
Flower development in garlic: the ups and downs of gaLFY expression
10
ORIGINAL ARTICLE Flower development in garlic: the ups and downs of gaLFY expression Rotem Neta • Rakefet David-Schwartz • Yuval Peretz • Ilan Sela • Haim D. Rabinowitch • Moshe Flaishman • Rina Kamenetsky Received: 28 October 2010 / Accepted: 12 January 2011 / Published online: 1 February 2011 Ó Springer-Verlag 2011 Abstract The lack of sexual processes prohibits genetic studies and conventional breeding in commercial cultivars of garlic. Recent restoration of garlic flowering ability by environmental manipulations has opened new avenues for physiological and genetic studies. The LEAFY homologue gaLFY has been shown to be involved in the floral devel- opment, while two alternatively spliced gaLFY transcripts are expressed in flowering genotypes. In the present work, quantitative real-time PCR and two techniques of RNA in situ hybridization were employed to analyze spatiotempo- ral expression patterns of the gaLFY during consequent stages of the garlic reproductive process. Temporal accu- mulation of gaLFY is strongly associated with reproductive organs, significantly increased during florogenesis and gametogenesis, and is down-regulated in the vegetative meristems and topsets in the inflorescence. The two alter- native transcripts of the gene show different expression patterns: a high level of the long gaLFY transcript coincided only with floral transition, while further up-regulation of this gene in the reproductive organs is associated mainly with the short gaLFY transcript. It is concluded that gaLFY is involved at different stages of the sexual reproduction of garlic. These new insights broaden our basic understanding of flower biology of garlic and help to establish conventional and molecular breeding systems for this important crop. Keywords Florogenesis Allium sativum qPCR FISH DIG-labeled In situ hybridization LEAFY homologue Introduction Modern cultivars of garlic (Allium sativum L.), one of the most economically important plants, are completely sterile and thus propagated only vegetatively. The lack of sexual propagation prohibits genetic studies and conventional breeding, and fertility restoration should significantly broaden variability and make the improvement of eco- nomically important traits possible (Kamenetsky 2007). Recent introductions of fertile A. sativum accessions from Central Asia (Etoh 1985; Etoh and Simon 2002), restora- tion of flowering ability and fertility by environmental manipulations (Kamenetsky et al. 2004b) and increased variation by seed propagation (Jenderek and Hannan 2004; Jenderek and Zewdie 2005; Kamenetsky et al. 2005; Shemesh et al. 2008) open the way for in-depth physio- logical and genetic studies, as well as conventional and molecular breeding in this plant. However, the regulation of floral development and fertility in geophytes remains obscure, and fundamental physiological and genetic knowledge is required for both basic and applied purposes. In this context, investigation of genes involved in the Electronic supplementary material The online version of this article (doi:10.1007/s00425-011-1361-8) contains supplementary material, which is available to authorized users. R. Neta Y. Peretz I. Sela H. D. Rabinowitch The Robert H. Smith Faculty of Agriculture, Food and Environment, The Hebrew University of Jerusalem, The Robert H. Smith Institute of Plant Science and Genetics in Agriculture, 76100 Rehovot, Israel R. David-Schwartz M. Flaishman R. Kamenetsky (&) Institute of Plant Sciences, ARO, The Volcani Center, 50250 Bet Dagan, Israel e-mail: [email protected] R. David-Schwartz The Mina and Everard Goodman Faculty of Life Sciences, Bar-Ilan University, Ramat Gan, Israel 123 Planta (2011) 233:1063–1072 DOI 10.1007/s00425-011-1361-8
Transcript of Flower development in garlic: the ups and downs of gaLFY expression

ORIGINAL ARTICLE
Flower development in garlic: the ups and downs of gaLFYexpression
Rotem Neta • Rakefet David-Schwartz •
Yuval Peretz • Ilan Sela • Haim D. Rabinowitch •
Moshe Flaishman • Rina Kamenetsky
Received: 28 October 2010 / Accepted: 12 January 2011 / Published online: 1 February 2011
� Springer-Verlag 2011
Abstract The lack of sexual processes prohibits genetic
studies and conventional breeding in commercial cultivars
of garlic. Recent restoration of garlic flowering ability by
environmental manipulations has opened new avenues for
physiological and genetic studies. The LEAFY homologue
gaLFY has been shown to be involved in the floral devel-
opment, while two alternatively spliced gaLFY transcripts
are expressed in flowering genotypes. In the present work,
quantitative real-time PCR and two techniques of RNA in
situ hybridization were employed to analyze spatiotempo-
ral expression patterns of the gaLFY during consequent
stages of the garlic reproductive process. Temporal accu-
mulation of gaLFY is strongly associated with reproductive
organs, significantly increased during florogenesis and
gametogenesis, and is down-regulated in the vegetative
meristems and topsets in the inflorescence. The two alter-
native transcripts of the gene show different expression
patterns: a high level of the long gaLFY transcript
coincided only with floral transition, while further
up-regulation of this gene in the reproductive organs is
associated mainly with the short gaLFY transcript. It is
concluded that gaLFY is involved at different stages of the
sexual reproduction of garlic. These new insights broaden
our basic understanding of flower biology of garlic and
help to establish conventional and molecular breeding
systems for this important crop.
Keywords Florogenesis � Allium sativum � qPCR � FISH �DIG-labeled � In situ hybridization � LEAFY homologue
Introduction
Modern cultivars of garlic (Allium sativum L.), one of the
most economically important plants, are completely sterile
and thus propagated only vegetatively. The lack of sexual
propagation prohibits genetic studies and conventional
breeding, and fertility restoration should significantly
broaden variability and make the improvement of eco-
nomically important traits possible (Kamenetsky 2007).
Recent introductions of fertile A. sativum accessions from
Central Asia (Etoh 1985; Etoh and Simon 2002), restora-
tion of flowering ability and fertility by environmental
manipulations (Kamenetsky et al. 2004b) and increased
variation by seed propagation (Jenderek and Hannan 2004;
Jenderek and Zewdie 2005; Kamenetsky et al. 2005;
Shemesh et al. 2008) open the way for in-depth physio-
logical and genetic studies, as well as conventional and
molecular breeding in this plant. However, the regulation
of floral development and fertility in geophytes remains
obscure, and fundamental physiological and genetic
knowledge is required for both basic and applied purposes.
In this context, investigation of genes involved in the
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00425-011-1361-8) contains supplementarymaterial, which is available to authorized users.
R. Neta � Y. Peretz � I. Sela � H. D. Rabinowitch
The Robert H. Smith Faculty of Agriculture,
Food and Environment, The Hebrew University of Jerusalem,
The Robert H. Smith Institute of Plant Science and Genetics
in Agriculture, 76100 Rehovot, Israel
R. David-Schwartz � M. Flaishman � R. Kamenetsky (&)
Institute of Plant Sciences, ARO, The Volcani Center,
50250 Bet Dagan, Israel
e-mail: [email protected]
R. David-Schwartz
The Mina and Everard Goodman Faculty of Life Sciences,
Bar-Ilan University, Ramat Gan, Israel
123
Planta (2011) 233:1063–1072
DOI 10.1007/s00425-011-1361-8

control of flowering is of great importance (Benschop et al.
2010; Noy-Porat et al. 2010; Rotem et al. 2007).
In general, the formation of inflorescence and flowers in
plants consists of a series of sequential steps. These pro-
cesses were studied in detail in Arabidopsis and other
model plants. The effect of environmental factors on
flowering involves several signaling pathways that con-
verge towards the regulation of floral meristem identity
(Mouradov et al. 2002). ‘Integrators’ genes, such as
FLOWERING LOCUS T (FT) and SUPPRESSOR OF
OVEREXPRESSION OF CO1 (SOC1) co-operate in the up-
regulation of floral meristem identity genes like LEAFY
(LFY) and APETALA1 (AP1) (Benlloch et al. 2007;
Cockram et al. 2007; Krizek and Fletcher 2005). These
genes up-regulate each other and are essentially expressed
in the shoot apical meristem (SAM), where the transition
from vegetative to reproductive development is followed
by the differentiation of inflorescence and individual
flowers. The activity of LFY/AP1 is antagonized by TFL1
(TERMINAL FLOWER 1) whose expression maintains the
indeterminacy of the SAM (Ratcliffe 1999; Bernier and
Perilleux 2005).
The meristem identity-genes activate the floral-organ
identity-genes in discrete meristematic regions of the
inflorescence, with the consequent differentiation of flower
primordia. In turn, the floral organ identity genes regulate
the various cell types and tissues’ specialization, including
sepals, petals, stamens and carpels (Benlloch et al. 2007;
Bernier and Perilleux 2005; Moon et al. 2005). Regulatory
genes, involved in this process, act either as activators or as
repressors of floral initiation, and the concerted action of
the regulatory network ensures the correct temporal and
spatial control of floral meristem specification, its mainte-
nance and development (Vijayraghavan et al. 2005).
The gene LFY of Arabidopsis encodes for a unique
plant-specific transcription factor that plays a key role in
plant development (Moyroud et al. 2010). It was proposed
that LFY combines properties of flowering-time and flower-
meristem-identity genes, and provides a direct link between
the global process of floral induction (Benlloch et al. 2007;
Blazquez and Weigel 2000; Nilsson et al. 1998), the
regional events associated with the initiation of individual
flowers (Vijayraghavan et al. 2005), and activation of the
ABC genes such as AGAMOUS (AG), APETALA1 (AP1)
and APETALA3 (AP3) that control floral organ identity
(Lamb et al. 2002; Zik and Irish 2003; Benlloch et al.
2007).
LFY and its homologue FLORICAULA (FLO) were first
discovered in Arabidopsis and in Antirrhinum, respectively
(Coen et al. 1990; Weigel et al. 1992), followed by a
number of homologous genes in rice (Oryza sativa: RFL,
Kyozuka et al. 1998); Darnel ryegrass (Lolium temulentum:
LtLFY, Gocal et al. 2001); Narcissus (NLF, Noy-Porat et al.
2010), and more. Recently, we have demonstrated that the
LFY homologue gaLFY is involved in the control of
florogenesis in garlic. Further, we showed that two alter-
natively spliced gaLFY transcripts are expressed in flow-
ering garlic plants (Rotem et al. 2007).
In the present work, we studied the temporal and spatial
expression patterns of gaLFY during several sequential
stages of garlic flower development. Three methodologies
were employed, to analyze the expression patterns of
gaLFY during the differentiation of garlic inflorescence,
individual flowers and floral organs.
Materials and methods
Plant material and sampling procedures
Bulbs of garlic clone #3028, originated from Central Asia,
were obtained from the Field Gene Bank for Vegetatively
Propagated Short-Day Allium spp., The Robert H. Smith
Faculty of Agriculture, Food and Environment, Rehovot,
Israel (FAHU). In Israel, these plants produce bulbs, fertile
flowers and seeds. In June 2008, mature bulbs were har-
vested, cured and stored in a roofed shed under ambient
conditions until September. Intact bulbs were then placed
in a temperature-controlled room at 4�C for 8 weeks, in
the dark. In November 2008, individual clean, healthy
cloves were sorted for size and planted in the FAHU
experimental farm in Rehovot, and standard agricultural
practices were applied throughout. Destructive morpho-
logical analyses on three randomly selected plants were
performed weekly, and apical meristems were sampled for
scanning electron microscopy (SEM), RNA analysis and
in situ studies.
Scanning electron microscopy
Freshly harvested plants were carefully stripped of their
leaves and the spathes removed from the developing
inflorescences. Samples were fixed in a 5:5:90 mixture of
glacial acetic-acid:formalin (40%):ethanol (70%), dehy-
drated in a graded ethanol series (25, 50, 75, 95, 100%),
dried using liquid CO2 in a Bio-Rad 750 critical-point
dryer, mounted on SEM stubs and gold coated. Images
were obtained directly from the SEM (JSM-35C, JEOL
Japan) at 15 kV.
RNA isolation and cDNA preparation
Entire shoot apices, including reproductive meristems and
inflorescences were collected weekly from three garlic
plants, under binocular (November 2008–June 2009).
Additionally, 30–40 flower buds and flowers at the same
1064 Planta (2011) 233:1063–1072
123

developmental stages were carefully detached from the
inflorescence (May and June 2009). All collected samples
were immediately frozen in liquid nitrogen, and RNA
extracted using RNeasy Mini Kit (Qiagen, Hilden,
Germany). Extracts were treated with DNase using
TURBO DNA-free KIT (Ambion Austin, TX, USA) and
tested for DNA contamination. Amount and quality of
RNA were determined by spectrophotometry (NanoDrop
Technologies, Wilmington, DE, USA). Only RNA samples
of sufficient purity with absorbance ratio A260/280 [1.8
were considered for synthesis of cDNA. Total RNA from
individual samples was reverse-transcribed to produce
cDNA in a 20 ll reaction volume containing 1.5 lg of
RNA sample, using EZ-first strand cDNA synthesis kit
(Biological Industries, Israel) according to the manufac-
turer protocol. Each sample of the obtained cDNA was
diluted in a ratio of 1:10 before amplification.
gaLFY expression patterns were analyzed by quantita-
tive real-time PCR (qPCR) and RNA in situ hybridization,
using primers and probes designed from the formerly
described 682-bp fragment of the gene (Rotem et al. 2007;
GenBank AY563104 http://www.ncbi.nlm.nih.gov/nuccore/
48476628).
Quantitative real-time PCR
Quantification of gaLFY (the sum of the two transcripts;
gaLFYl and gaLFYs, Rotem et al. 2007) was determined
using the commercial dual hybridization probe #144
(CCTCTTCC) designed using the Universal Probe Library
Software (http://www.roche-applied-science.com) and
provided as a master PCR mix by Roche. The primers used
for gaLFY are presented in Table S1 and Fig. S1. Several
reference genes were tested with similar results, and actin
(AY821677) transcript was selected for data normalization
and for calculation of the relative mRNA levels. The
commercial dual hybridization probe #64 (CCAGGCTG)
was used to detect qPCR of actin. The qPCR primers used
for forward and reverse actin transcript are depicted in
Table S1. qPCR was carried out in a 20-ll reaction volume
using FastStart Universal Probe Master (92) (Roche,
Indianapolis, IN, USA); the reaction mixture included
0.5 lM primer, and 4 ll cDNA.
Reactions were performed using Light-Cycler 480 Real-
Time PCR (Roche) in multi-well plate. In each PCR plate,
at both primer combinations, two internal controls were
included: a negative control without cDNA template and
a sample from the eighth leaf (February 2, 2009) as a
calibrator (Pfaffl et al. 2002). For each primer set, PCR
efficiency was determined by standard curve method
(Table S1).
The reaction was performed under the following
cycling conditions: an initial denaturing step of 10 min at
95�C, followed by 40 cycles consisting of 10 s at 95�C
and 25 s at 60�C. All qPCR experiments were repeated in
three biological and two technical replications. During
inflorescence development, gaLFY was quantified using
the DDCt method (Pfaffl 2001), and the n-fold change was
calculated with LightCycler 480 software, provided by
the manufacturer, relative to the vegetative sample as
described above.
Quantitative analysis of gaLFYl was performed by
qPCR using the SYBR Green fluorescent dye. Actin was
used as reference gene, and the eighth leaf served as cali-
brator in the QC with the negative control in each qPCR
plate. Primers were designed by Primer3 software (http://
frodo.wi.mit.edu/primer3). Primers for actin were descri-
bed above (Table S1). The reactions were carried out in a
20 ll RT-reaction mixture containing 10 ll of FastStart
SYBR green I Master 910 (Roche), 0.5 lM of each pri-
mer, and 4 ll cDNA template, and performed with the
following cycling conditions: 1 cycle for 10 min at 95�C,
followed by 40 cycles of 10 s at 95�C, 10 s at 58�C and
10 s at 72�C. PCR analyses were performed using 96
samples Light-Cycler 480 and Real-Time PCR software
(Roche) as described above. All PCR experiments were
carried out in three biological and two technical
replications.
RNA fluorescence in situ hybridization
Modified fluorescence in situ hybridization (FISH) tech-
nique was adapted from Gottlieb et al. (2006). Freshly
harvested plants were carefully stripped of leaves and
spathes. Apices’ samples for microscope analysis were
collected from November to June, and fixed overnight in
Carnoy’s fixative (6:3:1 mixture of chloroform:etha-
nol:glacial acetic acid). Thereafter, samples were decolor-
ized twice in 6% H2O2 solution in ethanol for 1 h.
Pre-hybridization in hybridization buffer [20 mM Tris–
HCl, pH 8.0, 0.9 M NaCl, 0. 01% sodium dodecyl sulfate
(SDS), 30% formamide] for 1 h was followed by overnight
hybridization at room temperature using the same buffer
supplemented with 1 pmol fluorescent probe/ml. The
hybridized samples were washed twice in 29 sodium
chlorides/sodium citrate (SSC), 0.1% SDS solution for
10 min, and once with a mixture of 0.59 SSC and 0.1%
SDS for 10 min at room temperature. Meristems or
reproductive tissues similarly hybridized except that the
fluorescent probe was replaced by water, served as con-
trols. Samples were then stored in 29 SSC at 4�C, until
studied by confocal microscope (LSM 510 Axiovert
100 M; ZEISS, USA). The labeled probe 50-CUAUGA
ACGGGUGCUCCCUUUGCCG-30 (Fig. S1) was designed
to detect the two gaLFY transcripts (Bioneer, Munpyeong-
dong, Daedeok-gu, Daejeon 306-220, Korea).
Planta (2011) 233:1063–1072 1065
123

DIG-labeled in situ hybridization
DIG-labeled in situ analysis was performed according to
Szymkowiak and Irish (2006). Meristems were fixed in
FAA (formaldehyde:acetic acid:70% ethanol, 10:5:85,
v/v), dehydrated in a series of ethanol solutions and
embedded in ParaPlast (McCormick Scientific, St. Louis,
MO, USA) according to Johansen (1940), then sliced
(10 lm) by a Leica RM2245 microtome (VectaMount-
Vector Laboratories, Burlingame, CA, USA), and placed
on SuperFrost Plus slides (Menzel-Glaser, Braunschweig,
Germany) for 2 days on a 37�C hot plate.
The 195-bp gaLFY segment (Fig. S1) isolated by RT–
PCR was cloned into a p-Drive cloning vector (Qiagen)
with a T7 promoter attached to the 50-end of the gene and
an SP6 promoter sequence attached to its 30-end. DIG-
labeled RNA sense and antisense probes were generated
using the MEGAscript kit (Ambion) and DIG RNA
labeling mix (Roche Applied Science), purified by
MEGAclear kit (Ambion) and quantified on agarose
gel, 1 ll per sample. Sections were deparaffinized in
Histoclear (Finkelman Chemicals, Yahood, Israel) for
2 9 10 min and rehydrated in a decreasing ethanol series,
followed by 20 min wash with 29 SSC, and by a 30-min
incubation at 37�C with 1 lg ml-1 proteinase K
(Fermentas) in TE buffer. The sections were then fixed
for 10 min in 4% paraformaldehyde in PBS buffer, and
re-dehydrated prior to hybridization. Sections were
hybridized overnight at 50�C with hybridization solution
containing 50% formamide, 10% dextran sulfate,
4 9 SSC, 0.4 mg ml-1 tRNA and 400 ng probe diluted in
100 ll 50% formamide. Following three washing steps
with 0.29 SSC at 50�C for 1 h each, slides were incu-
bated with 10 lg ml-1 RNase A (Sigma–Aldrich, St.
Louis, MO, USA) at 37�C for 30 min, followed by
washing with PBS for 10 min. Sections were blocked for
45 min in Blocking Reagent (Roche) at RT, followed by a
15 min wash. Slides were incubated in BSA solution (1%
BSA, 100 mM Tris pH 7.5, 150 mM NaCl and 0.3%
Triton X-100) for 15 min with mild agitation, followed by
incubation with anti-digoxigenin-AP Fab Fragments
(Roche) (diluted 1:500 in BSA solution) for 2 h and then
washed 3 9 10 min in TBS (100 mM Tris pH 7.5,
150 mM NaCl). Sections were then incubated 2 9 5 min
in detection solution (100 mM Tris pH 9.5, 50 mM
MgCl2, 100 mM NaCl) prior to overnight staining in
detection solution (without MgCl2) containing NBT/BCIP
(Roche). The staining reaction was stopped by washing
the slides with water. Sections were mounted in Vecta-
Mount (Vector Laboratories) and recorded under a
microscope (DMLB, Leica, Germany) using a digital
camera (NikonDs-Fs-fi1).
Results
Developmental morphology of garlic inflorescence
Developmental studies of growing garlic plants in Israel
reveal that florogenesis progresses in several sequential
stages, as follows:
1. The vegetative meristem produces leaf primordia
during the first 7–8 weeks after planting (Fig. 1a).
2. After differentiation of the first 10 leaves, the transi-
tion from vegetative to reproductive stage and initia-
tion of the inflorescence meristem occur (Fig. 1b).
3. Flower primordia are initiated in the inflorescence
meristem (Fig. 1b, c).
4. Differentiation of individual flowers, leafy bracts and
vegetative buds (topsets). Flowers and flower primor-
dia at various stages of differentiation are present
inside the bulb (Fig. 1d).
5. Scape elongates, flowers are differentiated,
26–28 weeks after planting (Fig. 1e).
6. Gametogenesis and anthesis occur ca. 30 weeks after
planting (Fig. 1f, g).
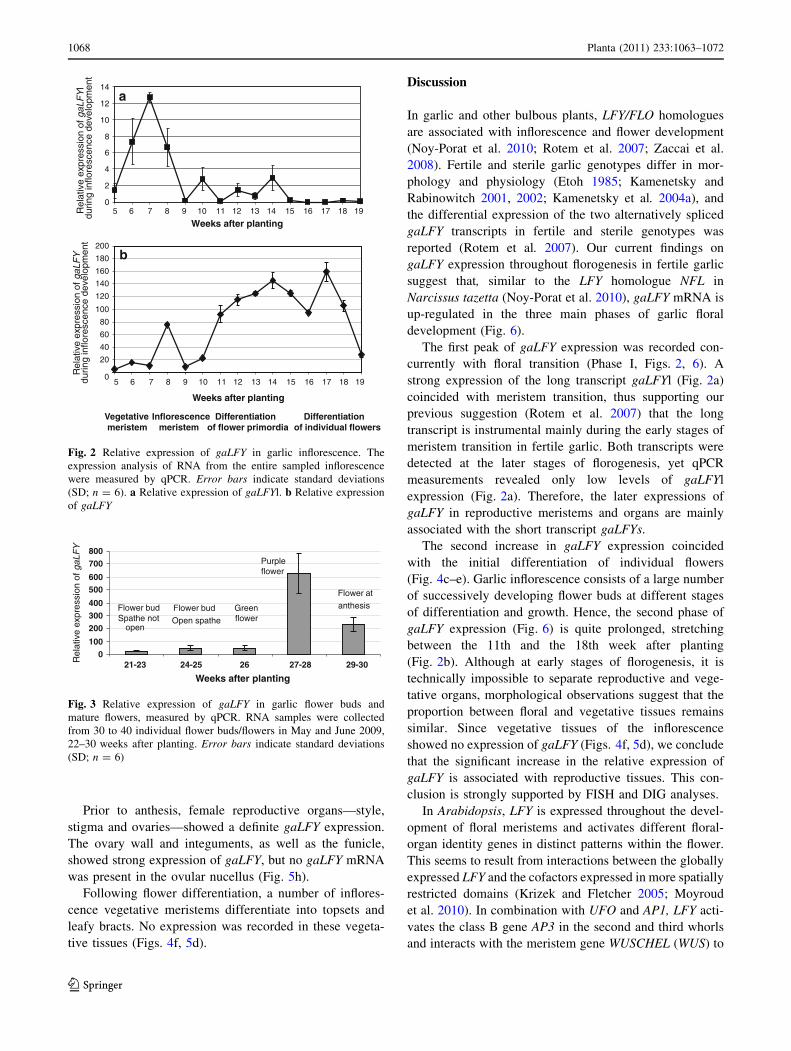
Quantification of gaLFY expression by qPCR
Microscope studies of the developing inflorescence were
accompanied by quantitative estimates of the relative
expression of total gaLFY and the long transcript gaLFYl,
using qPCR (Fig. 2). In general, the relative expression of
the gaLFYl was rather low throughout floral development,
when compared with the total gaLFY expression levels.
However, a sharp increase in gaLFYl expression was
recorded during the transition of the apical meristem from
the vegetative to the reproductive stage and at the begin-
ning of inflorescence differentiation. This peak declined
thereafter (Fig. 2a).
Between initiation and anthesis, total gaLFY is up-reg-
ulated at least three times (Figs. 2b, 3). The first peak
coincided with meristem transition to the reproductive
stage and initial differentiation of the inflorescence; the
second with differentiation of the individual flowers; and
the third with differentiation of anthers and gynoecia. It
should be noted that early in January (7–9 weeks after
planting)—during initial stages of the inflorescence
development—a sharp increase in gaLFYl expression
overlap the first peak of total gaLFY.
In individual flowers, gradual increase in gaLFY
expression was recorded from differentiation to maturation
(Fig. 3). At spathe break, flower buds are still closed,
anthers and ovaries are immature and gametophytes’ dif-
ferentiation continues until anthesis (Fig. 1e–g). During
1066 Planta (2011) 233:1063–1072
123

this period, flowers elongate and both perianth lobes and
anthers change color from green to purple. A 600-fold
increase in gaLFY expression was recorded during flower
maturation, compared with the values measured in vege-
tative tissues and sevenfold higher than in previous
developmental stage of flower buds. A few days later, at
anthesis, floral gaLFY expression dropped considerably
(Fig. 3).
Spatial expression of gaLFY in differentiating
inflorescence
In situ hybridization analysis of the vegetative apical
meristems at the fourth leaf stage of development, showed
a very weak fluorescence signal of gaLFY expression
(Fig. 4a). gaLFY expression was first recorded immediately
after the transition and at the early stages of inflorescence
differentiation, in the receptacle, inflorescence meristem
and flower primordia prior to flower differentiation
(Fig. 4b, c, d, f). At this stage, flower primordia consist of
small and dense meristematic cells with rather big nuclei
(Fig. 4g). DIG-labeled in situ hybridization revealed a
gaLFY expression in the cytoplasm of the outer cell layers
of flower primordia (Fig. 4g) concomitantly with the dif-
ferentiation of the first flower organs—perianth lobes and
anthers (Fig. 4d, e, h, i). Later, gaLFY expression became
visible in the pedicel (Fig. 4e, j).
Expression of gaLFY during the differentiation
of individual flowers and topsets
Following the differentiation of the perianth lobes and
anthers, the expression of gaLFY weakens (Fig. 5a, b),
and increases again with the differentiation of the anthers
and gynoecium (Fig. 5c). After spathe’s break, anthers
grow in size and change color from green to purple
(Fig. 1f, g). Strong expression of gaLFY was recorded in
the stamens’ filaments of the purple anthers, just before
pollen shedding (Fig. 5f, g), but not in pollen grains
(Fig. 5e–g).
Fig. 1 Stages of reproductive development in garlic accessions
#3028. a Vegetative meristem (vm) produces leaf primordia (lp),
6 weeks after planting. Bar 0.5 mm. b Inflorescence meristem (im),
9 weeks after planting. Differentiation of first flower meristems is
visible. Spathe (sp) removed. Bar 0.5 mm. c Inflorescence meristem
produces flower meristems (fm), 12 weeks after planting. Spathe (sp)
removed. Bar 0.3 mm. d Flower primordia at various stages of
differentiation (from 1 to 5) and vegetative topsets (arrows) can be
found simultaneously in the developing inflorescence within the bulb.
Bar 0.5 mm. e Flowers (f) and leaf bracts (lb) are formed, but spathe
is still not open, 25 weeks after planting. In this picture, spathe is
removed. Bar 0.5 cm. f Individual flower prior to anthesis, with
developing anthers (an) and gynoecium. Ovule (ov) is visible inside
carpels. Bar = 0.1 cm. g Anthesis, 30 weeks after planting. Game-
togenesis occurs prior to anthesis of individual flowers. Bar 1 cm
Planta (2011) 233:1063–1072 1067
123
Prior to anthesis, female reproductive organs—style,
stigma and ovaries—showed a definite gaLFY expression.
The ovary wall and integuments, as well as the funicle,
showed strong expression of gaLFY, but no gaLFY mRNA
was present in the ovular nucellus (Fig. 5h).
Following flower differentiation, a number of inflores-
cence vegetative meristems differentiate into topsets and
leafy bracts. No expression was recorded in these vegeta-
tive tissues (Figs. 4f, 5d).
Discussion
In garlic and other bulbous plants, LFY/FLO homologues
are associated with inflorescence and flower development
(Noy-Porat et al. 2010; Rotem et al. 2007; Zaccai et al.
2008). Fertile and sterile garlic genotypes differ in mor-
phology and physiology (Etoh 1985; Kamenetsky and
Rabinowitch 2001, 2002; Kamenetsky et al. 2004a), and
the differential expression of the two alternatively spliced
gaLFY transcripts in fertile and sterile genotypes was
reported (Rotem et al. 2007). Our current findings on
gaLFY expression throughout florogenesis in fertile garlic
suggest that, similar to the LFY homologue NFL in
Narcissus tazetta (Noy-Porat et al. 2010), gaLFY mRNA is
up-regulated in the three main phases of garlic floral
development (Fig. 6).
The first peak of gaLFY expression was recorded con-
currently with floral transition (Phase I, Figs. 2, 6). A
strong expression of the long transcript gaLFYl (Fig. 2a)
coincided with meristem transition, thus supporting our
previous suggestion (Rotem et al. 2007) that the long
transcript is instrumental mainly during the early stages of
meristem transition in fertile garlic. Both transcripts were
detected at the later stages of florogenesis, yet qPCR
measurements revealed only low levels of gaLFYl
expression (Fig. 2a). Therefore, the later expressions of
gaLFY in reproductive meristems and organs are mainly
associated with the short transcript gaLFYs.
The second increase in gaLFY expression coincided
with the initial differentiation of individual flowers
(Fig. 4c–e). Garlic inflorescence consists of a large number
of successively developing flower buds at different stages
of differentiation and growth. Hence, the second phase of
gaLFY expression (Fig. 6) is quite prolonged, stretching
between the 11th and the 18th week after planting
(Fig. 2b). Although at early stages of florogenesis, it is
technically impossible to separate reproductive and vege-
tative organs, morphological observations suggest that the
proportion between floral and vegetative tissues remains
similar. Since vegetative tissues of the inflorescence
showed no expression of gaLFY (Figs. 4f, 5d), we conclude
that the significant increase in the relative expression of
gaLFY is associated with reproductive tissues. This con-
clusion is strongly supported by FISH and DIG analyses.
In Arabidopsis, LFY is expressed throughout the devel-
opment of floral meristems and activates different floral-
organ identity genes in distinct patterns within the flower.
This seems to result from interactions between the globally
expressed LFY and the cofactors expressed in more spatially
restricted domains (Krizek and Fletcher 2005; Moyroud
et al. 2010). In combination with UFO and AP1, LFY acti-
vates the class B gene AP3 in the second and third whorls
and interacts with the meristem gene WUSCHEL (WUS) to
Vegetativemeristem
Differentiationof flower primordia
Differentiation of individual flowers
Inflorescencemeristem
0
20
40
60
80
100
120
140
160
180
200
5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
Rel
ativ
e ex
pres
sion
of g
aLF
Ydu
ring
inflo
resc
ence
dev
elop
men
t
Weeks after planting
0
2
4
6
8
10
12
14R
elat
ive
expr
essi
on o
f gaL
FY
l du
ring
inflo
resc
ence
dev
elop
men
t
5 6 7 8 9 10 11 12 13 14 15 16 17 18 19
Weeks after planting
a
b
Fig. 2 Relative expression of gaLFY in garlic inflorescence. The
expression analysis of RNA from the entire sampled inflorescence
were measured by qPCR. Error bars indicate standard deviations
(SD; n = 6). a Relative expression of gaLFYl. b Relative expression
of gaLFY
0100
200300400
500600
700800
Flower budSpathe not
open
Flower bud
Open spathe
Green flower
Purple flower
Flower at
anthesis
Rel
ativ
e ex
pres
sion
of g
aLF
Y
21-23 24-25 26 27-28 29-30
Weeks after planting
Fig. 3 Relative expression of gaLFY in garlic flower buds and
mature flowers, measured by qPCR. RNA samples were collected
from 30 to 40 individual flower buds/flowers in May and June 2009,
22–30 weeks after planting. Error bars indicate standard deviations
(SD; n = 6)
1068 Planta (2011) 233:1063–1072
123

turn on AG expression in the inner two whorls. In petunia,
the LFY homologue ALF actively regulates a wide spectrum
of B-, C-, D- and E-type organ identity genes (Souer et al.
2008). Similarly, in garlic, gaLFY is strongly expressed in
differentiating flower primordia, but is down-regulated
when most flowers are formed (Fig. 5b). These results are in
agreement with previously reported data from many plant
species such as, violet (Jonopsidium acaule), wheat
(Triticum spp.) and Monterey Pine (Pinus radiate), all of
which show a clear expression of LFY/FLO homologues
during flower initiation and differentiation (Mouradov et al.
1998; Shitsukawa et al. 2006; Shu et al. 2000).
The third peak of gaLFY expression was recorded prior
to anthesis, in developing stamen and pistils (Fig. 3). The
expression of gaLFY was visualized in stamen primordia
(Fig. 4j) and filaments, but not in mature anther (Fig. 5e–
g). It is therefore concluded that gaLFY is mainly active
at the early stages of stamen development. Expression
was also detected in most parts of the gynoecium,
including style, ovary, inner and outer integuments and
funicle but not in the ovular nucellus (Fig. 5e–h). The
results indicate a general role for gaLFY in gynoecium
development.
It is suggested that similar to Arabidopsis LFY, gaLFY
plays a key role in the early stages of garlic florogenesis:
floral initiation and the differentiation of individual flowers
and floral organs. However, unlike LFY, gaLFY may also
act as a regulator of gametogenesis. Similarly, in radish a
LFY homologue RsLFY is expressed in primordia of sepals,
petals, stamens and gynoecium, and is also detected in
ovule and developing seeds (Oshima and Nomura 2008).
GmLFY, a soybean LFY homologue, was detected in
developing seeds, suggesting that it plays an essential
regulatory role in seed development (Meng et al. 2007).
Our findings conclusively show that the temporal
expression of gaLFY is strongly associated with the
developmental stages of flowers, but not with the vegeta-
tive meristems, topsets and the inflorescence’s leafy bracts
(Fig. 5d). In other plant species, mutations or down-regu-
lation of LFY and its homologues was reported to cause
reversion in flower determination with the consequent
development of vegetative shoots (Coen et al. 1990;
Molinero-Rosales et al. 1999; Wang et al. 2004; Weigel
et al. 1992).
In the annual Arabidopsis, flowering is irreversible
(Amasino 2010); however, reversion to vegetative growth
Fig. 4 gaLFY expression in the vegetative meristem and during the
early florogenesis stages in garlic. a–e FISH, as observed by confocal
microscopy. f–j DIG-labeled in situ hybridization, as observed by
light microscopy. Bars a–e 200 lm, f, h–j 100 lm, g = 50 lm.
a Vegetative meristem (vm) in plants at the fourth foliage leaf stage of
development, November 2008. b Meristem transition and initiation of
the inflorescence. Note the high level of fluorescence signal in the
receptacle (rc), inflorescence meristem (im) and flower primordia (fp).
c Differentiation of flower primordia (fp). A weaker signal in the
receptacle (rc). d Early developmental stages of flower buds (stages
1–3). gaLFY expression is evident in the center of the reproductive
meristem at stage 1; in the apical and lateral parts of the primordium
at stage 2, and in flower organ primordia at stage 3. e Early
developmental stages of flower buds (stage 4). Perianth lobes (pr) and
anthers (an) are visible. f Initiation of flower primordia in the
inflorescence, comparable to b and c. Note absence of gaLFYexpression in the leafy bracts (lb). g Reproductive meristem prior to
differentiation of flower organs, comparable to stage 1 in d. gaLFYexpression is visible in the cytoplasm (blue staining). Note the small
meristematic cells with large nuclei. h Initial differentiation of
individual flower. The apical and lateral parts of primordium are
stained blue, comparable with stage 2 in d. i Flower with perianth and
anther primordia, comparable with stage 3 in d. j Individual flower:
perianth lobes (pr) and anthers (an) are differentiated, comparable
with stage 4 in e
Planta (2011) 233:1063–1072 1069
123

is common in quite a few plant species when inductive
environmental signals is not maintained (Zeevaart 1985;
Tooke et al. 2005). In garlic, topsets’ formation in the
inflorescence can be considered as a reversion in flower
determination, yet the role gaLFY plays in this process, as
well as the question of environmental regulation deserve
further elucidation.
In this study, gaLFY mRNA levels of expression were
recorded during reproductive developmental stages, using
three methodologies, as well as primers and probes from
Fig. 5 gaLFY expression pattern during development of individual
flower. a–d FISH, as observed by confocal microscopy. e–h DIG-
labeled in situ hybridization, as observed by light microscopy Bars a–
d 200 lm, e–g 100 lm, h 50 lm. a Individual flower—differentiation
of perianth lobes (pl) and anthers (a). A strong gaLFY expression is
obvious in the differentiating organs. b Following differentiation of
perianth lobes (pl), gaLFY expression decreases. c Individual flower
with differentiating anthers. gaLFY is expressed in the filament (fi)
and in the seam between the perianth lobes and the pedicel (pd).
d Differentiation of topsets (t) and leafy bracts (l) in the inflorescence.
Note absence of gaLFY expression. e gaLFY expression in maturing
flowers. f gaLFY expression in the ovary (ov) and the filaments (fi)
during microgametogenesis. No gaLFY expression is visible in pollen
grains (p). g gaLFY expression in the style (s), ovary and ovules. No
expression is detectable in the center of ovule. h Close-up of the
ovarian wall (ow) shows a strong gaLFY expression in both the outer
and inner integuments (oi and ii) and in the funicle (f), but not in the
nucellus (n)
Fig. 6 Scheme of the three
main phases of gaLFYexpression during florogenesis
of fertile garlic. Initially, mRNA
is detected during meristem
transition from the vegetative to
the reproductive phase (phaseI). Following initiation of the
individual flower primordia,
gaLFY is down-regulated in the
inflorescence meristem and
expressed again during organ
differentiation in the individual
flowers (phase II). The third
peak is detected in anthers and
ovules of the fully differentiated
mature flowers (phase III)
1070 Planta (2011) 233:1063–1072
123

various fragments of the gene. The results support our
assumption that gaLFY is in fact involved in the regulation
of a number of florogenetic processes in garlic, from floral
transition to flower maturation and gametogenesis. Indeed,
the up- and down-regulation of gaLFY coincided with tis-
sue differentiation and various physiological stages of
garlic inflorescence.
Numerous reports suggest that LFY and its homologues
are multi-functional genes, possibly involved in a variety of
morphogenetic stages, and thus can be considered as a
master regulator of flower development (Albert et al. 2002;
Moyroud et al. 2010). Our results obtained from flowering
garlic support this wisdom for florogenesis. Further studies,
however, are required to elucidate the additional (up- and
downstream) molecular and genetic mechanisms involved
in garlic flowering and seed production, as well as general
aspects of florogenesis in herbaceous perennial plants.
Acknowledgments We thank E. Belausov, E. Shemesh and
H. Zemach, The Institute of Plant Science, Agricultural Research
Organization, The Volcani Center, for their assistance in microscopic
observations.
References
Albert VA, Oppenheimer DG, Lindqvist C (2002) Pleiotropy,
redundancy and the evolution of flowers. Trends Plant Sci
7:289–301
Amasino R (2010) Seasonal and developmental timing of flowering.
Plant J 61:1001–1013
Benlloch R, Berbel A, Serrano-Mislata A, Madueno F (2007) Floral
initiation and inflorescence architecture: a comparative view.
Ann Bot 100:659–676
Benschop M, Kamenetsky R, Le Nard M, Okubo H, De Hertogh AA
(2010) The global flower bulb industry: production, utilization,
research. Hort Rev 36:1–115
Bernier G, Perilleux C (2005) A physiological overview of the
genetics of flowering time control. Plant Biotech J 3:3–16
Blazquez MA, Weigel D (2000) Integration of floral inductive signals
in Arabidopsis. Nature 404:889–892
Cockram J, Jones H, Leigh FJ, O’Sullivan D, Powell W, Laurie DA,
Greenland AJ (2007) Control of flowering time in temperate
cereals: genes, domestication, and sustainable productivity.
J Exp Bot 58:231–1244
Coen ES, Romero JM, Doyle S, Elliot R, Murphy G, Carpenter R
(1990) Floricaula: a homeotic gene required for flower devel-
opment in Antirrhinum majus. Cell 63:1311–1322
Etoh T (1985) Studies on the sterility in garlic, Allium sativum L.
Memoirs of the Faculty of Agriculture, Kagoshima University,
vol 21. pp 77–132
Etoh T, Simon PW (2002) Diversity, fertility and seed production of
garlic. In: Rabinowitch HD, Currah L (eds) Allium crop
sciences: recent advances. CAB International, Wallingford,
pp 101–117
Gocal GF, King RW, Blundell CA, Schwartz OM, Andersen CH,
Weigel D (2001) Evolution of floral meristem identity genes:
analysis of Lolium temulentum genes related to APETALA1 and
LEAFY of Arabidopsis. Plant Physiol 125:1788–1801
Gottlieb Y, Ghanim M, Chiel E, Gerling D, Portnoy V, Steinberg S,
Tzuri G, Horowitz AR, Belausov E, Mozes-Daube N, Kontsed-
alov S, Gershon M, Gal S, Katzir N, Zchori-Fein E (2006)
Identification and localization of a Rickettsia sp. in Bemisia
tabaci (Homoptera: Aleyrodidae). Appl Environ Microbiol
72:3646–3652
Jenderek MM, Hannan RM (2004) Variation in reproductive charac-
teristics and seed production in the USDA garlic germplasm
collection. HortScience 39:485–488
Jenderek MM, Zewdie Y (2005) Within- and between-family
variability for important bulb and plant traits among sexually
derived progenies of garlic. HortScience 40:1234–1236
Johansen DA (1940) Plant microtechnique. McGraw-Hill, New York
Kamenetsky R (2007) Garlic: Botany and Horticulture. Hort Rev
33:123–172
Kamenetsky R, Rabinowitch HD (2001) Floral development in
bolting garlic. Sex Plant Reprod 13:235–241
Kamenetsky R, Rabinowitch HD (2002) Florogenesis. In: Rabino-
witch HD, Currah L (eds) Allium crop sciences: recent advances.
CAB International, Wallingford, pp 31–58
Kamenetsky R, London Shafir I, Baizerman M, Khassanov F, Kik C,
Rabinowitch HD (2004a) Garlic (Allium sativum L.) and its wild
relatives from Central Asia: evaluation for fertility potential.
Acta Hort 637:83–91
Kamenetsky R, London Shafir I, Zemah H, Barzilay A, Rabinowitch
HD (2004b) Environmental control of garlic growth and
florogenesis. J Am Soc Hort Sci 129:144–151
Kamenetsky R, London Shafir I, Khassanov F, Kik C, van Heusden
AW, Vrielink-van Ginkel M, Burger-Meijer K, Auger J, Arnault
I, Rabinowitch HD (2005) Diversity in fertility potential and
organo-sulphur compounds among garlics from Central Asia.
Biodivers Conserv 14:281–295
Krizek BA, Fletcher JC (2005) Molecular mechanisms of flower
development: an armchair guide. Nat Rev Genet 6:688–698
Kyozuka J, Konishi S, Nemoto K, Izawa T, Shimamoto K (1998)
Down-regulation of RFL, the FLO/LFY homolog of rice,
accompanied with panicle branch initiation. Proc Natl Acad
Sci 95:1979–1982
Lamb RS, Hill TA, Tan OK, Irish VF (2002) Regulation of
APETALA3 floral homeotic gene expression by meristem
identity genes. Development 129:2079–2086
Meng Q, Zhang C, Huang F, Gai J, Yu D (2007) Molecular cloning
and characterization of a LEAFY-like gene highly expressed in
developing soybean seeds. Seed Sci Res 17:297–302
Molinero-Rosales N, Jamilena M, Zurita S, Gomez P, Capel J,
Lozano R (1999) FALSIFLORA, the tomato orthologue of
FLORICAULA and LEAFY, controls flowering time and floral
meristem identity. Plant J 20:685–693
Moon J, Lee H, Kim M, Lee I (2005) Analysis of flowering pathway
integrators in Arabidopsis. Plant Cell Physiol 46:292–299
Mouradov A, Glassick T, Hamdorf B, Murphy L, Fowler B, Marla S,
Teasdale RD (1998) NEEDLY, a Pinus radiate ortholog of
FLORICAULA/LEAFY genes, expressed in both reproductive
and vegetative meristems. Proc Natl Acad Sci 95:6537–6542
Mouradov A, Cremer F, Coupland G (2002) Control of flowering
time: interacting pathways as a basis for diversity. Plant Cell
14:S111–S130
Moyroud E, Kusters E, Monniaux M, Koes R, Parcy F (2010) LEAFY
blossoms. Trends Plant Sci 15:346–352
Nilsson O, Lee I, Blazquez MA, Weigel D (1998) Flowering-time
genes modulate the response to LEAFY activity. Genetics
150:403–410
Noy-Porat T, Kamenetsky R, Eshel A, Flaishman MA (2010)
Temporal and spatial expression patterns of the LEAFY homo-
logue NLF during florogenesis in Narcissus tazetta. Plant Sci
178:105–113
Planta (2011) 233:1063–1072 1071
123

Oshima S, Nomura K (2008) RsLFY, a LEAFY homologue gene in
radish (Raphanus sativus), is continuously expressed in vegeta-
tive, reproductive and seed development. Plant Biotechnol
25:579–582
Pfaffl MW (2001) A new mathematical model for relative quantifi-
cation in real-time RT-PCR. Nucleic Acids Res 29:e45
Pfaffl MW, Horgan GW, Dempfle L (2002) Relative expression
software tool (REST�) for group-wise comparison and statistical
analysis of relative expression results in real-time PCR. Nucleic
Acids Res 30:e36
Ratcliffe OJ (1999) Separation of shoot and floral identity in
Arabidopsis. Development 126:1109–1120
Rotem N, Shemesh E, Peretz Y, Akad F, Edelbaum O, Rabinowitch
HD, Sela I, Kamenetsky R (2007) Reproductive development
and phenotypic differences in garlic are associated with expres-
sion and splicing of LEAFY homologue gaLFY. J Exp Bot
58:1133–1141
Shemesh E, Scholten O, Rabinowitch HD, Kamenetsky R (2008)
Unlocking variability: inherent variation and developmental
traits of garlic plants originated from sexual reproduction. Planta
227:1013–1024
Shitsukawa N, Takagishi A, Ikari C, Takumi S, Murai K (2006) WFL,
a wheat FLORICAULA/LEAFY ortholog, is associated with
spikelet formation as lateral branch of the inflorescence meri-
stem. Genes Genet Syst 81:13–20
Shu GP, Amaral W, Hileman LC, Baum DA (2000) LEAFY and the
evolution of rosette flowering in violet cress (Ionopsidiumacaule, Brassicaceae). Am J Bot 87:634–641
Souer E, Rebocho AB, Bliek M, Kusters E, de Bruin RAM, Koes R
(2008) Patterning of inflorescences and flowers by the F-Box
protein DOUBLE TOP and the LEAFY Homolog ABERRANT
LEAF AND FLOWER of Petunia. Plant Cell 20:2033–2048
Szymkowiak EJ, Irish VE (2006) JOINTLESS suppresses sympodial
identity in inflorescence meristems of tomato. Planta 223:646–658
Tooke F, Ordidge M, Chiurugwi T, Battey N (2005) Mechanisms and
function of flower and inflorescence reversion. J Exp Bot
56:2587–2599
Vijayraghavan U, Prasad K, Meyerowitz E (2005) Specification and
maintenance of the floral meristem: interactions between posi-
tively acting promoters of flowering and negative regulators.
Curr Sci 89:1835–1843
Wang CN, Moller M, Cronk QCB (2004) Altered expression of
GFLO, the Gesneriaceae homologue of FLORICAULA/LEAFY,
is associated with the transition to bulbil formation in Titano-trichum oldhamii. Dev Genes Evol 3:122–127
Weigel D, Alvarez J, Smyth DR, Yanofsky MF, Meyerowitz EM
(1992) LEAFY controls floral meristem identity in Arabidopsis.
Cell 69:843–859
Zaccai M, Mazor I, Weingarten-Kenan E, Ram A (2008) Vernaliza-
tion and floral transition in the Madonna lily (Lilium candidum).
Abstracts of the Xth international symposium on flower bulbs
and herbaceous perennials. Lisse, The Netherlands, p 38
Zeevaart JAD (1985) Perilla. In: Halevy AH (ed) Handbook, CRC of
flowering. CRC Press, Raton Boca, pp 239–252
Zik M, Irish VF (2003) Flower development: initiation, differentia-
tion, and diversification. Ann Rev Cell Dev Biol 19:119–140
1072 Planta (2011) 233:1063–1072
123




















