
Going international versus going global. The Case of the European Steel Pipe SMES
Upload
independentCategory
view
2download
0
BIODIVERSITYRESEARCH
Going beyond taxonomic diversity:deconstructing biodiversity patternsreveals the true cost of iceplant invasionTommaso Jucker1, Marta Carboni2* and Alicia T. R. Acosta2
1Department of Plant Sciences, University of
Cambridge, Downing Street, Cambridge CB2
3EA, UK, 2Dipartimento di Biologia
Ambientale, Universit�a degli Studi di Roma
Tre, V.le Marconi 446, Roma 00146, Italy
*Correspondence: Marta Carboni,
Dipartimento di Biologia Ambientale,
Universit�a degli Studi di Roma Tre, V.le
Marconi 446, 00146 Roma, Italy.
E-mail: [email protected]
ABSTRACT
Aim Although invasion has been linked to species losses in native plant com-
munities, it is unclear how invasive species affect other important aspects of
native community biodiversity, such as the composition of functional traits or
the degree of phylogenetic relatedness. Here, we ask whether declines in taxo-
nomic diversity (TD) associated with the spread of a highly invasive South
African species (iceplant) are linked to similar losses in functional (FD) and
phylogenetic diversity (PD). Motivated by recent advances in coexistence the-
ory, we aim to infer the mechanisms involved in driving the exclusion of native
species following invasion.
Location Coastal dunes of central Italy.
Methods We sampled 2 9 2 m vegetation plots characterized by varying
degrees of abundance of iceplant (Carpobrotus spp.), and combined species
occurrence data with life-history trait information and a dated phylogeny of
the native species pool. Rao’s quadratic entropy index was used to quantify the
TD, FD and PD of each plot, which we then related to iceplant abundance via
linear models. Finally, to better understand the mechanisms driving changes in
diversity, we characterized both species and communities according to phyloge-
netic relatedness to iceplant and functional strategy.
Results We found that the negative association between the level of invasion
and native community TD is mirrored by quantitatively similar declines in FD,
as well as a shift in community phylogenetic structure. These changes appear to
result from the selective exclusion of specific functional groups/clades, likely via
a combination of niche- and fitness-related processes.
Main conclusions We found that iceplant poses a greater threat to biodiversity
than previously understood and that on top of causing declines in species rich-
ness invasion may also hamper ecosystem functioning and reduce evolutionary
potential. Accounting for FD and PD holds promise for gaining a better under-
standing of how invasive species alter the structure of native communities.
Keywords
Biological invasions, Carpobrotus spp., coastal dunes, competitive exclusion,
fitness hierarchy, functional and phylogenetic diversity, invasive species.
INTRODUCTION
Invasive species constitute a major threat to biodiversity, and
their impact on natural and semi-natural habitats is the
focus of much ecological research (Pimentel et al., 2001;
Gurevitch & Padilla, 2004; Ehrenfeld, 2010). Numerous studies
have explored how invasion affects patterns of native species
diversity (Davis, 2003; Vil�a et al., 2011; Py�sek et al., 2012).
Yet despite the advent of readily accessible molecular phylog-
enies and comprehensive trait databases which have helped
redefine how we quantify and study biodiversity (Purvis
& Hector, 2000; Pavoine & Bonsall, 2011), most studies
DOI: 10.1111/ddi.121241566 http://wileyonlinelibrary.com/journal/ddi ª 2013 John Wiley & Sons Ltd
Diversity and Distributions, (Diversity Distrib.) (2013) 19, 1566–1577A
Jou
rnal
of
Cons
erva
tion
Bio
geog
raph
yD
iver
sity
and
Dis
trib
utio
ns

continue to focus exclusively on how the number and rela-
tive abundance of native species (hereafter taxonomic diver-
sity, TD) is affected by invasion (Gaertner et al., 2009; Hejda
et al., 2009; Powell et al., 2011). In contrast, the impact of
invasives on other aspects of biodiversity such as functional
traits or phylogenetic relatedness remains unclear (e.g. Lamb-
don et al., 2008; Winter et al., 2009). Here, we investigate
how an invasive affects not only the TD of the native plant
communities it invades, but also their functional (FD) and
phylogenetic diversity (PD).
FD summarizes the diversity and variability of morpho-
logical, physiological and ecological traits present within a
community (Petchey & Gaston, 2006). The idea behind this
approach is to measure those traits which best capture a
species ecological niche. PD on the other hand is a measure
of the distance which separates each member of a commu-
nity when placed on a phylogeny (Purvis & Hector, 2000).
Assuming that closely related species are more functionally
similar than distant ones (Felsenstein, 1985; ‘phylogenetic
signal’ sensu Blomberg et al., 2001), the phylogenetic disper-
sion of a community should mirror its functional spectrum
(Cavender-Bares et al., 2004; Kraft et al., 2007; Vamosi
et al., 2009). Both the combination of functional traits and
the phylogenetic structure of a community have been shown
to play a key role in many ecological processes, such as
mediating species interactions and influencing community
assembly (Webb et al., 2002; HilleRisLambers et al., 2012),
determining responses to environmental change (Thuiller
et al., 2008; Swab et al., 2012) and driving ecosystem func-
tioning (Cadotte et al., 2011; Flynn et al., 2011). Conse-
quently, FD and PD are widely recognized as important and
complementary components of biodiversity and should be
studied alongside TD to better understand which factors
affect the structure and dynamics of plant communities
(Devictor et al., 2010), including responses to invasive
species.
Considerable effort has gone into understanding whether
certain traits make it more likely for species to become inva-
sive, as well as exploring how biodiversity (sensu lato) relates
to invasibility (Cadotte et al., 2009; MacDougall et al., 2009;
van Kleunen et al., 2010; Drenovsky et al., 2012). Less clear
is whether changes in competitive environment following
invasion, which have been linked to declines in TD, are also
altering the functional and phylogenetic structure of native
communities. Gerhold et al. (2011) reported that invasion
led to increased phylogenetic dispersion in recipient commu-
nities because of coexistence with, rather than replacement of
natives. It also appears that certain plant functional groups
may differ in their response to invasives (Davies, 2011). For
instance, Prunus serotina was shown to reduce FD in native
understorey plant communities by facilitating species with
specific trait combinations (Chabrerie et al., 2010). Overall,
these results suggest that focusing on multiple aspects of bio-
diversity can help us gain a more mechanistic understanding
of how invasives alter the structure of native plant
communities.
Determining how invasion might affect biodiversity pat-
terns requires an understanding of the processes which shape
FD and PD in plant communities. Traditionally, the
consensus has been that local scale diversity patterns essen-
tially depend on the combined effect of environmental filter-
ing (leading to the coexistence of ecologically similar
species), biotic interactions (leading to the exclusion of simi-
lar species, in accordance with niche theory) and neutral
processes related to dispersal and stochastic events (leading
to randomly assembled communities) (Hubbell, 2001; Webb
et al., 2002; Watkins & Wilson, 2003; Cavender-Bares et al.,
2004; Kraft et al., 2007). However, in the light of theoretical
advances in coexistence theory, this model of community
assembly has recently come into question (Mayfield &
Levine, 2010; HilleRisLambers et al., 2012). Crucially, as
Mayfield & Levine (2010) suggest, competitive interactions
often lead to the co-occurrence of functionally similar species
due to the exclusion of weaker competitors. To complicate
matters, further a number of other extrinsic factors also
affect local biodiversity patterns (e.g. spatial scales, regional
species pools, stress and disturbance; Pavoine & Bonsall,
2011; Brunbjerg et al., 2012; Mouillot et al., 2012). How the
disturbance caused by invasion affects the functional and
phylogenetic structure of communities is unknown.
We use Mediterranean coastal dune plant communities as
a model system to explore how the degree of invasion by the
South African succulent iceplant (Carpobrotus spp.) influ-
ences multiple aspects of native community biodiversity. Ice-
plant is considered a serious threat to biodiversity (Affre
et al., 2010) and has been associated with changes in coastal
ecosystems at a variety of scales (Vil�a et al., 2006; Conser &
Connor, 2009; Santoro et al., 2012b). Specifically, the pres-
ence of iceplant has been associated with a loss of TD in
native communities (Vil�a et al., 2006; Santoro et al., 2012a),
suggesting that the successful establishment of iceplant oper-
ates through the replacement and exclusion of incumbent
native species, rather than through coexistence with them.
However, it remains to be seen whether iceplant affects cer-
tain functional groups or phylogenetic lineages more than
others, thus resulting in altered patterns of FD and PD, or if
the loss of native species is generalized and independent of a
species ecological role. Assuming that iceplant invasion leads
to a decrease in TD (i.e. loss of species, as evidenced by pre-
vious studies), two alternative mechanisms of species exclu-
sion can be hypothesized, each leading to distinct FD and
PD patterns (Fig. 1):
H1)Generalized/Random impact: the loss of TD is not
associated with a decrease in FD and PD, which remain
unchanged. According to this scenario, iceplant randomly
excludes species from the community, independently of
their functional characteristics or phylogenetic clade. Ice-
plant competes equally with all species in the community,
probably for basic resources such as space, light or water.
H2)Directional/Selective impact: the decrease in TD is
coupled with a parallel decrease in FD and/or PD, as ice-
plant selectively excludes species that share a specific set
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1567
Invasion and biodiversity loss in plant communities

of functional traits or belong to a particular lineage. In
this scenario, the presence of iceplant could either lead to
the loss of species that share its ecological requirements
(limiting similarity hypothesis; H2a) or to the exclusion
of those that are competitively inferior (weaker
competitor exclusion hypothesis; H2b).
METHODS
Study area and vegetation sampling
We studied recent (Holocene) coastal dunes of central Italy
(Lazio Region) at six sites distributed along the length of
Figure 1 Conceptual diagram illustrating how alternative processes of species exclusion in response to invasion (left-hand side) can
lead to contrasting diversity patterns (right-hand side). Communities are characterized both in terms of their phylogenetic structure and
by a combination of two quantitative traits (symbolized by squares and triangles, which vary in sizes according to unitless values).
Following invasion, extant lineages are represented by filled symbols (black squares and triangles), while excluded species are shown as
empty symbols. According to the first hypothesis (H1; top row), invasion leads to the exclusion of a random subset of species (both in
terms of their position on the phylogeny and of their trait values). As a result, although TD declines in response to invasion, as long as
PD and FD are not correlated with TD, they both remain unaffected by the processes. Conversely, in the second scenario, the
introduction of an exotic species triggers the loss of specific lineages and traits from the species pool, leading to a decline in both PD
and FD. However, this same pattern can result from two very different processes. In the first instance (H2a), in accordance with the
limiting similarity hypothesis, the invader competes most strongly with those species that share its ecological requirements (in the
diagram, excluded species have a high degree of niche overlap with the invader in terms of their trait values). Instead, based on the
predictions of the weaker competitor exclusion hypothesis (H2b), the change in competitive regime brought on by invasion should lead
to the loss of competitively inferior species (here symbolized by smaller trait values).
1568 Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd
T. Jucker et al.

coastline (Fig. 2). The area is characterized by a Mediterra-
nean climate, with recent dunes generally occupying a nar-
row strip along the seashore. The dunes are not high
(<10 m) and are relatively simple in structure, with beaches
varying in breadth from a few metres to around 40 m, fol-
lowed by a section of low embryo-dunes, generally only one
mobile dune ridge, dune slacks and lastly a stabilized dune
zone (Acosta et al., 2003; Carboni et al., 2011). Vegetation
sampling was undertaken in spring (April–May) between
2006 and 2009. Randomly generated GPS coordinates were
used to define the location of 2 9 2 m vegetation plots
which we then sampled in the field. In each plot, we
recorded the presence of all vascular plant species (natives
and aliens), along with a measure of percentage cover for
each species using a 10% interval rank scale (for further
details on sampling and nomenclature, see Santoro et al.,
2012a).
A total of 414 plots were visited and 158 species recorded.
Based on previous work which identified six main vegetation
habitats along the sea-inland zonation (from pioneer com-
munities of the upper beach to shrubby communities in the
back dune; Carboni et al., 2011), each study plot was
assigned a habitat code (coding scheme follows that of the
European Habitats Directive; European Commission 1992).
Iceplant invasion occurs almost exclusively in the three habi-
tats which occupy the central sector of the zonation (EU
habitats 2110, 2210 and 2250; Carboni et al., 2011), which
are characterized by comparable levels of iceplant abundance
(habitat 2110: 37.7 � 4.5%; habitat 2210: 32.2 � 5.2%; habi-
tat 2250: 39.3 � 8.4%). For the purpose of this study, we
therefore restricted our analyses to plots found in this section
of the dunes. Furthermore, in order to test the extent to
which plot-level patterns of TD, FD and PD are affected by
the degree of iceplant invasion (quantified in terms of
percentage cover), we focused primarily on plots in which
iceplant was recorded (n = 58). Because exploring what
drives biodiversity patterns in native communities goes
beyond the focus of this study, data from non-invaded plots
were only used to provide context for certain results and
were excluded from further analyses unless explicitly stated.
Plant functional traits
Traits were compiled and measured for a subset of 46 domi-
nant species (Ricotta et al., 2012). Collectively, these selected
species account for ~80% of the standing live biomass in
each dune habitat. Seven life-history traits were selected
based on their relevance to the functional ecology of plants
in coastal dune environments: plant height (cm), SLA
(mm2 mg�1), LDMC, leaf size (cm2), life-form, dispersal
mode and pollination system. Quantitative traits (plant
height, SLA, LDMC, leaf size) were measured in the field or
laboratory by taking the mean of 10 different individuals.
Qualitative traits (life-form, dispersal mode, pollination sys-
tem) were obtained from the literature and were supple-
mented by personal observations in the field (for details see
Ricotta et al., 2012).
Phylogeny and phylogenetic signal
Using the complete species list, we constructed an aged phy-
logenetic tree using the open source software Phylomatic and
Phylocom (Webb et al., 2008). Branch lengths were assigned
using a branch length adjustment algorithm (BLADJ), based
on the minimum age of nodes estimated from the
fossil record. We then used a standard Mantel statistics
(Hardy & Pavoine, 2012) to test whether the multivariate
matrix of pair-wise functional dissimilarities between species
Figure 2 Study area. Locations where
vegetation sampling was undertaken are
marked by open squares.
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1569
Invasion and biodiversity loss in plant communities

(calculated with the Gower distance for mixed-variables) was
significantly correlated with the matrix of phylogenetic dis-
tances between species (summed branch lengths separating
pairs of species). Gower was chosen as it summarizes the
information of multiple traits (both quantitative and qualita-
tive) into a single metric, which for the purposes of this
study was preferable to analysing each trait individually. We
found a significant Mantel correlation (R = 0.215;
P = 0.001) between species’ pair-wise functional distances
and their phylogenetic relatedness, meaning that the traits we
selected exhibit strong phylogenetic signal at the species
level.
Quantifying taxonomic, functional and phylogenetic
diversity
Among the numerous metrics that exist to quantify biodiver-
sity patterns (Pavoine & Bonsall, 2011), we selected Rao’s
quadratic entropy index as it provides a framework for cal-
culating different biodiversity components (i.e. TD, FD and
PD) using a common methodological approach (de Bello
et al., 2010). This allows a much more robust comparison
between the different diversity measures. For a given plot, we
calculated Rao’s alpha diversity as follows:
aRao ¼Xs
i¼1
Xs
j¼1
dijpipj
where s is the number of species in the plot (species rich-
ness), dij is the distance between the species pair i and j,
weighted by the relative abundance pi and pj of the two spe-
cies (estimated here on the basis of percentage cover in the
plot). For FD and PD, the dissimilarity between species (dij)
is quantified using the multivariate functional distance
matrix (using the Gower distance) and the phylogenetic dis-
tance matrix, respectively (de Bello et al., 2010). In the case
of TD, where the distance between two species is fixed
(dij = 1, unless i = j in which case dij = 0), a Rao is equiva-
lent to the Simpson diversity index. All calculations were
performed in R (2.15; R Foundation for Statistical Comput-
ing, Vienna, Austria. http://www.R-project.org.) by adapting
code provided in the supplementary material of de Bello
et al. (2010).
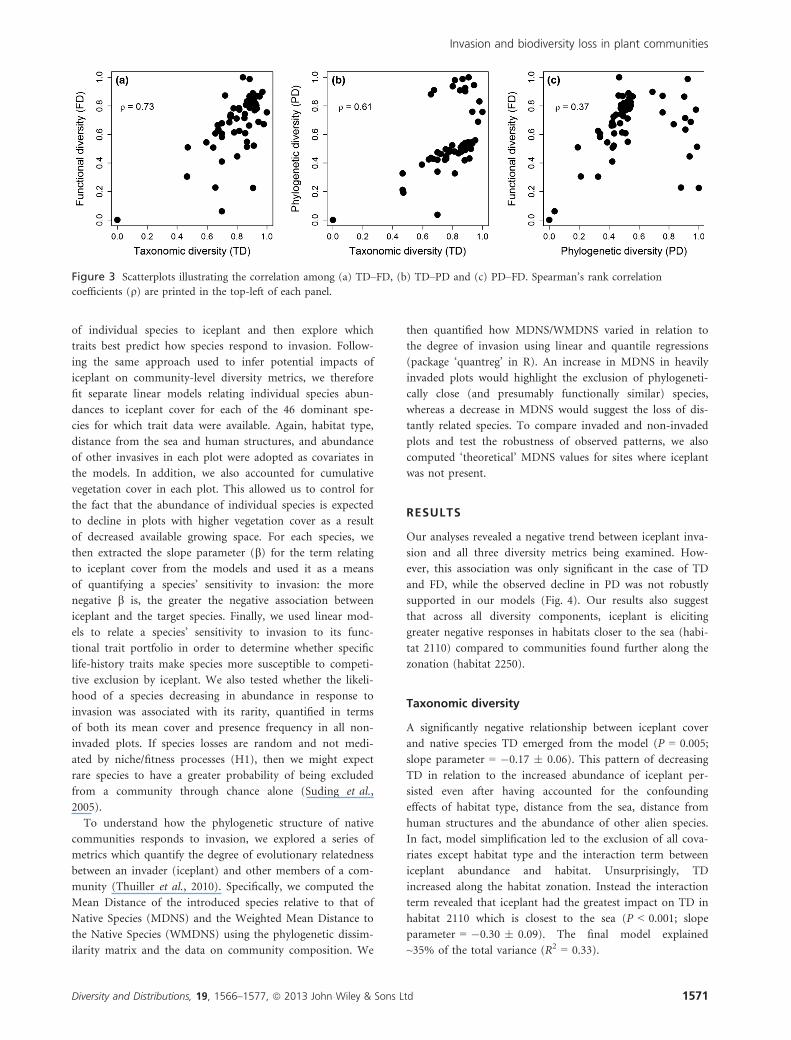
Accounting for collinearity among diversity metrics
As TD, FD and PD are derived from a common species
abundance matrix, they will inevitably exhibit a certain
degree of correlation between one another. Unsurprisingly,
Spearman’s rank correlation coefficients (q) among the three
diversity indices were positive (qTD:FD = 0.73; qTD:PD = 0.61;
qPD:FD = 0.37) and highly significant (P < 0.001; Fig. 3).
This strong collinearity poses a problem when attempting to
infer the mechanisms through which iceplant excludes native
species from a community (directional vs. generalized),
because any decline in TD will lead to a decrease in FD and
PD even if species loss is random. To account for this, we
first regressed FD and PD against TD (all logit-transformed)
and then adopted the residuals of these regressions as
response variables in our models (FDRES and PDRES). By
determining what portion of the residual variance is
explained by iceplant abundance, we were thus able to infer
the direct impact of invasion on a community’s functional
and phylogenetic structure (Devictor et al., 2010). As an
alternative approach, we accounted for variation in TD using
a null model that randomizes functional and phylogenetic
relationships among species to standardize FD and PD values
(see Appendix S1 in Supporting Information). Given that
both methods yielded almost identical results (Fig. S1), we
focus on the simpler regression residuals approach here.
Modelling TD, FD and PD as a function of iceplant
abundance
Linear models were fit for each of the three response vari-
ables (TD which we logit-transformed, FDRES and PDRES).
Iceplant percentage cover was adopted as the main explana-
tory variable. Alongside this, we accounted for the habitat
type of each plot to accommodate the fact that native plant
diversity differs markedly between habitats (hereafter referred
to using their respective European Habitats Directive codes:
2110, 2210 and 2250; Acosta et al., 2009). Importantly, an
interaction term between iceplant cover and habitat was also
fit to determine whether invasion impacts the three habitats
differently. Lastly, three further covariates were included in
the models to correct for additional sources of variation in
diversity patterns. A measure of distance from the sea of
each plot was calculated in a GIS environment (ArcGIS 9.2,
ESRI, Redlands, CA, USA) and used as a proxy of environ-
mental harshness (salt spray, sand burial, soil aridity, salinity
and nutrient shortage are all known to decrease with distance
from the sea; Carboni et al., 2011). Similarly, the distance
from the nearest human structure was also computed and
adopted as an indicator of human disturbance (Alston &
Richardson, 2006; Carboni et al., 2011). Finally, the abun-
dance of all other alien plants was used to account for the
potential influence of other invasive species. Model simplifi-
cation was performed by implementing stepwise model selec-
tion (forward and backward) based on AIC.
Determining which phylogenetic lineages and
functional traits are targeted by iceplant
Two complementary approaches were taken to determine
whether species being excluded by iceplant are more likely
to be functionally and phylogenetically similar (in accor-
dance with the limiting similarity hypothesis) or different
from the invader (suggesting the weaker competitor exclu-
sion hypothesis): one exploring individual species responses
and the other focusing on community-level patterns.
Assuming that iceplant selectively excludes species from a
community, it should be possible to quantify the sensitivity
1570 Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd
T. Jucker et al.
of individual species to iceplant and then explore which
traits best predict how species respond to invasion. Follow-
ing the same approach used to infer potential impacts of
iceplant on community-level diversity metrics, we therefore
fit separate linear models relating individual species abun-
dances to iceplant cover for each of the 46 dominant spe-
cies for which trait data were available. Again, habitat type,
distance from the sea and human structures, and abundance
of other invasives in each plot were adopted as covariates in
the models. In addition, we also accounted for cumulative
vegetation cover in each plot. This allowed us to control for
the fact that the abundance of individual species is expected
to decline in plots with higher vegetation cover as a result
of decreased available growing space. For each species, we
then extracted the slope parameter (b) for the term relating
to iceplant cover from the models and used it as a means
of quantifying a species’ sensitivity to invasion: the more
negative b is, the greater the negative association between
iceplant and the target species. Finally, we used linear mod-
els to relate a species’ sensitivity to invasion to its func-
tional trait portfolio in order to determine whether specific
life-history traits make species more susceptible to competi-
tive exclusion by iceplant. We also tested whether the likeli-
hood of a species decreasing in abundance in response to
invasion was associated with its rarity, quantified in terms
of both its mean cover and presence frequency in all non-
invaded plots. If species losses are random and not medi-
ated by niche/fitness processes (H1), then we might expect
rare species to have a greater probability of being excluded
from a community through chance alone (Suding et al.,
2005).
To understand how the phylogenetic structure of native
communities responds to invasion, we explored a series of
metrics which quantify the degree of evolutionary relatedness
between an invader (iceplant) and other members of a com-
munity (Thuiller et al., 2010). Specifically, we computed the
Mean Distance of the introduced species relative to that of
Native Species (MDNS) and the Weighted Mean Distance to
the Native Species (WMDNS) using the phylogenetic dissim-
ilarity matrix and the data on community composition. We
then quantified how MDNS/WMDNS varied in relation to
the degree of invasion using linear and quantile regressions
(package ‘quantreg’ in R). An increase in MDNS in heavily
invaded plots would highlight the exclusion of phylogeneti-
cally close (and presumably functionally similar) species,
whereas a decrease in MDNS would suggest the loss of dis-
tantly related species. To compare invaded and non-invaded
plots and test the robustness of observed patterns, we also
computed ‘theoretical’ MDNS values for sites where iceplant
was not present.
RESULTS
Our analyses revealed a negative trend between iceplant inva-
sion and all three diversity metrics being examined. How-
ever, this association was only significant in the case of TD
and FD, while the observed decline in PD was not robustly
supported in our models (Fig. 4). Our results also suggest
that across all diversity components, iceplant is eliciting
greater negative responses in habitats closer to the sea (habi-
tat 2110) compared to communities found further along the
zonation (habitat 2250).
Taxonomic diversity
A significantly negative relationship between iceplant cover
and native species TD emerged from the model (P = 0.005;
slope parameter = �0.17 � 0.06). This pattern of decreasing
TD in relation to the increased abundance of iceplant per-
sisted even after having accounted for the confounding
effects of habitat type, distance from the sea, distance from
human structures and the abundance of other alien species.
In fact, model simplification led to the exclusion of all cova-
riates except habitat type and the interaction term between
iceplant abundance and habitat. Unsurprisingly, TD
increased along the habitat zonation. Instead the interaction
term revealed that iceplant had the greatest impact on TD in
habitat 2110 which is closest to the sea (P < 0.001; slope
parameter = �0.30 � 0.09). The final model explained
~35% of the total variance (R2 = 0.33).
Figure 3 Scatterplots illustrating the correlation among (a) TD–FD, (b) TD–PD and (c) PD–FD. Spearman’s rank correlation
coefficients (q) are printed in the top-left of each panel.
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1571
Invasion and biodiversity loss in plant communities
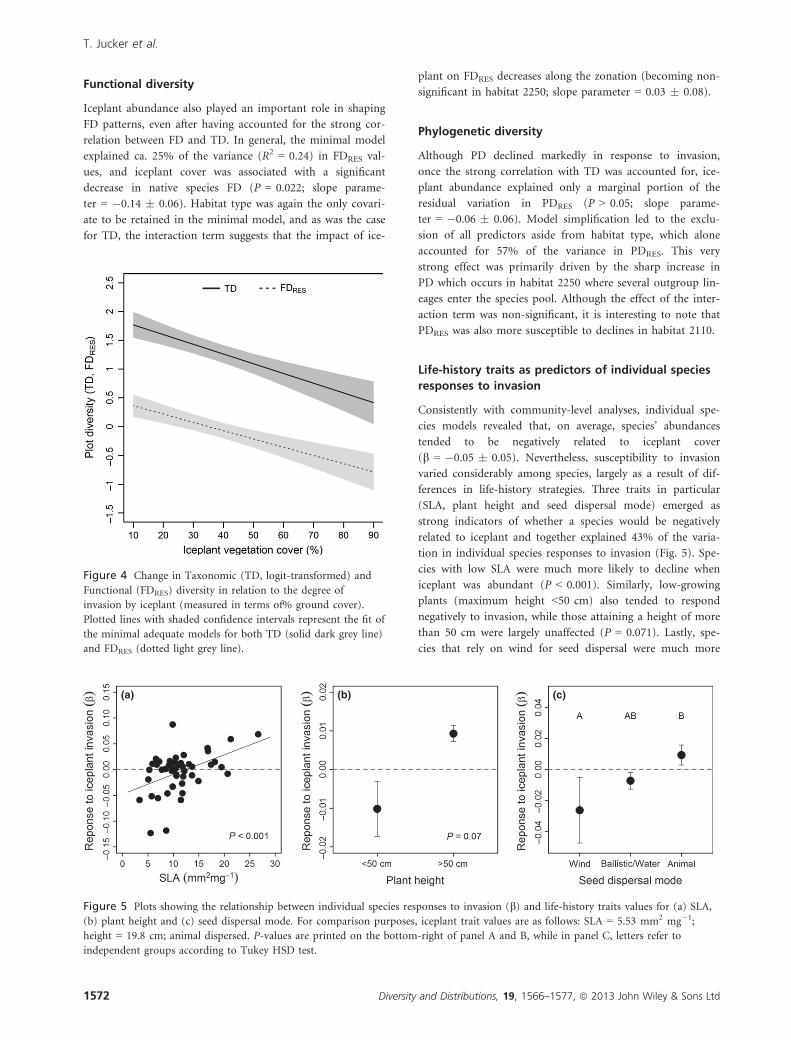
Functional diversity
Iceplant abundance also played an important role in shaping
FD patterns, even after having accounted for the strong cor-
relation between FD and TD. In general, the minimal model
explained ca. 25% of the variance (R2 = 0.24) in FDRES val-
ues, and iceplant cover was associated with a significant
decrease in native species FD (P = 0.022; slope parame-
ter = �0.14 � 0.06). Habitat type was again the only covari-
ate to be retained in the minimal model, and as was the case
for TD, the interaction term suggests that the impact of ice-
plant on FDRES decreases along the zonation (becoming non-
significant in habitat 2250; slope parameter = 0.03 � 0.08).
Phylogenetic diversity
Although PD declined markedly in response to invasion,
once the strong correlation with TD was accounted for, ice-
plant abundance explained only a marginal portion of the
residual variation in PDRES (P > 0.05; slope parame-
ter = �0.06 � 0.06). Model simplification led to the exclu-
sion of all predictors aside from habitat type, which alone
accounted for 57% of the variance in PDRES. This very
strong effect was primarily driven by the sharp increase in
PD which occurs in habitat 2250 where several outgroup lin-
eages enter the species pool. Although the effect of the inter-
action term was non-significant, it is interesting to note that
PDRES was also more susceptible to declines in habitat 2110.
Life-history traits as predictors of individual species
responses to invasion
Consistently with community-level analyses, individual spe-
cies models revealed that, on average, species’ abundances
tended to be negatively related to iceplant cover
(b = �0.05 � 0.05). Nevertheless, susceptibility to invasion
varied considerably among species, largely as a result of dif-
ferences in life-history strategies. Three traits in particular
(SLA, plant height and seed dispersal mode) emerged as
strong indicators of whether a species would be negatively
related to iceplant and together explained 43% of the varia-
tion in individual species responses to invasion (Fig. 5). Spe-
cies with low SLA were much more likely to decline when
iceplant was abundant (P < 0.001). Similarly, low-growing
plants (maximum height <50 cm) also tended to respond
negatively to invasion, while those attaining a height of more
than 50 cm were largely unaffected (P = 0.071). Lastly, spe-
cies that rely on wind for seed dispersal were much more
Figure 4 Change in Taxonomic (TD, logit-transformed) and
Functional (FDRES) diversity in relation to the degree of
invasion by iceplant (measured in terms of% ground cover).
Plotted lines with shaded confidence intervals represent the fit of
the minimal adequate models for both TD (solid dark grey line)
and FDRES (dotted light grey line).
(a) (b) (c)
Figure 5 Plots showing the relationship between individual species responses to invasion (b) and life-history traits values for (a) SLA,
(b) plant height and (c) seed dispersal mode. For comparison purposes, iceplant trait values are as follows: SLA = 5.53 mm2 mg�1;
height = 19.8 cm; animal dispersed. P-values are printed on the bottom-right of panel A and B, while in panel C, letters refer to
independent groups according to Tukey HSD test.
1572 Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd
T. Jucker et al.
strongly impacted than those that disperse through other
abiotic mechanisms (ballistic, water) or are spread by ani-
mals. In contrast to life-history traits, we found no evidence
to suggest that susceptibility to iceplant is associated with the
commonness or rarity of a species (Fig. S2).
Changes in community phylogenetic structure
following invasion
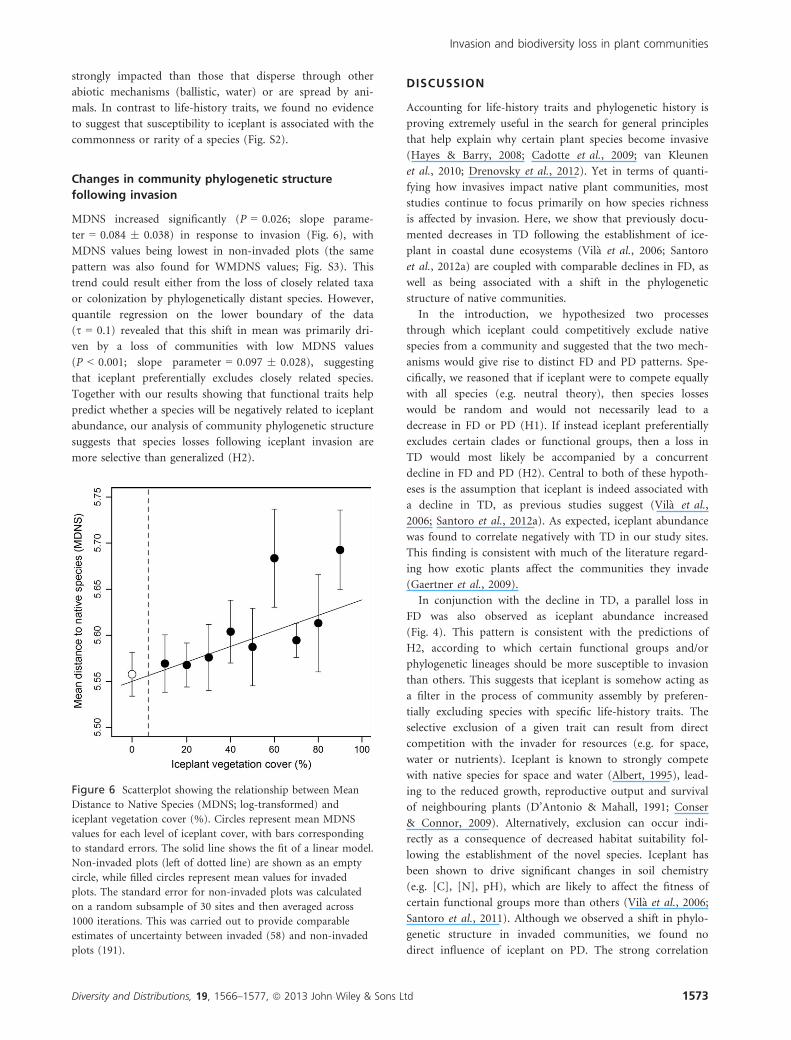
MDNS increased significantly (P = 0.026; slope parame-
ter = 0.084 � 0.038) in response to invasion (Fig. 6), with
MDNS values being lowest in non-invaded plots (the same
pattern was also found for WMDNS values; Fig. S3). This
trend could result either from the loss of closely related taxa
or colonization by phylogenetically distant species. However,
quantile regression on the lower boundary of the data
(s = 0.1) revealed that this shift in mean was primarily dri-
ven by a loss of communities with low MDNS values
(P < 0.001; slope parameter = 0.097 � 0.028), suggesting
that iceplant preferentially excludes closely related species.
Together with our results showing that functional traits help
predict whether a species will be negatively related to iceplant
abundance, our analysis of community phylogenetic structure
suggests that species losses following iceplant invasion are
more selective than generalized (H2).
DISCUSSION
Accounting for life-history traits and phylogenetic history is
proving extremely useful in the search for general principles
that help explain why certain plant species become invasive
(Hayes & Barry, 2008; Cadotte et al., 2009; van Kleunen
et al., 2010; Drenovsky et al., 2012). Yet in terms of quanti-
fying how invasives impact native plant communities, most
studies continue to focus primarily on how species richness
is affected by invasion. Here, we show that previously docu-
mented decreases in TD following the establishment of ice-
plant in coastal dune ecosystems (Vil�a et al., 2006; Santoro
et al., 2012a) are coupled with comparable declines in FD, as
well as being associated with a shift in the phylogenetic
structure of native communities.
In the introduction, we hypothesized two processes
through which iceplant could competitively exclude native
species from a community and suggested that the two mech-
anisms would give rise to distinct FD and PD patterns. Spe-
cifically, we reasoned that if iceplant were to compete equally
with all species (e.g. neutral theory), then species losses
would be random and would not necessarily lead to a
decrease in FD or PD (H1). If instead iceplant preferentially
excludes certain clades or functional groups, then a loss in
TD would most likely be accompanied by a concurrent
decline in FD and PD (H2). Central to both of these hypoth-
eses is the assumption that iceplant is indeed associated with
a decline in TD, as previous studies suggest (Vil�a et al.,
2006; Santoro et al., 2012a). As expected, iceplant abundance
was found to correlate negatively with TD in our study sites.
This finding is consistent with much of the literature regard-
ing how exotic plants affect the communities they invade
(Gaertner et al., 2009).
In conjunction with the decline in TD, a parallel loss in
FD was also observed as iceplant abundance increased
(Fig. 4). This pattern is consistent with the predictions of
H2, according to which certain functional groups and/or
phylogenetic lineages should be more susceptible to invasion
than others. This suggests that iceplant is somehow acting as
a filter in the process of community assembly by preferen-
tially excluding species with specific life-history traits. The
selective exclusion of a given trait can result from direct
competition with the invader for resources (e.g. for space,
water or nutrients). Iceplant is known to strongly compete
with native species for space and water (Albert, 1995), lead-
ing to the reduced growth, reproductive output and survival
of neighbouring plants (D’Antonio & Mahall, 1991; Conser
& Connor, 2009). Alternatively, exclusion can occur indi-
rectly as a consequence of decreased habitat suitability fol-
lowing the establishment of the novel species. Iceplant has
been shown to drive significant changes in soil chemistry
(e.g. [C], [N], pH), which are likely to affect the fitness of
certain functional groups more than others (Vil�a et al., 2006;
Santoro et al., 2011). Although we observed a shift in phylo-
genetic structure in invaded communities, we found no
direct influence of iceplant on PD. The strong correlation
Figure 6 Scatterplot showing the relationship between Mean
Distance to Native Species (MDNS; log-transformed) and
iceplant vegetation cover (%). Circles represent mean MDNS
values for each level of iceplant cover, with bars corresponding
to standard errors. The solid line shows the fit of a linear model.
Non-invaded plots (left of dotted line) are shown as an empty
circle, while filled circles represent mean values for invaded
plots. The standard error for non-invaded plots was calculated
on a random subsample of 30 sites and then averaged across
1000 iterations. This was carried out to provide comparable
estimates of uncertainty between invaded (58) and non-invaded
plots (191).
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1573
Invasion and biodiversity loss in plant communities

between PD and TD could partly account for this, because
little residual variation in PD remained for iceplant abun-
dance to explain. A better resolved molecular phylogeny
could potentially provide the phylogenetic precision needed
to detect shifts in PD.
Aside from revealing that iceplant influences more than just
TD, our results also hint to which processes are driving the
competitive exclusion of species. For much of the last decade,
the prevalent theory has been that regional species pools are
constrained by habitat filtering, while at the community-level
coexistence, patterns can be explained by the limiting similar-
ity hypothesis (H2a), which predicts that ecologically similar
species will exclude one another due to strong niche overlap
(Webb et al., 2002; Emerson & Gillespie, 2008). However, an
alternative model based on coexistence theory (Chesson,
2000) attempts to explain why species co-occurrence patterns
often clash with traditional expectations (HilleRisLambers
et al., 2012). This framework proposes that the outcome of
competition among species depends on the interplay between
niche differences (ecological similarity) and relative fitness dif-
ferences (competitive ability), thus explaining why it is often
the competitively weaker species that are excluded (H2b). Our
analyses suggest that changes in the functional and phyloge-
netic structure of native communities associated with high
levels of iceplant invasion may be the result of two separate
processes. In line with the predictions of limiting similarity
hypothesis (H2a), iceplant appears to preferentially exclude
closely related and ecologically similar taxa (Figs 5 & 6). Low-
growing species with small leaf surface area-to-weight ratios
(similar to iceplant: height = 19.8 cm; SLA = 5.53 mm2
mg�1) were much more likely to decrease in abundance in
response to invasion. In contrast, species that grow above ice-
plant mats and/or have high SLA were largely unaffected, pos-
sibly as a result of their ability to avoid excessive shading
(Weiner & Thomas, 1986; Lep�s, 1999).
However, iceplant also had a strong impact on species that
are wind dispersed. Limited seed reserves may make it harder
for wind dispersed species to germinate and grow under the
suboptimal conditions associated with iceplant mats (e.g.
altered soil chemistry, reduced water and light availability;
D’Antonio & Mahall, 1991; Vil�a et al., 2006; Conser & Con-
nor, 2009; Santoro et al., 2011). The fact that iceplant may
be limiting the distribution of wind dispersed species, many
of which are ephemeral and characterized by annual life
cycles, suggests that weaker competitors are also being
excluded from the species pool (H2b). This is consistent with
previous studies showing that annual grasses (therophytes)
are particularly sensitive to iceplant (Vil�a et al., 2006; Andreu
et al., 2010), a pattern which we also find (Fig. S4). More-
over, it is supported by the fact that iceplant had the strong-
est impact in foredune communities where many species
have annual life cycles (H2b) and/or adopt creeping growth
strategies (H2a). Although seemingly contradictory, the fact
that both weaker competitors and ecologically similar species
declined in the presence of iceplant is precisely what
coexistence theory would predict. Not only are niche- and
fitness-related processes non-mutually exclusive, it is their
combined effect which ultimately shapes community
assembly and species exclusion following invasion.
As with any correlative study, caution is needed when inter-
preting results (Renter�ıa et al., 2012). Because we have no
information on the floral composition of invaded plots prior
to invasion, it is impossible to determine whether the lower
diversity of these plots is a direct consequence of invasion or
simply a pre-existing pattern which facilitated iceplant estab-
lishment (MacDougall & Turkington, 2005; Carboni et al.,
2013). Nevertheless, we feel there are several reasons why our
interpretation of the data is well supported by our analysis.
First, ours is not the first study to identify a change in commu-
nity structure and diversity following invasion by iceplant,
even though previous work had not explored FD and PD pat-
terns. Furthermore, because our analysis relates the loss of
diversity to the relative abundance of iceplant (and not to its
presence/absence), we were able to show that TD and FD pro-
gressively decline as iceplant becomes more abundant. If ice-
plant is not driving species losses and is merely a passenger
along for the ride (MacDougall & Turkington, 2005), we might
instead expect little difference in diversity among weakly and
strongly invaded plots. Finally, iceplant abundance was nega-
tively associated with TD and FD even after having accounted
for environmental stress and human disturbance, suggesting
the patterns we find are not driven by external factors. Regard-
less, future experimental and observational work is needed to
settle this debate. Long-term monitoring, both in the field and
under controlled experimental settings, would allow changes
in community composition to be assessed over time. Con-
versely, removal trials could help determine whether currently
invaded plots shift back to baseline conditions, or if these com-
munities were in fact compositionally distinct to start with.
The need to redefine conservation priorities and adapt
management strategies to include aspects such as functional
and phylogenetic diversity is clear (Mace et al., 2003; Forest
et al., 2007; Cadotte et al., 2011; Rolland et al., 2012). Real-
izing that invasive species may affect not only patterns of
species richness, but also hamper ecosystem functioning and
reduce evolutionary potential is an important step in this
process. Conservation efforts assessing the impact of inva-
sives need to consider that native communities can be
affected in more ways than one, or underlying changes could
be inadvertently missed. Just as importantly, in order to
develop effective management strategies to safeguard biodi-
versity, the knowledge that some species may be more threa-
tened than others by invasion is crucial. In the hope of
preserving the functionality and uniqueness of coastal dune
ecosystems, mitigating the spread of exotic species such as
iceplant should be of special concern.
ACKNOWLEDGEMENTS
We are grateful to R. Santoro for helping us develop and
refine the questions explored here. Three anonymous
reviewers, along with editors David Richardson and Mark
1574 Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd
T. Jucker et al.

van Kleunen, were instrumental in helping us improve on
earlier drafts. Lastly, we thank S. Del Vecchio and I. Prisco
for helping with fieldwork.
REFERENCES
Acosta, A.T.R., Stanisci, A., Ercole, S. & Blasi, C. (2003)
Sandy coastal landscape of the Lazio region (Central Italy).
Phytocoenologia, 33, 715–726.
Acosta, A.T.R., Carranza, M.L. & Izzi, C.F. (2009) Are
there habitats that contribute best to plant species
diversity in coastal dunes? Biodiversity Conservation, 18,
1087–1098.
Affre, L., Suehs, C.M., Charpentier, S., Vil�a, M., Brundu, G.,
Lambdon, P., Traveset, A. & Hulme, P.E. (2010) Consis-
tency in the habitat degree of invasion for three invasive
plant species across Mediterranean islands. Biological Inva-
sions, 12, 2537–2548.
Albert, M.E. (1995) Portrait of an invader II: the ecology and
management of Carpobrotus edulis. CalEPPC News, 3, 4–6.
Alston, K.P. & Richardson, D.M. (2006) The roles of habitat
features, disturbance, and distance from putative source
populations in structuring alien plant invasions at the
urban/wildland interface on the Cape Peninsula, South
Africa. Biological Conservation, 132, 183–198.
Andreu, J., Manzano-Piedras, E., Bartomeus, I., Dana, E.D.
& Vil�a, M. (2010) Vegetation response after removal of the
invasive Carpobrotus hybrid complex in Andaluc�ıa, Spain.
Ecological Restoration, 28, 440–448.
de Bello, F., Lavergne1, S., Meynard, C.N., Lep�s, J. & Thuil-
ler, W. (2010) The partitioning of diversity: showing
Theseus a way out of the labyrinth. Journal of Vegetation
Science, 21, 992–1000.
Blomberg, S.P., Garland, T. & Ives, A.R. (2001) Testing for
phylogenetic signal in comparative data: behavioral traits
are more labile. Evolution, 57, 717–745.
Brunbjerg, A.K., Borchsenius, F., Eiserhardt, W.L., Ejrnæs, R.
& Svenning, J.C. (2012) Disturbance drives phylogenetic
community structure in coastal dune vegetation. Journal of
Vegetation Science, 23, 1082–1094.
Cadotte, M.W., Hamilton, M.A. & Murra, B.R. (2009) Phy-
logenetic relatedness and plant invader success across two
spatial scales. Diversity and Distributions, 15, 481–488.
Cadotte, M.W., Carscadden, K. & Mirotchnick, N. (2011)
Beyond species: functional diversity and the maintenance
of ecological processes and services. Journal of Applied Ecol-
ogy, 48, 1079–1087.
Carboni, M., Santoro, R. & Acosta, A.T.R. (2011) Dealing
with scarce data to understand how environmental gradi-
ents and propagule pressure shape fine-scale alien distribu-
tion patterns on coastal dunes. Journal of Vegetation
Science, 22, 751–765.
Carboni, M., M€unkem€uller, T., Gallien, L., Lavergne, S., Aco-
sta, A. & Thuiller, W. (2013) Darwin’s naturalization
hypothesis: scale matters in coastal plant communities.
Ecography, 36, 560–568.
Cavender-Bares, J., Ackerly, D.D., Baum, D.A. & Bazzaz, F.A.
(2004) Phylogenetic overdispersion in floridian oak com-
munities. The American Naturalist, 163, 823–843.
Chabrerie, O., Loinard, J., Perrin, S., Saguez, R. & Decocq,
G. (2010) Impact of Prunus serotina invasion on under-
story functional diversity in a European temperate forest.
Biological Invasions, 12, 1891–1907.
Chesson, P. (2000) Mechanisms of maintenance of species
diversity. Annual Review of Ecology and Systematics, 31,
343–366.
Conser, C. & Connor, E.F. (2009) Assessing the residual
effects of Carpobrotus edulis invasion, implications for res-
toration. Biological Invasions, 11, 349–358.
D’Antonio, C.M. & Mahall, B.E. (1991) Root profiles and
competition between the invasive, exotic perennial,
Carpobrotus edulis, and two native shrub species in
California coastal scrub. American Journal of Botany, 78,
885–894.
Davies, K.W. (2011) Plant community diversity and native
plant abundance decline with increasing abundance of an
exotic annual grass. Oecologia, 167, 481–491.
Davis, M.A. (2003) Biotic globalization: does competition
from introduced species threaten biodiversity? BioScience,
53, 482–489.
Devictor, V., Mouillot, D., Meynard, C., Jiguet, F., Thuiller,
W. & Mouquet, N. (2010) Spatial mismatch and congru-
ence between taxonomic, phylogenetic and functional
diversity: the need for integrative conservation strategies in
a changing world. Ecology Letters, 13, 1030–1040.
Drenovsky, R.E., Grewell, B.J., D’Antonio, C.M., Funk, J.L.,
James, J.J., Molinari, N., Parker, I.M. & Richards, C.L.
(2012) A functional trait perspective on plant invasion.
Annals of Botany, 110, 141–153.
Ehrenfeld, J.G. (2010) Ecosystem consequences of biological
invasions. Annual Review of Ecology, Evolution, and System-
atics, 41, 59–80.
Emerson, B.C. & Gillespie, R.G. (2008) Phylogenetic analysis
of community assembly and structure over space and time.
Trends in Ecology and Evolution, 23, 619–630.
Felsenstein, J. (1985) Phylogenies and the comparative
method. The American Naturalist, 125, 1–15.
Flynn, D.F.B., Mirotchnick, N., Jain, M., Palmer, M.I. &
Naeem, S. (2011) Functional and phylogenetic diversity as
predictors of biodiversity–ecosystem-function relationships.
Ecology, 92, 1573–1581.
Forest, F., Grenyer, R., Rouget, M., Davies, T.J., Cowling,
R.M., Faith, D.P., Balmford, A., Manning, J.C., Proche, S.,
van der Bank, M., Reeves, G., Hedderson, T.A.J. & Savolai-
nen, V. (2007) Preserving the evolutionary potential of flo-
ras in biodiversity hotspots. Nature, 445, 757–760.
Gaertner, M., Bree€yen, A.D., Hui, C. & Richardson, D.M.
(2009) Impacts of alien plant invasions on species richness
in Mediterranean-type ecosystems: a meta-analysis. Progress
in Physical Geography, 33, 319–338.
Gerhold, P., P€artel, M., Tackenberg, O., Hennekens, S.M.,
Bartish, I., Schamin�ee, J.H., Fergus, A.J., Ozinga, W.A. &
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1575
Invasion and biodiversity loss in plant communities

Prinzing, A. (2011) Phylogenetically poor plant communi-
ties receive more alien species, which more easily coexist
with natives. The American Naturalist, 177, 668–680.
Gurevitch, J. & Padilla, D.K. (2004) Are invasive species a
major cause of extinctions? Trends in Ecology and Evolu-
tion, 19, 470–474.
Hardy, O. & Pavoine, S. (2012) Assessing phylogenetic signal
with measurement error: a comparison of Mantel tests,
Blomberg et al’.s K and phylogenetic distograms. Evolution,
66, 2614–2621.
Hayes, K.R. & Barry, S.C. (2008) Are there any consistent
predictors of invasion success? Biological Invasions, 10,
483–506.
Hejda, M., Py�sek, P. & Jaro�s�ık, V. (2009) Impact of invasive
plants on the species richness, diversity and composition of
invaded communities. Journal of Ecology, 97, 393–403.
HilleRisLambers, J., Adler, P.B., Harpole, W.S., Levine, J.M.
& Mayfield, M.M. (2012) Rethinking community assembly
through the lens of coexistence theory. Annual Review of
Ecology, Evolution, and Systematics, 43, 227–248.
Hubbell, S.P. (2001) The unified neutral theory of biodiversity
and biogeography. Princeton University Press, Princeton,
NJ.
van Kleunen, M., Weber, E. & Fischer, M. (2010) A meta-
analysis of trait differences between invasive and non-inva-
sive plant species. Ecology Letters, 13, 235–245.
Kraft, N.J.B., Cornwell, W.K., Webb, C.O. & Ackerly, D.D.
(2007) Trait evolution, community assembly, and the phy-
logenetic structure of ecological communities. The Ameri-
can Naturalist, 170, 271–283.
Lambdon, P.W., Lloret, F. & Hulme, P.E. (2008) Do non-
native species invasions lead to biotic homogenization at
small scales? The similarity and functional diversity of
habitats compared for alien and native components
of Mediterranean floras. Diversity and Distributions, 14,
774–785.
Lep�s, J. (1999) Nutrient status, disturbance and competition:
an experimental test of relationships in a wet meadow
copy. Journal of Vegetation Science, 10, 219–230.
MacDougall, A.S. & Turkington, R. (2005) Are invasive spe-
cies the drivers or passengers of change in degraded ecosys-
tems? Ecology, 86, 42–55.
MacDougall, A.S., Gilbert, B. & Levine, J.M. (2009) Plant
invasions and the niche. Journal of Ecology, 97, 609–615.
Mace, G.M., Gittleman, J.L. & Purvis, A. (2003) Preserving
the tree of life. Science, 300, 1707–1709.
Mayfield, M.M. & Levine, J.M. (2010) Opposing effects of
competitive exclusion on the phylogenetic structure of
communities. Ecology Letters, 13, 1085–1093.
Mouillot, D., Graham, N.A.J., Vill�eger, S., Mason, N.W.H. &
Bellwood, D.R. (2012) A functional approach reveals com-
munity responses to disturbances. Trends in Ecology and
Evolution, 28, 167–177.
Pavoine, S. & Bonsall, M.B. (2011) Measuring biodiversity to
explain community assembly: a unified approach. Biological
Reviews, 86, 792–812.
Petchey, O.L. & Gaston, K.J. (2006) Functional diversity:
back to basics and looking forward. Ecology Letters, 9,
741–758.
Pimentel, D., McNair, S., Janecka, J., Wightman, J., Sim-
monds, C., O’Connell, C., Wong, E., Russel, E., Zern, J.,
Aquino, T. & Tsomondo, T. (2001) Economic and envi-
ronmental threats of alien plant, animal, and microbe inva-
sions. Agriculture Ecosystems & Environment, 84, 1–20.
Powell, K.I., Chase, J.M. & Knight, T.M. (2011) A synthesis
of plant invasion effects on biodiversity across spatial
scales. American Journal of Botany, 98, 539–548.
Purvis, A. & Hector, A. (2000) Getting the measure of biodi-
versity. Nature, 405, 212–219.
Py�sek, P., Jaro�s�ık, V., Hulme, P.E., Pergl, J., Hejda, M.,
Schaffner, U. & Vil�a, M. (2012) A global assessment of
invasive plant impacts on resident species, communities
and ecosystems: the interaction of impact measures, invad-
ing species’ traits and environment. Global Change Biology,
18, 1725–1737.
Renter�ıa, J.L., Gardener, M.R., Panetta, F.D., Atkinson, R. &
Crawley, M.J. (2012) Possible impacts of the invasive plant
Rubus niveus on the native vegetation of the Scalesia forest
in the Galapagos Islands. PLoS ONE, 7, e48106.
Ricotta, C., Pavoine, S., Bacaro, G. & Acosta, A.T.R. (2012)
Functional rarefaction for species abundance data. Methods
in Ecology and Evolution, 3, 519–525.
Rolland, J., Cadotte, M.W., Davies, J., Devictor, V., Lavergne,
S., Mouquet, N., Pavoine, S., Rodrigues, A., Thuiller, W.,
Turcati, L., Winter, M., Zupan, L., Jabot, F. & Morlon, H.
(2012) Using phylogenies in conservation: new perspec-
tives. Biology Letters, 8, 692–694.
Santoro, R., Jucker, T., Carranza, M.L. & Acosta, A.T.R.
(2011) Assessing the effects of Carpobrotus invasion on
coastal dune soils. Does the nature of the invaded habitat
matter? Community Ecology, 12, 234–240.
Santoro, R., Carboni, M., Carranza, M.L. & Acosta, A.T.R.
(2012a) Focal species diversity patterns can provide diag-
nostic information on plant invasions. Journal for Nature
Conservation, 20, 85–91.
Santoro, R., Jucker, T., Carboni, M. & Acosta, A.T.R.
(2012b) Patterns of plant community assembly in invaded
and non-invaded communities along a natural environ-
mental gradient. Journal of Vegetation Science, 23, 483–494.
Suding, K.N., Collins, S.L., Gough, L., Clark, C., Cleland,
E.E., Gross, K.L., Milchunas, D.G. & Pennings, S. (2005)
Functional- and abundance-based mechanisms explain
diversity loss due to N fertilization. Proceedings of the
National Academy of Sciences of the USA, 102, 4387–4392.
Swab, R.M., Regan, H.M., Keith, D.A., Regan, T.J. & Ooi,
M.K.J. (2012) Niche models tell half the story: spatial con-
text and life-history traits influence species responses to
global change. Journal of Biogeography, 39, 1266–1277.
Thuiller, W., Albert, C., Araujo, M.B., Berry, P.M., Cabeza,
M., Guisan, A., Hickler, T., Midgley, G.F., Paterson, J.,
Schurr, F.M., Sykes, M.T. & Zimmermann, N.E. (2008)
Predicting global change impacts on plant species’ distribu-
1576 Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd
T. Jucker et al.

tions: future challenges. Perspectives in Plant Ecology, Evolu-
tion and Systematics, 9, 137–152.
Thuiller, W., Gallien, L., Boulangeat, I., de Bello, F.,
M€unkem€uller, T., Roquet, C. & Lavergne, S. (2010) Resolv-
ing Darwin’s naturalization conundrum: a quest for evi-
dence. Diversity and Distributions, 16, 461–475.
Vamosi, S.M., Heard, S.B., Vamosi, J.C. & Webb, C.O.
(2009) Emerging patterns in the comparative analysis of
phylogenetic community structure. Molecular Ecology, 18,
572–592.
Vil�a, M., Tessier, M., Suehs, C.M., Brundu, G., Carta, L.,
Galanidis, A., Lambdon, P., Manca, M., Medail, F., Mo-
rogues, E., Traveset, A., Troumbis, A.Y. & Hulme, P.E.
(2006) Local and regional assessments of the impacts of
plant invaders on vegetation structure and soil properties of
Mediterranean islands. Journal of Biogeography, 33, 853–861.
Vil�a, M., Espinar, J.L., Hejda, M., Hulme, P.E., Jarosik, V.,
Maron, J.L., Pergl, J., Schaffner, U., Sun, Y. & Pysek, P.
(2011) Ecological impacts of invasive alien plants: a meta-
analysis of their effects on species, communities and eco-
systems. Ecology Letters, 14, 702–708.
Watkins, A.J. & Wilson, J.W. (2003) Local texture conver-
gence: a new approach to seeking assembly rules. Oikos,
102, 525–532.
Webb, C.O., Ackerly, D.D., McPeek, M.A. & Donoghue, M.J.
(2002) Phylogenies and community ecology. Annual Review
of Ecology, Evolution, and Systematics, 33, 475–505.
Webb, C.O., Ackerly, D.D. & Kembel, S.W. (2008) Phylo-
com: software for the analysis of phylogenetic community
structure and trait evolution. Bioinformatics, 24, 2098–
2100.
Weiner, J. & Thomas, S.C. (1986) Size variability and com-
petition in plant monocultures. Oikos, 47, 211–222.
Winter, M., Schweiger, O., Klotz, S., Nentwig, W., Andriopo-
ulos, P., Arianoutsou, M., Basnou, C., Delipetrou, P., Did-
ziulis, V., Hejda, M., Hulme, P.E., Lambdon, P.W., Pergl,
J., Pysek, P., Roy, D.B. & Kuhn, I. (2009) Plant extinctions
and introductions lead to phylogenetic and taxonomic
homogenization of the European flora. Proceedings of the
National Academy of Sciences of the USA, 106, 21721–21725.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the
online version of this article:
Appendix S1 Comparison of ‘regression residuals’ and ‘null
model’ approaches to removing the correlation between FD
and PD values with TD.
Appendix S2 Test of the ‘random loss scenario’ according to
which rare or low-abundance species are more likely to be
excluded from a community through chance alone.
Appendix S3 Comparison of Mean Distance to Native Spe-
cies (MDNS) and Weighted Mean Distance to Native Species
(WMDNS) metrics used to quantify the mean phylogenetic
distance between iceplant and all other species present in a
community.
Figure S1 Scatterplot of regression residuals (RES) and stan-
dardized effect sizes (SES) for FD and PD.
Figure S2 Relationship between species sensitivity to invasion
and two metrics of species rarity - mean vegetation cover
and frequency of plots in which the species is found.
Figure S3 Scatterplot showing the relationship between
Weighted Mean Distance to Native Species (WMDNS) and
iceplant vegetation cover.
Figure S4 Comparison of the mean number of annual grass
species (Therophytes) found in plots with varying degrees of
iceplant abundance.
BIOSKETCHES
Tommaso Jucker is a plant ecologist interested in the
mechanisms involved in promoting, maintaining and shaping
biodiversity patterns. His research has explored how plant
communities respond to drivers of global change and more
recently has focused on understanding how biodiversity loss
influences ecosystem functioning. He is currently pursuing a
PhD at the University of Cambridge, before which he was a
visiting researcher at the University of Roma Tre, in the
plant ecology laboratory lead by Alicia T.R. Acosta. Her
research team relies on coastal dune environments as model
systems to analyse distribution patterns and dynamics of
plant species/communities in relation to environmental and
human drivers and to agents of global change (particularly
invasive species) across spatial scales. Marta Carboni is a
post-doctoral researcher focusing on plant invasions and on
the mechanisms driving community assembly and species co-
existence.
Author contributions: A.A., M.C. and T.J. conceived the
ideas; M.C. and T.J. collated and analysed the data; all
authors contributed to the writing which was led by T.J.
Editor: Mark van Kleunen
Diversity and Distributions, 19, 1566–1577, ª 2013 John Wiley & Sons Ltd 1577
Invasion and biodiversity loss in plant communities
Copyright © 2022 FDOKUMEN




















