
Genes Related to Mitochondrial Functions, Protein Degradation, and Chromatin Folding Are...
18
Hindawi Publishing Corporation Mediators of Inflammation Volume 2013, Article ID 137629, 18 pages http://dx.doi.org/10.1155/2013/137629 Research Article Genes Related to Mitochondrial Functions, Protein Degradation, and Chromatin Folding Are Differentially Expressed in Lymphomonocytes of Rett Syndrome Patients Alessandra Pecorelli, 1,2 Guido Leoni, 3 Franco Cervellati, 4 Raffaella Canali, 3 Cinzia Signorini, 1 Silvia Leoncini, 1,2 Alessio Cortelazzo, 2,5 Claudio De Felice, 6 Lucia Ciccoli, 1 Joussef Hayek, 2 and Giuseppe Valacchi 4,7 1 Department of Molecular and Developmental Medicine, University of Siena, 53100 Siena, Italy 2 Child Neuropsychiatry Unit, University Hospital, Azienda Ospedaliera Universitaria Senese, 53100 Siena, Italy 3 National Research Institute on Food and Nutrition (INRAN), 00178 Rome, Italy 4 Department of Life Science and Biotechnologies, University of Ferrara, 44121 Ferrara, Italy 5 Department of Medical Biotechnologies, University of Siena, 53100 Siena, Italy 6 Neonatal Intensive Care Unit, University Hospital, Azienda Ospedaliera Universitaria Senese, 53100 Siena, Italy 7 Department of Food and Nutrition, Kyung Hee University, Seoul 130-701, Republic of Korea Correspondence should be addressed to Giuseppe Valacchi; [email protected] Received 11 October 2013; Accepted 7 November 2013 Academic Editor: Paul Ashwood Copyright © 2013 Alessandra Pecorelli et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. Rett syndrome (RTT) is mainly caused by mutations in the X-linked methyl-CpG binding protein (MeCP2) gene. By binding to methylated promoters on CpG islands, MeCP2 protein is able to modulate several genes and important cellular pathways. erefore, mutations in MeCP2 can seriously affect the cellular phenotype. Today, the pathways that MeCP2 mutations are able to affect in RTT are not clear yet. e aim of our study was to investigate the gene expression profiles in peripheral blood lymphomonocytes (PBMC) isolated from RTT patients to try to evidence new genes and new pathways that are involved in RTT pathophysiology. LIMMA (Linear Models for MicroArray) and SAM (Significance Analysis of Microarrays) analyses on microarray data from 12 RTT patients and 7 control subjects identified 482 genes modulated in RTT, of which 430 were upregulated and 52 were downregulated. Functional clustering of a total of 146 genes in RTT identified key biological pathways related to mitochondrial function and organization, cellular ubiquitination and proteosome degradation, RNA processing, and chromatin folding. Our microarray data reveal an overexpression of genes involved in ATP synthesis suggesting altered energy requirement that parallels with increased activities of protein degradation. In conclusion, these findings suggest that mitochondrial-ATP-proteasome functions are likely to be involved in RTT clinical features. 1. Introduction Rett syndrome (RTT) is a rare form of autism spectrum disor- der (ASD), which mostly affects girls with worldwide preva- lence rate ranges from 1 : 10,000 to 1 : 20,000 live births [1–5]. RTT is a clinically defined condition with a large spectrum of phenotypes associated with a wide genotypic variability [6, 7]. Classic or typical RTT, the most common form of the condition, is caused in about 90–95% of cases by de novo mutations in the MeCP2, a gene mapped on chromosome X and encoding methyl-CpG binding protein 2 [7, 8]. e clinical picture of classical form progresses through 4 stages and is characterized by normal development for the first 6 to 18 months, followed by loss of purposeful hand movements, failure of speech development, autistic-like behavior, slowed brain and head growth, and mental retardation [9]. To date, it is not known how MeCP2 mutations lead to RTT phenotypes; therefore the identification of the pathways
Transcript of Genes Related to Mitochondrial Functions, Protein Degradation, and Chromatin Folding Are...

Hindawi Publishing CorporationMediators of InflammationVolume 2013 Article ID 137629 18 pageshttpdxdoiorg1011552013137629
Research ArticleGenes Related to Mitochondrial Functions Protein Degradationand Chromatin Folding Are Differentially Expressed inLymphomonocytes of Rett Syndrome Patients
Alessandra Pecorelli12 Guido Leoni3 Franco Cervellati4 Raffaella Canali3
Cinzia Signorini1 Silvia Leoncini12 Alessio Cortelazzo25 Claudio De Felice6
Lucia Ciccoli1 Joussef Hayek2 and Giuseppe Valacchi47
1 Department of Molecular and Developmental Medicine University of Siena 53100 Siena Italy2 Child Neuropsychiatry Unit University Hospital Azienda Ospedaliera Universitaria Senese 53100 Siena Italy3 National Research Institute on Food and Nutrition (INRAN) 00178 Rome Italy4Department of Life Science and Biotechnologies University of Ferrara 44121 Ferrara Italy5 Department of Medical Biotechnologies University of Siena 53100 Siena Italy6Neonatal Intensive Care Unit University Hospital Azienda Ospedaliera Universitaria Senese 53100 Siena Italy7 Department of Food and Nutrition Kyung Hee University Seoul 130-701 Republic of Korea
Correspondence should be addressed to Giuseppe Valacchi giuseppevalacchiunifeit
Received 11 October 2013 Accepted 7 November 2013
Academic Editor Paul Ashwood
Copyright copy 2013 Alessandra Pecorelli et al This is an open access article distributed under the Creative Commons AttributionLicense which permits unrestricted use distribution and reproduction in any medium provided the original work is properlycited
Rett syndrome (RTT) is mainly caused by mutations in the X-linked methyl-CpG binding protein (MeCP2) gene By binding tomethylated promoters onCpG islandsMeCP2 protein is able tomodulate several genes and important cellular pathwaysThereforemutations in MeCP2 can seriously affect the cellular phenotype Today the pathways that MeCP2 mutations are able to affect inRTT are not clear yet The aim of our study was to investigate the gene expression profiles in peripheral blood lymphomonocytes(PBMC) isolated from RTT patients to try to evidence new genes and new pathways that are involved in RTT pathophysiologyLIMMA (LinearModels forMicroArray) and SAM (SignificanceAnalysis ofMicroarrays) analyses onmicroarray data from 12 RTTpatients and 7 control subjects identified 482 genes modulated in RTT of which 430 were upregulated and 52 were downregulatedFunctional clustering of a total of 146 genes in RTT identified key biological pathways related to mitochondrial function andorganization cellular ubiquitination and proteosome degradation RNA processing and chromatin folding Our microarray datareveal an overexpression of genes involved in ATP synthesis suggesting altered energy requirement that parallels with increasedactivities of protein degradation In conclusion these findings suggest that mitochondrial-ATP-proteasome functions are likely tobe involved in RTT clinical features
1 Introduction
Rett syndrome (RTT) is a rare formof autism spectrumdisor-der (ASD) which mostly affects girls with worldwide preva-lence rate ranges from 1 10000 to 1 20000 live births [1ndash5]RTT is a clinically defined condition with a large spectrumof phenotypes associated with a wide genotypic variability[6 7] Classic or typical RTT the most common form of thecondition is caused in about 90ndash95 of cases by de novo
mutations in the MeCP2 a gene mapped on chromosomeX and encoding methyl-CpG binding protein 2 [7 8] Theclinical picture of classical form progresses through 4 stagesand is characterized by normal development for the first 6 to18 months followed by loss of purposeful hand movementsfailure of speech development autistic-like behavior slowedbrain and head growth and mental retardation [9]
To date it is not known how MeCP2 mutations lead toRTT phenotypes therefore the identification of the pathways
2 Mediators of Inflammation
that are affected byMeCP2 functions could bring new insightin the RTT pathogenetic mechanisms MeCP2 was originallythought to function as a transcription repressor by bindingto methylated CpG dinucleotides but recent studies haveindividuatedmore functions related toMeCP2 [10 11] In factMeCP2 is now considered a multifunctional protein since itis implicated not only in genome transcriptional silencingbut also in transcriptional activation by regulating chromatinand nuclear architecture [11] therefore its malfunction ormutation can lead to severe cellular function alterations
Hence it is very difficult to understand the link betweenMeCP2mutation and the clinical feature present in RTT Oneof the most common approaches used to better understandthe molecular pathways involved in genetic disorders hasbeen the determination of gene expression profiling sinceit provides the opportunity to evaluate possible transcrip-tome alterations at both gene and gene-network levels Thisapproach should not be considered an end point but amagnifying lent where new aspects involved in the diseasescan be discovered and then studied
So far only a handful of studies have investigated thegene expression profiles of RTT children in tissues thatis postmortem brain samples [12] or in cells such asclones of fibroblasts isolated from skin biopsies [13 14] andimmortalized lymphoblastoid cell lines [14ndash16] Moreoverseveral studies have performed microarray gene expressionanalysis using in vitro cellular models representing MeCP2deficiency induced by siRNAs [17] or cells and tissues fromRTT mouse models [18] but to our knowledge there are nodata onmicroarray analysis from ldquoex vivordquo fresh samples Forthis reason the aim of this study was to evaluate the geneexpression patterns in PBMC isolated from RTT patientsThis approach lets us bypass some of the limitationsvariablesof the previous gene arrays studies on RTT such as the use ofpostmortem samples gene-modified cells andmurine tissuesthat do not always reflect all features of the human disease Infact studying ex vivo samples such as PBMC provides someadvantage that can be summarized by the fact PBMC are theonly readily available cells in humans various studies showeddisease-characteristic gene expression patterns in PBMC thatcan be easily obtained
Our results identified a clear difference in gene expressionprofile between control and RTT patients with almost 500genes being deregulated suggesting several new pathwaysinvolved in this disorder
2 Subjects and Methods
21 Subjects Population The study included 12 femalepatients with clinical diagnosis of typical RTT (mean age109 plusmn 49 years range 6ndash22) with demonstrated MeCP2gene mutation and 7 sex- and age-matched healthy controls(mean age 151 plusmn 903 years range 4ndash32) RTT diagnosisand inclusionexclusion criteria were based on the recentlyrevised RTT nomenclature consensus [4] All the patientswere consecutively admitted to the Rett Syndrome NationalReference Centre of the University Hospital of the AziendaOspedaliera Universitaria Senese (AOUS) Table 1 presents
Table 1 Demographic and genetic data for RTT patients enrolled instudy
No Age Stage Mutation type Nucleotidechange
Amino acidchange
1 7 3 ETMs2 10 3 missense c403AgtG pK135E3 9 4 missense c403AgtG pK135E4 9 4 missense c455CgtG pP152R5 12 3 missense c473CgtT pT158M6 19 3 nonsense c763CgtT pR255X
7 22 3frameshiftinsertion ordeletion
c806 807delG pG269fs
8 7 4 nonsense c808CgtT pR270X9 7 3 nonsense c808CgtT pR270X10 12 4 nonsense c880CgtT pR294X11 6 3 nonsense c880CgtT pR294X12 11 3 missense c916CgtT pR306CETMs early truncating mutations
the demographic and genetic characteristics of the enrolledpatients subjected to microarray analysis Blood samplingin the control group was carried out during routine healthchecks sports or blood donations while blood sample inpatients were obtained during the periodic clinical checksThe study was conducted with the approval of the Institu-tional Review Board and all informed consents were obtainedfrom either the parents or the legal tutors of the enrolledpatients
22 Blood Specimen Collection Peripheral Blood Lym-phomonocytes Isolation and RNA Extraction Blood wascollected in heparinized tubes and all manipulations werecarried out within 30 minutes after sample collection PBMCwere separated from whole blood by density gradient cen-trifugation using Ficoll-Paque PLUS (GE Healthcare EuropeGmbH Milan Italy) After PBMC isolation total RNA wasextracted from cells using RNeasy mini kit (Qiagen HildenGermany) according to the manufacturerrsquos instructions Thetotal nucleic acid concentration and purity were estimatedusing aNanoDrop spectrophotometer (NanoDropTechnolo-gies Wilmington DE) Quality of RNA was checked onAgilent bioanalyzer (Agilent Technologies Santa Clara CA)The isolated RNA samples were stored at minus80∘C until theanalysis
23 Microarray Processing For the microarray processingRNA was amplified and labeled using the AffymetrixWhole-Transcript (WT) Sense Target Labeling Protocol AffymetrixGeneChip Human Gene 10 ST arrays were hybridized withlabeled sense DNA washed stained and scanned accordingto the protocol described inWT Sense Target Labeling AssayManual
Briefly 100 ng of total RNA was reverse transcribed intodouble-stranded cDNA with random hexamers tagged with
Mediators of Inflammation 3
a T7 promoter sequence The double-stranded cDNA wassubsequently used as a template and amplified by T7 RNApolymerase producing many copies of antisense cRNA Inthe second cycle of cDNA synthesis random hexamers wereused to prime reverse transcription of the cRNA from the firstcycle to produce single-stranded DNA in the sense orienta-tion dUTP was incorporated in the DNA during the second-cycle first-strand reverse transcription reaction This single-stranded DNA sample was then treated with a combinationof uracil DNA glycosylase (UDG) and apurinicapyrimidinicendonuclease 1 (APE 1) that specifically recognized theunnatural dUTP residues and broke the DNA strand DNAwas labeled by terminal deoxynucleotidyl transferase (TdT)with the Affymetrix proprietary DNA Labeling Reagent thatis covalently linked to biotin 5 120583g of labeled cDNA washybridized to the Human Gene 10 ST Array at 45∘C for 17hoursThe arrays were washed and stained in Affymetrix Flu-idics Station 450 and scanned using theAffymetrixGeneChipScanner 3000
24 Microarray Data Analysis The microarray experimentswere analyzed with the oneChannelGUI package availablein R software The signal intensities from each chip werepreliminarily normalized with RMA method and filteredby IQR filter choosing as threshold a value of 025 Thisspecific filter removes the probesets that do not presentchanges in their expression across the analyzed microarrayThe threshold of 025 is an intermediate value that retainsthe probesets that show significative changes of their signal atleast in the 25 of analyzed microarray The microarray datawere analyzed by SAM and LIMMA analyses SAM approachuses permutation based statistics and is a valid method toanalyze data that may not follow a normal distributionoutperforming other techniques (eg ANOVA and classical119905-test) which assume equal variance andor independenceof genes LIMMA fit each gene expression to a linearregression model testing the significativity of the distancefrom the model with a 119905-test robust against nonnormalityand inequality of variances In order to characterize thebiological processes enriched in the list of modulated genesa statistical analysis of overrepresented Gene Ontology (GO)terms (BP level 5) was performed with DAVID (Databasefor Annotation Visualization and IntegratedDiscovery) webserver (DAVID Bioinformatics Resources) [19] Only the GOterms enriched with a 119875 value lt 005 corrected with theBenjamini and Hochberg method [20] were selected andhierarchically clusterized (Wardrsquos method) according to theirldquosemanticrdquo similarity estimated with the GOSIM R packagewith default parameters After clusterization the best numberof clusters was identified according to the analysis of thesilhouette scores and the medoid term in each cluster wasselected as the representative member of the cluster Eachcluster represents a group of biological processes with similareo related functions
25 Validation ofMicroarrayData by RT-qPCR (Reverse Tran-scription Quantitative Real-Time PCR) For confirmation ofAffymetrix expression microarray results RT-qPCR analysis
was performed as previously described [21] For validationsix of the differentially expressed genes 3 upregulatedmdashGSTO1 PSMB10 and COX8Amdashand 3 downregulatedmdashHIST1H1B MMP9 and ARHGAP11Bmdashby microarray werechosen Validation was done in a randomly selected subset ofthe original samples (submitted for microarray analysis) thatincluded 3 healthy controls and 3 RTT patients Primer pairswere obtained from the Real-time PCR GenBank Primer tohybridise unique regions of the appropriate gene sequenceGSTO1 (Fw 51015840-AGA GTT GTT TTC TAA GGT TCT GACT-31015840) and (RW 51015840-ACT TCA TTG CTT CCA GCC GT-31015840) product length 116 bp PSMB10 (Fw 51015840-ACA GAC GTGAAG CCT AGC AG-31015840) and (Rw 51015840-ACC GAA TCG TTAGTG GCT CG-31015840) product length 294 bp COX8A (Fw 51015840-GCC AAG ATC CAT TCG TTG CC-31015840) and (RW 51015840-TCTGGC CTC CTG TAG GTC TC-31015840) product length 137 bpHIST1H1B (Fw 51015840-CCC GGC TAA GAA GAA GGC AA-31015840) and (RW 51015840-ACA GCC TTG GTG ATC AGC TC-31015840)product length 99 bp MMP9 (Fw 51015840-GTC CGT GAG GGTGTT GAG TG-31015840) and (RW 51015840-ACT GCT CAA AGC CTCCAC AA-31015840) product length 145 bp ARHGAP11B (Fw 51015840-AAC TGC CAG AGC CCA TTC TC-31015840) and (RW 51015840-GTCTGGTACACGCCCTTCTT-31015840) product length 295 bpAllreactions were run in triplicate GAPDH (FW 51015840-TGA CGCTGG GGC TGG CAT TG-31015840 and RW 51015840-GGC TGG TGGTCC AGG GGT CT-31015840 134 pb) was used in our experimentsas internal standard As previously described samples werecompared using the relative cycle threshold (CT) method(Livak and Schmittgen 2001) After normalization to morestable mRNA GAPDH the fold increase or decrease wasdeterminedwith respect to control using the formula 2minusΔΔCTwhere ΔCT is (gene of interest CT)minus(reference gene CT)andΔΔCT is (ΔCT experimental)minus(ΔCT control) Results arethe means plusmn SEM of three independent experiments eachanalysed in triplicate lowast119875 lt 0001 versus control (one-wayANOVA followed by Bonferronirsquos posttest)
3 Results and Discussion
31 Differentially Regulated Genes in RTT Patients Giventhat RTT results from dysfunction of the transcriptionalmodulator MeCP2 several strategies have been developedto identify its target genes in order to gain insights into thedisease pathogenesis [12ndash18] In our study to identify geneexpression changes associated with and potentially related toMeCP2 mutations and to delineate alterations of pathwaysassociated with the disease we evaluated and comparedtranscriptomic profiles in PBMC from RTT patients andcontrol subjects
This work showed for the first time to our knowledgealtered expression of a large set of genes that may helpelucidating and explaining the link between MeCP2 andsome of the molecular and cellular aspects observed in RTTpatients In our previous studies on RTT we were able toshow increased levels of oxidative stress (OS)markers such asisoprostanes (IsoPs) and 4-hydroxynonenal protein adducts(4-HNE PAs) [22 23] and increased ubiquitination anddegradation of oxidatively modified proteins [24] but how
4 Mediators of Inflammation
Up
Down
LIMMA SAM
5
5
1 430
52
Figure 1 Comparison of two distinct approaches for screening ofdifferentially expressed genes in RTT PBMC Venn diagram repre-senting numbers of common and exclusively up- and downregulatedgenes for LIMMA (left) and SAM (right) analyses (FC plusmn 1 adj119875 value le 005) The 11 genes indicated by only one method wereexcluded by further analysis
MeCP2 mutation is able to affect cellular redox balance andproteins turn over still needs to be defined
Using the SAM and LIMMA methods we have defineda set of genes differentially expressed in RTT patients withrespect to controls (healthy subjects) and a significant overlapwas found by comparing results from two approaches Acut-off level based on a minimum of 1-fold change inexpression resulted in a list of 482 common deregulatedgenes while 10 genes were suggested only by SAM andonly 1 gene was indicated by LIMMA Among the sharedgenes 430 showed significant upregulation while 52 weredownregulated in RTT compared to controls (Figure 1 andSupplemental Tables 1 and 2) The 11 genes indicated by onlyone method were excluded for further analysis Genes withthe strongest changes in expression both upregulated (FC ge2) and downregulated (FC le minus12) are listed in Tables 2 and3 respectively The complete list of differentially expressedgenes both upregulated and downregulated (FC plusmn 1) isshown in Supplemental Tables 1 and 2
It is evident from this first set of data that mutations inMeCP2 influence more the genes upregulation with respectto the downregulation This is in part in line with the firstfunctions that have been attributed to MeCP2 such as a geneexpression repressor [10 11] In addition it is worth it tounderline that both screening approaches used in this study(LIMMA and SAM) almost overlap between them makingthe results more reliable
Next using the DAVID databases we performed thefunctional annotation of the significant genes and iden-tification of the biochemical pathways in which they areinvolved A comparison of the differentially regulated mRNAtranscripts in RTT PBMC compared to control group showssignificant changes in cellular pathways In particular weidentify 10 major clusters corresponding to 62 biologicalprocesses enriched by 146 genes (Table 4) These clustershighlight key biological pathways related to mitochondrialfunction and organization (ie mitochondrial ATP synthesis
coupled to electron transport inner mitochondrial mem-brane organization such asATP5A1COX6CETFAUQCRQTIMM10 and TSPO) cellular protein metabolic process (ieregulation of protein ubiquitination regulation of ubiquitin-protein ligase activity and proteasomal ubiquitin-dependentprotein catabolic process such as PSMA2 PSMD6 UBE2E3and UFC) RNA processing (ie nuclear mRNA splicingand spliceosome assembly and RNA elongation from RNApolymerase II promoter such asRPL15) DNAorganization inchromatin and cellular macromolecular complex assembly(ie nucleosome assembly DNA packaging and proteincomplex biogenesis such asHIST1H4L H2AFZ TOP2A andHMGB2)
32 Mitochondrial Related Genes in RTT Patients Amongthese clusters the most significantly regulated transcriptsinclude those encoding several subunits of mitochondrialrespiratory chain complexes and thus linked directly tomitochondrial ATP production and indirectly to potentialreactive oxygen species (ROS) generation In particularNDUFA1 NDUFAB1 NDUFA2 and NDUFB6 all compo-nents of mitochondrial complex I (NADH ubiquinone oxi-doreductase) showed the greater changes with a FC of morethan 2 Moreover other subunits of complex I (NDUFV2NDUFS4 NDUFA9 NDUFS6 NDUFB10 NDUFB4NDUFC2 NDUFB2 NDUFS5 NDUFC1 NDUFB9 andNDUFA8) were clearly upregulated in RTT group
Complex I plays a vital role in cellular ATP productionthe primary source of energy for many crucial processes inliving cells It removes electrons from NADH and passesthem by a series of different protein coupled redox centersto the electron acceptor ubiquinone Because complex I iscentral to energy production in the cell it is reported thatits malfunction results in a wide range of neuromusculardiseases [25] Some of them are due to mutations in themitochondrial genome but others which result from adecrease in the activity of complex I or an increase in theproduction of ROS are not well understood The productionof ROS by complex I is linked to Parkinsonrsquos disease and toageing [26 27] and this is in line with RTT since it is nowwelldocumented as an increased OS condition in this pathology[22 23]
Another gene involved in complex I function isNDUFV2that was also clearly upregulated in our study Mutationsin this gene are implicated in Parkinsonrsquos disease bipo-lar disorder and schizophrenia and have been found inone case of early onset hypertrophic cardiomyopathy andencephalopathy also it has been shown for NDUFA2 asubunit of the hydrophobic protein fraction of the complex IMutations in this gene are associated with Leigh syndromean early onset progressive neurodegenerative disorder Ofnote is NDUFAB1 which is a carrier of the growing fattyacid chain in fatty acid biosynthesis in mitochondria andalteration in fatty acid levels has been noted in ASD [28 29]
Not only complex I subunits were upregulated in RTTbut also we have detected an upregulation of genes involvedin all the five complexes of the electron transport chain Infact also SDHB (succinate dehydrogenase complex subunit
Mediators of Inflammation 5
Table 2 Genes upregulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0045413 NDUFA1NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 1 75 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone 31
NM 0064982 LGALS2Lectingalactoside-bindingsoluble 2
Carbohydrate binding mdash 28
NM 0143023 SEC61G Sec61 gamma subunit Protein transporteractivity
Protein targeting to ER antigenprocessing and presentation ofexogenous peptide antigen viaMHC class I
28
NM 0010404371 C6orf48 Chromosome 6 openreading frame 48 mdash mdash 27
NM 0010985772 RPL31 Ribosomal protein L31 RNA binding structuralconstituent of ribosome
Translational elongationtranslational initiationtranslational termination
27
NM 0069895 RASA4 RAS p21 proteinactivator 4
GTPase activator activityphospholipid binding
Intracellular signal transductionpositive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction
26
NM 0009833 RPL22 Ribosomal protein L22RNA binding heparinbinding structuralconstituent of ribosome
Alpha-beta T cell differentiationtranslational elongationtranslational initiationtranslational termination
26
NM 0190593 TOMM7
Translocase of outermitochondrialmembrane 7 homolog(yeast)
Protein transmembranetransporter activity
Cellular protein metabolic processprotein import into mitochondrialmatrix protein targeting tomitochondrion
26
NM 0311572 HNRNPA1 Heterogeneous nuclearribonucleoprotein A1
Nucleotide bindingsingle-stranded DNAbinding single-strandedRNA binding
RNA export from nucleus mRNAsplicing via spliceosome mRNAtransport nuclear import
25
NM 0018285 CLC Charcot-Leyden crystalgalectin
Carbohydrate bindingcarboxylesterase activitylysophospholipase activity
Lipid catabolic processmulticellular organismaldevelopment
24
NM 0329013 COX14Cytochrome c oxidaseassembly homolog 14(S cerevisiae)
Plays a role in theassembly or stability of thecytochrome c oxidasecomplex (COX)
Mitochondrial respiratory chaincomplex IV assembly 23
NM 0249604 PANK2 Pantothenate kinase 2ATP bindingpantothenate kinaseactivity
Cell death coenzyme Abiosynthetic process coenzymebiosynthetic process pantothenatemetabolic process
23
NR 0023091 RPS26P11 Ribosomal protein S26pseudogene 11
Structural constituent ofribosome Translation 23
NM 0050032 NDUFAB1
NADH dehydrogenase(ubiquinone) 1alphabeta subcomplex1 8 kDa
NADH dehydrogenase(ubiquinone) activityACP phosphopantetheineattachment site bindinginvolved in fatty acidbiosynthetic process fattyacid binding calcium ionbinding
Cellular metabolic process proteinlipoylation small moleculemetabolic process respiratoryelectron transport chain fatty acidbiosynthetic processmitochondrial electron transportNADH to ubiquinone
23
6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

2 Mediators of Inflammation
that are affected byMeCP2 functions could bring new insightin the RTT pathogenetic mechanisms MeCP2 was originallythought to function as a transcription repressor by bindingto methylated CpG dinucleotides but recent studies haveindividuatedmore functions related toMeCP2 [10 11] In factMeCP2 is now considered a multifunctional protein since itis implicated not only in genome transcriptional silencingbut also in transcriptional activation by regulating chromatinand nuclear architecture [11] therefore its malfunction ormutation can lead to severe cellular function alterations
Hence it is very difficult to understand the link betweenMeCP2mutation and the clinical feature present in RTT Oneof the most common approaches used to better understandthe molecular pathways involved in genetic disorders hasbeen the determination of gene expression profiling sinceit provides the opportunity to evaluate possible transcrip-tome alterations at both gene and gene-network levels Thisapproach should not be considered an end point but amagnifying lent where new aspects involved in the diseasescan be discovered and then studied
So far only a handful of studies have investigated thegene expression profiles of RTT children in tissues thatis postmortem brain samples [12] or in cells such asclones of fibroblasts isolated from skin biopsies [13 14] andimmortalized lymphoblastoid cell lines [14ndash16] Moreoverseveral studies have performed microarray gene expressionanalysis using in vitro cellular models representing MeCP2deficiency induced by siRNAs [17] or cells and tissues fromRTT mouse models [18] but to our knowledge there are nodata onmicroarray analysis from ldquoex vivordquo fresh samples Forthis reason the aim of this study was to evaluate the geneexpression patterns in PBMC isolated from RTT patientsThis approach lets us bypass some of the limitationsvariablesof the previous gene arrays studies on RTT such as the use ofpostmortem samples gene-modified cells andmurine tissuesthat do not always reflect all features of the human disease Infact studying ex vivo samples such as PBMC provides someadvantage that can be summarized by the fact PBMC are theonly readily available cells in humans various studies showeddisease-characteristic gene expression patterns in PBMC thatcan be easily obtained
Our results identified a clear difference in gene expressionprofile between control and RTT patients with almost 500genes being deregulated suggesting several new pathwaysinvolved in this disorder
2 Subjects and Methods
21 Subjects Population The study included 12 femalepatients with clinical diagnosis of typical RTT (mean age109 plusmn 49 years range 6ndash22) with demonstrated MeCP2gene mutation and 7 sex- and age-matched healthy controls(mean age 151 plusmn 903 years range 4ndash32) RTT diagnosisand inclusionexclusion criteria were based on the recentlyrevised RTT nomenclature consensus [4] All the patientswere consecutively admitted to the Rett Syndrome NationalReference Centre of the University Hospital of the AziendaOspedaliera Universitaria Senese (AOUS) Table 1 presents
Table 1 Demographic and genetic data for RTT patients enrolled instudy
No Age Stage Mutation type Nucleotidechange
Amino acidchange
1 7 3 ETMs2 10 3 missense c403AgtG pK135E3 9 4 missense c403AgtG pK135E4 9 4 missense c455CgtG pP152R5 12 3 missense c473CgtT pT158M6 19 3 nonsense c763CgtT pR255X
7 22 3frameshiftinsertion ordeletion
c806 807delG pG269fs
8 7 4 nonsense c808CgtT pR270X9 7 3 nonsense c808CgtT pR270X10 12 4 nonsense c880CgtT pR294X11 6 3 nonsense c880CgtT pR294X12 11 3 missense c916CgtT pR306CETMs early truncating mutations
the demographic and genetic characteristics of the enrolledpatients subjected to microarray analysis Blood samplingin the control group was carried out during routine healthchecks sports or blood donations while blood sample inpatients were obtained during the periodic clinical checksThe study was conducted with the approval of the Institu-tional Review Board and all informed consents were obtainedfrom either the parents or the legal tutors of the enrolledpatients
22 Blood Specimen Collection Peripheral Blood Lym-phomonocytes Isolation and RNA Extraction Blood wascollected in heparinized tubes and all manipulations werecarried out within 30 minutes after sample collection PBMCwere separated from whole blood by density gradient cen-trifugation using Ficoll-Paque PLUS (GE Healthcare EuropeGmbH Milan Italy) After PBMC isolation total RNA wasextracted from cells using RNeasy mini kit (Qiagen HildenGermany) according to the manufacturerrsquos instructions Thetotal nucleic acid concentration and purity were estimatedusing aNanoDrop spectrophotometer (NanoDropTechnolo-gies Wilmington DE) Quality of RNA was checked onAgilent bioanalyzer (Agilent Technologies Santa Clara CA)The isolated RNA samples were stored at minus80∘C until theanalysis
23 Microarray Processing For the microarray processingRNA was amplified and labeled using the AffymetrixWhole-Transcript (WT) Sense Target Labeling Protocol AffymetrixGeneChip Human Gene 10 ST arrays were hybridized withlabeled sense DNA washed stained and scanned accordingto the protocol described inWT Sense Target Labeling AssayManual
Briefly 100 ng of total RNA was reverse transcribed intodouble-stranded cDNA with random hexamers tagged with
Mediators of Inflammation 3
a T7 promoter sequence The double-stranded cDNA wassubsequently used as a template and amplified by T7 RNApolymerase producing many copies of antisense cRNA Inthe second cycle of cDNA synthesis random hexamers wereused to prime reverse transcription of the cRNA from the firstcycle to produce single-stranded DNA in the sense orienta-tion dUTP was incorporated in the DNA during the second-cycle first-strand reverse transcription reaction This single-stranded DNA sample was then treated with a combinationof uracil DNA glycosylase (UDG) and apurinicapyrimidinicendonuclease 1 (APE 1) that specifically recognized theunnatural dUTP residues and broke the DNA strand DNAwas labeled by terminal deoxynucleotidyl transferase (TdT)with the Affymetrix proprietary DNA Labeling Reagent thatis covalently linked to biotin 5 120583g of labeled cDNA washybridized to the Human Gene 10 ST Array at 45∘C for 17hoursThe arrays were washed and stained in Affymetrix Flu-idics Station 450 and scanned using theAffymetrixGeneChipScanner 3000
24 Microarray Data Analysis The microarray experimentswere analyzed with the oneChannelGUI package availablein R software The signal intensities from each chip werepreliminarily normalized with RMA method and filteredby IQR filter choosing as threshold a value of 025 Thisspecific filter removes the probesets that do not presentchanges in their expression across the analyzed microarrayThe threshold of 025 is an intermediate value that retainsthe probesets that show significative changes of their signal atleast in the 25 of analyzed microarray The microarray datawere analyzed by SAM and LIMMA analyses SAM approachuses permutation based statistics and is a valid method toanalyze data that may not follow a normal distributionoutperforming other techniques (eg ANOVA and classical119905-test) which assume equal variance andor independenceof genes LIMMA fit each gene expression to a linearregression model testing the significativity of the distancefrom the model with a 119905-test robust against nonnormalityand inequality of variances In order to characterize thebiological processes enriched in the list of modulated genesa statistical analysis of overrepresented Gene Ontology (GO)terms (BP level 5) was performed with DAVID (Databasefor Annotation Visualization and IntegratedDiscovery) webserver (DAVID Bioinformatics Resources) [19] Only the GOterms enriched with a 119875 value lt 005 corrected with theBenjamini and Hochberg method [20] were selected andhierarchically clusterized (Wardrsquos method) according to theirldquosemanticrdquo similarity estimated with the GOSIM R packagewith default parameters After clusterization the best numberof clusters was identified according to the analysis of thesilhouette scores and the medoid term in each cluster wasselected as the representative member of the cluster Eachcluster represents a group of biological processes with similareo related functions
25 Validation ofMicroarrayData by RT-qPCR (Reverse Tran-scription Quantitative Real-Time PCR) For confirmation ofAffymetrix expression microarray results RT-qPCR analysis
was performed as previously described [21] For validationsix of the differentially expressed genes 3 upregulatedmdashGSTO1 PSMB10 and COX8Amdashand 3 downregulatedmdashHIST1H1B MMP9 and ARHGAP11Bmdashby microarray werechosen Validation was done in a randomly selected subset ofthe original samples (submitted for microarray analysis) thatincluded 3 healthy controls and 3 RTT patients Primer pairswere obtained from the Real-time PCR GenBank Primer tohybridise unique regions of the appropriate gene sequenceGSTO1 (Fw 51015840-AGA GTT GTT TTC TAA GGT TCT GACT-31015840) and (RW 51015840-ACT TCA TTG CTT CCA GCC GT-31015840) product length 116 bp PSMB10 (Fw 51015840-ACA GAC GTGAAG CCT AGC AG-31015840) and (Rw 51015840-ACC GAA TCG TTAGTG GCT CG-31015840) product length 294 bp COX8A (Fw 51015840-GCC AAG ATC CAT TCG TTG CC-31015840) and (RW 51015840-TCTGGC CTC CTG TAG GTC TC-31015840) product length 137 bpHIST1H1B (Fw 51015840-CCC GGC TAA GAA GAA GGC AA-31015840) and (RW 51015840-ACA GCC TTG GTG ATC AGC TC-31015840)product length 99 bp MMP9 (Fw 51015840-GTC CGT GAG GGTGTT GAG TG-31015840) and (RW 51015840-ACT GCT CAA AGC CTCCAC AA-31015840) product length 145 bp ARHGAP11B (Fw 51015840-AAC TGC CAG AGC CCA TTC TC-31015840) and (RW 51015840-GTCTGGTACACGCCCTTCTT-31015840) product length 295 bpAllreactions were run in triplicate GAPDH (FW 51015840-TGA CGCTGG GGC TGG CAT TG-31015840 and RW 51015840-GGC TGG TGGTCC AGG GGT CT-31015840 134 pb) was used in our experimentsas internal standard As previously described samples werecompared using the relative cycle threshold (CT) method(Livak and Schmittgen 2001) After normalization to morestable mRNA GAPDH the fold increase or decrease wasdeterminedwith respect to control using the formula 2minusΔΔCTwhere ΔCT is (gene of interest CT)minus(reference gene CT)andΔΔCT is (ΔCT experimental)minus(ΔCT control) Results arethe means plusmn SEM of three independent experiments eachanalysed in triplicate lowast119875 lt 0001 versus control (one-wayANOVA followed by Bonferronirsquos posttest)
3 Results and Discussion
31 Differentially Regulated Genes in RTT Patients Giventhat RTT results from dysfunction of the transcriptionalmodulator MeCP2 several strategies have been developedto identify its target genes in order to gain insights into thedisease pathogenesis [12ndash18] In our study to identify geneexpression changes associated with and potentially related toMeCP2 mutations and to delineate alterations of pathwaysassociated with the disease we evaluated and comparedtranscriptomic profiles in PBMC from RTT patients andcontrol subjects
This work showed for the first time to our knowledgealtered expression of a large set of genes that may helpelucidating and explaining the link between MeCP2 andsome of the molecular and cellular aspects observed in RTTpatients In our previous studies on RTT we were able toshow increased levels of oxidative stress (OS)markers such asisoprostanes (IsoPs) and 4-hydroxynonenal protein adducts(4-HNE PAs) [22 23] and increased ubiquitination anddegradation of oxidatively modified proteins [24] but how
4 Mediators of Inflammation
Up
Down
LIMMA SAM
5
5
1 430
52
Figure 1 Comparison of two distinct approaches for screening ofdifferentially expressed genes in RTT PBMC Venn diagram repre-senting numbers of common and exclusively up- and downregulatedgenes for LIMMA (left) and SAM (right) analyses (FC plusmn 1 adj119875 value le 005) The 11 genes indicated by only one method wereexcluded by further analysis
MeCP2 mutation is able to affect cellular redox balance andproteins turn over still needs to be defined
Using the SAM and LIMMA methods we have defineda set of genes differentially expressed in RTT patients withrespect to controls (healthy subjects) and a significant overlapwas found by comparing results from two approaches Acut-off level based on a minimum of 1-fold change inexpression resulted in a list of 482 common deregulatedgenes while 10 genes were suggested only by SAM andonly 1 gene was indicated by LIMMA Among the sharedgenes 430 showed significant upregulation while 52 weredownregulated in RTT compared to controls (Figure 1 andSupplemental Tables 1 and 2) The 11 genes indicated by onlyone method were excluded for further analysis Genes withthe strongest changes in expression both upregulated (FC ge2) and downregulated (FC le minus12) are listed in Tables 2 and3 respectively The complete list of differentially expressedgenes both upregulated and downregulated (FC plusmn 1) isshown in Supplemental Tables 1 and 2
It is evident from this first set of data that mutations inMeCP2 influence more the genes upregulation with respectto the downregulation This is in part in line with the firstfunctions that have been attributed to MeCP2 such as a geneexpression repressor [10 11] In addition it is worth it tounderline that both screening approaches used in this study(LIMMA and SAM) almost overlap between them makingthe results more reliable
Next using the DAVID databases we performed thefunctional annotation of the significant genes and iden-tification of the biochemical pathways in which they areinvolved A comparison of the differentially regulated mRNAtranscripts in RTT PBMC compared to control group showssignificant changes in cellular pathways In particular weidentify 10 major clusters corresponding to 62 biologicalprocesses enriched by 146 genes (Table 4) These clustershighlight key biological pathways related to mitochondrialfunction and organization (ie mitochondrial ATP synthesis
coupled to electron transport inner mitochondrial mem-brane organization such asATP5A1COX6CETFAUQCRQTIMM10 and TSPO) cellular protein metabolic process (ieregulation of protein ubiquitination regulation of ubiquitin-protein ligase activity and proteasomal ubiquitin-dependentprotein catabolic process such as PSMA2 PSMD6 UBE2E3and UFC) RNA processing (ie nuclear mRNA splicingand spliceosome assembly and RNA elongation from RNApolymerase II promoter such asRPL15) DNAorganization inchromatin and cellular macromolecular complex assembly(ie nucleosome assembly DNA packaging and proteincomplex biogenesis such asHIST1H4L H2AFZ TOP2A andHMGB2)
32 Mitochondrial Related Genes in RTT Patients Amongthese clusters the most significantly regulated transcriptsinclude those encoding several subunits of mitochondrialrespiratory chain complexes and thus linked directly tomitochondrial ATP production and indirectly to potentialreactive oxygen species (ROS) generation In particularNDUFA1 NDUFAB1 NDUFA2 and NDUFB6 all compo-nents of mitochondrial complex I (NADH ubiquinone oxi-doreductase) showed the greater changes with a FC of morethan 2 Moreover other subunits of complex I (NDUFV2NDUFS4 NDUFA9 NDUFS6 NDUFB10 NDUFB4NDUFC2 NDUFB2 NDUFS5 NDUFC1 NDUFB9 andNDUFA8) were clearly upregulated in RTT group
Complex I plays a vital role in cellular ATP productionthe primary source of energy for many crucial processes inliving cells It removes electrons from NADH and passesthem by a series of different protein coupled redox centersto the electron acceptor ubiquinone Because complex I iscentral to energy production in the cell it is reported thatits malfunction results in a wide range of neuromusculardiseases [25] Some of them are due to mutations in themitochondrial genome but others which result from adecrease in the activity of complex I or an increase in theproduction of ROS are not well understood The productionof ROS by complex I is linked to Parkinsonrsquos disease and toageing [26 27] and this is in line with RTT since it is nowwelldocumented as an increased OS condition in this pathology[22 23]
Another gene involved in complex I function isNDUFV2that was also clearly upregulated in our study Mutationsin this gene are implicated in Parkinsonrsquos disease bipo-lar disorder and schizophrenia and have been found inone case of early onset hypertrophic cardiomyopathy andencephalopathy also it has been shown for NDUFA2 asubunit of the hydrophobic protein fraction of the complex IMutations in this gene are associated with Leigh syndromean early onset progressive neurodegenerative disorder Ofnote is NDUFAB1 which is a carrier of the growing fattyacid chain in fatty acid biosynthesis in mitochondria andalteration in fatty acid levels has been noted in ASD [28 29]
Not only complex I subunits were upregulated in RTTbut also we have detected an upregulation of genes involvedin all the five complexes of the electron transport chain Infact also SDHB (succinate dehydrogenase complex subunit
Mediators of Inflammation 5
Table 2 Genes upregulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0045413 NDUFA1NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 1 75 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone 31
NM 0064982 LGALS2Lectingalactoside-bindingsoluble 2
Carbohydrate binding mdash 28
NM 0143023 SEC61G Sec61 gamma subunit Protein transporteractivity
Protein targeting to ER antigenprocessing and presentation ofexogenous peptide antigen viaMHC class I
28
NM 0010404371 C6orf48 Chromosome 6 openreading frame 48 mdash mdash 27
NM 0010985772 RPL31 Ribosomal protein L31 RNA binding structuralconstituent of ribosome
Translational elongationtranslational initiationtranslational termination
27
NM 0069895 RASA4 RAS p21 proteinactivator 4
GTPase activator activityphospholipid binding
Intracellular signal transductionpositive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction
26
NM 0009833 RPL22 Ribosomal protein L22RNA binding heparinbinding structuralconstituent of ribosome
Alpha-beta T cell differentiationtranslational elongationtranslational initiationtranslational termination
26
NM 0190593 TOMM7
Translocase of outermitochondrialmembrane 7 homolog(yeast)
Protein transmembranetransporter activity
Cellular protein metabolic processprotein import into mitochondrialmatrix protein targeting tomitochondrion
26
NM 0311572 HNRNPA1 Heterogeneous nuclearribonucleoprotein A1
Nucleotide bindingsingle-stranded DNAbinding single-strandedRNA binding
RNA export from nucleus mRNAsplicing via spliceosome mRNAtransport nuclear import
25
NM 0018285 CLC Charcot-Leyden crystalgalectin
Carbohydrate bindingcarboxylesterase activitylysophospholipase activity
Lipid catabolic processmulticellular organismaldevelopment
24
NM 0329013 COX14Cytochrome c oxidaseassembly homolog 14(S cerevisiae)
Plays a role in theassembly or stability of thecytochrome c oxidasecomplex (COX)
Mitochondrial respiratory chaincomplex IV assembly 23
NM 0249604 PANK2 Pantothenate kinase 2ATP bindingpantothenate kinaseactivity
Cell death coenzyme Abiosynthetic process coenzymebiosynthetic process pantothenatemetabolic process
23
NR 0023091 RPS26P11 Ribosomal protein S26pseudogene 11
Structural constituent ofribosome Translation 23
NM 0050032 NDUFAB1
NADH dehydrogenase(ubiquinone) 1alphabeta subcomplex1 8 kDa
NADH dehydrogenase(ubiquinone) activityACP phosphopantetheineattachment site bindinginvolved in fatty acidbiosynthetic process fattyacid binding calcium ionbinding
Cellular metabolic process proteinlipoylation small moleculemetabolic process respiratoryelectron transport chain fatty acidbiosynthetic processmitochondrial electron transportNADH to ubiquinone
23
6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 3
a T7 promoter sequence The double-stranded cDNA wassubsequently used as a template and amplified by T7 RNApolymerase producing many copies of antisense cRNA Inthe second cycle of cDNA synthesis random hexamers wereused to prime reverse transcription of the cRNA from the firstcycle to produce single-stranded DNA in the sense orienta-tion dUTP was incorporated in the DNA during the second-cycle first-strand reverse transcription reaction This single-stranded DNA sample was then treated with a combinationof uracil DNA glycosylase (UDG) and apurinicapyrimidinicendonuclease 1 (APE 1) that specifically recognized theunnatural dUTP residues and broke the DNA strand DNAwas labeled by terminal deoxynucleotidyl transferase (TdT)with the Affymetrix proprietary DNA Labeling Reagent thatis covalently linked to biotin 5 120583g of labeled cDNA washybridized to the Human Gene 10 ST Array at 45∘C for 17hoursThe arrays were washed and stained in Affymetrix Flu-idics Station 450 and scanned using theAffymetrixGeneChipScanner 3000
24 Microarray Data Analysis The microarray experimentswere analyzed with the oneChannelGUI package availablein R software The signal intensities from each chip werepreliminarily normalized with RMA method and filteredby IQR filter choosing as threshold a value of 025 Thisspecific filter removes the probesets that do not presentchanges in their expression across the analyzed microarrayThe threshold of 025 is an intermediate value that retainsthe probesets that show significative changes of their signal atleast in the 25 of analyzed microarray The microarray datawere analyzed by SAM and LIMMA analyses SAM approachuses permutation based statistics and is a valid method toanalyze data that may not follow a normal distributionoutperforming other techniques (eg ANOVA and classical119905-test) which assume equal variance andor independenceof genes LIMMA fit each gene expression to a linearregression model testing the significativity of the distancefrom the model with a 119905-test robust against nonnormalityand inequality of variances In order to characterize thebiological processes enriched in the list of modulated genesa statistical analysis of overrepresented Gene Ontology (GO)terms (BP level 5) was performed with DAVID (Databasefor Annotation Visualization and IntegratedDiscovery) webserver (DAVID Bioinformatics Resources) [19] Only the GOterms enriched with a 119875 value lt 005 corrected with theBenjamini and Hochberg method [20] were selected andhierarchically clusterized (Wardrsquos method) according to theirldquosemanticrdquo similarity estimated with the GOSIM R packagewith default parameters After clusterization the best numberof clusters was identified according to the analysis of thesilhouette scores and the medoid term in each cluster wasselected as the representative member of the cluster Eachcluster represents a group of biological processes with similareo related functions
25 Validation ofMicroarrayData by RT-qPCR (Reverse Tran-scription Quantitative Real-Time PCR) For confirmation ofAffymetrix expression microarray results RT-qPCR analysis
was performed as previously described [21] For validationsix of the differentially expressed genes 3 upregulatedmdashGSTO1 PSMB10 and COX8Amdashand 3 downregulatedmdashHIST1H1B MMP9 and ARHGAP11Bmdashby microarray werechosen Validation was done in a randomly selected subset ofthe original samples (submitted for microarray analysis) thatincluded 3 healthy controls and 3 RTT patients Primer pairswere obtained from the Real-time PCR GenBank Primer tohybridise unique regions of the appropriate gene sequenceGSTO1 (Fw 51015840-AGA GTT GTT TTC TAA GGT TCT GACT-31015840) and (RW 51015840-ACT TCA TTG CTT CCA GCC GT-31015840) product length 116 bp PSMB10 (Fw 51015840-ACA GAC GTGAAG CCT AGC AG-31015840) and (Rw 51015840-ACC GAA TCG TTAGTG GCT CG-31015840) product length 294 bp COX8A (Fw 51015840-GCC AAG ATC CAT TCG TTG CC-31015840) and (RW 51015840-TCTGGC CTC CTG TAG GTC TC-31015840) product length 137 bpHIST1H1B (Fw 51015840-CCC GGC TAA GAA GAA GGC AA-31015840) and (RW 51015840-ACA GCC TTG GTG ATC AGC TC-31015840)product length 99 bp MMP9 (Fw 51015840-GTC CGT GAG GGTGTT GAG TG-31015840) and (RW 51015840-ACT GCT CAA AGC CTCCAC AA-31015840) product length 145 bp ARHGAP11B (Fw 51015840-AAC TGC CAG AGC CCA TTC TC-31015840) and (RW 51015840-GTCTGGTACACGCCCTTCTT-31015840) product length 295 bpAllreactions were run in triplicate GAPDH (FW 51015840-TGA CGCTGG GGC TGG CAT TG-31015840 and RW 51015840-GGC TGG TGGTCC AGG GGT CT-31015840 134 pb) was used in our experimentsas internal standard As previously described samples werecompared using the relative cycle threshold (CT) method(Livak and Schmittgen 2001) After normalization to morestable mRNA GAPDH the fold increase or decrease wasdeterminedwith respect to control using the formula 2minusΔΔCTwhere ΔCT is (gene of interest CT)minus(reference gene CT)andΔΔCT is (ΔCT experimental)minus(ΔCT control) Results arethe means plusmn SEM of three independent experiments eachanalysed in triplicate lowast119875 lt 0001 versus control (one-wayANOVA followed by Bonferronirsquos posttest)
3 Results and Discussion
31 Differentially Regulated Genes in RTT Patients Giventhat RTT results from dysfunction of the transcriptionalmodulator MeCP2 several strategies have been developedto identify its target genes in order to gain insights into thedisease pathogenesis [12ndash18] In our study to identify geneexpression changes associated with and potentially related toMeCP2 mutations and to delineate alterations of pathwaysassociated with the disease we evaluated and comparedtranscriptomic profiles in PBMC from RTT patients andcontrol subjects
This work showed for the first time to our knowledgealtered expression of a large set of genes that may helpelucidating and explaining the link between MeCP2 andsome of the molecular and cellular aspects observed in RTTpatients In our previous studies on RTT we were able toshow increased levels of oxidative stress (OS)markers such asisoprostanes (IsoPs) and 4-hydroxynonenal protein adducts(4-HNE PAs) [22 23] and increased ubiquitination anddegradation of oxidatively modified proteins [24] but how
4 Mediators of Inflammation
Up
Down
LIMMA SAM
5
5
1 430
52
Figure 1 Comparison of two distinct approaches for screening ofdifferentially expressed genes in RTT PBMC Venn diagram repre-senting numbers of common and exclusively up- and downregulatedgenes for LIMMA (left) and SAM (right) analyses (FC plusmn 1 adj119875 value le 005) The 11 genes indicated by only one method wereexcluded by further analysis
MeCP2 mutation is able to affect cellular redox balance andproteins turn over still needs to be defined
Using the SAM and LIMMA methods we have defineda set of genes differentially expressed in RTT patients withrespect to controls (healthy subjects) and a significant overlapwas found by comparing results from two approaches Acut-off level based on a minimum of 1-fold change inexpression resulted in a list of 482 common deregulatedgenes while 10 genes were suggested only by SAM andonly 1 gene was indicated by LIMMA Among the sharedgenes 430 showed significant upregulation while 52 weredownregulated in RTT compared to controls (Figure 1 andSupplemental Tables 1 and 2) The 11 genes indicated by onlyone method were excluded for further analysis Genes withthe strongest changes in expression both upregulated (FC ge2) and downregulated (FC le minus12) are listed in Tables 2 and3 respectively The complete list of differentially expressedgenes both upregulated and downregulated (FC plusmn 1) isshown in Supplemental Tables 1 and 2
It is evident from this first set of data that mutations inMeCP2 influence more the genes upregulation with respectto the downregulation This is in part in line with the firstfunctions that have been attributed to MeCP2 such as a geneexpression repressor [10 11] In addition it is worth it tounderline that both screening approaches used in this study(LIMMA and SAM) almost overlap between them makingthe results more reliable
Next using the DAVID databases we performed thefunctional annotation of the significant genes and iden-tification of the biochemical pathways in which they areinvolved A comparison of the differentially regulated mRNAtranscripts in RTT PBMC compared to control group showssignificant changes in cellular pathways In particular weidentify 10 major clusters corresponding to 62 biologicalprocesses enriched by 146 genes (Table 4) These clustershighlight key biological pathways related to mitochondrialfunction and organization (ie mitochondrial ATP synthesis
coupled to electron transport inner mitochondrial mem-brane organization such asATP5A1COX6CETFAUQCRQTIMM10 and TSPO) cellular protein metabolic process (ieregulation of protein ubiquitination regulation of ubiquitin-protein ligase activity and proteasomal ubiquitin-dependentprotein catabolic process such as PSMA2 PSMD6 UBE2E3and UFC) RNA processing (ie nuclear mRNA splicingand spliceosome assembly and RNA elongation from RNApolymerase II promoter such asRPL15) DNAorganization inchromatin and cellular macromolecular complex assembly(ie nucleosome assembly DNA packaging and proteincomplex biogenesis such asHIST1H4L H2AFZ TOP2A andHMGB2)
32 Mitochondrial Related Genes in RTT Patients Amongthese clusters the most significantly regulated transcriptsinclude those encoding several subunits of mitochondrialrespiratory chain complexes and thus linked directly tomitochondrial ATP production and indirectly to potentialreactive oxygen species (ROS) generation In particularNDUFA1 NDUFAB1 NDUFA2 and NDUFB6 all compo-nents of mitochondrial complex I (NADH ubiquinone oxi-doreductase) showed the greater changes with a FC of morethan 2 Moreover other subunits of complex I (NDUFV2NDUFS4 NDUFA9 NDUFS6 NDUFB10 NDUFB4NDUFC2 NDUFB2 NDUFS5 NDUFC1 NDUFB9 andNDUFA8) were clearly upregulated in RTT group
Complex I plays a vital role in cellular ATP productionthe primary source of energy for many crucial processes inliving cells It removes electrons from NADH and passesthem by a series of different protein coupled redox centersto the electron acceptor ubiquinone Because complex I iscentral to energy production in the cell it is reported thatits malfunction results in a wide range of neuromusculardiseases [25] Some of them are due to mutations in themitochondrial genome but others which result from adecrease in the activity of complex I or an increase in theproduction of ROS are not well understood The productionof ROS by complex I is linked to Parkinsonrsquos disease and toageing [26 27] and this is in line with RTT since it is nowwelldocumented as an increased OS condition in this pathology[22 23]
Another gene involved in complex I function isNDUFV2that was also clearly upregulated in our study Mutationsin this gene are implicated in Parkinsonrsquos disease bipo-lar disorder and schizophrenia and have been found inone case of early onset hypertrophic cardiomyopathy andencephalopathy also it has been shown for NDUFA2 asubunit of the hydrophobic protein fraction of the complex IMutations in this gene are associated with Leigh syndromean early onset progressive neurodegenerative disorder Ofnote is NDUFAB1 which is a carrier of the growing fattyacid chain in fatty acid biosynthesis in mitochondria andalteration in fatty acid levels has been noted in ASD [28 29]
Not only complex I subunits were upregulated in RTTbut also we have detected an upregulation of genes involvedin all the five complexes of the electron transport chain Infact also SDHB (succinate dehydrogenase complex subunit
Mediators of Inflammation 5
Table 2 Genes upregulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0045413 NDUFA1NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 1 75 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone 31
NM 0064982 LGALS2Lectingalactoside-bindingsoluble 2
Carbohydrate binding mdash 28
NM 0143023 SEC61G Sec61 gamma subunit Protein transporteractivity
Protein targeting to ER antigenprocessing and presentation ofexogenous peptide antigen viaMHC class I
28
NM 0010404371 C6orf48 Chromosome 6 openreading frame 48 mdash mdash 27
NM 0010985772 RPL31 Ribosomal protein L31 RNA binding structuralconstituent of ribosome
Translational elongationtranslational initiationtranslational termination
27
NM 0069895 RASA4 RAS p21 proteinactivator 4
GTPase activator activityphospholipid binding
Intracellular signal transductionpositive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction
26
NM 0009833 RPL22 Ribosomal protein L22RNA binding heparinbinding structuralconstituent of ribosome
Alpha-beta T cell differentiationtranslational elongationtranslational initiationtranslational termination
26
NM 0190593 TOMM7
Translocase of outermitochondrialmembrane 7 homolog(yeast)
Protein transmembranetransporter activity
Cellular protein metabolic processprotein import into mitochondrialmatrix protein targeting tomitochondrion
26
NM 0311572 HNRNPA1 Heterogeneous nuclearribonucleoprotein A1
Nucleotide bindingsingle-stranded DNAbinding single-strandedRNA binding
RNA export from nucleus mRNAsplicing via spliceosome mRNAtransport nuclear import
25
NM 0018285 CLC Charcot-Leyden crystalgalectin
Carbohydrate bindingcarboxylesterase activitylysophospholipase activity
Lipid catabolic processmulticellular organismaldevelopment
24
NM 0329013 COX14Cytochrome c oxidaseassembly homolog 14(S cerevisiae)
Plays a role in theassembly or stability of thecytochrome c oxidasecomplex (COX)
Mitochondrial respiratory chaincomplex IV assembly 23
NM 0249604 PANK2 Pantothenate kinase 2ATP bindingpantothenate kinaseactivity
Cell death coenzyme Abiosynthetic process coenzymebiosynthetic process pantothenatemetabolic process
23
NR 0023091 RPS26P11 Ribosomal protein S26pseudogene 11
Structural constituent ofribosome Translation 23
NM 0050032 NDUFAB1
NADH dehydrogenase(ubiquinone) 1alphabeta subcomplex1 8 kDa
NADH dehydrogenase(ubiquinone) activityACP phosphopantetheineattachment site bindinginvolved in fatty acidbiosynthetic process fattyacid binding calcium ionbinding
Cellular metabolic process proteinlipoylation small moleculemetabolic process respiratoryelectron transport chain fatty acidbiosynthetic processmitochondrial electron transportNADH to ubiquinone
23
6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

4 Mediators of Inflammation
Up
Down
LIMMA SAM
5
5
1 430
52
Figure 1 Comparison of two distinct approaches for screening ofdifferentially expressed genes in RTT PBMC Venn diagram repre-senting numbers of common and exclusively up- and downregulatedgenes for LIMMA (left) and SAM (right) analyses (FC plusmn 1 adj119875 value le 005) The 11 genes indicated by only one method wereexcluded by further analysis
MeCP2 mutation is able to affect cellular redox balance andproteins turn over still needs to be defined
Using the SAM and LIMMA methods we have defineda set of genes differentially expressed in RTT patients withrespect to controls (healthy subjects) and a significant overlapwas found by comparing results from two approaches Acut-off level based on a minimum of 1-fold change inexpression resulted in a list of 482 common deregulatedgenes while 10 genes were suggested only by SAM andonly 1 gene was indicated by LIMMA Among the sharedgenes 430 showed significant upregulation while 52 weredownregulated in RTT compared to controls (Figure 1 andSupplemental Tables 1 and 2) The 11 genes indicated by onlyone method were excluded for further analysis Genes withthe strongest changes in expression both upregulated (FC ge2) and downregulated (FC le minus12) are listed in Tables 2 and3 respectively The complete list of differentially expressedgenes both upregulated and downregulated (FC plusmn 1) isshown in Supplemental Tables 1 and 2
It is evident from this first set of data that mutations inMeCP2 influence more the genes upregulation with respectto the downregulation This is in part in line with the firstfunctions that have been attributed to MeCP2 such as a geneexpression repressor [10 11] In addition it is worth it tounderline that both screening approaches used in this study(LIMMA and SAM) almost overlap between them makingthe results more reliable
Next using the DAVID databases we performed thefunctional annotation of the significant genes and iden-tification of the biochemical pathways in which they areinvolved A comparison of the differentially regulated mRNAtranscripts in RTT PBMC compared to control group showssignificant changes in cellular pathways In particular weidentify 10 major clusters corresponding to 62 biologicalprocesses enriched by 146 genes (Table 4) These clustershighlight key biological pathways related to mitochondrialfunction and organization (ie mitochondrial ATP synthesis
coupled to electron transport inner mitochondrial mem-brane organization such asATP5A1COX6CETFAUQCRQTIMM10 and TSPO) cellular protein metabolic process (ieregulation of protein ubiquitination regulation of ubiquitin-protein ligase activity and proteasomal ubiquitin-dependentprotein catabolic process such as PSMA2 PSMD6 UBE2E3and UFC) RNA processing (ie nuclear mRNA splicingand spliceosome assembly and RNA elongation from RNApolymerase II promoter such asRPL15) DNAorganization inchromatin and cellular macromolecular complex assembly(ie nucleosome assembly DNA packaging and proteincomplex biogenesis such asHIST1H4L H2AFZ TOP2A andHMGB2)
32 Mitochondrial Related Genes in RTT Patients Amongthese clusters the most significantly regulated transcriptsinclude those encoding several subunits of mitochondrialrespiratory chain complexes and thus linked directly tomitochondrial ATP production and indirectly to potentialreactive oxygen species (ROS) generation In particularNDUFA1 NDUFAB1 NDUFA2 and NDUFB6 all compo-nents of mitochondrial complex I (NADH ubiquinone oxi-doreductase) showed the greater changes with a FC of morethan 2 Moreover other subunits of complex I (NDUFV2NDUFS4 NDUFA9 NDUFS6 NDUFB10 NDUFB4NDUFC2 NDUFB2 NDUFS5 NDUFC1 NDUFB9 andNDUFA8) were clearly upregulated in RTT group
Complex I plays a vital role in cellular ATP productionthe primary source of energy for many crucial processes inliving cells It removes electrons from NADH and passesthem by a series of different protein coupled redox centersto the electron acceptor ubiquinone Because complex I iscentral to energy production in the cell it is reported thatits malfunction results in a wide range of neuromusculardiseases [25] Some of them are due to mutations in themitochondrial genome but others which result from adecrease in the activity of complex I or an increase in theproduction of ROS are not well understood The productionof ROS by complex I is linked to Parkinsonrsquos disease and toageing [26 27] and this is in line with RTT since it is nowwelldocumented as an increased OS condition in this pathology[22 23]
Another gene involved in complex I function isNDUFV2that was also clearly upregulated in our study Mutationsin this gene are implicated in Parkinsonrsquos disease bipo-lar disorder and schizophrenia and have been found inone case of early onset hypertrophic cardiomyopathy andencephalopathy also it has been shown for NDUFA2 asubunit of the hydrophobic protein fraction of the complex IMutations in this gene are associated with Leigh syndromean early onset progressive neurodegenerative disorder Ofnote is NDUFAB1 which is a carrier of the growing fattyacid chain in fatty acid biosynthesis in mitochondria andalteration in fatty acid levels has been noted in ASD [28 29]
Not only complex I subunits were upregulated in RTTbut also we have detected an upregulation of genes involvedin all the five complexes of the electron transport chain Infact also SDHB (succinate dehydrogenase complex subunit
Mediators of Inflammation 5
Table 2 Genes upregulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0045413 NDUFA1NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 1 75 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone 31
NM 0064982 LGALS2Lectingalactoside-bindingsoluble 2
Carbohydrate binding mdash 28
NM 0143023 SEC61G Sec61 gamma subunit Protein transporteractivity
Protein targeting to ER antigenprocessing and presentation ofexogenous peptide antigen viaMHC class I
28
NM 0010404371 C6orf48 Chromosome 6 openreading frame 48 mdash mdash 27
NM 0010985772 RPL31 Ribosomal protein L31 RNA binding structuralconstituent of ribosome
Translational elongationtranslational initiationtranslational termination
27
NM 0069895 RASA4 RAS p21 proteinactivator 4
GTPase activator activityphospholipid binding
Intracellular signal transductionpositive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction
26
NM 0009833 RPL22 Ribosomal protein L22RNA binding heparinbinding structuralconstituent of ribosome
Alpha-beta T cell differentiationtranslational elongationtranslational initiationtranslational termination
26
NM 0190593 TOMM7
Translocase of outermitochondrialmembrane 7 homolog(yeast)
Protein transmembranetransporter activity
Cellular protein metabolic processprotein import into mitochondrialmatrix protein targeting tomitochondrion
26
NM 0311572 HNRNPA1 Heterogeneous nuclearribonucleoprotein A1
Nucleotide bindingsingle-stranded DNAbinding single-strandedRNA binding
RNA export from nucleus mRNAsplicing via spliceosome mRNAtransport nuclear import
25
NM 0018285 CLC Charcot-Leyden crystalgalectin
Carbohydrate bindingcarboxylesterase activitylysophospholipase activity
Lipid catabolic processmulticellular organismaldevelopment
24
NM 0329013 COX14Cytochrome c oxidaseassembly homolog 14(S cerevisiae)
Plays a role in theassembly or stability of thecytochrome c oxidasecomplex (COX)
Mitochondrial respiratory chaincomplex IV assembly 23
NM 0249604 PANK2 Pantothenate kinase 2ATP bindingpantothenate kinaseactivity
Cell death coenzyme Abiosynthetic process coenzymebiosynthetic process pantothenatemetabolic process
23
NR 0023091 RPS26P11 Ribosomal protein S26pseudogene 11
Structural constituent ofribosome Translation 23
NM 0050032 NDUFAB1
NADH dehydrogenase(ubiquinone) 1alphabeta subcomplex1 8 kDa
NADH dehydrogenase(ubiquinone) activityACP phosphopantetheineattachment site bindinginvolved in fatty acidbiosynthetic process fattyacid binding calcium ionbinding
Cellular metabolic process proteinlipoylation small moleculemetabolic process respiratoryelectron transport chain fatty acidbiosynthetic processmitochondrial electron transportNADH to ubiquinone
23
6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 5
Table 2 Genes upregulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0045413 NDUFA1NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 1 75 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone 31
NM 0064982 LGALS2Lectingalactoside-bindingsoluble 2
Carbohydrate binding mdash 28
NM 0143023 SEC61G Sec61 gamma subunit Protein transporteractivity
Protein targeting to ER antigenprocessing and presentation ofexogenous peptide antigen viaMHC class I
28
NM 0010404371 C6orf48 Chromosome 6 openreading frame 48 mdash mdash 27
NM 0010985772 RPL31 Ribosomal protein L31 RNA binding structuralconstituent of ribosome
Translational elongationtranslational initiationtranslational termination
27
NM 0069895 RASA4 RAS p21 proteinactivator 4
GTPase activator activityphospholipid binding
Intracellular signal transductionpositive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction
26
NM 0009833 RPL22 Ribosomal protein L22RNA binding heparinbinding structuralconstituent of ribosome
Alpha-beta T cell differentiationtranslational elongationtranslational initiationtranslational termination
26
NM 0190593 TOMM7
Translocase of outermitochondrialmembrane 7 homolog(yeast)
Protein transmembranetransporter activity
Cellular protein metabolic processprotein import into mitochondrialmatrix protein targeting tomitochondrion
26
NM 0311572 HNRNPA1 Heterogeneous nuclearribonucleoprotein A1
Nucleotide bindingsingle-stranded DNAbinding single-strandedRNA binding
RNA export from nucleus mRNAsplicing via spliceosome mRNAtransport nuclear import
25
NM 0018285 CLC Charcot-Leyden crystalgalectin
Carbohydrate bindingcarboxylesterase activitylysophospholipase activity
Lipid catabolic processmulticellular organismaldevelopment
24
NM 0329013 COX14Cytochrome c oxidaseassembly homolog 14(S cerevisiae)
Plays a role in theassembly or stability of thecytochrome c oxidasecomplex (COX)
Mitochondrial respiratory chaincomplex IV assembly 23
NM 0249604 PANK2 Pantothenate kinase 2ATP bindingpantothenate kinaseactivity
Cell death coenzyme Abiosynthetic process coenzymebiosynthetic process pantothenatemetabolic process
23
NR 0023091 RPS26P11 Ribosomal protein S26pseudogene 11
Structural constituent ofribosome Translation 23
NM 0050032 NDUFAB1
NADH dehydrogenase(ubiquinone) 1alphabeta subcomplex1 8 kDa
NADH dehydrogenase(ubiquinone) activityACP phosphopantetheineattachment site bindinginvolved in fatty acidbiosynthetic process fattyacid binding calcium ionbinding
Cellular metabolic process proteinlipoylation small moleculemetabolic process respiratoryelectron transport chain fatty acidbiosynthetic processmitochondrial electron transportNADH to ubiquinone
23
6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

6 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1528512 MS4A6AMembrane-spanning4-domains subfamilyA member 6A
May be involved in signaltransduction as acomponent of amultimeric receptorcomplex
mdash 23
NM 0042693 MED27 Mediator complexsubunit 27
Transcription coactivatoractivity
Regulation of transcription fromRNA polymerase II promoterstem cell maintenancetranscription initiation from RNApolymerase II promoter
22
NR 0154041 C12orf47 MAPKAPK5 antisenseRNA 1 mdash mdash 22
NM 0018653 COX7A2Cytochrome c oxidasesubunit VIIapolypeptide 2 (liver)
Cytochrome-c oxidaseactivity electron carrieractivity
Oxidative phosphorylation 22
NM 0043743 COX6C Cytochrome c oxidasesubunit VIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
22
NM 0029842 CCL4 Chemokine (C-Cmotif) ligand 4 Chemokine activity
Cell adhesion cell-cell signalingchemotaxis immune responseinflammatory response positiveregulation of calcium iontransport positive regulation ofcalcium-mediated signalingpositive regulation of natural killercell chemotaxisresponse to toxic substanceresponse to virus signaltransduction
22
NM 0140602 MCTS1 Malignant T cellamplified sequence 1 RNA binding
Cell cycle positive regulation ofcell proliferation regulation ofgrowth regulation oftranscription DNA-dependentresponse to DNA damage stimulustranscription DNA-dependent
21
NM 0048322 GSTO1 GlutathioneS-transferase omega 1
Glutathionedehydrogenase(ascorbate) activityglutathione transferaseactivity methylarsonatereductase activity
L-ascorbic acid metabolic processglutathione derivative biosyntheticprocess negative regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation ofryanodine-sensitivecalcium-release channel activitypositive regulation of skeletalmuscle contraction by regulationof release of sequestered calciumion regulation of cardiac musclecontraction by regulation of therelease of sequestered calcium ionxenobiotic catabolic process
21
NM 0018672 COX7C Cytochrome c oxidasesubunit VIIc
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
21
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001
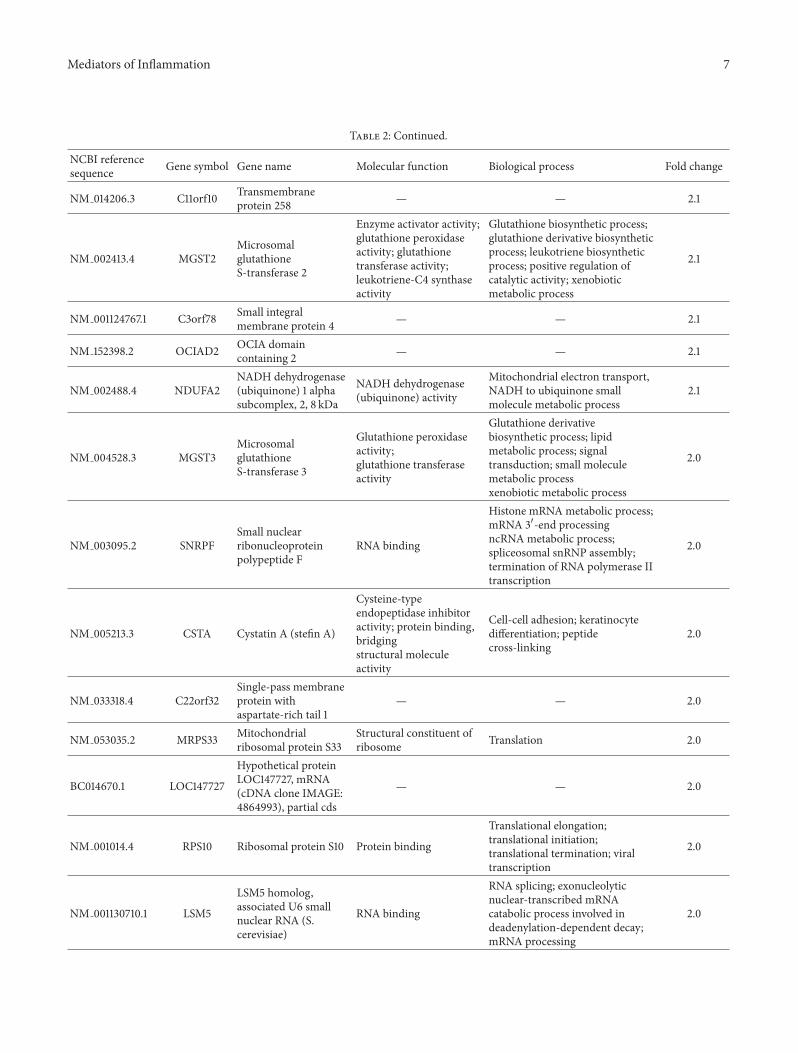
Mediators of Inflammation 7
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0142063 C11orf10 Transmembraneprotein 258 mdash mdash 21
NM 0024134 MGST2MicrosomalglutathioneS-transferase 2
Enzyme activator activityglutathione peroxidaseactivity glutathionetransferase activityleukotriene-C4 synthaseactivity
Glutathione biosynthetic processglutathione derivative biosyntheticprocess leukotriene biosyntheticprocess positive regulation ofcatalytic activity xenobioticmetabolic process
21
NM 0011247671 C3orf78 Small integralmembrane protein 4 mdash mdash 21
NM 1523982 OCIAD2 OCIA domaincontaining 2 mdash mdash 21
NM 0024884 NDUFA2NADH dehydrogenase(ubiquinone) 1 alphasubcomplex 2 8 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
21
NM 0045283 MGST3MicrosomalglutathioneS-transferase 3
Glutathione peroxidaseactivityglutathione transferaseactivity
Glutathione derivativebiosynthetic process lipidmetabolic process signaltransduction small moleculemetabolic processxenobiotic metabolic process
20
NM 0030952 SNRPFSmall nuclearribonucleoproteinpolypeptide F
RNA binding
Histone mRNA metabolic processmRNA 31015840-end processingncRNA metabolic processspliceosomal snRNP assemblytermination of RNA polymerase IItranscription
20
NM 0052133 CSTA Cystatin A (stefin A)
Cysteine-typeendopeptidase inhibitoractivity protein bindingbridgingstructural moleculeactivity
Cell-cell adhesion keratinocytedifferentiation peptidecross-linking
20
NM 0333184 C22orf32Single-pass membraneprotein withaspartate-rich tail 1
mdash mdash 20
NM 0530352 MRPS33 Mitochondrialribosomal protein S33
Structural constituent ofribosome Translation 20
BC0146701 LOC147727
Hypothetical proteinLOC147727 mRNA(cDNA clone IMAGE4864993) partial cds
mdash mdash 20
NM 0010144 RPS10 Ribosomal protein S10 Protein binding
Translational elongationtranslational initiationtranslational termination viraltranscription
20
NM 0011307101 LSM5
LSM5 homologassociated U6 smallnuclear RNA (Scerevisiae)
RNA binding
RNA splicing exonucleolyticnuclear-transcribed mRNAcatabolic process involved indeadenylation-dependent decaymRNA processing
20
8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

8 Mediators of Inflammation
Table 2 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0028013 PSMB10Proteasome (prosomemacropain) subunitbeta type 10
Threonine-typeendopeptidase activity
DNA damage response signaltransduction by p53 class mediatorresulting in cell cycle arrest G1Stransition of mitotic cell cycle Tcell proliferationanaphase-promotingcomplex-dependent proteasomalubiquitin-dependent proteincatabolic process apoptoticprocess cell morphogenesis geneexpression humoral immuneresponse mRNA metabolicprocess negative regulation ofapoptotic process negativeregulation of ubiquitin-proteinligase activity involved in mitoticcell cycle positive regulation ofubiquitin-protein ligase activityinvolved in mitotic cell cycleprotein polyubiquitinationregulation of cellular amino acidmetabolic process small moleculemetabolic process
20
NM 0140445 UNC50 Unc-50 homolog (Celegans) RNA binding Cell surface receptor signaling
pathway protein transport 20
NM 0327473 USMG5Up-regulated duringskeletal muscle growth5 homolog (mouse)
Plays a critical role inmaintaining the ATPsynthase population inmitochondria
mdash 20
NM 0010013302 REEP3 Receptor accessoryprotein 3
May enhance the cellsurface expression ofodorant receptors
mdash 20
NM 0040742 COX8ACytochrome c oxidasesubunit VIIIA(ubiquitous)
Cytochrome-c oxidaseactivity
Respiratory electron transportchain small molecule metabolicprocess
20
NM 0024934 NDUFB6NADH dehydrogenase(ubiquinone) 1 betasubcomplex 6 17 kDa
NADH dehydrogenase(ubiquinone) activity
Mitochondrial electron transportNADH to ubiquinone smallmolecule metabolic process
20
NM 0322733 TMEM126A Transmembraneprotein 126A mdash Optic nerve development 20
ldquomdashrdquo lacking item
B and iron sulfur (Ip)) gene encoding for a subunit of mito-chondrial complex II (succinate ubiquinone oxidoreductase)was significantly upregulated This subunit is responsiblefor transferring electrons from succinate to ubiquinone(coenzyme Q) Complex II of the respiratory chain whichis specifically involved in the oxidation of succinate carrieselectrons from FADH to CoQ Of note also 3 genesUQCRQUQCRFS1 andUQCRH encoding for subunits of complex III(ubiquinol-cytochrome c oxidoreductase) were upregulatedwith a mean FC = 160 This complex plays a critical role inbiochemical generation of ATP contributing to the genera-tion of electrochemical potential by catalyzing the electrontransfer reaction from ubiquinol to cytochrome c coupled
with proton translocation across the membrane Lines ofevidence report that in mouse models some of the promotersof ubiquinol-cytochrome c reductase subunit are able to betargeted byMECP2 protein contributing to the developmentof the pathology [30]
Furthermore the cytochrome c gene (CYCS) togetherwith genes encoding subunits of mitochondrial complex IV(cytochrome c oxidase) (COX14 COX7A2 COX6C COX7Cand COX8A) was upregulated with a mean FC of circa 15 Ofnote is the upregulation of cytochrome c gene The encodedprotein accepts electrons from cytochrome b and transfersthem to the cytochrome oxidase complexThis protein is alsoinvolved in initiation of apoptosis and this would be in line
Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 9
Table 3 Genes down-regulated in RTT patients by LIMMA and SAM analyses
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 1706014 SIAE Sialic acid acetylesterase Sialate O-acetylesteraseactivity mdash minus23
NR 0023121 RPPH1 Ribonuclease P RNAcomponent H1 mdash mdash minus22
NM 0053222 HIST1H1B Histone cluster 1 H1b DNA binding Nucleosome assembly minus18
NM 0009023 MME Membranemetalloendopeptidase
Metalloendopeptidaseactivity Proteolysis minus18
NM 0320474 B3GNT5
UDP-GlcNAc betaGalbeta-13-N-acetylglucosaminyltransferase5
Galactosyltransferaseactivity
Glycolipid biosynthetic processposttranslational proteinmodification
minus17
NM 0035132 HIST1H2AB Histone cluster 1 H2ab DNA binding Nucleosome assembly minus17
NR 0025621 SNORD28 Small nucleolar RNA CDbox 28 mdash mdash minus15
NM 0046682 MGAM Maltase-glucoamylase(alpha-glucosidase)
Alpha-glucosidaseactivity amylaseactivity
Carbohydrate metabolic process minus15
NM 0120815 ELL2 Elongation factor RNApolymerase II 2 mdash Regulation of transcription
DNA-dependent minus14
NM 0210662 HIST1H2AJ Histone cluster 1 H2aj DNA binding nucleosome assembly minus14
NM 0024242 MMP8 Matrix metallopeptidase 8(neutrophil collagenase)
Metalloendopeptidaseactivity zinc ionbinding
Proteolysis minus14
NM 0049942 MMP9
Matrix metallopeptidase 9(gelatinase B 92 kDagelatinase 92 kDa type IVcollagenase)
Collagen bindingmetalloendopeptidaseactivity zinc ionbinding
Collagen catabolic processextracellular matrix disassemblypositive regulation of apoptoticprocess proteolysis
minus14
NM 0035332 HIST1H3I Histone cluster 1 H3i DNA binding Nucleosome assembly regulationof gene silencing minus13
NG 0008614 GK3P Glycerol kinase 3 pseudogene ATP binding glycerolkinase activity
Catabolic process glycerolmetabolic process minus13
NR 0334231 LOC1720 Dihydrofolate reductasepseudogene mdash mdash minus13
NM 0035212 HIST1H2BM Histone cluster 1 H2bm DNA binding Nucleosome assembly minus13
NM 0024174 MKI67 Antigen identified bymonoclonal antibody Ki-67 ATP binding
DNA metabolic process cellproliferation cellular response toheat meiosis organ regeneration
minus12
NM 0204062 CD177 CD177 molecule mdash Blood coagulation leukocytemigration minus12
NM 0010398411 ARHGAP11B Rho GTPase activatingprotein 11B
GTPase activatoractivity
Positive regulation of GTPaseactivity regulation of smallGTPase mediated signaltransduction small GTPasemediated signal transduction
minus12
NM 0010046901 OR2M5 Olfactory receptor family 2subfamily M member 5
G-protein coupledreceptor activityolfactory receptoractivity
Detection of chemical stimulusinvolved in sensory perception ofsmell
minus12
NM 0529663 FAM129A Family with sequencesimilarity 129 member A mdash
Negative regulation of proteinphosphorylation positiveregulation of proteinphosphorylation positiveregulation of translation responseto endoplasmic reticulum stress
minus12
10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

10 Mediators of Inflammation
Table 3 Continued
NCBI referencesequence Gene symbol Gene name Molecular function Biological process Fold change
NM 0010673 TOP2A Topoisomerase (DNA) IIalpha 170 kDa
ATP binding DNAbinding bendingchromatin bindingdrug bindingmagnesium ionbinding ubiquitinbinding
DNA ligation DNA repair DNAtopological changeDNA-dependent DNA replicationapoptotic chromosomecondensation mitotic cell cyclephosphatidylinositol-mediatedsignaling positive regulation ofapoptotic process positiveregulation of retroviral genomereplication positive regulation oftranscription from RNApolymerase II promoter sisterchromatid segregation
minus12
NM 0210182 HIST1H3F Histone cluster 1 H3f DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 1827072 PSG8 Pregnancy specificbeta-1-glycoprotein 8
The humanpregnancy-specificglycoproteins (PSGs)are a group ofmolecules that aremainly produced by theplacentalsyncytiotrophoblastsduring pregnancy PSGscomprise a subgroup ofthe carcinoembryonicantigen (CEA) familywhichbelongs to theimmunoglobulinsuperfamily
Female pregnancy minus12
NM 0035352 HIST1H3J Histone cluster 1 H3j DNA bindingS phase blood coagulationnucleosome assemblyregulation of gene silencing
minus12
NM 0045663 PFKFB3
6-Phosphofructo-2-kinasefructose-26-biphosphatase3
6-Phosphofructo-2-kinase activity ATPbinding fructose-26-bisphosphate2-phosphatase activity
Fructose metabolic processglycolysis small moleculemetabolic process
minus12
NM 0164482 DTLDenticleless E3ubiquitin-protein ligasehomolog (Drosophila)
mdash
DNA replication G2 DNAdamage checkpoint proteinmonoubiquitination proteinpolyubiquitinationubiquitin-dependent proteincatabolic process
minus12
NG 0010195 IGHM Immunoglobulin heavyconstant mu Antigen binding Immune response minus12
NR 0029072 SNORA73A Small nucleolar RNA HACAbox 73A mdash mdash minus12
ldquomdashrdquo lacking item
with previous studies that have shown a possible involvementof apoptosis in RTT [31 32] although only few studies haveinvestigated the role of apoptosis in RTT and the currentliterature is still controversial In addition several subunits
of complex IV were upregulated It receives an electron fromeach of the four cytochrome c molecules and transfers themto one oxygenmolecule converting molecular oxygen to twomolecules of water In the process via a protons translocation
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001
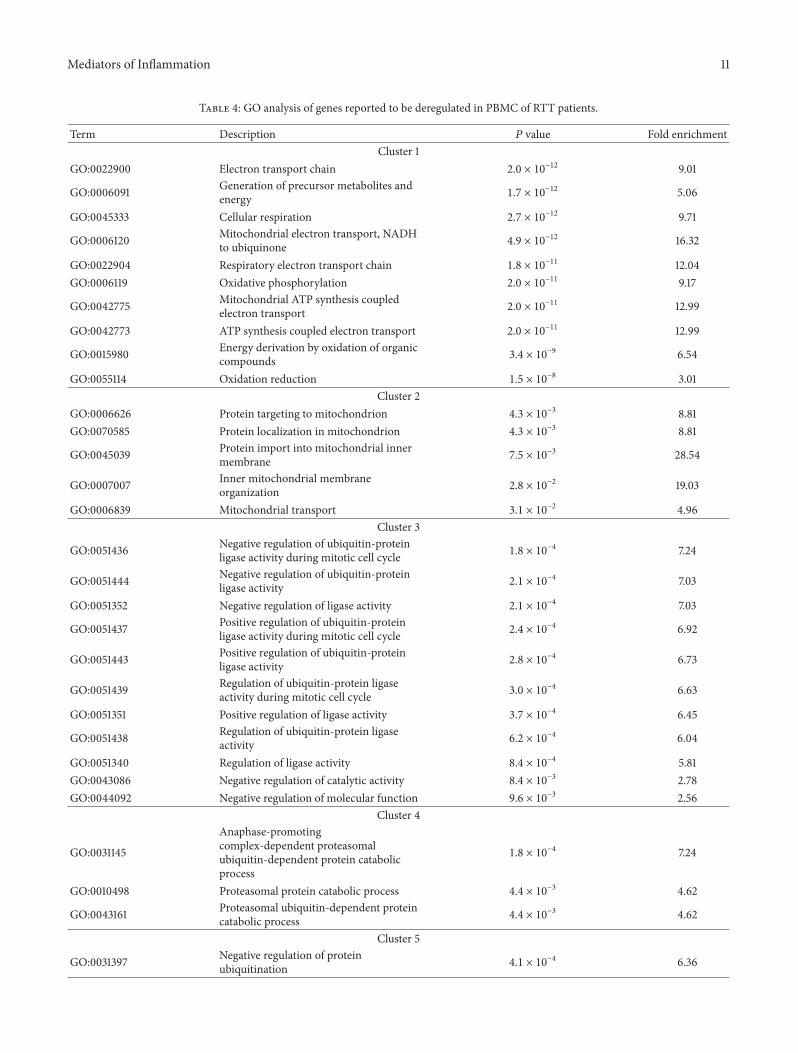
Mediators of Inflammation 11
Table 4 GO analysis of genes reported to be deregulated in PBMC of RTT patients
Term Description 119875 value Fold enrichmentCluster 1
GO0022900 Electron transport chain 20 times 10minus12 901
GO0006091 Generation of precursor metabolites andenergy 17 times 10
minus12 506
GO0045333 Cellular respiration 27 times 10minus12 971
GO0006120 Mitochondrial electron transport NADHto ubiquinone 49 times 10
minus12 1632
GO0022904 Respiratory electron transport chain 18 times 10minus11 1204
GO0006119 Oxidative phosphorylation 20 times 10minus11 917
GO0042775 Mitochondrial ATP synthesis coupledelectron transport 20 times 10
minus11 1299
GO0042773 ATP synthesis coupled electron transport 20 times 10minus11 1299
GO0015980 Energy derivation by oxidation of organiccompounds 34 times 10
minus9 654
GO0055114 Oxidation reduction 15 times 10minus8 301
Cluster 2GO0006626 Protein targeting to mitochondrion 43 times 10
minus3 881GO0070585 Protein localization in mitochondrion 43 times 10
minus3 881
GO0045039 Protein import into mitochondrial innermembrane 75 times 10
minus3 2854
GO0007007 Inner mitochondrial membraneorganization 28 times 10
minus2 1903
GO0006839 Mitochondrial transport 31 times 10minus2 496
Cluster 3
GO0051436 Negative regulation of ubiquitin-proteinligase activity during mitotic cell cycle 18 times 10
minus4 724
GO0051444 Negative regulation of ubiquitin-proteinligase activity 21 times 10
minus4 703
GO0051352 Negative regulation of ligase activity 21 times 10minus4 703
GO0051437 Positive regulation of ubiquitin-proteinligase activity during mitotic cell cycle 24 times 10
minus4 692
GO0051443 Positive regulation of ubiquitin-proteinligase activity 28 times 10
minus4 673
GO0051439 Regulation of ubiquitin-protein ligaseactivity during mitotic cell cycle 30 times 10
minus4 663
GO0051351 Positive regulation of ligase activity 37 times 10minus4 645
GO0051438 Regulation of ubiquitin-protein ligaseactivity 62 times 10
minus4 604
GO0051340 Regulation of ligase activity 84 times 10minus4 581
GO0043086 Negative regulation of catalytic activity 84 times 10minus3 278
GO0044092 Negative regulation of molecular function 96 times 10minus3 256
Cluster 4
GO0031145
Anaphase-promotingcomplex-dependent proteasomalubiquitin-dependent protein catabolicprocess
18 times 10minus4 724
GO0010498 Proteasomal protein catabolic process 44 times 10minus3 462
GO0043161 Proteasomal ubiquitin-dependent proteincatabolic process 44 times 10
minus3 462
Cluster 5
GO0031397 Negative regulation of proteinubiquitination 41 times 10
minus4 636
12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

12 Mediators of Inflammation
Table 4 Continued
Term Description 119875 value Fold enrichment
GO0031400 Negative regulation of proteinmodification process 92 times 10
minus4 468
GO0031398 Positive regulation of proteinubiquitination 10 times 10
minus3 561
GO0032269 Negative regulation of cellular proteinmetabolic process 32 times 10
minus3 358
GO0031396 Regulation of protein ubiquitination 42 times 10minus3 471
GO0051248 Negative regulation of protein metabolicprocess 44 times 10
minus3 343
GO0031401 Positive regulation of proteinmodification process 41 times 10
minus2 298
GO0032268 Regulation of cellular protein metabolicprocess 47 times 10
minus2 208
Cluster 6GO0065003 Macromolecular complex assembly 14 times 10
minus8 296
GO0043933 Macromolecular complex subunitorganization 99 times 10
minus8 277
GO0034622 Cellular macromolecular complexassembly 32 times 10
minus6 363
GO0034621 Cellular macromolecular complexsubunit organization 26 times 10
minus5 324
GO0065004 Protein-DNA complex assembly 11 times 10minus4 612
GO0034728 Nucleosome organization 52 times 10minus4 552
GO0006461 Protein complex assembly 21 times 10minus3 237
GO0070271 Protein complex biogenesis 21 times 10minus3 237
Cluster 7GO0006334 Nucleosome assembly 24 times 10
minus4 612GO0031497 Chromatin assembly 31 times 10
minus4 590GO0006323 DNA packaging 82 times 10
minus4 476GO0006333 Chromatin assembly or disassembly 60 times 10
minus3 404Cluster 8
GO0008380 RNA splicing 56 times 10minus6 377
GO0006396 RNA processing 57 times 10minus6 282
GO0006397 mRNA processing 15 times 10minus4 320
GO0000398 Nuclear mRNA splicing via spliceosome 20 times 10minus4 448
GO0000377RNA splicing via transesterificationreactions with bulged adenosine asnucleophile
20 times 10minus4 448
GO0000375 RNA splicing via transesterificationreactions 20 times 10
minus4 448
GO0016071 mRNA metabolic process 85 times 10minus4 278
Cluster 9GO0006412 Translation 26 times 10
minus8 401GO0006414 Translational elongation 54 times 10
minus5 593Cluster 10
GO0006367 Transcription initiation from RNApolymerase II promoter 11 times 10
minus3 630
GO0006352 Transcription initiation 45 times 10minus3 516
Cluster 11
GO0006368 RNA elongation from RNA polymerase IIpromoter 11 times 10
minus3 713
GO0006354 RNA elongation 45 times 10minus3 671
Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 13
it is able to generate ATP This data would suggest that RTTpatients are in continuous new ATP synthesis and this couldbe a consequence of new protein synthesis
Our data are in line with a previous work by Kriaucioniswhere the authors have analyzed the gene profile in the brainof RTT animal model [30] They have shown that there wereseveral mitochondrial abnormalities and an upregulation ofboth complexes I and III subunits In particular they werealso able to show a correlation between upregulation ofcomplexes I and III and the animal symptoms severity with asignificant increase inmitochondrial respiratory capacity anda reduction in respiratory efficiency The defect appears to beassociated with respiratory complex III which is also upregu-lated in our study and that containing the Uqcrc1 protein Inaddition it has been shown thatMeCP2 binds to the promoterof the Uqcr gene in vivo and that Uqcr mRNA expressionis elevated in brains of Mecp2-null mice that have acquiredneurological symptoms and this is in line with our results
Finally we also observed in RTT PBMC an upregula-tion of mitochondrial complex V (ATP synthase) subunits(ATP5A1 ATP5EP2 ATP5J2 and ATP5O) together withATPase inhibitory factor 1 gene (ATPIF1) Mitochondrialmembrane ATP synthase is a master regulator of energymetabolism and cell fate therefore a misregulation of thisgene can be associated with altered ATP production and cellmetabolism It is interesting to note that also the ATPaseinhibitory factor 1 (ATPIF1) that inhibits the activity of themitochondrial H+-ATP synthase was upregulated and thislets us speculate the existence of an aberrant loop betweenmaking new ATP and inhibiting its production In additionrecent findings indicate that ATPIF1 has additional functionsby promoting adaptive responses of cell to ROS [33] acondition (OS) that has been well documented to be presentin RTT [22 23]
Similarly other genes related to theATP synthesis showedsignificant expression changes (ie CYB5A CYB561D2ETFA LDHB PDHB and SURF1) For instance ETFA (elec-tron transfer flavoprotein alpha polypeptide) serves as aspecific electron acceptor for several dehydrogenases and inmitochondria it shuttles electrons between primary flavo-protein dehydrogenases and the membrane-bound electrontransfer flavoprotein ubiquinone oxidoreductase In additionLDHB encodes lactate dehydrogenase B an enzyme thatcatalyzes the reversible conversion of lactate and pyruvate andNAD and NADH in the glycolytic pathway being thereforecorrelatedwithATPgenerationOf note is the upregulation ofSURF1 (surfeit 1) gene that encodes a protein localized to theinner mitochondrial membrane and thought to be involvedin the biogenesis of the cytochrome c oxidase complex
Related to mitochondrial structureorganization wefound upregulation of 7 translocase genes (TIMM10 TSPOTIMM9 TIMM17A TOMM7 TIMM13 and TIMM8B)involved in proteins import into mitochondrion (mean FCof 134) and of several mitochondrial ribosomal proteins(MRPL13 MRPL20 MRPL21 MRPL33 MRPL51 MRPL52MRPS25 MRPS30 MRPS33 MRPS36 RPL10A RPL13RPL15 RPL22 RPL22L1 RPL26 RPL31 RPL32 RPL39LRPS10 RPS25 RPS26 RPS26P11 RPS29 RPS5 and RPS7)with a mean FC of 15
All together these lines of evidence seem to suggest anincreased mitochondrial activity that might be linked to theobservation of the pathologic phenotype Anyways at thisstage of the study it is not possible to determine whether ornot there is an increase inATP levels It is possible to speculatethat increased genes related to mitochondrial subunits couldbe a consequence of increased cells request of energy (ATP)This hypothesis is in line with recent study where the authorshave showndecreased levels of ATP in brainmouse RTT [34]
Recent discussions regarding a possible connectionbetween RTT andmitochondrial dysfunction have generatedsignificant interest The basis for these discussions is relatedin part to the common features of RTT on the one hand anddisorders of mitochondrial function on the other Of interestis the fact that a patient with symptoms normally associatedwith mitochondrial disorders harbored mutations in theMeCP2 gene [35] This overlap between symptoms of RTTandmitochondrial disorders recalls early reports of structuralabnormalities [36 37] and defects in the electron transportchain [37 38] inmitochondria from skin andmuscle biopsiesof RTT patients Moreover about half of RTT patients werereported to have elevated levels of circulatory lactic or pyruvicacid which might be caused by defects in the efficiencyof the respiratory chain and urea cycle complexes both ofwhich are mitochondrial [39ndash41] Several disorders relatedto the brain are a consequence of mitochondrial alterationwith the resultant increase of OS and in certain cases thecell apoptosis As RTT is not a neurodegenerative disorder[42] any contribution of mitochondrial dysfunction to RTTsymptoms may take the form of chronic mitochondrialunderperformance rather than catastrophic failure leading toneuronal death
To date no systematic study of mitochondrial function inindividuals with RTT has presented whether these findingsrepresent a primary or secondary effect that is are theyinvolved directly in the clinical features of RTT or do theyreflect effects of these clinical features onmitochondrial func-tion Prior to identification of mutations in MeCP2 in 1999several reports appeared to be related tomitochondrial struc-ture and function [36ndash54] However since 2001 publicationson a possible role ofmitochondria in the pathogenesis of RTThave been very few [36 54] In a recent work investigators inAustralia reported gene expression results from postmortembrain tissue of individualswithRTTandnormal controls [55]One gene related to a mitochondrial enzyme cytochromec oxidase subunit 1 had reduced expression in RTT tissueraising the possibility that loss of MeCP2 function could beresponsible However whether this is a primary or secondaryfinding remains to be established and provides an importanttarget for further investigation
In summary while mitochondrial abnormalities relatedto structure and functions have been reported sufficientinformation is lacking as to the precise role of such abnor-malities in RTT As mentioned alteration of mitochondrialfunctions is often correlated with OS and in particular themitochondrial sites that are often invoked as the most impor-tant mitochondrial superoxide producers are in respiratorycomplexes I and III [56 57] and this can explain the increasedOS levels found in RTT patients
14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

14 Mediators of Inflammation
10
20
30
40
COX8APSMB10GSTO1
mRN
A ex
pres
sion
leve
l (au
)
mRN
A ex
pres
sion
leve
l (au
)
00
05
10
15
ARHGAP11BMMP9HIST1H1B00
Healthy controls RTT patients
Healthy controls RTT patients
lowast
lowastlowast
lowast
lowast
lowast
Figure 2 Validation of relative gene expression levels for selected genes using RT-qPCRin PBMC from 12 RTT patients and 7 controls Resultsare themeansplusmn SEMof three independent experiments each analysed in triplicate lowast119875 valuelt 0001 versus control (one-wayANOVA followedby Bonferronirsquos post-test)
33 Oxidative Stress Related Genes in RTT Patients Thepresence of a redox unbalance in RTT is confirmed by theupregulations of several genes involved in redox homeostasissuch as superoxide dismutase 1 catalase and peroxiredoxin1 (SOD1 CAT and PRDX1) with a 16 114 and 112 FCrespectively Moreover glutathione S-transferase omega 1microsomal glutathione S-transferase 2 and microsomalglutathione S-transferase 3 (GSTO1MGST2 andMGST3) arealso overexpressed in RTTwith a FC= 21 For instance SOD1upregulation could be a consequence of increased superoxideproduction by aberrant activation of complex I and III and thedismutation of superoxide in H
2O2can explain the increased
expression of CAT and PRDX1 It is likely to believe that thecompensatory antioxidant system is not quite sufficient toquench ROS production and this could explain the high OSlevel present in RTT [22 23] For this reason the induction ofglutathione S-transferase omega-1 (GSTO1) is not surprisingThis enzyme is involved in the detoxification mechanismsvia conjugation of reduced glutathione (GSH) to oxidativellymodified proteins (carbonyls and 4-HNE PAs) In fact theGST genes are upregulated in response to OS In additionour results showed the upregulation of MGST2 and MGST3which are the microsomal glutathione S-transferases thisdata correlates very well with our previous findings whereRTT patients showed an increased level of 4-HNE PAs asMGST2 is able to also conjugate 4-HNE with GSH [58]We also observed an increased expression of mRNA foralcohol dehydrogenase 5 (ADH5) aldo-keto reductase family1 member A1 (AKR1A1) and aldehyde dehydrogenase 1family member A1 (ALDH1A1) all enzymes involved in lipidperoxidation products detoxification [59]
34 Ubiquitin-Proteasome Related Genes in RTT PatientsOur results evidenced also the upregulation of genes relatedto protein degradation and ubiquitination In fact RTTPBMC microarray data revealed increased expression lev-els of genes associated with protein turnover such asgenes encoding proteasome subunits (PSMA2 PSMA3
PSMA5 PSMA7 PSMB1 PSMB10 PSMC5 PSMC6 PSMD6and PSMD9) furthermore proteasome maturation protein(POMP) involved in proteasome assembly and genes reg-ulating the activity of the ubiquity ligases (RBX1 UFC1CCNB1IP1 and DAXX) are also up regulated (Table 2) sug-gesting an increase in cell and protein degradation processesThis could also be a consequence of oxidized proteins and thepresence of 4-HNE PAs This evidence is supported by theobserved upregulation of the ubiquitin-conjugating enzymeE2E3 (UBE2E3 FC = 119) that accepts ubiquitin from theE1 complex and catalyzes its covalent attachment to otherproteins However this is in contrast with some lines ofevidence that link RTT to the downregulation of ubiquitinconjugating enzymes (UBE3A) by MeCP2 [60] Overall theeffect of MeCP2 on UBE3A regulation is still controversialIn fact there is even a recent work that did not find anydifference inUBE3A expression betweenwild type and a RTTmouse with the mutation R168X [61] In general the levelsof protein ubiquitination that is one of the steps to degrademodified proteins are increased in RTT [24]
35 Chromatin Folding Related Genes in RTT PatientsIn addition several histone related genes (HIST1H1BHIST1H2AB HIST1H2AI HIST1H2AJ HIST1H2ALHIST1H2BB HIST1H2BH HIST1H2BM HIST1H3BHIST1H3F HIST1H3I HIST1H3J HIST1H4D HIST1H4Fand HIST1H4) were downregulated with a mean FC of minus128suggesting a reduced production of proteins necessary to theDNA chromatin assembly
In general histone modifications are very dynamic andinclude acetylation methylation isomerization phosphory-lation and ubiquitination [62] The combination of suchmodifications confers enormous variability of cellular signalsto environmental stimuli It is easy to understand that mod-ifications such as histone methylation can display additionalcomplexity since the degree of methylation is very variable(mono- di- or trimethylation) [63] Furthermore combina-tions or sequential additions of different histone marks can
Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 15
PSMB10PSMB1
PSMA2
UBE2E3
C O
4-HNE
4-HNE GSTO1
SOD1
CAT
PRDX1
44 H4
O
EEE
CCC
SDHB
NDUFA1UQCRQ
CYCS ATP5A1
LDHB
(2)Genes releted to
mitochondrial functions
(3)Genes releted to
ubiquitin-proteosome(2)
Gene expressionprofile deregulation
MeCP2
Sin3A
HDCA
CREB1
MeCMeCMeCMeCMeCeCMeCMeCMeCeCMeCeCeCCeCeCeCeCMeCeeCPPPP2PPPPPPPPPPP
MeCP2MeCMeCMeCMeCMeCeCeCM CMeCeCeCeCeCeCeCeCeCeCeCMeCMeCPPPP2PPPPPPPPPPPP
(4)Genes releted to
antioxidant defence
RTT PBMC
ROS
Co-activator
HIST1H1B(1)
Genes releted tochromatin folding
ST1H
Nucleus
Cytoplasm
Protein
Ubiquitin
Peptide fragments
Nucleosome
Scheme 1 Schematic summary related to the altered gene expression observed in RTT PBMC MeCP2 in a normal situation binds to severalcofactors (Sin3A CREB1 etc) to regulate gene transcription Mutation of MeCP2 (crossed boxes) will affect gene expression leading to agene profile deregulationThere are mainly 4 gene clusters significantly affected in RTT PBMC that is those related to chromatin folding (1)mitochondrial functions (2) ubiquitin-proteasome (3) and antioxidant defence (4) (green boxes) The overexpression of the genes involvedin ATP synthesis processes (2) can be interpreted as a possible energy requirement for an increment of cellular protein degradation (3)consequent to increasedmitochondrial ROS production and protein oxidationThe increased expression of the ldquoantioxidant cellular defencerdquogenes (4) is the possible compensatory mechanism activated by the cells to quench ROS production and protein oxidation (red arrows)
affect the chromatin organization and subsequently alter theexpression of the corresponding target genes
In our case several genes related to histone expressionwere downregulated and this can dramatically affect geneexpression For instance histone H1 protein binds to linkerDNA between nucleosomes forming the macromolecularstructure known as the chromatin fiber Histones H1 arenecessary for the condensation of nucleosome chains intohigher-order structured fibers Acts also as a regulator ofindividual gene transcription through chromatin remodel-ing nucleosome spacing and DNA methylation We havedetected a down-regulation of several H1 subunits rangingfrom 2- to 3-fold in RTT patients
36 Validation of Selected PBMC mRNAs by qPCR Analy-ses Next we wanted to confirm the differential expression
observed for selected mRNAs on an individual basis byRT-qPCR Six genes were selected based on their differentpatterns of expression (3 up- and 3 downregulated) Assess-ment of theirmRNAexpression levels byRT-qPCRaccuratelyreflected those obtained by microarray profiling (Figure 2)thereby confirming the validity of ourmicroarray resultsThelevels of threemRNAs encoding for glutathione S-transferaseomega 1 (GSTO1) proteasome (prosome macropain) sub-unit beta type-10 (PSMB10) and cytochrome c oxidasesubunit VIIIA (COX8A) were upregulated in RTT patients by25- 2- and 22-fold respectively (Figure 2) very similarly tothe levels measured by gene array In contrast histone cluster1 H1b (HIST1H1B) matrix metallopeptidase 9 (MMP9)and Rho GTPase activating protein 11B (ARHGAP11B) weredownregulated by sim50 in RTT patients as compared tohealthy subjects (Figure 2) and also in this case similarexpression was detected by gene array analysis
16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

16 Mediators of Inflammation
37 Conclusion Our microarray data reveal an alteredgene expression profile in RTT lymphomonocytes with theupregulation of genes related to mitochondrial biology andubiquitin-proteasome proteolytic pathway In particular theoverexpression of the genes involved in ATP synthesis pro-cesses means the tendency of cells to show an altered energyrequirement perhaps to cope with the increased activitiesof protein degradation On the other hand it should benoted that mitochondrion plays essential roles in mediatingthe production of ROS and these in turn cause damageto proteins as well as lipids and nucleic acids To removedamaged molecules in a kind of vicious circle increasedcellular proteolytic activity requires an extra mitochondrialATP production with a further ROS generation This pictureis consistent with our previous reports [24] indicating thealteration of redox status in RTT patients coupled withthe increased ubiquitination and degradation of oxidativelymodified proteins (Scheme 1)
In conclusion these findings on transcriptional pro-filing in RTT patients reveal new molecular mechanismsunderlying RTT phenotype suggesting that mitochondrial-ATP-proteosome are likely to have direct actions on redoxbalance in RTT syndrome Furthermore it confirmed apossible indirect role of OS in pathogenesis and progressionof disorder Thus RTT should be considered as possiblemitochondriopathy
Acknowledgments
The authors thank ldquoFondazione Monte Dei Paschirdquo Associ-azione Italiana Rett (AIRETT) and Tuscany Region (BandoSalute 2009 ldquoAntioxidants (120596-3 polyunsaturated fatty acidsLipoic Acid) Supplementation in Rett Syndrome a novelapproach to therapyrdquo) for partial support
References
[1] A Rett ldquoOn a unusual brain atrophy syndrome in hyperam-monemia in childhoodrdquo Wiener Medizinische Wochenschriftvol 116 no 37 pp 723ndash726 1966
[2] B Hagberg ldquoRettrsquos syndrome prevalence and impact on pro-gressive severe mental retardation in girlsrdquo Acta PaediatricaScandinavica vol 74 no 3 pp 405ndash408 1985
[3] C L Laurvick N de Klerk C Bower et al ldquoRett syndrome inAustralia a review of the epidemiologyrdquo Journal of Pediatricsvol 148 no 3 pp 347ndash352 2006
[4] J L Neul W E Kaufmann D G Glaze et al ldquoRett syndromerevised diagnostic criteria and nomenclaturerdquoAnnals of Neurol-ogy vol 68 no 6 pp 944ndash950 2010
[5] J L Neul ldquoThe relationship of Rett syndrome and MECP2disorders to autismrdquo Dialogues in Clinical Neuroscience vol 14no 3 pp 253ndash262 2012
[6] A Bebbington A Anderson D Ravine et al ldquoInvestigatinggenotype-phenotype relationships in Rett syndrome using aninternational data setrdquo Neurology vol 70 no 11 pp 868ndash8752008
[7] J L Neul P Fang J Barrish et al ldquoSpecific mutations inMethyl-CpG-Binding Protein 2 confer different severity in Rettsyndromerdquo Neurology vol 70 no 16 pp 1313ndash1321 2008
[8] R E Amir I B Van Den Veyver M Wan C Q TranU Francke and H Y Zoghbi ldquoRett syndrome is caused bymutations in X-linked MECP2 encoding methyl-CpG-bindingprotein 2rdquo Nature Genetics vol 23 no 2 pp 185ndash188 1999
[9] M Chahrour and H Y Zoghbi ldquoThe story of Rett syndromefrom clinic to neurobiologyrdquoNeuron vol 56 no 3 pp 422ndash4372007
[10] N L Adkins and P T Georgel ldquoMeCP2 structure and func-tionrdquo Biochemistry and Cell Biology vol 89 no 1 pp 1ndash11 2011
[11] J Guy H Cheval J Selfridge and A Bird ldquoThe role of MeCP2in the brainrdquo Annual Review of Cell and Developmental Biologyvol 27 pp 631ndash652 2011
[12] C Colantuoni O-H Jeon K Hyder et al ldquoGene expressionprofiling in postmortem Rett Syndrome brain differential geneexpression and patient classificationrdquo Neurobiology of Diseasevol 8 no 5 pp 847ndash865 2001
[13] J Nectoux Y Fichou H Rosas-Vargas et al ldquoCell cloning-based transcriptome analysis in Rett patients relevance to thepathogenesis of Rett syndrome of new human MeCP2 targetgenesrdquo Journal of Cellular andMolecular Medicine vol 14 no 7pp 1962ndash1974 2010
[14] J Traynor P Agarwal L Lazzeroni and U Francke ldquoGeneexpression patterns vary in clonal cell cultures from Rettsyndrome femaleswith eight differentMECP2mutationsrdquoBMCMedical Genetics vol 3 article 12 2002
[15] E Ballestar S Ropero M Alaminos et al ldquoThe impact ofMECP2 mutations in the expression patterns of Rett syndromepatientsrdquo Human Genetics vol 116 no 1-2 pp 91ndash104 2005
[16] I J Delgado D S Kim K N Thatcher J M LaSalle and I BVan den Veyver ldquoExpression profiling of clonal lymphocyte cellcultures from Rett syndrome patientsrdquo BMC Medical Geneticsvol 7 article 61 2006
[17] S Yakabe H Soejima H Yatsuki et al ldquoMeCP2 knockdownreveals DNA methylation-independent gene repression of tar-get genes in living cells and a bias in the cellular location oftarget gene productsrdquo Genes and Genetic Systems vol 83 no2 pp 199ndash208 2008
[18] D H Yasui H Xu K W Dunaway J M Lasalle L W Jin andI Maezawa ldquoMeCP2 modulates gene expression pathways inastrocytesrdquoMolecular Autism vol 4 no 1 p 3 2013
[19] G Dennis Jr B T Sherman D A Hosack et al ldquoDAVIDdatabase for annotation visualization and integrated discov-eryrdquo Genome Biology vol 4 no 5 p P3 2003
[20] Y Benjamini and Y Hochberg ldquoControlling the false discoveryrate a practical and powerful approach to multiple testingrdquoJournal of the Royal Statistical Society vol 57 no 1 pp 289ndash3001995
[21] F Cervellati G Valacchi L Lunghi et al ldquo17-120573-estradiolcounteracts the effects of high frequency electromagnetic fieldson trophoblastic connexins and integrinsrdquo Oxidative Medicineand Cellular Longevity vol 2013 Article ID 280850 11 pages2013
[22] C De Felice L Ciccoli S Leoncini et al ldquoSystemic oxidativestress in classic Rett syndromerdquo Free Radical Biology andMedicine vol 47 no 4 pp 440ndash448 2009
[23] A Pecorelli L Ciccoli C Signorini et al ldquoIncreased levelsof 4HNE-protein plasma adducts in Rett syndromerdquo ClinicalBiochemistry vol 44 no 5-6 pp 368ndash371 2011
[24] C Sticozzi G Belmonte and A Pecorelli ldquoScavenger receptorB1 post-translational modifications in Rett syndromerdquo FEBSLetters vol 587 no 14 pp 2199ndash2204 2013
Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

Mediators of Inflammation 17
[25] L Iommarini M A Calvaruso I Kurelac G Gasparre and AM Porcelli ldquoComplex I impairment in mitochondrial diseasesand cancer parallel roads leading to different outcomesrdquo TheInternational Journal of Biochemistry amp Cell Biology vol 45 no1 pp 47ndash63 2013
[26] R Stefanatos and A Sanz ldquoMitochondrial complex I a centralregulator of the aging processrdquo Cell Cycle vol 10 no 10 pp1528ndash1532 2011
[27] D N Hauser and T G Hastings ldquoMitochondrial dysfunctionand oxidative stress in Parkinsonrsquos disease and monogenicparkinsonismrdquoNeurobiology of Disease vol 51 pp 35ndash42 2013
[28] U N Das ldquoAutism as a disorder of deficiency of brain-derivedneurotrophic factor and altered metabolism of polyunsaturatedfatty acidsrdquo Nutrition vol 29 no 10 pp 1175ndash1185 2013
[29] T Clark-Taylor and B E Clark-Taylor ldquoIs autism a disorder offatty acid metabolism Possible dysfunction of mitochondrial120573-oxidation by long chain acyl-CoA dehydrogenaserdquo MedicalHypotheses vol 62 no 6 pp 970ndash975 2004
[30] S Kriaucionis A Paterson J Curtis J Guy N MacLeodand A Bird ldquoGene expression analysis exposes mitochondrialabnormalities in a mouse model of Rett syndromerdquo Molecularand Cellular Biology vol 26 no 13 pp 5033ndash5042 2006
[31] C Battisti P Formichi S A Tripodi et al ldquoLymphoblastoidcell lines of Rett syndrome patients exposed to oxidative-stress-induced apoptosisrdquo Brain and Development vol 26 no 6 pp384ndash388 2004
[32] M Anvret Z P Zhang and B Hagberg ldquoRett syndromethe bcl-2 genemdasha mediator of neurotrophic mechanismsrdquoNeuropediatrics vol 25 no 6 pp 323ndash324 1994
[33] M Sanchez-Arago L Formentini I Martınez-Reyes et alldquoExpression regulation and clinical relevance of the ATPaseinhibitory factor 1 in human cancersrdquoOncogenesis vol 2 articlee46 2013
[34] V Saywell A Viola S Confort-Gouny Y Le Fur L Villardand P J Cozzone ldquoBrain magnetic resonance study of Mecp2deletion effects on anatomy and metabolismrdquo Biochemical andBiophysical Research Communications vol 340 no 3 pp 776ndash783 2006
[35] H A Heilstedt M D Shahbazian and B Lee ldquoInfantile hypo-tonia as a presentation of Rett syndromerdquo American Journal ofMedical Genetics vol 111 no 3 pp 238ndash242 2002
[36] A Ruch T W Kurczynski and M E Velasco ldquoMitochondrialalterations in Rett syndromerdquo Pediatric Neurology vol 5 no 5pp 320ndash323 1989
[37] M T Dotti L Manneschi A Malandrini N De Stefano FCaznerale and A Federico ldquoMitochondrial dysfunction in RettsyndromeAnultrastructural and biochemical studyrdquoBrain andDevelopment vol 15 no 2 pp 103ndash106 1993
[38] S B Coker and A R Melnyk ldquoRett syndrome and mitochon-drial enzyme deficienciesrdquo Journal of ChildNeurology vol 6 no2 pp 164ndash166 1991
[39] T Matsuishi F Urabe A K Percy et al ldquoAbnormal carbo-hydrate metabolism in cerebrospinal fluid in Rett syndromerdquoJournal of Child Neurology vol 9 no 1 pp 26ndash30 1994
[40] R H Haas M Light M Rice and B A Barshop ldquoOxidativemetabolism in Rett syndromemdash1 Clinical studiesrdquo Neuropedi-atrics vol 26 no 2 pp 90ndash94 1995
[41] AAAl-JarallahMAM SalihMNAlNasser F AAl Zamiland J Al Gethmi ldquoRett syndrome in Saudi Arabia report of sixpatientsrdquo Annals of Tropical Paediatrics vol 16 no 4 pp 347ndash352 1996
[42] D D Armstrong ldquoNeuropathology of Rett syndromerdquo MentalRetardation and Developmental Disabilities Research Reviewsvol 8 no 2 pp 72ndash76 2002
[43] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoAbnormal mitochondria in the Rett syndromerdquo Brainand Development vol 10 no 4 pp 260ndash262 1988
[44] O Eeg-Olofsson A G H Al-Zuhair A S Teebi and M M NAl-Essa ldquoRett syndrome genetic clues based on mitochondrialchanges in musclerdquo American Journal of Medical Genetics vol32 no 1 pp 142ndash144 1989
[45] O Eeg-Olofsson A G H Al-Zuhair A S Teebi et alldquoRett syndrome a mitochondrial diseaserdquo Journal of ChildNeurology vol 5 no 3 pp 210ndash214 1990
[46] S Wakai K Kameda Y Ishikawa et al ldquoRett syndrome find-ings suggesting axonopathy and mitochondrial abnormalitiesrdquoPediatric Neurology vol 6 no 5 pp 339ndash343 1990
[47] S-C Mak C-S Chi C-H Chen and W-J Shian ldquoAbnormalmitochondria in Rett syndrome one case reportrdquo ChineseMedical Journal vol 52 no 2 pp 116ndash119 1993
[48] M E Cornford M Philippart B Jacobs A B Scheibel and HV Vinters ldquoNeuropathology of Rett syndrome case report withneuronal andmitochondrial abnormalities in the brainrdquo Journalof Child Neurology vol 9 no 4 pp 424ndash431 1994
[49] S Naidu S Hyman E L Harris V Narayanan D Johns andF Castora ldquoRett syndrome studies of natural history and searchfor a genetic markerrdquo Neuropediatrics vol 26 no 2 pp 63ndash661995
[50] J Tang Y Qi X-H Bao and X-R Wu ldquoMutational analysis ofmitochondrial DNA of children with Rett syndromerdquo PediatricNeurology vol 17 no 4 pp 327ndash330 1997
[51] C Ellaway and J Christodoulou ldquoRett syndrome clinicalupdate and review of recent genetic advancesrdquo Journal ofPaediatrics and Child Health vol 35 no 5 pp 419ndash426 1999
[52] Y Oi X Wu J Tang and X Bao ldquoComputerized ribosomalRNA secondary structure modeling of mutants found in Rettsyndrome patients and their mothersrdquo Chinese Journal ofMedical Genetics vol 16 no 3 pp 153ndash155 1999
[53] J Armstrong M Pineda and E Monros ldquoMutation analysis of16S rRNA in patients with rett syndromerdquo Pediatric Neurologyvol 23 no 1 pp 85ndash87 2000
[54] H Meng H Pan and Y Qi ldquoRole of mitochondrial lesion inpathogenesis of sporadic rett syndromerdquo Zhonghua yi xue zazhi vol 81 no 11 pp 662ndash664 2001
[55] J H Gibson B Slobedman H KN et al ldquoDownstream targetsofmethyl CpG binding protein 2 and their abnormal expressionin the frontal cortex of the human Rett syndrome brainrdquo BMCNeuroscience vol 11 article 53 2010
[56] MD Brand ldquoThe sites and topology ofmitochondrial superox-ide productionrdquo Experimental Gerontology vol 45 no 7-8 pp466ndash472 2010
[57] S Drose and U Brandt ldquoMolecular mechanisms of superoxideproduction by the mitochondrial respiratory chainrdquo Advancesin Experimental Medicine and Biology vol 748 pp 145ndash1692012
[58] S Ahmad D Niegowski A Wetterholm J Z HaeggstromR Morgenstern and A Rinaldo-Matthis ldquoCatalytic charac-terization of human microsomal glutathione s-transferase 2identification of rate-limiting stepsrdquo Biochemistry vol 52 no10 pp 1755ndash1764 2013
[59] G Poli R J Schaur W G Siems and G Leonarduzzildquo4-Hydroxynonenal a membrane lipid oxidation product of
18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001

18 Mediators of Inflammation
medicinal interestrdquo Medicinal Research Reviews vol 28 no 4pp 569ndash631 2008
[60] K Makedonski L Abuhatzira Y Kaufman A Razin and RShemer ldquoMeCP2 deficiency in Rett syndrome causes epige-netic aberrations at the PWSAS imprinting center that affectsUBE3A expressionrdquo Human Molecular Genetics vol 14 no 8pp 1049ndash1058 2005
[61] A Lawson-Yuen D Liu L Han et al ldquoUbe3a mRNA andprotein expression are not decreased in Mecp2R168X mutantmicerdquo Brain Research vol 1180 no 1 pp 1ndash6 2007
[62] G P Delcuve M Rastegar and J R Davie ldquoEpigenetic controlrdquoJournal of Cellular Physiology vol 219 no 2 pp 243ndash250 2009
[63] J C Rice and C D Allis ldquoHistone methylation versus histoneacetylation new insights into epigenetic regulationrdquo CurrentOpinion in Cell Biology vol 13 no 3 pp 263ndash273 2001




















