
Habitat Use and Life History as Predictors of Bird Responses to Habitat Change
12
Contributed Paper Habitat Use and Life History as Predictors of Bird Responses to Habitat Change NICOLA C. OKES, ∗ PHILIP A. R. HOCKEY, AND GRAEME S. CUMMING DST/NRF Centre of Excellence at the Percy FitzPatrick Institute, University of Cape Town, Rondebosch 7701, South Africa Abstract: In theory the consideration of life-history characteristics should provide a way of making predictive generalizations about the responses of different species to environmental modification. Nevertheless, few studies have tested the validity of this assumption or attempted to apply it across large numbers of related species. We explored both quantitative and qualitative contrasts between species of waterbirds that have either expanded or contracted their ranges in southern Africa over the past 40 years to test the hypothesis that expansionists and contractionists, respectively, should share life-history characteristics and/or ecological attributes. Similarities and differences in life history and ecology were explored through multivariate statistics. Overall, life-history traits provided an inadequate explanation of whether species would be range expansionists or contractionists. By contrast, ecological attributes of species that related to habitat use correlated well with range changes. In particular, waterbird species that inhabit pans seemed to be preadapted to using human- made dams and impoundments. The ability of many species to use artificial wetlands has aided their westward range expansions into arid regions of southern Africa. By contrast, species that rely on vegetated wetlands and that require reeds for nesting were predisposed to range contraction because their habitats have been severely affected by agricultural development and urbanization. In direct contrast to range expansions, most range contractions were west to east, the eastward contraction reflected the high level of wetland loss and degradation in the eastern lowlands of South Africa. Based on analysis of ecological attributes of regional contractionists, several additional species were identified as of potential conservation concern, although such concern may not as yet have been expressed. Keywords: life-history traits, range contraction, range expansion, waterbird conservation, wetland loss Uso de H´ abitat e Historia de Vida como Vaticinadores de las Respuestas de Aves al Cambio de H´ abitat Resumen: En teor´ ıa, la consideraci´ on de caracter´ ısticas de la historia de vida deber´ ıa proveer una v´ ıa para hacer generalizaciones vaticinadoras de las respuestas de diferentes especies a las modificaciones ambientales. Sin embargo, pocos estudios han probado la validez de esta suposici´ on o han intentado aplicarla en n´ umeros grandes de especies relacionadas. Exploramos contrastes cuantitativos y cualitativos entre especies de aves acu´ aticas que han expandido o contra´ ıdo sus rangos de distribuci´ on en el sur de ´ Africa a lo largo de los ´ ultimos 40 a˜ nos para probar la hip´ otesis de que expansionistas y contraccionistas deben compartir caracter´ ısticas de historia de vida y/o atributos ecol´ ogicos. Las similitudes y diferencias de historias de vida y ecolog´ ıa fueron exploradas mediante estad´ ıstica multivariada. En general, las caracter´ ısticas de la historia de vida no proporcionaron una explicaci´ on adecuada sobre s´ ı las especies ser´ ıan expansionistas o contraccionistas de rango de distribuci´ on. En contraste, los atributos ecol´ ogicos de las especies relacionados con el uso de h´ abitat se correlacionaron positivamente con los cambios en el rango de distribuci´ on. En particular, las especies de aves acu´ aticas que habitan en cuencas parecieron estar preadaptadas para utilizar diques y represas hechas por humanos. La habilidad de muchas especies para utilizar humedales ratif´ ıcales ha ayudado a su expansi´ on hacia regiones ´ aridas en el sur de ´ Africa. En contraste, las especies que dependen de humedales con vegetaci´ on y que requieren de juncos para anidar estuvieron predispuestas a la contracci´ on del rango de distribuci´ on debido a que sus h´ abitats han sido severamente afectados por el desarrollo agr´ ıcola y la urbanizaci´ on. ∗ email [email protected] Paper submitted January 23, 2007; revised manuscript accepted August 14, 2007. 151 Conservation Biology, Volume 22, No. 1, 151–162 C 2008 Society for Conservation Biology DOI: 10.1111/j.1523-1739.2007.00862.x
-
Upload
jamescookuniversity -
Category
Documents
-
view
6 -
download
0
Transcript of Habitat Use and Life History as Predictors of Bird Responses to Habitat Change

Contributed Paper
Habitat Use and Life History as Predictors of BirdResponses to Habitat ChangeNICOLA C. OKES,∗ PHILIP A. R. HOCKEY, AND GRAEME S. CUMMINGDST/NRF Centre of Excellence at the Percy FitzPatrick Institute, University of Cape Town, Rondebosch 7701, South Africa
Abstract: In theory the consideration of life-history characteristics should provide a way of making predictivegeneralizations about the responses of different species to environmental modification. Nevertheless, fewstudies have tested the validity of this assumption or attempted to apply it across large numbers of relatedspecies. We explored both quantitative and qualitative contrasts between species of waterbirds that haveeither expanded or contracted their ranges in southern Africa over the past 40 years to test the hypothesisthat expansionists and contractionists, respectively, should share life-history characteristics and/or ecologicalattributes. Similarities and differences in life history and ecology were explored through multivariate statistics.Overall, life-history traits provided an inadequate explanation of whether species would be range expansionistsor contractionists. By contrast, ecological attributes of species that related to habitat use correlated well withrange changes. In particular, waterbird species that inhabit pans seemed to be preadapted to using human-made dams and impoundments. The ability of many species to use artificial wetlands has aided their westwardrange expansions into arid regions of southern Africa. By contrast, species that rely on vegetated wetlandsand that require reeds for nesting were predisposed to range contraction because their habitats have beenseverely affected by agricultural development and urbanization. In direct contrast to range expansions, mostrange contractions were west to east, the eastward contraction reflected the high level of wetland loss anddegradation in the eastern lowlands of South Africa. Based on analysis of ecological attributes of regionalcontractionists, several additional species were identified as of potential conservation concern, although suchconcern may not as yet have been expressed.
Keywords: life-history traits, range contraction, range expansion, waterbird conservation, wetland loss
Uso de Habitat e Historia de Vida como Vaticinadores de las Respuestas de Aves al Cambio de Habitat
Resumen: En teorıa, la consideracion de caracterısticas de la historia de vida deberıa proveer una vıa parahacer generalizaciones vaticinadoras de las respuestas de diferentes especies a las modificaciones ambientales.Sin embargo, pocos estudios han probado la validez de esta suposicion o han intentado aplicarla en numerosgrandes de especies relacionadas. Exploramos contrastes cuantitativos y cualitativos entre especies de avesacuaticas que han expandido o contraıdo sus rangos de distribucion en el sur de Africa a lo largo de los ultimos40 anos para probar la hipotesis de que expansionistas y contraccionistas deben compartir caracterısticasde historia de vida y/o atributos ecologicos. Las similitudes y diferencias de historias de vida y ecologıafueron exploradas mediante estadıstica multivariada. En general, las caracterısticas de la historia de vida noproporcionaron una explicacion adecuada sobre sı las especies serıan expansionistas o contraccionistas derango de distribucion. En contraste, los atributos ecologicos de las especies relacionados con el uso de habitatse correlacionaron positivamente con los cambios en el rango de distribucion. En particular, las especies deaves acuaticas que habitan en cuencas parecieron estar preadaptadas para utilizar diques y represas hechaspor humanos. La habilidad de muchas especies para utilizar humedales ratifıcales ha ayudado a su expansionhacia regiones aridas en el sur de Africa. En contraste, las especies que dependen de humedales con vegetaciony que requieren de juncos para anidar estuvieron predispuestas a la contraccion del rango de distribuciondebido a que sus habitats han sido severamente afectados por el desarrollo agrıcola y la urbanizacion.
∗email [email protected] submitted January 23, 2007; revised manuscript accepted August 14, 2007.
151
Conservation Biology, Volume 22, No. 1, 151–162C©2008 Society for Conservation BiologyDOI: 10.1111/j.1523-1739.2007.00862.x

152 Bird Response to Habitat Change
En contraste directo con las expansiones de rango de distribucion, la mayorıa de las contracciones fueronde poniente a oriente, la contraccion hacia el oriente reflejo el alto nivel de perdida y degradacion dehumedales en las tierras bajas orientales de Africa del Sur. Con base en el analisis de atributos ecologicosde los contraccionistas regionales, varias especies adicionales fueron identificadas como potencial asunto depreocupacion para la conservacion, aunque tal preocupacion aun no ha sido expresada.
Palabras Clave: caracterısticas de la historia de vida, conservacion de aves acuaticas, contraccion de rango dedistribucion, expansion de rango de distribucion, perdida de humedales
Introduction
Conservation biology as a discipline has historicallyplaced a strong emphasis on understanding why ani-mal and plant populations decrease and on reversing de-creases. Although the causes of population and speciesloss have been studied widely, the problem of generaliza-tion remains a central scientific challenge. If conservationbiology is to avoid being a mere collection of case studiesand if it is ever to make accurate predictions about futurechanges in biodiversity, it is imperative that conservationbiologists develop a detailed, mechanistic understandingof why particular kinds of organism respond to anthro-pogenic habitat modification (and other influences) indifferent or similar ways.
One approach that has considerable potential as ameans of deriving conservation-relevant generalities is toconsider responses to habitat change based on a frame-work of life histories and ecological attributes across mul-tiple species of interest. If the responses of organisms todifferent influences can be predicted from what is knownabout their basic biology, one can in theory derive mecha-nistic relationships that will allow conservation biologiststo make reliable predictions about the effects of partic-ular habitat alterations on biodiversity and potentially toact in advance to reduce or mitigate threats to key popu-lations and species.
We tested the hypothesis that birds with similar life his-tories and/or ecological attributes respond in similar waysto environmental change. Because birds often respond toenvironmental change over large areas and long time pe-riods and because current trends in bird populations arean integrative response to a large number of environ-mental changes, we based our approach on a “naturalexperiment” rather than a manipulative one. We used adata set of bird distributions in southern Africa as a testcase. In the study area changes in the environment haveresulted in range expansions by some species of birdsand range contractions by others. Examples of expan-sionists include the Yellow-billed Oxpecker (Buphagusafricanus, Stutterheim & Brooke 1981), South AfricanCliff-Swallow (Hirundo spilodera, Rowan 1963), andSouthern Grey-headed Sparrow (Passer diffusus, Wardet al. 2004; Craig et al. 1987). Contractionists include theBlue Swallow (Hirundo atrocaerulea, Allan 1988), Red-
billed Oxpecker (Buphagus erythrorhynchus, Bezuiden-hout & Stutterheim 1980), Burchell’s Courser (Cursoriusrufus, Hockey 2005), Eurasian Bittern (Botaurus stel-laris, Dean 2005) and many raptors (Boshoff et al. 1983).Range expansions have been attributed mostly to theemergence of anthropogenic habitats, such as forestryplantations, which provide new habitat in previouslyunsuitable environments (Hockey 2003). Range contrac-tions, by contrast, are frequently attributed to habitat lossor degradation (e.g., Barnes 2000).
To narrow the range of possible habitats for consid-eration in the analysis, we focused on waterbirds. Thelocal extinction of waterbirds in southern Africa is of-ten a consequence of reclamation of wetland habitat foragricultural development (Hockey & Douie 1995). Theconstruction of impoundments on agricultural land hasfurther affected upstream and downstream habitats byinundating large areas and disrupting stream flow, re-sulting in the loss of some species (Hockey et al. 1989;Macdonald 1992). Nevertheless, together with salt andsewage works, the construction of farm dams and im-poundments has also created potential breeding and feed-ing sites (often in otherwise arid regions) into whichwaterbirds can move, and some species, including theBlack-winged Stilt (Himantopus himantopus) and Black-smith Lapwing (Vanellus armatus), have expanded theirranges over the last 50–100 years for this reason (Hockey& Douie 1995; Hockey 2003). Other species of water-birds that are thought to have benefited from anthro-pogenic habitat modification include the Hadeda Ibis(Bostrychia hagedash, Macdonald et al. 1986), GlossyIbis (Plegadis falcinellus, Hockey 2003), Goliath Heron(Ardea goliath, Harebottle & Harrison 2001), Sacred Ibis(Threskiornis aethiopicus), White-faced Duck (Dendro-cygna viduata), and Hottentot Teal (Anas hottentota,Harebottle & Harrison 2001).
If the life-history approach to generalization is useful,one would expect that range expansionists share life-history and/or ecological attributes that differ from thoseof contractionists. By the same token, the dissimilaritiesbetween range expansionists and range contractionistsshould shed light on the processes that drive changes inthe ranges of birds in southern Africa. These processesare of high interest to groups (such as SANBI, the SouthAfrican National Biodiversity Institute) that are actively
Conservation BiologyVolume 22, No. 1, 2008

Okes et al. 153
involved in conservation planning and policy develop-ment in South Africa. We tested these hypotheses forsouthern African waterbirds and concluded that the mainfactor separating contractionists and expansionists istheir ability to use human-made habitats in the form ofdams, impoundments, sewage works, and salt works.
Methods
Distribution Changes
A list of all waterbirds that breed in southern Africa wasobtained from Tarboton and Dennis (1993). We prelimi-narily classified species as potential expansionists (rangehas increased), contractionists (range has decreased), orstable (no change in range) by comparing distributionmaps for each species published in McLachlan and Liver-sidge (1978), Maclean (1993), and Hockey et al. (2005).The extent of the putative range change for each specieswas determined by overlaying distribution maps on aquarter degree (15 × 15′), equal-area projection grid mapof southern Africa. We expressed change over time aseither a positive or negative percentage. Nevertheless,using maps alone was deemed insufficient evidence foraccurately categorizing the nature of range changes orlack thereof. For some species, such as Cape Teal (A.capensis), the early distribution maps were clearly unre-liable, and these species were excluded from analyses apriori.
Because some early maps were identifiable as unreli-able, the potential existed for other maps to be in error(even though these errors were not obvious to us). There-fore, for example, a potential expansionist/contractionistto be classified as such in analyses (with a minimum rangechange of 5%), the range change had to be documentedin the primary literature. In six cases in which historicalmaps were deemed unreliable, range expansions wereso well documented in the literature that these specieswere included in the analyses (Table 1). Where appropri-ate, species with apparently stable ranges (as determinedfrom distribution maps) were further subclassified as lo-cal expansionists or contractionists on the basis of pub-lished evidence. A local contractionist was defined as aspecies that had been lost from a small part of its range(<5%) (e.g., local extinction along a river catchment ora small, but documented contraction at the peripheryof its range). A local expansionist was a species exhibit-ing the reverse pattern. These are therefore species po-tentially starting a regional contraction or expansion. Toidentify common patterns between species, we superim-posed maps on one another (separately for expansionistsand contractionists) with Arcview GIS 3.3 (ESRI 2002).We compared the resultant composite maps (for SouthAfrica alone) with the dispersion of natural and artificial
Table 1. Classification of species of waterbirds according to change intheir regional ranges and the percentage of range lost or gained insouthern Africa between 1978 and 2005 for contractionists andexpansionists.
RangeScientific name change (%)
Range expansionistsFulvous Duck Dendrocygna bicolor 9White-faced Duck∗ D. viduataMaccoa Duck Oxyura maccoa 16Egyptian Goose Alopochen aegyptiaca 15South African Shelduck Tadorna cana 12Spur-winged Goose Plectropterus gambensis 37Cape Shoveler∗ Anas smithiiHottentot Teal A. hottentota 11Common Moorhen Gallinula chloropusAfrican Jacana Actophilornis africanus 28Black-winged Stilt∗ Himantopus himantopusKittlitz’s Plover∗ Charadrius pecuariusBlacksmith Lapwing∗ Vanellus armatusGrey-headed Gull Larus cirrocephalus 27African Fish-Eagle Haliaeetus vocifer 17Goliath Heron Ardea goliath 21Black-crowned Nycticorax nycticorax 11
Night-HeronLittle Bittern Ixobrychus minutus 26Dwarf Bittern I. sturmii 10Glossy Ibis Plegadis falcinellus 19African Sacred Ibis Threskiornis aethiopicus 12African Spoonbill Platalea alba 20Woolly-necked Stork Ciconia episcopus 11African Pied Wagtail Motacilla aguimp 12
Range contractionistsHalf-collared Kingfisher Alcedo semitorquata −20African Grass-Owl Tyto capensis −18Pel’s Fishing-Owl Scotopelia peli −5Grey-crowned Crane Balearica regulorum −14Wattled Crane Bugeranus carunculatus −31African Finfoot Podica senegalensis −9Red-chested Flufftail Sarothrura rufa −17Baillons Crake Porzana pusilla −23Allen’s Gallinule Porphyrio alleni −32Lesser Jacana Microparra capensis −24Collared Pratincole Glareola pratincola −10Rufous-bellied Heron Ardeola rufiventris −5African Marsh-Harrier Circus ranivorus −19Eurasian Bittern Botaurus stellaris −9Yellow-billed Stork Mycteria ibis −13
Population decreasePied Kingfisher Nettapus auritusAfrican Pygmy-Goose Ceryle rudisAfrican Purple Porphyrio
Swamphen madagascariensisAfrican Skimmer Rynchops flavirostrisGreater Flamingo Phoenicopterus ruber
Population increaseBlack Heron Egretta ardesiacaWhiskered Tern Chlidonias hybridaBlack-necked Grebe Podiceps nigricollis
Stable populationWhite-backed Duck Thalassornis leuconotusComb Duck Sarkidiornis melanotosAfrican Black Duck A. sparsaYellow-billed Duck A. undulata
continued
Conservation BiologyVolume 22, No. 1, 2008

154 Bird Response to Habitat Change
Table 1. (continued)
RangeScientific name change (%)
Range expansionistsGiant Kingfisher Megaceryle maximusBlack Crake Amaurornis flavirostrisLesser Moorhen Gallinula angulataAfrican Wattled Vanellus senegallus
LapwingCaspian Tern Sterna caspiaLittle Grebe Tachybaptus ruficollisGreat Crested Grebe Podiceps cristatusWhite-breasted Phalacrocorax lucidus
CormorantSlaty Egret Egretta vinaceigulaYellow-billed Egret E. intermediaGreat Egret E. albaSquacco Heron Ardeola ralloidesGreen-backed Heron Butorides striataLesser Flamingo Phoenicopterus minorPink-backed Pelican Pelecanus rufescensAfrican Openbill Anastomus lamelligerusBlack Stork Ciconia nigraBrown-throated Martin Riparia paludicolaLittle Rush-Warbler Bradypterus baboecalaRufous-winged Cisticola Cisticola galactotesLevaillant’s Cisticola C. tinniensMountain Wagtail Motacilla clara
∗Species included in analyses based on >5% range increases thatare well documented in the literature but are not reflected incomparisons of distribution maps.
wetlands and patterns of wetland loss and degradation(Cowan 1995).
Ecological and Life-History Traits
Life-history and ecological traits of the 86 species of wa-terbirds for which reliable distribution maps could beobtained were extracted from the literature (Harrisonet al. 1997; Underhill et al. 1999; Hockey et al. 2005).We created two separate, binary matrices, one for life-history traits and one for ecological traits. The life-historytraits we examined included developmental mode (al-tricial, precocial), body mass, clutch size, nest disper-sion (solitary, colonial), residency (extensive vs. limitedmovement), and diet (herbivorous, carnivorous, and in-sectivorous). Extensive movements encompass migrationor long-distance (in a regional context) dispersal or no-madism. The classification of limited movement was ac-corded to essentially resident species for which no ev-idence (e.g., band recoveries) exists to suggest move-ment of individuals over long distances. We compiled thisinformation as part of a database that accompanies theregion’s definitive bird handbook (Hockey et al. 2005).The ecological traits related primarily to habitat use andincluded habitat types used by birds (breeds in modifiedhabitat; feeds in modified habitat; vegetated wetlands andmarshes, rivers, floodplains, grassland, and pans) and nest
sites (reeds, water, ground, and tree). Data were recordedin binary format as presence or absence. Although somevariables could have been treated as continuous (e.g.,body mass), we recorded all data in binary form so wecould include all variables in the same analyses. There-fore, species could occur in more than one category. Forexample, a species could use pans, marshes, and human-modified habitats. Both life-history and habitat matricesare available (see Supplementary Material).
Statistical Analyses
We analyzed the data with ANOSIM, multidimensionalscaling (MDS), and cluster analysis in the statistical soft-ware package Primer 6 (Clarke & Warwick 1994). Specieswere placed in groups a priori as expansionists, con-tractionists, or species with stable ranges. An analysisof similarity (ANOSIM) was performed on each matrixseparately to determine whether there was a significantdifference between the predefined groupings based firston life-history traits alone and then on ecological traits.If there was a significant difference between groups,we performed a cluster analysis with Bray–Curtis simi-larity and group-average sorting, which groups speciestogether based on how similar they are to one another.To determine which species grouped together on thebasis of either their life-history or ecological traits, theclusters were complemented by a nonlinear MDS thatprovided a three-dimensional visual of the separation ofgroups (MDS, Clarke & Warwick 1994).
We determined the source of the differences amonggroups with a SIMPER test, which compares the contri-bution of each variable to the similarity and dissimilaritywithin and between groups. For SIMPER, groups can bedefined a priori or a posteriori on the basis of the clusteranalysis. The analysis was initially performed with groupsdefined a priori. The variables that contributed most tothe observed differences between species were then se-lected, and cluster analysis, MDS, and SIMPER analysiswere subsequently performed with only these variablesand in a posteriori groupings based on the cluster analy-sis. This process makes it possible to isolate the variablesthat distinguish the groups and to determine their relativeimportance in grouping particular species together.
Results
The species we analyzed came from 22 families. Eachfamily was represented by up to 6 species, with threeexceptions: Anatidae (10 species), Ardeidae (14), andRallidae (9). There were 24 regional expansionists, 18regional contractionists, 28 stable species, 8 local ex-pansionists, and 8 local contractionists (Table 1). Six-teen species were excluded from the analyses be-cause historical maps were deemed unreliable (Table 1;
Conservation BiologyVolume 22, No. 1, 2008
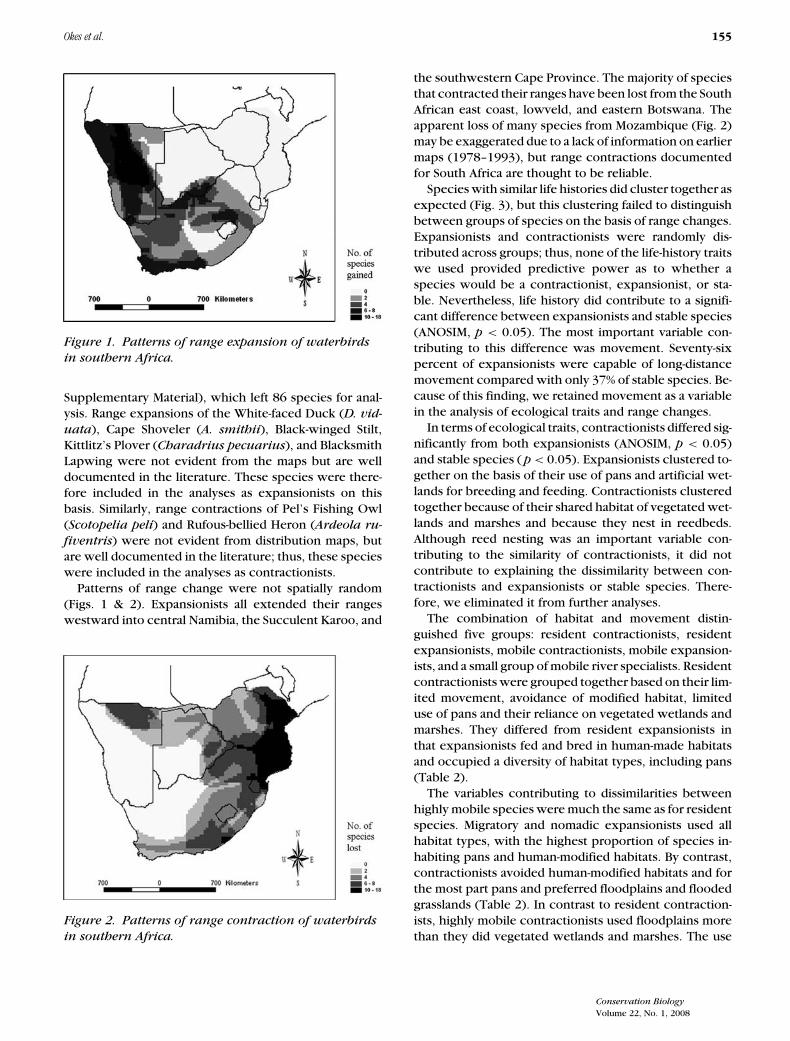
Okes et al. 155
Figure 1. Patterns of range expansion of waterbirdsin southern Africa.
Supplementary Material), which left 86 species for anal-ysis. Range expansions of the White-faced Duck (D. vid-uata), Cape Shoveler (A. smithii), Black-winged Stilt,Kittlitz’s Plover (Charadrius pecuarius), and BlacksmithLapwing were not evident from the maps but are welldocumented in the literature. These species were there-fore included in the analyses as expansionists on thisbasis. Similarly, range contractions of Pel’s Fishing Owl(Scotopelia peli) and Rufous-bellied Heron (Ardeola ru-fiventris) were not evident from distribution maps, butare well documented in the literature; thus, these specieswere included in the analyses as contractionists.
Patterns of range change were not spatially random(Figs. 1 & 2). Expansionists all extended their rangeswestward into central Namibia, the Succulent Karoo, and
Figure 2. Patterns of range contraction of waterbirdsin southern Africa.
the southwestern Cape Province. The majority of speciesthat contracted their ranges have been lost from the SouthAfrican east coast, lowveld, and eastern Botswana. Theapparent loss of many species from Mozambique (Fig. 2)may be exaggerated due to a lack of information on earliermaps (1978–1993), but range contractions documentedfor South Africa are thought to be reliable.
Species with similar life histories did cluster together asexpected (Fig. 3), but this clustering failed to distinguishbetween groups of species on the basis of range changes.Expansionists and contractionists were randomly dis-tributed across groups; thus, none of the life-history traitswe used provided predictive power as to whether aspecies would be a contractionist, expansionist, or sta-ble. Nevertheless, life history did contribute to a signifi-cant difference between expansionists and stable species(ANOSIM, p < 0.05). The most important variable con-tributing to this difference was movement. Seventy-sixpercent of expansionists were capable of long-distancemovement compared with only 37% of stable species. Be-cause of this finding, we retained movement as a variablein the analysis of ecological traits and range changes.
In terms of ecological traits, contractionists differed sig-nificantly from both expansionists (ANOSIM, p < 0.05)and stable species ( p < 0.05). Expansionists clustered to-gether on the basis of their use of pans and artificial wet-lands for breeding and feeding. Contractionists clusteredtogether because of their shared habitat of vegetated wet-lands and marshes and because they nest in reedbeds.Although reed nesting was an important variable con-tributing to the similarity of contractionists, it did notcontribute to explaining the dissimilarity between con-tractionists and expansionists or stable species. There-fore, we eliminated it from further analyses.
The combination of habitat and movement distin-guished five groups: resident contractionists, residentexpansionists, mobile contractionists, mobile expansion-ists, and a small group of mobile river specialists. Residentcontractionists were grouped together based on their lim-ited movement, avoidance of modified habitat, limiteduse of pans and their reliance on vegetated wetlands andmarshes. They differed from resident expansionists inthat expansionists fed and bred in human-made habitatsand occupied a diversity of habitat types, including pans(Table 2).
The variables contributing to dissimilarities betweenhighly mobile species were much the same as for residentspecies. Migratory and nomadic expansionists used allhabitat types, with the highest proportion of species in-habiting pans and human-modified habitats. By contrast,contractionists avoided human-modified habitats and forthe most part pans and preferred floodplains and floodedgrasslands (Table 2). In contrast to resident contraction-ists, highly mobile contractionists used floodplains morethan they did vegetated wetlands and marshes. The use
Conservation BiologyVolume 22, No. 1, 2008

156 Bird Response to Habitat Change
Figure 3. Cluster analysis of southern African waterbirds on the basis of life-history characteristics (exp,expansionist; con, contractionist; stable, no change in range).
Conservation BiologyVolume 22, No. 1, 2008
Okes et al. 157
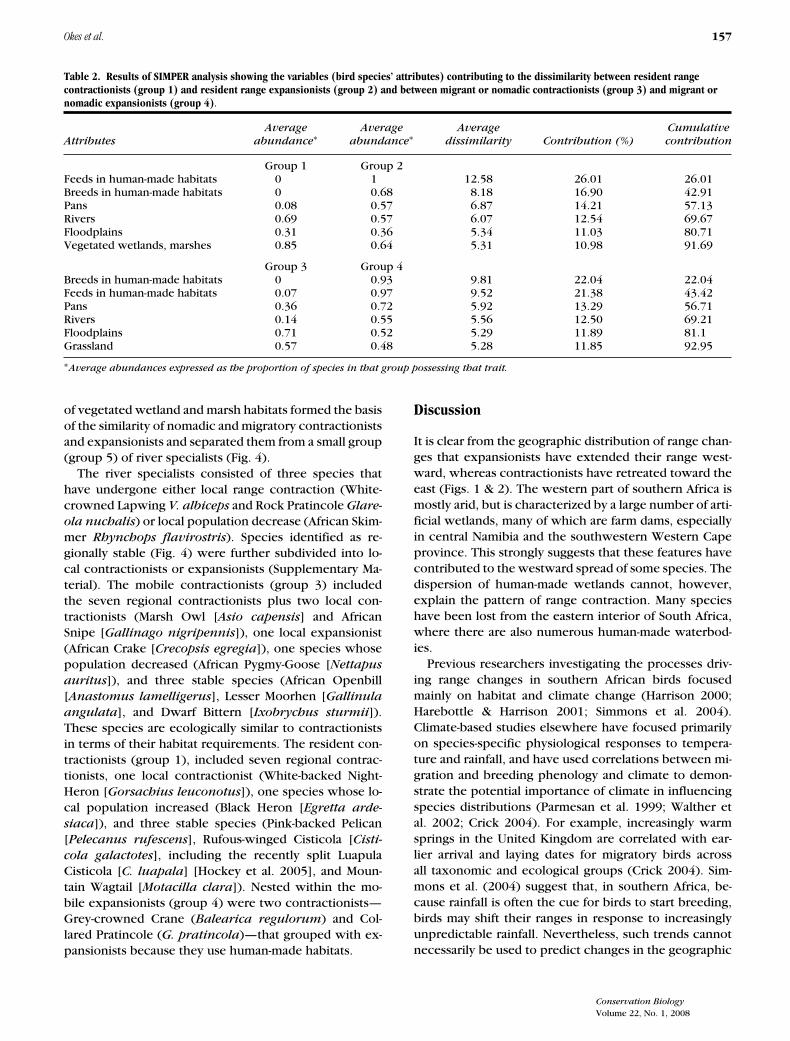
Table 2. Results of SIMPER analysis showing the variables (bird species’ attributes) contributing to the dissimilarity between resident rangecontractionists (group 1) and resident range expansionists (group 2) and between migrant or nomadic contractionists (group 3) and migrant ornomadic expansionists (group 4).
Average Average Average CumulativeAttributes abundance∗ abundance∗ dissimilarity Contribution (%) contribution
Group 1 Group 2Feeds in human-made habitats 0 1 12.58 26.01 26.01Breeds in human-made habitats 0 0.68 8.18 16.90 42.91Pans 0.08 0.57 6.87 14.21 57.13Rivers 0.69 0.57 6.07 12.54 69.67Floodplains 0.31 0.36 5.34 11.03 80.71Vegetated wetlands, marshes 0.85 0.64 5.31 10.98 91.69
Group 3 Group 4Breeds in human-made habitats 0 0.93 9.81 22.04 22.04Feeds in human-made habitats 0.07 0.97 9.52 21.38 43.42Pans 0.36 0.72 5.92 13.29 56.71Rivers 0.14 0.55 5.56 12.50 69.21Floodplains 0.71 0.52 5.29 11.89 81.1Grassland 0.57 0.48 5.28 11.85 92.95
∗Average abundances expressed as the proportion of species in that group possessing that trait.
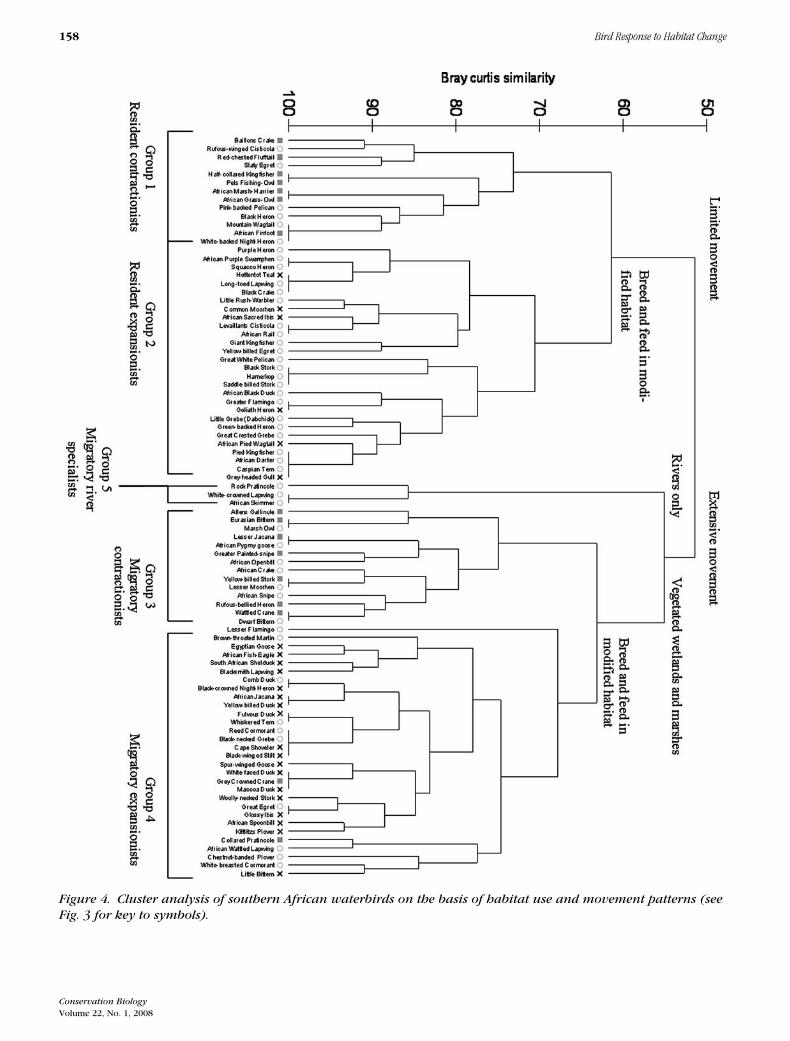
of vegetated wetland and marsh habitats formed the basisof the similarity of nomadic and migratory contractionistsand expansionists and separated them from a small group(group 5) of river specialists (Fig. 4).
The river specialists consisted of three species thathave undergone either local range contraction (White-crowned Lapwing V. albiceps and Rock Pratincole Glare-ola nuchalis) or local population decrease (African Skim-mer Rhynchops flavirostris). Species identified as re-gionally stable (Fig. 4) were further subdivided into lo-cal contractionists or expansionists (Supplementary Ma-terial). The mobile contractionists (group 3) includedthe seven regional contractionists plus two local con-tractionists (Marsh Owl [Asio capensis] and AfricanSnipe [Gallinago nigripennis]), one local expansionist(African Crake [Crecopsis egregia]), one species whosepopulation decreased (African Pygmy-Goose [Nettapusauritus]), and three stable species (African Openbill[Anastomus lamelligerus], Lesser Moorhen [Gallinulaangulata], and Dwarf Bittern [Ixobrychus sturmii]).These species are ecologically similar to contractionistsin terms of their habitat requirements. The resident con-tractionists (group 1), included seven regional contrac-tionists, one local contractionist (White-backed Night-Heron [Gorsachius leuconotus]), one species whose lo-cal population increased (Black Heron [Egretta arde-siaca]), and three stable species (Pink-backed Pelican[Pelecanus rufescens], Rufous-winged Cisticola [Cisti-cola galactotes], including the recently split LuapulaCisticola [C. luapala] [Hockey et al. 2005], and Moun-tain Wagtail [Motacilla clara]). Nested within the mo-bile expansionists (group 4) were two contractionists—Grey-crowned Crane (Balearica regulorum) and Col-lared Pratincole (G. pratincola)—that grouped with ex-pansionists because they use human-made habitats.
Discussion
It is clear from the geographic distribution of range chan-ges that expansionists have extended their range west-ward, whereas contractionists have retreated toward theeast (Figs. 1 & 2). The western part of southern Africa ismostly arid, but is characterized by a large number of arti-ficial wetlands, many of which are farm dams, especiallyin central Namibia and the southwestern Western Capeprovince. This strongly suggests that these features havecontributed to the westward spread of some species. Thedispersion of human-made wetlands cannot, however,explain the pattern of range contraction. Many specieshave been lost from the eastern interior of South Africa,where there are also numerous human-made waterbod-ies.
Previous researchers investigating the processes driv-ing range changes in southern African birds focusedmainly on habitat and climate change (Harrison 2000;Harebottle & Harrison 2001; Simmons et al. 2004).Climate-based studies elsewhere have focused primarilyon species-specific physiological responses to tempera-ture and rainfall, and have used correlations between mi-gration and breeding phenology and climate to demon-strate the potential importance of climate in influencingspecies distributions (Parmesan et al. 1999; Walther etal. 2002; Crick 2004). For example, increasingly warmsprings in the United Kingdom are correlated with ear-lier arrival and laying dates for migratory birds acrossall taxonomic and ecological groups (Crick 2004). Sim-mons et al. (2004) suggest that, in southern Africa, be-cause rainfall is often the cue for birds to start breeding,birds may shift their ranges in response to increasinglyunpredictable rainfall. Nevertheless, such trends cannotnecessarily be used to predict changes in the geographic
Conservation BiologyVolume 22, No. 1, 2008
158 Bird Response to Habitat Change
Figure 4. Cluster analysis of southern African waterbirds on the basis of habitat use and movement patterns (seeFig. 3 for key to symbols).
Conservation BiologyVolume 22, No. 1, 2008

Okes et al. 159
distribution of a species without an accurate assessmentof the behavior and habitat requirements of the species inquestion (Parmesan et al. 1999; Crick 2004). Factors thatmight influence whether a species can change its range inresponse to rainfall include habitat suitability (e.g., avail-ability and fragmentation of habitat and ability to live indisturbed or modified habitats) and the structure of theecological community (e.g., predator–prey relationshipsand competition for nest sites).
In southern Africa there is some evidence that indige-nous birds with characteristics similar to those of success-ful invaders have increased in range and abundance in re-sponse to environmental change (Macdonald 1992; Dean2000). For example, the Pied Crow (Corvus albus), a gre-garious, adaptable omnivore, uses telephone poles, wind-mill platforms, and trees as nest sites and has increasedin abundance and range (Dean 2000). The South AfricanCliff-Swallow, Speckled Pigeon (Columba guinea), andRed-winged Starling (Onychognathus morio) are addi-tional examples of species that habitually use human-made structures for nest sites: all three species have un-dergone marked increases in range and abundance withinthe last century (Dean 2000).
The hypothesis that certain life-history traits predis-pose waterbirds to range change was not supported byour results despite the large number of traits included.Analyses based on life-history traits did not result in anyconsistent groupings of species that had undergone sim-ilar range changes. By contrast, analysis of ecologicaltraits highlighted the importance of habitat use, and, forcontractionists, the importance of reedbed nest sites.Nevertheless, expansionists occupy a variety of nest sites,including reedbeds, and therefore are not consistentlydistinguishable from contractionists on this basis.
The main factor separating contractionists and expan-sionists was the use by expansionists of human-madehabitats in the form of impoundments, sewage works,and salt works. Expansionists use human-made habitatshabitually, whereas contractionists avoid them. The gainof artificial habitat in the form of impoundments (for ex-ample) therefore does not benefit species that never orrarely use them. This trend is reflected in the distribu-tion of contractionists (Fig. 2) that have been lost fromthe South African east coast and lowveld despite the largenumber of impoundments in this area. It is far more likelythat contractionists are responding to a loss or degrada-tion of natural wetlands.
Although the species we examined inhabit natural wet-lands, the natural habitats of expansionists and contrac-tionists differed. The natural habitats of resident con-tractionists are rivers, vegetated wetlands and marshes,and, to a lesser degree, floodplains. Migratory and no-madic contractionists favored floodplains and floodedgrassland. Neither made extensive use of pans, a habitatfrequented by both resident and migratory expansionists.Besides pans and dams, expansionists also made use of
all other habitat types including vegetated wetlands andmarshes, rivers, floodplains, and grassland. The fact thatthe use of pans is the key variable distinguishing expan-sionists and contractionists suggests that expansionistsare preadapted to using impoundments, which providethe same open-water habitat as pans. Species that relyon vegetated wetlands, nest in reeds, and avoid pans andhuman-made wetlands are the most vulnerable to rangecontraction.
The geographical pattern of natural wetland loss showsthat the greatest loss of wetlands is in the east (see Cowan1995) and is remarkably well correlated with the distri-bution of waterbird range contraction (Fig. 2). Fairbankset al. (2002) provide further support for this argumentby demonstrating that the most land transformation—through commercial and subsistence agriculture, de-graded vegetation, and soil erosion—has occurred in thenortheastern parts of South Africa. Anthropogenic ratherthan climate-mediated habitat change therefore appearsto be the most influential factor driving both range expan-sion and contraction. The proliferation of human-madewetland habitats is facilitating the westward range expan-sion of many waterbirds, whereas the loss of vegetatedwetlands is driving considerable range contraction in theeastern half of southern Africa.
One reviewer suggested we compare our results topredictions generated by a null model of habitat change.In our case, however, it makes no ecological sense todevelop a null model as these should generally be aimedat testing mechanisms. In this study the expansion of ar-tificial wetlands in western South Africa is indisputable,as is the current use of these wetlands by waterbirds.Common sense is adequate to tell us that without thechange in habitat, no range expansion would have oc-curred because without water, there would be no water-birds. The question is not whether waterbirds use thesenew habitats, but which waterbirds use them and why.A null model based on the assumptions we made wouldhave to make a decision about whether or not wetlandbirds could hypothetically exist in the arid western areasof South Africa without human influences. We do notbelieve we need a null model in order to make that deci-sion. Our existing analyses are sufficient to conclude thatthere are distinct, nonrandom patterns in terms of whichspecies have expanded their ranges to take advantage ofthe new habitats.
Conservation Implications
The wetlands that are most vulnerable to degradation andloss in southern Africa include vegetated wetlands andmarshes, rivers, and floodplains, especially those with ri-parian vegetation (Barnes 2000). The degradation of vege-tated wetlands and marshes is usually the result of damconstruction, erosion, and drainage and conversion toagriculture (Cowan & van Riet 1998). These negative
Conservation BiologyVolume 22, No. 1, 2008

160 Bird Response to Habitat Change
effects may be exacerbated by a predicted decrease inregional rainfall of up to 10% (30% in the arid west) dur-ing the 21st Century (de Wit & Stankiewicz 2006). Forexample, the Eurasian Bittern owes its critically endan-gered status in South Africa (Barnes 2000) to its need fortall, dense emergent vegetation. It has undergone signif-icant range contraction over much of South Africa andis locally extinct in several areas where extensive wet-land drainage has occurred (Keil 1938; Barnes 2000; Fair-banks et al. 2002). Another critically endangered speciesin South Africa, the Wattled Crane (Bugeranus caruncu-latus) (Barnes 2000), relies on permanent wetlands withshort emergent vegetation surrounded by grasslands forbreeding (Hockey et al. 2005). An estimated 50% lossof wetlands from the KwaZulu-Natal province midlands(the species’ main stronghold) through damming (Bento2002), drainage, and the conversion of grassland to agri-culture has undoubtedly contributed to the decrease inrange and abundance of this species (Johnson & Barnes1991).
Anthropogenic threats to rivers and floodplains includecatchment degradation and the associated increase in sed-iment load, pollution, water abstraction, and regulationby impoundments (Davies et al. 1998). Species relyingon rivers have suffered as a result. For example, the Half-collared Kingfisher (Alcedo semitorquata), African Fin-foot (Podica senegalensis), and Pel’s Fishing-Owl are allrange contractionists classified as being either vulnerableor near threatened (Barnes 2000). They rely on rivers withriparian vegetation, prefer clear water, and have been lostto some extent from the east coast of South Africa andsouthern Mozambique. Riparian deforestation, whetherthrough the removal of alien trees, destruction of man-groves, or inundation caused by damming, results in in-creased riverine silt loads (Harrison et al. 1997). Siltationleads to the eutrophication of rivers, which facilitates al-gal growth that in turn affects the food chain (Harrison etal. 1997). The overall effect is a loss of overhanging veg-etation for nesting and roosting, and a reduction in clear,fast-flowing water—a trend most severe in the EasternCape and KwaZulu-Natal provinces (Harrison et al. 1997).
Two species, the Grey-crowned Crane (Balearica reg-ulorum) and Collared Pratincole (G. pratincola), haveundergone historical range contractions yet grouped inanalyses with expansionists. Although the Grey CrownedCrane inhabits and breeds in marshes, requiring fairly tallemergent vegetation (Morris 1987), it also breeds at damsand feeds in agricultural land, with 56% of the SouthAfrican population occupying cultivated lands (Filmer& Haultshausen 1992). Despite its vulnerable classifica-tion (Barnes 2000) and well-documented range reduc-tions (Vernon et al. 1992; Morrison 1998; Oschadleus2000), there are reports of apparent population increasesin Zimbabwe and the Free State province of South Africa.These expansions are thought to be in response to cropfarming (Geldenhuys 1984). Similarly, the Collared Pratin-
cole is near threatened (Barnes 2000) in South Africa andhas undergone range contraction due to developmentand urbanization (Maclean 1973). Its use of large damshas facilitated recent local expansion in Eastern Botswana(Brewster 2000) and Zimbabwe (Tree 1987). These exam-ples highlight the importance of the use of human-madehabitats in facilitating range expansions.
Some species appear not to have experienced regionalrange contractions, yet grouped with contractionistsbased on similar habitat requirements. Some of thesespecies have undergone local contractions or populationdecreases, and need to be monitored if future range con-tractions are to be prevented. These include the MarshOwl, African Snipe, African Pygmy-Goose, White-backedNight-Heron, and the nomadic/migratory river specialists(White-crowned Lapwing, African Skimmer, and RockPratincole). Other species that may require monitoring,based on the fact that they grouped with contractionists,include the African Openbill, Lesser Moorhen, Dwarf Bit-tern, Pink-backed Pelican, Rufous-winged Cisticola, andMountain Wagtail. The recent inclusion of the AfricanOpenbill in the regional red data book as near threat-ened (Barnes 2000) highlights the very real possibilitythat some of these species may be facing similar as yetundetected problems. The conservation of natural, vege-tated wetlands should, therefore, remain a top conserva-tion priority for waterbirds.
Our results emphasize the importance of biological andbehavioral data in giving an improved representation ofhow anthropogenic changes affect biodiversity. Recentstudies assessing the impact of anthropogenic changeson habitat in southern Africa (Fairbanks et al. 2002) andNorth America (La Sorte 2006) have focused on speciesrichness, diversity, and evenness. Nevertheless, the lossof rare species may be countered by a gain in commonspecies, which results in a net maintenance of speciesrichness at the expense of species composition (La Sorte2006). In South Africa species richness and diversity ofbirds is highest in the northeastern regions (Fairbankset al. 2002), yet this is the very area where the major-ity of range contraction has occurred. Measures such asspecies diversity, richness, and evenness may be attrac-tive to ecologists because of their mathematical tractabil-ity, but they provide little information about the structureof communities and have no capacity to identify whichspecies within these assemblages might be predisposedto local or regional extinction.
If conservation biologists are to be able to offer pre-dictions about imminent conservation problems at thespecies level, the approach followed here should bevaluable in many different contexts because it allowsthe identification of pattern while providing strong in-ferences about process. With further refinement andmore widespread application, characterization of poten-tial range contractionists should facilitate a shift from con-servation that is merely responsive to a more predictive,
Conservation BiologyVolume 22, No. 1, 2008

Okes et al. 161
proactive approach in which potential species declinesare responded to before the population has attained anendangered status.
Acknowledgments
We are grateful to colleagues at the Percy FitzPatrickInstitute for their inputs to this research through variousdiscussions and debates. J. Houlahan, C. Elphick, and athird anonymous referee provided valuable commentson an earlier manuscript. This work was supported bythe University of Cape Town and the DST/NRF Centre ofExcellence at the Percy FitzPatrick Institute.
Supplementary Material
Justification for the classification of species as contrac-tionists and expansionists (Appendix S1), classificationof species (Appendix S2), scientific names of species in-cluded in analyses (Appendix S3), the binary matrices ofraw data (Appendices S4 and S5), and literature cited inthe appendices (Appendix S6) are available as part of theon-line article from http://www.blackwell-synergy.com/.The author is responsible for the content and function-ality of these materials. Queries (other than absence ofthe material) should be directed to the correspondingauthor.
Literature Cited
Allan, D. G. 1988. The Blue Swallow in with a chance. Quagga 22:5–7.Barnes, K. N. 2000. The Eskom red data book of birds of South Africa,
Lesotho and Swaziland. Birdlife South Africa, Johannesburg.Bento, C. M. 2002. The status and prospects of Wattled Cranes Grus
carunculatus in the Marromeu complex of the Zambezi Delta. MSthesis. University of Cape Town, Cape Town, South Africa.
Bezuidenhout, J. D., and C. J. Stutterheim. 1980. A critical evalua-tion of the role played by the Red-billed Oxpecker Buphagus ery-throrhynchus in the biological control of ticks. Onderstepoort Jour-nal of Veterinary Research 47:51–75.
Boshoff, A. F., C. J. Vernon, and R. K. Brooke. 1983. Historical atlas ofdiurnal raptors (Aves: Falconiformes). Annals of the Cape ProvincialMuseums (Natural History) 14:173–297.
Brewster, C. A. 2000. Breeding of Collared (Red-winged) PratincolesGlareola pratincola at Shashe Dam. Babbler (Botswana) 36:21.
Clarke, K. R., and R. M. Warwick. 1994. Change in marine communi-ties: an approach to statistical analysis and interpretation. PlymouthMarine Laboratory, Plymouth, United Kingdom.
Cowan, G. I., editor. 1995. Wetlands of South Africa. Department ofEnvironmental Affairs and Tourism, Pretoria, South Africa.
Cowan, G. I., and W. van Riet. 1998. A directory of South African wet-lands. Department of Environmental Affairs and Tourism, Pretoria,South Africa.
Craig, A., B. Every, and D. Summers-Smith. 1987. The spread of theSouthern Greyheaded Sparrow in the Cape Province. Annals of theCape Provincial Museums (Natural History) 16:191–200.
Crick, H. Q. P. 2004. The impact of climate change on birds. Ibis 146(supplement 1):48–56.
Davies, B. R., J. H. O’Keefe, and C. D. Snaddon. 1998. A synthesis ofthe ecological functioning, conservation and management of SouthAfrican river ecosystems. Water Research Commission. Page 44 in
G. I. Cowan and W. van Riet, editors. Report TT 62/93, Pretoria. Adirectory of South African wetlands. Department of EnvironmentalAffairs and Tourism, Pretoria, South Africa.
Dean, W. R. J. 2000. Alien birds in southern Africa: what factors deter-mine success? South African Journal of Science 96:9–14.
Dean, W. R. J. 2005. Eurasian Bittern. Pages 602–603 in P. A. R. Hockey,W. R. J. Dean, and P. G. Ryan, editors. Roberts–birds of SouthernAfrica. 7th edition. The Trustees of the John Voelcker Bird BookFund, Cape Town, South Africa.
De Wit, M., and J. Stankiewicz. 2006. Changes in surface water supplyacross Africa with predicted climate change. Science 311:1917–1920.
Fairbanks, D. H. K., M. Kshatriya, A. S. van Jaarsveld, and L. G. Underhill.2002. Scales and consequences of human land transformation onSouth African avian diversity and structure. Animal Conservation5:61–73.
Filmer, R. J., and G. R. Haultshausen. 1992. The South African cranecensus, 1985/1986. Pages 132–153 in D. G. Porter, H. S. Craven,D. N. Johnson, and M. J. Porter, editors. Proceedings of the 1stSouth African Crane Conference. South African Crane Foundation,Durban, South Africa.
Geldenhuys, J. N. 1984. The status of cranes (Aves: Gruidae) in theOrange Free State. South African Journal of Wildlife Research 14:15–18.
Harebottle, D. M., and J. A. Harrison. 2001. Changes in distribution andabundance of waterbirds in South Africa. Bird Numbers 10:1–7.
Harrison, J. A. 2000. Changing suburban birds—terrestrial species. BirdNumbers 9:1–7.
Harrison, J. A., D. G. Allan, L. G. Underhill, S. M. Herremans, A. J.Tree, V. Parker, and C. J. Brown, editors. 1997. The atlas of South-ern African birds. Volume 1. Non-passerines. Birdlife South Africa,Johannesburg, South Africa.
Hockey, P. A. R. 2003. Land invasions: are indigenous expansionistscause for concern? Africa Birds and Birding 8:35–41
Hockey, P. A. R. 2005. Burchell’s courser. Pages 424–426 in P. A. R.Hockey, W. R. J. Dean, and P. G. Ryan, editors. Roberts–birds ofSouthern Africa. 7th edition. The Trustees of the John Voelcker BirdBook Fund, Cape Town, South Africa.
Hockey, P. A. R., and C. Douie. 1995. Waders of Southern Africa. StruikWinchester, Cape Town.
Hockey, P. A. R., L. G. Underhill, M. Neatherway, and P. G. Ryan. 1989.Atlas of the birds of the southwestern Cape. Cape Bird Club, CapeTown, South Africa.
Hockey, P. A. R., W. R. J. Dean, and P. G. Ryan. 2005. Roberts–birdsof Southern Africa. 7th edition. The Trustees of the John VoelckerBird Book Fund, Cape Town, South Africa.
Johnson, D. N., and P. R. Barnes. 1991. The breeding biology of theWattled Crane in Natal. Pages 377–386 in J. T. Harris, editor. Pro-ceedings of the 1987 International Crane Workshop. InternationalCrane Foundation, Baraboo, Wisconsin.
Keil, S. F. 1938. The Cape Bittern. Ostrich 9:87–92.La Sorte, F. 2006. Geographical expansion and increased prevalence
of common species in avian assemblages: implications for large-scale patterns of species richness. Journal of Biogeography 33:1183–1191.
Macdonald, I. A. W. 1992. Vertebrate populations as indicators of en-vironmental change in southern Africa. Transactions of the RoyalSociety of South Africa 481:87–123.
Macdonald, I. A. W., D. M. Richardson, and F. J. Powrie. 1986. Range ex-pansion of the hadeda ibis Bostrychia hagedash in southern Africa.South African Journal of Zoology 21:331–342.
McLachlan, G. R., and R. Liversidge. 1978. Roberts birds of South Africa.4th edition. The Trustees of the John Voelcker Bird Book Fund, CapeTown, South Africa.
Maclean, G. L. 1973. Red-winged pratincoles nesting in Natal. Bok-makierie 25:61–63.
Conservation BiologyVolume 22, No. 1, 2008

162 Bird Response to Habitat Change
Maclean, G. L. 1993. Roberts’ birds of Southern Africa. 7th edition. TheTrustees of the John Voelcker Bird Book Fund, Cape Town, SouthAfrica.
Morris, A. 1987. The status of cranes in Zimbabwe. Pages 345–348in G. W. Archibald and R. F. Pasquier, editors. Proceedings of the1983 International Crane Worshop. International Crane Foundation,Baraboo, Wisconsin.
Morrison, K. L. 1998. Habitat utilisation and the population ecologyof cranes in the Dullstroom area of the Mpumalanga province. MSthesis. University of Pretoria, Pretoria, South Africa.
Oschadleus, H. D. 2000. Report on recoveries received at SAFRING:July 1999– June 2000. Safring News 29:93–99.
Parmesan, C., et al. 1999. Poleward shifts in geographic ranges of butter-fly species associated with regional warming. Nature 399:579–583.
Rowan, M. K. 1963. Range of the Cliff Swallow. Ostrich 34:181–182.Simmons, R. E., P. Barnard, W. R. J. Dean, G. F. Midgley, W. Thuiller,
and G. Hughes. 2004. Climate change and birds: the perspectivesand prospects from southern Africa. Ostrich 754:295–306.
Stutterheim, C. J., and R. K. Brooke. 1981. Past and present ecologicaldistribution of the Yellow-billed Oxpecker in South Africa. SouthAfrican Journal of Zoology 16:44–49.
Tarboton, W. R., and N. Dennis. 1993. Waterbirds: birds of southernAfrica’s wetlands. Struik, Cape Town, South Africa.
Tree, A. J. 1987. Redwinged Pratincoles breeding at Darwendale Dam.Honeyguide 33:15–17.
Underhill, L. G., A. J. Tree, H. D. Oschadleus, and V. Parker. 1999.Review of ring recoveries of waterbirds in southern Africa. AvianDemography Unit, University of Cape Town, Cape Town, SouthAfrica.
Vernon, C. J., A. F. Boshoff, and W. S. Stretton. 1992. The sta-tus and conservation of cranes in the Eastern Cape Province.Pages 47–58 in D. J. Porter, H. S. Craven, D. N. Johnson,and M. J. Porter, editors. Proceedings of the 1st South AfricanCrane Conference. South African Crane Foundation, Durban, SouthAfrica.
Walther, G., E. Post, P. Convey, A. Menzel, C. Parmesan, T. J. C. Bee-bee, J. M. Fromentin, O. Hoegh-Guldberg, and F. Bairlein. 2002.Ecological responses to recent climate change. Nature 416:389–395.
Ward, V. L., H. D. Oschadleus, and L. Upfold. 2004. Southern Grey-headed Sparrows Passer diffusus range expansion in the WesternCape. Afring News 33:18–20.
Conservation BiologyVolume 22, No. 1, 2008




















