
Gamma Interferon Positively Modulates Actinobacillus actinomycetemcomitans-Specific RANKL+ CD4+...
10
10.1128/IAI.73.6.3453-3461.2005. 2005, 73(6):3453. DOI: Infect. Immun. Yen-Tung A. Teng, Deeqa Mahamed and Bhagirath Singh Alveolar Bone Destruction In Vivo Th-Cell-Mediated + CD4 + -Specific RANKL Actinobacillus actinomycetemcomitans Gamma Interferon Positively Modulates http://iai.asm.org/content/73/6/3453 Updated information and services can be found at: These include: REFERENCES http://iai.asm.org/content/73/6/3453#ref-list-1 at: This article cites 51 articles, 16 of which can be accessed free CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on November 11, 2014 by guest http://iai.asm.org/ Downloaded from on November 11, 2014 by guest http://iai.asm.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Gamma Interferon Positively Modulates Actinobacillus actinomycetemcomitans-Specific RANKL+ CD4+...

10.1128/IAI.73.6.3453-3461.2005.
2005, 73(6):3453. DOI:Infect. Immun. Yen-Tung A. Teng, Deeqa Mahamed and Bhagirath Singh Alveolar Bone Destruction In Vivo
Th-Cell-Mediated+ CD4+-Specific RANKLActinobacillus actinomycetemcomitansGamma Interferon Positively Modulates
http://iai.asm.org/content/73/6/3453Updated information and services can be found at:
These include:
REFERENCEShttp://iai.asm.org/content/73/6/3453#ref-list-1at:
This article cites 51 articles, 16 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
on N
ovember 11, 2014 by guest
http://iai.asm.org/
Dow
nloaded from

INFECTION AND IMMUNITY, June 2005, p. 3453–3461 Vol. 73, No. 60019-9567/05/$08.00�0 doi:10.1128/IAI.73.6.3453–3461.2005Copyright © 2005, American Society for Microbiology. All Rights Reserved.
Gamma Interferon Positively Modulates Actinobacillusactinomycetemcomitans-Specific RANKL� CD4� Th-Cell-Mediated
Alveolar Bone Destruction In VivoYen-Tung A. Teng,1* Deeqa Mahamed,1 and Bhagirath Singh2
Division of Periodontics, Eastman Department of Dentistry, and Centre for Oral Biology, Department of Microbiology &Immunology, School of Medicine and Dentistry, the University of Rochester, Rochester, New York, 14620,1 and
Department of Microbiology & Immunology, School of Medicine and Dentistry, the University ofWestern Ontario, London, Ontario, N6A 5C1, Canada2
Received 5 November 2004/Returned for modification 27 December 2004/Accepted 26 January 2005
Recent studies have shown the biological and clinical significance of signaling pathways of osteogeniccytokines RANKL-RANK/OPG in controlling osteoclastogenesis associated with bone pathologies, includingrheumatoid arthritis, osteoporosis, and other osteolytic disorders. In contrast to the inhibitory effect of gammainterferon (IFN-�) on RANKL-mediated osteoclastogenesis reported recently, alternative new evidence isdemonstrated via studies of experimental periodontitis using humanized NOD/SCID and diabetic NOD miceand clinical human T-cell isolates from diseased periodontal tissues, where the presence of increasing IFN-�is clearly associated with (i) enhanced Actinobacillus actinomycetemcomitans-specific RANKL-expressing CD4�
Th cell-mediated alveolar bone loss during the progression of periodontal disease and (ii) a concomitant andsignificantly increased coexpression of IFN-� in RANKL(�) CD4� Th cells. Therefore, there are more complexnetworks in regulating RANKL-RANK/OPG signaling pathways for osteoclastogenesis in vivo than have beensuggested to date.
Human periodontal disease (i.e., periodontitis) results from theinterplay between the specific subgingival microorganisms andthe host’s immune and inflammatory response (9). Periodontitis ischaracterized by loss of attachments including the periodontalconnective tissues and supporting alveolar bone. Actinobacillusactinomycetemcomitans (A. actinomycetemcomitans) is a G(�)facultative anaerobe strongly associated with human aggressiveperiodontitis (AgP) and also involved in medical diseases such asurinary tract and hip-joint prosthetics infections, thyroid andbrain abscesses, and subacute endocarditis (54). Although severalcarbohydrate and protein antigens (Ags), fimbriae, and polysac-charides were identified, much less is known about critical Ags ofA. actinomycetemcomitans that sensitize T or B cells involved inthe periodontal pathogenesis (16).
The recently identified tumor necrosis factor (TNF) familymolecule, receptor activator of NF-�B ligand (RANKL [orTRANCE, OPGL, or ODF]), its receptor RANK, and thenatural antagonist, osteoprotegerin (OPG), have been shownto be the key regulators of bone remodeling and are directlyinvolved in the differentiation, activation, and survival of oste-oclasts (OC) and OC precursors (27, 40, 52–53). Previously, weshowed that activated CD4� T cells express RANKL, whichcan directly trigger osteoclastogenesis and alveolar bone lossassociated with periodontitis in vivo (46). Further, blockingRANKL activity via OPG injections results in significantly re-duced bone loss in arthritis (25, 49), periodontitis (46, 50),
osteoporosis (19, 30), cancer-related bone metastasis (5, 20),and enhanced alveolar bone loss associated with type 1 diabe-tes in vivo (29). RANKL-RANK signaling is also critical forsurvival of dendritic cells, lymph node formation, and organo-genesis and involved in dendritic cell/T-cell interactions (1, 26).Genetic mutations of RANKL and RANK demonstrate similarphenotypes in OC development with severe osteopetrosis, sug-gesting that they are essential for osteoclastogenesis duringbone remodeling (26, 28, 50). OPG-transgenic mice are osteo-petrotic with defective OC activity, and OPG-deficient miceare severely osteoporotic (30, 40, 50). Thus, RANKL, RANK,and OPG are essential for controlling OC development andfunctions in bone remodeling. These studies have supportedthe new paradigm of linking adaptive immunity and bone re-modeling (termed osteo-immunology) associated with variousinflammatory bone disorders (1, 5, 19–20, 25–26, 28–30, 40, 46,50, 53). More recently, studies have shown that periodontalresiden T cells and tissues (i.e., periodontal ligament and fi-broblast tissues) can also be induced to express RANKL/OPGunder microbe- or microbial product-induced inflammatoryconditions in vivo or/and in vitro (15, 32–33, 37), suggesting thebroad contributions of cytokine RANKL-RANK/OPG signal-ing network in periodontal disease.
We have established a humanized mouse model to studyhuman immune-parasite interactions in periodontitis in vivo byusing immunodeficient NOD/SCID mice (called: HuPBL-NOD/SCID mice) where microbial A. actinomycetemcomitans-specific human CD4� T cells are functionally active and theirTCR immune repertoire overlaps significantly (83 to 91%)with those of human clinical isolates from active disease sites(14, 44, 46, 47). Interestingly, these microorganism-specificRANKL�CD4� T cells manifest a mixed Th1-Th2 cytokine
* Corresponding author. Mailing address: Lab. of Molecular Micro-bial Immunity, Eastman Dental Center, University of Rochester Med-ical Centre, Box 683, 625 Elmwood Ave., Rochester, NY 14620.Phone: (585) 275-7309. Fax: (585) 473-5254. E-mail: [email protected].
3453
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

profile in active periodontal lesions, in which specific cytokinessuch as gamma interferon (IFN-�) and interleukin-10 (IL-10)are significantly coexpressed with RANKL during osteoclasto-genesis in vivo (44). It has been shown that both TNF-� and/orIL-1 can work synergistically or independently with RANKL tomodulate bone resorption in arthritis and osteoporotic disor-ders in vitro and in vivo (2, 34, 18, 38, 41, 45, 50). Despitestudies that have suggested inhibitory effect of IFN-� onRANKL-associated osteoclastogenesis and bone remodelingin vitro and in vivo (11, 13, 42), it has also been shown thatIFN-�� Th1 cells are strongly associated with enhanced alve-olar bone loss during periodontal infections (3, 29, 22, 43, 51)and that RANKL is often highly coexpressed in Th1 cells (7,21). Furthermore, there is strong evidence suggesting that thisRANKL and IFN-� coexpression exists in active arthritic le-sions in vivo (4, 6, 35–36, 39) and that deficient IFN-� expres-sion significantly reduces the severity of periodontal bone lossin mice after mounting a microbial challenge (3).
To date, it is not clear what the exact role and contributionof IFN-� cytokines are in terms of modulating RANKL ex-pression and associated osteoclastogenesis under the inflam-matory periodontal lesions “in vivo.” We thus hypothesizedthat a Th1 cytokine, IFN-�, can positively modulate RANKL-mediated alveolar bone loss under inflammatory conditions invivo. This issue was addressed by several approaches: (i) de-tecting early cytokine expression profile and injecting low-dose hIFN-� into A. actinomycetemcomitans-infected HuPBL-NOD/SCID mice followed by monitoring the coexpression ofIFN-� and RANKL and alveolar bone loss over time in vivo,(ii) analyzing human clinical T-cell isolates extracted from thediseased periodontal tissues of AgP subjects, and (iii) assessingcytokine expression profile in diabetic NOD mice with en-hanced alveolar bone loss in vivo. The results show that, incontrast to what has been suggested in recent studies (11, 13,42), IFN-� can indeed positively modulate its coexpressionwith RANKL in periodontal microorganism-reactive CD4� Thcells, which can further mediate osteoclastogenesis associatedwith alveolar bone loss in vivo.
MATERIALS AND METHODS
Human subjects, CD4� T-cell purification, tissue cultures and reagents. Fourconsenting AgP subjects (two female and two male; mean age � 21 � 4) whoseclinical diagnosis were confirmed based on clinical, X-ray, and microbiologicalcriteria described previously (14, 44, 45) and two consenting age-matched healthypatients (N1 and N2) (one female and one male; age � 20 and 21) were recruitedfor the present study. The diseased periodontal tissues obtained from the surgicaldiscards of aggressive periodontitis (AgP) subjects undergoing periodontal sur-gical treatment, and periodontally healthy tissues from normal control subjectsundergoing prescribed periodontal surgeries for different reasons, were col-lected. These procedures were all approved by the human ethics committees ofthe University of Western Ontario, London, Ontario, Canada, and the Universityof Rochester, Rochester, N.Y. Later, hCD4� T cells were enriched and purifiedfrom the above-described surgical discards by collagenase treatment and in vitrodirect panning as described previously (14, 44, 46, 47). Meanwhile, to generatemCD4� T cells, a splenic single-cell suspension was prepared after lysing redblood cells. Then, total splenocytes were passed through a nylon-wool column toenrich T cells, after which mCD4� T cells were further purified via directpanning on an anti-mCD4 GK1.5 monoclonal antibody (MAb)-coated petri dish(44, 46). The purity of both kinds of CD4� T cells was 95 to 97% (by fluores-cence-activated cell sorter [FACS]) (14, 44, 46, 47).
All primary T-cell cultures in the present study were performed in completeRPMI 1640 medium supplemented with 10% heat inactivated fetal bovine serum(GIBCO, Ontario, Canada), 50 �M 2-mercaptoethanol, 100 �g/ml streptomy-cin, and 100 U/ml penicillin, and cells were incubated at 37°C in a humidified 5%
CO2 incubator as described previously (46). The following reagents and MAbswere purchased from commercial sources: recombinant hIFN-� cytokine, anti-mFc-R (CD16/32), anti-mCD4, phosphatidylethanolamine (PE)-conjugatedanti-h/m CD4, fluorescein isothiocyanate (FITC)- or antigen-presenting cell(APC)-conjugated goat anti-hFc-� and anti-h/m IL-4, PE-, or PerCP-Cy5.5-conjugated anti-h/m IFN-� (BD Pharmingen, Toronto, Canada), goat anti-rabbitbiotinylated immunoglobulin G (IgG), and streptavidin-PE molecules (VectorLaboratories, Calif.), and recombinant OPG-hu-Fc and OPG-FITC fusion pro-teins were kindly provided by J. M. Penninger (Austria) (25, 44).
Mice and A. actinomycetemcomitans oral inoculation in vivo. Female BALB/c(H-2d) and NOD/LtJ mice (H-2g7 � Kd, Aad, Abg7, Enull, Db) (4 to 6 weeks old)were purchased from Jax Mice (Bar Harbor, Maine), and the NOD/SCID micewere generated and maintained in our own mouse-breeding suites at both Uni-versities. All mice were housed under specific-pathogen-free environmental con-ditions in the Animal Care Facility of the University of Rochester, Rochester,New York, and the University of Western Ontario, Ontario, Canada. All animalprotocols were conducted based on each institution’s guidelines and approved bythe Animal Experimentation and Use Committees of the universities.
In addition, NOD mice studied here were monitored for type 1 diabetes bydaily monitoring of urine ketone and glucose levels by Diastix strips and aGlucometer Elite XL meter (Bayer, Ontario, Canada) (29). Mice were consid-ered diabetic when whole-blood glucose levels exceeded 200 mg/dl on 2 to 3consecutive days, with the histological evidence of severe lymphocytic infiltrationin pancreatic -islets (insulitis). Typically, 70 to 80% female NOD mice devel-oped diabetes by 16 to 20 weeks (29). Keto-Diastix strips (Bayer, Ontario,Canada) were used for daily monitoring and diagnosis. Diabetic mice weretreated for hyperglycemia with Humulin U insulin (Eli Lilly, Indianapolis) (1 to4 units/day) to maintain urine ketones (0 to 0.5 unit) and glucose at 2 units onthe Keto-Diastix strips. All mice were categorized based on their age and thestate of diabetes with random pancreatic histology as follows: mice 6 to 8 weekswere considered prediabetic with peri-insulitis, mice �12 to 16 week with bloodglucose levels 140 mg/dl that did not develop diabetes were considered non-diabetic, and mice more than 16 to 20 weeks old with blood glucose levels �200mg/dl and insulitis were considered diabetic (29).
The A. actinomycetemcomitans JP2 (ATCC-29523) strain was originally pur-chased from the American Type Culture Collection (Rockville, MD) and grownanaerobically (80% N2, 10% H2, 10% CO2) in tryptic soy broth-yeast extractculture broth (Sigma Chemical, St. Louis, Missouri) supplemented with 0.75%glucose and 0.4% NaHCO3. Experimental mice, including BALB/c, NOD/SCID,and NOD strains, were orally inoculated with freshly prepared A. actinomyce-temcomitans (109 CFU/100 �l broth mixed with 2% carboxy-methylcellulose inphosphate-buffered saline [PBS]) twice per week for three consecutive weeks asdescribed previously (46). The mice were then sacrificed at different time pointsbetween week 3 and week 7 to 8. Age-matched NOD mice without bacterialinoculation served as controls and were kept under the same specific-pathogen-free environmental conditions. For the immunization study, 6-week-old BALB/cmice were immunized intraperitoneally (i.p.) with 20 �g A. actinomycetemcomi-tans-CagE-homologue Ag (48–49) in colonization factor antigen (CFA) (1:1ratio) or with CFA alone or were sham immunized 2 weeks before the start of A.actinomycetemcomitans oral inoculation as described above and elsewhere (14,44, 46, 47). All mice were then sacrificed at the end of week 7.
Quantitative reverse transcriptase PCR (RT-PCR) and FACS analysis. Thelevels of HuPBL engraftment into NOD/SCID mice obtained in the presentstudy were comparable (�30%) to those reported in our previous studies (46,47). The generation of different sources of tissues, cells, and cDNA samples usedin this study were described previously (14, 44, 47). Briefly, at different timepoints of the experiments, periodontal or/and cervical lymph node (CLN) CD4�
T cells of the mice were purified by positive panning (�95 to 97%) and thensubjected to in vitro stimulation with autologously irradiated human monocytes/macrophages (Mo/MQ) or mouse total splenocytes and A. actinomycetemcomi-tans sonicate Ags (JP2 strain) in 24-well plates (14, 44, 47). After 2, 3, or 4 daysin cultures, 2 � 106 to 3 � 106 A. actinomycetemcomitans-reactive CD4� T cellswere collected by Ficoll-gradient centrifugation, followed by fluorescence-acti-vated cell scanning and sorting (FACSVantage, BD Biosciences) for RANKL(�)CD4� T cells by using OPG-hu-Fc plus 20-conjugate MAb (purity �95 to 97%;CD25� CD69�) (14, 44, 46). Further purification of the CD4� T cells studiedhere by sorting CD69/CD25-positive cells followed by labeling with CFSE (car-boxyfluorescein diacetate succinimidyl ester), a FITC-based dye, for a 3-dayrestimulation assay in the presence of 25 Gy-irradiated human Mo/MQ or mousesplenocytes and A. actinomycetemcomitans-Ags rendered �85 to 90% CFSE-positive T cells in active division, indicating their specific reactivity to A. actino-mycetemcomitans Ags (references 14, 29, 44, and 46 and data not shown).
To quantify the cytokine expression levels, total RNA was prepared from an
3454 TENG ET AL. INFECT. IMMUN.
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

individual pool of 1 � 106 A. actinomycetemcomitans-reactive RANKL(�) peri-odontal CD4� Th cells derived from the above-described cultures representingindividual groups of A. actinomycetemcomitans-HuPBL-NOD/SCID mice forquantitative RT-PCR analysis (Q-PCR) (14, 44). The first-strand cDNA wasprepared from 2 �g of total RNA and then precipitated and diluted in 40 �l ofTris-EDTA (14, 44). cDNA from each sample (2 �l) was aliquoted into PCRtubes containing specific forward and reverse primers of hIFN-� and hIL-4 genesas described previously (10, 44). The amplified h-actin gene products were usedas the internal control for all PCRs and subsequent quantitation. Based on ourprevious studies (14, 44), all RT-PCRs were carried out for 30 cycles under thefollowing conditions: 94°C denaturation for 1 min, 60°C annealing for 1 min, and72°C extension for 1.5 min (RoboCycler 96 Gradient; Stratagene, CA) with anadditional 7 min at 72°C after the last cycle. The resulting PCR products wereanalyzed by electrophoresis in 2% agarose gels for their respective sizes. AllPCRs and subsequent quantification were performed at least two to three timesfrom the same cDNA sources to ensure consistent and reproducible measure-ments. Fluorescent intensities of the amplified PCR products were captured bya UVP digital camera and quantified via image acquisition-and-analysis software(v3.0.2; LabWorks, Upland, CA). The resulting signal intensities were thennormalized to the mean values of the internal control h-actin set as 1. Anotherendogenous control, hTCR-C� primers (14, 44), was included to compare thetotal amounts of transcripts derived from each of 1 � 106 A. actinomycetem-comitans-reactive RANKL(�) CD4� T cells as described above for Q-PCRanalyses. To this end, repeated PCR analyses of individually amplified cytokinetranscripts performed two to three times did not change the results obtained,indicating high reproducibility.
In parallel, 1 � 106 to 2 � 106 purified A. actinomycetemcomitans-reactiveperiodontal CD4� T cells derived in vitro as described above were harvested andsubjected to immunostaining for their cell surface expression of RANKL (OPG-Fc-FITC or APC labeled) (44, 46) and intracellular staining after being fixed by4% formaldehyde and permeabilized with 0.2% Triton X-100 followed by incu-bation with IgG-conjugate for the expression of IFN-� (PE-coupled or PerCP-Cy5.5: a True-Red fluorochrome conjugate for flow cytometry analysis with Ex at490 and 675 nm and Em at 695 nm) and IL-4 (FITC coupled) by FACS analysisdescribed previously (29, 44, 46). Briefly, to block nonspecific immunostaining,cells were first incubated with anti-FcR-IgG followed by incubation with OPG-hu-Fc to label RANKL molecules and PerCP-Cy5.5-conjugated rat anti-CD4-MAb to label CD4 molecules on T cells, respectively. Then, samples were washedtwice and incubated with goat anti-hFc-IgG-FITC 20-conjugate. The isotypiccontrol was incubated with goat anti-hFc-IgG-FITC. For FACS scanning, cellswere gated on live lymphocytes and analyzed for RANKL expression in CD4� Tcells using FACSCalibur and CellQuest software (BD Biosciences) (29).
Measurement of alveolar bone loss via a digital histomorphometry. Themouse jaw samples were defleshed and stained with methylene blue to define thearea between the cementum-enamel junction and the alveolar bone crest (44,46). The surface areas represent measurement of the total amount of alveolarbone loss on the jaws, in square micrometers, which was carried out with acalibrated Leica MZ95 stereo microscope and a Hamamatsu Orca digital camera(44, 46). The jaw images were captured under �16 magnification; the right andleft maxillary first two molars (i.e., M1 and M2) perpendicular to the optical lightsource were scanned and automatically enumerated by using the density-slicefeatures of the OpenLab for full quantitation (29). To ensure reproducible andaccurate images captured for quantification, independent jaw images were takenby two calibrated members (MA and XZ) whose results showed consistentmeasurements for the exposed surfaces between cementum-enamel junction-alveolar bone crest. The results of alveolar bone loss detected in the presentstudy were consistent with our previous findings using the immunocompetentBALB/c, diabetic NOD (29), and humanized NOD/SCID mice where activealveolar bone loss and/or significantly inflamed periodontal lesions did not occuruntil 6 to 8 weeks post-A. actinomycetemcomitans inoculation (14, 44, 46, 47).Thus, the resulting values for alveolar bone loss are expressed as the means ofthe surface areas � standard errors (S.E.) (in square micrometers) of M1 andM2 from each mouse in each group.
Statistical analysis. Statistical analysis was performed using the two-sidedStudent t test, and the difference between various groups was considered statis-tically significant when the P value was 0.05.
RESULTS
hIFN-�, not hIL-4, is detected in A. actinomycetemcomitans-reactive RAMKL� Th cells at the early stage (week 3), andincreasing hIFN-� is associated with enhanced alveolar bone
loss by week 7 post-microbial infection in vivo. We previouslyreported that RANKL and some Th1-Th2 cytokines can becoexpressed in A. actinomycetemcomitans-reactive CD4� Tcells during active alveolar bone loss in HuPBL-NOD/SCIDmice between week 6 and 8 post-microbial infection in vivo(44). Therefore, in order to establish an early temporal rela-tionship between RANKL and expression of key Th1-Th2 (i.e.,IFN-� and IL-4) cytokines prior to active alveolar bone de-struction in vivo, we studied their expression by quantitativePCR (Q-PCR) using diseased periodontal tissues of four dif-ferent groups of A. actinomycetemcomitans-infected HuPBL-NOD/SCID mice (10 to 12 mice per donor group) whoseautologous HuPBL samples were obtained from 4 AgP sub-jects (mean age, 21 � 4), individually. Interestingly, the resultsof Q-PCR showed that there were significantly higher expres-sion of hIFN-� than hIL-4 in A. actinomycetemcomitans-reac-tive RANKL(�) periodontal CD4� T cells at as early as week3 after restimulating these cells for 2 to 4 days in vitro (Fig.1A). Similar results were also detected at the end of week 7,based on the results of FACS analysis (Fig. 1B). These dataindicate (i) that IFN-� does coexpress with RANKL in micro-organism-reactive periodontal hCD4� Th cells in vivo, consis-tent with our previous study (44) and (ii) that increasing coex-pression of IFN-� in the microorganism-reactive RANKL(�)periodontal hCD4� Th cells is associated with increased alve-olar bone loss over time, as significantly higher alveolar boneloss occurred post-A. actinomycetemcomitans infection be-tween week 6 to 8 in the current human-mouse chimeras (44,46). To confirm the impact of increasing hIFN-� on A. actino-mycetemcomitans-specific RANKL(�) periodontal Th1 cellsand alveolar bone remodeling, a low-level dose of rh-IFN-�(200 ng/mouse) or PBS (sham control) was i.p. injected into A.actinomycetemcomitans-HuPBL-NOD/SCID mice (n � fiveper group) once a week for the first 3 weeks. These mice weresacrificed at various time points until week 7 to 8 for assessingthe coexpression of IFN-� and RANKL by Q-PCR and FACSand the mean alveolar bone loss via a digital imaging forhistomorphometry. The results of studying samples collectedat the end of week 7 clearly showed that increasing IFN-� inthe environment had led to significantly increased A. actino-mycetemcomitans-reactive RANKL(�) periodontal CD4� Thcells coexpressing IFN-� in vivo compared to those seen withPBS-injected sham-control A. actinomycetemcomitans-HuPBL-NOD/SCID mice (Fig. 2A and B). Meanwhile, injection ofrh-IFN-� also yielded significantly higher alveolar bone loss invivo compared to the sham-control results (P � 0.017; Fig. 2Cand D). These data confirm the above hypothesis that a specificTh1 cytokine, hIFN-�, can indeed positively modulateRANKL-medicated osteoclastogenesis in vivo where there isan associated increase of periodontal microorganism-reactiveRANKL(�) CD4� Th cells coexpressing hIFN-� in the dis-eased tissues.
Higher IFN-� expression is strongly associated with thediseased human periodontal tissues and significant RANKL-mediated alveolar bone loss in diabetic NOD mice in vivo.Since other studies have suggested an opposite effect of IFN-�on bone remodeling (11, 13, 42), we though it was necessary toseek further evidence by using different approaches or models.A. actinomycetemcomitans-reactive CD4� T cells collectedfrom periodontal tissues and CLN were purified, respectively
VOL. 73, 2005 IFN-� POSITIVELY REGULATES RANKL-MEDIATED BONE LOSS 3455
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

(14, 44, 46), from (i) surgical discard samples of three AgPsubjects who were diagnosed to have active A. actinomycetem-comitans infection (14, 44, 47) and (ii) diabetic NOD micewhich were orally infected with freshly prepared A. actinomy-cetemcomitans and monitored for up to 8 weeks for assessing“enhanced” alveolar bone loss after comparison to the predi-abetic and nondiabetic NOD mice (data not shown) (29). Fur-ther, it is worth mentioning that the “enhanced” alveolar bonedestruction observed and detected in “diabetic” NOD micepost-A. actinomycetemcomitans oral inoculation by week 8 issignificantly associated with the severity of autoimmune insu-lititis (i.e., lymphocytic infiltration in -islets) where signifi-cantly increased A. actinomycetemcomitans-specific CD4� Tcells proliferation and associated RANKL expression are noted(29). Since a NOD mouse is the analogue of human type 1diabetes, hence, “diabetic” NOD mice can serve as a goodmodel to study the molecular interactions between microbe-induced immune responses and exacerbated alveolar bone loss
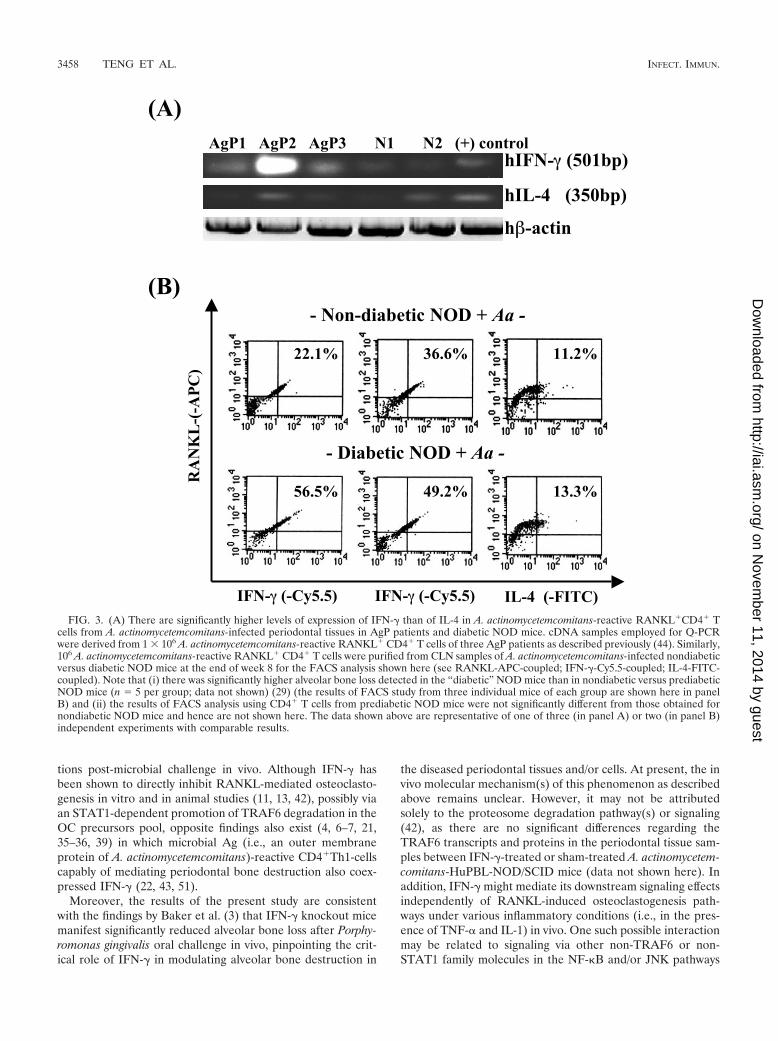
in vivo. As expected, there was significantly higher coexpres-sion of IFN-� in A. actinomycetemcomitans-reactive RANKL(�)periodontal CD4� Th cells in clinical human T-cell isolates(Fig. 3A) (Q-PCR) and in “diabetic” NOD mice where therewas concomitantly higher alveolar bone loss (for RANKL�
murine IFN-�� (mIFN-��) at 22 to 36% in “nondiabetic” and49 to 56% in “diabetic” NOD mice, see FACS in Fig. 3B) (29).In contrast, there were no significant differences regarding IL-4expression in RANKL� CD4� Th cells in both cases. Theseresults strongly suggest that coexpression of IFN-� in micro-organism-reactive RANKL(�) periodontal Th1 cells is signif-icantly associated with alveolar bone destruction in vivo, basedon all three different lines of evidence (Figs. 1 to 3).
Microbial Ag (CagE)-specific RANKL� IFN-�-expressingTh cells are strongly associated with increasing alveolar boneloss in vivo. Finally, to examine whether the above-describedevent of cytokine regulation in response to A. actinomycetem-comitans challenge can also be induced by specific microbial
hIFN-� (501bp)
hIL-4 (350bp)
h�-actin
day 2 day3 day4 (+)-control
(Aa-reactive CD4+T-cells re-stimulated)
I ) II) III)
Un-stained control hIL-4 (-FITC) hIFN-� (-PE)
(A)
(B)
<6% 41% 83%
FIG. 1. Increased coexpression of hIFN-� in A. actinomycetemcomitans-reactive RANKL(�) CD4� T cells before and during periodontaldisease progression in vivo. There were significantly increased levels of hIFN-� expression in A. actinomycetemcomitans-reactive RANKL� CD4�
T cells at both the third week (Q-PCR in panel A) and the seventh week (FACS analysis in panel B: hIL-4-FITC coupled and hIFN-�-PE coupled)of the periodontal lesions in four groups of A. actinomycetemcomitans-HuPBL-NOD/SCID mice (n � 10 to 12 per group) receiving autologousHuPBL samples from four AgP subjects. There was no IL-4 signal detected at the third week, based on Q-PCR. The actual IFN-� and IL-4 signalintensities detected (via LabWorks software) in the days 2, 3, and 4 in vitro stimulation were 0.06, 0.28, and 0.42, and �0.001, �0.001, and �0.001,respectively. The actual signals for the positive control were from 0.61 to 1.24 and for the -actin were from 0.93 to 1.35. Note that there were nosignificant differences between different groups of A. actinomycetemcomitans-HUPBL-NOD/SCID mice receiving autologous HuPBL samplesregarding the intensity of PCR amplicon signals detected (see Materials and Methods and reference 44). The results shown are for A.actinomycetemcomitans-reactive CD4� T cells restimulated in vitro for 2, 3, and 4 days before being subjected to Q-PCR, and concanavalinA-stimulated CD4� T cells were used as the positive control. The data shown are representative of one of three independent experiments withcomparable results.
3456 TENG ET AL. INFECT. IMMUN.
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

Ag in vivo, we employed an A. actinomycetemcomitans-associ-ated CagE homologue (in short, CagE) (48, 49) recognized byhuman CD4� T cells as a model Ag for testing via an immu-nization strategy. Based on our previous study, CagE is a crit-ical microbial virulence factor associated with human CD4�
T-cell-mediated destructive immunity for alveolar bone lossand induction of apoptosis of different human cell types in theperiodontal tissues (46, 48–49). Therefore, BALB/c mice werei.p. immunized with CagE Ag (group 3; n � 5 per group) priorto oral infection with A. actinomycetemcomitans (for 3 weeks),which resulted in a significantly enhanced alveolar bone loss byweek 7 to 8 in vivo (Fig. 4A and B). The controls included A.actinomycetemcomitans oral inoculation with sham-treatedmice (group 1) and CFA-immunized BALB/c mice (group 2;Fig. 4A and B). Meanwhile, the same protocol also resulted ina significantly increased coexpression of mIFN-� in A. actino-
mycetemcomitans-reactive RANKL(�) periodontal CD4� Thcells in CagE-immunized BALB/c mice by week 7 to 8, sug-gesting that modulation of RANKL-mediated osteoclastogen-esis by mIFN-� can be induced by the whole microorganismand a single microbial Ag (i.e., CagE) in vivo. Together, theabove data are in concordance with our hypothesis that IFN-�,a Th1 signature cytokine, can positively modulate RANKL-mediated osteoclastogenesis in A. actinomycetemcomitans-induced periodontal bone loss in human-mouse chimeras, clin-ical periodontal T-cell isolates of AgP patients, and diabeticNOD mice during periodontal disease progression in vivo.
DISCUSSIONThe present report clearly describes the positive role of
IFN-� (for human and mouse) in modulating RANKL� Thcell-mediated alveolar bone loss under inflammatory condi-
IFN-� (-PE)
IFN-� (-PE)
IFN-� (-PE)
RA
NK
L-
(FIT
C)
RA
NK
L-
(FIT
C)
RA
NK
L-
(FIT
C)
un-stained control
hIFN-�-treated Aa-
inoculated HuPBL-
NOD/SCID mice
Sham-treated Aa-
inoculated HuPBL-
NOD/SCID mice
50 % 21 %
5 %37%
<1 %
2 %
hIFN-�
hTCR-C�
h�-actin
(A)
IFN-�-treated*(N1, N3) sham-treated (N2, N4)
N1* N2 N3* N4
hIFN-�-treated Aa-infected
HuPBL-NOD/SCID mice (n=4)
Sham-treated Aa-infected
HuPBL-NOD/SCID mice (n=4)
(C)
(B)
(D)
Mean alveolar bone loss
0
50
100
150
200
250
300
Sham-treated humanized mice IFN-g-treated humanized mice
p=0.017A
rea (
um
2)
FIG. 2. Injection of rh-IFN-� results in an increased coexpression of IFN-� in A. actinomycetemcomitans-reactive RANKL� periodontal CD4�
T cells with a concomitantly higher alveolar bone loss in the diseased periodontal tissues in vivo. (A) Based on Q-PCR, there were significantlyhigher hIFN-� signals detected in A. actinomycetemcomitans-reactive RANKL� CD4� T cells of the diseased periodontal tissues at week 8 fromIFN-�-treated A. actinomycetemcomitans-HuPBL-NOD/SCID mice (see labeled N1* and N3*) than from PBS-treated sham-control mice (labeledN2 and N4; total n � four per group) (only two samples per group are shown here). (B) Based on FACS data, there were significantly higher IFN-�levels coexpressed in 1.5 � 106 A. actinomycetemcomitans-reactive RANKL� CD4� T cells in the diseased periodontal tissues from IFN-�-treatedA. actinomycetemcomitans-HuPBL-NOD/SCID mice than from PBS-treated sham-control mice (total n � 4 per group). Meanwhile, there was alsosignificantly higher alveolar bone loss detected in IFN-�-treated A. actinomycetemcomitans-HuPBL-NOD/SCID than PBS-treated sham-controlmice (week 8; total n � four per group), as shown by the images of the jaw samples (panel C; two jaw samples of the molar areas shown per group)and statistical analysis for comparison between the two groups (see panel D; mean � S.E.; paired Student t test, P � 0.017). The data shown arerepresentative of one of two independent experiments with comparable results.
VOL. 73, 2005 IFN-� POSITIVELY REGULATES RANKL-MEDIATED BONE LOSS 3457
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from
tions post-microbial challenge in vivo. Although IFN-� hasbeen shown to directly inhibit RANKL-mediated osteoclasto-genesis in vitro and in animal studies (11, 13, 42), possibly viaan STAT1-dependent promotion of TRAF6 degradation in theOC precursors pool, opposite findings also exist (4, 6–7, 21,35–36, 39) in which microbial Ag (i.e., an outer membraneprotein of A. actinomycetemcomitans)-reactive CD4�Th1-cellscapably of mediating periodontal bone destruction also coex-pressed IFN-� (22, 43, 51).
Moreover, the results of the present study are consistentwith the findings by Baker et al. (3) that IFN-� knockout micemanifest significantly reduced alveolar bone loss after Porphy-romonas gingivalis oral challenge in vivo, pinpointing the crit-ical role of IFN-� in modulating alveolar bone destruction in
the diseased periodontal tissues and/or cells. At present, the invivo molecular mechanism(s) of this phenomenon as describedabove remains unclear. However, it may not be attributedsolely to the proteosome degradation pathway(s) or signaling(42), as there are no significant differences regarding theTRAF6 transcripts and proteins in the periodontal tissue sam-ples between IFN-�-treated or sham-treated A. actinomycetem-comitans-HuPBL-NOD/SCID mice (data not shown here). Inaddition, IFN-� might mediate its downstream signaling effectsindependently of RANKL-induced osteoclastogenesis path-ways under various inflammatory conditions (i.e., in the pres-ence of TNF-� and IL-1) in vivo. One such possible interactionmay be related to signaling via other non-TRAF6 or non-STAT1 family molecules in the NF-�B and/or JNK pathways
(A)
(B)
AgP1 AgP2 AgP3 N1 N2 (+) controlhIFN-� (501bp)
hIL-4 (350bp)
h�-actin
RA
NK
L-(
-AP
C)
IFN-� (-Cy5.5) IL-4 (-FITC)IFN-� (-Cy5.5)
- Non-diabetic NOD + Aa -
- Diabetic NOD + Aa -
22.1% 36.6% 11.2%
56.5% 49.2% 13.3%
FIG. 3. (A) There are significantly higher levels of expression of IFN-� than of IL-4 in A. actinomycetemcomitans-reactive RANKL�CD4� Tcells from A. actinomycetemcomitans-infected periodontal tissues in AgP patients and diabetic NOD mice. cDNA samples employed for Q-PCRwere derived from 1 � 106 A. actinomycetemcomitans-reactive RANKL� CD4� T cells of three AgP patients as described previously (44). Similarly,106 A. actinomycetemcomitans-reactive RANKL� CD4� T cells were purified from CLN samples of A. actinomycetemcomitans-infected nondiabeticversus diabetic NOD mice at the end of week 8 for the FACS analysis shown here (see RANKL-APC-coupled; IFN-�-Cy5.5-coupled; IL-4-FITC-coupled). Note that (i) there was significantly higher alveolar bone loss detected in the “diabetic” NOD mice than in nondiabetic versus prediabeticNOD mice (n � 5 per group; data not shown) (29) (the results of FACS study from three individual mice of each group are shown here in panelB) and (ii) the results of FACS analysis using CD4� T cells from prediabetic NOD mice were not significantly different from those obtained fornondiabetic NOD mice and hence are not shown here. The data shown above are representative of one of three (in panel A) or two (in panel B)independent experiments with comparable results.
3458 TENG ET AL. INFECT. IMMUN.
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

(11, 24, 41–42). Alternatively, there are regulatory interactionsmediated by non-T-cell sources (i.e., B and NK cells) (8, 23) ordifferent cytokines (i.e., TNF-� and IL-1) (2, 34, 18, 38, 41, 45)that may, at least in part, contribute to the above phenomenon,as it has been shown that B and NK cells can express RANKLand IFN-�, respectively, associated with bone remodeling (8,23). On the same token, it requires further study to seekwhether periodontal resident in tissues or cells can potentiallyinfluence RANKL-mediated osteoclastogenesis in vivo (15, 32,33, 37). Lastly, it is known that IFN-� can up-regulate theexpression of major histocompatibility complex class II andother accessory molecules on the antigen-presenting cells, leu-kocytes, and mesenchymal cells, which may further recruitother signaling molecules and/or immune effectors associated
with bone remodeling (12, 17, 31). Ultimately, the fine balancebetween IFN-� and RANKL under various inflammatory con-ditions may also contribute to determining the outcome oftheir coexpression on Th1 cells for osteoclastogenesis associ-ated with bone remodeling in vivo.
In summary, the present study provides new lines of evi-dence describing a positive coexpression relationship and in-teractions between IFN-� and RANKL-mediated osteoclasto-genesis for alveolar bone loss in periodontal microorganism-specific CD4�T-cell subpopulation(s) in vivo. Our studystrongly suggests that there are complex networks of cytokineregulation associated with RANKL-RANK/OPG signalingpathways for osteoclastogenesis in vivo that are much morecomplicated than what we currently have explored and under-
Group-1
Group-2
Group-3
30% 6% 12% 28%
RA
NK
L /
FIT
C
IFN-� / PE
RA
NK
L /
FIT
C
IFN-� / PE
11% 4 % 10% 3%
Wk 3
Wk 7
Group-1 Group-3
<1% <1%
Un-stained
control
(C)(A)
(B) Mean of the alveolar bone loss
0
50
100
150
200
250
300
Group 1 Group 2 Group 3
Are
a (
um
2)
p=0.048
p=0.031
FIG. 4. Microbial CagE Ag-mediated higher coexpression of IFN-� in A. actinomycetemcomitans-reactive RANKL�CD4� T cells fromCagE-immunized BALB/c mice after A. actinomycetemcomitans oral infection. At day 0, 15 naı̈ve BALB/c mice were sham-treated (group 1),CFA-immunized (group 2), and CagE-immunized (group 3: 20 �g in PBS-CFA; 1:1 ratio), respectively (n � five per group). At 2 weeks later, allmice were orally infected by A. actinomycetemcomitans twice per week for next 3 weeks. Later, all mice were sacrificed at the end of week 7. Jawsamples were collected for digital histomorphometry (44) (images in panels A and B: mean � S.E.) and then A. actinomycetemcomitans-reactiveCD4� T cells were further enriched by in vitro restimulation as described previously (44) followed by FACS analysis for RANKL (FITC-labeled)-positive cells (106 per sample) and intracellular staining for IFN-� (PE-coupled) expression (see panel C). Note that there were significantly higherlevels of IFN-� coexpressed in A. actinomycetemcomitans-reactive RANKL� CD4� T cells in group 3 (CagE-immunized) than group 1 (sham-immunized) and group 2 (CFA-immunized) BALB/c mice. The results of FACS analysis from group 2 were not significantly different from group1 and hence are not shown here. The data shown above are representative of one of the three independent experiments with comparable results.
VOL. 73, 2005 IFN-� POSITIVELY REGULATES RANKL-MEDIATED BONE LOSS 3459
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

stood. Further study of these critical signaling networks isessential for better understanding and developing new treat-ment protocols when dealing with inflammatory bone disor-ders such as human periodontitis.
ACKNOWLEDGMENTS
We thank X. Zhang and M. Alnaeeli for their help in the work.This work was supported by grants to Y.-T.A.T. from the Ministry of
Health of Ontario, Canada; the Canadian Institute of Health Research(CIHR), Canada (MOP-37960); the University of Rochester; and theNational Institutes of Health of the United States (DE12969 andDE14473).
REFERENCES
1. Anderson, D. M., E. Maraskovsky, W. L. Billingsley, W. Dougall, M. E.Tometsko, E. R. Roux, M. C. Teepe, R. F. DuBose. D. Cosman, and L.Galibert. 1997. A homologue of the TNF receptor and its ligand enhanceT-cell growth and dendritic-cell function. Nature 390:175–179.
2. Azuma, Y., K. Kaji, R. Katogi, S. Takeshita, and A. Kudo. 2000. Tumornecrosis factor-alpha induces differentiation of and bone resorption by os-teoclasts. J. Biol. Chem. 275:4858–4864.
3. Baker, P. J., M. Dixon, R. T. Evans, L. Dufour, and D. C. Roopenian. 1999.CD4� T cells and the proinflammatory cytokines gamma interferon andinterleukin-6 contribute to alveolar bone loss in mice. Infect. Immun. 67:2804–2809.
4. Boissier, M. C., G. Chiocchia, N. Bessis, J. Hajnal, G. Garotta, F. Nicoletti,and C. Fournier. 1995. Biphasic effect of interferon-gamma in murine col-lagen-induced arthritis. Eur. J. Immunol. 25:1184–1190.
5. Brown, J. M., J. Zhang, and E. T. Keller. 2004. Opg, RANKL and RANK incancer metastasis: expression and regulation. Cancer Treat. Res. 118:149–172.
6. Canete, J. D., S. E. Martinez, J. Farres, R. Sanmarti, M. Blay, A. Gomez, G.Salvador, and J. Munoz-Gomez. 2000. Differential Th1/Th2 cytokine pat-terns in chronic arthritis: interferon gamma is highly expressed in synoviumof rheumatoid arthritis compared with seronegative spondyloarthropathies.Ann. Rheum. Dis. 59:263–268.
7. Chen, N. J., M. W. Huang, and S. L. Hsieh. 2001. Enhanced secretion ofIFN-� by activated Th1 cells occurs via reverse signaling through TNF-related activation-induced cytokine. J. Immunol. 166:270–276.
8. Choi, Y., K. M. Woo, S. H. Ko, Y. J. Lee, S. J. Park, H. M. Kim, and B. S.Kwon. 2001. Osteoclastogenesis is enhanced by activated B cells but sup-pressed by activated CD8(�) T cells. Eur. J. Immunol. 31:2179–2188.
9. Clark, W. B., and H. Loe. 1993. Mechanisms of initiation and progression ofperiodontal disease. Periodontology 2000. 2:72–82.
10. Coligan, J. E., A. M. Kruisbeek, D. H. Margulies, E. M. Shevach, and W.Strober. 1996. Current protocols in immunology, vol. 2, suppl. 10, p. 10.23.2.John Wiley & Sons, Inc., New York, N.Y.
11. De Klerck, B., I. Carpentier, R. J. Lories, Y. Habraken, J. Piette, G. Car-meliet, R. Beyaert, A. Billiau, and P. Matthys. 2004. Enhanced osteoclasticdevelopment in collagen-induced arthritis in interferon-� receptor knock-outmice as related to increased splenic CD11b� myelopoiesis. Arthritis Res.Ther. 6:R220–R231.
12. Ellis, T. N., and B. L. Beaman. 2004. Interferon-gamma activation of poly-morphonuclear neutrophil function. Immunology 112:2–12.
13. Fox, S. W., and T. J. Chambers. 2000. Interferon-� directly inhibitsTRANCE-induced osteoclastogenesis. Biochem. Biophys. Res. Commun.276:868–872.
14. Gao, X., and Y.-T. A. Teng. 2002. T-cell-receptor gene usage of Actinobacillusactinomycetemcomitans-reactive periodontal CD4� T cells from localizedjuvenile periodontitis patients and human peripheral blood leukocyte-recon-stituted NOD/SCID mice. J. Periodontol. Res. 37:399–404.
15. Hasegawa, T., Y. Yoshimura, T. Kikuiri, Y. Yawaka, S. Takeyama, A. Mat-sumoto, H. Oguchi, and T. Shirakawa. 2002. Expression of receptor activatorof NF-kappa B ligand and osteoprotegerin in culture of human periodontalligamen T cells. J. Periodontal Res. 37:405–411.
16. Henderson, B., S. P. Nair, J. M. Ward, and M. Wilson. 2003. Molecularpathogenicity of the oral opportunistic pathogen Actinobacillus actinomyce-temcomitans. Annu. Rev. Microbiol. 57:29–55.
17. Herold, K. C. 2004. Achieving antigen-specific immune regulation. J. Clin.Investig. 113:346–349.
18. Hofbauer, L. C., D. L. Lacey, C. R. Dunstan, T. C. Spelsberg, B. L. Riggs, andS. Khosla. 1999. Interleukin-1-beta and tumor necrosis factor-alpha, but notinterleukin-6, stimulate osteoprotegerin ligand gene expression in humanosteoblastic cells. Bone 25:255–259.
19. Hofbauer, L. C., and M. Schoppet. 2004. Clinical implication of the osteo-protegerin/RANKL/RANK system for bone and vascular diseases. JAMA28:490–495.
20. Honore, P., N. M. Luger, M. A. C. Sabino, M. J. Schwei, S. D. Rogers, andD. B. Mach. 2000. Osteoprotegerin blocks bone cancer-induced skeletal
destruction, skeletal pain and pain-related neurochemical reorganization ofthe spinal cord. Nat. Med. 6:521–528.
21. Josien, R., B. R. Wong, H. L. Li, R. M. Steinman, and Y. Choi. 1999.TRANCE, a TNF family member, is differentially expressed on T cell subsetsand induces cytokine production in dendritic cells. J. Immunol. 162:2562–2568.
22. Kawai, T., R. Eisen-Lev, M. Seki, J. W. Eastcott, M. E. Wilson, and M. A.Taubman. 2000. Requirement of B7 costimulation for Th1-mediated inflam-matory bone resorption in experimental periodontitis. J. Immunol. 164:2102–2109.
23. Kikuchi, T., C. L. Hahn, S. Tanaka, S. E. Barbour, H. A. Schenkein, andJ. G. Tew. 2004. Dendritic cells stimulated with Actinobacillus actinomyce-temcomitans elicit rapid gamma interferon responses by natural killer cells.Infect. Immun. 72:5089–5096.
24. Koga, T., M. Inui, K. Inoue, S. Kim, A. Suematsu, E. Kobayashi, T. Iwata,H. Ohnishi, T. Matozaki, T. Kodama, T. Taniguchi, H. Takayanagi, and T.Takai. 2004. Costimulatory signals mediated by the ITAM motif cooperatewith RANKL for bone homeostasis. Nature 428:758–763.
25. Kong, Y. Y., U. Feige, I. Sarosi, B. Bolon, A. Tafuri, S. Morony, C. Cappar-elli, J. Li, R. Elliott, S. McCabe, T. Wong, G. Campagnuolo, E. Moran, E. R.Bogoch, G. Van, L. T. Nguyen, P. S. Ohashi, D. L. Lacey, E. Fish, W. J. Boyle,and J. M. Penninger. 1999. Activated T cells regulate bone loss and jointdestruction in adjuvant arthritis through osteoprotegerin ligand. Nature 402:304–308.
26. Kong, Y. Y., H. Yoshida, I. Sarosi, H. L. Tan, E. Timms, C. Capparelli, S.Morony, A. J. Oliveira-dos-Santos, G. Van, A. Itie, W. Khoo, A. Wakeham,C. R. Dunstan, D. L. Lacey, T. W. Mak, W. J. Boyle, and J. M. Penninger.1999. OPGL is a key regulator of osteoclastogenesis, lymphocyte develop-ment and lymph-node organogenesis. Nature 397:315–323.
27. Lacey, D. L., E. Timms, H. L. Tan, M. J. Kelly, C. R. Dunstan, T. Burgess,R. Elliott, A. Colombero, G. Elliott, S. Scully, H. Hsu, J. Sullivan, N.Hawkins, E. Davy, C. Capparelli, A. Eli, Y. X. Qian, S. Kaufman, I. Sarosi,V. Shalhoub, G. Senaldi, J. Guo, J. Delaney, and W. J. Boyle. 1998. Osteo-protegerin ligand is a cytokine that regulates osteoclast differentiation andactivation. Cell 93:165–176.
28. Li, J., I. Sarosi, X. Q. Yan, S. Morony, C. Capparelli, H. L. Tan, S. McCabe,R. Elliott, S. Scully, G. Van, S. Kaufman, S. C. Juan, Y. Sun, J. Tarpley, L.Martin, K. Christensen, J. McCabe, P. Kostenuik, H. Hsu, F. Fletcher, C. R.Dunstan, D. L. Lacey, and W. J. Boyle. 2000. RANK is the intrinsic hema-topoietic cell surface receptor that controls osteoclastogenesis and regula-tion of bone mass and calcium metabolism. Proc. Natl. Acad. Sci. USA97:1566–1571.
29. Mahamed, D. A., A. Marleau, M. Alnaeeli, S. Bhagirath, X. Zhang, J. M.Penninger, and Y.-T. A. Teng. G(�) microbes-reactive CD4� T cells triggerRANKL-mediated enhanced alveolar bone loss in diabetic NOD mice. Di-abetes, in press.
30. Mizuno, A., N. Amizuka, K. Irie, A. Murakami, N. Fujise, T. Kanno, Y. Sato,N. Nakagawa, H. Yasuda, S. Mochizuki, T. Gomibuchi, K. Yano, N. Shima,N. Washida, E. Tsuda, T. Morinaga, K. Higashio, and H. Ozawa. 1998.Severe osteoporosis in mice lacking osteoclastogenesis inhibitory factor/osteoprotegerin. Biochem. Biophys. Res. Commun. 247:610–615.
31. Mochizuki, S., M. Kobayashi, T. Suzuki, A. Oikawa, T. Koseki, T. Nishihara,and K. Hasegawa. 2004. Gamma-interferon enhances expression of CD14/MyD88 and subsequent responsiveness to lipopolysaccharide from Actinoba-cillus actinomycetemcomitans in human gingival fibroblasts. J. PeriodontalRes. 39:333–343.
32. Nagasawa, T., H. Kobayashi, M. Kiji, M. Aramaki, R. Mahanonda, T.Kojima, Y. Murakami, M. Saito, Y. Morotome, and I. Ishikawa. 2002. LPS-stimulated human gingival fibroblasts inhibit the differentiation of monocytesinto osteoclasts through the production of osteoprotegerin. Clin. Exp. Im-munol. 130:338–344.
33. Nukaga, J., M. Kobayashi, T. Shinki, H. Song, T. Takada, T. Takiguchi, R.Kamijo, and K. Hasegawa. 2004. Regulatory effects of interleukin-1beta andprostaglandin E2 on expression of receptor activator of nuclear factor-kappaB ligand in human periodontal ligamen T cells. J. Periodontol. 75:249–259.
34. O’Gradaigh, D., D. Ireland, S. Bord, and J. E. Compston. 2004. Joint erosionin rheumatoid arthritis: interactions between tumor necrosis factor alpha,interleukin 1, and receptor activator of nuclear factor kappa B ligand(RANKL) regulate osteoclasts. Ann. Rheum. Dis. 63:354–359.
35. Ortmann, R. A., and E. M. Shevach. 2001. Susceptibility to collagen-inducedarthritis: cytokine-mediated regulation. Clin. Immunol. 98:109–118.
36. Park, S. H., D. J. Min, M. L. Cho, W. U. Kim, J. Youn, W. Park, et al. 2001.Shift toward T helper 1 cytokines by type II collagen-reactive T cells inpatients with rheumatoid arthritis. Arthritis Rheum. 44:561–569.
37. Rani, C. S., and M. MacDougall. 2000. Dental cells express factors thatregulate bone resorption. Mol. Cell Biol. Res. Commun. 3:145–152.
38. Romas, E., M. T. Gillespie, M. T., and T. J. Martin. 2002. Involvement ofreceptor activator of NF�B ligand and tumor necrosis factor-� in bonedestruction in rheumatoid arthritis. Bone 30:340–346.
39. Ronaghy, A., B. J. Prakken, K. Takabayashi, G. S. Firestein, D. Boyle, N. J.Zvailfler, S. T. Roord, S. Albani, D. A. Carson, and E. Raz. 2002. Immuno-
3460 TENG ET AL. INFECT. IMMUN.
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from

stimulatory DNA sequences influence the course of adjuvant arthritis. J. Im-munol. 168:51–56.
40. Simonet, W. S., D. L. Lacey, C. R. Dunstan, M. Kelley, M. S. Chang, R.Luthy, H. Q. Nguyen, S. Wooden, L. Bennett, T. Boone, G. Shimamoto, M.DeRose, M., R. Elliott, A. Colombero, H. L. Tan, G. Trail, J. Sullivan, E.Davy, N. Bucay, L. Renshaw-Gegg, T. M. Hughes, D. Hill, W. Pattison, P.Campbell, S. Sander, G. Van, J. Tarpley, P. Derby, R. Lee, and W. J. Boyle.1997. Osteoprotegerin: a novel secreted protein involved in the regulation ofbone density. Cell 89:309–319.
41. Takada, Y., and B. B. Aggarwal. Posting date, 17 August 2004. Evidence thatgenetic deletion of the TNF receptor p60 or p80 in macrophages modulatesRANKL-induced signaling. Blood 104:4113–4121. [Online.]
42. Takayanagi, H., K. Ogasawara, S. Hida, T. Chiba, S. Murata, K. Sato, et al.2000. T cell mediated regulation of osteoclastogenesis by signaling cross-talkbetween RANKL and IFN-�. Nature 408:600–605.
43. Taubman, M. A., and T. Kawai. 2001. Involvement of T-lymphocytes inperiodontal disease and in direct and indirect induction of bone resorption.Crit. Rev. Oral Biol. Med. 12:125–135.
44. Teng, Y.-T. A. 2002. Mixed periodontal Th1-Th2 cytokine profile inActinobacillus actinomycetemcomitans-specific osteoprotegerin ligand (orRANK-L)-mediated alveolar bone destruction in vivo. Infect. Immun. 70:5269–5273.
45. Teng, Y.-T. A. 2003. The role of acquired immunity and periodontal diseaseprogression. Crit. Rev. Oral Biol. Med. 14:237–252.
46. Teng, Y.-T. A., H. Nguyen, X. Gao, Y. Y. Kong, R. M. Gorczynski, B. Singh,R. P. Ellen, and J. M. Penninger. 2000. Functional human T-cell immunityand osteoprotegerin ligand control alveolar bone destruction in periodontalinfection. J. Clin. Investig. 106:R59–R67.
47. Teng, Y.-T. A., H. Nguyen, A. Hassanloo, R. P. Ellen, H. Hozumi, and R. M.
Gorczynski. 1999. Periodontal immune responses of human lymphocytes inActinobacillus actinomycetemcomitans-inoculated NOD/SCID mice en-grafted with peripheral blood leukocytes of periodontitis patients. J. Peri-odontol. Res. 34:54–61.
48. Teng, Y.-T. A., and W. Hu. 2003. Expression cloning of a periodontitis-associated apoptotic effector, cage homologue, in Actinobacillus actinomyce-temcomitans. Biochem. Biophys. Res. Commun. 303:1086–1094.
49. Teng, Y.-T. A., and X. Zhang. 2005. Apoptotic activity and sub-cellularlocalization of a T4SS-associated CagE-homologue in Actinobacillus actino-mycetemcomitans. Microb. Pathog. 38:125–132.
50. Theill, L. E., W. J. Boyle, and J. M. Penninger. 2002. RANK-L and RANK:T cell, bone loss and mammalian evolution. Annu. Rev. Immunol. 20:795–823.
51. Valverde, P., T. Kawai, and M. A. Taubman. 2004. Selective blockade ofvoltage-gated potassium channels reduces inflammatory bone resorption inexperimental periodontal disease. J. Bone Miner. Res. 19:155–164.
52. Yasuda, H., N. Shima, N. Nakagawa, K. Yamaguchi, M. Kinosaki, S.Mochizuki, A. Tomoyasu, K. Yano, M. Goto, and A. Murakami. 1998. Os-teoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogen-esis-inhibitory factor and is identical to TRANCE/RANKL. Proc. Natl.Acad. Sci. USA 95:3597–3602.
53. Wong, B. R., R. Josien, S. Y. Lee, B. Sauter, H. L. Li, R. M. Steinman, andY. Choi. 1997. TRANCE (tumor necrosis factor [TNF]-related activation-induced cytokine), a new TNF family member predominantly expressed in Tcells, is a dendritic cell-specific survival factor. J. Exp. Med. 186:2075–2080.
54. Zambon, J. J., T. Umemoto, E. De Nardin, F. Nakazawa, J. A. Christersson,and R. J. Genco. 1988. Actinobacillus actinomycetemcomitans in the patho-genesis of human periodontal disease. Adv. Dent. Res. 2:269–274.
Editor: J. D. Clements
VOL. 73, 2005 IFN-� POSITIVELY REGULATES RANKL-MEDIATED BONE LOSS 3461
on Novem
ber 11, 2014 by guesthttp://iai.asm
.org/D
ownloaded from


















![Silica grafted with a silsesquioxane containing the positively charged 1,4-diazoniabicyclo[2.2.2]octane group used as adsorbent for anionic dye removal](https://static.fdokumen.com/doc/165x107/63228038807dc363600a7986/silica-grafted-with-a-silsesquioxane-containing-the-positively-charged-14-diazoniabicyclo222octane.jpg)

