
Growth and Mortality of the Red Shrimp Aristaeomorpha Foliacea in the Sicilian Channel...
14
Crustareana 67 (3) 1994,@ E.J. Brill, Leiden GROW'TH AND MORI-ALITY OF THE RED SHRIMP ARISTAEOMORPHA FOLIACEA IN THE SICILIAN CHANNEL (MEDITERRANEAN SEA) BY SERGIO RAGONESE'), MARCO L. BIANCHINIz,:r) and VINCENT F. GALLUCCI3) r) Ist. Tecnologia Pesca e Pescato, CNR, Via Vaccara, N{azara ('I'P), Italy; 2) P.F. RAISA, CNR, Via Tiburtina 770, Roma, Italy; 3) School of Fisheries, University of Washington, Seattle, WA, U,S.A, ABSTRACl' 'l-he deep water red shrimp Aristaeomorpha.foliatea is an important commercial species lor the trarvl lìsheries in the Sicilian Channel (Central \'Iediterranean Sea; FAO statistical area 37.5). An assessment program was recently implemented bv ITPP-IINR. Two years of seasonal research trau'l sun'eys lvere carried out on the Italian side of the Siciìian flhannel. A time seriesoflength-frequencv distributions has been analyzed using different techniques; the results are used to estimate growth and mortality parameters of thc stock. In females, two modal pulses can be discriminated for the.juveniles w-hile two-to-three "years" are visible for the adults. Female gros'th can rvell be represented by a Von Bertalanffy function: as rvith other species, too, there are ts'o possible growth hypotheses, slower and faster; A. foliacea Ièmales appear to grow relatively fast any,hou'. Femaìe total annual mortality (Z) is estimated around 0.65, with a natural mortalitv (X,I) at 0.42, resulting in an exploitation rate (E) of'0.38. \'Iale results appear similar to those ol'lèmales, but the models are less wcll-defined and valid; male srowth data are best fittecl with a linear model. RE,SU\,fEN La gamba de prolìndidad Ari.staeomorpha Jòliacea es una especie comercial importante en la pesqueria de arrastre del Canal de Sicilia (N'IarMediterróneo central; órea estadística 37.5 de la FAO). En el marco de un programa de evaluación realizado por el ITPP-CNR, se han llevado a cabo muestreos estacionales de arrastre durante dos afros cubricndo la parte italiana del Canal de Sicilia. El crecimiento y la mortalidad de la población de A.foliacease ha estimado a partir del anólisis temporal de sus frecuencias de tallas por métodos distintos. Las hembras.jór'enes y adultas se separan claramente, en las primeras pueden identificarse dos picos modales, mientras que en las segundaspueden observarsedos o tres "aiios". El crecimiento es expresado, según la functión clàsica de Von Bertalanffy, con dos hipótesis posibles de creci- miento, mós o menos rapido. La mortalidad total (2, anual) para hembras es 0,65 y la natural (Nf) de 0,42 las cuales resultan de una tasa de explotaciòn (E) del 0,38. Los machos presentan unos patrones similares a las hembras pero menos claros, siendo el crecimiento de los machos me..jor representado por un modeìo linear. INTRODT-ÌCTION 'fhe deep-sea red shrimp Aristaeomorpha foliacea (Risso, 1827) (Decapoda, Aristeidae) is of great economic interest,actual or potential, over all of its wide area of distribution (Holthuis, 1980; Dore & Frimodt, 1987).
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Growth and Mortality of the Red Shrimp Aristaeomorpha Foliacea in the Sicilian Channel...

Crustareana 67 (3) 1994, @ E.J. Bri l l , Leiden
GROW'TH AND MORI-ALITY OF THE RED SHRIMPARISTAEOMORPHA FOLIACEA IN THE SICILIAN CHANNEL
(MEDITERRANEAN SEA)
BY
SERGIO RAGONESE') , MARCO L. BIANCHINIz, : r ) and VINCENT F. GALLUCCI3)
r) Is t . Tecnologia Pesca e Pescato, CNR, Via Vaccara, N{azara ( ' I 'P) , I ta ly; 2) P.F. RAISA, CNR,
Via Tiburt ina 770, Roma, I ta ly; 3) School of Fisher ies, Univers i ty of Washington, Seatt le, WA,
U , S . A ,
ABSTRACl '' l -he
deep water red shr imp Ar istaeomorpha.fo l iatea is an important commercia l species lor thetrarv l l ìsher ies in the Sic i l ian Channel (Central \ ' Iedi terranean Sea; FAO stat is t ical area 37.5) . An
assessment program was recent ly implemented bv ITPP-I INR. Two years of seasonal research
trau' l sun'eys lvere carr ied out on the I ta l ian s ide of the Sic i ì ian f lhannel .A t ime ser ies of length- f requencv dist r ibut ions has been analyzed using di f ferent techniques; the
resul ts are used to est imate growth and morta l i ty parameters of thc stock.In females, two modal pulses can be discr iminated for the. juveni les w-hi le two-to- three "years"
are v is ib le for the adul ts. Female gros ' th can rvel l be represented by a Von Berta lanf fy funct ion: asrv i th other species, too, there are ts 'o possib le growth hypotheses, s lower and faster ; A. fo l iaceaIèmales appear to grow relat ively fast any,hou' .
Femaìe tota l annual morta l i ty (Z) is est imated around 0.65, wi th a natural morta l i tv (X, I ) at 0.42,resul t ing in an exploi tat ion rate (E) of '0.38.
\ ' Ia le resul ts appear s imi lar to those ol ' lèmales, but the models are less wcl l -def ined and val id;male srowth data are best f i t tec l wi th a l inear model .
RE,SU\,fEN
La gamba de prol ìndidad Ar i .s taeomorpha Jòl iacea es una especie comercia l importante en la
pesquer ia de arrastre del Canal de Sic i l ia (N' Iar Medi terróneo centra l ; órea estadíst ica 37.5 de la
FAO). En el marco de un programa de evaluación real izado por e l ITPP-CNR, se han l levado a
cabo muestreos estacionales de arrastre durante dos afros cubr icndo la parte i ta l iana del Canal de
Sic i l ia .El crecimiento y la morta l idad de la población de A. fo l iacea se ha est imado a part i r del anól is is
temporal de sus f recuencias de ta l las por métodos dist intos.Las hembras. jór 'enes y adul tas se separan c laramente, en las pr imeras pueden ident i f icarse dos
picos modales, mientras que en las segundas pueden observarse dos o t res "a i ios" . El crecimientoes expresado, según la funct ión c làsica de Von Berta lanf fy, con dos hipótesis posib les de creci-miento, mós o menos rapido.
La morta l idad tota l (2, anual) para hembras es 0,65 y la natural (Nf) de 0,42 las cuales resul tan
de una tasa de explotaciòn (E) del 0,38.Los machos presentan unos patrones s imi lares a las hembras pero menos c laros, s iendo el
crecimiento de los machos me.. jor representado por un modeìo l inear.
INTRODT-ÌCTION
' fhe deep-sea red shrimp Aristaeomorpha foliacea (Risso, 1827) (Decapoda,
Ar is te idae) is of great economic in terest , actual or potent ia l , over a l l o f i ts wide
area of d is t r ibut ion (Hol thuis , 1980; Dore & Fr imodt , 1987).

GROWTH AND MORTALITY OF RED SHRIMP 3+9
Particularly in the Mediterranean, it represents, together with the other red
shrimp Aristeus antennatus (Risso, lBl6) (cf. Demestre, 1990) and the Norway
lobster, the target for the demersal trawl f isheries operating between 200 and
800 m, i .e . , over the cont inenta l s lope (Ors i Rel in i & Rel in i , l9B5; Sardà, l9BB;
Ragonese, l9B9; Ragonese & Bianchin i , 1992).
Besides having a scientif ic interest, the need to evaluate the red shrimp
resource arises from its economic importance, since the total landings from the
Sicil ian Channel are of the order of several hundred tons per year, i.e., a value
\\ 'ell over 10,000,000 US$. In fact, the fishing activity truly resembles a mono-
speci f ic f ishery (Sardà, l9BB; Demestre, 1990).
The life cycle of the red shrimp in the Sicil ian Channel occurs in a deep
water envi ronment , homeothermic (about 13.5"C), considered o l igotrophic and
stable. At present, the red shrimp occupies one of the top positions of its trophic
web, because larger predators, such as angierfish, hake, sharks, etc., are scarce
due to over f ish ing of the demersal resources (CGPM, l9B9).
This paper describes the size structure of the populations ol Aristaeomorpha
.foliacea living in the Italian part of the Sicil ian Channel; afterwards, its temporal
evolution is analyzed in order to estimate growth and mortality parameters.
The ultimate objective is to uti l ize these preliminary estimates to develop
production and population dynamics models to be used for a rational exploita-
t ion of these resources.
MATERIALS AND N,{E,THODS
The red shr imps were col lected us ing an ad hoc char tered commercia l
trawler in the Sicil ian Channel (f ig. l), an area characterized by irregular
grounds, strong hydrodynamics (actual deep water gyros have been recorded)
and by the presence of shallow banks that separate the mesobathyal bottoms
into an Eastern and a Western basin (Ragonese, 1993).
Eight seasonal trawl surveys (Levi, 1990) have been carried out from spring
l9B5 to winter l9B7 using a commercia l t rawl net (net mouth of 0.7 x l2 m;
terminal bag, or cod-end, wi th mesh of lB-20 mm of s ide) .All the hauls were made in daylight, during the period of maximum availabil-
itv of these shrimps (Sardà, l9BB), which are presumed to swirn in the water
column dur ing the n ight (Lagardère, l97 l ) . The length of each haul was about
5000 m (1 h at 2.8 knots, in a random di rect ion) . Whenever possib le, the
shrimps were measured on board using a pair of calipers (precision 1 mm),
recordins the oblique length of the carapace (CL; Mayrat, 1964) and also the
S C X .' f
he total catch oî A.foliacea ín the eight surveys was 463 ke, corresponding to
19,+00 animals, coming l rom 164 val id hauls, o{ 'which 132 were posi t ive.
Length frequency distributions (LFSs) by sex and survey were produced
thc or , 'era l l d is t r ibut ion area, i .e . , 17,000 km2 in a depth range of 400-800
for
m.
350 S. RAGONESE, M. L. BIANCHINI & V. F, GALLUCCI
1 0 ' E
Fig. l . Study area idoued).
Bhat tacharya 's method (MPA), as implemented in the Compleat ELEFAN(Gayani lo et a l . , l9B9), was used for a pre l iminarv d iscr iminat ion of theGaussian components; the select ion points have been chosen subject ive ly , nottak ing in to considerat ion the suggest ion of Pauly & Caddv (1985).
The evolut ion in t ime of the mean lengths, est imated for " representat ive
components" ( in general , components wi th a f requency greater than l0 and aseparation index sreater than 2) has been qualitatively analyzed. Since theseasonal LFDs exhib i t considerable var iabi l i ty over the years, two methodslr'ere applied to damp the differences: f irst, LFDs were combined bv l ike-seasons ( i .e . , summer l9B5 p lus summer 1986, and so on) , and, second, theywere smoothed over three points.
An age was assigned to each Gaussian component on the basis of the surveyseason, and of previous information on the reproductive pattern (Ragonese,1e89) .
The procedure NONLIN of SYSTAT (Wilkinson, lgBB)was used to estimarethe srowth parameters; the ratio of the number and the standard deviation wasused as a weighting coefficient.
The re lat ionship between CL and age has been model led wi th the Brody-Von Berta lanf f l 'equat ion, e i ther in i ts specia l (C = 0; Ricker , 1979) or seasonalform (Somers, in : Sparre et a l . , l9B9). I t is important to note that the two curvesare only homolosous, not equal or s imi lar : that means that they produceestimates of "different" CL- and K, and therefore a strict comparison of theparametcrs is impossib le.

GROWTH AND MORTALITY OF RED SHRINIP 3 5 1
The procedure ELEFAN I was a lso used on the smoothed LFDs, to obta in an
est imate of the growth parameters independent f rom the previous method.
The "best curve " was chosen on the basis of an index of f itness (R,), and of its
apparent overall biological reasonableness.
Different growth models, such as the quadratic, potential, and l inear curves
as \ ! 'e l l as the Gompertz 's model (Ricker , 1979), have been essayed too.
The mean square error (MSE) and residuals pattern have been used to
evaluate the goodness-oÈfit of the data.
The Munro 's rp ' (phi pr ime) (Sparre et a l . , l9B9) has a lso been computed' for
broad comparison of the overall growth performance of other populations.
Total mortality rates (Z/year') have been estimated by sex using Heincke's
method (Garc ia & Le Reste, lg8 l ) , which is based on the hypothesis of
demographic s tat ionar i ty (Ebert , l981) , on the LFDs combined by l ike-seasons.
The Beverton and Hol t 's method, based on length moments, implemented
on the overall pooled data in order to reduce the bias associated with the
discrete recru i tment (Ralston, l9Bg), has been ut i l ized as an a l ternat ive
approach.The longevity (T0.01) estimation for thc exploited stock was computed using
the 'faylor's
approximation (Taylor, l95B).
A point value for the natural mortality of the lèmales has been estimated with
the Rikhter and Efanov 's equat ion (Garc ia & Le Reste, 198l ) us ing lB months
as a value for T, , .0 (Ragonese & Bianchin i , 1995); in the meant ime' a ser ies of
natural mortality values (M/year) was derived along with the empirical method
of Alagaraya (Sparre et a l . , l9B9), based on longevi tv est imates for unexplo i ted
stocks.
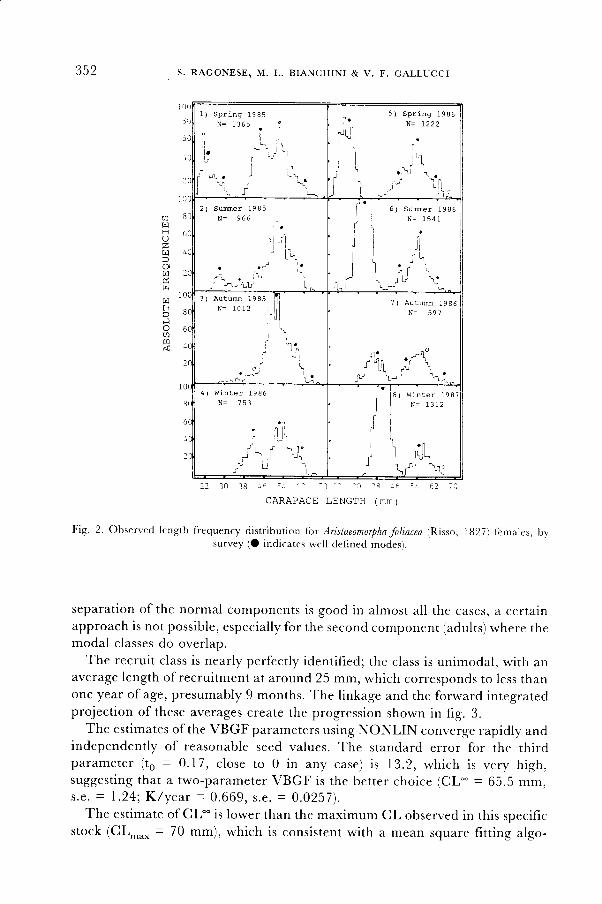
Size distributionThe LFDs for the B surveys,
N{PA, are reported in fig. 2.The surveyed population is made up of two major components: the first
major component, that of the recruits, appears in the sprinstime with one or
two modes (in this last case , of different strengths and substantially overlap-
ping); the recruits are completely separated from the second component' which
shows 2 to 3 representative modes. Despite the large variabil ity (especially the
much smaller recruitment in the frrst year) it is possible to distinguish a basic
scheme where the first major component quickly progresses up to the second,
unti l the merging in the following spring, when a new recruitment appears.
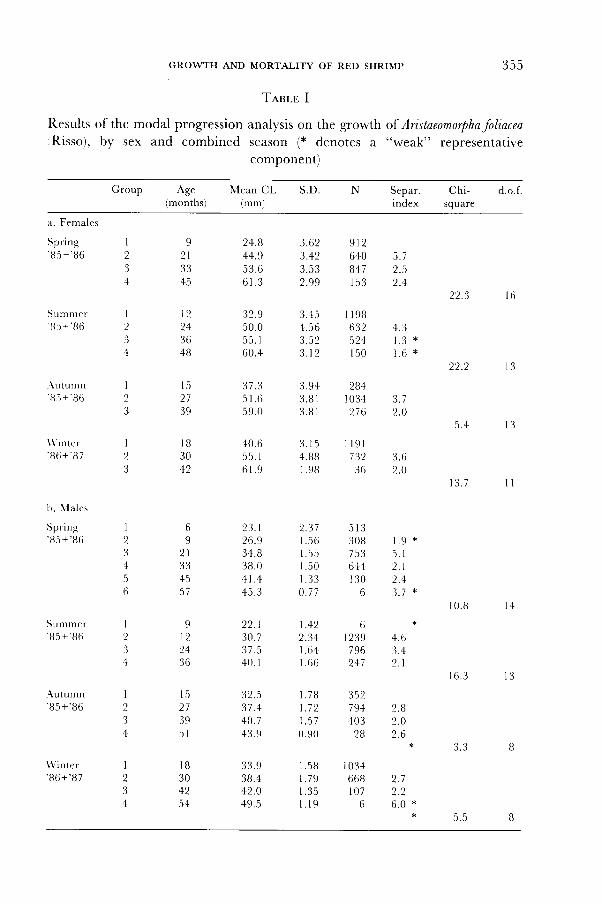
Results from the MPA for combined like-seasons are reported in table Ia,
together with the level of significance of the curves. Although the degree of
RESULTS
Female s
together with the modal classes resuiting from
352 S . R A G O N E S E , M . L . B I A N C H I N I & V . F . G A L L U C C I
3 0 3 8 . 6 . . \ - 0 " " r î r q . . 6 :
C A R A P A C E L E N G T H ( n . n )
Fig. 2 Observed length frequency distribution for Aristaeomorphafoliacea (Risso, ltl27i females, bvsurvey (O indicates well defined modes).
separation of the normal components is good in almost all the cases, a certainapproach is not possible, especially for the second component (adults) where themodal classes do overlap.
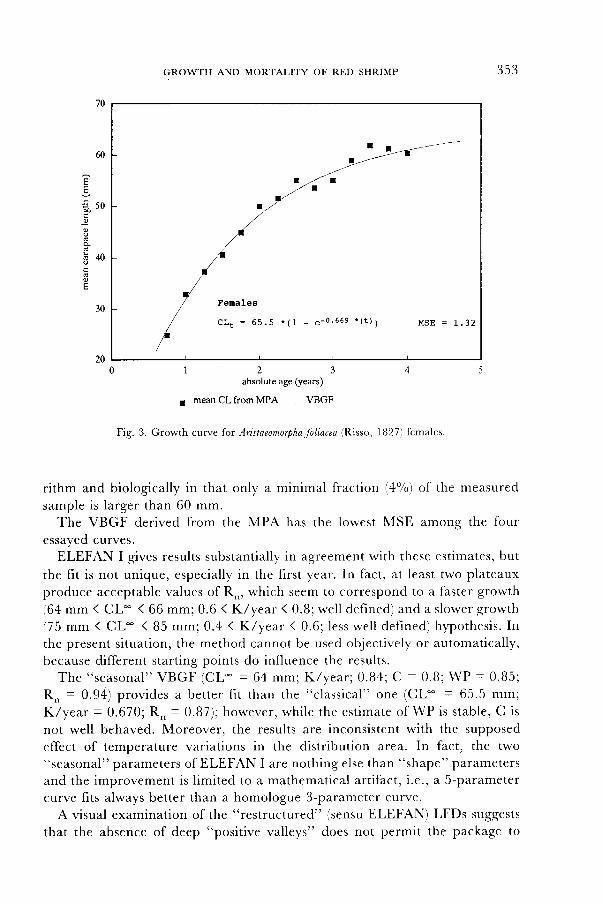
The recruit class is nearly perfectly identif ied; the class is unimodal, with anaverage length of recruitment at around 25 mm, which corresponds to less thanone year of age, presumably 9 months. The linkage and the forward intesratedprojection of these averages create the progression shown in fig. 3.
The e stimates of the VBGF parameters using NONLIN converge rapidly andindependently of reasonable seed values. The standard error for the thirdparameter ( to = 0.17, c lose to 0 in any case) is 13.2, which is very h igh,suggest ing that a two-parameter VBGF is the bet ter choice (CL- = 65.5 mm,s .e . = I . 24 ; K / yea r = 0 .669 , s .e . = 0 .0257 ) .
The estimate of CL- is lower than the maximum cL observed in this specificstock (CL^u*= 70 mm), which is consistent wi th a mean square f i t t ing a lgo-
a
OzDortrl
I
-ÉU]
6
8 r
l i\ i
l . r - 1l . (
jf
/ i .
- " - l L - -
1 ) S p r i n g t 9 8 5N = 1 3 6 5
5 ) s p r i n g 1 9 8 6
.r qlì \
: \I tt l
,lr f l
y L t: L ì i
2 ) S u m e r L 9 8 5
N = 9 6 6 -t.i , i-
LL' ', . / , '
f ' l
6 ) s u m e r 1 e 8 6
J Ì N = t s 4 1
l l rI I :'r"I I . . r \
, l \ - I J \3) Autumn 1e85 fJ l
N = 1 0 1 2 I
I ' rl l; t
, . / !
-'*íii- . l
7 ) A u t u m n 1 9 8 6N = 5 9 7
n . J al Ì n - . l - \
a ' l I :
4 ) W r n t e r 1 9 8 6N = 7 5 3
. î 1I I l r ì, - ') i i \ f ] "
i l ! \" i I I- !
, / ;
w r n t e r 1 9 8 7N = 1 . 3 1 . 2
. i ]R l l
GROWTH AND MORTALITY OF RED SHRIMP 3s3
Fenales
C L . = 6 5 . 5
absolute age (years)
I mean CL from MPA - VBGF
Fig. 3. Growth curve lor Aristaeomorphafoliarra (Risso, l827) females.
rithm and biologically in that only a minimal fraction (,4o/o) of the measured
sample is larger than 60 mm.The VBGF derived from the MPA has the lowest MSE among the four
essayed curves.ELEFAN I gives results substantially in agreement with these estimates, but
the fit is not unique, especially in the first year. In fact, at least two plateaux
produce acceptable values of R,, which seem to correspond to a laster growth
(64 mm < CL- ( 66 mm; 0.6 ( K/year ( 0.8; wel l def ined) and a s lower growth(75 mm < CL- ( 85 mm; 0.4 ( K/year ( 0.6; less wel l def ined) hypothesis . In
the present situation, the method cannot be used objectively or automatically,
because different starting points do influence the results.
The " seasona l " VBGF (CL- = 64 mm; K /yea r ;0 .84 ; C = 0 .B i WP = 0 .85 ,
R, , = 0.94) prov ides a bet ter f i t than the "c lass ical" one (CL- = 65.5 mm;
K/year = 0.670, R" = 0.87) ; however, whi le the est imate of WP is s table, C is
not well behaved. Moreover, the results are inconsistent with the supposed
effect of temperature variations in the distribution area. In fact, the two"seasonal" parameters of ELEFAN I are nothing else than "shape" parameters
and the improvement is l imited to a mathematical artifact, i.e., a S-parameter
curve fits always better than a homologue 3-parameter curve.
A visual examination of the "restructured" (sensu ELEFAN) LFDs sugge sts
that the absence of deep "positive valleys" does not permit the package to
E
€ s oú - -0
h a o
o

3 5 4 S. RAGONESL, . À , I , L . B IANCHINI & V. F . GALLUCCI
univocally orient the curl 'e, also because pe aks lvith just a lèw animals are toostrongìy emphasized. Thc tr 'vo hypothese s and the intercycle differences can becxplained by the fact that the program finds only two "strong" representativecohorts , i .e . , just the two major components ( recru i ts and adul ts) ; as a conse-quence, rvithout forcing the curve ol'er the suggested three-cohorts hypothesis,the proeram tends to overest imate the growth rate.
f 'he index of per formance is p ' - - 1 .7 + and g ' = 3.46 for K expressed in weeksand years, respect ive ly .
NÍor ta l i tv
The ra t i o o f r ec ru i t s t o t o ta lpopu la t i on i s 0 .357 , 0 .478 ,0 .178 , and 0 .608 fo rspr ing, summer) fa l l , and lv inter , respect ive ly .
The Heincke's morta l i ty est imate based on the summer surveys (Z = 0.65 l ) isthe best su i ted, because the juveni les are fu l ly recru i ted to the area and to thegear (CL, , . . . = 33 mm); i t is lower than the value iZ = 0.786) obta ined l rom themethod o lmoments (CL'= 40 mm, CL, , , . , . , , = 51.7 mm) and of the growthpa rame te rs (CL - = 65 .5 mrn , K /yea r = 0 .669 ) .
The lonsevity computed with Taylor's approximation based on the abor.'eVBGF parameters and on a ' Io .or is around 7 years, qui te a h igh value; anyhow,assumins a longevity range from 6 to B years, it is possible to estimate a naturalmortaÌ i tv of 0.92,0.77, and 0.58 for age 6, 7 and B vears, respect ive ly .
On the other hand, the estimate of \,{,/ycar from the reproductive pattern is0 . 4 1 B .
Gir , 'en that th is s tock is in tcnsely explo i ted (Z >> N,{ ) and that the overal lpooled LFD shape suggests a type "k" confisuration rvith Z/K ( I (Barry &' f epne r ,
1990 ) . i . e . , 2 < 0 .669 i n t h i s case , t he coup led es t ima tes o f Z = 0 .65 Iand N{ = 0.418 appears to be the most probable, a lso in v iew of rvhat a l readyhas bce n repor ted or assumed in the l i terature for other " temperate" shr imps(Pauly et a l . , 19841 Yahiaoui et a l . , l986; N, f oht t & Polor , ' ina, 1987; Sardà, 19BB;Demestre, 1990; Ragonese & Bianchin i , 1992).
' fhe explo i tat ion rat io (E = I ' /21 is therefore 0.38, a reasonable est imate
given the fact that this species is actively hshed since early ages.
N{ales
Size d is t r ibut ionBesides the dilferent length ranee (males are smaller, with CL-^* = 50 mm),
the size structure and its evolution broadly resemble that of the females.I-rom a qualitative analysis of the LFDs, and especially in the spring, it re sults
that the overlap in size be twee n the adult components of female s and males isvery smal l .
The adul t component car ìnot be easi ly separated in c lasse s, even i f 2-3 modesare sketched. I 'hc presence of two modes for the first major component tends to
GROWTH AND MORTALITY OF RED SHRIMP 355
TesrE, I
Results of the modal progression analysis on the growth ol Aristaeomorphafoliacea,Risso), by sex and combined season (* denotes a "weak" representative
component)
Group S.DAg" Mean CL(months) (mm)
Sepa r . Ch i - d . o . findex square
Surnmer I' fh*
86 234
- \ r r t umn' t 3 i + ' 96
\ \ ' i r . r ter'B t i + ' 87
b. Nlales
Spr ing' u5+ '86
a. Females
Sprinu'85 + ' �86
òummer' 8 5 + ' 8 6
\utumnB5+ '86
\\ i inter' 86+ 'B 7
t2 32 .92+ s0.03 6 5 5 . 148 60 .4
1 5 3 7 . 32 7 5 1 . 639 59 .0
22.3 16
: ) -+ l 3
1 6 . 3
I
23+
I23
92 l3 3+J
I B3012
24.844.95 3 . 66 1 . 3
3 . 6 23.123 . 5 32 .99
3.454.563 . s 23 . 1 2
3.943 . 8 l3.8 r
3 . t 54 . BB1 . 9 8
2 .37t . 5 6r . 5 5r .5 t )l . 3 30 . 7 7
t . 4 22 .34I . (i.11 . 6 6
I . 7 ut . i 2| . 5 70.90
9 1 26408471 5 3
I l 9 86 3 25211 5 0
28+I 034276
(ìr 2397962+7
35279+.t032B
( f
2 .52 .1
1 .31 . 3 *l . t i *
22.2 13
I23
It 2
3 6
l 52 7395 t
2 3 . I2 6 . 934.83{Ì.0+ t . +45 .3
22.130.73 7 . 540 . I
32.537 .1+0 .743 .9
33 .938..++2 .049.5
3 . 72 .0
1 . 95 . 12 . 1l . t
3 . 7
+. t )3 .+2 . 1
2 .82 .02 . 6
40.65 5 . I6 l . 9
I l 9 l73' �2 3.63 6 2 . 0
6I
2 l3 3+ J
5t-
I,2
3I
5(ì
I B3012)+
5 1 33087 5 364.11 3 0
6
II
3+
I234
I23+
t 3
3 . 3
5 8 1 0 3 479 66835 107t 9 6
2 . 72.'�26 .0 *
* 5 . 5 8
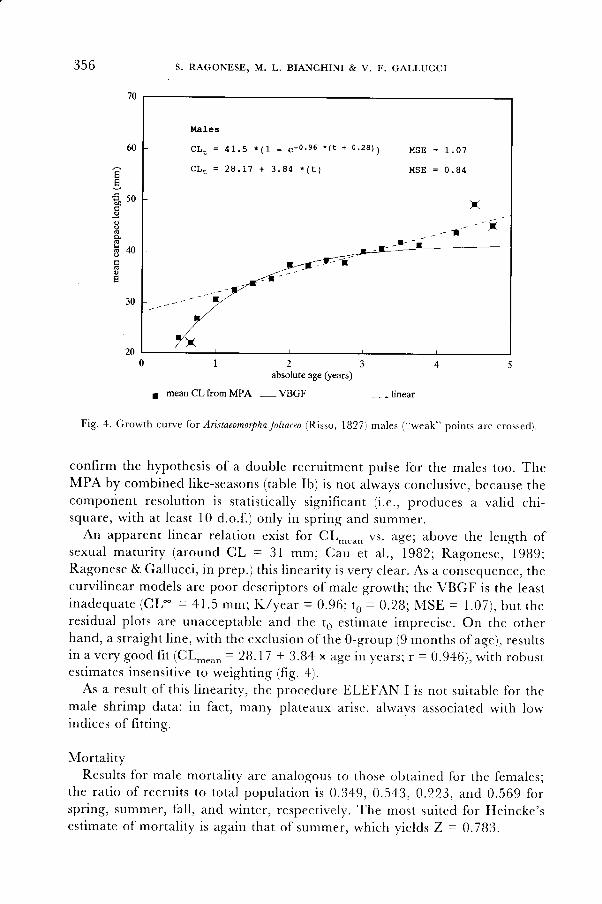
356 S. RAGONESE, M.L, BIANCHINI & V. F. GALLUCCI
Males
C L . = 4 1 . 5 * ( 1 - e - 0 . s 6 * ( t + 0 . 2 8 ) ) M s E = l . 0 7
C L . = 2 9 . 1 7 + 3 . 8 4 * ( t ) M S E = 0 . 8 4
absolute age (years)
mean CL from MPA - VBGF
Fis.4. Growth curve lor Ar istaeomorphafol lacea (Risso, l827) males ("weak" points are crossed).
confirm the hypothesis of a double recruitment pulse for the males too. TheMPA by combined like-seasons (table Ib) is not always conclusive, because thecomponent resolution is statistically significant (i.e., produces a valid chi-square, wi th at least l0 d.o. f . ) only in spr ing and summer.
An apparent l inear relation exist for CL-... vs. age; above the length ofsexual matur i ty (around CL = 3 l mm; Cau et a l . , 1982; Ragonese, l9B9;Ragonese & Gallucci, in prep.) this l inearity is very clear. As a consequence, thecurvil inear models are poor descriptors of male growth; the VBGF is the leasti n a d e q u a t e ( C L - = 4 l . 5 m m ; K / y e a r = 0 . 9 6 ; t o = 0 . 2 8 ; M S E = 1 . 0 7 ) , b u t t h eresidual p lots are unacceptable and the t6 est imate imprecise. on the otherhand, a straight l ine, with the exclusion of the 0-group (9 months of aee), resultsin a very good f i t (CL-.o, = 28. l7 + 3.84 x age in years, r = 0.946) , wi th robustestimates insensitive to weighting (fiS. a).
As a result of this l inearity, the procedure ELEFAN I is not suitable for themale shrimp data: in fact, many plateaux arise, always associated with lowindices of frtt ins.
MortalityResults for male mortality are analosous to those obtained for the females;
the rat io of recru i ts to tota l populat ion is 0.349, 0.5+3, 0.223, and 0.569 forspring, summer) fall, and winter, respectively. I 'he most suited for Heincke'sest imate of morta l i ty is again that of summer, which y ie lds Z = 0.783.
ó0
= \ { l@ - -
Io
qo
k a o
oE
20

GROWTH AND MOR'IALITY OF RED SHRIMP 357
The shape of the overall pooled Ll'D again suggests a type "k" confisuration
but the estimates of N,I could be reasonable onlv under the hypothesis of a
longer l ife, of the same orde r as that of the fèmale s.It is worth noting that the observed CL,..* of 50 mm corresponds to an age of
5-6 years, when the l inear model is used.
DISCUSSION
Size structure
Size structure in a unit stock is the inteerated result of'growth and mortality;
sti l l , problems in modeling and interpretation arisc from the fact that, even with
the constra ints of s tat ionar i ty ( i .e . , random f luctuat ions in reproduct ion,recruitment, growth, and mortality), the relationship betr,veen length structl lreand demographic parameters is not l incar (Rosenberg & Beddington, l9BB).
A d iscrete recru i tment , wi th one or two unequal over lapping peaks, seems
the ru le for deep water shr imps (Pauly et a l . , l9B4).The presence of abundant adult animals vs. the recruits suggests a consider-
able l ife span.Beside (unknown) selectivity problems, the lack of r. 'ery small animals in the
samples is probably ascribable to the campaien period, that started in the late
spr ing. Nevertheless, g iven the pract ica i iso lat ion of the recru i ts component ,
the subsequent b ias and changes of the parameter est imates should be
negligible.The observed differences in recruitment between the two years of study
confirm the aleatory nature of this process, whose factors are sti l l unknown.
GrowthDespite their exoskeleton, shrimps are suited to continuous growth models,
given that the carapace is fairly elastic and that moltins is frequent andasynchronous (Cobb & Caddy, l9B9; Dai l e t a l . , 1990): moreover, a VBGF canbe used to model shrimp growth (Parrack, 1979), at least lèmale growth.
A frequently used tool for LFD analysis is FILEFAN (Hilborn & Walters,1992), notwi thstanding i ts in t r ins ic weakness, among which the subject iv i ty ofthe procedures and the necessity to introduce external biological information in
the interpretation of the results; this tool, however, is ver1, handy to fit a VBGF
to lensth data.More sophisticated growth curves could be used for a better fìt, providing a
more "flexible " shape or seasonality, but such complicated curves wil l seldombe just i f ied (MacDonald, 1987).
Wurtz (unpubl ished), us ing LFDs l rom Cau et a l . (1982), produces the onlyknown est imates for A. fo l iacea, i .e . , CL- = 66.3 mm or 45.3 mm and K/year =
0.51 or 0.79 for females and males, respect ive ly ; i t appears that females values

358 s. RAGONESE, M. L.BIANCHINI & V. F. GALLUCCI
(g' = 1.6; computed for K/week) are in general agreement with the results of'
the present study.
Growth of males, captured in this study, is better modeled with a l inear
relation; while other authors continue to use a curvil inear model for both sexes
(Sardà & Demestre, l9B7), it appears from a rough analysis that a l inear
relation may apply for their male data too.Comparing the growth performance of the red shrimp in the Sicil ian Chan-
nel with that of other shrimps, the g' = 1.7 (K expressed in weeks) puts thefemales of A. fo l iaceainthecategoryof " largerMetapenaeus" (1 .62!0.27: Dal le t
a l . , 1990) and near the values of i ts companion species, Ar is teus antennatus (1 .5:
Yah iaou i e t a l . , l 986 ; Sa rdà & Demes t re , l 9B7 ;1 .7 : Ragonese & B ianch in i ,
1992; but a lso 2.1: Ors i Rel in i & Rel in i , 1985). Unfor tunate ly , th is index,
despite its popular use, does not allow for an estimate of variabil ity and,consequently, differences cannot be tested statistically, in contrast to the possi-
bil i ty offered by the omega test (Gallucci & Quinn, 1979)
MortalityMortality is the most variable and diff icult parameter to estimate (Neal &
Maris, l9B5), especially when different exploitation hypotheses (Mathews et al.,l 987 ) a re cons ide red .
Moreover, the constraints for the estimation of mortality are stronger thanthose for the estimation of srowth; in fact, the classical methodologies require
that the sample is representative of each age class and that the stationarityhypothesis is fulf i l led, conditions that are only rarely met, given fluctuations inrecruitment, varying levels of exploitation, and non-uniform catchabil it ies. Anapproach that simply uses the ratio between recruits and total catch, such asHeincke's, is probably the best suited for the estimation of the total mortality
when constraints are seriously violated.The est imate of Z (Z/year = 0.651) is consistent wi th the hypotheses of a l i fe
span Ionger than that of coastal shrimp, and ol a not (too) high F; in fact, the(trawl) f ishing mortality does not act over the whole distribution area norcontinuously, given the presence of canyons and other ruggedness and the redshrimp supposed behaviour of dispersal over the demersopelagic domain.
The estimation of the natural mortality is more diff icult, because the empiri-cal approaches based on correlations (Vetter, l9BB) could not be applied to
shrimps (Sparre et al., l9B9); when improperly used (such as the Rikhter and
Efanov's method used in the present study), they tend to grossly overestimatethe na tu ra l mor ta l i t y .
The use of the empirical approach of Alagaraya (Mo.o,) suggests a longer l ifespan (for unexploited shrimp stocks) longer than the results which are derivedfrom growth parameters; in fact, the exploited segment may have a compensa-tion effect to grow faster (Fréchette & Parsons, l983), or mortality may slow
down f ior the larger specimens.

GROWTH AND MORTALITY OF RED SHRIMP 3 5 9
The present value of M could be smaller than that expected for unexploitedpopulat ions (Mathews et a l . , l987) i f the hypothesis of reduced predat ion (dueto the removal of predators by the commercial f ishery) is accepted, togetherlvith the opportunistic, f lexible role of the red shrimp, thus placed at the top ofi ts t rophic web (Lagardère, 1971) in a pract ica l ly densi ty- independenrs i tuat ion.
CONCI,USIONS
Results, similar in the broad life history strategies ro those shown, have beenalready obtained for the companion spe cies Aristeus antennatus in the same areaiRagonese & Bianchin i , 1992).
The results of the present study, coupled with more general biologicalobservations on the fisheries in the Sicil ian Channel, contribute to consolidatethe feeling that, while the shrimp resources are probably not yet exceedinglyexploited, some sort of growth overfishing may be present and that the fishingactivit ies are not operating in accordance to an eumetric profi le.
A preliminary, conservative management intervention would require replac-ing the compulsory general f ishing halt, presently at the end of summer, withmore "targeted" interruptions, such as shift ing the closure for the red shrimpfisheries to the spring recruitment period.
Furthermore , the enforcement of the minimum lawful mesh size (40 mm),a l ready below the "opt imal" s ize for demersal shr imps (60 mm; Caddy, lgB2),is long overdue.
BIBLIOGRAPHY
B,+nnv,J. P. & M.J. TtcNnt, 1990. Inferr ing demographic processes f rom size-f requencv dist r ibu-t i ons : s imp le mode l s i nd i ca te spec i f i c pa r re rns o f s row th and mor ta l i t y . F i sh . Bu l l . , 88 ( l ) :l 3 - 1 9 .
Caoov,J. F. , 1992. Management of shr imp f isher ies. In: .F ish by-catch bonus l rom the sea. Rep.Techn . Cons . Sh r imp By -ca t ch U t i l . (Geo ree tow-n , Guyana ) , IDRC Or rawa : 120 -123 .
Cau, A. , A. M. DEIarve & M. Mune, 1982. Nuovi dat i sul l 'accrescimcnto e sul la matur i tà sessualedi Ar isteomorphafol iacea (Risso, 1827) (Decapoda Peneidae). Natural is ta Sic i l . , (a) 6 (suppl . 2) :429-+3+.
CGPM, 1989. Evolut ion récente des pèches médi terranéennes. CGPM XIX Sess. , FAO, RN'I /VI I :t - 3 4 .
Conn ,J . S . &J . F . Ceonv , 1989 . Thepopu la t i onb io l ogyo f decapods . I n : J . F . C ìeoov (ed . ) , N , l a r i neinvertebrate f isher ies: their assessment and managcment: 327-374. (Wi lev & Sons, NewYork).
De r r , w . , B . J . Hn r , P . c . Ro rn r r sennc & D .J . Snane r -es , 1990 . The b i o l ogy o f t he pene idae .Adv . ma r . B i o l . , 27 : l - 489 .
Dnnnsrne, M., 1990. Bio logia pesquera de la gamba Ar isteus antennatus (Risso, lBl6) en al MarCatalan: xxxiv, l - .1.13. (Doctoral thesis, L i r . r i r ' . of Barcelona).
Done, I . & c. Fnnroor, 1987. An i l lustrated guide to shr imp of the wor ld: l -229. (osprey Books,Hunt ington, NY).
Ennxr, T. , { . , l98l . Est imat ing morta l i ty f rom srowth parameters and a s ize dist r ibut ion whenrecrui tment is per iodic. L imnol . Oceanogr. , 26 ( ,4 '1: 761-7Gg.

360 S. RAGONESE. M. I , . BIANCHINI & V. F. GALLUCCI
FnÉcurrr t , .J . & D. G. Pensoxs, 1983. Report of shr imp ageing workshop held at Ste. Foy,
Quebec , i n \ { a l ' and i n Da r tmou th , Nova Sco t i a , i n November 1981 . NAFO Sc i . Coun .S tud ies , 6 : 79 -100 .
Ga.I - t . t rcct , V. 1" . & ' l ' .J . QurNN, I I . 1979. Reparameter iz ing. f ì t t ing, and test ing a s imple growthmode l . T rans . Amer i can F i sh . Soc . . l 0B : l 4 -25 .
G,+ncta, S. & L. Lr l ' Rnsrn, 1981. L i fè c,vc lcs. dynamics, explo i tat ion and management of coastalpenae id sh r imp s tock . FAO F i sh . t echn . Pap . , 203 : l - 215 .
Gavasrro. F- . ( -1. , .Jr . NI . SonreNo & D. Paurv. 1989. A draf t guide to the Compleat ELEFAN. Int .C ìen te r l i r " i ng ac l ua t i c Res . N Ianasem. Con t r i b . . 435 : l - 65 .
HILsoRN. R. & C.J. \Ver. lnns, 1982. Quant i tat ive f isher ies stock assessment: choice, dynamics anduncc r t a i n t v : l - 570 . iC lhapman and Ha l l ) .
Holrurr ls . L. B. , 1980. FAO specics catalogue. Shr imps and prawns of the wor ld: an annotatedca ta l ogue o f spec ies o f i n t e res t t o f i shc r i c s . FAO F i sh . Synop . , 125 ( l ) : l - 261 .
LecenoÈnn ,J . -P . , 1971 . Reche rches su r l ' a l imen ta t i on des c reve t t es de l a pen te con t i nen ta l emaroca ine . Te thvs . 3 i 3 ) : t ì 55 -675 .
LrvI , D. . 1990. Relazione sul l 'a t t iv i tà svol ta dal la U.O. Ist . d i Tecnolosia del la Pesca e delPesca to , N laza ra de l Va l l o . N .T .R . - I . T ' .P .P . , l 5 ( b i s ) : l - 21 l .
NIatrDoNeI-r . P. D. N, I . , 1987. Analysis of leneth l requencv dist r ibut ion. In: R. C. Suulrn 'nnnr"r &G. E. H, t r - r - (eds.) , Thc age and growth ol f ish: 371-38.1. ( Iowa State Univ. Press).
Ma' lsnws. Cl . P. , M. Al-Hosserur, A. R. AsluI- Gserren & N{. Ar, -SorrsNaNl, 1987. Assessment ofshort- l ived stocks wi th specia l re lèrence to Kuwai t 's shr imp f isher ies: a contrast of the resul tsobtained f iom tradi t ional and recent s ize-based techniques. In: D. Peulv & G. R. MonceN(eds.) , Length-based methods in f isher ies research. ICLAR\,{ Conf. Proc. , l3: 147-166.
N {evne r , A . , 1964 . C ro i ssance e t déve loppemen t chez l es c rus tacés . C .S .A . Spec ia l i s t Mee t . ,OAU/S ' IRC , 96 : 502 -648 .
.N{orrrr r , R. B. &J.J. PolovrNe, 1987. Distr ibut ion and y ie ld of deepwater shr imp Heterctcarpusresou rce i n t he N , I a r i anas . F i sh . Bu l l . . 85 i 2 ) : 339 -349 .
Naer. R. A. & R. C. N, IenIs, 1985. Fisher ies bio logy of shr imps and shr impl ike animals. In: Theb io l oev o f 'C rus tacea : l - l 10 . (Acad . P ress ) .
Onst Rnl IxI , L. & G. Rr,r . I r t , 1985. The red shr imps f ishery in the Ligur ian Sea: mismanasemento r no t? FAO F i sh . Rep . , 336 : 99 -106 .
P e t , r - r ' . D . & J . C a o o v , l 9 B 5 . A m o d i f i c a t i o n o f B h a t t a c h a r y a ' s m e t h o d f o r t h e a n a l v s i s o f m i x t u r eo l ' no rma l d i s t r i bu t i ons . FAO F i sh . ( l i r c . . 781 : l - 16 .
Perrr .v, D. ,J. INcr-ns & R. Nnel . t9t ì4. Appl i r :at ion to shr imp stocks of object ive methods for theest imat ion ol 'growth, morta l i ty and recrui tment-re lated parameters f rom length- f requencydata IELEFAN I and I I ) . In:J. A. Gur-r . lNr & B.J. Rornscnrro (eds.) , Penaeid shr imps: theirb io logv and manasement: 220-23.1. (Fishing News Books).
Reco:vese , S. , 19U9. Pr ima valutazione del lo stock di Ar istaeomorphafol iarea Risso, lB27 (Crustacea,A r i s t e i dae ) de l Cana le d i S i c i l i a . Nova Tha lass i a , l 0 ( supp l . l ) : 457 -465 .
, 1993. Geoeraphical d ist r ibut ion of Ar istaeomorpha fo l iacea (Crustacear Ar iste idae) in theSic i l ian Channel (Nfedi terrane an Sca). Rapp. Proc.-verb. Réun. ICES, ( in press). (Atso NTR-ITPP , 28 : l - 9 ) .
RecoNnsn, S. & \ { . L. BtarcnrNl , 1992. St ima dei parametr i d l cresci ta d i Ar isteus antennatus nelf l abna l c d i S i c i l i a . Oeba l i a , l 7 ( 2 ) : l 0 l - 107 .& , 1995. Size at sexual matur i ty in red shr imp females, Ar istaeomorphafol iacea, f romtheS i c i l i an Channe l (Med i t e r ranean Sea ) . f l r us taceana , 68 ( l ) .
RelsroN S., 1989. Fl f fect of seasonal recrui r -ment on bias of the Beverton-Hol t length-basedmor ta l i t y es t ima to r . Amer i can F i sh . Soc . Symp . , 6 : 190 -197 .
Rtcrr ,n, \ \ ' . E. , 1979. Grorvth rates and models. In: W. S. Hoen, D.J. ReNoar l &J. R. Bnarr
1eds.1, Fish physio logy, 3 (Bioenerget ics and growth:) : 677-7+3. (Academic Press, New York) .Rosnrnrnc, A. A. &J. R. Btol rucrou, 1988. Length-based methods of f ish stock assessment. In:J.
A. Gr i r .LeNo (ed.) , F ish populat ion d_vnamics: 83-103. (Wi ley & Sons, New York) .SeroÀ, F. , l9BB. Proeresos real izados en el conocimiento de la b io logia pesquera de la gamba,
Ar i . r taeus antennatus (Risso, lBl6) : ensayo de ela luación de bancos en el Medi terràneo Occi-den ta l . F -AO F i sh . Rep . , 395 : 76 -83 .
Sexoe. F. & N' I . DalrasrnE, lgB7. Estudio b io logico de la gamba Ar isteus antennatus (Risso, lBl6) enel Mar Catalan (NE de Espafra) . Invest . Pesq. (Barcelona), 5 l (suppl . 1) :213-232.

GROWTH AND MORTALITY OF RED SHRIMP
Seennl , P. , E. UnsIN & S. C. Vr;NEna, 1989. Introduct ion to t ropical l ìsh stock asscssmenr. Part l ,manua l . FAO F i sh . t echn . Pap . , 306 ( l ) : l - 337
Tav ron , C . C . . 1958 . Cod g row th and t empe ra ru re . Jou rn . Cons . C IE \ f , 23 : 366 -370 .Vlrrcn, E. F. , l9BB. Est imat ion of natural morta l i ty in f ish stocks: a re l ' ierv. Fish. Bul l . , 86 ( l ) :
2 5 - + 3 .\ \ ' r rxr :vsor, r , L. , l9BB. SYSTAT: rhe svstem f 'or stat is t ics: l -822. (Svstat Inc. , I ì l 'anston, IL) .\ ' . ' rHIeoul , M., A. Nouen & A. NInssr l l , 1986. Evalutat ion des stocks de deux espèces c le crevct tes
profondes de la famille des pénéidés: Aristeu.; anlennatu: et Paraltenaeus longira.rtris. FAO Fish.Rep . . 347 : 221 -231 .
36r
Received for publ icat ion I 2 Nlarch, I 993




















