
Cytotype distribution in Empetrum ( Ericaceae ) at various spatial scales in the Czech Republic

A taxonomic revision of Leucothoë (Ericaceae; Tribe Gaultherieae)
WALTER S. JUDD1, NORMAN C. MELVIN III2,3, KATHERINE WASELKOV4, AND
KATHLEEN A. KRON5
1Department of Biology, University of Florida, 220 Bartram Hall, P.O. Box 118525, Gainesville,FL 32611-8525, USA; e-mail: [email protected]
2Central National Technology Support Center, USDA-NRCS, 501 W Felix St., FWFC, Building23, Fort Worth, TX 76115, USA; e-mail: [email protected]
3Botanical Research Institute of Texas, 1700 University Drive, Fort Worth, TX 76107, USA4Department of Biology, Washington University in St. Louis, One Brookings Dr, Box 1137, St.Louis, MO 63130, USA; e-mail: [email protected]
5Department of Biology, Wake Forest University, Winston-Salem, NC 27109, USA; e-mail:[email protected]
Abstract. Leucothoë is here defined as a genus of five species (L. axillaris,L. fontanesiana, L. griffithiana, L. keiskei, and L. davisiae), occurring in easternAsia and western and eastern North America. Generic autapomorphies includeautumn-formed inflorescences with meiosis occurring in the autumn. Members ofthe Leucothoë clade, which are evergreen, are sister to the deciduousEubotryoides grayana, sharing the apomorphies of winged seeds with a distinctlyprotuberant hilum, the former of which is homoplasious. Leucothoë axillaris andL. fontanesiana, both of eastern North America, form a closely related speciespair, as do the eastern Asian L. griffithiana and L. keiskei. Leucothoë davisiae,occurring in western North America, is likely sister to the remaining species, andit differs in having erect branches and inflorescences, minutely bullate capsules,and in the lack of awns on the anthers. Species descriptions, nomenclaturalinformation, specimen citations, and eco-geographical characterizations for thesefive species are presented, along with an identification key.
Key Words: Leucothoë, Eubotryoides, Gaultherieae, Vaccinioideae, Ericaceae.
Recent phylogenetic analyses based on mor-phology (Waselkov & Judd, 2008), molecularsequence data from matK, ndhF and nrITS(Bush et al., 2009), and combined morpholog-ical and molecular data (Bush et al., 2010)indicate that Leucothoë D. Don, as broadly andtraditionally circumscribed (see Sleumer, 1959;Stevens, 1971; Melvin, 1980; Luteyn et al.,1996), is not monophyletic. However, the fiveevergreen species, i.e., L. axillaris (Lam.) D.Don, L. fontanesiana (Steud.) Sleumer, L.griffithiana C. B. Clarke, L. keiskei Miq., andL. davisiae Torr., form a strongly-supportedclade, and these species are recognized here asLeucothoë s.s. The deciduous species formerlyplaced in Leucothoë are now recognized asEubotrys Nutt., including Eubotrys racemosa
(L.) Nutt. and Eubotrys recurva (Buckley)Britton, and Eubotryoides (Nakai) Hara, in-cluding only Eubotryoides grayana (Maxim.)Hara (Waselkov & Judd, 2008; Bush et al.,2010; Judd et al., 2012).Autapomorphies of Leucothoë include mei-
osis occurring in the fall and autumn-formedinflorescences of the L. axillaris type, havingthe appearance of an elongated bud, theindividual floral units being covered by over-lapping, non-leafy bracts (Bush et al., 2010).Leucothoë is probably most closely related toEubotryoides grayana, and they share theapomorphies of winged seeds with a distinct-ly protuberant hilum, but we note that theseed-wing is sometimes lost (Waselkov &Judd, 2008; Lu et al., 2010). (Stevens et al.
Published online: 9 July 2013
Brittonia 65(4): 417–438 (2013), DOI 10.1007/s12228-012-9295-4ISSN: 0007-196X (print) ISSN: 1938-436X (electronic)© 2013, by The New York Botanical Garden Press, Bronx, NY 10458-5126 U.S.A.

[2004] included the extremely pheneticallydivergent E. grayana within Leucothoë.)Together, Leucothoë and Eubotryoides aresister to the remaining members of thetribe Gaultherieae (including. Eubotrys,Chamaedaphne Moench, Gaultheria L.,Diplycosia Blume, and Tepuia Camp).Leucothoë is easily distinguished from both
Eubotryoides and Eubotrys, with which it hasoften been united, by its evergreen leaves; theevergreen condition of Leucothoë is likely aretained plesiomorphy, with deciduous leaveshaving evolved in parallel in Eubotryoidesand Eubotrys (see Waselkov & Judd, 2008;Bush et al., 2010). Additionally, Leucothoëcan be distinguished from Eubotryoides by itsmore or less ovate to elliptic leaves (vs.elliptic to obovate leaves in Eubotryoides),axillary inflorescences with small, scale-likebracts (vs. inflorescences terminal or at leastappearing terminal due to the presence ofwell-developed leafy bracts in the proximalportion of the reproductive shoot), inflores-cence emerging in the autumn and of the L.axillaris type (see Waselkov & Judd, 2008;vs. inflorescences emerging in the spring),bracteoles (0.5–) 1–2.5 mm wide (vs. 0.1–0.8 mm wide), corolla white and cylindric tolong-urceolate, i.e., 4–25 mm long, withlength/width quotient of 1.3–2.4 (vs. corollagreenish, shortly and strongly urceolate, i.e.,3.5–5.5 mm long, with length/width quotientof 0.8–1.5), corolla lacking unicellular hairson the inner surface (vs. such hairs present),and stamens usually with awns of variouslengths (vs. lacking awns). Finally, the ovaryis papillose in Eubotryoides, while Leucothoëhas more or less smooth ovaries except for L.davisiae, which has ovaries with bullateepidermal cells.In addition to being evergreen, the species
of Leucothoë can be distinguished fromEubotrys by their inflorescences that areautumn-emerging, but have the appearanceof an elongated bud, the individual floralunits being covered by overlapping, nonleafybracts (vs. inflorescences that are autumn-emerging, but resemble a vegetative shoot,with long, somewhat leaf-like bracts notenclosing the floral buds, which are insteadprotected by the sepals), by the bracteolespositioned more or less basally on the floral
pedicel (vs. apical bracteoles), and by anat-ropous ovules (vs. campylotropous ovules).We also note that the species of Leucothoëhave Calluna-type pith (or heterogeneouspith, in L. davisiae), while the pith ishomogeneous in Eubotrys. Finally, the inflor-escences of most species of Leucothoë areusually numerous and positioned distally toall along the shoots of the previous season,but in Eubotrys the inflorescences are usuallyreduced to only one to three or four inflor-escences per shoot, and clustered at the distal-most nodes of the previous year’s branches.Within the Leucothoë clade, it is apparent
that the eastern North American species L.axillaris and L. fontanesiana form a closelyrelated pair, as do the Eastern Asian L.griffithiana and L. keiskei (Waselkov &Judd, 2008; Bush et al., 2009, 2010). The L.axillaris+L. fontanesiana clade is supportedby the synapomorphies of stigmas prominent-ly expanded and capitate to peltate and thepresence of a sclerified band of cells associ-ated with the leaf margin (the former uniquein the tribe, and the latter only occurring in afew species of Gaultheria; Waselkov & Judd,2008). Additional, more homoplasious syna-pomorphies of these two species includebracts lacking marginal unicellular hairs(and instead with a characteristic fringe ofgland-headed or eglandular multicellularhairs, or at least with several such hairs alongmargin) and the presence of a bare spot on theadaxial surface of the placenta, the latter alsooccurring in Chamaedaphne and Eubotrys(Palser, 1952; Waselkov & Judd, 2008). TheL. griffithiana+L. keiskei clade is supportedby the homoplasious synapomorphies of longacuminate leaf apices (a character thatevolved in parallel in L. fontanesiana), andleaves lacking unicellular hairs adaxially (acharacter that is also found in some popula-tions of Eubotryoides grayana) (Waselkov &Judd, 2008). These two species also differfrom L. axillaris and L. fontanesiana in theirlonger staminal awns, although this is prob-ably a retained plesiomorphy. The monophylyof both of these species pairs is alsosupported by matK, ndhF and nrITS nucleo-tide sequence data (Bush et al., 2009, 2010).The position of L. davisiae has varied in
phylogenetic analyses. The morphology-based
418 BRITTONIA [VOL 65

cladistic analysis of Waselkov and Judd (2008)placed this species sister to the L. axillaris+L.fontanesiana clade, while the molecular-basedanalysis of Bush et al. (2009) placed it as sisterto a L. griffithiana+L. fontanesiana clade (withL. axillaris and L. keiskei excluded from theanalysis). In the combined morphological andmolecular analysis of Bush et al. (2010), whichincluded all species of Leucothoë, L. davisiaewas hypothesized to be sister to the cladecontaining the remaining species of the genus(with bootstrap support of 94 %). Potentialsynapomorphies of the L. axillaris+L. fonta-nesiana+L. griffithiana+L. keiskei clade in thiscombined analysis include the characteristicarching pattern of their branches and theCalluna-type pith.Within Leucothoë, L. davisiae and L.
keiskei are each noteworthy due to theirnumerous autapomorphies. Leucothoë davi-siae has rigidly ascending inflorescences,staminal filaments that are merely papillate,anthers lacking awns, and minutely bullateepidermal cells of the ovary. In the analysis ofWaselkov and Judd (2008) the additionalautapomorphies of erect branches and hetero-geneous pith were suggested, but thesecharacter state placements were based onuse of Agarista populifolia (Lam.) Judd asthe outgroup, and given the cladogramtopology of Kron et al. (1999) it is moreparsimonious to consider these as retainedplesiomorphies. Leucothoë keiskei is distinc-tive because of its thick lamina, racemes witha few large flowers, elongate and subtubularcorollas that have elongate papillae on themargins of the lobes, and seeds with elongat-ed testa cells (Waselkov & Judd, 2008). Incontrast, L. fontanesiana and L. axillaris haveonly a few morphological autapomorphies,i.e., long-acuminate leaf apices and staminalfilaments that are merely papillose in L.fontanesiana, and bracteole margins lackingunicellular hairs in L. axillaris (Waselkov& Judd, 2008). Leucothoë griffithianaappears to have no morphological autapo-morphies, although it does possess molec-ular apomorphies (K. Kron, unpublisheddata). Morphological and molecular evi-dence, therefore, support the preliminaryhypothesis that the species of Leucothoërepresent cladospecies (i.e., they are hy-
pothesized to represent monophyletic groups,see Donoghue, 1985, and Mishler & Theriot,2000, for definitions of this phylogeneticapproach to species circumscription).Melvin (1980) conducted phenetic analyses
of the Leucothoë species, employing bothprincipal components analysis (PCA) andhierarchical agglomerative cluster analysis,using 91 morphological characters. Theseanalyses indicate that each of the five speciesdiscussed above is phenetically distinct, withthe partial exception of L. axillaris and L.fontanesiana, which are better separated inthe phenogram than in the PCA analysis.These phenetic analyses also indicate that L.axillaris and L. fontanesiana are very similarin appearance, as are L. griffithiana and L.keiskei, with L. davisiae being the mostmorphologically distinct species. Leucothoëdavisiae is well separated from the other fourspecies on the first principal component (seeMelvin, 1980, Figure III-2). These pheneticanalyses are congruent with our phylogeneticresults – they support the recognition of fivespecies within Leucothoë (employing thetraditionally used, phenetic or morphologicalspecies concept; see Judd, 1981, 2007) andalso suggest the close relationship of L.axillaris and L. fontanesiana, as well as thatof L. griffithiana and L. keiskei.We have closely studied the pattern of
variation within Leucothoë, based on anexamination of nearly 1350 herbarium speci-mens, as well as field studies of the NorthAmerican taxa, and this work lends support tothe explicit phenetic studies of Melvin(1980), again indicating that the recognitionof five species within the genus is appropri-ate, i.e., L. axillaris (occurring predominantlyin the coastal plain of the eastern UnitedStates, from southeastern Virginia toLouisiana), L. fontanesiana (occurring in thesouthern Appalachian mountains, fromVirginia to northern Georgia and northeasternAlabama, in the eastern United States), L.griffithiana (occurring in the Himalayanregion, from Bhutan and northwestern Indiato Burma, southwestern China, Laos andVietnam), L. keiskei (occurring in centralJapan), and L. davisiae (occurring fromcentral California northward into southernOregon, in the Sierra Nevada mountains and
419JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

the Coast Range). The geographical separa-tion of each of these species suggests thatthey are reproductively isolated, and thusshould also be considered biological species(Mayr, 1969).
Taxonomic history and treatment of fossils
The early-discovered temperate species,Leucothoë axillaris, was originally describedwithin Andromeda L. because these plantshave long-urceolate flowers with superiorovaries and capsular fruits. The process ofbreaking up this broadly circumscribedLinnaean genus began with D. Don (1834),who described Leucothoë (based on L. axil-laris), and several other genera now placed inthe tribes Gaultherieae or Lyonieae (see alsoKron et al., 2002). As similar evergreenspecies were discovered, they were added toLeucothoë as follows (using their currentspecific epithets): L. keiskei (in 1863), L.davisiae (in 1868), L. griffithiana (in 1882)and L. fontanesiana (in 1959; previouslyconsidered under the misapplied name L.catesbaei (Walter) A. Gray, 1856). Thesetemperate species were consistently placedwithin Leucothoë from the late 1800s to fairlyrecently (Hooker, 1876; Gray, 1856, 1886;Niedenzu, 1889; Drude, 1897; Sleumer,1959; Melvin, 1980; Luteyn et al., 1996).As noted above, most systematic botanists
also included the deciduous species (i.e.,Eubotryoides grayana, Eubotrys racemosa,and Eubotrys recurva) in a broadly circum-scribed Leucothoë, although their distinctive-ness was early noticed, and led to their genericsegregation into Eubotrys by Nuttall (1843),Britton and Brown (1913), and Small (1914,1933) and Eubotryoides by Hara (1935; basedon Leucothoë sect. Eubotryoides Nakai, 1922).See more detailed discussion of the taxonomicplacement of the species of Eubotrys in Judd etal. (2012).Finally, a group of tropical species was also
usually included in Leucothoë, as sect.Agastia DC. (Neotropical species) or sect.Agauria DC. (Paleotropical species)(Sleumer, 1959). The species in these sectionsare now recognized as not closely related tothe temperate species of Leucothoë, whetherdeciduous or evergreen, but instead aremembers of the tribe Lyonieae (Kron &
Judd, 1997; Kron et al., 1999, 2002) andplaced in Agarista. For more detailed dis-cussion of the species of Agarista see Judd(1979, 1984).The evergreen clade was further divided
into genera when Small (1914) recognizedthe new genus Oreocallis Small, containingonly O. davisiae. Oreocallis was recognizedbecause of its erect stems, distinctive inflo-rescence position (i.e., inflorescences bornetowards the end of the branches, instead ofdistributed along the shoots), and minordifferences in bracteole position and shapeof the calyx lobes. Leucothoë keiskei wastransferred by Honda (1949) to the newlyerected genus Paraleucothoë (Nakai) Honda,a name based on Leucothoë sect .Paraleucothoë Nakai (1922), based on theelongate pedicels, tubular corolla, pubescentstaminal filaments, and long, paired antherawns of this species. Our phylogeneticanalyses (Waselkov & Judd, 2008; Bush etal., 2009, 2010) indicate that the segregationof the phenetically distinctive L. davisiaeand L. keiskei would result in a non-monophyletic Leucothoë, and therefore, thegenera Oreocallis and Paraleucothoë areconsidered within Leucothoë.Likewise, our phylogenetic results (as well
as the phenetic analyses of Melvin, 1980)indicate that the recognition of several sec-tions within Leucothoë, i.e., the evergreenclade, cannot be justified. Within thesespecies, Sleumer (1959) treated L. axillarisand L. fontanesiana as sect. Leucothoë; L.griffithiana within sect. Oligarista Sleumer(including in this section also L. tonkinensisDop, which we consider conspecific with L.griffithiana); L. keiskei as sect. ParaleucothoëNakai; and L. davisiae as sect. AcranthesSleumer. Such extreme sectional splitting islargely redundant, because it merely mirrorsthe recognized species, except for the closelyrelated pair L. axillaris and L. fontanesiana,and obscures phylogenetic relationships, suchas the sister group relationship of L. griffithi-ana and L. keiskei. Wood (1961) and Melvin(1980) came to the same conclusion, andtreated all of these evergreen species within asingle supraspecific taxon: Leucothoë sect.Leucothoë. As noted above, the retention ofthe deciduous species within Leucothoë is notphylogenetically justifiable, and thus their
420 BRITTONIA [VOL 65

sect. Leucothoë becomes Leucothoë s.s., as herecircumscribed (with L. axillaris as the type).Consideration of the numerous fossil spe-
cies of Leucothoë is beyond the scope of thistreatment, but we note that Leucothoë prae-cox, represented by small 5-loculed capsules,is known from the Mastrichian (LateCretaceous) of central Europe (Knoblock &Mai 1986), indicating that the clade has along evolutionary history. It is unfortunatethat seeds are not known for this fossil, astheir morphology would strengthen its tax-onomic placement. Capsular fruits of generaof tribes Lyonieae Kron & Judd andGaultherieae Nied. (Kron et al., 2002) arequite similar, so some care in interpretationis required. Leucothoë lusatica Mai, forexample, based on similar capsules, whichis known from the lower to middleMiocene of Germany and Poland, has seedsthat are unwinged with elongated testa cells,and it has been compared (Kowalski, 2008)to Agarista populifolia (Lam.) Judd (whichKowalski incorrectly treated as a species ofLeucothoë). So L. lusatica may actually bea fossil species of Agarista (in the tribeLyonieae), and not a species of Leucothoë(in the Gaultherieae).An unidentified Leucothoë is reported (and
illustrated) by Van der Burgh (1978) from thePliocene of Germany, and it is represented by acapsule and seeds. These do appear to be well-placed within the genus based on their mor-phology. Thus, it is likely that Leucothoë onceoccurred in Europe, a pattern also seen in othergenera of Ericaceae, e.g., Agarista, Eubotrys,Lyonia, and Zenobia, that are now morerestricted in distribution (Gerrienne et al.,1999; Stevens et al., 2004; Friis et al., 2011).
Horticultural Importance
Leucothoë is known in the horticulturaltrade, both as cultivated native collectionsand by named cultivars. The first reference tocultivation is by Aiton (1789) who cited acollection of from North Carolina and pro-vided a common name: Notch’d leav’dAndromeda. Liberty H. Bailey (1924) in hisManual of Cultivated Plants, treated twospecies, L. axillaris and L. fontanesiana (thelatter considered as L. catesbaei) but in laterworks (Bailey & Bailey 1941, 1976) listed
four taxa, omitting only L. griffithiana. Inaddition, Rehder (1960) identified the time ofinitial horticultural introduction for four ofthe species: L. axillaris (1765), L. fontanesi-ana (1793), L. davisiae (1915), and L. keiskei(1915). We note that the related Eubotryoidesgrayana entered the horticultural trade in1890 (Rehder, 1960).The first named horticultural cultivar,
Leucothoë catesbaei (0 L. fontanesiana)‘Girard’s Rainbow’ was developed by theGirard’s Nursery in Geneva, Ohio in 1949; ithas variegated leaves and other variable leafcolorations (Wyman, 1960). Since that timenumerous cultivars of L. fontanesiana and L.axillaris have been released (plant patentnumbers: L. fontanesiana, 5229, 11088; L.axillaris, 10842, 17666; however, confusionin the horticultural trade between L. axillarisand L fontanesiana exists (Melvin, 1976).Leucothoë catesbaei (0 L. fontanesiana)‘Zeblid’ (plant patent number 5195) devel-oped by A. J. Zebehazy, Perry, Ohio in 1984is attributed as being a hybrid between L.fontanesiana (pistillate) and L. axillaris (sta-minate); however, the plant patent informa-tion indicates that the hybridization cannot besubstantiated since the breeder was unsure ifthe staminate plant was identified as L.axillaris or L. axillaris ‘Nana’, and thepistillate flower was not bagged or otherwiseprotected for possible contamination by otherpollen. Several named cultivars (‘Scarletta,’‘Lipsbolwi,’ and ‘Curly Red’) have beenderived as sports from the purported hybridL. fontanesiana x axillaris ‘Zeblid’.
Taxonomic treatment
Given that the Leucothoë s.s. clade com-prising the evergreen species is now stronglysupported as a distinct monophyletic groupwithin the tribe Gaultherieae (Waselkov &Judd, 2008; Bush et al., 2010), it is appropri-ate that the genus receive careful revisionarystudy. We have thus provided detailed speciesdescriptions, nomenclatural information,specimen citations, and eco-geographic char-acterizations for the five species ofLeucothoë, along with an identification key.In this revision we have employed thediagnostic (Nixon & Wheeler, 1990;Wheeler & Platnick, 2000), morphological-
421JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

phenetic (Judd, 1981, 2007), and phylogenet-ic-apomorphic (Donoghue, 1985; Mishler &Theriot, 2000) species concepts.Although our work is based on observa-
tions of nearly 1350 specimens (from 26different herbaria, see Acknowledgments)in order to save space we have cited inmost cases only a single specimen percounty for the three North Americanspecies, and only a single specimen fromeach geographical locality in Leucothoekeiskei and L. griffithiana.
Leucothoë D. Don, Edinburgh New Philos. J.7: 159. 1834. Type: L. axillaris (Lam.) D.Don (0 Andromeda axillaris Lam.).
Oreocallis Small, Flora North Amer. 29:58. 1914. Type: L. davisiae Torrey.
Paraleucothoë (Nakai) Honda, J. Japan. Bot.24: 29. 1949. Leucothoë sect.Paraleucothoë Nakai, Trees and Shrubs ofJapan, 1st ed., 127. 1922. Type: L. keiskeiMiq.
Evergreen shrubs with horizontal under-ground rhizomes and usually±arching todrooping branches (but erect in Leucothoëdavisiae); pith without diaphragms, of theCalluna-type (but heterogeneous in L. davi-siae); methyl-salicylate absent. Twigs±terete,with scattered gland-headed hairs (but theseabsent in L. keiskei), otherwise glabrous tomoderately unicellular-pubescent. Buds ovateto conical or shallowly hemispherical, usuallywith 4 to several imbricate scales (includingthe 2 lateral prophylls), glabrous, the marginsof the scales entire to erose, with unicellularand/or glandular hairs. Leaves alternate,exstipulate, simple, petiolate, coriaceous tostrongly coriaceous, flat, dark green andlustrous above and dull and lighter beneath,with scattered, multicellular, gland-headedhairs on abaxial surface, but these lackingadaxially, with unicellular hairs absent, onlyon the midvein of both surfaces, or occasion-ally also on secondary veins and evenscattered near base of lamina adaxially;margin clearly to obscurely serrate, with eachtooth associated with a gland-headed hair;venation brochidodromous with midveinprominent and tertiary veins reticulate; petiolevascular bundle unifacial. Inflorescences ax-illary, usually±pendulous racemes (but erect
racemes in L. davisiae), these usually severalper shoot, positioned only distally or extend-ing along the majority of the nodes of shootsof previous season, emerging in the latesummer, i.e., meiosis in the early fall, andexposed in winter, having the appearance ofan elongated bud, the individual floral unitsbeing covered by overlapping, persistent,nonleafy bracts, the flowers opening in thespring, with each flower perfect, pendulous,usually fragrant, and in the axil of a scale-like, ovate to ovate-triangular, and persistentbract; bracteoles 2, persistent, alternate toopposite, near base to mid-pedicel. Calyx of 5imbricate lobes, ± white at anthesis, bothsurfaces glabrous, articulate with the pedicel,persistent in fruit; corolla±long-urceolate (butvery elongate and±cylindrical-urceolate in L.keiskei), with 5 short, reflexed, triangular,imbricate lobes, white (sometimes pink-tinged in bud), glabrous on both surfaces;stamens 10, in 2 whorls, isomorphic, butslightly dimorphic in L. keiskei, inserted onreceptacle, the filaments flattened, basallydilated, ± straight, unicellular-pubescent ormerely papillose, without spurs, the anthers±ovoid, papillose, with 4 elongate to veryreduced awns (but these essentially lackingin L. davisiae) positioned dorso-apically,dehiscing by large, introrse-terminal, ellipticpores, with disintegration tissue on the backof each theca near bases of the awns, thepollen tricolporate, in tetrads, without viscinstrands; the nectariferous disk a slightly tostrongly lobed enlargement of the base of theovary wall; stigma truncate to expanded andcapitate or peltate, the style columnar, withfluted central canal, impressed into apex ofovary, slightly exserted, the ovary superior, 5-locular, glabrous and smooth (but cellsminutely bullate in L. davisiae); placentationaxile, positioned subapically on columella,but central in locule (due to impression ofstyle), extended into the locule, completelycovered by ovules (or with a bare spotadaxially in L. axillaris and L. fontanesiana),bearing numerous anatropous ovules, themegagametophyte development of thePolygonum-type, the megagametophyte withthe micropylar end slightly enlarged and theantipodals not dividing. Capsules loculicidal,erect, subglobose, glabrous, each valve with alongitudinal groove, the sutures pale, not
422 BRITTONIA [VOL 65

thickened, the placentae persistent on col-umella and extending into the locule.Seeds numerous, small, brown, the testathick to thin, composed of isodiametric to±elongated cells, wing absent, or presentand formed by flattened, bulging marginaltesta cells, with a distinctly protuberanthilum; embryo small, straight, white, with2 small cotyledons, central in cross section,nearest micropylar end, surrounded byfleshy endosperm. 2n022 (based on severalcounts from L. axillaris and L. fontanesi-ana; Rudenberg, 1963; Middleton &Wilcock, 1990).
Distribution.—Eastern Asia, and disjunctlyin eastern and western North America (fivespecies).Measurements.—All observations and
measurements (except plant height and flowercolor, which were taken from informationgiven on specimen labels or observed in thefield) included in the key and descriptionscome directly from dried herbarium material(see acknowledgments). We note that theinformation pertaining to the megagameto-phyte is derived from observations limited toLeucothoë axillaris and L. fontanesiana(Palser, 1952).
Key to species of Leucothoë
1. Plants with erect branches; inflorescences erect or ascending; ovary minutely bullate; anther±lacking awns; plantsof California and southern Oregon..…………………………………………………………………….1. L. davisiae.
1. Plants with±arching or drooping branches; inflorescences±pendulous; ovary±smooth; anthers with 4 awns, butthese sometimes inconspicuous; plants of Eastern North America or eastern Asia.2. Awns≤0.2 mm long; leaf margin associated with sclerified band of cells; stigma prominently capitate to
peltate; inflorescence bracts lacking unicellular hairs, but with multicellular, gland-headed or eglandular hairssparse and scattered to numerous and forming a marginal multicellular fringe; seeds unwinged, the testa cellsover the seed-body isodiametric; plants of eastern North America.3. Leaves obtuse or acute to short-acuminate, petiole 2–10 cm long; staminal filaments almost always
with at least a few unicellular hairs; calyx lobes ovate-triangular to ovate or widely so, 1.2–2.3 mm wide;plants of the coastal plain and lower piedmont.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. 2. L. axillaris
3. Leaves long-acuminate, petiole (5–)6–16 mm long; staminal filaments merely papillose; calyx lobesovate or ovate-triangular to oblong-triangular, 0.7–1.6 mm wide; plants of the Appalachian Mountains andupper piedmont.. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . 3. L. fontanesiana
2. Awns≥0.3 mm long; leaf margin lacking a sclerified band of cells; stigma truncate to slightly capitate;inflorescence bracts with a fringe of unicellular hairs, and multicellular, gland-headed or eglandular hairs absentor few and scattered along margin; seeds unwinged, with an inconspicuous wing, or with a conspicuous marginalwing formed from balloon-like, bulging cells, the testa cells over the seed-body isodiametric to±elongate; plantsof eastern Asia.4. Corolla±cylindrical to narrowly campanulate, 12–25 mm long; anther with awns 0.9–1.8 mm long;
pedicels 7.5–27 mm long; 2–12-flowered racemes; seeds with wing absent or inconspicuous (and composedof poorly developed and/or only a few bulging cells); plants of Japan. . . . . . . . . . . . . . . . . . . 4. L. keiskei
4. Corolla long-urceolate, 4–7 mm long; anther with awns 0.3–0.8 mm long; pedicels 3–7 mm long; 6–30-flowered racemes; seeds with conspicuous wing of numerous bulging and balloon-like marginal testa cells;plants of the Himalayan Mountains of Bhutan and NW India east to southern China, Laos and Vietnam.. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .. . . . . . . . . 5. L. griffithiana
1. Leucothoë davisiae Torr. in A. Gray, Proc.Amer. Acad. Arts 7: 400. 1868.Andromedadavisiae (Torr.) C. K. Schneid., Ill. Hand.Laubh. 2: 530, f. 346, a–d. 1911.Oreocallisdavisiae (Torr.) Small, North Amer. Flora29: 58. 1914. Type: U.S.A.. California.Nevada Co.: Near Eureka, Jul 1867, N. J.Davis 6 (lectotype, here designated: NY).Remaining syntypes: Nevada Co.: nearEureka, Sierra Nevada, 1867, N. J. Daviss.n. (NY, PH 2 sheets, online images seen,PH00015153, PH00015154).
Leucothoë cusickii M. E. Jones, Contr. W. Bot. 11: 1.1903. Type: U.S.A. Oregon. Siskiyou Mountains,on the border of California, 25–30mi from the coast,on the Illinois River, 30 Jul 1902, W. C. Cusick2925b (holotype: POM).
Shrub to 1.5 m tall, with erect or ascendingbranches, the new shoots arising from loweraxils of the branches to those just belowinflorescence-producing axils. Twigs withsparsely scattered multicellular gland-headedhairs, otherwise glabrous or with sparse tomoderate covering of short unicellular hairs.
423JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

Leaf blades oblong to elliptic or ovate, 1.5–8×0.8–3.3 cm, with length/width quotient of1.2–3.4; base rounded to obtuse (rarelyacute); apex acute to rounded-mucronate;adaxial surface with very sparse to sparseunicellular hairs on proximal portion, occa-sionally along entire length, of midvein, andoccasionally also with a few unicellular hairson secondary veins or even lamina surface;abaxial surface with scattered multicellulargland-headed hairs, these±persistent, other-wise glabrous or with very sparse to sparseunicellular hairs on proximal portion ofmidvein; margin serrate or serrulate, withthe serrations distributed all along margin,slightly revolute, especially proximally, toplane, and margin not associated with asclerified band of cells; petiole 2–8 mm long,glabrous or with sparse to moderate unicellu-lar hairs. Racemes axillary, erect or ascend-ing, several and produced from the uppermostaxils of each shoot, thus appearing paniculate,10–40 flowered, 4.5–14.5 cm long, theinflorescence axis glabrous or sparsely tomoderately covered with unicellular hairs;bracts ovate-triangular to broadly ovate, 1.5–3.3 mm long, the apex acute to rounded orshortly acuminate, the margin entire toundulate or erose, with very few unicellularhairs to unicellular-fringed, occasionally alsowith a few multicellular gland-headed (oreglandular) hairs. Flowers with pedicels 2.5–8 mm long, glabrous to sparsely pubescent;bracteoles opposite to alternate, inserted±basally to 1/3 of the way up pedicel, ovate-triangular, 1–1.8 mm long, the apex acute toobtuse or acuminate, the margin entire toundulate or erose, with very few unicellularhairs to unicellular-fringed, occasionally alsowith a few multicellular gland-headed (oreglandular) hairs. Calyx lobes triangular toovate-triangular, 2.2–4×1.3–1.8 mm, theapex acute to obtuse, the margin entire,usually with at least a few unicellular and/ormulticellular gland-headed hairs, occasionallyboth absent; corolla long-urceolate, 5.2–8.5×3–5 mm, glabrous abaxially; filaments 2.5–4 mm long, papillose, the anthers 1.3–1.8 mmlong, the awns±absent. Ovary minutely bul-late; style columnar; stigma capitate orlobate-capitate. Capsules 2.5–3 mm long, 5–6 mm in diam., each valve with a stronglyimpressed, median, longitudinal groove;
seeds ellipsoid to ovoid, 0.7–1.2 mm long,with a conspicuous wing formed by bulgingand balloon-like marginal testa cells; the testacells over the seed-body isodiametric toelongate.Distribution and ecology.—U.S.A., central
and northern California in the Sierra NevadaMountains from El Dorado County north intothe Klamath and Siskiyou Ranges and intoadjacent Oregon (Fig. 1). Occurring in bogsand acidic wetlands, along lake or streammargins, from 400–3000 m.Phenology. Flowering from May through
August.Common names.—Sierra laurel, sierra dog-
hobble, black laurel, western leucothoe.Illustrations.—Abrams (1951, fig. 3687),
Baker (1876, fig. 6247), Judd (2012, fig. onp. 705), Stuart and Sawyer (2001, fig. 127),and Tucker (2009, fig. on p. 507).
FIG. 1. Distribution of Leucothoë davisiae in Oregonand California.
424 BRITTONIA [VOL 65

Selected specimens examined. U.S.A. California:Butte Co.: Inskip, 1520 m, 20 Jul 1911, Eggleston 7334(GH). Calaveras Co.: 2.5 mi E of Creighton, 5200 ft, 25Oct 1935, Howden 215 (A). Del Norte Co.: Baker Flat,Jun 1939, Black 39-784 (NY). El Dorado Co.: streamnear head of Long Canyon, 2–3 mi W of Pollock Pines,3200 ft, 20 Jun 1943, Robbins 1190 (A). Fresno Co.:7000–8000 ft., Line Creek near Silver Fir Camp, justbelow Cedar Creek, Carter 84 (US). Madera Co.:Shuteye Mt, 7600 ft, 19 Jul 1914, Smiley 558 (GH).Mariposa Co.: summit of old Wawona Rd, overChowchilla Mt, E of Miami, 3 Oct 1934, Bacigalupi2332 (A); Yosemite National Park, vicinity of Sequoiaand Hazel Green, 6000 ft, Jul 1902, Hall & Babcock3404 (A). Nevada Co.: Castle Peak, near small lake,8300 ft, 10 Aug 1913, Smiley 475 (GH). Placer Co.:Robertson Flat region, Tahoe Forest, Aug 1927,Eggleston 21570 (NY). Plumas Co.: Without definitelocality, R. M. Austin s.n., 1876 (GH, K, online imageseen K000780220); Bear Lake, Mt Elwell region,7000 ft, 27 Jun 1934, Ewan 8188 (GH). Sierra Co.: Sof Mountain Mine, Sierra Buttes N of Sierra City,2015 m, 29 Jul 1964, Clausen 641121 (GH). SiskiyouCo.: Caribou Basin, Salmon-Trinity Alps, 6500–7500 ft,24 Jul 1937, Howell 13387 (A). Trinity Co.: E shore ofupper Canyon Creek Lake, 11 mi N of Dedrick, 11 Jul1939, Hitchcock & Martin 5409 (A). County unknown(within California): Sierra Nevada, 5000 ft, W. Lobb448 (K, online image seen K000780221), withoutdefinite locality, W. Lobb s.n. (K, online image seenK000780217) Oregon: Curry Co.: Pyramid Rock andGold Basin Trail, 1971, Leach 2926 (ORE).Josephine Co.: without definite locality, 26 Jun1950, Kruckeburg 1971 (WTU).
Leucothoë davisiae is quite uniform mor-phologically and (as discussed in the intro-duction) phenetically distinctive. It is easilydistinguished from the other species ofLeucothoë by the characters indicated in thekey.The Lobb specimens (see specimens ex-
amined) have sometimes been treated assyntypes, but should not be considered typespecimens because only the N. J. Davismaterial is cited in the protologue, eventhough A. Gray (1886) later alluded to thematerials used as “…collections of Lobb,Davis, and etc.” The Austin specimens,likewise, although sometimes considered tobe syntypes, are not cited in the protologue,and thus are not type material.
2. Leucothoë axillaris (Lam.) D. Don,Edinburgh New Philos. J. 17: 159. 1834.Andromeda axillaris Lam., Encycl. 1 (1):157. 1783. Andromeda walteri Willd.,Enum. Pl. 1: 453. 1809, nom. superfl.
Lyonia axillaris (Lam.) K. Koch,Dendrol. 2 (1): 124. 1872.Type:Amérique septentrionale, on la cultive auJardin du Roi, s.d., J. B. Lamarck s.n.(holotype: P n.v., but photo [Plate 900,Fig. 1, in Fernald, 1945] seen; isotype: P,online image seen P00715903).
Andromeda catesbaei Walt., Fl. Carol. 137. 1788.Andromeda spinulosa Pursh, Fl. Am. Sept. 1: 293.1814, nom. superfl. Leucothoë spinulosa (Pursh) G.Don, Gen. Syst. 3: 832. 1834. Leucothoë catesbaei(Walt.) A. Gray, Manual, ed. 2. 252. 1856. Lyoniacatesbaei (Walt.) K. Koch, Dendrol. 2 (1): 125.1872. Type: U.S.A.[South Carolina], T. Walter s.n.[6-H] (holotype: BM, in the Fraser/WalterHerbarium).Leucothoë platyphylla Small, Bull. Torrey Bot.Club 28: 290. 1901. Type: U.S.A. Georgia.Thomas Co.: Along or near the OchlockoneeRiver, near Thomasville, 28 May to 6 Jun 1895,J. K. Small s.n. (holotype: NY).Leucothoë axillaris (Lam.) D. Don var. ambigensFernald, Rhodora 47: 168. 1945. Type: U.S.A.Virginia. Nansemond Co.: Pine barrens, E of CoxLanding, S of South Quay, 10 May 1940, M. L.Fernald & B. Long 11887 (holotype: GH; isotype:MO, online image seen, MO-345562).
Shrub to 1(–2) m tall, with arching ordrooping branches, the new shoots arisingfrom lower to middle axils of branches.Twigs with scattered multicellular gland-headed hairs, and with sparse to moderatecovering of short to±elongate unicellularhairs, or rarely such hairs absent. Leaf bladesnarrowly to broadly ovate to elliptic, oblong,or obovate, (2.5–)4–14.2×1–5(–8)cm, withlength/width quotient of 1.1–5.5; base round-ed to obtuse or acute, broadly to narrowlycuneate; apex obtuse or acute to shortacuminate; adaxial surface with very sparseto sparse unicellular hairs on midvein, andsometimes also secondary veins and/or ex-treme basal portion of lamina near margin;abaxial surface with scattered multicellulargland-headed hairs, these±persistent, other-wise glabrous or with sparse unicellular hairson proximal portion of midvein; marginobscurely to clearly serrate, with serrationsall along margin to restricted to distal 1/3 ofblade, slightly revolute to plane, and marginassociated with a sclerified band of cells;petiole 2–10(–11) mm, with unicellular hairs.Racemes axillary, ± pendulous, usually sev-eral and positioned along the branches, 8–44
425JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

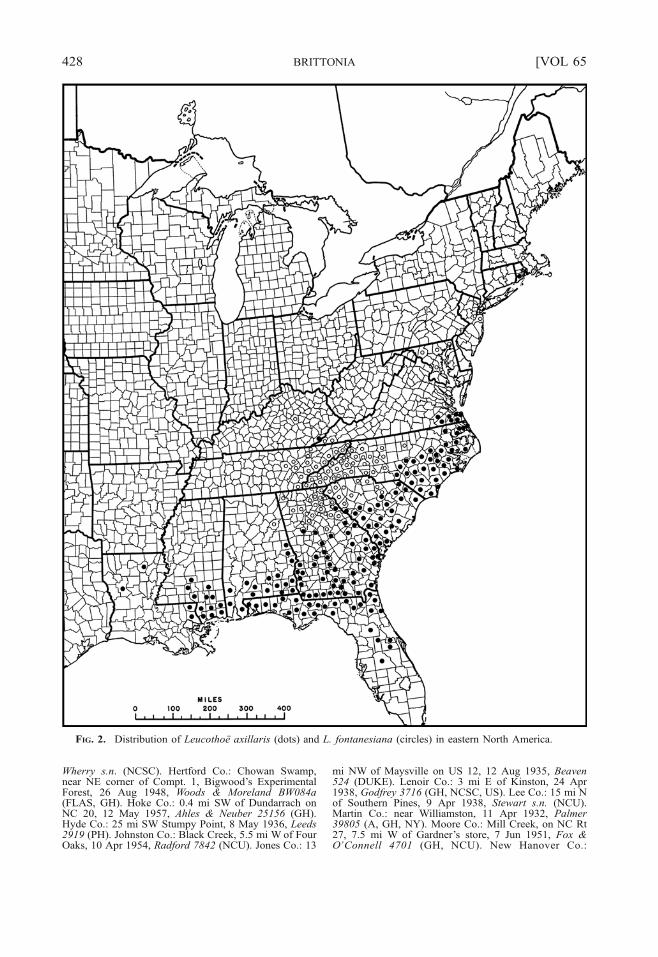
flowered, 0.9–5.5 cm long, the inflorescenceaxis sparsely to densely covered with unicel-lular hairs, rarely also with gland-headedhairs; bracts ovate-triangular, 1.5–4.1 mmlong, the apex acute to obtuse, the marginentire to undulate or erose, lacking unicellularhairs, usually with multicellular gland-headed(or eglandular) hairs, these only few, orsparse and scattered, to numerous and form-ing a multicellular-fringe. Flowers with ped-icels 1–7 mm long, sparsely to denselypubescent, rarely also with gland-headedhairs; bracteoles opposite to alternate,inserted±basally to 1/3 of the way up thepedicel, ovate-triangular, 1–2 mm long, theapex acute to obtuse, the margin entire toundulate or erose, lacking unicellular hairs,often with multicellular gland-headed (oreglandular) hairs, these sparse and scatteredto numerous and forming a fringe. Calyxlobes ovate to ovate triangular, often broadlyso, to rarely orbicular, 1.3–2.7×1.2–2.3 mm,the apex obtuse or acute to acuminate, themargins entire to erose, the undulations orprojections sometimes ending in a multicel-lular gland-headed hair, and such hairs one toseveral, also with a few to numerous unicel-lular hairs, or such hairs absent; corolla long-urceolate, (4.5–)5–8×2.5–5 mm, glabrousabaxially; filaments 2–3 mm long, papilloseand usually also with few to numerousunicellular hairs, especially on proximalportion, the anthers 0.8–1.6 mm long, with2 inconspicuous awns per theca, these 0.03–0.2 mm long, rarely±obsolete. Ovary smooth,the style columnar with stigma prominentlycapitate to peltate. Capsules 2.5–3.5 mmlong, 4–6 mm in diam., each valve with aslightly to moderately impressed, median,longitudinal groove; seeds angular, ovoid to±ellipsoid, 1–1.8 mm long, unwinged, thetesta cells over the seed-body isodiametric,flat to slightly bullate or distinctly bulging.2n022 (Middleton & Wilcock, 1990).Distribution and ecology.—U.S.A., on the
coastal plain and lower Piedmont, fromsoutheastern Virginia south to Florida, andthen west to Louisiana (Fig. 2). Occurring inbayheads, pocosins, swamp forests and wetacidic depressions, and along streams, from0–200 m. The species is naturalized in RhodeIsland (Angelo & Boufford, 2011). TheHarlan Co., Kentucky, population (Abbott et
al., 2001) likely represents plants persistingfrom cultivation.Phenology.—Flowering from March to May
(sometimes in early June).Common names.—Coastal doghobble, coastal
leucothoe, coast leucothoe, swamp dog-laurel.Illustrations.—Britton and Brown (1913,
fig. on p. 687), Catesby (1729-1747, vol. 2, y.52; see also Reveal 2009), Gleason (1952,fig. on p. 17), Holmgren (1998, fig. on p.188), Nelson (1996, fig. on p. 108). Photos ofthis species have been published in Taylor(1992, fig. on p. 78, 1998, fig. 288).
Selected specimens examined. U.S.A. Alabama:Baldwin Co.: Fairhope, 29 Mar 1924, Jack 3020 (A,US). Chambers Co.: Edge of Creek, 2 mi N StandingRock, 11 Jul 1969, Miller 103 (AUA). Coffee Co.: Creekbottoms by AL 189, 4.2 mi N of Kingston, 8 May 2000,Kral 89581 (VDB). Conecuh Co.: Rd on E side ofMurder Creek, 9 Oct 2008, Diamond 20263 (VDB).Covington Co.: Bottoms of Five Run Creek on AL 95, 7Apr 1997, MacDonald 10196 (BRIT). Crenshaw Co.:Along streams near Brantley, 1 Apr 1950, Norris 44(AUA). Dale Co.: Choctawhatchee River, W of Newton,6 Apr 1957, Godfrey 55479 (FSU, GH, NCSC, NY).Escambia Co.: Atmore, 15 Mar 1935, Blanton 225 (GH).Geneva Co.: 3.7 mi E of Geneva, 29 Mar 1953, Duncan& Hardin 14886 (GA, NCSC). Houston Co.: Creekbottoms 2 mi W of Cottonwood, 20 Aug 1981, Kral67760 (VDB). Henry Co.: Hidden Falls Creek on dirt rdbetween AL 95 and Henry County Hwy 12 at NE cornerof Haleburg, 31 Mar 1988, Diamond 4331 (AUA). LeeCo.: Chewacla Park, 5 Apr 1963, Coats 133 (AUA).Macon Co.: Swamp S of Hwy 80, 28 Mar 1954, Rauschs.n. (AUA). Mobile Co.: Mobile, 18 Apr 1890, Mohr s.n.(A). Monroe Co.: Little River Park S of Uriah, 29 Mar1981, Kral 66787 (VDB). Pike Co.: banks of WhitewaterCreek, 5 mi ESE Springhill on AL 6, 30 Mar 1968, Kral29962 (VDB). Russell Co.: just S Seale along US 431,31 Mar 1967, Kral 28015 (VDB). Washington Co.:Magnolia Branch bottoms 10 mi SSW Chatom by Co. 1,26 Mar 1971, Kral 41812 (VDB). Florida: Baker Co.:E1/4 SW1/4 Sec 21, T1S, R20E, near S margin offloodplain of Middle Prong of St. Marys River, 13 May1977, Martin 1064 (FLAS); Glen St. Mary, 25 Apr 1940,West & Arnold s.n. (FLAS). Bay Co.: 14 mi W ofWewahitchka, near Sandy Creek, 24 Mar 1975,McDaniel 19789 (FLAS). Calhoun Co.: On ChipolaRiver at Scott Ferry, 21 Mar 1949, Hood 1636 (FLAS).Clay Co.: T6S, R24E, Sec 27, 1/4 mi E of Rt 21 on sandrd, along small creek feeding into Black Creek, May1983, Judd 3328 (FLAS, USF). Duval Co.: NearJacksonville, Apr & Nov, Curtiss 1690 (A, FLAS, GH).Escambia Co.: Barrineau Park, 26 Mar 1956, Godfrey54615 (FLAS, GH). Gadsden Co.: Ravine of CrookedCreek, N of FL Rt 270, 8 mi W of Greensboro, Sec 7,T2N, R6W, 6 Nov 1959, Ward 1634 (FLAS). HamiltonCo.: 30° 36.5′N, 83° 3.5′W, 2.2 mi E of Jennings,floodplain S of Alapaha River, 7 Mar 2001, Abbott
426 BRITTONIA [VOL 65

14190 (FLAS). Holmes Co.: Along West Pittman Creek,0.35 mi ESE of FL 179A bridge over creek, Sec 23,T6N, R17W, 30° 54′ 44″N, 85° 54′ 37″W, 70–130 ft,8 Apr 1993, Orzell & Bridges 21433 (FLAS, USF).Jackson Co.: Half way between Cottondale and Alford,20 Aug 1940, Hocking s.n. (FLAS). Lake Co.: Creekalong Sulfur Run, 0.6 mi SE of Lake Jordan, 1.5 mi ESEof Cassia, Seminole State Forest, Sec 31, T18S, R29E,28° 52′ 36″N, 81° 26′ 37″W, 18 Oct 1994, Orzell &Bridges 23330 (FLAS, USF). Leon Co.: 2 mi E ofTallahassee, 28 Mar 1955, Godfrey 53113 (FLAS, GH,USF). Liberty Co.: Torreya State Park, 3 Jun 1953,Laessle s.n. (FLAS). Marion Co.: Mormon Branch ofJuniper Creek, 1.2 mi S of Juniper Creek bridge on Rt19, then 1/3 mi E on forest rd; Ocala National Forest, 6Apr 1986, Judd 5066 (FLAS); ibid., 15 Jun 2005,Waselkov 3 (FLAS). Nassau Co.: near Callahan, 31 Mar1931, Palmer 38291 (A, FSU, PH, US). Okaloosa Co.: 1mi S of Dorcas on FL Rt 393 (along Shoal River), Sec 2,T3N, R22W, 11 Sep 1979, Judd 2296 (FLAS, USF).Orange Co.: near Orlando, Mar 1932, Couch s.n. (NCU).Polk Co.: W edge of Reedy Creek Swamp near mouth ofHorse Creek, ca 2.6 mi E of center of Davenport, T27S,R27E, Sec. 12, 28° 9′ 15″ N, 81° 33′ 33″ W, 73–80 ft, 15Apr 1991, Orzell & Bridges 16370 (USF). St. Johns Co.:Without definite locality, 14 Apr 1879, Smith s.n. (US).Santa Rosa Co.: 4 mi E of Berrydale, N side of FL Rt 4,on W side of East Fork Big Coldwater Creek, Sec 5,T4N, R27W, 11 Sep 1979, Judd 2306 (FLAS). SeminoleCo.: 6 mi N of Longwood, T20S, R29E, Sec. 15, 7 Oct1998, Grey s.n. (USF, online image seen). Wakulla Co.:W side of Black Creek, on N side of FL 267, 0.8 mi SEof jct with FL 373 at Hilliardville, Sec 25, T2S, R2W,30° 16′ 44″N, 84° 23′ 11″W, 30 Mar 1995, Bridges23515 (FLAS, USF). Walton Co.: Near Alaqua Creek, atjct of creek and FL 282, 4 mi NE of Portland, Sec 30,T1N, R19W, 20 Jun 1967, Smith 1523 (FLAS). Georgia:Bacon Co.: USGS Crawley Quad., UTM 17363773 E,3479289 N (WGS84/NAD83); 8 mi S Alma jct US 1 andGA 32, 18 Aug 1988, Carter 7412 (VDB). Baldwin Co.:SE of Millegeville, 30 Apr 1942, Duncan 309 (GA).Bibb Co.: Seven Bridges, 6 Apr 1898, Andrews 569(AUA). Brooks Co.: Along Rt 94 SE of Morven and 8.9mi NW of Little River, 9 Sep 1973, Judd 279 (MSC).Bryan Co.: SE of Blitchton, 20 Apr 1937, Pryon &McVaugh 1371 (GA). Bulloch Co.: 6 mi ESE ofStatesboro, De Loach place, 5 Jul 1936, Wherry s.n.(GH). Chatham Co.: 4 mi WSW of Savannah, 23 Aug1953, Duncan 17275 (GA). Clay Co.: Ft Gaines, 27 Dec1952, Wigginton s.n. (FLAS). Cook Co.: along MorrisonMill Creek, 15.6 mi. SW of Adel, 20 July 1965,Faircloth 2436 (VSC). Decatur Co.: 3 mi NE ofFaceville, 31 Mar 1947, Thorne 2559 (GH). Early Co.:1 mi E of Blakely, 17 Nov 1947, Thorne 75264 (PH).Echols Co.: Branch Swamp, Co. rd E of Howell, 2 mi Wof Alpaha River, 22 Jun 1966, Bozeman 4689 (NCU).Effingham Co.: between Gayton and Springfield, 22 Jun1901, Harper 935 (GH). Elbert Co.: Deep Creek, 8.9 miW 2°N of Elberton, 17 Mar 1939, Duncan 11677 (GA).Emanuel Co.: swamp of Little Ohoopee River, 5 Apr1904, Harper 2093 (A, NY, US). Fayette Co.: W ofFayetteville, 24 Feb 1939, Henry 1429 (PH). Grady Co.:5 mi W of Cairo, 20 Mar 1938, Pryon & McVaugh 2199(GA, PH). Houston Co.: 3.1 mi N of Unadilla, 4 Sep1954, Hardin & Duncan 18169 (GA). Irwin Co.: 3 mi
SW of Irwinville, 19 Mar 1938, Pryon & McVaugh 2166(GA). Jefferson Co.: Duhart Creek 1 mi SW of Stapleton,20 Jun 1941, Duncan 3367 (GH). Liberty Co.: Ludovia,22 Mar 1910, Smith 2830 (F). Lanier Co.: bluff along theAlapaha River at Union Church, 2.6 mi SE of Lanier, 30July 1965, Faircloth 2586 (VSC). Lowndes Co.: ClyattCreek, 0.2 mi W of Clyattsville, 15 Apr 1964, Cotrans.n. (MISS). Macon Co.: Flint River, 4 Apr 1936, Pryon& McVaugh 487 (GA). McDuffie Co.: Brier Creek, ca 9mi S of Thompson, 2 May 1933, Leeds 2243 (DUKE,PH). Pulaski Co.: E side of Ocmubee River E ofHawkinsville, 4 Apr 1966, Duncan 22593 (GA).Quitman Co.: N of Georgetown, 9 Aug 1953, Duncan17174 (GA). Randolph Co.: 8 mi S of Cuthbert, 5 Apr1950, Hansen 2047 (GA). Richmond Co.: near Augusta,14 Apr 1900, Sargent s.n. (A). Screven Co.: near BlueSprings, 24 Apr 1940, Eyles 6882 (GH). Stewart Co.:Pataula Creek and just E of US Rt 27, 0.9 mi N ofRandolph Co line, 29 Apr 1984, Garland 250 (FLAS).Tattnall Co.: Without definite locality, 1923, HarveyE1575 (FLAS). Thomas Co.: Ochlocknee River, 6 Jun1895, Small s.n. (GH). Tift Co.: Along Rough Creek, 12mi. NE of Tifton, 24 Aug 1965, Faircloth 2832 (VSC).Ware Co.: Waycross Georgia, Apr 1897, Williamson s.n.(PH). Wayne Co.: SE of Jessup, 11 Apr 1942, Duncan4679 (MISS, TENN). Kentucky: Harlan Co.: 3.1 mi SSEfrom Big Laurel on KY 221 at jct with KY 510, 5 May2001, Thompson 01-51 (BEREA, scanned image seen).Louisiana: Natchitoches Parish: Briarwood near LA 9and Readhimer, 6 Jun 1969, Thomas 13859 (NLU). StTammany Parish: 4 mi NNW of Folsom, 31 Jan 1966,Thieret 21845 (DUKE, LAF). Washington Parish: besidePushepatapa Creek and county rd, Sec 12, T2S, R13E, 9May 1979, Thomas 18522 (FLAS, GH). Mississippi:Forrest Co.: Providence Creek, 2 mi NW of Eatonville,25 Jul 1956, Ray 7157 (FLAS). George Co.: Wyatt Hills,swamp of Black Creek, 25 mi E of Lucedale, 14 Mar1957, Ray 7764 (FLAS). Hancock Co.: creek bottoms,31 Oct 1954, Demaree 36305 (LAF). Harrison Co.:Canal Rd, 4 mi N of Hwy I-10, 13 Apr 1979, Darwin &Sundell 1331 (FLAS). Jackson Co.: 1 mi S of Rice Bluff,Vancleave, Ocean Springs, 28 Feb 1954, Demaree 34720(GH). Jones Co.: R13W, T7N, Sec 33, Lear River W ofMoselle, 11 Apr 1966, Jones 4472 (MISS). LawrenceCo.: 2 mi N of Monticello, Hwy 27, 6 Jul 1966, Jones7779 (MISS). Marion Co.: Columbia, 30 Mar 1966,Jones 3484 (MISS). Pearl River Co.: Hibolochitta Creek,8 mi W of Poplarville, 28 Feb 1927, Coville 97 (GH).Perry Co.: Janice rec. area 1 mi S of Janice at BlackCreek, 7 Jun 1967, Jones 13978 (MISS). Simpson Co.:Saratoga, 1 Apr 1903, Tracy 8653 (GH, NY). Stone Co.:S of Perkinston, 30 Apr 1966, Jones 4967 (MISS).North Carolina: Beaufort Co.: Bayview, 25 May 1947,Fox 242 (GA, NCSC). Bladen Co.: 2.75 mi NE ofDublin, 12 Apr 1953, Radford 6847 (GH). BrunswickCo.: Hwy 1 S of Exum, 26 Apr 1958, Bell 11591 (NCU).Camden Co.: 5 mi S of South Mills, 16 Jun 1927,Wiegand & Manning 2372 (GH). Carteret Co.: GatesCreek, S of Newport, 6 Aug 1949, Godfrey 49822 (FSU,NCSC). Craven Co.: 3 mi W of New Bern, 24 Apr 1938,Godfrey 3727 (GH). Columbus Co.: NE of Tabor City, 1Dec 1961, Hardin 2393 (NCSC). Cumberland Co.: 3 miSE of Fayetteville, 24 Mar 1954, Rossbach s.n. (FLAS).Currituck Co.: Mayock, 9 Apr 1929, Bagley s.n.(NCSC). Duplin Co.: N of Calypso, 15 Apr 1933, Fogg5469 (PH). Harnett Co.: 2 mi E of Erwin, 2 Jul 1936,
427JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]
Wherry s.n. (NCSC). Hertford Co.: Chowan Swamp,near NE corner of Compt. 1, Bigwood’s ExperimentalForest, 26 Aug 1948, Woods & Moreland BW084a(FLAS, GH). Hoke Co.: 0.4 mi SW of Dundarrach onNC 20, 12 May 1957, Ahles & Neuber 25156 (GH).Hyde Co.: 25 mi SW Stumpy Point, 8 May 1936, Leeds2919 (PH). Johnston Co.: Black Creek, 5.5 mi W of FourOaks, 10 Apr 1954, Radford 7842 (NCU). Jones Co.: 13
mi NW of Maysville on US 12, 12 Aug 1935, Beaven524 (DUKE). Lenoir Co.: 3 mi E of Kinston, 24 Apr1938, Godfrey 3716 (GH, NCSC, US). Lee Co.: 15 mi Nof Southern Pines, 9 Apr 1938, Stewart s.n. (NCU).Martin Co.: near Williamston, 11 Apr 1932, Palmer39805 (A, GH, NY). Moore Co.: Mill Creek, on NC Rt27, 7.5 mi W of Gardner’s store, 7 Jun 1951, Fox &O’Connell 4701 (GH, NCU). New Hanover Co.:
FIG. 2. Distribution of Leucothoë axillaris (dots) and L. fontanesiana (circles) in eastern North America.
428 BRITTONIA [VOL 65

Wilmington, 2 Apr 1880, Sargent s.n. (A). Onslow Co.:Cowhorn Creek, 13 Apr 1948, Woods s.n. (FLAS, GH).Pasquotank Co.: Dismal Swamp, 17 Dec 1948, Shearins.n. (NCSC). Pender Co.: 6 mi S of Player, 6 Feb 1956,Henry 6610 (PH). Perquimans Co.: Moist woods, Apr1932, Glasson s.n. (DUKE). Pitt Co.: E of ECC campushwy to Greenville TV station, 8 Apr 1956, Leonard s.n.(NCU). Richmond Co.: Hoffman, 1928, Roller s.n.(NCSC). Robeson Co.: 3.4 mi NW of Parkton, nearcounty line, 1 May 1959, Britt 2887 (GA). Sampson Co.:9 mi N of Salemburg on Rt 242, 22 Apr 1957, Wilbur5208 (DUKE). Washington Co.: near Roper, 8 May1938, Godfrey 3898 (GH). Wayne Co.: jct of NC 55 andrural rd 1937, 3 mi E of Mt. Olive, 3 May 1965, Barwick72 (AUA). Wilson Co.: 2 mi NE of Kenly, 11 Mar 1956,Bell 1573 (NCU). South Carolina: Aiken Co.: Flat RockBranch on W side of Flat Rock Pond, 1.75 mi NE of jctS-33 and S-144, N side of Graniteville, 83 m, 13 Apr1995, Nelson 16331 (FLAS). Barnwell Co.: 1.2 mi W ofjct Co. 217 & US 78 on 217 near Rosemary Creek, SWof Williston, 28 Oct 1961, Ahles & Crutchfield 55753(NCU). Bamburg Co.: 0.3 mi SW of jct Co. 26 & LittleSalkehatchie River, SW of Denmark, 6 Apr 1957 Ahles& Haesloop 22194 (NCU). Beaufort Co.: 6.5 mi S ofWashington Co. line on Rt 32, 17 Jun 1947, Wood &Clement 6946 (GH). Berkeley Co.: Daisy Swamp, 0.5 miN of Rt 2 near Summerville, 28 Jun 1945, Hunt 3280(NY). Calhoun Co.: 2 mi N of St. Mathews, then W offCo. 24 on private rd, 31, Mar, 1957, Ahles & Haesloop21746 (NCU). Clarendon Co.: Black River Swamp, 5 miE Manning, 15 May 1914, Stone 15 (PH). Colleton Co.:2.2 mi NE of Hampton Co. line, 5 Apr 1956, Ahles &Bell 10513 (NCU). Darlington Co.: Darlington, 22 Nov1919, Harbison 34 (A). Dillon Co.: 7.8 mi SW ofKemper on SC 41, 18 Apr 1957, Ahles & Ramseur23313 (NCU). Dorchester Co.: Summerville, 25 Mar1922, Hunnewell 8175 (GH). Edgefield Co.: NE of Rt230, 0.3 mi NW of Aiken Co. line, 200-250 m, 5 Apr1990, Hill 21383 (GH). Florence Co.: Tributary of LakeSwamp, 5 mi E of Lake City, 24 Apr 1924, Coker s.n.(NCU). Hampton Co.: 4.1 mi NNE of Early Branch onCo. 13, 1.4 mi S of Co. 42 & Co. 13 jct, 11 Apr 1956,Ahles & Bell 12468 (NCU). Horry Co.: Myrtle Beach, 19Apr 1932, Weatherby & Griscom 16609 (A, GH). JasperCo.: 1 mi SW of Gillesonville on Co. 23, 2 Apr 1956,Bell 1673 (NCU). Kershaw Co.: Camden, 29 Dec 1921,McKelvey s.n. (A). Lexington Co.: North Fork of EdistoRiver SW of Steedman, 5 Apr 1957, Radford 20547(NCU). Marlboro Co.: 5 mi NNE of Wallace, 10 Aug1956, Radford 15568 (NCU). McCormick Co.: LittleRiver, 12 1/4 mi N55°W of McCormick, 24 Feb 1962,McComb 119 (GA). Orangeburg Co.: Goodland Creek, Nof S-132, N of Springfield, 10 Apr 1990, Nelson 8791(FLAS). Richland Co.: Columbia, 7 Apr 1936, Philsons.n. (GA). Saluda Co.: SC 932 1.4 mi S of RidgesSpring, 26 May 1957, Radford 23105 (NCU). SumterCo.: Sumter, 5 Dec 1914, Bartram 3742 (NCU).Virginia: Chesapeake Co.: Johnstown Rd and DardenAve, Great Bridge, 3 May 1969, Gates 8 (AUA). City ofSuffolk: trail along Feeder Ditch in the Great DismalSwamp Refuge, 4 Feb 2004, Bolin 04-01 (ODU). City ofVirginia Beach: swamp along North Landing River, 0.5mi SW if Creeds School and 2.3 mi NW of Creeds, 2 Jul1996, Fleming 11819 (WILLI). City of Chesapeake: Bog,near Johnstown Rd. and Darden Ave., Great Bridge,Gates 8 (WILLI). Isle of Wright Co.: S of Zuni, 15, Apr1975, Harville 12051 (NCU). Nansemond Co.: Nearlumber camp of Camp Lumber Co., Great Dismal Swamp,SE of Whitemarsh School, 19 Oct 1939, Fernald & Long11600 (GH, PH). Norfolk Co.: S of Yadkin, 21 Apr 1942,Fernald et al. 14213 (GH). Southampton Co.: Swamp ofNottoway River, Smith’s Ferry, 9 Apr 1938, Fernald &
Long 7935 (GH). Suffolk Co.: Dismal Swamp, 27 May1920, Trudell s.n. (A, FLAS, GH). Sussex Co.: BelowKello’s Dam, Lightwood Swamp, 13 Sep 1945, Fernald &Long 14974 (GH, NY, PH, US).
Several vegetative characters are conspicu-ously variable in Leucothoë axillaris. The leafshape varies from narrowly ovate to elliptic tovery broadly so, and the apex ranges fromacute, obtuse to shortly acuminate. Some leavesare conspicuously serrate, while in others theserrations are inconspicuous and often restrict-ed to the distal margin. This variation has led tothe description of taxonomic entities hereconsidered in synonymy, i.e., L. axillaris var.ambigens (Fernald, 1945) for plants character-ized by narrow leaves with acute apices (andsuch plants are most common in the extremenortheastern portion of the range; see Fernald,1945; Melvin, 1980, 1981), and L. platyphylla(Small, 1901) for plants with widely ovateleaves with obtuse to broadly and shortlyacuminate apices (which are most common inthe western and southern parts of the range).Needless to say, these features vary continu-ously and show only weak geographical corre-lation; thus they do not support recognition oftaxonomic entities within L. axillaris (see alsoMelvin, 1980, Fig. III-5).Leucothoë axillaris is closely related to L.
fontanesiana, and these species sometimeshave been recognized as conspecific, e.g.,Radford et al. (1968), who considered themonly at the varietal level. However, mostsystematists have treated them as specificallydistinct (e.g., Sleumer, 1959; Wood, 1961;Melvin, 1980, 1981; Luteyn et al., 1996;Tucker, 2009; Weakley, 2011). These speciescan be distinguished best by the character-istics given in the key (see above); we notethat the broader calyx lobes of L. axillarisalso tend to be blunter at the apex, but thischaracter is not as useful as the width, itself.The two species also differ in the number ofserrations per leaf (ca. 5 to 50 in L. axillaris vs.25 to 75 in L. fontanesiana; Green, 1963a), andinflorescence length (0.9 to 5.5 cm long with 8 to44 flowers in L. axillaris vs. 2 to 10 cm long with17 to 80 flowers in L. fontanesiana). The lattercharacter was used by Tucker (2009), along withthe shape and apex of the calyx lobes, todistinguish the two species, and we note thatthese features are not as reliable in differentiating
429JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

the species as are the characters employed in ourkey. As can be seen in the distribution map(Fig. 2), these species are allopatric, and are thuslikely reproductively isolated (although a puta-tive hybrid has been produced in cultivatedmaterial, i.e., the cultivar ‘Zeblid’).It is of interest that even in the original
description of the species, Lamarck (1783)noted that the plant was variable in leaf shapeand apex, stating that it typically had widelyelliptic leaves with abruptly acuminate apices,while other specimens had narrower leavesand acute apices, and these constituted hisunnamed “foliis lanceolatis” variety (see alsoFernald, 1945, who considered these narrowleaved plants as var. ambigens).The name Andromeda catesbaeiWalt. is one
of only 15 of the 403 species described byThomasWalter as new that are represented by aspecimen in the Fraser/Walter Herbariumwith alabel bearing the full name (genus and epithet)in Walter’s hand (see Ward, 2007). It is clearthat the specimen in this collection is theholotype and that this name is to be applied tothe coastal plain species Leucothoë axillaris;see also discussion under L. fontanesiana.
3. Leucothoë fontanesiana (Steud.) Sleumer,Bot. Jahrb. Syst. 78: 438. 1959.AndromedalanceolataDesf., Cat. Pl. Horti Paris. Ed. 3:398. 1829., nom illeg., non AndromedalanceolataWall. Asiat. Res. 13: 390. 1820.Andromeda fontanesiana Steud., Nomencl.Bot., ed 2, 1: 88. 1840. Leucothoë editorumFernald & B. G. Schub., Rhodora 50: 218.1948. Leucothoë axillaris var. editorum(Fernald & B. G. Schub.) H. E. Ahles, J.Elisha Mitchell Sci. Soc. 80: 173. 1964.Type: U.S.A. Without definite locality,[either L.C. or P.] Noisette s.n. (holotype:FI-W, xerographic copy at GH seen).
Leucothoë catesbaei (Walt.) A. Gray var. rollisoniiBean, Trees & Shrubs of the British Isles 2: 19. 1914.Neotype, here designated: U.S.A. Maryland.Montgomery Co.: Brookside Gardens, 1500Glenallan [Ave.], Wheaton, 9 May 1986, F. G.Meyer & P. M. Mazzeo 21344 (A).
Shrub to 1.5(–3) m tall, with arching ordrooping branches, the new shoots arisingfrom lower to middle axils of branches.
Twigs with scattered multicellular gland-headed hairs, and otherwise glabrous or withsparse to moderate covering of short to±elongate unicellular hairs. Leaf blades nar-rowly to broadly ovate to oblong or elliptic,(3–)4–15×(1–)1.5–5(–6) cm, with length/width quotient of 1.8–6.2; base cuneate torounded or acute; apex long-acuminate; ad-axial surface with very sparse to sparseunicellular hairs on midvein, and occasionallya few hairs on extreme basal portion oflamina near margin; abaxial surface withscattered multicellular gland-headed hairs,these±persistent, otherwise glabrous; marginobscurely to clearly and sharply serrate, withserrations all along margin to restricted todistal 3/4 of blade, slightly revolute to plane,and margin associated with a sclerified bandof cells; petiole (5–)6–16 mm, with unicellu-lar hairs, especially adaxially. Racemes axil-lary (rarely terminal, and then paniculate), ±pendulous, several and positioned±along thebranches, 17–80 flowered, (2–)3–10 cm long,the inflorescence axis sparsely to moderatelycovered with unicellular hairs, occasionallyalso with multicellular gland-headed hairs;bracts ovate-triangular, 1.5–4 mm long, theapex acute to obtuse, the margin entire toundulate or erose, lacking unicellular hairs,usually with multicellular gland-headed (oreglandular) hairs, these only few, or sparseand scattered, to numerous and forming amulticellular-fringe. Flowers with pedicels1.5–8 mm long, glabrous to densely pubes-cent, often also with multicellular gland-headed hairs; bracteoles opposite to alternate,inserted±basally to 1/4 of the way up thepedicel, ovate-triangular, 1–2.5 mm long, theapex acute to obtuse, the margin entire to±undulate, lacking unicellular hairs or some-times with a few such hairs, often withmulticellular gland-headed (or eglandular)hairs, these sparse and scattered to numerousand forming a multicellular-fringe. Calyxlobes ovate or ovate-triangular to oblongtriangular, 1.4–3×0.7–1.6 mm, the apex(obtuse-) acute to acuminate, the marginsentire to erose, with a few to numerousunicellular hairs, or such hairs absent; corollalong-urceolate, 5–8×2.5–5 mm, glabrousabaxially; filaments 1.8–3 mm long, merelypapillose (extremely rarely with a few unicel-lular hairs), the anthers 0.8–1.5 mm long,
430 BRITTONIA [VOL 65

with 2 inconspicuous awns per theca, these0.03–0.2 mm long, rarely±obsolete. Ovarysmooth, the style columnar with stigmaprominently capitate to peltate. Capsules2.2–3.5 mm long, 4–6 mm in diam., eachvalve with a slightly to moderately impressed,median, longitudinal groove; seeds angular,ovoid to ± ellipsoid, 0.8–2 mm long,unwinged, although the angles sometimesthin and appearing wing-like, the testa cellsover the seed-body isodiametric, flat toslightly bullate. 2n022 (Rudenberg, 1963;Middleton & Wilcock, 1990).Distribution and ecology.— U.S.A., in the
upper Piedmont and southern Appalachiansfrom Virginia south to northern Georgia andAlabama (Fig. 2), occurring in woods alongmountain streams and ravines, and moistslopes, from 150–2000 m. The species isnaturalized in Massachusetts (Nantucket Co.,see specimens examined; Norfolk Co.; Angelo& Boufford, 2011) and NewYork (Suffolk Co.;Greller & Clemants, 2001). Steve Clemants, onhis “New York Metropolitan Flora Project”webpage (nymf.bbg.org), also reported a visualrecord from Nassau Co., N.Y., as well as visualrecords (representing either persistence aftercultivation or naturalized occurrences) fromseveral counties in northeastern New Jersey,and Steve Glenn (personal comm., N.Y.Metropolitan Flora Project, has provided spec-imen label data documenting occurrence inMonmouth and Middlesex Cos., New Jersey,and Suffolk Co., New York. Steve Young(personal comm., N.Y. Natural HeritageProgram) also reports the species from NassauCo. (in Coffin Woods), New York. The reportfrom Madison Co., Kentucky (Abbott et al.,2001) likely represents plants persisting fromcultivation. Finally, the Maryland records(Fig. 2) probably also represent naturalizedpopulations, although Riefner (1979) consid-ered the Baltimore Co. record to represent anative occurrence.Phenology.—Flowering from April to June.Common names.—Mountain doghobble,
mountain dog-laurel, highland doghobble,drooping fetterbush, drooping leucothoe,switch-ivy.Illustrations.—Britton and Brown (1913, fig.
on p. 688. as L. catesbaei), Gleason (1952, fig.on p. 17, as L. editorum), Holmgren (1998, fig.on p. 188, as L. walteri), Small (1919, t. 151, as
L. catesbaei). Photos of the species have beenpublished in Green (1963b, pl. 4), Symonds(1963, fig. on p. 284, as L. catesbaei), Kron etal. (2002, figs. 99, 100), and Waselkov andJudd (2008, fig. 5).
Selected specimens examined. U.S.A. Alabama:Calhoun Co.: Jacksonville, 6 Apr 1956, Line J107(TENN). Georgia: Banks Co.: Tributary of MiddleBroad River near jct of Habersham & Stephens Cos.,29 Jun 1950, Duncan 11185 (GA). Bartow Co.: EtowahRiver 5 ¾ mi S 80° E of Cartersville, 17 Apr 1948,Duncan 8021 (GA, GH, MO, NCSC, NY, TENN, US).Cherokee Co.: Shoal Creek 1 2/3 mi W 5° N of Canton,2 May 1948, Duncan 8217 (GA). Clark Co.: Whitehall,17 Apr 1938 (FLAS). Dawson Co.: Amicalola Creek 6mi W of Dawsonville, 8 Apr 1936, Pyron & McVaugh450 (DUKE, GA). DeKalb Co.: Tributary of NancyCreek, 8.8 mi N 15° W of Decatur, 2 May 1952, Duncan13466 (NCSC). Elbert Co.: S bank of Broad River, S ofSR 172, 34° 09.294′N, 83° 04.811 W, 150-210 m, 25 Jun2005, Waselkov 5 (FLAS). Fannin Co.: Ocoee River nearBlueridge, 30 Jun 1899, Unknown Collector 1280 (US).Fulton Co.: Chattahoochee River near White WaterCreek, 21 Apr 1945, Henry 4031 (PH). Gwinnett Co.:Steep hillside, 24 Apr 1909, Allard 185 (US). HabershamCo.: Bottom of Tallulah Gorge, 14 Apr 1936, Eyles 1381(DUKE, GA). Hall Co.: Gainesville, 25 Apr 1916, Pessins.n. (GA). Hart Co.: Tugaloo River, 8 Apr 1941, Duncan4876 (GA). Jackson Co.: Tallassee Shoals, 15 mi NW ofAthens, 10 Aug 1941, Duncan 251 (GA). Lumpkin Co.:Creek banks, S slopes of Blood Mountain, 10 Apr 1976,Kral 57576 (VDB). Murray Co.: Cohutta WildlifeManagement Area, E of Cisco, Alaculsy Valley besideConasauga River, 7 May 1983, Coile 3514 (FLAS).Putnam Co.: 13 mi E of Eatonton, 20 Feb 1938, Pyron &McVaugh 2067 (PH). Rabun Co.: 2 mi W of Tiger(boro)on Tiger Creek Rd, E side of Glassy Mountain, 650 m,13 May 1982, Utech et al. 82-072 (GH). Stephens Co.:Panther Creek, 4 May 1963, Fitzgerald 166 (GA).Townes Co.: Near Young Harris, 1 May 1930, Harveys.n. (GA). White Co.: W bank of Chattahoochee River,26 May 1953, Duncan & Hardin 16372 (GA). WilkesCo.: Broad River, Anthony Shoals, 26 Aug 1939,Duncan 1621 (GA). Kentucky: Madison Co.: LowerDogfoot Springs, Berea College Forest, 1.6 mi S ofBighill and 0.2 mi S of Grant House entrance, 12 Jul1995, Thompson 95-407 (BEREA, scanned image seen).Maryland: Baltimore Co.: Along a stream valley, 0.5 mi.N from Dulaney Valley Rd. and Jarrettsville Pike,Gunpowder Falls State Park, 8 May 1977, Riefner77-187 (MARY). Calvert Co.: W of Lot 371, Scientist’sCliffs, Port Republic, 7 May 1968, Johnson s.n.(MARY). Frederick Co.: E side of Sugarloaf Mountain,27 Sep 1948, Roller 917 (US). Prince Georges Co.:Behind J. Enos Ray Elementary School, 25 Apr 1977,Brannigan 13 (MARY). Massachusetts: Nantucket Co.:Nantucket Island, 8 July 1964, MacKeever 806 (NEBC,NY). Norfolk Co.: Chestnut Run, 0.5 mi Wof Unquity Rd,Blue Hills, Milton, 10 Sep 1992, Wells & Palmer s.n.(NEBC). New Jersey: Middlesex Co.: Black Horse Lane,0.2 mi NE of Deans Lane, Apr 2001, Glenn 5404 (BLK,
431JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

n.v.). Monmouth Co.: Thompson Park, Oct 2003, Glenn8897 (BKL, n.v.). New York: Suffolk Co.: Rocky PointNRM area, SE corner, Aug 2008, Rockermann s.n. (BKL,n.v.).NorthCarolina:Alexandria Co.:Middle Little River,7 mi SW of Taylorsville, 15 Sep 1956, Radford 18146(NCU). Avery Co.: Linville, 11 Jul 1924, Hunnewell 9254(GH). Buncombe Co.: along NC Rt 191 at Bent Creek, 10mi SWofAshville, 30Apr 1941,McVaugh 5635 (A). BurkeCo.: Linville River below falls, 12 Jun 1972, Currie 663(NCSU). Caldwell Co.: Richland Valley, 31 Jul 1891, Small& Heller 111 (F). Catawba Co.: Catawba River W ofMooresville, 17 Apr 1949, Fox & Godfrey 2233 (GH,NCSC, US). Cherokee Co.: stream margins, 1 Jun 1963,Rogers 133 (AUA). Clay Co.: 1.8 mi W of Hayesville, 16Nov 1956, Radford & Ahles 13844 (NCU). Cleveland Co.:Broad River, 3.5 mi S of Boiling Springs, 12 Jul 1950, Fox3906 (FSU, NCSC). Davidson Co.: Hartley Mill siteYadkin River, 15 Jun 1914, Totten s.n. (NCU). ForsythCo.: Rich woods, 5 May 1940, Schallert s.n. (TENN).Gaston Co.: Crowders Creek, 1 mi E of Mt. View, 17 Sep1956, Ahles 18919 (NCU, NY). Graham Co.: Rt 28, alongWolf Creek, 26 May 1973, Fowler s.n. (FLAS). HaywoodCo.:W bank of Pigeon River, 9.5 mi S of Lake Logan and 3mi N of Beech Gap, 12 Jun 1964, Wilbur 7113 (DUKE,FSU, GA, LAF). Henderson Co.: Hendersonville, 5 mi outSaluda Rd, 8 Jul 1947, Hood 69 (FLAS). Iredell Co.:Statesville, Hyanis 4993 (NY). Jackson Co.: NearDillsboro, 22 May 1934, Palmer 42485 (A, MO). MaconCo.: Highlands Biological Station, Cashiers, 23 Jul 1975,Nelson 327 (FLAS). Madison Co.: Shut-in Creek, 2.7 miWof Hot Springs on US 25-70, and 2.5 mi S on Rd #1183, 29Apr 1967, Bozeman et al. 9081 (FLAS, FSU, GA, GH,NCU, NY, TENN, LAF). McDowell Co.: Mackey Creek,on US Rt 70, 5 mi E of Old Fort, 14 Apr 1949, Fox &Godfrey 2159 (GA, GH, MO, NCU, NCSC, TENN).Mitchell Co.: jct Roses Creek and North Tow River,Altapass, 28 Sep 1962, Harrison 522 (GH, NCSC).Orange Co.: E of bridge over Morgan’s Creek, 21 Apr1946, Bell 79 (NCU, NY). Polk Co.: Stream on N side ofTryon city limit, 26 Apr 1963, Hardin 2581 (GA, NCSC).Rowan Co.: South Yadkin River about 0.7 mi fromconfluence with Yadkin River, 29 Apr 1957, Horton 936(NCU, TENN). Rutherford Co.: 2 mi E of Lake Lure, 1May 1954, Blomquest 16367 (DUKE). Stanly Co.: AlongYadkin River, NE of Richfield, 6May 1951,Godfrey & Fox51135 (DUKE, GA, GH, MO, NCSC, TENN). Swain Co.:2 mi NWof Cherokee, 26 Apr 1969, Roberson 103 (AUA).Transylvania Co.: along US Rt 276, just N of Sliding Rock,2800 ft, 25 May 1982, Judd 3134 (FLAS). Watauga Co.:near Blowing Rock, 14 Aug 1890, Heller 112 (PH). WilkesCo.:Wilkesboro, Feb 1894,Cowles 2471 (US). Yadkin Co.:Winfrey’s Rock, 1 May 1940, Schallert s.n. (MO). YanceyCo.: 7 Mile Ridge, 4 Aug 1896, Leeds 2240 (DUKE, PH).South Carolina: Abbeville Co.: Little River just belowHwy 185, 15 Jul 1940, Ellison 639 (NCU). Anderson Co.:8 mi N of Pendleton, 20 Aug 1927, Wiegand & Manning2378 (GH). Cherokee Co.: 2.7 mi E of jct Rt 59 and SC Rt110 on 59, 13Apr 1957, Ahles 22753 (GA). Darlington Co.:Black Creek, SE of Darlington, 15 Jul 1940, Smith 574(NCU). Greenville Co.: Near Marietta, 28 Apr 1932,Weatherby 6152 (A, GH). Lancaster Co.: Catawba River,13.7 mi NNW of Lancaster, 21 Apr 1953, Hardin &Duncan 15613 (GA). Laurens Co.: N slope of Reedy Riverat BoydMill Pond, 12mi SWof Laurens, 12 Apr 1957, Bell
5574 (FLAS). Oconee Co.:W sideWest Village Creek, NWside Old Rocky Gap Rd, Mountain Rest, 1700 ft, 28 Apr1991, Hill 22184 (GH). Pickens Co.: SC Rt 11, E side ofLake Keowee, 23 Apr 1986, Hill 16614 (GH); entrance rdto Table Rock State Park, private inholding, 0.1 mi N of SCRt 11, 35° 01.265′N, 82° 41.963′W, 350 m, 26 Jun 2005,Waselkov 7 (FLAS). York Co.: 0.5 mi S of Catawba, 20 Jun1949, Wherry s.n. (PH). Tennessee: Blount Co.: MontvaleSprings, 5 May1934,Underwood 552 (TENN). Carter Co.:Froghead, Laurel Park, 21 Jun 1951, Barclay 15976(TENN). Claiborne Co.: S of Harrogate, W of PowellRiver Bridge, 20 Apr 1960, Sharp 26995 (TENN). CockeCo.: Without definite locality, 31 Aug 1935, Jennison 17(TENN). Cumberland Co.: Walders Ridge, 3 Oct 1889,Coffman 26 (US). Green Co.: S end of Horse Creek Rd, 19Jun 1957, Sherman & Sharp 22277 (TENN). Johnson Co.:NE of Cartersville above Watauga River, 23 Jul 1949, Sharp& Shanks 7234 (TENN). Knox Co.: without definite locality,Gaylon 20443 (TENN). Monroe Co.: Near Beech Gap inCherokee National Forest, 30 May 1940, Sharp 698 (TENN).Morgan Co.: Hwy 11, 7 mi SE of Wartburg, 9 Jan 1938,Jennison 4 (TENN). Polk Co.: Rt 68 near Farner, 17 Mar1955, Shanks 19045 (TENN). Roane Co.: Along BriggsChapel Rd, near Clifty Creek, 12 Apr 1998, McNeilus 98-94(A). Sevier Co: Pigeon River, along streams, 24 Apr 1927,Braun s.n. (US); along Little S slope of Mt Le Conte. 3000 ft,16 May 1943, Sharp 1723 (GH, MO). Unicoi Co.: BaldMountain near Erwin, 2 Oct 1931, Core s.n. (TENN). UnionCo.: near Gatlinburg, 20 Oct 1935, Daubenmire 1935-14(TENN).Washington Co.: JohnsonCity, 21 Jun 1879,Gray etal. 294 (GH). Virginia: Lee Co.: 3.9 mi NE of jct Rt 720 andRt 606 on 606, 17 May 1968, James 9952 (NCU). Scott Co.:0.9 mi E of jct of Joel Branch and Little Stony Creek, 2.9 miNNW of Dungannon, 2400 ft, 10 Jun 1979, Peake 538(FLAS).
Leucothoë fontanesiana is less variablethan the closely related L. axillaris, althoughit does vary in the size and shape of its leavesand extent of prominence of the marginalserrations (see description, also Green, 1963a;Melvin, 1980). The characters best differen-tiating the two species are discussed under L.axillaris.The species is frequently used as an
ornamental because of its evergreen foliageand arching branches with beautiful clustersof white flowers in the spring; severalcultivars are available, including variegatedand dwarf forms (Green, 1963b; Melvin,1981; Blazich et al., 1991). But despite itsattractiveness, it has had a confusing nomen-clatural history. We note that the nameLeucothoe catesbaei (Walt.) A. Gray wasmisapplied by Asa Gray and considered asthe name of L. fontanesiana in his manual(Gray, 1856). This error unfortunately hasbeen perpetuated for these montane plants in
432 BRITTONIA [VOL 65

the horticultural literature. The nameLeucothoe walteri N. C. Melvin (Melvin1978) also appears in the horticultural trade;this name is illegitimate and thus cannot beused (see Melvin, 1979). Finally, we note thatthe correct name of this species, L. fontanesi-ana, is based on the type of the nameAndromeda lanceolata Desf., which is a laterhomonym of Andromeda lanceolata Wall.The collector of the holotype of
Andromeda lanceolata is either L. C.Noisette, who collected in North America(as documented by some of his specimens atP), or his brother, P. Noisette, who was for atime Michaux’s assistant (Sleumer, 7 Oct1976 letter attached to sheet at GH withxerographic copy of holotype).
4. Leucothoë keiskei Miq. Ann. Mus. Bot.Lugduno-Batavum 1: 32 . 1863 .Paraleucothoë keiskei (Miq.) Honda, J.Jap. Bot. 24: 29. 1949. Type: Japan. Prov.Kusji, Keiske 903 (holotype: L).
Shrub to 1.5 m tall, often sprawling, witharching or drooping branches, the new shootsarising from lower axils of branches. Twigsglabrous. Leaf blades ovate or narrowly so,occasionally elliptic, 3.5–9.5×1.5–4 cm, withlength/width quotient of 2–4.7; base broadlycuneate to rounded; apex long acuminate;adaxial surface glabrous; abaxial surface withscattered multicellular gland-headed hairs,these persistent to deciduous, otherwise gla-brous; margin serrate, with serrations along±entire margin, revolute, especially proximally,to plane, and not associated with a sclerifiedband of cells; petiole 2–9 mm long, glabrous.Racemes usually axillary (occasionally termi-nal, and then usually paniculate), ± pendu-lous, several and positioned±along thebranches or restricted to the upper axils, 2–12 flowered, 0.8–7.2 cm long, the inflores-cence axis glabrous to moderately coveredwith unicellular hairs; bracts ovate-triangular,1–2 (–4.5) mm long, the apex rounded-mucronate to acuminate, the margin entire toundulate, with fringe of unicellular hairs andoccasionally also a few multicellular gland-headed (or eglandular) hairs, occasionallyintergrading with leaves (when inflorescencesterminal). Flowers with pedicels 7.5–27 mm
long, glabrous to sparsely pubescent; brac-teoles subopposite to alternate, inserted±basally to 1/4 of the way up the pedicel,ovate-triangular, 0.5–1.5 mm long, the apexrounded to acute; the margin±entire, withfringe of unicellular hairs, occasionally alsowith a few multicellular gland-headed (oreglandular) hairs. Calyx lobes triangular,ovate-triangular or ovate, 1.5–3.5×1.3–2 mm, the apex acute to obtuse or shortlyacuminate, the margins entire, with a fringeof unicellular hairs; corolla±cylindrical tonarrowly campanulate, 12–25×6–11 mm,glabrous abaxially, but the lobes often witha fringe of elongate papillae; filaments 7–12 mm long, papillose and with numerousunicellular hairs, the anthers 3-4 mm long,with 2 conspicuous awns per theca, these0.9–1.8 mm long. Ovary smooth, the stylecolumnar with stigma truncate. Capsules3–4 mm long, 6–7 mm in diam., eachvalve with a strongly impressed, median,longitudinal groove; seeds ovoid to ellip-soid, 0.8–1 mm long, with a thin wing orat least a few bulging marginal cells, orwing absent; the testa cells over the seed-body±elongated.Distribution and ecology.—Leucothoë
keiskei occurs in shaded, moist habitats,especially on wet rocky cliffs from 150–1700 m, throughout central and southernareas of Honshu Island, Japan (Fig. 3).Phenology.—Flowering from June through
August.Common name.—Iwa-nanten.Illustrations.—Makino (1956, Fig. 741),
Matsumura (1912, Fig. 34). A photo waspublished in Hayashi (1985, fig. on p. 586).
Selected specimens examined. JAPAN. Honshu:Gifu Pref.: without definite locality, 9 Aug 1925, Shiota2459 (A). Kanagawa Pref.: Mt. Futago, Hakone, 8 Sep1951, Mizushima 1499 (MO); Sagami, Hakone, Sakunais.n., 2 Jul 1908 (A). Mie Pref.: Locality unknown, Hiroe7270 (P); Itaya – Ookochi, Kiwa-cho, Minamimuro-gun,200-300 m, 17 Nov 1977, Mimoro et al. 4306 (MO);Uotobi, between Kotsu and Yadokoro, Miyama-cho,Kitamuro-gun, 25 Jun 1983, Seto 29715 (MO). KyotoPref.: Mt. Atago, 10 Nov 1950, Mizushima 830 (A). NaraPref.: Mt. Daifugen-dake, from Mt Wasamata in MtOhmine, Yoshino-gun, 1700 m, 12 Aug 1977, Koyama &Hotta 5401 (A); Koka, Totsukawa-mura, Yoshino-gun,150 m, 23 Oct 1996, Tsugaru & Takahashi 24271 (MO).Saitama Pref.: Chichibu Basin, Mt. Shiraiwa, Matsumotos.n., 2 Aug 1921 (A). Shizuoka Pref.: Tagata-gun,
433JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

Amagiyugashima-machi, Joren-no-taki Falls, 34° 52′N,138° 56′E, 280–350 m, 14 Sep 1994, Kobayashi 2702(A). Tokyo Pref.: Mt. Kawanori-yama [Okutama-machi],27 Jul 1948, Yamazaki & Ono 2504 (A). Wakayama Pref.:Kobiro-toge – Kogomori, Hong-cho, Higashimuro-gun, 13Oct 1977, Mimoro et al. 4104 (MO); Muju, Hongo-cho,Higashimuro-gun, 28 Jul 2003,Murakami 710 (MO); KuboDani, upstream Hirai River, Kozagawa Cho, HigashimuroGun, 33° 39′N, 135° 40′E, Ogata et al. TWTw-14209 (A);near Kita-yadori, Hashimoto-shi, along the Nyu-river,300 m, 15 Oct 1989, Takahashi et al 1354 (MO).Yamanashi Pref.: Nishiyatsushiro-gun, Shimobe-cho,Tojiro-Mt Kenashi-yama, 35° 25–26′N, 138° 33′E,1200 m, 22 May 1994, Kobayashi 2457 (A).
Leucothoë keiskei is limited to south-central Honshu and is the only species ofLeucothoë restricted to Japan, although wenote that the related Eubotryoides grayana isalso endemic to Japan (and is more broadlydistributed within these islands). The speciesis morphologically quite uniform, unlike theextremely variable E. grayana, in whichnumerous infraspecific taxa have been de-scribed (see taxonomic treatment in Melvin,1980). Leucothoe keiskei is easily distin-guished from the other species of Leucothoëby its cylindrical to narrowly campanulateflowers that are 12–25 mm long, elongatestaminal awns (i.e., 0.9–1.8 mm), and rela-tively long pedicels (i.e., 7.5–27 mm).The species is occasionally cultivated as
an ornamental because of its showy flow-ers and the often-bright red young archingshoots.
5. Leucothoë griffithiana C. B. Clarke inHook. f., Fl. Br. India 3: 460. 1882.Type: Bhutan. Towards Sanah, 6800 ft,
W. Griffith 3485 [02256] (holotype: K;isotypes: E, K, P, online image seenP00715885).
Pieris cavaleriei H Lév. & Vaniot, Bull. Soc.Bot. France 51: 292. 1904. Type: China.Guizhou: Kouy-tchéou, Pin-fa, precipices, 15Oct 1902, P. J. Cavalerie 780 (holotype: P;isotypes: A, frag. ex E, E, online image seenE00327713, L, online image seen L0007235).
Leucothoë tonkinensis Dop, Fl. Indo-Chine 3:730, f. 83. 1930. Type: Tonkin [Vietnam].Massif du Phan-si-Pan, 2500 m, 17 Aug 1926,M. E. Poilane 12990 (holotype: P, specimenwith accession P00715887; isotypes: L, onlineimage seen L0007238. P, online image seenP00715888).
Shrub to 4 m tall, often±scandent, witharching or drooping branches, the new shootsarising from lower axils of branches. Twigswith sparsely scattered multicellular gland-headed hairs, otherwise glabrous or withsparse to moderate covering of short unicel-lular hairs. Leaf blades ovate to elliptic, 5–16×2.1–6 cm, with length/width quotient of2.5–4.1; base broadly cuneate to rounded oracute; apex long acuminate; adaxial surfaceglabrous; abaxial surface with scattered mul-ticellular gland-headed hairs, these±persis-tent, otherwise glabrous; margin serrate,with serrations often widely spaced andrestricted to distal 1/3 to 2/3 of blade,revolute, especially proximally, and some ofthe teeth obscured, to plane, and margin notassociated with a sclerified band of cells;petiole 5–17 mm long, glabrous. Racemesaxillary, ± pendulous, several and positioned±along the branches, 6–30 flowered, 2–7 cmlong, the inflorescence axis sparsely to dense-ly covered with unicellular hairs; bractsovate-triangular, 2–4.5 mm long, the apexacuminate, the margin entire, with a fringe ofunicellular hairs, and occasionally also with afew multicellular gland-headed (or eglandu-lar) hairs. Flowers with pedicels 3–7 mmlong, glabrous to densely pubescent; brac-teoles opposite to alternate, inserted±basallyto near midpoint of pedicel, ovate-triangular,1–2 mm long, the apex acute to acuminate,the margin±entire, with a fringe of unicellularhairs, sometimes also with a few multicellulargland-headed (or eglandular) hairs. Calyxlobes triangular to ovate-triangular, 1.1–2×
FIG. 3. Distribution of Leucothoë keiskei in Japan.
434 BRITTONIA [VOL 65

1.3–2.5 mm, the apex acute to slightlyacuminate, the margins entire, with a fringeof unicellular hairs distally; corolla long-urceolate, 4–7×3–5 mm, glabrous abaxially;filaments 1.7–2.7 mm long, papillose andusually also with few to numerous unicellularhairs on proximal portion, the anthers 0.9–1.2 mm long, with 2±conspicuous awns pertheca, these 0.3–0.8 mm long. Ovary smooth,the style columnar with stigma truncate toslightly capitate. Capsules 2.7–3.5 mm long,4–6.5 mm in diam., each valve with a stronglyimpressed, median, longitudinal groove; seeds±ellipsoid, 1–1.5 mm long, with a conspicuouswing formed by bulging and balloon-likemarginal testa cells; the testa cells over theseed-body isodiametric to±elongated.Distribution and ecology.—Primarily in the
Himalayan Mountains of Bhutan, northwest-ern India, and Burma, to southwestern China(Xizang, Yunnan, Guizhou), Laos, andVietnam (Fig. 4); occurring in shaded orexposed, rocky slopes or ledges in damp,acidic soils. It is a component of the highelevation shrub belts dominated byRhododendron L. and also of Abies forests,from 1000–3000(–4000) m elevation.Phenology.—Flowering in April and May.Common name.—Jian-ji-mu-li-lu (China).Illustrations.—Academia Sinica (1974,
Figs. 4299, 4300), Fang and Stevens (2005,Fig. 646).
Selected specimens examined. BHUTAN:[Trashigang District], towards Sanah, 6800 ft, Feb 1836,Griffith 2256 (E online image seen E00438089, K).
INDIA. Arunachal Pradesh: Pange to Talle Valle,2400-2800 m, Panda 30844 (CAL, n.v.).
BURMA. Adung Valley, 6000 ft, 28 Apr 1931,Ward 9434 (A); ibid., 8000-9000 ft, 31 May 1931,Ward 9548 (A); North Triangle, 9 Nov 1953, Ward9250 (A).
CHINA. Xizang: Mt. Kenyichunpo and region ofChamputong, Salween – Irrawaddy watershed,10,000 ft, 1923, Rock 11520 (US); Mt. Kenichunpo,E and W slopes, Salwin and Irrawaddy Divide,11,500 ft, May-Jun 1932, Rock 22036 (A, US); MtKenichunpo, mountains of Champutong, upper SalwinRiver, 12,000 ft, Oct 1932, Rock 22479 (A). Yunnan:Without definite locality, Aug 1917, Forrest 15944(E, 3 sheets, epitypes); Nujiang Lisu Aut. Pref.,Gongsham Co., track from Qi Qi Nature ReserveStation towards Dong Shao Fang, 21 Sep 1997, LiHeng 9513 (MO); Shang-pa Hsien, 19 Oct 1933, H.T. Tsai 54467 (A); Chi-na-tung, Cham-pu-tung, Bar-roa-tung, 2800 m, C. W. Wang 67342 (A); Champu-tung, Bar-ru-lah, Salween-chu-kiang Divide, 3200 m,Oct 1935, C. W. Wang 67523 (A); Kiukiang Valley, Eof Monting, 2500 m, 13 Sep 1938, T. T. Yu 20207(A); Kiukiang Valley, Muchietu to Chiengen, 1600 m,10 Nov 1938, T. T. Yu 21027 (A).
LAOS. Without definite locality, Apr 1932, Kerr21029 (P).
Although Leucothoë griffithiana is wide-spread its overall morphology is not toovariable, although it does show interestingvariation in leaf characters. The form of theleaf base (i.e., broadly cuneate to rounded oracute) and number and prominence of the leafserrations often have been used to distinguishL. tonkinensis (Dop, 1930; Academia Sinica,1974; Fang & Stevens, 2005), but we find thepattern of variation to be overlapping andtherefore recognize only a single species (seealso Melvin, 1980, Fig. III-3).We note that Leucothoë griffithiana var.
sessilifolia C. Y. Wu & T. Z. Hsu is actuallyGaultheria brevistipes (C. Y. Wu & T. Z.Hsu) R. C. Fang (Fang, 1999).
An ambiguous name
Andromeda axillaris Lam. var. longifoliaPursh, Fl. Am. Sept. 1: 293. 1814.Leucothoë axillaris (Lam.) D. Don var.longifolia (Pursh) DC., Prodr. 7(2): 602.1839. Type: U.S.A. On the mountains ofVirginia to Georgia, F. T. Pursh, not seen.
Pursh’s very brief description of this taxon isnot adequate to clearly indicate whether he was
FIG. 4. Distribution of Leucothoë griffithiana ineastern Asia.
435JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

referring to Leucothoe axillaris or L. fontanesi-ana, but it does state that Andromeda axillarisvar. longifolia possesses “short spikes” (sic,short racemes). This calls the geographicalrange given in the protologue, i.e., “On themountains of Virginia to Georgia,” into ques-tion because the montane L. fontanesianausually has longer inflorescences (i.e., 3–10 cm) than the coastal L. axillaris (i.e., 0.9–5.5 cm). Therefore, Melvin (1980) suggestedthat perhaps Pursh transposed the habitats of hisA. axillaris var. longifolia with that of his A.spinulosa, which was stated by Pursh to becoastal. Our lack of confidence regarding theaccuracy of the stated geographical range ofvar. longifolia removes the only definitivecharacteristic that could be used to identify thisplant. Unfortunately, we have not been able tolocate the type of A. axillaris var. longifolia,and thus the identity of this name cannot bedetermined with certainty.
Acknowledgments
We thank the curators of A, AUA, BRIT,DUKE, FLAS, FSU, GA, LAF, LSU, MISS,MSC, MU, NEBC, NCSC, NCU, NLU, NY,ODU, ORE, PH, TENN, US, VDB, VSC,WILLI, and WTU for their loans of speci-mens, and Norris Williams and Kent Perkinsof the University of Florida Herbarium andAmanda Neill and Tiana Rehman of theherbarium of the Botanical Research Instituteof Texas for assistance with specimen pro-cessing. We thank Bruce Hansen for kindlymaking available digital images of all theUSF specimens of Leucothoe axillaris. Wealso thank Ralph Thompson for his assistancein tracking down two specimens at BEREA,Steve Glenn (BKL) for specimen records ofL. fontanesiana from New Jersey and NewYork; and Steve Young for informationregarding the distribution of Leucothoe fon-tanesiana in New York. We appreciateGretchen Ionta’s assistance with the distribu-tion maps, and we value the helpful com-ments of Daniel Atha and an anonymousreviewer on an earlier version of this paper.NCM thanks the administrators of the WillardSherman Turrell Herbarium Fund and theMiami University Graduate School Achieve-ment Fund for their support, and this revisionreflects, in part, work done as part of his
doctoral dissertation. WSJ thanks his wife,Beverly, for assistance in field work in theeastern United States. Finally, KAK and WSJacknowledge (and are grateful for) supportfrom the National Science Foundation (grants:DEB-0717724, DEB-0717711), Wake ForestUniversity, and the University of Florida.
Literature Cited
Abbott, J. R., R. L. Thompson & R. A. Gelis. 2001.Vascular plants new to Kentucky. Sida 19: 1199–1202.
Abrams, L. 1951. Illustrated flora of the Pacific states.Vol. 3. Stanford Univ. Press, Stanford, CA.
Academia Sinica. 1974. Iconographia CormophytorumSinicorum. Vol. 3.
Aiton, W. 1789. Hortus Kewensis, ed. 1. Vol. 2. GeorgeNicol, bookseller to his majesty, Pall Mall. London.
Angelo, R. & D. E. Boufford. 2011. Atlas of the flora ofNew England: Paeoniaceae to Ericaceae. Phyto-neuron 2011–53: 1–13+map pages 1–18.
Bailey, L. H. 1924. Manual of cultivated plants.MacMillian Co., New York.
——— & E. Z. Bailey. 1941. Hortus Second. MacMil-lian Co., New York.
——— & ———. 1976. Hortus Third. MacMillian Co.,New York.
Baker, J. G. 1876. Leucothoe davisiae. Curtis’s Botan-ical Magazine. Vol. 102. t. 6247.
Blazich, F. A., S. L. Warren, J. R. Acedo & R. O.Whitehead. 1991. Seed germination of Leucothoëfontanesiana as influenced by light and temperature.Journal of Environmental Horticulture 9: 72–75.
Britton, N. L. & A. Brown. 1913. An illustrated flora ofthe northern United States and Canada, ed. 2. part 2.Charles Scribner’s Sons, New York.
Bush, C. M., L. Lu, P. F. Fritsch, D. Li & K. A. Kron.2009. Phylogeny of Gaultherieae (Ericaceae: Vacci-nioideae) based on DNA sequence data from matK,ndhF, and nrITS. International Journal of PlantSciences 170: 355–364.
———, W. S. Judd, L. Frazier & K. A. Kron. 2010.The phylogeny of Leucothoë s.l. (Ericaceae: Vaccinioi-deae) based on morphological and molecular (ndhF,matK, and nrITS) data. Systematic Botany 35: 201–206.
Catesby, M. 1729-1747. The natural history of Carolina,Florida, and the Bahama Islands. Publ. by the author,London, U.K.
Don, D. 1834. An attempt at a new arrangement of theEricaceae. Edinburgh New Philosophical Journal 17:150–160.
Dop, P. 1930. Treatment of Leucothoë and description ofL. tonkinensis. In: H. Lecomte, H. Humbert & F.Gagnepain (eds.), Flore générale de l’Indo-Chine 3:729–730, f. 83. Masson et C. Ėditeurs, Paris.
Donoghue, M. J. 1985. A critique of the biologicalspecies concept and recommendations for a phyloge-netic alternative. Bryologist 88: 172–181.
Drude, O. 1897. Ericaceae. In: Engler, A. & K. Prantl(eds.), Die natürlichen Pflanzenfamilien, ed. 4, 1:115–165. Wilhelm Englemann, Leipzig.
436 BRITTONIA [VOL 65

Fang, R.-C. 1999. New taxa of Ericaceae from China.Novon 9: 162–178.
——— & P. F. Stevens. 2005. Leucothoë. Pp. 458–459.Fig. 646. In: Z. Wu & P. H. Raven (eds.), Flora ofChina, Vol. 14. Science Press, Beijing, and MissouriBotanical Garden Press, St. Louis.
Fernald, M. L. 1945. Botanical specialties of the SewardForest and adjacent areas of southeastern Virginia.Rhodora 47: 93–142, 149–182, 191–204. [Description ofLeucothoë axillaris var. ambigens on pp. 168–169.]
Friis, E. M., P. R. Crane & K. R. Pedersen. 2011.Early flowers and angiosperm evolution. CambridgeUniversity Press, Cambridge, U.K.
Gerrienne, P., H. Beekman, F. Damblon, H. Doutrelepont,M. Fairon-Demaret & T. Smith. 1999. Agaristoxylongarennicum Gerrienne et al., gen. et sp. nov., anarborescent Ericaceae from the Belgian Upper Paleo-cene: palaeoenvironmental implications. Review ofPalaeobotany and Palynology 104: 299–307.
Gleason, H. A. 1952. Britton and Brown illustrated floraof the northeastern United States and adjacentCanada. Vol. 3. New York Botanical Garden, Lancas-ter Press, Inc., Lancaster, PA.
Gray, A. 1856. A manual of botany of the northern United
States, ed. 2, Ivison, Blakeman, Taylor andCo., NewYork.———. 1886. Synoptical flora of North America. Vol. 2,
Part 1. Gamopetalae after Compositae. Ed. 2. Ivison,Blakeman, Taylor, & Co., New York.
Green, P. S. 1963a. Leucothoë axillaris and L. fonta-nesiana. Castanea 28: 93–100.
Green, P. S. 1963b. Leucothoë fontanesiana. Arnoldia23: 93–99.
Greller, A. M. & S. E. Clemants. 2001. Flora of WestHills Park, Suffolk County, New York, with consid-erations of provenance of some long-distance dis-juncts. Journal of the Torrey Botanical Society 128:76–89.
Hara, H. 1935. Eubotryoides. Journal of JapaneseBotany 11: 626–627.
Hayashi, Y. 1985. Woody plants of Japan. Yama-keiPubl. Co.. Tokyo, Japan.
Holmgren, N. H. 1998. Illustrated companion toGleason and Cronquist’s manual. New York Botan-ical Garden, Bronx, N.Y.
Honda, M. 1949. Paraleucothoë. Journal of JapaneseBotany 24: 29.
Hooker, J. D. 1876. Ericaceae. Pp. 577–604. In: G.Bentham & J. D. Hooker (eds.), Genera plantarum 2,part 2. Lovell Reeve and Co., London.
Judd, W. S. 1979. Generic relationships in the Androme-deae (Ericaceae). Journal of the Arnold Arboretum 60:477–503.
———. 1984. A taxonomic revision of the Americanspecies of Agarista (Ericaceae). Journal of the ArnoldArboretum 65: 225–342.
———. 1981. A monograph of Lyonia (Ericaceae).Journal of the Arnold Arboretum 62: 63–209, 315–436.
———. 2007. Revision of Miconia sect. Chaenopleura(Miconieae: Melastomataceae) in the Greater Antil-les. Systematic Botany Monographs 81: 1–235.
———. 2012. Leucothoë. Pp. 703, 705. In: Baldwin, B.G. (convening ed.), The Jepson manual: Vascularplants of California, 2nd ed. University of CaliforniaPress, Berkeley, CA.
———, N. C. Melvin, K. Waselkov & K. A. Kron.2012. Taxonomic revision of Eubotrys (Ericaceae;Gaultherieae). Brittonia 64: 165–178.
Knoblock, E. & D. H. Mai. 1986. Monographie derFrüchte und Samen in der Kreide von Mitteleuropa.Rozpravy Ústredního Ústavu Geologické no, Praha47: 1–219.
Kowalski, R. 2008. Contribution to the knowledge ofthe Middle Miocene flora from Konin Brown CoalBasin (Central Poland). Acta Palaeobotanica 48: 277–299.
Kron, K. A. & W. S. Judd. 1997. Systematics of theLyonia group (Andromedeae, Ericaceae) and the useof species as terminals in higher-level cladisticanalysis. Systematic Botany 22: 479–492.
———, ——— & D. M. Crayn. 1999. Phylogeneticanalyses of Andromedeae (Ericaceae subfam. Vacci-nioideae). American Journal of Botany 86: 1290–1300.
———, ———, P. F. Stevens, D. M. Crayn, A. A.Anderberg, P. A. Gadek, C. J. Quinn & J. L.Luteyn. 2002. Phylogenetic classification of Erica-ceae: molecular and morphological evidence. TheBotanical Review 68: 335–423.
Lamarck, J.-B. 1783. Description of Andromeda axil-laris. Encyclopédie Méthodique: Botanique 1(1):157.
Lu, L., P. W. Fritsch, C. M. Bush, L.-N. Dong, H.Wang & D.-Z. Li. 2010. Systematic implications ofseed coat diversity in Gaultherieae (Ericaceae). Bot.J. Linn. Soc. 162: 477–495.
Luteyn, J. L., W. S. Judd, S. P. Vander Kloet, L. J.Dorr, G. D. Wallace, K. A. Kron, P. F. Stevens &S. E. Clemants. 1996. Ericaceae of the southeasternUnited States. Castanea 61: 101–144.
Makino, T. 1956. An illustrated flora of Japan. EnlargedEd. Hokuryukan Co., Tokyo.
Matsumura, J. 1912. Icones Plantarum Koisikavensis.Vol. 1. t. 34.
Mayr, E. 1969. Principles of systematic zoology.McGraw-Hill Book Co., New York.
Melvin, N. C. 1976. A systematic account of theLeucothoë axillaris complex. M.S. Thesis, ClemsonUniversity, Clemson, South Carolina.
———. 1978. The new combination Leucothoë walteri(Willd.) Melvin. Castanea 42: 304–308.
———. 1979. Reflections on the name Leucothoëfontanesiana versus L. walteri. Castanea 43: 265–266.
———. 1980. A systematic investigation of the genusLeucothoë (Ericaceae). Ph.D. Dissertation, MiamiUniversity, Oxford, Ohio.
———. 1981. Additional comments on Leucothoë[Ericaceae]. Baileya 21: 127–130.
Middleton, D. J. & C. C. Wilcock. 1990. Chromosomecounts in Gaultheria and related genera. EdinburghJournal of Botany 47: 303–313.
Mishler, B. D. & E. C. Theriot. 2000. The phylogeneticspecies concept (sensu Mishler and Theirot): mono-phyly, apomorphy, and phylogenetic species con-cepts. Pp. 44–54. In: Q. D. Wheeler & R. Meier(eds.), Species concepts and phylogenetic theory.Columbia University Press, New York.
Nakai, T. 1922. Trees and shrubs indigenous to Japanproper, part 1. Seibido Shoten, Nihombashi, Tokyo.
437JUDD ET AL.: LEUCOTHOË (ERICACEAE)2013]

Nelson, G. 1996. The shrubs and woody vines ofFlorida. Pineapple Press, Sarasota, FL.
Niedenzu, F. 1889. Über den anatomischen Bau derLaubblätter der Arbutoideae und Vaccinioideae inBeziehung zu ihrer systematischen Gruppierung undgeographischen Verbreitung Botanische Jahrbücher11: 134–263.
Nixon, K. C. & Q. D. Wheeler. 1990. An amplificationof the phylogenetic species concept. Cladistics 6:211–223.
Nuttall, T. 1843. Description and notices of new or rareplants in the natural orders Lobeliaceae, Campanula-ceae, Vaccinieae, Ericaceae, collected in a journeyover the continent of North America, and during avisit to the Sandwich Islands, and Upper California.Transactions of the American Philosophical Society8: 251–272.
Palser, B. F. 1952. Studies of floral morphology in theEricales: II. Megasporogenesis and megagametophytedevelopment in the Andromedeae. Botanical Gazette114: 33–52.
Radford, A. E., H. E. Ahles & C. R. Bell. 1968.Manual of the vascular flora of the Carolinas. TheUniversity of North Carolina Press. Chapel Hill,NC.
Rehder, A. 1960. Manual of cultivated trees and shrubshardy in North America. MacMillian Co., New York.
Reveal, J. L. 2009. Identification of the plant andassociated animal images in Catesby’s Natural Histo-ry, with nomenclatural notes and comments. Rhodora111: 273-388.
Riefner, R. E. 1979. Leucothoë axillaris var. editorum(Fernald and Schubert) Ahles, new to Maryland.Castanea 44: 59.
Rudenberg, L. 1963. Documented chromosome numb-ers of plants. Madroño 17: 116–117.
Sleumer, H. 1959. Studien über die Gattung LeucothoëD. Don. Botanische Jahrbücher 78: 435–480.
Small, J. K. 1901. Shrubs and trees of the southernstates. III. Bulletin of the Torrey Botanical Club 28:290–294. [Description of Leucothoë platyphylla onpp. 290-291.]
———. 1914. Ericaceae. North American Flora 29: 33–102.
———. 1919. Leucothoe catesbaei. Addisonia 4 (4): 61-62, t. 151.
———. 1933. Manual of the southeastern flora. Univer-sity of North Carolina Press. Chapel Hill. NorthCarolina.
Stevens, P. F. 1971. A classification of the Ericaceae:subfamilies and tribes. Botanical Journal of theLinnean Society 64: 1–53.
———, J. L. Luteyn, E. G. H. Oliver, T. L. Bell, E. A.Brown, R. K. Crowden, A. S. George, G. J.Jordan, P. Ladd, K. Lemson, C. B. McClean, Y.Menadue, J. S. Pate, H. M. Stace & C. M. Weiller.2004. Ericaceae. Pp. 145–194. In: K. Kubitzki (ed.),The families and genera of vascular plants: VI. Flower-ing plants: Dicotyledons: Celastrales, Oxalidales,Rosales, Cornales, Ericales. Springer-Verlag. Berlin.
Stuart, J. D. & J. O. Sawyer. 2001. Trees and shrubs ofCalifornia. Univ. of California Press, Berkeley, CA.
Symonds, G. W. D. 1963. The shrub identification book.William Morrow & Co., New York.
Taylor, W. K. 1992. The guide to Florida wildflowers.Taylor Publ. Co., Dallas, TX.
———. 1998. Florida wildflowers in their naturalcommunities. Univ. Press of Florida, Gainesville, FL.
Tucker, G. C. 2009. Leucothoë D. Don. Pp. 508–510,and fig. on p. 507. In: Flora of North AmericaEditorial Committee, Flora of North America, Vol. 8.Magnoliophyta: Paeoniaceae to Ericaceae. OxfordUniv. Press, New York.
Van der Burgh, J. 1978. The Pliocene flora ofFortuna-Garsdorf. I. Fruits and seeds of angio-sperms. Review of Palaeobotany and Palynology26: 173–211.
Ward, D. B. 2007. The Thomas Walter Herbarium is notthe herbarium of Thomas Walter. Taxon 56: 917–926.
Waselkov, K. & W. S. Judd. 2008. A phylogeneticanalysis of Leucothoë s.l. (Ericaceae; tribe Gaulther-ieae) based on phenotypic characters. Brittonia 60:382–397.
Weakley, A. S. 2011. Flora of the southern and mid-Atlantic states. “Working draft of 15 May 2011.”UNC Herbarium, North Carolina Botanical Gar-den, University of North Carolina at Chapel Hill.Document available online: www.herbarium.unc.edu/flora.htm.
Wheeler, Q. D. & N. I. Platnick. 2000. The phyloge-netic species concept (sensu Wheeler and Platnick).Pp. 55–69. In: Q. D. Wheeler and R. Meier, eds.Species concepts and phylogenetic theory. ColumbiaUniversity Press, New York.
Wood, C. E. 1961. The genera of Ericaceae in theSoutheastern United States. Journal of the ArnoldArboretum 42: 10–18.
Wyman, D. 1960. Plants of possible merit. Arnoldia 20: 12.
438 BRITTONIA [VOL 65
Copyright © 2022 FDOKUMEN




















