
Further observations on the primate community at Rudabánya II (late Miocene, early Vallesian age),...
11
Further observations on the primate community at Rudaba´nya II (late Miocene, early Vallesian age), Hungary Miranda Armour-Chelu a, * , Peter Andrews b , Raymond L. Bernor a a College of Medicine, Department of Anatomy, Laboratory of Evolutionary Biology, Howard University, Washington, DC 20059, USA b Department of Palaeontology, Natural History Museum, Cromwell Rd, London, SW7 5BD, UK Received 25 March 2004; accepted 21 March 2005 Abstract It has been proposed that the pliopithecid Anapithecus hernyaki was hunted by the ape Dryopithecus brancoi based on analyses of the age and sex distribution of 25 individuals from the late Miocene site of Rudaba´nya, Hungary (ca. 10 Ma). In this study, the minimum number of individuals (MNI) and age distribution of the assemblage are recalculated and considered in relation to their sedimentary context. We suggest that Dryopithecus juveniles may be underrepresented in the assemblage as a consequence of taphonomic loss of deciduous teeth. Furthermore, we find that while Dryopithecus and Anapithecus were contemporaneous, there was little spatial overlap between them, as each taxon is primarily associated with a separate lithofacies. We conclude that when the revised estimates of MNI and age distribution of these primates are reconsidered in conjunction with their context of burial, the case for the Dryopithecus hunting hypothesis cannot be sustained. Ó 2005 Elsevier Ltd. All rights reserved. Keywords: Dryopithecus; Anapithecus; Age distribution; Tooth loss; Paleoecology; Paleodemography; Taphonomy; Rudaba´nya Introduction Rudaba´nya is a late Miocene site in northeast- ern Hungary first made famous by its assemblage of diverse plant and vertebrate remains (Kretzoi, 1969). The fauna is early Vallesian age, best correlated with late MN 9 faunas of Europe, ca. 9.7e10 Ma (Kordos, 1991; Bernor et al., 2004). Vertebrates are represented by over 100 taxa, with more than 70 species of small and large mammal, including Dryopithecus and Anapithecus (Bernor * Corresponding author. E-mail address: [email protected] (M. Armour-Chelu). 0047-2484/$ - see front matter Ó 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.jhevol.2005.03.007 Journal of Human Evolution 49 (2005) 88e98
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Further observations on the primate community at Rudabánya II (late Miocene, early Vallesian age),...

Journal of Human Evolution 49 (2005) 88e98
Further observations on the primate communityat Rudabanya II (late Miocene, early
Vallesian age), Hungary
Miranda Armour-Chelu a,*, Peter Andrews b, Raymond L. Bernor a
a College of Medicine, Department of Anatomy, Laboratory of Evolutionary Biology,
Howard University, Washington, DC 20059, USAb Department of Palaeontology, Natural History Museum,
Cromwell Rd, London, SW7 5BD, UK
Received 25 March 2004; accepted 21 March 2005
Abstract
It has been proposed that the pliopithecid Anapithecus hernyaki was hunted by the ape Dryopithecus brancoi based
on analyses of the age and sex distribution of 25 individuals from the late Miocene site of Rudabanya, Hungary (ca.10 Ma). In this study, the minimum number of individuals (MNI) and age distribution of the assemblage arerecalculated and considered in relation to their sedimentary context. We suggest that Dryopithecus juveniles may be
underrepresented in the assemblage as a consequence of taphonomic loss of deciduous teeth. Furthermore, we find thatwhile Dryopithecus and Anapithecus were contemporaneous, there was little spatial overlap between them, as each taxonis primarily associated with a separate lithofacies. We conclude that when the revised estimates of MNI and age
distribution of these primates are reconsidered in conjunction with their context of burial, the case for the Dryopithecushunting hypothesis cannot be sustained.� 2005 Elsevier Ltd. All rights reserved.
Keywords: Dryopithecus; Anapithecus; Age distribution; Tooth loss; Paleoecology; Paleodemography; Taphonomy; Rudabanya
Introduction
Rudabanya is a late Miocene site in northeast-ern Hungary first made famous by its assemblage
* Corresponding author.
E-mail address: [email protected] (M. Armour-Chelu).
0047-2484/$ - see front matter � 2005 Elsevier Ltd. All rights reserv
doi:10.1016/j.jhevol.2005.03.007
of diverse plant and vertebrate remains (Kretzoi,1969). The fauna is early Vallesian age, bestcorrelated with late MN 9 faunas of Europe, ca.9.7e10 Ma (Kordos, 1991; Bernor et al., 2004).Vertebrates are represented by over 100 taxa, withmore than 70 species of small and large mammal,including Dryopithecus and Anapithecus (Bernor
ed.
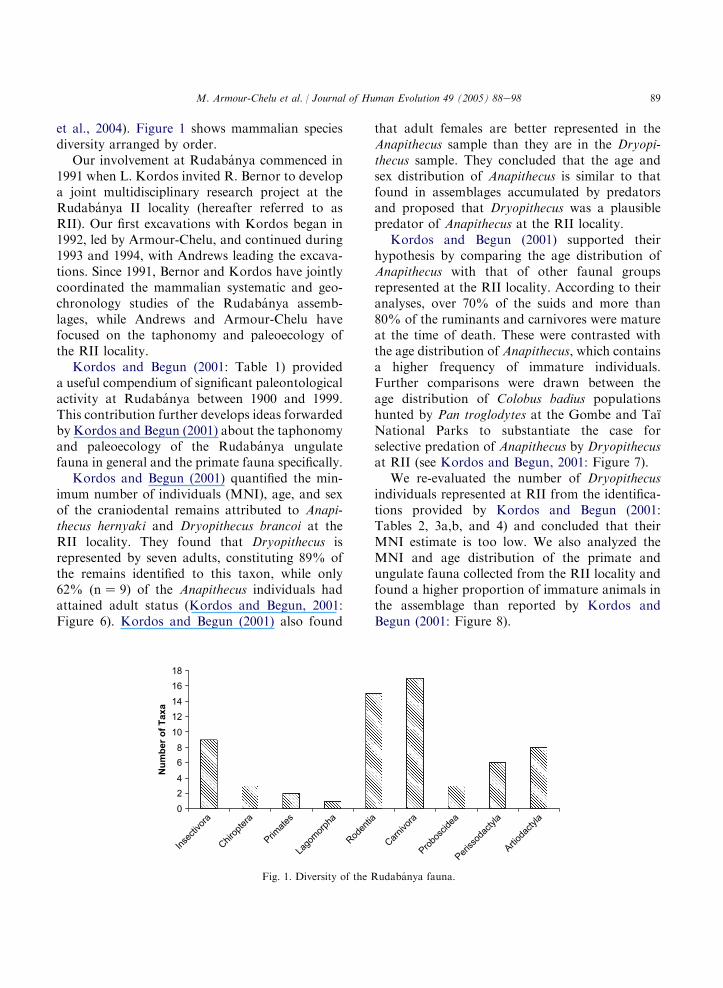
89M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
et al., 2004). Figure 1 shows mammalian speciesdiversity arranged by order.
Our involvement at Rudabanya commenced in1991 when L. Kordos invited R. Bernor to developa joint multidisciplinary research project at theRudabanya II locality (hereafter referred to asRII). Our first excavations with Kordos began in1992, led by Armour-Chelu, and continued during1993 and 1994, with Andrews leading the excava-tions. Since 1991, Bernor and Kordos have jointlycoordinated the mammalian systematic and geo-chronology studies of the Rudabanya assemb-lages, while Andrews and Armour-Chelu havefocused on the taphonomy and paleoecology ofthe RII locality.
Kordos and Begun (2001: Table 1) provideda useful compendium of significant paleontologicalactivity at Rudabanya between 1900 and 1999.This contribution further develops ideas forwardedby Kordos and Begun (2001) about the taphonomyand paleoecology of the Rudabanya ungulatefauna in general and the primate fauna specifically.
Kordos and Begun (2001) quantified the min-imum number of individuals (MNI), age, and sexof the craniodental remains attributed to Anapi-thecus hernyaki and Dryopithecus brancoi at theRII locality. They found that Dryopithecus isrepresented by seven adults, constituting 89% ofthe remains identified to this taxon, while only62% (nZ 9) of the Anapithecus individuals hadattained adult status (Kordos and Begun, 2001:Figure 6). Kordos and Begun (2001) also found
that adult females are better represented in theAnapithecus sample than they are in the Dryopi-thecus sample. They concluded that the age andsex distribution of Anapithecus is similar to thatfound in assemblages accumulated by predatorsand proposed that Dryopithecus was a plausiblepredator of Anapithecus at the RII locality.
Kordos and Begun (2001) supported theirhypothesis by comparing the age distribution ofAnapithecus with that of other faunal groupsrepresented at the RII locality. According to theiranalyses, over 70% of the suids and more than80% of the ruminants and carnivores were matureat the time of death. These were contrasted withthe age distribution of Anapithecus, which containsa higher frequency of immature individuals.Further comparisons were drawn between theage distribution of Colobus badius populationshunted by Pan troglodytes at the Gombe and TaıNational Parks to substantiate the case forselective predation of Anapithecus by Dryopithecusat RII (see Kordos and Begun, 2001: Figure 7).
We re-evaluated the number of Dryopithecusindividuals represented at RII from the identifica-tions provided by Kordos and Begun (2001:Tables 2, 3a,b, and 4) and concluded that theirMNI estimate is too low. We also analyzed theMNI and age distribution of the primate andungulate fauna collected from the RII locality andfound a higher proportion of immature animals inthe assemblage than reported by Kordos andBegun (2001: Figure 8).
024
681012141618
Insectivora
Chiroptera
Primates
Lagomorpha
Rodentia
Carnivora
Proboscidea
Perissodactyla
Artiodactyla
Nu
mb
er o
f T
axa
Fig. 1. Diversity of the Rudabanya fauna.

90 M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
MNI and age distribution of Dryopithecus
The case for selective predation of Anapithecusby Dryopithecus largely rests on counts of MNIand comparisons of the age distribution of thefauna from the RII locality (primates, suids,ruminants, and carnivores). Kordos and Begun(2001: Tables 3a,b) listed the criteria by which theyallocated dental remains to each primate individ-ual to calculate the MNI. This entailed identifyingantimeres, developmental criteria, and tooth wear.Isolated teeth were also allocated to specificindividuals according to their spatial proximitywithin the deposits.
Each individual was assigned to one of four ageclasses (infant, juvenile, subadult, and adult).Animals with the first molar in the process ofemergence were classified as infants and those witha mixture of deciduous and permanent teeth wereawarded juvenile status. Individuals that wereassigned to the subadult category had shed theirdeciduous teeth but the permanent dentition wasnot fully erupted. Individuals with the third molarin the process of emergence or at full occlusalheight were defined as adults.
Nine Dryopithecus individuals (D4-D12) wereidentified from RII by Kordos and Begun (2001:Table 3a), although it should be noted that whileD13 was plotted as an RII find in their Figure 2a, itwas listed as an RIII specimen in their Table 2.Four individuals were identified as juvenile (D3,D9, D13, and D14) from the whole Rudabanyasite, but D9 is the only juvenile Dryopithecusidentified at the RII locality. Three additionalindividuals (D3, D13, and D14) from RI, RIII, andRVII, respectively, were assigned to this age class.Individual D9 is represented by both deciduous(left dp3 and dp4, right dc) and permanent teeth(right P3 germ and upper incisor). Individual D3comprises a premolar tooth germ (P3), and D13 isrepresented by a deciduous canine and two toothgerms (P4 and M2). Individual D14 is representedby a right permanent lower second incisor crown,and does not seem to meet any of the criteriadescribed by Kordos and Begun (2001: 28) forassigning specimens to the juvenile age class.
One individual, D4, was identified as a subadultmale, and it comprises associated maxillary and
mandibular fragments and teeth (RUD 7, 11, 14,and 70). In this individual, the lower permanentincisors and premolars are at full occlusal heightand M2 (RUD 14) is in occlusion. An incompletelyerupted mandibular canine and M3 tooth germ(RUD 11) are also present. Individual D4 is animportant specimen because it provides someevidence for the timing and sequence of toothemergence in Dryopithecus. In D4, emergence ofthe lower premolars and second molar occurred atapproximately the same time, as is usual in apes(Dean, 1985). Unfortunately, it is not possible todetermine the exact order of emergence of thepremolars and second molar. It is possible that M2
erupted slightly earlier than the premolars giventhat it shows slight occlusal wear, but this is notcertain. The type specimen of Dryopithecus fontani(mandible with P3-M2) from Saint Gaudens,France, is of a similar age to D4. In this individual,emergence of the second molar was slightlyadvanced over the premolars (Simons and Meinel,1983), the same as reported for Pan (Fenart andDeblock, 1973; Dean and Wood, 1981; Kinzey,1984).
Seven individuals were identified by Kordosand Begun (2001) as adults. We agree with most ofthese attributions, but Individual D10 appears tobe both more than one individual and not adult. Itcomprises a left P3 germ (RUD 10) and a rightM2 germ (RUD 16), and was assigned adult status(Kordos and Begun, 2001: Table 3a). The presenceof a premolar tooth germ shows that D10 isontogenetically younger than D4, in which thepremolars (RUD 7, RUD 70, and RUD 14) arefully erupted (Kretzoi, 1975: Figure 4; Kordos andBegun, 2001: Figure 3a). Further, it also indicatesthat the deciduous premolars were present at thetime of death and this individual would thereforehave been at a similar stage of development asIndividual D9 (juvenile). In addition to this,however, there are problems with the associationof RUD 10 and RUD 16.
Anemone et al. (1991) found that in Pantroglodytes there is overlap in the development ofadjacent molar tooth germs, so that mineralizationof M3 cusps is initiated before M2 is completelyerupted. This is distinct from Homo, whereemergence of M2 occurs some time before the

91M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
initiation of crown formation in M3 (Dean andWood, 1981). In D. fontani, there is also overlap inthe developmental sequence of M2 and M3, asshown in the radiograph figured in Simons andMeinel (1983: Figure 1). The M2 had recentlyattained full occlusal height, as shown by theabsence of wear facets, and the onset of M3
development is attested to by the presence ofa tooth crypt within the mandibular ramus.Simons and Meinel (1983) suggested that the lossof the M3 tooth germ in this specimen may beattributed to its immaturity (i.e., incompletemineralization).
The relative state of development of M2 and M3
in D. fontani seems similar to that found inP. troglodytes, where M2 crown formation iscomplete at around stage D (48 months in thechimpanzee) and initiation of cusp mineralization(stage A) has commenced in M3 (Anemone et al.,1991). Molars at this early stage of mineralization(stage A) would be extremely unlikely to bepreserved in the fossil record.
If the timing of cheek teeth formation anderuption in Dryopithecus from Rudabanya bearsany resemblance to that of Pan or D. fontani, thenit is unlikely that RUD 10 and RUD 16 (In-dividual D10) are derived from the same animalbecause the M3 is too advanced in its development.The M3 tooth germ RUD 16 exhibits completecrown formation (Begun and Kordos, 1993: 273,Figure 1), and would be assigned to stage C or Dof Anemone’s dental development scheme. At thisstage of development, the associated premolarsand second molar would be in occlusion.
Finally, the allocation to one individual of oneupper tooth (P3) and one lower non-opposingtooth (M3) (RUD 10 and RUD 16, respectively),which were separated by a distance of at least fourmeters at the site, seems tenuous. Accordingly, wesuggest that D10 comprises two individuals, RUD10 (a P3 germ), a juvenile (Individual D10), andRUD 16 (M3 germ), a subadult individual, whichwe assign to New Individual B in Table 1.
Three Dryopithecus teeth from the RII localityare not included in the counts of MNI by Kordosand Begun (2001). These are a right fragmentaryP4 (RUD 46), a left M3 (RUD 13), and a left M2
germ (RUD 19). Specimen RUD 13 was collected
in 1974, as was the left maxilla with CeM1
(RUD 12), but we do not have any informationas to whether these teeth were associated. Speci-men RUD 19 is clearly derived from a younganimal and thus cannot be associated withindividuals assigned to the subadult or adult agecategories. Developmentally, RUD 19 is at a sim-ilar stage as Individual D9 (a juvenile). SpecimenRUD 19 was excavated in 1975 (Kordos andBegun, 2001), and separated by a distance of over6 meters from the D9 individual, which wasrecovered in 1993 and 1994. Accordingly, wesuggest that there are no criteria to attribute thesespecimens to the same individual and we proposethat RUD 19 represents another Dryopithecusindividual, which should be assigned to thejuvenile age class (New Individual A).
In our revision of the MNI and age distributionof Dryopithecus from the RII locality, we concludethat at least eleven individuals are representedin the assemblage, and five of these were immatureat the time of death (Table 1). We could notfurther evaluate the age profile of Anapithecusbecause tooth replacement is rapid in this taxonand there is no clear separation in the timing oferuption between individual cheek teeth, as seen inDryopithecus.
Why are deciduous teeth of Dryopithecuspoorly represented at the RII site?
Only four deciduous teeth attributed to Dry-opithecus are known from Rudabanya (Kordos
Table 1Revised count of Dryopithecus MNI from the RII site
Age group Individual Specimens
Juvenile D9 RUD 124, 126, 147, 153, ?150
Juvenile D10 RUD 10
Juvenile New Individual A RUD 19
Subadult D4 RUD 7, 11, 14, 70
Subadult New Individual B RUD 16
Adult male D7 RUD 44, 47, 144, 45, 140-3
Adult male D11 RUD 82, 85
Adult female D5 RUD 12, 17
Adult female D6 RUD 15, 18
Adult female D8 RUD 77
Adult female D12 RUD 121, 197-200

92 M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
and Begun, 2001). These are two associated upperpremolars and a canine from RII and one uppercanine from RIII. Kordos and Begun (2001:Figure 6) concluded that the Dryopithecus assem-blage was largely composed of adult individualsbased on the low frequency of deciduous teeth. Wehave shown that the presence of permanentpremolar or first and second molar germs atteststo the former existence of deciduous teeth, whichwere lost, probably due to taphonomic or othercircumstances (Table 2).
Some measure of the degree of tooth loss can becalculated for the sample. The MNI for juvenileDryopithecus from RII is three (D9, D10, and NewIndividual A). Given that only two deciduouspremolars were recovered out of the 24 that wereoriginally present (i.e., MNI! total number ofpremolars per individual, or 3! 8), we cancalculate that 92% (i.e., 22/24) of deciduouspremolars are missing.
There are several reasons for non-recovery ofdeciduous teeth from fossil assemblages. Benefit(1994) has shown how retrieval of deciduous teethmay be correlated with collection technique. Lackof screening may result in the loss of small items.However, all matrixes from the RII locality werecarefully sieved to optimize the recovery rate of thefauna (Kordos, pers. comm.). It seems unlikelythat Dryopithecus deciduous teeth are underrepre-sented as a result of excavation techniques. Weinfer that other processes are responsible for theirrarity in this assemblage.
Differential preservation of juvenile versus adultskeletal remains is a common circumstance in thefossil record. Immature bones are poorly mineral-ized compared to mature elements, and thus more
Table 2Dryopithecus tooth germs from Rudabanya
Individual Specimen Locality Lithofacies Tooth
D3 RUD 5 RI d RP3 germ
D4 RUD 11 RII Black mud RM3 germ
D9 RUD 147 RII Black mud RP3 germ
D10 RUD 10 RII Black mud LP3 germ
New Individual A RUD 19 RII Black mud LM2 germ
New Individual B RUD 16 RII Black mud RM3 germ
D13 RUD 79 RIII d LP4 germ
D13 RUD 58 RIII d RM2 germ
susceptible to decay (Benefit, 1994; Andrews andArmour-Chelu, 1998). We believe that deciduousteeth of Dryopithecus are underrepresented atRudabanya because of their inherent structuralproperties combined with the context of deposition.Human tooth enamel is primarily composed of themineral hydroxyapatite, which has a hardness of250e360 VHN (Vickers Hardness Test, kg/sq mm),as opposed to the underlying dentin which hasa hardness of 50e70 (Gutierrez-Salazar and Reyes-Gasga, 2003). It seems plausible that survivorshipof teeth is correlated with enamel thickness becausethicker enamel would be less susceptible to chemicalor biological attack and confer greater protection tothe weaker underlying dentin. The deciduous teethof chimpanzees, orang-utans, and modern humansare more thinly enameled compared to theirpermanent successors (Aiello et al., 1991; Grine,2002). If deciduous tooth enamel is generallythinner than that found in permanent teeth, onewould anticipate greater taphonomic loss of de-ciduous teeth. Further wear (thinning) of enamel byattrition would also reduce thickness, especially onthe occlusal surface of the tooth. Tooth enamelvaries in thickness across taxonomic groups andDryopithecus was a thin-enameled species (Martin,1983). If Dryopithecus deciduous teeth were selec-tively destroyed as a consequence of their structuralqualities, then similar losses should be evident inother fossil assemblages.
Dryopithecus is usually a rather rare componentof late Miocene fauna, but is well represented atCan Llobateres and Can Ponsic, Spain (Crusafont-Pairo and Golpe-Posse, 1973). Harrison (1991:Table 1) published an inventory of all craniodentalspecimens identified as Dryopithecus laietanusfrom the Can Llobateres locality (49 specimens).Most, if not all, of this assemblage was derivedfrom the lowest level of the site, which comprisesblack muds deposited under low-energy conditions(Begun et al., 1990; Moya Sola and Kohler, 1995).Immature Dryopithecus individuals are repre-sented by ten deciduous teeth (7 canines and 3premolars), and two permanent premolar and fivemolar germs (Harrison, 1991). We calculated thatthere are a minimum number of at least threejuvenile individuals in this assemblage. There aretwo right upper first molar germs (IPS 1844, one

93M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
without an IPS accession number) that would havederived from very young, pre-weaned individuals(Smith, 1991). There is also a left M2 germ (IPS1782) from an older individual. All of theseindividuals would have had their deciduouspremolars in use but only 3 out of a potential of24 were recovered (IPS 1784, IPS 1839, IPS 1846).Percentage loss of deciduous premolars in the CanLlobateres assemblage is similar to that found atRII (87.5%, i.e., 21/24).
Twenty Dryopithecus permanent teeth derivedfrom three or four individuals are known fromCan Ponsic (Harrison, 1991; Begun, 1992; Ribotet al., 1996). The sole evidence for immatureDryopithecus individuals in this assemblage isindicated by the P4 germ (IPS 1781) and M1 germ(IPS 1815). Loss of deciduous teeth in thisassemblage is thus 100%.
It seems that Dryopithecus deciduous teeth arepoorly preserved in the fossil record, although ourobservations are limited by the low number ofassemblages containing an adequate sample of thistaxon. Experimental taphonomic studies haveshown that deciduous teeth are more susceptibleto loss when subjected to consumption by carni-vores or weathering (Binford and Bertram, 1977;Munson and Garniewicz, 2003). These factors mayhave been responsible for the losses evident fromthe assemblages described above. The context ofburial also determines the survivorship of faunalremains. Unfavorable conditions such as low pHand leaching will selectively destroy the weakercomponents of an assemblage (Walker et al.,1988).
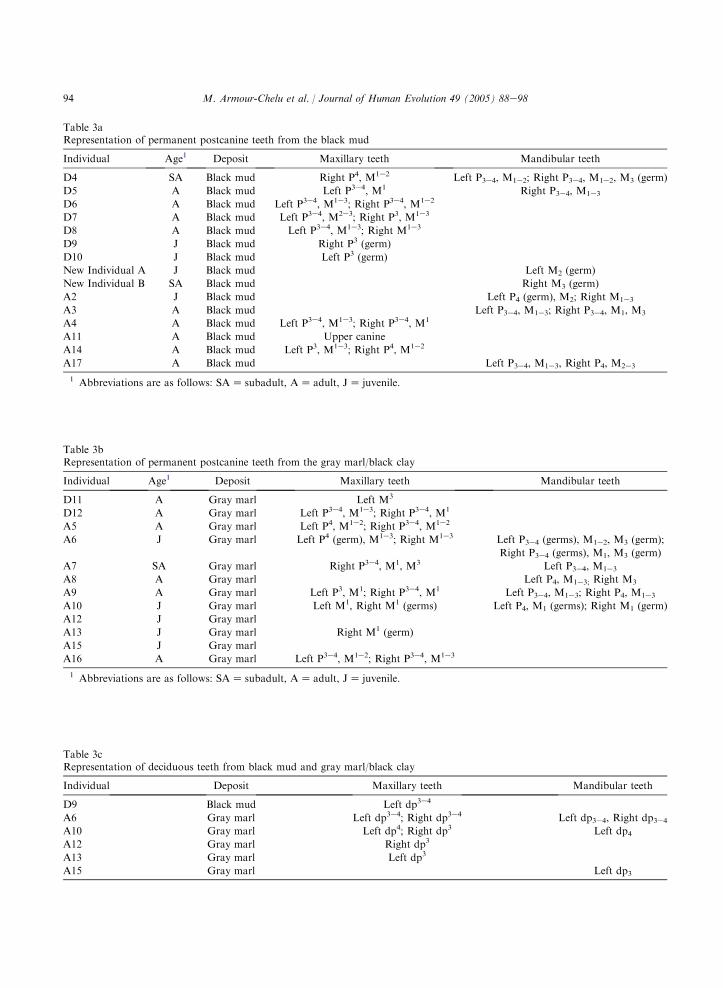
Two major depositional regimes have beenidentified at Rudabanya (Kordos, 1982; P. Andrews,personal observation). The black mud level isfound in the upper part of the sequence and isoverlain by lignites. The lower part of the sequenceconsists of gray marl/black clay that was depositedduring a rise in lake level (Kordos, 1982). Fifteenindividuals of Anapithecus and Dryopithecusare known from the black mud and twelveindividuals are identified from the gray marl/blackclay. Table 3 (a, b, c) itemizes the age andpostcanine dentitions of Anapithecus and Dryopi-thecus individuals and sedimentary context. Boththe black mud and the gray marl/black clay contain
approximately the same number of juvenile indi-viduals, but deciduous teeth are poorly representedin the black mud (3 specimens). Anapithecusjuveniles are rather better represented in the graymarl/black clay, with a total of 14 deciduouspremolars recovered. This represents a loss of 65%of the original number present (Table 3c).
The black mud accumulated under conditionsof higher energy in comparison to the underlyinggray marl/black clay (Kordos and Begun, 2001).Kordos and Begun (2001) also noted that speci-mens recovered from the area of contact betweenthe black mud and lignites are in poor condition.Individuals A4 and D6 show areas of bone lossconsistent with low pH values. The deciduouspremolar crowns of individual A2 are not pre-served, although the tooth roots are intact withinthe alveoli of the mandible (Kordos and Begun,2001). It seems plausible that vertebrate remainsfrom the black mud were subjected to greatermechanical breakage and chemical attack incomparison to material from the gray marl/blackclay.
We conclude that Dryopithecus deciduousdentitions are underrepresented in the black mudas a consequence of the inherent weakness of theseteeth and the unfavorable circumstances of de-position.
Calculation of MNI and age distribution
of the ungulates
Kordos and Begun (2001) figured the age profileof carnivores, suids, and ruminants to supporttheir argument that Anapithecus shows unusuallyhigh juvenile mortality. We calculated the MNIand age distribution of ungulates from the RIIlocality from associated cheek teeth rows andisolated teeth (collected between 1974 and 1994and housed at the Geological Institute of Hungary).Figure 2 shows the age distribution ofDryopithecus,the ruminants [Micromeryx, Euprox, a small bovidspecies, Dorcatherium, Miotragocerus (see Gentry,2004)], and suids (Propotamochoerus) plottedaccording to our analyses. Our results show that48% (nZ 30) of the ruminants died before theirdeciduous teeth were replaced, while the pro-portion of suids in the immature category is 58%
94 M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
Table 3bRepresentation of permanent postcanine teeth from the gray marl/black clay
Individual Age1 Deposit Maxillary teeth Mandibular teeth
D11 A Gray marl Left M3
D12 A Gray marl Left P3e4, M1e3; Right P3e4, M1
A5 A Gray marl Left P4, M1e2; Right P3e4, M1e2
A6 J Gray marl Left P4 (germ), M1e3; Right M1e3 Left P3e4 (germs), M1e2, M3 (germ);
Right P3e4 (germs), M1, M3 (germ)
A7 SA Gray marl Right P3e4, M1, M3 Left P3e4, M1e3
A8 A Gray marl Left P4, M1e3; Right M3
A9 A Gray marl Left P3, M1; Right P3e4, M1 Left P3e4, M1e3; Right P4, M1e3
A10 J Gray marl Left M1, Right M1 (germs) Left P4, M1 (germs); Right M1 (germ)
A12 J Gray marl
A13 J Gray marl Right M1 (germ)
A15 J Gray marl
A16 A Gray marl Left P3e4, M1e2; Right P3e4, M1e3
1 Abbreviations are as follows: SAZ subadult, AZ adult, JZ juvenile.
Table 3cRepresentation of deciduous teeth from black mud and gray marl/black clay
Individual Deposit Maxillary teeth Mandibular teeth
D9 Black mud Left dp3e4
A6 Gray marl Left dp3e4; Right dp3e4 Left dp3e4, Right dp3e4A10 Gray marl Left dp4; Right dp3 Left dp4A12 Gray marl Right dp3
A13 Gray marl Left dp3
A15 Gray marl Left dp3
Table 3aRepresentation of permanent postcanine teeth from the black mud
Individual Age1 Deposit Maxillary teeth Mandibular teeth
D4 SA Black mud Right P4, M1e2 Left P3e4, M1e2; Right P3e4, M1e2, M3 (germ)
D5 A Black mud Left P3e4, M1 Right P3e4, M1e3
D6 A Black mud Left P3e4, M1e3; Right P3e4, M1e2
D7 A Black mud Left P3e4, M2e3; Right P3, M1e3
D8 A Black mud Left P3e4, M1e3; Right M1e3
D9 J Black mud Right P3 (germ)
D10 J Black mud Left P3 (germ)
New Individual A J Black mud Left M2 (germ)
New Individual B SA Black mud Right M3 (germ)
A2 J Black mud Left P4 (germ), M2; Right M1e3
A3 A Black mud Left P3e4, M1e3; Right P3e4, M1, M3
A4 A Black mud Left P3e4, M1e3; Right P3e4, M1
A11 A Black mud Upper canine
A14 A Black mud Left P3, M1e3; Right P4, M1e2
A17 A Black mud Left P3e4, M1e3, Right P4, M2e3
1 Abbreviations are as follows: SAZ subadult, AZ adult, JZ juvenile.

95M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
(nZ 17). We also recorded the age distribution ofnon-ruminant taxa (Proboscidea and Perissodactyla)and found that the proportion of immatureanimals constitutes 43% (nZ 14) of the assem-blage. When the age distribution of Anapithecus(data from Kordos and Begun, 2001) and theungulates are compared, the differences betweenthe percentage of immature animals within anyspecific group are slight (Fig. 2). These data do notsupport the contention of Kordos and Begun(2001: Figure 8) that Anapithecus shows anelevated level of juvenile mortality.
Stratigraphic considerations
Most of the fauna from the RII locality isderived from two levels known as the black mudand gray marl/black clay. Stratigraphically, theblack mud overlies the gray marl/black clay, andKordos and Begun (2001) suggested that thesedeposits represent different lithofacies. The taxo-nomic composition of the fauna from these twolevels is similar, although diversity is lower in theblack mud, possibly because a lesser volume of thisdeposit was excavated. Anapithecus predominatesin the gray marl/black clay, while Dryopithecus ismore abundant in the black mud (Kordos andBegun, 2001). Despite the difference between therelative abundance of the primate taxa in theselevels, Kordos and Begun combined the sample intheir analyses of the age and sex structure of the
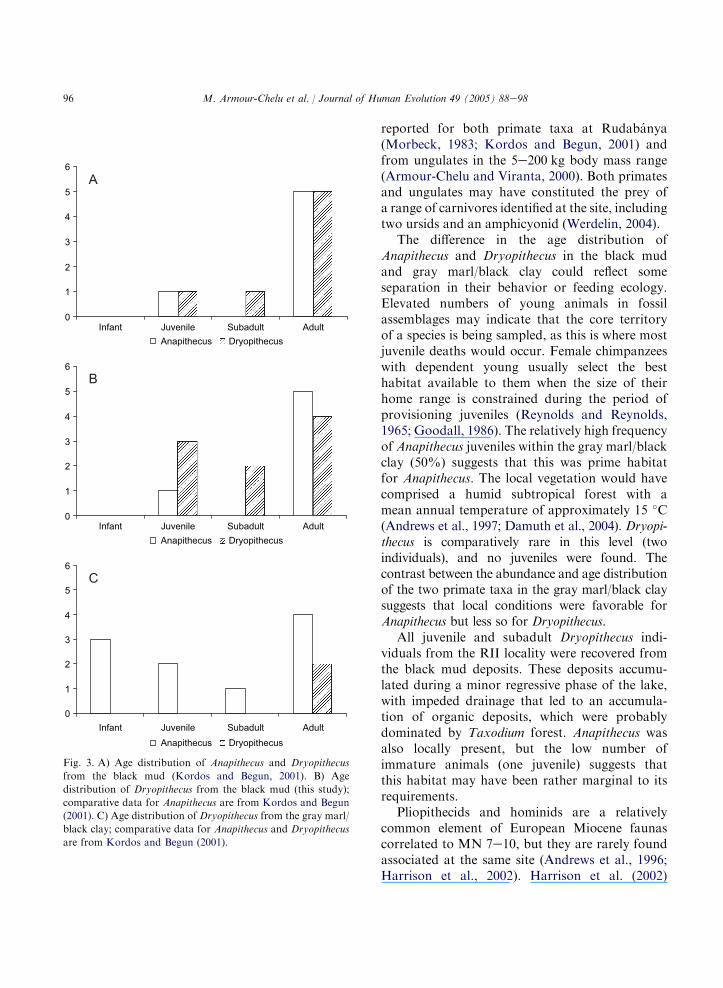
primate remains (see Kordos and Begun, 2001:Figure 7). We plotted the age distribution of thetwo primate species by stratigraphic unit usingboth the data from Kordos and Begun (2001)and with our revised estimates for Dryopithecusdescribed above (Table 1).
Figure 3a shows the age distribution ofAnapithecus and Dryopithecus from the blackmud according to Kordos and Begun’s (2001)data set and Fig. 3b shows our revised estimate.Both plots show that the number of immatureDryopithecus exceeds that of Anapithecus in theblack mud, and according to our calculations, onethird of Dryopithecus individuals were immature atthe time of death. Conversely, Anapithecus isrepresented by five adult individuals (83%) andonly one juvenile in this level. Figure 3c shows theage distribution of Anapithecus and Dryopithecusfrom the gray marl/black clay. In this lithofacies,Anapithecus is represented by 10 individuals, halfof which are immature, while Dryopithecus isrepresented by two adult animals.
Conclusions
We conclude that the age distribution ofAnapithecus and Dryopithecus at the RII localityprovides little support for the hypothesis thatAnapithecus was hunted by Dryopithecus (Kordosand Begun, 2001). Evidence of predation orscavenging (carnivore tooth marks) has been
0
20
40
60
80
SuidsAnapithecus Dryopithecus RuminantsImmatures Adults
Fig. 2. Age distribution of primates, ruminants, and suids. Comparative data for Anapithecus are from Kordos and Begun (2001).
Compare with Figure 8 from Kordos and Begun (2001).
96 M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
0
1
2
3
4
5
6
Infant Juvenile Subadult Adult
Anapithecus Dryopithecus
Infant Juvenile Subadult Adult
Anapithecus Dryopithecus
Infant Juvenile Subadult Adult
Anapithecus Dryopithecus
0
1
2
3
4
5
6
0
1
2
3
4
5
6
A
B
C
Fig. 3. A) Age distribution of Anapithecus and Dryopithecus
from the black mud (Kordos and Begun, 2001). B) Age
distribution of Dryopithecus from the black mud (this study);
comparative data for Anapithecus are from Kordos and Begun
(2001). C) Age distribution of Dryopithecus from the gray marl/
black clay; comparative data for Anapithecus and Dryopithecus
are from Kordos and Begun (2001).
reported for both primate taxa at Rudabanya(Morbeck, 1983; Kordos and Begun, 2001) andfrom ungulates in the 5e200 kg body mass range(Armour-Chelu and Viranta, 2000). Both primatesand ungulates may have constituted the prey ofa range of carnivores identified at the site, includingtwo ursids and an amphicyonid (Werdelin, 2004).
The difference in the age distribution ofAnapithecus and Dryopithecus in the black mudand gray marl/black clay could reflect someseparation in their behavior or feeding ecology.Elevated numbers of young animals in fossilassemblages may indicate that the core territoryof a species is being sampled, as this is where mostjuvenile deaths would occur. Female chimpanzeeswith dependent young usually select the besthabitat available to them when the size of theirhome range is constrained during the period ofprovisioning juveniles (Reynolds and Reynolds,1965; Goodall, 1986). The relatively high frequencyof Anapithecus juveniles within the gray marl/blackclay (50%) suggests that this was prime habitatfor Anapithecus. The local vegetation would havecomprised a humid subtropical forest with amean annual temperature of approximately 15 �C(Andrews et al., 1997; Damuth et al., 2004). Dryopi-thecus is comparatively rare in this level (twoindividuals), and no juveniles were found. Thecontrast between the abundance and age distributionof the two primate taxa in the gray marl/black claysuggests that local conditions were favorable forAnapithecus but less so for Dryopithecus.
All juvenile and subadult Dryopithecus indi-viduals from the RII locality were recovered fromthe black mud deposits. These deposits accumu-lated during a minor regressive phase of the lake,with impeded drainage that led to an accumula-tion of organic deposits, which were probablydominated by Taxodium forest. Anapithecus wasalso locally present, but the low number ofimmature animals (one juvenile) suggests thatthis habitat may have been rather marginal to itsrequirements.
Pliopithecids and hominids are a relativelycommon element of European Miocene faunascorrelated to MN 7e10, but they are rarely foundassociated at the same site (Andrews et al., 1996;Harrison et al., 2002). Harrison et al. (2002)

97M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
proposed that the distribution pattern of pliopi-thecids and hominids in middle to late MioceneEuropean deposits reflects their disparate ecolog-ical requirements. This hypothesis is supported bythe paleodemography of the primate taxa at theRudabanya locality. Immature individuals ofAnapithecus are largely found in lake marginlithofacies while Dryopithecus favored the swampyareas that developed during a period of lakeregression.
Acknowledgements
We gratefully acknowledge the advice andcomments provided by David Cameron, AlanGentry, Terry Harrison, Bill Kimbel, LaszloKordos, Lorenzo Rook, Lars Werdelin, and twoanonymous reviewers. Terry Harrison deservesa special thank you for all the effort he put intothis paper. We thank the National GeographicSociety and L.S.B. Leakey Foundation for grantsupport (to Bernor and Kordos).
Bibliography
Aiello, L.C., Montgomery, C., Dean, C., 1991. The natural
history of deciduous tooth attrition in hominoids. J. Hum.
Evol. 21, 397e412.Andrews, P.A., Armour-Chelu, M.J., 1998. Taphonomic
observations on a surface bone assemblage in a temperate
environment. Bull. Soc. Geol. Fr. 169, 433e442.
Andrews, P.A., Harrison, T., Delson, E., Bernor, R.L.,
Martin, L., 1996. Distribution and biochronology of
European and southwest Asian Miocene catarrhines. In:
Bernor, R.L., Fahlbusch, V., Mittmann, H.-W. (Eds.), The
Evolution of Western Eurasian Neogene Mammal Faunas.
Columbia University Press, New York, pp. 168e207.
Andrews, P.A., Begun, D.R., Zylstra, M., 1997. Interrelation-
ships between functional morphology and paleoenviron-
ments in Miocene hominoids. In: Begun, D.R., Ward, C.V.,
Rose, M.D. (Eds.), Function, Phylogeny and Fossils:
Miocene Hominoid Evolution and Adaptations. Plenum
Press, New York, pp. 29e58.Anemone, R.L., Watts, E., Swindler, D.R., 1991. Dental
development of known-age chimpanzees, Pan troglodytes
(Primates, Pongidae). Am. J. Phys. Anthropol. 86, 229e241.
Armour-Chelu, M.J., Viranta, S., 2000. Carnivore modification
to Rudabanya bones. Carolinea 58, 93e102.
Begun, D.R., 1992. New species of Dryopithecus from the
Vallesian of Can Ponsic (northeastern Spain). Am. J. Phys.
Anthropol. 87, 291e309.
Begun, D.R., Moya-Sola, S., Kohler, M., 1990. New Miocene
hominoid specimens from Can Llobateres (Valles Penedes,
Spain) and their geological and paleoecological context. J.
Hum. Evol. 19, 255e268.
Begun, D.R., Kordos, L., 1993. Revision of Dryopithecus
brancoi SCHLOSSER, 1901 based on the fossil hominoid
material from Rudabanya. J. Hum. Evol. 25, 271e285.Benefit, B.R., 1994. Phylogenetic, paleodemographic, and
taphonomic implications of Victoriapithecus deciduous teeth
fromMaboko,Kenya. Am. J. Phys. Anthropol. 95, 277e327.
Bernor, R.L., Kordos, L., Rook, L. (Eds.), 2004. Recent
advances on multidisciplinary research at Rudabanya, late
Miocene (MN9), Hungary: a compendium. Palaeontol. Ital.
89, pp. 3e36 (2002).
Binford, L.R., Bertram, J.B., 1977. Bone frequenciesdand
attritional processes. In: Binford, L.R. (Ed.), For Theory
Building in Archaeology. Academic Press, New York,
pp. 77e153.Crusafont-Pairo, M., Golpe-Posse, J.M., 1973. New pongids
from the Miocene of Valles Penedes Basin (Catalonia,
Spain). J. Hum. Evol. 2, 17e23.
Dean, M.C., 1985. The eruption pattern of the permanent
incisors and first permanent molars in Australopithecus
(Paranthropus) robustus. Am. J. Phys. Anthropol. 67, 251e
257.
Dean, M.C., Wood, B.A., 1981. Developing pongid dentition
and its use for ageing individual crania in comparative
cross-sectional growth studies. Folia Primatol. 36, 111e127.
Damuth, J., van Dam, J.A., Utescher, T., 2004. Palaeoclimate
estimates from biotic proxies. In: Bernor, R.L., Kordos, L.,
Rook, L. (Eds.), Recent advances on multidisciplinary
research at Rudabanya, late Miocene (MN9), Hungary:
a compendium. Palaeont. Ital. 89, pp. 3e36 (2002).
Fenart, R., Deblock, R., 1973. Pan paniscus and Pan troglodytes
e Craniometrie e Etude comparative et ontogenique selon
les methods classique et vestibulaire. Tome 1. Musee Royal
de l’ Afrique Centralee Turvuren, Belgique Annalese Serie
IN- 8 Sciences Zoologiques e No. 204.
Gentry, A., 2004. Ruminants. In: Bernor, R.L., Kordos, L.,
Rook, L. (Eds.), Recent advances on multidisciplinary
research at Rudabanya, late Miocene (MN9), Hungary:
a compendium. Palaeont. Ital. 89, pp. 3e36 (2002).
Goodall, J., 1986. The Chimpanzees of Gombe: Patterns of
Behavior. Belknap Harvard University Press, MA.
Grine, F.E., 2002. Scaling of tooth enamel thickness, and molar
crown size reduction in modern humans. S. Afr. J. Sci. 98,
503e508.
Gutierrez-Salazar, M.d.P., Reyes-Gasga, J., 2003. Microhard-
ness and chemical composition of human tooth. Mater. Res.
6, 367e373.
Harrison, T., 1991. Some observations on the Miocene
hominoids from Spain. J. Hum. Evol. 20, 515e520.
Harrison, T., van der Made, J., Ribot, F., 2002. A new middle
Miocene pliopithecid from Sant Quirze, northern Spain.
J. Hum. Evol. 42, 371e377.
Kinzey, W.G., 1984. The dentition of the pygmy chim-
panzee, Pan paniscus. In: Susman, R.L. (Ed.), The Pygmy

98 M. Armour-Chelu et al. / Journal of Human Evolution 49 (2005) 88e98
Chimpanzee: Evolutionary Biology and Behavior. Plenum
Press, New York, pp. 65e88.
Kordos, L., 1982. The prehominid locality of Rudabanya
(NE Hungary) and its neighbourhood: a palaeogeographic
reconstruction. Foldt. Int. Evi. Jel. 1980, 395e406.
Kordos, L., 1991. Le Rudapithecus hungaricus de Rudabanya
(Hongrie). L’Anthropologie (Paris) 95, 343e362.Kordos, L., Begun, D.R., 2001. Primates from Rudabanya:
allocation of specimens to individuals, sex and age
categories. J. Hum. Evol. 40, 17e39.
Kretzoi, M., 1969. Geschichte der primaten und der hominisa-
tion. Symp. Biol. Hung. 9, 3e11.
Kretzoi, M., 1975. New ramapithecines and Pliopithecus from
the lower Pliocene of Rudabanya in north-eastern Hungary.
Nature 257, 578e581.
Martin, L., 1983. The relationships of the later Miocene
Hominoidea. Ph.D. Dissertation, University College London.
Moya Sola, S., Kohler, M., 1995. New partial cranium of
Dryopithecus Lartet, 1863 (Hominoidea, Primates) from the
upper Miocene of Can Llobateres, Barcelona, Spain.
J. Hum. Evol. 29, 101e139.
Morbeck, M.E., 1983. Miocene hominoid discoveries from
Rudabanya: implications from the postcranial skeleton. In:
Ciochon, R.L., Corruccini, R.S. (Eds.), New Interpretations
of Ape and Human Ancestry. Plenum Press, New York,
pp. 369e404.
Munson, P.J., Garniewicz, R.C., 2003. Age-mediated survivor-
ship of ungulate mandibles and teeth in canid-ravaged
faunal assemblages. J. Archaeol. Sci. 30, 405e416.
Ribot, F., Gibert, J., Harrison, T., 1996. A reinterpretation
of the taxonomy of Dryopithecus from Valles-Penedes,
Catalonia (Spain). J. Hum. Evol. 31, 129e141.
Reynolds, V., Reynolds, F., 1965. Chimpanzees of the Budongo
Forest. In: DeVore, I. (Ed.), Primate Behavior. Holt,
Rinehart and Winston, New York, pp. 368e424.Simons, E.L., Meinel, W., 1983. Mandibular ontogeny in
the Miocene great ape Dryopithecus. Int. J. Primatol. 4,
331e337.
Smith, B.H., 1991. Dental development and the evolution of
life history in Hominidae. Am. J. Phys. Anthropol. 86,
157e174.
Walker, P.L., Johnson, J.R., Lambert, P.M., 1988. Age and sex
biases in the preservation of human skeletal remains. Am. J.
Phys. Anthropol. 76, 183e188.
Werdelin, L., 2004. Carnivora. In: Bernor, R.L., Kordos, L.,
Rook, L. (Eds.), Recent Advances on Multidisciplinary
Research at Rudabanya Late Miocene (MN9), Hungary:
a compendium. Palaeont. Ital. 89 (2002), 3e36.




















