bothast 1996; Yanand Lin 1997) or even stimulated by (Nascimento et al.2010; Sonia et al. 2008; Zanoelo et al. 2004) glucose. Also,bacterial β-glucosidases were found to be tolerant (Pérez-Pons et al. 1995; Wright et al. 1992). Having β-glucosidasewith one of these characteristics in an industrial process,such as in commercial preparations, can increase thecellulose hydrolysis ability, which is very interesting interms of biotechnological purposes (Lynd et al. 2002).
Considering that β-glucosidase is essential for cellulosedegradation, several studies have been done on β-glucosidase-producing fungi and bacteria for biomassutilization (Kotaka et al. 2008; Nascimento et al. 2010;Saha et al. 1994; Saha and Bothast 1996). In contrast, thereare only a few studies about β-glucosidase produced byanimal species (Ni et al. 2007; Scharf et al. 2010) that focuson bioethanol production. It is well-known that termites areefficient decomposers of cellulosic materials: 74–99% ofthe cellulose ingested is hydrolyzed (Prins and Kreulen1991) due to the existence of symbiotic protists living intheir hindguts or to their own cellulolytic system to degradethose materials (Watanabe and Tokuda 2001). Termites arethus called as “microscale bioconversion systems” (Watanabeand Tokuda 2010). Despite this high efficiency, the produc-tion level is too low to be used for commercial exploitation.For further production and characterization of termite-producing cellulases, molecular techniques have beenemployed to increase the enzyme production. In this work,we have used the expression system of Aspergillus oryzae, afilamentous fungus that can produce and secrete large
amounts of proteins, to produce endogenous β-glucosidasefound in the salivary glands of the lower termite Neotermeskoshunensis. This enzyme, termed hereafter G1NkBG,belongs to the glycoside hydrolase family 1 (refer to CAZy,http://www.cazy.org/). A. oryzae has a Generally Recognizedas Safe (GRAS) status, since it is widely used in Japanesetraditional food fermentation (Kitamoto 2002). Thus, thisfungus is considered as a suitable host for the production ofheterologous proteins.
Although there is a study regarding the heterologousexpression of β-glucosidase produced by N. koshunensis(Ni et al. 2007), this work was focused only in the basiccharacterization of the enzyme. Besides purification and abetter characterization of the enzyme, in this work, we alsoshow the most interesting characteristic that was notanalyzed before: the discovery of an unusual stimulationof β-glucosidase from animal origin by glucose.
Materials and methods
Microorganisms
N. koshunensis cDNA was used as a donor of the targetgene (GenBank accession no. AB073638). For G1NkBGproduction, a double-protease disruptant, A. oryzae strainNS-tApE (niaD- sC- ΔtppA ΔpepE) (Nemoto et al. 2009),was used as a host and in all other DNA manipulationsEscherichia coli DH5α was used.
Plasmid construction and transformation of A. oryzae
The expression plasmid was constructed by using MultiSiteGateway system (Invitrogen, Carlsbad, CA, USA) accordingto our previous study (Mabashi et al. 2006). g1nkbg wasexpressed with glucoamylase as a carrier partner, under glaApromoter, and with A. nidulans sC as a selectable marker.The sC gene encodes ATP sulfurylase, which catalyzes theactivation of inorganic sulfate by ATP and is essential forsulfate assimilation. The putative mature region (fromnucleotide 103 to 1,539 and from amino acid 35 to 513) ofG1NkBG was preceded by Kex2 cleavage site plus threeconsecutive glycine residues. Transformation of A. oryzaewas done according to the transformation method previouslydescribed (Kitamoto 2002).
Media and culture conditions
A. oryzae transformants were grown on minimal (M)medium (0.2% NH4Cl, 0.1% (NH4)2SO4, 0.05% NaCl,0.1% KH2PO4, 0.05% MgSO4·7H2O, 0.002% of 2%FeSO4·7H2O, and 2% glucose, pH 5.5) with 1.2 M sorbitol.Colonies were grown at 30 °C for 3–4 days and purified
1762 Appl Microbiol Biotechnol (2011) 89:1761–1771

three times on M medium plates. For β-glucosidaseproduction, 500-ml flasks containing 100 ml of 5 × DPYliquid medium (10% dextrin, 5% polypeptone, 2.5% yeastextract, 0.5% K2HPO4, and 0.05% MgSO4·7H2O, pH 8.0)were inoculated with approximately 1×106 conidia. Thecultures were incubated at 30 °C for 5 days on shaker(approximately 150 rpm). After the 5th day, the cultureswere harvested by filtration with Miracloth (Calbiochem,CA, USA).
Enzyme assay and protein determination
As a standard assay procedure, a standard curve wasconstructed using p-nitrophenol in five different concen-trations (62.5, 125, 250, 500, and 1,000 μM), and theabsorbance was measured at A410. Then, the β-glucosidaseactivity was measured at 45 °C with 10 mM p-nitrophenyl-β-D-glucopyranoside (pNPG; Sigma, St. Louis, MO, USA)as a substrate in 50 mM sodium acetate buffer (pH 5.5) andappropriately diluted enzyme preparation. After 30 min ofincubation, the reaction was stopped by adding 1 ml of0.6 M Na2CO3, and the release of p-nitrophenol (pNP)was measured at A410. The results were calculated byusing the equation obtained in the standard curve. Forsubstrate specificity analyses, 10 mM of p-nitrophenyl-β-D-fucopyranoside (pNPFuc), p-nitrophenyl-β-D-lactopyranoside(pNPLac), and p-nitrophenyl-β-D-galactopyranoside (pNPGal)was assayed in the same condition. The β-glucosidaseactivities were also determined by the release of glucosefrom natural substrates using glucose oxidase–mutarotasereagent (Glucose CII Test Wako; Wako Pure ChemicalCo., Tokyo, Japan). A standard curve was constructedusing five different concentrations of glucose (0.25, 0.5,1.0, 1.25, and 1.5 mg/ml). The substrates used were: 2%(w/v) laminaribiose, 2% laminarin, 2% gentibiose, 2%sophorose, 2% salicin, 2% lactose, 1% carboxymethylcellulose (CMC), 30 mM cellobiose, 30 mM cellotriose,30 mM cellotetraose, 15 mM cellopentaose, and 15 mMcelloxaose. Each substrate was dissolved in 50 mMsodium acetate buffer (pH 5.5) and incubated with theenzyme at appropriate dilutions. After 5 min of incubationat 37 °C, 1 ml of the glucose oxidase–mutarotase reagentwas added and the release of glucose was measured atA505. In all analyses, one unit (U) was defined as theamount of enzyme that releases 1 μmol of product perminute under those conditions.
Protein concentration was measured by absorbance at595 nm (A595) by using the Bio-Rad protein assay (Bio-Rad), according to the Bradford method (1976), withbovine serum albumin (Takara, Japan) as the standard.Specific activity was expressed as units per milligram ofprotein (U/mg). In chromatographic profiles, proteins in thecolumn effluents were monitored by measuring A280.
Purification of G1NkBG
Ammonium sulfate precipitation
Using a saturated solution of (NH4)2SO4, most contaminantproteins were removed at 45% saturation. The β-glucosidase activity in the supernatant was recovered at65% saturation. In the last step, after stirring for 1 h at 4 °C,the resulting precipitate was collected by centrifugation at9,000×g for 30 min and concentrated by dissolving inproper volume of 50 mM Tris–HCl (pH 7.5). This samplewas dialyzed overnight against the same buffer.
Ion exchange chromatography
The dialyzed enzyme solution was applied on a column(1.6×2.5 cm) of HiTrap DEAE-Sepharose anion exchangeequilibrated with 175 mM NaCl in 50 mM Tris–HClbuffer, pH 7.5. Proteins were eluted with a gradient of175 to 500 mM NaCl in the same buffer at a flow rateof 0.3 ml/min. Each fraction contained 0.5 ml and theywere used for enzyme activity assay and sodiumdodecylsulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analyses. Active fractions (from 21 to 31) werepooled and then concentrated from 5.5 ml to 550 μl, andthe buffer was also exchanged using an ultrafiltration cell(10-kDa cutoff, Sartorius Stedim, Germany).
Hydrophobic chromatography
The concentrated sample after ultrafiltration was loaded onHiTrap Phenyl FF hydrophobic column (0.7×2.5 cm)equilibrated with 1 M (NH4)2SO4 in 50 mM Tris–HClbuffer, pH 7.5. The proteins were then eluted by lineargradient from 1 to 0 M (NH4)2SO4 in the same buffer at aflow rate of 0.3 ml/min. Fractions of 0.5 ml were collectedand assayed for the β-glucosidase activity, followed bySDS-PAGE analyses. Active fractions (from 37 to 49) werepooled and then concentrated from 6.5 ml to 550 μl.
Gel filtration
The purification of G1NkBG was followed by gel filtrationchromatography (HiLoad 16/60 Superdex-75 prep grade;60 cm×16 mm). The column was equilibrated and elutedwith 150 mM NaCl in 50 mM Tris–HCl buffer (pH 7.5) at aflow rate of 0.5 ml/min, with the collection of 2-mlfractions that were used for enzyme activity assay, followedby SDS-PAGE analyses. Active fractions (from 25 to 27)were pooled and then concentrated from 6.0 ml to 550 μl.
All of the columns were from GE Healthcare and thechromatography procedures were performed usingÄKTAprime plus (GE Healthcare).
Appl Microbiol Biotechnol (2011) 89:1761–1771 1763

Enzyme properties
SDS-PAGE and Western blotting analysis
After each step of purification, proteins were mixed with2 × sample buffer and boiled for 3 min (Laemmli 1970).Then, electrophoresis was done using 10% polyacrylamidegels. Proteins were stained with Coomassie brilliant blue(CBB) or used in Western blotting analysis. In Westernblotting, anti-β-glucosidase antibody raised against the C-terminal peptide of β-glucosidase from the higher termiteNasutitermes takasagoensis (Tokuda et al. 2009), whichshares 72% amino acid sequence homology with G1NkBG,was used as the primary antibody. The peroxidase-labeledrabbit immunoglobulin G antibody (Cell Signaling) was usedas the secondary antibody. Protein was detected through aluminescent image analyzer LAS-4000miniEPUV (Fujifilm,Japan) using ELC detection reagents (Pierce).
Molecular weight determination
The native molecular weight of the purified G1NkBG wasdetermined using a calibration curve made by a mixture ofprotein markers (Sigma) loaded in gel filtration columnHiLoad 16/60 Superdex 75 (GE Healthcare). Blue dextran(2,000,000 Da) was used to determine the void volume ofthe column and albumin (66,000 Da), carbonic anhydrase(29,000 Da), cytochrome C (12,400 Da), and aprotinin(6,500 Da) were the proteins used for column calibration.The column was equilibrated and eluted with 150 mMNaCl in 50mMTris–HCl (pH 7.5) at a flow rate of 0.5 ml/minwith collection of 2-ml fractions.
Post-translational modification
Glycosylation analysis was done by digestion with glyco-peptidase F (Takara, Japan) according to the instructionsgiven by Takara.
Effect of temperature on β-glucosidase activity and stability
In order to determine the optimum temperature ofG1NkBG, an appropriate concentration of the enzymewas incubated with 10 mM pNPG in 50 mM sodiumacetate buffer (pH 5.5) for 30 min at different temperaturesranging from 25 to 60 °C, and the activity was measured asthe standard assay method. To determine the thermostabilityof G1NkBG, the purified enzyme was pre-incubated atdifferent temperatures ranging from 25 °C to 80 °C in50 mM sodium acetate buffer (pH 5.5) without substrate.After incubating for 30 min, the residual β-glucosidaseactivity was measured according to the standard assayprocedure using pNPG as the substrate and the maximum
activity obtained was taken to be 100%. The resultspresented are the average of three different experiments.
Effect of pH on β-glucosidase activity and stability
The optimum pH of G1NkBG was determined by incubatingthe purified enzyme at 45 °C for 30 min in different buffers inthe range 3.0–8.0. To determine the pH stability of theG1NkBG, the purified enzyme was incubated in differentbuffers in the range 3–10 without substrate at 40 °C for30 min. The buffers used in this experiment were 50 mMcitrate (pH 3 to 6), 50 mM sodium acetate (pH 4.5 to 6.5),50 mM phosphate (pH 6 to 8), and 50 mM glycine–NaOH(pH 8 to 10). The remaining activities were measuredaccording to the standard assay procedure using pNPG asthe substrate and the maximum activity obtained was takento be 100%. The results presented are the average of threedifferent experiments.
Effect of cations and reagents on β-glucosidase activity
The effects of various metal ions (5 mM each of MgCl2,MnCl2, CaCl2, FeSO4, ZnSO4, and CuSO4) and reagents(10 mM SDS, dimethyl sulfoxide, dithiothreitol, glycerol,and EDTA) on G1NkBG activity were studied. The enzymewas pre-incubated with reagents at 50 °C for 10 min. Theremaining activities were measured as the standard assayprocedures using pNPG as the substrate, and the controlenzyme activity without cations/reagents was taken as100%. The results presented are the average of threedifferent experiments.
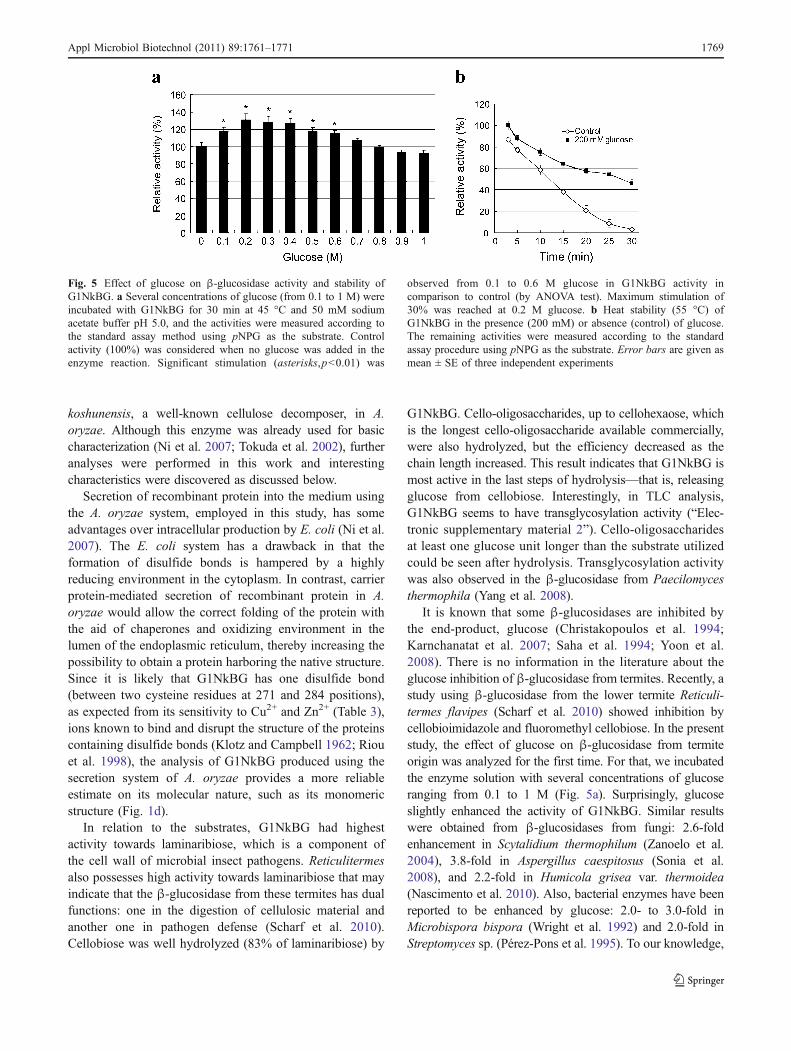
Effect of glucose on β-glucosidase activity and stability
Different concentrations of glucose (0.1 to 1 M) wereincubated with purified G1NkBG at 45 °C for 30 min in50 mM sodium acetate buffer (pH 5.0). After that, the β-glucosidase activities were measured as the standard assayprocedures using pNPG as the substrate. The activity of theenzyme without glucose in the reaction mixture was takenas 100%. For checking β-glucosidase stability, enzymesolution was pre-incubated with 200 mM glucose in 50 mMsodium acetate buffer at 55 °C for 3, 5, 10, 15, 20, 25, and30 min. Enzyme solution without addition of glucose wasused as control. The remaining activities were measuredaccording to the standard assay method. The resultspresented are the average of three different experiments.
Kinetics analysis
Kinetic constants (Km and Vmax) were analyzed at 50 °C in50 mM sodium acetate buffer (pH 5.0). pNPG was used asthe substrate at concentrations ranging from 0.25 to 20 mM.
1764 Appl Microbiol Biotechnol (2011) 89:1761–1771

The initial velocity (v0) was determined by taking samplesafter 15 min of incubation at different substrate concen-trations. The Michaelis constants were calculated fromHanes–Woolf plot using linear regression techniques.
Statistical analysis
In Tables 2 and 3, data were analyzed using a two-tailedStudent’s t test. The data of Fig. 5 were analyzed byANOVA. Differences were regarded as significant at the p<0.05 level.
Results
Expression in A. oryzae
To produce G1NkBG in A. oryzae, G1NkBG was expressedas a fusion protein with glucoamylase, a protein that ishighly secreted by A. oryzae, under the control of the strongpromoter glaA. The expression plasmid pgA-G1NkBG,which contains A. nidulans sC as a selectable marker, was
introduced to A. oryzae (sC− strain). It allowed multiple andrandom integration of the plasmid into the fungus genome.Sixteen transformants were obtained and the production ofthe protein in each transformant was confirmed by Westernblotting analysis. It is clear that the β-glucosidase is fromheterologous origin and not from an endogenous enzyme ofthe host strain (“Electronic supplementary material 1”). Theenzyme activity was also measured, and the one thatpresented the highest β-glucosidase activity was selectedfor large-scale culture and enzyme purification. Theinsertion of the Kex2 cleavage (Lys-Arg) sequence betweenglucoamylase and the putative mature region of G1NkBGled to the release of these two proteins by Kex2-likeprotease in the A. oryzae secretory pathway.
Enzyme purification
From the culture supernatant, proteins were concentrated byammonium sulfate precipitation, and the dialyzed enzymesolution was used in the next step of purification. Figure 1shows the typical elution profile of each chromatography.G1NkBG was purified by anion exchange (Fig. 1a),
Fig. 1 Chromatographic profiles of G1NkBG. a First chromatographyon HiTrap DEAE-Sepharose column. b Second chromatography onHiTrap Phenyl FF. c Third chromatography on HiLoad 16/60 Superdex.Details of chromatography are given in “Materials and methods”. d
Graphic obtained by plotting the logarithmic of molecular weights ofprotein markers (open diamonds) and the ratio Ve/Vo (elution volume/void volume). G1NkBG was eluted at 60 kDa (star)
Appl Microbiol Biotechnol (2011) 89:1761–1771 1765

followed by hydrophobic (Fig. 1b) and gel filtration(Fig. 1c) chromatographies. Samples after each step wererun side by side on SDS-PAGE (Fig. 2). In anion exchangechromatography (Fig. 1a), two protein peaks were observed:the first peak corresponds to the elution of α-amylase,according to Western blotting analysis done with anti-α-amylase antibody (data not shown). β-glucosidase activitywas detected in the second peak after checking by enzymeassay and Western blotting (data not shown). As shown inSDS-PAGE gel stained with CBB (Fig. 2), although the bandof α-amylase at around 50 kDa (asterisk) disappeared afteranion exchange chromatography, some contaminant proteinswere still detected. Hydrophobic chromatography waschosen for the second step of purification. β-Glucosidaseactivity was detected in the second peak (Fig. 1b), but onecontaminant protein at around 30 kDa (dot) still elutedtogether with G1NkBG (Fig. 2). Since the difference in sizebetween β-glucosidase and the contaminant protein wasaround 26 kDa, gel filtration chromatography (molecularweight ranges between 3,000 and 70,000 Da) was performed.
G1NkBG was eluted in the first peak (Fig. 1c) and thehomogeneity of the purified enzyme was confirmed afterCBB staining and Western blotting analysis (Fig. 2a). Asummary of the purification procedures is presented inTable 1; the purity increased by 48-fold and the recoverywas 18% as compared to the culture supernatant. Thespecific activity was 12 U/mg.
Molecular weight determination and post-translationalmodification
In the SDS-PAGE analysis under denaturing conditions,two closely migrating bands could be observed afterpurification (56 and 54 kDa; closed and open arrowheads,respectively, in Fig. 2b). The size of the upper band isslightly higher than what was calculated (54.4 kDa). Onereason for such difference could be glycosylation since onepotential site for N-glycosylation exists near the C-terminusof G1NkBG. To investigate this, the purified enzymewas treated with glycopeptidase F, an enzyme thatcleaves N-linked carbohydrates between asparagine andN-acetylglucosamine (Fig. 2b). This treatment decreasedthe apparent molecular weight of G1NkBG from 56 kDa(closed arrowhead) to around 54 kDa (open arrowhead)and only a single lower band was observed. This resultsuggests that G1NkBG is produced by A. oryzae in twoheterogeneous forms: one is non-glycosylated (openarrowhead) and the other is glycosylated (closed arrowhead)(Fig. 2b). Gel filtration analysis demonstrated that G1NkBGeluted at around 60 kDa, indicating that G1NkBG is amonomeric protein (Fig. 1d).
Effect of temperature and pH on G1NkBG activityand stability
The effects of temperature on enzyme activity and stabilityare shown in Fig. 3. Optimum temperature was 50 °C, andat 60 °C the activity decreased to 15% of the maximumactivity (Fig. 3a). G1NkBG was stable, i.e., retained morethan 80% of its maximum activity up to 45 °C, but wasinactivated at temperatures above 55 °C (Fig. 3b). RegardingpH, G1NkBG presented optimum value at pH 5.0–5.5(Fig. 4a) and was inactivated at extreme pH (pH 3.0 and8.0). G1NkBG retained more than 60% of its maximumactivity from pH 5.0 to 9.0 when pre-incubated at 40 °C for30 min (Fig. 4b). A comparative table with other micro-organisms regarding the temperature and pH is shown in“Electronic supplementary material 3”.
Substrate specificities
In order to investigate the substrate specificities ofG1NkBG, the enzyme was incubated with several aryl-
Fig. 2 SDS-PAGE analyses. a After each step of purification ofG1NkBG, samples were concentrated, loaded on SDS-PAGE gel, andstained with CBB. The right-most column shows Western blottinganalysis performed with anti-β-glucosidase antibody using the samesample as GF. The arrow shows the band of β-glucosidase, theasterisk indicates the band of glucoamylase, and the dot shows acontaminant protein. M, protein marker; ASP, ammonium sulfateprecipitation; AEC, anion exchange chromatography;HC, hydrophobicchromatography; GF, gel filtration; W, Western blotting. b G1NkBGwas treated (+) or not (−) with glycopeptidase F loaded on SDS-PAGEgel and stained with CBB. Reduction in size from 56 kDa (closedarrowhead) to 54 kDa (open arrowhead) indicates the presence of thepost-translational modification by N-glycosylation
1766 Appl Microbiol Biotechnol (2011) 89:1761–1771

glycosides, saccharides, and some oligosaccharides (Table 2).The preferred saccharide and aryl-glycoside substrates forG1NkBG were pNPFuc (βFuc) and laminaribiose (β-1,3-linked glucose), respectively; then, their respective relativeactivity was taken as 100%. pNPG, the substrate used in thestandard assay procedures, was hydrolyzed at 18.0% of thatof pNPFuc. The other two aryl-glycosides tested, pNPLacand pNPGal, were also hydrolyzed (4.3% and 2.9% ofpNPFuc, respectively).
Regarding saccharides, sophorose (β-1,2-linked glucose)was more easily hydrolyzed than gentibiose (β-1,6-linkedglucose). Usually, β-1,6 glycosidic linkage is less susceptibleto hydrolysis by β-glucosidases (Pérez-Pons et al. 1995).G1NkBG hydrolyzed lactose, albeit very poorly (3.8%of laminaribiose), and had very little activity towardsalicin (3.0%) and CMC (2.8%). CMC was the uniqueβ-1,4 polyglycan tested. No activity was detected whenlaminarin (β-1,3 glucan) was used as a substrate. Cello-oligosaccharides up to cellohexaose were readily hydro-lyzed. The release of glucose from cello-oligosaccharides wasalso analyzed by thin-layer chromatography (“Electronicsupplementary material 2”). Interestingly, the products withat least one glucose unit longer than the original substrateswere observed when cello-oligosaccharides were reactedwith G1NkBG, which indicates that the enzyme may havesome transglycosylation activity.
All values were statistically (Student’s t test) significantwith p<0.005 in relation to the substrate most hydrolyzed.
Effect of cations and other reagents on the purifiedG1NkBG
The effect of cations and other potential inhibitors on thepurified β-glucosidase activity was studied and shown inTable 3. Significant inactivation (Student’s t test, p<0.001)was observed with Zn2+ and Cu2+ (74.8% and 94.6%,respectively), probably by reacting with cysteine sidechains. The enzyme activity was not affected by somepotential inhibitors, such as EDTA and dimethyl sulfoxide(p>0.1 and p>0.2, respectively). G1NkBG was slightlyaffected by DTT, Mg2+, Ca2+, and Fe2+ (p<0.01). Theactivity was stimulated by the addition of Mn2+ andglycerol (12.3% and 5.2% and p<0.02 and p<0.005,respectively), probably due to the stabilization of theenzyme, and totally inhibited by SDS.
Effect of glucose on β-glucosidase activity and stabilityof G1NkBG
To check the effect of a possible inhibitor, glucose, on theβ-glucosidase activity of G1NkBG, several concentrationsof glucose (from 0.1 to 1 M) were incubated with the
Fig. 3 Effect of temperature on β-glucosidase activity and stability ofG1NkBG. a Effect of temperature on enzyme activity. Proteins wereincubated at different temperatures and the activities were measured bythe standard assay procedure. b Effect of temperature on the stability of
G1NkBG. The enzyme solution was incubated with 50 mM sodiumacetate buffer (pH 5.5; without substrate) at different temperatures for30 min and the residual activities were measured. Error bars are givenas mean ± SE of three independent experiments
Table 1 Summary of purification of G1NkBG
Total activity (unit, U) Total protein (mg) Specific activity (U/mg) Recovery (%) Purification fold
Culture supernatant 22.0 85.0 0.3 100 1.0
(NH4)2SO4 precipitation 21.6 43.0 0.5 98 1.9
HiTrap DEAE-Sepharose 7.4 2.0 3.8 33 14.5
HiTrap Phenyl FF 5.2 1.0 5.4 23 20.7
HiLoad 16/60 Superdex 4.0 0.3 12.4 18 47.7
Appl Microbiol Biotechnol (2011) 89:1761–1771 1767

G1NkBG (Fig. 5a). Glucose slightly enhanced the activityof G1NkBG from 0.1 to 0.6 M glucose (ANOVA, p<0.01),reaching a maximum value (30%) at 0.2 M. G1NkBGpreserved 100% of its activity at up to 600 mM of glucoseand still showed 92% of its activity in the presence of 1 Mglucose. Regarding the heat stability, while the enzyme(control; without glucose) lost its activity after 30 min ofincubation at 55 °C, addition of 200 mM glucose to the
enzyme solution retarded its inactivation (Fig. 5b). Around46% of the total activity was still present in such condition.
Kinetic analysis
The reaction kinetics of the purified G1NkBG wasdetermined from Hanes–Woolf plot, using pNPG as asubstrate, at the optimum conditions (pH 5.0 and 50 °C).The Km and Vmax for pNPG were 0.77 mM (or 0.23 mg/ml)and 16 U/mg of protein, respectively.
Discussion
In the present study, we reported the successful heterologousexpression of β-glucosidase from the lower termite N.
Fig. 4 Effect of pH on β-glucosidase activity and stability ofG1NkBG. a Effect of pH on enzyme activity. Proteins were incubatedat different pH and the activities were measured by the standard assayprocedure. b Effect of pH on the stability of purified β-glucosidase.The enzyme solution was incubated with buffer (without substrate) atdifferent pH and the residual activities were measured according to the
standard assay procedure. Error bars are given as mean ± SE of threeindependent experiments. The buffers used were 50 mM citrate buffer(pH 3.0 to 6.0; closed circles), 50 mM sodium acetate buffer (pH 4.5to 6.5; open squares), 50 mM phosphate buffer (pH 6.0 to 8.0; closedtriangles), 50 mM glycine–NaOH buffer (pH 8.0 to 10; opendiamonds)
Table 2 Relative activity (%) of G1NkBG on various substrates
Substrates Linkage ofglycosyl group
Relativeactivity (%)a
Saccharides
Laminaribiose (β-1,3) Glc 100.0±2.18
Sophorose (β-1,2) Glc 32.1±0.97
Gentibiose (β-1,6) Glc 6.0±0.75
Lactose (β-1,4) Gal 3.8±0.83
Laminarin (β-1,3) Glc 0.0±0.00
CMC (β-1,4) Glc 2.8±0.50
Salicin βGlc 3.0±0.51
Cellobiose (β-1,4) Glc 82.9±2.15
Cellotriose (β-1,4) Glc 54.9±1.84
Cellotetraose (β-1,4) Glc 47.7±0.69
Cellopentaose (β-1,4) Glc 45.6±0.51
Cellohexaose (β-1,4) Glc 42.7±0.61
Aryl-glycosides
p-Nitrophenyl-β-D-fucopyranoside βFuc 100.0±2.52
p-Nitrophenyl-β-D-glucopyranoside βGlc 18.0±0.29
p-Nitrophenyl-β-D-lactopyranoside βLac 4.29±0.37
p-Nitrophenyl-β-D-galactopyranoside βGal 2.85±0.29
a The release of sugars were measured at A505 for saccharides and at A410
for aryl-glycosides. The relative activity of the most preferentiallyhydrolyzed substrates was taken as 100%. Each value is the mean oftriplicate experiments
Table 3 The effect of various cations and reagents on β-glucosidaseactivity of G1NkBG
Cation or reagent Relative activity (%)a
Control 100.0±1.79
MnCl2 112.3±5.14
MgCl2 92.0±2.14
CaCl2 91.2±1.12
FeSO4 83.0±2.19
ZnSO4 25.2±2.47
CuSO4 5.4±0.73
Glycerol 105.2±2.46
EDTA 99.4±1.84
Dimethyl sulfoxide 98.3±1.89
Dithiothreitol 81.0±0.69
SDS 0.0±00
a Control activity (100%) was determined when none of the substanceswere added. Each value is the mean of triplicate experiments
1768 Appl Microbiol Biotechnol (2011) 89:1761–1771
koshunensis, a well-known cellulose decomposer, in A.oryzae. Although this enzyme was already used for basiccharacterization (Ni et al. 2007; Tokuda et al. 2002), furtheranalyses were performed in this work and interestingcharacteristics were discovered as discussed below.
Secretion of recombinant protein into the medium usingthe A. oryzae system, employed in this study, has someadvantages over intracellular production by E. coli (Ni et al.2007). The E. coli system has a drawback in that theformation of disulfide bonds is hampered by a highlyreducing environment in the cytoplasm. In contrast, carrierprotein-mediated secretion of recombinant protein in A.oryzae would allow the correct folding of the protein withthe aid of chaperones and oxidizing environment in thelumen of the endoplasmic reticulum, thereby increasing thepossibility to obtain a protein harboring the native structure.Since it is likely that G1NkBG has one disulfide bond(between two cysteine residues at 271 and 284 positions),as expected from its sensitivity to Cu2+ and Zn2+ (Table 3),ions known to bind and disrupt the structure of the proteinscontaining disulfide bonds (Klotz and Campbell 1962; Riouet al. 1998), the analysis of G1NkBG produced using thesecretion system of A. oryzae provides a more reliableestimate on its molecular nature, such as its monomericstructure (Fig. 1d).
In relation to the substrates, G1NkBG had highestactivity towards laminaribiose, which is a component ofthe cell wall of microbial insect pathogens. Reticulitermesalso possesses high activity towards laminaribiose that mayindicate that the β-glucosidase from these termites has dualfunctions: one in the digestion of cellulosic material andanother one in pathogen defense (Scharf et al. 2010).Cellobiose was well hydrolyzed (83% of laminaribiose) by
G1NkBG. Cello-oligosaccharides, up to cellohexaose, whichis the longest cello-oligosaccharide available commercially,were also hydrolyzed, but the efficiency decreased as thechain length increased. This result indicates that G1NkBG ismost active in the last steps of hydrolysis—that is, releasingglucose from cellobiose. Interestingly, in TLC analysis,G1NkBG seems to have transglycosylation activity (“Elec-tronic supplementary material 2”). Cello-oligosaccharidesat least one glucose unit longer than the substrate utilizedcould be seen after hydrolysis. Transglycosylation activitywas also observed in the β-glucosidase from Paecilomycesthermophila (Yang et al. 2008).
It is known that some β-glucosidases are inhibited bythe end-product, glucose (Christakopoulos et al. 1994;Karnchanatat et al. 2007; Saha et al. 1994; Yoon et al.2008). There is no information in the literature about theglucose inhibition of β-glucosidase from termites. Recently, astudy using β-glucosidase from the lower termite Reticuli-termes flavipes (Scharf et al. 2010) showed inhibition bycellobioimidazole and fluoromethyl cellobiose. In the presentstudy, the effect of glucose on β-glucosidase from termiteorigin was analyzed for the first time. For that, we incubatedthe enzyme solution with several concentrations of glucoseranging from 0.1 to 1 M (Fig. 5a). Surprisingly, glucoseslightly enhanced the activity of G1NkBG. Similar resultswere obtained from β-glucosidases from fungi: 2.6-foldenhancement in Scytalidium thermophilum (Zanoelo et al.2004), 3.8-fold in Aspergillus caespitosus (Sonia et al.2008), and 2.2-fold in Humicola grisea var. thermoidea(Nascimento et al. 2010). Also, bacterial enzymes have beenreported to be enhanced by glucose: 2.0- to 3.0-fold inMicrobispora bispora (Wright et al. 1992) and 2.0-fold inStreptomyces sp. (Pérez-Pons et al. 1995). To our knowledge,
Fig. 5 Effect of glucose on β-glucosidase activity and stability ofG1NkBG. a Several concentrations of glucose (from 0.1 to 1 M) wereincubated with G1NkBG for 30 min at 45 °C and 50 mM sodiumacetate buffer pH 5.0, and the activities were measured according tothe standard assay method using pNPG as the substrate. Controlactivity (100%) was considered when no glucose was added in theenzyme reaction. Significant stimulation (asterisks,p<0.01) was
observed from 0.1 to 0.6 M glucose in G1NkBG activity incomparison to control (by ANOVA test). Maximum stimulation of30% was reached at 0.2 M glucose. b Heat stability (55 °C) ofG1NkBG in the presence (200 mM) or absence (control) of glucose.The remaining activities were measured according to the standardassay procedure using pNPG as the substrate. Error bars are given asmean ± SE of three independent experiments
Appl Microbiol Biotechnol (2011) 89:1761–1771 1769

however, this is the first time that such stimulation isobserved in β-glucosidase from animal origin.
In addition, when G1NkBG was incubated with 200 mMglucose, its inactivation by heat (55 °C) was delayed,while in the absence of glucose, G1NkBG was completelyinactivated after 30 min. In the presence of glucose, theenzyme still had 46% of its maximum activity (Fig. 5b).Protection from inactivation was also observed when β-glucosidase from H. grisea was incubated with 50 mM ofglucose (Nascimento et al. 2010). Differently, β-glucosidasefrom R. flavipes had its temperature stability increasedaround 20% in the presence of 30 mM calcium chloride(Scharf et al. 2010).
In conclusion, A. oryzae was confirmed as a suitable hostfor heterologous protein production. The high activity ofG1NkBG towards cellobiose and cello-oligosaccharides aswell as the stimulatory effect of glucose on its enzymeactivity/stability are some of the characteristics that renderthis enzyme of great interest in the process of enzymatichydrolysis of cellulose to glucose. In addition, the trans-glycosylation activity of G1NkBG widens the possibilitiesof its usage in another area of commercial interest. Furtherstudies must be done using endogenous β-glucosidasesfrom termite origin in order to compare their applicability inbiotechnological processes.
Acknowledgments This work was supported in part by a grant fromBio-oriented Technology Research Advancement Institution(BRAIN), Japan. We thank the Ministry of Education, Culture, Sports,Science, and Technology of Japan for the scholarship received by C.A.U. The authors are also thankful to the anonymous reviewers fortheir constructive comments, which helped us to improve themanuscript.
References
Balat M, Balat H, Öz C (2008) Progress in bioethanol processing.Prog Energy Comb Sci 34:551–573
Béguin P, Aubert JP (1994) The biological degradation on cellulose.FEMS Microbiol Rev 13:25–58
Bhatia Y,Mishra S, Bisaria VS (2002)Microbialβ-glucosidases: cloning,properties, and applications. Crit Rev Biotechnol 22:375–407
Bothast RJ, Saha BC (1997) Ethanol production from agriculturalbiomass substrates. Adv Appl Microbiol 44:261–286
Bradford MM (1976) A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal Biochem 72:248–254
Carroll A, Somerville C (2009) Cellulosic biofuels. Annu Rev PlantBiol 60:165–182
Christakopoulos P, Goodenough PW, Kekos D, Macris BJ, ClaeyssensM, Bhat MK (1994) Purification and characterisation of anextracellular β-glucosidase with transglycosylation and exo-glucosidase activities from Fusarium oxysporum. Eur J Biochem224:379–385
Dashtban M, Schraft H, Qin W (2009) Fungal bioconversion oflignocellulosic residues; opportunities & perspectives. Int J BiolSci 5:578–595
Günata Z, Vallier MJ (1999) Production of a highly glucose-tolerantextracellular β-glucosidase by threeAspergillus strains. BiotechnolLett 21:219–223
Karnchanatat A, Petsom A, Sangvanich P, Piaphukiew J, Whalley AJ,Reynolds CD, Sihanonth P (2007) Purification and biochemicalcharacterization of an extracellular β-glucosidase from the wood-decaying fungus Daldinia eschscholzii (Ehrenb.:Fr.) Rehm.FEMS Microbiol Lett 270:162–170
Kaur J, Chadha BS, Kumar BA, Kaur GS, Saini HS (2007)Purification and characterization of β-glucosidase from Melano-carpus sp. MTCC 3922. Eletron J Biotechnol 10:260–270
Kitamoto K (2002) Molecular biology of the Koji molds. Adv ApplMicrobiol 51:129–153
Klotz IM, Campbell BJ (1962) Copper-induced hydrolysis of thedisulfide bond. Arch Biochem Biophys 96:92–99
Kotaka A, Bando H, Kaya M, Kato-Murai M, Kuroda K, Sahara H,Hata Y, Kondo A, Ueda M (2008) Direct ethanol productionfrom barley β-glucan by sake yeast displaying Aspergillus oryzaeβ-glucosidase and endoglucanase. J Biosci Bioeng 105:622–627
Laemmli UK (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227:680–685
Lynd LR, Weimer PJ, van Zyl WH, Pretorius IS (2002) Microbialcellulose utilization: fundamentals and biotechnology. MicrobiolMol Biol Rev 66:506–577
Mabashi Y, Kikuma T, Maruyama J, Arioka M, Kitamoto K (2006)Development of a versatile expression plasmid constructionsystem for Aspergillus oryzae and its application to visualizationof mitochondria. Biosci Biotechnol Biochem 70:1882–1889
Nascimento CV, Souza FH, Masui DC, Leone FA, Peralta RM, JorgeJA, Furriel RP (2010) Purification and biochemical properties ofa glucose-stimulated β-D-glucosidase produced by Humicolagrisea var. thermoidea grown on sugarcane bagasse. J Microbiol48:53–62
Nemoto T, Watanabe T, Mizogami Y, Maruyama J, Kitamoto K (2009)Isolation of Aspergillus oryzae mutants for heterologous proteinproduction from a double proteinase gene disruptant. ApplMicrobiol Biotechnol 82:1105–1114
Ni J, Tokuda G, Takehara M, Watanabe H (2007) Heterologousexpression and enzymatic characterization of β-glucosidase fromthe drywood-eating termite, Neotermes koshunensis. Appl Ento-mol Zool 42:457–463
Pérez-Pons JA, Rebordosa X, Querol E (1995) Properties of a novelglucose-enhanced β-glucosidase purified from Streptomyces sp.(ATCC 11238). Biochim Biophys Acta 1251:145–153
Prins RA, Kreulen DA (1991) Comparative aspects of plant cell walldigestion in insects. Anim Feed Sci Technol 32:101–118
Riou C, Salmon JM, Vallier MJ, Gunata Z, Barre P (1998)Purification, characterization, and substrate specificity of a novelhighly glucose-tolerant β-glucosidase from Aspergillus oryzae.Appl Environ Microbiol 64:3607–3614
Saha BC, Bothast RJ (1996) Production, purification, and character-ization of a highly glucose-tolerant novel β-glucosidase fromCandida peltata. Appl Environ Microbiol 62:3165–3170
Saha BC, Freer SN, Bothast RJ (1994) Production, purification, andproperties of a thermostable β-glucosidase from a color variantstrain of Aureobasidium pullulans. Appl Environ Microbiol60:3774–3780
Scharf ME, Kovaleva ES, Jadhao S, Campbell JH, Buchman GW,Boucias DG (2010) Functional and translational analyses of a β-glucosidase gene (glycosyl hydrolase family 1) isolated from thegut of the lower termite Reticulitermes flavipes. Insect BiochemMol Biol 40:611–620
Sonia KG, Chadha BS, Badhan AK, Saini HS, Bhat MK (2008)Identification of glucose tolerant acid active β-glucosidases fromthermophilic and thermotolerant fungi. World J MicrobiolBiotechnol 24:599–604
1770 Appl Microbiol Biotechnol (2011) 89:1761–1771

Tokuda G, Saito H, Watanabe H (2002) A digestive β-glucosidasefrom the salivary glands of the termite, Neotermes koshunensis(Shiraki): distribution, characterization and isolation of itsprecursor cDNA by 5′- and 3′-RACE amplifications withdegenerate primers. Insect Biochem Mol Biol 32:1681–1689
Tokuda G, Miyagi M, Makiya H, Watanabe H, Arakawa G (2009)Digestive β-glucosidases from the wood-feeding higher termite,Nasutitermes takasagoensis: intestinal distribution, molecularcharacterization, and alteration in sites of expression. InsectBiochem Mol Biol 39:931–937
Watanabe H, Tokuda G (2001) Animal cellulases. Cell Mol Life Sci58:1167–1178
Watanabe H, Tokuda G (2010) Cellulolytic systems in insects. AnnuRev Entomol 55:609–632
Wright RM, Yablonsky MD, Shalita ZP, Goyal AK, Eveleigh DE(1992) Cloning, characterization, and nucleotide sequence of agene encoding Microbispora bispora BglB, a thermostable β-
glucosidase expressed in Escherichia coli. Appl Environ Microbiol58:3455–3465
Yan TR, Lin CL (1997) Purification and characterization of a glucose-tolerant β-glucosidase from Aspergillus niger CCRC 31494.Biosci Biotechnol Biochem 61:965–970
Yang S, Jiang Z, Yan Q, Zhu H (2008) Characterization of athermostable extracellular β-glucosidase with activities of exo-glucanase and transglycosylation from Paecilomyces thermo-phila. J Agric Food Chem 56:602–608
Yoon JJ, Kim KY, Cha CJ (2008) Purification and characterization ofthermostable β-glucosidase from the brown-rot basidiomyceteFomitopsis palustris grown on microcrystalline cellulose. JMicrobiol 46:51–55
Zanoelo FF, Polizeli ML, Terenzi HF, Jorge JA (2004) β-Glucosidaseactivity from the thermophilic fungus Scytalidium thermophilumis stimulated by glucose and xylose. FEMS Microbiol Lett240:137–143
Appl Microbiol Biotechnol (2011) 89:1761–1771 1771
Copyright © 2022 FDOKUMEN




















