
Pandey et al 2012 Perisphinctes from the Oxfordian of Kachchh

International Journal of Botany and
Research (IJBR)
ISSN 2277-4815
Vol. 3, Issue 4, Oct 2013, 39-50
© TJPRC Pvt. Ltd.
FLORAL BIOLOGY AND POLLINATION ECOLOGY OF RHIZOPHORA MUCRONATA
LAMK. IN GULF OF KACHCHH, GUJARAT, INDIA
R. PANDEY1 & PANDEY
C. N
2
1Project Coordinator-CCS, Gujarat, India
2Principal Chief Conservator of Forests (Wildlife), Gujarat, India
ABSTRACT
Rizophora mucronata is a viviparous mangrove species of the Rhizophoraceae family. The species is reported to
be wind pollinated. However, the floral biology and pollination ecology of the species have not been studied in detail in
Gulf of Kachchh. To fill this information gap, the present work has involved detailed field and laboratory investigations
regarding the flower biology and pollination ecology of R. mucronata. The major objective of the research was to
document the floral biology and pollination ecology of R. mucronata by examining the temporal relation between the
anthesis, anther dehiscence, stigma receptivity and pollinator visitation pattern. Further, the study also investigated the
chemical nature of the floral exudate. The visitation pattern of floral visitors was observed at different levels to classify
them into floral visitors and pollinators. The flowers of R. mucronata have been found to exhibit protandry. Further, the
flower is inverted and exhibits herkogamy. These features enhance the possibilities of cross breeding over self breeding.
A crucial role played by petals has been found in regulating, with help of corona, the pollen dispersal for a longer period
after anther dehiscence. Further, the observed asynchrony between the anther dehiscence and stigma receptivity also appear
to be promoting cross breeding. Wind appears to be a major pollinator. However, some insects have also been found to
play a definite role as pollinators. The sugar content of the floral exudate indicates that these exudates are nectar and
further accounts for the bees and ants in later floral stages when the exudate is secreted.
KEYWORDS: Anther Dehiscence, Floral Phenology, Only Four Mangrove Species
INTRODUCTION
Mangrove species, in general, show structural and functional features which favor outbreeding
(Primack et al 1980). Mangroves are often pollinated by animals and the classes of flower visitors are remarkably diverse.
The floral biology, to some extent, determines the breeding type (viz. self or cross breeding) in mangroves. Recently the
reproductive biology of some mangrove species such as Avicennia marina (Clarke & Myerscough 1991, Coupland et al
2005), Aegiceras corniculatum (Pandit & Chaudhary 2001), Rhizophora stylosa, Ceriops australis Sonneratia alba
(Coupland et al 2005 ), , and C. decandra (Raju et al 2006) has been studied in different mangrove forests of the world.
The floral biology and pollination ecology of R. mucronata, however, has not been studied in details for Gulf of Kachchh.
The reproductive biology, in general, depends on the local biotic and abiotic factors such as climatic condition,
pollinator resources etc. It is in this context that the present study focuses on the floral biology and pollination ecology of R.
mucronata in Gulf of Kachchh (GOK), Gujarat, India. The genus is represented by eight species (including 3 hybrids).
Although the genus is quite common, none of the species shows a worldwide distribution. The genus Rhizophora
belonging to the family Rhizophoraceae is generally considered wind pollinated. The floral features which indicate the
occurrence of wind pollination are a high pollen/ovule ratio and light powdery pollens. Chai (1982) reported that
Rhizophora sets fruits when animal visitors are denied access to flowers enclosed in fine mesh bags that does not exclude

40 R. Pandey & Pandey C. N
wind born pollens and Tomlinson (1980) recognized the probability of self- compatibility in the genus due to the above
mentioned features. However, the exact pollination strategy has not been experimentally tested. Notwithstanding the strong
possibility of wind pollination, the absence of an elaborate stigma to catch wind born pollens, the observed frequent visits
by bees and wasp to the flowers and the significant presence of floral exudates definitely suggest the possibility of biotic
vectors having some role in the pollination process. Hence, there is a need to have a closer examination of the pollination
biology of the genus of which a particular member - R. mucronata - was selected for this study. R. mucronata
inflorescences are in axillary cymes bearing 2-4 pedicellate, tetramerous flowers. The flower has white corolla, eight
epipetalous anthers and inferior ovary with bifid stigma The objectives of the present work on R. mucronata was to
investigate: (i) floral phenology, (ii) floral biology, (iii)pollination ecology, (iv)synchrony among anthesis, anther
dehiscence, stigma receptivity, exudate secretion and visitation pattern of floral visitors, and (v) the chemical composition
of floral exudates.
MATERIALS AND METHODS
Study Area
The present work has been carried out in the Marine National Park and Sanctuary, located in the Gulf of Kachchh,
Gujarat, India. Gujarat has the longest coastline (1650 Km.) and hosts two of the three gulfs of India viz. Gulf of Kachchh
and the Gulf of Khambhat. Gulf of Kachchh is located between the latitudes of 22015’N and 23
040’N and the longitudes of
68020’ E and 70
040’ E. The coastline is marked by very high tidal amplitudes of up to 12 ms. As a semiarid to arid zone,
the rate of evaporation in most parts of coastal Gujarat (particularly around Gulf of Kachchh) exceeds the rate of
precipitation. The mangrove forests in the state have relatively less species diversity as compared to many other mangrove
regions of the country. So far, 13 mangrove species have been reported from Gujarat. Further, the mangrove species are not
uniformly distributed in the state and the community distribution is highly skewed in favour of Avicennia which is
represented by three species A. marina, A. alba and A. Officinalis. Out of these, only A. marina is abundant and other two
species are conspicuously rare (Pandey and Pandey 2008). Therefore, even at the species level, the mangrove community
of Gujarat shows a highly skewed distribution in favour of only one species, A. marina.The two islands Pirotan and Bhens
Bid and the coastal site Sikka were selected as study sites. The islands are located in the eastern part of Gulf of Kachchh
(Figure 1). During the field observations, only four mangrove species A marina, R. mucronata, C.. tagal and A.
corniculatum could be found on the two islands of which, A marina was the most dominant and A. corniculatum was the
rarest species. Since the selected islands are quite remotely located and situated in a protected area, the anthropogenic
pressure is limited.
Source: Status of Magrove in Gulf of Kachchh(2005)
Figure 1: Location of Study Area in MNP&S, in GOK

Floral Biology and Pollination Ecology of Rhizophora mucronata Lamk. in Gulf of Kachchh, Gujarat, India 41
METHODOLOGY
Reproductively matured trees of different heights were selected for making field observations. The field
observations were made between March 2007 and December 2008 covering two flowering seasons. The phenology and
floral biology of R. mucronata was studied by systematically observing all the present inflorescence on 30 selected trees
(10 trees on each site) and studying temporal relation between various flowering processes such as anthesis, anther
dehiscence, stigma receptivity and nectar secretion. For this, the flowers and buds were categorized into different floral
stages based upon morphological observations and the current phnological process such as anthesis, anther dehiscence,
pollination etc.
To study the process of anthesis, 100 buds were selected and tagged at each of the three sites (total 300 buds).
The 100 buds were selected in a way that the initiation of anthesis occurred equally in different periods of the day
including mornings, evenings and nights. The pollen presentation duration was calculated by noting the average time gap
between the anthesis and the petal fall. It was noted that anther dehiscence commenced and almost completed before the
initiation of anthesis. However, pollen dispersal was observed to start after anthesis. Generally, the duration of anther
dehiscence corresponds to the pollen dispersal period (also known as the pollen presentation time or pollen availability
period). However, in case of R. mucronata anther dehiscence takes place before anthesis. Therefore, although the pollen
have been released from anther they do not disperse as the anthesis has not initiated. The pollen dispersal takes place only
after initiation of anthesis. Hence, the duration of pollen dispersal was taken to represent the pollen presentation time.
The role of petals in pollen dispersal was examined by enumerating the pollen load on petal hair (corona).
The petals of t 102 flowers of different floral stages were examined for the pollen load by using light microscopy method.
To study the stigma receptivity, 2000 buds/flowers of different floral stages were examined by the H2O2 method
(Dafni et al 1998) where the formation of bubbles indicated receptivity of stigma and vice versa. The observed intensity of
bubbling- measured by number of bubbles produced- was then directly proportional to stigma receptivity. To estimate
pollen production per flower, all the anthers of each matured bud (N=120 buds) were put in one ml of distilled water before
anther dehiscence and a homogenous solution was prepared. Pollens from 0.1 µl of this solution were then quantified in a
heamocytometer (B S. 748-I. S. 10269 ROHEM India). Ovule production per flower was separately examined by
dissecting the same 120 flowers. The P/O ratio (pollen produced per ovule) was calculated to examine the reproductive
success of the species. Further, the pollination success was examined by the ratio of pollen load on stigma and pollen
production.
The floral visitors were examined at the three levels. At the first level, the 10-15 trees per site were examined for
floral visitors (the visitor on other parts of the tree were not included) for 15 minutes. This observation was taken at the
interval of 2 hrs and it was carried out for two continuous days. Further, the whole set of observation was carried out twice
in each flowering season for two consecutive flowering season. Among all observed floral visitors, those which were
observed more than five times were treated as major floral visitors. At the second level observation, the visitation pattern of
major floral visitors, identified at the first level, was observed to assess the duration and frequency of visits, movement
pattern among the inflorescences (on a single tree or between different trees) and the foraging location etc. Lastly, at the
third level, the major floral visitors were collected and examined for the pollen load on their body.
42 R. Pandey & Pandey C. N
RESULTS
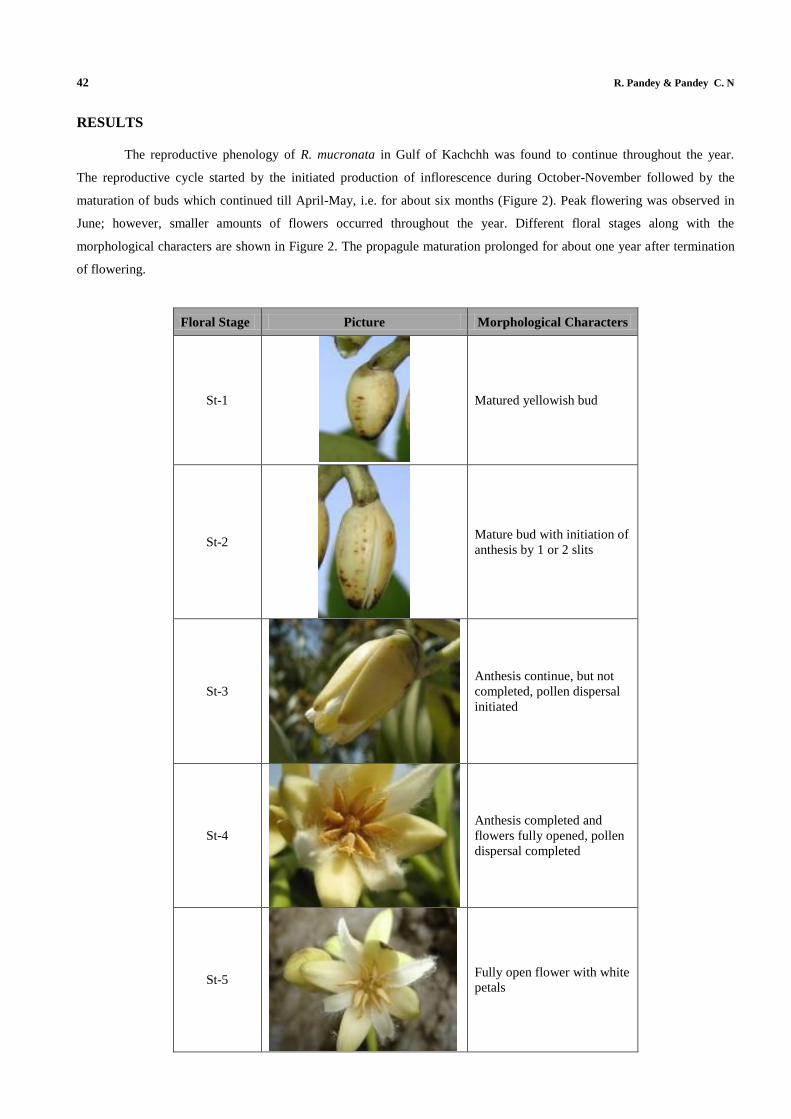
The reproductive phenology of R. mucronata in Gulf of Kachchh was found to continue throughout the year.
The reproductive cycle started by the initiated production of inflorescence during October-November followed by the
maturation of buds which continued till April-May, i.e. for about six months (Figure 2). Peak flowering was observed in
June; however, smaller amounts of flowers occurred throughout the year. Different floral stages along with the
morphological characters are shown in Figure 2. The propagule maturation prolonged for about one year after termination
of flowering.
Floral Stage Picture Morphological Characters
St-1
Matured yellowish bud
St-2
Mature bud with initiation of
anthesis by 1 or 2 slits
St-3
Anthesis continue, but not
completed, pollen dispersal
initiated
St-4
Anthesis completed and
flowers fully opened, pollen
dispersal completed
St-5
Fully open flower with white
petals

Floral Biology and Pollination Ecology of Rhizophora mucronata Lamk. in Gulf of Kachchh, Gujarat, India 43
St-6
Fully open flower with
petals becoming slightly
brown
St-7
Initiation of senescence of
anther and petals
St-8
Completion of senescence of
anther and petals, the sepal
base spotless and
St-9
Sepals are yellow and the
sepal base are light brown
St-10
Sepals are yellow and the
sepal base are dark brown
St-11
Sepals are green and
perpendicular to the
orientation of ovary
Figure 2: The 11 Distinct Floral Stages of R. mucronata
44 R. Pandey & Pandey C. N
FLORAL BIOLOGY
The floral biology was studied by examining process of anthesis, anther dehiscence, pollen presentation, stigma
receptivity and nectar secretion and exploring the temporal relation among them. Anthesis was initiated in St-2 and
observed to be completed in St-4. As mentioned above, to study the different flowering processes, the 24 hours was
divided into four time zones 6 am-12 pm, 12 pm-6 pm, 6 pm-12 am and 12 am-6 am, however, field observations between
12 am and 6 am was not possible.
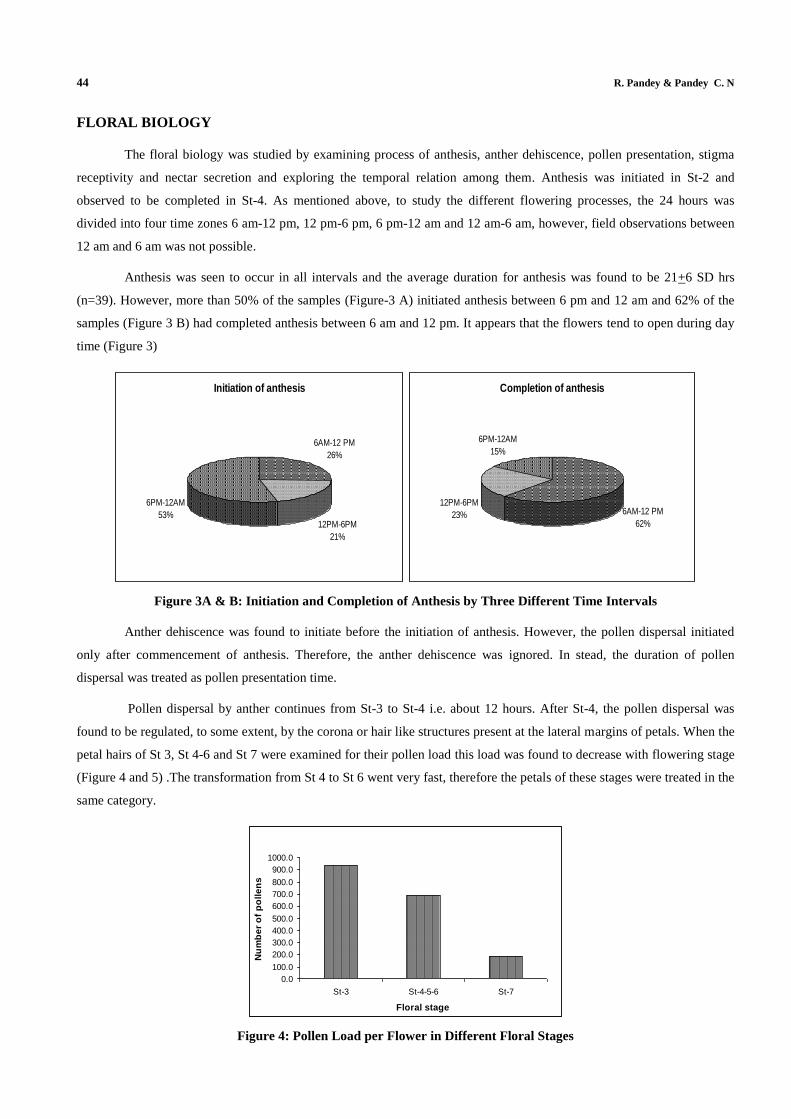
Anthesis was seen to occur in all intervals and the average duration for anthesis was found to be 21+6 SD hrs
(n=39). However, more than 50% of the samples (Figure-3 A) initiated anthesis between 6 pm and 12 am and 62% of the
samples (Figure 3 B) had completed anthesis between 6 am and 12 pm. It appears that the flowers tend to open during day
time (Figure 3)
Initiation of anthesis
6AM-12 PM
26%
12PM-6PM
21%
6PM-12AM
53%
Completion of anthesis
6AM-12 PM
62%
12PM-6PM
23%
6PM-12AM
15%
Figure 3A & B: Initiation and Completion of Anthesis by Three Different Time Intervals
Anther dehiscence was found to initiate before the initiation of anthesis. However, the pollen dispersal initiated
only after commencement of anthesis. Therefore, the anther dehiscence was ignored. In stead, the duration of pollen
dispersal was treated as pollen presentation time.
Pollen dispersal by anther continues from St-3 to St-4 i.e. about 12 hours. After St-4, the pollen dispersal was
found to be regulated, to some extent, by the corona or hair like structures present at the lateral margins of petals. When the
petal hairs of St 3, St 4-6 and St 7 were examined for their pollen load this load was found to decrease with flowering stage
(Figure 4 and 5) .The transformation from St 4 to St 6 went very fast, therefore the petals of these stages were treated in the
same category.
0.0
100.0
200.0
300.0
400.0
500.0
600.0
700.0
800.0
900.0
1000.0
St-3 St-4-5-6 St-7
Floral stage
Nu
mb
er
of
po
lle
ns
Figure 4: Pollen Load per Flower in Different Floral Stages

Floral Biology and Pollination Ecology of Rhizophora mucronata Lamk. in Gulf of Kachchh, Gujarat, India 45
0.0
100.0
200.0
300.0
400.0
500.0
600.0
St- 3-4 St-4-7
Floral stage
Nu
mb
er
of
po
llen
s
Figure 5: Relative % of Pollen Dispersal per Flower in Different Floral Stages
The stigma receptivity initiated in St-4, and continued till St-10, totaling 109 hours. The intensity of receptivity
was low, however up to St 6 (Figure 6) but showed maximum values from St 7 to St 10 (i.e. for about 80 hours). It may be
noted that pollen dispersal from anthers terminated in St 5. However, some pollen mingled in the petal hair and, therefore,
the pollen presentation continues till St 7, but the quantum of pollens available (loaded on petal hair) was very low,
compared to pollen production.
0.0
10.0
20.0
30.0
40.0
50.0
60.0
70.0
80.0
90.0
100.0
St 1
St 2
St 3
St 4
St 5
St 6
St 7
St 8
St 9
St 10
St 11
Floral stages
Re
lati
ve
% o
f in
ten
cit
y o
f s
tig
ma
re
ce
pti
vit
y
Figure 6: Relative % of Stigma Receptivity and its Intensity in Different Floral Stages (the Full Line Indicate the
Value While the Broken Line Indicate the Polynomial Trend Line of 5th
Order)
All the eight anthers were found to produce pollens. The pollen production ranged between 5 and 30 x 105 per
flower with an average production of 12+4 x 105 SD (n=60) (Figure 7). Ovule production, on the other hand, showed no
variation; it was in all cases four ovules per flower (n=60).
0
2
4
6
8
10
12
14
16
18
20
6-8 8-10 10-12 12-14 14-16 16-18 18-20 20-22 22-24
Number of pollen (in lakhs)
Fre
qu
en
cy o
f O
ccu
rren
ce
Figure 7: The Distribution of Pollen Production per Flower

46 R. Pandey & Pandey C. N
All the eight anthers were found to produce pollens. The pollen production ranged between 5 and 30 x 105 per
flower with an average production of 12+4 x 105 SD (n=60) (Figure 7). Ovule production, on the other hand, showed no
variation; it was in all cases four ovules per flower (n=60).
0.0
1.0
2.0
3.0
4.0
St 4/6 St 7 St 8 St 9 St 10
Floral stage
Nu
mb
er
of
po
llen
s
Figure 8: Pollen Grains Observed on Stigma in Different Floral Stages
As mentioned above, reproductive success was calculated as the number of pollen produced per ovule and
showed a high value of 3 X 105. Further, the pollen load on stigma was investigated to examine pollen availability which
was found to be very low compared to pollen production,(Figure 9). Further, the pollen loads on stigma in different floral
stages were also examined (Figure 8). The pollen load on stigma did not show significant variation among the floral stages.
0
5
10
15
20
25
30
35
40
0 1 2 3 4 5 6 7 8 9 10
No of pollens
Fre
qu
en
cy
Figure 9: The Distribution of the Number of Pollen Grains on Stigma
after St- 9
9%St- 9
5%St- 8
9%St- 7
0%
St 4-6
9% St- 3
68%
Figure 10: The Visitation Frequencies of Bees on Flowers in Different Floral Stages

Floral Biology and Pollination Ecology of Rhizophora mucronata Lamk. in Gulf of Kachchh, Gujarat, India 47
Floral Visitors
Though, the spectrum of floral visitors of R. mucronata was found to be diverse, the visitation frequencies of most
species were low. Among various floral visitors, honey bees and ants were found to be the most frequent visitors. Although
beetles were not very frequent visitors they showed the longest visitation durations among all floral visitors. The range of
floral visitation duration was between 2 s and 3 min, 2 s and 75 min, and 2 sec and about 6 hours for honey bees, ants and
beetles, respectively. Further, the floral visitors were most frequently recorded on St 3, St 8, St 9, and St 10. Bees were
found to visit St 3 more frequently (68%) than other floral stages (Figure 10). However, it was also reported to visit flowers
in St 8, St 9 and St 10. The bees were found to forage upon the floral rewards in several floral stages. During visit to the
opening flowers of St 3 they foraged upon anther while in the later floral stages (St 8-10), they foraged upon nectar. Unlike
bees, ants were more frequently observed in the later floral stages (St 8-10) foraging upon the nectar (Figure 11).
St 4-6
6%St- 7
13%
St- 3
6%After St 9
25%
St- 9
31%
St- 8
19%
Figure 11: The Visitation Frequencies of Ants on Flowers in Different Floral Stages
The visitation patterns of the floral visitors were examined in terms of their physical contact with anther and/or
stigma. It was found that in more than 45% (Figure 12 ) of the visits, bees physically contacted only anthers, in more than
25% of the visits they contacted only stigma and in about 30% of the visits they touched both. It may be noted that these
bees have pollen pockets on their hind legs in which the pollens are stored. During landing on the flowers these pollen
pockets come in contact with the floral parts such as anther stigma etc. Hence, in most cases pollens would be present on
their bodies which lead to pollination of stigma when it touch stigma. Therefore in more than 50% of visits
(when they contact stigma or stigma and anther both) there would be a high probability of pollinating the flowers
(Figure 12).
Unlike bees, in about 80% of their visits, ants did not directly contact either anther or stigma. However, in 13% of the
visits they were found to climb on stigma (Figure-13). Since high number of pollens were present at the base of the ovaries,
pollen was recorded on the bodies of the ants. Hence, it is possible that ants too can pollinate stigma.
Figure 12: Relative Visitation Pattern of Bee Figure 13: Relative % Visitation Pattern of Ant

48 R. Pandey & Pandey C. N
The three frequent floral visitors were examined for the pollen load on their body and ants, bees and beetles all
showed positive results.
It may be noted here that ants were mostly reported to forage upon the flowers without physically touching anther
or stigma. Therefore, it appears that ant would be poor pollinators, despite their potential to transport pollen. However,
organisms that prey upon ants may pollinate the flowers.
The chemical composition of the floral exudate was examined by the paper chromatography method
(Harborne 1964) which showed a significant presence of monosaccharides such as glucose and fructose and the
disaccharide sucrose. It showed that the floral exudate is nothing but the floral nectar. The nectar secretion initiated in St 7
and peaked in St 8. After St 8 nectar secretion was almost terminated.
DISCUSSIONS
Pollen dispersal by anthers was found to terminate at St 4 while the pollen dispersal by petals continued till St 6
(less than 10%). Thus pollen was deposited on the petal hairs, forming a bridge between the anther dehiscence and early
stigma receptivity, though, the quantum of pollen present on the petal hair was found to range between 0.06 and 0.01% of
the pollen production by anthers.
Further, stigma receptivity was found to initiate in St 7 (more than 10%). After St 7 stigma receptivity increased
and was found in more than 50% of the flowers. Therefore, pollen dispersal and stigma receptivity were out of phase
indicating a strong possibility of cross pollination (Figure 14). This finding counters the generalized theory that mangroves
are generally self pollination species (Tomlinson 1980).
0
10
20
30
40
50
60
70
80
ST
1
ST
2
ST
3
St
4
ST
5
ST
6
ST
7
ST
8
ST
9
ST
10
ST
11
Floral stages
Perc
en
t o
ccu
rren
ce
A
B
C
Figure 14: Distribution and Temporal Relation among Pollen Dispersal by Anthers (A),
Pollen Dispersal by Petal Hair (B), and Stigma Receptivity (C)
It was observed that bees visited flowers more frequently during St 3 (about 70%) compared to any other floral
stages. In St 3 they foraged upon the anthers but bees were never observed to forage on St 4 and St 5 flowers (Figure 15).
During these stages, pollen dispersal by anthers was less than 30% and pollens were transferred to the petal hair or blown
away with wind and water.
It thus appears that bees would not prefer to forage upon pollens present on petal hairs. However, they certainly
visited the flowers at the later stages.

Floral Biology and Pollination Ecology of Rhizophora mucronata Lamk. in Gulf of Kachchh, Gujarat, India 49
Figure 15: Distriibution and Temporal Relation among Bee Visits (A), ant Visits (B) and Nectar Secretion (C)
Unlike bees, ants were mostly seen to forage on later floral stages such as St 6 to St 11 (Figure 15). During our
observations we found that ants would not allow bees to forage on the same flower. Even if the ant arrived on the flower
after the bee, it would chase away the bee within less than a second. Therefore, as the frequency of visits by ant increased
the frequency of visits by bees decreased.
CONCLUSIONS
R. mucronata is known to be a wind pollinated species. However, the role of floral exudates and frequent floral
visits by insects has not been examined hitherto. The present work suggest a strong possibility of insect pollination and yet
it does not exclude the possibility of wind pollination. In Gulf of Kachchh, the flowering of R. mucronata was taking place
just before the monsoon, when the wind velocity would was very high (15-30 km/hr) Such strong winds may play a
positive as well as a negative role in pollen transfer. Therefore, it appears that the plant has developed the possibility of
insect pollinations and yet retained wind as an alternative pollen vector.
ACKNOWLEDGEMENTS
PCCF Wildlife Gujarat, Staff of marine National Park and Sanctuary. Dr. Salvi Harshad, Mr. Dodiya, Ms. Anjali,
Ms Urvi, Mr. Nilesh, Mr. Amit, Mr Jayesh, Mr. Harshad solanki, Mr Irshad, Mr Arif, Mr. Neeraj. Mr. Chetan, Mr. Prateek,
Ms Puja. Ms Reshama, Mr Jusus Bhai and his team.
REFERENCES
1. Chai P. K. (1982) “Ecological Studies of Mangrove Forest in Sarawak” Ph. D. Thesis University of Malaysia,
Kaula Lumpur.
2. Clarke J. Peter and Myerscough (1991) “Floral biology and reproductive phenology of Avicennia marina in south
eastern Australia” Aust. J. Bot. 39:283-93
3. Coupland G. T. , Paling E. I. and McGuinness K. A. (2005) “Floral abortion and pollination in four species of
tropical from northern Australia” Aquatic Botany 84:2:151-157.
4. Dafni A et. Al. (1998) “A Rapid and Simple Procedure to Determine Stigma receptivity” Plant Sexual Production
1:177-180.
5. Harborne, J. B. (1964) “Phytochemical Methods- A guide to modern techniques of plant analysis” Pub. Springer
(india )Pvt.Ltd pp.238-245

50 R. Pandey & Pandey C. N
6. Pandey C. N. and Pandey R. (2008) “The Status of mangrove in Gujarat” The proceedings of workshop on
Conservation and Management of Mangrove Ecosystem in India: - Stake holder Consultation for Assessment of
Training and Capacity Building Needs and Design of National Strategy and Action Plan.
7. Pandit S. and B. C. Choudhury (2001) “Factors affecting pollinator visitation and Reproductive success in
Sonneratia caseolaris and Aegiceras corniculatum in a mangrove forest in India” Journal of Tropical Ecology
17:3:431-447.
8. Primack, R. B. and p. B. Tomlinson (1980) “Variation in Tropical forests breeding systems” Biotropica
12:229-231.
9. Solomon Raju A. J. , Jonathan K. Henry, Lakshmi A. Vijaya (2006) “Polination Biology of Ceriops decandra
(Griff.) Ding Hou (Rhizophoraceae), an important true viviparous mangrove tree species” Current Science
91:9:1235-1238.
10. Tomlinson P. B. (1980) “The Botany of Mangroves” Pub. Camb. Univ. Pr
Copyright © 2022 FDOKUMEN




















