
Heat denaturation and aggregation of β-lactoglobulin enriched WPI in the presence of arginine HCl,...
14
FOOD HYDROCOLLOIDS Food Hydrocolloids 20 (2006) 1006–1019 Heat denaturation and aggregation of b-lactoglobulin enriched WPI in the presence of arginine HCl, NaCl and guanidinium HCl at pH 4.0 and 7.0 G. Unterhaslberger a,1 , C. Schmitt a, , C. Sanchez b , C. Appolonia-Nouzille a , A. Raemy a a Department of Food Science, Nestle´Research Center, PO Box 44, CH 1000 Lausanne 26, Switzerland b Laboratoire de Physico-Chimie et Ge´nie Alimentaires, INPL-ENSAIA, 2 Avenue de la Foreˆt de Haye, BP 172, F-54505 Vandoeuvre-le`s-Nancy Cedex, France Received 9 May 2005; accepted 28 October 2005 Abstract The heat denaturation and aggregation behaviour of b-lactoglobulin (b-LG) enriched WPI was investigated at pH 4.0 and 7.0 in the presence of arginine HCl, NaCl and GdnHCl using differential scanning calorimetry (DSC) and dynamic light scattering (DLS). Beside the classical endothermic signal attributed to protein heat denaturation, DSC thermograms displayed appearance of an additional exothermic peak in the presence of cosolute. Using in situ DLS, it was shown that the appearance of the exothermic peaks is linked to protein aggregation, in particular to a strong increase in aggregate size. Upon increased cosolute concentration at pH 4.0 the exothermic peak occurred at temperatures lower than the actual denaturation peak (85 1C). At this pH, negatively charged chloride anions interact with b-LG leading to charge screening and physical aggregation. At pH 7.0, exothermic peaks appeared at higher temperatures than the denaturation peak (75 1C). Upon increased cosolute concentration the exothermic peak was shifted to lower temperatures, indicating protein destabilisation in the presence of cosolutes. Charge screening of b-LG by the positively charged cations (arginine, Na and guanidinium) reduced repulsion forces and promoted aggregation. r 2005 Elsevier Ltd. All rights reserved. Keywords: DSC; Dynamic light scattering; Cosolutes; Amino acids; Exothermic peak; Hofmeister Series 1. Introduction Globular proteins are widely used in food industry, health care and pharmacology. Due to their unique folded structure, these proteins express specific functional proper- ties like enzyme catalysis, ligand binding and transport of biologically active molecules, and also water solubility, flavour binding, gelation, emulsification and foaming properties. Biological functions of proteins in general require correct protein folding and protein stability. However, protein stability is strongly influenced by environmental conditions like pH, ionic strength, protein concentration, pressure, heating temperature, heating time and type of solvent. As a consequence, depending on the environmental conditions, globular proteins rather unfold partially, leading to many energetically equivalent con- formations in a so-called ‘‘quasi-native state’’, or the protein can be completely unfolded. These conformational changes and the unfolding process result in aggregation, precipitation, coagulation or gelation (Boye, Ma, & Harwalkar, 1997; Boye, Ma, Ismail, Harwalkar, & Kalab, 1997; McClements, 2002; Mulvihill & Donovan, 1987). In the presence of cosolutes (salts, amino acids), protein stability is influenced due to changes in the hydration of the protein. This results in stabilisation or destabilisation of the protein structure against thermal unfolding. Complex mechanisms like preferential hydration or preferential interaction of the protein with the salt component ARTICLE IN PRESS www.elsevier.com/locate/foodhyd 0268-005X/$ - see front matter r 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.foodhyd.2005.10.017 Corresponding author. Tel.: +41 21 785 89 36; fax: +41 21 785 85 54. E-mail address: [email protected] (C. Schmitt). 1 Present address: Unternehmensgruppe Theo Mu¨ ller GmbH & Co KG, R&D-Food Science & Sensory, MilchstraXe 4, G-85354 Freising, Germany.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Heat denaturation and aggregation of β-lactoglobulin enriched WPI in the presence of arginine HCl,...

ARTICLE IN PRESS
FOODHYDROCOLLOIDS
0268-005X/$ - s
doi:10.1016/j.fo
�CorrespondE-mail addr
1Present addr
R&D-Food Sc
Germany.
Food Hydrocolloids 20 (2006) 1006–1019
www.elsevier.com/locate/foodhyd
Heat denaturation and aggregation of b-lactoglobulin enrichedWPI in the presence of arginine HCl, NaCl and guanidinium
HCl at pH 4.0 and 7.0
G. Unterhaslbergera,1, C. Schmitta,�, C. Sanchezb, C. Appolonia-Nouzillea, A. Raemya
aDepartment of Food Science, Nestle Research Center, PO Box 44, CH 1000 Lausanne 26, SwitzerlandbLaboratoire de Physico-Chimie et Genie Alimentaires, INPL-ENSAIA, 2 Avenue de la Foret de Haye, BP 172,
F-54505 Vandoeuvre-les-Nancy Cedex, France
Received 9 May 2005; accepted 28 October 2005
Abstract
The heat denaturation and aggregation behaviour of b-lactoglobulin (b-LG) enriched WPI was investigated at pH 4.0 and 7.0 in the
presence of arginine HCl, NaCl and GdnHCl using differential scanning calorimetry (DSC) and dynamic light scattering (DLS). Beside
the classical endothermic signal attributed to protein heat denaturation, DSC thermograms displayed appearance of an additional
exothermic peak in the presence of cosolute. Using in situ DLS, it was shown that the appearance of the exothermic peaks is linked to
protein aggregation, in particular to a strong increase in aggregate size. Upon increased cosolute concentration at pH 4.0 the exothermic
peak occurred at temperatures lower than the actual denaturation peak (�85 1C). At this pH, negatively charged chloride anions interact
with b-LG leading to charge screening and physical aggregation. At pH 7.0, exothermic peaks appeared at higher temperatures than the
denaturation peak (�75 1C). Upon increased cosolute concentration the exothermic peak was shifted to lower temperatures, indicating
protein destabilisation in the presence of cosolutes. Charge screening of b-LG by the positively charged cations (arginine, Na and
guanidinium) reduced repulsion forces and promoted aggregation.
r 2005 Elsevier Ltd. All rights reserved.
Keywords: DSC; Dynamic light scattering; Cosolutes; Amino acids; Exothermic peak; Hofmeister Series
1. Introduction
Globular proteins are widely used in food industry,health care and pharmacology. Due to their unique foldedstructure, these proteins express specific functional proper-ties like enzyme catalysis, ligand binding and transport ofbiologically active molecules, and also water solubility,flavour binding, gelation, emulsification and foamingproperties. Biological functions of proteins in generalrequire correct protein folding and protein stability.However, protein stability is strongly influenced by
ee front matter r 2005 Elsevier Ltd. All rights reserved.
odhyd.2005.10.017
ing author. Tel.: +4121 785 89 36; fax: +4121 785 85 54.
ess: [email protected] (C. Schmitt).
ess: Unternehmensgruppe Theo Muller GmbH & Co KG,
ience & Sensory, MilchstraXe 4, G-85354 Freising,
environmental conditions like pH, ionic strength, proteinconcentration, pressure, heating temperature, heating timeand type of solvent. As a consequence, depending on theenvironmental conditions, globular proteins rather unfoldpartially, leading to many energetically equivalent con-formations in a so-called ‘‘quasi-native state’’, or theprotein can be completely unfolded. These conformationalchanges and the unfolding process result in aggregation,precipitation, coagulation or gelation (Boye, Ma, &Harwalkar, 1997; Boye, Ma, Ismail, Harwalkar, & Kalab,1997; McClements, 2002; Mulvihill & Donovan, 1987).In the presence of cosolutes (salts, amino acids), protein
stability is influenced due to changes in the hydration of theprotein. This results in stabilisation or destabilisation ofthe protein structure against thermal unfolding. Complexmechanisms like preferential hydration or preferentialinteraction of the protein with the salt component

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1007
(cosolute) influence the equilibrium between the native andthe unfolded state (Arakawa, Prestrelski, Kenney, &Carpenter, 2001; Timasheff, 1992). Aggregation is stronglyinfluenced by attraction and repulsion forces, which aredependent on pH, nature of cosolute and solvent, ionicstrength, temperature, charge and concentration of theprotein (McClements, 2002; von Hippel & Schleich, 1969).The presence of ions in the system reduces the repulsionforces at pH values far from the IEP due to chargeshielding and induces heat aggregation more easily(Schokker, Singh, Pinder, & Creamer, 2000; Verheul,Roefs, & de Kruif, 1998). The efficiency of arginine HCland GdnHCl as protein denaturants and refolding agentswas demonstrated for various enzymes and proteins(Arakawa & Timasheff, 1984; De Bernardez Clark, 2001;Hagihara, Tan, & Goto, 1994; Span, Lenarcic, & Lapanje,1974). These cosolutes display strong potential in perturb-ing the structure of proteins by interacting with the peptidebonds of the protein, hindering incorrect protein refoldingand suppressing heat aggregation (Shiraki, Kudou, Fuji-wara, Imanaka, & Takagi, 2002; von Hippel & Schleich,1969; Xie, Guo, Lu, & Zhou, 2004).
b-Lactoglobulin (b-LG) is a well-characterised globularwhey protein with a molecular weight of 36,800 gmol�1
(dimeric form) and an IEP of 5.1 (Hambling, McAlpine, &Sawyer, 1992; McKenzie, 1971). It is a small protein with ahydrodynamic diameter, Dh, of 3.58 nm for the non-heatedmonomeric protein (b-LG variant A). Between pH 3.0 and7.0 the protein exists predominantly in a dimeric form(Dh ¼ 6:2 nm) (Griffin & Griffin, 1993; Pessen, Purcell, &Farrell, 1985). Beside two disulphide bonds, b-LG containsalso a free SH group. This free SH group is activated byconformational changes of the protein between pH 6.5 and8.0 (Tanford Transition) and dominates the heat aggrega-tion mechanism in neutral pH conditions (Brownlow et al.,1997; Surroca, Haverkamp, & Heck, 2002; Watanabe &Klostermeyer, 1976). Previous studies have been carriedout to describe the heat denaturation and aggregationmechanism of b-LG (Aymard, Gimel, Nicolai, & Durand,1996b; Hoffmann, 1997; Roefs & de Kruif, 1994). Besidethe influence of pH and ionic strength on the aggregationbehaviour, additional parameters like protein concentra-tion, heating rate, heating time and temperature have beeninvestigated (Aymard, Durand, & Nicolai, 1996a; Boye,Ma, & Harwalkar, 1997; Boye, Ma, Ismail et al., 1997;Dannenberg & Kessler, 1988; de Wit, 1981; Durand,Gimel, & Nicolai, 2002; Elofsson, Dejmek, & Paulsson,1996; Gimel, Durand, & Nicolai, 1994; Nielsen, Singh, &Latham, 1996; Surroca et al., 2002; Verheul et al., 1998).
Despite the fundamental knowledge on b-LG heataggregation found in the literature, the effects of lowconcentrations of cosolutes (o1M) have been scarcelystudied, especially in acidic conditions. The aim of thisstudy was to investigate the effects of low concentrations ofarginine HCl, NaCl and GdnHCl, on the thermal stabilityand aggregation behaviour of b-LG at pH 4.0 and 7.0.Differential scanning calorimetry (DSC) is a common tool
to monitor protein unfolding and denaturation. However,no clear differentiation can be made between proteinunfolding and the beginning of aggregation. Using in situdynamic light scattering (DLS), protein aggregation can befollowed by measuring intensity fluctuations of scatteredlight due to the presence of protein particles. Therefore, thecombination of DSC and DLS should give insight on bothmechanisms, protein unfolding and aggregation, duringheat treatment of b-LG in the presence of cosolutes.
2. Material and methods
2.1. Material
b-LG enriched whey protein isolate (BioPURE b-LG,lot JE002-8-922, 13-12-2000) was obtained from DaviscoFoods International Inc. (Le Sueur, MN, USA). Theprotein was purified from sweet whey by filtration and ionexchange chromatography. The composition of the powder(wet basis) was 89.7% protein, 8.85% moisture, fat(o1%), lactose (o1%) and 1.36% ash (major componentswere 0.079% Ca2+, 0.013% Mg2+, 0.097% K+, 0.576%Na+, 0.05% Cl�). The protein profile was determined byRP-HPLC and major proteins were as follows: 12.9% a-lactalbumin (a-LA), 35% b-LG A, 32.4% b-LG B, 1.2%bovine serum albumin (BSA). The amount of non-nativeprotein in the powder was about 4% as measured at 20 1Cfrom the ratio between total soluble protein at pH 4.6 and7.0 after centrifugation (26,900g for 15min) (Zhu &Damodaran, 1994). Due to the high content of b-LG(467%) in the powder, the b-LG enriched whey protein ismainly referred to as b-LG or protein instead, unlessotherwise noted (a-LA, BSA).Arginine HCl (BioChemika, lot 431391/1) and GdnHCl
(BioChemika, lot 450290/1) were obtained from FlukaChemie (Buchs, Switzerland). All other reagents used wereanalytical or HPLC grade (Merck, Darmstadt, Germany)unless otherwise stated.
2.2. Sample preparation
The protein dispersion was prepared by dispersion ofb-LG powder in Millipore water, and stirring at 20 1C for2 h. The protein dispersion was stored for 12 h at 4 1C toallow complete hydration. Afterwards, the dispersion(natural pH 7.070.1) was centrifuged at 10,800g
(Rotor SLA-3000, Sorvall Instruments RC5C, Kendro,Switzerland) for 60min to remove insoluble matter.Protein solution was then filtered on paper filter(Schleicher & Schuell filter 5971
2, Germany). The protein
concentration was measured by UV/VIS spectroscopy(e278 ¼ 1:013 cm�1 g�1 l as determined experimentally in acuvette with a pathlength of 1 cm) (UVIKON 810,Kontron Instruments, Flowspec, Switzerland) and adjustedto a b-LG concentration of 40 g l�1. The cosolute solutions(arginine HCl, NaCl and GdnHCl; 0–2000mM) wereprepared in Millipore water. Mixtures of protein and

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191008
cosolute solutions were prepared by mixing equal volumesof protein and cosolute to a final protein concentration of20 g l�1 and 0–1000mM of cosolute and were adjusted topH 4.0 and 7.0 by addition of 1M HCl or NaOH. Theamount of acid and base (additional Na+ and Cl� ions)needed to adjust the pH of the solutions was less than 2%of the final molarity for pH 7 and less than 3–4% for pH4.0. These amounts were neglected in the final cosolutemolarity calculation.
For the sake of clarity, the following nomenclature wasused for sample coding: A, arginine HCl; N, NaCl; G,GdnHCl with the subsequent specification 0 for b-LGalone, 10 for a cosolute concentration of 10mM and soforth.
2.3. Differential scanning calorimetry
DSC experiments were carried out using a micro-DSCIII (Setaram, Caluire, France). The sample (�500mg) wasmeasured against a reference sample of Millipore water in atemperature range between 20 and 110 1C at a heating rateof 0.5 1Cmin�1. Measurements were performed twice inmixtures of 20 g l�1 of protein and 0–200mM of cosolute(arginine HCl, GdnHCl or NaCl) at pH 4.0 and0–1000mM of cosolute at pH 7.0.
2.4. Dynamic light scattering
b-LG heat aggregation was followed using DLS. ANanosizer ZS apparatus (Malvern Instruments, UK)equipped with a 4mW laser emitting at 633 nm was used.The instrument was used in the backscattering configura-tion where detection is done at a scattering angle of 1731.This allows considerable reduction of multiple lightscattering in turbid samples. Samples were placed in asquared quartz cell (Hellma, pathlength 1 cm). Thepathlength of the light beam was automatically set by theapparatus, depending on the sample turbidity (attenua-tion). The in situ kinetics of aggregation were studied byheating with a Peltier heating device. The protein disper-sions were filtered in a two-step process, using 0.1 and0.025 mm filters (NC 03 Membrane Filters, Schleicher &Schuell, Germany) prior to heating to eliminate dustparticles and protein aggregates larger than 25 nm. The insitu heating kinetics were performed predominantly twicein a temperature range between 60 and 9071 1C with aheating rate of 0.5 1Cmin�1 as done with DSC. Theautocorrelation function was calculated from the fluctua-tion of the scattered intensity. From the fit of thecorrelation function using the ‘‘Cumulant’’ technique(Stepanek, 1993), the hydrodynamic diameter (Dh) wascalculated using the Stokes–Einstein equation:
Dapp ¼kBT
3pZDh
,
where kB (1.38� 10�23NmK�1) represents the Boltzmannconstant, T (K) the temperature, Z (N sm�2) the solvent
viscosity and Dapp the translational diffusion coefficient ofparticles (m2 s�1) (Berne & Pecora, 1976; Liu & Chu, 2002).
3. Results and discussion
3.1. Protein stability measured by DSC
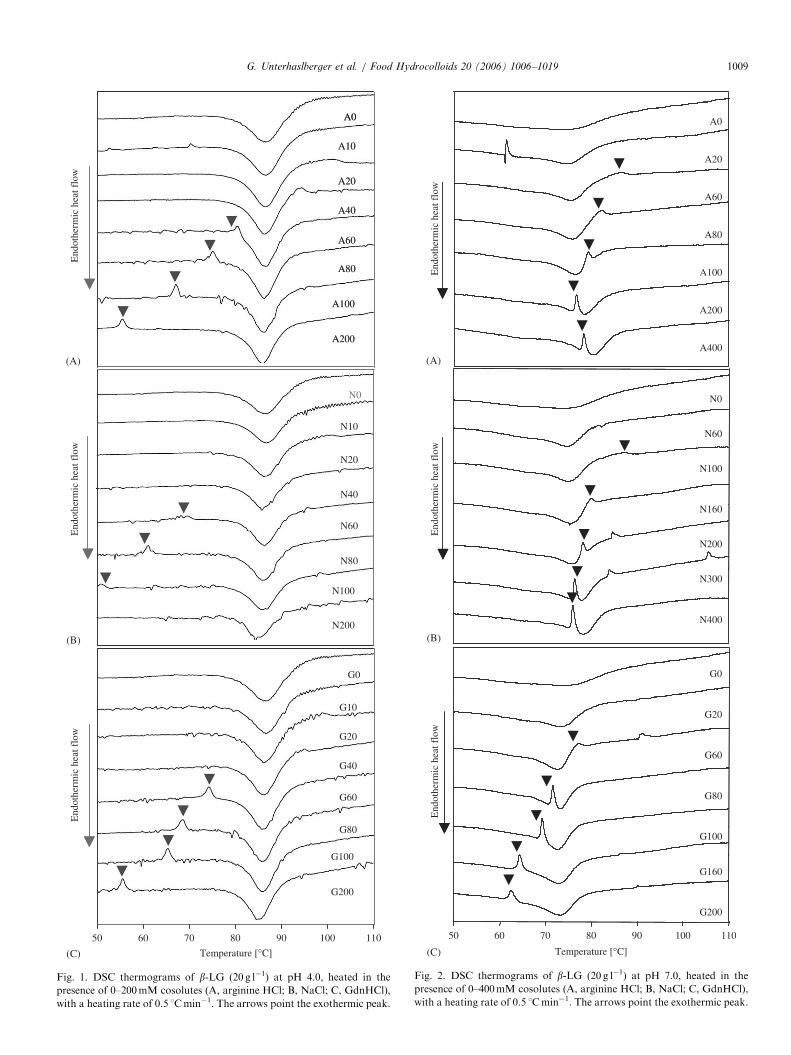
Fig. 1A–C shows thermograms of the heat denaturationof b-LG in the presence of increasing concentrations ofcosolutes at pH 4.0. A characteristic broad endothermicpeak for b-LG heat denaturation was observed at 85 1C.This indicated high heat stability of the protein, inagreement with values reported earlier (Boye, Ma, &Harwalkar, 1997; Boye, Ma, Ismail et al., 1997; Hegg,1980; Verheul et al., 1998). Interestingly, the increase incosolute concentration did not decrease the denaturationtemperature (Td) of b-LG significantly (DTd ¼ 2 1C). Atcosolute concentrations higher than 40mM an exothermicpeak appeared at lower heating temperatures than theendothermic denaturation peak (arrows in Fig. 1A–C).With increasing cosolute concentration this exothermicpeak shifted to lower temperatures. It is worth mentioningthat the exothermic peaks occurred at lower cosoluteconcentrations and temperatures for NaCl than forGdnHCl and arginine HCl.At pH 7.0, b-LG (A0, N0, G0) showed a broad
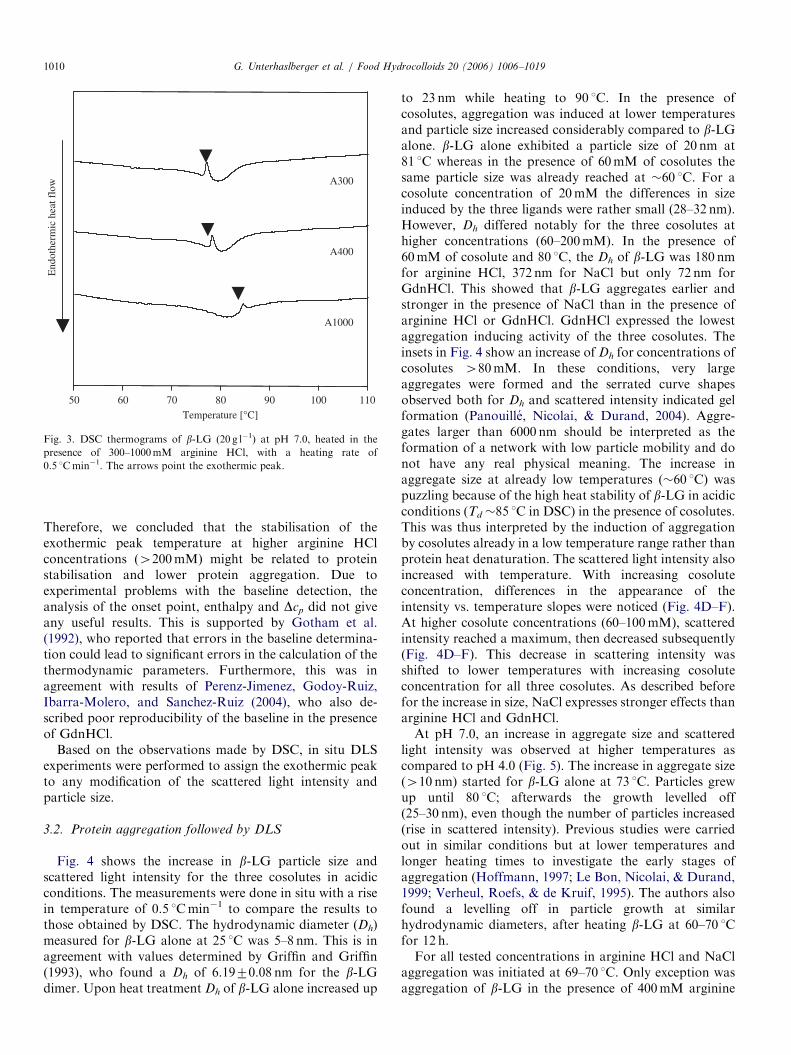
denaturation profile with one endothermic peak (Fig.2A–C). In the presence of arginine HCl, NaCl andGdnHCl, the profile of the endothermic denaturation peakwas more pronounced, possibly due to the combined endo-and exothermic processes. At higher cosolute concentra-tions a second peak appeared. However, it could not beclearly identified to be exothermic. For all three cosolutes,the peak appeared as a small shoulder at high temperatures(85–90 1C). With increased cosolute concentration the peakshifted to lower temperatures and got more defined. Itsexothermic nature was then clearly expressed. GdnHClshowed a distinctively stronger effect than arginine HCland NaCl on the shift of the exothermic peak temperature.Interestingly, for arginine HCl concentrations higher than200mM, the exothermic peak temperature was shifted tohigh values (76 1C for 200mM vs. 85 1C for 1000mM)(Fig. 3). This stabilisation effect was not found for NaCland GdnHCl in the experimental conditions tested.The appearance of exothermic peaks in DSC thermo-
grams of protein solutions is often associated with crystal-lisation processes, as well as Maillard reactions in thepresence of reducing sugars (de Wit, 1981; de Wit &Klarenbeek, 1981; Gotham, Fryer, & Pritchard, 1992).However, the latter could be excluded as possible mechan-ism, because of the low amount of lactose in the sample(o1%). Nevertheless, as protein aggregation and crystal-lisation basically proceed from similar mechanisms (over-coming of the energetic barrier, nucleation and growth),the exothermic peak could be considered to be associatedwith protein aggregation (Georgalis, Umbach, Saenger,Ihmels, & Soumpasis, 1999; Marshall & Zarins, 1989).
ARTICLE IN PRESS
A200
A100
A80
A60
A40
A20
A10
A0
N0
N10
N20
N40
N60
N80
N100
N200
Temperature [°C]
50 60 70 80 90 100 110
End
othe
rmic
hea
t flo
wE
ndot
herm
ic h
eat f
low
End
othe
rmic
hea
t flo
w
G0
G20
G40
G60
G80
G100
G200
G10
A200
A100
A80
A60
A40
A20
A10
A0
(A)
(B)
(C)
Fig. 1. DSC thermograms of b-LG (20 g l�1) at pH 4.0, heated in the
presence of 0–200mM cosolutes (A, arginine HCl; B, NaCl; C, GdnHCl),
with a heating rate of 0.5 1Cmin�1. The arrows point the exothermic peak.
Temperature [°C]
50 60 70 80 90 100 110
G200
G100
G80
G60
G20
G0
G160
N400
N300
N200
N160
N100
N60
N0
End
othe
rmic
hea
t flo
wE
ndot
herm
ic h
eat f
low
End
othe
rmic
hea
t flo
w
A400
A200
A100
A80
A60
A20
A0
(A)
(B)
(C)
Fig. 2. DSC thermograms of b-LG (20 g l�1) at pH 7.0, heated in the
presence of 0–400mM cosolutes (A, arginine HCl; B, NaCl; C, GdnHCl),
with a heating rate of 0.5 1Cmin�1. The arrows point the exothermic peak.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1009
ARTICLE IN PRESS
A300
Temperature [°C]
50 60 70 80 90 100 110
End
othe
rmic
hea
t flo
w
A400
A1000
Fig. 3. DSC thermograms of b-LG (20 g l�1) at pH 7.0, heated in the
presence of 300–1000mM arginine HCl, with a heating rate of
0.5 1Cmin�1. The arrows point the exothermic peak.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191010
Therefore, we concluded that the stabilisation of theexothermic peak temperature at higher arginine HClconcentrations (4200mM) might be related to proteinstabilisation and lower protein aggregation. Due toexperimental problems with the baseline detection, theanalysis of the onset point, enthalpy and Dcp did not giveany useful results. This is supported by Gotham et al.(1992), who reported that errors in the baseline determina-tion could lead to significant errors in the calculation of thethermodynamic parameters. Furthermore, this was inagreement with results of Perenz-Jimenez, Godoy-Ruiz,Ibarra-Molero, and Sanchez-Ruiz (2004), who also de-scribed poor reproducibility of the baseline in the presenceof GdnHCl.
Based on the observations made by DSC, in situ DLSexperiments were performed to assign the exothermic peakto any modification of the scattered light intensity andparticle size.
3.2. Protein aggregation followed by DLS
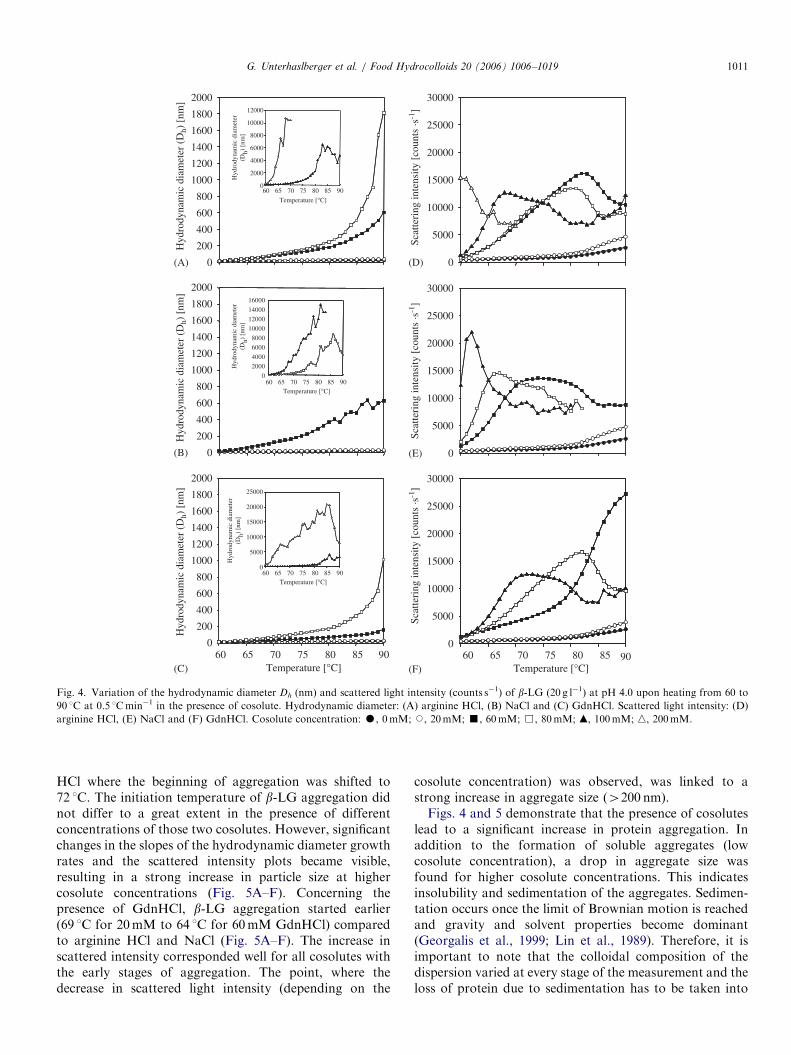
Fig. 4 shows the increase in b-LG particle size andscattered light intensity for the three cosolutes in acidicconditions. The measurements were done in situ with a risein temperature of 0.5 1Cmin�1 to compare the results tothose obtained by DSC. The hydrodynamic diameter (Dh)measured for b-LG alone at 25 1C was 5–8 nm. This is inagreement with values determined by Griffin and Griffin(1993), who found a Dh of 6.1970.08 nm for the b-LGdimer. Upon heat treatment Dh of b-LG alone increased up
to 23 nm while heating to 90 1C. In the presence ofcosolutes, aggregation was induced at lower temperaturesand particle size increased considerably compared to b-LGalone. b-LG alone exhibited a particle size of 20 nm at81 1C whereas in the presence of 60mM of cosolutes thesame particle size was already reached at �60 1C. For acosolute concentration of 20mM the differences in sizeinduced by the three ligands were rather small (28–32 nm).However, Dh differed notably for the three cosolutes athigher concentrations (60–200mM). In the presence of60mM of cosolute and 80 1C, the Dh of b-LG was 180 nmfor arginine HCl, 372 nm for NaCl but only 72 nm forGdnHCl. This showed that b-LG aggregates earlier andstronger in the presence of NaCl than in the presence ofarginine HCl or GdnHCl. GdnHCl expressed the lowestaggregation inducing activity of the three cosolutes. Theinsets in Fig. 4 show an increase of Dh for concentrations ofcosolutes 480mM. In these conditions, very largeaggregates were formed and the serrated curve shapesobserved both for Dh and scattered intensity indicated gelformation (Panouille, Nicolai, & Durand, 2004). Aggre-gates larger than 6000 nm should be interpreted as theformation of a network with low particle mobility and donot have any real physical meaning. The increase inaggregate size at already low temperatures (�60 1C) waspuzzling because of the high heat stability of b-LG in acidicconditions (Td �85 1C in DSC) in the presence of cosolutes.This was thus interpreted by the induction of aggregationby cosolutes already in a low temperature range rather thanprotein heat denaturation. The scattered light intensity alsoincreased with temperature. With increasing cosoluteconcentration, differences in the appearance of theintensity vs. temperature slopes were noticed (Fig. 4D–F).At higher cosolute concentrations (60–100mM), scatteredintensity reached a maximum, then decreased subsequently(Fig. 4D–F). This decrease in scattering intensity wasshifted to lower temperatures with increasing cosoluteconcentration for all three cosolutes. As described beforefor the increase in size, NaCl expresses stronger effects thanarginine HCl and GdnHCl.At pH 7.0, an increase in aggregate size and scattered
light intensity was observed at higher temperatures ascompared to pH 4.0 (Fig. 5). The increase in aggregate size(410 nm) started for b-LG alone at 73 1C. Particles grewup until 80 1C; afterwards the growth levelled off(25–30 nm), even though the number of particles increased(rise in scattered intensity). Previous studies were carriedout in similar conditions but at lower temperatures andlonger heating times to investigate the early stages ofaggregation (Hoffmann, 1997; Le Bon, Nicolai, & Durand,1999; Verheul, Roefs, & de Kruif, 1995). The authors alsofound a levelling off in particle growth at similarhydrodynamic diameters, after heating b-LG at 60–70 1Cfor 12 h.For all tested concentrations in arginine HCl and NaCl
aggregation was initiated at 69–70 1C. Only exception wasaggregation of b-LG in the presence of 400mM arginine
ARTICLE IN PRESS
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000
0
200
400
600
800
1000
1200
1400
1600
1800
2000
Temperature [°C]
Temperature [°C]
60 65 70 75 80 85 90
Hyd
rody
nam
ic d
iam
eter
(Dh)
[nm
]H
ydro
dyna
mic
dia
met
er(D
h) [n
m]
Hyd
rody
nam
ic d
iam
eter
(Dh)
[nm
]
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
0
2000
4000
6000
8000
10000
12000
(A)
60 65 70 75 80 85 90
Temperature [°C]60 65 70 75 80 85 90
0
2000
4000
6000
8000
10000
12000
14000
16000
(B)
Temperature [°C]60 65 70 75 80 85 90
Temperature [°C]60 65 70 75 80 85 90
0
5000
10000
15000
20000
25000
(C)
(E)
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
5000
10000
15000
20000
25000
30000
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
5000
10000
15000
20000
25000
30000
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
5000
10000
15000
20000
25000
30000
(D)
(F)
Fig. 4. Variation of the hydrodynamic diameter Dh (nm) and scattered light intensity (counts s�1) of b-LG (20 g l�1) at pH 4.0 upon heating from 60 to
90 1C at 0.5 1Cmin�1 in the presence of cosolute. Hydrodynamic diameter: (A) arginine HCl, (B) NaCl and (C) GdnHCl. Scattered light intensity: (D)
arginine HCl, (E) NaCl and (F) GdnHCl. Cosolute concentration: K, 0mM; J, 20mM; ’, 60mM; &, 80mM; m, 100mM; n, 200mM.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1011
HCl where the beginning of aggregation was shifted to72 1C. The initiation temperature of b-LG aggregation didnot differ to a great extent in the presence of differentconcentrations of those two cosolutes. However, significantchanges in the slopes of the hydrodynamic diameter growthrates and the scattered intensity plots became visible,resulting in a strong increase in particle size at highercosolute concentrations (Fig. 5A–F). Concerning thepresence of GdnHCl, b-LG aggregation started earlier(69 1C for 20mM to 64 1C for 60mM GdnHCl) comparedto arginine HCl and NaCl (Fig. 5A–F). The increase inscattered intensity corresponded well for all cosolutes withthe early stages of aggregation. The point, where thedecrease in scattered light intensity (depending on the
cosolute concentration) was observed, was linked to astrong increase in aggregate size (4200 nm).Figs. 4 and 5 demonstrate that the presence of cosolutes
lead to a significant increase in protein aggregation. Inaddition to the formation of soluble aggregates (lowcosolute concentration), a drop in aggregate size wasfound for higher cosolute concentrations. This indicatesinsolubility and sedimentation of the aggregates. Sedimen-tation occurs once the limit of Brownian motion is reachedand gravity and solvent properties become dominant(Georgalis et al., 1999; Lin et al., 1989). Therefore, it isimportant to note that the colloidal composition of thedispersion varied at every stage of the measurement and theloss of protein due to sedimentation has to be taken into

ARTICLE IN PRESS
0
1500
1000
500
2000
Temperature [°C]60 65 70 75 80 85 90
Temperature [°C]
Temperature [°C]
60 65 70 75 80 85 90
Temperature [°C]60 65 70 75 80 85 90
Temperature [°C]60 65 70 75 80 85 90
Hyd
rody
nam
ic d
iam
eter
(Dh)
[nm
]
Hyd
rody
nam
ic d
iam
eter
(Dh)
[nm
]
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
0
5000
10000
15000
20000
25000
30000
35000
0
1000
4000
3000
2000
Hyd
rody
nam
ic d
iam
eter
(D
h) [
nm]
0
1000
4000
3000
2000
0
2000
4000
6000
8000
10000
12000
14000
0
2000
4000
6000
8000
10000
12000
14000
16000
60 65 70 75 80 85 90
Temperature [°C]
60 65 70 75 80 85 90
0
10000
20000
30000
40000
30000
(A)
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
Scat
teri
ng in
tens
ity[c
ount
s ·s
-1]
Temperature [°C]60 65 70 75 80 85 90
0
2000
4000
6000
8000
10000
12000
14000
Scat
teri
ng in
tens
ity[c
ount
s ·s
-1]
Scat
teri
ng in
tens
ity[c
ount
s ·s
-1]
(F)
(E)
(C)
(B)
(D)
20000
15000
10000
5000
25000
30000
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
20000
15000
10000
5000
25000
30000
Scat
teri
ng in
tens
ity [
coun
ts ·s
-1]
0
20000
15000
10000
5000
25000
Fig. 5. Variation of the hydrodynamic diameter Dh (nm) and scattered light intensity (counts s�1) of b-LG (20 g l�1) at pH 7.0 upon heating from 60 to
90 1C at 0.5 1Cmin�1 in the presence of cosolute. Hydrodynamic diameter: (A) arginine HCl, (B) NaCl and (C) GdnHCl. Scattered light intensity: (D)
arginine HCl, (E) NaCl and (F) GdnHCl. Arginine HCl concentration: K, 0mM; J, 20mM; ’, 60mM; &, 70mM; m, 80mM; n, 400mM. NaCl
concentration: K, 0mM; J, 20mM; ’, 100mM; &, 140mM; m, 150mM; n, 400mM. GdnHCl concentration: K, 0mM; J, 20mM; ’, 40mM; &,
50mM; m, 60mM.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191012
account. The DLS measurement is generally carried on thesoluble fraction of the solution. However, once weak gelstructures or large particles (46000 nm) are formed theaccuracy of the measurement is strongly influenced. Thiscan be seen in the serrated curve shapes (insets in Figs. 4and 5).
Fig. 6 allows distinguishing between the aggregationeffect (formation of soluble aggregates) and insolubilityfollowed by sedimentation. Samples containing arginineHCl were chosen here, but similar results were found forNaCl and GdnHCl at both acidic and neutral pH. Thecomparison of conditions (20mM arginine HCl, Dh,
90 1C ¼ 30 nm) where no sedimentation occurs at pH 4.0(Fig. 6A) and pH 7.0 (Fig. 6C) shows strong similarities.Depending on pH, temperature and cosolute concentra-tion, reactive b-LG particles were formed. Due to the heattreatment the protein unfolded leading to an increase insize. Reaching a critical denaturation point, proteinsstarted to aggregate with each other, resulting in a gradualdecrease of scatterers in the dispersion. Less particles weredetected even though they are larger in size, causing thedrop of the intensity of the autocorrelation function(Fig. 6A and C). This showed that the decrease inscattering intensity cannot be directly correlated with

ARTICLE IN PRESS
0.0
0.2
0.1
0.3
0.4
0.5
0.6
0.7
0.8
0.9
g 2 (
t)-1
90°C
90°C
85°C
85°C
80°C
80°C
75°C
75°C
70°C70°C
65°C65°C60°C
60°C25°C
90°C85°C
80°C
75°C
70°C65°C60°C25°C
90°C
85°C
80°C
75°C
70°C65°C60°C
25°C
25°C
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
10710-1 101 103 105
Time (µs) Time (µs)10710-1 101 103 105
(A)
(C)
(B)
(D)
g 2 (t
)-1
Fig. 6. Autocorrelation function as determined by dynamic light scattering (20 g l�1 b-LG, heating rate 0.5 1Cmin�1): (A) pH 4.0, 20mM arginine HCl,
(B) pH 4.0, 60mM arginine HCl, (C) pH 7.0, 20mM arginine HCl and (D) pH 7.0, 60mM arginine HCl.
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
0 20 40 60 80 100
Temperature (°C)
Initi
al in
tens
ity o
f g 2
(t) loss of scatterers
Fig. 7. Representation of the initial intensity of g2(t) derived from the
autocorrelation function shown in Fig. 6A–D as a function of the
temperature and the cosolute concentration (n) pH 4.0 60mM arginine
HCl and (m) pH 7.0 60mM arginine HCl. Lines are only guide for the
eyes.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1013
insolubility. Further, the autocorrelation function wasslightly shifted to higher relaxation times, indicating adecrease in mobility of the particles in both cases (Berne &Pecora, 1976; Horne & Davidson, 1993; Ten Grotenhuis,Paques, & van Aken, 2000).
In conditions where insolubility occurred (60mMarginine HCl), the differences in acidic and neutral pHconditions were significant (Fig. 6B, D and 7). AtpH 7.0, the increase in cosolute concentration did notchange the intensity of the autocorrelation function to agreat extent at low temperatures (25–70 1C) (Fig. 6D).However, at higher temperatures (475 1C), the majority ofthe signal was lost. This could be explained by the loss ofscatterers due to the formation of soluble aggregatesfollowed by insolubilisation and sedimentation. Thedecrease in the initial intensity of the autocorrelationfunction was lower for pH 4.0 than for pH 7.0 and shiftedto higher temperatures, which might be due to theconsiderably lower degree of aggregation and the highprotein stability in acidic conditions (Fig. 7). Additionally,the autocorrelation function shifted to higher relaxationtimes, which could be explained by a strong decreasein particle mobility, probably induced by aggregationand the formation of weak network-like structures(Ten Grotenhuis et al., 2000).

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191014
It has to be taken into account that the autocorrelationfunction and the derived particle size depend on theturbidity of the sample and as a consequence on theattenuation at which the measurement was performed. Dueto changes in the sample composition, formation ofaggregates or sedimentation the attenuation varies duringthe measurement (automatically adapted to sample turbid-ity by the apparatus). This means that the calculatedparticle size (Dh) and the autocorrelation function (Fig. 6)reflect properties specific to the appropriate measurementconditions, whereas scattering intensity values (Figs. 4 and5D–F) are corrected for the respective attenuation andconsider the average properties of the solution. As aconsequence of the correction step, the scattered intensitydid not prove to be the suitable parameter to link theappearance of exothermic peaks in DSC with the forma-tion of aggregates as it was detected by DLS. In contrast,the increase in Dh, (properties of the soluble fraction)showed good agreement with the appearance of theexothermic peaks and was chosen for the followingargumentation.
As described above, and as expected, aggregation inacidic pH conditions was different from aggregation inneutral pH conditions. Therefore, it was necessary toseparate these two pH conditions for further considera-tions on the mechanism of protein aggregation in thepresence of cosolutes and the appearance of exothermicpeaks.
3.3. Comparison of DSC and DLS results
3.3.1. Protein denaturation and aggregation at pH 4.0
Few studies have been carried out on b-LG aggregationat pH 4.0. Generally, aggregation in acidic conditions hasto be divided into conditions close to (low net charge of theprotein, positive charges o5) and far from the isoelectricpoint (protein strongly positively charged, positive charges410) (Fogolari et al., 2000). In the following discussion,pH 4.0 will be considered as far from the IEP. Between pH5.0 and 3.0 the proteins carboxyl groups are titrated, whichleads to a strong decrease in negative charges (Arakawa &Timasheff, 1987). Intermolecular repulsion forces domi-nate between the protein molecules, resulting in a lowtendency for aggregation for b-LG alone, as observed inFig. 4 (Basch & Timasheff, 1967; Schokker et al., 2000). Noexothermic peaks appeared for low cosolute concentrations(0–40mM). This could be due to the fact that theaggregates react one by one with activated monomers,which is in line with observations made by Aymard,Nicolai, and Durand (1999). These authors demonstratedthat in acidic conditions and low ionic strength, proteinaggregation follows a nucleation and growth mechanism,where b-LG monomers associate preferentially withaggregates. Due to large repulsion forces between themonomers, the nucleation process should be rate limited,but once a certain size is reached, the growth processproceeds relatively fast. Further, in acidic pH conditions,
aggregates are mainly due to physical aggregation and areformed by non-covalent bonds like electrostatic interac-tions and hydrogen bonding (Aymard et al., 1996a, 1999;Schokker et al., 2000). The expected release of exothermicenergy due to aggregation is therefore quite small. Athigher cosolute concentrations (460mM) exothermicpeaks (Texo) occurred below the actual protein denatura-tion peak (Td) for the three cosolutes. These peaks shiftedto lower temperatures with increasing cosolute concentra-tion. In general, increasing the ionic strength leads to twoopposite effects. First, the aggregation rate increases due tocharge shielding and a decrease of intermolecular repulsionforces. The second effect is the stabilisation of the proteinstructure against heat denaturation. Hence, the reductionof intramolecular repulsion forces increases the conforma-tional stability of the protein and the tendency of proteinunfolding is reduced, leading to a decrease of theaggregation rate (Boye, Ma, & Harwalkar, 1997; Boye,Ma, Ismail et al., 1997; Schokker et al., 2000). If these twoeffects are considered for b-LG heat aggregation in thepresence of the three cosolutes (Figs. 1 and 4) themechanism can be explained as follows.Since the aggregation exothermic peak was observed for
the same molar concentration for the three differentcosolutes (60mM), the likely explanation is that thechloride counter-ions are responsible for the neutralisationof the positive charges at the surface of the b-LG, leadingto aggregation. If arginine HCl and guanidinium HClthermograms were similar as far as the shift of exothermicpeak to lower temperature is concerned, a markeddifference was observed in the presence of NaCl. Hence,this cosolute induced a specific effect on b-LG asprecipitation and increase in turbidity was observed uponmixing b-LG and NaCl at concentrations higher than60mM at pH 4.0 and 20 1C. It can be concluded that thiscosolute induces a salting-out effect on the protein,enhancing protein aggregation. Arakawa and Timasheff(1987) reported similar specific effects of NaCl or glycineon b-LG. By contrast, lower cosolute concentrationsinduced a salting-in effect, reducing the tendency foraggregation. The likely explanation for this behaviour ofNaCl resides in the balance between the specific ionicbinding to the protein surface and its preferential hydrationby the solvent. Due to the low dissociation of thecarboxylic groups of the b-LG at pH 4.0, it is positivelycharged and exhibits a low dipolar moment. When lowamounts of NaCl are added (below 60mM in our case),insufficient number of chloride ions screens the positivecharges of the protein so that the preferential hydrationforces are prevalent. Upon heating, b-LG slightly dena-tures as traduced by the endothermic peak on DSC curves,but does not aggregate. When higher amounts of NaCl areconsidered, more chloride ions bind to the protein surfaceso that preferential hydration by water is overcome,leading to the observed salting-out (appearance of theexothermic aggregation peak). The specific effect of Na+
on the on the shift of the exothermic peak to low
ARTICLE IN PRESS
Temperature [°C]
Temperature [°C]
50 60 70 80 90 100 110
End
othe
rmic
hea
t flo
w
Tonset Texo
TonsetTexo
60 65 70 75 80 85 90
Hyd
rody
nam
ic d
iam
eter
[nm
]
0
100
200
300
400
(A)
(B)
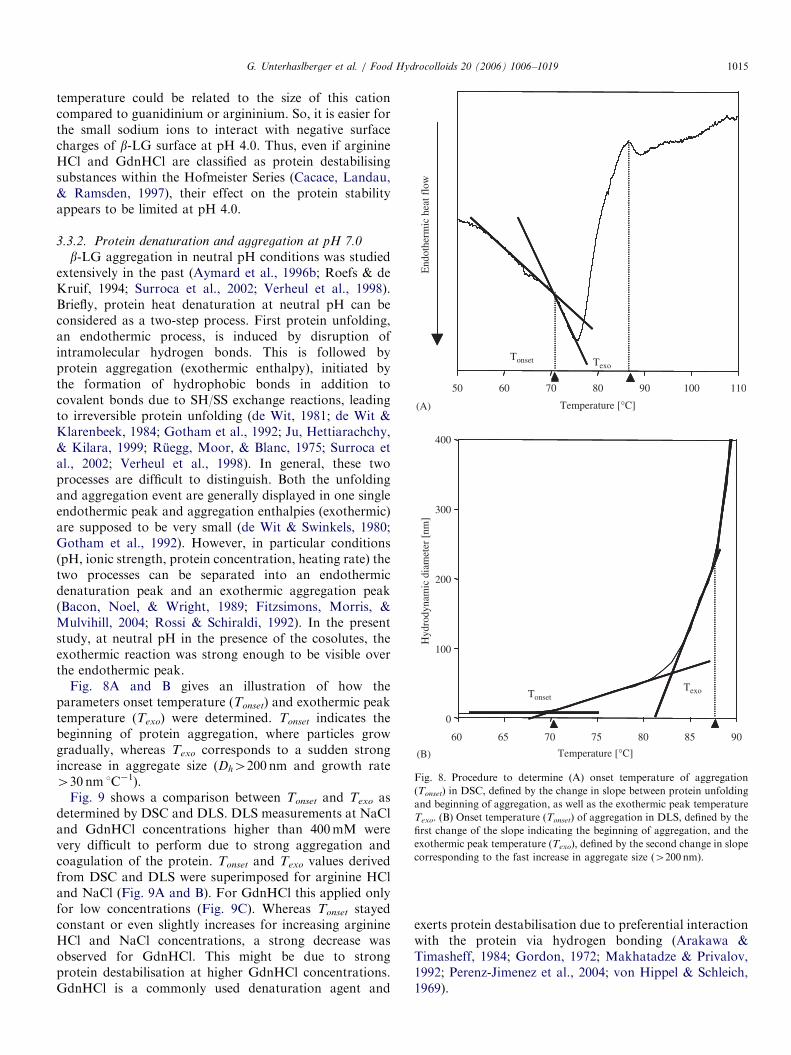
Fig. 8. Procedure to determine (A) onset temperature of aggregation
(Tonset) in DSC, defined by the change in slope between protein unfolding
and beginning of aggregation, as well as the exothermic peak temperature
Texo. (B) Onset temperature (Tonset) of aggregation in DLS, defined by the
first change of the slope indicating the beginning of aggregation, and the
exothermic peak temperature (Texo), defined by the second change in slope
corresponding to the fast increase in aggregate size (4200 nm).
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1015
temperature could be related to the size of this cationcompared to guanidinium or argininium. So, it is easier forthe small sodium ions to interact with negative surfacecharges of b-LG surface at pH 4.0. Thus, even if arginineHCl and GdnHCl are classified as protein destabilisingsubstances within the Hofmeister Series (Cacace, Landau,& Ramsden, 1997), their effect on the protein stabilityappears to be limited at pH 4.0.
3.3.2. Protein denaturation and aggregation at pH 7.0
b-LG aggregation in neutral pH conditions was studiedextensively in the past (Aymard et al., 1996b; Roefs & deKruif, 1994; Surroca et al., 2002; Verheul et al., 1998).Briefly, protein heat denaturation at neutral pH can beconsidered as a two-step process. First protein unfolding,an endothermic process, is induced by disruption ofintramolecular hydrogen bonds. This is followed byprotein aggregation (exothermic enthalpy), initiated bythe formation of hydrophobic bonds in addition tocovalent bonds due to SH/SS exchange reactions, leadingto irreversible protein unfolding (de Wit, 1981; de Wit &Klarenbeek, 1984; Gotham et al., 1992; Ju, Hettiarachchy,& Kilara, 1999; Ruegg, Moor, & Blanc, 1975; Surroca etal., 2002; Verheul et al., 1998). In general, these twoprocesses are difficult to distinguish. Both the unfoldingand aggregation event are generally displayed in one singleendothermic peak and aggregation enthalpies (exothermic)are supposed to be very small (de Wit & Swinkels, 1980;Gotham et al., 1992). However, in particular conditions(pH, ionic strength, protein concentration, heating rate) thetwo processes can be separated into an endothermicdenaturation peak and an exothermic aggregation peak(Bacon, Noel, & Wright, 1989; Fitzsimons, Morris, &Mulvihill, 2004; Rossi & Schiraldi, 1992). In the presentstudy, at neutral pH in the presence of the cosolutes, theexothermic reaction was strong enough to be visible overthe endothermic peak.
Fig. 8A and B gives an illustration of how theparameters onset temperature (Tonset) and exothermic peaktemperature (Texo) were determined. Tonset indicates thebeginning of protein aggregation, where particles growgradually, whereas Texo corresponds to a sudden strongincrease in aggregate size (Dh4200 nm and growth rate430 nm 1C�1).
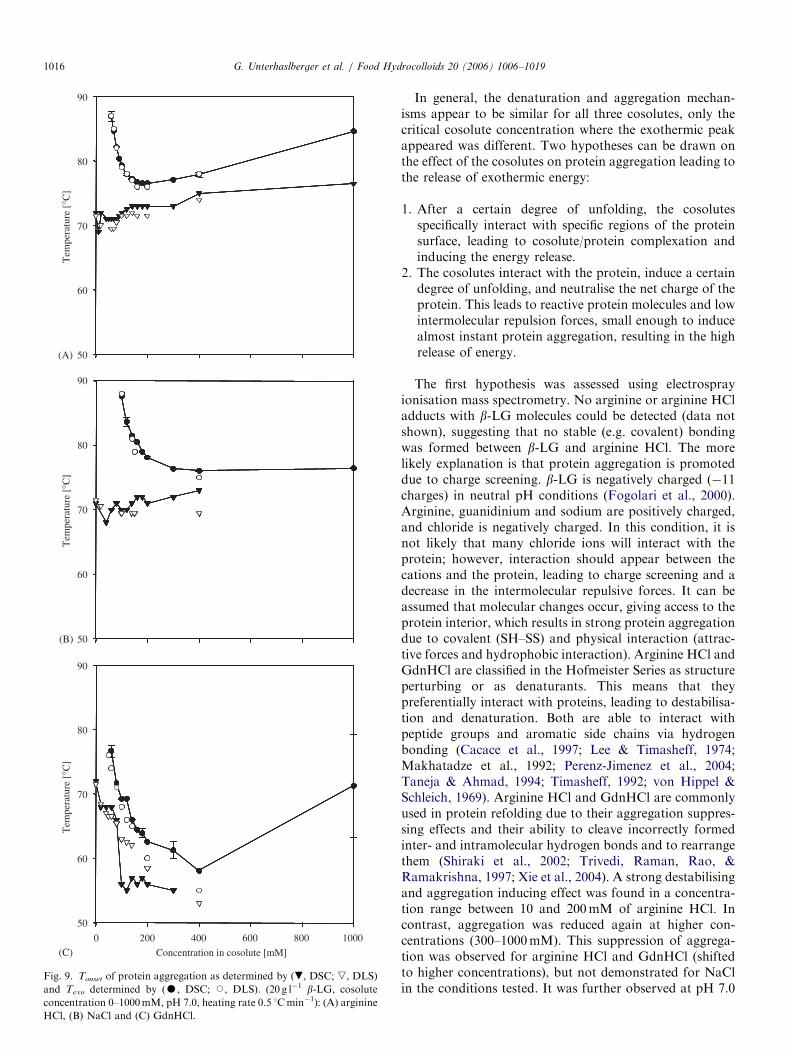
Fig. 9 shows a comparison between Tonset and Texo asdetermined by DSC and DLS. DLS measurements at NaCland GdnHCl concentrations higher than 400mM werevery difficult to perform due to strong aggregation andcoagulation of the protein. Tonset and Texo values derivedfrom DSC and DLS were superimposed for arginine HCland NaCl (Fig. 9A and B). For GdnHCl this applied onlyfor low concentrations (Fig. 9C). Whereas Tonset stayedconstant or even slightly increases for increasing arginineHCl and NaCl concentrations, a strong decrease wasobserved for GdnHCl. This might be due to strongprotein destabilisation at higher GdnHCl concentrations.GdnHCl is a commonly used denaturation agent and
exerts protein destabilisation due to preferential interactionwith the protein via hydrogen bonding (Arakawa &Timasheff, 1984; Gordon, 1972; Makhatadze & Privalov,1992; Perenz-Jimenez et al., 2004; von Hippel & Schleich,1969).
ARTICLE IN PRESST
empe
ratu
re [
°C]
70
80
90
Concentration in cosolute [mM]
(A)
(C)
(B)
50
60
Tem
pera
ture
[°C
]
70
80
90
50
60
Tem
pera
ture
[°C
]
70
80
90
50
60
0 200 400 600 800 1000
Fig. 9. Tonset of protein aggregation as determined by (., DSC; ,, DLS)
and Texo determined by (K, DSC; J, DLS). (20 g l�1 b-LG, cosolute
concentration 0–1000mM, pH 7.0, heating rate 0.5 1Cmin�1): (A) arginine
HCl, (B) NaCl and (C) GdnHCl.
G. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191016
In general, the denaturation and aggregation mechan-isms appear to be similar for all three cosolutes, only thecritical cosolute concentration where the exothermic peakappeared was different. Two hypotheses can be drawn onthe effect of the cosolutes on protein aggregation leading tothe release of exothermic energy:
1.
After a certain degree of unfolding, the cosolutesspecifically interact with specific regions of the proteinsurface, leading to cosolute/protein complexation andinducing the energy release.2.
The cosolutes interact with the protein, induce a certaindegree of unfolding, and neutralise the net charge of theprotein. This leads to reactive protein molecules and lowintermolecular repulsion forces, small enough to inducealmost instant protein aggregation, resulting in the highrelease of energy.The first hypothesis was assessed using electrosprayionisation mass spectrometry. No arginine or arginine HCladducts with b-LG molecules could be detected (data notshown), suggesting that no stable (e.g. covalent) bondingwas formed between b-LG and arginine HCl. The morelikely explanation is that protein aggregation is promoteddue to charge screening. b-LG is negatively charged (�11charges) in neutral pH conditions (Fogolari et al., 2000).Arginine, guanidinium and sodium are positively charged,and chloride is negatively charged. In this condition, it isnot likely that many chloride ions will interact with theprotein; however, interaction should appear between thecations and the protein, leading to charge screening and adecrease in the intermolecular repulsive forces. It can beassumed that molecular changes occur, giving access to theprotein interior, which results in strong protein aggregationdue to covalent (SH–SS) and physical interaction (attrac-tive forces and hydrophobic interaction). Arginine HCl andGdnHCl are classified in the Hofmeister Series as structureperturbing or as denaturants. This means that theypreferentially interact with proteins, leading to destabilisa-tion and denaturation. Both are able to interact withpeptide groups and aromatic side chains via hydrogenbonding (Cacace et al., 1997; Lee & Timasheff, 1974;Makhatadze et al., 1992; Perenz-Jimenez et al., 2004;Taneja & Ahmad, 1994; Timasheff, 1992; von Hippel &Schleich, 1969). Arginine HCl and GdnHCl are commonlyused in protein refolding due to their aggregation suppres-sing effects and their ability to cleave incorrectly formedinter- and intramolecular hydrogen bonds and to rearrangethem (Shiraki et al., 2002; Trivedi, Raman, Rao, &Ramakrishna, 1997; Xie et al., 2004). A strong destabilisingand aggregation inducing effect was found in a concentra-tion range between 10 and 200mM of arginine HCl. Incontrast, aggregation was reduced again at higher con-centrations (300–1000mM). This suppression of aggrega-tion was observed for arginine HCl and GdnHCl (shiftedto higher concentrations), but not demonstrated for NaClin the conditions tested. It was further observed at pH 7.0

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1017
that the presence of GdnHCl induced strongest aggrega-tion followed by arginine HCl and NaCl. This might beexplained by studies showing, that GdnHCl is moreefficient in charge screening than NaCl due to preferentialinteraction with the protein (Arakawa & Timasheff, 1984;Makhatadze & Privalov, 1992; Perenz-Jimenez et al., 2004).However, NaCl was shown to exert a salting-in effect on b-LG in neutral pH conditions, which strongly depends onthe cosolute concentration (Arakawa & Timasheff, 1987).This might explain the higher concentrations of NaClneeded for equivalent protein aggregation.
4. Conclusions
At pH 4.0, protein aggregation is based on physicalaggregation (electrostatic interactions) more than on proteinunfolding (hydrophobic and covalent interactions). Physicalaggregation is promoted by the presence of cosolutes due tocharge screening effects. The intermolecular repulsion forcesare decreased by negatively charged chloride ions, whichinteract with the positively charged protein (�8 positivecharges) leading to aggregation (Fogolari et al., 2000). Dueto the high stability of the protein regarding unfolding inacidic conditions no additional driving force for aggregationwas present (hydrophobic interactions). The effect ofarginine, guanidinium and sodium on protein destabilisationseems to be secondary in those conditions, explaining theonly small differences in aggregation between the cosolutes(NaCl4arginine HCl4GdnHCl).
At pH 7.0, protein aggregation is more complex, due tothe preliminary step of protein unfolding. The positivelycharged cations (arginine, guanidinium and Na) interactwith the negatively charged protein. Concentration effectsplay a strong role in the generally destabilising effects of allthree cosolutes on the native protein structure. However,arginine HCl and GdnHCl act at low concentrations bydestabilising or even denaturing the protein, leading tostrong aggregation. This effect is reversed at higherconcentrations, which results in aggregation suppression.NaCl shows less efficiency in charge screening of theprotein, protein destabilisation is reduced and higherconcentrations are needed to compensate for this effect.In neutral conditions, the efficiency of the cosolutes in theheat aggregation of b-LG can be described asGdnHCl4arginine HCl4NaCl.
The overall mechanism of b-LG heat denaturation oraggregation is not specifically changed by the presence ofarginine HCl, GdnHCl or NaCl at acidic or neutral pH.However, rate and intensity of protein aggregation isreinforced due to charge screening effects. The cosolutesexpress specific stabilising or destabilising effects onprotein stability following the Hofmeister Series. However,stabilising or destabilising mechanisms never exist sepa-rately. These mechanisms are strongly influenced by factorslike pH, ionic strength, solvent or cosolute affinity due toprotein surface charges (polar, non-polar, charged or non-charged), temperature and protein concentration.
Acknowledgements
Dr. M. Affolter from Nestle Research Center is thankedfor his assistance in carrying out the electrospray ionisationmass spectrometry. Fruitful discussion with Dr. L. Bovettoand Dr. P. Lambelet from Nestle Research Center isgratefully acknowledged.
References
Arakawa, T., Prestrelski, S. J., Kenney, W. C., & Carpenter, J. F. (2001).
Factors affecting short-term und long-term stabilities of proteins.
Advances in Drug Delivery Reviews, 46, 307–326.
Arakawa, T., & Timasheff, S. N. (1984). Protein stabilisation and
destabilisation by guanidinium salts. Biochemistry, 23, 5924–5929.
Arakawa, T., & Timasheff, S. N. (1987). Abnormal solubility behaviour of
b-lactoglobulin: Salting-in by glycine and NaCl. Biochemistry, 26,
5147–5153.
Aymard, P., Durand, D., & Nicolai, T. (1996a). A comparison of the
structure of b-lactoglobulin aggregates formed at pH 7.0 and pH 2.0.
International Journal of Polymer Analysis & Characterisation, 2,
115–119.
Aymard, P., Gimel, J.-C., Nicolai, T., & Durand, D. (1996b).
Experimental evidence for a two-step process in the aggregation of
b-lactoglobulin at pH 7. Journal de Chimie Physique, 93, 987–997.
Aymard, P., Nicolai, T., & Durand, D. (1999). Static and dynamic
scattering of b-lactoglobulin aggregates formed after heat-induced
denaturation at pH 2. Macromolecules, 32, 2542–2552.
Bacon, J. R., Noel, T. R., & Wright, D. J. (1989). Studies on the thermal
behaviour of pea (Pisum sativum) vicilin. Journal of Science in Food &
Agriculture, 49, 335–345.
Basch, J. J., & Timasheff, S. N. (1967). Hydrogen ion equilibria of the
genetic variants of bovine b-lactoglobulin. Archives of Biochemistry
and Biophysics, 118, 37–47.
Berne, B. J., & Pecora, R. (1976). Dynamic light scattering. New York:
Wiley.
Boye, J. I., Ma, C.-Y., & Harwalkar, V. R. (1997). Thermal denaturation
and coagulation of proteins. In S. Damodaran, & A. Paraf (Eds.),
Food proteins and their applications (pp. 25–56). New York: Marcel
Dekker.
Boye, J. I., Ma, C.-Y., Ismail, A., Harwalkar, V. R., & Kalab, M. (1997).
Molecular and microstructural studies of thermal denaturation and
gelation of b-lactoglobulins A and B. Journal of Agricultural and Food
Chemistry, 45, 1608–1618.
Brownlow, S., Cabral, J. H. M., Cooper, R., Flower, D. R., Yewdall, S. J.,
Polikarpov, I., et al. (1997). Bovine b-lactoglobulin at 1.8 A resolu-
tion—Still an enigmatic lipocalin. Structure, 5, 481–495.
Cacace, M. G., Landau, E. M., & Ramsden, J. J. (1997). The Hofmeister
Series: Salt and solvent effects on interfacial phenomena. Quarterly
Reviews of Biophysics, 30, 241–277.
Dannenberg, F., & Kessler, H.-G. (1988). Reaction kinetics of the
denaturation of whey proteins in milk. Journal of Food Science, 53,
258–263.
De Bernardez Clark, E. (2001). Protein refolding for industrial processes.
Current Opinion in Biotechnology, 12, 202–207.
De Wit, J. N. (1981). Structure and functional behaviour of whey proteins.
Netherlands Milk Dairy Journal, 35, 47–64.
De Wit, J. N., & Klarenbeek, G. (1981). A differential scanning
calorimetric study of the thermal behaviour of bovine b-lactoglobulinat temperatures up to 160 1C. Journal of Dairy Research, 48, 293–302.
De Wit, J. N., & Klarenbeek, G. (1984). Effects of various heat treatments
on structure and solubility of whey proteins. Journal of Dairy Science,
67, 2701–2710.
De Wit, J. N., & Swinkels, G. A. M. (1980). A differential scanning
calorimetric study of the thermal denaturation of bovine b-lactoglo-

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–10191018
bulin thermal behaviour at temperatures up to 100 1C. Biochimica et
Biophysica Acta, 624, 40–50.
Durand, D., Gimel, J.-C., & Nicolai, T. (2002). Aggregation, gelation and
phase separation of heat denatured globular proteins. Physica A, 304,
253–265.
Elofsson, U. M., Dejmek, P., & Paulsson, M. A. (1996). Heat-induced
aggregation of b-lactoglobulin studied by dynamic light scattering.
International Dairy Journal, 6, 343–357.
Fitzsimons, S. M., Morris, E. R., & Mulvihill, D. M. (2004). Resolution of
the denaturation and aggregation processes during whey protein
gelation by differential scanning calorimetry. Irish Journal of
Agricultural and Food Research, 43(1), 114.
Fogolari, F., Ragona, L., Licciardi, S., Romagnoli, S., Michelutti, R.,
Ugolini, R., et al. (2000). Electrostatic properties of bovine
b-lactoglobulin. Proteins: Structure, Function and Genetics, 39,
317–330.
Georgalis, Y., Umbach, P., Saenger, W., Ihmels, B., & Soumpasis, D. M.
(1999). Ordering of fractal clusters in crystallising lysozyme solutions.
Journal of the American Chemical Society, 121, 1627–1635.
Gimel, J.-C., Durand, D., & Nicolai, T. (1994). Structure and distribution
of aggregates formed after heat-induced denaturation of globular
proteins. Macromolecules, 27, 583–589.
Gordon, J. A. (1972). Denaturation of globular proteins. Interaction
of guanidinium salts with three proteins. Biochemistry, 11,
1862–1871.
Gotham, S. M., Fryer, P. J., & Pritchard, A. M. (1992). b-Lactoglobulindenaturation and aggregation reactions and fouling deposit formation:
A DSC study. International Journal of Food Science and Technology,
27, 313–327.
Griffin, W. G., & Griffin, M. C. A. (1993). Molecular basis of thermal
aggregation of bovine b-lactoglobulin A. Journal of the Chemical
Society—Faraday Transactions, 89, 3395–3406.
Hagihara, Y., Tan, Y., & Goto, Y. (1994). Comparison of the
conformational stability of the molten globule and native states of
horse cytochrome C. Effects of acetylation, heat, urea and guanidi-
ne–hydrochloride. Journal of Molecular Biology, 237, 336–348.
Hambling, S. G., McAlpine, A. S., & Sawyer, L. (1992). b-Lactoglobulin.In P. F. Fox (Ed.), Advanced dairy chemistry, proteins, Vol. 1 (pp.
141–190). London: Blackie Academic & Professional.
Hegg, P. O. (1980). Thermal stability of b-lactoglobulin as a function of
pH and the relative concentration of sodium dodecylsulphate. Acta
Agriculturae Scandinavica, 30, 401–404.
Hoffmann, M. (1997). b-Lactoglobulin: Denaturation and aggregation.
PhD dissertation, University of Utrecht, The Netherlands.
Horne, D. S., & Davidson, C. M. (1993). Application of diffusing-wave
spectroscopy to particle sizing in concentrated dispersions.
Colloids and Surfaces A: Physicochemical and Engineering Aspects,
77, 1–8.
Ju, Z. Y., Hettiarachchy, N., & Kilara, A. (1999). Thermal properties of
whey protein aggregates. Journal of Dairy Science, 82, 1882–1889.
Le Bon, C., Nicolai, T., & Durand, D. (1999). Growth and structure of
aggregates of heat-denatured b-lactoglobulin. International Journal of
Food Science and Technology, 34, 451–465.
Lee, J. C., & Timasheff, S. N. (1974). Partial specific volumes and
interactions with solvent components of proteins in guanidine
hydrochloride. Biochemistry, 13, 257–265.
Lin, M. Y., Lindsay, H. M., Weitz, D. A., Ball, R. C., Klein, R., &
Meakin, P. (1989). Universality of fractal aggregates as probed
by light scattering. Proceedings of the Royal Society of London A,
423, 71–87.
Liu, T., & Chu, B. (2002). Light-scattering of proteins. Encyclopedia of
Surface and Colloid Science, L, 3023–3043.
Makhatadze, G. I., & Privalov, P. L. (1992). Protein interactions with urea
and guanidinium chloride. A calorimetric study. Journal of Molecular
Biology, 226, 491–505.
Marshall, W. E., & Zarins, Z. M. (1989). Exothermic transitions of
glycinin determined by differential scanning calorimetry. Journal of
Agricultural and Food Chemistry, 37, 869–873.
McClements, D. J. (2002). Modulation of globular protein functionality
by weakly interacting cosolvents. Critical Reviews in Food Science and
Nutrition, 42, 417–471.
McKenzie, H. A. (1971). b-Lactoglobulins. In H. A. McKenzie (Ed.), Milk
proteins chemistry and molecular biology, Vol. 2 (pp. 257–330). New
York: Academic Press.
Mulvihill, D. M., & Donovan, M. (1987). Whey proteins and their thermal
denaturation—A review. Irish Journal of Food Science and Technology,
11, 43–75.
Nielsen, B. T., Singh, H., & Latham, J. M. (1996). Aggregation of bovine
b-lactoglobulins A and B on heating at 75 1C. International Dairy
Journal, 6, 519–527.
Panouille, M., Nicolai, T., & Durand, D. (2004). Heat induced
aggregation and gelation of casein submicelles. International Dairy
Journal, 14, 297–303.
Perenz-Jimenez, R., Godoy-Ruiz, R., Ibarra-Molero, B., & Sanchez-Ruiz,
J. M. (2004). The efficiency of different salts to screen charge
interactions in proteins: A Hofmeister effect? Biophysical Journal, 86,
2414–2429.
Pessen, H., Purcell, J. M., & Farrell, H. M., Jr. (1985). Proton relaxation
rates of water in dilute solutions of b-lactoglobulin. Determination of
cross relaxation and correlation with structural changes by the use of
two genetic variants of a self-associating globular protein. Biochimica
et Biophysica Acta, 828, 1–12.
Roefs, S. P. F. M., & de Kruif, C. G. (1994). A model for the denaturation
and aggregation of b-lactoglobulin. European Journal of Biochemistry,
226, 883–889.
Rossi, M., & Schiraldi, A. (1992). Thermal denaturation and aggregation
of egg proteins. Thermochimica Acta, 199, 115–123.
Ruegg, M., Moor, U., & Blanc, B. (1975). Hydration and thermal
denaturation of b-lactoglobulin. A calorimetric study. Biochimica et
Biophysica Acta, 400, 334–342.
Schokker, E. P., Singh, H., Pinder, D. N., & Creamer, L. K. (2000). Heat-
induced aggregation of b-lactoglobulin AB at pH 2.5 as influenced by
ionic strength and protein concentration. International Dairy Journal,
10, 233–240.
Shiraki, K., Kudou, M., Fujiwara, S., Imanaka, T., & Takagi, M. (2002).
Biophysical effect of amino acids on the prevention of protein
aggregation. Journal of Biochemistry, 132, 591–595.
Span, J., Lenarcic, S., & Lapanje, S. (1974). Solvation of lysozyme and b-lactoglobulin in aqueous guanidine hydrochloride solutions. Biochimi-
ca et Biophysica Acta, 359, 311–319.
Stepanek, P. (1993). Data analysis in dynamic light scattering. In W.
Brown (Ed.), Dynamic light scattering. The methods and some
applications (pp. 177–241). London: Clarendon Press.
Surroca, Y., Haverkamp, J., & Heck, A. J. R. (2002). Towards the
understanding of molecular mechanisms in the early stages of heat-
induced aggregation of b-lactoglobulin AB. Journal of Chromatogra-
phy A, 1, 1–11.
Taneja, S., & Ahmad, F. (1994). Increased thermal stability of proteins in
the presence of amino acids. Biochemical Journal, 303, 147–153.
Ten Grotenhuis, E., Paques, M., & van Aken, G. A. (2000). The
application of diffusing-wave spectroscopy to monitor the phase
behaviour of emulsion-polysaccharide systems. Journal of Colloid and
Interface Science, 227, 495–504.
Timasheff, S. N. (1992). A physicochemical basis for the selection of
osmolytes by nature. In G. N. Somero, C. B. Osmond, & C. L. Bolis
(Eds.), Water and life (pp. 70–84). Berlin: Springer.
Trivedi, V. D., Raman, B., Rao, Ch. M., & Ramakrishna, T. (1997). Co-
refolding denatured-reduced hen egg white lysozyme with acidic and
basic proteins. FEBS Letters, 418, 363–366.
Verheul, M., Roefs, S. P. F. M., & de Kruif, C. G. (1995). Heat-induced
denaturation and aggregation of b-lactoglobulin: Influence of sodium
chloride. In E. Dickinson, & D. Lorient (Eds.), Food macromolecules
and colloids (pp. 437–444). Cambridge: The Royal Society of Chemistry.
Verheul, M., Roefs, S. P. F., & de Kruif, C. G. (1998). Kinetics of heat-
induced aggregation of b-lactoglobulin. Journal of Agricultural and
Food Chemistry, 46, 896–903.

ARTICLE IN PRESSG. Unterhaslberger et al. / Food Hydrocolloids 20 (2006) 1006–1019 1019
Von Hippel, P. H., & Schleich, T. (1969). The effects of neutral salts on the
structure and conformational stability of macromolecules in solution. In S.
N. Timasheff, & G. D. Fasman (Eds.), Structure and stability of biological
macromolecules, Vol. 2 (pp. 417–574). New York: Marcel Dekker.
Watanabe, K., & Klostermeyer, H. (1976). Heat-induced changes in
sulphydryl and disulphide levels of b-lactoglobulin A and the
formation of polymers. Journal of Dairy Research, 43, 411–418.
Xie, Q., Guo, T., Lu, J., & Zhou, H.-M. (2004). The guanidine like
effects of arginine on aminoacylase and salt-induced molten globule
state. The International Journal of Biochemistry and Cell Biology, 36,
296–306.
Zhu, H., & Damodaran, S. (1994). Heat-induced conformational changes
in whey protein isolate and its relation to foaming properties. Journal
of Agricultural and Food Chemistry, 42, 846–855.

![Effect of protein aggregates on the complex coacervation between [beta]-lactoglobulin and acacia gum at pH 4.2](https://static.fdokumen.com/doc/165x107/6331c19f83bb92fe980414c8/effect-of-protein-aggregates-on-the-complex-coacervation-between-beta-lactoglobulin.jpg)


















