
Gynostegium Morphology of Mesechiteae Miers (Apocynaceae, Apocynoideae) as It Pertains to the...
14
GYNOSTEGIUM MORPHOLOGY OF MESECHITEAE MIERS (APOCYNACEAE, APOCYNOIDEAE) AS IT PERTAINS TO THE CLASSIFICATION OF THE TRIBE Andre ´ Olmos Simo ˜ es, 1, * Maria Carolina Scatolin do Rio,* Marı ´lia de Moraes Castro, y and Luiza Sumiko Kinoshita y *Programa de Po ´ s-Graduac ¸a ˜o em Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, Cep 13083-970, Campinas, Sa ˜o Paulo, Brazil; and y Departamento de Bota ˆnica, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109, Cep 13083-970, Campinas, Sa ˜o Paulo, Brazil In order to investigate morphological patterns in Mesechiteae and test the new circumscription of the tribe, the gynostegium structure of nine species belonging to Forsteronia, Mandevilla, and Mesechites, with two species from Secondatia as outgroup, were comparatively studied. Our results support the most recent circum- scription of Mesechiteae, including Forsteronia, Mandevilla, and Mesechites and excluding Secondatia. The gynostegia of Forsteronia, Mandevilla, and Mesechites have the same basic organization: a proliferation of parenchyma cells in the style head forms five projecting ribs that are adnate to an expanded portion of the connective below the thecae, here named the ‘‘staminal retinacle.’’ This adnation is very tenuous in Forsteronia (few cells of the staminal retinacle are effectively connected to the style head) but much stronger in Mandevilla and Mesechites (several cells of the staminal retinacle are connected to the style head). The observed differences in the size of the style head ribs and the degree of adnation to the stamens also support the three major clades recognized within the tribe. The gynostegium of Secondatia has a different organization, of the type reported for representatives of the tribe Apocyneae: the style head has no projecting ribs, and its epidermal cells are interdigitated with, but not adnate to, unicellular trichomes of the staminal retinacle. Keywords: Apocynaceae, anatomy, Forsteronia, gynostegium, Mandevilla, Mesechiteae, Mesechites, Secondatia, style head. Introduction Apocynaceae s.l. is one of the largest angiosperm families, with ca. 335 genera and 3700 species (Judd et al. 2002). The family is characterized by a complex flower structure, with morphological and functional specialization of its parts (Woodson 1933; Ezcurra 1981; Fallen 1986; Rosatti 1989; Nicholas and Baijnath 1994; Endress and Bruyns 2000; Simo ˜ es and Kinoshita 2002). A major evolutionary trend in Apocynaceae is the progressive synorganization of gynoecium and androecium, which are postgenitally united to different de- grees to form a single functional unit, the gynostegium (Endress and Bruyns 2000) (fig. 1A). Within the family, the term ‘‘gyno- stegium’’ was originally used only in the former Asclepiadaceae and Periplocaceae (corresponding to subfamilies Asclepiadoideae, Secamonoideae, and Periplocoideae sensu Endress and Bruyns 2000), but Fallen (1986) expanded the use of this term to the subfamily Apocynoideae. She argued that even though post- genital fusion in Apocynoideae is less complete than in the former Asclepiadaceae, the basic design and function of the reproductive parts are the same in both groups. We agree with Fallen (1986), and the term ‘‘gynostegium’’ is used in this ar- ticle hereafter. The structure of the gynostegium has traditionally been used as a key character in the taxonomy of Apocynoideae (Woodson 1933, 1935, 1936; Pichon 1950; Ezcurra 1981; Fallen 1986; Simo ˜ es and Kinoshita 2002) and was the basis of the tribal classification proposed by Endress and Bruyns (2000). In contrast to its great relevance in systematics, few studies focused on anatomical aspects of the gynostegium have been made (Woodson and Moore 1938; Rao and Ganguli 1963; Allorge 1976; Schick 1980, 1982; Fallen 1986; Galetto 1997). Description of the morphology of a structure as complex as the gynostegium requires a well-defined set of terms. How- ever, a profuse terminology has been used by many authors over time to describe and characterize the gynostegium of Apocynaceae. One of the clearest examples of this are the terms used to describe the stamens of Apocynoideae, which are highly specialized organs composed of a combination of sterile and fertile tissues. Woodson (1933) used the term ‘‘pel- tate connective’’ for the dorsal, sclerified part of the stamens that extends below the thecae to form the two side wings. Pichon (1948) considered this tissue to be a specialization of the anthers and proposed the term ‘‘retinacle’’ to denote the sterile, nonsclerified portion of the stamens on the ventral sur- face of the connective that is adjacent or attached to the style head, forming the gynostegium (fig. 1B,1C). The name ‘‘reti- nacle,’’ however, has been widely used in Orchidaceae to refer to the upper portion of the translator arm of the polli- naria since its first use by Darwin (1895). Robertson (1887) adopted the same term to describe an analogous structure of 1 Author for correspondence; current address: Universidade de Sa ˜ o Paulo, Escola de Artes, Cie ˆncias e Humanidades (EACH), Avenida Arlindo Bettio 1000, Ermelino Matarazzo, Cep 03828-000, Sa ˜o Paulo, Brazil; e-mail [email protected]. Manuscript received May 2006; revised manuscript received April 2007. 999 Int. J. Plant Sci. 168(7):999–1012. 2007. Ó 2007 by The University of Chicago. All rights reserved. 1058-5893/2007/16807-0003$15.00
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Gynostegium Morphology of Mesechiteae Miers (Apocynaceae, Apocynoideae) as It Pertains to the...

GYNOSTEGIUM MORPHOLOGY OF MESECHITEAE MIERS (APOCYNACEAE,APOCYNOIDEAE) AS IT PERTAINS TO THE CLASSIFICATION OF THE TRIBE
Andre Olmos Simoes,1,* Maria Carolina Scatolin do Rio,* Marılia de Moraes Castro,y and Luiza Sumiko Kinoshitay
*Programa de Pos-Graduacao em Biologia Vegetal, Instituto de Biologia, Universidade Estadual de Campinas, Caixa Postal 6109,Cep 13083-970, Campinas, Sao Paulo, Brazil; and yDepartamento de Botanica, Instituto de Biologia, Universidade
Estadual de Campinas, Caixa Postal 6109, Cep 13083-970, Campinas, Sao Paulo, Brazil
In order to investigate morphological patterns in Mesechiteae and test the new circumscription of the tribe,the gynostegium structure of nine species belonging to Forsteronia, Mandevilla, and Mesechites, with twospecies from Secondatia as outgroup, were comparatively studied. Our results support the most recent circum-scription of Mesechiteae, including Forsteronia, Mandevilla, and Mesechites and excluding Secondatia. Thegynostegia of Forsteronia, Mandevilla, and Mesechites have the same basic organization: a proliferation ofparenchyma cells in the style head forms five projecting ribs that are adnate to an expanded portion of theconnective below the thecae, here named the ‘‘staminal retinacle.’’ This adnation is very tenuous in Forsteronia(few cells of the staminal retinacle are effectively connected to the style head) but much stronger in MandevillaandMesechites (several cells of the staminal retinacle are connected to the style head). The observed differencesin the size of the style head ribs and the degree of adnation to the stamens also support the three major cladesrecognized within the tribe. The gynostegium of Secondatia has a different organization, of the type reportedfor representatives of the tribe Apocyneae: the style head has no projecting ribs, and its epidermal cells areinterdigitated with, but not adnate to, unicellular trichomes of the staminal retinacle.
Keywords: Apocynaceae, anatomy, Forsteronia, gynostegium, Mandevilla, Mesechiteae, Mesechites,Secondatia, style head.
Introduction
Apocynaceae s.l. is one of the largest angiosperm families,with ca. 335 genera and 3700 species (Judd et al. 2002). Thefamily is characterized by a complex flower structure, withmorphological and functional specialization of its parts(Woodson 1933; Ezcurra 1981; Fallen 1986; Rosatti 1989;Nicholas and Baijnath 1994; Endress and Bruyns 2000;Simoes and Kinoshita 2002). A major evolutionary trend inApocynaceae is the progressive synorganization of gynoeciumand androecium, which are postgenitally united to different de-grees to form a single functional unit, the gynostegium (Endressand Bruyns 2000) (fig. 1A). Within the family, the term ‘‘gyno-stegium’’ was originally used only in the former Asclepiadaceaeand Periplocaceae (corresponding to subfamilies Asclepiadoideae,Secamonoideae, and Periplocoideae sensu Endress and Bruyns2000), but Fallen (1986) expanded the use of this term to thesubfamily Apocynoideae. She argued that even though post-genital fusion in Apocynoideae is less complete than in theformer Asclepiadaceae, the basic design and function of thereproductive parts are the same in both groups. We agree withFallen (1986), and the term ‘‘gynostegium’’ is used in this ar-ticle hereafter.
The structure of the gynostegium has traditionally beenused as a key character in the taxonomy of Apocynoideae(Woodson 1933, 1935, 1936; Pichon 1950; Ezcurra 1981;Fallen 1986; Simoes and Kinoshita 2002) and was the basisof the tribal classification proposed by Endress and Bruyns(2000). In contrast to its great relevance in systematics, fewstudies focused on anatomical aspects of the gynostegiumhave been made (Woodson and Moore 1938; Rao and Ganguli1963; Allorge 1976; Schick 1980, 1982; Fallen 1986; Galetto1997).Description of the morphology of a structure as complex
as the gynostegium requires a well-defined set of terms. How-ever, a profuse terminology has been used by many authorsover time to describe and characterize the gynostegium ofApocynaceae. One of the clearest examples of this are theterms used to describe the stamens of Apocynoideae, whichare highly specialized organs composed of a combination ofsterile and fertile tissues. Woodson (1933) used the term ‘‘pel-tate connective’’ for the dorsal, sclerified part of the stamensthat extends below the thecae to form the two side wings.Pichon (1948) considered this tissue to be a specialization ofthe anthers and proposed the term ‘‘retinacle’’ to denote thesterile, nonsclerified portion of the stamens on the ventral sur-face of the connective that is adjacent or attached to the stylehead, forming the gynostegium (fig. 1B, 1C). The name ‘‘reti-nacle,’’ however, has been widely used in Orchidaceae torefer to the upper portion of the translator arm of the polli-naria since its first use by Darwin (1895). Robertson (1887)adopted the same term to describe an analogous structure of
1 Author for correspondence; current address: Universidade deSao Paulo, Escola de Artes, Ciencias e Humanidades (EACH), Avenida
Arlindo Bettio 1000, Ermelino Matarazzo, Cep 03828-000, Sao Paulo,
Brazil; e-mail [email protected].
Manuscript received May 2006; revised manuscript received April 2007.
999
Int. J. Plant Sci. 168(7):999–1012. 2007.
� 2007 by The University of Chicago. All rights reserved.
1058-5893/2007/16807-0003$15.00

the pollinaria in the former Asclepiadaceae, and he was fol-lowed by other authors (Moore 1946; Vijayaraghavan andCheema 1977; a more complete list was provided by Book-man [1981]). The terminology used to characterize the cellsof the retinacle of Apocynoideae and their relation to thestyle head is also confusing. Terms like ‘‘hairs,’’ ‘‘trichomes,’’and ‘‘brushlike cells’’ all refer to the same thing and have beenextensively used in the taxonomic literature by many authors(e.g., Pichon 1950; Fallen 1986; Endress et al. 1996; Sennbladet al. 1998). In contrast, ‘‘adnation,’’ ‘‘agglutination,’’ and ‘‘con-tact,’’ even though poorly defined, probably represent different,nonhomologous sorts of union between stamens and style head.Mesechiteae, as circumscribed by Endress and Bruyns (2000),
is one of the tribes of Apocynoideae and comprises nine genera:Allomarkgrafia Woodson, Galactophora Woodson, QuiotaniaZarucchi, Macrosiphonia Mull. Arg., Mandevilla Lindl., Me-sechites A. DC., Secondatia A.DC., Telosiphonia (Woodson)Henr., and Tintinnabularia Woodson. In their classification,Mesechiteae was characterized mainly on the basis of the gyno-stegium structure, with two diagnostic characters: the stylehead without a soft, membranous basal collar but having fivestrongly projecting vertical ribs (fig. 1D) and the anthers beingattached to the style head by very short hairs or by cellularfusion. In the latter case, fusion occurs between the smoothepidermal surface of the two organs and is accompanied by in-creased periclinal cell division.In a broad phylogenetic study of the tribe, Simoes et al.
(2004) proposed a recircumscription of Mesechiteae, exclud-ing Galactophora and Secondatia and including ForsteroniaMull. Arg., the latter having been placed in the tribe Apocy-neae by Endress and Bruyns (2000). This modified circum-
scription of Mesechiteae by Simoes et al. (2004) relied mainlyon molecular characters but was also supported by morpho-logical evidence and showed that the two morphological char-acters used by Endress and Bruyns (2000) to characterizeMesechiteae (style head with five strongly projecting verticalribs and anthers attached to the style head by cellular fusionor very short hairs) were diagnostic for the tribe. Recently,Simoes et al. (2006) provided a more complete phylogeny ofMesechiteae based on a broad taxon sampling and sequencingof additional genetic regions. Their results corroborated thefindings of Simoes et al. (2004) and provided a better under-standing of generic relationships within Mesechiteae, espe-cially for the large genus Mandevilla. In the light of theseresults, Simoes et al. (2007) proposed significant amendmentsto the classification of the tribe by broadening the circum-scription of Mandevilla, with the inclusion of Quiotania,Macrosiphonia, and Telosiphonia into its synonymy. Thusdefined, Mesechiteae comprises five genera (Allomarkgrafia,Forsteronia, Mandevilla, Mesechites, and Tintinnabularia),a classification that is followed in this article hereafter.A comparison between the gynostegium morphology of
genera from Mesechiteae and that of Secondatia can provideuseful information on homology assessment in the tribe andtest whether the observed morphological and anatomicalcharacteristics of that organ are congruent with the phyloge-netic evidence derived from molecular characters. The aimsof this study are (1) to characterize the gynostegium structureof 11 species belonging to four genera, of which three (Man-devilla, Mesechites, and Secondatia) were previously includedin Mesechiteae by Endress and Bruyns (2000) and one (For-steronia) was later transferred to the tribe by Simoes et al.
Fig. 1 Diagrammatic drawings depicting morphological and functional parts of the gynostegium of Apocynoideae. A–C, Secondatia densiflora.D, Mandevilla syrinx Woodson. A, Partially schematic longitudinal section of flower. B, Anther in ventral view. C, Anther in lateral view. D, Style
head. aa ¼ apical appendages; aw ¼ lignified anther wings; co ¼ corolla; f ¼ filament; gz ¼ glue zone; lr ¼ longitudinal ribs; o ¼ ovary;
pd ¼ pollen depository; r ¼ retinacle; s ¼ stamen; sc ¼ stigmatic chamber; sh ¼ style head; st ¼ style; t ¼ thecae. Scale bars ¼ 1 mm. A detailedexplanation of the three functional zones of the gynostegium (pollen depository, glue zone, and stigmatic chamber) is provided in the text (see
‘‘Organization of the Gynostegium’’).
1000 INTERNATIONAL JOURNAL OF PLANT SCIENCES

(2004), (2) to revise the terminology required to describe thegynostegium in the studied species, (3) to verify the existenceof structural patterns of the gynostegium in the tribe, and (4)to compare the classifications of Endress and Bruyns (2000)and Simoes et al. (2004, 2006) with the results obtained here.
Material and Methods
Nine species from three genera included in Mesechiteaeby Simoes et al. (2004), as amended by Simoes et al. (2006,2007), were selected for this study: four from Forsteronia,four from Mandevilla, and one from Mesechites. In addition,two species of Secondatia (Apocyneae) were included for com-parative purposes, since these were placed in Mesechiteaeby Endress and Bruyns (2000). The four studied species ofMandevilla were selected to represent three distinct taxo-nomic groups within the genus, as delimited by Simoes et al.(2006). Representatives from the other two genera includedin Mesechiteae by Simoes et al. (2004), Allomarkgrafia andTintinnabularia, were not included in this study because ofthe lack of available material for anatomical investigation.Only anthetic flowers were selected. Samples were col-
lected in their natural habitat in southeastern Brazil and lateridentified by the first author, and vouchers were deposited inthe herbarium UEC. Voucher information is provided in theappendix. Flowers of Mandevilla tenuifolia were obtainedfrom cultivated specimens maintained at the greenhouse of theDepartment of Botany at the University of Campinas, Brazil.Flowers were collected and fixed in FAA (Johansen 1940)
for at least 24 h at room temperature and placed under a lowvacuum to aid the penetration of the fixative. The flowerswere then stored in 70% ethanol. For anatomical sections,only the portion of the corolla surrounding the gynostegiumat the transition zone between the lower and upper corollatube was used. In Apocynaceae, two distinct parts of thecorolla tube can be discerned: the congenitally fused lowertube, which is composed of the united stamen and petal pri-mordia, and the postgenitally fused upper tube, which is thepart of the corolla above stamen insertion up to the base ofthe corolla lobes (Nishino 1982; Endress et al. 1996; Simoesand Kinoshita 2002). In Mandevilla longiflora and Mandevillascabra, the whole upper corolla tube was removed in order toexpose the gynostegium. All materials were dehydrated usingthe tertiary butyl alcohol series, embedded in Paraplast, andthen sectioned (Johansen 1940). Longitudinal and transverseserial sections 10–12 mm thick were cut with a rotary micro-tome. Deparaffinized sections were stained with Safranin Oand Astra Blue (Gerlach 1969). Light microscope observationswere carried out with an Olympus BX51 microscope. Photo-micrographs were made using Kodak Pro Image (100 ASA)film and were electronically processed using the softwareAdobe Photoshop 7.0.
Results
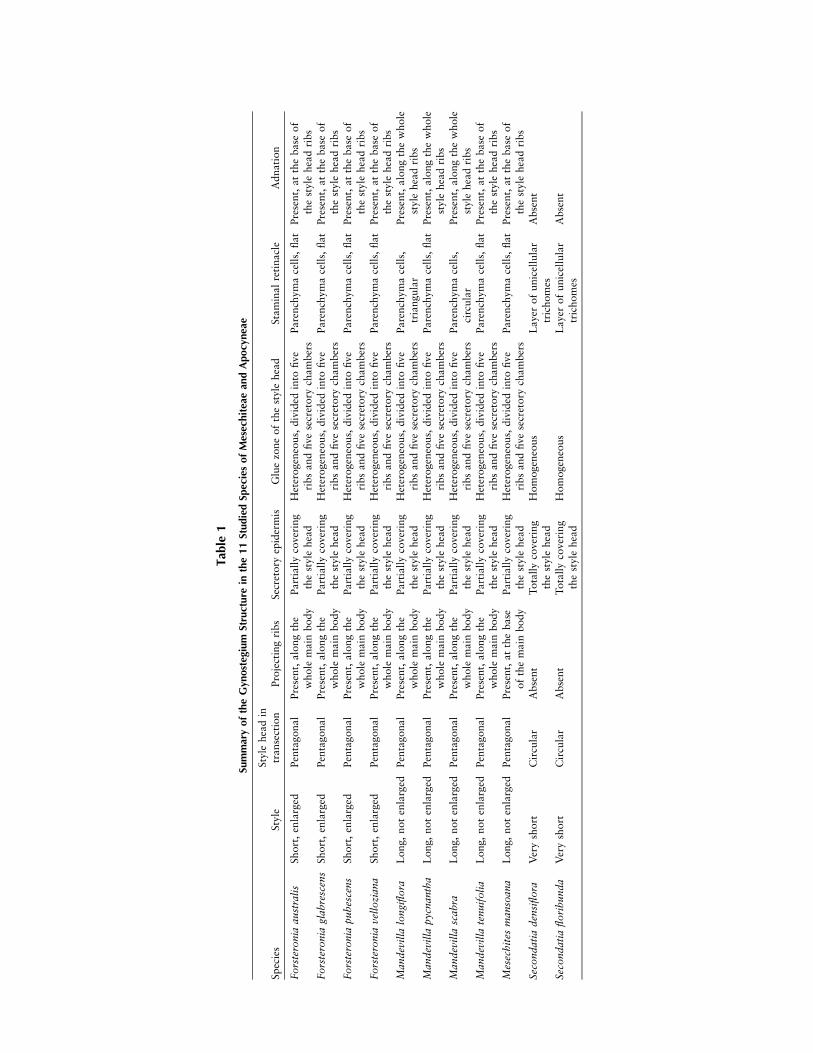
A comparison between the gynostegium structures of allstudied species is summarized in table 1. The style forms aslender tube, becoming enlarged in the upper portion at thebase of the style head. The style is long and cylindrical inMandevilla and Mesechites and short and somewhat enlarged
in Forsteronia. In Secondatia floribunda, the style is greatlyreduced and not clearly discernible from the style head. In allspecies, the style head is composed of two parts: the mainbody and the apical appendages, the former corresponding tothe conical to cylindrical structure located above the styleand the latter representing the uppermost, unfused apices ofthe two carpels, free from each other and positioned on thetop of the main body (figs. 1D, 2A). In Forsteronia, the stylehead main body is broader in its middle part, whereas inSecondatia, it is broader in its lower part, forming a ringlikestructure.In all species, an epidermis formed by a single layer of pali-
sade cells, sometimes very elongated, covers the style head.The epidermal cells are longitudinally elongated and havethin cell walls, a relatively large nucleus, and dense cytoplasm,a set of characteristics common to secretory cells. This secretoryepidermis is absent only at the top of the apical appendages.In Secondatia, the whole surface of the style head is coveredby a secretory epidermis, even in its lower portion below thebasal ring and close to the ovary (fig. 3A). Secretion was ob-served inside these cells and also covering the main body ofthe style head and adjacent portions of the stamens. On theupper half of the main body, close to the base of the antherthecae, these cells reach their maximum length, and in twospecies (Mandevilla pycnantha, Mesechites mansoana), a con-siderable amount of secretion is accumulated outside (figs. 3B,4C; fig. 5B, 5C; fig. 6B).The apical appendages show great variation in size, from
very small and inconspicuous in Mandevilla longiflora andMandevilla scabra (fig. 2B) to well developed and almost aslong as the main body of the style head in species of Forstero-nia (fig. 6A), M. pycnantha (fig. 2A), and M. mansoana (fig.5A). They are composed, for the most part, of parenchymacells covered by a single epidermal layer, secretory in Seconda-tia and nonsecretory in the other genera. In M. pycnantha,Mandevilla tenuifolia, M. mansoana, and the four studied spe-cies of Forsteronia, cells from the parenchyma core stained in-tensely red or purple, a condition particularly marked in M.pycnantha, in which the entire appendages were deeply redstained (fig. 2A; fig. 4A, 4B).In Forsteronia, Mandevilla, and Mesechites, the paren-
chyma cells of the main body of the style head proliferate infive parts and grow toward the stamens, forming fivestrongly projecting ribs. In Mandevilla, the ribs are well de-veloped and extend along the entire length of the main body,reaching their maximum size at the base (fig. 2; fig. 4A; fig.7C, 7G). In Forsteronia, the ribs also extend along the mainbody but are much less developed and reach their maximumsize at the middle part (fig. 6A; fig. 7D, 7H). In Mesechites,the ribs are shorter and are restricted to the base of the mainbody (fig. 5; fig. 7B, 7F). Each of the five spaces between ad-jacent ribs and the surrounding stamens forms a chamberclosed at the bottom by the extended base of the style headribs and open at the top, partially to completely filled withsecretion (secretory chambers of the gynostegium; see detailsin ‘‘Discussion’’). In Mesechites, these chambers are well de-limited only at the base of the style head, at the level of theribs, but are not completely separated from each other abovethat level. In M. tenuifolia, M. pycnantha, and M. mansoana,the secretion can surpass the limits of the chambers and was
1001SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)
Tab
le1
SummaryoftheGyn
osteg
ium
Structure
inthe11StudiedSp
eciesofMesechitea
ean
dApocy
nea
e
Species
Style
Style
headin
transection
Projectingribs
Secretory
epidermis
Gluezoneofthestyle
head
Staminalretinacle
Adnation
Forsteronia
australis
Short,enlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Forsteronia
glab
rescens
Short,enlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Forsteronia
pubescens
Short,enlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Forsteronia
vellozian
aShort,enlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Man
devilla
longiflora
Long,notenlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,
triangular
Present,alongthewhole
style
headribs
Man
devilla
pycnan
tha
Long,notenlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,alongthewhole
style
headribs
Man
devilla
scab
raLong,notenlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,
circular
Present,alongthewhole
style
headribs
Man
devilla
tenuifolia
Long,notenlarged
Pentagonal
Present,alongthe
whole
main
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Mesechites
man
soan
aLong,notenlarged
Pentagonal
Present,atthebase
ofthemain
body
Partiallycovering
thestyle
head
Heterogeneous,divided
into
five
ribsandfivesecretory
chambers
Parenchymacells,flat
Present,atthebase
of
thestyle
headribs
Secondatia
densiflora
Veryshort
Circular
Absent
Totallycovering
thestyle
head
Homogeneous
Layer
ofunicellular
trichomes
Absent
Secondatia
floribunda
Veryshort
Circular
Absent
Totallycovering
thestyle
head
Homogeneous
Layer
ofunicellular
trichomes
Absent

observed to fill the space between the apical appendages ofthe style head and the anther thecae.The style head of Secondatia has a morphological pattern
different from those of the other studied genera (fig. 3; fig.7A, 7E). The main body is long, cylindrical, and smooth,with no discernible projecting ribs. In cross sections, the stylehead has a circular shape (figs. 3B–3D, 7E), in contrast tothe other genera, in which the style head is strongly pentago-nal, at least at the base (figs. 4C–4E, 7B–7D, 7F–7H).
The stamens are inserted in the corolla at the level corre-sponding to the base of the upper tube, forming a cone aroundthe style head in all genera. They are divided into three mainparts: anther, connective, and filament. The anthers are com-plex structures, composed of both sterile and fertile regions.The dorsal part is mainly composed of sclerenchyma and ex-tends below the thecae to form two side wings, which aresagittate in Secondatia and more or less cordate in Forstero-nia, Mandevilla, and Mesechites; the thecae are reduced and
Fig. 2 Longitudinal sections of the gynostegium of Mandevilla. A, M. pycnantha. B, M. scabra. C, M. tenuifolia. D, M. longiflora. a ¼ anther;
ap ¼ style head apical appendages; c ¼ connective; f ¼ filament; mb ¼ style head main body; s ¼ style; sh ¼ style head. The transition betweenconnective and filament is indicated by a dashed line.
1003SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)

restricted to the upper half of the ventral part of the anther.The connective is composed of two main parts: a narrow stripof parenchyma cells between the two thecae on top of an en-larged portion that extends down to the region in which the
anther side wings are differentiated (fig. 1B, 1C; fig. 8). In thisenlarged portion, the ventral surface is covered by unicellulartrichomes (Secondatia; fig. 3C, 3E) or epidermal parenchymacells (Forsteronia, Mandevilla, and Mesechites; figs. 4–6, 9) that
Fig. 3 Gynostegium structure of Secondatia. A–C, S. densiflora. D, E, S. floribunda. A, General view. B, Upper portion of the gynostegium.
C, Lower portion of the gynostegium, showing the proximity between the staminal retinacle and the epidermal cells of the style head. D, Base of
the style head. E, Trichomes of the staminal retinacle, interdigitated with the epidermis of the style head. A, E, Longitudinal sections; B, C, D,
cross sections. a ¼ anther; c ¼ connective; f ¼ filament; o ¼ ovary; s ¼ style; sh ¼ style head; sr ¼ staminal retinacle. The transition betweenconnective and filament is indicated by a dashed line.
1004 INTERNATIONAL JOURNAL OF PLANT SCIENCES

are in close contact with the epidermal cells of the style head.In M. pycnantha, M. tenuifolia, M. mansoana, and Forsteronia,this enlarged portion is flat, and the ventral surface is com-posed of only a few layers of epidermal parenchyma cells and
is not clearly distinct from the sterile tissue of the anther (fig.9A, 9B). In M. longiflora and M. scabra, in contrast, the ven-tral surface is rounded (M. scabra; fig. 9C, 9D) or triangularin cross section (M. longiflora; fig. 9E, 9F) and composed of
Fig. 4 Gynostegium structure of Mandevilla pycnantha. A, Overview; 1, 2, 3, and 4 correspond to the levels of the transverse sections
illustrated in B, C, D, and E, respectively. B, Upper portion of the gynostegium, with the anther thecae and the style head held togetherby secretion. C, Zone of adnation at the top of the style head ribs. D, Middle portion of the zone of adnation. E, Base of style head, in which the
style head ribs reach their maximum size and adnation is strongest. A, Longitudinal section; B–E, cross sections. a ¼ anther; c ¼ connective;
f ¼ filament; sh ¼ style head. The transition between connective and filament is indicated by a dashed line.
1005SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)

multiple layers of parenchyma cells that are clearly distinctfrom the sterile tissue of the anther. The filaments are locatedjust below the connective and are densely covered by long, uni-cellular trichomes directed toward the base of the style headand style. The upper part of the filaments and the connective
have a similar histological composition, with no clear distinc-tion between them. No secretory activity was detected in thestamens in any of the studied species.In Forsteronia, Mandevilla, and Mesechites, the androecium
and gynoecium are adnate at the level in which the style head
Fig. 5 Gynostegium structure ofMesechites mansoana. A, Overview. B, C, The anther thecae and the style head are held together by secretion.
D, E, Zone of adnation. A, B, E, Longitudinal sections; C, D, cross sections. a ¼ anther; c ¼ connective; f ¼ filament; sh ¼ style head. Secretedproducts in B and C are indicated by arrows. The transition between connective and filament is indicated by a dashed line.
1006 INTERNATIONAL JOURNAL OF PLANT SCIENCES
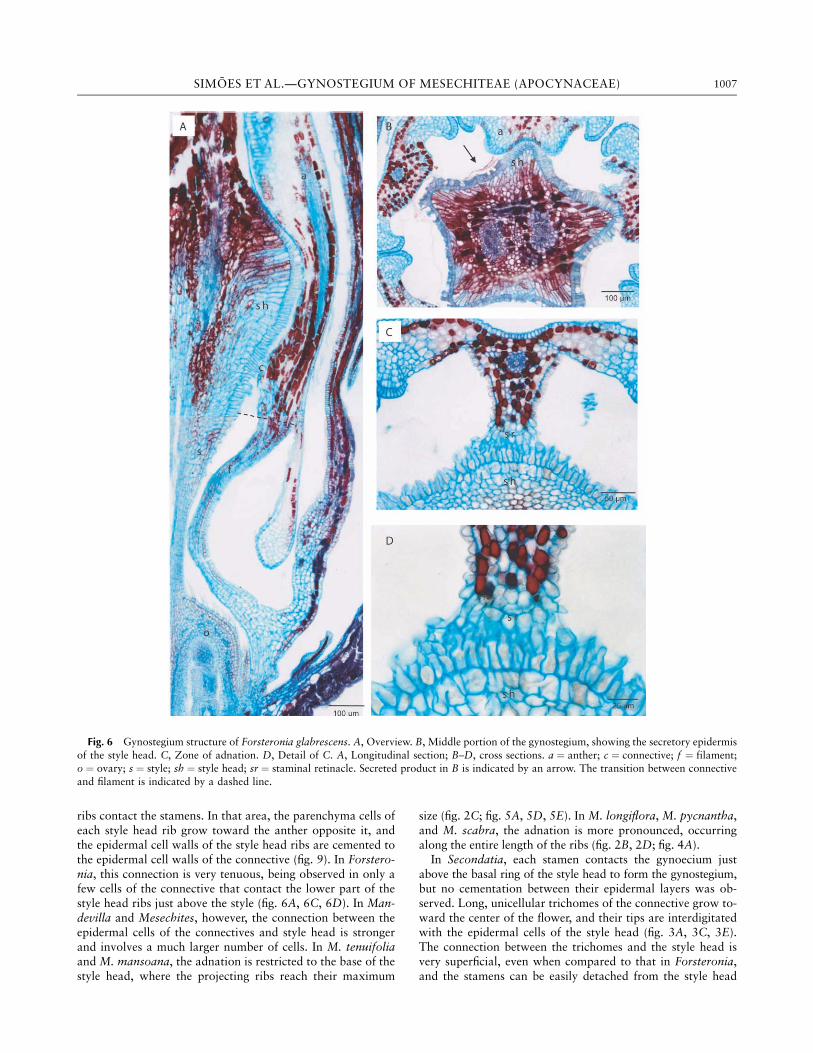
ribs contact the stamens. In that area, the parenchyma cells ofeach style head rib grow toward the anther opposite it, andthe epidermal cell walls of the style head ribs are cemented tothe epidermal cell walls of the connective (fig. 9). In Forstero-nia, this connection is very tenuous, being observed in only afew cells of the connective that contact the lower part of thestyle head ribs just above the style (fig. 6A, 6C, 6D). In Man-devilla and Mesechites, however, the connection between theepidermal cells of the connectives and style head is strongerand involves a much larger number of cells. In M. tenuifoliaand M. mansoana, the adnation is restricted to the base of thestyle head, where the projecting ribs reach their maximum
size (fig. 2C; fig. 5A, 5D, 5E). In M. longiflora, M. pycnantha,and M. scabra, the adnation is more pronounced, occurringalong the entire length of the ribs (fig. 2B, 2D; fig. 4A).In Secondatia, each stamen contacts the gynoecium just
above the basal ring of the style head to form the gynostegium,but no cementation between their epidermal layers was ob-served. Long, unicellular trichomes of the connective grow to-ward the center of the flower, and their tips are interdigitatedwith the epidermal cells of the style head (fig. 3A, 3C, 3E).The connection between the trichomes and the style head isvery superficial, even when compared to that in Forsteronia,and the stamens can be easily detached from the style head
Fig. 6 Gynostegium structure of Forsteronia glabrescens. A, Overview. B, Middle portion of the gynostegium, showing the secretory epidermis
of the style head. C, Zone of adnation. D, Detail of C. A, Longitudinal section; B–D, cross sections. a ¼ anther; c ¼ connective; f ¼ filament;o ¼ ovary; s ¼ style; sh ¼ style head; sr ¼ staminal retinacle. Secreted product in B is indicated by an arrow. The transition between connective
and filament is indicated by a dashed line.
1007SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)
when flowers are dissected. No proliferation of parenchymacells was observed in the main body of the style head or inthe connective (fig. 3A, 3D; fig. 7A, 7E).A second region in which the androecium and the gynoe-
cium are in close contact was observed in all species. In thatregion, the basal part of the anther thecae and the upper partof the style head below the apical appendages are held to-gether by the secretion produced by the epidermal cells of thestyle head (figs. 3B, 5B). These secretory cells are particularlywell developed and reach their maximum size in that region,and the cytoplasm therein is intensely stained pink or blue.Evidence for the location of the stigmatic region was ob-
served only in M. scabra. In that species, pollen tube growthwas detected at the transition zone between the style headand the style, in the concavity at the base of the style head. Nopollen tube growth was detected in any of the other species.
Discussion
Stamen Structure and Terminology
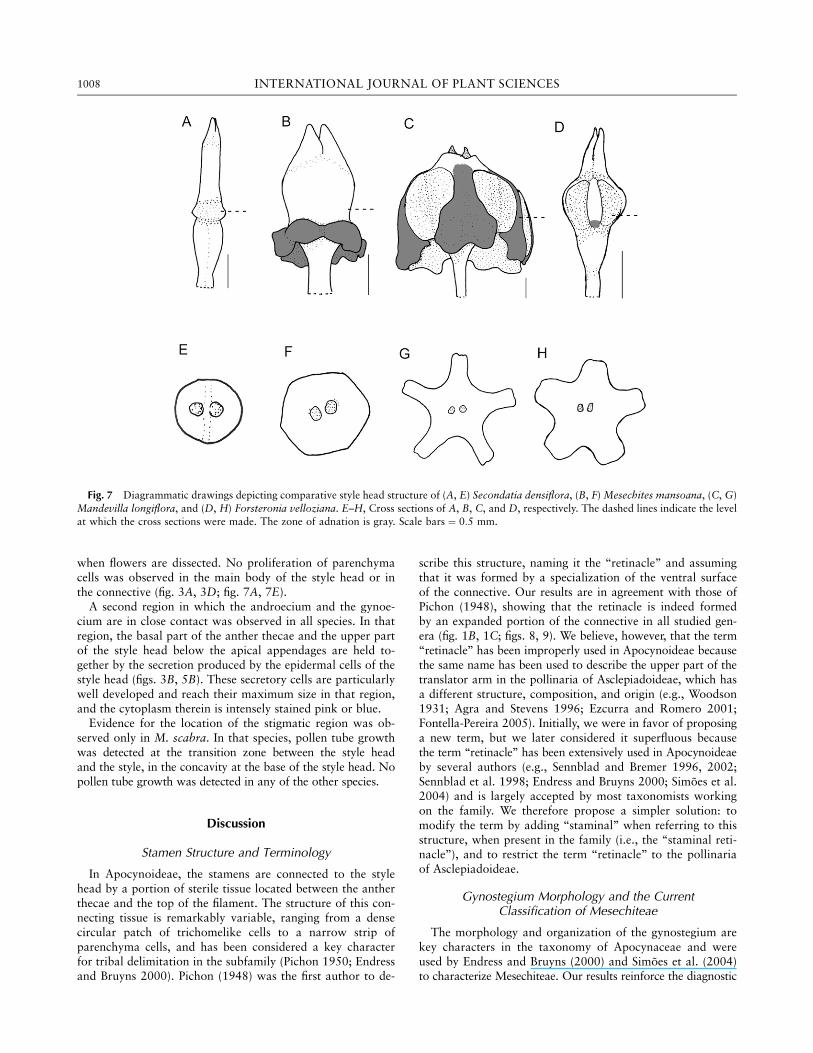
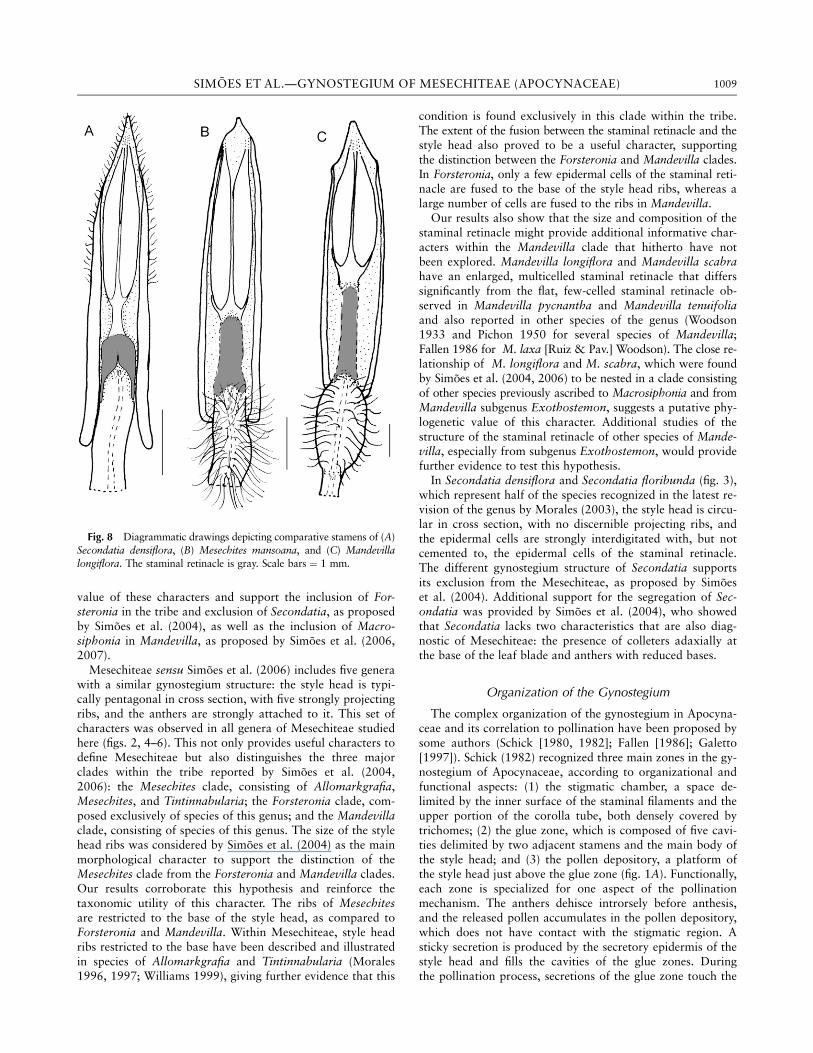
In Apocynoideae, the stamens are connected to the stylehead by a portion of sterile tissue located between the antherthecae and the top of the filament. The structure of this con-necting tissue is remarkably variable, ranging from a densecircular patch of trichomelike cells to a narrow strip ofparenchyma cells, and has been considered a key characterfor tribal delimitation in the subfamily (Pichon 1950; Endressand Bruyns 2000). Pichon (1948) was the first author to de-
scribe this structure, naming it the ‘‘retinacle’’ and assumingthat it was formed by a specialization of the ventral surfaceof the connective. Our results are in agreement with those ofPichon (1948), showing that the retinacle is indeed formedby an expanded portion of the connective in all studied gen-era (fig. 1B, 1C; figs. 8, 9). We believe, however, that the term‘‘retinacle’’ has been improperly used in Apocynoideae becausethe same name has been used to describe the upper part of thetranslator arm in the pollinaria of Asclepiadoideae, which hasa different structure, composition, and origin (e.g., Woodson1931; Agra and Stevens 1996; Ezcurra and Romero 2001;Fontella-Pereira 2005). Initially, we were in favor of proposinga new term, but we later considered it superfluous becausethe term ‘‘retinacle’’ has been extensively used in Apocynoideaeby several authors (e.g., Sennblad and Bremer 1996, 2002;Sennblad et al. 1998; Endress and Bruyns 2000; Simoes et al.2004) and is largely accepted by most taxonomists workingon the family. We therefore propose a simpler solution: tomodify the term by adding ‘‘staminal’’ when referring to thisstructure, when present in the family (i.e., the ‘‘staminal reti-nacle’’), and to restrict the term ‘‘retinacle’’ to the pollinariaof Asclepiadoideae.
Gynostegium Morphology and the CurrentClassification of Mesechiteae
The morphology and organization of the gynostegium arekey characters in the taxonomy of Apocynaceae and wereused by Endress and Bruyns (2000) and Simoes et al. (2004)to characterize Mesechiteae. Our results reinforce the diagnostic
Fig. 7 Diagrammatic drawings depicting comparative style head structure of (A, E) Secondatia densiflora, (B, F) Mesechites mansoana, (C, G)
Mandevilla longiflora, and (D, H) Forsteronia velloziana. E–H, Cross sections of A, B, C, and D, respectively. The dashed lines indicate the levelat which the cross sections were made. The zone of adnation is gray. Scale bars ¼ 0:5 mm.
1008 INTERNATIONAL JOURNAL OF PLANT SCIENCES
value of these characters and support the inclusion of For-steronia in the tribe and exclusion of Secondatia, as proposedby Simoes et al. (2004), as well as the inclusion of Macro-siphonia in Mandevilla, as proposed by Simoes et al. (2006,2007).Mesechiteae sensu Simoes et al. (2006) includes five genera
with a similar gynostegium structure: the style head is typi-cally pentagonal in cross section, with five strongly projectingribs, and the anthers are strongly attached to it. This set ofcharacters was observed in all genera of Mesechiteae studiedhere (figs. 2, 4–6). This not only provides useful characters todefine Mesechiteae but also distinguishes the three majorclades within the tribe reported by Simoes et al. (2004,2006): the Mesechites clade, consisting of Allomarkgrafia,Mesechites, and Tintinnabularia; the Forsteronia clade, com-posed exclusively of species of this genus; and the Mandevillaclade, consisting of species of this genus. The size of the stylehead ribs was considered by Simoes et al. (2004) as the mainmorphological character to support the distinction of theMesechites clade from the Forsteronia and Mandevilla clades.Our results corroborate this hypothesis and reinforce thetaxonomic utility of this character. The ribs of Mesechitesare restricted to the base of the style head, as compared toForsteronia and Mandevilla. Within Mesechiteae, style headribs restricted to the base have been described and illustratedin species of Allomarkgrafia and Tintinnabularia (Morales1996, 1997; Williams 1999), giving further evidence that this
condition is found exclusively in this clade within the tribe.The extent of the fusion between the staminal retinacle and thestyle head also proved to be a useful character, supportingthe distinction between the Forsteronia and Mandevilla clades.In Forsteronia, only a few epidermal cells of the staminal reti-nacle are fused to the base of the style head ribs, whereas alarge number of cells are fused to the ribs in Mandevilla.Our results also show that the size and composition of the
staminal retinacle might provide additional informative char-acters within the Mandevilla clade that hitherto have notbeen explored. Mandevilla longiflora and Mandevilla scabrahave an enlarged, multicelled staminal retinacle that differssignificantly from the flat, few-celled staminal retinacle ob-served in Mandevilla pycnantha and Mandevilla tenuifoliaand also reported in other species of the genus (Woodson1933 and Pichon 1950 for several species of Mandevilla;Fallen 1986 for M. laxa [Ruiz & Pav.] Woodson). The close re-lationship of M. longiflora and M. scabra, which were foundby Simoes et al. (2004, 2006) to be nested in a clade consistingof other species previously ascribed to Macrosiphonia and fromMandevilla subgenus Exothostemon, suggests a putative phy-logenetic value of this character. Additional studies of thestructure of the staminal retinacle of other species of Mande-villa, especially from subgenus Exothostemon, would providefurther evidence to test this hypothesis.In Secondatia densiflora and Secondatia floribunda (fig. 3),
which represent half of the species recognized in the latest re-vision of the genus by Morales (2003), the style head is circu-lar in cross section, with no discernible projecting ribs, andthe epidermal cells are strongly interdigitated with, but notcemented to, the epidermal cells of the staminal retinacle.The different gynostegium structure of Secondatia supportsits exclusion from the Mesechiteae, as proposed by Simoeset al. (2004). Additional support for the segregation of Sec-ondatia was provided by Simoes et al. (2004), who showedthat Secondatia lacks two characteristics that are also diag-nostic of Mesechiteae: the presence of colleters adaxially atthe base of the leaf blade and anthers with reduced bases.
Organization of the Gynostegium
The complex organization of the gynostegium in Apocyna-ceae and its correlation to pollination have been proposed bysome authors (Schick [1980, 1982]; Fallen [1986]; Galetto[1997]). Schick (1982) recognized three main zones in the gy-nostegium of Apocynaceae, according to organizational andfunctional aspects: (1) the stigmatic chamber, a space de-limited by the inner surface of the staminal filaments and theupper portion of the corolla tube, both densely covered bytrichomes; (2) the glue zone, which is composed of five cavi-ties delimited by two adjacent stamens and the main body ofthe style head; and (3) the pollen depository, a platform ofthe style head just above the glue zone (fig. 1A). Functionally,each zone is specialized for one aspect of the pollinationmechanism. The anthers dehisce introrsely before anthesis,and the released pollen accumulates in the pollen depository,which does not have contact with the stigmatic region. Asticky secretion is produced by the secretory epidermis of thestyle head and fills the cavities of the glue zones. Duringthe pollination process, secretions of the glue zone touch the
Fig. 8 Diagrammatic drawings depicting comparative stamens of (A)Secondatia densiflora, (B) Mesechites mansoana, and (C) Mandevillalongiflora. The staminal retinacle is gray. Scale bars ¼ 1 mm.
1009SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)

pollinator and stick on a mass of pollen in the pollen deposi-tory, with the whole unit being carried by the pollinator toanother flower. The donor pollen carried by the pollinator isbrushed off by the staminal and corolline trichomes of thestigmatic chamber of the recipient flower.The same organizational pattern was seen in all species,
with the three regions of the gynostegium clearly differenti-ated. We observed, however, that in Forsteronia, Mandevilla,and Mesechites, the glue zone of the gynostegium is not ho-mogeneous but instead is formed by two structural and func-
tional parts: the secretory chambers and the five alternatingribs. No subdivision of the glue zone was observed in Secon-datia, again supporting its exclusion from Mesechiteae.The five secretory chambers are formed by the concave
space between two adjacent style head ribs and their attachedstamens. The chambers act as storage areas for the secretionproduced by the epidermis of the main body of the style headin the five regions located between the ribs. The epidermal se-cretory cells reach their maximum size in that region, especiallyat the upper part of the chambers that contacts the base of the
Fig. 9 Cross sections of the gynostegium of Mandevilla, showing the zone of adnation between epidermal cells in detail. A, B, M. pycnantha,staminal retinacle flat. C, D, M. scabra, staminal retinacle enlarged. E, F, M. longiflora, staminal retinacle enlarged. a ¼ anther; sh ¼ style head;
sr ¼ staminal retinacle.
1010 INTERNATIONAL JOURNAL OF PLANT SCIENCES

anther thecae, and are less developed or even absent in thestyle head ribs. The observed differences in the size and distri-bution of this epidermal tissue suggest that the secretory activityon the style head is not homogeneous but instead is concen-trated or even restricted to the epidermis that covers the innerwalls of the secretory chambers close to the pollen depository.The five alternating ribs are the most enlarged portion of
the main body and function primarily as a contacting surfaceto the androecium. In Mandevilla and Mesechites, there is aprobable correlation between the length of the ribs and degreeof adnation to the stamens. In these genera, the whole longitu-dinal surface of the ribs is strongly adnate to the stamens; theonly exception is M. tenuifolia, in which only the epidermalcells at the base of the ribs are cemented to the staminal reti-nacle epidermis. This correlation was not observed in Forstero-nia, however, with most of the style head rib free from thestamens and only the basalmost cells effectively adnate to thestaminal retinacle. We believe that even though the primaryfunction of the style head ribs is to increase the area of contactbetween the gynoecium and the androecium, they can also bespecialized for other functions, e.g., acting as a wall to delimitthe areas of production and storage of secretion or also servingas a support for the stamens in the gynostegium architecture.Observations of the gynoecium of other species from Mesechi-teae, especially from Forsteronia and Mandevilla, are necessaryto achieve a better understanding of the evolution of this organwithin the tribe.
Conclusions
The main conclusions of our study are as follows. (1) Thegynostegium structure of the species we studied corroborate
the new circumscription of Mesechiteae proposed by Simoeset al. (2004), with the inclusion of Forsteronia and the exclu-sion of Secondatia. The gynostegia of Forsteronia, Mandevilla,and Mesechites have the same organization, with five project-ing longitudinal ribs adnate to the epidermal cells of thestaminal retinacle, whereas in Secondatia, the style head hasno projecting ribs, and the epidermal cells are interdigitatedwith the staminal retinacle. (2) Differences in the size of thestyle head ribs and their degree of adnation to the stamensalso support the three major clades (Forsteronia, Mesechites,and Mandevilla, respectively) within the tribe observed bySimoes et al. (2004, 2006). (3) The division of the gynoste-gium into three organizational and functional regions (stig-matic chamber, glue zone, and pollen depository) proposedby Schick (1982) was observed in all studied species. In For-steronia, Mandevilla, and Mesechites, however, the glue zoneis specialized into two structural and functional parts: thestyle head ribs, involved in the adnation process with the an-droecium, and the secretory chambers, involved in the pro-duction and storage of secretion.
Acknowledgments
We thank Dr. Mary E. Endress for her critical review ofthe manuscript and two anonymous reviewers for their valu-able comments and suggestions. We are also grateful to RicardoLourenco for skillfully preparing figure 1. This study was sup-ported by grants to A. O. Simoes from CAPES (Coordenacaoe Aperfeicoamento de Pessoal de Nıvel Superior, Brazil) andto A. O. Simoes andM. C. S. do Rio from FAPESP (Fundacao deAmparo a Pesquisa do Estado de Sao Paulo, Brazil; 00/04994-0and 01/05341-3, respectively).
Appendix
Literature Cited
Agra MF, WD Stevens 1996 Matelea roulinioides, uma nova especieda tribo Gonolobeae (Asclepiadaceae) do Nordeste brasileiro. Novon
6:6–8.
Allorge L 1976 Morphologie et biologie florale des Apocynacees:aplications taxonomiques. PhD diss. Ecole Pratique des Hautes
Etudes, Paris.
Table A1
Voucher Information for the Taxa Used in This Article
Species Localitya Collector
Forsteronia australis Mull. Arg Sao Paulo, Moji Guacu M. C. S. do Rio et al. 26
Forsteronia glabrescens Mull. Arg. Sao Paulo, Moji Guacu M. C. S. do Rio et al. 22
Forsteronia pubescens A. DC. Sao Paulo, Sao Jose do Rio Pardo L. S. Kinoshita 94-19 and A. SartoriForsteronia velloziana (A. DC.) Woodson Sao Paulo, Moji Guacu M. C. S. do Rio et al. 29
Mandevilla longiflora (Desf.) Pichon Minas Gerais, Carrancas R. S. Rodrigues et al. 1227
Mandevilla pycnantha (Steud. ex A.DC.)
Woodson Minas Gerais, Grao-Mogol A. O. Simoes et al. 1176Mandevilla scabra (Hoffmanns. ex
Roem. & Schult.) Bahia, Andaraı A. O. Simoes et al. 1127
Mandevilla tenuifolia (J.C. Mikan)
Woodson Sao Paulo, Atibaia A. O. Simoes et al. 1048Mesechites mansoana (A. DC.) Woodson Minas Gerais, Grao-Mogol A. O. Simoes 1087 and E. R. Pansarin
Secondatia densiflora A. DC. Minas Gerais, Serra do Cipo L. S. Kinoshita et al. 02/106
Secondatia floribunda A. DC. Minas Gerais, Pedra Azul A. O. Simoes 1288 and R. B. Singer
a All localities are in Brazil.
1011SIMOES ET AL.—GYNOSTEGIUM OF MESECHITEAE (APOCYNACEAE)

Bookman SS 1981 The floral morphology of Asclepias speciosa(Asclepiadaceae) in relation to pollination and a clarification in
terminology of the genus. Am J Bot 68:675–679.
Darwin C 1895 The various contrivances by which orchids arefertilized by insects. Appleton, New York. 316 pp.
Endress ME, PV Bruyns 2000 A revised classification of the
Apocynaceae s.l. Bot Rev 66:1–56.
Endress ME, B Sennblad, S Nilsson, L Civeyrel, MW Chase, SHuysmans, E Grafstrom, B Bremer 1996 A phylogenetic analysis
of Apocynaceae s. str. and some related taxa in Gentianales: a multi-
disciplinary approach. Opera Bot Belg 7:59–102.
Ezcurra C 1981 Revision de las Apocinaceas de la Argentina. Dar-winiana 23:367–474.
Ezcurra C, MEM Romero 2001 Sobre la presencia de Ditassaacerosa (Asclepiadaceae) en la Argentina. Hickenia 3:95–97.
Fallen ME 1986 Floral structure in Apocynaceae: morphologi-
cal, functional and evolutionary aspects. Bot Jahrb Syst 106:245–
286.
Fontella-Pereira J 2005 Asclepiadaceae. Pages 93–156 inMGLWanderley,GJ Shepherd, TS Mehlem, AM Guilietti, eds. Flora Fanerogamica
de Sao Paulo. RiMa, Sao Paulo.
Galetto E 1997 Flower structure and nectar chemical composition in
three Argentine Apocynaceae. Flora 192:197–207.Gerlach D 1969 Botanische Mikrotechnik: eine Einfuhrung. Thieme,
Stuttgart. 311 pp.
Johansen DA 1940 Plant microtechnique. McGraw-Hill, New York.523 pp.
Judd WS, CS Campbell, EA Kellogg, PF Stevens 2002 Plant system-
atics: a phylogenetic approach. Sinauer, Sunderland, MA. 464 pp.
Moore RJ 1946 Investigation on rubber-bearing plants. III. Devel-opment of normal and aborting seeds in Asclepias syriaca. Can J
Res Sect C Bot Sci 24:55–65.
Morales JF 1996 Una nueva especie de Tintinnabularia (Apocyna-
ceae). Novon 6:392–394.——— 1997 A synopsis of the genus Allomarkgrafia (Apocynaceae).
Brittonia 49:337–345.
——— 2003 Studies in neotropical Apocynaceae. III. A revision of
the genus Secondatia A. DC., with discussion of its genericclassification. Candollea 58:305–319.
Nicholas A, H Baijnath 1994 A consensus classification for the order
Gentianales with additional details on the suborder Apocynineae.Bot Rev 60:440–482.
Nishino E 1982 Corolla tube formation in six species of Apocyna-
ceae. Bot Mag Tokyo 95:1–17.
Pichon M. 1948 Classification des Apocynacees. XIX. Le retinacledes Echitoıdees. Bull Mus Natl Hist Nat 22:211–216.
——— 1950 Classification des Apocynacees. XXV. Echitoıdees.
Mem Mus Natl Hist Nat Ser B Bot 1:1–143.
Rao VS, A Ganguli 1963 Studies in the floral anatomy of the Apo-cynaceae. J Indian Bot Soc 42:419–433.
Robertson C 1887 Insect relations of certain asclepiads. II. Bot Gaz
12:244–250.Rosatti TJ 1989 The genera of suborder Apocyninae (Apocynaceae
and Asclepiadaceae) in the southeastern United States. J Arnold
Arbor Harv Univ 70:307–401.
Schick B 1980 Untersuchungen uber die Biotechnik der Apocyna-ceenblute. I. Morphologie und Funktion des Narbenkopfes. Flora
170:394–432.
——— 1982 Untersuchungen uber die Biotechnik der Apocynaceenblute.
II. Bau und Funktion des Bestaubungsapparates. Flora 172:347–371.Sennblad B, B Bremer 1996 The familial and subfamilial relation-
ships of Apocynaceae and Asclepiadaceae evaluated with rbcL data.
Plant Syst Evol 202:153–175.——— 2002 Classification of Apocynaceae s.l. according to a new
approach combining Linnaean and phylogenetic taxonomy. Syst
Biol 51:389–409.
Sennblad B, ME Endress, B Bremer 1998 Morphology and moleculardata in phylogenetic fraternity: the tribe Wrightieae (Apocynaceae)
revisited. Am J Bot 85:1143–1158.
Simoes AO, ME Endress, T van der Niet, LS Kinoshita, E Conti 2004
Tribal and intergeneric relationships of Mesechiteae (Apocynaceae,Apocynoideae): evidence from three noncoding plastid regions and
morphology. Am J Bot 91:1409–1418.
——— 2006 Is Mandevilla monophyletic? evidence from five plastidDNA loci and morphology. Ann Mo Bot Gard 93:565–591.
Simoes AO, LS Kinoshita 2002 The Apocynaceae s. str. of the Carrancas
region, Minas Gerais, Brazil. Darwiniana 40:127–169.
Simoes AO, LS Kinoshita, ME Endress 2007 New combinations inMandevilla Lindley (Apocynaceae, Apocynoideae). Novon 17:87–90.
Vijayaraghavan MR, K Cheema 1977 Ontogenetical and histochem-
ical studies on the translator apparatus in Calotropis procera R. Br.
I. The retinaculum. Acta Histochem 59:15–20.Williams JK 1999 A new species of Tintinnabularia (Apocynaceae,
Apocynoideae) from Honduras, with taxonomical notes. Lundellia
2:136–141.
Woodson RE Jr 1931 New South American Asclepiadaceae. Ann MoBot Gard 18:557–563.
——— 1933 Studies in the Apocynaceae. IV. The American genera of
Echitoideae. Ann Mo Bot Gard 20:605–790.——— 1935 Studies in the Apocynaceae. IV. The American genera of
Echitoideae. Ann Mo Bot Gard 22:153–306.
——— 1936 Studies in the Apocynaceae. IV. The American genera of
Echitoideae. Ann Mo Bot Gard 23:169–548.Woodson RE, Jr, JA Moore 1938 The vascular anatomy and com-
parative morphology of apocynaceous flowers. Bull Torrey Bot Club
65:79–134.
1012 INTERNATIONAL JOURNAL OF PLANT SCIENCES




















