
On the receptivity of the by-pass transition to the length scale of the outer stream turbulence
Upload
worldagroforestryCategory
view
3download
0
Annals of Botany 86: 133±148, 2000doi:10.1006/anbo.2000.1170, available online at http://www.idealibrary.com on
Floral Structure, Stigma Receptivity and Pollen Viability in Relation to Protandryand Self-incompatibility in Silky Oak (Grevillea robusta A. Cunn.)
A. KALINGANIRE*{, C. E. HARWOOD{, M. U. SLEE{ and A. J. SIMONS}
{Department of Forestry, Australian National University, ACT 0200, Australia, {CSIRO Forestry and ForestProducts, PO Box E4008 Kingston, ACT 2604, Australia and }International Centre for Research in Agroforestry,
PO Box 30677 Nairobi, Kenya
Received: 24 December 1999 Returned for revision: 11 February 2000 Accepted: 24 March 2000
has receive
0305-7364/0
* For corrPO Box E400e-mail Antoin
The reproductive biology of Grevillea robusta growing under exotic conditions in Kenya and Australia is reported.The species showed both protandry and a self-incompatibility mechanism. The stigma was wet and papillate with adistinct groove in the middle. The anthers dehisced prior to anthesis, when the perianth opened. Stigmatic receptivitybegan 1 d after anthesis, with the greatest pollen germination rates and longest pollen tubes obtained 2 d afteranthesis. Nectar secretion commenced with pollen dehiscence and was abundant at anthesis. Most stigmatic groovesopened widely 1±2 d after anthesis and stigmas showed taller papillae and abundant secretion. Controlled pollina-tions gave a greater fruit set from cross-pollination (5.9% in April and 17.5% in July) than open-pollination (0.1%in April and 3.3% in July). No fruit set from self-pollination was obtained in April, and very few fruit set forgeitonogamous (two out of 1622; 0.1%) or for autogamous (one out of 2707 ¯owers; 0.04%) pollination treatmentsin July. Following self-pollination, growth of pollen tubes was poorer than in other treatments, and was generallyarrested in the upper style. Cross-pollinated ¯owers produced normal and straight pollen tubes, while self-pollentubes had growth abnormalities. Most of the open-pollinated ¯owers were found without pollen or with only self-pollen on their stigmas indicating that the amount of cross-pollen reaching the stigma under open-pollination may bea factor limiting seed production. Flowers shed soon after the fertilization phase were those with ungerminated pollenor no pollen. Although a very low rate of sel®ng may occur, G. robusta presents a self-incompatibility system andallogamy is its primary breeding behaviour. # 2000 Annals of Botany Company
Key words: Grevillea robusta, silky oak, Proteaceae, protandry, controlled pollinations, receptivity, pollen-tubegrowth, self-incompatibility, pollination.
INTRODUCTION
Grevillea robusta A. Cunn. ex R. Br. (silky oak) is a treebelonging to the dicotyledonous plant family Proteaceae,subfamily Grevilleoideae in the tribe Grevilleeae (McGil-livray, 1993). The species has a restricted natural range onthe east coast of Australia from latitude 228500 S to 308100 S(Harwood, 1992; McGillivray, 1993). Grevillea robusta hasbeen introduced to many countries in south and centralAmerica, south Asia, and in the highlands of eastern andcentral Africa (Harwood, 1989) where it is very commonand popular for farm plantings. It is grown as a shade treefor tea and co�ee plantations, in agroforestry plantings forfuel wood and timber and also as an ornamental tree.
Detailed studies of the reproductive biology of theProteaceae have been mainly limited to species of thegenus Banksia L.f. (e.g. Carthew, 1993; Sedgley et al., 1993,1994; Goldingay and Carthew, 1998) and MacadamiaMuell. (e.g. Sedgley et al., 1985; Wallace et al., 1996) andrecently Dryandra (Matthews and Sedgley, 1998). GrevilleaR. Br. ex Knight, the largest genus in the family Proteaceae,
d less attention and existing information has
0/070133+16 $35.00/00
espondence at: CSIRO Forestry and Forest Products,8, Kingston ACT 2604, Australia. Fax �61 2 6281 8266,e.Kalinganire@�p.csiro.au
focused mainly on levels of self-incompatibility (Hersco-vitch and Martin, 1990; Harriss and Whelan, 1993; Ayreet al., 1994; Hermanutz et al., 1998; Hogbin et al., 1998).The most comprehensive work on the biology of G. robustais that of Brough (1933), who reported in detail itslife history, ¯oral morphology and seed development,and that of McGillivray (1993) who reported its taxonomy.Harwood (1992) and Kalinganire et al. (1996) havereported the ¯owering pattern and pollen vectors forG. robusta.
Proteaceous species are generally considered protandrous(see Johnson and Briggs, 1975; Collins and Rebelo, 1987;Ladd et al., 1998). The protandrous behaviour of G. robustahas been previously reported by Brough (1933) for plantedtrees in Sydney and by Venkata Rao (1971) for landraces inIndia; anthers dehisce prior to stigma receptivity and pollenis deposited onto the pollen presenter. In contrast, the studyof Owuor and Oduol (1992) concluded that in Kenya thespecies is protogynous. There have been no reported studiesof stigma development and pollen tube growth and viabilitybefore, during and after anthesis in G. robusta.
Although most species of Proteaceae are largely self-incompatible, there is some indication that several self-compatible species preferentially out cross (Goldingay and
Carthew, 1998). Grevillea shows highly variable breeding# 2000 Annals of Botany Company

lf
systems (Harriss and Whelan, 1993; Ayre et al., 1994;Hermanutz et al., 1998), although McGillivray (1993)stated that self-compatibility is probably predominant inthe genus. For G. robusta, Brough (1933) noted allogamyappeared to be its predominant behaviour, although someself-fertilization did occur. However, Owuor and Oduol(1992) found it to have an autogamous breeding system inexotic stands in Kenya with 63% fruit set from self-pollination and 69% from open-pollination. Using isozymestudies, Harwood et al. (1992) reported G. robusta to bepredominantly outcrossing in two natural populations. Thecontradictory results reported for the species to date suggesta need for more research to elucidate its breeding system.
The availability of seed is often a limiting factor forplanting programmes in most East African countries and anunderstanding of the factors a�ecting seed production hasimportant practical applications, for both genetic improve-ment and operational seed production. This study investi-gates important aspects of the reproductive biology ofG. robusta: protandrous behaviour including ¯ower struc-ture and development, and the self-incompatibility mech-anism dealing with pollen tube growth and fruit setfollowing self-, outcross- and open-pollination. Moreover,the study attempts to de®ne the temporal sequence ofstigma receptivity, the relationship of stigma receptivity to
134 Kalinganire et al.ÐProtandry and Se
pollen release and the degree of pollen viability.
MATERIALS AND METHODS
Plant material and study site
The study was conducted at two sites: Malava, westernKenya, from January 1996 to April 1997; and Canberra,southern Australia, in December 1997.
The Malava site (08280 N, 348510 E; 1600 m a.s.l.) is partof a ®eld trial site maintained by the International Centre
for Research in Agroforestry (ICRAF) and Kenya ForestryTABLE 1. Details of the natural provenances of Grevillea ro(western
Seedlot no.CSIRO
seedlot no.No. of families
included Provenanc
1 17694 6 Porters Ga2 15872 4 Linville3 17693 8 Bunya Mo4 17610 4 Mudgereeb5 17699 5 Albert Riv6 15873 4 Emu Vale7 17612 5 Nimbin8 17185 1 Woodenbo9 17613 3 Grevillea10 17611 4 Tyalgum11 17614 4 Duck Cree12 17616 2 Paddys Fla13 15615 3 Bottle Cre14 17617 4 Mummulg15 17618 4 Rappville16 17620 10 Mann Riv17 17619 4 Fine Flow18 17621 4 McPherso19 17622 11 Boyd Rive
Research Institute. The site has a slope of around 5% witha south-westerly aspect. The adjacent vegetation is mainlynatural evergreen forest with some cleared farmland nearby.The mean annual rainfall (years 1990 to 1995) is 2413 mmwith a bimodal distribution. Rain falls in every month ofthe year, with May and September the wettest months.During the study period, the monthly means of dailytemperature maxima ranged from 278C (August) to 338C(March) and corresponding minima from 148C (July) to168C (November). The monthly means of daily relativehumidity at 0700 h ranged from 76% (November) to 97%(July). The soil is classi®ed as very-®ne, kaolinitic, isohyper-thermic, udic kandic ustalf, with a pH of 6.2 (Matungulu,1994).
A trial of 90 open-pollinated G. robusta families from19 Australian provenances was planted on the site in April1991, in a design which maintained both family andprovenance identity. Provenance details (Table 1) spannedthe range of the species' natural distribution. The trial,within 1.1 ha, used a randomized complete block designwith seven complete replicates and single-tree plots. Spacingbetween trees was 4 m � 4 m. At the time of the study,mean height of trees was around 14 m, mean diameter atbreast height 18 cm, and overall survival exceeded 95%.
The plant material used for the timing of stigmaticexudates, measurements of groove and papillae size and foran in-vitro pollen germination test, as part of the ¯oralreceptive study, was collected on planted trees growing inthe Australian National Botanic Gardens (ANBG), atCanberra. Preliminary observations had indicated theduration of ¯owering in Canberra was very similar to thatin Kenya and the ambient temperature in the two locationsat ¯owering was also similar. Thus the Canberra observa-tions should be applicable to Kenya. At Canberrathe average daily maximum temperature for December,
-incompatibility of Grevillea robusta
the ¯owering and study period, was 26.28C and the
busta planted in the provenance±progeny trial at MalavaKenya)
e nameAlt(m)
Lat(8S)
Long(8E)
p 650 26 45 151 30140 26 29 152 16
untains 1000 26 54 151 37a 20 28 5 153 22er 280 28 16 153 6
550 28 14 152 1750 28 38 153 13
ng 200 28 26 152 45180 28 26 152 4780 28 22 153 11
k 160 28 43 152 33t 180 28 44 152 26ek 200 28 48 152 39um 100 28 50 152 49
40 29 7 152 58er 60 29 24 152 29er 60 29 33 152 29ns 40 29 48 152 54r 200 29 53 152 27

pollen.
conflorescenceor raceme
mainraceme
primarypeduncle
Len
gth
of
infl
ores
cen
ce
length of raceme
FIG. 1. A four-branched in¯orescence of Grevillea robusta shows
lf-incompatibility of Grevillea robusta 135
corresponding average daily minimum was 11.28C. Theaverage daily relative humidity recorded during the studyperiod at 0700 h was 62%. The trees were 35-years-old and¯owering heavily at the time of the study. The naturalprovenance origin of the trees is uncertain; they originatedfrom seeds collected from one or more mother trees in aplanted stand in Western Australia.
Floral structure and development
In¯orescences of G. robusta are terminal and may formup to six branches each bearing hermaphroditic ¯owersgrouped into racemes (Fig. 1) (Johnson and Briggs, 1975;McGillivray, 1993). In this study, each individual branch isreferred to as a raceme. Floral development in G. robustahas been described by Brough (1933) and the main stagesare illustrated in Fig. 2. Each ¯ower of G. robusta containstwo ovules (McGillivray, 1993; Hermanutz et al., 1998).
Flower structure and development were studied at theMalava site. In¯orescence length, number of racemes perin¯orescence and raceme length were recorded. Dailyobservations began on 25 Mar. 1996 and ended on 26Jul. 1996. Basic stages of ¯ower development were de®nedby detailed examination of ®ve trees. Flower parameterswere measured on 43 trees ( four in¯orescences per tree andfour racemes per in¯orescence sampled) representing 16families in six provenances (Emu Vale, Mudgereeba, AlbertRiver, Nimbin, Rappville, Mann River; details in Table 1)distributed across the species' natural range. Style lengthwas measured on ®ve ¯owers selected at random aroundeach raceme. Only the length of the style was recorded asthe pedicel does not change signi®cantly during ¯oraldevelopment. The number of ¯owers per raceme wererecorded at all stages. Open ¯owers were counted and
Kalinganire et al.ÐProtandry and Se
recorded as were the number of ¯owers with nectar.
Time (no. of days from 0d):Stage:
10 20 31(i) (ii) (iii)
FIG. 2. Individual ¯ower development stages of Grevillea robusta and the minitiation. The main stages represented in sequence are from left to righ
(iv and v) two stages of early looping; (vi) advanced loopin
Furthermore, observations on capped ¯owers were madeon 12 trees (three in¯orescences per tree and two racemesper in¯orescence sampled). Capped ¯owers are individual¯owers with the staminal structure remaining in place overthe stigma holding the ¯ower's pollen and excluding other
di�erent con¯orescences or racemes.
Fruit set at di�erent stages of stigmatic development
Timing of stigma receptivity was investigated by assess-ment of fruit set following controlled pollination at Malava.Preliminary observations showed that cross-pollinationbetween unrelated trees produced fruit with viable seeds.Consequently, crosses were made between unrelated trees,
i.e. from di�erent families, for the experiments. Flowers33 35 38 39 54(viii)(vii)(vi)(v)(iv)
ean number of days to reach each ¯oral stage from ¯ower macroscopict: (i) ¯ower bud macroscopic initiation; (ii) early buds; (iii) late buds;g; (vii) pollen presentation (anthesis); and (viii) fruit set.

any pollen tubes were scored as ungerminated.
lf
were emasculated to remove self-pollen and isolated, thenhand pollinated.
The emasculation and isolation technique used was amodi®cation of the procedure described by Owuor andOduol (1992). Emasculation was carried out at the latelooping stage (Fig. 2; stage vi), approximately 48 h beforethe style elongated and was freed from the perianths. At thisstage pollen presenters are easily pulled out by hand orusing a pin, thus being separated from the perianths enclos-ing the anthers. The undehisced anthers were then removed.All other ¯owers were trimmed o� using ®ne scissors. Allgroups were emasculated at the same time.
In¯orescences were isolated from pollinators by the useof muslin bags. Crosses were carried out by applyingpollen-laden stigmas to stigmas of emasculated ¯owers.Bags were retained until the end of the receptive phase, i.e.the fall of the perianths, about 20 d after cross-pollination.
For this experiment, three trees from three di�erentfamilies were used. Flowers from the main raceme for onein¯orescence per tree and per treatment were tagged. Flowerstages and the corresponding number of ¯owers (given inbrackets) assessed were: ÿ2 d (170); ÿ1 d (141) beforeanthesis; at 0 d (132) or anthesis; and 1 d (159); 2 d (127);3 d (156); 4 d (132); and 5 d (149) after anthesis. Thenumber of ¯owers used for each treatment ranged from 21 to64 ¯owers per tree. Di�erent sources of pollen were appliedon the three trees, thus avoiding any chance of incompat-
136 Kalinganire et al.ÐProtandry and Se
ibility. Fruit set was recorded 60 d after pollination.
Pollen germination at di�erent stages of stigmaticdevelopment
To determine the ¯oral stage which corresponds tooptimum stigmatic receptivity, pollen germination onstigmas and subsequent pollen tube growth at various¯oral stages was assessed. Two 35-year-old trees growing inthe ANBG, Canberra, were used as mother trees for thistrial. One raceme per tree was tagged for each stigmaticdevelopment stage. Flower stages assessed were: ÿ2 d priorto anthesis; ÿ1 d prior to anthesis; at 0 d or anthesis; and1 d; 2 d; 3 d; and 4 d after anthesis. In this study anthesis isde®ned as the time when the perianth opens and pollen ispresented to ¯oral visitors. Flowers were emasculatedapprox. 2 d before anthesis (Fig. 2; stage vi) and isolatedfrom visitors using polyester pollination bags (bag type PBS10-1; PBS International, UK). Flowers for ÿ2 d, ÿ1 dprior to anthesis and on the day of anthesis (0 d) had theirself-pollen removed and were pollinated immediately withcross-pollen from two di�erent trees, whose relatedness isuncertain. For other stages the pollination time was referredto as the emasculation day, which corresponds approximat-ely to 2 d before anthesis, and ¯owers were cross-pollinatedas above. Emasculated ¯owers were cross-pollinated at theappropriate stage using pollen of fresh 0 d ¯owers fromdi�erent trees as above. The isolation bags were replacedimmediately after pollination. Pollen could be seen onstigmas after application.
At least 25 ¯owers per stage and per tree were harvested24 h after pollination and styles processed for ¯uorescence
microscopy for pollen germination and tube growth asfollows. Immediately after harvesting pistils were ®xed in`Carnoy' ®xative (absolute ethanol : chloroform:acetic acid;6 :3 :1) for at least 2 h and then transferred to 70% ethanolfor storage. Pistils were later cleared and softened withsodium hydroxide (0.8 M NaOH) for up to 1.5 h at 608C,rinsed in distilled water and then stained overnight withwater-soluble aniline blue in potassium phosphate (K3PO4)bu�er. This method causes callose in the pollen grains andtubes to be stained and to ¯uoresce brightly under a short-wave (UV) light. Pistils were then placed on a microscopeslide with a drop of 80% glycerol and squashed beneath acoverslip. Pollen tubes were examined by ¯uorescencemicroscopy and photographed or stored in the dark forfurther viewing. Samples were viewed through a NikonOptiphot epi¯uorescence microcope with a ¯uorescenesource.
Twenty-®ve ¯owers for each treatment on each tree wereexamined microscopically. A ¯ower with at least onegerminated pollen grain was scored as receptive. A pollengrain was considered germinated when the length of its tubewas more than the diameter of the pollen grain (Shivannaand Rangaswamy, 1992). Fluorescing pollen grains without
-incompatibility of Grevillea robusta
Timing of stigmatic exudates, and groove and papillae size
The stigmatic features of unmanipulated ¯owers atdi�erent stages were assessed and recorded. Flower stagesassessed were: ÿ2 d; ÿ1 d before anthesis; at 0 d oranthesis; and 1 d; 2 d; 3 d; and 4 d after anthesis. One35-year-old tree growing in the ANBG, Canberra, was usedfor this experiment. One in¯orescence per treatment wastagged and bagged as above. For stages ÿ2 d, ÿ1 d and0 d, ¯owers were identi®ed, tagged and immediatelycollected for scanning electron microscopy (SEM). Fortreatments 1 d, 2 d, 3 d, 4 d and 5 d after anthesis, on theday of anthesis (0 d), all ¯owers at looping stage wereclipped o� the raceme using ®ne scissors. The open ¯owersremained untouched for collection at the appropriate stage.Five fresh pistils were collected per stage for examination.All ¯owers from di�erent stages used in this experimentwere collected at the same time.
The timing of stigmatic exudation and associated grooveand papillae size and adhesion of pollen on the stigma weredetermined using SEM. Samples for SEM were preparedfollowing a cryo-SEM method (Craig and Beaton, 1996;R. Heady, pers. comm., 1998). This method allowed freshstigmatic exudates to be checked, which would have driedout using other methods. Fresh sample material wasattached to a mounting plate using tissue freezing mediumand rapidly frozen by plunging into liquid nitrogen slush atÿ2308C. The plate with attached sample was then insertedinto the preparation chamber of an Oxford CT1500 CryoPreparation System and slowly warmed to ÿ808C in orderto remove ice crystals on the surface of the specimen. Thefrozen sample was then coated with a 10 nm layer of goldand transferred to a cryo stage at ÿ1858C ®tted inside thechamber of a Cambridge Instruments S360 SEM. Theelectron optics system of the SEM was optimized for high
resolution, but with su�cient depth-of-®eld to enable the
straight pollen tubes were measured in each ®eld.
lf
entire selected image to be focused. This required a 30 mmdiameter ®nal aperture, a working distance of approx.18 mm, electron beam current of 80 pA and an acceleratingvoltage of 15 kV. The sample was maintained at ÿ1858Cthroughout the SEM viewing operation.
The papillae length, the pollen size and the width of thestigmatic groove opening at its widest point, half way alongits length, were estimated from micrographs of ®ve di�erentstigmas per stage using the `NIH Image Program' version1.57 (Public Domain Image Processing and AnalysisProgram, National Institute of Health, USA). However,groove and papillae size could not be measured withprecision as pollen and stigmatic secretions obstructed thereading for most stages. Estimates for groove width wererecorded for up to 2 d after anthesis, while other stageswere only checked for stigmatic ¯uids. Five stigmas wereobserved and at least 30 pollen grains were measured per¯oral stage.
In order to check for any e�ect of emasculation on thestigmatic receptivity, ®ve ¯owers were emasculated atapprox. 1 d before anthesis and ®ve other ¯owers on thesame in¯orescence were left unmanipulated. The pistils ofthese ten ¯owers were collected 24 h later and examined bySEM for stigmatic exudates, a probable indication of
Kalinganire et al.ÐProtandry and Se
stigmatic receptivity.
Duration of viability of pollen grains on the stigma
Pollen grains of G. robusta were tested for their capacityto germinate and produce a normal pollen tube afterexposure on the pollen presenter and stigmatic region under®eld conditions. Pollen was collected from ¯owers of thesame stage (age) from three trees at the ANBG, Canberra,at ÿ2 d, ÿ1 d, at 0 d (anthesis), and 1 d, 2 d, 3 d and 4 dafter anthesis.
Racemes of ¯owers at di�erent stages were identi®ed,tagged and isolated using polyester bags to avoid any pollencontamination by visitors until the pollen was collected forviability testing. On a tagged raceme, as di�erent ¯owerstages are present on a single raceme, only ¯owers of thesame stage following a given treatment were left and otherswere removed. Flowers at stages ÿ2 d, ÿ1 d and 0 d weretagged and pollen collected immediately. Pollen from¯owers at the same stage was mixed for a germination test.
For other stages, ¯owers were tagged on the day ofanthesis and then collected at the appropriate timedepending on the treatment. At least 100 ¯owers per stageand per tree were collected and pollen was removed fromthe stigmas using a ®ne sterile pin. The fresh pollen wasmixed in a vial and tested immediately for germination. Afresh pin was used for each stage.
Preliminary studies indicated that media containing onlysucrose might not result in optimum pollen germination. Inthe study 100 ml of aqueous nutrient agar solution wasmade up using 20 g sucrose, 1 g agar, 0.01 g boric acid,0.03 g calcium nitrate, 0.02 g magnesium sulphate, and0.01 g potassium nitrate. Two drops of the medium wereplaced on each end of a microscope slide. As pollen grainsof many species exhibit a population e�ect (e.g. Mulcahy
et al., 1992) the density of pollen grains in the culturemedium may be critical (Shivanna and Rangaswamy,1992). Consequently when the medium had set, a suitableamount of pollen (not too small or too large; about 100pollen grains) was added over each drop and mixedthoroughly with a needle to obtain homogenous pollendistribution. The microscope slide was then placed in aPetri dish on two glass rods placed parallel at about 50 mmapart on moist ®lter paper and then incubated at 228C.After 24 h the microscope slide was viewed under themicroscope at �160 magni®cation. A coverglass waslowered on the preparation prior to the microscopicexamination. The number of pollen grains was countedand germination assessed.
Germination was scored and recorded following Shi-vanna and Rangaswamy (1992). Pollen grains in tenrandomly selected microscope ®elds were observed. Toavoid repeated scoring of the same group of pollen, thepreparation was moved under the microscope to viewadjacent but non-overlapping ®elds. From each ®eld, thetotal number of pollen grains and the number of germin-ated grains were recorded, and the length of pollen tubesmeasured with an ocular micrometer. Five individual
-incompatibility of Grevillea robusta 137
Assessment of self-compatibility and barriers to sel®ng
A combination of ®eld work and ¯uorescence microscopywas used to investigate incompatibility mechanisms and todetermine barriers to sel®ng. Data from ®eld observationand plant material for microscopy were collectedbetween March and September 1996 at Malava andbetween November 1997 and January 1998 at Canberra.
A controlled pollination technique was used in order toinvestigate the e�ect of di�erent pollination treatments onfruit set. This involved 27 trees, selected from nine di�erentfamilies (i.e. three trees per family) from three di�erentnatural provenances (Bunya Mountains, Boyd River andDuck Creek; details in Table 1). The above provenancesand families were selected randomly from the heavily¯owering families in the stand. The same trees were usedin April and July to check for seasonal variation.
Four di�erent pollination treatments were applied toeither one or two in¯orescences on each tree. A minimum of30 ¯owers per in¯orescence was used. In¯orescences weretagged and randomly allocated to one of four experimentaltreatments: (1) no experimental manipulation (left for open-pollination): in¯orescences were left uncovered to allowvisits by pollen vectors; (2) xenogamy (cross-pollination):self-pollen removed (by emasculation as described above)and in¯orescences bagged, and pollen from trees of adi�erent provenance applied to stigmas 48 h after emascu-lation. Pollen was collected randomly from ¯owering trees,maintaining the same source of pollen for the trees of eachfamily at one period, but not necessarily being the sametrees for both experimental periods (April and July); (3)autogamy (natural self-pollination; spontaneous sel®ng):in¯orescences were bagged and left unmanipulated; (4)geitonogamy (induced self-pollination): self-pollen removedand in¯orescences bagged; pollen taken from other in¯o-
rescences on the same tree, just after pollen dehisces from
microscopy.
1993).
lf
the anthers onto the pollen presenter but prior to anthesis,was transferred to stigmatic areas.
For treatments (2) and (4), ¯owers were emasculated,isolated and pollinated as described earlier. Fruit set wasevaluated 60 d after anthesis. The presence or absence offruit developed from each ¯ower was recorded.
The level of incompatibility among families was assessedby controlled cross-pollination. At Malava, ®ve familieseach from Boyd River (BR) (only three families arereported) and Bunya Mountains (BM) provenances wereused in this study. The two populations are widelyseparated, being at opposite ends of the natural distributionof the species. The choice for cross mating presumablyavoided any possibility of recent common ancestry. One to®ve trees per family were crossed with pollen from one to®ve other families in the provenance and/or crossed withone to ®ve families in the other provenance.
The e�ectiveness of di�erent pollination treatments wasassessed by detection of pollen tubes in treated ¯owers. Theexperiment involved eight trees of di�erent families (onetree per family) in July 1996. In¯orescences with at least 120¯owers were identi®ed and tagged. These were randomlyallocated to one of the four experimental treatments (open,xenogamy, autogamy and geitonogamy) as describedabove. The isolation, emasculation and hand-pollinationprocedures were the same as described under fruit set.
Samples of the ®ve ¯owers were randomly collected fromover the whole raceme for each treatment of each tree(40 ¯owers per treatment) 48 h after pollination. Prelimin-ary observations showed growth of the pollen to takebetween 24 and 48 h from pollen germination to fertiliza-tion. In the case of the autogamy treatment, ¯owers weresampled 72 h after even-aged ¯owers had their pollenpresented, giving self-pollen a chance to germinate. For theopen-pollination treatment (1), ¯owers were sampled afterthey had started to fade and the stigmas to turn brown(i.e. more than 72 h after anthesis). The fading of the¯owers was known to have no e�ect on the visibility ofpollen tubes in other Proteaceae (e.g. Goldingay et al.,1991).
Sampled ¯owers were ®xed and processed for ¯uor-escence microscopy as described above. Forty pistils pertreatment were processed and examined microscopicallyusing aniline blue as a callose stain. Pollen tubes werecounted close to the stigmatic region, where individualpollen grains and attached tubes could be observed. Addi-tional observations were made in the transmission tissue,further down the style, to check for continued growth ofpollen tubes. It was di�cult to observe pollen tubes in thelower style, thus observations for the lower style could notbe scored with precision and were discarded from analysis.However, cross-pollen tubes were occasionally observedentering the ovary, and self-pollen tubes did not appear toenter the lower style region. Counts were made of thenumber of pollen grains per ¯ower, pollen grains germi-nated on the stigma and the number of grains with pollentubes in the upper style. Fluorescing pollen grains withoutany pollen tubes were scored ungerminated. Fluorescing
138 Kalinganire et al.ÐProtandry and Se
pollen tubes were scored as normal tubes if they appeared
straight, or scored abnormal if they had either bulbousswellings or directionless growth.
Flowers which fell 5 to 10 d after anthesis were checkedfor pollen germination and pollen tube growth. These werecollected fresh under two di�erent trees at Malava (Kenya)and 56 single ¯owers were examined for pollen germinationon the stigmas. At Canberra, dropping ¯owers weresampled in the crown of three trees in the ANBG. A totalof 720 ¯owers were collected fresh from three hangingplastic dishes per tree on four di�erent occasions; they were®xed immediately and later processed for ¯uorescence
-incompatibility of Grevillea robusta
Statistical analysis
Most data are categoral and were approximated by abinomial distribution. A logistic regression model with alink function (logit) was ®tted to analyse data on fruit setunder di�erent pollination treatments and the proportionof pollen tubes (presence or absence of tubes) in the style.For pollen viability on stigmas, the signi®cance of treat-ment di�erences in the proportion of germinated pollengrains and pollen tube length and the size of the stigmaticgroove and pollen on di�erent stigmatic phases wasassessed by analysis of variance (ANOVA) (McCullaghand Nelder, 1989; Mead et al., 1993). Log transformationsof the original data were applied to the number of pollengrains per stigma and the number of germinated grains(compatibility experiment and dropping ¯owers), andsquare-root transformations to the proportion of pollengermination (pollen germination on di�erent stigmaticphases) before ANOVA was carried out.
For traits observed for ¯oral structure and development,the signi®cance of provenance and family di�erences weretested by analysis of variance (GLM procedure; Mead et al.,
RESULTS
Flower structure and development
In¯orescence length averaged 151 mm (range 50 to 264;Table 2). Signi®cant (P 5 0.05) provenance and familyvariation was evident. Mudgereeba and Mann River hadlonger in¯orescences than other observed provenances withan average of 168 mm and 167 mm, respectively. Theoverall mean number of racemes per in¯orescence was 3.9,with provenances di�ering signi®cantly in number. Therewas signi®cant variation in in¯orescence length at theprovenance and family level. The total number of ¯owersper raceme was the only trait with signi®cant tree-to-treevariation within families.
Racemes were on average 105 mm (range 10 to 175 mm;Table 2) in length when fully developed, bearing 84 (range15 to 159 ¯owers) early ¯oral green buds and 40 (range 0 to134) golden-yellow or orange coloured ¯owers at anthesis.The open-pollinated ¯owers set few fruits with a lowfruit :¯ower ratio (0.1%; range 0 to 18.2%).
Flowers developed over an average period of 39 d from
bud initiation to anthesis, and a further 60 d was required
the stigmas.
obtained.
TABLE 2. Family means for ¯oral parameters at the time of anthesis for Grevillea robusta
Family name{ No. of treesIn¯or. length
(mm)No. of racemes
per in¯or.Raceme length
(mm)No. of ¯owersper raceme
Final lengthof style (mm)
Loss of ¯owers toanthesis{ (%)
Emu Vale1 4 135 4.1 92 22 21 70Emu Vale2 2 136 3.2 107 55 22 41
Mudgereeba1 2 169 3.6 118 50 25 68Mudgereeba2 1 167 4.8 95 40 24 52
Albert River1 4 148 4.1 109 44 24 49Albert River2 3 149 4.7 82 31 22 64Albert River3 4 164 4.1 123 57 22 48
Nimbin1 4 194 4.5 126 59 22 43Nimbin2 2 145 4.3 71 32 21 72Nimbin3 3 117 4.0 92 29 21 58
Rappville1 3 141 3.0 95 26 25 58Rappville2 3 130 3.2 85 26 23 75Rappville3 2 126 2.8 79 27 24 49
Mann River1 2 183 3.8 107 32 24 48Mann River2 1 156 3.8 110 58 20 52Mann River3 3 161 3.9 119 38 26 61
Mean (43) 151 3.9 105 40 23 56Range 50±264 1±6 10±175 0±134 13±30 3±100F-values:Provenance 3.79** 5.99** 1.14ns 1.83ns 6.11*** 0.60nsFamily 2.89* 1.64ns 2.82* 0.06ns 1.05ns 0.37nsTree in family 0.36ns 0.55ns 0.28ns 3.14* 0.74ns 2.05ns
*** P 5 0.001; ** P 5 0.01; * P 5 0.05; ns, non-signi®cant.{ Details for families are given in Table 1.{ Di�erence between the number of ¯owers at the early bud stage and the number of ¯owers at anthesis, as a percentage of the number of
¯owers observed at early bud stage.
Kalinganire et al.ÐProtandry and Self-incompatibility of Grevillea robusta 139
for fruits to reach maturity. The mean number of days from¯ower bud initiation to di�erent ¯ower stages, including thenumber of days between stages, is given in Table 5 andillustrated in Fig. 2. As the ¯ower matures the style elon-gates nearly three-fold from 8.6 + 0.5 mm at early bud, to23 + 3.1 mm (up to 30 mm; Table 2). The pollen presenterof the mature ¯ower is thus about 25 mm away from thenectar (the main attractant for pollinators), which isproduced near the base of the ¯ower. Flower developmentis accompanied by a gradual change in style colour fromlight green at initiation, orange at anthesis to green/brownat fruit maturity.
An average of 99% of the ¯owers in a raceme may abort(range 100 to 82%). Many were lost between initiation andearly looping (mean of 42% of ¯owers initiated). More¯owers were lost between early looping and anthesis(overall 14%) and another 43% from fruit initiation tofruit maturity. There was no signi®cant correlation betweenthe number of ¯owers at anthesis and the number of fruitset.
The ¯owers opened acropetally, mostly towards dawn(Kalinganire, unpubl. res., 1996) with opening lasting 3 to5 d for each whole raceme. Generally, nectar ¯ow startedwithin 24 h of anthesis of the ®rst ¯ower to present pollenon a raceme and the ¯ow lasted for about 5 d for the raceme(range 4±7 d; Table 3), until about 24 h after the last ¯owerhad presented its pollen. Occasional ¯owers were observed
open without any nectar until 1 d after opening.For each individual ¯ower, observations indicated thatnectar secretion occurred within 24 h of the stigma beingfreed from the perianth and continued for 2 d. Within a dayafter anthesis, nectar secretion occurred together with theproduction of stigmatic exudates, the stigma appearingshiny and wet. However, from these observations it was notpossible to establish when the exudates started ¯owing from
Fruit set at di�erent stages of stigmatic development
Results of the controlled pollination experiment (Fig. 3)indicated that fruit set was possible for the period between1 d before and 2 d after anthesis. No fruits were set on¯owers pollinated 2 d before anthesis and 3 to 5 d afteranthesis, suggesting non-receptivity of the stigma at theseperiods. However, it was not possible to determine thetiming of stigma receptivity from this study as depositedpollen may remain viable to germinate when the stigmabecomes receptive. For example stigmas might not bereceptive 1 d before anthesis although fruit set was
Pollen germination at di�erent stages of stigmaticdevelopment
The percentage of receptive stigmas, de®ned by the
germination of pollen, at di�erent stigmatic stages was
lacked clarity of appearance and direction of growth.
TABLE 3. The main ¯owering stages de®ned for Grevillea robusta at Malava
Stage Main stages Day* s.d. Description
1 Initiation 0 ± Start of formation of ¯ower bud; green2 Early bud 10 0.1 Individual buds visible; purple/green3 Early looping 31 0.4 Style elongates with a small loop; purple/orange4 Advanced looping 35 0.4 Style elongates with a very large loop and gradual emergence between
two perianth segments; orange5 Anthesis/nectar 39 0.5 Freeing of the stigmatic region, stigma shiny; orange6 Fertilization 44 0.6 Perianth segments bearing the dehisced anthers fall; orange7 Fruit development 54 0.4 Embryo starts to swell; orange/green8 Fruit maturity 99 11.0 Fruit from green to brown
e
140 Kalinganire et al.ÐProtandry and Self-incompatibility of Grevillea robusta
signi®cantly di�erent among treatments (P 5 0.05). Therewas no pollen germination on stigmas observed 2 d or 1 dbefore anthesis (Fig. 4).
On the day of anthesis the few pollen grains germinatedhad very short pollen tubes in the styles. Peak stigmareceptivity in G. robusta was recorded 2 d post-anthesis with98% of observed stigmas being receptive and pollen grainshaving long tubes. Stigmas observed 3 and 4 d post-
* The start (mean day) of a given stage and the end (mean day) of th
anthesis, showed high pollen germination but with short
14
12
10
8
6
4
2
0−2 0−1 1 2 3 4
Time of pollination
Fru
it s
et (
%)
FIG. 3. Fruit set from cross-controlled pollinations carried out atvarying times [days before (ÿ) and after (�) anthesis] on emasculated
¯owers at Malava. Bars show s.e.
−2 −1 0 1 2 3 4 5
10090
80
70
60
5040
30
20
10
0
Days before (−) and after (+) anthesis
Per
cen
tage
of
rece
ptiv
e st
igm
as
FIG. 4. Percentage of receptive stigmas (de®ned by the germination ofpollen) following compatible pollen germination on stigmas at di�erent
¯oral stages at Canberra. Bars show s.e.
pollen tubes and a messy aspect, i.e. pollen tubes which
preceding stage.
Timing of stigmatic exudates, and groove and papillae size
Stigma development over a 7 d period and receptivityfeatures of individual G. robusta ¯oral periods are given inTable 4. Groove width was at a maximum 1 d after anthesis(419 + 36 mm; range 379 to 491 mm). There was no pollenadhering to the stigma 2 d before anthesis (Fig. 5); there-after all stages examined had stigmas covered by pollen.Overall pollen size was 40.8+ 2.7mm. Pollen sizes atdi�erent stigmatic stages were not signi®cantly di�erent.
One day and 2 d before anthesis, papillae were visible andgrooves apparently open (Figs 5 and 6) but no stigmaticexudates were observed. Anther dehiscence was 1 d beforeanthesis and pollen was observed adhering to the stigma(Fig. 6). At anthesis (Fig. 7), papillae are taller than at theprevious stages, but no stigmatic ¯uids were observed. Allstigmas examined 1 d after anthesis had their pollen andpapillae immersed in heavy droplets of exudates (Fig. 8),and the width of the stigmatic groove increased to itsmaximum at 1 d post-anthesis. Exudates ran freely (Fig. 8)out of the stigmatic groove at the early period of this stage.Stigmas observed 3 and 4 d after anthesis were characterizedby shorter papillae than in other stages (Fig. 9), a closinggroove and drying out of stigmatic ¯uids. Few stigmashad remnants of exudates at these stages, which were hardlyever seen 5 d after anthesis, and had faded and wrinkledpapillae (Fig. 10). Under ®eld conditions the stigma looksdry during these stages, with collapsed brown papillae.
SEM examination revealed early stigmatic secretions onall 20 emasculated ¯owers 24 h after emasculation, and noexudates for the untreated ¯owers (nine out of ten ¯owers)for the same period. These results suggest that emasculation
slightly enhances an early stigma receptivity.Duration of viability of pollen grains on the stigma
Treatment means for pollen germination and the corres-ponding pollen tube growth are given in Fig. 11. This studyfound that pollen collected about 2 d before anthesis isnot mature as no germination was obtained. Signi®cantdi�erences (P 5 0.001) were obtained among treatments
(treatment ÿ2 d excluded from analysis) for pollen germin-
TABLE 4. Flower development at various times during the stigma receptivity period of Grevillea robusta
Days before (ÿ) orafter (�) anthesis{
Groove size (mm) Pollen size (mm)
Exudate (yes/no)Mean+ s.e. Range Mean+ s.e. Range
ÿ2 290+ 20 232±338 No pollen Nÿ1 274+ 32 218±330 37.9+ 5.2 31±48 Nÿ0 369+ 47 259±491 43.2+ 2.6 35±50 N�1 419+ 36 379±491 40.3+ 3.3 34±45 Y�2 268+ 13 265±271 40.5+ 2.5 38±43 Y/N�3 n.a. Y/N�4 n.a. N
ANOVA F(4,13) � 2.81ns F(3,9) � 0.63ns
{ Five ¯owers were observed at each time and the total number of stigmas measured were three for 1 d before anthesis (on 1 and 2 d afteranthesis) and ®ve stigmas for other stages. At least 30 pollen grains were measured each time.
n.a., Data not available as stigmas looked dry with few or no pollen grains at these stages.
TABLE 5. Pollination success parameters following pollen grain germination and pollen tube growth for di�erent pollinationtreatments in Grevillea robusta
Treatment Empty ¯owers{ (%) Pollen grains per ¯ower Germinated grains per ¯ower Tubes in style per ¯ower{
Open-pollination 55 7.1 2.1 0.6Autogamy 27 6.7 3.2 0.2Geitonogamy 25 11.6 2.6 0.5Cross-pollination 25 19.7 9 4.9
ANOVA 10.4*** 7.2*** 7.4***Logistic regression model 31.81***
***P 5 0.001.n � 40 ¯owers from eight trees.{ Empty ¯owers have stigmas without pollen grains as observed under ¯uorescence microscopy.{ Pollen tubes were counted from the upper part of the style.
TABLE 6. Frequency distribution of pollen deposition onstigmas for Grevillea robusta at Malava, under di�erent
types of pollination
Number ofpollen grainson stigma
Type of pollination (¯owers %)
Open Autogamy Geitonogamy Cross
0±1 62.2 40.9 48.1 34.92±10 24.4 22.8 31.9 30.1
11±20 4.4 31.8 6.5 4.721±30 4.4 4.5 9.5 14.031±60 2.2 0 4.0 14.0
Kalinganire et al.ÐProtandry and Self-incompatibility of Grevillea robusta 141
ated on stigmas of various phases and their correspondingpollen tube growth under ®eld conditions. For pollencollected 1 d before anthesis, 52% of pollen grains germin-ated with short (17.6 mm) but normal pollen tubes. On theday of anthesis 49% of the pollen germinated with a meanpollen tube length of 31.4 mm. The adhesion of pollen onthe presenter 1 d before anthesis and pollen germinationcon®rm that pollen is mature 1 d before ¯ower opening.The best results were obtained with pollen collected 1 dafter anthesis, when 95% of pollen germinated and hadlonger tubes (mean length 46.2 mm). Later pollen graincollections (mostly 3 d after anthesis) showed poor
germination and shorter tubes with a messy appearance. 60±100 2.2 0 0 2.3Assessment of self-compatibility and barriers to sel®ng
The e�ectiveness of natural open-pollination (open), self-(autogamy and geitonogamy) and cross-pollination wasexamined by scoring pollen germinated on the stigma,pollen growth in the style and subsequent fruit set. Thedi�erent types of pollination resulted in signi®cantly di�er-ent numbers of pollen grains on the stigma (P 5 0.001;Tables 5 and 6 for ranges) with 7.1 (range 0 to 100), 6.7(range 0 to 28), 9.2 (range 0 to 47), and 19.7 (range 0 to 81),pollen grains per ¯ower respectively for open, autogamy,
geitonogamy and cross-pollination treatments. Nearlytwo-thirds (65%) of cross-pollinated ¯owers had morethan two pollen grains per ¯ower compared with only 38%of the ¯owers which were naturally open-pollinated(Table 6). The number of ¯owers observed to have nopollen on stigmas di�ered signi®cantly between treatments(P 5 0.001; Table 5); open-pollinated ¯owers had thehighest proportion of bare stigmas (55%).
There were signi®cant (P 5 0.001) di�erences betweenthe numbers of pollen grains germinated on the stigma forall treatments. Although some (25±55%) of the ¯owers had
no pollen germinated on their stigma, cross-pollinated
FIGS 5±10. Scanning electron micrographs (SEM) of Grevillea robusta stigmas. Fig. 5. Papillae (p) 2 d before anthesis. Note the stigmatic groove(g) free of pollen grains and exudates. Fig. 6. Pollen presenter (pp) and papillae (p) covered with pollen grains 1 d before anthesis. Fig. 7. Stigmashowing papillae (p) and pollen grains (po) deposited over the stigmatic groove on the day of anthesis. No stigmatic secretion observed. Fig. 8.Receptive stigma of Grevillea robusta 1 d after anthesis showing pollen grains (po) submerged by the stigmatic secretion (ss) running from thestigmatic groove. Fig. 9. Stigma showing fading papillae with little remains of stigmatic ¯uids (ss) 4 d after anthesis. Fig. 10. Dry stigma and
collapsed papillae 5 d after anthesis. Remnants of exudates were not seen at this stage.
142 Kalinganire et al.ÐProtandry and Self-incompatibility of Grevillea robusta
¯owers had the most germinated pollen grains (nine pollengrains per ¯ower) and open-pollinated ¯owers the fewest(2.1 pollen grains per ¯ower) (Table 5). For autogamous
and geitonogamous treatments, pollen grains germinated,but most were unable to penetrate the stigma, and fewpollen tubes grew down the style towards the ovary. Cross-pollinated ¯owers had signi®cantly more pollen tubes than
other treatments (P 5 0.001). No signi®cant di�erences
(Table 8).
−2 0−1 1 2 3 4 5
germinationtube length
1009080706050403020100
Days before (−) and after (+) anthesis
Pol
len
ger
min
atio
n (
%)
and
poll
en t
ube
len
gth
(m
)
FIG. 11. Pollen germination and pollen tube growth in vitro forGrevillea robusta pollen exposed to ®eld conditions for various lengths
of time. Pollen was tested on a nutrient agar. Bars show s.e.
Kalinganire et al.ÐProtandry and Self
between trees or families were obtained for these observa-tions (P 5 0.05).
Pollen for cross-pollinated ¯owers germinated well withapparently healthy and straight tube growth (Fig. 12). Tubelengths in control cross-pollinations were similar anduniform. However, pollen tubes that grew into the stylesfollowing self-pollination were short and did not progresspast the upper style just next to the stigmatic region. Mostpollen for self- and open-pollinated ¯owers either did notgerminate or showed some pollen tube abnormalities.Tubes were either distorted, presenting a messy growth,with irregular or spiralling tubes (Fig. 13) or with a bulbousappearance at the tube end.
Microscopic examination of dropped ¯owers collectedsoon after the fertilization stage at Malava showed 75%(42 ¯owers out of 56 collected) had no pollen germinated ontheir stigmas; 21% had bare stigmas and 4% had germ-inated pollen. This result suggested that dropping ¯owersare generally not fertilized. A study on ¯owers collected inthe ANBG con®rmed the Kenyan results. A total of 490dropped ¯owers was observed. Although 90% of the ¯owershad pollen grains on their stigmas (mean 13.7 + 1.0 pollengrains), pollen grains germinated on only 35, or 7%, of theobserved ¯owers (mean 4.5+ 1.9%). Only 1.4% (seven outof 490 ¯owers) had pollen tubes growing in the upper style.
In the main controlled pollination experiment there were
signi®cant (P 5 0.001) di�erences in the probability of fruitFIGS 12 and 13. Fluorescence micrographs of Grevillea robusta upperdisplaying abundant pollen germination and growth of normal and straigh
pollen showing inhibition of pollen tube with a dire
set between pollination treatments in both the April andJuly experimental periods (Table 7). The results indicatedfamily di�erences in April (P 5 0.05) but not in July, andprovenance di�erences in both April (P 5 0.01) and July(P 5 0.05).
In July, very low fruit set was obtained for autogamoussel®ng (0.04%; one fruit set out of 2707 ¯owers; 27 trees);and for geitonogamous sel®ng (mean 0.1%; two fruits out of1622 ¯owers obtained from only one tree; 27 trees). No fruitwas set from either autogamy or geitonogamy treatmentsin April. The overall fruit set observed was dependent on thetime of the pollination period (P 5 0.001), with higher fruitset in July than in April. The fruit set following open-pollination was considerably lower (mean 0.1% in Apriland 3.3% in July; range 0±27.1%) than that resulting fromhand cross-pollination (mean 5.9%; range 0±21.4 in Apriland 17.5%, range 0±50 in July).
There was strong evidence of family in¯uence, for bothmale and female parents, on fruit set (Table 8). The averagesuccess rate of the best ®ve crosses was 45% (range 35±55).Thus, some pollen sources promoted higher fruit set. More-over, there was a high overall number of inter-family crosseswithout fruit set (27%). No fruits were set following self-pollination of any of the 18 trees in the crossing programme
-incompatibility of Grevillea robusta 143
DISCUSSION
Fruit set in relation to ¯oral development
Grevillea robusta trees bear approx. 50 000 hermaphroditic¯owers, opening acropetally. The period from ¯oralinitiation to fruit maturity is approx. 3 months. Dependingon the size of an in¯orescence, it may take from 3 to 10 dfor all the ¯owers on an in¯orescence to open and inmost in¯orescences nectar ¯ows continuously from one ormore of the racemes over this period. As anthesis is accom-panied by nectar secretion, at least within 24 h ofpresentation, this strategy encourages ¯oral visitors andhence self-pollen removal and pollen deposition from other¯ower sources, thus increasing the chances of cross-
pollination.style. Fig. 12. Style showing compatible controlled cross-pollinationt pollen tubes 48 h after pollination. Fig. 13. Style with controlled self-ctionless and messy aspect 48 h after pollination.

by the timing of anthesis and nectar ¯ow.
TABLE 7. Fruit set of Grevillea robusta from di�erent pollination treatments at Malava
Provenance (CSIRO no.) Family code
April{ July{ July
Cross-pollination(% fruit set)
Cross-pollination(% fruit set)
Open-pollination(% fruit set)
Boyd River (17622) CEH 89 2.8 16.7 0CEH 91 3.0 9.2 6.9CEH 980 5.2 10.8 0.4
Duck Creek (17614) CEH 26 7.7 10.7 0CEH 28 2.8 16.8 4.8CEH 29 6.0 15.3 9.1
Bunya Mountains (17693) T 5 12.8 31.9 0T 3 6.8 32.4 4.0T 2 5.7 13.3 4.2
Overall mean (no. ¯owers observed) 5.9 (2112) 17.5 (2416) 3.3 (2718)
Variation (logistic regression model):Among treatments 78.48*** 62.83***Inter-provenance 5.93** 3.91*Inter-family 2.86* 0.85nsFruit set over periods 56.41***
*** P 5 0.001; ** P 5 0.01; * P 5 0.05; ns, non-signi®cant.{ April, No fruit set for self-pollination treatments and very low fruit set for open-pollination (two out of 2078 ¯owers; 0.1%).
(
144 Kalinganire et al.ÐProtandry and Self-incompatibility of Grevillea robusta
On the day of anthesis the pollen presenter, a specializedstructure, serves as both a pollen-reception zone and apresenter of pollen for transfer to pollinating agents(Collins and Rebelo, 1987). Pollen can be seen on thepresenters of individual ¯owers for up to 5 d. The peak of¯ower opening on a raceme level coincides with the peak ofnectar production. This presumably maximizes the chancesof ¯ower visitation and cross-pollination. Also, a longperiod of nectar secretion should encourage more visits perraceme, leading to higher pollen removal and transfer.However, the process does not exclude the possibility ofgeitonogamy (the transfer of self-pollen to other ¯owers onthe same individual; Snow et al., 1996) or the existence of aself-incompatibility mechanism.
Many ¯owers were aborted from initiation to early budstage and less than 44% reached anthesis. Five to 10 d afteranthesis, ¯owers abort en masse (43% of the total numberof ¯owers), with very few remaining until fruit maturity.This ¯ower drop might be a result of competition forassimilates. The heavy losses after anthesis suggest unsuc-cessful fertilization. After anthesis the ovary starts swellingif fertilization takes place.
It is not clear from the above observations whether cross-pollen or other resources are factors limiting fruit set fornatural open-pollinated ¯owers of G. robusta. This studyfound that dropping ¯owers are not fertilized, and thatthere was no fruit set from capped ¯owers. This is contraryto the report by Brough (1933) who found fruit develop-ment on capped ¯owers. The absence of fruit set ondropping and capped ¯owers indicates the presence ofincompatible, probably self-pollen on stigmas. The aboveresult, together with the large variation in the fruit :¯owerratio, indicates poor fertilization, most probably related tothe lack of suitable pollinators. The very low fruit set (5.9 to
{ July, For geitonogamy (two out of 1622; 0.1%) and for autogamy
17.5%) from controlled hand-pollination is due in part to
failure of some individual crosses to set any fruit. Othercrosses displayed higher levels of fruit set. As the pollenparents used were successful in at least some crosses(Table 8), this points to incompatibility in some crosses.
The clustering of ¯owers on the ends of branches,brush-type presentation, bright colour and copious nectarproduction suggest G. robusta is pollinated by animalvectors. The structure of the in¯orescences and the nectarreward suggest the species is most probably bird- ormammal-pollinated, similar to other proteaceous species(Collins and Rebelo, 1987). Individual ¯owers are adaptedto early-morning presentation for pollination, as suggested
one out of 2707 ¯owers; 0.04%).
The protandrous behaviour of Grevillea robusta
The study showed G. robusta to have a wet papillatestigma, contrary to the dry stigma reported for Grevilleaspecies by Heslop-Harrison and Shivanna (1977). Someother members of the Proteaceae have been reported tohave wet stigmas at the time of stigma receptivity; forexample Banksia (Fuss and Sedgley, 1991), Macadamia(Sedgley et al., 1985), Dryandra (Matthews and Sedgley,1998) and Grevillea leucopteris (Lamont, 1982). The SEMexamination revealed that stigmas were dry at anthesis, buthad copious secretion commencing 1 d thereafter for aperiod of 3 d. The pollen germination studies indicated thatstigmatic receptivity of G. robusta began to develop 1 dpost-anthesis, but the greatest pollen germination rates onstigmas and longest pollen tubes were observed 2 d post-anthesis, and individual stigmas remained receptive forapprox. 3 d post-anthesis.
In the studies reported here, G. robusta pollen was notviable until 1 d before its presentation to ¯oral visitors.
Pollen viability was retained for 4 d, providing a good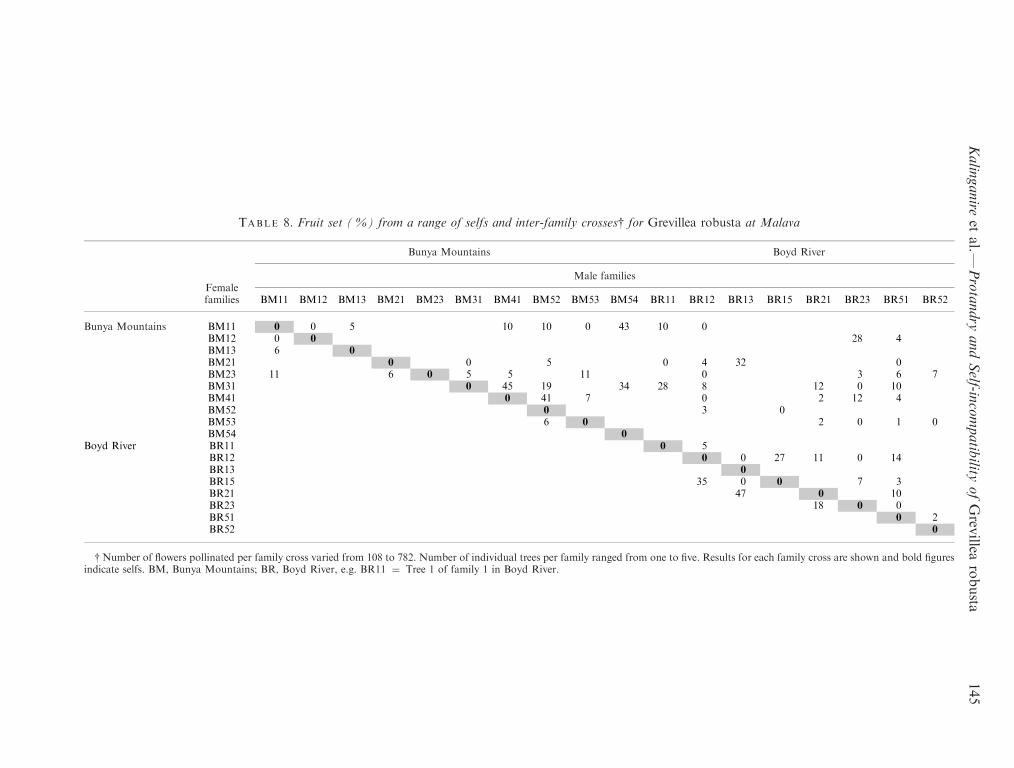
TABLE 8. Fruit set (%) from a range of selfs and inter-family crosses{ for Grevillea robusta at Malava
Bunya Mountains Boyd River
Femalefamilies
Male families
BM11 BM12 BM13 BM21 BM23 BM31 BM41 BM52 BM53 BM54 BR11 BR12 BR13 BR15 BR21 BR23 BR51 BR52
Bunya Mountains BM11 0 0 5 10 10 0 43 10 0BM12 0 0 28 4BM13 6 0BM21 0 0 5 0 4 32 0BM23 11 6 0 5 5 11 0 3 6 7BM31 0 45 19 34 28 8 12 0 10BM41 0 41 7 0 2 12 4BM52 0 3 0BM53 6 0 2 0 1 0BM54 0
Boyd River BR11 0 5BR12 0 0 27 11 0 14BR13 0BR15 35 0 0 7 3BR21 47 0 10BR23 18 0 0BR51 0 2BR52 0
{ Number of ¯owers pollinated per family cross varied from 108 to 782. Number of individual trees per family ranged from one to ®ve. Results for each family cross are shown and bold ®guresindicate selfs. BM, Bunya Mountains; BR, Boyd River, e.g. BR11 � Tree 1 of family 1 in Boyd River.
Kalin
ganire
etal.Ð
Protandry
andSelf-in
compatib
ilityofGrev
illearobusta
145

prevent autogamy in G. robusta.
lf
opportunity for its removal and deposition on di�erent¯owers. Pollen collected on stigmas 3 d after anthesis andgerminated in-vitro had a messy appearance and shortertubes. This poor vigour casts doubt on its ability to growand fertilize ovules. The results conform with Thomson andThomson's (1989) statement that pollen may lose viabilityover time, such that pollen removed late in the ¯ower's lifehas less chance of fertilizing ovules than pollen removedearlier. At anthesis, the stigma is probably not receptive.Results obtained at Malava (Fig. 3) indicated that pollendeposited before the period of receptivity remained viablelong enough to germinate when the stigma became recep-tive. Under ®eld conditions the stigma appears shiny andwet 1 d after anthesis. Moreover, there is evidence of poorpollen germination and pollen tube growth for pollencollected more than 2 d after anthesis. Also, there is lesslikelihood of collection taking place, as the ¯ower ceasesnectar production, thus becoming less attractive to visitors.
To maximize the likelihood of pollen germination, andsubsequent seed set, hand-pollinations should be made onthe day of peak stigma receptivity. This peak was 1±2 d afteranthesis when the highest rates of pollen grain germinationwith normal pollen tubes in the styles were obtained. Thisperiod was accompanied by morphological changes to thestigma including longer papillae and the production ofstigmatic exudate. In Grevillea wilsonii (Collins and Grey,1988) and G. barklyana (Vaughton, 1996), the stigmaticpapillae have also been shown to enlarge as receptivityincreases. The current study showed that no fruit setoccurred on stigmas hand-pollinated 4 and 5 d afteranthesis, indicating loss of stigma receptivity at this stage.
The stigmatic groove was apparently open and widerthan the diameter of the pollen grain before anthesis, but nosuccessful pollination was e�ected. Thus, groove width didnot limit pollen deposition within the groove, but thestigma was not yet receptive. Therefore, the timing ofgroove opening in G. robusta is not su�cient to determinestigma receptivity.
Within a ¯ower, the anthers of G. robusta dehisce prior toanthesis and stigma receptivity. One day before, and on theday of anthesis, self-pollen completely covers the pollenpresenter and the stigmatic groove. Thus, successful naturalcross-pollination may depend upon the removal of self-pollen before the stigma becomes receptive within 1 d ofanthesis. Consequently, protandry makes it possible formost pollen to be dispersed before the stigma becomes mostreceptive. Therefore, the protandrous ¯owering habit ofG. robusta makes individual ¯owers well adapted to out-breeding. But geitonogamy is always possible within thesame raceme, the same in¯orescence or the same tree as theycontain both male and female phase ¯owers at a range ofdevelopmental stages.
The study has shown the existence of protandry inG. robusta, with less than 2 d separation between male andfemale maturity phases for individual ¯owers. Thisprotandrous mechanism was reported by Brough (1933)and Venkata Rao (1971) but disputed by Owuor and Oduol(1992). The latter reported G. robusta to have simultaneousanther dehiscence and stigma receptivity. Protandry in
146 Kalinganire et al.ÐProtandry and Se
G. robusta allows some time for self-pollen removal. If self-
pollen is not removed by ¯oral visitors, self-pollination mayoccur autogamously. Since viable self-pollen will still bepresent on ¯owers when the stigma becomes receptiveÐunless removed by pollinators or rainÐ protandry will not
-incompatibility of Grevillea robusta
Fertilization and sel®ng barriers in Grevillea robusta
In this study, the stigmatic surface was the ®rst site ofpollen grain inhibition for self-pollinated ¯owers. Thesecond site of inhibition was the upper part of the style:most self-pollen tubes (Table 5) failed to penetrate to thelower part of the style. Few self-pollen tubes grew normallytowards the ovary. Cross-pollen tubes were occasionallyseen penetrating the ovules, but no self-pollen could bedetected doing this. However, although there was no fruitset following self-pollination, self-pollen tubes were occa-sionally seen extending beyond the upper style; and becauseof di�culties in observing fully the lower style it could notbe determined whether self-incompatibility occurs in theovary as well. Further investigation of the cytology(pollen±ovule interaction) of fertilization might clarifythis question.
Fewer pollen grains were observed on open- and self-pollinated ¯owers than on cross-pollinated stigmas, andopen-pollinated ¯owers had a higher incidence of empty¯owers (Tables 6 and 7). Together with the poor pollengermination obtained on self- and natural open-pollinated¯owers at Malava, these observations suggest self-pollenfails to adhere and germinate on stigmas before it is washedo� by frequent heavy rains (see Lamont, 1982). Most cross-pollinated stigmas presented more pollen grains than theovule number (two per ¯ower), giving a better chance ofsuccessful fertilization.
Grevillea robusta set few fruit by self-pollination(50.1%), while cross-pollinated ¯owers set 5.9% and17.5%, respectively, in April and July (Table 7). Most self-pollen tubes failed to fertilize the ovule after exhibitingdistorted growth patterns (Fig. 13), suggesting that thisfailure resulted from self-incompatibility reactions. Theresults demonstrated that G. robusta possesses strongbarriers to sel®ng with a likely gametophytic self-incompatibility system (de Nettancourt, 1977). This kindof inhibition in the upper style has been reported in othermembers of the Proteaceae (Herscovitch and Martin, 1990;Fuss and Sedgley, 1991; Goldingay and Whelan, 1993).
The natural fruit set following open-pollination was low(0.1 to 3.3%), and was considerably lower than that whichresulted from controlled cross-pollination. Brough (1933)observed variable fruit production (meagre to high) in anexotic population at Sydney, Australia depending on theweather conditions prevailing at the time of pollination,while Owuor and Oduol (1992) reported very high fruit set(69%) with G. robusta landraces at Maseno in westernKenya. The present results are consistent with those forother members of the Proteaceae (e.g. Vaughton, 1991;Harriss and Whelan, 1993) in general, and with Grevilleaspecies (0.01 to 0.09 fruit set; Hermanutz et al., 1998) in
particular.
example, eucalypts (Eldridge et al., 1993).
lf
In this study, fruit set in April was much lower thanin July for both open-pollination and cross-pollination.This variation is probably due to heavy rains which fellin April±May, which may have had an impact on fertil-ization of the ¯owers. Such conditions, i.e. rainy and cloudyweather, naturally restrict pollinator activity and in generalpresent an unfavourable environment for pollination, andconsequently result in poor fruit production. Moreover,pollination is adversely a�ected following the displacementof pollen which is washed o� the stigmas [see Boland andOwuor (1996) for Calliandra calothyrsus under similarenvironmental conditions], and probably by an increasedpercentage of sterile pollen which occurs as a result ofrepeated wetting and drying (Brough, 1933). The aboveresults emphasize the e�ects of climatic conditions at thetime of fertilization.
A signi®cant variation between trees in the rate of fruitset for both cross- and open-pollinated ¯owers is evidentfrom this study. For natural open-pollinated ¯owers, suchvariation may be due in part to di�erences in nectarconcentration and production, and hence energy rewards(Heinrich and Raven, 1972), or to other factors such as thenumber of in¯orescences displayed per tree or the colour ofthe ¯owers. This may a�ect movement of pollinatorsamongst trees and hence the rate of pollen transfer. Di�er-ences seen between cross-pollinated ¯owers within familiesmay also arise from di�ering compatibilities of di�erentmale parents for di�erent individual crosses. Furthermore,this study revealed that few of the ¯owers that droppedsoon after natural fertilization had germinated pollen andvery few had pollen-tubes in their styles. The germinationfailure is probably due to a self-incompatibility mechanism,indicating that the ¯ower's own pollen was probably notremoved by ¯oral visitors. This was observed in otherAustralian Proteaceae where self- and outcross-pollen areoften transported by pollinators but self-pollen is expectedto predominate on stigmas (Goldingay et al., 1991;Carthew, 1993; Ayre et al., 1994). Thus, the unfertilized¯owers, probably bearing self-pollen, dry up and drop o�the raceme soon after the fertilization phase.
The poor fruit set following natural open-pollination isdue mainly to the absence of outcrossed pollen. This isprobably due to low visitation rates by e�ective pollinators(Kalinganire et al., 1996). Flowers of G. robusta contain twoovules and it was established (Kalinganire, unpubl. res.,1996) that whenever there is fruit set the follicle alwayscontains two seeds, suggesting a low abortion rate once a¯ower receives adequate pollen. As found in the Grevilleaspecies studied (Herscovitch and Martin, 1990; Hermanutzet al., 1998), only two or three pollen tubes penetrate thestylar transmission tissue and these have access to the twoovules.
Fruit production per in¯orescence showed no relation-ship to the number of ¯owers remaining after anthesis andappeared to be primarily dependent on the proportion of¯owers with compatible pollen. This suggests a weak e�ectof other resources limiting fruit set. Inadequate receipt ofcross-pollen on open-pollinated ¯owers contributed con-siderably to the low fruit set. If pollination was not a
Kalinganire et al.ÐProtandry and Se
limiting factor, higher rates of fruit set might be expected on
open-pollinated ¯owers than on cross-pollinated ¯owersbecause of the stress experienced during emasculation. Lowrates of cross-pollination appear to be a primary cause oflow fecundity in perennial plants, which are often self-incompatible (Burd, 1994). Therefore a lack of cross-pollenwas mainly responsible for a low fruit set in G. robusta.However, post-anthesis loss in cross-pollinated ¯owerssuggests resource limitation should not be excluded. Assuggested by Vaughton (1991) and Campbell and Halama(1993), pollen and nutrients may interact in determininglevels of fruit set, suggesting that fecundity may depend onthe availability of both factors. It is unlikely that there is astrict separation between the e�ects of pollen and resourceallocation on the reproductive output. Yet, as reported byBierzychudek (1981), if hand-pollinated plants producemore seeds than naturally pollinated controls, thenreproduction is being limited by pollinator activity.
The present study has clearly shown the natural low fruitset to be largely due to a lack of cross-pollen on stigmas anda self-incompatibility mechanism. Cross-pollen transferseems likely to be a factor limiting fruit production becauseof a lack of suitable pollinators at Malava (Kalinganireet al., 1996) and probably high levels of incompatibilityamong families. Misty weather conditions might a�ect theoverall fruit production at Malava, by cross- or naturalpollination, and it is suggested that breeding work shouldbe undertaken during the drier parts of the year even if¯owering is poor. Therefore, it is likely that the lowfruit :¯ower ratio is due to complex genetic factors, includ-ing self-incompatibility, as well as being in¯uenced byenvironmental conditions, pollinator abundance andpossibly to a lesser extent by the availability of resources.
The study shows G. robusta to be highly self-incompatibleand outcrossing. The species might be capable of a very lowrate of sel®ng, but fruit set results indicate very high rates ofoutcrossing for all genotypes studied. This is consistent withthe high outcrossing rates obtained by Harwood et al. (1992)from isozyme studies of two natural populations. Allogamyis con®rmed to be the primary breeding behaviour ofG. robusta. Self-incompatibility may help to explain why thespecies has not shown any substantial genetic deteriorationas an exotic, especially in the eastern and central Africanhighlands. Very few selfed plants are produced in progeny;therefore inbreeding does not develop as rapidly as in, for
-incompatibility of Grevillea robusta 147
ACKNOWLEDGEMENTS
This study comprised part of the PhD research undertakenby Antoine Kalinganire and was funded by Intercoopera-tion Suisse. The Australian National University (ANU)Department of Forestry, CSIRO and ICRAF also providedresearch support. The Australian National BotanicGardens (Canberra) supplied some of the plant materialfor this study under permit number 20. Thanks to SallyStowe and Roger Heady of the ANU Electron MicroscopyUnit; Ross Cunningham and Christine Donnelly of theStatistical Consulting Unit of the Graduate School (ANU)and David Boshier (University of Oxford) for their
assistance, and Peter Kanowski (ANU), Mike Moncur,
lf
Alan Brown and Doug Boland (CSIRO) for helpfulcomments. Thanks to Stephen Ruigu, Agnes Yobterikand Amadou Niang (ICRAF Maseno) for their support,and to Walter Adongo, Joseph Njeri and their team for
148 Kalinganire et al.ÐProtandry and Se
technical assistance during the ®eld work at Malava.
LITERATURE CITED
Ayre DJ, Whelan RJ, Reid A. 1994. Unexpectedly high levels of sel®ngin the Australian shrub Grevillea barklyana (Proteaceae). Heredity72: 168±174.
Bierzychudek P. 1981. Pollination limitation of plant reproductivee�ort. The American Naturalist 117: 838±840.
Boland DJ, Owuor B. 1996. Some aspects of ¯oral biology and seedproduction in exotic Calliandra calothyrsus at Maseno, Kenya. In:Evans DO, ed. International workshop on the genus Calliandra.Forest, Farm, and Community Tree Research Reports (SpecialIssue). Morrilton, Arkansas: Winrock International, 49±61.
Brough P. 1933. The life history of Grevillea robusta (Cunn). Proceed-ings of the Linnaean Society of New South Wales 58: 33±73.
Burd M. 1994. A probabilistic analysis of pollinator foraging behaviourand female reproductive success in Lobelia deckenii. Ecology 75:1635±1646.
Campbell DR, Halama KJ. 1993. Resource and pollen limitation tolifetime seed production in a natural plant population. Ecology 74:1043±1051.
Carthew SM. 1993. An assessment of pollinator visitation to Banksiaspinulosa. Australian Journal of Ecology 18: 257±268.
Collins BG, Grey J. 1988. Pollination and seed set in Grevillea wilsonii.In: Singh MB, Troiani LF, eds. Pollination, `88'. Melbourne:School of Botany, University of Melbourne, 67±70.
Collins BG, Rebelo T. 1987. Pollination biology of the Proteaceae inAustralia and southern Africa. Australian Journal of Ecology 12:387±422.
Craig S, Beaton CD. 1996. A simple cryo-SEM method for delicateplant tissues. Journal of Microscopy 182(2): 102±105.
de Nettancourt D. 1977. Incompatibility in angiosperms. Berlin:Springer-Verlag.
Eldridge KG, Davidson J, Harwood CE, Van Wyk G. 1993. Eucalyptdomestication and breeding. Clarendon: Oxford University Press.
Fuss AM, Sedgley M. 1991. Pollen tube growth and seed set of Banksiacoccinea R.Br. (Proteaceae). Annals of Botany 68: 377±384.
Goldingay RL, Carthew SM. 1998. Breeding and mating systems ofAustralian Proteaceae. Australian Journal of Botany 46: 421±437.
Goldingay RL, Whelan RJ. 1993. The in¯uence of pollinators on fruitpositioning in the Australian proteaceous shrub Telopea specio-sissima. Oikos 68: 501±509.
Goldingay RL, Schibeci SM, Walker BA. 1991. Breeding system andpollination levels of Banksia ericifolia. Australian Journal ofBotany 39: 365±372.
Harriss F, Whelan RJ. 1993. Selective fruit abortion in Grevilleabarklyana (Proteaceae). Australian Journal of Botany 41: 499±509.
Harwood CE. 1989. Grevillea robustaÐan annotated bibliography.Nairobi: International Centre for Research in Agroforestry.
Harwood CE. 1992. Natural distribution and ecology of Grevillearobusta. In: Harwood CE, ed. Grevillea robusta in agroforestry andforestry. Nairobi: International Centre for Research in Agro-forestry, 21±28.
Harwood CE, Bell JC, Moran GF. 1992. Isozyme studies on geneticvariation and the breeding system inGrevillea robusta. In: HarwoodCE, ed. Grevillea robusta in agroforestry and forestry. Nairobi:International Centre for Research in Agroforestry, 165±176.
Heinrich B, Raven PH. 1972. Energetics and pollen ecology. Science176: 598±602.
Hermanutz L, Innes D, Denham A, Whelan R. 1998. Very lowfruit :¯ower ratios in Grevillea (Proteaceae) are independent ofbreeding system. Australian Journal of Botany 46: 465±478.
Herscovitch JC, Martin AR. 1990. Pollen±pistil interactions in
Grevillea banksii II. Pollen tube ultrastructure and interactions,and results of ®eld experiments. Grana 29: 5±17.Heslop-Harrison Y, Shivanna KR. 1977. The receptive surface ofangiosperm stigma. Annals of Botany 41: 1233±1258.
Hogbin PM, Ayre DJ, Whelan RJ. 1998. Genetic variation andreproductive success of road verge populations of the rare shrubGrevillea barklyana (Proteaceae). Heredity 80: 180±186.
Johnson LAS, Briggs BG. 1975. On the ProteaceaeÐthe evolution andclassi®cation of a southern family. Botanical Journal of LinnaeanSociety 70: 83±182.
Kalinganire A, Harwood CE, Simons AJ, Moncur MW, Slee M. 1996.Reproductive ecology of Grevillea robusta in western Kenya. In:Dieters MJ, Matheson AC, Nikles DG, Harwood CE, WalkerSM, eds. Tree improvement for sustainable forestry. Proceedings ofthe QFRI±IUFRO Conference, Caloundra, Queensland, Austra-lia. 27 October±1 November 1996, 238±243.
Ladd PG, Nanni I, Thomson GJ. 1998. Unique stigmatic structure inthree genera of Proteaceae. Australian Journal of Botany 46:479±488.
Lamont B. 1982. The reproductive biology of Grevillea leucopteris(Proteaceae), including reference to its glandular hairs andcolonizing potentials. Flora 172: 1±20.
McCullagh P, Nelder JA. 1989. Generalized linear models. 2nd edn.Monographs on statistics and applied probability 37. New York:Chapman & Hall.
McGillivray DJ. 1993. Grevillea, Proteaceae: a taxonomic revision.Melbourne: Melbourne University Press.
Matthews ML, Sedgley M. 1998. Breeding system of Dryandraquercifolia and D. formosa (Proteaceae). Australian Journal ofBotany 46: 439±452.
Matungulu K. 1994. Soils and climate: Eastern and Central AfricaAFRENAS Research Sites. Nairobi: International Centre forResearch in Agroforestry.
Mead R, Curnow RN, Hasted AM. 1993. Statistical methods inagriculture and experimental biology. 2nd edn. London: Chapman& Hall.
Mulcahy DL, Mulcahy GB, Searcy KB. 1992. Evolutionary geneticscompetition. In: Wyatt R, ed. Ecology and evolution of plantreproduction. New York: Chapman & Hall, 25±36.
Owuor B, Oduol P. 1992. Early observations on ¯oral biology ofGrevillea robusta (A. Cunn.) in relation to controlled cross-pollination. In: Harwood CE, ed. Grevillea robusta in agroforestryand forestry. Nairobi: International Centre for Research inAgroforestry, 177±182.
Sedgley M, Blesing MA, Vithanage HIMV. 1985. A developmentalstudy of the structure and pollen receptivity of the macadamiapistil in relation to protandry and self-incompatibility. BotanicalGazette 146: 6±14.
Sedgley M, Sierp MG, Maguire TL. 1994. Interspeci®c hybridizationinvolving Banksia prionotes Lind. and B. menziesii R.Br.(Proteaceae). International Journal of Plant Science 155: 755±762.
Sedgley M, Sierp MG, Wallwork MA, Fuss AM, Thiele K. 1993. Pollenpresenter and pollen morphology of Banksia L.f. (Proteaceae).Australian Journal of Botany 41: 439±464.
Shivanna KR, Rangaswamy NS. 1992. Pollen biology: a laboratorymanual. Berlin, Heidelberg, New York: Springer-Verlag.
Snow AA, Spira TP, Simpson R, Klips RA. 1996. The ecology ofgeitonogamous pollination. In: Lloyd DG, Barrett SCH, eds.Floral biology: studies on ¯oral evolution in animal-pollinatedplants. New York: Chapman & Hall, 191±216.
Thomson JD, Thomson BA. 1989. Dispersal of Erythronium grandi-¯orum pollen by bumblebees: implications for gene ¯ow andreproductive success. Evolution 43: 657±661.
Vaughton G. 1991. Variation between years in pollen and nutrientlimitation of fruit-set in Banksia spinulosa. Journal of Ecology 78:389±400.
Vaughton G. 1996. Pollination disruption by European honeybeesin the Australian bird-pollinated shrub, Grevillea barklyana(Proteaceae). Plant Systematics and Evolution 200: 89±100.
Venkata Rao C. 1971. Proteaceae. New Delhi, India: CSIR.Wallace HM, Vithanage V, Exley EM. 1996. The e�ect of supple-
-incompatibility of Grevillea robusta
mentary pollination on nut of Macadamia (Proteaceae). Annals ofBotany 78: 765±773.
Copyright © 2022 FDOKUMEN




















