
Fish Genetics Research in Member Countries and Institutions ...
189
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Fish Genetics Research in Member Countries and Institutions ...


Fish Genetics Researchin Member Countries and Institutionsof the International Networkon Genetics in Aquaculture
Edited byModadugu V. GuptaBelen O. Acosta
2 0 0 1

Fish Genetics Research in Member Countries and Institutions of theInternational Network on Genetics in Aquaculture
Proceedings of the Fifth Steering Committee Meeting of INGAheld on 3-5 March 1999, Kuala Lumpur, Malaysia
Edited byModadugu V. GuptaBelen O. Acosta
2001
Published by ICLARM – The World Fish CenterPO Box 500 GPO10670 Penang, Malaysia
Gupta, M.V. and B.O. Acosta, Editors. 2001. Fish Genetics Research in Member Countriesand Institutions of the International Network on Genetics in Aquaculture. ICLARM Conf.Proc. 64, 179 p.
Perpustakaan Negara Malaysia. Cataloguing in Publication Data
Fish genetics research in member countries and institutions of the internationalnetwork on genetics in aquaculture/[research coordinator Modadugu V. Gupta].ISBN 983-2346-05-31. Fisheries - Research. 2. Aquaculture - Research.I. Gupta, Modadugu V.338.372707
ISBN 983-2346-05-3
ICLARM Contribution No. 1614
Printed by Juta Print, Penang, Malaysia
ICLARM – The World Fish Center is one of the 16international research centers of the ConsultativeGroup on International Agricultural Research(CGIAR) that has initiated the public awarenesscampaign, Future Harvest.

CONTENTS
FOREWORD vM.J. Williams
PREFACE viiM.V. Gupta
Networking in aquaculture genetics research 1M.V. Gupta and B.O. Acosta
Aquaculture genetics research in Bangladesh 7M.G. Hussain and M.A. Mazid
Aquaculture genetics research in China 15Li Sifa
Fish genetics research in Côte d’Ivoire 2 5V. Yapi-Gnaore
Aquaculture genetics research in Egypt 29H. Elghobashy
A comparative evaluation of two tilapia strains in Fiji 35S. Nandlal, C.W. Morris, M. Lagibalavu and E. Ledua
Aquaculture genetics research in India: an overview 43S. Ayyappan, A.G. Ponniah, P.V.G.K. Reddy, R.K. Jana,K.D. Mahapatra and Y. Basavaraju
Aquaculture genetics research in Indonesia 51A. Hardjamulia, F. Sukadi, Subagyo and R. Gustiano
Aquaculture genetics research in Malawi 61A. Ambali
Genetics for improvement of fish in Malaysia 65T.K. Mukherjee
Fish genetics research and development in the Philippines 71A. Camacho, T. Abella and M. Tayamen
Aquaculture genetics research in Thailand 77N. Pongthana

Review on fish genetics and breeding research in Vietnam 9 1T. M. Thien, N. C. Dan and P. A. Tuan
Fish genetics research at ICLARM - The World Fish Centre 9 7M.V. Gupta, B.O. Acosta, R. Dunham and P.R. Gardiner
Israeli aquaculture genetic improvement programs 103G. Hulata
Gene mapping, marker-assisted selection, gene-cloning, genetic engineering 109and integrated genetic improvement programs at Auburn UniversityZ. J. Liu
Gene banking and common carp breeding program in Hungary 119S. Gorda and L. Varadi
Aquatic genetic resource activities of the Fisheries Department, 1 2 3Food and Agriculture OrganizationD. Bartley
Activities at AKVAFORSK of importance to the INGA 129T. Gjedrem and T. Svealv
Overview of Fish Genetics Research at Queensland University of Technology 133P. B. Mather
Genetics research at SEAFDEC Aquaculture Department 141Z. Basiao
Overview of aquaculture genetics research in the Institute of Aquaculture, 145University of StirlingD.J. Penman
Research on the genetics of aquatic organisms at the School of Biological 1 5 1Sciences, University of Wales SwanseaG.C. Mair and J.A. Beardmore
Genetics research at Wageningen University and Research Centre, 161the NetherlandsH. Komen, M. Tanck and N. Ruane
SPECIES INDEX 169
LIST OF PARTICIPANTS 1 7 5

v
FOREWORD
With a growing world population, especially in developing countries,achieving and improving food security calls for well-targeted actions. Fishare an important food, and fisheries provide employment for many.Declining fish stocks in oceans, rivers and lakes pose a threat to those whodepend on catching fish for daily sustenance, and who depend onemployment in fisheries, particularly artisanal fisheries.
Aquaculture is one sector that contributes significantly to world foodproduction. As natural fish stocks decline, it is crucial to improveproduction from aquaculture, to improve production efficiency, and toimprove aquaculture species by genetic enhancement.
The International Network on Genetics in Aquaculture (INGA) wasestablished in 1993 to increase the quantity of fish protein consumed bylow-income rural and urban people in tropical developing countries throughefficient breeding and selection programs that will improve aquacultureproduction and the income of resource-poor fish farmers. Thirteendeveloping countries, nine advanced scientific institutions and threeinternational and regional organizations make up INGA’s membership.
The INGA network has been a catalyst for substantial progress in geneticsresearch in member countries. These 13 countries produce 83 per cent ofworld aquaculture output. Some of them are already reaping the benefitsfrom their investment in genetics research in the form of more productivestrains of fish.
This proceedings provides a detailed and valuable insight into currentaquaculture genetics. The papers were prepared for the Fifth INGA SteeringCommittee meeting held in March 1999 in Kuala Lumpur, Malaysia. Themeeting, kindly hosted by the Malaysian Department of Fisheries and theUniversity of Malaya, was significant because it brought together scientistsfrom both developing and developed countries, permitting a wide exchangeof the latest advances in genetics research in aquaculture.
Meryl J. WilliamsDirector GeneralICLARM – The World Fish Center

vi

vii
PREFACE
This publication incorporates the information drawn from aquaculturegenetics research undertaken or in progress in Member Countries andAssociate Member Institutions of the International Network on Geneticsin Aquaculture (INGA). INGA has a membership of 13 countries in Asia,the Pacific and Africa, and 12 advanced scientific institutions. The SteeringCommittee of INGA consists of senior research planners from membercountries and institutions as well as ICLARM. The Committee meets atregular intervals to discuss progress in aquaculture genetics, identify areasfor possible collaboration, and share information.
The Fifth Steering Committee of INGA hosted by the Malaysian Departmentof Fisheries and the University of Malaysia was held in Kuala Lumpur,from 3-5 March 1999. The meeting was attended by all members andassociate members of the network. The papers presented at this meetingwere informative and contained a wealth of information on the state ofaquaculture genetics research in the respective countries and institutions .Hence, INGA/ICLARM undertook to publish the papers presented at themeeting. Papers presented at the meeting in 1999 were revised by theirrespective authors to incorporate the developments that have taken placesince then.
Aquaculture genetics research is in its infancy in many developing countriesand there is an urgent need for collaboration among institutions in thedeveloping and developed countries for synergy in developing better breedsof fish that can contribute to increased production and food security whileensuring the conservation of aquatic biodiversity. All the members andassociate members of INGA have been playing an active and importantrole in this and in supporting network activities. Close collaboration amongnetwork members is evident from the exchange of germplasm followingmaterial transfer agreements and quarantine protocols and collaborativeresearch projects that have been developed. On behalf of INGA I thankall those who have contributed to this volume.
Modadugu V. GuptaResearch Coordinator, INGA

viii

1Networking in Aquaculture Genetics Research
NETWORKING INAQUACULTURE
GENETICS RESEARCHa
M.V. Gupta and B.O. AcostaICLARM – The World Fish Center
P.O. Box 500, GPO 10670, Penang, Malaysia
GUPTA, M.V. and B.O. ACOSTA. 2001. Networking in aquaculture genetics research, p. 1-5 In M.V. Gupta and B.O. Acosta(eds.) Fish genetics research in member countries and institutions of the International Network on Genetics inAquaculture. ICLARM Conf. Proc. 64, 179 p.
ABSTRACT
Aquaculture genetics research is in its infancy compared to that of crops and livestock. Multidisciplinary researchcoupled with networking are effective tools in genetics research especially in developing countries.
Variability in fish species, farming systems and production environments made it imperative for genetic enhancementof aquaculture species to be done through regional and international cooperation. Realizing this, ICLARM-the WorldFish Center initiated the International Network on Genetics in Aquaculture (INGA) in 1993. At present, the networkhas a membership of 13 countries from Asia, the Pacific and Africa and 12 advanced scientific institutions.
In the short time since its inception, the network has made significant progress in: assisting member countries indeveloping national breeding programs; initiating two regional research programs for genetic improvement of carpsand Nile tilapia; assisting the transfer of germplasm between member countries for research and to disseminateimproved strains; assisting the formation of national genetics networks; assisting in capacity building among developingcountry scientists; and providing information on genetics research and policy issues in biodiversity conservation,intellectual property rights, etc.
Background
About a billion people, mostly in developing countries,depend on fish as a primary source of animal protein.Global fish production reached 122 million t in 1996,of which only 90 million t were used for humanconsumption (FAO 1999). FAO estimates that by theyear 2010, demand for food fish would increase by13.5-18.0% or to about 105-110 million t. Annualgrowth in capture fisheries production decreased from
6% in the 1950s and 1960s to 0.6% in 1995 and 1996(FAO 1999), suggesting an increase in dependence onproduction from aquaculture.
New (1991) estimated that the world’s projectedrequirement for aquaculture products would increasefrom 19.6 million t in the year 2000 to 62.4 million tin 2025. Based on FAO statistics (1998), the 13 membercountries (Bangladesh, China, Côte d’Ivoire, Egypt,Fiji, Ghana, India, Indonesia, Malaysia, Malawi,
a ICLARM Contribution No. 1612

Fish Genetics Research in Member Countries and Institutions of INGA2
Philippines, Thailand and Vietnam) of the Inter-national Network on Genetics in Aquaculture (INGA)collectively produced 25.7 million t of fish fromaquaculture in 1998, accounting for 83.3% of world’saquaculture production.
Concomitant with technological advances inaquaculture are the problems associated withenvironmental degradation, genetic deterioration ofcultured stocks and decline in aquatic biodiversitylimiting options for new species and strains foraquaculture and diminishing natural productivity andresilience of aquatic ecosystems. A review of world’sfisheries indicate that the potential contribution ofaquaculture could only be realized if a number ofissues, including genetic improvement of aquaculturespecies and biodiversity, are addressed (FAO 1995).
Remarkable progress has been made in improving theproductivity of crops and livestock during the last 3-4decades through breeding and selection. However, inspite of the long history of fish farming, it was only inrecent years that efforts have been made to harnessthe benefits of genetic enhancement in fish.Norwegian scientists have amply demonstrated thepossibility of increasing productivity of farmed salmonand trout through an efficient breeding program. In1995, salmon farming produced a combined outputgreater than the sum of the country’s meat productionfrom swine, cattle and poultry (Gjoen and Bentsen1997).
It was ascertained, however, whether the same benefitscould be obtained in the case of tropical and sub-tropical finfish which contribute about 90% of theglobal aquaculture production. This uncertainty poseda challenge to scientists to develop techniques for thegenetic improvement of tropical finfish used inaquaculture.
The collaborative research project for the geneticimprovement of Nile tilapia (Oreochromis niloticus)implemented by ICLARM in collaboration with thePhilippine and Norwegian institutions, indicated thepotential of increased production through selectivebreeding. In its sixth generation of selection, the GIFTstrain had demonstrated 77% faster growth and 60%higher survival compared with farmed strains in thePhilippines (Eknath and Acosta 1998; Eknath et al.1998). Subsequent evaluation of the second generationGIFT strain in Bangladesh, China, Thailand andVietnam showed that the GIFT strain had highergrowth rates than local strains, the difference ranging
from 18% in China to 66% in Bangladesh. Analysis ofoverall potential impact of improved fish in the fivecountries indicated that the adoption of improvedstrain would result in reduced cost of production andincreased fish production (Dey 2000).
Overall, the genetic gains per generation and per yearof coho, rainbow trout, Atlantic salmon, channelcatfish and tilapia was estimated at about 15% and5%, respectively (Gjedrem 1997). Gjedrem’sassumption was that if efficient breeding programswere introduced throughout the aquaculture industry,the aquaculture production by the year 2025 wouldbe higher than the projected requirement foraquaculture products.
In Asia, evidences of inbreeding and unintentionalselection among hatchery-based and establishedfarmed stocks have been found (Pullin and Capili1988; McAndrew et al. 1993; Eknath 1995). In Africa,freshwater species are on the verge of extinction,despite it being considered as the world’s repositoryof diverse freshwater fish fauna. There are alsouncontrolled fish transfers and introduction of exoticfish in many countries, threatening local aquaticresources.
Realizing that multidisciplinary research is an effectivetool in genetics and breeding and networking is aproven mechanism for international cooperation,national institutions in developing countries in Asia,Africa and ICLARM formally established theInternational Network on Genetics in Aquaculture(INGA) in 1993 (Seshu et al. 1994). At present, INGAhas a membership of 13 countries from Asia, Africaand the Pacific (Bangladesh, China, Côte d’Ivoire,Egypt, Fiji, Ghana, India, Indonesia, Malawi, Malaysia,Philippines, Thailand and Vietnam) and 11 advancedscientific institutions (Fig. 1). The following is a briefreport on activities being undertaken by INGA andthe important role it has been playing throughconsolidation of strengths of its members.
Network Activities
INGA acts as catalyst for genetics research beingundertaken by national programs. In 1997, the 13member countries reviewed the declining fish catchesfrom natural resources and came up with the “ManilaResolution” (Box 1) stressing the need for concertedregional and international efforts for advancing fishbreeding and genetics through cooperation (The INGAPlanning Meeting 1997). To accomplish this, the

3
regularly reviewed by the network (Table 1).
Initiation of regional research programs
Genetic improvement of carps in Asia
In carp culture, diverse farming systems andsocioeconomic scenarios prevail in various countriesin Asia. About 20 carp species are extensively culturedand for each species, several traits have been identifiedas having improved (Gupta et al. 1998). Since many ofthe carp species are common to aquaculture systems inAsian countries, the six major carp-producing membercountries of INGA (Bangladesh, China, India,Indonesia, Thailand and Vietnam) recognized that acollaborative effort would be better than each countrytrying to improve all their species. This has resultedin the implementation of the regional project forgenetic improvement of 4 species of carps (C. carpio,M. amblycephala, Labeo rohita, B. gonionotus) with fundingfrom the Asian Development Bank and ICLARM.
Collaborative tilapia genetic research in Africa
In Africa, INGA/ICLARM with start-up funds providedby the International Development Research Center(IDRC), Canada, initiated a regional collaborative
following major activities have been and are beingcarried out.
Development of nationalbreeding programs
INGA has been assisting member countries in thedevelopment of plans and strategies for theimplementation of national fish breeding programsbased on their respective needs and resources. Thenetwork and the Norway’s Institute of AquacultureResearch (AKVAFORSK) have assisted the AquacultureResearch Institute Nos. 1 and 2 in Vietnam indeveloping breeding plans for: (1) Nile tilapia (O.niloticus) through combined multitrait selection; (2)silver barb (Barbodes gonionotus) through individual(mass) selection to improve growth; and (3) mrigal(Cirrhinus mrigala) through individual selection toimprove growth rate.
Assistance was also provided to the Research Institutefor Freshwater Fisheries, Indonesia, in developingbreeding plans for: (1) common carp (Cyprinus carpio)using multitrait selection; (2) Nile tilapia (GIFT strain)using individual selection; and (3) milkfish (Chanoschanos) using individual selection.The breeding programs in member countries are also
Fig. 1. Member countries and associate member institutions of INGA.
Networking in Aquaculture Genetics Research

Fish Genetics Research in Member Countries and Institutions of INGA4
research project aimed at documentation of tilapiagenetic resources, including indigenous knowledge,genetic characterization of tilapia strains and initiationof genetic improvement of tilapias. Institutions in Côted’Ivoire, Egypt, Ghana and Malawi are implementingthe research.
Strengthening of national research capacity
Since genetics research started only in recent years inmember countries of the network, emphasis is beinglaid in capacity building through organization oftraining programs in quantitative genetics by ICLARM/INGA in collaboration with AKVAFORSK.
Member countries of INGA do not have the same levelof scientific training and expertise. Aquaculture andgenetics research is generally more advanced in Asiathan in Africa. In view of this, INGA has beenorganizing cross-country visits for senior scientists ofparticipating countries.
Exchange of fish germplasm
INGA has been assisting the member countries inexchange of germplasm for evaluation, direct use inaquaculture or utilization in breeding programs.Protocols and quarantine procedures formulated bythe network based on international codes of practiceand material transfer agreements (Box 2) are beingfollowed in transfer of germplasm through thenetwork.
The network has coordinated the following exchangesof genetic materials:• Improved O. niloticus (GIFT strain) from the
Philippines to Bangladesh, China, Fiji, India,Indonesia, Thailand and Vietnam;
• O. niloticus strains from Côte d’Ivoire and Kenyato Egypt;
• O. aureus and O. niloticus from Egypt to China;• Improved C. carpio strain from Vietnam to
Bangladesh, India, Philippines and Thailand;• Diverse stocks of B. gonionotus from Indonesia and
Thailand to Bangladesh;• C. mrigala from India to Vietnam; and• Improved L. rohita strain from India to Thailand.
Country Species Activity
Bangladesh B. gonionotus Selective breeding, line crossing and development of monosex female populationC. catla, L. rohita Selective breeding and line crossing experiments for genetic improvement
China C. carpio, M. amblycephala Mass selection for improvement of growthCôte d’Ivoire O. niloticus Evaluation of local strains for selective breedingEgypt O. niloticus Evaluation of local strains for selective breedingFiji O. niloticus Performance evaluation of Fijian tilapia and GIFT strainGhana O. niloticus Evaluation of local strains for selective breedingIndia L. rohita Combined family and within family selection for further genetic improvement
C. carpio Production of sterile and monosex populationIndonesia C. carpio Diallele crossing experiment of gynogenetic common carpMalawi O. shiranus Mass selection for genetic improvementMalaysia Red tilapia, O. niloticus Selective breedingPhilippines O. niloticus Continuation of multitrait (growth and frequency of spawning females) selectionThailand B. gonionotus Mass selection for improvement of growth
C. carpio Mass selection for improvement of growthVietnam C. carpio Family selection for improvement of growth
B. gonionotus Establishment of base population for selective breeding
Table 1. Progress of breeding programs in member countries of INGA.
Box 1. Manila Resolution adopted by INGA members.
The participants of the Planning Meeting of INGA resolve toencourage the INGA member countries, donors and otherinstitutions/organizations to give due emphasis for conservationand sustainable use of aquatic genetic resources andbiodiversity by:
• organizing national aquaculture genetics networks andproviding support for coordination of activities;
• providing support to national genetics research programs;• strictly adhering to the germplasm transfer and quarantine
protocols approved by the network members which conformto wider international standards and rigorously implementingthe guidelines;
• sharing knowledge and methodologies;• sharing germplasm, with prior informed consent and on
mutually agreed terms;• providing technical and policy inputs to national and
international efforts to maintain aquatic biodiversity andgenetic resource conservation;
• assisting in the implementation of the Convention onBiological Diversity (CBD); and
• providing support to INGA activities.

5
Box 2. Material Transfer Agreement.
A country planning to import new or exotic species has to signa Material Transfer Agreement which states that the recipientagrees:
• to abide by the provisions of the Convention on BiologicalDiversity;
• to preclude further distribution of germplasm to locations atwhich it could have adverse environmental impact;
• to not claim ownership over the material received; nor seekintellectual property rights over the germplasm or relatedinformation;
• to ensure that any subsequent person or institution to whomthey make samples of germplasm available is bound by thesame provision;
• that the responsibility to comply with the country’s biosafetyand import regulations and any of the recipient country’srules governing the release of genetic materials is entirelyits own;
• to follow the quarantine protocols suggested by ICLARM/INGA; and
• to abide by the International Code of Transfer of Germplasmin case germplasm is transferred beyond the boundaries ofthe country.
Formation of national networks
INGA assisted in the formation of national networks,e.g., in India, Indonesia, Malawi, Malaysia and thePhilippines. The international network also providesits local counterparts opportunities and linkagesnecessary for research and development.
Information dissemination
There is a fairly large amount of information availableon genetics research. However, many of the institutionsin member countries do not have access to thisinformation. Hence the network has been assisting themembers with dissemination of information. Throughthe network, member countries can have access toreports, reprints and other materials on fish geneticsresearch. The network has also been publishing itsNewsletter in Naga, ICLARM quarterly.
References
Dey, M.M. 2000. The impact of genetically improved farmed Niletilapia in Asia. Aquacult. Econ. Manage. 4(1&2): 107-123.
Eknath, A.E. 1995. Managing aquatic genetic resources.Management example 4: the Nile tilapia, p.176–194. In J.Thorpe, G. Gall, J.E. Lannan and C.E. Nash (eds.)Conservation of fish and shellfish resources: managingdiversity. Academic Press, Harcourt Brace Company, London.
Eknath, A.E. and B.O. Acosta. 1998. Genetic Improvement ofFarmed Tilapias (GIFT) Project Final Report, March 1998 toDecember 1997. International Center for Living AquaticResources Management, Makati City, Philippines.
Eknath, A.E. M.M. Dey, M. Rye, B. Gjerde, T.A. Abella, R. Sevilleja,M.M. Tayamen, R.A. Reyes and H.B. Bentsen. 1998. Selectivebreeding of Nile tilapia for Asia, p. 89-96. In Proceedings ofthe Sixth World Congress of Genetics Applied to LivestockProduction. Armidale, Australia.
FAO. 1995. Review of the state of world fishery resources:aquaculture. FAO Fish. Circ. No. 886. Food and AgricultureOrganization of the United Nations, Rome, Italy.
FAO. 1998. Fishery statistics – aquaculture production. Vol. 86/2.Food and Agriculture Organization, Rome, Italy. 169 p.
FAO. 1999. The state of world fisheries and aquaculture: 1998. Foodand Agriculture Organization, Rome, Italy. 112 p.
Gjedrem, T. 1997. Selective breeding to improve aquacultureproduction. World Aquacult. 28 (1) : 33 - 45.
Gjoen, H.M. and H.B. Bentsen. 1997. Past, present and future ofgenetic improvement in salmon aquaculture. ICES J. Mar. Sci.54: 1009–1014.
Gupta, M.V., M.M. Dey, R. Dunham and G. Bimbao, Editors. 1997.Proceedings of the Collaborative Research and Training onGenetic Improvement of Carp Species in Asia. Bhubaneswar,India. ICLARM Work. Doc. No. 1.
The INGA Planning Meeting. 1997. Manila Resolution:strengthening partnerships to advance the science of fishbreeding and genetics and development of national fishbreeding programs. ICLARM Conf. Proc. 54, 12 p.
McAndrew, B.J., K,J. Rana and D.J. Penman. 1993. Conservationand preservation of genetic variation in aquatic organisms, p.295-336. In J.F. Muir and R.J. Roberts (eds.) Recent advancesin aquaculture IV. Blackwell Scientific Publications, Oxford,UK.
New, M.B. 1991. Turn of the millenium aquaculture: navigatingtroubled waters or riding the crest of the wave? World Aquacult.22(3): 28-49.
Pullin, R.S.V. and J.B. Capili. 1988. Genetic improvement of tilapias:problems and prospects, p. 259-266. In R.S.V. Pullin, T.Bhukaswan, K. Tonguthai and J.L. Maclean (eds.) The SecondInternational Symposium on Tilapia in Aquaculture. ICLARMConf. Proc. 15, 623 p.
Seshu, D.V., A.E. Eknath and R.S.V. Pullin. 1994. InternationalNetwork on Genetics in Aquaculture. ICLARM, Manila,Philippines.
Networking in Aquaculture Genetics Research

Fish Genetics Research in Member Countries and Institutions of INGA6

7Aquaculture Genetics Research in Bangladesh
AQUACULTUREGENETICS RESEARCH
IN BANGLADESH
M.G. Hussain and M.A. MazidBangladesh Fisheries Research Institute
Mymensingh 2201, Bangladesh
HUSSAIN, M.G. and M.A. MAZID. 2001. Aquaculture genetics research in Bangladesh, p. 7-14. In M.V. Gupta and B.O.Acosta (eds.) Fish genetics research in member countries and institutions of the International Network on Geneticsin Aquaculture. ICLARM Conf. Proc. 64, 179 p.
ABSTRACT
Aquaculture productivity has declined in recent years in Bangladesh. The reasons for this are habitat degradation,loss of genetic diversity and stock deterioration in hatchery populations due to poor broodstock management andinbreeding. In consonance with the government’s plans to rehabilitate inland fisheries in the country, the BangladeshFisheries Research Institute through its Freshwater Station, initiated fish genetics research in 1986.
Making effective breeding plans for commercially important carps and other fish species is an important researcharea for the development of genetically improved broodstock. A new dimension of this research has begun with theinvolvement of international agencies such as the International Center for Living Aquatic Resources Management,Australian Centre for International Agricultural Research/Commonwealth Scientific and Industrial ResearchOrganization and Institute of Aquaculture, University of Stirling, Scotland.
The major research programs being implemented are: genetic stock improvement of endemic (Catla catla andLabeo rohita) and exotic (Barbodes gonionotus) carps, catfish (Heteropneustes fossilis) and genetically improvedGIFT strain Nile tilapia (Oreochromis niloticus); population genetics of an anadromous clupeid, hilsa shad (Tenualosailisha); and genetic conservation of some endemic threatened fish species. This paper presents the highlights ofthese research programs in Bangladesh and future plans.
Introduction
Stock deterioration in hatchery populations causedby poor broodstock management and inbreeding hasbeen observed in recent years in Bangladesh.Retarded growth, reduction in reproductiveperformance, morphological deformities, increasedincidence of diseases and mortalities of hatchery-produced seed have been reported. This has resultedin deterioration of carps and barb seed quality. There
is widespread concern that stocking of such geneticallypoor quality seed in floodplains and related openwater bodies for fishery enhancement might causeserious feral gene introgression in the wild stocks.
To avoid loss of genetic diversity and inbreedingdepression in hatchery populations, the developmentof improved broodstocks through implementation ofeffective breeding plans for commercially importantcarps and other fish species has been identified by

Fish Genetics Research in Member Countries and Institutions of INGA8
Table 1. Endemic fish species of Bangladesh (Hussain 1998).
Family Species Common name Local name
Cyprinidae Labeo rohita Rohu RuiCatla catla Catla KatlaCirrhinus mrigala Mrigal MrigalLabeo calbasu Calbashu KalibaushLabeo bata Bata BataLabeo gonious Gonious GonniaPuntius sarana Barb SarpuntiTor putitora Tor mahseer Mahashoal
Claridae Clarias batrachus Asian catfish MagurSiluridae Ompok pabda Pabda PabdaBagridae Mystus cavasius Gulsha GolshaAnabantidae Anabas testudineus Climbing perch KoiHeteropneustidae Heteropneustes fossilis Asian catfish Shingi
the Bangladesh Fisheries Research Institute (BFRI)as an important area for research. Research wasinitiated with technical and financial support fromthe International Center for Living AquaticResources Management (ICLARM); AustralianCentre for International Agricultural Research(ACIAR)/Commonwealth Scientific and IndustrialResearch Organization (CSIRO), Australia; andInstitute of Aquaculture (IOA), University ofStirling, Scotland. Genetic evaluation of improvedGIFT strain Nile tilapia (Oreochromis niloticus) wasundertaken during 1994-1996. Genetic stockimprovement of two selected carp species, silverbarb (Barbodes gonionotus) and catla (Catla catla), hasbeen in progress since January 1997 under theauspices of ICLARM. A collaborative researchprogram with IOA to produce all-female populationof B. gonionotus , through gynogenesis and sexreversal, has been initiated. The population geneticstructure of hilsa shad (Tenualosa ilisha) inBangladesh is being investigated in collaborationwith ACIAR/CSIRO, Australia. Other programsbeing implemented include the selective breedingof rohu (Labeo rohita), further genetic selection anddevelopment of all-male population of GIFT strainO. niloticus , genetic manipulation in catfish(Heteropneustes fossilis) and genetic conservation ofendangered fish species like Labeo gonius, Cirrhinusreba, Channa striata and others.
Endemic carps and other fish species
There are at least 13 species of carps and otherspecies under six genera inhabiting Halda and
Padma-Brahmaputra River systems. Of thesespecies, eight species of carps and barbs are usedfor artificial seed production in hatcheries (Table1). Breeding techniques have been developed forthe other five species: Clarias batrachus, Ompok pabda,Mystus cavasius, Anabas testudineus, and H. fossilis.
Exotic carps and other fish species
Several exotic fish species have been introducedin Bangladesh since 1960. Such introductionswere not properly recorded. Table 2 lists theintroduced fish species which are bred in hatcheriesfor seed production.
Genetics Research in Progress
Carp species
Breeding plans for stock improvement of catlaand rohu
Land races of C. catla and L. rohita were collectedfrom Halda, Jamuna and Brahmaputra riversystems, and were investigated for differences inextrinsic genetic traits by means of morphologicaland growth assessment. During the spawning seasonof 1999, two wild L. rohita stocks (from Jamuna andBrahmaputra Rivers) were mated with existinghatchery stocks to produce three crossbreds (i.e.,hatchery! x Jamuna"; Jamuna! x Brahmaputra";and Brahmaputra! x Jamuna") and three purebredlines (i.e., hatchery! x hatchery" ; Jamuna! xJamuna" ; and Brahmaputra! x Brahmaputra" ).With all these lines, selective breeding and linecrossing program will be continued, which will resultin the development of genetically improved strain,
Freshwater Fish Species Used forArtificial Seed Production
9Aquaculture Genetics Research in Bangladesh
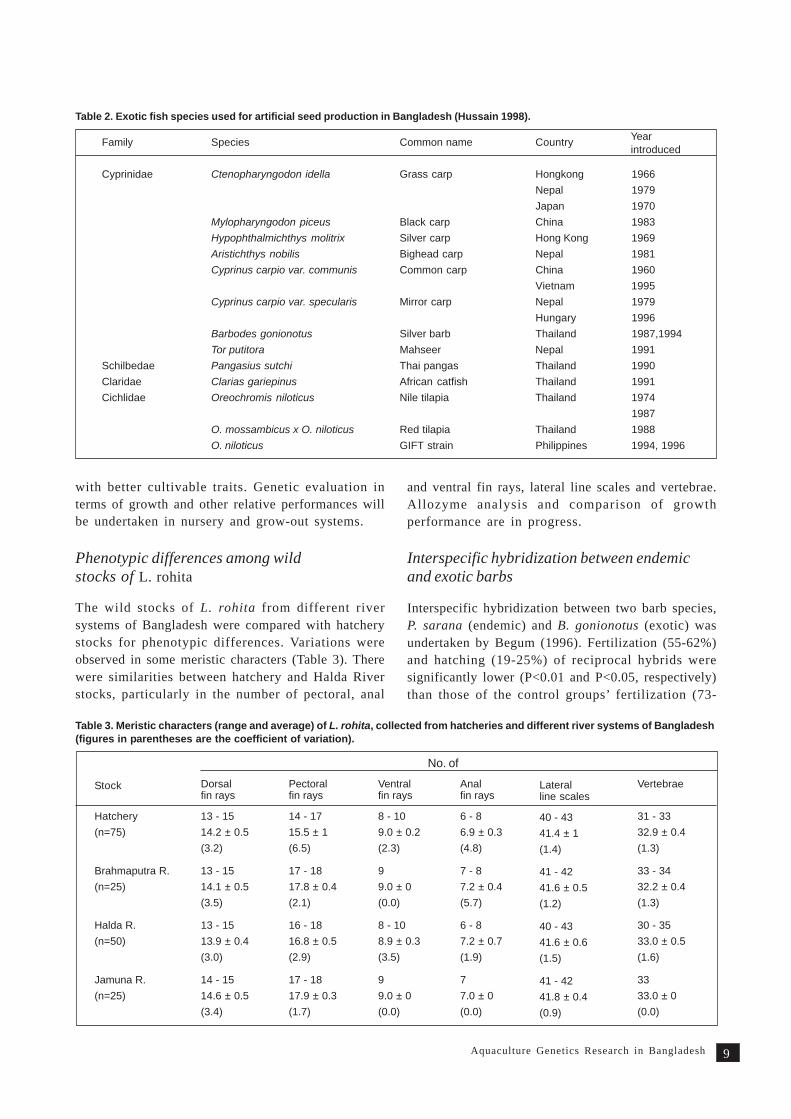
Table 3. Meristic characters (range and average) of L. rohita, collected from hatcheries and different river systems of Bangladesh(figures in parentheses are the coefficient of variation).
Stock
Hatchery(n=75)
Brahmaputra R.(n=25)
Halda R.(n=50)
Jamuna R.(n=25)
Dorsalfin rays
13 - 1514.2 ± 0.5(3.2)
13 - 1514.1 ± 0.5(3.5)
13 - 1513.9 ± 0.4(3.0)
14 - 1514.6 ± 0.5(3.4)
Pectoralfin rays
14 - 1715.5 ± 1(6.5)
17 - 1817.8 ± 0.4(2.1)
16 - 1816.8 ± 0.5(2.9)
17 - 1817.9 ± 0.3(1.7)
Ventralfin rays
8 - 109.0 ± 0.2(2.3)
99.0 ± 0(0.0)
8 - 108.9 ± 0.3(3.5)
99.0 ± 0(0.0)
No. ofAnalfin rays
6 - 86.9 ± 0.3(4.8)
7 - 87.2 ± 0.4(5.7)
6 - 87.2 ± 0.7(1.9)
77.0 ± 0(0.0)
Lateralline scales
40 - 4341.4 ± 1(1.4)
41 - 4241.6 ± 0.5(1.2)
40 - 4341.6 ± 0.6(1.5)
41 - 4241.8 ± 0.4(0.9)
Vertebrae
31 - 3332.9 ± 0.4(1.3)
33 - 3432.2 ± 0.4(1.3)
30 - 3533.0 ± 0.5(1.6)
3333.0 ± 0(0.0)
Family Species Common name Country
Cyprinidae Ctenopharyngodon idella Grass carp Hongkong 1966Nepal 1979Japan 1970
Mylopharyngodon piceus Black carp China 1983Hypophthalmichthys molitrix Silver carp Hong Kong 1969Aristichthys nobilis Bighead carp Nepal 1981Cyprinus carpio var. communis Common carp China 1960
Vietnam 1995Cyprinus carpio var. specularis Mirror carp Nepal 1979
Hungary 1996Barbodes gonionotus Silver barb Thailand 1987,1994Tor putitora Mahseer Nepal 1991
Schilbedae Pangasius sutchi Thai pangas Thailand 1990Claridae Clarias gariepinus African catfish Thailand 1991Cichlidae Oreochromis niloticus Nile tilapia Thailand 1974
1987O. mossambicus x O. niloticus Red tilapia Thailand 1988O. niloticus GIFT strain Philippines 1994, 1996
Yearintroduced
Table 2. Exotic fish species used for artificial seed production in Bangladesh (Hussain 1998).
with better cultivable traits. Genetic evaluation interms of growth and other relative performances willbe undertaken in nursery and grow-out systems.
The wild stocks of L. rohita from different riversystems of Bangladesh were compared with hatcherystocks for phenotypic differences. Variations wereobserved in some meristic characters (Table 3). Therewere similarities between hatchery and Halda Riverstocks, particularly in the number of pectoral, anal
and ventral fin rays, lateral line scales and vertebrae.Allozyme analysis and comparison of growthperformance are in progress.
Interspecific hybridization between two barb species,P. sarana (endemic) and B. gonionotus (exotic) wasundertaken by Begum (1996). Fertilization (55-62%)and hatching (19-25%) of reciprocal hybrids weresignificantly lower (P<0.01 and P<0.05, respectively)than those of the control groups’ fertilization (73-
Phenotypic differences among wildstocks of L. rohita
Interspecific hybridization between endemicand exotic barbs

Fish Genetics Research in Member Countries and Institutions of INGA10
Fig.1. Design for mating and production of F1 broodstock from the base population derived from reciprocal cross
of B. gonionotus.
Communal stocking with 125larvae per pair and rearing in
nursery / rearing ponds
Mating of 3-4 pairs of breedersfor each 6 reciprocal crosses
Incubation of eggs inseparate funnel units
Communal stocking with 695larvae per pair and rearing in
nursery / rearing ponds
Communal stocking offingerlings in grow-out
ponds
Growth evaluation ofcommunal stock with
control group
Mass selection of at least20% fish as broodstock forfurther selective breeding
Mass selection of at least20% fish as broodstock forfurther selective breeding
Communal stocking offingerlings in grow-out ponds
81%) and hatching (49-56%). Both control groupsproduced significantly higher (P<0.05) percentageof normal embryos (52-59%) compared with bothhybrids (23-24%). No variat ions were foundbetween the two control and hybrid groups.
Genetic improvement of B. gonionotus
B. gonionotus was first introduced from Thailandin 1977, and subsequently in 1994 wild germplasmwas introduced from Thailand and Indonesiawith the help of ICLARM. Studies are in progressto develop a genetically improved and al l-female population of B. gonionotus using thesethree founder stocks through selective breedingand genetic manipulation techniques underprojects supported by ICLARM and theDepartment for International Development (UK),respectively (Hussain 1997).
For genetic improvement of B. gonionotus throughselective breeding and line crossing techniques,germplasm obtained from Thailand and Indonesiaand exist ing local stocks were used. Therepresentative breeders of F1 generations of thethree unrelated stocks were crossed through 3 x 3diallele crossing to produce nine genetic groups.The rationale of this crossing scheme is to form aheterogeneous outbred base population for the
breeding program. For each of the reciprocalcrosses, five to eight pairs were mated separatelyand the best three progeny (larvae) groups wereselected to make 18 F1 full sib progeny familieswhich were then communally stocked by mixing 125and/or 695 larvae from each family. Subsequently,they were transferred to communal grow-out pondswhere stocking density was maintained at 0.75 to1.0 fish/m 2 (Fig. 1) . A separate growth tr ialexperiment was conducted for eight months withcommunal F1 crossbred and exist ing purebred(Bangla x Bangla) strain in four chambered earthenponds with two replicates at a stocking density of3 fish/m3. At harvest, the maximum and minimumaverage weight gains of communal crossbredgroups were 77.26 g and 49.21 g, respectively,whereas those for the control groups were 61.47 gand 56.13 g, respectively (Table 4). The data ofmonthly mean weight showed superior growthrate in crossbreeds but was not significantlydifferent (P>0.05).
During the spawning season of 1998, 20% of largestfemales (at least 150 pairs) were selected frommature communal F1 crossbreeds and used in theproduction of F2 generation (Fig. 2). A comparativegrowth evaluation of F 2 crossbred and controlgroups (Bangla x Bangla) in nursery and grow-outsystems indicated that the former had higher

11Aquaculture Genetics Research in Bangladesh
growth in terms of weight at an early age than thelatter, but this was not statistically significant. Ingrow-out ponds, the crossbred group showedsignificantly higher growth (7-8%).
During the spawning season of 1999, 15% of largestfemales and males were selected from the matureF2 crossbred progenies and used them (182 pairs)for the generation of F3 progenies. At the same time,78 pairs of nonselected control (existing Bangla xBangla) breeders were separately used to producecontrol progeny group. The on-station comparativegrowth evaluation trial data revealed that the
average genetic gain in terms of body weightachieved by the selected group (F3) was 22-25% overnonselected control group.
Like other cyprinids, female B. gonionotus hasbetter growth than males; therefore, a programfor mass production of all female populationthrough gynogenesis and sex reversal was initiatedin 1995 in collaboration with the University ofStirling, Scotland, and the National AquacultureGenetic Research Institute (NAGRI), Thailand.The protocol for such approach was to produceneomale (phenotypic male having XX genotype)
Fig. 2. Design for mating of selected broodstock and production of F2 generation of B. gonionotus and theirevaluation for growth.
Pool breeding of at least150 pairs of mass selected breeders
in several batches (25 - 30 pairs per batch)
Hatching of eggs
Rearing of progenies ingrow-out ponds
Pool breeding of at least50 pairs of nonselected breeders
(existing stock)
Hatching of eggs
Nursing of early fry at astocking density of 25/m2
Evaluation for growth inpartitioned ponds
Nursing of early fry at astocking density of 25/m2
Rearing ofprogenies in
grow-out ponds
On-station andon-farm evaluation of growth
performance of progeniesbetween selected and
nonselected control pairs
Sale of hatchlings/fry tointerested grow-out farmers
Table 4. Comparison of performance between crossbred and control groups of B. gonionotus .
Month
InitialOctoberNovemberDecemberJanuaryFebruaryMarchAprilMay
Group 13.86 ± 1.04
17.41 ± 3.7025.17 ± 6.4032.50 ± 17.8940.97 ± 10.1343.56 ± 10.0646.25 ± 14.5755.24 ± 12.4777.26 ± 29.49
Group 2 3.86 ± 1.0416.58 ± 4.0921.53 ± 4.1423.79 ± 7.6033.42 ± 9.6036.11 ± 8.8939.76 ± 12.1045.40 ± 11.6249.21 ± 13.56
Average Weight (g)
Group 1 3.49 ± 0.8820.01 ± 3.9424.02 ± 4.7728.48 ± 5.1136.18 ± 11.0040.23 ± 9.7542.56 ± 13.4545.60 ± 14.4961.47 ± 14.05
Group 2 3.49 ± 0.8813.43 ± 4.0117.68 ± 3.3321.92 ± 8.5132.07 ± 9.2435.58 ± 11.2239.37 ± 12.1848.80 ± 11.7156.13 ± 13.96
Communal F1 hybrid Control group
Fish Genetics Research in Member Countries and Institutions of INGA12
through feeding androgen hormone treated feedto the gynogenetic fish and then crossing neo-males with normal females for mass production ofall female seeds.
Neomales of B. gonionotus were imported fromNAGRI for mating them with females belongingto wi ld germplasm of Thai land and Indones ia( in t roduced in 1994 for se lec t ive breedingprogram). Product ion of addi t ional batches ofneomale through gynogenesis and sex reversal wasinitiated (Pongthana et al. 1998).
The putative monosex female progeny groupsproduced were tested on-station and on-farm.Putative monosex females produced from newlyproduced neomales and mixed sex control fishattained weights of 55-105 g in on-station trials, while40-80 g in on-farm trials. The sex ratio in theharvested neomale progenies varied from 50% to 90%female. The male individuals present in the neomaleprogenies could be of second generation neomale orthere might have been some complexities in the sexdetermination, XY systems.
Sexual dimorphism for weight in B. gonionotuswas initially done through sampling of differentpopulations from different culture systems. Thesexual dimorphism indices (ratio of mean weightof females to mean weight of males , whichis expressed as percentage) for weight inB. gonionotus at the age of one year class rangedbetween 1.1 and 1.7 in different populations andcul ture systems. Proximate composi t ion of the
carcass of female and male f i sh indica tedinsignif icant differences (P>0.05) in moisture ,protein and ash content, except fat (Azad 1998).
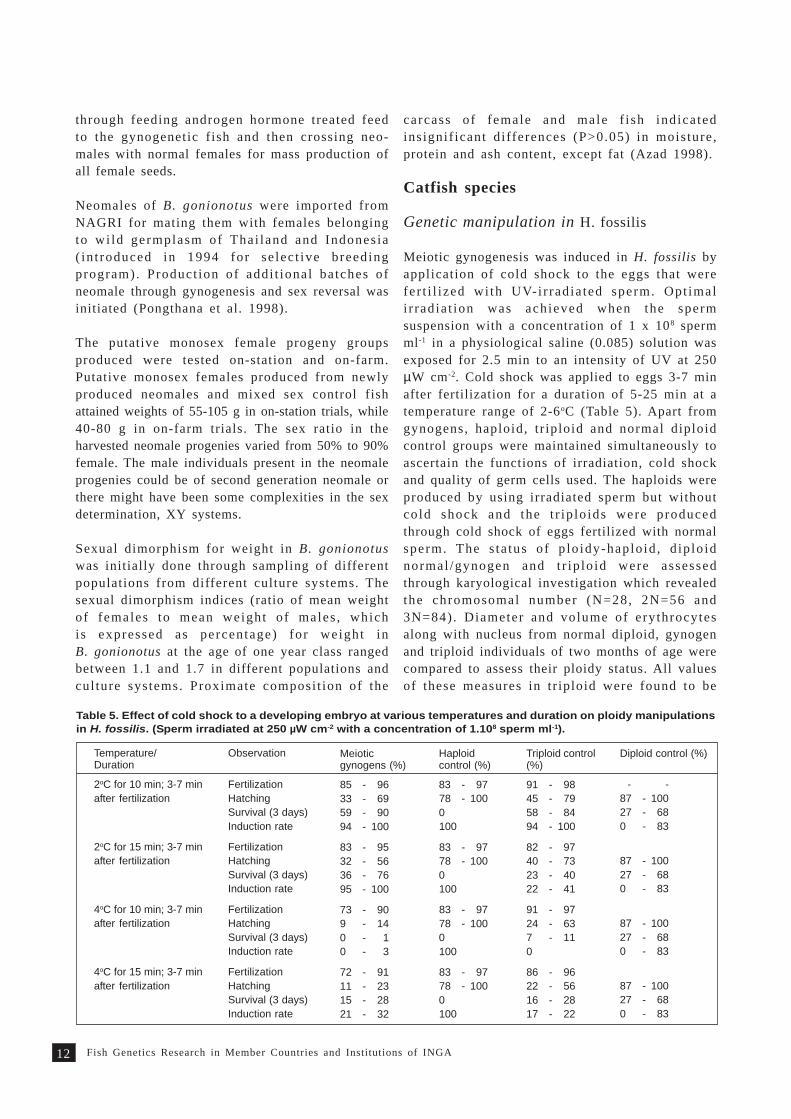
Catfish species
Genetic manipulation in H. fossilis
Meiotic gynogenesis was induced in H. fossilis byapplication of cold shock to the eggs that werefer t i l ized wi th UV-i r radia ted sperm. Opt imali r radia t ion was achieved when the spermsuspension with a concentration of 1 x 108 spermml-1 in a physiological saline (0.085) solution wasexposed for 2.5 min to an intensity of UV at 250µW cm-2. Cold shock was applied to eggs 3-7 minafter fertilization for a duration of 5-25 min at atemperature range of 2-6oC (Table 5). Apart fromgynogens, haploid, t r iploid and normal diploidcontrol groups were maintained simultaneously toascertain the functions of irradiation, cold shockand quality of germ cells used. The haploids wereproduced by using irradiated sperm but withoutcold shock and the t r ip lo ids were producedthrough cold shock of eggs fertilized with normalsperm. The s ta tus of p lo idy-haplo id , d ip lo idnormal /gynogen and t r ip lo id were assessedthrough karyological investigation which revealedthe chromosomal number (N=28, 2N=56 and3N=84) . Diameter and volume of erythrocytesalong with nucleus from normal diploid, gynogenand triploid individuals of two months of age werecompared to assess their ploidy status. All valuesof these measures in tr iploid were found to be
Table 5. Effect of cold shock to a developing embryo at various temperatures and duration on ploidy manipulationsin H. fossilis. (Sperm irradiated at 250 µW cm-2 with a concentration of 1.108 sperm ml-1).
Temperature/Duration
2oC for 10 min; 3-7 minafter fertilization
2oC for 15 min; 3-7 minafter fertilization
4oC for 10 min; 3-7 minafter fertilization
4oC for 15 min; 3-7 minafter fertilization
Observation
FertilizationHatchingSurvival (3 days)Induction rate
FertilizationHatchingSurvival (3 days)Induction rate
FertilizationHatchingSurvival (3 days)Induction rate
FertilizationHatchingSurvival (3 days)Induction rate
Meioticgynogens (%)
85 - 9633 - 6959 - 9094 - 100
83 - 9532 - 5636 - 7695 - 100
73 - 909 - 140 - 10 - 3
72 - 9111 - 2315 - 2821 - 32
Haploidcontrol (%)
83 - 9778 - 1000100
83 - 9778 - 1000100
83 - 9778 - 1000100
83 - 9778 - 1000100
Triploid control(%)
91 - 9845 - 7958 - 8494 - 100
82 - 9740 - 7323 - 4022 - 41
91 - 9724 - 637 - 110
86 - 9622 - 5616 - 2817 - 22
Diploid control (%)
- -87 - 10027 - 680 - 83
87 - 10027 - 680 - 83
87 - 10027 - 680 - 83
87 - 10027 - 680 - 83

13Aquaculture Genetics Research in Bangladesh
s igni f icant ly h igher (P<0.05) than tha t ofcorresponding normal d ip lo id and gyno-genet ic d ip lo id , but ins igni f icant d i fferences(P>0.05) were found between the last two groups.A slow development of embryo and also delayedhatching were observed in gynogenet ic andtriploid groups (Gheyas 1998).
Administration of androgen hormone 17 alpha-methyltestosterone (MT) orally to the fry ofO. niloticus from first feeding stage to 40 days atdifferent doses ranging from 60 to 100 µg/g of feedin hapas, pond and aquarium resulted in 75-95%males. Seven to 10-day old fry of GIFT strainO. niloticus, when administered orally with MThormone at a dose of 60 and 100 µg/g of feed andSRT-99 (MT 10%) feed obtained from thePhilippines for 3 weeks resulted in 95-100% males(n=50) in the batch that were administered with adose of MT 100 µg/g feed whereas 85% males wereobtained in the batch fed with MT 60 µg/g feed. SRT-99 hormone treated feed also resulted in 95-100%males (n=50).
The population genetic structure of T. ilisha whichconstitutes 25% of the total fish production inBangladesh was investigated using starch gelelectrophoresis. The research was done incollaboration with ACIAR/CSIRO, Australia. Fishsamples were collected from different geographicalareas within and beyond Bangladesh (Table 6).Starch gel electrophoresis of different enzymes ofsamples of various tissues was initially attempted toinvestigate allozyme variation. Five loci - Idh-l, Idh-m, Mdh-l, Mdh-m and Pgm - were polymorphic overthe six locations studied. Levels of variation werelow but significant differences in allele frequencieswere detected among Kuwait, Bangladesh andIndonesia (Table 7). However, allozyme data, so far,cannot discriminate genetically distinct stocks ofhilsa within Bangladesh (Hussain et al. 1998).
The International Union for the Conservation ofNature, Bangladesh (1998) documented 11 cyprinid
species as critically or fairly endangered in thecountry (Table 8), indicating a need for thedevelopment of artificial breeding techniques for theconservation of the carp’s “gene pool”. Attemptswere made to develop artificial propagationtechniques for Tor putitora and Puntius sarana, andvery recently for L. gonius and C. reba.
Future Research Plans
Following are plans for aquaculture genetics researchin Bangladesh:• Genetic characterization of wild land races of
C. catla and L. rohita;• Initiation of genetic stock improvement of C.
catla and L. rohita using selective breeding and
Conservation of endangeredcarp species
Production of al l -male populat ionof GIFT strain O. niloticus
Populat ion genet ic structure ofhilsa shad, Tenualosa ilisha Table 7. Results of G statistics tests (similar to contingency
X2 tests) for homogeneity of alleles. Ho: all alleles are drawnfrom the same homogeneous population.
Comparisons
Within BangladeshKuwait vs. Bangladesh vs.IndonesiaKuwait vs. IndonesiaKuwait vs. BangladeshIndonesia vs. Bangladesh
P
NSP<0.001
P<0.001P<0.001P<0.001
Significant loci
NoneMdh-m, Pgm, Idh-l
Idh-1Mdh-m, PgmMdh-m, Pgm, Idh-l
Table 6. Locations and sample sizes used in theelectrophoretic study.
Location
KuwaitBangladesh
KhulnaAricha GhatChandpurCox’s Bazar
Indonesia
Number
4765
70698233
Table 8. List of endangered carp and barb species ofBangladesh.
Scientific name
Labeo nandinaL. bogaL. goniusL. bataL. pangusiaL. calbasuCirrhinus rebaPuntius saranaP. tictoTor torT. putitora
Local name
NandinaBhanganGhoniaBataGhora maachKalbausLaachu, bataSarpuntiTit puntiMahasholMahashol
Fairlyendangered
XXXXX
X
Criticallyendangered
XX
X
XX

Fish Genetics Research in Member Countries and Institutions of INGA14
line crossing techniques;• Evaluation of growth performance of F4 putative
genetically improved B. gonionotus;• Genetic sex determination and production of all-
female population of B. gonionotus throughgynogenesis and sex reversal;
• Continuation of selection of GIFT strainO. niloticus for further genetic improvement;
• Optimization of sex reversal technique for GIFTstrain O. niloticus;
• Continuation of genetic manipulation in H. fossilis,particularly production of all-female populationthrough inducing homogametic neomales; and
• Continuation of population genetics study ofT. ilisha.
References
Azad, M.A.K.1998. A preliminary study on the sexual dimorphismfor weight of silver barb, Puntius gonionotus Bleeker.Bangladesh Agricultural University, Mymensingh,Bangladesh. 97 p. M.S. thesis.
Begum, S. 1996. Studies on the hybridization between Puntiussarana and Puntius gonionotus and their embryonicdevelopment. Bangladesh Agricultural University,Mymensingh, Bangladesh. 80 p. M.S. thesis.
Gheyas, A.A. 1998. Studies on cold shock induced gynogenesisand artificial breeding performance in Heteroneustes fossilisBloch. Bangladesh Agricultural University, Mymensingh,Bangladesh. 132 p. M.S. thesis.
Hussain, M.G. 1997. Current status of carp genetic research andbreeding practices in Bangladesh, annex 2.1. In M.V. Gupta,M.M. Dey, R. Dunham and G. Bimbao (eds.) Proceedings ofthe Collaborative Research and Training on GeneticImprovement of Carp Species in Asia, 26-29 July 1997,Central Institute of Freshwater Aquaculture, Bhubaneswar,India. ICLARM Work. Doc. 1, 12 p. (Unpublished).
Hussain, M.G. 1998. Broodstock management and breeding plansto control negative selection and inbreeding in hatcherystocks: BFRI concepts. Paper presented at the Workshop onBroodstock Management for Genetic Maintenance andImprovement, 27-28 April 1998, Dhaka, Bangladesh.
Hussain, M.G., J.P. Salini, M.S. Islam and M.A. Mazid. 1998.Genetic structure of hilsa shad population: preliminary resultsusing starch gel allozymes. In M.A. Mazid and S.J.M. Blaber(eds.) Proceedings of Hilsa Research Project Workshop. (Inpress).
IUCN, Bangladesh. 1998. List of threatened animals ofBangladesh. Paper presented at the Special Workshop onBangladesh Red Book of Threatened Animals, 22 February1998, Dhaka, Bangladesh.
Pongthana, N., D.J. Penman, P. Baoprasertkul, M.G. Hussain, M.S.Islam, S.F. Powel and B.J. McAndrew. 1998. Monosex femaleproduction in the silver barb (Puntius gonionotus Bleeker).Aquaculture 173: 247-256.

15
ABSTRACT
China is the largest aquaculture country. The total output of fisheries in 1998 reached 38.2 million t,in which the share of aquaculture was 57%. This paper gives a brief review of the aquaculture genetics researchin China, including genetic characterization, breeding for genetic improvement and conservation. The majorachievements were breeding of common carp (Cyprinus carpio) (such as red carp, jing carp) and crucian carp(Carassius auratus) (such as allogynogenetic crucian carp) in early 1980s; gene engineering in late 1980s; andgenetic conservation efforts in 1990s. These efforts contributed to the development of aquaculture in China.
AQUACULTUREGENETICS RESEARCH
IN CHINA
S. LiShanghai Fisheries University
Shanghai, 200090 China
LI, S. 2001. Aquaculture genetics research in China, p. 15-24. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research inmember countries and institutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc.64, 179 p.
Introduction
Fish production in China has increased rapidly since1979. It reached 38 million t or about 30% of theworld production in 1998. Developing more rapidlythan capture fisheries, aquaculture’s contribution tototal fish production increased from 26% in 1978 to57% in 1998.
Since marine capture fisheries has maintained zerogrowth since 1999, the increase in fish productioncomes entirely from aquaculture. Thus, geneticimprovement of aquaculture species is expected toplay a major role in the coming century. This papersummarizes the status of genetic research in China.
Genetic Characterization
Chinese carps
The genetic characterization of silver carp (Hypo-pthalmichthys molitrix), bighead carp (Aristichthys
Aquaculture Genetics Research in China
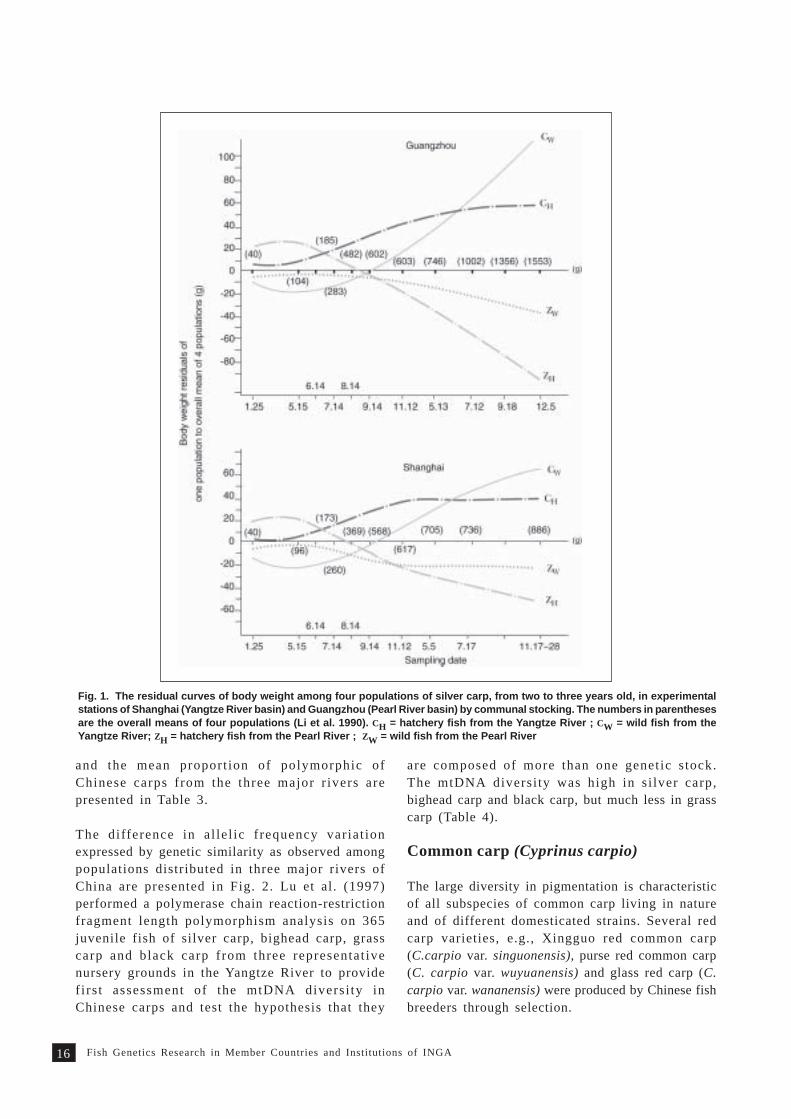
nobilis), grass carp (Ctenopharyngodon idella) andblack carp (Mylopharyngodon piceus) has been studiedin detail since 1980s (Li et al. 1990; Li 1998). Thegrass carp, silver carp and bighead carp from threemajor rivers of China, i.e., Yangtze River, Pearl Riverand Amur River, were studied. Results indicated thatfish in Yangtze River, grow fastest. The growthdifferences in wild populations of silver carp fromYangtze River and Pearl River and hatcherypopulations from Yangtze River and Pearl Riverbasins are presented in Fig. 1. This finding has ledto understanding of selection.
The diploid number of chromosomes in Chinesecarps is 48, but the composition of chromosomes isdifferent (Table 1). By biochemical genetic study,the LDH is coded by two loci, Ldh-A and Ldh-B,and shows five bands in muscle, eye, heart andbrain. Besides loci Ldh-A and Ldh-B, there is Ldh-C in liver showing one major band. The relativeactivity and mobili ty of bands are different inChinese carps (Table 2). The average heterozygosity
Fish Genetics Research in Member Countries and Institutions of INGA16
and the mean proport ion of polymorphic ofChinese carps from the three major r ivers arepresented in Table 3.
The difference in al le l ic f requency variat ionexpressed by genetic similarity as observed amongpopulations distributed in three major rivers ofChina are presented in Fig. 2. Lu et al. (1997)performed a polymerase chain reaction-restrictionfragment length polymorphism analysis on 365juvenile fish of silver carp, bighead carp, grasscarp and black carp from three representat ivenursery grounds in the Yangtze River to providefirs t assessment of the mtDNA diversi ty inChinese carps and test the hypothesis that they
are composed of more than one genetic stock.The mtDNA diversi ty was high in si lver carp,bighead carp and black carp, but much less in grasscarp (Table 4).
Common carp (Cyprinus carpio)
The large diversity in pigmentation is characteristicof all subspecies of common carp living in natureand of different domesticated strains. Several redcarp varieties, e.g., Xingguo red common carp(C.carpio var. singuonensis), purse red common carp(C. carpio var. wuyuanensis) and glass red carp (C.carpio var. wananensis) were produced by Chinese fishbreeders through selection.
Fig. 1. The residual curves of body weight among four populations of silver carp, from two to three years old, in experimentalstations of Shanghai (Yangtze River basin) and Guangzhou (Pearl River basin) by communal stocking. The numbers in parenthesesare the overall means of four populations (Li et al. 1990). CH = hatchery fish from the Yangtze River ; CW = wild fish from theYangtze River; ZH = hatchery fish from the Pearl River ; ZW = wild fish from the Pearl River

17
The biochemical genetic study indicated that theaverage heterozygosity and the mean proportionof polymorphism of Chinese carps of the abovevarieties are slightly different (Table 5).
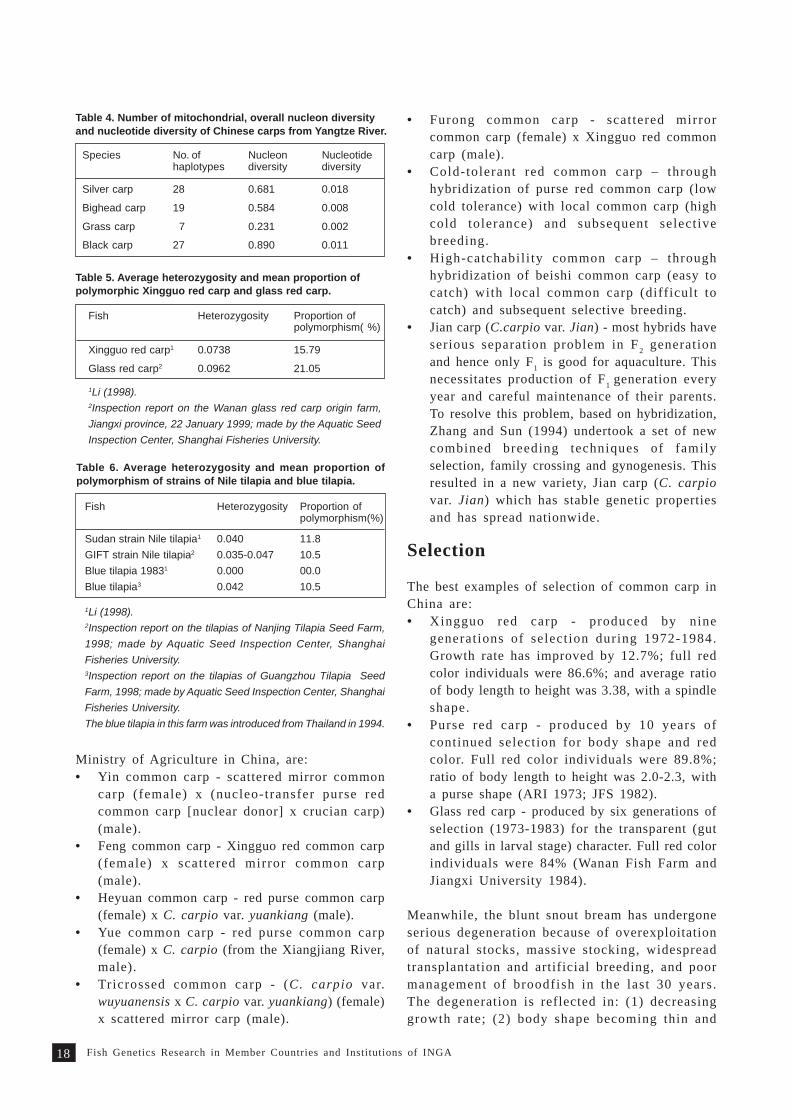
Crucian carp (Carassius auratus)
Crucian carp is the fifth most important speciesin freshwater aquaculture in China. In 1996, itsproduction reached 690 000 t. Silver crucian carp(C. auratus gibelio) and Penzhe crucian carp (C. a.var pengzenensis) are widely used in aquacultureproduction and genetic studies in China. In silvercrucian carp, the average heterozygosity and meanproportion of polymorphism are zero (Li 1998).
Blunt snout bream(Megalobrama amblycephala)
Blunt snout bream (Wuchang fish) was first recordedin 1960s from the natural fauna of Liangtze Lakein the middle-reach of Yangtze River. Subsequently,it was introduced into large areas as a principalcultured species. Its production reached 380 000 tin 1996. Its average heterozygosity is 0.756 andmean proportion of polymorphism is 19.6% (Li 1997).
Tilapias
The ti lapias cultured in China are the Sudanstrain (introduced in 1978), GIFT strain of Niletilapia (Oreochromis niloticus) (1994, from thePhilippines), and blue tilapia (O. aureus) (1983, fromUSA). The average heterozygosity and meanproportion of polymorphism of the above strainsare different (Table 6).
Breeding for Genetic Improvement
Hybridization
Artificial hybridization among cyprinids of differentfamilies and genera was carried out extensivelyduring the last three decades in China. Availableinformation indicates that 112 crosses were carriedout among fish of three orders, five families, and32 species. Of these, hybrids among populations ofcommon carp usually showed some heterosis. Someof these hybrids with valuable characteristics foraquaculture, and have been certificated by the
Aquaculture Genetics Research in China
Heilongjiang R.
Changjiang R.
Heilongjiang R.
Changjiang R.
A
B
Zhujiang R.
Zhujiang R.
(%)
(%)100 80 60 40
Relative genetic similarity
100 80 60 40 20
Fig. 2. Dendrogram showing genetic similarity amongpopulations of silver carp (a) and grass carp (b) in Yangtze(Changjiang), Pearl (Zhujiang) and Amur (Heilongjiang) Rivers(Li et al. 1990).
Species Heterozygosity Proportion ofpolymorphism ( %)
Silver carp 0.0484 - 0.0591 11.8 - 23.5
Bighead carp 0.0772 - 0.1133 15.8 - 29.4
Grass carp 0.0454 - 0.1076 20.0 - 33.3
Black carp 0.0010 - 0.0350 5.9 - 11.8
Table 3. Average heterozygosity and mean proportion ofpolymorphism in Chinese carps from three major river systemsof China (Li 1998).
Table 2. Relative activity (RA, %) and relative mobility (RM, %)of LDH in muscle of Chinese carps (Li 1998).
Species BandA4 A3B1 A2B2 A1B3 B4
Silver carp RA 45.0 32.3 18.6 2.8 1.4RM 46.8 61.0 72.2 87.0 100.0
Bighead carp RA 54.1 26.0 15.9 2.0 1.9RM 46.7 57.9 68.8 85.7 100.0
Grass carp RA 47.2 19.1 13.5 5.2 1.9RM 7.1 36.3 47.5 77.8 100.0
Black carp RA 41.9 20.2 11.5 2.9 2.6
RM 25.3 41.4 61.6 79.8 100.0
Table1. Composition of chromosomes of Chinesecarps (Li 1998).
Species Metacentrics Submetacentrics Subtelocentrics
Silver carp 10 9 5Bighead carp 12 9 3Grass carp 13 7 4Black carp 14 4 6
Fish Genetics Research in Member Countries and Institutions of INGA18
Ministry of Agriculture in China, are:• Yin common carp - scattered mirror common
carp (female) x (nucleo-transfer purse redcommon carp [nuclear donor] x crucian carp)(male).
• Feng common carp - Xingguo red common carp(female) x scattered mirror common carp(male).
• Heyuan common carp - red purse common carp(female) x C. carpio var. yuankiang (male).
• Yue common carp - red purse common carp(female) x C. carpio (from the Xiangjiang River,male).
• Tricrossed common carp - (C. carpio var.wuyuanensis x C. carpio var. yuankiang) (female)x scattered mirror carp (male).
• Furong common carp - scattered mirrorcommon carp (female) x Xingguo red commoncarp (male).
• Cold-tolerant red common carp – throughhybridization of purse red common carp (lowcold tolerance) with local common carp (highcold tolerance) and subsequent selectivebreeding.
• High-catchabil i ty common carp – throughhybridization of beishi common carp (easy tocatch) with local common carp (difficult tocatch) and subsequent selective breeding.
• Jian carp (C.carpio var. Jian) - most hybrids haveserious separation problem in F2 generationand hence only F1 is good for aquaculture. Thisnecessitates production of F1 generation everyyear and careful maintenance of their parents.To resolve this problem, based on hybridization,Zhang and Sun (1994) undertook a set of newcombined breeding techniques of familyselection, family crossing and gynogenesis. Thisresulted in a new variety, Jian carp (C. carpiovar. Jian) which has stable genetic propertiesand has spread nationwide.
Selection
The best examples of selection of common carp inChina are:• Xingguo red carp - produced by nine
generations of selection during 1972-1984.Growth rate has improved by 12.7%; full redcolor individuals were 86.6%; and average ratioof body length to height was 3.38, with a spindleshape.
• Purse red carp - produced by 10 years ofcontinued selection for body shape and redcolor. Full red color individuals were 89.8%;ratio of body length to height was 2.0-2.3, witha purse shape (ARI 1973; JFS 1982).
• Glass red carp - produced by six generations ofselection (1973-1983) for the transparent (gutand gills in larval stage) character. Full red colorindividuals were 84% (Wanan Fish Farm andJiangxi University 1984).
Meanwhile, the blunt snout bream has undergoneserious degeneration because of overexploitationof natural stocks, massive stocking, widespreadtransplantation and artificial breeding, and poormanagement of broodfish in the last 30 years.The degeneration is reflected in: (1) decreasinggrowth rate; (2) body shape becoming thin and
Fish Heterozygosity Proportion ofpolymorphism(%)
Sudan strain Nile tilapia1 0.040 11.8GIFT strain Nile tilapia2 0.035-0.047 10.5Blue tilapia 19831 0.000 00.0Blue tilapia3 0.042 10.5
1Li (1998).2Inspection report on the tilapias of Nanjing Tilapia Seed Farm,1998; made by Aquatic Seed Inspection Center, ShanghaiFisheries University.3Inspection report on the tilapias of Guangzhou Tilapia SeedFarm, 1998; made by Aquatic Seed Inspection Center, ShanghaiFisheries University.The blue tilapia in this farm was introduced from Thailand in 1994.
Table 6. Average heterozygosity and mean proportion ofpolymorphism of strains of Nile tilapia and blue tilapia.
Species No. of Nucleon Nucleotidehaplotypes diversity diversity
Silver carp 28 0.681 0.018
Bighead carp 19 0.584 0.008
Grass carp 7 0.231 0.002
Black carp 27 0.890 0.011
Table 4. Number of mitochondrial, overall nucleon diversityand nucleotide diversity of Chinese carps from Yangtze River.
Fish Heterozygosity Proportion ofpolymorphism( %)
Xingguo red carp1 0.0738 15.79
Glass red carp2 0.0962 21.05
1Li (1998).2Inspection report on the Wanan glass red carp origin farm,Jiangxi province, 22 January 1999; made by the Aquatic SeedInspection Center, Shanghai Fisheries University.
Table 5. Average heterozygosity and mean proportion ofpolymorphic Xingguo red carp and glass red carp.

19
long; (3) decrease in proportion of edible part; and(4) early maturity. In 1986, a selection programwas started, which aimed at protecting this fish’sgenetic resource, preventing degeneration andimproving performance in aquaculture. Theobjective is to produce a new strain with bettergrowth and body shape.
The foundation population of bream was obtainedfrom Yuli Lake in Hubei Province in 1985 and 1986.From this population, two lines were established,and mass selection was undertaken. As a result, thefifth generation from the two lines showed 29.1%higher growth than the base population. Theselection response was 35.8% for each generation.The ratio of standard length to height of body was2.27, similar to the original strain.
A study indicated that improper managementand breeding result in decreased growth rate. Forexample, a blunt snout bream inbred in threeconsecutive generations had a growth rate 16% lowerthan the original fish. Heterozygosity also decreased.
Polyploid breeding
In China, polyploid breeding research was initiatedin the middle 1970s. Hydrobiology Institute,Academia Sinica, was the first to use cold shock andchemical induction to obtain triploids of grass carpand grass carp x blunt snout bream hybrid.Subsequently, heat shock and hydropressure wereapplied to induce triploids but very few were used inproduction. There is interest in the production oftetraploid fish. Crossing tetraploid fish with diploidfish results in triploid fish that could be used inaquaculture production. So long as tetraploid fishcould mate with each other and get tetraploidprogeny, the former could continue generating. Wuet al. (1981, 1988) produced a tetraploid carp bycrossbreeding Xingguo red carp (female) x grass carp(male), then backcrossing the hybrid and grass carp.The resultant tetraploid carp showed strong diseaseresistance. Gui et al. (1990,1991) produced tetraploidcrystal-colored crucian carp by hydropressure andcold shock. This was a creative idea in polyploidinduction. However, the two carps were not used inaquaculture production. Liu Yun and his colleagues(pers. comm.) found several tetraploid crucian carps
during their selection study, which were used toproduce the triploid crucian carp, commerciallycalled the Xiangyuan crucian carp.
Haploid breeding
Since the 1970s, China has produced more than 10gynogenetic species, including grass carp, silver carp,common carp, loach (Misgurnus anguillicaudatus),crucian carp, rainbow trout (Oncorhynchus mykiss), etc.
The crucian carp, C. auratus gibelio, is a naturalgynogenetic fish. Jiang et al. (1982) used sperm ofXingguo red common carp to “fertilize” the ova ofsilver crucian carp, and obtained allogynogeneticcrucian carp, with fast growth and good shape. Thisvariety is highly preferred by producers andconsumers in China. This practice of gynogenesisproved successful.
Androgenesis is a more valuable technique in theory,but produces little success in practice. Liu et al.(1987) used nucleo-transfer in loach and got purediploid mature fish from androgenesis. Yi et al.(1990) used gamma rays to irradiate ripe egg, matedwith normal sperm, and got diploid common carp-grass carp fish via androgenesis.
Sexual control
China started research on sex control in the 1970s.Several methods were used:• Interspecific crossbreeding - such as O. niloticus
(female) x O. aureus (male), resulting in about95% males.
• Sex reversal hormone treatment - sex reversedO. mossambicus , Nile t i lapia, red t i lapia,common carp, grouper, Clarias leather, cruciancarp and black porgy (Sparus macrocephalus)have been produced successfully by feeding thefry with estradiol or methyltestosterone treatedfeed. However, sex reversed fish are seldomused in commercial production in Chinabecause of food safety considerations.
• YY supermale technique - Yang et al. (1980)used estradiol benzoate to turn XY O. mossambicusinto XY female, mated it with XY fish, selectedYY super female from progenies and thenmated YY with normal XX, producing 100%male fish. These fish have significant growthsuperiority with 35% increase in yield.
• Gynogenesis and sex reversal technique -Wu et al. (1981) used methyltestosterone or
Aquaculture Genetics Research in China
Genome manipulations(chromosomal engineering)

Fish Genetics Research in Member Countries and Institutions of INGA20
testosterone propionate to change gynogeneticcommon carp into XX physiological female fish.This female was mated with XX male, resultingin gynogenetic inbred strain of carp. Using F1 ofgynogenetic Xingguo red carp as female parentand male gynogenetic mirror carp, an all-femalehybrid with higher production was produced.
Cell engineering
Nucleo-transfer
Fish nucleo-transfer technique was used first in Chinaby Tong Dizhou and his team (1963, 1973a, 1973b,1973c). They succeeded in transferring the nucleusof crucian carp vesicular germ cell into the nucleusof the removed egg of Rhodeus spp. This techniquecould bind the different fish’s nucleus and cytoplasm,creating a new body. Since 1970s, nucleo-transfer hasbeen applied in breeding common carp and cruciancarp, and grass carp and blunt snout bream (Tong1973a, 1973b; Yan et al. 1984; Yan and Wang 1985).Nuclear cytoplasm hybrids of common carp-cruciancarp, crucian carp-common carp, grass carp-bluntsnout bream, and blunt snout bream-grass carp, etc.,were produced. Of these, common carp-crucian carpnucleo-transfer fish reproduced three generations,and showed 20% faster growth than parental redcommon carp.
Cell fusion
Tong (1973a, 1973b) used Xiantai virus to inducethe cell fusion of crucian carp vesicular germ cellsand Ehrlich ascites cells. Yan et al. (1984) tested themain parameters of cell fusion by polyethylene glycol(PEG) in crucian carp. Liu et al. (1988) used electricalfusion method and got a fusion ratio of 47% and asurvival ratio of 88.6% in vesicular germ cell ofParamisgurnus dabryanus.
Yi et al. (1988), using P. dabryanus, M. anguillicaudatus,common carp and red crucian carp as materials forelectric fusion, produced six fusion fish of P. dabryanusand four fusion fish of common carp and red cruciancarp. Zhang et al. (1988) used laser to fuse the eggsof M. anguillicaudatus and P. dabryanus with a fusionrate of 53%.
Gene engineering
Tong et al. (1963) and Tong (1973a, 1973b) foundout that injecting mRNA from the ripe egg of crucian
carp, or DNA from liver and testis of crucian carpinto goldfish fertilized egg, could change thegoldfish’s genetic properties.
With the development of cloning and DNArecombinant technique, gene transfer research wasinitiated. Zhu et al. (1985, 1986) recombined humanGH gene and mouse MT gene promoter, and thenmicroinjected it into a fertilized egg of goldfish, andobserved heterogene integration. Subsequently,many institutes have undertaken studies on gene-transfer using mainly mammalian (human and cow)GH gene. Gene-transferred common carp, cruciancarp, blunt snout bream, Paramisgurnus sp. and troutwere produced. Some of them showed significantgrowth superiority. The gene-transfer fish model(Zhu et al. 1989) indicated that once the heterogeneexpresses, it could improve the performance of gene-transfer fish significantly. Heterogene could bepassed on by generative propagation and expressedin offspring. With further research, all-fish genetransfer was proposed, i.e., to produce gene-transferfish with all the gene components from fishthemselves. Zhu et al. (1990) successfully cloned theGH from common carp and grass carp and actin genefrom common carp, and got functional actinpromoter. Sun et al. (1993a, 1993b, 1995) separatedand cloned MT gene promoter from northerncommon carp gene library, and cloned GH gene fromsalmon gene library. Then they transferred the all-fish gene of common carp MT promoter and salmonGH gene into carp, producing a new common carpstrain with faster growth.
While microinjection is the major method used ingene transfer, some new methods such as sperm
Species Promoter Gene IG References
Carassius auratus mMT hGH + Zhu et al. 1985C. auratus RSV neo ? Yoon et al. 1989C. auratus ß-act CAT ? Liu et al. 1990Cyprinus carpio RSV rtGH + Zhang et al. 1990
RSV csGH + Zhang et al. 1990mMT hGH + Zhu et al. 1990
mMT hGH ? Xia et al. 1992
Table 7. Gene transfer experiments on cyprinids in China.
Megalobramaamblycephala
Notes:IG - increased growth ; mMT - mouse metallothionein promotor ;h - human ; GH - growth hormone ; RSV - rous sarcoma virus ;neo - neomycin resistance ß-act - ß-actin ; CAT - chloramphenicolacetyltreansferase ; rt - rainbow trout ; cs - coho salmon

21Aquaculture Genetics Research in China
vehicle, electrical pulse, injection method, in vitrocultivation, are also being explored. Gene-transferredfish has been investigated by many people (Table 7).
Conservation
Aquatic biodiversity in China is under stress due tohigh demand for fish and rapid development ofaquaculture industry. Hence, genetic conservation ofaquatic organisms is essential not only for the countrybut also for the world. Efforts are being made by thegovernment, fisheries scientists and communes, butthese are not enough.
The application of genetic principles to aqua-culture is far behind that of agriculture or animalhusbandry. There is no single management strategyfor conservation of genetic resources. It must beconsidered on species-by-species and case-by-case basis. For each species, appropriate in situ or exsitu conservation needs to be undertaken. Somespecies that need to be conserved in China are listedin Table 8.
In situ conservation
Protecting the genetic diversity at populationlevel involves ecosystem maintenance and speciesmanagement.
The Yangtze River basin accounts for 60% of the totalfreshwater fish production in China. It is not only
the cradle of Chinese fish culture, but also the primeworld site for genetic diversity of Chinese carps andthe major source of wild brooders for artificialbreeding.
The Swan Oxbow Lake (Fig. 3), formed in the 1970sand has an area of 1 200 ha, remains open to YangtzeRiver and has been selected for major Chinese carpconservation. The carps migrate between the OxbowLake and Yangtze River. It is expected that the SwanOxbow Lake can keep sufficient fish to supplybrooders to hatcheries. A 2 000 ha conservation areafor blunt snout bream was set up in Yuli Lake in 1990.A 300 ha natural spawning ground has beenrehabilitated, and a fish screen has been built to
Table 8. Priority species for aquatic genetic resourcesconservation in China.
Family Species
Cyprinidae Cyprinus carpio
Carasius auratusCtenopharyngodon idellaHypophthalmichthys molitrixAristichthys nobilisMegalobrama amblycephalaMyxocyprinus asiaticus
Cichlidae Oreochromis niloticusO. aureus
Acipenseridae Acipenser sinensisA. dabryanus
Clupeidae Macrura reevesiGrapsidae Eriocher sinensisTrionychidae Trionyx sinensis
Fig. 3. Swan Oxbow, an open conservation area for majorChinese carps in Yangtze River.
Fish Genetics Research in Member Countries and Institutions of INGA22
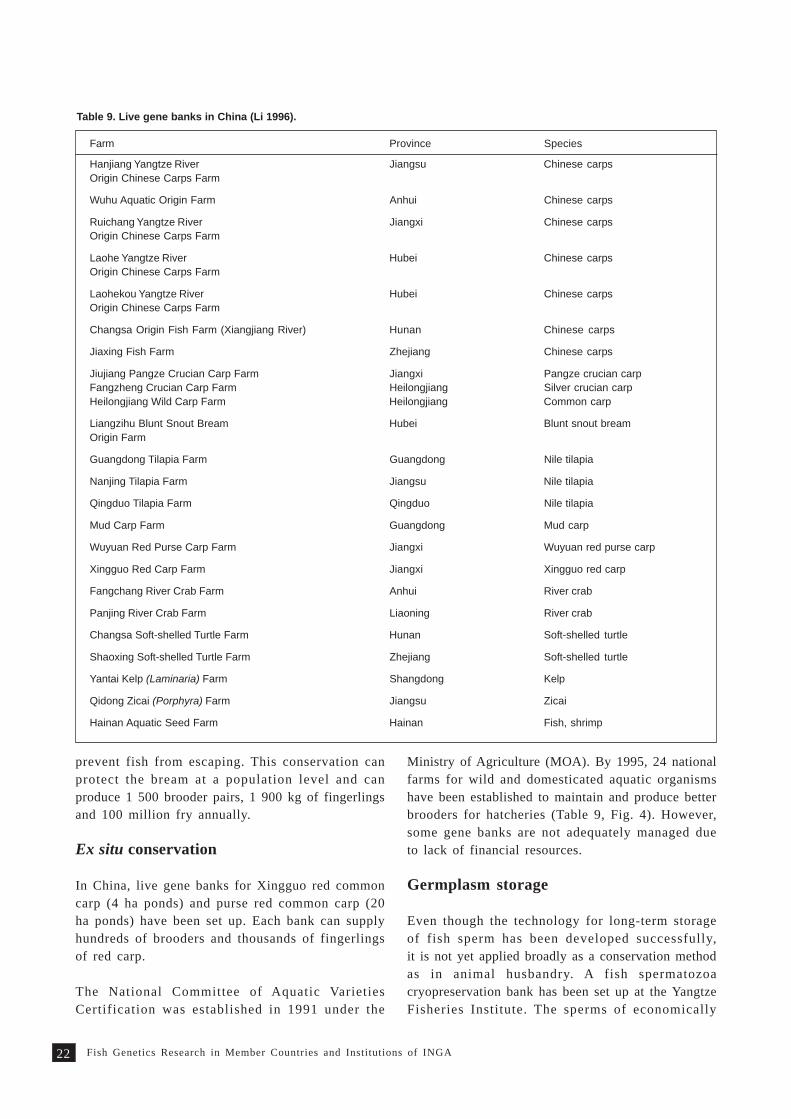
Table 9. Live gene banks in China (Li 1996).
Farm Province Species
Hanjiang Yangtze River Jiangsu Chinese carpsOrigin Chinese Carps Farm
Wuhu Aquatic Origin Farm Anhui Chinese carps
Ruichang Yangtze River Jiangxi Chinese carpsOrigin Chinese Carps Farm
Laohe Yangtze River Hubei Chinese carpsOrigin Chinese Carps Farm
Laohekou Yangtze River Hubei Chinese carpsOrigin Chinese Carps Farm
Changsa Origin Fish Farm (Xiangjiang River) Hunan Chinese carps
Jiaxing Fish Farm Zhejiang Chinese carps
Jiujiang Pangze Crucian Carp Farm Jiangxi Pangze crucian carpFangzheng Crucian Carp Farm Heilongjiang Silver crucian carpHeilongjiang Wild Carp Farm Heilongjiang Common carp
Liangzihu Blunt Snout Bream Hubei Blunt snout breamOrigin Farm
Guangdong Tilapia Farm Guangdong Nile tilapia
Nanjing Tilapia Farm Jiangsu Nile tilapia
Qingduo Tilapia Farm Qingduo Nile tilapia
Mud Carp Farm Guangdong Mud carp
Wuyuan Red Purse Carp Farm Jiangxi Wuyuan red purse carp
Xingguo Red Carp Farm Jiangxi Xingguo red carp
Fangchang River Crab Farm Anhui River crab
Panjing River Crab Farm Liaoning River crab
Changsa Soft-shelled Turtle Farm Hunan Soft-shelled turtle
Shaoxing Soft-shelled Turtle Farm Zhejiang Soft-shelled turtle
Yantai Kelp (Laminaria) Farm Shangdong Kelp
Qidong Zicai (Porphyra) Farm Jiangsu Zicai
Hainan Aquatic Seed Farm Hainan Fish, shrimp
prevent fish from escaping. This conservation canprotect the bream at a population level and canproduce 1 500 brooder pairs, 1 900 kg of fingerlingsand 100 million fry annually.
Ex situ conservation
In China, live gene banks for Xingguo red commoncarp (4 ha ponds) and purse red common carp (20ha ponds) have been set up. Each bank can supplyhundreds of brooders and thousands of fingerlingsof red carp.
The National Committee of Aquatic VarietiesCertification was established in 1991 under the
Ministry of Agriculture (MOA). By 1995, 24 nationalfarms for wild and domesticated aquatic organismshave been established to maintain and produce betterbrooders for hatcheries (Table 9, Fig. 4). However,some gene banks are not adequately managed dueto lack of financial resources.
Germplasm storage
Even though the technology for long-term storageof fish sperm has been developed successfully,it is not yet applied broadly as a conservation methodas in animal husbandry. A fish spermatozoacryopreservation bank has been set up at the YangtzeFisheries Institute. The sperms of economically

23
important species, such as grass carp, black carp,silver carp, bighead carp, blunt snout bream,Xingguo red common carp, mirror common carp andsilver crucian carp are stored there.
Current Major Research
Fisheries research in China is organized by theMinistry of Science and Technology, NationalFoundation for Natural Science and MOA. Currently,research is conducted on the development of fast-growing strains of carps, pure strains of silver carp,bighead carp and grass carp, and conservation ofgenetic resources.
References
ARI (Agriculture Research Institute), Wuhan County, JiangxiProvince. 1973. Report on the selection of red purse carp.Freshwat. Fish. Sci.10: 3-6. (In Chinese).
Gui, J.F., J.M. Sun, S.C. Liang, W.Y. Huang and Y.G. Jiang. 1991.Studies on genome manipulation in fish. II. Tetraploidyinduced by hydrostatic pressure treatment and acombination of hydrostatic pressure and cold treatments intransparent colored crucian carp. Acta Hydrobiol. Sin. 15(4):333-342. (In Chinese with English abstract).
Gui, J.F., S.C. Liang, J.M. Sun, W.Y. Huang and Y.G. Jiang. 1990.Studies on genome manipulation in fish. I. Induction oftriploid transparent colored crucian carp (Carassius auratustransparent colored variety) by hydrostatic pressure. ActaHydrobiol. Sin. 14(4): 336-344. (In Chinese with Englishabstract).
JFS (Jiangxi Fisheries Society). 1982. Selection of red purse carpin Wuyuan. Freshwat. Fish. (1): 29-31. (In Chinese).
Jiang, Y.G., H.C. Yu, B.D. Chen, S.C. Liang, D.L. Yang and S.A.Ling. 1982. Artificial and natural gynogenesis in cruciancarp. Acta Hydrobiol. Sin.7(4): 471-487. (In Chinese withEnglish abstract).
Li, S.F. 1995. Germplasm resources and conservation offreshwater fishes in China. China Agriculture Press. 186 p.(In Chinese with English preface).
Li, S.F., editor. 1998. Genetic characterization of major freshwaterculture fishes in China. Shanghai Scientific and TechnologyPress, Shanghai. (In Chinese with English preface).
Li, S.F., L.Z. Wu, J. Wang, Q.R. Chou and Y.L. Chen. 1990.Comprehensive genetic study on Chinese carps. ShanghaiScientific and Technical Publishers, Shanghai. 228 p. (InChinese with English abstract).
Liu, H.Q., Y.L. Yi and H.Q. Chen. 1987. The production ofdiploid androgenetic loach. Acta Hydrobiol. Sin. 13(3): 241-246. (In Chinese with English abstract).
Liu, P.L., Y.L. Yi, H.Q. Liu and H.Q. Chen. 1988. Preliminarystudy on electric fusion of fish cells. Acta Hydrobiol. Sin.12(1): 94-95. (In Chinese with English abstract).
Liu, Z., Z. Zhu, K. Roberg, A.J. Faras, K.S. Guise, A.R. Kapuscinskiand P.B. Hackett. 1990. Isolation and characterization ofthe beta-actin gene of carp (Cyprinus carpio). DNA Seq. J.DNA Seq. Map.1: 125-136.
Aquaculture Genetics Research in China
Fig. 4. Locations in China of live fish gene banks.
LEGENDsilver carp, bighead, grass carpand black carpblunt snout breamcommon carpcrucian carptilapiariver crabsoft-shelled turtlekelp (Laminaria)and Porphyra.

Fish Genetics Research in Member Countries and Institutions of INGA24
Lu, G.Q., S.F. Li and L. Bernatchez.1997. Mitochondrial DNAdiversity, population structure, and conservation geneticsof four native carps within the Yangtze River, China. Can.J. Fish. Aquat. Sci. 54(1): 47-58.
Sun, X.W., J.B. Shen, X.C. Yan, L.Q. Liang and P. Wang. 1995.The research of engineering common carp of transferringbovine and orine growth hormone gene. J. Fish. Sci. China2(2): 23-33. (In Chinese with English abstract).
Sun, X.W., P. Wang, L.Q. Liang, X.C. Yan and J.B. Shen. 1993a.Construction of all gene transfer fish. Bull. High Tech. 3(9):23-26. (In Chinese with English abstract).
Sun, X.W., X.C. Yan, L.Q. Liang, D.C. Cao and L.M. Zhang.1993b. Production of gene transfer fish by directedinjection. Biotechniques 3(3): 12-14. (In Chinese withEnglish abstract).
Tong, D.Z. and M.J. Niu. 1973c. Character variations of goldfishinduced by nucleic acid. Scient. Sin. (4): 389-394. (InChinese with English abstract).
Tong, D.Z., C.C. Wu, Y.F. Yi, F.M. Tong and M. Du. 1963. Nucleustransplantation of fishes. Sci. Bull. (7): 61-62. (In Chinesewith English abstract).
Tong, D.Z., Y.F. Yi, D.Y. Lu, F.M. Tong and M. Du. 1973. Nucleustransplantation between two fishes of different subfamilies.Acta Zool. Sin. 19(3): 201-209. (In Chinese with Englishabstract).
Wanan Fish Farm and Jiangxi University. 1984. Breeding studyon the Wanan glass red carp. Wanan Fish Farm and JiangxiUniversity. (In Chinese).
Wu, W.X., C.W. Li, G.A. Lie, D.Y. Xu, S.C. Liu, Y. Xai and C.X.Dan. 1988. Studies on tetraploid hybrid between redcommon carp and grass carp and its backcross triploid. ActaHydrobiol. Sin. 12(4): 355-356. (In Chinese with Englishabstract).
Wu, W.X., L.L. Lin and D.Y. Xu. 1981. A tetraploidy hybridcrossing red carp Cyprinus carpio L. with grass carpCtenopharyngoden idella Cuv. et val. Acta Hydrobiol. Sin. 7(3):433-436. (In Chinese with English abstract).
Wu, T.T., H. Yang, Z. J. Dong. D.Q. Xia, Y.X.Shi, X.D. Ji, Y. Shenand W. X. Sun. 1994. The integration and expression ofhuman growth hormone gene in blunt snout bream andcommon carp. J. Fish. China/Shuichan Xuebao 18(4):284-289. (In Chinese)
Yan, C.Y. and Y. Wang. 1985. Nucleus transplantation in teleosts.J. Bioeng. (4): 1-11. (In Chinese with English abstract).
Yan, C.Y., D.Y. Lu, M. Du, G.S. Li, L.T. Lin, G.Q. Jin, H. Wang,Y.Q. Yang, D.Q. Xia, A.Z. Liu, Z.Y. Zhu, Y.N. Yi, M.Y. Chen,X.Q. Yang and H.X. Chen. 1984. Nucleus transplantationin teleosts. Sci. Sin. (edition B) (8): 729-731. (In Chinesewith English abstract).
Yan, K., M.Y. Chen and H.X. Chen. 1984. Study on the cell fusionof fish. Heredity 6(6): 19-21. (In Chinese).
Yang, Y.Z., Z. Zhang, K. Lin, Y. Wei, E. Huang, Z. Xu, S. Ke andJ. Wei. 1980. Use of three line combinations for productionof genetic all-male Tilapia mossambica. Acta Genet. Sin. 7(3):240-246. (In Chinese with English abstract).
Yi, Y., C. Wu and R. Chen. 1990. Study on gamma ray inducedandrogenesis in fishes. Acta Hydrobiol. Sin. 14(1): 91-92.(In Chinese with English abstract).
Yi, Y.L., P.L. Liu, H.Q. Liu and H.X. Chen. 1988. Electric fusionbetween blastula cells and unfertilized eggs in fish. ActaHydrobiol. Sin. 12(2): 189-192. (In Chinese with Englishabstract).
Yoon, S.J., Z. Liu, A.R. Kapuscinski, P.B. Hackett, A.J. Faras andK.S. Guise. 1989. Successful gene transfer in fish, p. 29-34.In A.L. Beadeaut, R. Mulligan and I.M. Verma (eds.) Genetransfer and therapy. Alan R. Liss, Inc., New York.
Zhang, P., M. Hayat, C. Joyce, L.I. Gonzales-Villasenor, R.A. Lin,R.A. Dunham, T.T. Chen and D.A. Powers. 1990. Genetransfer, expression and inheritance of pRSV-rainbow trout-GH cDNA in the common carp, Cyprinus carpio (Linnaeus).Mol, Reprod. Dev. 25:3-13.
Zhang, W., B. Zhao, Q. Wang, G. Ge, Z. Li, J. Zou. 1988. Laserinducing cell fusion of fertilized loach eggs: a study of newmethod of cell fusion. Chin. J. Biotechnol. 4(4):298-303.
Zhang, Z.S. and X.Y. Sun. 1994. A new technique of the combinedbreeding developed in Jian carp (Cyprinus carpio var. Jian),p. 22-26. In J.S. Zhang and X.Y. Sun (eds.) The selectedpaper of breeding in Jian carp (Cyprinus carpio var. Jian).Science Press, Beijing.
Zhu, Z.Y, G. Li, L. He and S. Chen. 1985. Novel gene transferinto fertilized eggs of gold fish (Carassius auratus L. 1758).Z. Angew. Ichthyol. 1: 31-34.
Zhu, Z.Y., L. He, Y.F. Xie and G.H. Li. 1990. Construction ofgenome library of carp (Cyprinus carpio) and grass carp(Ctenopharyngodon idellus) and isolation of their growthhormone gene and actin gene. Acta Hydrobiol. Sin. 14(2):176-178. (In Chinese with English abstract).
Zhu, Z.Y, K.F. Xu, Y.F. Shi, G. Li and L. He. 1989. Modelestablishment of transgenic fish. Sci. Sin. 2: 147-155. (InChinese with English abstract).
Zhu, Z.Y., K.S. Xu, G.H. Li, Y.F. Shi and L. He. 1986. Biologicaleffect of human growth hormone in the loach fertilized eggsafter microinjection. Sci. Bull. 31(5): 387-389. (In Chinesewith English abstract).

25Fish Genetics Research in Côte d’Ivoire
ABSTRACT
Fish genetics research conducted in two research centers and other institutions in Côte d’Ivoire is presented.Most of this research so far conducted focused on population genetics. In addition to electrophoresischaracterization, morphometric and meristic characteristics were used to evaluate and describe fish populationin Côte d’Ivoire. Recent genetic research tools, such as microsatellites, restriction fragment length polymorphismof mitochondrial DNA techniques have been used. Research activities dealt mainly with tilapias (Sarotherodonmelanotheron, Oreochromis niloticus and O. aureus) and Siluriformes (Clarias gariepinus, C. anguillaris,Heterobranchus longifilis, H. bidorsalis and Chrysichthys nigrodigitatus) which were mostly used for their potentialfor aquaculture. One of the research centers, the Fish Station, attached to the Centre National de RechercheAgronomique-Bouaké regional office, is participating in the International Network on Genetics in AquacultureProject.
FISHGENETICS RESEARCH
IN CÔTE D’IVOIRE
V. Yapi-GnaoréCentre National de Recherche Agronomique
01 BP 633 Bouaké 01, Côte d’Ivoire
YAPI-GNAORE, V. 2001. Fish genetics research in Côte d’Ivoire, p. 25-28. In M.V. Gupta and B.O. Acosta (eds.) Fish geneticsresearch in member countries and institutions of the International Network on Genetics in Aquaculture. ICLARMConf. Proc. 64, 179 p.
Introduction
Dependence on catch from the wild as source ofanimal protein is a tradition for majority of thepopula t ion in Côte d’ Ivoi re . Consequent ly,overexploitation of fish population, degradationand des t ruc t ion of f i sh habi ta t s a re majorconcerns for planners. The need for aquaculturedevelopment in the country has become morecrucial especially because 60% of the national fishconsumption is covered by importation.
For many years , the Ivor ian Government hasgiven mandate to two specia l ized centers toconduct fisheries and aquaculture research: theCentre de Recherche Océanologique (CRO) andthe Fish Sta t ion of the newly crea ted Cent re
National de Recherche Agronomique (CNRA),former ly known as the Ins t i tu t des Savannes(IDESSA) Fish Station. This paper gives a briefoverview of the genetics research undertaken anda description of the CNRA structure.
CNRA was created in April 1998 by merging threeformer research centers: IDESSA, Inst i tut desForê ts ( IDEFOR) and Cent re Ivoi r ien deRecherche Technologique (CIRT) . The newinstitution, with headquarters in Abidjan, receivedmandate to continue activities undertaken by thethree centers. It is mainly funded by the privatesector. In order to be ful ly operat ional and tocover smallholders’ concerns in its activities, CNRApartitioned the country into five regions, i.e.,Korhogo (northern), Bouaké (central), Gagnoa (west-

Fish Genetics Research in Member Countries and Institutions of INGA26
central), Man (western) and Abidjan (southern). Oneor more stations are located in each region. The FishStation is attached to CNRA Bouaké regional officeand is participating in the International Network onGenetics in Aquaculture (INGA) Project.
Genetic Research Tools
Most of the genetics research so far conducted inCôte d’Ivoire focused on population genetics. Themain objectives were to characterize wild andcultured stocks of fish in terms of genetic variabilitybetween and within populations and differentiationin natural populations; make biogeographicalinferences; and identify populations with specialpotential and of interest to fish farmers. Theseresearch activities are important because they canlead to the development of strains for aquacultureor the protection of fish populations which are atrisk of extinction.
Genetic tools developed in advanced countriesare adapted, or new ones are established for researchconducted in Côte d’Ivoire. Species studied includenatural populations and their cultured strains fromCôte d’Ivoire and other regions in Africa. Enzymaticprotein electrophoresis, microsatellites andrestriction fragment length polymorphism (RFLP)of mitochondrial DNA (mtDNA) techniques havebeen used. In a very recent collaborative projectfunded by the European Community, 57 fishpopulations collected from 23 African countries wereevaluated using these three tools. The project called“GENETICS” (GENETic Investigations on Cichlidsand Siluriformes) dealt mainly with tilapias(Sarotherodon melanotheron, Oreochromis niloticus andO. aureus) and Siluriformes (Clarias gariepinus,C. anguillaris, Heterobranchus longifilis, H. bidorsalis andChrysichthys nigrodigitatus) which have potential andmostly used for aquaculture in Africa (Adépo-Gourène et al. 1997; Agnèse et al. 1997, 1998; Anon.1997; and Vreven et al. 1998). The genetic evaluationwas extended to some fisheries species - Ethmalosafimbriata, Sardinella aurita, S. maderensis, Carcharinusspp., Marcusenius ussheri and M. furcidens (Gouréne etal. 1993; Adépo-Gouréne, pers. comm.). In additionto these molecular genetic tools, morphometric andmeristic analysis and comparison of zootechnicalperformances were used.
Electrophoresis characterization
Electrophoresis has been used in Bouaké Fish Stationsince 1989 on two species, O. niloticus (Bouaké and
Burkina Faso strains) and O. aureus (Israel and Egyptstrains). The main objective is to define the geneticidentity of the domestic stocks and developmanagement strategies for these (Rognon 1993).CRO has also recently undertaken electrophoresis,through the GENETICS project. Agnèse (1989 and1991) and Agnèse et al. (1989) have publishedelectrophoresis evaluations on C. nigrodigitatus. Forexample, 16 populations of O. niloticus collected in17 locations were clustered in three major groups,the West African populations being in the samecluster (Agnèse et al. 1998).
Microsatellites and RFLP of mtDNA
Fish genetics research using microsatellites and RFLPof mtDNA, as part of the GENETICS Project (Anon.1997), was conducted at CRO in collaboration withthe Institut Francais de Recherche Scientifique pourle Developpement en Cooperation, Université deMontpellier, Institut National de la RechercheAgronomique, Musée Royal de l’Afrique Centrale,Institute of Marine Biology-Greece and UniversitéChatholique de Louvain. The above aquaculturespecies were evaluated and results from the analysesavailable, e.g., Agnèse et al. (1998), found that mtDNAof West African O. niloticus cannot be distinguishedfrom that of O. aureus.
Morphometric and meristic analyses
Recent analyses include those of Adépo-Gourèneet al. (1997) on C. nigrodigitatus and C. maurus andof Vreven et al. (1998) on O. niloticus. Similar researchis being conducted at both university campuses inAbidjan (Abobo-Adjamé and Cocody). Taxonomicand biogeographical implications were derived fromthe results of the various morphometric and meristicanalyses (Gourène and Teugels 1998; Teugels andGourène 1998). Production performances (growth,survival rate, fecundity, feed conversion ratio etc.),adaptability to capture breeding or localenvironments and production systems werecompared or evaluated among cultured strains(Otémé 1993; Da Costa and Assémien 1998). Thesestudies provide complementary information tothe molecular analysis.
Comparison of the growth performance of O. niloticusfrom four African countries, Côte d’Ivoire (BKE),Ghana (GHA), Mali (NIG) and Senegal (SEN) wasundertaken (Tables 1 and 2). Weights of thefingerlings, initially varying between 60 and 75 g,reached 330-460 g in 150 days of rearing. The Mali
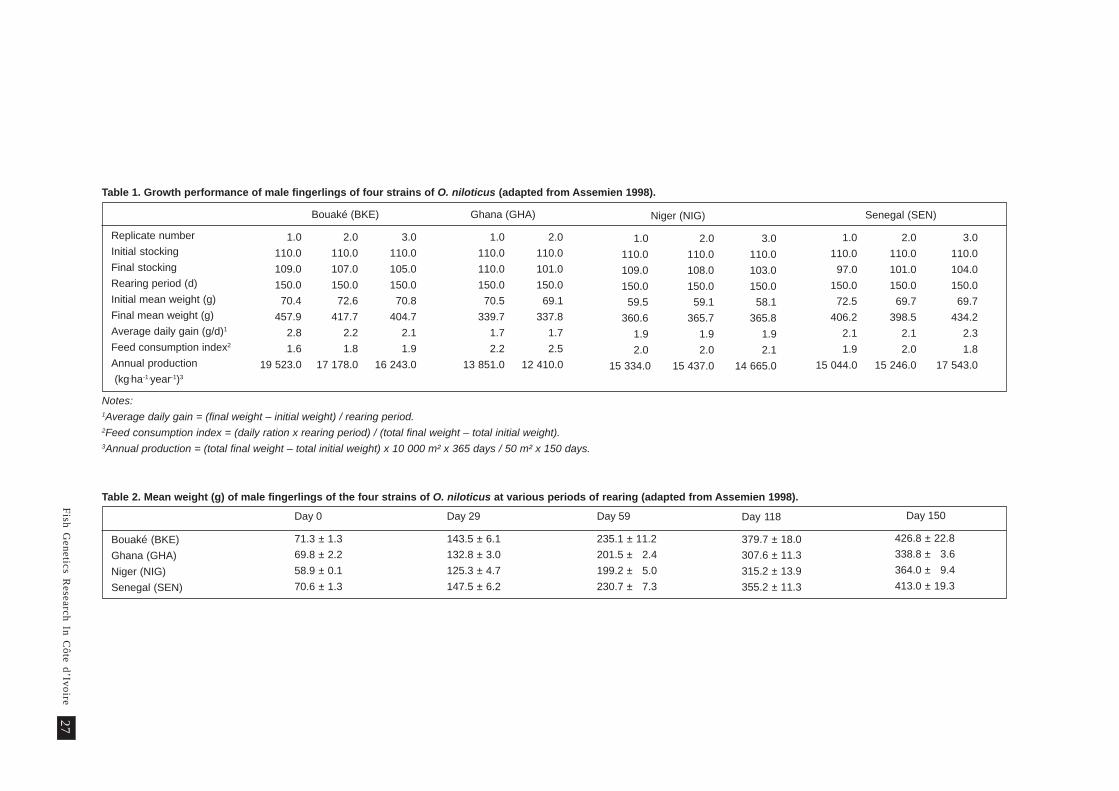
27Fish G
enetics Research In C
ôte d’Ivoire
Table 1. Growth performance of male fingerlings of four strains of O. niloticus (adapted from Assemien 1998).
Replicate numberInitial stockingFinal stockingRearing period (d)Initial mean weight (g)Final mean weight (g)Average daily gain (g/d)1
Feed consumption index2
Annual production (kg.ha-1.year-1)3
Bouaké (BKE)
1.0 2.0 3.0110.0 110.0 110.0109.0 107.0 105.0150.0 150.0 150.070.4 72.6 70.8
457.9 417.7 404.72.8 2.2 2.11.6 1.8 1.9
19 523.0 17 178.0 16 243.0
Ghana (GHA)
1.0 2.0110.0 110.0110.0 101.0150.0 150.070.5 69.1
339.7 337.81.7 1.72.2 2.5
13 851.0 12 410.0
Niger (NIG)
1.0 2.0 3.0110.0 110.0 110.0109.0 108.0 103.0150.0 150.0 150.059.5 59.1 58.1
360.6 365.7 365.81.9 1.9 1.92.0 2.0 2.1
15 334.0 15 437.0 14 665.0
Senegal (SEN)
1.0 2.0 3.0110.0 110.0 110.0 97.0 101.0 104.0150.0 150.0 150.072.5 69.7 69.7
406.2 398.5 434.22.1 2.1 2.3
1.9 2.0 1.815 044.0 15 246.0 17 543.0
Notes:1Average daily gain = (final weight – initial weight) / rearing period.2Feed consumption index = (daily ration x rearing period) / (total final weight – total initial weight).3Annual production = (total final weight – total initial weight) x 10 000 m² x 365 days / 50 m² x 150 days.
Table 2. Mean weight (g) of male fingerlings of the four strains of O. niloticus at various periods of rearing (adapted from Assemien 1998).
Bouaké (BKE)Ghana (GHA)Niger (NIG)Senegal (SEN)
Day 0
71.3 ± 1.369.8 ± 2.258.9 ± 0.170.6 ± 1.3
Day 29
143.5 ± 6.1132.8 ± 3.0125.3 ± 4.7147.5 ± 6.2
Day 59
235.1 ± 11.2201.5 ± 2.4199.2 ± 5.0230.7 ± 7.3
Day 118
379.7 ± 18.0307.6 ± 11.3315.2 ± 13.9355.2 ± 11.3
Day 150
426.8 ± 22.8338.8 ± 3.6364.0 ± 9.4413.0 ± 19.3

Fish Genetics Research in Member Countries and Institutions of INGA28
selective breeding program for tilapia; test this on-station and on-farm; and fully implement it in CNRAproduction strategies. In the long-term, thedomestication and breeding in captive conditions ofinland water fish, such as Lates niloticus, Labeo coubie,Parachana obscura and Distichodus rostratus, are planned.
References
Adépo-Gourène, B., G.G. Teugels, L.M. Risch, M.M. Hanssens andJ.-F. Agnèse. 1997. Morphometrical and genetic differentiationof 11 populations of African catfish Chrysichthys nigrodigitatus(Siluroidei, Claroteidae), with consideration of theirbiogeography. Can. J. Zool. 75:102-109.
Agnèse, J.-F. 1989. Différenciation génétique de plusieurs espècesde Siluriformes Ouest Africain ayant un intérêt pourl’aquaculture. University of Montpellier, France. Ph. D. thesis.
Agnèse, J.-F. 1991. Taxonomic status and genetic differentiationamong West African Chrysichthys auratus complex (Pisces,Siluriformes), based on protein electrophoresis. Aquacult. Fish.Manage. 22: 229-237.
Agnèse, J.-F., B. Adépo-Gourène and L. Pouyaud. 1998. Geneticdifferentiation among natural populations of the Nile tilapiaOreochromis niloticus (Teleostei, Cichlidae), p. 153-163. In J.-F.Agnèse (ed.) Genetics and aquaculture in Africa. ORSTOM,France.
Agnèse, J.-F., N. Pasteur and C. Lévêque. 1989. Différenciationgénétique de quelques populations de Chrysichthys nigrodigitatuset de C. johnelsi (Pisces, Bagridae) de Côte d’Ivoire et du Mali.Rev. Hydrobiol. Trop. 22: 101-106.
Agnèse, J.-F., B. Adépo-Gourène, E.K. Abban and Y. Fermon. 1997.Genetic differentiation among natural populations of Niletilapia Oreochromis niloticus. Heredity 79: 88-96.
Anon. 1997. Mise en évidence et valorisation de la diversité despopulations naturelles des espèces de poissons utilisées enpisciculture Ouest Africaine. Rapp. Final du ProgrammeGENETICS. Contrat C.E.E. No. ERBTS3*CT920079.
Assemien, O.S.A. 1998. Contribution à l’élevage d’un tilapiad’intérêt éconmique Oreochromis niloticus (Linnaeus, 1758):caractérisation zootechnique de quatre souches. Memoire pourle Diplôme d’Etudes Approfondies d’Ecologie Tropicale.Université de Cocody, Abidjan, Côte d’Ivoire.
Da Costa, K.S. and O.S.A. Assemien. 1998. Caractérisationzootechnique (croissance et fécondité) de différentes souchesde Clarias anguillaris et Oreochromis niloticus. Rapp. FinalProgramme GENETICS. Convention STD-3 ORSTOM/IDESSA. IDESSA, Bouaké, Côte d’Ivoire.
Gourène, A.B., L. Pouyaud and J.-F. Agnèse. 1993. Importance decertaines caractéristiques biologiques dans lan structurationgénétique des espèces de poissons: le cas de Etmalosa fimbriataet Sarotherodon melanotheron. J. Ivoir. Océanol. Limnol. 2: 55-69.
Otémé, Z.J. 1993. Reproduction et fécondité du mâchoironChrysichthys nigrodigitatus en élevage. J. Ivoir. Océanol. Limnol.2:53-59.
Rognon, X. 1993. Diversité génétique et relations phylogénétiqueschez les tilapias (Pisces: Cichlidae): comparaison des donnéesdu polymorphisme enzymatique et mitochondrial. UniversitéParis-Sud Orsay, France. Thèse de doctorat en science.
Vreven, E.J., B. Adépo-Gourène, J.-F. Agnèse and G.G. Teugels.1998. Morphometric and allozyme variation in naturalpopulations and cultured strains of Nile tilapia Oreochromisniloticus (Teleostei, Cichlidae), p. 175-182. In J.-F. Agnèse (ed.)Genetics and aquaculture in Africa. ORSTOM, France.
strain, which had the lowest initial weight, had a finalweight higher than that of the Ghana strain.
Bouake strain showed the highest weight at variousperiods. The Bouake and Senegal strains appearedto have similar growth performance compared to Maliand Ghana strains. Although the Bouake and Ghanastrains are most likely to be related genetically, theydiffer in growth performance.
Through INGA, the project, Genetic Characterizationand Improvement of Native Tilapia for Culture,is being undertaken by the CNRA Fish Station inBouake and its local partners. Preparations areunderway for the collection of broodstocks ofO. niloticus from areas where these were previouslyintroduced such as the Niger Basin in north-west part,Volta Basin in north-east region, dams or smallwater bodies of the northern region, Buyo Lake andKossou Lake. The strains collected and O. niloticusBouake strain will be conditioned and bred. Theprogenies will be used for genetic characterization andstrain evaluation experiments prior to initiation ofselective breeding work.
Fish Genetics Research in the Future
Despite tremendous efforts devoted to geneticsresearch, the present fish production level is far frombeing optimal. Selective breeding program aimed atrapid genetic gain on economically importantproduction characters is lacking. One of the majorconstraints is the absence of qualified personnel inquantitative genetics in various research teams.
Plans for the future include training in quantitativegenetics and improvement of infrastructures at theFish Station. A quantitative geneticist will soon berecruited. The research team will be reinforced withthe increase of senior fish scientists to seven by theyear 2001.
The CNRA Fish Station intends to contribute, throughpartnerships with other institutions and competencein the country and elsewhere, to the characterization,evaluation, conservation and utilization of fish geneticresources. Plans are made for CNRA fish scientists toactively participate in national, regional andinternational fora where these topics are discussed.A medium-term goal is to develop, through INGA, a
Research on Genetic Improvementof Local Tilapia Populations

29
ABSTRACT
Fish production in Egypt has significantly increased over the last few years reaching 546 000 t in 1998 comparedto 306 000 t in 1988. The contribution of aquaculture in the overall fish production has increased as well reachingits utmost share in 1998 (26%). Research on aquaculture topics was conducted in several institutes targeting theincrease of productivity as well as working on problems facing this promising sector. However, research on fishgenetics in Egypt only started in recent years. The genetic improvement of tilapias and African catfish representsthe focus of most genetics research in Egypt. This paper presents the status of genetics research within theInternational Network on Genetics in Aquaculture and in other national institutes in Egypt.
AQUACULTUREGENETICS RESEARCH
IN EGYPT
H. ElghobashyCentral Laboratory for Aquaculture Research
Agricultural Research CenterAbbassa, Abou Hammad, Sharkia, Egypt
ELGHOBASHY, H. 2001. Aquaculture genetics research in Egypt, p. 29-34. In M.V. Gupta and B.O. Acosta (eds.) Fishgenetics research in member countries and institutions of the International Network on Genetics in Aquaculture.ICLARM Conf. Proc. 64, 179 p.
Introduction
Aquaculture in Egypt has become an increasinglyimportant activity, as an immediate source of animalprotein required for the country’s growing population.The total fish production in 1998 was estimated at546 000 t, of which 26% is from aquaculture.Aquaculture is being undertaken at extensive andsemi-intensive levels. Most fish farms practicepolyculture where tilapia represents about 38% of thetotal production. Along with tilapia (Oreochromisniloticus and O. aureus), mullets and carps are alsostocked. Fish farms in Egypt include seven governmentfarms (total area of 4 000 ha) and private fish farms(total area of about 50 000 ha) that are located aroundthe lakes. All fish farms rely to some extent onhatcheries for obtaining their seed requirements. Mostfish hatcheries belong to the General Authority of Fish
Aquaculture Genetics Research in Egypt
Resources Development. The main fish seed producedare common carp, Chinese carps and tilapia. However,new tilapia hatcheries are now emerging. The totalproduction of tilapia fry from hatcheries or fish farmsin 1998 was estimated at about 49.9 million fry. O.niloticus and O. aureus dominate the fry production.
Most consumers in Egypt prefer tilapia comparedto other freshwater fish. There are four tilapia speciesin Egypt: Nile tilapia (O. niloticus), blue tilapia (O.aureus), white tilapia (Sarotherodon galilaeus) and greentilapia (Tilapia zillii).
Fish Genetics Research
Fish genetics research in Egypt has a relatively shorthistory concentrating initially on the biochemicalidentification and strain evaluation. The institutes/
Fish Genetics Research in Member Countries and Institutions of INGA30
universities working on fish genetics in aquaculturein Egypt are the: Central Laboratory for AquacultureResearch (CLAR), Abbassa; International Center forLiving Aquatic Resources Management RegionalResearch Center for Africa and West Asia, Abbassa;National Institute of Oceanography and Fisheries;Veterinary College-Idfena, Alexandria University;Genetics Department, Faculty of Agriculture, AinShams University; Genetics Department, Faculty ofAgriculture, Assuit University; Zoology Department,Faculty of Science, Zagazig University; ZoologyDepartment, Faculty of Girls, Ain Shams University;and Genetic Engineering Research Institute, SadatCity, Monofaya University.
Performance evaluationand genetic characterization
CLAR is leading collaborative research along withtwo schools in Ain Shams University, i.e., Zoologyand Genetics. The objectives of the research are to:• determine whether Nile tilapia originated from
a single strain or several strains;• compare the reproductive performance among
different stocks of Nile tilapia collected fromthree geographical locations: Maryout (MR) -Alexandria at North of Egypt (which has lowertemperature); Zawia (ZW) - Kafer El Sheikh atthe Delta area (which has saline soil); andAbbassa (AB) middle of delta area as a referencestrain;
• compare the growth performances of the threestocks; and
• carry out a biochemical genetic analysis todetermine the genetic variations within andamong stocks collected from different locations.
This research was undertaken since strain differences,inbreeding and crossings could affect reproductiveperformance in cultured fishes. Large straindifferences influenced reproductive performance inchannel catfish (Dunham and Smitherman 1984) andrainbow trout (Kincaid 1976). If similar geneticdifferences exist for reproductive performance intilapia, selection of a species or strain of tilapia withlower fecundity might be desirable for minimizingreproduction in rearing ponds. In contrast, strainswith higher fecundity may be desirable for enhancingreproduction in hatcheries.
Abbassa stocks with an average weight of 150 g werecollected from the production ponds. Maryout stocks
were collected as fry (1 g) and then nursed to largersize in earthen ponds. Zawia stocks (150 g) werebrought to Abbassa and maintained in ponds untilused for spawning. All fish groups were acclimatizedto Abbassa conditions upon their arrival.
Spawning activities
The spawning activities of the three strains andtheir crosses were carried out in circular earthen pondsof 100 m2 each during 1996 and 1997 (Table 1).Therewere differences in the total number of fry producedfrom strains evaluated. During 1996 and 1997 studies,Abbassa strain was highly reproductive, while Maryoutand Zawia strains had lower reproduction (Tables 2and 3). There were differences in the total number offry produced from each crossing. The cross betweenAbbassa x Zawia was highly reproductive, whileMaryout x Abbassa came intermediate and Zawia xMaryout was the lowest (Table 4). Crossing withAbbassa strain indicated its superiority with regard tofry production.
Growth performance
To evaluate the growth performance of three stocksand their hybrids, the fish were stocked in 0.1 haearthen ponds at a rate of one fish per square meter.Supplementary feed (25% protein) was given at arate of 10% of body weight, six days a week for 45days and later decreased to 5% of body weight.Biweekly samplings were carried out when randomsamples were individually measured for total lengthand weight. At the end of the growing season,harvesting was done and individual lengths andweights were estimated. The total yield for each pondwas determined.
The growth rate is certainly a key element regardlessof the production system. Studies undertaken during
Table 1. Details of spawning to evaluate performance of threestrains of O. niloticus, 1996-1997.
Spawning
PeriodSex ratioNo. of females/pondsNo. of replicationHarvest intervalsFish stocksCrossesNo. of harvestTemperature range (oC)
1996 season
6 Jun – 30 Sep1.5 : 16623 weeks3 stocks3 crosses721-25
1997 season
20 Jul – 15 Sep1.5 : 16623 weeks3 stocks3 crosses222-25
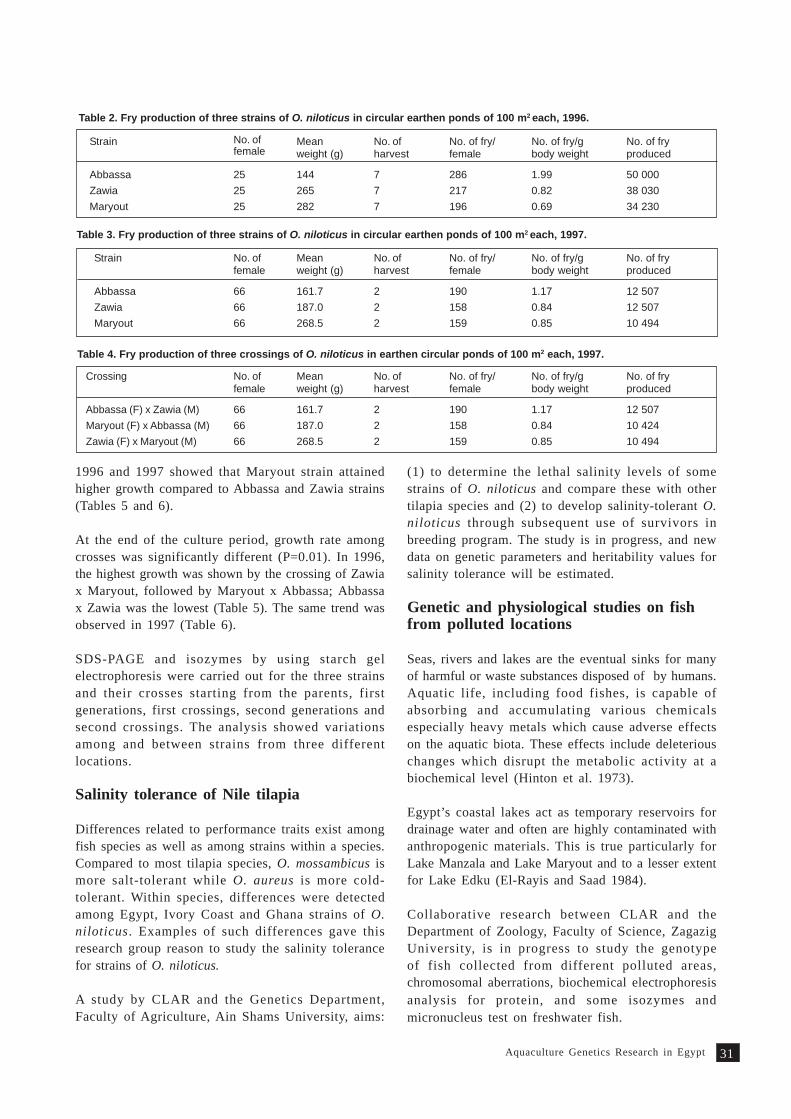
31
1996 and 1997 showed that Maryout strain attainedhigher growth compared to Abbassa and Zawia strains(Tables 5 and 6).
At the end of the culture period, growth rate amongcrosses was significantly different (P=0.01). In 1996,the highest growth was shown by the crossing of Zawiax Maryout, followed by Maryout x Abbassa; Abbassax Zawia was the lowest (Table 5). The same trend wasobserved in 1997 (Table 6).
SDS-PAGE and isozymes by using starch gelelectrophoresis were carried out for the three strainsand their crosses starting from the parents, firstgenerations, first crossings, second generations andsecond crossings. The analysis showed variationsamong and between strains from three differentlocations.
Salinity tolerance of Nile tilapia
Differences related to performance traits exist amongfish species as well as among strains within a species.Compared to most tilapia species, O. mossambicus ismore salt-tolerant while O. aureus is more cold-tolerant. Within species, differences were detectedamong Egypt, Ivory Coast and Ghana strains of O.niloticus. Examples of such differences gave thisresearch group reason to study the salinity tolerancefor strains of O. niloticus.
A study by CLAR and the Genetics Department,Faculty of Agriculture, Ain Shams University, aims:
Aquaculture Genetics Research in Egypt
Table 4. Fry production of three crossings of O. niloticus in earthen circular ponds of 100 m2 each, 1997.
Meanweight (g)
161.7187.0268.5
Crossing
Abbassa (F) x Zawia (M)Maryout (F) x Abbassa (M)Zawia (F) x Maryout (M)
No. offemale
666666
No. ofharvest
222
No. of fry/female
190158159
No. of fry/gbody weight
1.170.840.85
No. of fryproduced
12 50710 42410 494
Table 3. Fry production of three strains of O. niloticus in circular earthen ponds of 100 m2 each, 1997.
Strain
AbbassaZawiaMaryout
No. offemale
666666
Meanweight (g)
161.7187.0268.5
No. ofharvest
222
No. of fry/female
190158159
No. of fry/gbody weight
1.170.840.85
No. of fryproduced
12 50712 50710 494
Table 2. Fry production of three strains of O. niloticus in circular earthen ponds of 100 m2 each, 1996.
Strain
AbbassaZawiaMaryout
No. offemale
252525
Meanweight (g)
144265282
No. ofharvest
777
No. of fry/female
286217196
No. of fry/gbody weight
1.990.820.69
No. of fryproduced
50 00038 03034 230
(1) to determine the lethal salinity levels of somestrains of O. niloticus and compare these with othertilapia species and (2) to develop salinity-tolerant O.niloticus through subsequent use of survivors inbreeding program. The study is in progress, and newdata on genetic parameters and heritability values forsalinity tolerance will be estimated.
Genetic and physiological studies on fishfrom polluted locations
Seas, rivers and lakes are the eventual sinks for manyof harmful or waste substances disposed of by humans.Aquatic life, including food fishes, is capable ofabsorbing and accumulating various chemicalsespecially heavy metals which cause adverse effectson the aquatic biota. These effects include deleteriouschanges which disrupt the metabolic activity at abiochemical level (Hinton et al. 1973).
Egypt’s coastal lakes act as temporary reservoirs fordrainage water and often are highly contaminated withanthropogenic materials. This is true particularly forLake Manzala and Lake Maryout and to a lesser extentfor Lake Edku (El-Rayis and Saad 1984).
Collaborative research between CLAR and theDepartment of Zoology, Faculty of Science, ZagazigUniversity, is in progress to study the genotypeof fish collected from different polluted areas,chromosomal aberrations, biochemical electrophoresisanalysis for protein, and some isozymes andmicronucleus test on freshwater fish.
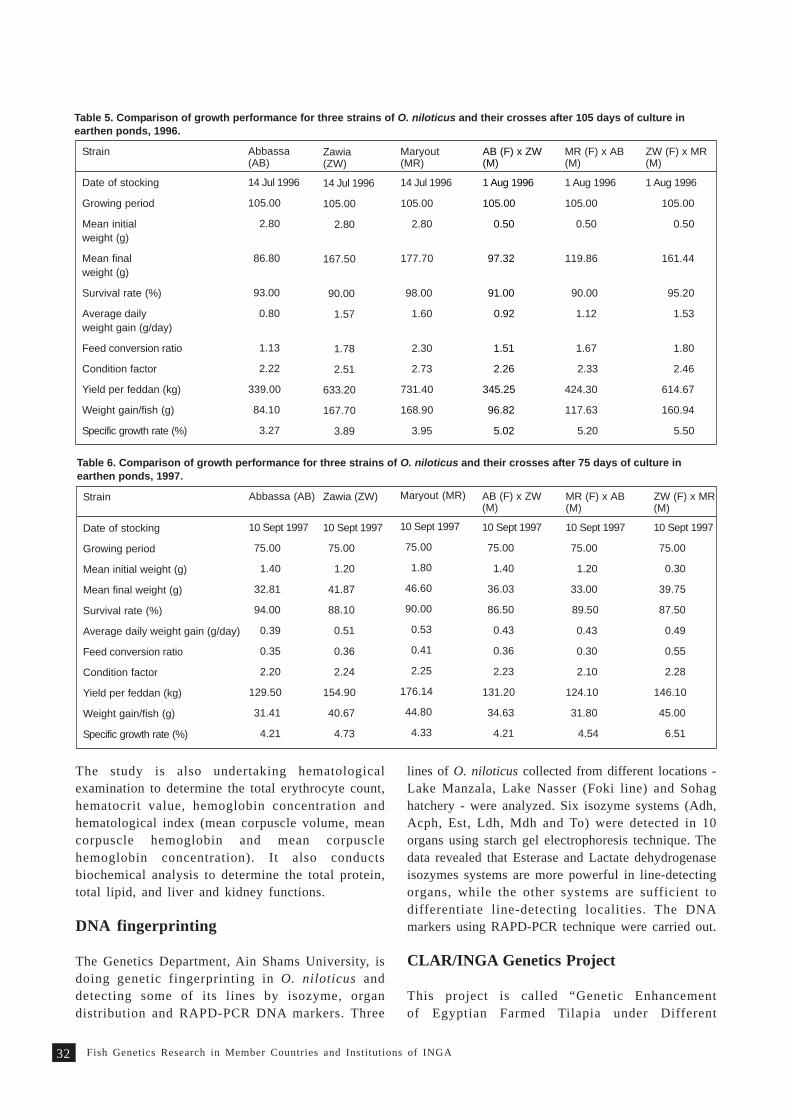
Fish Genetics Research in Member Countries and Institutions of INGA32
The study is also undertaking hematologicalexamination to determine the total erythrocyte count,hematocrit value, hemoglobin concentration andhematological index (mean corpuscle volume, meancorpuscle hemoglobin and mean corpusclehemoglobin concentration). It also conductsbiochemical analysis to determine the total protein,total lipid, and liver and kidney functions.
DNA fingerprinting
The Genetics Department, Ain Shams University, isdoing genetic fingerprinting in O. niloticus anddetecting some of its lines by isozyme, organdistribution and RAPD-PCR DNA markers. Three
lines of O. niloticus collected from different locations -Lake Manzala, Lake Nasser (Foki line) and Sohaghatchery - were analyzed. Six isozyme systems (Adh,Acph, Est, Ldh, Mdh and To) were detected in 10organs using starch gel electrophoresis technique. Thedata revealed that Esterase and Lactate dehydrogenaseisozymes systems are more powerful in line-detectingorgans, while the other systems are sufficient todifferentiate line-detecting localities. The DNAmarkers using RAPD-PCR technique were carried out.
CLAR/INGA Genetics Project
This project is called “Genetic Enhancementof Egyptian Farmed Tilapia under Different
Table 6. Comparison of growth performance for three strains of O. niloticus and their crosses after 75 days of culture inearthen ponds, 1997.
Zawia (ZW)
10 Sept 1997
75.00
1.20
41.87
88.10
0.51
0.36
2.24
154.90
40.67
4.73
Strain
Date of stocking
Growing period
Mean initial weight (g)
Mean final weight (g)
Survival rate (%)
Average daily weight gain (g/day)
Feed conversion ratio
Condition factor
Yield per feddan (kg)
Weight gain/fish (g)
Specific growth rate (%)
Abbassa (AB)
10 Sept 1997
75.00
1.40
32.81
94.00
0.39
0.35
2.20
129.50
31.41
4.21
AB (F) x ZW(M)
10 Sept 1997
75.00
1.40
36.03
86.50
0.43
0.36
2.23
131.20
34.63
4.21
Maryout (MR)
10 Sept 1997
75.00
1.80
46.60
90.00
0.53
0.41
2.25
176.14
44.80
4.33
ZW (F) x MR(M)
10 Sept 1997
75.00
0.30
39.75
87.50
0.49
0.55
2.28
146.10
45.00
6.51
MR (F) x AB(M)
10 Sept 1997
75.00
1.20
33.00
89.50
0.43
0.30
2.10
124.10
31.80
4.54
Zawia(ZW)
14 Jul 1996
105.00
2.80
167.50
90.00
1.57
1.78
2.51
633.20
167.70
3.89
Table 5. Comparison of growth performance for three strains of O. niloticus and their crosses after 105 days of culture inearthen ponds, 1996.
Strain
Date of stocking
Growing period
Mean initialweight (g)
Mean finalweight (g)
Survival rate (%)
Average dailyweight gain (g/day)
Feed conversion ratio
Condition factor
Yield per feddan (kg)
Weight gain/fish (g)
Specific growth rate (%)
Abbassa(AB)
14 Jul 1996
105.00
2.80
86.80
93.00
0.80
1.13
2.22
339.00
84.10
3.27
AB (F) x ZW(M)
1 Aug 1996
105.00
0.50
97.32
91.00
0.92
1.51
2.26
345.25
96.82
5.02
Maryout(MR)
14 Jul 1996
105.00
2.80
177.70
98.00
1.60
2.30
2.73
731.40
168.90
3.95
ZW (F) x MR(M)
1 Aug 1996
105.00
0.50
161.44
95.20
1.53
1.80
2.46
614.67
160.94
5.50
MR (F) x AB(M)
1 Aug 1996
105.00
0.50
119.86
90.00
1.12
1.67
2.33
424.30
117.63
5.20

33
Environmental and Culture Conditions”. Its researchincludes:• evaluation of strains of O. niloticus to identify the
ones that perform better under differentenvironmental and climatic conditions (ponds,cages, ricefields);
• genetic characterization of different strains; and• production of new breeds that are more suitable
for Egyptian aquaculture conditions.The data collected from evaluation work will be usedto establish an effective selective breeding program.
For this research, four different stocks of O. niloticuswere collected from Ismailia canal at the center ofDelta area (around Abbassa area) with moderatetemperature, Lake Manzala in the northern part ofDelta, and Lake Nasser, south of Egypt, with hightemperature climate and Maryout strain fromMaryout hatchery, originally from north of Egyptwith cold temperature climate. Reproductiveperformance of Ismailia, Manzala and Maryout stockshas been studied.
Winter growth evaluation of Nile tilapia strains
Temperature tolerance is a key factor in tilapiaproduction in temperate zones and to some extent insubtropical zones where tilapia aquaculture ischaracterized by seasonal changes in watertemperature (Lahav and Ra’anan 1997). Duringwinter, water temperature may drop to levels that causesevere growth inhibitions and sometimes, mortality.A species or a hybrid with greater cold tolerancebecomes more valuable and may significantly improvethe profitability of the industry. Moreover, therelatively short growing season in temperate climatescould be extended as species and/or strains of cold-tolerant tilapia are surviving and/or able to continuegrowing at temperatures few degrees less than theirnoncold tolerant counterparts. In a three-replicateexperiment, three strains of O. niloticus were stockedin 0.1 ha earthen ponds starting 15 December 1998,as temperature was declining in order to compare thewinter growth for the three strains. As the workprogresses, new data on genetic parameters andheritability values for cold tolerance will be developed.
Genetic evaluation of African catfish
Taxonomists and systematists attempt to distinguishspecies and hypothesize lineage by conducting studiesof genetically based differences and similarities among
populations. Historically, genetic differentiation hasusually been inferred from a comparison ofmorphological characters. There is, however, anincreasing trend in taxonomy to supplementmorphological, analogical, ethalogical, biochemicalor karyotypic characters among populations as ameans of detecting genetic divergence. The Africancatfish is widely distributed throughout Africa. Itinhabits tropical swamps, lakes and rivers, some ofwhich are subject to seasonal drying. In the northenand central part of Africa, it has been described asClarias lazera; in the eastern part as C. senegalensis; inthe western part as C. mossambicus; and in the southernpart as C. gariepinus.
The research at CLAR aims to:• evaluate different catfish strains collected from
various environmental conditions;• make a biochemical genetic analysis (SDS-PAGE
and isozyme electrophoresis) and a RAPD-DNAmarker analysis.
It plans to collect broodstocks of Clarias from threelocations, Abbassa, Kafer El-Seikh and Nile River, andevaluate the origin of these strains.
Other Research Highlights
The following research activities are being undertakenat the National Institute of Oceanography andFisheries, Alexandria:• karyological analysis of two species of family
Sparidae (Sparus auratus and Lithognathusmormyrus);
• isolation of salinity-resistant gene from Artemiasalina;
• genotype analysis of Tilapia spp. at Lake Manzala;• determination of the effect of water pollution in
Lake Manzala on RNA/DNA ratio in Tilapia spp.in Egypt;
• genetic differentiation of sarcoplasmic protein infamily Mugilidae at different habitats;
• determination of genetic variability of Mugilcephalus in freshwater and marine habitats;
• determination of the effect of two herbicides,Saturn and Ronstar, on the cytological characterof and on electrophoretic protein polymorphismof C. lazera.
At the General Authority for Fish ResourcesDevelopment, genetic development within tilapiastrains in Egypt was carried out through a selection
Aquaculture Genetics Research in Egypt

Fish Genetics Research in Member Countries and Institutions of INGA34
program in government hatcheries. O. niloticus(parent, first and second generations) was selected forbody weight and total body length of each of the threelines (Saft–Khaled, Sohag and Foki lines).
At the Idfena Veterinary College, AlexandriaUniversity, research on tilapia includes commercialproduction of monosex tilapia and supermaleproduction of O. niloticus.
Most research activities at the Genetic EngineeringResearch Institute, Monofaya University, Sadat City,deal with fingerprinting and gene transfer.
ReferencesDunham, R.A. and R.O. Smitherman 1984. Ancestry and breeding
of catfish in the United States. Circ. 273, Auburn University,Alabama, USA. 100 p.
El Rayis, O.A. and A.H. Saad. 1984. Concentration of Cu, Cd, Zn,Fe, and Mn in the Nile River water and its contribution tothe Mediterranean. J. Etud. Pollut. (Lucerne) CLESM: 45-52.
Hinton, D.E., M.W. Kendall and B.B. Silver. 1973. Use of histologicand histochemical assessments in the prognosis of the effectsof aquatic pollutions, p. 528. In Biological methods for theassessment of water quality. ASTM.STP, Washington, USA.
Kincaid, H.L. 1976. Trout strain registry, FWS/NFC-L/81-1. USFisheries and Wildlife Service, Kearneysville, WV, USA. 118p .
Lahav, E. and Z. Ra’anan. 1997. Cold tolerance of geneticallyproduced all-male tilapia hybrids (Oreochromis aureus), p. 662-670. In: K. Fitzsimmons (ed.) Tilapia aquaculture:proceedings from the Fourth International Symposium onTilapia in Aquaculture, Orlando, Florida, USA, 9-12November 1997. NRAES-106. Northeast RegionalAgricultural Engineering Science, Ithaca, N.Y.

35
ABSTRACT
The reproductive, survival and growth performances of Nile tilapia (Oreochromis niloticus), Fijian Chitraladastrain and that developed from the Genetic Improvement of Farmed Tilapia (GIFT) program in thePhilippines, were evaluated in two culture environments, integrated and nonintegrated.
Results showed that the Chitralada strain had more participating breeders and higher fry survival thanthe GIFT strain. The latter had higher fecundity but the brooders showed a high incidence of fry cannibalism.It also had a slightly better growth than that of Chitralada strain. Growth of the GIFT strain in the integratedsystem was better than in the nonintegrated one, while there was no significant difference in growthbetween the two systems for Chitralada strain.
A COMPARATIVE EVALUATIONOF TWO TILAPIA STRAINS
IN FIJI
S. Nandlal, C.W. Morris, M. Lagibalavu, E. LeduaFiji Fisheries Division
Ministry of Agriculture, Fisheries and ForestsP.O. Box 819, Nausori, Fiji
P.B. MatherSchool of Natural Resource SciencesQueensland University of Technology
GPO Box 2434, Brisbane, Qld 4001, Australia
NANDLAL, S., C.W. MORRIS, M. LAGIBALAVU, E. LEDUA and P.B. MATHER. 2001. A comparative evaluation of twotilapia strains in Fiji, p. 35-41. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries andinstitutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
Tilapia genetics research started in 1993 in Fiji bythe Fisheries Division, Ministry of Agriculture,Fisheries and Forests (MAFF) in collaboration withthe Queensland University of Technology, Brisbane,Australia, and the Institute of Advanced Studies,University of Malaya, Kuala Lumpur, Malaysia,under a project entitled Genetic Identification andStock Improvement of Tilapia in Malaysia and Fiji.Phase1 of this project was funded by the AustralianCentre for International Agricultural Research(ACIAR) and was completed in December 1995.
A Comparative Evaluation of Two Tilapia Strains in Fiji
During the first phase, the culture performance offour tilapias in Fiji, namely Oreochromis mossambicus,Israel and Chitralada strains of Nile tilapia (O.niloticus) and red tilapia hybrid, was assessed forthree consecutive generations under twoaquaculture production systems (integrated andnonintegrated ponds) to determine which of thestrains was performing best. Results of the studyshowed that Chitralada strain was the bestperforming strain in Fiji.
This led to the second phase of the project onGenetic Improvement of Tilapia in Fiji and Redclaw

Fish Genetics Research in Member Countries and Institutions of INGA36
in Australia, funded also by ACIAR. The projectcommenced in January 1998 and aims in the firstinstance to compare the performance of theChitralada strain with the GIFT strain, thenundertake selection for three generations of bestperforming strain under Fijian conditions.Concurrently, microsatellite markers will also bedeveloped through collaboration betweenQueensland University of Technology and theCommonwealth Scientific and Industrial ResearchOrganization (CSIRO), Australia. Individualperformance in culture will be correlated with allelicvariation to determine the potential for marker-assisted selection for further improvement ofcultured strains.
Another objective is to provide training for researchstaff to enhance skills in experimental designtechniques and statistical methodology and intagging and related techniques for selectionprogams. The project also aims to develop extensionprocedures for farmers for the effectivedissemination of genetic gain in improved breeds.
Future research will be towards repetition of thecomparative breeding and growout trials of the twotilapia strains (GIFT vs. Chitralada).
Materials and Methods
Tilapia strains
The GIFT strain of O. niloticus, was imported fromthe Philippines in August 1997 and maintained atthe Naduruloulou Fisheries Station. Fifthgeneration of Chitralada was imported from theNational Inland Fisheries Institute, Thailand, inAugust 1988 and is currently being used by farmersthroughout the country.
Evaluation of culture performance
Breeding of the two strains was carried out atNaduruloulou Aquaculture Station from 1 May toJune 1998. In a breeding design structured togenerate as many families per strain, 25 fine-mesh breeding hapas (1m x 1m x 1m) were eachstocked with four females and two males, and wereset up in a 2 500 m2 pond. Brooders were fed with agrower mix feed twice daily until they spawned.Dead brooders resulting from damage fromhandling during stocking were replaced only duringthe first week.
Fry collection began soon after swim-up fry wereobserved. Spawned females were identified andtransferred into separate holding pondsimmediately after fry collection. For each strain, frywere collected on the same day and their numberwas estimated. They were stocked in nursery hapasat a density of approximately 200 fry per hapa. Thefry were fed a fry mix feed comprising 50% fish mealand 50% rice bran three times a day. Collection wascarried out every fourth day or whenever swim-upfry were observed. Fry ages were carefully noted,and upon transfer to larger mesh-size hapas, onlyfry spawned within 40 days of collection were pooledas a single batch. Early fry survival counts were madeon days 20-24.
The fry were then nursed for 2 months until theyreached fingerling size (10–11 g). The nurseryhapas were changed every 14 days for the firstmonth and thereafter transferred to bigger meshsize hapas (2 m x 2 m x 1.5 m) to allow for easymovement of water through the hapa.
Breeding efficiency (the proportion of femalesspawned), average fecundity (the total number offry produced divided by the number of femalespawners in the breeding cycle) and early frysurvival (after 20 days) were calculated for eachstrain at the end of the breeding trial.
Growth performance trials were undertaken duringSeptember 1998 - January 1999. Trials were carriedout in three replicate ponds integrated with ducksand in three nonintegrated ponds eachapproximately 600 m2 in size and 1m in depth. Eachintegrated pond had a bamboo hut which housed40 ducklings (2 - 4 weeks old) at the beginningof the growth cycle. Both types of ponds werelimed (1 250 kg.ha-1 of hydrated lime) and manuredwith chicken manure (1 250 kg.ha-1) before stockingof fry. Using a communal pond concept whilekeeping the strains separate, 100 representativesfrom each strain were stocked into rearing hapas (2m x 2 m x 1.5 m with mesh size 0.5 cm) in each ofthe six ponds in a randomized design. Initialweights were taken before stocking. A commercialtilapia pellet (29.9% crude protein) was used forfeeding twice daily.
Sampling of individual weights and lengths wasmade on 30 fish per strain in each pond every 23days (except for the first sampling which was 30days). Sex was noted as soon as i t could be
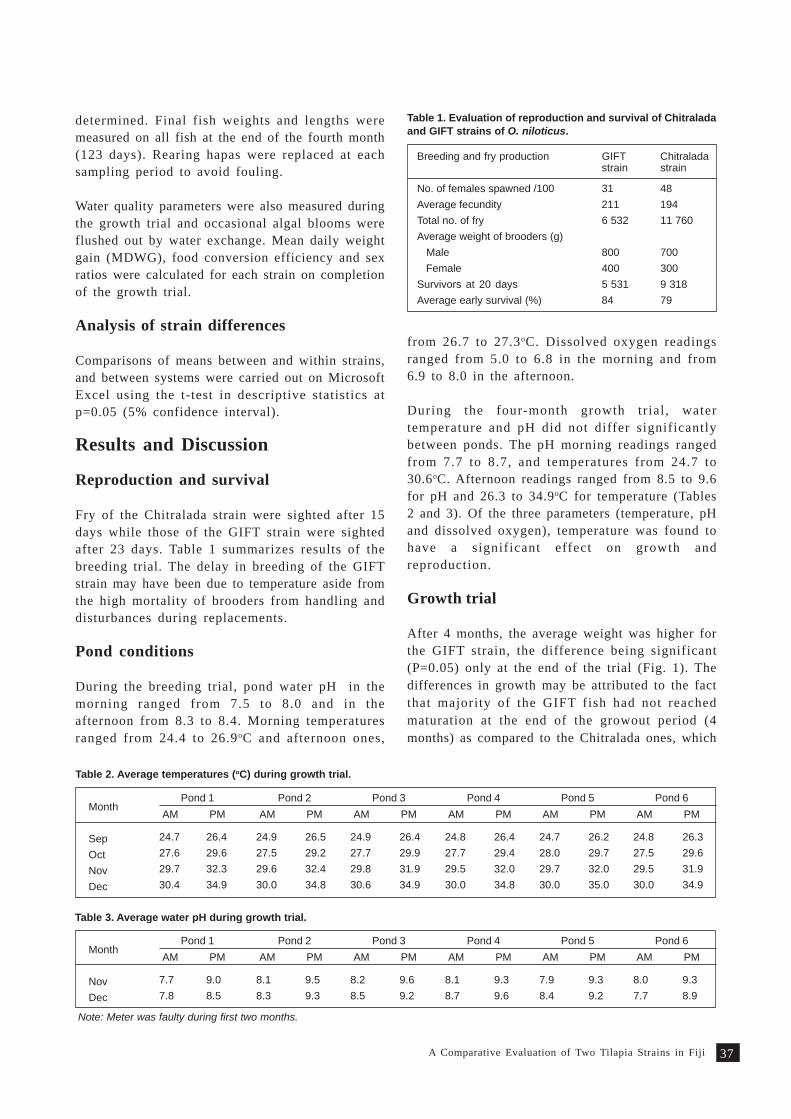
37
determined. Final fish weights and lengths weremeasured on all fish at the end of the fourth month(123 days). Rearing hapas were replaced at eachsampling period to avoid fouling.
Water quality parameters were also measured duringthe growth trial and occasional algal blooms wereflushed out by water exchange. Mean daily weightgain (MDWG), food conversion efficiency and sexratios were calculated for each strain on completionof the growth trial.
Analysis of strain differences
Comparisons of means between and within strains,and between systems were carried out on MicrosoftExcel using the t-test in descriptive statistics atp=0.05 (5% confidence interval).
Results and Discussion
Reproduction and survival
Fry of the Chitralada strain were sighted after 15days while those of the GIFT strain were sightedafter 23 days. Table 1 summarizes results of thebreeding trial. The delay in breeding of the GIFTstrain may have been due to temperature aside fromthe high mortality of brooders from handling anddisturbances during replacements.
Pond conditions
During the breeding trial, pond water pH in themorning ranged from 7.5 to 8.0 and in theafternoon from 8.3 to 8.4. Morning temperaturesranged from 24.4 to 26.9oC and afternoon ones,
from 26.7 to 27.3oC. Dissolved oxygen readingsranged from 5.0 to 6.8 in the morning and from6.9 to 8.0 in the afternoon.
During the four-month growth trial , watertemperature and pH did not differ significantlybetween ponds. The pH morning readings rangedfrom 7.7 to 8.7, and temperatures from 24.7 to30.6oC. Afternoon readings ranged from 8.5 to 9.6for pH and 26.3 to 34.9oC for temperature (Tables2 and 3). Of the three parameters (temperature, pHand dissolved oxygen), temperature was found tohave a significant effect on growth andreproduction.
Growth trial
After 4 months, the average weight was higher forthe GIFT strain, the difference being significant(P=0.05) only at the end of the trial (Fig. 1). Thedifferences in growth may be attributed to the factthat majority of the GIFT fish had not reachedmaturation at the end of the growout period (4months) as compared to the Chitralada ones, which
Breeding and fry production GIFT Chitraladastrain strain
No. of females spawned /100 31 48Average fecundity 211 194Total no. of fry 6 532 11 760Average weight of brooders (g) Male 800 700 Female 400 300Survivors at 20 days 5 531 9 318Average early survival (%) 84 79
Table 1. Evaluation of reproduction and survival of Chitraladaand GIFT strains of O. niloticus.
Month
SepOctNovDec
Table 2. Average temperatures (oC) during growth trial.
Pond 6 AM PM
24.8 26.327.5 29.629.5 31.930.0 34.9
Pond 5 AM PM
24.7 26.228.0 29.729.7 32.030.0 35.0
Pond 4 AM PM
24.8 26.427.7 29.429.5 32.030.0 34.8
Pond 3 AM PM
24.9 26.427.7 29.929.8 31.930.6 34.9
Pond 2 AM PM
24.9 26.527.5 29.229.6 32.430.0 34.8
Pond 1 AM PM
24.7 26.427.6 29.629.7 32.330.4 34.9
A Comparative Evaluation of Two Tilapia Strains in Fiji
Note: Meter was faulty during first two months.
Table 3. Average water pH during growth trial.
Month
NovDec
Pond 6 AM PM
8.0 9.37.7 8.9
Pond 5 AM PM
7.9 9.38.4 9.2
Pond 4 AM PM
8.1 9.38.7 9.6
Pond 3 AM PM
8.2 9.68.5 9.2
Pond 2 AM PM
8.1 9.58.3 9.3
Pond 1 AM PM
7.7 9.07.8 8.5
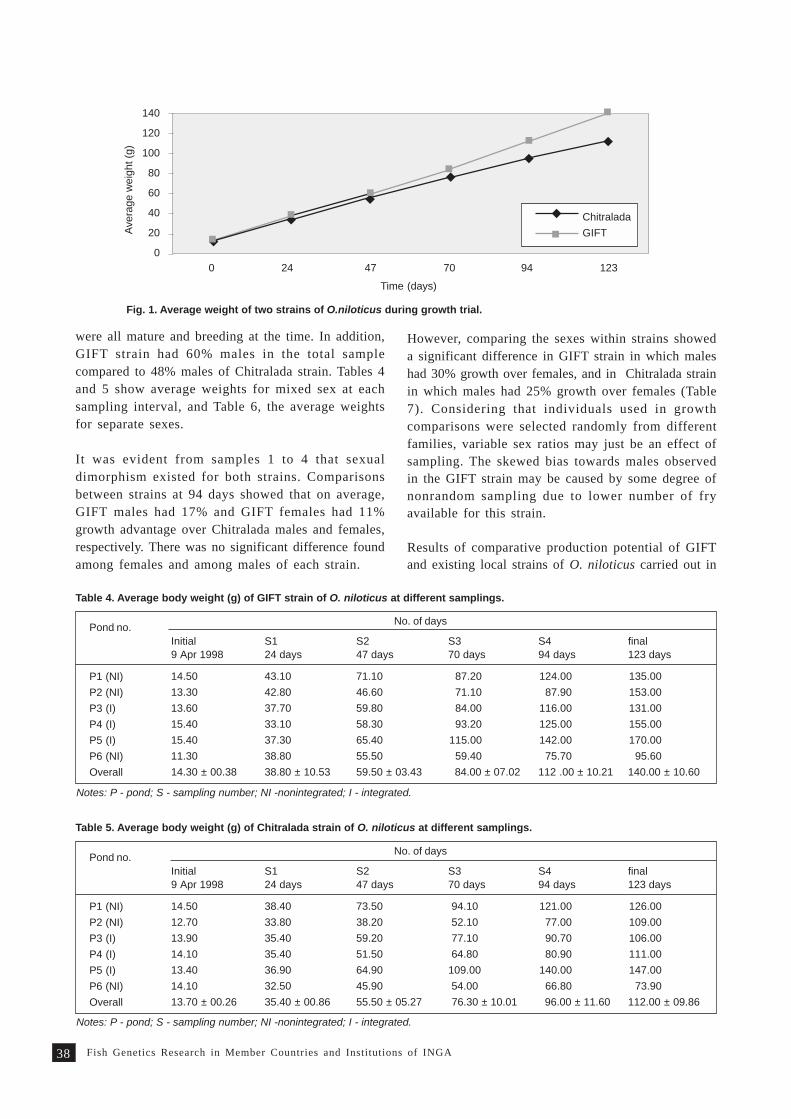
Fish Genetics Research in Member Countries and Institutions of INGA38
were all mature and breeding at the time. In addition,GIFT strain had 60% males in the total samplecompared to 48% males of Chitralada strain. Tables 4and 5 show average weights for mixed sex at eachsampling interval, and Table 6, the average weightsfor separate sexes.
It was evident from samples 1 to 4 that sexualdimorphism existed for both strains. Comparisonsbetween strains at 94 days showed that on average,GIFT males had 17% and GIFT females had 11%growth advantage over Chitralada males and females,respectively. There was no significant difference foundamong females and among males of each strain.
However, comparing the sexes within strains showeda significant difference in GIFT strain in which maleshad 30% growth over females, and in Chitralada strainin which males had 25% growth over females (Table7). Considering that individuals used in growthcomparisons were selected randomly from differentfamilies, variable sex ratios may just be an effect ofsampling. The skewed bias towards males observedin the GIFT strain may be caused by some degree ofnonrandom sampling due to lower number of fryavailable for this strain.
Results of comparative production potential of GIFTand existing local strains of O. niloticus carried out in
Pond no.
P1 (NI)P2 (NI)P3 (I)P4 (I)P5 (I)P6 (NI)Overall
Table 4. Average body weight (g) of GIFT strain of O. niloticus at different samplings.
Notes: P - pond; S - sampling number; NI -nonintegrated; I - integrated.
Initial9 Apr 1998
14.5013.3013.6015.4015.4011.3014.30 ± 00.38
No. of days
S124 days
43.1042.8037.7033.1037.3038.8038.80 ± 10.53
S247 days
71.1046.6059.8058.3065.4055.5059.50 ± 03.43
S370 days
87.2071.1084.0093.20
115.0059.40
84.00 ± 07.02
S494 days
124.0087.90
116.00125.00142.0075.70
112 .00 ± 10.21
final123 days
135.00153.00131.00155.00170.00
95.60140.00 ± 10.60
Pond no.
P1 (NI)P2 (NI)P3 (I)P4 (I)P5 (I)P6 (NI)Overall
Table 5. Average body weight (g) of Chitralada strain of O. niloticus at different samplings.
Notes: P - pond; S - sampling number; NI -nonintegrated; I - integrated.
Initial9 Apr 1998
14.5012.7013.9014.1013.4014.1013.70 ± 00.26
No. of days
S124 days
38.4033.8035.4035.4036.9032.5035.40 ± 00.86
S247 days
73.5038.2059.2051.5064.9045.9055.50 ± 05.27
S370 days
94.1052.1077.1064.80
109.0054.00
76.30 ± 10.01
S494 days
121.0077.0090.7080.90
140.0066.80
96.00 ± 11.60
final123 days
126.00109.00106.00111.00147.00
73.90112.00 ± 09.86
Fig. 1. Average weight of two strains of O.niloticus during growth trial.
Time (days)
140
120
100
80
60
40
20
0
Aver
age
wei
ght (
g)
0 24 47 70 94 123
ChitraladaGIFT
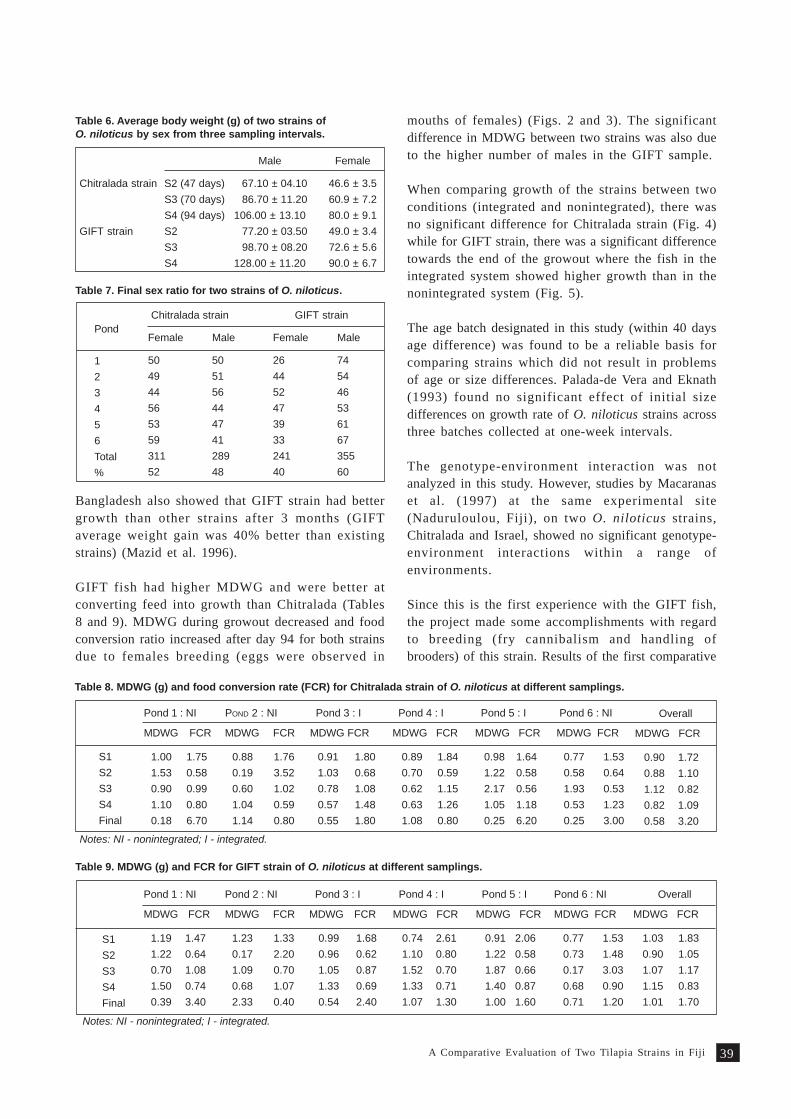
39
Bangladesh also showed that GIFT strain had bettergrowth than other strains after 3 months (GIFTaverage weight gain was 40% better than existingstrains) (Mazid et al. 1996).
GIFT fish had higher MDWG and were better atconverting feed into growth than Chitralada (Tables8 and 9). MDWG during growout decreased and foodconversion ratio increased after day 94 for both strainsdue to females breeding (eggs were observed in
mouths of females) (Figs. 2 and 3). The significantdifference in MDWG between two strains was also dueto the higher number of males in the GIFT sample.
When comparing growth of the strains between twoconditions (integrated and nonintegrated), there wasno significant difference for Chitralada strain (Fig. 4)while for GIFT strain, there was a significant differencetowards the end of the growout where the fish in theintegrated system showed higher growth than in thenonintegrated system (Fig. 5).
The age batch designated in this study (within 40 daysage difference) was found to be a reliable basis forcomparing strains which did not result in problemsof age or size differences. Palada-de Vera and Eknath(1993) found no significant effect of initial sizedifferences on growth rate of O. niloticus strains acrossthree batches collected at one-week intervals.
The genotype-environment interaction was notanalyzed in this study. However, studies by Macaranaset al. (1997) at the same experimental site(Naduruloulou, Fiji), on two O. niloticus strains,Chitralada and Israel, showed no significant genotype-environment interactions within a range ofenvironments.
Since this is the first experience with the GIFT fish,the project made some accomplishments with regardto breeding (fry cannibalism and handling ofbrooders) of this strain. Results of the first comparative
Pond
123456Total%
Table 7. Final sex ratio for two strains of O. niloticus.
Chitralada strain
Female Male
50 5049 5144 5656 4453 4759 41311 28952 48
GIFT strain
Female Male
26 7444 5452 4647 5339 6133 67241 35540 60
A Comparative Evaluation of Two Tilapia Strains in Fiji
Male Female
Chitralada strain S2 (47 days) 67.10 ± 04.10 46.6 ± 3.5S3 (70 days) 86.70 ± 11.20 60.9 ± 7.2S4 (94 days) 106.00 ± 13.10 80.0 ± 9.1
GIFT strain S2 77.20 ± 03.50 49.0 ± 3.4S3 98.70 ± 08.20 72.6 ± 5.6S4 128.00 ± 11.20 90.0 ± 6.7
Table 6. Average body weight (g) of two strains ofO. niloticus by sex from three sampling intervals.
Table 8. MDWG (g) and food conversion rate (FCR) for Chitralada strain of O. niloticus at different samplings.
Notes: NI - nonintegrated; I - integrated.
S1S2S3S4Final
Pond 6 : NI
MDWG FCR
0.77 1.530.58 0.641.93 0.530.53 1.230.25 3.00
Pond 5 : I
MDWG FCR
0.98 1.641.22 0.582.17 0.561.05 1.180.25 6.20
Pond 4 : I
MDWG FCR
0.89 1.840.70 0.590.62 1.150.63 1.261.08 0.80
Pond 3 : I
MDWG FCR
0.91 1.801.03 0.680.78 1.080.57 1.480.55 1.80
POND 2 : NI
MDWG FCR
0.88 1.760.19 3.520.60 1.021.04 0.591.14 0.80
Pond 1 : NI
MDWG FCR
1.00 1.751.53 0.580.90 0.991.10 0.800.18 6.70
Overall
MDWG FCR
0.90 1.720.88 1.101.12 0.820.82 1.090.58 3.20
Table 9. MDWG (g) and FCR for GIFT strain of O. niloticus at different samplings.
Notes: NI - nonintegrated; I - integrated.
S1S2S3S4Final
Pond 6 : NI
MDWG FCR
0.77 1.530.73 1.480.17 3.030.68 0.900.71 1.20
Pond 5 : I
MDWG FCR
0.91 2.061.22 0.581.87 0.661.40 0.871.00 1.60
Pond 4 : I
MDWG FCR
0.74 2.611.10 0.801.52 0.701.33 0.711.07 1.30
Pond 3 : I
MDWG FCR
0.99 1.680.96 0.621.05 0.871.33 0.690.54 2.40
Pond 2 : NI
MDWG FCR
1.23 1.330.17 2.201.09 0.700.68 1.072.33 0.40
Pond 1 : NI
MDWG FCR
1.19 1.471.22 0.640.70 1.081.50 0.740.39 3.40
Overall
MDWG FCR
1.03 1.830.90 1.051.07 1.171.15 0.831.01 1.70
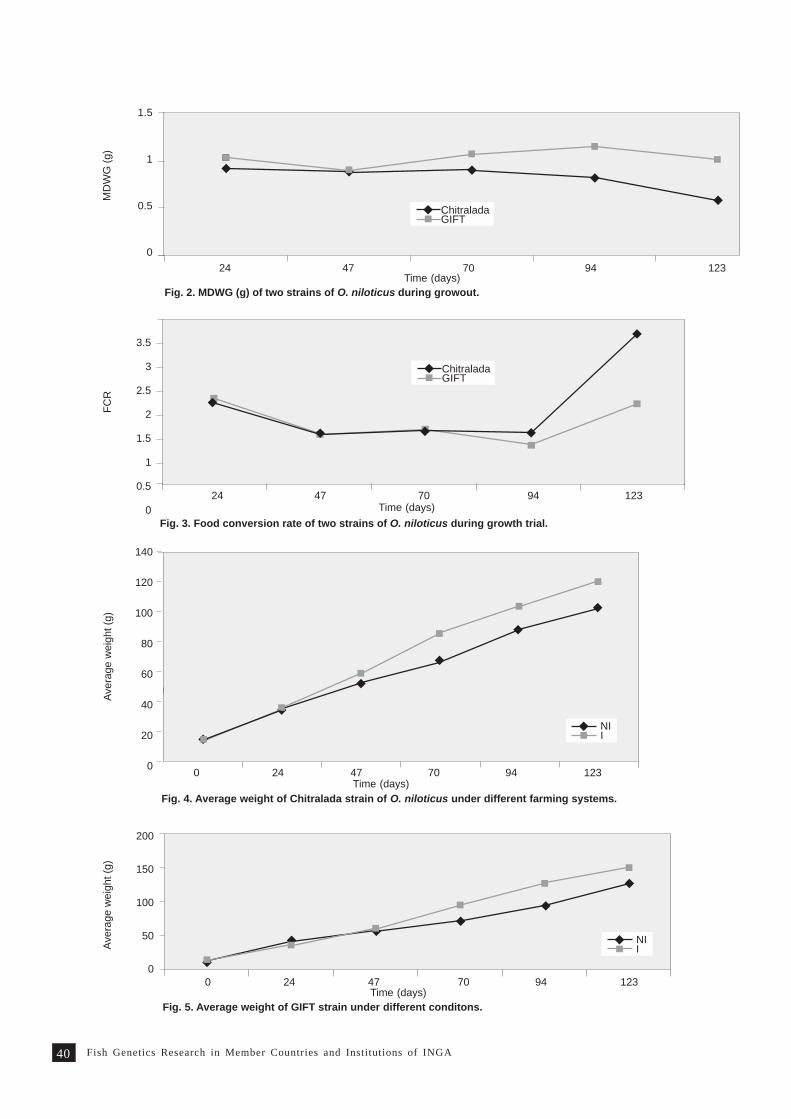
Fish Genetics Research in Member Countries and Institutions of INGA40
0 24 47 70 94 123
Aver
age
wei
ght (
g)
Time (days)Fig. 4. Average weight of Chitralada strain of O. niloticus under different farming systems.
140
120
100
80
60
40
20
0
NII
0 24 47 70 94 123
Aver
age
wei
ght (
g)
Time (days)Fig. 5. Average weight of GIFT strain under different conditons.
200
150
100
50
0
NII
1.5
1
0.5
024 47 70 94 123
MD
WG
(g)
Time (days)
ChitraladaGIFT
Fig. 2. MDWG (g) of two strains of O. niloticus during growout.
Fig. 3. Food conversion rate of two strains of O. niloticus during growth trial.
3.5
3
2.5
2
1.5
1
0.5
024 47 70 94 123
Time (days)
ChitraladaGIFT
FCR

41
breeding and growth trial indicated that the GIFTstrain performed slightly better than the Chitraladastrain. A more substantial statistical analysis of datafrom this trial is being carried out. To complete theevaluation of performance of the two strains, the trialwas repeated in April-May 1999 for breeding and inJune-September 1999 for growth.
Acknowledgements
This study is part of a three-year project entitledGenetic Improvement of Cultured Tilapia andRedclaw Crayfish in Fiji and Australia which is fundedby ACIAR and fully endorsed by MAFF, Fiji. We aregrateful for the cooperation of Technical Assistants,Jone Ratubalevu and Litiana Seravatu, who wereinvolved in the previous phase of the project. Supportwas also provided by other fieldworkers namely, RajenPrasad, Simione Lawe, Kameli Lea, Ilisoni Kalou,Amini Vuicolo and Peni Vunisau, especially during thebreeding cycle and sampling for the growth trial.
A Comparative Evaluation of Two Tilapia Strains in Fiji
ReferencesMacaranas, J., P.B. Mather, S.N. Lal, T. Vereivalu, M. Lagibalavu
and M.F. Capra. 1997. Genotype and environment: acomparative evaluation of four tilapia stocks in Fiji.Aquaculture 150: 11-24.
Mazid, M.A., M. Kamal and M.G. Hussain. 1996. Fish geneticresearch progress and planned activities in Bangladesh. InProceedings from the Third Steering Committee Meeting,International Network on Genetics in Aquaculture, 8-11 July1996, Cairo, Egypt.
Palada-de Vera, M.S. and A.E. Eknath. 1993. Predictability ofindividual growth rates in tilapia. Aquaculture 111: 147-158.

Fish Genetics Research in Member Countries and Institutions of INGA42

43Aquaculture Genetics Research in India: an Overview
AQUACULTUREGENETICS RESEARCH
IN INDIA : AN OVERVIEW
S. Ayyappana
Central Institute of Freshwater AquacultureKausalyaganga, Bhubaneswar-751002, Orissa, India
A.G. PonniahNational Bureau of Fish Genetic ResourcesCanal Ring Road, P.O. Diskusha, Telibagh
Lucknow 226002, U.P., India
P.V.G.K. Reddy, R.K. Jana, K.D. MahapatraCentral Institute of Freshwater Aquaculture
Kausalyaganga, Bhubaneswar – 751002, Orissa, Indiaand
Y. BasavarajuUniversity of Agricultural Sciences, Fisheries Research Station
Hesaraghatta, Bangalore, 560 089, Karnataka, India
AYYAPPAN, S., A.G. PONNIAH, P.V.G.K. REDDY, R.K. JANA and K.D. MAHAPATRA and Y. BASAVARAJU 2001. Aquaculturegenetics research in India: an overview, p. 43-50. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research inmember countries and institutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc.64, 179 p.
ABSTRACT
Aquaculture genetics research in India is relatively recent. Initially, research was limited to production and evaluationof carp hybrids. Among the Indian carp species, as many as six interspecific and 13 intergeneric hybrids have beenproduced. Besides these, hybrid crosses were also made between Indian and Chinese grass carp, silver carp andcommon carp. The crosses between common carp and Indian carp resulted in sterile hybrids.
Methodologies for chromosomal engineering, leading to gynogenesis and polyploidy (triploidy and tetraploidy), werealso developed for Indian and Chinese carps for application in aquaculture. Sex manipulation of fish, particularly intilapia and common carp, were also carried out successfully.
A survey of some carp seed producing hatcheries in India indicated inbreeding. Selective breeding work was undertakenin Rohu (Labeo rohita) and Catla (Catla catla) to produce genetically improved seed and to develop proper breedingprocedures for the hatchery managers in the country to avoid inbreeding in hatchery stocks.
An overview of these research efforts in genetic improvement is discussed in this paper.
a Current address: Central Institute of Fisheries Education, Jai Prakash Road Versova, Mumbai – 400 061 India

Fish Genetics Research in Member Countries and Institutions of INGA44
Introduction
Inland aquaculture in India is centered on carps, i.e.,Indian major carps, catla (Catla catla), rohu (Labeorohita), mrigal (Cirrhinus mrigala), Chinese carps andcommon carp (Cyprinus carpio). Until the mid-1950s,fish culturists depended on wild collections of Indiancarps seed from rivers. However, the success ofhypophysation technique made spawning these fishunder captivity possible. This has helped farmers inprocuring seed from hatcheries.
However, in an attempt to produce more seed to meetthe increasing demand, the seed producer (i.e., aqualified hatchery manager or a fish farmer) did notpay attention to the quality. Consequently, the carpseed produced from many hatcheries was reported toshow signs of inbreeding depression, such as poorsurvival and growth, susceptibility to disease,deformities, etc.
Hence, it became essential to carry out research ongenetic improvement of these carps through variousapproaches. In the 1960s and 1970s, fish geneticresearch was mainly restricted to cytogenetic studiesand hybridization. Later, chromosome manipulation,developing breeding programs for important carpspecies and application of molecular techniques wereemphasized.
Several national and central institutions and stateagricultural and other universities undertake fishgenetic research. Some of these are the CentralInstitute of Freshwater Aquaculture (CIFA), NationalBureau of Fish Genetic Resources (NBFGR), CentralInstitute of Fisheries Education (CIFE), University ofAgricultural Sciences (UAS), Bangalore (at FisheriesResearch Station, Hesaraghatta, and College ofFisheries, Mangalore), Centre for Cellular andMolecular Biology (CCMB), Madhurai KamrajUniversity (MKU) and Bose Institute.
Genetics Research
Hybridization
In any genetic/stock improvement program, it isnecessary to breed the candidate species artificiallyunder controlled conditions. The success of inducedbreeding of Indian major carps paved the way forresearch on genetic improvement of carps. In the initialstages, simple interspecific and intergeneric
hybridization was done to produce and evaluate thecarps’ useful traits for aquaculture. Hybridization isone of the methods used for combining desirable traitsof selected species, with experiments demonstrating ahigh level of compatibility among Indian carps.
Altogether, six interspecific and 13 intergeneric hybridswere produced among the four species of Indian majorcarps belonging to three genera, i.e., Catla, Labeo andCirrhinus (Reddy 1999). Over three decades of researchon hybridization showed that these major carps withdistinct morphological characters are highlycompatible and able to produce viable and fertilehybrid progenies. Mature interspecific and intergenerichybrids could be induced to produce F2 progeny orbackcross and triplecross hybrids.
The growth exhibited by most of these hybrids wasintermediate, i.e., better than the slow-growing parentas in the case of interspecific hybrid between L. rohitaand L. calbasu or intergeneric hybrids between L. rohitaand C. catla or C. catla and C. mrigala, etc.
Hybridization between Indian major carps (C. catla andL. rohita) and medium carp L. fimbriatus also had asimilar trend in growth with hybrids showingsuperiority over slow-growing parents (Basavaraju etal. 1989, 1990 and 1995).
With regard to hybridization between Indian majorcarps and Chinese grass carp (Ctenopharyngodon idella),silver carp (Hypophthalmichthys molitrix) and bigheadcarp (Aristichthys nobilis), no compatibility was observed.Almost all of these died either during the embryonicdevelopment or soon after hatching. Only a few hybridsbetween C. catla and H. molitrix survived.
Relatively better survival of the hybrid progeny wasobserved in the crosses between female common carp(C. carpio) and males of C. catla, L. rohita and C. mrigalacompared with the reciprocal hybrid crossesparticularly between female L. rohita and male C. carpio(Kowtal and Gupta 1985; John and Reddy 1987;Khan et al. 1989).
It is necessary to prevent indiscriminate hybridization.These carps are highly compatible to interbreed. Also,since almost all interspecific and intergeneric hybridswere found fertile, one should be more cautious of theircrossbreeding as this may cause serious damage to theoriginal gene pool through genetic introgression(Padhi and Mandal 1994).

45
Chromosome manipulation
Chromosome manipulation as a tool to improvegenetic status of fish was initiated at CIFA and in otherinstitutes.
Meiotic gynogenesis was successfully induced in Indianmajor carps for the first time in L. rohita, C. catla (Johnet al. 1984) and C. mrigala (John et al. 1988). Efficacyof shock, i.e., thermal and pressure, and duration oftreatment seem to vary from species to species.Effective intensity lies at some point around thesublethal level. Thus, in all species of Indian carps,cold shock below 10oC and heat shock above 42oCproved to be lethal (John et al. 1984; Reddy et al. 1993).
The first successful induction of mitotic gynogenesisin L. rohita was reported by Reddy et al. (1993). Theuse of milt from heterologous species preferably havinglow compatibility to hybridize may be more desirablefor easy elimination of paternal genetic input withirradiation.
Induction of androgenesis in Indian major carps wasnot very successful as egg consists of huge mass ofcytoplasm and yolk that prevent the proper exposureof the chromosomes to UV rays.Some preliminaryattempts have been made to induce artificial inductionof triploidy and tetraploidy in Indian major carps withvarying degrees of success (Reddy et al. 1987; 1990).The genetics division of CIFA succeeded in inducingtriploidy in L. rohita and tetraploidy in L. rohita and C.catla. Common carp triploid showed 60-100% moregrowth than its diploid siblings (Reddy et al. 1998).Triploid C. idella can be utilized in openwater reservoirsto control aquatic weeds without propagating itself asit is supposed to be sterile.
Triploidy was successfully induced in C. carpio throughheat shock treatment to produce sterile individuals asan approach to control unwanted reproduction in grow-out ponds. Heat shock at 40°C, 1-3 min afterfertilization for 1.5 min resulted in 100% triploidy. Thetriploid C. carpio was found to grow as fast as diploidand yield 15-18% extra edible meat because of lowGSI in case of triploids (Basavaraju et al. 1998).
An attempt to induce sterility in freshwater catfishHeteropneustis fossillis through heat shock was partiallysuccessful. Heat shock for 3 min at 40°C, 4 min afterfertilization yielded 46% triploids.
Current research in chromosomal engineering in Indianmajor carps aims to produce allotriploids of carphybrids (C. catla x L. rohita and L. calbasu x L. rohita)and to evaluate them for resistance against parasiticinfection. In C. carpio, the objective of chromosomemanipulation is to produce sterile fish to optimizeproduction in aquaculture.
Sex manipulation
Research on manipulation of sex (production of sterileand monosex population) as a tool for increasingproduction has been in progress in many institutes inIndia. Administration of 17-α methyltestosterone (MT)through diet to eight-day old fry for a period of 45days induced complete sterility in C. carpio. Comparedwith normal fish, sterile fish grew faster (21%) and hadbetter resistance against bacteria (Mohire et al. 1999).
In progress at UAS, Bangalore, are studies on theproduction of monosex population of C. carpio throughhormone sex reversal and progeny testing as analternative approach to overcome sexual maturationand unwanted reproduction. UAS, in collaborationwith the International Center for Living AquaticResources Management (ICLARM), is carrying outresearch to evaluate performance of crossbreeds ofIndian major carp and to assess inbreeding of C. catlafrom different hatcheries of Karnataka state and theusefulness of sterile triploid and monosex C. carpio inaquaculture.
Selective breeding
CIFA, in collaboration with the Institute of AquacultureResearch of Norway (AKVAFORSK), undertook geneticimprovement of L. rohita through selective breeding.The project started in 1992, and the first phase wascompleted in 1997. The second phase is in progress.L. rohita was chosen as the model fish, not only becauseof consumer preference, but also because of its poorgrowth in many polyculture systems. Besides, L. rohitais also prone to disease, particularly parasitic infection.
The founder population of L. rohita was collected asfry/fingerlings from five different rivers, i.e., Ganga,Gomati, Yamuna, Sutlej and Brahmaputra, in additionto CIFA hatchery stocks. The stocks were marked byM-prucian blue dye marking/fin clipping forcommunal rearing in the same pond to raise them tobroodfish.
Aquaculture Genetics Research in India: an Overview

Fish Genetics Research in Member Countries and Institutions of INGA46
Full-sib families were produced as maternal/paternalhalf-sibs. Full-sib families were reared separately innursery ponds until they reached 10-15 g size.Individual tagging was done by using passiveintegrated transponder (PIT) tag and stocked incommunal ponds for rearing in monoculture orpolyculture with C. catla and C. mrigala.
After nearly one year of rearing, estimation of breedingvalue was done following combined selection.Individuals were ranked according to their breedingvalue and the top 300-500 individuals were selectedas broodfish for the next generation. Individuals withbreeding value around the mean value were taken ascontrol groups.
Full-sib progeny groups were produced from selectedgroup of broodfishes after avoiding full-sib/half-sibmating. Mass breeding was conducted for productionof control group progeny from control broodfishes.After one year of rearing, realized response wascalculated taking growth as the trait of selection (Reddyet al. 1998).
Large differences in growth between full-sib familieswithin stocks were observed in both monoculture andpolyculture systems, which prove that the founderstocks of L. rohita have large genetic variation and theselective breeding program did not start from a closedpopulation (Table 1).
Growth and survival of wild (riverine) stocks were equalor better than that of the farmed (local) stocks,indicating that the present procedure followed byhatcheries for seed production is improper (Reddy etal. in press).
Ranking of full-sib groups is highly consistent forgrowth in monoculture and polyculture systems,showing that different breeding plans are not requiredfor different production systems.
Diallele crosses of four riverine and farmed stocks wereconducted. Negative heterosis effect was observed in
growth of L. rohita, indicating that pure breeding mightbe more suitable for selection (Gjerde et al. 1999).
Realized selection responses were calculated after eachgeneration of selection for first and second line basepopulations (Fig. 1). Since genotype interactssensitively with environment, these findings have tobe tested under different agroclimatic conditions inthe country. Field experiments to this effect were startedin Ludhiana (Punjab), Bangalore (Karnataka),Vijaywada (Andhra Pradesh) and Kausalyaganga statefish farm (Orissa).
Genetically improved L. rohita was supplied to somecarp hatcheries in India to replace their broodstock.Studies for formulation of effective feed for improvedL. rohita are in progress.
In 1995, a research program to assess the genetic statusof Indian major carps with reference to C. catla and tostart a sustainable breeding program was initiated atthe Fisheries Research Station, UAS, in collaborationwith the Fish Genetic Program of the University ofWales, UK. A survey on broodstock management andbreeding practices by major carp hatcheries revealedthat these hatcheries function as isolated units. Thesurvey also indicated that the stocks are likely to beinbred (Table 2) and that their performance willpossibly decline if corrective measures are not taken.Germplasm from four wild stocks were collected andwill be evaluated. Progress has been slow due to latematurity (3+ years) and difficulty in gettingsimultaneously ripe breeders of all stocks.
Genetic characterization
Identification and genetic characterization of wild andhatchery populations are important since broodstockwith good genetic background is necessary for asuccessful breeding program. The knowledge fromsuch investigations can be used in optimizing andsustaining yield, stock management and conservationof genetic diversity. The work being undertaken in thebiotechnology division of CIFA involves molecular
Pure stocksCrossesHeterosis
Table 1. Average heterosis for body weight and survival of L. rohita.
Body weight (g)
Monoculture Polyculture
445.3 446.5437.7 414.7-7.7 + 10.3 -31.8 + 13.5
Survival (%)
Monoculture Polyculture
90.0 68.890.0 74.70.00 + 3.1 +5.9 + 4.3

47
genetic characterization of Indian major carps usingdifferent types of DNA markers.
Gene mapping
Gene mapping is done for localization and functionalcharacterization of economically important trait genesthat will improve the breeding program throughmarker-assisted selection (MAS). The work involvesisolation and development of highly polymorphic type-II DNA markers and, subsequently, development of agenetic linkage map for Indian major carps withparticular reference to L. rohita. The long-termobjective is the identification of trait-associated genessuch as disease resistance and body growth.
At present, CIFA, in collaboration with ICLARM, isundertaking studies for the utilization of DNAfingerprinting in L. rohita breeding program to:(1) determine DNA profiles of selected, unselected andcontrol stocks of L. rohita being investigated under theselective breeding program; (2) correlate performancewith DNA markers within the sampled groups forfuture use in MAS; and (3) determine genetic tags forbreeding programs.
Attempts are being made to identify RAPD markers, ifany, that could differentiate between growth-selectedand control animals. Simultaneously, probe-basedDNA fingerprinting was also initiated on selected,unselected parent and progeny DNA restrictionenzymes.
Table 2. Estimated effective population sizes and rates of inbreeding for major stocks of C. catla inKarnataka state fish hatcheries.
Hatchery
Tunga Bhadra Dam
Bhadra Reservoir Project
Kabini Reservoir Project
Shivapura Fish FarmVanivilas Sagar Fish FarmMunirabad
Species
catlarohumrigal
catlarohu
catla
Insufficient dataInsufficient dataInsufficient data
Ne DF (% per annum)
55.5 (10.5)a 0.9 (04.7)23.5 (12.3) 2.1 (04.0)18.4 (13.5) 2.7 (03.7)
18.4 (08.5) 3.5 (05.9)14.2 (06.9) 3.5 (07.2)
11.4 (04.7) 4.4 (10.6)
Base population(Second line)
47.5%
1st generation
(1996)
2nd generation
(1997)
1st generation
(1995)8.6%
Base population(First line)
(1993)
34.5%
(1994)
Fig. 1. Realized selection response of L. rohita (Das Mahapatra et al. 2000).
Aquaculture Genetics Research in India: an Overview

Fish Genetics Research in Member Countries and Institutions of INGA48
The Department of Biochemistry and the FisheriesResearch Station, UAS, have jointly taken up researchon population genetics of peninsular carp L. fimbriatususing genetic markers. This study, using RAPD marker,indicated 37% polymorphism in different populationswhile studies with 21 enzyme systems did not showpolymorphism. The use of mt-DNA PCR-RFLP as analternative marker is under study.
Biochemical genetics of two reciprocal hybrids betweenL. fimbriatus and C. catla and their parents was studiedelectrophoretically using several protein and enzymemarkers. The myogen and serum protein patternshowed high degree of homology among the parentsand absolute homology among the reciprocal hybrids.The ontogenetic expression of LDH showed somedegree of gene expression in the early developmentstudies (Basavaraju and Keshavappa et al. 1996).
Conservation of Endangered Species
To replenish the declining population of mahseer (Torputitora), a seed ranching program was initiated atGarampani in Kosi stream in Uttar Pradesh hills jointlyby NBFGR, Lucknow; National Research Center (NRC)on Coldwater Fisheries, G.P. Pant University and StateFisheries Department.
Studies on short-term preservation of spermatozoa ofDeccan mahseer (T. khudree) were also done. The trialswere conducted using plastic vial/polythene bags atambient temperature (AT) and 4°C, with or withoutoxygen. In general, the spermatozoa of mahseer rearedat 18-24°C were motile for a longer period (132 hours).Spermatozoa stored at 4°C showed better motility thanthose maintained at AT (Basavaraja et al. 1999).
Genetic markers
Screening of L. rohita founder stocks used for geneticimprovement undertaken by CIFA in collaboration withNBFGR indicated the presence of adequate geneticvariability and absence of genetic contamination.
Screening of two populations of L. rohita, from Gangaand Gomati Rivers with isozymes and isoelectricfocusing of eye lens proteins revealed polymorphismin XDH, PGM-1, PGM-2, EST-1G6PDH and ACP outof 20 isoenzymes studied at NBFGR.
With the use of a cost-effective package of geneticmarkers to quantify genetic contamination of hatcherystocks of Indian major carps, significantly higher levels
of introgressions than with phenotypic markers wererevealed. In Indian major carps, two mt-DNA geneswere successfully amplified with polymerase chainreaction and amplified reaction fragment pattern for2.5 kb was obtained for five restriction enzymes.Polymorphism was indicated in the pattern obtainedwith Eco R1 (NBFGR).
Genebanking
A compendium entitled “Fish biodiversity of India”containing information on systematics, habitat anddistribution of 2118 finfishes has been made byNBFGR. The bureau has also achieved some successin using cryopreserved sperm to transfer germplasmfor crossbreeding program between distant anddiscrete populations. The farmed C. carpio at Bilaspur(Himachal Pradesh) was crossed with frozen spermsof wild stocks of Ooty (Tamil Nadu) and Rewalsar.Rewalsar stock was found to be superior, and HimachalPradesh State Fisheries was advised to replace existingfarmed stocks of C. carpio, which was found to becontaminated with goldfish genome. A similarprogram is in progress for rainbow trout at Ooty, TamilNadu.
The NBFGR genebank has sperms of eleven species:C. catla, L. rohita, C. mrigal, C. carpio, Salmo gairdneri,Salmo trutta, T. putitora, T. khudree, Tenualosa ilisha, Labeodussumieri and Horabagus brachysoma.. The improvementof the technique over time resulted in hatching with arange of 65%-100% with majority between 80%-90%.Hatchlings have been produced from five-year oldfrozen sperms of L. rohita.
Crossbreeding of Nilgiri’s rainbow trout with faster-growing stock from Himachal Pradesh to achieve bettergrowth was successfully undertaken. The fingerlingsproduced from crossbreeding of Tamil Nadu rainbowtrout with Himachal Pradesh stocks usingcryopreserved milt have exhibited higher growth.These results showed that cryopreserved milt couldbe utilized to transfer genome.
Sperm cryopreservation protocols were developed forthe first time for the above species by NBFGR.Viablelarvae were produced from cryopreserved milt raisingthe possibility of stock enhancement in depleted areas.
Other institutes like CCMB and the National Instituteof Immunology and universities such as MKU andJawaharlal Nehru University (JNU) are developingtransgenic fish technology, including construction of

49
genomic and C-DNA libraries of Indian carps C. catla,L. rohita and Indian catfish H. fossilis and Clariasbatrachus, and isolation of growth hormone gene fromthese libraries.
At CCMB, construction of c-DNA libraries frompituitary glands of C. catla and L. rohita and sequencingof c-DNA clones of C. catla were carried out. At JNU,construction of genomic DNA libraries of C. catla andL. rohita, cloning and characterization of genomicclones and encoding interferon gamma gene(s) in thesespecies are being undertaken. MKU is also undertakingc-DNA cloning of growth hormone (GH) gene of H.fossilis and L. rohita; screening and isolation of full-length c-DNA of growth hormone of H. fossilis and L.rohita; and mapping and sequencing of full length c-DNA of GH gene of these two species.
Future Genetics Research
Following are key areas for genetics research in thefuture:• Development of breeding programs for important
aquaculture species;• Extra traits for selection in L. rohita breeding
program, such as disease resistance and quality offlesh;
• Determination of suitable age and size ofbroodfishes for production of quality seed anddevelopment of suitable plan for geneticimprovement of carps for hatchery managers;
• Application of molecular markers and genetictechniques for breeding programs;
• Continued research on genetic engineering; and• Testing and dissemination of research outputs to
end-users.
References
Basavaraja, N. 1999. Hormonal manipulation of sex in fishes.Abstract presented at the National Workshop on the Use ofGonadal Hormones in the Manipulation of Sex and Growthin Fishes, University of Agricultural Sciences, FisheriesResearch Station, Hesaraghatta, Bangalore.
Basavaraju, Y. and G.Y. Keshavappa. 1996. Evaluation of tworeciprocal hybrids Catla catla and Labeo fimbriatus for importantaquaculture traits. In The Fourth Indian Fisheries Forum heldat Kochi, India on 24-28 November 1996 (Abstract).
Basavaraju,Y., K.V. Devaraj and S.P. Ayyar. 1995. Comparative growthof two carp hybrids between Catla catla and Labeo fimbriatus.Aquaculture 129:187-191.
Basavaraju,Y., G.Y. Keshavappa, D. Seenappa and K.V. Devaraj.1989. First report on an interspecific hybrid between Labeorohita and Labeo fimbriatus. Curr. Res.18: 164-165.
Basavaraju, Y., G.Y. Keshavappa, D. Seenappa, K.V. Devaraj andK.V. Mohire. 1990. Growth of Labeo rohita x Labeo fimbriatus
hybrid in comparison with parental species, p. 23-25. In M.Joseph (ed.). The Second Indian Fisheries Forum. AsianFisheries Society Indian Branch, Mangalore.
Basavaraju, Y., S. Pradeep Kumar, G.Y. Keshavappa, D.J. Penmanand G.C. Mair. 1998. Triploid common carp Cyprinus carpio: apotential solution to the problem of precocious sexualmaturation? Paper presented at the Fifth Asian FisheriesForum: International Conference on Fisheries and FoodSecurity Beyond the Year 2000, 11-14 November 1998, ChiangMai, Thailand.
Das Mahapatra, K., P.K. Meher, J.N. Saha, B. Gjerde, P.V.G.K. Reddy,R.K. Jana, M. Sahoo and M. Rye. 2000. Selection response ofrohu, Labeo rohita (Ham.) after two generations of selectivebreeding. In. The Fifth Indian Fisheries Forum held at CIFA,Bhubaneswar, India on 17-20 Jan 2000 (Abstract).
Eknath, A.E. and R.W. Doyle. 1990. Effective population size andrate of inbreeding in aquaculture of Indian major carps.Aquaculture 85: 293-305.
Gjerde, B., P.V.G.K. Reddy, M. Rye, K.D. Mahapatra, J.N. Saha,R.K. Jana, S.D. Gupta, M. Sahoo, S. Lenka and P.Govindassammy. 1999. Average heterosis for growth andsurvival in two diallele crosses with five stocks of rohu carp, L.rohita, p. 27. In Proceedings of Aquaculture, Europe-99,Trondheim, Norway.
John, G. and P.V.G.K. Reddy.1987. A note on the Labeo rohita (Ham.)x Cyprinus carpio L. hybrid, p. 88-90. In Proceedings of theWorld Symposium on Selection, Hybridization and GeneticEngineering in Aquaculture, 27-30 May 1986, Bordeaux. Vol.II.
John, G., P.V.G.K. Reddy and R.K. Jana. 1984. Artificial gynogenesisin two Indian major carps, Labeo rohita (Ham.) and Catla catla(Ham.). Aquaculture 42: 161-168.
John, G., P.V.G.K. Reddy and R.K. Jana. 1988. Induced gynogenesisin the Indian major carp, Cirrhinus mrigala (Ham.), p. 107-109. In M. Joseph (ed.) The First Indian Fisheries Forum:Proceedings of the Asian Fisheries Society, Indian Branch,Mangalore.
Khan, H.A., P.V.G.K. Reddy and S.D. Gupta. 1989. Studies on someaspects of biology and morphology of the intergeneric hybrid,Cyprinus carpio var. communis L. x Labeo rohita (Ham.), p. 155-162. In Proceedings of the National Academy of India 59 (B).
Kowtal, G.V. and S.D. Gupta. 1985. A note on the hybrid mrigal,Cirrhinus mrigala (Ham.) x common carp, Cyprinus carpio (L.).Aquaculture 49: 179-183.
Mohire, K.V., C.V. Mohan, G.P. Sathyanarayana Rao, K.M. Shankarand K. Sushmitha. 1999. Susceptibility and inflammatoryresponse to Aeromanus hydrophilla in 17-a methyltestosterone-induced sterile common carp. Abstract presented at theNational Workshop on the Use of Gonadal Hormones in theManipulation of Sex and Growth in Fishes, University ofAgricultural Sciences, Fisheries Research Station,Hesaraghatta, Bangalore.
Padhi, B.K. and R.K. Mandal. 1994. Improper fish breedingpractices and their impact on aquaculture and fish biodiversity.Curr. Sci. 66(9): 624-626.
Reddy, P.V.G.K. 1999. Genetic resources of Indian major carps. FAOFish. Tech. Pap. 387, 76 p.
Reddy, P.V.G.K., G. John and R.K. Jana. 1987. Induced polyploidymosaics in Indian carp, Labeo rohita (Ham.). J. Inland Fish.Soc. India 19(1): 9-12.
Reddy, P.V.G.K., G.V. Kowtal and M.S. Tantia. 1990. Preliminaryobservation on induced polyploidy in Indian major carps,Labeo rohita (Ham.) and Catla catla (Ham.). Aquaculture 87:279-287.
Reddy, P.V.G.K., R.K. Jana and S.D. Tripathi. 1993. Induction ofmitotic gynogenesis in rohu (Labeo rohita, Ham.) throughendomitosis. Nucleus 36(3): 106-109.
Reddy, P.V.G.K., B. Gjerde, S.D. Gupta, K.D. Mahapatra, J.N. Saha
Aquaculture Genetics Research in India: an Overview

Fish Genetics Research in Member Countries and Institutions of INGA50
and R.K. Jana. 1998. Selective breeding and mating designfor selection in fishes with special reference to Indian majorcarps. pp. 449-456. In : Proceedings of Fish Biodiversity andConservation. Fish. Gen. Biodiversity Conserv. Natcon. Pub.05:
Reddy, P.V.G.K., B. Gjerde, S.D. Tripathi, R.K. Jana, K.D. Mahapatra,S.D. Gupta, J.N. Saha, M. Sahoo, S. Lenka, P. Govindssammy,M. Rye and T. Gjedrem. Growth and survival of six stocks ofrohu (Labeo rohita) in mono and polyculture productionsystems. Aquaculture (In press).

51
ABSTRACT
This paper presents genetics research conducted by the institutional members of the Indonesian Network of FishGenetics Research and Development (INFIGRAD) during 1998. The research included: (1) identification and geneticcharacterization of freshwater and brackishwater fish and (2) genetic improvement through selection andhybridization. This paper also reports on the contribution of the GIFT strain Nile tilapia (Oreochromis niloticus) tothe development of floating net culture in reservoirs and on the research collaborations with the InternationalNetwork on Genetics in Aquaculture/ International Center for Living Aquatic Resources Management on commoncarp and with the L’Institut Francais de Recherche Scientifique pour le Developpement en Cooperation on geneticcharacterization of catfish.
AQUACULTUREGENETICS RESEARCH
IN INDONESIA
A. Hardjamulia1, F. Sukadi2, Subagyo1 and R. Gustiano1
1 Research Institute for Freshwater FisheriesJl. Raya Sukamandi, Subang 412563 Indonesia
2 Central Research Institute for FisheriesJl. K.S. Tubun Petamburan VI, Slipi, Jakarta 11410A Indonesia
HARDJAMULIA, A., F. SUKADI, SUBAGYO and R. GUSTIANO. 2001. Aquaculture genetics research in Indonesia, p. 51-59. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of theInternational Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
Fresh and brackishwater aquaculture contributed733 000 t (16.5%) to the total fish production of about4.452 million t in Indonesia (DCFA 1998). Of this,freshwater aquaculture production was about 328 800t, of which common carp (Cyprinus carpio) was 54.3%(Table 1). Nile tilapia (Oreochromis niloticus), which wasfirst introduced in Indonesia in 1969 from Taiwan hasbeen widely accepted by fish farmers. Common carpand Nile tilapia (recently the GIFT strain) arecultivated intensively, especially in floating net cagesin reservoirs.
Common carp culture started in West Java in themiddle of 19th century (Ardiwinata 1971) and
Aquaculture Genetics Research in Indonesia
subsequently, was taken up in other parts of Java,Sumatra and Sulawesi islands since early 20th century.There are at least ten stocks of common carps knownin Indonesia: Punten, Sinyonya, Majalaya, Domas,Merah, Kancra-domas, Kaca (mirror carp), Kumpai,Taiwan carp and Koi (Japanese fancy carp) (Taniguchiet al. 1992). Recent collections include Rajadanu,Wildan Cianjur and Sutisna Kuningan (Nugroho andWahyudi 1991).
In recent years, the genetic improvement of commoncarp and Nile tilapia has been considered importantin aquaculture operations. Research on geneticcharacterization of both fresh and marine fishes is alsogaining importance and has been part of thegovernment’s conservation efforts.
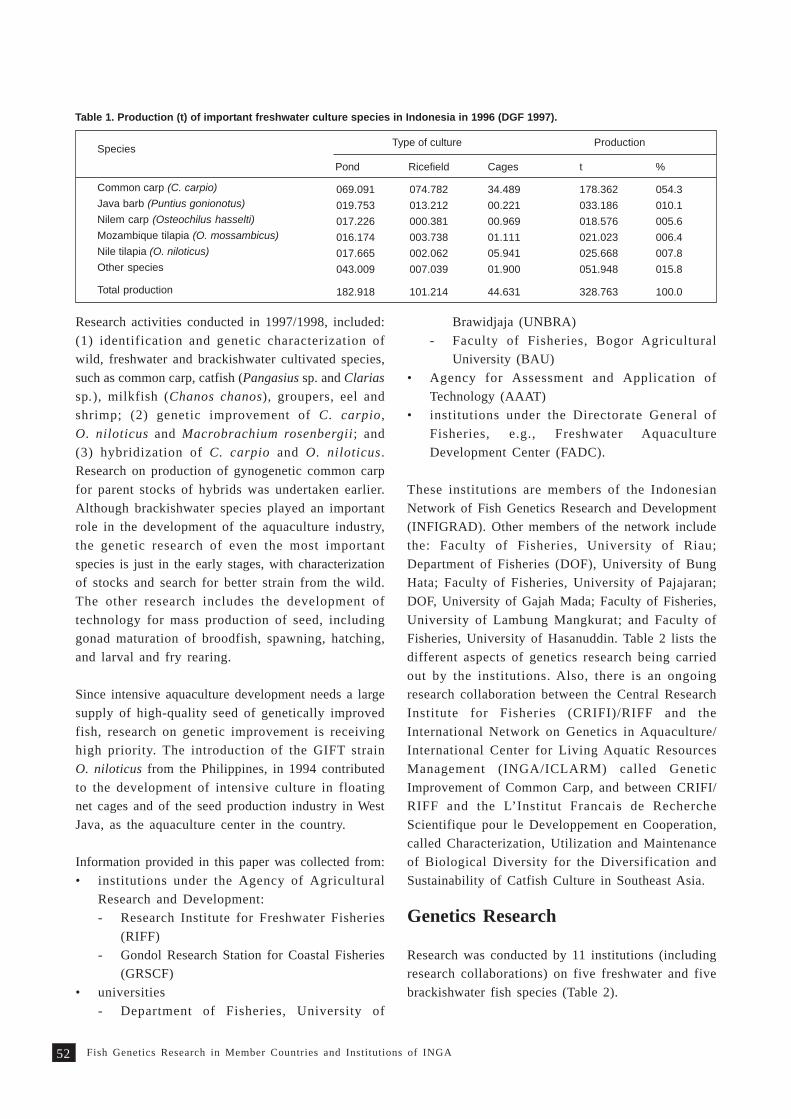
Fish Genetics Research in Member Countries and Institutions of INGA52
Research activities conducted in 1997/1998, included:(1) identification and genetic characterization ofwild, freshwater and brackishwater cultivated species,such as common carp, catfish (Pangasius sp. and Clariassp.), milkfish (Chanos chanos), groupers, eel andshrimp; (2) genetic improvement of C. carpio,O. niloticus and Macrobrachium rosenbergii; and(3) hybridization of C. carpio and O. niloticus.Research on production of gynogenetic common carpfor parent stocks of hybrids was undertaken earlier.Although brackishwater species played an importantrole in the development of the aquaculture industry,the genetic research of even the most importantspecies is just in the early stages, with characterizationof stocks and search for better strain from the wild.The other research includes the development oftechnology for mass production of seed, includinggonad maturation of broodfish, spawning, hatching,and larval and fry rearing.
Since intensive aquaculture development needs a largesupply of high-quality seed of genetically improvedfish, research on genetic improvement is receivinghigh priority. The introduction of the GIFT strainO. niloticus from the Philippines, in 1994 contributedto the development of intensive culture in floatingnet cages and of the seed production industry in WestJava, as the aquaculture center in the country.
Information provided in this paper was collected from:• institutions under the Agency of Agricultural
Research and Development:- Research Institute for Freshwater Fisheries
(RIFF)- Gondol Research Station for Coastal Fisheries
(GRSCF)• universities
- Department of Fisheries, University of
Brawidjaja (UNBRA)- Faculty of Fisheries, Bogor Agricultural
University (BAU)• Agency for Assessment and Application of
Technology (AAAT)• institutions under the Directorate General of
Fisheries, e.g., Freshwater AquacultureDevelopment Center (FADC).
These institutions are members of the IndonesianNetwork of Fish Genetics Research and Development(INFIGRAD). Other members of the network includethe: Faculty of Fisheries, University of Riau;Department of Fisheries (DOF), University of BungHata; Faculty of Fisheries, University of Pajajaran;DOF, University of Gajah Mada; Faculty of Fisheries,University of Lambung Mangkurat; and Faculty ofFisheries, University of Hasanuddin. Table 2 lists thedifferent aspects of genetics research being carriedout by the institutions. Also, there is an ongoingresearch collaboration between the Central ResearchInstitute for Fisheries (CRIFI)/RIFF and theInternational Network on Genetics in Aquaculture/International Center for Living Aquatic ResourcesManagement (INGA/ICLARM) called GeneticImprovement of Common Carp, and between CRIFI/RIFF and the L’Institut Francais de RechercheScientifique pour le Developpement en Cooperation,called Characterization, Utilization and Maintenanceof Biological Diversity for the Diversification andSustainability of Catfish Culture in Southeast Asia.
Genetics Research
Research was conducted by 11 institutions (includingresearch collaborations) on five freshwater and fivebrackishwater fish species (Table 2).
Table 1. Production (t) of important freshwater culture species in Indonesia in 1996 (DGF 1997).
Species
Common carp (C. carpio)Java barb (Puntius gonionotus)Nilem carp (Osteochilus hasselti)Mozambique tilapia (O. mossambicus)Nile tilapia (O. niloticus)Other species
Total production
Type of culture
Pond
069.091019.753017.226016.174017.665043.009
182.918
Ricefield
074.782013.212000.381003.738002.062007.039
101.214
Cages
34.48900.22100.96901.11105.94101.900
44.631
Production
t
178.362033.186018.576021.023025.668051.948
328.763
%
054.3010.1005.6006.4007.8015.8
100.0

53
Carps
Documentation of the genetic resources of carps(Cyprinus carpio, Barbodes gonionotus,Osteochilus hasselti, Leptobarbus hoeveniand Neolissochilus thienemanni) in Indonesia
This activity was a collaboration between CRIFI/RIFFand INGA/ICLARM. Literature searches were madefrom 12 faculties or divisions of fisheries ofuniversities in West, Central and East Java and WestSumatra, and from related fisheries institutions, suchas the Institute for Evaluation and Application ofTechnology, Provincial Fishery Extension Services,Central Hatchery and RIFF. Questionnaires weredistributed to the Provincial and Regional FisheryExtension Services and Central Hatcheries in Westand Central Java. Common carp genetic resources,which focused on strains of Rajadanu, Wildan Cianjur,Sutisna Kuningan and Majalaya, were documented.Information was collected on the biology of species;status of genetic quality evaluation and utilization ofgenotypically different common carp.
Establishment of synthetic base population ofcommon carp from four strains
This study is being carried out by CRIFI/RIFF andINGA/ICLARM. At the first stage, complete diallelecrossing of three strains (Rajadanu, Wildan Cianjurand Majalaya) and one cross of Sutisna Kuningan wasmade, with a total of 10 crossings. Each strainconsisted of five females and five males. Crossing wasaccomplished by artificial fertilization, where eggs ofeach female were fertilized with sperm of five males.Simultaneous ovulation occurred after two injectionswith Ovaprim (combination of GnRH anddomperidone). Fertilized eggs were distributed tokakabans (Arenga fiber egg collectors) set in hapas. Theeggs of each female were hatched in fivehapas.Rearing of eight-day old larvae was carried outin fine-mesh (1-2 mm) net cages set in two ponds, inwhich 50 net cages of 2 x 1.2 x 1 m in dimensionwere set in each pond. The stocking density was1 300 larvae per cage. In one month, the larvae grewto fry with individual weight of about 1 g. The nextstep was rearing of fry to fingerling in net cages with
Aquaculture Genetics Research in Indonesia
Species
Freshwater
C. carpio
Clarias spp. (1)
C. batrachus (2)
Pangasius spp.
O. niloticus
M. rosenbergii
BrackishwaterC. chanos
Epinephelus spp.
Chromileptesaltivelis
Anguilla bicolor
Penaeus monodon
Table 2. Genetics research on freshwater and brackishwater fish species and the institutions involved in Indonesia.
Characterization
RAPD
Allozyme
Isozyme electrophoresis
Allozyme
Isozyme electrophoresis
Isozyme electrophoresis
Isozyme electrophoresis
RAPD
Isozyme electrophoresis
Gynogenesis
Punten strain
Majalaya, Sinyonya
Majalaya, Sinyonya,mirror carp
Selection
Combinedmultitrait
Heritability,family
Family
Hybridization
Diallele crossing
Institutions involved
AAAT
UNBRA and PuntenHatcheryFADC
BAU
CRIFI/RIFF and INGA/ICLARM
ORSTOM and RIFF
RIFF
ORSTOM and RIFF
FADC
RIFF
GRSCF
GRSCF
GRSCF
AAAT
GRSCF

Fish Genetics Research in Member Countries and Institutions of INGA54
larger mesh. The base population will be establishedthrough 150 tagged fingerlings per full-sib family.This will be conducted in the second year.
Data collected from each breeder were fecundity,egg diameter, hatching rate, larval size, fry size andsurvival rate.
Gynogenesis of Punten common carp
Punten common carp is a well-known strain inIndonesia. Its phenotype characteristics include bigbelly and relatively wide body. The original Puntenstrain was the result of selection done by Goossens in1928-1930 in Punten Hatchery, West Java (Ardiwinata1971). This strain is not popular with fish farmersand competes with the widely spread Majalaya strain.In view of this, the Punten strain has become rareand is difficult to find. Punten Hatchery, incollaboration with UNBRA in East Java, has tried toconserve and make gynogenesis of the existing strain.This effort produced gynogenetic Punten withdifferent color variation, red, green and blue, withthe ratio of standard length to body depth 1:2.4; 1:2.5-2.7; and 1:2.6-2.9, respectively (Hasanudin 1998).
Gynogenesis of Majalaya and Sinyonya strainsof common carp and their diallele crossing
The Majalaya strain is a big belly type one, with widebody depth and greyish-green scales. The Sinyonyastrain is elongated and yellow. The eyes of the youngfish are normal, while those of the old fish are coveredby the eyelid membranes. The second generationof mitotic-gynogenetic diploid was produced forboth strains. This diploid has a larger degree ofphenotypic variation than the normal fish(Sumantadinata et al. 1990).
Diallele crossing was made to obtain better results.Progeny produced by crossing male Majalaya andfemale Sinyonya showed better growth and survivalrates than the reciprocal and pure lines. The colorsof the progeny were 70% green; yellow, 20%; and inbetween both colors, 10% (Hadadi et al. 1998).
Qualitative and quantitative characters of threeC. carpio strains
The second generation of gynogenetic diploidcommon carp strains of Majalaya, Sinyonya andmirror carp were produced in 1992. Some fish of thesestrains were sex-reversed by oral administration of
17α-methyltestosterone. Females and the sex-reversedmales were matured in early 1994. The fish were grownin earthen pond and in running water system withintensive feeding. Characteristics of the young fish tomarket size were observed and analyzed .
Slight differences among strains were found in theegg size before and after ovulation and in their furtherdevelopment. Fecundity of Sinyonya was higher thanthat of the mirror and Majalaya strains, but theirfertilization and hatching rate were relatively similar.The growth rate of mirror and Sinyonya strains weresimilar under still or running water conditions, butthey were better than that of Majalaya strain. Thesurvival rate of mirror carp was the lowest. As regardsbody composition, the Sinyonya strain had the highestpercentage of fillet; Majalaya strain, biggest viscera;and mirror carp, fillet remnant. The total quantity ofmuscular spines of the Sinyonya carp was the lowest.Proximate analysis of flesh showed it had the highestprotein content and the lowest lipid content (Arfahand Sumantadinata 1998).
Application of RAPD-PCR method for analysesof DNA polymorphism of C. carpio
Six strains of C. carpio (Majalaya, Sinyonya, Punten,Domas, mirror and Merah [red]) were collected fromWanayasa Hatchery, fish farmer ponds and floatingnet cages. Blood samples were collected for extractionof total DNA, visualization of DNA by electrophoreticgel agaros and ethidium bromide staining, test andassessment of DNA quality, and reaction of PCR byRAPD method. The results showed no difference intypical band pattern for the four strains, exceptMajalaya and Sinyonya. For these two strains, DNApolymorphism can be detected with the applicationof RAPD-PCR (Faizal et al. 1998).
Catfish
Identification and characterization of Clariasand Pangasius sp.
Collaborative research involving six institutions hasbeen carried out, since November 1997. Theinstitutions were ORSTOM and Centre deCooperation Internationale en Recherche Agro-nomique pour le Developpement (France); KatholiekeUniversiteit Leuven (Belgium); Cantho University(Vietnam) and CRIFI/RIFF (Indonesia). The researchproject was entitled “Characterization, Utilization andMaintenance of Biological Diversity for the
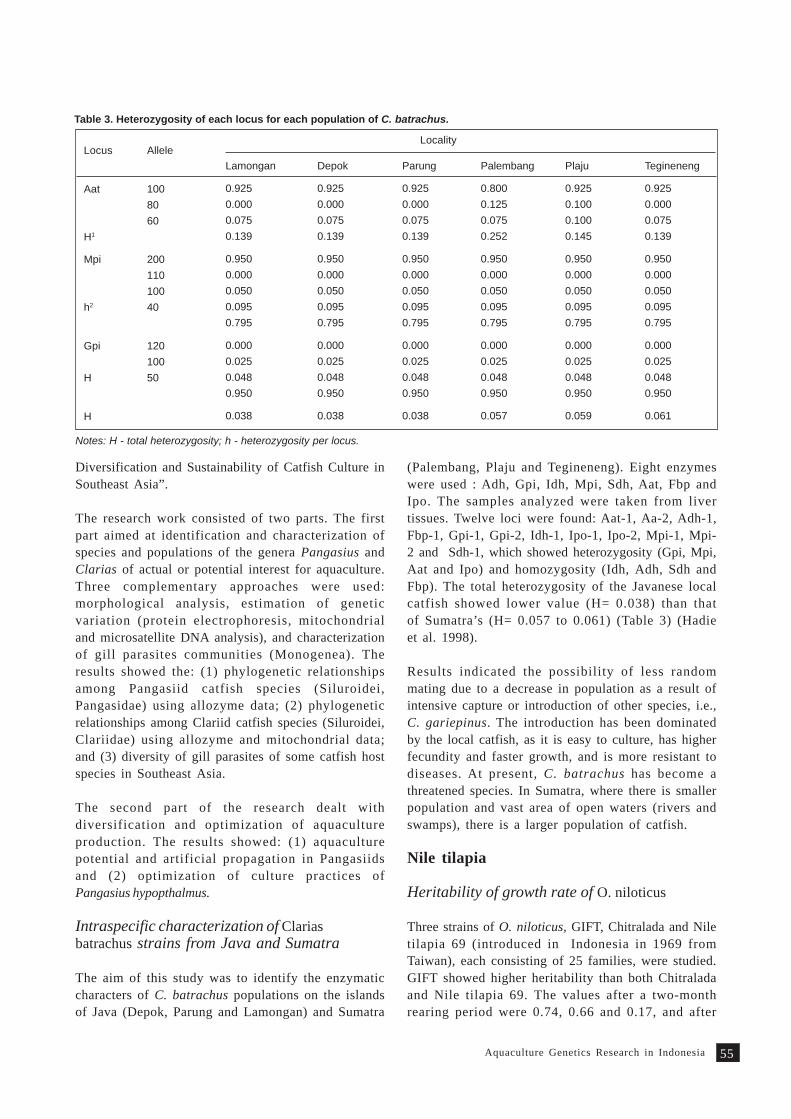
55
Diversification and Sustainability of Catfish Culture inSoutheast Asia”.
The research work consisted of two parts. The firstpart aimed at identification and characterization ofspecies and populations of the genera Pangasius andClarias of actual or potential interest for aquaculture.Three complementary approaches were used:morphological analysis, estimation of geneticvariation (protein electrophoresis, mitochondrialand microsatellite DNA analysis), and characterizationof gill parasites communities (Monogenea). Theresults showed the: (1) phylogenetic relationshipsamong Pangasiid catfish species (Siluroidei,Pangasidae) using allozyme data; (2) phylogeneticrelationships among Clariid catfish species (Siluroidei,Clariidae) using allozyme and mitochondrial data;and (3) diversity of gill parasites of some catfish hostspecies in Southeast Asia.
The second part of the research dealt withdiversification and optimization of aquacultureproduction. The results showed: (1) aquaculturepotential and artificial propagation in Pangasiidsand (2) optimization of culture practices ofPangasius hypopthalmus.
Intraspecific characterization of Clariasbatrachus strains from Java and Sumatra
The aim of this study was to identify the enzymaticcharacters of C. batrachus populations on the islandsof Java (Depok, Parung and Lamongan) and Sumatra
(Palembang, Plaju and Tegineneng). Eight enzymeswere used : Adh, Gpi, Idh, Mpi, Sdh, Aat, Fbp andIpo. The samples analyzed were taken from livertissues. Twelve loci were found: Aat-1, Aa-2, Adh-1,Fbp-1, Gpi-1, Gpi-2, Idh-1, Ipo-1, Ipo-2, Mpi-1, Mpi-2 and Sdh-1, which showed heterozygosity (Gpi, Mpi,Aat and Ipo) and homozygosity (Idh, Adh, Sdh andFbp). The total heterozygosity of the Javanese localcatfish showed lower value (H= 0.038) than thatof Sumatra’s (H= 0.057 to 0.061) (Table 3) (Hadieet al. 1998).
Results indicated the possibility of less randommating due to a decrease in population as a result ofintensive capture or introduction of other species, i.e.,C. gariepinus. The introduction has been dominatedby the local catfish, as it is easy to culture, has higherfecundity and faster growth, and is more resistant todiseases. At present, C. batrachus has become athreatened species. In Sumatra, where there is smallerpopulation and vast area of open waters (rivers andswamps), there is a larger population of catfish.
Nile tilapia
Heritability of growth rate of O. niloticus
Three strains of O. niloticus, GIFT, Chitralada and Niletilapia 69 (introduced in Indonesia in 1969 fromTaiwan), each consisting of 25 families, were studied.GIFT showed higher heritability than both Chitraladaand Nile tilapia 69. The values after a two-monthrearing period were 0.74, 0.66 and 0.17, and after
Aquaculture Genetics Research in Indonesia
Table 3. Heterozygosity of each locus for each population of C. batrachus.
Locus
Aat
H1
Mpi
h2
Gpi
H
H
Locality
Lamongan
0.9250.0000.0750.139
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.038
Allele
1008060
20011010040
12010050
Depok
0.9250.0000.0750.139
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.038
Parung
0.9250.0000.0750.139
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.038
Palembang
0.8000.1250.0750.252
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.057
Plaju
0.9250.1000.1000.145
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.059
Tegineneng
0.9250.0000.0750.139
0.9500.0000.0500.0950.795
0.0000.0250.0480.950
0.061
Notes: H - total heterozygosity; h - heterozygosity per locus.

Fish Genetics Research in Member Countries and Institutions of INGA56
nine months, 0.45, 0.37 and 0.05, respectively (Widiyatiet al. 1998).
Evaluation of diallele crossed tilapias
Diallele crossing of tilapias was carried out for thefollowing strains: (1) Chitralada (black) strain of O.niloticus and Philippine red tilapia; (2) GIFT strain ofO. niloticus (black) and red tilapia (NIFI – NationalInland Fisheries Institute, Thailand); (3) GIFT strainand Philippine red tilapia; (4) Nile tilapia 69 (black)and NIFI strain; (5) Nile tilapia 69 and Philippinered tilapia; (6) Chitralada strain and GIFT strain; (7)Chitralada strain and Nile tilapia 69; and (8) GIFTstrain and Nile tilapia 69.
The growth rate and color composition were observedfor 16 weeks. Crossing male GIFT strain with otherstrains showed better growth rate (117-123 g) thaneither crossing male Chitralada strain with otherstrains (89-100 g) or pure GIFT strain (average weight,98 g). The highest percentage of red tilapia (46%)was obtained from crossing male GIFT strain andfemale Philippine red tilapia, followed by yellow (42%)and black (11%) types (Hadadi et al. 1998).
Milkfish
Genetic variation of Chanos chanos samples (naturaland hatcheries F1 and F2) and performance of F1 andF2 were examined. Among 15 enzymes, 29 loci weredetected. Eleven were polymorphic, namely, Adh, Aat-1, Est-2, Gpd, Gpi-1, Gpi-2, Idh-1, Ldh-1, Mdh-1, 6-Pgd, Pgm-2. Reduction in genetic variability was foundin the hatchery stock (F2). The reductions were 50%in the number of polymorphic loci, 22.36% in thenumber of allele per locus and 37.78% in theheterozygosity. The actual number of broodstockcontributing in F2 was 14 out of 50 fish individuals inthe spawning tanks. GPI locus among the mostlypolymorphic loci could be used as a marker in geneticimprovement program. In order to avoid reductionin genetic variability in hatchery stocks, it is proposedto increase the number of broodstock for massproduction of milkfish fry (Sugama and Prijono 1998).
Grouper
The study of genetics and characteristics of groupersEpinephelus spp. and Chromileptes altivelis wasconducted by the Research Station of ResearchInstitute for Coastal Aquaculture in Gondol, Bali. Fourspecies of groupers (E. fuscogutatus, E. coioides,
E. microdon and E. bonthoides) and one species ofC. altivelis were analyzed by isozyme electrophoresisfor 13 enzymes, such as Adh, Aat, Est, Gpi, Idh, Ldh,Mdh, Me, 6-Pgd, Pgm, Sdh, Sod and Sp. Using TC-8 and CAPM-6 buffer systems combined with skeletalmuscle and liver tissues, six polymorphic loci weredetected in E. coioides and C. altivelis, four inE. fuscogutatus and E microdon and two in E. bonthoides.Genetic variability was highest in E. coioides, 1.78allele per locus and 0.89 heterozygosity. The shortestgenetic distance (D) was found between E. coioidesand E. bonthoides with D value of 0.0832. D valuesbetween C. altivelis and other species were significant(D±0.8). The results also suggest that Adh locus is areliable marker for species identification (Sugamaand Trijoko 1998).
Eel
RAPD-DNA method on Anguilla bicolor elver wasapplied. Elvers were collected from Cimandiri River(1-2 km from the river mouth) on south coast of WestJava. The samples of tissue were extracted for the totalDNA assessment of the concentration of DNA, test ofthe quality of DNA with restriction enzyme, RAPD-PCR. The results showed that RAPD-PCR method canbe applied to distinguish different populations ofelvers (Amarullah et al. 1998). The study wasconducted by AAAT.
Giant freshwater prawn
A study was conducted to evaluate response toselection on dress-out characteristics of the syntheticpopulation of Macrobrachium rosenbergii. The characterevaluated was the percentage proportion of carapaceto the standard body length. Base population wasestablished with three collections from Kalipucang,Tanjung Air (both located in West Java) and Musi(South Sumatra) through half-sib mating. Larval andpostlarval rearings were conducted in concrete tanksusing clear water system with temperature of 28-31°C.Growout was done in earthen ponds of 400 m2. TheF1 synthetic population of five months old showedaverage body weight of 23.56 ± 10.37; maximum andminimum body weight of 53.30 and 6.20 g,respectively; coefficient of variation, 44.21%;heritability, 0.56% (standard error 0.07); averagedress-out 56.63±3.39%; coefficient of variation, 6.0%(Emmawati et al. 1998). The average dressing washigher compared to the average of the character atfarmer level (45%) (Hadie et al. 1991). The selectionindicated the possibility of improving dress-out

57
percentage in M. rosenbergii. Response of selection,selection differential and intensity, and predictedaverage dress-out of F2 are presented in Table 4.
Shrimp
The association between heterozygosity at two lociof isocitrate dehydrogenase (Idh) and glucosephosphate isomerase (Gpi) and the development ratein P. monodon were examined. The purpose of thisstudy, which was conducted by GRSCF, was to test theprediction that heterozygous prawn develops fasterthan the homozygous one. There were significantdifferences in fecundity, hatching rate and vitalitybetween heterozygous and homozygous individuals.Larvae derived from heterozygous broodstockdeveloped faster than those from homozygous one.The results suggest that heterozygous individuals aresuperior to homozygous ones (Sugama et al.1998).
The study of genetic variation and populationstructure of P. monodon was another collaborativeresearch among GRSCF, Faculty of Agriculture, KochiUniversity, and Faculty of Fisheries, BAU. Fivepopulations of P. monodon in the coastal waters ofIndonesia (Aceh, Madura, Bali, West Nusa Tenggaraand South Sulawesi) were sampled electrophoreticallyfor evaluation of genetic variation at 21 loci. Six loci(Est-2, ±- Gpd, Gpi, Idh, Ldh-1 and Mdh-1) werepolymorphic in at least one of the samples. Allpolymorphic loci at all localities were in Hardy-Weinberg equilibrium. The genetic variability of thespecies, as indicated by number of alleles per locus(Na) and heterozygosity (H), was low, with average(Na=1.428), ranging from 1.333 to 1.523 andH=0.034, ranging from 0.023 to 0.047. The meangenetic distance between population pairs was0.00045, ranging from 0.00004 to 0.00112. Clusteringsamples according to their paired genetic distancesshowed that the population of P. monodon consisted ofthree geographical groups. However, the absoluteamount of genetic variation among populationssampled appeared to be very low (Sugama et al. 1998).
The GIFT strain O. niloticus was introduced inIndonesia in 1994 as part of the INGA program. Thefry were studied in three locations, RIFF at Bogor,FADC at Sukabumi and Cangkringan Hatchery atYogyakarta. Studies on growth rate, conducted inthree locations in West Java, showed that GIFTperforms better compared to O. niloticus strains alreadyexisting in Indonesia.
The economic crisis during 1998 had a strong impacton intensive fish culture, primarily because the highprice of pelleted feed, which increased by 70% (fromRp 1 400 to Rp 2 400 or from US$0.15 to US$0.30).However, the number of pellet feed industriesalso increased; trademarks increased from four to12, with different qualities. The quality of some feedsis poor resulting in low feed conversion. Efforts weremade for the intensive fish culture to survive andbecome profitable through the use of fish speciesthat are easy to handle and have fast growth rate,good marketability, and efficient feed conversion.GIFT strain has met these requirements particularlywith regard to rearing in floating net cages inSaguling, Cirata and Jatiluhur reservoirs in theprovince of West Java.
On the other hand, the use of C. carpio, the lead speciescultured in cages, has decreased dramatically; atpresent, it is estimated to be less than 10%. A total of25 560 cages (7 x 7 x 3 m3) need a large supply ofGIFT fry. The high demand is due to the shift fromC. carpio carp fry to GIFT production, and theestablishment of large-scale GIFT hatcheries by newinvestors.
In single-bottom floating net cages, only GIFT fishor C. carpio are stocked, while in double-bottom cages,C. carpio are stocked in the inner cage and GIFT fishin the outer cage. Feeds are applied only to C. carpio,while GIFT fish are fed food droppings from the innerlayer, in addition to periphyton growing on net walls.The rearing period varies from 60 to 110 days. InJatiluhur reservoir, the stocking densities per cage insingle-bottom net cages varied from 60 to 130 kg ofC. carpio or 100 to 150 kg of O. niloticus . Theproduction was from 875 to 1 940 kg of C. carpio andfrom 626 to 1 200 kg of O. niloticus per cage. In double-bottom cages, stocking densities of C. carpio were50-60 kg in inner cages and 18-50 kg of O. niloticus inlower cages. The production was 1 080-1 326 kg and
Table 4. Response, selection differential, intensity, anddress-out of M. rosenbergii.
Parameter
Parental response, F1 (%)Selection differentialSelection intensityAverage dress-out (%)Prediction of average dress-out (%)
Value
07.6913.7404.0556.6364.32
Aquaculture Genetics Research in Indonesia
O. niloticus (GIFT strain)culture in Indonesia

Fish Genetics Research in Member Countries and Institutions of INGA58
160-313 kg, respectively (Krismono 1998). The fish areusually harvested at the individual size of250-300 g with the price of GIFT strain at the producerlevel of Rp 3 500 (US$0.44) and for individual fishsize over 500g the price was Rp 4 500 (US$0.56).Production of C. carpio and O. niloticus in floating netcages in Cirata reservoirs is shown in Table 5.
Brackishwater ponds, with a maximum salinity of 15ppt, is another source for culture of GIFT strain. InKarawang area (West Java), such culture has beencarried out since 1997. There are 130 units of pondsof 4 000 m2 each that can produce 1.7-2.0 t/pond in110-day rearing period with a stocking density of 10000 fingerlings (3-5 cm). Fish are harvested at theaverage individual size of 200 g, and the survival rateis about 80-85%. A trial in ponds of 3 000 m2 each,with a stocking density of 75 000 fry of 1 g /fishproduced fingerlings of 15-20 g/fish with a survivalrate of 80% in 110-day period. To maintain the geneticquality of GIFT strain, broodstock managementshould be done by seed producers, with assistancefrom authorities and the scientific community.
Future Research
Following are various future research collaborationson genetic research in Indonesia:• CRIFI/RIFF and INGA/ICLARM - genetic
improvement of carp in Asia: establishment ofbase population of the second ten crossingsamong strains of Rajadanu, Wildan Cianjur,Majalaya and Sutisna Kuningan. During 2000/
2001, the project will continue to undertakeestimation of the breeding value of 100 half-siband 200 full-sib families from the best cross ofthe first generation.
• CRIFI/RIFF and ORSTOM - characterization,utilization and maintenance of biological diversityfor the diversification and sustainability of catfishculture in Southeast Asia. For geneticcharacterization of Clariidae and Pangasidae,more samples will be collected from rivers in Java,Sumatra and Kalimantan, and analyzed thorughprotein electrophoresis and mtDNA.
• INFIGRAD (Fig. 1):- enzymatic characterization of C. carpio – to be
conducted by RIFF for four strains of C. carpio,i.e., Majalaya, Rajadanu, Sutisna Kuninganand Wildan Cianjur;
- DNA polymorphism analysis for carp species– to be carried out by AAAT;
- hybridization of C. carpio strains – to be done
Table 5. Production of C. carpio and O. niloticus in floating netcages in Cirata reservoirs.1
YEAR
199619971998
Number of cages (unit)
Available
10 95025 55817 477
Operating
8 70015 28910 485
C. carpio
15 89636 532 9 349
Production (t)
O. niloticus
9 2182
12 6393
5 9164
Total
25 11449 17115 265
1Data collected from Fish Culture Station of Saguling-Ciratareservoirs.2Consisted of Nile tilapia 69.3Mostly Nile tilapia 69.4Mostly GIFT strain.
Fig. 1. Distribution of members of Indonesia Network of Fish Genetic Reseach and Development (INFIGRAD)

59
by FADC at Sukabumi;- evaluation of heterosis of Pangasius hybrids
(P. jambal and P. nasutus) – to be conducted byRIFF at Sukamandi;
- family selection of the sixth generation ofGIFT strain of O. niloticus – to be conductedby RIFF, Sukamandi;
- study on the application of RNA:DNA ratio toevaluate growth of tilapia – to be done byAAAT, Serpong;
- hybridization of tilapia – to be done by FADC,Sukabumi;
- morphological and DNA characterization ofP. monodon - to be carried out by GRSCF andFaculty of Fisheries, BAU; and
- studies on allozyme heterozygosity and growthrate of C. chanos fry – to be done by GRSCF.
References
Amrullah, M.H., I. Faizal and I.T. Makagiansar. 1998. Theapplication of RAPD technique for identification of DNApolymorphism on elver Anguilla bicolor. Paper presented atthe INFIGRAD Meeting, 8 February 1998, CRIFI, Jakarta,Indonesia. (In Indonesian).
Ardiwinata, R.O. 1971. Cultivation of common carp (tambera).Sumur Bandung, Indonesia. (In Indonesian).
Arfah, H. and K. Sumantadinata. 1998. Qualitative andquantitative characters of three common carp (Cyprinus carpio)strains in Indonesia. Department of Aquaculture, Faculty ofFisheries, Bogor Agricultural University, Indonesia.
DCFA (Development Centre for Freshwater Aquaculture). 1998.Progress report of research result 1997/1998. DirectorateGeneral of Fisheries, Indonesia. (In Indonesian).
Emmawati, L., W. Hadie, I. Iriana and Jaelani. 1998. Familyselection of synthetic population to increase genetic variationof freshwater giant prawn (Macrobrachium rosenbergii). Res.Prog. Rep. RIFF, Indonesia. (In Indonesian).
Faizal, I., D. Irawan, R.S. Aliah, I.T. Makagiansar and M.H.Amarullah.1998. Preliminary study on polymorphism DNAanalyses of common carp using RAPD-PCR. Paper presentedat the INFIGRAD Meeting, 8 February 1998, CRIFI, Jakarta,Indonesia. (In Indonesian).
Hadadi, A., J. Trenggana, J. Rasyidik, D. Suganda, C. Muharam,A. Kusuma and A. Santika. 1998. The application of commoncarp broodstock pure line and seed quality improvement.Proj. Rep. FADC, Directorate General of Fisheries, Indonesia.(In Indonesian).
Hadie, W., Sudarto, Jaelani and L. Emmawati. 1998.Characterization of local catfish Clarias batrachus (islands ofJava and Sumatra) with intraspecific analyses. Res. Prog. Rep.RIFF, Indonesia. (In Indonesian).
Hadie, L.E., M.F. Sukadi and W. Hadie. 1991. Selection of giantfreshwater prawn (Macrobrachium rosenbergii) to increase theproduction performance in three different collections. RIFFReport to IDRC Canada, p. 71-90.
Hasanudin, I.1998 Gynogenesis and cryopreservation to improvethe quality of broodstock and fry of common carp. PuntenHatchery, Indonesia. (In Indonesian).
Krismono. 1998. Fish farming in floating net cage using geneticallyimproved fish. RIFF, Indonesia. (In Indonesian).
Nugroho, E and N.A. Wahyudi. 1991. Selection of commoncarp races collected from some Indonesian region using Z-score. Bul. Penel. Perik. Darat 10(2): 49-54 (in Indonesian)
Sugama, K. and A. Prijono. 1998. Genetic variability andperformance of F1 and F2 in milkfish Chanos chanos. GondolRSCF Report. (in Indonesian).
Sugama, K. and Trijoko. 1998. Genetic and characteristics ofgroupers Epinephelus spp. and Chromileptes altivelis. GondolRSCF Report (in Indonesian).
Sugama, K., Haryanti, S. Tsumura, N. Taniguchi and K.Sumantadinata. 1998. Genetic variation and populationstructure of tiger shrimp Penaeus monodon in Indonesia.Zuriat, 9(1): 25:32.
Sumantadinata K, N. Taniguchi and Sugiarto. 1990. Increasedvariance of quantitative characters in the two types ofgynogenetic diploids of Indonesian common carp. NipponSuisan Gakkaishi, Japan.
Taniguchi, N., K. Sumantadinata and K. Sugama. 1992. Geneticassessments of common carp stocks in Indonesia. Indon. J.Trop. Agric.3(2): 91-96.
Widiyati, A., L. Emmawati and T. Kadarini. 1998. Heritabilitiesand response of family selection on growth of N ile tilapia.RIFF, Indonesia. (In Indonesian).
Aquaculture Genetics Research in Indonesia

Fish Genetics Research in Member Countries and Institutions of INGA60

61
ABSTRACT
Aquaculture genetics research is carried out at the National Aquaculture Centre at Domasi, Malawi, by staff of theUniversity of Malawi (UM) and the Department of Fisheries. The main species cultured are from genera Tilapia andOreochromis. The genus Oreochromis was domesticated without proper identification of the various speciesespecially in the subgenus Nyasalapia where three species (O. karongae, O. squamipinnis, and O. lidole) aredifficult to identify before they reach breeding size. It is, however, observed that local fishers can identify thesespecies using morphological characters. There have been no attempts to carry out deliberate domestication selectionin the small-scale farms with the prevailing practices probably leading to deterioration of stock performance anddecline in genetic diversity. Studies showed that wild populations grow faster than domesticated populations, andresults of mass selection on collimated individuals suggest that selected individuals grow faster than unselectedindividuals. Studies of population genetics of domesticated and wild tilapias are underway at UM.
AQUACULTUREGENETICS RESEARCH
IN MALAWI
A. AmbaliUniversity of Malawi
Department of Biology, Chancellor CollegeBox 280, Zomba, Malawi
AMBALI, A. 2001. Aquaculture genetics research in Malawi, p. 61-64. In M.V. Gupta and B.O. Acosta (eds.) Fish geneticsresearch in member countries and institutions of the International Network on Genetics in Aquaculture. ICLARMConf. Proc. 64, 179 p.
Introduction
The aquaculture industry in Malawi is characterizedby small-scale farmers, and tilapia is the major fishspecies cultured. The Government has put a banon importation of exotic species, which have beendomesticated and selected, to fairly greater extentthan the indigenous species. The ban is intended toprotect the genetic integrity of fish populations in thenatural water bodies.
Although aquaculture has developed to considerableextent in Malawi, compared to other African countries,it is still faced with the problem of culturing species
Aquaculture Genetics Research in Malawi
with low growth rate. This, among other factors,is due to poor domestication protocols in thegovernment and private farms. There is little regardfor quality of germplasm being cultured. Small-scalefarmers and government stations do not practicedeliberate and effective genetic selection. They reservea portion of fingerlings from grow-out ponds at thetime of harvest for the next grow-out cycle. Althoughno criteria are known to be used in selecting thefingerlings, strong indirect or domestication selectioncan nevertheless occur, e.g., if the fingerlings comefrom very few parents or from particularly early orlate maturing parental stock. Not much is known atpresent if such a system depletes genetic variation or

Fish Genetics Research in Member Countries and Institutions of INGA62
not, but DNA analysis of farm populations indicatesthat depletion is actually rather rapid.
The species cultured have not been properlyidentified or characterized. As a result, informationhas not been reproducible, consistent or easilytransferable among programs. The domesticationefforts of the Nyasalapia species have been carriedout without sorting out the confusion surroundingthe identity of the various species of Oreochromis(Nyasalapia) sp. Lack of documentation of theavailable genetic material is due not merely to lackof facilities and trained personnel to identify thespecies, but also to lack of knowledge of the structureof the gene pool of species relevant to aquaculture.
The increasing pressure from human populationgrowth and activities associated with the lakes onaquatic biodiversity in Malawi is of particular concern.Cyprinids, like Opsaridium microleps and Labeo mesops,have disappeared from their traditional naturalhabitats where they used to occur in abundance.O. (Nyasalapia) sp. catches have declined considerablyin the wild. The country is currently faced with thechallenge of conserving the aquatic biodiversity inthe lakes. Ex situ conservation is not economicallyfeasible in such a country where financial resourcesare limited; the economic feasibility of in situconservation offers achievable promise as analternative means of preserving the otherwisethreatened biodiversity. Several reservoirs exist in thecountry, but most of them are underutilized for fisheryproduction.
Tilapia Genetic Resources in Malawi
In Malawi, the genus Tilapia is represented byT. rendalli while the genus Oreochromis is divided intotwo subgenera: O. Oreochromis and O. Nyasalapia.Oreochromis is composed of O. shiranus sp. (subdividedinto O. shiranus shiranus and O. shiranus chilwae),O. mossambicus and O. placidus. The O. (Nyasalapia)species flock is composed of O. Ny. karongae,O. Ny. lidole, O. Ny. squamipinnis and O. Ny. saka. Thetaxonomy of O. Nyasalapia sp. is based almost entirelyon morphometric data, some of which are extremelypreliminary. Contrary to the species division listedabove, multivariate morphometric analysis carriedout recently suggests that there are only three speciesof O. (Nyasalapia), i.e., O. karongae, O. lidole andO. squamipinnis. O. saka is classified as a juniorsynonym of O. karongae (Turner and Robinson 1990).Difficulties in identifying the O. Nyasalapia species
have deterred proper broodstock handlingprocedures and development of improved strains foraquaculture.
Affinities
The O. Nyasalapia species flock appearsmorphologically to be the derivatives of S. galilaeusgroup (Fryer and Iles 1972) and they have affinitieswith O. machrochir and O. rukwaensis. Thys vanAudernaerde (1968, cited in Sodsuk et al. 1995)proposed a separate subgenus Nyasalapia as partof the genus Tilapia which would be only restrictedto the Lake Malawi species and subgenus Lowuwialato include all other tasseled male species, for instanceO. (Ny.) machrochir.
O. shiranus sp. is divided into two subspecies basedon morphological and meristic characteristics(Trewavas 1983), O. sh. chilwae and O. sh. shiranus. Thesubspecies are from a common ancestor O. shiranuswhich existed in Rovuma River into which the ancientLakes Chilwa-Chiuta drained to the Indian Oceanvia Lujenda River. The most nearly related species isO. rovumae, and together with O. mossambicus andO. placidus, they belong to the mossambicus group oftilapias (Fryer and Iles 1972).
Zoogeographical distribution
The O. Nyasalapia sp. is endemic to Lakes Malawiand Malombe only. The fishes are distributedthroughout Lake Malawi from north to south. Theirpopulation in the lake is reported to be declining. O.shiranus and T. rendalli are the most widely distributedtilapias in the country. They are found in Lake Malawiand Malombe, lagoons and rivers. O. mossambicus andO. placidus are only found in the Lower Shire (Table 1).
In aquaculture, the species have been moved withoutregard to their natural geographical distribution. TheO. Nyasalapia subgenus has been stocked in reservoirsand fishponds outside their natural regions ofdistribution.
Indigenous knowledge on characterizationof tilapias
It has been observed that fishers at the lakes andalong the Shire River have their own methods ofdistinguishing among fish species based onmorphometric characters and fish behavior. Thisknowledge has been passed on from several

63
generations but has not been well documented. Anationwide survey of indigenous knowledge ofthe fishing communities on the taxonomy andbehavior of tilapias has been completed and dataare being analyzed.
Also, a survey of local names of tilapia species wascarried out throughout the country. The main interestwas in determining the local classification system forspecies in the O. Nyasalapia subgenus which aredifficult to identify before breeding stages. Thesespecies are as follows:• O. squamipinnis - deep body and sharp bend on
the forehead; breeding males have characteristicvertical blue band on the forehead;
• O. karongae - deep body with depression on theforehead; males do not have vertical bands onthe forehead as in O. squamipinnis; and
• O. lidole - big head and small body; generallyslender in appearance.
Quantitative Genetics
Comparison of growth between wild anddomesticated populations
Wild broodstock populations of O. shiranus chilwa werecollected from Lakes Chilwa and Chiuta for
domestication. F1 progenies were compared topopulations that have been at the NationalAquaculture Centre (NAC) for several years. Theexperiment was carried out between August 1997 andApril 1998. The results obtained in 1998 indicatedthat wild population grew faster than domesticatedone, 16.6 g and 13.3 g, respectively (Table 2).
Mass selection on collimated populations
Broodstock of O. shiranus were collected fromdomesticated populations at the NAC. These werestocked in three ponds where they bred naturally.Fingerlings were collected and stocked randomlyinto 16 200 m2 ponds at stocking rate of 5 fish m-2.After one month, standard length (SL) measurements
Aquaculture Genetics Research in Malawi
Table 1. Distribution of tilapia in the water bodies of Malawi.
Water bodies Species
Lake Malawi, Lake Malombe and Upper Shire O. shiranus shiranusO. Nyasalapia species flockT. rendalli
Middle Shire O. shiranus shiranusT. rendalli
Lower Shire O. mossambicusO. placidusT. rendalli
Reservoirs O. shiranus hybrids and pure strainsO. Nyasalapia sp.O. mossambicus/O. placidus hybrids
Rivers and lagoons O. shiranus sp.T. rendalli
Aquaculture O. Nyasalapia sp.(distributed in all regions of the country but only recentlydomesticated)
O. shiranus shiranus (common in all regions)O. shiranus chilwae (common in southern and central regions)O. mossambicus (common at Kasinthula Fish Farm, the species hashybridized with O. shiranus in Zomba district)
O. placidus (rare, mainly hybridized with O. mossambicus)T. rendalli (common in all regions)
Table 2. Growth comparison between wild and domesticatedpopulations of O. shiranus.
Month
AugOctNovJanMarApr
Wild fish
05.3 ± 0.0006.2 ± 0.0010.0 ± 0.0612.2 ± 0.0316.5 ± 0.0321.9 ± 0.03
Mean weight (g)
Domesticated fish
03.8 ± 0.0004.9 ± 0.0106.9 ± 0.0309.7 ± 0.0612.3 ± 0.0317.4 ± 0.05

Fish Genetics Research in Member Countries and Institutions of INGA64
were taken from fish in eight of the 16 ponds. Alllength measurements were converted to weight (W)using the formula, W = 2.46x10-2SL2.79. Individualsof the weight range 7.3-14.1 g were selected andrestocked at stocking rate of 800 fish per pond. Thefish were grown for another 3 months before SLmeasurements were taken. The average length was17.2 g and range was 11.4-30.4 g. Fish of the range20.7-30.4 g were selected for grow-out for 6 months,August 1998-January 1999. The mean size of selectedpopulation was 24.7 g and selection differential was7.2 g. Stocking rate was 300 fish per pond. The restof the fish were discarded. Length measurementswere taken monthly.
The second group formed a control population whichwas stocked in eight ponds at 5 fish m-2 and thenrandomly trimmed to 800 fish per pond after a month.Two months later, the stocking density was reducedto 300 fish per pond and grown for 6 months, fromAugust 1998 to January 1999, to compare the growthbetween selected and unselected populations. Thestocking size range in this control population was 4.9-37.1 g and mean was 23.1 g.
In both populations, fish were fed 10% body weightwith maize bran; chicken manure purchased fromnearby chicken farms was applied to the ponds topromote primary productivity.
Selected populations grew faster than unselected ones(Table 3). Overall, the population had very low growthrate, probably supporting the fact that domesticatedtilapias in Malawi have poor growth rate than theirwild counterparts.
Population genetics
Studies of genetic characterization and populationstructure of O. shiranus sp. have been carried out atUM in collaboration with Dalhousie University ofCanada with funding from the International
Development Research Centre. In these studies,microsatellite DNA markers were used to determinethe level of genetic diversity in wild and domesticatedpopulations. Considerable decline in genetic diversitywas reported in populations which were domesticatedfor longer period (Ambali 1996; Ambali et al. 1999).
A DNA analysis laboratory has been established atChancellor College, UM, with funding from theJapanese International Cooperation Agency. Activitiesin the laboratory include characterization ofindigenous tilapias and cichlid species with potentialfor aquaculture.
References
Ambali, A.J.D. 1996. The relationship between domestication andgenetic biodiversity of mouthbrooding tilapia species inMalawi: Oreochromis shiranus shiranus (Boulenger) andOreochromis shiranus chilwae (Trewavas). Department ofBiology, Dalhousie University. 216 p. Ph.D. thesis.
Ambali, A.J.D., R.W. Doyle and D.I. Cook. 1999. Genetic changesin Oreochromis shiranus (Trewavas) associated with the earlystages of national aquaculture development in Malawi.Aquacult. Res. 30: 1-10.
Fryer, G., and T.D. Iles. 1972. The cichlid fishes of the Great Lakesof Africa: their biology and evolution. Oliver and Boyd,Edinburgh. 641 p.
Sodsuk, P.K., B.J. McAndrew and G.F. Turner. 1995. Evolutionaryrelationships of the Lake Malawi Oreochromis species: evidencefrom allozymes. J. Fish Biol. 47:321-333.
Trewavas, E. 1983. Tilapiine fishes of the genera Sarotherodon,Oreochromis and Danakilia. Natural History, BritishMuseum, London.
Turner, G. F., and R. L. Robinson. 1990. Ecology, morphologyand taxonomy of the Lake Malawi Oreochromis (Nyasalapia)species. Ann. Mus. Roy. Afr. Centr. Sci. Zool. 262: 243-28.
Table 3. Growth comparison between selected andunselected populations of O. shiranus.
Month
AugNovDecJanFebMar
Unselected fish
23.12 ± 0.0423.46 ± 0.0025.77 ± 0.0029.00 ± 0.0129.40 ± 0.0131.02 ± 0.00
Mean weight (g)
Selected fish
24.69 ± 0.0234.06 ± 0.0337.68 ± 0.0439.02 ± 0.0439.02 ± 0.0342.04 ± 0.00

65
ABSTRACT
The application of genetics in aquaculture in Malaysia and the realization of the need for conservationand development of genetic resources are very recent. Many of the ongoing projects on these have yet to obtainresults that are therefore still unpublished. This paper discusses briefly some relevant work on documentation ofgenetic resources, evaluation of resources, quantitative genetics and biotechnological developments. Research primarilycarried out by the University of Malaya (UM) is presented in this paper. Research carried out by other projects iseither unpublished or unavailable at the time of writing this paper.
GENETICSFOR IMPROVEMENT
OF FISH IN MALAYSIA
T.K. MukherjeeInstitute of Biological Sciences
University of Malaya50603 Kuala Lumpur, Malaysia
MUKHERJEE, T.K. 2001. Genetics for improvement of fish in Malaysia, p. 65-70. In M.V. Gupta and B.O. Acosta (eds.) Fishgenetics research in member countries and institutions of the International Network on Genetics in Aquaculture.ICLARM Conf. Proc. 64, 179 p.
Genetics for Improvement of Fish in Malaysia
Introduction
In the National Agriculture Policy (January 1999)announced by the Minister of Agriculture, Governmentof Malaysia, the need for sustainable development offisheries industry was well emphasized.
The demand for fish increases with a growingpopulation. According to Mazlan (1997), Malaysia,with an average per capita fish consumption of 35 kg,requires approximately 796 100 t of fish by the end ofthis century. To meet this demand, integrated planningfor land resource management, protection of aquaticenvironment and development of an efficient fishindustry are vital steps.
The development of a modern industry, as envisagedby the government, requires the establishment ofsound breeding programs for important food species
and this is usually accompanied by an expansion ofprograms for genetics research. This is the focus ofthis paper. Ornamental fish breeding is equallyimportant though in the developing market, bothwithin and outside the country.
Genetic diversity of freshwater fish has beendocumented. The dominant commercial species arecarps, catfish and tilapia. Carps and catfish have alsothe most diverse species. There are nearly 300 fishspecies in Malaysia, the conservation status of whichare not exactly known. The IUCN Red Data Book listsonly two species (Scleropages formosus and Probarbusjullieni) from Malaysia as endangered. However, thereare at least seven other species which may becategorized as rare or endangered (Table 1). Others
Conservation and ConservationGenetics

Fish Genetics Research in Member Countries and Institutions of INGA66
(e.g., Asian Wetland Bureau, Kuala Lumpur) believethat at least 17 species are rare. ICLARM alsodeveloped a global database, which has informationon Malaysian species, but the exact status of geneticresources is insufficiently known and can only beascertained through research on documentation andevaluation. For the rare species, methods ofconservation need to be developed, which includestock identification, monitoring population size andlive conservation or cryopreservation.
While the Department of Fisheries (DOF) hasextensive programs for turtle conservation in the EastCoast of Malaysia (Ibrahim 1998, pers. comm.),efforts have been initiated in cryopreservation ofgametes and embryos for a few endemic species andmarine finfish and in in-vitro culture of Oreochromisspp. (pers. comm.: Ali 1999, Khadijah 1999 andNorhanizan 1999).
A collaborative research on genetic identification ofvarious strains of Oreochromis spp., carried out byQueensland University of Technology, University ofMalaya and Universiti Putra Malaysia (UPM), willpave the way for future conservation genetics workin Malaysia.
Electrophoresis of 32 protein loci, 18 of which arepolymorphic, in seven tilapia strains (O. mossambicus;O. niloticus, Chitralada strain; O. niloticus, Philippinesstrain; O. niloticus, Israel strain; O. niloticus, localstrain; O. aureus, Singapore strain; and a Taiwanesehybrid strain) showed very close genetic relationshipamong the strains (Selvaraj et al. 1994).
Genetic similarity or distance index may be useful inconservation work. Only one of a set of strains maybe conserved if they are genetically similar in termsof performance. Laboratories at UPM are nowengaged in similar studies using molecular markers,
instead of protein markers (pers. comm.: Selvaraj1999 and Tan 1999). Preliminary results are presentedbelow, under “Molecular genetics” portion.
Genetics and Breeding
Genetics research and its application in Malaysia arestill conspicuously inadequate. A survey of governmentand private breeding farms and hatcheries showed thatmost seed stocks for finfish production are imported(Chew and Mukherjee 1988).
Currently, species produced locally in hatcheriesinclude shrimp, tilapia, Javanese carp, bighead carp,common carp, local catfish and seabass. For cultureof cockles, mussels, grouper and snapper, seeds arealways collected from the wild. Also, imports ofexotic carps, especially grass carp, red tilapia, catfish,seabass and red snapper seedlings continue. Onlyseabass hatchlings and tiger prawn fry are exported(DOF 1996).
Research
Hatcheries have almost no research anddocumentation is scanty. Whatever little research hasbeen done in genetics and breeding during the 1980sand 1990s is presented below.
Electrophoretic markers
There have been various studies on electrophoreticmarkers. Genetic relationships have been studied onOreochromis spp. (Tan et al. 1989; Selvaraj et al. 1994);Trichogaster pectoralis (Tan et al. 1980); andMacrobrachium rosenbergii (Patimah, pers. comm.).Temporal variation (Yao 1980; Hasnah 1982;Surinderpal 1984; and Chew 1986), genetic controlof isozymes (Tan et al. 1980; Patimah et al. 1989)and discriminating loci between Penaeus monodon andP. merguiensis (Daud et al. 1995) have been studied.
Karyotyping
There were several contributions in the creation of adatabase of karyograms of locally available fish(Bongso et al. 1980; Jamuna 1989; Das 1990). Workby Das (1990) illustrates the qualitative andquantitative variations between carp and tilapiaspecies with respect to macrochromosomes. Jamuna’s(1980) work relates the differences among ornamentalspecies (guppy and koi).
Table 1. Some rare (R) or endangered and extinct (E)species in Malaysia.
Species
Balantiocheilos melanopterusDischerondontus haleiLuciosoma trinemaParachela maculicaudaProbarbus jullieniPhallostethus dunckeriScleropages formosusHelicophagus wandersiiCeratoglanis scleronema
Status
RRRREEEEE

67
Chromosome engineering
Induction of polyploidy in walking catfish, Clariasbatrachus, by controlling water temperature wassuccessful (Siraj et al. 1992). Growth performances andgonadal development of diploid and triploid werecompared with a 30% protein feed. While the growthpattern was not significantly different (P>0.05),gonads of male and female triploids were poorlydeveloped compared to those of diploid.
Siraj et al. (1993) achieved 67% gynogenetic fry inLampan Jawa Barbodes gonionotus, using ultraviolet(UV)-irradiated sperm of P. schwanenfeldii followed bycold shock at 2oC.
Triploidy induction in the hybrid catfish (female Clariasgariepinus x male C. macrocephalus) has been successfulusing cold shock (Mohidin 1995). Production ofgynogenetic female of C. macrocephalus with UVradiation and subsequently 92% sex reversal ofgynogenetic females treated with oral administrationof several dose levels of synthetic androgen (10, 50and 100 mg/kg of feed) were attempted by Mohidin(1995). The presence of hermaphrodites, as expressedby both testicular and ovarian tissues, proved that thetreatments did not result in total sex reversal.
Genetic evaluation
Genetic evaluation of six random-mating, parentaltilapia strains in Malaysia (O. niloticus, Thai strain; O.aureus, Singapore strain; O. niloticus, Israel strain; O.mossambicus, Thai strain; and O. niloticus, local strain)for three generations in integrated (with ducks) andnonintegrated ponds showed consistent superiority ofO. niloticus, Thai strain, in terms of body weight atdifferent ages and carcass composition at 24 weeks.Performance in integrated ponds was better than thatin nonintegrated ones (ACIAR 1997).
Hybridization
During the 1950s and 1960s, the Fisheries ResearchInstitute at Batu Berendam, was involved incrossbreeding of various tilapia species. Performanceof crossbreds, in general, was found to be better thantheir corresponding parental strains. Hickling’s (1960)study first showed the possibility of all male hybridsby interspecific hybridization. The variation in sex ratioof progeny in different interspecific crosses from theusual 1:1 ratio was later explained by suggesting that
S. mossambica (O. mossambicus) has a homogameticfemale and a heterogametic male while the reverseis true in the case of S. honorum (Chen 1969).However, this could not totally explain the resultsof subsequent crosses.
Intraspecific and interspecific reciprocal crossesinvolving O. aureus and O. niloticus suggested significantincrease in growth and fecundity of hybrids comparedto those of the parental species. Significant reciprocaldifference (O. niloticus males x O. aureus females vs. O.aureus males x O. niloticus females) in terms of the abovetwo characters and the departure from 1:1 sex ratiosuggested that the O. aureus strain may havehomogametic males (Mukherjee and Geeta 1992).Results of the chromosome study of Das (1990)partially confirmed this.
Phenotypic selection of families of O. niloticusPhilippine strain, and of O. aureus that produce moremales on crossing, and subsequent mating betweenthese families produced 67% male progeny and 33%female progeny. Similar selection for anothergeneration did not improve further the sex ratio infavor of males (Table 2). Therefore, it was not possibleto produce all male progeny which was originallyplanned in a three-year experiment.
Geeta (1995), using four populations from Universityof Malaya (two populations of O. niloticus and one eachof O. aureus and O. mossambicus), performed a reciprocaldiallele cross to estimate general combining ability(GCA), specific combining ability (SCA) and reciprocaleffects for such traits as growth, fecundity, hatchability,fry survival, fry growth and various carcasscharacteristics. The following conclusions were drawnfrom her study:
1. Body weight at different ages had significantGCA which had greater influence than SCA.
2. Survival rate showed significant reciprocaleffects, mainly when O. niloticus females wereused in the reciprocal crosses.
The above study also showed VA (additive geneticvariance) for body weight was much higher than VD
(dominance genetic variance) and thereforeintrapopulation selection may be effective in tilapiapopulations.
In another experiment with tilapia, two pure strainsof O. niloticus obtained directly from a breeding farmin Taiwan were crossed (Khadijah 1999, pers. comm.).Mean birth weight and standard length at 24 weeks of
Genetics for Improvement of Fish in Malaysia

Fish Genetics Research in Member Countries and Institutions of INGA68
two pure strains, their F1 and F2 hybrids were recorded.Taiwan B was significantly heavier and longer thanTaiwan A in three subsequent generations of testing,when the strains were not subjected to any selectionpressure. Mean of F1 hybrids was intermediate betweenthe mean of the putative parents, but intermediacywas not exact. To measure this, hybrid index over twogenerations was estimated (Table 2) using the formula:
2Vh = 1/g ∑ (hi-Mli)/(U2I-Uli),
i = lwhere g is no. of generations; hi is mean of thereciprocal F1 ; M1i is mean of two parent groups; Uli ismean of the inferior parent group; and U2I is mean ofthe superior parent group.
Hybrid index of body weight was much higher thansimilar index for total length and standard length.
Genotype x diet interaction has also been observedfor almost all morphometric traits in experimentsconducted at UM (Eriyusni et al. 1998), with twoO. niloticus and one O. mossambicus strains and two diets(commercial and UM diets prepared from mainly localingredients).
Apart from tilapia hybridization, crossing betweenspecies is being carried on Clarias spp. The local Clariasbatrachus, although has low growth and is difficultto culture, is tasty. This species has been crossedwith exotic African catfish C. gariepinus and withC. macrocephalus. Hybrids of these two crosses havehigher survival rate than that of their parent species(Ong 1992b).
Giant freshwater prawn-breeders from hatcheryversus wild stocks
A study was undertaken on giant freshwater prawn(Macrobrachium rosenbergii) to compare the performanceof wild stock and pond breeders (Zainoddin 1995).This study, involving four generations, showed thatgrowth and survival rates of stocks derived from pondbreeders during larval stage were better than those
from wild stocks. On the other hand, growthperformance of the stocks from wild breeders duringpostlarval stage was better than those derived frompond breeders.
Larviculture of genotype from wild breeders duringthe parental and next three generations did not showany significant difference but the fourth generationlarvae showed faster metamorphosis and highersurvival rate.
Selection
Genetic selection was conducted to improve overallquality of the red tilapia in the following traits:production of fry with intense red body color, fastgrowth and desirable morphological characteristicssuch as smaller head size, higher body girth and higherweight per unit length. Based on results of the study,red tilapia fry may be produced by spawningindividuals which originated from fry with completelyred color. The selection of “supergrowers” was foundto give rise to faster growing fry as compared tounselected wild stock (Chuah and Nor Azam 1992).
The project is still progressing at the Central FisheriesResearch Station (CFRS), Batu Berendam, andquantitative analysis of the response to geneticselection is expected in near future.
Ornamental Fish
A great deal of interest exists among breeders ofornamental fish in genetic variation of color andmeristic characteristics. Private breeders in Malaysiaand Singapore have developed many new varieties ofguppy, tiger barbs and koi (Japanese carp) but due totheir own secrecy, published papers are scant. Ofsignificance to mention is the publication fromShahreza et al. (1998) on crosses among three varietiesof tiger barb, Puntius tetrazona - normal (N), green (G)and yellow (Y). Results of their crosses indicate thedominance of N over G and Y, and there was nosegregation of the yellow varieties.
Color genetic studies of ornamental fish are also beingconducted by CFRS and will be undertaken by UM(Selvaraj, pers. comm.).
Molecular Genetics
Intensive fish molecular genetic studies were initiatedat UPM. Other universities are also planning theirresearch programs on these.
Table 2. Hybrid index of some traits in crosses of tilapiastrains from Taiwan after 24 months.
Trait
Weight (g)Total length (cm)Standard length (cm)
Mean
P2
145.0019.2015.0
F1reciprocals
17919.515.7
Hybridindex
703532
P1
207.0020.7017.0

69
Tan et. al. (1998) developed RAPD (randomlyamplified polymorphic DNA) and AFLP (amplifiedfragment length polymorphism) techniques to typefor DNA markers in river catfish, Mystus numerus andin several strains of Oreochromis spp. (Chong et al. 1998;Usmani et al. 1998).
In M. numerus, genetic similarity index (SI) valuesamong five sampled populations indicate almostidentical results with RAPD and AFLP. High similaritybetween populations and existence of sufficientheterogeneity within populations were observed. Thisgroup now aims to search for molecular markers inthe populations which may have influence onquantitative traits.
At UPM, research work has generated a total of 73RAPD markers in three strains of tiger barbs.Significant polymorphism observed in threepopulations (N, Y and G) was followed by work on theassessment of genetic variability within and betweenpopulations, and estimation of Nei and Li’s SI. Resultsshowed significant genetic differences among tigerbarb populations (Asma et al. 1998).
Preliminary results on DNA fingerprinting in aMalaysian strain of B. gonionotus using a YNZ22 DNAprobe on genomic DNA digested with Hae III werereported (Siraj et al. 1998). As there were only a lownumber of bands shown in the fingerprint, geneticvariability in their 15 samples was minimum.
Molecular genetic work is also planned on someornamental fish by Dr. Selvaraj (pers. comm.) of UMin collaboration with UPM and Central FisheriesResearch Institute.
Future Outlook
For a country placing importance on aquaculture,Malaysia cannot afford to neglect the application ofgenetics in aquaculture. With the problems of artificialpropagation being overcome, there is increasing useof hatchery-produced stocks in aquaculture. Hatcheriesfunction as genetically isolated closed units raisingtheir own stock of breeders and producing seed fordistribution in grow-out areas. In fish and shellfish,where fecundity is very high with thousands of eggsper female, the breeding can be concentrated on lessthan 1% of the fish. Management practices thus exertstrong selection pressures.
Breeding of fish with high local demand anddevelopment of modern hatchery techniques shall
further reduce the dependence on imported fry.Already one of the mainstays of the industry, prawnculture, is showing a decline in the quality of stock.The size obtained from wild broodstock is larger thanthose cultivated from hatchery stock, probably becauseof inbreeding (Ong 1992a). Here again the selectionprogram for higher body weight and genetic resistanceto disease may be practiced.
Restocking of hatchery-bred fish in public waterbodiescould reduce the genetic pool more than in closedaquaculture environments. Some rare or endangeredfish are being stocked in public waters throughhatchery production while commercial fishingcontinues. Without control of the number of parentsto maintain variability, there could be loss of importantgenetic characteristics such as disease resistance.
Besides inbreeding, changes in the endemic gene poolcould also be occurring through introductions ofspecies or varieties for aquaculture. The consequencesof such introductions are not yet known.
There is a need for definition of the effectivepopulation size and the rate of loss of variability tocontrol inbreeding through hatchery propagation.Consequently, maintenance of a reservoir of geneticvariability, so that cultured populations are notvulnerable to change, is important. However, the useof holding facilities is expensive so cryopreservationof sperm and ova if/when possible could be analternative worth looking into. Moreover, aquaticorganisms introduced for culture could be made sterilethrough hybridization, ploidy or hormonal means.
Fish molecular biology research is gaining importancein UM. This research must be linked with that ofinstitutional or industrial breeders whose coopera-tion makes it possible to include many fish samplesin the experiments, and to develop proper breedingstrategies.
Conclusion
Genetic research is no longer an option but arequirement if aquaculture in Malaysia is to improve.However, techniques adopted should be feasible withinthe infrastructure available at national and farm levelsand relatively safe environmentally. Fish species ofeconomic importance should be bred and selected forfurther genetic improvement. Molecular genetics mayplay an important role in identification and selectionfor genetic improvement in the future.
Genetics for Improvement of Fish in Malaysia

Fish Genetics Research in Member Countries and Institutions of INGA70
References
ACIAR.1997. Genetic identification and stock improvement oftilapia in Malaysia. ACIAR Proj. Rep. (Project No. 9026).(Unpubl.).
Asma, A.N., J. Patimah, S.G. Tan and M.S. Shahreza. 1998. Geneticdifferences of tiger barb (Puntius tetrazona) in Perakpopulation using random amplified polymorphic DNAmarkers, p. 291-294. In Proceedings of the Tenth NationalBiotechnology Seminar, Sirim Berhad, Shah Alam.
Chen, F.Y. 1969. Preliminary studies on the sex determiningmechanism of Tilapia mossambica Peters and T. hornorum Trewas.Verh. Int. Ver. Limnol. 17: 719-724.
Chew, Y.F. 1986. Distribution of the Glucose Phosphate Isomerase-2 (GPI-2) and Phosphoglucomutase (PGM) genotypes inpopulation of guppies (Poecilia reticulata). University of Malaya,Malaysia. B.S. thesis.
Chew, Y.F. and T.K. Mukherjee, T.K. A survey of breeding farmsand hatcheries in Malaysia. (Unpubl.).
Chong, L.K., S.G. Tan, K. Yusoff and S.S. Siraj. 1998. Amplifiedfragment length polymorphism analysis of Malaysian catfish,Mystus nemurus, p. 134-136. In Proceedings of the NationalCongress on Genetics, Bangi, Malaysia.
Chuah, H.P. and A.G. Nor Azam. 1992. Genetic selection on thered tilapia, p. 247-248. In Y.W. Ho, M.K. Vidyadaran, N.Abdullah, M.R. Jainudeen and Abd. Rani Bahaman (eds.)Proceedings of the National Intensification of Research inPriority Areas (Agricultural Sector) Seminar. Vol. II.
Das, K. 1990. Chromosomal studies of eight Malaysian freshwaterfish species of economic importance. University of Malaya,Malaysia. M.S. thesis.
Daud, S.K., B.J. McAndrew, D. Penman and K.J. Ang. 1995.Discriminating loci between Penaeus monodon and P. merguiensis(abstract), p. 26. In Proceedings of the First Asian AustralasianFish Genetics Workshop, University of Malaya, Malaysia.
DOF. 1996. Annual report. Department of Fisheries, Ministry ofAgriculture, Government of Malaysia.
Geeta, S. 1995. An evaluation of economic traits in a diallele crossamong four populations of the genus Oreochromis. 203 p.University of Malaya, Malaysia. Ph. D. thesis.
Eriyusni, N., T.K. Mukherjee, W.E. Wan Khadijah, O. Selvaraj andS.A. Hj. Baba. 1998. Effect of two diets on growth and carcassin three strains of Oreochromis (abstract), p. 89-90. InProceedings of the Twentieth Malaysian Society for AnimalProduction, Malaysia.
Hasnah, N. 1982. Electrophoretic studies of phosphoglucomutaseand glucose phosphate isomerase in Poecilia reticulata.University Malaya, Malaysia. B.S. thesis.
Hickling, C.F. 1960. The Malacca tilapia hybrids. J. Genet. 57: 1-10.
Jamuna, L.S. 1989. Cytogenetic studies of freshwater speciesof ornamental importance. University of Malaya, Malaysia. B.S.thesis.
Mazlan, J. 1997. Malaysian fisheries policy beyond 2000, p. 15-20.In Sidik et al. (eds.) Fisheries and the environment beyond2000. University Pertanian Malaysia, Serdang, Malaysia.
Mohidin, M. 1995. Sex reversal of gynogenetic female of Clariusmacrocephalus using synthetic androgen (abstract). InProceedings of the First Asian Australian Fish GeneticsWorkshop, University of Malaya, Kuala Lumpur.
Mukherjee, T.K. and Geeta, S. 1992. Selection for the productionof all male tilapia progeny, p. 537-538. In Y.W. Ho, M.K.Vidyadaran, N. Abdullah, M.R. Jaunudeen and Abd. RaniBahaman (eds.) Proceedings of the National Intensification ofResearch in Priority Areas (Agricultural Sector) Seminar. Vol.2.
Ong, K.S. 1992a. Aquaculture development in Malaysia in the1990’s. Department of Fisheries, Ministry of Agriculture. 59p .
Ong, K.S. 1992b. Keynote address at the Inception Workshop onConservation and Management of Freshwater Fish and TheirHabitats in Peninsular Malaysia, 18-20 June 1992, Institutefor Advanced Studies-Asian Wetland Bureau, University ofMalaya. 9 p.
Selvaraj, O.S., M. Yahya, K. Pathmaja, T.K. Mukherjee and S.G. Tan.1994. Genetic distance among four strains of tilapia(Oreochromis spp.), p. 130-134. In Proceedings of the FirstNational Congress on Genetics, Bangi, Malaysia.
Siraj, S.S., S. Seki and K.J. Ang. 1993. Diploid gynogenesis inLampam Jawa Puntius gonionotus using UV irradiated spermof Puntius schwanenfeldii followed by temperature shock. NipponSuisan Gakkaishi 59(6): 957-962.
Siraj, S.S., Y.K. Law, S.K. Cheah and K.J. Ang. 1992. Inducedtriploidy in walking catfish, Clarius batrachus, by watertemperature shock. Malays. Appl. Biol. 21(1): 65-69.
Surinderpal, S. 1984. Genetics and variation of the glucosephosphate loci in two populations of guppies, Poecilia reticulata.56 p. University of Malaya, Malaysia. B.S. thesis.
Tan, S.G., M.Yahaya, K. Padmajah, O.S. Selvaraj and T.K. Mukherjee.1989. Electrophoretic characterisation of and geneticrelationships among tilapia (Oreochromis species) strains inMalaysia, p. 12. In Proceedings of the First Asian AustralasianFish Genetics Workshop, Kuala Lumpur, Malaysia.
Tan, S.G., K. Yusof, S.S. Siraj, L.K. Chong, S. Bhassu and S. Usmani.1998. Population genetics for management and conservationof aquatic resources and DNA fingerprinting in fishes: rivercatfish and tilapia (extended abstract). Res. Rep. UniversitiPutra Malaysia.
Tan, S.K., S.G. Tan and Y.Y. Gan. 1980. Liver esterasepolymorphisms in Sepat Siam (Trichogaster pectoralis). Pertanika3(2): 92-96.
Usmani, S., S.G. Tan and K.M. Yusoff. 1998. Development of directamplification of length polymorphisms in Mystus nemurus, p.93-95. In Proceedings of the Third National Congress onGenetics, Bangi, Malaysia.
Yao, L. 1980. Electrophoretic studies of protein variation in Poeciliareticulata and other Poecilia fishes (Teleostei, Poeciliidae). 70 p.University of Malaya, Malaysia. M.S. thesis.
Zainoddin, A.J. 1995. The different performance within pond grownbreeders and wild stocks and their generations in the hatcheryand pond grow-out of giant freshwater prawn (Macrobrachiumrosenbergii), p. 21. In Proceedings of the First Asian AustralianFish Genetics Workshop, University of Malaya, Kuala Lumpur.

71
ABSTRACT
Fish genetics research in the Philippines is still in the nascent stage. It started in the early 1980s with concentrationon the Nile tilapia (Oreochromis niloticus), which is most popularly used in freshwater aquaculture. Classicalbreeding programs to develop fast-growing strains of O. niloticus were initiated and have resulted in increasedgrowth rate. Other genetic studies were focused more on the production of monosex tilapia using the YY-maletechnology, hybridization and sex reversal. Selection program is underway for the development of saline-toleranttilapia for brackishwater aquaculture. Further genetic work is also carried out along with conservation of biodiversityof freshwater fishes.
FISH GENETICS RESEARCHAND DEVELOPMENTIN THE PHILIPPINES
A.S. Camachoa
Bureau of Fisheries and Aquatic Resources860 Quezon Avenue, Quezon City, Philippines
T. AbellaFreshwater Aquaculture Center, Central Luzon State University
Muñoz, Nueva Ecija, Philippines
and
M.M. TayamenNational Freshwater Fisheries Technology Research Center
Bureau of Fisheries and Aquatic ResourcesMuñoz, Nueva Ecija, Philippines
CAMACHO, A.S., T. ABELLA and M.M. TAYAMEN. 2001. Fish genetics research and development in the Philippines, p.71-76. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of theInternational Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
The average annual growth rate achieved by thePhilippine fisheries during 1987-1997 was 2.2%. Whileaquaculture and commercial fisheries have shown 5.3%and 4.4% annual growth, respectively, municipalfisheries declined by 1.6%.
In 1997, fish production was 2.7 million t valued atPhP80.7 billion. Of this, 0.885 million t (32%), valuedat PhP25.9 billionb , was from marine fisheries, and0.958 million t (35%), valued at PhP27.4 billion, was
Introduction
The Philippine archipelago has a wide expanse ofwater resources, approximately seven times bigger thanits land resources. The total marine area is about 220million ha, including 200-mile exclusive economiczone. Inland fishery resources, which include fresh andbrackishwater, swamps, ponds, rivers, lakes andreservoirs, make up 842 247 ha.
Fish Genetics Research and Development in the Philippines
a Current address: No. 220 NE 175th St. Seattle, Washington D.C., 98155 USA.b As of 1999: US$1 = PhP38.
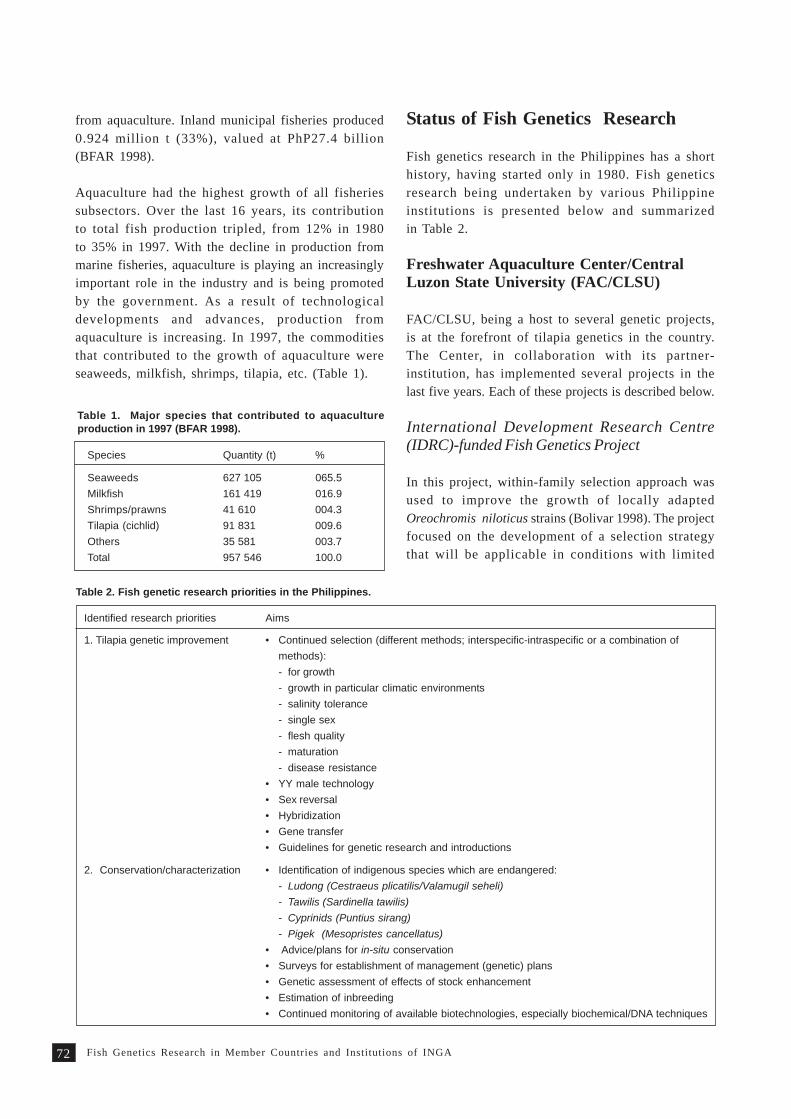
Fish Genetics Research in Member Countries and Institutions of INGA72
from aquaculture. Inland municipal fisheries produced0.924 million t (33%), valued at PhP27.4 billion(BFAR 1998).
Aquaculture had the highest growth of all fisheriessubsectors. Over the last 16 years, its contributionto total fish production tripled, from 12% in 1980to 35% in 1997. With the decline in production frommarine fisheries, aquaculture is playing an increasinglyimportant role in the industry and is being promotedby the government. As a result of technologicaldevelopments and advances, production fromaquaculture is increasing. In 1997, the commoditiesthat contributed to the growth of aquaculture wereseaweeds, milkfish, shrimps, tilapia, etc. (Table 1).
Status of Fish Genetics Research
Fish genetics research in the Philippines has a shorthistory, having started only in 1980. Fish geneticsresearch being undertaken by various Philippineinstitutions is presented below and summarizedin Table 2.
Freshwater Aquaculture Center/CentralLuzon State University (FAC/CLSU)
FAC/CLSU, being a host to several genetic projects,is at the forefront of tilapia genetics in the country.The Center, in collaboration with its partner-institution, has implemented several projects in thelast five years. Each of these projects is described below.
International Development Research Centre(IDRC)-funded Fish Genetics Project
In this project, within-family selection approach wasused to improve the growth of locally adaptedOreochromis niloticus strains (Bolivar 1998). The projectfocused on the development of a selection strategythat will be applicable in conditions with limited
Table 1. Major species that contributed to aquacultureproduction in 1997 (BFAR 1998).
Species
SeaweedsMilkfishShrimps/prawnsTilapia (cichlid)OthersTotal
Quantity (t) %
627 105 065.5161 419 016.941 610 004.391 831 009.635 581 003.7957 546 100.0
Identified research priorities
1. Tilapia genetic improvement
2. Conservation/characterization
Aims
• Continued selection (different methods; interspecific-intraspecific or a combination ofmethods):- for growth- growth in particular climatic environments- salinity tolerance- single sex- flesh quality- maturation- disease resistance
• YY male technology• Sex reversal• Hybridization• Gene transfer• Guidelines for genetic research and introductions
• Identification of indigenous species which are endangered:- Ludong (Cestraeus plicatilis/Valamugil seheli)- Tawilis (Sardinella tawilis)- Cyprinids (Puntius sirang)- Pigek (Mesopristes cancellatus)
• Advice/plans for in-situ conservation• Surveys for establishment of management (genetic) plans• Genetic assessment of effects of stock enhancement• Estimation of inbreeding• Continued monitoring of available biotechnologies, especially biochemical/DNA techniques
Table 2. Fish genetic research priorities in the Philippines.

73
higher than mixed sex tilapia. Dissemination of YYtechnology is being done through 49 accreditedhatcheries in the Philippines.
Genetic Improvement of Farmed Tilapias (GIFT)
FAC/CLSU was one of the research partners in theICLARM-coordinated and implemented Project forthe Genetic Improvement of Farmed Tilapia (GIFT).Apart from participation in major breedingexperiments, the FAC/CLSU also completed severalcomplementary studies and graduate theses research.Among these were the response of GIFT strain tovarious feed formulations; growth performance in rice-fish environments and with bagasse (sugarcane waste)use in freshwater aquaculture.
Fish Biodiversity with Focus on Ex-situConservation of Freshwater Fishes
For conservation and protection of biological diversity,an inventory of indigenous freshwater fishes in thePhilippines was made. This was followed by collectionand propagation of these fishes by induced spawning.Artificially bred fishes were cultured in ponds andhapas. Some of the indigenous freshwater fish thatwere bred successfully in captivity were Anabastestudineus, Trichogaster pectoralis, T. trichopterus, Clariasmacrocephalus and Misgurnus anguillicaudatus.
National Freshwater Fisheries TechnologyResearch Center / Bureau of Fisheries andAquatic Resources (NFFTRC/BFAR)
For ten years, NFFTRC/BFAR has been a collaboratorin UNDP-funded GIFT Project and the ODA-fundedGMIT Project. Ongoing activities of the center are:• Application of YY male technology to an
improved breed of O. niloticus and fingerlingproduction of GMT from crosses of YY males withdifferent maternal strains of O. niloticus, under acollaborative project with UWS with funding fromODA; and
• Tilapia broodstock development for saline watersin collaboration with FAC/CLSU and thePhilippine Council for Aquatic and MarineResearch and Development (PCAMRD).
University of the Philippines at Los Baños(UPLB)
The Genetic and Molecular Biology Division of UPLBis working mainly on conservation and identificationof stocks/genetic variations using biochemical method
facilities. Twelve generations of within-family selectionhave shown that this approach is effective, asdemonstrated by the selection response. Theregression of mean breeding values against a numberof generations indicated that the expected genetic gainwould be about 12% per generation. Based on mixedmodel methodology, the estimate of heritability in thebase population was 0.38. Not much genotype-environment interaction was observed in ponds, tanksand cages. Although the selection was done in tankenvironment, good response was also observed inhapas and ponds. The study has indicated routineselection activities can be undertaken on small facilitieslike tanks while the production of stock and grow-outcan proceed normally in ponds.
From a managerial perspective, it has been found thatwithin-family selection is easy to manage andinbreeding can be kept to a minimum if a structuredmating scheme, like a rotational mating plan is used.Rotational mating has proven to be easy to apply inassociation with within-family selection scheme wherea complete pedigree is maintained. Within-familyselection does not require extensive facilities as wouldbe needed for a presumably more efficient selectionapproach like combined selection. The choice of aselection procedure, particularly for tilapia, is a matterto be decided not only on genetic aspects but also oneconomic grounds, given the prevalent scale of tilapiaindustry in Asia, which is highly diverse and small-scale. Small-scale farmers can practice on-farmselective breeding using a simple, low-cost within-family selection scheme. This will empower farmersto use strains of their choice and not to depend oncommercial hatcheries.
Genetic Manipulation for Improved Tilapia(GMIT)
Through the application of simple geneticmanipulations, the University of Wales, Swansea(UWS), in collaboration with FAC-CLSU and fundingfrom the Overseas Development Administration ofU.K. (ODA/UK), has developed a new genetictechnology for producing all or nearly all-maleprogeny in O. niloticus. Known as the “YY maletechnology”, this takes the form of a breeding programcombining feminization and progeny testing toproduce novel males with YY genotypes (i.e., with twomale sex chromosomes) instead of the usual XY malegenotype (Mair and Abella 1997). These YY males areknown as “supermales” and have the unique propertyof siring only male progeny. The growth performanceof the genetically male tilapia (GMT) is 30%-50%
Fish Genetics Research and Development in the Philippines

Fish Genetics Research in Member Countries and Institutions of INGA74
and DNA fingerprinting. Some of the studiesconducted include comparison of the protein bandingpatterns of milkfish (Chanos chanos), O. niloticus andother fish in polluted and less polluted areas in Lagunade Bay. Initial results indicate that heterogenousbanding exists in stocks found in Laguna de Baypolluted areas.
University of the Philippines in the Visayas(UPV)
Genetics research at UPV is being carried out in thefollowing aspects:• survival mechanisms of selected marine finfish
larvae (DNA/RNA composition of milkfish larvalforms, seabass; DNA/RNA ratio for determininggrowth of fish);
• database of genetically identified commercialspecies (being conducted in collaboration with aVenezuelan scientist);
• genetic profiling of commercially importantfinfish species and species karyotyping (beingconducted in collaboration with a Japanesescientist); and
• DNA fingerprinting and electrophoreticidentification.
PCAMRD
PCARMD, together with the Centre de CooperationInternationale En Recherche Agronomique Pour LeDeveloppement (CIRAD), France, and BFAR, haveinitiated a four-year project for developing a highlysalinity-tolerant tilapia strain for brackishwaterenvironments. The project is financed by thePhilippine and French governments and is conductedat the BFAR National Integrated Fisheries TechnologyDevelopment Center, Pangasinan.
The technique used is repeated backcrossing of F1
progeny of O. niloticus and O. mossambicus. From thesalinity-tolerant hybrid population, selection for fastgrowth under saline water conditions will be done,followed by rotational backcrossing scheme protocol.
In France, a similar project using hybrids obtainedfrom crossing fast-growing O. niloticus with highsalinity-tolerant species Sarotherodon melanotheron is alsounderway. The project is a collaboration amongCIRAD, Institut National de la Recherche Agro-nomique, L’Institut Francais de Recherche Scientifiquepour le Developpement en Cooperation, Institut
Francais de Recherche pour l’Exploitation de la Merand Liège University in Belgium.
GIFT Foundation International Inc. (GFII)
For ten years (ending in December 1997), GIFT projecthas demonstrated how stepwise progression fromsystematic documentation of genetic resources to theirutilization can generate rapid benefits for both farmersand consumers (Abella 1998; GIFT 1998). The projectevolved into a self-sustaining tilapia breeding programknown as GFII. This nonprofit foundation wasestablished to serve as custodian of the breedingnucleus and to continue the selective breeding startedand developed under the GIFT Project. Thefoundation’s research and breeding activities arefunded by contributions from hatchery operators whouse GIFT broodstock as well as “royalties” collectedfrom the sale of fingerlings by the hatcheries.
San Miguel Corporation (SMC)
SMC, a profit-oriented private company in thePhilippines has been doing selection experiments toimprove the growth of O. niloticus. In 1995, it hasdeveloped a special B-Meg strain, called Best 200. Thisstrain showed superior growth compared withcommercially available O. niloticus.
National Research Priorities
New Philippine National Tilapia BreedingProgram (NTBP) of BFAR
The broad objective of the national breeding program(Eknath et al. 1991; Sevilleja et al. 1993) is to enhancegenetic resources for aquaculture productivity. Thespecific objectives are to: (1) sustain a long-termselection program in improving performance traits(growth, survival, maturation, disease resistance andother economic traits); (2) institute broodstockmanagement practices to avoid inbreeding; and (3)disseminate efficiently the genetic gains to targetbeneficiaries of the breeding program, especially theresource-poor farmers.
Efficient dissemination of genetic gains is possible onlywhen there are organized channels for production anddistribution of fish seed to fishfarmers (Morales et al.1994). The NTBP operates as the National BroodstockCenter (NBC) for tilapia in the country. The NBC hasthe primary function as first-level multiplier/producer

75
of quality broodstock and fingerlings for distributionto the second-level multiplier stations. The frameworkof NTBP includes the participation of differentRegional Outreach Stations of the Department ofAgriculture (DA) and of Local Government FishHatchery Units in provinces, municipalities andvillages. The program also includes partnership withprivate tilapia hatcheries assisted by BFAR which willbe accredited to serve as the second-level multiplier/producer of broodstock and fingerlings for the tilapiahatchery operators and grow-out operators,respectively. Various state colleges and universities alsoserve as collaborators in freshwater aquacultureresearch, development and extension.
The national fish breeding program was establishedin the Philippines, following the experience of theGIFT Project. The present efforts of NFFTRC/BFARand of FAC/CLSU in formulating organizationalstructure, strategies and management plan for a self-sustaining tilapia breeding program will be furtherstrengthened and given support.
Development of saline-tolerant tilapia
In 1997, tilapia production from 14 000 ha freshwaterfishponds was reported at 39 005 t. Brackishwaterfishponds with an area of 239 323 ha contributed only5 939 t (BFAR 1997) because more than half of theseare underutilized. There is room for increasingproductivity of brackishwater ponds by using otheraquaculture species. Tilapia has been identified as oneof the species for the purpose, and breeds could bedeveloped to withstand saline and high watertemperature conditions.
The need for a salinity-tolerant tilapia species is dueto these reasons: (i) increased consumer acceptanceof tilapia; and (ii) high demand for tilapia fingerlingsdue to inadequate supply of milkfish fry for stockingbrackishwater ponds where shrimp culture has beenabandoned due to disease problems. At present, onlyO. mossambicus and tilapia hybrid can thrive well inbrackishwater fishponds.
At FAC/CLSU, a study was undertaken to assess thesalinity tolerance of different strains of geneticallyimproved O. niloticus. Three strains of O. niloticus(GIFT, IDRC and Israel) were mated following a 3 x 3diallele cross. The saline-tolerant O. mossambicus, wasused as the reference line.
Progenies from three purebreds and six reciprocalcrosses of O. niloticus and purebred O. mossambicus weretested for tolerance at different salinities. Three salinitytolerance indices, i.e., median lethal salinity (MLS-96), mean survival time (MST) and median survivaltime (ST50), were used for evaluation. Group variationsof these indices were subjected to statistical analysis.The reciprocal effect, heterosis and general combiningability were estimated based on the differences insalinity tolerance of different cross progenies.
Results revealed that salinity tolerance, based on themeasurements of MLS-96, MST and ST50, variedamong the crosses. Purebred IDRC, IDRC x GIFT,purebred Israel and O. mossambicus exhibited highestMLS-96 of 22.43, 22.33, 22.27 and 22.21 ppt,respectively. Crossbred IDRC (female) x Israel (male)(and reciprocal) showed relatively high MLS-96 (20.93and 20.09 ppt) while purebred GIFT and crossbredsof GIFT (female) x Israel (male), GIFT (female) xIDRC (male) and Israel (female) x GIFT (male) werelow-tolerant crosses, and exhibited MLS-96 lower than20 ppt. For MST and ST50, O. mossambicus exhibitedthe highest tolerance which were 145.16 min and143.75 min, respectively. Results of the study indicatedthat IDRC strain is a potential strain for saline water.
Reciprocal effects for MLS-96 and MST were foundin the crossbreds between GIFT and IDRC strains. TheIDRC (female) x GIFT (male) cross has significantlyhigher MLS-96 and MST (P<0.01) compared to itsreciprocal cross. Reciprocal effects were also observedin MLS-96 of GIFT (female) x Israel (male) crossbred(P<0.01) and MST (P<0.05) and ST50 (P<0.01) ofIDRC (female) x Israel (male) crossbred.
Percentage heterosis was negative for every crossbred.The estimated values for overall crosses were –8.36%,-17.26% and –17.41% for MLS-96, MST and ST50,respectively, indicating no hybrid vigor for salinitytolerance in crossbreds.
Estimates for general combining ability based on theindices measured showed that IDRC strain gavepositive values when used as maternal strain andsignificantly differed from Israel and GIFT strains.This reflects IDRC strain’s merit for hybridization.
Based on the results on MLS-96, MST and ST50,O. mossambicus, it was observed that the salt-toleranttilapia that can survive in seawater, cannot survive insalinity over 25 ppt following direct transfer. This could
Fish Genetics Research and Development in the Philippines

Fish Genetics Research in Member Countries and Institutions of INGA76
be attributed to its progenies that were spawned andhatched in freshwater.
Fish genetics research in the Philippines has alreadyachieved a certain level of competency. Whilelimitations exist, these are manageable, and if attendedto, the research can be accelerated.
On 24 September 1996, BFAR convened a meeting ofresearch scientists from various Philippine institutionsto initiate the formation of the Philippine nationalnetwork which is the local counterpart of INGA. Thisnational network serves as a forum for informationexchange and for facilitating national and internationalcollaborative research.
References
Abella, T.A., A.E. Eknath, B.O. Acosta, R.A. Reyes, R.R. Velascoand B.M. Rodriguez, Jr. 1998. Genetic Improvement ofFarmed Tilapias: a Philippine experience. Paper presentedat the Fifth Asian Fisheries Forum, 11-14 November 1998,Chiangmai, Thailand.
BFAR. 1998. 1997 Philippine fisheries profile. Bureau of Fisheriesand Aquatic Resources, Quezon City, Metro Manila,Philippines.
Bolivar, R.B. 1998. Estimation of response to within-familyselection for growth in Nile tilapia (Oreochromis niloticus).Dalhousie University, Halifax, Nova Scotia, 166 p. Ph.D.thesis.
Eknath, A.E., H.B. Bentsen, B. Gjerde, M.N. Tayamen, T.A. Abella,T. Gjedrem and R.S.V. Pullin. 1991. Approach to nationalbreeding programs: pointers from a tilapia pilot study. Naga:ICLARM Q. 14(2):10-12.
GIFT. 1998. Genetic Improvement of Farmed Tilapia (GIFT)Project. Final Rep. Part 1. ICLARM. Makati, Philippines.
Mair, G.C. and T.A. Abella, Editors. 1997. Technoguide on theproduction of genetically male tilapia (GMT). FreshwaterAquaculture Center, Central Luzon State University, NuevaEcija, Philippines.
Morales, G.A., A.E. Eknath, R.C. Sevilleja, M.M. Tayamen, R.B.Bolivar and R.A. Reyes. 1994. An evolving National TilapiaBreeding Program for the Philippines. Poster paperpresented at the Fifth International Symposium in Geneticsin Aquaculture, 19-25 June 1994, Dalhousie University,Halifax, Canada.
Sevilleja, R.C., M.M. Tayamen, R.B. Bolivar, R.A. Reyes and A.E.Eknath. 1993. Philippine National Tilapia Breeding Program.Paper presented at the Third National Symposium-Workshopon Tilapia Farming, 25-27 November 1993, Miag-ao, Iloilo,Philippines.
Strengths and Limitations inConducting Fish Genetics Research

77
ABSTRACT
The paper gives a brief review of the aquaculture genetic research in Thailand, including genetic characterizationof populations or species, selective breeding, sex control, genetic manipulation, cytogenetic studies, cryopreservationand genetic engineering.
AQUACULTUREGENETICS RESEARCH
IN THAILAND
N. PongthanaNational Aquaculture Genetics Research Institute
Pathum Thani 12120, Thailand
PONGTHANA, N. 2001. Aquaculture genetics research in Thailand, p. 77-89. In M.V. Gupta and B.O. Acosta (eds.) Fishgenetics research in member countries and institutions of the International Network on Genetics in Aquaculture.ICLARM Conf. Proc. 64, 179 p.
Introduction
The major inland resources in Thailand availablefor freshwater aquaculture and fisheries developmentare composed of 66 rivers, 10 233 lakes and swamps,685 reservoirs and human-made lakes with a surfacearea of 566 400 ha. The Gulf of Thailand on the eastcoast has an area of 320 000 km2 and a coastline of 1 874km. The Andaman Sea to the west has an area of116 280 km2 and a coastline of 740 km. The distancealong both coastlines is 2 614 km, with a total area of272 000 km2.
With these natural resources, fisheries industries playan important role in the economy of Thailand. Fishproduction in 1993 was estimated at 3.385 million t,with a total value of US$3 136.27 million. In 1993,the total value of exported fish and fishery productswas valued at 91 018.326 million baht (US$3 640.73million), ranking fourth among export items.Moreover, the fisheries industries have contributeddirectly to the development of other relatedbusinesses, of which there are about 7 000 units, suchas ice plants, cold storage, fish processing, shipbuilding and fishing gear.
Aquaculture Genetic Research in Thailand
Overview
Genetic characterization of populations/species
Genetic characterization of populations or species inblack tiger shrimp (Penaeus monodon), banana shrimp(P. merguiensis), oyster (Crassostrea belcheri), giantfreshwater prawn (Macrobrachium rosenbergii),snakeskin gourami (Trichogaster pectoralis), silver barb(Barbodes gonionotus), catfish, tilapia and frog (Ranarugulosa) was carried out by molecular genetic studies,using either isozyme electrophoresis, polymerasechain reaction (PCR) and restriction enzyme analysisor microsatellite DNA marker techniques.
Black tiger shrimp
Genetic variation was electrophoretically examined inpopulations of P. monodon from four locations alongthe coasts of Thailand (Trad, Suratthani, Phuket andSatun) to determine the extent of genetic isolation.The average heterozygosity of the species is relativelylow (0.066±0.028) based on 46 loci. Results showedsignificant difference between populations from

Fish Genetics Research in Member Countries and Institutions of INGA78
Andaman Sea and Gulf of Thailand, and between thosefrom Trad and Suratthani (Sodsuk 1996).
Studies to characterize microsatellite DNA inP. monodon genome and to develop genetic markersfor use in various applications, including populationgenetic studies, selective breeding and genomemapping (Tiptawonnukul 1996) were completed. Atotal of 131 microsatellites were isolated andcharacterized from partial genomic libraries byscreening with an oligonucleotide probe (GT)15. Thesemicrosatellites were classified as perfect, imperfectand compound repeats. With 51.9%, imperfectrepeats were the predominant category ofmicrosatellites. A total of 97 (GT)n microsatellites wereisolated and the frequency of the (GT)n repeats in thegenome was estimated. The average distance betweenneighboring (GT)n microsatellites was 92.8 kb. Themost common size class in all categories containedsequences between 30 and 35 repeats in length. PCRprimers were designed from the unique flankingsequences of seven microsatellites, two pairs for (AT)n
and five pairs for (GT)n microsatellites. By testingeight individual shrimps from Kruntung, AndamanSea and Angsila, Gulf of Thailand, with PCRamplification, it was found that three of sevenmicrosatellite primer sets, namely, Pmo 18, Pmo 36and Pmo 14, produced fragments which sizes were inthe expected range; the others gave nonspecificamplifications. Two microsatellite loci, Pmo18 andPmo386, yielded scorable PCR products and showedhigh level of polymorphism indicating their potentialuse in DNA typing of P. monodon.
Randomly amplified polymorphic DNA analysis(RAPD) was used to examine genetic variation of fourgeographically separated wild populations of P.monodon from two regions, Andaman Sea and Gulf ofThailand (Pongsomboon 1996). A total of 75individuals were tested. A screen of 300 randomdecaoligonucleotide primers identified sevenprimers, which were selected for the analysis of DNA.A total of 80 reproducible RAPD fragments rangingin size from 200 to 2 000 bp were scored and 62fragments (77.5%) were polymorphic. Thepercentages of polymorphic bands were 57.77, 52.2,45.6 and 53.4% for samples from Satun–Trang, Trad,Angsila and Medan, respectively. The similarity index(SI) within and between populations and geneticdistances was performed based on the RAPD data.The results of SI within population illustrated that
the Angsila sample was the most similar amongthemselves. The results of SI between populations andUPGMA dendrograms showed that the Medan samplewas significantly different from the three samples ofThai P. monodon. Primer 428 detected a RAPD markerthat was found only in samples from Andaman Sea,Satun–Trang and Medan, suggesting the use of thismarker as region–specific. RAPD patterns among thefour samples gave 214 genotypes. Of these, 160 werepopulation–specific genotypes and 10 were region–specific; 97 of these genotypes were represented byan individual. Chi–square analysis of the genotypesshowed that Thai and Indonesian P. monodon weresignificantly different for all primers (P<0.0001), andThai P. monodon from Andaman Sea and Gulf ofThailand were also different for primers 101, 268,428, 457 and 459 (P = 0.0049, <0.0001, < 0.0014and = 0.0156, respectively).
cDNA library of abdominal muscles has beenconstructed in lambda ZAPII vector using E. coliXL–1 Blue cost in order to obtain quickly DNAsequences (Boonyawan 1998). A set of 93 partialP. monodon cDNA sequences (expressed sequence tags,ESTs) was obtained. Forty-one sequences were foundto share significant matching to known genesequences from other organisms. Their putativeidentities are: actin, myosin heavy chain, myosin lightchain, acylphosphatease, arginine kinase, Ca2-
ATPase, cytochrome b, gelsolin, glutaryl–CoAdehydrogenase, heat shock protein, initiation factor,phosphopyruvate hydratase and pyrophosphatase.Two putative actin clones, designated as clonesPMM048 and PMM099, were then subjected tocomplete sequencing. Open reading frames (ORF)of 376 and 377 amino acids were found, respectively.The predicted amino acid sequences showed highdegree of similarity to those of insect and crustaceanactins as well as of other vertebrate actins. Theidentities of those sequences were over 90%. Thesteady state mRNA actins were also analyzed byNorthern blot. One 1.2 kb band was found whenprobed, with each probe corresponding well to itspredicted size. When Southern blots, containing P.monodon’s genomic DNA digested with EcoR I, HindIII, Pst I and Xho I, were probed with DIG labeledprobe, 8, 11, 5 and 4 DNA bands larger than 1.2 kbcould be detected in autoradiograms. Based on thenumber of hybridized bands and their intensities, thenumber of actin genes in the P. monodon genome wasestimated at five to ten copies per haploid genome.

79
Banana shrimp
Genetic variation was electrophoretically examined inpopulations of P. merguiensis from three locations alongthe coasts of Thailand (Chonburi, Suratthani andSatun) to determine the extent of genetic isolation.The average heterozygosity of the species is relativelylow (0.066±0.028) based on 26 loci. Results indicatedthat all three populations are different. The twopopulations from Gulf of Thailand (Chonburi andSuratthani) were more closely related (Sodsuk andSodsuk 1998a).
Oyster
Genetic variation was electrophoretically examined inpopulations of C. belcheri from five locations along thecoasts of Thailand (Suratthani, Krabi, Ranong,Chantaburi and Trad) to determine the extent ofgenetic isolation. The average heterozygosity rangedfrom 0.424 to 0.584, and the mean number of allelesper locus ranged from 2.82 to 3.55, based on 11 loci.The most genetically similar populations were thosefrom Krabi and Ranong, while the most different werethose from Suratthani and Ranong (Pongthana et al.1999a).
Genetic analyses of growth and survival rates werecarried out in three C. belcheri populations fromSuratthani, Ranong and Trad, grown in two naturalenvironments (Ban Don Bay, Suratthani and Kao YouaBay, Ranong). There were significant genetic effectson growth and survival rates. Significant location effectexisted only in survival rate. The growth and survivalrates of the genetic groups were site-dependent, andsignificant genotype by environment interactions werepresent for both parameters. The difference in rankingof performance in various environments wasresponsible for this interaction (Pongthana et al1999a).
Giant freshwater prawn
Genetic variation in natural populations ofM. rosenbergii was studied to identify races in differentlocations of Thailand (Yaitavorn 1989). The study wasbased on analysis of mitochondria DNA (mtDNA)variation, which was isolated from hepatopancreas andcut with restriction enzyme Sau3A1 to generate DNAfragments ranging from 0.2-2.0 kb. These DNAfragments were cloned into vector pUC12 at BamH1site and transformed into E. coli strain JM107. After
colony hybridization and Southern blot hybridization,51 recombinant clones were obtained. These cloneswere further selected for strongly hybridized signalwith α -32 P dATP labeled mtDNA, and used asprobes to analyze mtDNA variation in prawns.Therecombinant DNA no. 1 showed significant differencein restriction fragment length polymorphism (RFLP)pattern of mtDNA between specimens from KraburiRiver and from Bangpakong River by Southern blothybridization. The specimens from the latter showedstrong discrete band at 1.1 kb, but those from theformer was at 0.7 kb. By using this clone, RFLP patternof Bangpakong River was similar to that from KungKam Thong Farm. The recombinant DNA no. 1 hadinserted fragment about 1.1 kb and restrictionendonuclease sites for Rsa1, Hha1, Hae111 andBstU1.
Genetic variation was electrophoretically examined inpopulations of M. rosenbergii from three locations alongthe coasts of Thailand (Chachoengsao, Suratthani andSongkla) to determine the extent of genetic isolation.The average heterozygosity of the species is relativelylow (0.031±0.018) based on 24 loci. Results indicatedthat all individuals in three areas belong to the samepopulation (Sodsuk and Sodsuk 1998b).
DNA probe was developed using RFLP techniqueand used for assaying DNA of M. rosenbergii(Prakongsai 1991). The extracted DNA has Tm
(melting temperature) about 86°C corresponding to43% G–C content. The genomic library of M.rosenbergii was constructed from Sau3AI–digestedhepatopancreas DNA of a male prawn and ligated toBamHI site of pUC18. The transformants wereselected through ampicillin resistance, their colorlesscharacteristic on the medium containing IPTG andX-gal, and screening by hybridization with total DNAof either male or female DNA of 20 recombinantcolonies selected for RFLP analysis. Thirteencontained repetitive sequences and could generate 120bp repeating fragments as the common RFLP pattern.The repetitive sequences were divided into two typesbased on their sequence homologies. Sixty-one prawnscollected from three provinces were classified into 13groups based on their HinfI–RFLP patterns againstpMR3 probe and suggested strain variation of M.rosenbergii. MR3–A fragment of pMR3 could generateHinfI–RFLP patterns and showed homology to DNAof P. monodon and P. merguiensis, whereas MR3–B andMR3–C hybridized specifically to M. rosenbergii DNA.
Aquaculture Genetic Research in Thailand

Fish Genetics Research in Member Countries and Institutions of INGA80
Snakeskin gourami
Population structures of T. pectoralis collected fromfive locations in Thailand (Samutprakan, Pitsanulok,Suphanburi, Ubonratchathani and Pattanee) werestudied by detecting isozyme differences using Tris–Citrate buffer pH 7.0 and 8.0 and Tris–EDTA–Boratebuffer pH 8.0 and pH 8.3 to test 14 enzymes, AAT,AK, EST, FUM, GCDH, PGI, HK, IDDH, MDH, ME,MPI, PGDH, PGM and PK, in five organs (muscle,liver, kidney, blood and heart). The highest geneticidentity coefficient of 0.985 was found betweenSamutprakan and Pitsanulok populations; the lowest,0.923, between Pitsanulok and Pattanee populations.From the estimates of genetic identity and distancecoefficients among the five populations, theSamutprakan and Pitsanulok populations had thelowest genetic distance, while the Pitsanulok andPattanee populations had the highest. Morphometricstudy was also carried out by comparing the ratio ofhead length to standard length. The fish fromPitsanulok was different from others. The fish fromSamutprakan and Supanburi had the same headlength to standard length ratio of 0.28, and the fishfrom Ubonratchathani and Pattanee had the sameratio of 0.27 (Pongthana et al. 1999b).
Silver barb
Spatial genetic structure of B. gonionotus populationsin Thailand was reported by Kamonrat (1996).Microsatellite DNA markers were developed from aThai B. gonionotus genomic library and used to studyvarious aspects of its population genetics in Thailandfor evaluating management policies on conservationand genetic improvement. Twelve natural populationsfrom three rivers and 29 hatchery stocks from centraland northeast regions of Thailand were studied.Genetic variability was high in both groups ofpopulations. Multidimensional scaling analysis ofgenetic distances revealed the discreteness apparentbetween watersheds among natural populations, andbetween geographic regions among hatchery stocks.High genetic variability within populations andsignificant genetic differentiation between populationsboth in native and hatchery stocks indicated richgenetic resources of this species in Thailand. However,there were evidences that stock management may posea threat of losing or altering genetic integrity of bothnatural and hatchery populations. Mixed stockanalysis of the fish sampled from the rivers indicated75%-96% were from hatchery populations. This high
genetic contamination of natural populations wasundoubtedly the consequence of restocking programsin which millions of fry/fingerlings of this species arereleased each year in rivers. Evidence of reduction ofgenetic integrity between regions was also observeddue to stock transfer. The results suggested an urgentneed for genetically based stock management policiesfor both natural and hatchery populations.
The potential use of microsatellites for broodstockimprovement in aquaculture was also studied(Kamonrat 1996). Pedigrees of individuals weresuccessfully established in a large communal rearingby using one to five microsatellites. The ability toidentify individuals allowed a complicated geneticexperiment and selective breeding to be conductedin places where facilities were limited. Results areconsidered to be more reliable because environmentalvariances are accounted for as fish are grown togetherfrom birth. In this study, heritability of growth traitsin three stocks was estimated where all families werereared together. The estimates ranged from 0.193 to0.523, suggesting that selective breeding in this speciesshould result in good progress. However,heterozygosity in the largest individuals was greatlyreduced, indicating that rapid inbreeding is very likelyin simple means of selection strategies.
Catfish species
Genomic identification of catfish species by PCRand restriction enzyme analysis of the gene encodingthe immunoglobulin M heavy chain constant regionwere carried out by Thongpan et al. (1994). NuclearDNA was isolated from blood of catfish representingthree families (Clariidae, Pangasiidae and Ictaluriidae)for analysis by PCR and restriction enzymes. Thepurity of DNA (A260/A280) from all fish samples was inthe range of 1.8 to 2.0 primers, specific for the CH4exon of the gene encoding the immunoglobulin heavychain of channel catfish (Ictalurus punctatus) used.Nuclear DNA when amplified with these primersyielded a single band of about 300 base pairs forClarias macrocephalus, Pangasius gigas, P. hypophthalmusand the hybrid of P. gigas x P. hypophthalmus. However,the same primers yielded two DNA bands of about300 and 340 base pairs in C. gariepinus and in thehybrid of C. macrocephalus x C. gariepinus. Hence, thegenetic identity of C. gariepinus could be identified atmolecular level. Nucleotide sequences of the amplifiedDNA were identified for I. punctatus, C. macrocephalus,P. gigas and P. hypophthalmus. Based on the DNA
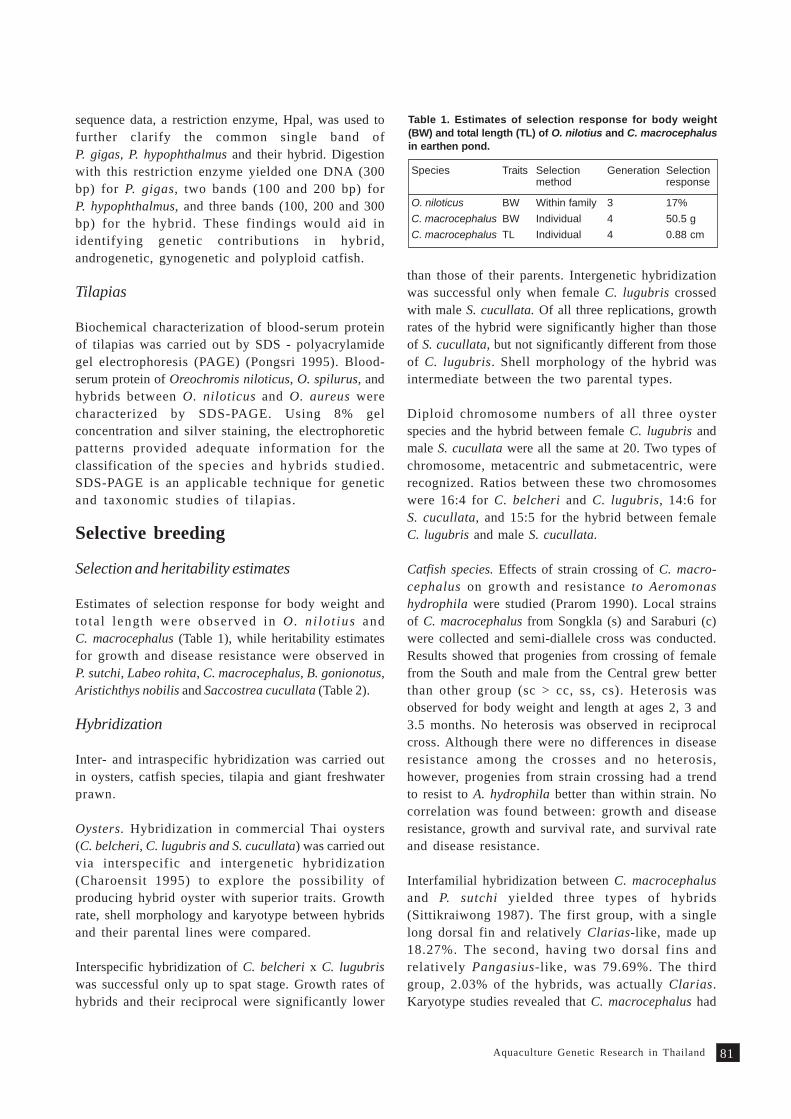
81
sequence data, a restriction enzyme, Hpal, was used tofurther clarify the common single band ofP. gigas, P. hypophthalmus and their hybrid. Digestionwith this restriction enzyme yielded one DNA (300bp) for P. gigas, two bands (100 and 200 bp) forP. hypophthalmus, and three bands (100, 200 and 300bp) for the hybrid. These findings would aid inidentifying genetic contributions in hybrid,androgenetic, gynogenetic and polyploid catfish.
Tilapias
Biochemical characterization of blood-serum proteinof tilapias was carried out by SDS - polyacrylamidegel electrophoresis (PAGE) (Pongsri 1995). Blood-serum protein of Oreochromis niloticus, O. spilurus, andhybrids between O. niloticus and O. aureus werecharacterized by SDS-PAGE. Using 8% gelconcentration and silver staining, the electrophoreticpatterns provided adequate information for theclassification of the species and hybrids studied.SDS-PAGE is an applicable technique for geneticand taxonomic studies of tilapias.
Selective breeding
Selection and heritability estimates
Estimates of selection response for body weight andtota l length were observed in O. ni lo t ius andC. macrocephalus (Table 1), while heritability estimatesfor growth and disease resistance were observed inP. sutchi, Labeo rohita, C. macrocephalus, B. gonionotus,Aristichthys nobilis and Saccostrea cucullata (Table 2).
Hybridization
Inter- and intraspecific hybridization was carried outin oysters, catfish species, tilapia and giant freshwaterprawn.
Oysters. Hybridization in commercial Thai oysters(C. belcheri, C. lugubris and S. cucullata) was carried outvia interspecific and intergenetic hybridization(Charoensit 1995) to explore the possibility ofproducing hybrid oyster with superior traits. Growthrate, shell morphology and karyotype between hybridsand their parental lines were compared.
Interspecific hybridization of C. belcheri x C. lugubriswas successful only up to spat stage. Growth rates ofhybrids and their reciprocal were significantly lower
than those of their parents. Intergenetic hybridizationwas successful only when female C. lugubris crossedwith male S. cucullata. Of all three replications, growthrates of the hybrid were significantly higher than thoseof S. cucullata, but not significantly different from thoseof C. lugubris. Shell morphology of the hybrid wasintermediate between the two parental types.
Diploid chromosome numbers of all three oysterspecies and the hybrid between female C. lugubris andmale S. cucullata were all the same at 20. Two types ofchromosome, metacentric and submetacentric, wererecognized. Ratios between these two chromosomeswere 16:4 for C. belcheri and C. lugubris, 14:6 forS. cucullata, and 15:5 for the hybrid between femaleC. lugubris and male S. cucullata.
Catfish species. Effects of strain crossing of C. macro-cephalus on growth and resistance to Aeromonashydrophila were studied (Prarom 1990). Local strainsof C. macrocephalus from Songkla (s) and Saraburi (c)were collected and semi-diallele cross was conducted.Results showed that progenies from crossing of femalefrom the South and male from the Central grew betterthan other group (sc > cc, ss, cs). Heterosis wasobserved for body weight and length at ages 2, 3 and3.5 months. No heterosis was observed in reciprocalcross. Although there were no differences in diseaseresistance among the crosses and no heterosis,however, progenies from strain crossing had a trendto resist to A. hydrophila better than within strain. Nocorrelation was found between: growth and diseaseresistance, growth and survival rate, and survival rateand disease resistance.
Interfamilial hybridization between C. macrocephalusand P. sutchi yielded three types of hybrids(Sittikraiwong 1987). The first group, with a singlelong dorsal fin and relatively Clarias-like, made up18.27%. The second, having two dorsal fins andrelatively Pangasius-like, was 79.69%. The thirdgroup, 2.03% of the hybrids, was actually Clarias.Karyotype studies revealed that C. macrocephalus had
Aquaculture Genetic Research in Thailand
Table 1. Estimates of selection response for body weight(BW) and total length (TL) of O. nilotius and C. macrocephalusin earthen pond.
Species
O. niloticusC. macrocephalusC. macrocephalus
Selectionmethod
Within familyIndividualIndividual
Generation
344
Selectionresponse
17%50.5 g0.88 cm
Traits
BWBWTL
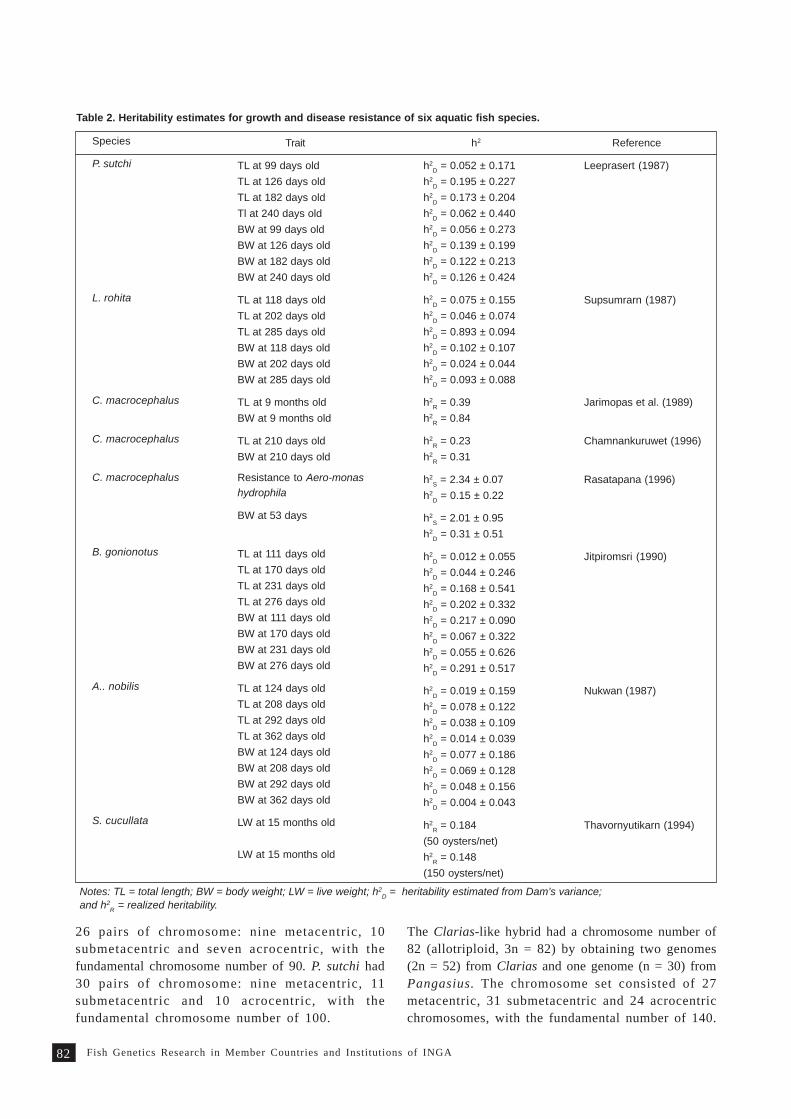
Fish Genetics Research in Member Countries and Institutions of INGA82
26 pairs of chromosome: nine metacentric, 10submetacentric and seven acrocentric, with thefundamental chromosome number of 90. P. sutchi had30 pairs of chromosome: nine metacentric, 11submetacentric and 10 acrocentric, with thefundamental chromosome number of 100.
Species
P. sutchi
L. rohita
C. macrocephalus
C. macrocephalus
C. macrocephalus
B. gonionotus
A.. nobilis
S. cucullata
Trait
TL at 99 days oldTL at 126 days oldTL at 182 days oldTl at 240 days oldBW at 99 days oldBW at 126 days oldBW at 182 days oldBW at 240 days old
TL at 118 days oldTL at 202 days oldTL at 285 days oldBW at 118 days oldBW at 202 days oldBW at 285 days old
TL at 9 months oldBW at 9 months old
TL at 210 days oldBW at 210 days old
Resistance to Aero-monashydrophila
BW at 53 days
TL at 111 days oldTL at 170 days oldTL at 231 days oldTL at 276 days oldBW at 111 days oldBW at 170 days oldBW at 231 days oldBW at 276 days old
TL at 124 days oldTL at 208 days oldTL at 292 days oldTL at 362 days oldBW at 124 days oldBW at 208 days oldBW at 292 days oldBW at 362 days old
LW at 15 months old
LW at 15 months old
h2
h2D = 0.052 ± 0.171
h2D = 0.195 ± 0.227
h2D = 0.173 ± 0.204
h2D = 0.062 ± 0.440
h2D = 0.056 ± 0.273
h2D = 0.139 ± 0.199
h2D = 0.122 ± 0.213
h2D = 0.126 ± 0.424
h2D = 0.075 ± 0.155
h2D = 0.046 ± 0.074
h2D = 0.893 ± 0.094
h2D = 0.102 ± 0.107
h2D = 0.024 ± 0.044
h2D = 0.093 ± 0.088
h2R = 0.39
h2R = 0.84
h2R = 0.23
h2R = 0.31
h2S = 2.34 ± 0.07
h2D = 0.15 ± 0.22
h2S = 2.01 ± 0.95
h2D = 0.31 ± 0.51
h2D = 0.012 ± 0.055
h2D = 0.044 ± 0.246
h2D = 0.168 ± 0.541
h2D = 0.202 ± 0.332
h2D = 0.217 ± 0.090
h2D = 0.067 ± 0.322
h2D = 0.055 ± 0.626
h2D = 0.291 ± 0.517
h2D = 0.019 ± 0.159
h2D = 0.078 ± 0.122
h2D = 0.038 ± 0.109
h2D = 0.014 ± 0.039
h2D = 0.077 ± 0.186
h2D = 0.069 ± 0.128
h2D = 0.048 ± 0.156
h2D = 0.004 ± 0.043
h2R = 0.184
(50 oysters/net)h2
R = 0.148(150 oysters/net)
Reference
Leeprasert (1987)
Supsumrarn (1987)
Jarimopas et al. (1989)
Chamnankuruwet (1996)
Rasatapana (1996)
Jitpiromsri (1990)
Nukwan (1987)
Thavornyutikarn (1994)
Table 2. Heritability estimates for growth and disease resistance of six aquatic fish species.
Notes: TL = total length; BW = body weight; LW = live weight; h2D = heritability estimated from Dam’s variance;
and h2R = realized heritability.
The Clarias-like hybrid had a chromosome number of82 (allotriploid, 3n = 82) by obtaining two genomes(2n = 52) from Clarias and one genome (n = 30) fromPangasius. The chromosome set consisted of 27metacentric, 31 submetacentric and 24 acrocentricchromosomes, with the fundamental number of 140.

83Aquaculture Genetic Research in Thailand
The Pangasius-like hybrid had a chromosome numberof 56 (amphidiploid, 2n = 56) by obtaining onegenome (n = 26) from Clarias and one genome(n = 30) from Pangasius. The chromosome setconsisted of 18 metacentric, 21 submetacentric and17 acrocentric chromosomes, with the fundamentalnumber of 95. The “actual Clarias” hybrids weresupposed to arise from gynogenesis.
Tilapias. Improvement in salinity tolerance ofO. niloticus by means of interspecific hybridization withO. mossambicus was studied (Thanakijkorn 1997). Thehybrid having O. niloticus as a mother (NxM) hadhigher survival rate after salinity challenge test at 20ppt than O. niloticus purebred, but lower than that ofthe reciprocal cross and O. mossambicus. At a 30 pptsalinity, a direct transfer killed all of O. niloticuspurebred and the hybrid with O. mossambicus as amother. The reciprocal hybrid (MxN) andO. mossambicus purebred had similar survivalpercentages (97.78±2.08 and 99.44±0.79%,respectively). An acclimated transfer at 30 ppt resultedin survival percentages of 34.45±11.00 and43.30±7.20% respective to O. niloticus and hybridNxM which were significantly lower than those ofhybrid MxN and O. mossambicus (98.33±7.20 and99.44±0.79%, respectively). Growth rate of the hybridNxM was comparable to that of Nile tilapia while thatof the hybrid NxM and Java tilapia was alsocomparable and lower than that of the first two groups.No statistical differences were observed in carcasspercentages of the four groups. Based on the directtransfer results, backcross hybrid MNxN showed thehighest salinity tolerance which was comparable tothat of O. mossambicus. No significant differences wereobserved among salinity tolerances of the remainingcrosses (NxNM, NMxN, NxMN) and O. niloticus.Carcass percentage of the backcross hybrids tendedto be higher than that of the purebreds. Differencesin some morphological traits were observed amonghybrids and parental species.
Giant freshwater prawn. The life history anddevelopment of larvae in intraspecific crosses ofM. rosenbergii from Kraburi River, Ranong province,and Chaopraya River, Ayutthaya province, werestudied (Rattikhansukha 1993). Four treatmentgroups, pure lines and their crosses, were included.Differences in egg and larval sizes, postlarvalincubation time and larval number were observed. Themorphological differences indicated that the ratiobetween dactylus of the second claw and body weightof female could be used to separate the stock with
73.24% positive prediction. The ratio between headwet weight and propodus of the second claw of malescould be used to separate the stock with 79.28%positive prediction.
Sex control
Snakeskin gourami. All-female T. pectoralis fry weresuccessfully induced by oral administration of 17β-estradiol (EST) at 200 mg/kg diet to the two-week oldpost-hatch fry for the duration of 3 weeks (Pongthanaet al. 1995a).
Catfish species. Effects of 17α-methyltestosterone (MT)on growth, survival and sex reversal of C. macrocephaluswere studied by Na-Nakorn et al. (1993). Growth wasnot affected at 30 days (P>0.05) but was retarded at60 days. Growth of treated groups was increasinglysuppressed with increased dose and duration oftreatment. Survival rate decreased at 30 days (P<0.05),but was unaffected from 31 to 60 days. Partial sexreversal was indicated by presence of ovotestes in somefish treated with MT for 60 days. In general, sexreversal to maleness was not accomplished, and withincreasing dose and duration there appeared to beparadoxical feminization.
Wattanakul (1993) experimented on the effects of 17β-estradiol and 11β-hydroxyandrostenedione on sexreversal of C. macrocephalus. Sex reversal to female wasaccomplished in the groups treated with 30 and 55mg/kg of 17β-estradiol, and not with the 5 mg/kggroup. Few intersex gonads were observed in thegroups receiving 30 and 55 mg/kg, and few sterilegonads in the 55 mg/kg group. Oral administrationof 11β-hydroxyandrostenedione (at 5, 30 and 55 mg/kg feed) to 20 and 30 days old gynogenetic fry resultedin normal growth and survival rates compared to theuntreated control group. Only the group treated with55 mg/kg at 20 and 30 days showed sex reversal.
No reduction was observed in survival, growth rateor GSI (ovarian size in relation to total body weight)relative to control groups in all female C. macrocephalusproduced through administration of 17β-estradiol.Hormone-treated fish also produced eggs which hadno apparent reduction in viability compared tocontrols in fertilization trials using sperm of C.macrocephalus and C. gariepinus (Pongthana et al. 1994).
Studies on effects of 17α -MT on sex reversal ofC. gariepinus revealed the highest female sex ratio of85.43±9.06% in three-week old post-hatch fish

Fish Genetics Research in Member Countries and Institutions of INGA84
immersed in dosages of 20 µg/l for 4 weeks. Averagesurvival rate was 55.83±5.20%. The highest male sexratio of 59.66±10.48% was observed in three-weekold post-hatch fish at dosages of 50 µg/l for 3 weeks.Average survival rate was 45.56±31.51% (Pongthanaand Tangthongpairoj 1999).
No effect on sex ratio was detected when C. gariepinus(three and four-week old post-hatch) were immersedin water treated with 11β-hydroxyandrostenedione(HAS) (Pongthana and Tangthongpairoj 1999).
Tilapia. Thassananukulkit (1979) reported inducingsex reversal using MT and increasing production ofmale O. niloticus, at 60, 40 and 20 mg/kg in diet to fryaged 3-4 weeks - male percentages of 84.5, 81.0 and72.0%, respectively, as against 57.5% in the controlgroup. Due to hormone effects, abnormal males werefound in 1.0, 3.0 and 0.5% cases, respectively, andhermaphrodites, in 2.5, 3.5 and 2.5% cases,respectively. For hermaphrodites, the ratios of thelength of the fins (dorsal, pelvic and anal) to the totalbody length were relatively close to those males. Themorphology of hermaphrodites’ genital papillae wasbetween that of male and female. The growth rates offish were significantly different among the threehormone levels and the control. The MT 40 mg/kgdiet produced the highest coefficient of condition(3.34), compared with that for other treated groups(3.16 and 3.20) and control group (2.95). Respectively,the survival rates were 96.1, 96.4, 94.7 and 97.8%;and the feed conversion rates were 1.76, 1.80, 1.61and 1.81.
Sunsetplaty. Feminization was induced by immersiontreatment of three-day old Xiphophorus variatus fry in17β-estradiol at a dose of 100 µg/l for 28 days resultingin 82.25% female fish (Kungcharoenvattana 1999).Masculinization was induced by immersion treatmentof three-day post-hatch fry in 17α-methyltestosteroneat a dose of 50 µg/l for 30 days (Pongthana et al.1999c).
Studies on the sex determination mechanismindicated that females are homogametic XX and malesare heterogametic XY. Trials to indirectly produce all-males were carried out by crossing YY males withnormal XX females resulting in average male ratio of95.37±17.97% (Pongthana et al. 1999c).
Siamese fighting fish. The effects of MT at differentconcentrations on change of morphology, sex organ,
survival rate, mating and behavior of Betta splendensfemales were studied (Suthitep 1982). A total of 1 200six-week old female fish were cultured in freshwaterwith 0.5x10-4, 1.0x10-4, 1.5x10-4, 2.0x10-4 and2.5x10-4 ppm of 17α .MT. Changes were observedin terms of body color. Compared with the control,dorsal fin was longer and body standard lengthwas shorter. The ratio between the length of dorsalfin and standard length had linear correlation to theamount of hormone applied with 99% statisticalsignificance. The survival rate decreased with theincrease of hormonal concentration. The treatedfemale behaved as an ordinary male. The develop-ment of the ovary was abnormal; the ovary lobeswere rudimentary, unequally developed andcontained abortive eggs and fluid. All treated femaleswere sterile.
Sex reversal was induced by fluoxymesterone(commercially known as Halotestin) treatments(Tangthongpairoj et al. 1988). Three-day old fry, fedwith Moina sp. immersed in 200 ppm Halotestin for20 min and treated for 14 days, were morphologicallymale as distinguished by color and fin shapes. Theirgonads couldn’t develop to the secondary oocyte stage.
Marine shrimps. Study on the effects of exogenous malehormone, 17α-MT and testosterone proprionate onspermatogenesis and microscopic structures ofP. indicus showed that 17α-MT at a dose of 1 mg/100gbody weight is able to maintain spermatogenesissignificantly (Chansawang 1994).
Genetic manipulation
Gynogenesis
Silver barb. Neomale or XX-male (genetically femalebut functionally male) silver barb (B. gonionotus) wereproduced by hormonally masculinized gynogeneticdiploid fry using 17α-MT at 25 mg/kg in the diet, fora duration of 4 or 5 weeks, starting from two weekspost-hatch. Gynogenetic diploid fish was induced byUV irradiation of sperm at 196 uW/cm2, cold shockingof 2°C for 10 min, starting from 1.5 min afterfertilization (Pongthana et al. 1995b). The neomalesproduced all female offspring when crossed withordinary females (Pongthana et al. 1999d).
Catfish species. Induction of gynogenesis inC. macrocephalus was studied by Leuangpromporn(1990). Irradiation of P. sutchi sperm using either 15 or

85
30 watts UV–tube at distance of either 20 or 30 cm(intensity of 201.38-604.14 erg/mm2–second) and 2 or7 min exposure could inactivate sperm resulting inthe highest hatching rates of haploid gynogenetic fry.Experiments to study the effects of temperature, timeafter insemination and shock duration onpolyploidization in C. macrocephalus showed triploidfish (3n = 81) in the group, which was shocked at 7°Cfor 20 min. In C. macrocephalus, gynogenesis could beinduced using heterologous sperms irradiated with UV–ray. The hatching and survival rates of diploidgynogenetic fish were low especially from sperm treatedwith 30 watts UV–tube and 20 cm distance (intensityof 604.14 erg/mm2–second). Gynogenetic fish of C.macrocephalus obtained from the group using lowest doseof UV–ray (15 watts, 20 cm and 2 min) grew faster thanthe others, including hybrid fish. This was probablydue to paternal effect.
Gynogenetic diploid in C. macrocephalus was inducedby UV irradiation of sperm at 4.8x104 ergs/mm2, coldshocking the fertilized eggs at 7°C for 14 min startingfrom 4.5 min after fertilization which resulted in 60%viable diploid gynogenetic fry (Na-Nakorn et al. 1993).
Rungsin (1994) studied the optimum condition forinduction of diploid gynogenesis in C. macrocephalusby retention of second polar body. UV–irradiatedsperms of P. sutchi was used to activate the eggs.Cold shock (7°C for 20 min) was most effective whenapplied to activated eggs at 3.5-5 min after mixingeggs with irradiated sperm. Cold shock at 8-10°C(initiated 4.5 min after activation, lasting 20 min) wasequally effective and gave satisfactory hatching ofdiploid gynogenesis. Shock duration of 14 min wasfound to be optimum to induce diploidization ofgynogenesis (at shock temperature of 10°C, initiated4.5 min after activation).
Diploid meiot ic gynogens were induced in C.macrocephalus aimed at creating all female stocks(Na-Nakorn 1995). Induction was performed using UV-irradiated sperm of P. sutchi followed by either cold orheat shock. Cold shock (7°C, lasting 14 min) gavesignificant higher percentage of meiotic diploid frythan that of heat shock. Six-month old meioticgynogens were all female and showed slightly inferiorgrowth to male and female control. Survivals of thegynogens (73%) were slightly inferior to the mixed sexcontrol (87.3%). The gonadosomatic index ofthe gynogens and the control female were 5.66 and8.82, respectively.
Gynogenesis was induced in C. macrocephalus by mixingeggs with UV-irradiated sperms of P. sutchi followingeither cold or heat shock (Vichsonkul 1996). Coldshock at 6-11°C (initiated 4.5 min after activation,lasting 4 min) was equally effective and gave satisfactoryhatching rates of diploid gynogenesis (30.65-48.24%).Heat shock at 42°C for 1 min was more effective thanother high temperature treatments (35, 37, 40 and 42°Cfor 1-7 min). Heat shock (42°C for 1 min) was mosteffective when applied to activated eggs at 3.5-5.5 minafter mixing eggs with UV-irradiated sperms. Coldshock (7°C for 14 min) gave significantly higherpercentage (2.52-10.88%) of diploid gynogenesis(13.40-30.47%) than the heat shock (42°C for 1 min).
Ploidy manipulation
Generally, it is believed that polyploid fish has highergrowth rate and stronger adaptability than those ofdiploid. The techniques of inducing polyploid werestudied in catfish and oysters.
Catfish species. Polyploid in C. macrocephalus was inducedby cold shock at 7°C for 25 min startingafter fertilization, resulting in 80% polyploidy(Na-Nakorn and Legrand 1992). Survival rates ofdiploid and triploid fish were not significantly differentin the first 2 months. But in the third, fourth and fifthmonths, triploid fish showed lower survival rates andbody weight compared to diploid group. Foodconversion ratio (FCR) of diploid was better than thatof triploid in the first month. Both diploid and triploidfish showed similar FCR in the second and fourthmonths. There was no significant difference betweenboth treatments as regards carcass percentages andresistance to Aeromonas hydrophila (Lakhaanantakun1992).
Oysters. Polyploidy was induced in small oyster(S. cucullata) by cold shock at 4°C for 6 and 15 minstarting from 20 min after fertilization, resulting in40% polyploid (Jarayabhand and Jindalikit 1992).Triploidy was successfully induced in the mangroveoyster (C. lugubris) with 0.1 mg/l cytochalasin B for 15min starting 15 min after fertilization resulting in 77%triploids (Roongratri and Youngvanichset 1988).
Polyploidy was induced in C. belcheri, by thermalshock (Silapajarn 1994). The polyploidizationusing heat shock at 35, 38, 40 and 42°C with 5 and10 min treatment duration 10, 30 and 40 min afterinsemination had average triploidy of around
Aquaculture Genetic Research in Thailand

Fish Genetics Research in Member Countries and Institutions of INGA86
0-43.3%, and survival at the straight hinge stagewas around 1.19-27.85%. The lowest polyploidywas achieved at shocking temperature of 35°C (P<0.01),while the lowest survival rate occurred at 42°C(P<0.01). The cold shock at 3, 7 and 10°C seawatertemperature, 10, 30 and 40 min after inseminationwith 15 and 20 min treatment duration increased thetriploidy to 8.45-35.28% and survival rate at the straighthinge stage was 0-20.3%. The lowest polyploid wasobtained at 10°C (P<0.01). The survival rate of treatedlarvae declined with the decrease in shockingtemperature. Oyster treated with heat shock at 35°Cstarting 30 min after insemination for 5 and 10 minperformed better in terms of survival at the straighthinge stage and of growth at the setting stage (P<0.05).
Cytogenetic studies
Cytogenetic studies (chromosome number, karyotypeand arm number) have been carried out in somefreshwater fish species (Table 3).
Cryopreservation
Catfish species
Chairak (1996) studied cryopreservation of milt ofC. macrocephalus. 0.85% NaCl (282 mOsm/kg, pH 7.0)and modified Cortland’s #1 solution (MC#1-2, 292mOsm/kg pH 7.0) were suitable extenders in terms ofsperm motility and fertilizing ability. These twoextenders were used for studying the toxicity(decreasing percentage of motile sperm) of eitherglycerol, dimethylsulfoxide (DMSO) or methanol atthe final concentrations of 0, 6, 8, 10, 12, 14 and 16%in both extenders and motility of the sperm wasdetermined at 30, 60 and 120 min after diluting andstoring. Glycerol at all concentrations elicited highertoxicity than DMSO and methanol. Freezing spermin 8, 10 or 12% DMSO and 6, 10 or 14% methanolbefore storing in liquid nitrogen by the programmablefreezing unit at rates of -10, -20 and -30°C/min showedthat DMSO was a better cryoprotectant than methanol.In another experiment, 8, 10 and 12% DMSO was usedto freeze the sperm in above liquid nitrogen vapor atrates of -5 and -10°C/min. The results indicated nosignificant difference among treatments and betweentwo thawing temperatures (50 or 70°C).
Red snapper
Hirunchaiyapuck (1996) reported that spermcharacteristics of Lutjanus argentimaculatus are similar
to most teleosts with round head and lack of acrosome.Average sperm density was 20.94 x 106 cells/ml withsperm motility ratings between 9 and 10. Averagesperm motility and live sperm detected by Eosin-Nigrosin stain were 50.99% and 69.43%, respectively.The effect of dilulent on sperm preservation was notsignificant (P=0.233). However, modified Cortland’ssolution, Alsever’s solution and glucose normal salinesolution as dilulent gave higher percentage of livesperm than others. The cryoprotectant (DMSO) gavea higher percentage of live sperm than glycerol (P <0.001). A study on freezing methods was alsoconducted. The pelleting method resulted in thehighest percentage of live sperm (P<0.001), comparedwith the rapid and the slow freezing methods. Therewas no significant difference among thawing methods,i.e., thawing in low temperature, room temperature andhigh temperature (60°C) (P = 0.215). Alsever’s solution+ 10% DMSO freezing by pelleting method andthawing in 60°C is the best procedure for spermcryopreservation, resulting in 75.67±10.89% of livesperm.
Genetic engineering
Catfish
Transfer of growth hormone gene into fertilized eggof C. macrocephalus by microinjection was carried outby Boonanuntanasarn (1995). Fish derived frommicroinjected egg had significantly higher growth rate(P<0.05). Integration rate of introduced gene in one,two and four-cell stages microinjected fish was 5.05%(5 of 99 samples), 6.45% (2 of 31 samples) and 8.5% (1of 12 samples), respectively. Detection of theintroduced gene in various tissues of microinjected fishindicated mosaicism in various tissues of fish, derivedfrom all stages of microinjected eggs. This studyrevealed that embryonic development of yellow C.macrocephalus at one and two-cell stages were appropriatefor gene transferring by microinjection.
Future Plans
Following are the plans for aquaculture genetics researchin Thailand:• Set up national breeding plans for all hatchery-
produced aquatic species.• Develop high-yielding strains of all hatchery-
produced aquatic species.• Study the genetic diversity of aquatic species in
Thailand.• Develop a genebank for germplasm preservation
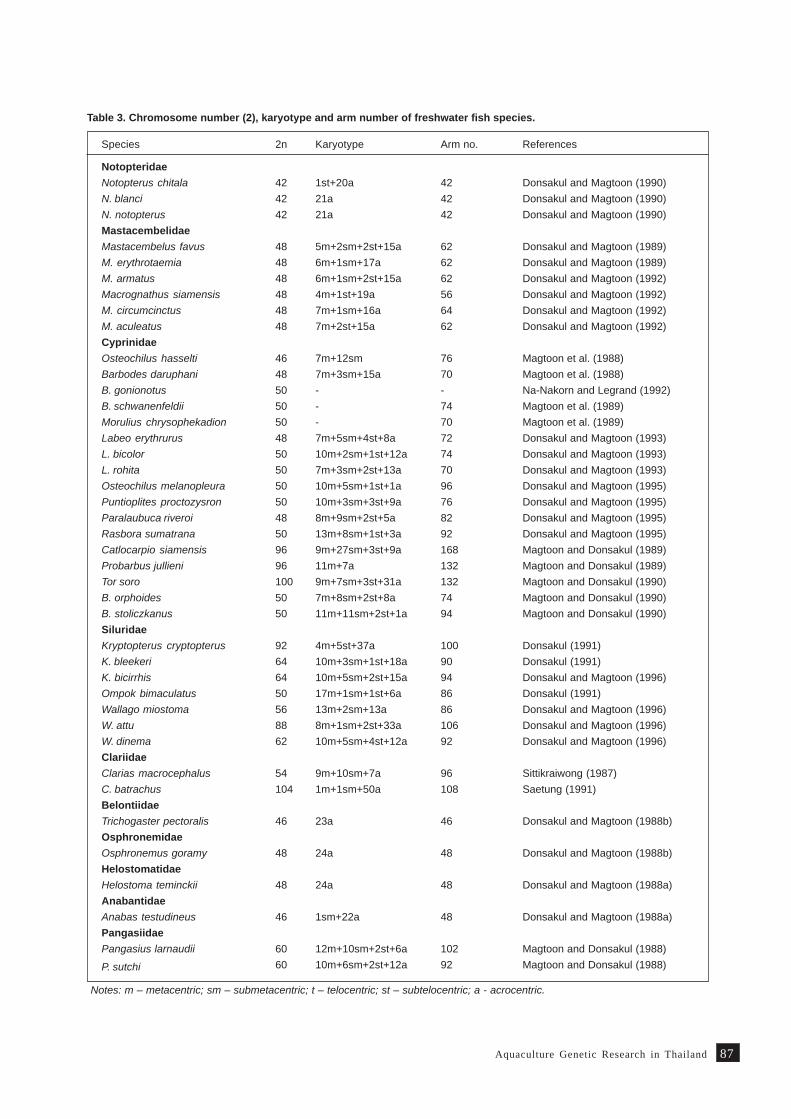
87Aquaculture Genetic Research in Thailand
2n
424242
484848484848
46485050504850505050485096961005050
92646450568862
54104
46
48
48
46
6060
Karyotype
1st+20a21a21a
5m+2sm+2st+15a6m+1sm+17a6m+1sm+2st+15a4m+1st+19a7m+1sm+16a7m+2st+15a
7m+12sm7m+3sm+15a---7m+5sm+4st+8a10m+2sm+1st+12a7m+3sm+2st+13a10m+5sm+1st+1a10m+3sm+3st+9a8m+9sm+2st+5a13m+8sm+1st+3a9m+27sm+3st+9a11m+7a9m+7sm+3st+31a7m+8sm+2st+8a11m+11sm+2st+1a
4m+5st+37a10m+3sm+1st+18a10m+5sm+2st+15a17m+1sm+1st+6a13m+2sm+13a8m+1sm+2st+33a10m+5sm+4st+12a
9m+10sm+7a1m+1sm+50a
23a
24a
24a
1sm+22a
12m+10sm+2st+6a10m+6sm+2st+12a
Species
NotopteridaeNotopterus chitalaN. blanciN. notopterusMastacembelidaeMastacembelus favusM. erythrotaemiaM. armatusMacrognathus siamensisM. circumcinctusM. aculeatusCyprinidaeOsteochilus hasseltiBarbodes daruphaniB. gonionotusB. schwanenfeldiiMorulius chrysophekadionLabeo erythrurusL. bicolorL. rohitaOsteochilus melanopleuraPuntioplites proctozysronParalaubuca riveroiRasbora sumatranaCatlocarpio siamensisProbarbus jullieniTor soroB. orphoidesB. stoliczkanusSiluridaeKryptopterus cryptopterusK. bleekeriK. bicirrhisOmpok bimaculatusWallago miostomaW. attuW. dinemaClariidaeClarias macrocephalusC. batrachusBelontiidaeTrichogaster pectoralisOsphronemidaeOsphronemus goramyHelostomatidaeHelostoma teminckiiAnabantidaeAnabas testudineusPangasiidaePangasius larnaudii
P. sutchi
Arm no.
424242
626262566462
7670-7470727470967682921681321327494
1009094868610692
96108
46
48
48
48
10292
References
Donsakul and Magtoon (1990)Donsakul and Magtoon (1990)Donsakul and Magtoon (1990)
Donsakul and Magtoon (1989)Donsakul and Magtoon (1989)Donsakul and Magtoon (1992)Donsakul and Magtoon (1992)Donsakul and Magtoon (1992)Donsakul and Magtoon (1992)
Magtoon et al. (1988)Magtoon et al. (1988)Na-Nakorn and Legrand (1992)Magtoon et al. (1989)Magtoon et al. (1989)Donsakul and Magtoon (1993)Donsakul and Magtoon (1993)Donsakul and Magtoon (1993)Donsakul and Magtoon (1995)Donsakul and Magtoon (1995)Donsakul and Magtoon (1995)Donsakul and Magtoon (1995)Magtoon and Donsakul (1989)Magtoon and Donsakul (1989)Magtoon and Donsakul (1990)Magtoon and Donsakul (1990)Magtoon and Donsakul (1990)
Donsakul (1991)Donsakul (1991)Donsakul and Magtoon (1996)Donsakul (1991)Donsakul and Magtoon (1996)Donsakul and Magtoon (1996)Donsakul and Magtoon (1996)
Sittikraiwong (1987)Saetung (1991)
Donsakul and Magtoon (1988b)
Donsakul and Magtoon (1988b)
Donsakul and Magtoon (1988a)
Donsakul and Magtoon (1988a)
Magtoon and Donsakul (1988)Magtoon and Donsakul (1988)
Table 3. Chromosome number (2), karyotype and arm number of freshwater fish species.
Notes: m – metacentric; sm – submetacentric; t – telocentric; st – subtelocentric; a - acrocentric.

Fish Genetics Research in Member Countries and Institutions of INGA88
of aquatic species.• Distribute genetically improved broodstocks and
seeds to public and private sectors.
References
Boonanuntanasarn, S. 1995. Transfer of growth hormone geneinto fertilized egg of catfish, Clarias macrocephalus, bymicroinjection. Chulalongkorn University, Bangkok,Thailand. 150 p. M.S. thesis.
Boonyawan, B. 1998. Sequence characterization of randomlyisolated cDNA from black tiger prawn (Penaeus monodon).Mahidol University, Bangkok, Thailand. M.S. thesis.
Chairak, N. 1996. Cryopreservation of milt of Gunther’s walkingcatfish, Clarias macrocephalus Gunther. Kasetsart University,Bangkok, Thailand. 142 p. M.S. thesis.
Chamnankuruwet, A. 1996. Response to bidirectional massselection for growth rate in Clarias macrocephalus Gunther.Kasetsart University, Bangkok, Thailand. 61 p. M.S. thesis.
Chansawang, N. 1994. Effects of 17α -methyltestosterone andtestosterone–proprionate on spermatogenesis of giant tigerprawn (Penaeus monodon Fabricuis). Kasetsart University,Bangkok, Thailand. M.S. thesis.
Charoensit, P. 1995. Hybridization in commercial oyster.Chulalongkorn University, Bangkok, Thailand. 115 p. M.S.thesis.
Donsakul, T. 1991. A chromosome study on three species of Siluridfishes, Kryptopterus cryptopterus, K. bleekeri and Ompokbimaculatus of Thailand . Tech. Pap. Agric. Biol.Srinakharinwirot University, Thailand.
Donsakul, T. and W. Magtoon. 1988a. A chromosome study ontwo species of Anabantid fishes, Anabas testudineus andHelostoma temminckii (Cuvier and Valenciennes). Tech. Pap.Agric. Biol. Srinakharinwirot University, Thailand.
Donsakul, T. and W. Magtoon. 1988b. A chromosome study ontwo species of Anabantid fishes, Trichogaster pectoralis andOsphronemus gouramy. Tech. Pap. Agric. Biol. SrinakharinwirotUniversity, Thailand.
Donsakul, T. and W. Magtoon. 1989. A chromosome study on twospecies of Mastacembelid fishes, Mastacembelus favus and M.erythrotaemia from Thailand . Tech. Pap. Agric. Biol.Srinakharinwirot University, Thailand.
Donsakul, T. and W. Magtoon. 1990. A chromosome study on threespecies of Notopterid fishes, Notopterus notopterus, N. chitalaand N. blanci from Thailand . Tech. Pap. Agric. Biol.Srinakharinwirot University, Thailand.
Donsakul, T. and W. Magtoon. 1992. A chromosome study on fourspecies of Mastacembelid fishes, Macrognathus siamensis, M.circumcinctus, M. aculeatus and Mastacembelus armatus fromThailand. Tech. Pap. Agric. Biol. Srinakharinwirot University,Thailand.
Donsakul, T. and W. Magtoon. 1993. A chromosome study of threeCyprinid fishes, Labeo erythrurus, L. bicolor and L. rohita,p.221-226. In Proceedings of the Thirty-first KasetsartUniversity Annual Conference, Bangkok, Thailand.
Donsakul, T. and W. Magtoon. 1995. Karyotypes of four Cyprinidfishes, Osteochilus melanopleura, Puntioplites proctozysron,Paralaubuca riveri and Rasbora sumatrana. In Proceedings ofthe Thirty-third Kasetsart University Annual Conference,Bangkok, Thailand.
Donsakul, T. and W. Magtoon. 1996. A chromosome study onfour species of Silurid fishes, Wallago miostoma, W. attu, W.dinema and Kryptopterus bicirihis. In Proceedings of theThirty-fourth Kasetsart University Annual Conference,Bangkok, Thailand.
Hirunchaiyapuck, W. 1996. Sperm cryopreservation of red snapper
Lutjanus argentimaculatus Forsskal, 1755. ChulalongkornUniversity, Bangkok, Thailand. 76 p. M.S. thesis.
Jarayabhand, P. and Jindalikit. 1992. Chromosome manipulationin small oyster, Saccostrea cucullata by thermal shock: evidencesfrom trochophore larvae, p.85-91. In Proceedings of the FirstInternational Workshop on Genetics in Aquaculture andFisheries Management.
Jarimopas, P., A. Kumnan and J. Wongchan. 1989. Mass selectionof Clarias macrocephalus Gunther for growth (fourgenerations), p. 177-191. In Fish genetics final report (PhaseII). International Development Research Centre and NationalAquaculture Genetics Research Institute, Department ofFisheries, Thailand.
Kamonrat, W. 1996. Spatial genetic structure of Thai silver barbPuntius gonionotus (Bleeker) populations in Thailand.Dalhousie University, Halifax, Nova Scotia, Canada. 193 p.Ph. D. dissertation.
Kungcharoenvattana, S. 1999. Sex control in sunsetplaty(Xiphophorus variatus, Meek, 1904). Kasetsart University,Bangkok, Thailand. 79 p. M.S. thesis.
Lakhaanantakun, A. 1992. The effects of triploidy on survival rate,growth rate and feed conversation ratio of walking catfish(Clarias macrocephalus Gunther). Kasetsart University,Bangkok, Thailand. 74 p. M.S. thesis.
Leeprasert, K. 1987. Genetic parameters of some quantitative traitsin Pangasius sutchi Fowler. Kasetsart University, Bangkok,Thailand. 75 p. M.S. thesis.
Leuangpromporn, W. 1990. Induction of gynogenesis in Clariasmacrocephalus Gunther. Kasetsart University, Bangkok,Thailand. M.S. thesis.
Magtoon, W. and T. Donsakul. 1988. Karyotypes of Pangasiidcatfishes, Pangasius larnaudii and P. sutchi. In Proceedings ofthe Twenty-sixth Kasetsart University Annual Conference,Bangkok, Thailand.
Magtoon, W. and T. Donsakul. 1989. Karyotypes of two Cyprinidfishes, Catlocarpio siamensis (Boulenger) and Probarbus jullieni(Sauvage). Tech. Pap. Agric. Biol. Srinakharinwirot University,Thailand.
Magtoon, W. and T. Donsakul. 1990. Morphology and karyotypesof Cyprinid fishes, Tor soro, Puntius orphoides and P. stoliczkanusin Pai River, Mae Hong Son, North, Thailand. Tech. Pap.Agric. Biol. Srinakharinwirot University, Thailand.
Magtoon, W., T. Donsakul and Y. Taki. 1989. Karyotype of Cyprinidfishes, P. schwanenfeldii and Morulius chrysophekadion. InProceedings of the Twenty-seventh Kasetsart UniversityAnnual Conference, Bangkok, Thailand.
Magtoon, W., U. Thipsana and Y. Taki. 1988. Karyotype ofCyprinid fishes, Osteochilus hasseltii and Puntius daruphani. InProceedings of the Twenty-sixth Kasetsart University AnnualConference, Bangkok, Thailand.
Meesukko, C. 1996. Karyotype of freshwater Amblemid musselsin Yom and Nan watersheds. Chulalongkorn University,Bangkok, Thailand. 84 p. M.S. thesis.
Na-Nakorn, U. 1995. Comparison of cold and heat shock to inducediploid gynogenesis in Thai walking catfish (Clariasmacrocephalus). In Proceedings of the Thirty-third KasetsartUniversity Annual Conference, Bangkok, Thailand.
Na-Nakorn, U. and E. Legrand. 1992. Induction of triploidy inPuntius gonionotus (Bleeker) by cold shock. Kasetsart Univ.Fish. Res. Bull. 18: 10 p.
Na-Nakorn, U., R.A. Dunham and P. Tabthipwon. 1993. Effectsof 17 Alpha-Methyltestosterone on growth, survival and sexreversal of Thai walking catfish, Clarias macrocephalus. InProceedings of the Thirty-first Kasetsart University AnnualConference, Bangkok, Thailand.
Nukwan, S. 1987. Preliminary study on genetic parameters ofbighead carp Aristichthys nobilis (Richardson). KasetsartUniversity, Bangkok, Thailand. 69 p. M.S. thesis.

89Aquaculture Genetic Research in Thailand
Pongsomboon, S. 1996. Detection of genetic variation inpopulations of black tiger prawn Penaeus monodon by DNAfingerprinting. Chulalongkorn University, Bangkok,Thailand. M.S. thesis.
Pongsri, C. 1995. Biochemical characterization of blood-serumprotein of tilapias by SDS- polyacrylamide gel electrophoresis.In Proceedings of the Thirty-third Kasetsart UniversityAnnual Conference, Bangkok, Thailand.
Pongthana, N. and M. Tangthongpairoj. 1999. Effects of 17α -methyltestosterone and 11β-hydroxyandrostenedione on sexreversal of the African catfish, Clarias gariepinus (Burchell,1822). Tech. Pap. No. 24, 16 p. National Aquaculture GeneticsResearch Institute, Department of Fisheries, Thailand.
Pongthana, N., B. Baoprasertkul and B. Tongmee. 1995a.Hormonal feminization of snakeskin gourami (Trichogasterpectoralis). Thai Fish. Gaz. 48(4): 303-318.
Pongthana, N., K. Amponsak and P. Baoprasertkul. 1999a. Geneticcharacteristics and performances among populations ofoyster (Crassostrea belcheri, Sowerby) along the coasts ofThailand. In Proceedings of the Fifth Asian Fisheries Forum,Special Session on Molluscs Research in Asia.
Pongthana, N., S. Prasertwiriyakul and P. Baoprasertkul. 1999b.Population genetic structures of the snakeskin gourami,Trichogaster pectoralis (Regan, 1910) in Thailand. Tech. Pap.No. 22, 59 p. National Aquaculture Genetics ResearchInstitute, Department of Fisheries, Thailand.
Pongthana, N., D.J. Penman, J. Karnasuta and B.J. McAndrew.1995b. Induced gynogenesis in the silver barb (Puntiusgonionotus Bleeker) and evidence for female homogamety.Aquaculture 135: 267-276.
Pongthana, N., P. Baoprasertkul, U. Ruangnarong and M. Niroth.1994. All-female broodstock production of Pla duk oui(Clarias macrocephalus). Tech. Pap. No. 6, 17 p. NationalAquaculture Genetics Research Institute, Department ofFisheries, Thailand.
Pongthana, N., S. Kungcharoenwattana, P. Baoprasertkul and M.Niroth. 1999c. Sex determination system and monosexproduction in sunsetplaty (Xiphophorus variatus, Meek, 1904).Tech. Pap. No. 22, 26 p. National Aquaculture GeneticsResearch Institute, Department of Fisheries, Thailand.
Pongthana, N., D.J. Penman, P. Baoprasertkul, M.G. Hussain, M.S.Islam, S.F. Powell and B.J. McAndrew. 1999d. Monosex femaleproduction in the silver barb (Puntius gonionotus Bleeker).Aquaculture 173: 247-256.
Prakongsai, S. 1991. Development of DNA probe for the RELPanalysis of DNA from Macrobrachium rosenbergii (de Man).Mahidol University, Bangkok, Thailand. M.S. thesis.
Prarom, W. 1990. The effect of strain crossing of Gunther’s walkingcatfish (Clarias macrocephalus) on growth and diseaseresistance. Kasetsart University, Bangkok, Thailand. M.S.thesis.
Rasatapana, D. 1996. Estimation of heritability of diseaseresistance, body weight and body length of Thai walkingcatfish, Clarias macrocephalus. Kasetsart University, Bangkok,Thailand. M.S. thesis.
Rattikhansukha, C. 1993. Intraspecific hybridization inMacrobrachium rosenbergii de Man. Chulalongkorn University,Bangkok, Thailand. M.S. thesis.
Roongratri, N. and K. Youngvanichset. 1988. Triploidy inductionin the mangrove oyster (Crassostrea lugubris) with cytochalasinB. Thai Fish. Gaz. 44(3): 223-227.
Rungsin, W. 1994. Study on optimum time after insemination,temperature and shock duration for induction of gynogenesisin walking catfish (Clarias macrocephalus Gunther). KasetsartUniversity, Bangkok, Thailand. M.S. thesis.
Saetung, C. 1991. The effects of cold shock on polyploidizationin Clarias batrachus Linn. Kasetsart University, Bangkok,Thailand. M.S. thesis.
Silapajarn, K. 1994. The study on polyploidy induction in Takromoyster Crassostrea belcheri (Sowerby) by thermal shock. MahidolUniversity, Bangkok, Thailand. M.S. thesis.
Sittikraiwong, P. 1987. Karyotype of the hybrid between Clariasmacrocephalus Gunther and Pangasius sutchi Fowler. KasetsartUniversity, Bangkok, Thailand. M.S. thesis.
Sodsuk, S. 1996. Genetic differentiation and population structureof Penaeus monodon in Thailand. Tech. Pap. No. 12, 23 p.National Aquaculture Genetics Research Institute,Department of Fisheries, Thailand.
Sodsuk, S. and P.K. Sodsuk. 1998a. Genetic diversity of Bananashrimp from three locations in Thailand. Tech. Pap. No. 17,45 p. National Aquaculture Genetics Research Institute,Department of Fisheries, Thailand.
Sodsuk, S. and P.K. Sodsuk. 1998b. Genetic diversity of giantfreshwater prawn from three locations in Thailand. Tech. Pap.No. 18, 40 p. National Aquaculture Genetics ResearchInstitute, Department of Fisheries, Thailand.
Supsumrarn, M. 1987. Preliminary study on genetics of Labeo rohita(Boche). Kasetsart University, Bangkok, Thailand. M.S. thesis.
Suthitep, P. 1982. Effects of methyltestosterone on morphologicalchanges of female Siamese fighting fish (Betta splendensRegan). Chulalongkorn University, Bangkok, Thailand. M.S.thesis.
Tangthongpairoj, M., K. Lawanyawut, S. Nukwan and P.Chanwatchanakul. 1988. Induced sex reversal in Bettasplendens by fluoxymesterone treatments. Thai Fish. Gaz.41(6): 25-32.
Thanakijkorn, J. 1997. Improvement of salinity tolerance in Niletilapia (Oreochromis niloticus) by means of interspecific-hybridization with Java tilapia (Oreochromis mossambicus).Kasetsart University, Bangkok, Thailand. M.S. thesis.
Thassananukulkit, W. 1979. Use of methyltestosterone for inducedsex reversal and increasing production of Nile tilapia, Tilapianilotica Linn. Kasetsart University, Bangkok, Thailand. M.S.thesis.
Thavornyutikarn, M. 1994. Realized heritability estimation ongrowth of oyster Saccostrea cucullata. ChulalongkornUniversity, Bangkok, Thailand. M.S. thesis.
Thongpan, A., M. Mingmuang, S. Thinchant, R.K. Cooper, T.R.Tiersch and K. Mongkonpunya. 1994. Genomic identificationof catfish species by polymerase chain reaction and restrictionenzyme analysis of the gene encoding the immunoglobulinM heavy chain constant region. In Proceedings of the Thirty-second Kasetsart University Annual Conference, Bangkok,Thailand.
Tiptawonnukul, A. 1996. Characterization of microsatellite DNAin giant tiger prawn Penaeus monodon genome. ChulalongkornUniversity, Bangkok, Thailand. M.S. thesis.
Uraiwan, S., R.W. Doyle and M. Meewan. 1992. Evaluation ofresponse to within-family selection of Oreochromis niloticus(Linn.) by line performance growth test. In Proceedings ofthe Third Asian Fisheries Forum, Manila, Philippines.
Vichsonkul, S. 1996. Efficiency of cold shock and heat shock ininducing diploid gynogenesis of Clarias macrocephalus .Kasetsart University, Bangkok, Thailand. M.S. thesis.
Yaitavorn, P. 1989. Mitochondrial DNA variation in giantfreshwater prawn (Macrobrachium rosenbergii de Man).Chulalongkorn University, Bangkok, Thailand. M.S. thesis.

Fish Genetics Research in Member Countries and Institutions of INGA90

91Review of Fish Genetics and Breeding Research in Vietnam
ABSTRACT
A series of studies on fish genetics and breeding programs were conducted in Vietnam in 1998. These programsincluded genetic control and manipulation of sex ratio; production of monosex Oreochromis aureus and Barbodesgonionotus; chromosome analysis of Macrobrachium rosenbergii; induction of triploids in Clarias macrocephalus;generating transgenic Carassius auratus; dechorionation of one-cell stage embryos for DNA microinjection intoMisgurnus anguillicaudatus; and breeding of Cyprinus carpio, O. niloticus and B. gonionotus. Details of the resultsof these programs are presented below.
REVIEW OF FISH GENETICSAND BREEDING RESEARCH
IN VIETNAM
T.M. Thien, N.C. Dan and P.A. TuanResearch Institute for Aquaculture No.1
Bac Ninh, Vietnam
THIEN, T.M., N.C. DAN and P.A. TUAN. 2001. Review of fish genetics and breeding research in Vietnam, p. 91-96. In M.V.Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of the InternationalNetwork on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
The Ministry of Fisheries (MOF, 1998) estimated thetotal fish production in Vietnam in 1998 at 1.67million t, of which 538 000 t came from aquaculture,which contributed nearly 35% of total animal proteinintake of the nation.
In Vietnam, traditional fish culture has been practicedas early as hundreds of years ago (Dan et al. 1997).Fish culture in ricefields and village ponds was firstpromoted by rice farmers to supplement theirnutritional requirement. The main species culturedare local carps and other indigenous fish. During the1970s and 1980s, with the introduction of exotic fishspecies from other countries and the success ofinduced fish breeding, aquaculture production in thecountry very much improved (Thien and Dan 1997).From 1990 to present, with the development of shrimpindustry and sea farming, aquaculture contributes
considerably to fish production for domesticconsumption and export. It has become an importanteconomic sector of the country.
Aquaculture is moving from extensive to semi-intensive culture system and commercial scale. Themain components of production, such as quality ofseed, feed and fish health management, need to beimproved. Of these, seed quality is the most importantfactor to enhance productivity. Unfortunately, thedeterioration of genetic quality in several cultured fishspecies has been very striking since the late 1970s(Thien 1996). Genetics research should be undertakento create high-quality strains, particularly of valuablespecies, to improve aquaculture production andupgrade market value of the products. In recent years,the government has provided more funding for researchon selective breeding and genetic technology. However,due to lack of qualified human resources andinstitutions, the research programs are limited, scattered
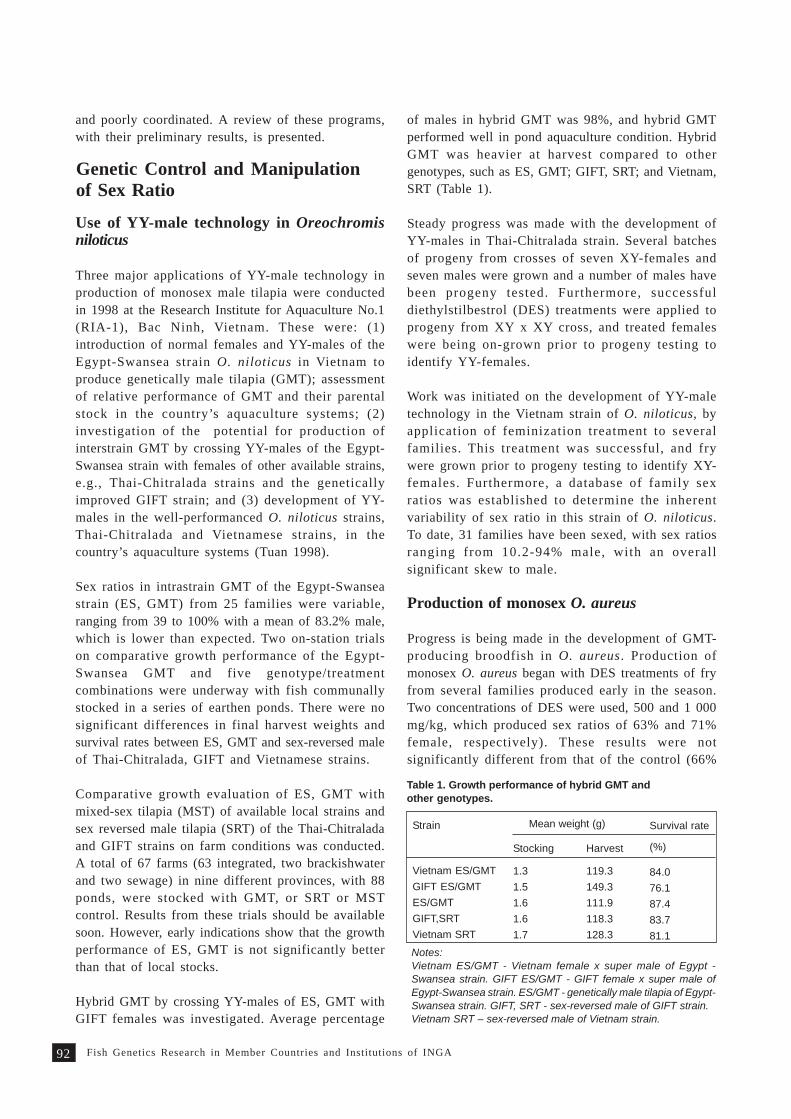
Fish Genetics Research in Member Countries and Institutions of INGA92
and poorly coordinated. A review of these programs,with their preliminary results, is presented.
Use of YY-male technology in Oreochromisniloticus
Three major applications of YY-male technology inproduction of monosex male tilapia were conductedin 1998 at the Research Institute for Aquaculture No.1(RIA-1), Bac Ninh, Vietnam. These were: (1)introduction of normal females and YY-males of theEgypt-Swansea strain O. niloticus in Vietnam toproduce genetically male tilapia (GMT); assessmentof relative performance of GMT and their parentalstock in the country’s aquaculture systems; (2)investigation of the potential for production ofinterstrain GMT by crossing YY-males of the Egypt-Swansea strain with females of other available strains,e.g., Thai-Chitralada strains and the geneticallyimproved GIFT strain; and (3) development of YY-males in the well-performanced O. niloticus strains,Thai-Chitralada and Vietnamese strains, in thecountry’s aquaculture systems (Tuan 1998).
Sex ratios in intrastrain GMT of the Egypt-Swanseastrain (ES, GMT) from 25 families were variable,ranging from 39 to 100% with a mean of 83.2% male,which is lower than expected. Two on-station trialson comparative growth performance of the Egypt-Swansea GMT and five genotype/treatmentcombinations were underway with fish communallystocked in a series of earthen ponds. There were nosignificant differences in final harvest weights andsurvival rates between ES, GMT and sex-reversed maleof Thai-Chitralada, GIFT and Vietnamese strains.
Comparative growth evaluation of ES, GMT withmixed-sex tilapia (MST) of available local strains andsex reversed male tilapia (SRT) of the Thai-Chitraladaand GIFT strains on farm conditions was conducted.A total of 67 farms (63 integrated, two brackishwaterand two sewage) in nine different provinces, with 88ponds, were stocked with GMT, or SRT or MSTcontrol. Results from these trials should be availablesoon. However, early indications show that the growthperformance of ES, GMT is not significantly betterthan that of local stocks.
Hybrid GMT by crossing YY-males of ES, GMT withGIFT females was investigated. Average percentage
of males in hybrid GMT was 98%, and hybrid GMTperformed well in pond aquaculture condition. HybridGMT was heavier at harvest compared to othergenotypes, such as ES, GMT; GIFT, SRT; and Vietnam,SRT (Table 1).
Steady progress was made with the development ofYY-males in Thai-Chitralada strain. Several batchesof progeny from crosses of seven XY-females andseven males were grown and a number of males havebeen progeny tested. Furthermore, successfuldiethylstilbestrol (DES) treatments were applied toprogeny from XY x XY cross, and treated femaleswere being on-grown prior to progeny testing toidentify YY-females.
Work was initiated on the development of YY-maletechnology in the Vietnam strain of O. niloticus, byapplication of feminization treatment to severalfamilies. This treatment was successful, and frywere grown prior to progeny testing to identify XY-females. Furthermore, a database of family sexratios was established to determine the inherentvariability of sex ratio in this strain of O. niloticus.To date, 31 families have been sexed, with sex ratiosranging from 10.2-94% male, with an overallsignificant skew to male.
Production of monosex O. aureus
Progress is being made in the development of GMT-producing broodfish in O. aureus. Production ofmonosex O. aureus began with DES treatments of fryfrom several families produced early in the season.Two concentrations of DES were used, 500 and 1 000mg/kg, which produced sex ratios of 63% and 71%female, respectively). These results were notsignificantly different from that of the control (66%
Table 1. Growth performance of hybrid GMT andother genotypes.
Strain
Vietnam ES/GMTGIFT ES/GMTES/GMTGIFT,SRTVietnam SRT
Mean weight (g) Survival rate
(%)
84.076.187.483.781.1
Stocking Harvest
1.3 119.31.5 149.31.6 111.91.6 118.31.7 128.3
Notes:Vietnam ES/GMT - Vietnam female x super male of Egypt -Swansea strain. GIFT ES/GMT - GIFT female x super male ofEgypt-Swansea strain. ES/GMT - genetically male tilapia of Egypt-Swansea strain. GIFT, SRT - sex-reversed male of GIFT strain.Vietnam SRT – sex-reversed male of Vietnam strain.
Genetic Control and Manipulationof Sex Ratio

93Review of Fish Genetics and Breeding Research in Vietnam
female), indicating poor rates of sex reversal. Furtherprogenies were treated with an alternative estrogen,ethylnylestradiol, at same doses. The high dose of1 000 mg/kg-1 caused total mortality but fry from thelower dose of 500 mg/kg-1 survived (86.9%-94.3%,with a sex ratio of 63.1% female). It is hoped thatsex-reversed ZZ-females can be identified andtheir progeny subjected to a second generation offeminization treatment.
All-female production of Barbodes gonionotus
At RIA-1 in 1998, progress was made in all-femaleproduction using technique for induction ofgynogenesis and hormone sex reversal in B. gonionotus.The sperm was diluted with 0.85% salt solution toconcentration of 8 x 108 sperm/ml, and 10 ml ofdiluted sperm was irradiated in a small petri dish for60 seconds using ultraviolet lamp. A cold shock at2oC was applied 90 sec after fertilization at 30oC toeggs of B. gonionotus fertilized with UV-irradiatedsperm for 10 min. Non-irradiated sperm was used inthe diploid control group, and one group of eggsfertilized with irradiated sperm was not cold shocked,serving as a haploid control group.
In haploid control group, all hatchlings at two daysold eventually died. On the other hand, the averagesurvival rates in diploid control and mitoticgynogenesis groups were 66.7% and 11.5%,respectively. Sex ratios in gynogenetic fish were 91.3-97.4% with a mean of 95.8% female.
Androgen-treatment (17α-methyltestosterone or MT)was orally applied to gynogenetic fish to produce sex-reversed male B. gonionotus. Gynogenetic fish were fedwith 25 mg MT/kg diets for 5 weeks. Percentages ofmales in androgen treated-fish varied from 8.1 to 36%,with a mean of 16.7% male (Thang 1998).
Chromosome analysis and sex-specificmarker in Macrobrachium rosenbergii
In 1998, at the Research Institute of AnimalHusbandry (RIAH), Hanoi, an attempt to analyzechromosome in M. rosenbergii was made. Chromosomeswere prepared from cells collected from abdominalmuscle of 50 individuals. A metaphase of M. rosenbergiishowed variation in chromosome numbers. Themethod for preparing the prawn chromosome needsto be refined (Sac 1998).
Also in 1998, the University of Hanoi carried out sexdetermination in M. rosenbergii. Through heat shock,
Chelex and kit methods, DNA was extracted from shelland muscle. DNA quantity in male and females wasassessed. Concentration of DNA obtained from femalewas remarkably higher than those from males.
The extracted DNA was cut by 18 different restrictionenzymes for analysis of restriction fragment lengthpolymorphism (RFLP). Restriction enzyme Mnl I andPvu I cut the DNA of male prawn at specific nucleotidesequences, whereas these restriction enzymes did notcut the DNA of female.
Random amplified polymorphic DNA (RAPD)technique was used to detect DNA variation betweenmale and females. Several random primers were used:M13 (F), M13 (F and R) and human Y-gene specificprimers. However, variability of RAPD profilesgenerated by these primers in male and females wasnot sex-specific (Luong et al. 1998).
Induction of triploids in Clarias macrocephalus
Induction of triploids in C. macrocephalus wasconducted in the Southern Branch of Vietnam-RussiaTropical Center and the Mekong River Delta ResearchStation of RIA-2 for two years (Nga et al. 1998).
Matured breeders were spawned using hormoneinjection and stripping method. Dry fertilization ofeggs was applied. To induce triploid embryos, 2-3 minafter fertilization, the eggs were cold shocked at3-5oC for 30-40 min. (Table 2).
After 28 days of rearing in tanks, the survival rate,mean weight and total length of cold shocked andnormal fry were similar (Table 3). Table 4 shows thegrowth and survival rates between triploid and normalC. macrocephalus.
Using cold shock to induce triploid catfish gavepreliminary results of 36.2% larval survival and 48.1%triploids. Growth and survival of triploid (3n) catfishfrom fry to adult stages are normal as those of 2ncatfish. After 5 months of rearing in grow-out ponds,the final weight of triploid fish was 16.4% higher thanthat of normal fish.
DNA microinjection in Carassius auratus andMisgurnus anguillicaudatus
An initial study on gene transfer in transgenicC. auratus was undertaken recently at the Institute ofBiological Technology (IBT), Hanoi (Do et al. 1998).After injection of hormone to female and male
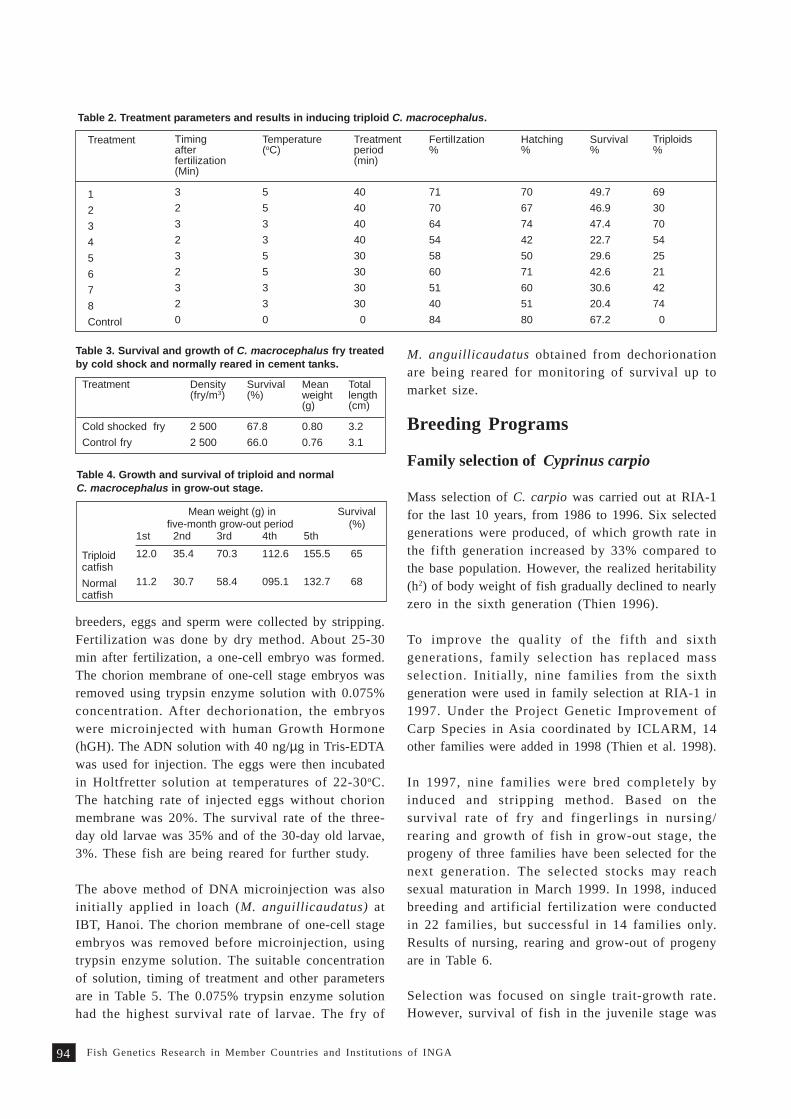
Fish Genetics Research in Member Countries and Institutions of INGA94
breeders, eggs and sperm were collected by stripping.Fertilization was done by dry method. About 25-30min after fertilization, a one-cell embryo was formed.The chorion membrane of one-cell stage embryos wasremoved using trypsin enzyme solution with 0.075%concentration. After dechorionation, the embryoswere microinjected with human Growth Hormone(hGH). The ADN solution with 40 ng/µg in Tris-EDTAwas used for injection. The eggs were then incubatedin Holtfretter solution at temperatures of 22-30oC.The hatching rate of injected eggs without chorionmembrane was 20%. The survival rate of the three-day old larvae was 35% and of the 30-day old larvae,3%. These fish are being reared for further study.
The above method of DNA microinjection was alsoinitially applied in loach (M. anguillicaudatus) atIBT, Hanoi. The chorion membrane of one-cell stageembryos was removed before microinjection, usingtrypsin enzyme solution. The suitable concentrationof solution, timing of treatment and other parametersare in Table 5. The 0.075% trypsin enzyme solutionhad the highest survival rate of larvae. The fry of
M. anguillicaudatus obtained from dechorionationare being reared for monitoring of survival up tomarket size.
Breeding Programs
Family selection of Cyprinus carpio
Mass selection of C. carpio was carried out at RIA-1for the last 10 years, from 1986 to 1996. Six selectedgenerations were produced, of which growth rate inthe fifth generation increased by 33% compared tothe base population. However, the realized heritability(h2) of body weight of fish gradually declined to nearlyzero in the sixth generation (Thien 1996).
To improve the quality of the fifth and sixthgenerations, family selection has replaced massselection. Initially, nine families from the sixthgeneration were used in family selection at RIA-1 in1997. Under the Project Genetic Improvement ofCarp Species in Asia coordinated by ICLARM, 14other families were added in 1998 (Thien et al. 1998).
In 1997, nine families were bred completely byinduced and stripping method. Based on thesurvival rate of fry and fingerlings in nursing/rearing and growth of fish in grow-out stage, theprogeny of three families have been selected for thenext generation. The selected stocks may reachsexual maturation in March 1999. In 1998, inducedbreeding and artificial fertilization were conductedin 22 families, but successful in 14 families only.Results of nursing, rearing and grow-out of progenyare in Table 6.
Selection was focused on single trait-growth rate.However, survival of fish in the juvenile stage was
Table 4. Growth and survival of triploid and normalC. macrocephalus in grow-out stage.
TriploidcatfishNormalcatfish
Mean weight (g) in Survival five-month grow-out period (%)1st 2nd 3rd 4th 5th12.0 35.4 70.3 112.6 155.5 65
11.2 30.7 58.4 095.1 132.7 68
Table 3. Survival and growth of C. macrocephalus fry treatedby cold shock and normally reared in cement tanks.
Treatment
Cold shocked fryControl fry
Density(fry/m3)
2 5002 500
Survival(%)
67.866.0
Meanweight(g)
0.800.76
Totallength(cm)
3.23.1
Table 2. Treatment parameters and results in inducing triploid C. macrocephalus.
Treatment
12345678Control
Timingafterfertilization(Min)
323232320
Temperature(oC)
553355330
Treatmentperiod(min)
4040404030303030 0
FertilIzation%
717064545860514084
Hatching%
706774425071605180
Survival%
49.746.947.422.729.642.630.620.467.2
Triploids%
6930705425214274 0
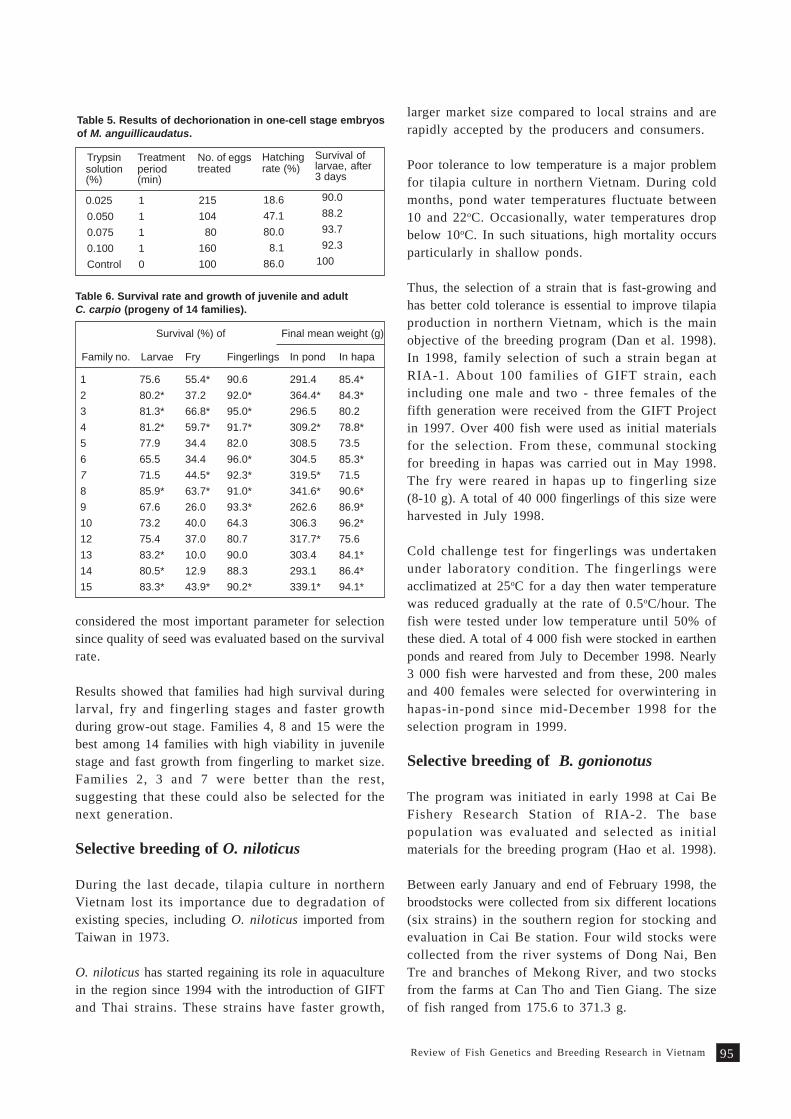
95Review of Fish Genetics and Breeding Research in Vietnam
considered the most important parameter for selectionsince quality of seed was evaluated based on the survivalrate.
Results showed that families had high survival duringlarval, fry and fingerling stages and faster growthduring grow-out stage. Families 4, 8 and 15 were thebest among 14 families with high viability in juvenilestage and fast growth from fingerling to market size.Families 2, 3 and 7 were better than the rest,suggesting that these could also be selected for thenext generation.
Selective breeding of O. niloticus
During the last decade, tilapia culture in northernVietnam lost its importance due to degradation ofexisting species, including O. niloticus imported fromTaiwan in 1973.
O. niloticus has started regaining its role in aquaculturein the region since 1994 with the introduction of GIFTand Thai strains. These strains have faster growth,
larger market size compared to local strains and arerapidly accepted by the producers and consumers.
Poor tolerance to low temperature is a major problemfor tilapia culture in northern Vietnam. During coldmonths, pond water temperatures fluctuate between10 and 22oC. Occasionally, water temperatures dropbelow 10oC. In such situations, high mortality occursparticularly in shallow ponds.
Thus, the selection of a strain that is fast-growing andhas better cold tolerance is essential to improve tilapiaproduction in northern Vietnam, which is the mainobjective of the breeding program (Dan et al. 1998).In 1998, family selection of such a strain began atRIA-1. About 100 families of GIFT strain, eachincluding one male and two - three females of thefifth generation were received from the GIFT Projectin 1997. Over 400 fish were used as initial materialsfor the selection. From these, communal stockingfor breeding in hapas was carried out in May 1998.The fry were reared in hapas up to fingerling size(8-10 g). A total of 40 000 fingerlings of this size wereharvested in July 1998.
Cold challenge test for fingerlings was undertakenunder laboratory condition. The fingerlings wereacclimatized at 25oC for a day then water temperaturewas reduced gradually at the rate of 0.5oC/hour. Thefish were tested under low temperature until 50% ofthese died. A total of 4 000 fish were stocked in earthenponds and reared from July to December 1998. Nearly3 000 fish were harvested and from these, 200 malesand 400 females were selected for overwintering inhapas-in-pond since mid-December 1998 for theselection program in 1999.
Selective breeding of B. gonionotus
The program was initiated in early 1998 at Cai BeFishery Research Station of RIA-2. The basepopulation was evaluated and selected as initialmaterials for the breeding program (Hao et al. 1998).
Between early January and end of February 1998, thebroodstocks were collected from six different locations(six strains) in the southern region for stocking andevaluation in Cai Be station. Four wild stocks werecollected from the river systems of Dong Nai, BenTre and branches of Mekong River, and two stocksfrom the farms at Can Tho and Tien Giang. The sizeof fish ranged from 175.6 to 371.3 g.
Table 5. Results of dechorionation in one-cell stage embryosof M. anguillicaudatus.
Trypsinsolution(%)
0.0250.0500.0750.100Control
Treatmentperiod(min)
11110
No. of eggstreated
215104 80160100
Hatchingrate (%)
18.647.180.08.1
86.0
Survival oflarvae, after3 days
90.088.293.792.3
100
Table 6. Survival rate and growth of juvenile and adultC. carpio (progeny of 14 families).
Family no.
1234567891012131415
Larvae
75.680.2*81.3*81.2*77.965.571.585.9*67.673.275.483.2*80.5*83.3*
Fry
55.4*37.266.8*59.7*34.434.444.5*63.7*26.040.037.010.012.943.9*
In pond
291.4364.4*296.5309.2*308.5304.5319.5*341.6*262.6306.3317.7*303.4293.1339.1*
In hapa
85.4*84.3*80.278.8*73.585.3*71.590.6*86.9*96.2*75.684.1*86.4*94.1*
Fingerlings
90.692.0*95.0*91.7*82.096.0*92.3*91.0*93.3*64.380.790.088.390.2*
Survival (%) of Final mean weight (g)

Fish Genetics Research in Member Countries and Institutions of INGA96
References
Dan, N.C. 1998. Family selection of Nile tiliapia (Oreochromisniloticus), Research Institute for Aquaculture No 1, Bac Ninh,Vietnam. 12 p.
Dan, N.C, T.M. Thien and D.V. Trung. 1997. Tilapia breeding inricefields in Vietnam. Naga, the ICLARM Q. 20 (1): 23-25.
Do, N.K., T.Q. Dung and T.H. Hang. 1997. Preliminary result ofgenerating transgenic goldfish (Carassius auratus). VietnamNational Center for National Science and Technology, HuePedagogic Technology, University of Hanoi. 7 p.
Hao, N.V. 1998. Collection of initial materials for selectivebreeding of silver barb Puntius gonionotus, Research Institutefor Aquaculture No. 2, Ho Chi Minh City, Vietnam. 15 p.
Luong, L.D. 1998. Preliminary result on sex specific markers ingiant freshwater shrimp (M. rosenbergii). University of Hanoi,Vietnam. 14 p.
MOF. 1998. Total fish production, p. 1-15. In Annual report.Ministry of Fisheries, Hanoi, Vietnam.
Nga, N.T., T.T.L. Chi and P.V. Khanh. 1998. Research results oninduction of triploids in catfish (Clarias macrocephalus), theSouthern Branch of Vietnam-Russia Tropical Center and theMekong River Delta Research Station, Research Institute forAquaculture No 2, Ho Chi Minh City, Vietnam. 23 p.
Sac, N.V. 1998. Preliminary result on chromosome analysis in giantfreshwater shrimp (Macrobrachium rosenbergii). ResearchInstitute of Animal Husbandry, Hanoi, Vietnam. 11 p.
Thang, N.C. 1998. All-female production of silver barb Puntiusgonionotus, 17-20. In Annual report. Research Institute forAquaculture No.1, Bac Ninh, Vietnam.
Thien, T.M. 1996. Carp breeding in Vietnam. Final Report. IFS.Thien, T.M and N.C. Dan. 1998. Carp genetics resources in
Vietnam. Paper presented at the Planning Workshop onRegional Collaborative Project for Genetic Improvement ofCarps in Asia 26-28 July 1997, Bhubaneswar, India.
Thien, T.M, N.C. Dan and H.D. Tu. 1998. Preliminary result onfamily selection of common carp (Cyprinus carpio). ResearchInstitute for Aquaculture No. 1, Bac Ninh, Vietnam. 9 p.
Tuan, P.A. 1998. Genetic control of sex ratio in Nile tilapia (O.niloticus) using YY-male technology, p. 12-15. In Annualreport. Research Institute for Aquaculture No.1, Bac Ninh,Vietnam.
Six stocks were conditioned in earthen ponds formaturity. Induced breeding, through hormoneinjection, was done within the same stock and crossingbetween two different stocks. There were twobreedings and success was achieved on the secondtime, when over 70% of larvae survived.
The fry obtained from six stocks and three crossingswere reared in earthen ponds and cement tanksfor evaluation and comparison of performance. Afterfour months, the fish reached 6.5-7.8 g, and therewas no significant difference in growth rate amongthe progenies.
Fin clipping and tagging with the use of fluorescencewere tried. Survival and retention of color marks aftertagging need to be tested again.
The broodstocks and their progenies are being rearedin controlled condition for further evaluation.
Future PlansFuture Plans
The following research activities are ongoing or willbe carried out at RIA-1 and other relevant institutions:• Genetic control of sex ratio in O. niloticus (Thai,
Vietnamese and GIFT strains), using YY-maletechnology (RIA-1);
• Mass seed production of all female B. gonionotus,based on the success of producing neo-male (x x)(RIA-1);
• Sex reversal of M. rosenbergii using hormonetreatment technology (RIA-1);
• Induction of triploids in C. macrocephalus andconsideration for production (Southern Branchof Vietnam-Russia Tropical Center);
• Gene transfer of some fish species (Institute ofBiotechnology);
• Cryopreservation of some species (RIA-1); and• Use of DNA technique for diagnosis of diseases
in fish.The following breeding programs are planned:
• Family selection of GIFT strain O. niloticus forsecond generation. (RIA-1);
• Mass selection of B. gonionotus (RIA-2); and• Family selection of C. carpio for second generation
(RIA-1).

97Fish Genetics Research atICLARM - the World Fish Center
ABSTRACT
ICLARM - The World Fish Center has a mandate to improve the food security of people in developing countries byproviding better management advice and methods to conserve current fish stocks and developing efficientaquaculture technologies. ICLARM pursues this through its various programs which include, among others, researchon germplasm enhancement and breeding, biodiversity and genetic resources.
A major research project aimed at developing methods for genetic improvement of finfish used Nile tilapia (Oreochromisniloticus) as a test species and was followed through six generations of selection. The improved strain (popularlyknown as GIFT strain) outperformed in growth and survival traits the widely farmed tilapia strains in the Philippinesand other parts of Asia. The overall potential impact of the GIFT strain has been assessed in Bangladesh, People’sRepublic of China, Philippines, Thailand and Vietnam.
The lessons learned and experiences gained from the GIFT project are now being applied for genetic improvementof six species of carp in Asia (Bangladesh, China, India, Indonesia, Thailand, and Vietnam) and the indigenous tilapiaspecies in Africa, in collaboration with national partners.
ICLARM addresses research and policy issues that have impact on the management of aquatic genetic resources.The Center also contributes to capacity building of national institutions in the areas of germplasm enhancement andbreeding and conservation of genetic resources through the International Network on Genetics in Aquaculture (INGA)and project-related training programs.
FISH GENETICS RESEARCH ATICLARM - THE WORLD FISH CENTERa
M.V. Gupta, B.O. Acosta, R. Dunhamb and P.R. GardinerICLARM - The World Fish Center
P.O. Box 500, 10670 Penang, Malaysia
GUPTA, M.V., B.O. ACOSTA, R. DUNHAM and P.R. GARDINER. 2001. Fish Genetics Research at the International Centerfor Living Aquatic Resources Management, p. 97-102. In M.V. Gupta and B.O. Acosta (eds). Fish genetics researchin member countries and institutions of the International Network on Genetics in Aquaculture. ICLARM Conf.Proc., 64, 179 p.
a ICLARM Contribution no. 1610.b Current Address: Department of Fisheries and Allied Aquaculture, Auburn University, Alabama 36848, USA.

Fish Genetics Research in Member Countries and Institutions of INGA98
Introduction
The mandate to improve the well-being and livelihoodof present and future generations of poor people inthe developing world through improved production,management and conservation of living aquaticresources has been the basis for ICLARM’s work forthe past two decades. As the only fisheries Center ofthe Consultative Group on International AgriculturalResearch Systems (CGIAR), ICLARM has a global rolein providing better management advice and methodsto conserve genetic resources and develop efficienttechnologies for aquaculture/fisheries managementnecessary to augment fish productivity and to meetthe world’s growing demand for fish.
Pullin et al (1991) summarized the results of studieson tilapias by ICLARM and research partners in theearly/mid 1980s. The overall observation was that Asianfarmed tilapia stocks had poor growth performance andgenetic quality. This could be due to introgressivehybridization of farmed stocks of Nile tilapia (Oreochromisniloticus) with feral populations of the much lessdesirable species for farming – the Mozambique tilapia(O. mossambicus). The workshop on Tilapia GeneticResources for Aquaculture organized by ICLARM inThailand in 1987 confirmed this and concluded thatthe presently available farmed stocks of O.niloticus inAsia were inadequate for genetic improvement programsbecause of their small founder populations, inbreeding,and introgression of genes from other less desirablespecies (Pullin 1988; Pullin and Capili 1988). Further,it was found that the indigenous tilapia populations inAfrica are under threat due to indiscriminate fishtransfers and habitat disturbance. There was obviouslyan urgent need to document tilapia genetic resourcesfor the twin goals of conservation and for futureaquaculture development in the region.
The above findings formed the basis of ICLARM’smajor strategic research on fish genetics. The Centerrealized that interactive research is essential indeveloping countries for sustained fish productivityand that this is best done in collaboration with itspartner institutions.
Germplasm enhancement and breeding and theconservation of biodiversity and genetic resources arethe two major thrusts of ICLARM’s genetics research.The genetic enhancement thrust aims at developingtechniques/methods for improving fish breeds,disseminating the methods developed and trainingdeveloping country scientists in their use. The Center’s
major effort on conservation of fish genetic resources isdirected at enhancing the capacity of national programsfor fisheries and biodiversity management and indemonstrating how to document aquatic geneticresources for their sustainable use and conservation.
Germplasm enhancement andbreeding
Genetic improvement of farmed tilapias. Investigations ofthe potential for genetic enhancement of tropical finfishthrough selective breeding using O.niloticus as the testspecies, was the first major strategic quantitative geneticsresearch undertaken by ICLARM. It was anticipated thatsuccessful results with this species would result in notonly better breeds of tilapia for farmers, but also newbreeding methods for use with other tropical finfishspecies.
ICLARM in collaboration with its partner institutionsin the Philippines (Bureau of Fisheries and AquaticResources, Freshwater Aquaculture Center of theCentral Luzon State University, and the Marine ScienceInstitute of the University of the Philippines) andNorway (Institute of Aquaculture Research Ltd.),implemented the Genetic Improvement of FarmedTilapia (GIFT) project to develop effective methodsfor producing improved breeds of O. niloticus for lowcost sustainable aquaculture in developing countries.In view of the poor quality of cultured stocks in Asia,pure strains of O. niloticus were collected from diversecountries: Egypt, Ghana, Senegal and Kenya. Thesewere used as founder stocks along with the four existingstrains in the Philippines, known as Singapore,Thailand, Israel and Taiwan strains (Eknath et al 1993).Based on the growth performance of the differentstrain combinations, individuals from the bestperforming groups were selected for a selective breedingprogram. Over ten years (1988-1997), selection wasundertaken through five successive generations of thebase population. Superior performance in terms ofgrowth and survival under on-station and on-farmconditions was observed even after one generation ofselection (Eknath 1993). The first generation of theimproved strain (which came to be known as GIFTstrain) grew 26% faster in on-station trials than theprevious generation and 75% faster than the mostcommonly farmed strain in the Philippines (Eknath1995). On average, the genetic gain per generationacross five generations of selection was about 12-17%(Fig. 1) (Eknath and Acosta 1998; Eknath et al 1998).
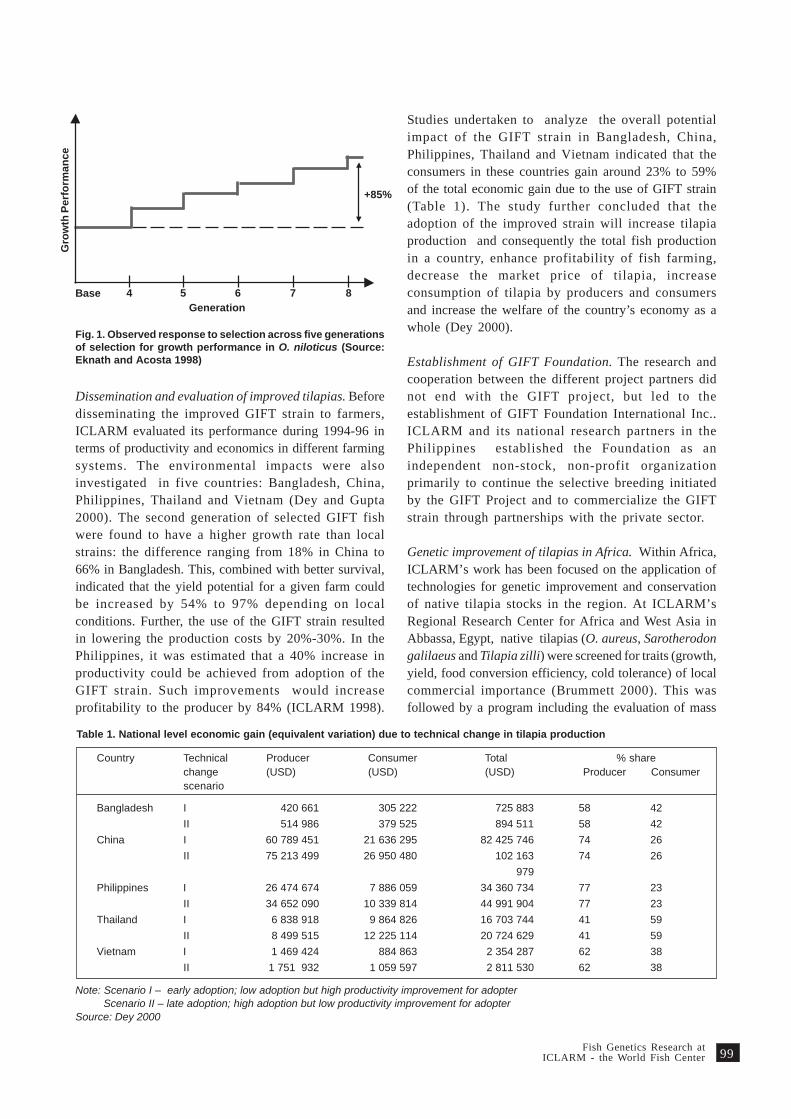
99
Dissemination and evaluation of improved tilapias. Beforedisseminating the improved GIFT strain to farmers,ICLARM evaluated its performance during 1994-96 interms of productivity and economics in different farmingsystems. The environmental impacts were alsoinvestigated in five countries: Bangladesh, China,Philippines, Thailand and Vietnam (Dey and Gupta2000). The second generation of selected GIFT fishwere found to have a higher growth rate than localstrains: the difference ranging from 18% in China to66% in Bangladesh. This, combined with better survival,indicated that the yield potential for a given farm couldbe increased by 54% to 97% depending on localconditions. Further, the use of the GIFT strain resultedin lowering the production costs by 20%-30%. In thePhilippines, it was estimated that a 40% increase inproductivity could be achieved from adoption of theGIFT strain. Such improvements would increaseprofitability to the producer by 84% (ICLARM 1998).
Studies undertaken to analyze the overall potentialimpact of the GIFT strain in Bangladesh, China,Philippines, Thailand and Vietnam indicated that theconsumers in these countries gain around 23% to 59%of the total economic gain due to the use of GIFT strain(Table 1). The study further concluded that theadoption of the improved strain will increase tilapiaproduction and consequently the total fish productionin a country, enhance profitability of fish farming,decrease the market price of tilapia, increaseconsumption of tilapia by producers and consumersand increase the welfare of the country’s economy as awhole (Dey 2000).
Establishment of GIFT Foundation. The research andcooperation between the different project partners didnot end with the GIFT project, but led to theestablishment of GIFT Foundation International Inc..ICLARM and its national research partners in thePhilippines established the Foundation as anindependent non-stock, non-profit organizationprimarily to continue the selective breeding initiatedby the GIFT Project and to commercialize the GIFTstrain through partnerships with the private sector.
Genetic improvement of tilapias in Africa. Within Africa,ICLARM’s work has been focused on the application oftechnologies for genetic improvement and conservationof native tilapia stocks in the region. At ICLARM’sRegional Research Center for Africa and West Asia inAbbassa, Egypt, native tilapias (O. aureus, Sarotherodongalilaeus and Tilapia zilli) were screened for traits (growth,yield, food conversion efficiency, cold tolerance) of localcommercial importance (Brummett 2000). This wasfollowed by a program including the evaluation of mass
Fish Genetics Research atICLARM - the World Fish Center
GenerationBase 4 5 6 7 8
Fig. 1. Observed response to selection across five generationsof selection for growth performance in O. niloticus (Source:Eknath and Acosta 1998)
+85%
Gro
wth
Per
form
ance
Table 1. National level economic gain (equivalent variation) due to technical change in tilapia production
Country Technical Producer Consumer Total % sharechange (USD) (USD) (USD) Producer Consumerscenario
Bangladesh I 420 661 305 222 725 883 58 42II 514 986 379 525 894 511 58 42
China I 60 789 451 21 636 295 82 425 746 74 26II 75 213 499 26 950 480 102 163 74 26
979Philippines I 26 474 674 7 886 059 34 360 734 77 23
II 34 652 090 10 339 814 44 991 904 77 23Thailand I 6 838 918 9 864 826 16 703 744 41 59
II 8 499 515 12 225 114 20 724 629 41 59Vietnam I 1 469 424 884 863 2 354 287 62 38
II 1 751 932 1 059 597 2 811 530 62 38
Note: Scenario I – early adoption; low adoption but high productivity improvement for adopter Scenario II – late adoption; high adoption but low productivity improvement for adopterSource: Dey 2000

Fish Genetics Research in Member Countries and Institutions of INGA100
selection to improve growth and food conversionefficiency of O. niloticus and O. aureus. Currently, thesespecies are being bred for another round of evaluationand alternative selection experiments (ICLARM 1999b,1999c, Brummett 2000).
In collaboration with Auburn University, Alabama,USA, a project has been initiated to evaluate thepotential usefulness of marker- assisted selection andquantitative trait loci (QTL) mapping in geneticenhancement of tilapias in Africa (Liu 2001; ICLARM2000a).
The research at ICLARM in Abbassa is complementedby a collaborative project being coordinated by theInternational Network on Genetics in Aquaculture(INGA)/ICLARM on genetic enhancement andconservation of tilapia genetic resources in Côted’Ivoire, Egypt, Ghana and Malawi. With start-up fundsfrom the International Development Research Centreof Canada, the participating institutions (CentreNational de Recherche Agronomique, Côte d’Ivoire;Central Laboratory of Aquaculture Research, Egypt;Water Research Institute, Ghana; and University ofMalawi, Malawi,) are undertaking the followingactivities: evaluation of local tilapia populationssuitable for genetic improvement/aquaculture; geneticcharacterization to determine the population structureand genetic diversity of native tilapia stocks;documenting traditional knowledge of tilapia species/stocks and farming systems; identifying breeding goalsand initiating studies for genetic improvement.
To further address the specific requirements of Africanaquaculture, ICLARM will initiate a project aimed attransferring the selective breeding technologydeveloped in the GIFT project to Sub-Saharan Africaand Egypt. This will be accomplished through theestablishment of a Regional Tilapia GeneticEnhancement Center in Egypt, training of Egyptianand sub-Saharan African scientists by Philippinescientists and scientific exchange between Egypt,Philippines and other regions of Africa (ICLARM2000b).
ICLARM is seeking funds to organize a conference onbiosafety issues pertaining to the dissemination ofgenetically enhanced fish, including geneticallymodified organisms (GMOs), with special reference tothe tilapias within Africa.
Genetic improvement of carps. The selective breedingtechnique developed for improving the growth andproductivity of O. niloticus in Asia is now being appliedto the other major farmed tropical finfish in Asia, thecarps, which make up 43.6% of global aquacultureproduction (FAO 1998). Bangladesh, China, India,Indonesia, Thailand and Vietnam are the major carpproducers in Asia and contribute more than 90% to theglobal production of carps.
ICLARM in 1997 initiated a project for the geneticimprovement of carps, in collaboration with its partnerinstitutions in Bangladesh (Bangladesh FisheriesResearch Institute and Bangladesh AgriculturalUniversity), China (Shanghai Fisheries University andFreshwater Fisheries Research Center of the ChineseAcademy of Fishery Sciences), India (Central Instituteof Freshwater Aquaculture, National Bureau of FishGenetic Resources and University of AgriculturalSciences), Indonesia (Research Institute for FreshwaterFisheries), Thailand (National Aquaculture GeneticsResearch Institute and the Department of Fisheries),and Vietnam (Research Institute for Aquaculture Nos.1 and 2). The first phase of the project focuses onassessing the current status of carp genetic resourcesincluding their systematic documentation andevaluation, developing criteria for prioritizing carpgenetic research, identifying research priorities andapproaches including species, traits, farming systemsand breeding strategies and initiating location - specificstrategic research and training based on identifiedresearch priorities for development of high yieldingcarp strains (Gupta et al. 1997). Extensive surveys ofcarp producers, consumers and hatchery operators havebeen completed in all the six participating countries toprioritize species, traits and farming systems for geneticimprovement. Carp genetic resources in theparticipating countries have been documented. Geneticenhancement of six species of carps: common carp(Cyprinus carpio), silver barb (Barbodes gonionotus), rohu(Labeo rohita), catla (Catla catla), Mrigal (CirrhinusMrigala) and Chinese bream (Megalobrama amblycephala)is in progress. Initially the selection is for growth. Someof the improved strains are presently being tested infarmers’ fields (ICLARM 1999a).
Biodiversity and Genetic Resources
Conserving fish biodiversity and protecting theenvironment are two of the major research goals ofICLARM. Since knowledge of genetic diversity andpopulation structure of commercially important species

101Fish Genetics Research atICLARM - the World Fish Center
are vital for the management of farmed and wildpopulations, ICLARM and its research partners areimplementing studies in Africa and Asia to developmethods for assessing genetic diversity. Two projectsfor documenting genetic resources for their sustainableuse and conservation were initiated in 1997. Thesestudies focus on black-chinned tilapia (Sarotherodonmelanotheron), a coastal species inhabiting brackish-water lagoons and water courses in West Africa and onan Asian carp, B.goniontous, a species that is rapidlygaining popularity for culture among resource-poorfish farmers in South and South East Asia .
In collaboration with Zoologisches Institut undZoologisches Museum, University of Hamburg,Germany and Water Research Institute, Ghana, studiesare in progress to assist in the conservation andsustainable use in aquaculture and fisheries of S.melanotheron in West Africa (Pullin 2000). Samples ofthis species were collected over the entire range inSenegal, Sierra Leone, Liberia, Côte d’Ivoire, Ghana,Togo, Benin and Congo. Extensive data were obtainedon the genetic diversity of S. melanotheron populationsand examined using molecular genetic methods andpartly with morphometric analysis. Preliminaryanalysis revealed 17 control region mtDNA haplotypesthat clustered samples into three regional groups: (i)Senegal – Liberia; (ii) Côte d’Ivoire – Ghana; and(iii) Bas Kouilou, Congo (ICLARM 1999b). Data werealso gathered on the biology, ecology and use of S.melanotheron in West African coastal lagoons andwatercourses. Culture trials of genetically determinedGhanaian S.melanotheron populations are in progress(ICLARM 1999b).
In Asia, research on the genetic diversity of B.gonionotus, a species that is gaining importance for low-input pond culture and integrated systems, is beingundertaken in collaboration with the University ofWales, Swansea, U.K. Samples were collected from wildpopulations in central, northern and southernThailand and in central, east and west Java, Indonesiaand from wild and farmed populations in Lao PDRand Vietnam. Initial results from molecular genetic andmeristic studies, indicate that B. gonionotus is a singlespecies across its range. Samples collected forcomparison included B. altus, B. belleroides and B.schwanenfeldii (ICLARM 1999b).
ICLARM addresses research and policy issues thathave impacts on the management of aquatic geneticresources through its contributions to the work of: the
Convention on Biological Diversity (CBD) and itsSubsidiary Body on Scientific, Technical andTechnological Advice and Clearing House Mechanism;Global Biodiversity Fora; Species 2000; Food andAgriculture Organization of the United Nations (FAO)Fisheries Division and Commission on GeneticResources for Food and Agriculture; and the WorldConservation Union (ICLARM 1999b). In 1998,ICLARM, in collaboration with FAO, convened aBellagio Conference entitled ‘Towards Policies forConservation and Sustainable Use of Aquatic GeneticResources’. The main output of this meeting was aconsensus statement which was distributed at theFourth Conference of the Parties to the CBD held inSlovakia in May 1998 (Pullin et al 1999).
ICLARM also participates in the CGIAR’s System-wideGenetic Resources Program and System-wideInformation Network on Genetic Resources.
Networking in Genetics Research
ICLARM coordinates the genetics research network -INGA. The network has been playing an importantrole in national, regional and international geneticsresearch aimed at improving production fromaquaculture operations and conservation of aquaticgenetic resources and biodiversity (Gupta and Acosta1999). With a present membership of 13 countries fromAsia, the Pacific and Africa and 12 advanced scientificinstitutions as Associate Members, the network isundertaking the: (i) development of national breedingprograms; (ii) initiation of regional research programsfor genetic improvement of carps and tilapias; (iii)transfer of germplasm among member countries,following strict quarantine protocols and materialtransfer agreements; (iv) assistance in formation ofnational genetics networks; and (v) strengthening ofnational research capacity (Gupta and Acosta 2001).Details of the network activities are elsewhere in thisproceedings.
Training of National Scientists
Through INGA and project - related trainingprograms, ICLARM will endeavor to form a cadre ofscientists in INGA member countries in the area ofquantitative genetics and selective breeding. Since thisarea is relatively new for the majority of nationalinstitutions in developing countries in Asia, Africa andthe Pacific, ICLARM/INGA, in collaboration withNorway’s Institute of Aquaculture Research Ltd.

Fish Genetics Research in Member Countries and Institutions of INGA102
(AKVAFORSK), has organized training programs forscientists from national institutions in these regions.ICLARM/INGA will also address the specific trainingneeds of national institutions in Africa through trainingprograms that will be organized under the project‘Transfer of Selective breeding technology foraquaculture improvement from Asia to Sub-SaharanAfrica and Egypt’ (ICLARM 2000b).
References
Brummett, RE. 2000. Research on improved germplasm foraquaculture at the ICLARM Regional Research Center forAfrica and West Asia and a note on the domestication ofHeterotis niloticus, p. 56-58. In E.K. Abban, C.M.V. Casal, T.M.Falk and R.S.V. Pullin (eds). Biodiversity and sustainable useof fish in the coastal zone. ICLARM Conf. Proc. 63, 71p.
Dey, M.M. 2000. The impact of genetically improved farmedNile tilapia in Asia. Aquacult. Econ. and Manage. 4(1&2):107 –123.
Dey, M.M. and M.V. Gupta 2000. Socio-economic aspects ofdisseminating improved tilapia in Asia: an introduction.Aquacult. Econ. and Manage. 4(1&2): 5-12.
Eknath, A.E. 1993. The Project of Genetic Improvement ofFarmed Tilapias (GIFT): from modest beginnings to anInternational Network on Genetics in Aquaculture. p. 9-13.In Villacorta, L.G., and L.A. Dureza (eds.). Tilapia FarmingGenetics Improvement and Advances on Culture Technology:Proceedings of the Third National Symposium andWorkshop on Tilapia Farming. Los Banos, Laguna,Philippines. PCAMRD Book Series No. 18/1994.
Eknath, A.E., M.M. Tayamen, M.S. Palada-de-Vera, J.C. Danting,R.A. Reyes, E.E. Dionisio, J.B. Capili, H.L. Bolivar, T.A. Abella,A.V. Circa, H.B. Bentsen, B. Gjerde, T. Gjedrem and R.S.V.Pullin. 1993. Genetic improvement of farmed tilapias: thegrowth performance of eight strains of Oreochromis niloticustested in different farm environments. Aquaculture. 111:171-188.
Eknath, A.E. 1995. Managing aquatic genetic resources.Management Example 4: The Nile tilapia. p. 176-194. In J.E. Thorpe, G.A.E. Gall, J.E. Lannan and C.E. Nash (eds.)Conservation of Fish and Shellfish Resources: ManagingDiversity. Academic Press, Harcourt Brace & Company,London.
Eknath, A.E. and B.O. Acosta. 1998. Genetic Improvement ofFarmed Tilapias (GIFT) Project” Final Report, March 1988to December 1997. International Center for Living AquaticResources Management, Manila, Philippines
Eknath, A.E., Dey, M.M., Rye, M., Gjerde, B., Abella, T.A., Sevilleja,R.A., Tayamen, M.M., Reyes, R.A. and Bentse, H.B. 1998.Selective breeding of Nile tilapia for Asia. In p. 89-96.Proceedings of the Sixth World Congress on Genetics Appliedto Livestock Production, 27, Armidale, Australia.
FAO 1998. Aquaculture production statistics 1987 – 1996. FAOFish. Circ. No. 815. Rev. 10. FAO, Rome. Italy. 197p.
Gupta, M.V. and B.O. Acosta. 1999. International Network onGenetics in Aquaculture: A global forum for collaborativeresearch and training in applied fish breeding and genetics.ICLARM, Makati City, Metro Manila, Philippines. 17 p.
Gupta, M.V. and B.O. Acosta. 2001. Networking in fish geneticsresearch. p. 1-5. In M.V. Gupta and B.O. Acosta.Fish Genetics Research in Member Countries and Institutionsof the International Network on Genetics in Aquaculture.ICLARM Conf. Proc. 64, 174p., ICLARM – the World FishCenter, Penang, Malaysia.
Gupta, M.V., M.M. Dey, R. Dunham and G. Bimbao, Editors.1997. Proceedings of the collaborative research and trainingon genetic improvement of carp species in Asia. ICLARMWork. Doc. No. 1 (Unpublished).
ICLARM. 1998. Dissemination and Evaluation of GeneticallyImproved Tilapia Species in Asia: Final Report. AsianDevelopment Bank Regional Technical Assistance No, 5558.International Center for Living Aquatic ResourcesManagement, Manila, Philippines
ICLARM. 1999a. Genetic improvement of carp species in Asia.Project Progress Report: July 1998 – January 1999.(Unpublished).
ICLARM. 1999b. Operational plan. International Center forLiving Aquatic Resources Management. Manila, Philippines.102 p.
ICLARM. 1999c. Report to the Arab fund for economic andsocial development :
1 January 1998 - 31 December 1999. (Unpublished).ICLARM. 2000a. Abbassa Update. 3(1): 1-4.ICLARM. 2000b. Transfer of selective breeding technology for
aquaculture improvement from Asia to Sub-Saharan Africaand Egypt. United Nations Development Programme -Project Document (Unpublished).
Liu, Z.J. 2001. Gene-mapping, marker-assisted selection, genecloning, genetic engineering and integrated geneticimprovement programs at Auburn University. p. 109-118.In M.V. Gupta and B.O. Acosta. Fish Genetics Research inMember Countries and Institutions of the InternationalNetwork on Genetics in Aquaculture. ICLARM Conf. Proc.64, 174p., ICLARM – the World Fish Center, Penang,Malaysia.
Pullin, R.S.V., Editor. 1988. Tilapia genetic resources foraquaculture. ICLARM Conf. Proc. 16, 108 p. InternationalCenter for Living Aquatic Resources Management, Manila,Philippines.
Pullin, R.S.V. 2000. A history of productive partnerships, p. 59-60. In E.K. Abban, C.M.V. Casal, T.M. Falk and R.S.V. Pullin(eds.). Biodiversity and sustainable use of fish in the coastalzone: ICLARM Conf. Proc. 63, 71p.
Pullin, R.S.V. and J.B. Capili. 1988. Genetic improvement oftilapias: problems and prospects, p. 259-266. In R.S.V. Pullin,T. Bhukaswan, K. Tonguthai and J.L. Maclean (eds.) TheSecond International Symposium on Tilapia in Aquaculture.ICLARM Conf. Proc. 15, 623p. Department of Fisheries,Bangkok, Thailand, and International Center for LivingAquatic Resources Management, Manila, Philippines.
Pullin, R.S.V., D.M. Bartley and J. Kooiman, Editors. 1999.Towards policies for conservation and sustainable use ofaquatic genetic resources. ICLARM Conf. Proc. 59, 277 p.
Pullin, R.S.V., A.E. Eknath, T. Gjedrem, M.M. Tayamen, J.M.Macaranas and T.A. Abella. 1991. The genetic improvementof farmed tilapias (GIFT) Project: The story so far. NAGA .ICLARM Q. 14(2): 31p.

103
ABSTRACT
Genetic improvement programs in Israel have focused mainly on common carp, tilapias, and recently on marinefishes. A breeding program to develop tilapia strains that are cold tolerant and fast growing in fresh and saltwaterenvironments is in progress. Efforts are also being made to produce disease-resistant carp seed. Selective breedingprogram for sea bream and sea bass has been initiated. Interspecific hybridization experiments were also carriedout to improve their culture performance.
Investigations on color pattern inheritance, chromosome-set and sex manipulations of various aquaculture specieshave been conducted or are in progress. Genetic engineering and transgenesis of a few candidate genes;development of DNA markers and the use of QTL mapping to support the fish breeding programs are underway.Other efforts include development of methods for gamete cryopreservation of common carp, marine fishes andshrimp and initiation of gene bank.
ISRAELI AQUACULTUREGENETIC IMPROVEMENT
PROGRAMS
G. HulataDepartment of AquacultureInstitute of Animal Science
Agricultural Research OrganizationVolcani Center, PO Box 6Bet Dagan 50250, Israel
HULATA, G. 2001. Israeli aquaculture genetic improvement programs, p. 103-108. In M.V. Gupta and B. O. Acosta (eds.)Fish genetics research in member countries and institutions of the International Network on Genetics in Aquacul-ture. ICLARM Conf. Proc. 64, 179 p.
Introduction
The history and status of aquaculture genetics in Israelwere reviewed a few years ago (Hulata 1995). Thecurrent report reviews trends and progress of the secondhalf of the 1990s, and provides the vision andperspective for the next couple of years. Selectivebreeding work is currently focused mainly on tilapias(Oreochromis sp.) and gilthead seabream (Sparus aurata).Edible and ornamental common carp (Cyprinus carpio)are used, among other species, in the application ofbiotechnological methods.
Israeli Aquaculture Genetic Inprovement Programs
Breeding programs
A program aiming at breeding new, syntheticpopulations of tilapia specifically adapted to temperateclimates and saline environments was initiated in 1995at the Department of Aquaculture (ARO, VolcaniCenter). It is based on plant breeders’ approach, andtakes benefit of the ease of producing interspecifichybrids among tilapias. The use of interspecificcomposite (or complex) crosses is an established practicein plant breeding for achieving wide genetic andphenotypic variability. This approach is based on the

Fish Genetics Research in Member Countries and Institutions of INGA104
creation of an artificial center of origin (ACO) viacomposite interspecific crosses. The ACO contains widegenetic diversity and presents opportunities for genesto recombine and interact with other genes originatingfrom different species, combinations and interactionswhich are impossible in any of the pure species. Israelicut flower breeders recently applied an adaptation ofthis approach, termed Multiple Re-Speciation, todevelop new cultivars of carnations.
The ACO (Hulata et al. 1999; Agresti et al. 2000) wasproduced by inter-crossing four tilapiine species:Oreochromis niloticus [wild type (On) and red (ROn)strains], O. aureus (Oa), O. mossambicus (Om), andSarotherodon galilaeus (Sg). All hybrids were obtainedby natural spawning, except for the S. galilaeus xOreochromis sp. F1 hybrids that were produced byartificial fertilization. A series of cold-water challengetests were performed, involving O. mossambicus, O.aureus and their F1 and F2 hybrids, to study the geneticbasis of cold tolerance in tilapias (Cnaani et al. 1999,2000). All of the different two-way (F1 hybrids) crossesrequired to establish the synthetic stock of tilapia (ACO)have been obtained, as well as a set of three four-way-crosses (4WC) derived from five strains in four speciesand two genera. Full-sibs of the [(Om x Oa) x (Sg xOn)] 4WC have been successfully bred to enable mixingof the inherited gene blocks; thus, we are ready to startselective breeding for cold tolerance and growth ratein freshwater and for growth rate in saltwater, fromthis base population. Selection will be based on acombination of individual and family performance;an approach that was used successfully for the GIFTbreeding program.
A tilapia breeding program is also conducted at theNir David Fish Breeding Farm, aiming at creating afast growing, salinity tolerant, uniformly red tilapiahybrid (Lahav and Ra’anan 1997), and othercommercial strains.
A first step in a breeding program to improve growthrate in gilthead seabream (Sparus auratus) carried outat the National Center for Mariculture (IOLR, Eilat)was strain evaluation (Knibb et al. 1997a). The researchteam then moved on to prepare for family selection,encountering difficulties in efficiently producingsingle-pair offspring groups (full- and half-sibs) andleading to the conclusion that family mating designsare inappropriate for the group spawning S. aurata(Gorshkov et al. 1997). Progeny testing, where singlemales were stocked with groups of females, which moreclosely simulate the natural group-spawning behavior
of the species, yielded genetically related groups moresuccessfully. The small number of families obtained (4each for full- and half-sibs) did not allow obtainingreliable estimates of heritability for growth, yet large(14-29%) sire components of the offspring weightvariance were evident (Knibb et al. 1998a). Massselection proved more effective and resulted insignificant heritability estimates for growth (Knibb etal. 1997b; 1998a,b). A new program has recently beeninitiated to assess strains and crossbred among strainsof cultured sea bass (Dicentrarchus labrax) in Israel. Thisinvolves comparative assessment of the performanceand culture potential of a new Egyptian and existingdomesticated strains, in order to identify the mostsuitable to commence a long-term selective breedingprogram for sea bass.
Interspecific hybrids between S. aurata and redseabream, Pagrus major (both belonging to the Sparidaefamily) have been produced (Knibb et al. 1998a). Thehybrids developed only vestigial gonads at the age 2and 3 years and were sterile. Subsequently, similarvestigial gonads were observed in offspring of thereciprocal crosses. No consistent growth (and survival)superiority until sexual maturity was detected in thereciprocal crosses, compared with parental species.Hybridization between European sea bass (D. labrax)females and striped bass (Morone saxatilis) males wascarried out. Viable hybrid larvae were produced, butsurprisingly 28% were triploids, and apparently onlytriploids survived to age of 6 months. At the age 8months, surviving (triploid) hybrids showed poorgrowth compared to diploid D. labrax. Interspecificsterile hybrids of marine cultured fish might be ofcommercial interest when production of fertile fish isrestricted for ecological reasons (Gorshkov, pers.comm.).
A severe viral disease has affected Israeli (edible andornamental) common carp (Cyprinus carpio) stocks acouple of years ago. Commercial farms as well as theDor station, which is holding the breeding nuclei ofthe parental stocks used for producing thecommercially cultured crossbred, have reportedmassive mortalities. No final census is available yet ofsurviving broodstocks, and it may well be that years ofgenetic improvement work had been lost. So far it isnot clear whether survivors from affected ponds caninherit tolerance to this disease to their offspring.Farmers are currently trying to produce resistant seedby hybridizing surviving carp broodstocks with Cruciancarp (Carassius carassius), even at the cost of obtaininga slower growing fish. It is possible that a breeding

105Israeli Aquaculture Genetic Inprovement Programs
program for common carp will have to be restarted tocope with the catastrophe.
Color inheritance
Investigations on color pattern inheritance are beingconducted in various aquaculture species, includingornamental (koi) carp (Gomelsky et al. 1995; 1996;1998a; David et al., work in progress); goldfish(Rothbard et al. 1999a); grass carp, Ctenopharyngodonidella (Rothbard and Shelton 1999); tilapia (Shirak etal. 2000) and guppies, Poecilia reticulata (Froyman andHulata, work in progress).
Various mutations which affect body coloration (e.g.,‘golden’, ‘albino’ and ‘ebony’) were isolated in S. aurata,some having pleiotropic effects (Knibb et al. 1996;1998a). While ‘ebony’ homozygotes are semi-lethal,heterozygotes show strong heterosis for growth.
Chromosome-set and sexmanipulations
Methods of chromosome-set manipulations areemployed for genetic improvement of variousaquaculture species: ornamental (koi) and ediblecommon carp, black carp (Mylopharyngodon piceus),tilapias (O. aureus and O. niloticus), white bass (M.chrysops), gilthead seabream (S. aurata), European seabass (D. labrax) and white grouper (Epinephelus aeneus).Work on edible common carp, carried out at the Dorstation (ARO), was directed towards establishingbroodstock producing all-female populations (Cherfaset al. 1996). This involved sex reversing XXgynogenetic females to males (Gomelsky et al. 1994),and using these XX males for breeding. Such all-femaleseed was released to commercial farms and resulted in10-15% yield improvement over existing commercialstocks. Triploid common carp were produced and theirculture potential evaluated (Cherfas et al. 1994). Thesurvival in shock treated progenies (predominantlytriploids) was about 70% of that in diploid control.Most of the one-year-old triploid males and femaleshad undeveloped gonads and were sterile. Triploidsgrew slower than their diploid sibs under allinvestigated conditions. The results did not reveal theexpected positive effect of sterility on somatic growthin triploid common carp, thence the potential of steriletriploid common carp for aquaculture is questionable.
Work on ornamental (koi) carp was aimed atinvestigating the inheritance of color patterns(Gomelsky et al. 1995; 1996; 1998a). Manipulations
are aimed at producing inbred lines, and as a tool inapplication of DNA-markers for establishment ofimproved koi broodstock (research in progress).Successful meiotic and mitotic gynogenesis wasachieved when the eggs were exposed to, either, heat/cold, or to pressure shocks. An interesting pheno-menon detected during the investigation was thepresence of males among gynogenetically producedfish. These, when sib-mated with their sisters, producedall-female progeny [Rothbard (Gan Shmuel FishBreeding Center), pers. comm.]. The objectives ofandrogenesis were to produce homozygous koiindividuals (XX and YY) of both sexes, and to preservekoi genetic traits in a sperm bank. Androgenesis ofkoi was induced in edible carp eggs, immersed in carpovarian fluid to avoid stickiness, by UV-irradiationfollowed by insemination with koi sperm. Low numbers(2-12) of diploid androgenotes, recognizable at the frystage by light pigmentation, were obtained (Rothbardet al. 1999b).
Studies involving black carp (M. piceus) are aimed atproducing triploid, presumably sexually sterile fish,as well as mono-sex female or male populations(through application of chromosome-set mani-pulations, e.g. gynogenesis, androgenesis, triploidyand tetraploidy). Combined technology of gynogenesisand hormonal sex-reversal, enables production ofXXX-triploid fish that are completely sterile, unlikethe XXY- or YYX-triploids that possess fragments oftestes, and are thus able to produce some active sperm(Rothbard et al., 1997). The black carp is a high qualityedible fish, which is also highly valued for its ability tocontrol snail populations [e.g., the golden snail(Pomacea sp.), zebra mussel (Dreissena polymorpha), orsnail infestation in the Israeli National Water Carriersystem, causing filter and pump clogging](Rothbardand Rubinstein 1999). The methodology enablesproducing ecologically and economically importantexotic species that can potentially serve for biocontrol,without endangering local fish fauna.
Gynogenesis and sex-reversal were successfully inducedin Morone sp. in an attempt to obtain broodstocksproducing monosex populations to avoid limitationson transfers of this exotic species (Gomelsky et al.1998b; 1999). This project was unfortunatelyterminated before reaching commercial application.
A meiogynogenetic line of O. aureus was establishedand gynogenetically propagated for 5 generations atthe Faculty of Life Sciences, Bar-Ilan University. Asuccessive increase in viability recorded over these 5

Fish Genetics Research in Member Countries and Institutions of INGA106
generations was interpreted as a gradual elimination oflethal genes. Mitogynogenetic O. aureus were produced(Shirak et al. 1998) using third generationmeiogynogenetic females from this stock. Threegenerations of gynogenetic O. niloticus were alsoproduced. Males from the gynogenetic O. aureus linewere used for hybridization with gynogenetic O.niloticus females, resulting in consistent production of100% male hybrids (Avtalion, pers. comm.).
Triploids and meiotic gynogenetic D. labrax and S.aurata were efficiently produced using temperatureshocks (Gorshkova et al. 1995; 1996; 1998). Gorshkovaet al. (1996) found nearly 20% males in meioticgynogenetic D. labrax cohorts. Recently, a highincidence (up to 60%) of severe cranial bonedeformities were recorded in gynogenetic cohorts atage 10 months (Knibb et al. 1998a). Fertilizing heat-shocked S. aurata eggs with untreated heterologoussperm from Pagrus major produced triploid hybrids(Gorshkova et al., 1995). However, no growthsuperiority of triploid hybrids, compared to eitherparent, was evident for fish up to age 2 and 3 years. D.labrax is characterized with sexual dimorphism, withfemales tending to grow faster than males, so there isconsiderable interest in culturing only females.Monosex populations were produced by administrationof estradiol (yielding all-females) and 17a-methyltestosterone (yielding all-males) to mixed-sexjuveniles (Gorshkova et al. 1996).
Karyological analyses of the major maricultured speciesunder cultivation in Israel are being conducted.Recently, series of experiments were undertaken toattempt the cytological technique for examination ofearly embryogenesis for the purpose of understandingthe genetic reasons of high egg and larval mortality inthe white grouper (E. aeneus). The proportion ofcytogenetically abnormal embryos carrying differenttypes of chromosomal aberrations vary significantlyin different spawnings of the parental fish. Cytologicalmonitoring of early embryogenesis is carried tosupport the selective breeding program and the geneticmanagement (stock identification and production ofmonosex populations) in the white grouper and othercultured marine species (Gorshkova et al. inpreparation).
No karyological evidence for a chromosomalmechanism responsible for sexual differentiation wasfound in European sea bass. However, males usuallyhave a heteromorphic pair of subtelocentricchromosomes, while females usually have a
homologous pair of those chromosomes. Temperaturemanipulation experiments are considerably effectivein changing sex ratio in offspring (at the age of 11months). While data could not identify any particulargenetic and/or environmental reason for the observeddeviations of sea bass sex ratios, they do illustratepossible plasticity of sex determination in this species(work in progress; Gorshkov and Gorshkova, pers.comm.).
Genetic engineering andtransgenesis
Investigations on a few candidate genes have beeninitiated and/or are in progress. Most noteworthy isthe work on the effect of growth hormone (GH),engineered for expression using various promoters,on common carp and gilthead seabream growth(Cavari et al. 1993a,b; Moav et al. 1995; Fine et al.1996; Hinits and Moav, 1999). Moav (Dept. of Zoology,Tel Aviv university) and coworkers have demonstrated20% growth improvement in transgenic carp rearedin experimental earthen ponds. Current efforts areaimed at constructing new expression vectors toimprove efficiency of producing transgenic fish.Further potential economically important genesconsidered for genetic engineering in marine fishinclude genes for D5 and D6 fatty acid desaturaseenzymes (Knibb et al. 1996).
DNA markers and QTL mapping
Collaborative work with US scientists (Hulata et al.1999; Agresti et al. 2000) using the [Om x (Oa x ROn)]cross resulted in mapping 191 AFLP and 26 UNHmicrosatellite markers to 24 linkage groups. TwentyUNH microsatellite markers were used to search forquantitative trait loci (QTL) associated with coldtolerance and body weight in an F2 family of O. aureusx O. mossambicus hybrid. Two markers putativelyassociated with cold tolerance and three markersputatively associated with body weight were found. Twoof these markers are located on the same linkage group,which is apparently a chromosomal region, whichaffects growth and survival in tilapia (Cnaani et al.1999, 2000). In a similar study (Agresti et al., inpreparation) using cold tolerance-tested [(Om x Oa) x(Sg x On)] 4WC, one UNH microsatellite and two AFLPmarkers from the female gave evidence of associationwith body weight; these were all from different linkagegroups. Seven markers (3 UNH microsatellite and 4AFLP) from the male also were found to be associatedwith body weight and six of them were restricted to

107Israeli Aquaculture Genetic Inprovement Programs
two linkage groups. Based on shared microsatellitemarkers, 12 composite linkage groups have beenidentified from the combined mapping data of Kocheret al. (1998), Agresti et al. (2000) and the 4WC family.These likely represent 12 of the 22 tilapia chromosomes.
Attempts are also being made at detecting sex linkageand non-Mendelian segregation of microsatellite DNAmarkers in a meiogynogenetic family of O. aureus (Paltiet al. 1999, 2000), and at analyzing genetic variationin immunological parameters associated with stressresistance (Palti et al. 2000, and work in progress).
Work is currently under way for development of DNAmarkers in edible and ornamental (koi) carp. This workis aimed at detecting linkage between DNA markersand color patterns in the ornamental carp (David andRothbard, pers. comm.).
Studies are under way to apply molecular biology tosupport the classical breeding program (specificallyfor determining parenthood in mass-spawning of S.aurata and D. labrax, and for genetic testing programs)by developing AFLP and microsatellite DNA profilingtechniques (Gorshkova, pers. comm.).
Gamete cryopreservation and genebank
Methods have been developed at the National instituteof Oceanography (IOLR, Haifa), for cryopreservationof carp sperm (Lubzens et al. 1993; 1997), leading tothe formation of an operating ‘sperm bank’ for C. carpio(mainly ornamental carp) in Israel. Subsequently,methods for cryopreservation of gilthead seabream,mullet (Mugil cephalus) and grouper (E. aeneus) spermhave also been developed (Lubzens, pers. comm.).
Attempts are currently directed towards developingcryopreservation methods for gilthead seabream(Lubzens and Pekarsky 1998) and marine shrimp(Penaneus semisulcatus) embryos. Successfulcryopreservation of yolk-laden eggs containingdeveloping fish embryos has yet to be achieved.Obstacles encountered so far are the low permeabilityof cryoprotectants into the embryos and sensitivity offertilized eggs to concentrations above 0.15M of DMSOor methanol. Attempts are currently being made atevaluating possible cryopreservation of ovarianoocytes, as an alternative (Lubzens and Pekarsky 1998).
References
Agresti, J.J., S. Seki, A. Cnaani, S. Poompuang, E.M. Hallerman,N. Umiel, G. Hulata, G.A.E. Gall, and B. May. 2000. Breedingnew strains of tilapia: development of an artificial center oforigin and linkage map based on AFLP and microsatelliteloci. Aquaculture. 185: 43-56.
Ben-Dom, N., N.B. Cherfas, B. Gomelsky and G. Hulata. 2000.Genetic stability of Israeli common carp stocks inferred fromelectrophoretic analysis of transferrin, phosphoglucomutaseand glucose-6-phosphate isomerase. Isr. J. of Aquacult./Bamidgeh. 52: 30-35.
Cavari, B., B. Funkenstein, T.T. Chen, L.I. Gonzales-Villasenor andM. Schartl. 1993a. Effect of growth hormone on the growthrate of the gilthead seabream (Sparus aurata), and use ofdifferent constructs for the production of transgenic fish.Aquaculture. 111: 189-197.
Cavari, B., B. Funkenstein, B. Moav and M. Schartl . 1993b. All-fish gene constructs for growth hormone gene transfer in fish.Fish Physiol. and Biochem. 11: 345-352.
Cherfas, N.B., B. Gomelsky, N. Ben-Dom, Y. Peretz and G. Hulata.1994. Assessment of triploid common carp for culture.Aquaculture 127: 11 - 18.
Cherfas, N.B., B. Gomelsky, N. Ben-Dom, D. Joseph, S. Cohen, I.Israel, M. Kabessa, G. Zohar, Y. Peretz, D. Mires and G. Hulata.1996. Assessment of all-female common carp progenies forfish culture, p.30-31. Isr. J. of Aquacult./Bamidgeh. 48: 149-157.
Cnaani, A., G. Hulata, E.M. Hallerman and G.A.E. Gall. 1999.Characterizing genetic variation in cold tolerance in tilapias,p.30-31. In Towards Predictable Quality, Aquaculture Europe99, Eur. Aquacult. Soc. Spec. Publ. No. 27.
Cnaani, A., G.A.E. Gall and G. Hulata. 2000. Cold tolerance oftilapia species and hybrids. Aquacult. Int. 8: 289-298.
Fine, M., D. Zilberg, Z. Cohen, G. Degani, B. Moav and A. Gertler.1996. The effect of dietary protein level, water temperatureand growth hormone administration on growth andmetabolism in the common carp (Cyprinus carpio). Comp.Biochem. Physiol. 114A: 35-42.
Gomelsky, B., N. Cherfas and G. Hulata. 1998a. Studies on theinheritance of black patches in ornamental (koi) carp. Isr. J.Aquacult./Bamidgeh. 50: 134-139.
Gomelsky, B.I., N.B. Cherfas, G. Hulata and N. Ben-Dom. 1995.Color variability in normal and gynogenetic progenies ofornamental (Koi) common carp (Cyprinus carpio L.).Aquaculture. 137: 102 (Abstract).
Gomelsky, B., N.B., Cherfas, N. Ben-Dom and G. Hulata. 1996.Color inheritance in ornamental (koi) carp (Cyprinus carpioL.) inferred from color variability in normal and gynogeneticprogenies. Isr. J. Aquacult./Bamidgeh. 48: 219-230.
Gomelsky, B., N. Cherfas and G. Hulata. 1998a. Studies on theinheritance of black patches in ornamental (koi) carp. Isr. J.of Aquacult/Bamidgeh. 50: 134-139.
Gomelsky, B., N. Cherfas, A. Gisis and G. Hulata. 1998b. Induceddiploid gynogenesis in white bass, Morone chrysops. Prog. Fish-Cult. 60: 288-292.
Gomelsky, B., N. Cherfas, A. Gisis and G. Hulata. 1999. Hormonalsex inversion in striped bass and white x striped bass hybrids.N. Am. J. Aquacult. 61: 199-205.
Gomelsky, B., N.B. Cherfas, Y. Peretz, N. Ben-Dom and G. Hulata.1994. Hormonal sex inversion in the common carp (Cyprinuscarpio L.). Aquaculture, 126: 265 - 270.
Gorshkov, S., H. Gordin, G. Gorshkova and W. Knibb. 1997.Reproductive constraints for family selection of the giltheadseabream (Sparus aurata L.). Isr. J. Aquacult./ Bamidgeh. 49:124-134.

Fish Genetics Research in Member Countries and Institutions of INGA108
Gorshkov, S., G. Gorshkova, A. Hadani, H. Gordin and W.R.Knibb. 1998. Chromosome set manipulations andhybridization experiments in gilthead seabream (Sparusaurata). I. Induced gynogenesis and intergenerichybridization using males of the red seabream (Pagrus major).Isr. J. Aquacult./Bamidgeh. 50: 99-110.
Gorshkov, S., G. Gorshkova, W. Knibb and H. Gordin. 1999. Analysisof sex ratios and growth performance of the European seabass (Dicentrarchus labrax L.) reared in mariculture in Eilat (RedSea). Isr. J. Aquacult./ Bamidgeh. 51: 91-105.
Gorshkova, G., S. Gorshkov, A. Hadani, H. Gordin and W.R. Knibb.1995. Chromosome set manipulations in marine fish.Aquaculture 137: 157-158.
Gorshkova, G., S. Gorshkov, H. Gordin and W. Knibb. 1996. Sexcontrol and gynogenetic production in European sea bassDicentrarchus labrax. p. 288-290. In B. Chatain, B. Saroglia,M. Sweetman and P. Lavens (eds.). Handbook of contributionsand short communications presented at the InternationalWorkshop on Seabass and Seabream Culture: Problems andProspects, 16-18 October 1996, Verona, Italy. Eur. Aquacult.Soc. Spec. Publ., Belgium.
Hinits, Y. and B. Moav. 1999. Growth performance studies intransgenic Cyprinus carpio. Aquaculture 173: 285-296.
Hulata, G. 1995. The history and current status of aquaculturegenetics in Israel. Isr. J. Aquacult./Bamidgeh.. 47: 142-154.
Hulata, G., A. Cnaani, N. Umiel, J.J. Agresti, S. Seki, B. May, S.Poompuang, E.M. Hallerman and G.A.E. Gall. 1999.Development of a tilapia artificial center of origin and geneticlinkage map based on AFLP and microsatellite loci. p. 93-94.In Towards predictable quality, Aquaculture Europe 99. Eur.Aquacult. Soc. Spec. Publ. No. 27.
Knibb, W., G. Gorshkova and S. Gorshkov. 1996. Potential gainsthrough genetic improvement: selection and transgenesis. p.175-188. In B. Chatain, B. Saroglia, M. Sweetman and P.Lavens (eds.). Seabass and Seabream Culture: Problems andProspects. Eur. Aquacult. Soc. Spec. Publ., Belgium.
Knibb, W., G. Gorshkova and S. Gorshkov. 1997a. Growth in strainsof gilthead seabream (Sparus aurata L.).Isr. J. Aquacult.Bamidgeh. 49: 43-56.
Knibb, W., G. Gorshkova and S. Gorshkov. 1997b. Selection forgrowth in the gilthead seabream (Sparus aurata L.). Isr. J.Aquacult./ Bamidgeh. 49: 57-66.
Knibb, W., G. Gorshkova and S. Gorshkov. 1998a. Geneticimprovement of cultured marine finfish: case studies. p. 111-149. In S. De Silva (ed.). Tropical Mariculture Academic PressLtd., San Diego, California.
Knibb, W., G. Gorshkova and S. Gorshkov. 1998b. Selection andcrossbreeding in Mediterranean cultured marine fish. p. 47-60. In D.M. Bartley and B. Basurco (eds.). Proc. TECAMSeminar on Genetics and Breeding of MediterraneanAquacultured Species, 34. Zaragoza, Spain.
Kocher, T.D., W-J. Lee, H. Sobalewska, D. Penman and B.J.McAndrew. 1998. A genetic linkage map of a cichlid fish,the tilapia (Oreochromis niloticus). Genetics 148: 1225-1232.
Lahav, E. and Z. Ra’anan. 1997. Salinity tolerance of geneticallyproduced tilapia (Oreochromis) hybrids. Isr. J. Aquacult./Bamidgeh. 49: 160-165.
Lubzens, E. and I. Pekarsky. 1998. Cryopreservation of marine fishembryos: obstacles and future directions. Isr. J. Aquacult./Bamidgeh. 50: 207.
Lubzens, E., S. Rothbard and A. Hadani. 1993. Cryopreservationand viability of spermatozoa from the ornamental Japanesecarp (Nishikigoi). Isr. J. Aquacult./Bamidgeh. 45: 169-174.
Lubzens, E., N. Daube, I. Pekarsky, Y. Magnus, A. Cohen, F.Yusefovich and P. Feigin. 1997. Carp (Cyprinus carpio L.)spermatozoa cryobanks: Strategies in research andapplication. Aquaculture 155: 13-30.
Moav, B., Y. Hinits, Y. groll and S. Rothbard. 1995. Inheritance ofrecombinant carp b-actin/GH cDNA gene in transgenic carp.Aquaculture 137: 179-189.
Palti, Y., A. Shirak, A. Cnaani, M. Ron, R.R. Avtalion and G. Hulata.1999. Sex linkage and non-Mendelian segregation ofmicrosatellite DNA markers in a meiogenetic family anddomesic strains of tilapia (Oreochromis aureus and O. niloticus).Paper presented at the Plant & Animal Genome VII, 17-21January 1999. Conference, San Diego, California, USA.
Palti, Y., S. Tinman, A. Cnaani, Y. Avidar, M. Ron and G. Hulata.2000. Comparative study of biochemical and non-specificimmunological parameters in two tilapia species (Oreochromisaureus and O. mossambicus). Isr. J. Aquacult./Bamidgeh, 51: 148-156.
Palti, Y., A. Shirak, A. Cnaani, E. Feldmesser, R.R. Avtalion, G. Hulataand M. Ron. 2001.A microsatellite locus has more than onecopy in the genome of two tilapia species (Oreochromis aureusand O. niloticus). Animal Genetics (In press).
Rothbard, S. and I. Rubinstein. 1999. The black carp, a potentialbiocontrol of mollusks. Isr. J. Aquacult./Bamidgeh 51: 74-83.
Rothbard, S and W.L. Shelton. 1999. The albino grass carp. Trop.Fish. Hobbyist. 45(6): 150-159.
Rothbard, S., I Rubinstein and L. David. 1999a. Coloration incrosses between two variants of goldfish, Carassius auratus. Isr.J. Aquacult./Bamidgeh. 51: 133-140.
Rothbard, S., I. Rubinstein, L. David and W.L. Shelton. 1999b.Ploidy manipulations aimed to produce androgeneticJapanese ornamental (koi) carp, Cyprinus carpio L. Isr. J.Aquacult./Bamidgeh. 51: 26-39.
Rothbard, S., W.L. Shelton, Z. Kulikovsky, I. Rubinstein, Y. Haganiand B. Moav. 1997. Chromosome-set manipulations in theblack carp. Aquacult. Int. 5: 51-56.
Shirak, A., J. Vartin, J. Don and R.R. Avtalion. 1998. Production ofviable mitogynogenetic Oreochromis aureus using cold shockand its optimization through definition of cleavage time. Isr.J. Aquacult./Bamidgeh. 50: 140-150.
Shirak, A., A. Shmarina, Y. Kuperman and R.R. Avtalion. 2000.Inheritance of body and peritoneum color in hybrids ofOreochromis aureus and red O. niloticus. Isr. J. Aquacult./Bamidgeh. 52: 21-29.

109
ABSTRACT
Auburn University’s fish genetics program has four major research projects in aquaculture genetics, namely: (1)
selective breeding, (2) genomic mapping and marker-assisted selection (MAS), (3) gene cloning and (4) transgenic
fish production, all with the long-term goal to produce fish broodstocks with improved economic traits. In addition
to aquaculture interest, the university has a population genetics program with the goal of proper management and
preservation of natural fisheries resources. However, this program will not be discussed in this report.
The aquaculture genetics projects were developed based on long-term efforts of selective breeding programs.
These projects are dependent upon one another so that the maximum benefits will be obtained by using the
integrated genetic improvement approaches. For instance, gene mapping will produce more economically important
genes for cloning and application in biotechnology or gene-assisted selection (GAS), and produce markers that
are linked to important performance traits for MAS. Biotechnology should use strains that are already developed
through selective breeding to obtain the best performance traits. Further genetic improvements by gene transfer
into an already superior strain provide the best “starting point” for genetic engineering. Regardless of the selection
approaches to be used, traditional or marker-assisted, selection of one trait should not take the expense of another.
Thus, a comprehensive selection index should be developed based on relative economic importance of each trait.
This report focuses on genomic mapping, marker-assisted selection and genetic engineering. Auburn University
has a strong selective breeding program and the genetics team works collaboratively in all projects, directed by a
philosophy of integrated genetic improvement. For the most part, this paper will only discuss some of the progress
on the work with catfish and, to a less extent, tilapia.
Z. J. LiuFish Molecular Genetics and Biotechnology Laboratory
Department of Fisheries and Allied AquacultureAuburn University, Alabama 36848, USA
LIU, Z. 2001. Gene mapping, marker-assisted selection, gene cloning, genetic engineering and integrated genetic improvementprograms at Auburn University, p. 109-118. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in membercountries and institutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179p.
GENE MAPPING, MARKER-ASSISTEDSELECTION, GENE CLONING,
GENETIC ENGINEERINGAND INTEGRATED GENETIC IMPROVEMENT
PROGRAMS AT AUBURN UNIVERSITY
Gene Mapping, Marker-Assisted Selection, Gene Cloning, Genetic Engineeringand Integrated Genetic Improvement Programs at Auburn University

Fish Genetics Research in Member Countries and Institutions of INGA110
Catfish is the most important cultured fish in the USand accounts for over 50% of all US aquacultureproduction. The catfish industry is valued at over US$2billion and production in 1999 should exceed 750million pounds. It is the only agricultural sector witha steady annual growth rate of over 8%. In Mississippi,Alabama, Arkansas, Louisiana, Georgia and severalother southern states, catfish is one of the topagricultural commodities. Tilapia culture is relativelysmall in the country, although ranked as the secondor third largest cultured finfish.
Despite the development of the aquaculture and catfishindustry in the US, a large trade deficit, US$3-6 billion,exists annually for seafood products. This deficit isstill increasing. Aquaculture appears to be more andmore important, especially considering the collapsingnatural fisheries (Anon. 1996, 1997). According to theUS Department of Agriculture (DA) estimates, thedemand for seafood is increasing steadily and wildfisheries will be able to supply only 25-30% of theadditional demand (USDA ERS 1995). Futureprojections predict a steadily widening gap betweenthe world’s demand for fish and the ability of theoceans to meet it (Anon. 1996). Development of aprofitable, productive, environmentally sound andsustainable aquaculture industry, therefore, providesan alternative to the already overexploited, collapsingnatural fisheries.
Despite its importance, the catfish industry is still ayoung industry and suffers from various productionproblems. A recent survey by Auburn UniversityExtension Systems indicated that disease problems areranked as the top concern of the catfish industry. “Tokeep my fish alive” is the greatest wish of all farmers(Jensen, unpubl.). Disease problems cause over 30%loss of the industry each year, equivalent to a loss ofseveral hundred million dollars annually. Otherconcerns include traits in growth, feed conversionefficiency, carcass and fillet yields, tolerance to lowdissolved oxygen, tolerance to poor water quality,reproductive success and harvestability. Broodstockswith enhanced culture traits are urgently needed for asustainable catfish aquaculture. However, most, if notall, economic targets are polygenic and quantitativeand controlled by quantitative trait loci (QTL).Understanding genomic organization and geneticlinkage and QTL mapping is required for marker-
assisted selection, improvements of economic traitsthrough biotechnology and retrogression of beneficialgenes from channel catfish (Ictalurus punctatus) andblue catfish (I. furcatus). Auburn University’s researchaddresses all three areas.
The research goal is to provide necessary scientific andtechnological information for improving the catfishbroodstocks by MAS, introgression of important QTLsfrom channel catfish and blue catfish and geneticengineering using beneficial genes. To reach this goal,initial steps are to construct genetic linkage maps usingvarious polymorphic markers and establish linkagesof QTLs with markers. This linkage informationshould be useful for MAS programs. Development ofthe catfish genetic linkage map has been identified asone of the most important research for aquaculture bythe USDA Aquaculture Steering Committee and it,among mapping of four other aquaculture species, wasjust approved by USDA as a regional project (NE-186)(Liu and Dunham 1998; Waldbieser et al. 1998).
Second, physical maps of catfish will be constructedso that major genes for important QTLs can beeventually isolated and used for improving broodstocksthrough the more effective GAS or biotechnology.Beneficial genes from various sources have been usedto improve genetic stocks of catfish and developtechnology needed for production of improved stocksthrough genetic engineering (Hallerman et al. 1990;Yoon et al. 1990a, 1990b; Hayat et al. 1991; Chen etal. 1992, 1993; Dunham et al. 1992; Gross et al. 1992;Powers et al. 1992; Chatakondi et al. 1994, 1998a,1998b; and Dunham 1995).
Third, linkage analysis using polymorphic DNAmarkers and channel catfish x blue catfish interspecifichybrids should produce information on QTLs that canbe introgressed from both species to generate syntheticbreeds that can grow faster, have higher feedconversion efficiencies and higher carcass yields, andare more resistant to diseases and other environmentalstresses.
In spite of dramatic development in many livestockanimals, genomic mapping of aquaculture species isstill at its infancy (Liu and Dunham 1998; Waldbieseret al. 1998). However, gene mapping in catfish istimely since well-developed, efficient marker systemsare now available. The plan is to generate a catfishgene map with a resolution of 2-5 cM from the USDA-supported project, which would have been virtually
Current Status of Catfishand Tilapia Aquaculture

111
impossible, or required much more resources severalyears ago. In addition, background researchconcerning genetics of economic trait loci (ETL)has been completed in the university’s laboratory andelsewhere (Dunham and Smitherman 1983a, 1983b,1987; Tiersch et al. 1990; Dunham et al. 1993b;Tiersch and Goudi 1993; Wolters and Johnson 1994;Dunham 1996; Wolters et al. 1996; Liu et al. 1998a,1998b, 1998c). The catfish genetics research andbreeding programs, heritability studies and availabilityof large numbers of markers, make mapping ofimportant ETLs in catfish immediately feasible.Development of linkage maps is certainly important,but QTLs will also be mapped for four importanteconomic traits that will be immediately useful for MASprograms in catfish, in addition to generating a geneticlinkage map of catfish.
Genetic improvement of catfish is a proven method ofaddressing production problems such as diseases.Previous research in the areas of traditional selectivebreeding and molecular genetics has resulted ingenetically improved catfish (Dunham et al. 1983a,1983b, 1987, 1990, 1992; Dunham 1996) and fourreleases of it to the industry. In addition to diseaseresistance, growth, feed conversion efficiency andprocessing yields are also addressed. Faster growth ratewould allow for more crops per unit of time, thus savingmanagement cost while increasing productivity. Feedaccounts for 60% of the variable costs of catfish farmingand, therefore, any improvement would significantlyimprove profit margins; processing yields areimportant since a 1% increase translates into US$20-30 million net income annually.
Economic traits, genetic variation andheritability
Domestic channel and blue catfish exhibit significantphenotypic and genetic variation for economic traitssuch as disease resistance, growth rate, feed conversionefficiency (found highly correlated with growth),environmental stress tolerance, carcass yield, seinabilityand reproduction (Dunham et al. 1982, 1983, 1984,1985, 1987a, 1990, 1993; Dunham and Smitherman1983a, 1983b, 1987; Bondari 1984; Hallerman et al.1986; Wolters and Johnson 1994; Dunham 1996;Wolters et al. 1996). Auburn University established acatfish genetics research program in 1969 to evaluate
traditional selective breeding and molecular geneticsfor improving these quantitative traits. Growth rateand feed conversion efficiency have been improvedby as much as 50% through selection (Bondari 1983;Dunham and Smitherman 1983a, 1987; Rezk 1993;Padi 1995); intraspecific crossbreeding (Bondari 1983,1984; Dunham and Smitherman 1983b); interspecifichybridization (Dunham et al. 1990; Dunham 1996);and genetic engineering (Dunham et al. 1992, 1995;Dunham 1996). Disease resistance has also beenimproved primarily through interspecific hybridization(Dunham et al. 1990; Argue 1996; Dunham 1996;Wolters et al. 1996), intraspecific crossbreeding andstrain selection (Wolters and Johnson 1994). Toleranceto low oxygen was enhanced primarily by interspecifichybridization (Dunham et al. 1983; Dunham 1996);seinability, by interspecific hybridization (Dunham1996) and strain selection; and carcass yield by strainselection (Dunham et al. 1984), hybridization(Dunham 1996; Argue and Dunham 1998) and indirectselection (Rezk 1993; Dunham et al. 1985).Heritabilities and genetic correlations have beencalculated (Argue 1996; Argue and Dunham 1998).The rationale for creating genetic maps of catfish is toincrease the efficiency of selection (Waldbieser et al.1998). Breeders wish to find molecular markerscorrelated with genetic loci controlling economic traitsand use these markers to select superior broodstocks.Because traits, such as growth rate, are relatively easyto measure with traditional selection, a genetic mapwill be more useful to select fish for traits for whichmeasurement is difficult or expensive (e.g., diseaseresistance) and is lethal to broodstocks (e.g., carcasscomposition). Such map will also be useful forintrogression of alleles into channel catfish from otherspecies with which hybrid production is feasible suchas blue catfish.
Polymorphic marker development andevaluation
In the last three years, various polymorphic markerswere evaluated for their usefulness in catfish genemapping. These markers include allozymes, restrictionfragment length polymorphism (RFLP), expressedsequence tags (EST), random amplified polymorphicDNA (RAPD), microsatellites, amplified fragmentlength polymorphism (AFLP) and single nucleotidepolymorphism (SNuPs). Allozyme markers are type Imarkers and should be highly useful as anchoragepoints for comparative mapping. Total numbers ofpolymorphic loci are small and polymorphic rates are
Gene Mapping, Marker-Assisted Selection, Gene Cloning, Genetic Engineeringand Integrated Genetic Improvement Programs at Auburn University
Research Progress in GenomicMapping

Fish Genetics Research in Member Countries and Institutions of INGA112
low at each locus. RFLP markers are codominantmarkers. However, previous knowledge is required,such as probes for Southern analysis or sequences forpolymerase chain reaction (PCR) facilitated RFLPanalysis. Only a few RFLP markers are available foruse in catfish mapping. We do not anticipate majorincreases in numbers of RFLPs because developingRFLP markers is slow and costly. Our results usingEST from a pituitary cDNA library indicated lowpolymorphism between channel and blue catfish(about 10%). After sequencing and PCR analysisof 100 cDNA clones, 11 EST markers were obtained(Karsi et al. 1998; Liu et al. 1999e). RAPD markerswere evaluated (Welsh and McClelland 1990; Williamset al. 1990) for their application in catfish genemapping. Polymorphic rates are low among strains ofchannel catfish, but high between channel andblue catfish (Liu et al. 1999a). Therefore, RAPD shouldbe only useful for mapping using the interspecifichybrid system. Inheritance of RAPD markers wasnormal following Mendelian expectations (Liu et al.1998b). To date, 142 primers have been tested and682 polymorphic markers have been identified usingthe channel catfish x blue catfish hybrids (Liu et al.1998b, 1999a). Reproducible results with RAPDmarkers of sizes 200-1 500 bp were obtained. Thesemarkers should be useful for gene mapping analysisin the future.
Microsatellites are abundant, highly polymorphic,easy for genotyping with PCR and are reliable becauseof their codominance. The progress on developingmicrosatellite markers from channel catfish issummarized in Table 1, and efforts on this are beingcontinued. Six microsatellite-enriched, small-insertlibraries were constructed with channel catfish genomicDNA using the procedure of Orstrander et al. (1992).Two of the six libraries were screened using radio-actively labeled (CA)15 and (GA)15 oligonucleotideprimers. A total of 1 530 microsatellite-containingclones (900 CA and 630 GA) were purified. PlasmidDNA containing microsatellites was prepared from 890clones (500 [CA], 390 [GA]). To date, 590 clones havebeen sequenced, of which 403 generated enoughflanking sequences to design PCR primers (Liu et al.1999d; Tan et al. 1999, in review). In more than 100clones, microsatellite sequences were immediately atthe cloning site making them useless. There were 403pairs of primers for their performance in amplifyingchannel catfish genomic DNA. Of this total, 257 pairssuccessfully amplified genomic DNA and generatedPCR products of expected sizes. High levels ofheterozygosity were observed.
As will be discussed below, the channel catfish x bluecatfish hybrid system offers great advantages for genemapping. However, one requirement for usingmicrosatellite markers with the interspecific system isthat the microsatellite flanking sequences must beconserved between the two species. This issue wasaddressed by determination of amplifiability of bluecatfish DNA using PCR primers designed fromchannel catfish microsatellite sequences. Over 90%loci were amplified from both species using channelcatfish primers. In fact, most of the microsatelliteflanking sequences have been conserved across thegenus borders of Ictalurid catfish (Liu et al. 1999d).This indicates that more than 230 microsatelliteswill be useful for mapping with the catfish interspecificsystem. The conservation of microsatellite loci betweenchannel and blue catfish is important for practicalapplications. This will allow a comprehensive linkagemap to be constructed using the interspecific hybridsystem and various types of markers: allozymes, RFLP,EST, RAPD, AFLP, microsatellites and SNuPs.
In addition to type II microsatellite markers, type Imicrosatellite markers were also developed. Type Imarkers represent genes of known functions, thus,these are more useful for comparative gene mapping.A pituitary cDNA library was enriched formicrosatellite-containing clones. Fifty clones of cDNAscontaining microsatellite CA or GA repeats wereobtained, which belong to 15 unique cDNAs aftersequencing. Most clones harbor microsatellitesequences at their 3’ nontranslated regions (NTR)or 5’-NTR, but two clones harbor CCA and CA repeatsin their coding regions (Liu et al., in review).
AFLP markers (Vos et al. 1995) combine the strengthsof RFLP and RAPD markers and overcome theirproblems. The approach is PCR-based and requiresno probe or previous sequence information as neededby RFLP. It is reliable because of high stringent PCRin contrast to RAPD’s problem of low reproducibility.The weakness is that they are dominant markers thuson average half of which are useful for a givenbackcross reference family. For gene mapping in
Table 1. Summary of progress on microsatellite markerdevelopment from I. punctatus.
Microsatellite-enriched library 6Microsatellite clones purified 1530Clones plasmid DNA prepared 890Clones sequenced 590Loci primers ordered 403Loci PCR worked 257

113
catfish, AFLP polymorphic markers are highlyabundant between channel catfish and blue catfish.They are inherited in the interspecific hybrids asdominant markers and segregated normally accordingto Mendelian ratios (Liu et al. 1998c). Some 64 primercombinations were tested and over 3 000 AFLPmarkers suitable for genetic mapping of catfish wereproduced (Liu et al. 1999c).
SNuP markers were evaluated several months ago. Todate, the university has sequenced 100 each of cDNAsfrom the channel and blue catfish muscle cDNAlibraries. Preliminary results indicate that SNuPs(type 1 in this case since they are cDNAs) are abundantusing the catfish interspecific hybrid system.
In summary, feasibility of using seven types ofpolymorphic markers in gene mapping of catfish wasevaluated. Microsatellite and AFLP are the two mostuseful types of markers for catfish mapping. Mappingthe available microsatellite and AFLP markers shouldgenerate a map with less than 2 cM resolution.
Resource/reference families
Although channel catfish is the major cultured catfish,channel catfish x blue catfish hybrid system offers greatadvantages. The F1 hybrid is fertile and in fact hasproduced F2, F3 and various backcrosses (Argue 1996;Liu et al. 1997; Dunham and Argue 1998a, 1998b,1998c, in press). They are a major resource for QTLmapping and for MAS. Backcross progeny designedfor this QTL mapping project was successfullyproduced. Sixteen backcross families (eight fromchannel and eight from blue backcrossed withheterozygous F1) were produced. These families arereared in 0.1acre ponds ready for QTL evaluation.The interspecific hybrid system for gene mapping ofcatfish is advantageous because: (1) high rates ofpolymorphic markers are assured to exist between blueand channel catfish; (2) several important ETLs arepossessed by blue catfish, mainly, the disease resistancegene(s) to enteric septicemia of catfish (ESC), carcassyield genes and genes controlling better seinability(Dunham et al. 1993); mapping these important genesis of great importance by itself; (3) mapping ETLs inblue catfish is important to selective breedingprograms using backcrossing to introgress beneficialgenes from blue catfish into channel catfish; and (4)drastic phenotypic variation of the hybrid system offerstools to be exploited for easy QTL evaluation andsegregation.
Genotyping
Genotyping, using AFLP and microsatellite markers,is in progress now. It is reasonable to assume thatin about a year, a linkage map for catfish should beproduced.
Two types of experiments toward marker-assistedselection have been initiated. The first is within theefforts for genetic linkage mapping. Phenotypes wereevaluated for all the reference families to be used formapping with the following traits: growth, bodyconformation (including body length, width anddepth; head length, width and depth; caudal widthand depth), disease resistance to ESC and tocolumnaris. The selective genotyping approach is usedto have a strong phenotype selection pressure on thepotential genotype differences. The top and bottom12.5% of the individuals in each trait will be genotyped.It is reasonable to expect that markers linked to QTLscontrolling these traits will be differential between thebest and the worst performers.
The second approach is a whole-genome QTL scanfrom outcrossing populations. Again, if markers arelinked to a specific QTL, they should harbor variablealleles between the best and the worst performers.To date, markers that are linked to QTLs controllinggrowth and QTLs controlling feed conversionefficiency have been identified. These identifiedmarkers, upon confirmation, will be highly usefulfor marker-assisted selection programs. Equallyimportant, they will be useful for checking the successof traditional selective breeding programs. Selectionfor growth, for instance, should produce populationsthat are highly enriched with alleles linked withfast-growing QTLs.
Genomic research has two ultimate applied goals:marker-assisted selection and cloning genes ofeconomic importance. The Auburn Universitylaboratory has been active in isolating channel catfishgenes involved in growth, development andreproduction (Tang et al. 1993; Liu et al. 1997; Karsiet al. 1998).
Gene Mapping, Marker-Assisted Selection, Gene Cloning, Genetic Engineeringand Integrated Genetic Improvement Programs at Auburn University
Research Progress in QTL Mappingand Marker-assisted Selection
Research Progress in Gene CloningEfforts and Aquaculture GenomicsOther than Mapping

Fish Genetics Research in Member Countries and Institutions of INGA114
In the last four years, genes for gonadotropin alpha-subunit (Liu et al. 1997), gonadotropin beta-subunit Iand beta-subunit II (Liu et al. 2001) were cloned. Twoclasses of Tc1-like transposable elements named IpTc1and IpTc2 (Liu et al., in press) and a class ofnonautonomous transposons from channel catfish wereisolated and characterized (Liu et al. 1999b). Anotherclass of highly repetitive elements, named as Xbaelements, was characterized, accounting for about 5%of the catfish genome (Liu et al. 1998a). Over 300cDNA clones were sequenced for EST markerdevelopment. These efforts are being continued, andin the near future, it can be expected that over 1 000catfish genes will be cloned and partially sequenced.
Research Progress in Transgenic Fish
The Auburn University laboratory was one of thepioneers in the world to transfer foreign genes in fish(Dunham et al. 1987b; Dunham 1990; Liu et al. 1990a,1990b, 1991). It has demonstrated the effects of thetransfer of growth hormone genes into warmwater fishin terms of growth, protein content, carcass yield,oxygen tolerance, disease resistance, fat reduction, andenhancement of flavor and texture (Zhang et al. 1990;Hayat et al. 1991; Dunham 1992; Dunham et al. 1992;Powers et al. 1992; Chen et al. 1993; Chatakondi et al.1994). The laboratory has also demonstrated thattransgenic catfish does not pose an environmental riskin regards to growth without supplemental feed,predator avoidance and reproduction (Dunham1995, 1996). There is greater potential for geneticimprovement through the use of genes identified fromAuburn’s mapping project, which applied traditionalselective breeding and molecular genetics. Maximumprogress will likely be made by combining the toolsof selective breeding with genetic engineering(Dunham 1992), using information obtained fromgenomic research.
Auburn University is currently working on homologoustransfer of growth hormone gene from channel catfishinto channel catfish, with promoters of various kinds,including metallothineine, beta-actin, elongationfactor 1 and histone H3 (Moav et al. 1991, 1992a,1992b). Also ongoing is work on gene transfer ofantibacterial peptide genes to fish to enhanceresistance against bacterial pathogens.
The two most significant problems in transgenicstudies are high levels of mosaicism and inactivationof transgene expression after gene transfer. The
university is developing expression vectors using theborder elements to prevent gene inactivation, andusing the Sleeping Beauty transposase system toenhance temporal integration. Upon development,these technologies should circumvent much of thecurrent problems in transgenic fish production. Asdiscussed above, the gene mapping projects willidentify and essentially clone more economicallyimportant genes, including disease-resistant ones.
In collaboration with the International Centerfor Living Aquatic Resources Management, a tilapiaproject entitled “Genetic Enhancement of Tilapiain Africa by Combined Selection, QTL Mappingand Marker-assisted Selection” is being undertaken.The goal is to genetically enhance Nile tilapia(Oreochromis niloticus), the most widespread farmedtilapia in sub-Saharan Africa, to increase production,efficiency, profit margins and quality of life of the poorfish farmers and consumers. Thus, the project willalso meet the needs for food security in Africa andinitiate decreasing child and infant malnutrition. Thespecific objectives of this research are:(1) determinationof heritabilities and genetic correlations for importantperformance traits such as growth, feed conversionefficiency, stress tolerance and sexual maturation; (2)development of selection indices; (3) gene mappingof important QTL; (4) initial evaluation of thefeasibility of marker-assisted selection; and (5) trainingof African scientists in quantitative and moleculargenetic techniques. This three-year project isbeginning.
Future Perspectives
Radiation hybrid mapping
Although the idea of irradiation and fusion genetransfer was published over 20 years ago (Goss andHarris 1975), the technology was underutilized untilresurrected by Cox et al. (1990) for applications ingenomic research by combining it with efficientgenotyping using PCR. Because of its extreme powerfor gene mapping, radiation hybrid (RH) panels arerecognized as a milestone in human genomicresearch and is now regarded as the ultimate tool forcorrection of marker orders for linkage mappingin mammals (Womack et al. 1997). Extremely finechromosome maps have been constructed in humans
Genetic Enhancement ofTilapia in Africa

115
(e.g., Schuler et al. 1996, reviewed by McCarthy1996), mouse (McCarthy et al. 1997), and bovine(Womak et al. 1997). Recently, RH panels have beenreported for rat, baboon, dog (Priat et al. 1998),pig (Yerle et al. 1998), chicken and zebrafish (Chevretteet al. 1997; Kwok et al. 1998). No RH panels areavailable for aquaculture species yet. In catfish, RHpanels are desperately needed as an initial geneticlinkage map is nearing. Undoubtedly, the develop-ment of RH in catfish will accelerate its genomicmapping and progress in genetic improvement ofcatfish broodstocks. A tilapia RH panel is underdevelopment elsewhere.
Physical mapping using large insert libraries
The second step after construction of a genetic linkagemap is to focus on chromosomes that containimportant QTLs controlling disease resistance, growth,feed conversion efficiency and processing yields.Catfish have 29 pairs of homogenous chromosomes(LeGrande et al. 1984) which make it extremelydifficult to microdissect for development ofchromosome-specific markers. Therefore, the strategyis to first identify the chromosomes containingimportant QTLs of interest. Then, large insertlibraries, such as BAC library, will be used to tagmarkers linked to specific QTLs to specific BAC clones.Once the anchorage points are found, contigs of theBAC clones can be established rapidly surroundingthe linked markers. This would allow development ofchromosome-specific or regional markers for finemapping of QTLs from the associated BAC clones.The work on physical mapping started in late 1999.
Research Teams and Facilities
A major strength of Auburn University’s research isthat it combines the expertise of two complementaryresearch teams specialized in molecular genetics formolecular marker development, evaluation, andgenotyping, gene cloning, sequencing and expressionvector development, and in catfish aquaculture forstrain selection, phenotypic evaluation, fish culture andmaintenance, QTL measurement and evaluation,population and quantitative genetic analysis,transgenic fish production and rearing, and evaluationof environmental impact of transgenic fish.
Auburn University has the best facility in the nationfor simulated commercial conditions for catfish geneticresearch such as QTL mapping. A large earthen pond
facility, with 128 experimental research ponds, isavailable for evaluation of performance under closelysimulated commercial conditions. This is importantsince genotype-environment interactions can occur ingenetic evaluations of catfish requiring pondexperiments to obtain realistic results.
Auburn University’s Fish Genetics Research Unit alsohas the best facility to develop transgenic fish foraquaculture application. A large earthen pond facilitywith 28 ponds, secured with nets and filters approvedby USDA for transgenic fish studies, is available forevaluation of performance of transgenic fish. This isthe only government-approved outdoor testing facilityfor transgenic fish in the world. Its features includeconcrete stabilized walls, chain-link fence and barbedwire surrounding the complex, bird netting, commondrain line, quadruple filtration, static conditions andFrench drain located in a seepage pond. The pondsare located 35 feet above the 100-year flood level, anda 17- acre barrier pond is located below these ponds.
A 6 500 ft2 hatchery contains 300 tanks for indoorspawning, incubation and initial QTL evaluations.
The 6 500 ft2 Molecular Genetics and BiotechnologyLaboratory includes a computer room, main laboratory(radioactive safety approved), refrigerated room, darkroom and tissue culture room. Two offices are attachedto the laboratory. The recently acquired automatedDNA sequencer (LI-COR IR2 system) is equipped withgenotyping apparatus for both microsatellite and AFLPanalysis and genotyping software. The laboratory alsohas all the equipment for molecular biology: molecularcloning, PCR, restriction analysis, gel electrophoresis,DNA sequencing, blotting, hybridization, geneexpression studies of RNA and proteins,radioimmunoassays (RIA), ELISA and Westernblotting. Other major equipment include: four PCRcyclers, a microinjection system with Brinkman MM33micromanipulator controlled by a Medical SystemsCorporation pico-injector Model PLI-100, a BackeonModel 2000 for electroporation, a Beckman DU20spectrophotometer, a Sorvall RC-6B clinical centrifuge,a Beckman J2-21 centrifuge, a Beckman GS-15centrifuge, five microfuges, computerized low and highvoltage power supplies for electrophoresis, two 37°CCO2/humidified incubators, two -80°C freezers, one -
20°C freezer, one analytical balance, one 37°C shakerincubator, one 37°C incubator for dish culture of E.coli, UV and visible transilluminators with camera, aLeitz microscope, two dissecting microscopes, one ice
Gene Mapping, Marker-Assisted Selection, Gene Cloning, Genetic Engineeringand Integrated Genetic Improvement Programs at Auburn University

Fish Genetics Research in Member Countries and Institutions of INGA116
maker, autoclave, three biosafety cabinets, one of whichis devoted as a radioactive work station, CAT assaytanks and a scintillation counter. The followingcomputer hardware are available - Power Mac 7100,Power Book Mac and two IBM computers, with twolaser printers. Available are DNA analysis softwarepackages, such as DNAStar and DNASIS and freeaccess to GenBank, EMBO and other databases, andto GCG software packages with minimal cost. AuburnUniversity also has excellent core facilities for oligosynthesis, sequencing, TaqMan PCR, confocalmicroscope and EM facilities with minimal costs.
References
Anon. 1997. Fisheries science review. Nature 386: 105-110.Argue, B. 1996. Performance of channel catfish Ictalurus punctatus,
blue catfish I. furcatus, and their F1, F2, F3 and backcross hybrids.Auburn University, Auburn, Alabama. Ph.D. dissertation.
Bondari, K. 1983. Response to bidirectional selection for bodyweight in channel catfish. Aquaculture 33: 73-81.
Bondari, K. 1984. Reproductive performance, growth, and survivalof selected and wild x selected channel catfish. Theor. Appl.Genet. 68: 391-395.
Chatakondi, N., A.C. Ramboux, A. Nichols, M. Hayat, P.L. Duncan,T.T. Chen, D.A. Powers and R.A. Dunham. 1994. The effect ofrainbow trout growth hormone gene on the morphology,dressing percentage and condition factor in the common carp,Cyprinus carpio, p. 481-484. In Proceedings of the Fifth WorldCongress on Genetics Application and Livestock Production.Vol. 17.
Chen, T.T., C.M. Lin, R.A. Dunham and D.A. Powers. 1992.Integration, expression and inheritance of foreign fish growthhormone gene in transgenic fish, p. 164-175. In C.L. Hewand G.L. Fletcher (eds.) Transgenic fish. World ScientificPublishing Co., River Edge, New Jersey.
Chen, T.T., D.A. Powers, C.M. Lin, M. Hayat, N. Chatakondi, A.Ramboux, P.L. Duncan and R.A. Dunham. 1993. Expressionand inheritance of pRSVrtGH1 cDNA in common carp,Cyprinus carpio. Mol. Mar. Biol. Biotech. 2: 88-95.
Chevrette, M., L. Joly, P. Tellis and M. Ekker. 1997. Contribution ofzebrafish-mouse cell hybrids to the mapping of the zebrafishgenome. Biochem. Cell Biol. 75: 641-649.
Cox, D.R., M. Burmeister, E. Royden Price, S. Kim and R.M. Myers.1990. Radiation hybrid mapping: a somatic cell geneticmethod for constructing high-resolution map of mammalianchromosomes. Science 250: 245-250.
Dunham, R.A. 1990. Genetic engineering in aquaculture. AgBiotechNews Info. 2: 401-406.
Dunham, R.A. 1992. Outlook for genetics research and applicationin aquaculture, p. 137-148. In Proceedings of AgricultureOutlook ’92: New Opportunities for Agriculture. USDepartment of Agriculture. Vol. 68.
Dunham, R.A. 1995. Predator avoidance, spawning and foragingability of transgenic catfish, p. 151-169. In M. Lerin, C. Grimand J.S. Angle (eds.) Proceedings of the Biotechnology RiskAssessment Symposium, 22-24 June 1994. US Department ofAgriculture, EPA, College Park, Maryland.
Dunham, R.A. 1996. Contribution of genetically improved aquaticorganisms to global food security. Paper presented at theInternational Conference on Sustainable Contribution ofFisheries to Food Security. Government of Japan and FAO,Rome, Italy. 4-9 Dec. 1995, Kyeto, Japan.
Dunham, R.A. and B.J. Argue. Anal fin ray count and anal fin shape
of channel catfish Ictalurus punctatus, blue catfish I. furcatusand their F1, F2, F3 and backcross hybrids. (In press, a).
Dunham, R.A. and B.J. Argue. Evaluation of reproduction amongchannel catfish Ictalurus punctatus, blue catfish I. furcatus andtheir F1 and F2 hybrids. (In press, b).
Dunham, R.A. and B.J. Argue. Seinability of channel catfish, bluecatfish and their F1, F2, F3 and backcross hybrids in ponds.Prog. Fish Culturist. (In press, c).
Dunham, R.A. and R.O. Smitherman. 1983a. Crossbreedingchannel catfish for improvement of body weight in earthenponds. Growth. 47: 97-103.
Dunham, R.A. and R.O. Smitherman. 1983b. Response to selectionand realized heritability for body weight in three strains ofchannel catfish, Ictalurus punctatus, grown in earthen ponds.Aquaculture 33: 88-96.
Dunham, R.A. and R.O. Smitherman. 1987. Genetics and breedingof catfish. Reg. Res. Bull. 325. South. Coop. Ser. AlabamaAgricultural Experiment Station, Auburn University, Alabama.
Dunham, R.A., R.O. Smitherman and C. Webber. 1983. Relativetolerance of channel x blue hybrid and channel catfish to lowoxygen concentrations. Prog. Fish-Culturist 45: 55-56.
Dunham, R.A., R.O. Smitherman and R.K. Goodman. 1987a.Comparison of mass selection, crossbreeding and hybridizationfor improving body weight in channel catfish. Prog. Fish-Culturist 49: 293-296.
Dunham, R.A., J.A. Joyce, K. Bondari and S.P. Malvestuto. 1985.Evaluation of body conformation, composition and density astraits for indirect selection for dress-out percentage of channelcatfish. Prog. Fish Culturist 47: 169-175.
Dunham, R.A., J. Eash, J. Askins and T.M. Townes. 1987b. Transferof metallothionein-human growth hormone fusion gene intochannel catfish. Trans. Am. Fish. Soc. 116: 87-91.
Dunham, R.A., M. Benchakan, R.O. Smitherman and J.A. Chappell.1984. Correlations among morphometric traits of 11-month-old blue, channel, white, and hybrid catfishes and therelationship to dressing percentage at 18-months of age. J.World Maricult. Soc. 14: 668-675.
Dunham, R.A., R.E. Brummett, M.O. Ella and R.O. Smitherman.1990. Genotype-environment interactions for growth of blue,channel and hybrid catfish in ponds and cages at varyingdensities. Aquaculture 85: 143-151.
Dunham, R.A., R.O. Smitherman, M.J. Brooks, M. Benchakan andJ.A. Chapell. 1982. Paternal predominance in channel-bluehybrid catfish. Aquaculture 29: 389-396.
Dunham, R.A., C. Hyde, M. Masser, R.O. Smitherman, R. Perez, J.Plumb and A.C. Ramboux. 1993. Comparison of culture traitsof channel catfish, Ictalurus punctatus, and blue catfish, I.furcatus. J. Appl. Aquacult. 3: 257-267.
Dunham, R.A., A.C. Ramboux, P.L. Duncan, M. Hayat, T.T. Chen,C.M. Lin, K. Kight, I. Gonzalez-Villasenor and D.A. Powers.1992. Transfer, expression and inheritance of salmonid growthhormone genes in channel catfish, Ictalurus punctatus, andeffects on performance traits. Mol. Mar. Biol. and Biotech. 1:380-389.
Goss, S.J. and H. Harris. 1975. New method for mapping genes inhuman chromosomes. Nature 255: 680-684.
Gross, M., J. Schneider, N. Moav, C. Alvarez, S. Myster, Z. Liu, E.Hallerman, P. Hackett, K. Guise, A. Faras and A. Kapuscinski.1992. Molecular analysis and growth evaluation of transgenicnorthern pike. Aquaculture 103: 253-273.
Hallerman, E., J. Schneider, M. Gross, Z. Liu, S.J. Yoon, L. He, P.Hackett, A. Faras, A. Kapuscinski and K. Guise. 1990. Geneexpression promoted by the RSV long terminal repeat elementin transgenic gold fish. Anim. Biotechnol. 1: 79-93.
Hayat, M., C.P. Joyce, T.M. Townes, T.T. Chen, D.A. Powers andR.A. Dunham. 1991. Survival and integration rate of channelcatfish and common carp embryos microinjected with DNA atvarious developmental stages. Aquaculture 99: 249-255.

117
Karsi, A., P. Li, R. Dunham and Z.J. Liu. 1998. Transcriptionalactivities in the pituitaries of channel catfish (Ictalurus punctatus)before and after induced ovulation as revealed by expressedsequence tag analysis. J. Mol. Endocrinol. 21: 121-129.
Kwok, C., R.M. Korn, M.E. Davis, D.W. Burt, R. Critcher, L.McCarthy, B.H. Paw, L.I. Zon, P.N. Goodfellow and K. Schmitt.1998. Characterization of whole genome radiation hybridmapping resources for non-mammalian vertebrates. Nucl.Acids Res. 26: 3562-3566.
LeGrande, W., R.A. Dunham and R.O. Smitherman. 1984.Comparative karyology of three species of north Americancatfishes (Siluriformes: Ictaluridae: Ictalurus) and four of theirhybrid combinations. Copeia 1984: 873-878.
Liu, Z.J. and R. Dunham. 1998. Genetic linkage and QTL mappingof ictalurid catfish. Circ. Bull. 321: 1-19. Alabama AgriculturalExperiment Station, USA.
Liu, Z.J., A. Karsi and R.A. Dunham. 1999e. Development ofpolymorphic EST markers suitable for genetic linkagemapping of catfish. Mar. Biotechnol. 5(i):437-447.
Liu, Z.J., P. Li and R. Dunham. 1998a. Characterization of an A/T-rich family of sequences from the channel catfish (Ictaluruspunctatus). Mol. Mar. Biol. Biotechnol. 7: 232-239.
Liu, Z.J., A. Nichols, P. Li and R. Dunham. 1998c. Inheritance andusefulness of AFLP markers in channel catfish (Ictaluruspunctatus), blue catfish (I. furcatus) and their F1, F2 andbackcross hybrids. Mol. Gen. Genet. 258: 260-268.
Liu, Z.J., P. Li, B. Argue and R. Dunham. 1997. Gonadotropinsubunit glycoprotein from channel catfish (Ictalurus punctatus)and its expression during hormone-induced ovulation. Mol.Mar. Biol. Biotechnol. 6: 221-231.
Liu, Z.J., P. Li, B. Argue and R. Dunham. 1998b. Inheritance ofRAPD markers in channel catfish (Ictalurus punctatus), bluecatfish (I. furcatus) and their F1, F2 and backcross hybrids. Anim.Genet. 29: 1-5.
Liu, Z.J., P. Li, B.J. Argue and R.A. Dunham. 1999a. Randomamplified polymorphic DNA markers: usefulness for genemapping and analysis of genetic variation of catfish.Aquaculture. 174:59-65.
Liu, Z.J., P. Li, H. Kucuktas and R.A. Dunham. 1999b.Nonautonomous Tc1-like transposable elements of channelcatfish (Ictalurus punctatus) contain sequence blocks similar tothe para-type sodium channel proteins. Fish Physiol. Biochem.21: 65-72
Liu, Z., B. Moav, A. Faras, K. Guise, A. Kapuscinski and P. Hackett.1990a. Development of expression vectors for transgenic fish.BioTechnology 8: 1268-1272.
Liu, Z., B. Moav, A. Faras, K. Guise, A. Kapuscinski and P. Hackett.1990b. Functional analysis of elements affecting expressionof the actin gene of carp. Mol. Cell Biol. 10: 3432-3440.
Liu, Z., B. Moav, A. Faras, K. Guise, A. Kapuscinski and P. Hackett.1991. Importance of CArG boxes in regulation of the carp-actin gene expression. Gene 108: 211-216.
Liu, Z.J., S. Kim, P. Li, G. Tan and A. Karsi 2001. Channel catfishgonadotropin-subunits: cDNA cloning and their expressionduring ovulation. Mar. Biotechnol.(In press).
Liu, Z.J., G. Tan, H. Kucuktas, P. Li, A. Karsi, D.R. Yant and R.A.Dunham. 1999d. High levels of conservation at microsatelliteloci among Ictalurid catfishes. J. Heredity. (90)2: 307-311.
Liu, Z.J., P. Li, H. Kucuktas, A. Nichols, G. Tan, X. Zheng, B.J.Argue, R. Yant and R.A. Dunham. 1999c. Development ofAFLP markers for genetic linkage mapping analysis usingchannel catfish and blue catfish interspecific hybrids. Trans.Am. Soc. Fish. 128(2): 317-327.
McCarthy, L.C. 1996. Whole genome radiation hybrid mapping.Trends Genet. 12(12): 491-493.
McCarthy, L.C., J. Terrett, M.E. Davis, C.J. Knights, A.L. Smith, R.Critcher, K. Schmitt, J. Hudson, N.K. Spurr and P.N.Goodfellow. 1997. A first-generation whole genome-radiation
hybrid map spanning the mouse genome. Genome Res. 7:1153-1161.
Moav, B., Z. Liu and P. Hackett. 1992a. Selection of promoters forgene transfer into fish. Mol. Mar. Biol. Biotechnol. 1: 338-345.
Moav, B., Z. Liu, A. Faras, K. Guise, A. Kapuscinski and P.B. Hackett.1992b. Regulation of early expression of transgenes indeveloping fish. Transgen. Res. 2: 153-161.
Moav, B., Z. Liu, N. Moav, A. Faras, K. Guise, A. Kapuscinski and P.Hackett. 1991. Expression of heterologous genes in transgenicfish, p. 121-140. In C. Hew (ed.) Transgenic fish. WorldScientific Publishing Co., New Jersey.
Ostrander, E.A., P.M. Jong, J. Rine and G. Duyk. 1992. Constructionof small-insert genomic DNA libraries highly enriched formicrosatellite repeat sequences. Proc. Natl. Acad. Sci. 89:3419-3423.
Padi, J.N. 1995. Response and correlated responses to fourgenerations of selection for increased body weight in theKansas strain channel catfish, Ictalurus punctatus, grown inearthen ponds. Auburn University, Alabama, USA. M.S. thesis.
Powers, D.A., T. Cole, K. Creech, T.T. Chen, C.M. Lin, K. Kightand R.A. Dunham.1992. Electroporation: a method oftransferring genes into the gametes of zebrafish, Brachydaniorerio, channel catfish, Ictalurus punctatus, and common carp,Cyprinus carpio. Mol. Mar. Biol. Biotech. 1: 301-308.
Priat, C., C. Hitte, F. Vignaux, C. Renier, Z. Jiang, S. Jouquand, A.Cheron, C. Andre and F. Galibert. 1998. A whole-genomeradiation hybrid map of the dog genome. Genomics 54: 361-378.
Rezk, M.S. 1993. Response and correlated responses to threegenerations of selection for increased body weight in channelcatfish, Ictalurus punctatus. Auburn University, Alabama, USA.Ph.D. dissertation.
Schuler, G.D., M.S. Boguski, E.A. Stewart, L.D. Stein, G. Gyapay,K. Rice, R.E. White, P. Rodrigueztome, A. Aggarwal, E. Bajorekand 94 coauthors. 1996. A gene map of the humanchromosome. Science 274: 540-546.
Tang, Y., C.M. Lin, T.T. Chen, H. Kawauchi, R.A. Dunham andD.A. Powers. 1993. Structure of the channel catfish (Ictaluruspunctatus) growth hormone gene and its evolutionaryimplications. Mol. Mar. Biol. Biotechnol. 2: 198-206.
Tiersh, T.R. and C.A. Goudie. 1993. Inheritance and variation ofgenome size in half-sib families of hybrid ictalurid catfishes.J. Heredity 84: 122-125.
Tiersh, T.R., B.A. Simco, K.B. Davis, R.W. Chandler, S.S. Wachteland G.J. Carmicheal. 1990. Stability of genome size amongstocks of the channel catfish. Aquaculture 87: 15-22.
USDA. 1995. Aquacultural situation and outlook report. USDepartment of Agriculture Economic Research Service.
USDA. 1996. Aquacultural situation and outlook report. USDepartment of Agriculture Economic Research Service.
Vos, P., R. Hogers, M. Bleeker, M. Reijans, T. van de Lee, M. Hornes,A. Frijters, J. Pot, J. Peleman, M. Kuiper and M. Zabeay. 1995.AFLP: a new technique for DNA fingerprinting. Nucl. AcidsRes. 23: 4407-4414.
Waldbieser, G., Z.J. Liu, W. Wolters and R. Dunham.1998. Genetic linkage and QTL mapping ofcatfish, p. 31-38. In Proceedings of theNational Aquaculture Species Gene MappingWorkshop.
Welsh, J. and M. McClelland 1990. Fingerprinting genomes usingPCR with arbitrary primers. Nucl. Acids Res. 18: 7213-7218.
Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski and S.V.Tingey. 1990. DNA polymorphisms amplified by arbitraryprimers are useful as genetic markers. Nucl. Acids. Res. 18:6531-6535.
Wolters, W.R. and M.R. Johnson. 1994. Enteric septicemiaresistance in blue catfish and three channel catfish strains. J.
Gene Mapping, Marker-Assisted Selection, Gene Cloning, Genetic Engineeringand Integrated Genetic Improvement Programs at Auburn University

Fish Genetics Research in Member Countries and Institutions of INGA118
Aquat. Anim. Health 6: 329-334.Wolters, W.R., D.J. Wise and P.H. Klesius. 1996. Survival and
antibody response of channel catfish, blue catfish, and catfishhybrids after exposure to E. ictaluri. J. Aquat. Anim. Health8: 249-254.
Womack, J.E., J.S. Johnson, E.K. Owens, C.E. Rexroad III, J.Schläpfer and Y.-P. Yang. 1997. A whole-genome radiationhybrid panel for bovine gene mapping. Mammalian Genome8: 854-856.
Yerle, M., P. Pinton, A. Robic, A. Alfonso, Y. Palvadeau, C. Delcros,R. Hawken, L. Alexander, C. Beattie, L. Schook, D. Milanand J. Gellin. 1998. Construction of a whole-genome radiationhybrid panel for high-resolution gene mapping in pigs.Cytogenet. Cell Genet. 82: 182-188.
Yoon, S.J., Z. Liu, A. Kapuscinski, P. Hackett, A. Faras and K. Guise.1990a. Successful gene transfer in fish. Proceedings of theSixteenth US-Japan Natural Resources Panel on AquacultureSymposium. NOAA Tech. Rep. NMFS 92: 39-44.
Yoon, S.J., E. Hallerman, M. Gross, Z. Liu, J. Schneider, A. Faras, P.Hackett, A. Kapuscinski and K. Guise. 1990b. Transfer of the
gene for neomycin resistance into goldfish, Carassius auratus.Aquaculture 85: 21-33.
Zhang, P., M. Hayat, C. Joyce, L.I. Gonzalez-Villasenor, C.M. Lin,R. Dunham, T.T. Chen and D.A. Powers. 1990. Gene transfer,expression and inheritance of pRSV-rainbow trout-GHcDNAin the carp, Cyprinus carpio (Linnaeus). Mol. Reprod. Dev. 25:3-13.

119
ABSTRACT
The development of a live gene bank of domesticated carps was started in the Fish Culture Research Institute,Szarvas, Hungary, in 1963, when common carp (Cyprinus carpio) strains were collected from different regions of thecountry. Later, the stock of the live gene bank was enlarged with common carp strains from different parts of Europeand Asia. At present, there are 20 Hungarian and 15 foreign races and strains in the gene bank. Besides the commonlycultivated carps, wild races are also maintained; those are native in the two major Hungarian Rivers, Danube andTisza. The broodstocks of 15 foreign races and strains are also available in the gene bank, mostly from Europe, e.g.,the Czech mirror and scaly carps, the Croatian Nasic and Poliana carps, the scaly carp of Ukraine and the RussianRopsha carp that has a long history in production.
Some Asian common carp races can also be found in the gene bank: scaly carps from Vietnam and Thailand, wildcarp from River Amur, and Japanese koi carp. Besides common carps, the pure lines of silver carp (Hypophthalmichthysmolitrix) and bighead carp (Aristichthys nobilis) are also available here.
The original goals of collection of common carp strains were: (i) to compare their characteristics within identicalenvironmental and farming conditions; (ii) to use them in the selection and hybridization work; and (iii) to produce andsupply high-value races, strains and parental lines of hybrids to preserve the strains with their original genetic structureas a potential gene reserve.
GENE BANKINGAND COMMON CARP
BREEDING PROGRAM IN HUNGARY
S. Gorda and L. VáradiFish Culture Research Institute
5541, Szarvas, P.O. Box 47, Hungary
GORDA, S and L. VARADI. 2001. Gene banking and common carp breeding program in Hungary, p. 119-122. In M.V.Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of the InternationalNetwork on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
In the early stage of the institute’s experimental work,comparison tests on the productivity and characteristicsof the collected Hungarian strains were carried out.The performance of strains was determined on thebasis of the performance of their progenies. Significantqualitative and quantitative differences have beenfound among the various strains.
Gene Banking and Common Carp Breeding Program in Hungary
Broodstocks of strains are kept in eight earthen pondsof 1 ha each, with an average depth of 1.2 m. Eachpond with continuous water exchange throughout theyear holds about 200-250 breeders. The breeders arefed ad libitum except for those chosen forreproduction. These breeders are fed with special feedhaving high animal protein content, for 2-2.5 monthsbefore the spawning season.

Fish Genetics Research in Member Countries and Institutions of INGA120
The minimum population size of the breeding stockof each race and strain in the gene bank is 50individuals. When the size of the breeding stock isnot less than 50 individuals, the inbreeding coefficient(F) is quite low (F= 0.01%) per generation.
Each strain of the gene bank is renewed in 8-10years or the population is supplemented. Eggs andsperms of a minimum of 10 females and 10 males areused during reproduction, simulating panmicticpopulation conditions.
For the identification of the given races and strains inthe gene bank, group marking is applied. The mirrorcarps are marked with brand marking, while the scalyones by fin clipping. PIT tagging system is also usedfor the identification of individuals.
The establishment of cryopreserved sperm bank wasstarted in 1997, which was based on the maleindividuals of live gene bank. One male is representedwith 15-20 ml (4 x 5 ml straw) frozen sperm in liquidnitrogen. At present, there are cryopreserved spermof 68 male broodstocks.
Breeding Programs
Using the strains preserved with original gene pool,several crossing combinations and inbred lines wereproduced. Gynogenesis was also used to produce high-level inbred individuals. Since 1963, more than 400crossings have been applied to determine the bestcrossing partners and produce hybrid common carpwith high productivity. As the races, strains and hybridsare different in their qualitative and quantitativecharacteristics, a progeny performance test waselaborated for the evaluation of their productivity.
During the tests, the scale of heterosis effect wasanalyzed in the crossbred progeny. The progenygroups in the performance tests were kept in identicalenvironmental conditions for reproduction until theend of the investigation. These conditions are similarto commercial fish farms in Hungary. The progenygroups are always produced by artificial reproduction,at the same time, during the natural spawning season.After hatching, the larvae are kept in larval rearingtanks for 2-3 days until the beginning of externalfeeding. In the first year, the larvae are reared in 1 hasize ponds with a low stocking density, until they reach100-150 g body weight. At the end of the first growingseason, the individuals of the experimental groups aremarked one by one, and each group has its own mark
(fin clipping and brand marking). After marking, thegroups are stocked in the same pond. The weight lossand mortality occurring during winter are evaluated.In spring, at the beginning of the following growingseason, the fish groups are stocked in grow-out pondsfor market size fish production.
The evaluation of experimental groups is carried outnot only in the experimental ponds of the institute,but also in three commercial production farms. In thetests, five main features are evaluated which determinethe economic value of races, strains and hybrids. Theseparameters are survival, weight gain, feed conversionratio, slaughtering value (dressing yield) and fatcontent of the fish flesh. To evaluate the overallperformance of the progenies, and to make thecomparison easier, a point system is elaborated. Thirty,25, 20, 15 and 10 points can be given as a maximumfor weight gain, survival, feed conversion ratio,slaughtering value and fat content, respectively. If allfive characteristics of a tested group get the highestscore, the sum of the points equals to 100. Themaximum point for a characteristic is always given toa group that has reached the highest performanceduring the test, and the points for the other groupsare decreased proportionally.
As a result of the breeding work, three highproductivity hybrids were produced, which received astate certificate. The first hybrid was the mirror Szarvas215, produced in 1977. This hybrid was introducedto commercial production, and has been cultivatedwith good results in intensive farming conditions andcage culture in Hungary. The second hybrid, the scalySzarvas P. 31, was produced in 1978. It is aheterozygote scaly hybrid which has also been widelycultivated in Hungarian fish farms.
As a recent result of the institute’s breeding work, thethird hybrid, the two-line scaly hybrid, called SzarvasP. 34, was produced in 1989. This hybrid inherited itsexcellent feed conversion ratio and growth rate fromthe domesticated maternal line, and its high resistanceand good ability to select natural food, from the wildpaternal line. The hybrid has been produced for thestocking of angling waters, reservoirs and extensivelyutilized fishponds.
The productivity of parental lines is surpassed by morethan 20-25% by the productivity of these hybrids.
Maternal and paternal lines of high productivityhybrids have also been supplied to different countries

121
(Croatia, Czech Republic, Brazil, Bulgaria, Greece,Lao PDR, Madagascar, Poland, Rumania, Russia andVietnam), and the hybrids have proven their highproductivity and good adaptibility even in tropicalconditions.
After the political and economic changes in Hungaryin the early 1990s, several new acts and regulationshave been issued, among them the Animal BreedingAct which regulates activities related to breedingof cultured animals, including fish, through theMinistry of Agriculture and Regional Policy (MARP),National Institute for Agricultural Quality Control(NIAQC) and approved breeding organizations. Thereare specific regulations on the operation ofinsemination stations, embryo-transfer stations,poultry and fish hatcheries, and of export and importof breeders and breeding materials.
Operation of fish hatcheries is controlled by NIAQC.The basic preconditions to operate a fish hatchery,which produces larvae for sale are: license fromNIAQC, qualified hatchery manager and certificatefrom the local veterinary authority. Only pure strainsor parental lines of hybrids, with state approval andcertificate of origin, can be propagated. The hatcheryshould keep a hatchery record book containing thenumber of spawners, certificate of origin, hormonetreatment, mass of stripped and fertilized eggs,number of produced larvae and registry number ofcertificate of origin if the larvae is sold.
At present, there are 20 licensed carp hatcheries inthe country and this number is increasing. Theoperation of hatcheries is checked randomly byNIAQC. Common carp strains can get state approvalby undergoing a standardized code of carpperformance test supervised by NIAQC. During thefirst year of the two-year test, the broodstocks ofthe various strains are transported to a centralbreeding station where propagation and fingerlingrearing take place under identical conditions. Duringthe second year, the fingerling groups are testedtogether on three fish farms in different regions ofthe country. The tested traits are survival, weight gain,food conversion ratio, slaughtering value (dressingyield) and fat content.
The feed conversion ratio is tested in five wire-cagesof 165 m2 each, placed in an earthen pond stockedwith the test groups of fish. About 25 individuals ofknown body weight are stocked in each cage andfed with a feeding rate of 5% body weight throughout
the growing season. Natural food is available for thefish in the cage, just like the other fish in the pond.The food conversion ratio is also examined underlaboratory conditions in closed recycling water systemwhere feeding is based on artificial food only. Aftersecond growing season, at the time of harvesting,the fish are counted one by one and survival isexpressed in a percentage of the number of fish atstocking. The weight gain is calculated as a differencebetween weight at stocking and harvesting, when eachindividual is weighed.
Slaughtering value means the rate of edible meatof the fish mass in percentage. For determination ofthe slaughtering value, 20 individuals of each groupare tested in laboratory conditions. The fat content ofthe fish flesh is determined from the edible part byacido-butirometer analysis and expressed inpercentage. The abnormalities and body proportionsare also recorded during the harvesting andexamination of slaughtering value.
NIAQC publishes the recorded data for agriculturalquality control in the official newsletter of MARP. Todate, 17 Hungarian strains (including the above-mentioned three hybrids) have been tested and havelicense for breeding and marketing of their progenies.
In 1996 the Hungarian Carp Breeding Organisationwas established to assist and coordinate breedingprograms, organize meetings and workshops, provideconsultancy to breeding farms and present the interestof the breeders in various fora.
Other new breeding organizations, legal measuresand regulations have contributed to the improve-ment of carp breeding and product quality inHungary in the past five years. The exchange ofinformation on these specific aspects would alsocontribute to the improvement of carp breeding onan international scale.
Gene Banking and Common Carp Breeding Program in Hungary

Fish Genetics Research in Member Countries and Institutions of INGA122

123
AQUATIC GENETIC RESOURCEACTIVITIES OF THE FISHERIES
DEPARTMENT, FOOD ANDAGRICULTURE ORGANIZATION
D.M. BartleyFood and Agriculture Organization
Rome, 00100 Italy
BARTLEY, D.M. 2001. Aquatic genetics resource activities of the Fisheries Department, Food and Agriculture Organization,p. 123-127. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of theInternational Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
ABSTRACT
The sustainable use and conservation of aquatic genetic resources are key activities of the Fisheries Department inthe implementation of the Code of Conduct for Responsible Fisheries (CCRF) and other mandates of the Food andAgriculture Organization (FAO) of the United Nations. Currently, the emphasis is on normative activities under theRegular Programme. However, there are a small number of projects dealing with genetic resources under the FieldProgramme. Main areas of specialization include:(i) promotion of efficient, sustainable and responsible fisheriessector management at the global, regional and national levels in the framework of CCRF; (ii) global monitoring andstrategic analysis of fisheries to provide a sound basis for protection of production potentials, resource trends andimpacts of fisheries practices; (iii) promotion of increased contribution of responsible fisheries and aquaculture toworld food supplies and food security; and (iv) improvement of biological data on marine resources.
Introduction
The CCRF is the most prominent document providingboth the policy framework for FAO, as well as thefundamental reference and guidance document togovernments, private sector and most NGOs,regarding aquatic genetic resources in the context ofglobal fisheries and aquaculture and their sustainabledevelopment and conservation. Currently, theemphasis within the Fisheries Department is onnormative activities under the Regular Programme(Table 1). However, there are a small number of projectsdealing with genetic resources under the FieldProgramme (Table 2) and an increased number ofactivities in support of the implementation of CCRFat national level.
Regular Program
Within the Fisheries Department of FAO, the FisheryResources Division (FIR) is the lead division dealingwith issues of aquatic genetic resources; the majorityof the work is handled by the Inland Water Resourcesand Aquaculture Service (FIRI) with assistance fromthe Marine Resources Service (FIRM), the FisheryInformation, Data and Statistics Unit (FIDI) and theFishery Development Planning Service (FIPP). Thepriorities concerning fishery genetic resources are:(i) Promotion of efficient, sustainable and responsiblefisheries sector management at the global, regionaland national levels in the framework of CCRF;(ii) Global monitoring and strategic analysis of fisheriesto provide a sound basis for protection of production
Aquatic Genetic Resource Activitiesof the Fisheries Development, Food and Aqriculture Organization
Fish Genetics Research in Member Countries and Institutions of INGA124
potentials, resource trends and impacts of fisheriespractices; (iii) promotion of increased contribution ofresponsible fisheries and aquaculture to world foodsupplies and food security; and (iv) Improvement ofbiological data on marine resources.
Information on these priorities is distributed tomembers and interested parties in specializedinternational fora and in normative publications, suchas guidelines, codes of conduct and protocols,technical publications (Fisheries Technical Papers andCirculars), external scientific publications andconference proceedings, and increasingly in the FAOAquaculture Newsletter and the Fishery Department’sinternet site on WAICENT.
Promotion of responsible fisheriesand aquaculture
The Fisheries Department continues to support theimplementation of FAO CCRF and Convention onBiological Diversity (CBD) through a variety ofactivities, such as participation in specialized meetingsof CBD, publication of technical guidelines on fisheriesand aquaculture, for example on the precautionaryapproach to the use of new species in aquaculture, andorganization of international fora on fishery geneticresources. The “Towards Policies for the Conservationand Sustainable Use of Aquatic Genetic ResourcesConference” was successfully convened in 1998, withthe support of the Sustainable DevelopmentDepartment of FAO and the International Center forLiving Aquatic Resources Management (ICLARM).The conference aimed to provide guidance on policydevelopment for fishery genetic resources.
Global monitoring and strategic analysis ofinland fisheries and aquaculture
Activities under this element concern analysis of newspecies being farmed or fished and their geneticresources. A specialized online and searchabledatabase on the introduction of aquatic species (DIAS)has been added to FAO’s fisheries internet site thatprovides basic information on exotic species andsummarizes important issues. FAO fishery statistics,data from DIAS, and information and illustrationsfrom the Species Identification Programme have beenincorporated into FishBase, a relational databasedistributed on CD-ROM, by ICLARM. With theincrease in number of genetically improved speciesbeing farmed and the growing interest in geneticengineering, efforts are underway to document newtechniques, and evaluate existing legislation that may
be required for sustainable use of fisheries andequitable benefit sharing.
FAO statistics on capture fisheries and aquaculturecontinue to provide valuable information on geneticresources. There are efforts to improve these datathrough more accurate reporting and inclusion ofinformation on fish stocks and genetic resources. Afisheries global information system (FIGIS) iscurrently being designed that will integrate many ofthe components of the department which deal withfishery genetic resources.
Table 2. FAO field projects that have a significant componenton fishery genetic resources.
Project Budget Estimated(US$000) weight ofand dates fishery genetic
resourcescomponent
Genetic improvement of tilapia in 225 Very highVenezuela (TCP/VEN/6611) 1997-1998
Research for the management 1 054 Medium-lowof the fisheries on Lake 1998 – 2001Tanganyika (GCP/ AF/271/FIN)
Sustainable contribution of 900 Medium-lowfisheries to food security II 1997- 2000(GCP/INT/643/ PN)
Regional Aquaculture 797 LowDevelopment for the 1997-1998South Pacific(GCP/RAS/116/ PN)
Table 1. 1997-1998 budget allocations to regular programelements with components relevant to fishery geneticresources, and estimated weight of these components.a
Programme element Budget Estimated(US$000) weight of
fishery geneticresourcescomponents
Promotion of responsible 1 700 Medium-lowfisheries and aquaculture
Global monitoring and 1 300 Medium-lowstrategic analysis ofinland fisheries andaquaculture
Increased contribution of 2 200 Medium-lowinland fisheries andaquaculture to worldfood supplies
Improvement of biological 300 Mediumdata on marine resources (not including
staff salaries)
aProgram elements were redefined in 1998 for the 1998/1999biennium.

125
Increased contribution of inlandfisheriesand aquaculture to world food supplies
The main activities in this element involve thepublication of technical documents and consultationsthat document, characterize, and assess fisherygenetic resources and related technologies importantfor food and agriculture. For example, throughcollaboration with the International Center forAdvanced Mediterranean Agronomic Studies, areview of genetic technologies in the region waspublished, and further work involving a survey of theprivate aquaculture sector is ongoing. In cooperationwith ICLARM and the World Fisheries Trust (Victoria,British Columbia), a global information andcommunication system on aquatic animal diversityand a global strategy for the management of aquaticanimal diversity are being formulated.
Improvement of biological dataon marine resources
The Species Identification and Data Programmecontinues to promote upgrading of marine fisheriesdata and reliable species identification throughpublication of species inventories, species diagnostickeys, reference system and a readily accessibleinformation system.
Participation in interagencyand interdepartmental activities
The Fisheries Department continues to supportregional and national fisheries bodies, networks andscientific associations in promoting sustainable use andconservation of fishery genetic resources. Principalexternal partners include the Secretariat to CBD,SBSTTA (Subsidiary Body for Scientific, Technical andTechnological Advice) and expert groups of CBD, asappropriate, ICLARM, the International Network onGenetics in Aquaculture, the Consultative Group onInternational Agricultural Research System-wideGenetic Resources Programme, the World FisheriesTrust, the Asian Fisheries Society and the AmericanFisheries Society. Internally, the departmentcollaborates with interdepartmental working groups
Aquatic Genetic Resource Activitiesof the Fisheries Development, Food and Aqriculture Organization
on biosafety, ethics in food and agriculture, biologicaldiversity, inland water management, integrated coastalarea management and integrated pest management.Several activities and outputs (Annex 1) related tosustainable use and conservation of fishery geneticresources have been generated over the past threeyears. It is now time to incorporate this backgroundinformation into planning a global strategy, elementsof which will be developed in consultation withdomestic animal, crop plant and forestry sectors, aswell as within the fisheries sector in the near future.The roles of a global information system and anintergovernmental body, i.e., the Commission, will beincluded in the resulting strategy.
Table 1 lists the major allocations to program elementswithin FAO’s 1997-1998 Regular Programme budgetfor the Fisheries Department, in which substantialfishery genetic resources activities are pursued.
Annex 1. FAO-supported technicalconsultations, training courses andworkshops, and publications related tofishery genetic resources and aquaticanimal diversity.
Technical consultations supported by FAO
Technical Consultation on Inland FisheryEnhancement, in cooperation with theDepartment for International Development of theUnited Kingdom, 7-11 April 1997, Dhaka,Bangladesh.
Genetics and Breeding of Mediterranean AquacultureSpecies, in cooperation with the InternationalCenter for Advanced Mediterranean AgronomicStudies and the Network on Technology ofAquaculture in the Mediterranean, 28-29 April1997, Zaragoza, Spain.
FAO Technical Consultation on Policies for SustainableShrimp Culture, 8-11 December 1997, Bangkok,Thailand.a
Towards Policies for the Conservation and SustainableUse of Aquatic Genetic Resources, in cooperationwith the Rockefeller Foundation, ICLARM andSustainable Development Department, 14-17April 1998, Bellagio, Italy.
aFAO. 1998. Report of the Bangkok FAO Technical Consultation on Policies for Sustainable Shrimp Culture,8-11 December 1997, Bangkok, Thailand. FAO Fish. Rep. 572. Rome, Italy. Also, visit http://www.fao.org/WAICENT/FAOINFO/FISHERY/faocons/shrimp/bangk.htm

Fish Genetics Research in Member Countries and Institutions of INGA126
Training courses and workshops supported by FAO
Regional Workshop on Health and QuarantineGuidelines for the Responsible Movement(Introduction and Transfer) of Aquatic Organisms,28 January 1996, Bangkok, Thailand.
First Regional Workshop of the FAO/NACA/OIERegional Programme for the Development ofTechnical Guidelines on Quarantine and HealthCertification, and Establishment of InformationSystems for the Responsible Movement of LiveAquatic Animals in Asia, 6-20 January 1998,Bangkok, Thailand.
Ad-hoc Expert Meeting on Indicators and Criteria ofSustainable Shrimp Culture, 28-30 April 1998,Rome, Italy.
Development of an Information and CommunicationSystem on Aquatic Animal Diversity – with CGIARSystem-wide Genetic Resources Programme andWorld Fisheries Trust (British Columbia), 16-17November 1998, Rome, Italy.
Publications
1 9 9 7
Bartley, D.M. and D. Minchin. Precautionary approachto species introductions. Sweden/FAO Fish. Tech.Pap. No. 350/2: 159-189.
Bartley, D.M., K. Rana and A.J. Immink. The use ofinter-species hybrids in aquaculture and theirreporting to FAO. FAO Aquacult. Newsl. 17:7-13.
Carpenter, K.E., F. Krupp, D.A. Jones and U. Zajons.FAO species identification field guide for fisherypurposes. The living marine resources of Kuwait,Eastern Saudi Arabia, Bahrain, Qatar, and theUnited Arab Emirates. 293 p.
FAO. Review of the state of the world aquaculture. FAOFish. Circ. No. 886.
FAO. Review of the state of the world fishery resources:marine fisheries. FAO Fish. Circ. No. 929.
Humphrey, J., J.R. Arthur, R.P. Subasinghe and M.J.Phillips. Aquatic animal quarantine and healthcertification in Asia. Proceedings of the RegionalWorkshop on Health and Quarantine Guidelinesfor the Responsible Movement (Introduction andTransfer) of Aquatic Organisms, 28 January 1996,Bangkok, Thailand. FAO Fish. Tech. Pap. No. 373.
McKay, R.J. 1997. FAO species catalogue. Vol. 17. Pearlperches of the world (Family Glaucosomatidae).An annotated and illustrated catalogue of the pearlperches known to date. FAO Fish. Synop. (125)17:26 p .
1 9 9 8
Bartley, D.M. Ex situ conservation, genebanks andresponsible fisheries, p. 45–56. In B. Harvey, C.Ross, D. Greer and J. Carolsfeld (eds.) Actionbefore extinction: conserving fish genetic diversity.World Fisheries Trust, Vancouver, B.C., Canada.
Bartley, D.M. Genetics and breeding in aquaculture:current status and trends, p. 13-30. In D.M. Bartleyand B. Basurco (eds.) Genetics and breeding ofMediterranean aquaculture species. Cah. OPTIONS34.
Bartley, D. and L. Garibaldi. Database on introductionsof aquatic species website on World AgricultureInformation Centre. http://www.fao.org/waicent/faoinfo/ fishery/statist/fisoft/dias/index.htm
Bartley, D.M. and B. Basurco, Editors. Genetics andbreeding of Mediterranean aquaculture species.Cah. OPTIONS 34: 297 p.
Bartley, D.M. and K. Rana. Evaluation of artificialrehabilitation of the Caspian Sea fisheries andgenetic resource management in aquaculture andfisheries of the Islamic Republic of Iran. Reportsubmitted to Shilat, Islamic Republic of Iran. FAOField Doc.
Carpenter, K.E. and V.H. Niem. FAO speciesidentification guide for fishery purposes. Theliving marine resources of the Western CentralPacific. Vol. 1. Seaweeds, corals, bivalves andgastropods.
Carpenter, K.E. and V.H. Niem. FAO speciesidentification guide for fishery purposes. Theliving marine resources of the Western CentralPacific. Vol. 2. Cephalopods, crustaceans,holothurians and sharks.
FAO. Aquaculture production statistics 1987-1996.FAO Fish. Circ. No. 815.
FAO. Fishery statistics-catches and landings. FAOyearbook. Vol. 82.
FAO/NACA/OIE. Regional Programme for theDevelopment of Technical Guidelines onQuarantine and Health Certification, andEstablishment of Information Systems for theResponsible Movement of Live Aquatic Animalsin Asia, 16-20 January 1998, Bangkok, Thailand.TCP/RAS/6714. FAO Field Doc. No. 1.
Lloris, D. and J. Rucabado. Guide FAO d’identificationdes espèces pour les besoins de la pechê. Guidedídentification des ressources marines vivantes duMaroc. 263 p., 28 planches en couleur.
Petr, T., Editor. Inland fishery enhancements. FAO/ODA Expert Consultation on Inland FisheryEnhancement, 7-11 April 1997, Dhaka,

127
Bangladesh. FAO Fish. Tech. Pap. 374.Satia, B. and D. Bartley. Paradox of international
introductions of aquatic organisms in Africa, p.115-124. In J.F. Agnèse (ed.) Genetics andaquaculture in Africa. ORSTOM, Paris.
Subasinghe, R.P., D.M. Bartley, S. McGladdery and U.Barg. Sustainable shrimp culture development:biotechnological issues and challenges. In T.W.Flegel (ed.) Advances in shrimp biotechnology.National Centre for Genetic Engineering andBiotechnology, Bangkok.
1 9 9 9
Bartley, D.M. and R.S.V. Pullin. Towards policies foraquatic genetic resources, p. 1-16. In R.S.V. Pullin,D.M. Bartley and J. Kooiman (eds.) Towardspolicies for conservation and sustainable use ofaquatic genetic resources. ICLARM Conf. Proc. 59,277 p.
In press/in review
Bartley, D.M. A precautionary approach for theintroduction of new species in aquaculture.Thirty-third International Symposium on NewSpecies for Mediterranean Aquaculture.Societa Italiana per il Progresso dellaZootecnica, Sardinia, Italy. (In press).
Bartley, D.M. and C.V. Casal. Impact of introductionson the conservation and sustainable use ofaquatic biodiversity. European Union andAsia-Caribbean-Pacific Research Initiative,Lisbon, Portugal. (In review).
Bartley, D.M., L. Garibaldi and R.L. Welcomme.Introductions of aquatic organisms: a globalperspective and database. Paper presented tothe American Fisheries Society Symposium onImpacts, Threats and Control of IntroducedSpecies in Coastal Waters, 28 August 1997,Monterey, California. Univ. Calif. Sea GrantPubl. (In press).
Winkler, F.M., D.M. Bartley and N.F. Dias. Geneticdifferences in year class in a population ofcoho salmon (Walbaum) in Chile. Aquaculture.(In press).
Aquatic Genetic Resource Activitiesof the Fisheries Development, Food and Aqriculture Organization

Fish Genetics Research in Member Countries and Institutions of INGA128

129Activities at AKVAFORSK of Importanceto the International Network on Genetics in Aquaculture
ABSTRACT
One of the main objectives of the Institute of Aquaculture Research of Norway, Ltd. (AKVAFORSK) since itsestablishment in 1971 is to study the theoretical, biological and practical bases to start, develop and runbreeding programs. The work started with Atlantic salmon and rainbow trout in Norway. It was later transferredto a breeding company, Aqua Gen, which today supplies 70% of the Norwegian industries with improved eyedeggs. AKVAFORSK has been and is offering its competence and experience in developing breeding programsfor members of the International Network on Genetics in Aquaculture (INGA) as well as for others. In thefuture, services will be provided through AKVAFORSK Genetics Center.
The programs so far undertaken by AKVAFORSK have clearly shown that there is a large unexploited potentialfor genetic gain in all populations of fish and shellfish studied. A genetic gain of 10-20% in selection responsefor growth rate is usual, and a considerable genetic gain is also obtained by selection for disease resistanceand age at sexual maturation. By applying selective breeding, it is possible to increase productivity andsustainability in aquaculture.
ACTIVITIES AT AKVAFORSKOF IMPORTANCE TO
THE INTERNATIONAL NETWORKON GENETICS IN AQUACULTURE
T. Gjedrem and T. SveälvAKVAFORSK, Institute of Aquaculture Research Ltd.
P.O. Box 5010, 1430 Aas, Norway
GJEDREM, T. and T. SVEÄLV. 2001. Activities at AKVAFORSK of importance to the International Network on Genetics inAquaculture, p. 129-132. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries andinstitutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
AKVAFORSK is a nonprofit research institute ownedby public shareholders. Its main office is located atthe Agricultural University of Norway (AUN), 30 kmsouth of capital Oslo, while the two research stationsat Sunndalsøra and Averøy are located in the coastsome 500 km northwest of Oslo. As of 1999, theInstitute has 85 employees, of whom 28 are scientistsand 7 are Ph.D. students. One-third of the budget isbasic funding from the Research Council of Norway,
and most of the rest comes from competitive-grantresearch projects. AKVAFORSK scientists teachaquaculture at AUN and at Sunndalsøra College.
The main fields of research are nutrition, quantitativegenetics and breeding, and flesh quality; and to aless extent, preventive health care, environmentalissues and technology. The main activity in Norway isresearch on coldwater species, such as Atlantic salmon,rainbow trout and Atlantic halibut; work with oystersand scallops is planned. Internationally, AKVAFORSK

Fish Genetics Research in Member Countries and Institutions of INGA130
is engaged in many countries in Europe, SoutheastAsia and South America. The projects are in the fieldsof selective breeding and nutrition.
Simple Breeding Programs
Genetic variation has been demonstrated foreconomically important traits in several species. Aselection response in the order of 10 to 20% pergeneration is obtained for growth rate and, in someinstances, also for disease resistance. It can thereforebe concluded that it is possible by means of efficientbreeding programs to increase productivity and obtaindomestication in aquatic species.
The simplest procedure for selective breeding is touse mass selection or individual selection, which hasproved to be very efficient under certain conditions:(i) It must be possible to measure or record the traitof interest in live animals. In practice, this is true forbody weight or body size. (ii)The trait should have amoderate or high heritability, higher than 20%.
In prolific breeding animals, such as fish and shellfish,inbreeding is frequently encountered. Since theinbreeding depression is considerable for fitnesstraits as well as for growth rate, steps must be taken tokeep inbreeding low. A method to control it in abreeding program has been described for Cirrhinusmrigala (Bentsen et al. 1996b), Barbodes gonionotus(Bentsen et al. 1996a), Oreochromis niloticus (Gjedremet al. 1997b) and Chanos chanos (Gjedrem et al. 1997a).To keep inbreeding low, the key element is to use manyparents per generation. Bentsen et al. (1996a, 1996b)described how to produce families and to sample outcertain number of progenies from each family to reachabout the same number per family during the grow-out period.
From a simulation trial, it is concluded that: (i) inindividual selection one should use many families,more than 50 pairs. (ii) since facilities for grow-outtesting will be limited, only few animals per familyshould be allowed.(iii) this strategy will keepinbreeding low, avoid reduction of diversity and securelong-term response to selection.
A breeding program must continually be improvedand developed to utilize new knowledge as well as tomeet the requirements of the industry for moreproductive animals. If more traits are to be includedin the breeding goal in the future, family selection
must often be applied. The estimation of breedingvalues becomes more complicated as the number oftraits increases, making it necessary to optimizedifferent components in the breeding program.
Traits of economic importance will vary from speciesto species and among production systems. In mostinstances, growth rate and disease resistance will bethe important breeding goals. Age at maturation isimportant in species like salmonids and tilapia. Fleshquality will vary among species as well as over time,and is usually difficult to measure, particularly in liveanimals. Temperature and salinity tolerance may beimportant in tilapia to expand the areas for cultivation.
During the grow-out period, test animals should bereared under conditions similar to those used by theindustry. It must be defined how and when each traitshould be measured or recorded. Growth rate, bodyweight or body length, should be recorded atmarketing. This is easy to apply: age of sexualmaturation can be recorded as frequencies of matureanimals at a certain age, flesh quality can normallyonly be measured on carcasses. To obtain a measureof disease resistance, challenge tests against specificpathogens have proved to be efficient. Challenge testscould also be used to test for temperature tolerance,while salinity tolerance should be evaluated by growingtest animals in water with the wanted salinity whilerecording survival and growth rates.
Lessons from the GIFT Project
The Genetically Improved Farmed Tilapia (GIFT)Project of ICLARM was started in 1988, and the basepopulation was formed by a complete diallele crossbetween eight strains of O. niloticus. This basepopulation had broad genetic variation and possibleinbreeding in the domesticated strains was eliminated.In each generation, more than 100 families wereproduced while avoiding mating of close relatives.During testing period, efforts were made tostandardize the environmental conditions. In eachgeneration, strong selection was practised except inthe first. These procedures were key elements that ledto the great success of GIFT project.
The selection response in growth rate during fivegenerations averaged 13.2% per generation. This
Economic Traits for GeneticImprovement

131
genetic gain is similar to results obtained for severalfish species. These results show that it is possibleto double the growth rate in six generations in tilapia,since the genetic gain is cumulative over generations.When these results reach the industry, it will causea big change in production and productivity. Therewill be a dramatic reduction in the turnover rate andproduction per unit will increase. The feed conversionrate will become much lower. This increasein productivity will also increase the survival rate. Insummary, the production cost will markedlyreduce and total production will increase. Everyfarmer in the whole country, rich and poor, can obtainthis benefit.
The cooperation among all partners in the GIFTproject - ICLARM, AKVAFORSK, and the Philippineinstitutions, Bureau of Fisheries and Aquatic Resources(BFAR) and Central Luzon State University (CLSU) -has been very productive and educational.
Back in the 1970s, AKVAFORSK formed thetheoretical basis for a breeding program for Atlanticsalmon and rainbow trout through a series of researchprojects. It also actually started and developed theprogram in Norway. After some years, AKVAFORSKinvited the industry to participate in running theprogram, and in 1992, a breeding company, AquaGen, was established. From then on, this company hasbeen running national breeding programs for salmonand trout in Norway. AKVAFORSK participates infurther developing the breeding program by includingadditional economically important traits, optimizingthe different components of the program andcontinuously estimating breeding values.
In 1993, the partners in the GIFT project started abreeding program for O. niloticus in the Philippines.Later, this led to the establishment of the GIFTFoundation, which is now running a commercialbreeding program for O. niloticus in the Philippines.GIFT Foundation builds on the GIFT strain andcontinues the selection for growth rate and latematuration. It is located in the facilities built by thegovernment at Munoz, Nueva Ecija, and hasaccredited eight private hatcheries for production offry for the industry.
These are two good examples of how breeding projectshave developed into breeding programs run bycompanies to benefit the industry. The most difficultaspects in this development have been to: (i) establisha breeding organization; (ii) raise financialsupport for investments and to start the breedingprogram; and (iii) raise financial support to scaleup production and market the improved product.
Other international breeding projects in whichAKVAFORSK has been involved with are listedin Table 1.
In cooperation with INGA, AKVAFORSK is involvedin designing breeding plans for tilapia, mrigal carp(C. mrigala) and silver barb (B. gonionotus) in Vietnam,and for Nile tilapia, common carp and milkfish(C. chanos) in Indonesia. The collaboration with INGAcontinues, and AKVAFORSK plans to expand this.
In 1995, ICLARM and INGA organized the AdvancedCourse in Quantitative Genetics and Breeding forscientists from INGA member countries. Held inCavite, Philippines, some of the lecturers were fromAKVAFORSK. In 1998, INGA held a similar coursein India, with lecturers from AKVAFORSK.
AKVAFORSK also plans to offer short courses in theseareas. It supervises Ph.D. students in aquaculture inthe fields of nutrition, breeding and selection, fleshquality and disease prevention. However, it cannotoffer fellowships or financial support.
Table 1. International breeding projects with whichAKVAFORSK has involvement.
Country Species Period
Completed projectsIceland Atlantic salmon 1987-1993Philippines Nile Tilapia 1988-1997Hawaii Shrimpa 1994-1997Brazil Shrimpa 1997-1998
Ongoing projectsIndia Rohu carp 1993-
Chile Coho, rainbow trout, Atlantic salmon 1996-
Columbia Shrimpa 1997-Vietnam Nile Tilapia 1999-Scotland Atlantic salmon 1999-Italy Rainbow trout 1999-
aPenaeus vannamei.
Activities at AKVAFORSK of Importanceto the International Network on Genetics in Aquaculture
Developing Breeding Programsfor the Industry

Fish Genetics Research in Member Countries and Institutions of INGA132
References
Bentsen, H.B., T. Gjedrem and N. Van Hao. 1996a. Breeding planfor silver barb (Barbodes gonionotus) in Vietnam: individual(mass) selection to improve growth rate. Rep. No. 3.International Network on Genetics in Aquaculture, ICLARM,Manila, Philippines.
Bentsen, H.B., T. Gjedrem, T. Mai Thien and N. Cong Dan. 1996b.Breeding plans for mrigal (Cirrhinus mrigala) in Vietnam:individual (mass) selection to improve growth rate. Rep. No.2. International Network on Genetics in Aquaculture,ICLARM, Manila, Philippines. (Unpublished).
Gjedrem, T., H.M. Gjoen, A. Hardjamulia, Sudarto, A. Widiaty, R.Gustiano, A.H. Kristanto, L. Emmawati and W. Hadie. 1997a.Breeding plans for milkfish (Chanos chanos) in Indonesia:individual selection to improve growth rate. Rep. No. 5.International Network on Genetics in Aquaculture, ICLARM,Manila, Philippines.
Gjedrem, T., H.M. Gjoen, A. Hardjamulia, Sudarto, A. Widiaty, R.Gustiano, A.H. Kristanto, L. Emmawati and W. Hadie. 1997b.Breeding plans for Nile tilapia (Oreochromis niloticus) inIndonesia: individual selection. Rep. No. 4. InternationalNetwork on Genetics in Aquaculture, ICLARM, Manila,Philippines.

133
ABSTRACT
This paper documents the focus of applied genetics research on aquatic species at Queensland University ofTechnology (QUT), Brisbane, Australia. Studies have focused on exotic tilapias and native freshwater crayfish,with emphasis on development of genetic markers and their application to breeding programs designed to improvecultured stocks. Since many cultured stocks have been exposed to significant bottlenecks in the past, studies havealso focused on comparisons of levels and patterns of genetic diversity in wild stocks, as they represent aconsiderable genetic resource which can be used to provide novel genetic traits in culture. These studies undertakenin parallel with those of collaborating institutions using animal breeding approaches (selective breeding) haveproduced better culture strains of tilapia in Fiji and redclaw in Australia.
OVERVIEW OF FISHGENETICS RESEARCH AT
QUEENSLAND UNIVERSITYOF TECHNOLOGY
P.B. MatherSchool of Natural Resources SciencesQueensland University of Technology
2 George St. Brisbane, Australia, 4001
MATHER, P.B. 2001. Overview of fish genetics research at Queensland University of Technology, p. 133-139. In M.V. Guptaand B.O. Acosta (eds.) Fish genetics research in member countries and institutions of the International Network onGenetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
Genetic projects at QUT on species of interest tothe aquaculture industry have focused largely ontwo groups of organism, tilapias and Australiannative freshwater crayfish.
Tilapia Genetics
Tilapias are considered noxious species in Australiaand therefore cannot be cultured for legal reasons.Genetic research on tilapias at QUT has focused,therefore, on the development and application ofappropriate genetic markers for tilapia breedingprograms conducted outside Australia in developingnations. QUT’s major research project on tilapias
Overview of Fish Genetics Research at Queensland University of Technology
is based in Fiji in collaboration with the FisheriesDivision of the Ministry of Agriculture, Fisheries andForests (MAFF). This project commenced in 1993and was directed at assisting the Fijians to developbetter culture stocks for their tilapia farmers. Initialresearch was on evaluating the culture performanceof the four indigenous ti lapia stocks in Fiji(Oreochromis niloticus Chitralada strain, O. niloticusIsrael strain, O. mossambicus and a red hybrid) undertwo production systems (integrated andnonintegrated), and identifying the stock whichperformed best there under most environmentalconditions. This work was completed in 1996 andbetter stock management practices developed andput into practice to maintain stock quality(Macaranas et al. 1997).

Fish Genetics Research in Member Countries and Institutions of INGA134
In parallel, a range of molecular markers (allozyme,randomly amplified polymorphic DNA [RAPD], mtDNA and microsatellite loci) were developed at QUTto characterize tilapia stocks and to monitor theirgenetic integrity over time. The second phase of theproject has the objectives of first comparing therelative performances in culture of the best-performing stock in Fiji (O. niloticus Chitralda -determined in the earlier work) against thegenetically improved GIFT O. niloticus. Once theevaluation is complete, QUT intends to take the best-performing stock through three generations of familyselection to further improve growth performance.At the end of the program, it is hoped that these willbe achieved: a better performing stock for Fijianfarmers; a set of markers which will allow monitoringof stock quality over time; and highly trained staffin Fiji who can maintain fry quality. Stock evaluationtrials are conducted under near quarantineconditions so that only better-performing stocks areretained for fry production. Other stocks will beculled at a later date reducing their potential forhybridizing with high-performing lines. Since mostfry used by farmers in Fiji are provided by theFisheries Division hatcheries to farmers at low cost,this should allow strict control of fry quality.
Development of genetic markers
Much of the tilapia genetic work at QUT has focusedon development and application of genetic markers.Initially, allozyme markers were developed using thecellulose acetate system. Currently, there is an excessof 50 allozyme coding loci which are used to routinelyscreen O. niloticus and O. mossambicus stocks (Table1). As other studies have reported, however, allozymevariation is generally quite low in tilapiine specieswith percent polymorphic loci estimates commonlybelow 5% and average heterozygosity per locusestimates often below 10%. The marker work wasexpanded, therefore, to include DNA sequences thatcommonly show higher levels of genetic diversity.Thus, the focus in recent years has been on assessingvariation in the mtDNA control region and at RAPDand microsatellite loci (Table 2). MtDNA controlregion sequence variation screened usingTemperature Gradient Gel Electrophoresis (TGGE)and variation at RAPD loci have been used todescribe phylogenetic and systematic relationshipsamong cultured stocks of O. niloticus and wild andferal stocks of O. mossambicus. Patterns of variation inthe mtDNA control region sequence in wildO. mossambicus stocks in southern Africa indicate that
disjunct wild stocks have experienced long periodsof isolation which has led to separate evolution ofstocks in different parts of the species’ natural range.The extent of differentiation observed could indicatethe presence of discrete evolutionary lineages or evensubspecies (Table 3).
The publication by Kocher’s laboratory (Lee andKocher 1996) recently of >150 microsatellite primersequences for O. niloticus as a starting point fordeveloping a saturated gene map for the species hasallowed the expansion of the marker work todocument microsatellite variation in cultured andwild tilapia stocks. The conditions have beenoptimized for approximately 30 microsatellite lociin O. niloticus and screened variation in bothO. niloticus and O. mossambicus. Primers developedfor O. niloticus generally work equally well inO. mossambicus. After initial trials and characterizationof variation at specific microsatellite loci, nine lociwith high allelic diversity (>12 alleles per locus) wereselected to examine the relationship between averageheterozygosity and relative growth performance inthe O. niloticus Chitralada stock in Fiji, as heterosishas been linked to growth performance in someaquaculture stocks (e.g., Zouros et al. 1980).
Currently, (with collaborators in Fiji) the cultureperformance of GIFT and Chitralada strains ofO. niloticus is being compared under identical cultureconditions. In parallel, the two stocks for microsatelliteallelic diversity have been characterized, and theirrelative heterozygosity levels as a component of trialshave been directed at identifying the best performingO. niloticus stock for Fiji. After the best performingstock has been identified, the markers will be used tomonitor genetic diversity across three generations offamily selection. The same markers will be used overthe long term to monitor the genetic health of theculture stocks that are used to generate fry for Fijianfarmers. Poor management and husbandry practicescan often lead to changes in the genetic characteristicsof culture stocks that may compromise their long-termproductivity. Since genetic marker technologies havebeen applied to tilapia culture in Fiji, the quality offry provided by government hatcheries to farmers hasimproved noticeably. Despite the increased costs anddifficulties with shipping tissue samples for geneticanalysis among collaborating partners (Australia andFiji), the benefits in terms of stock quality have beenvery evident. This outcome demonstrates that it is notnecessary for all groups involved in geneticimprovement programs on aquatic organisms to
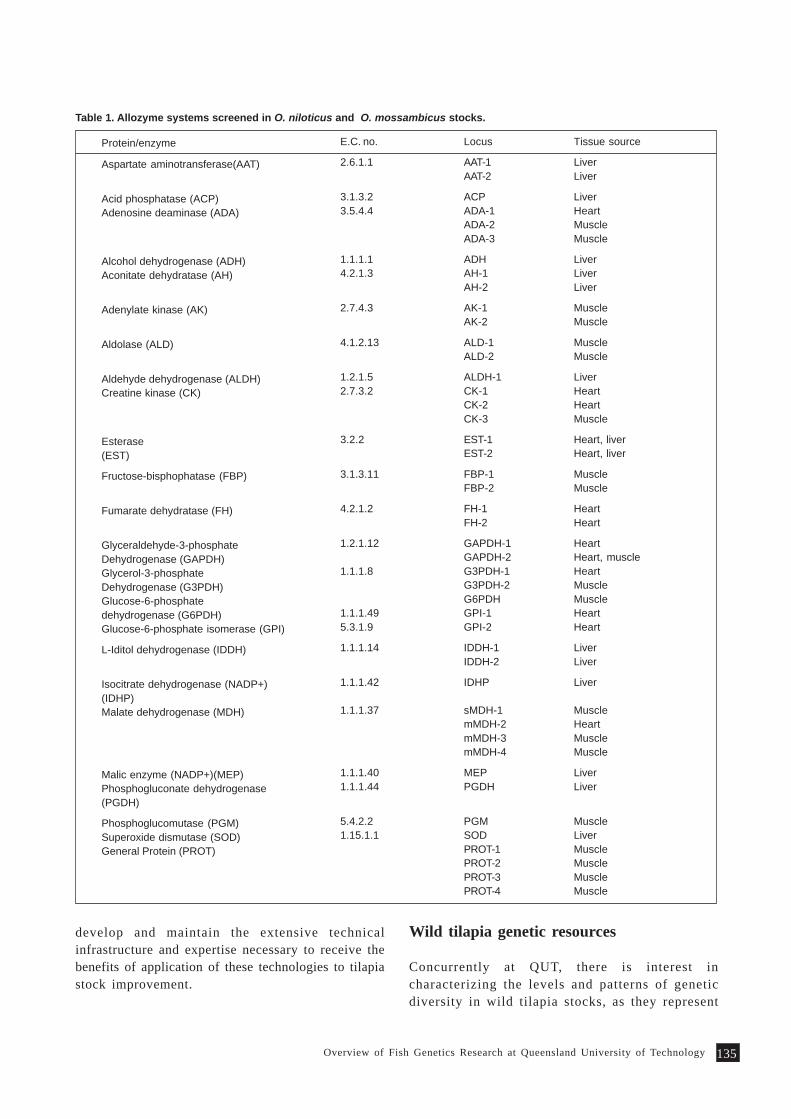
135
develop and maintain the extensive technicalinfrastructure and expertise necessary to receive thebenefits of application of these technologies to tilapiastock improvement.
Wild tilapia genetic resources
Concurrently at QUT, there is interest incharacterizing the levels and patterns of geneticdiversity in wild tilapia stocks, as they represent
Overview of Fish Genetics Research at Queensland University of Technology
Table 1. Allozyme systems screened in O. niloticus and O. mossambicus stocks.
Protein/enzyme
Aspartate aminotransferase(AAT)
Acid phosphatase (ACP)Adenosine deaminase (ADA)
Alcohol dehydrogenase (ADH)Aconitate dehydratase (AH)
Adenylate kinase (AK)
Aldolase (ALD)
Aldehyde dehydrogenase (ALDH)Creatine kinase (CK)
Esterase(EST)
Fructose-bisphophatase (FBP)
Fumarate dehydratase (FH)
Glyceraldehyde-3-phosphateDehydrogenase (GAPDH)Glycerol-3-phosphateDehydrogenase (G3PDH)Glucose-6-phosphatedehydrogenase (G6PDH)Glucose-6-phosphate isomerase (GPI)
L-Iditol dehydrogenase (IDDH)
Isocitrate dehydrogenase (NADP+)(IDHP)Malate dehydrogenase (MDH)
Malic enzyme (NADP+)(MEP)Phosphogluconate dehydrogenase(PGDH)
Phosphoglucomutase (PGM)Superoxide dismutase (SOD)General Protein (PROT)
E.C. no.
2.6.1.1
3.1.3.23.5.4.4
1.1.1.14.2.1.3
2.7.4.3
4.1.2.13
1.2.1.52.7.3.2
3.2.2
3.1.3.11
4.2.1.2
1.2.1.12
1.1.1.8
1.1.1.495.3.1.9
1.1.1.14
1.1.1.42
1.1.1.37
1.1.1.401.1.1.44
5.4.2.21.15.1.1
Locus
AAT-1AAT-2
ACPADA-1ADA-2ADA-3
ADHAH-1AH-2
AK-1AK-2
ALD-1ALD-2
ALDH-1CK-1CK-2CK-3
EST-1EST-2
FBP-1FBP-2
FH-1FH-2
GAPDH-1GAPDH-2G3PDH-1G3PDH-2G6PDHGPI-1GPI-2
IDDH-1IDDH-2
IDHP
sMDH-1mMDH-2mMDH-3mMDH-4
MEPPGDH
PGMSODPROT-1PROT-2PROT-3PROT-4
Tissue source
LiverLiver
LiverHeartMuscleMuscle
LiverLiverLiver
MuscleMuscle
MuscleMuscle
LiverHeartHeartMuscle
Heart, liverHeart, liver
MuscleMuscle
HeartHeart
HeartHeart, muscleHeartMuscleMuscleHeartHeart
LiverLiver
Liver
MuscleHeartMuscleMuscle
LiverLiver
MuscleLiverMuscleMuscleMuscleMuscle
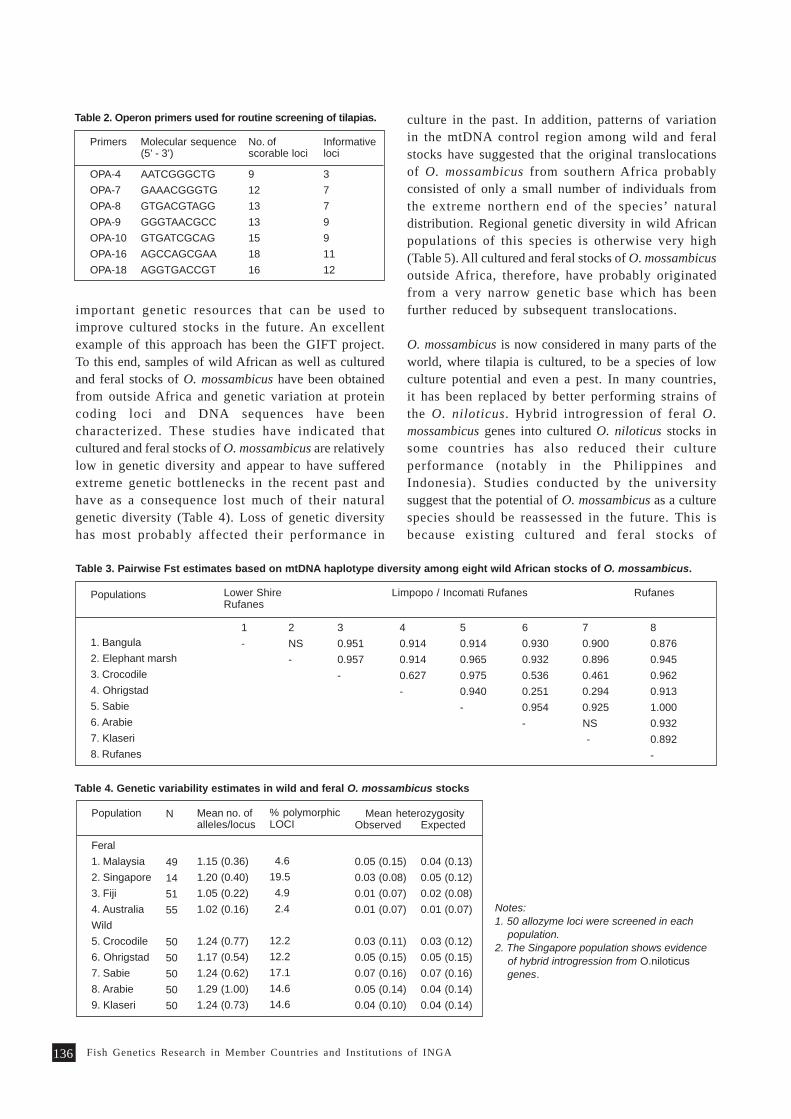
Fish Genetics Research in Member Countries and Institutions of INGA136
Table 4. Genetic variability estimates in wild and feral O. mossambicus stocks
Notes:1. 50 allozyme loci were screened in each population.2. The Singapore population shows evidence of hybrid introgression from O.niloticus genes.
Population
Feral1. Malaysia2. Singapore3. Fiji4. AustraliaWild5. Crocodile6. Ohrigstad7. Sabie8. Arabie9. Klaseri
% polymorphicLOCI
4.619.54.92.4
12.212.217.114.614.6
Mean heterozygosityObserved Expected
0.05 (0.15) 0.04 (0.13)0.03 (0.08) 0.05 (0.12)0.01 (0.07) 0.02 (0.08)0.01 (0.07) 0.01 (0.07)
0.03 (0.11) 0.03 (0.12)0.05 (0.15) 0.05 (0.15)0.07 (0.16) 0.07 (0.16)0.05 (0.14) 0.04 (0.14)0.04 (0.10) 0.04 (0.14)
N
49145155
5050505050
Mean no. ofalleles/locus
1.15 (0.36)1.20 (0.40)1.05 (0.22)1.02 (0.16)
1.24 (0.77)1.17 (0.54)1.24 (0.62)1.29 (1.00)1.24 (0.73)
important genetic resources that can be used toimprove cultured stocks in the future. An excellentexample of this approach has been the GIFT project.To this end, samples of wild African as well as culturedand feral stocks of O. mossambicus have been obtainedfrom outside Africa and genetic variation at proteincoding loci and DNA sequences have beencharacterized. These studies have indicated thatcultured and feral stocks of O. mossambicus are relativelylow in genetic diversity and appear to have sufferedextreme genetic bottlenecks in the recent past andhave as a consequence lost much of their naturalgenetic diversity (Table 4). Loss of genetic diversityhas most probably affected their performance in
culture in the past. In addition, patterns of variationin the mtDNA control region among wild and feralstocks have suggested that the original translocationsof O. mossambicus from southern Africa probablyconsisted of only a small number of individuals fromthe extreme northern end of the species’ naturaldistribution. Regional genetic diversity in wild Africanpopulations of this species is otherwise very high(Table 5). All cultured and feral stocks of O. mossambicusoutside Africa, therefore, have probably originatedfrom a very narrow genetic base which has beenfurther reduced by subsequent translocations.
O. mossambicus is now considered in many parts of theworld, where tilapia is cultured, to be a species of lowculture potential and even a pest. In many countries,it has been replaced by better performing strains ofthe O. niloticus. Hybrid introgression of feral O.mossambicus genes into cultured O. niloticus stocks insome countries has also reduced their cultureperformance (notably in the Philippines andIndonesia). Studies conducted by the universitysuggest that the potential of O. mossambicus as a culturespecies should be reassessed in the future. This isbecause existing cultured and feral stocks of
Table 2. Operon primers used for routine screening of tilapias.
Primers Molecular sequence No. of Informative(5’ - 3’) scorable loci loci
OPA-4 AATCGGGCTG 9 3OPA-7 GAAACGGGTG 12 7OPA-8 GTGACGTAGG 13 7OPA-9 GGGTAACGCC 13 9OPA-10 GTGATCGCAG 15 9OPA-16 AGCCAGCGAA 18 11OPA-18 AGGTGACCGT 16 12
Table 3. Pairwise Fst estimates based on mtDNA haplotype diversity among eight wild African stocks of O. mossambicus.
1 2 3 4 5 6 7 8- NS 0.951 0.914 0.914 0.930 0.900 0.876
- 0.957 0.914 0.965 0.932 0.896 0.945- 0.627 0.975 0.536 0.461 0.962
- 0.940 0.251 0.294 0.913- 0.954 0.925 1.000
- NS 0.932- 0.892
-
Populations
1. Bangula2. Elephant marsh3. Crocodile4. Ohrigstad5. Sabie6. Arabie7. Klaseri8. Rufanes
Lower Shire Limpopo / Incomati Rufanes RufanesRufanes
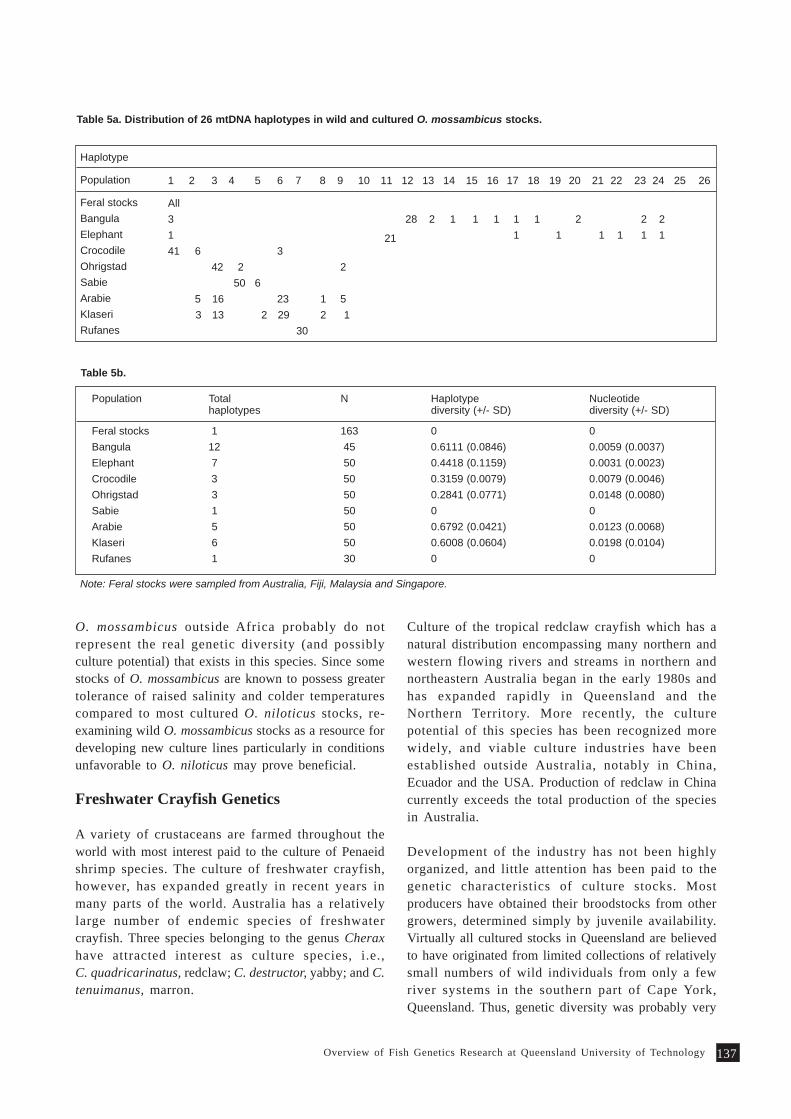
137
O. mossambicus outside Africa probably do notrepresent the real genetic diversity (and possiblyculture potential) that exists in this species. Since somestocks of O. mossambicus are known to possess greatertolerance of raised salinity and colder temperaturescompared to most cultured O. niloticus stocks, re-examining wild O. mossambicus stocks as a resource fordeveloping new culture lines particularly in conditionsunfavorable to O. niloticus may prove beneficial.
Freshwater Crayfish Genetics
A variety of crustaceans are farmed throughout theworld with most interest paid to the culture of Penaeidshrimp species. The culture of freshwater crayfish,however, has expanded greatly in recent years inmany parts of the world. Australia has a relativelylarge number of endemic species of freshwatercrayfish. Three species belonging to the genus Cheraxhave attracted interest as culture species, i.e.,C. quadricarinatus, redclaw; C. destructor, yabby; and C.tenuimanus, marron.
Culture of the tropical redclaw crayfish which has anatural distribution encompassing many northern andwestern flowing rivers and streams in northern andnortheastern Australia began in the early 1980s andhas expanded rapidly in Queensland and theNorthern Territory. More recently, the culturepotential of this species has been recognized morewidely, and viable culture industries have beenestablished outside Australia, notably in China,Ecuador and the USA. Production of redclaw in Chinacurrently exceeds the total production of the speciesin Australia.
Development of the industry has not been highlyorganized, and little attention has been paid to thegenetic characteristics of culture stocks. Mostproducers have obtained their broodstocks from othergrowers, determined simply by juvenile availability.Virtually all cultured stocks in Queensland are believedto have originated from limited collections of relativelysmall numbers of wild individuals from only a fewriver systems in the southern part of Cape York,Queensland. Thus, genetic diversity was probably very
Overview of Fish Genetics Research at Queensland University of Technology
Table 5a. Distribution of 26 mtDNA haplotypes in wild and cultured O. mossambicus stocks.
Haplotype
Population
Feral stocksBangulaElephantCrocodileOhrigstadSabieArabieKlaseriRufanes
1
All3141
2
6
53
3
42
1613
4
2 50
5
6
2
6
3
2329
7
30
8
12
9
2
51
10 11
21
12
28
13
2
14
1
15
1
16
1
17
11
18
1
19
1
20
2
21
1
22
1
23
21
24
21
25 26
Table 5b.
Population Total N Haplotype Nucleotidehaplotypes diversity (+/- SD) diversity (+/- SD)
Feral stocks 1 163 0 0Bangula 12 45 0.6111 (0.0846) 0.0059 (0.0037)Elephant 7 50 0.4418 (0.1159) 0.0031 (0.0023)Crocodile 3 50 0.3159 (0.0079) 0.0079 (0.0046)Ohrigstad 3 50 0.2841 (0.0771) 0.0148 (0.0080)Sabie 1 50 0 0Arabie 5 50 0.6792 (0.0421) 0.0123 (0.0068)Klaseri 6 50 0.6008 (0.0604) 0.0198 (0.0104)Rufanes 1 30 0 0
Note: Feral stocks were sampled from Australia, Fiji, Malaysia and Singapore.
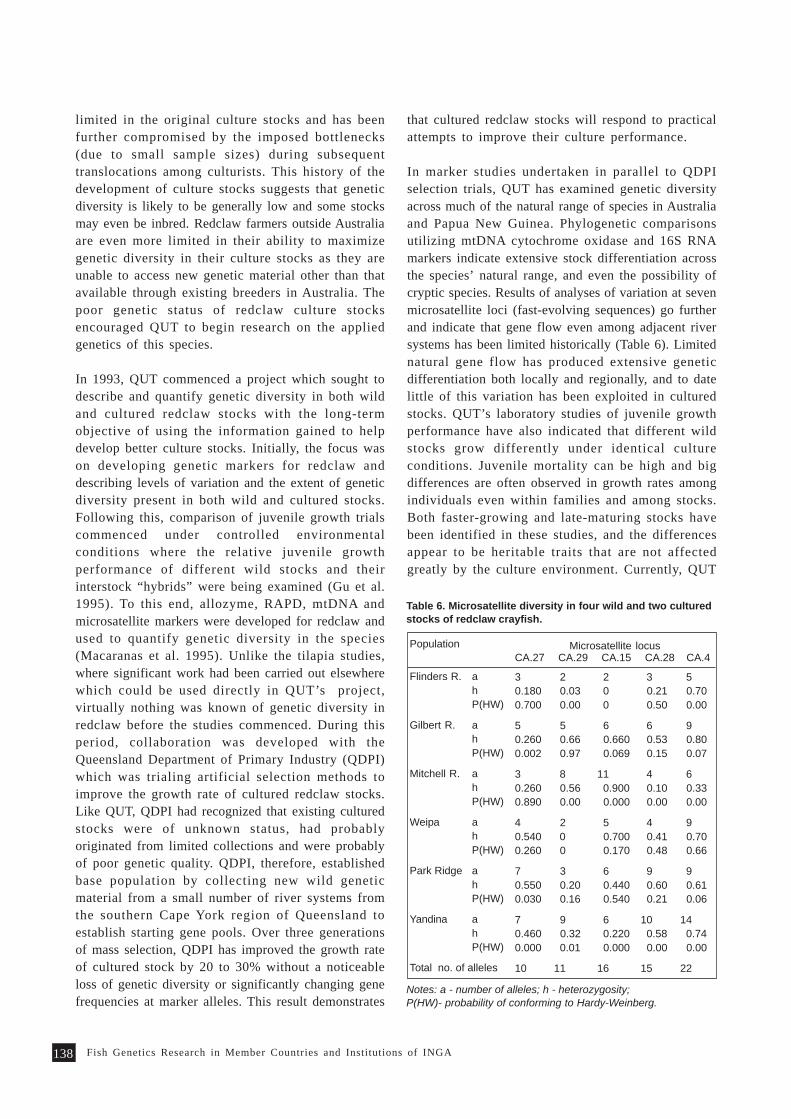
Fish Genetics Research in Member Countries and Institutions of INGA138
Table 6. Microsatellite diversity in four wild and two culturedstocks of redclaw crayfish.
Population
Flinders R. ahP(HW)
Gilbert R. ahP(HW)
Mitchell R. ahP(HW)
Weipa ahP(HW)
Park Ridge ahP(HW)
Yandina ahP(HW)
Total no. of alleles
Microsatellite locusCA.27 CA.29 CA.15 CA.28 CA.4
3 2 2 3 50.180 0.03 0 0.21 0.700.700 0.00 0 0.50 0.00
5 5 6 6 90.260 0.66 0.660 0.53 0.800.002 0.97 0.069 0.15 0.07
3 8 11 4 60.260 0.56 0.900 0.10 0.330.890 0.00 0.000 0.00 0.00
4 2 5 4 90.540 0 0.700 0.41 0.700.260 0 0.170 0.48 0.66
7 3 6 9 90.550 0.20 0.440 0.60 0.610.030 0.16 0.540 0.21 0.06
7 9 6 10 140.460 0.32 0.220 0.58 0.740.000 0.01 0.000 0.00 0.00
10 11 16 15 22
Notes: a - number of alleles; h - heterozygosity;P(HW)- probability of conforming to Hardy-Weinberg.
limited in the original culture stocks and has beenfurther compromised by the imposed bottlenecks(due to small sample sizes) during subsequenttranslocations among culturists. This history of thedevelopment of culture stocks suggests that geneticdiversity is likely to be generally low and some stocksmay even be inbred. Redclaw farmers outside Australiaare even more limited in their ability to maximizegenetic diversity in their culture stocks as they areunable to access new genetic material other than thatavailable through existing breeders in Australia. Thepoor genetic status of redclaw culture stocksencouraged QUT to begin research on the appliedgenetics of this species.
In 1993, QUT commenced a project which sought todescribe and quantify genetic diversity in both wildand cultured redclaw stocks with the long-termobjective of using the information gained to helpdevelop better culture stocks. Initially, the focus wason developing genetic markers for redclaw anddescribing levels of variation and the extent of geneticdiversity present in both wild and cultured stocks.Following this, comparison of juvenile growth trialscommenced under controlled environmentalconditions where the relative juvenile growthperformance of different wild stocks and theirinterstock “hybrids” were being examined (Gu et al.1995). To this end, allozyme, RAPD, mtDNA andmicrosatellite markers were developed for redclaw andused to quantify genetic diversity in the species(Macaranas et al. 1995). Unlike the tilapia studies,where significant work had been carried out elsewherewhich could be used directly in QUT’s project,virtually nothing was known of genetic diversity inredclaw before the studies commenced. During thisperiod, collaboration was developed with theQueensland Department of Primary Industry (QDPI)which was trialing artificial selection methods toimprove the growth rate of cultured redclaw stocks.Like QUT, QDPI had recognized that existing culturedstocks were of unknown status, had probablyoriginated from limited collections and were probablyof poor genetic quality. QDPI, therefore, establishedbase population by collecting new wild geneticmaterial from a small number of river systems fromthe southern Cape York region of Queensland toestablish starting gene pools. Over three generationsof mass selection, QDPI has improved the growth rateof cultured stock by 20 to 30% without a noticeableloss of genetic diversity or significantly changing genefrequencies at marker alleles. This result demonstrates
that cultured redclaw stocks will respond to practicalattempts to improve their culture performance.
In marker studies undertaken in parallel to QDPIselection trials, QUT has examined genetic diversityacross much of the natural range of species in Australiaand Papua New Guinea. Phylogenetic comparisonsutilizing mtDNA cytochrome oxidase and 16S RNAmarkers indicate extensive stock differentiation acrossthe species’ natural range, and even the possibility ofcryptic species. Results of analyses of variation at sevenmicrosatellite loci (fast-evolving sequences) go furtherand indicate that gene flow even among adjacent riversystems has been limited historically (Table 6). Limitednatural gene flow has produced extensive geneticdifferentiation both locally and regionally, and to datelittle of this variation has been exploited in culturedstocks. QUT’s laboratory studies of juvenile growthperformance have also indicated that different wildstocks grow differently under identical cultureconditions. Juvenile mortality can be high and bigdifferences are often observed in growth rates amongindividuals even within families and among stocks.Both faster-growing and late-maturing stocks havebeen identified in these studies, and the differencesappear to be heritable traits that are not affectedgreatly by the culture environment. Currently, QUT

139Overview of Fish Genetics Research at Queensland University of Technology
is comparing relative juvenile growth rates in threegenetically divergent stocks and their reciprocalinterstock hybrids. The three stocks were collectedfrom three geographically most disjunct wild streamswhere redclaw occur naturally in Australia. Thisexperiment is assessing the potential that hybrids mayshow for expressing hybrid vigor or even outbreedingdepression. A recent study conducted on the relatedspecies, C. destructor (yabby), which used a similarapproach, identified some crosses which yielded all-male progeny. This may prove to be a significantadvantage to the yabby culture industry because malefreshwater yabbies commonly grow faster and largerthan do females.
Future Directions
QUT’s collaborative projects with the Fiji FisheriesDivision on tilapia and with QDPI on redclaw geneticimprovement have approximately two years to run.Genetic marker studies on both species are welladvanced, and progress has been made towardsdeveloping faster-growing lines for both Fijian tilapiaand Australian redclaw farmers. This year, QUT iscommencing a project on strain/stock evaluations onfreshwater prawns (Macrobrachium australiense) whichwill utilize similar approaches to those outlined above.It is also currently exploring the possibility of applyingthe genetic markers developed for tilapia more widelyin countries where tilapia culture is practiced throughdeveloping collaboration with appropriate agencies.It is a costly and technologically demanding processto develop and utilize marker technology to assist stockimprovement so it is important to use it as widely aspossible once available. Existing markers andexpertise can be applied directly to cultured stocks ofO. niloticus, O. mossambicus and their interspecieshybrids basically anywhere genetic improvement orstock management practices are being trialed fortilapias. It is therefore important in the future toidentify appropriate collaborations betweenorganizations to maximize the technologies alreadydeveloped to improve tilapia (and crayfish) culture.This will reduce the costs of research programsdesigned to produce better stock, stop unnecessaryduplication and help promote aquaculture researchin developing nations.
References
Gu, H., P.B. Mather and M.F. Capra. 1995. Juvenile growthperformance among stocks and families of the redclaw crayfish(Cherax quadricarinatus von Martens). Aquaculture 134: 29-36.
Lee, W.-J. and T.D. Kocher. 1996. Microsatellite markers for geneticmapping in the tilapia, Oreochromis niloticus. J. Fish Biol. 49:169-71.
Macaranas, J.M., P.B. Mather, P. Hoeben and M.F. Capra. 1995.Assessment of genetic variation in wild populations of theredclaw crayfish (Cherax quadricarinatus, von Martens 1868)by means of allozyme and RAPD-PCR markers. Mar. Freshwat.Res. 46: 1217-1228.
Macaranas, J.M., P.B. Mather, S.N. Lal, T. Vereivalu, M. Lagibalavuand M.F. Capra. 1997. Genotype and environment: acomparative evaluation of four tilapia stocks in Fiji.Aquaculture 150: 11-24.
Zouros, E., S.M. Singh and H.E. Miles. 1980. Growth rate inoysters: an overdominant phenotype and its possibleexplanations. Evolution 34: 856-867.

Fish Genetics Research in Member Countries and Institutions of INGA140

141
ABSTRACT
Southeast Asia is endowed with a variety of species that can be cultured as food fish. Since 1973, the Southeast
Asian Fisheries Development Center/Aquaculture Department (SEAFDEC/AQD) has devoted research efforts
to establish broodstock for the tiger prawn (Penaeus monodon), milkfish (Chanos chanos), and other
economically important fish and crustaceans in the region. The objective is to ensure the availability and
reliability of seed supply for fish culture.
SEAFDEC’s pioneering effort on genetics was a collaborative study with the University of the Philippines
Marine Science Institute on the genetic variation of milkfish populations in the Philippines. Towards the end of
the 1980s, genetic activities, co-funded by the International Development Research Centre (IDRC) of Canada
in collaboration with Dalhousie University of Canada, were focused on tilapia.
GENETICS RESEARCH AT THESOUTHEAST ASIAN FISHERIES
DEVELOPMENT CENTER/AQUACULTURE DEPARTMENT
Z.U. BasiaoSoutheast Asian Fisheries Development Center
Aquaculture Department, Binangonan Freshwater StationBinangonan, Rizal, Philippines
BASIAO, Z.U. 2001. Genetics research at the Southeast Asian Fisheries Development Center/Aquaculture Department, p.141-144. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of theInternational Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
Compared to agriculture and livestock, aquacultureis sti l l behind in the management of geneticresources. Majority of the genetic resources ofaquatic animals is still found in wild populations asvery little domestication has taken place. Exceptionsinclude strains and varieties of the common carp,rainbow trout and, during the last decade, tilapia.
Genetics Research at the Southeast Asian FisheriesDevelopment Center/Aquaculture Department
Genetic variation in milkfish populations
Chanos chanos is one of the most important andexploited marine resources in the Philippines.Twelve populations analyzed for eletrophoreticvariation showed significant variations (Macaranaset al. 1990). There are at least two subpopulationsin the Philippine waters.

Fish Genetics Research in Member Countries and Institutions of INGA142
Development of salinity-tolerant strain ofNile tilapia hybrid
The objective of the project implemented at theSEAFDEC/AQD Tigbauan Station was to produce asalinity-tolerant strain of Oreochromis niloticus that canspawn and grow in regions with inadequate suppliesof freshwater. The procedure involved hybridizationof O. niloticus with O. mossambicus followed by repeatedbackcrossing of the hybrid with O. niloticus.
The mean survival time (MST) and median survivaltime (ST50), of freshwater-spawned and reared O.mossambicus and O. mossambicus and O. niloticus hybridsshowed higher salinity tolerance than O. niloticus(Villlegas 1990a). Increased salinity tolerance with ageor body size was also evident in the study. The highestgrowth for O. mossambicus was at 15 and 32 ppt whilethe optimum salinity range for growth of O. niloticuswas 0-10 ppt (Villegas 1990 b).
Experimental and statistical straincomparison procedures
One major objective of any fish breeding programis to develop genetically improved strains. Sincegenetic improvement programs are expensive,it is essential that proper strain testing methodologiesthat can detect an economically important diferencebetween two strains be developed. In aquaculturedecisionmaking, the magnitude of difference that theresearcher wants to detect can be decided a priori onpurely economic grounds. Economic theory or resultsof previous strain-comparison studies can help indicatethe magnitude of differences that would be importantbetween the growth rates of fish. Comparisonexperiments at SEAFDEC/AQD involve rigorous sizegrading (collimation), use of full-sib groups andinclusion of internal reference fish in each replicatetank/cage for statistical control.
The usual strain comparison procedure involvesanalysis of variance (ANOVA). The internal referenceprocedure uses analysis of covariance (ANCOVA) wheregrowth of the internal reference fish (red tilapia) is theconcomitant variable to reduce error variance causedby environmental variation like population density andfood supply (Basiao and Doyle 1990a).
Strain comparison in diverse environments
The efficiency of using an internal referencepopulation in detecting strain differences has beendemonstrated in high-input, well-managed environ-
ments as well as low-input marginal or artisanalconditions. Tilapia strains showed significant genotypeeffects during a three-week temporary crowding period(Basiao and Doyle 1990a, 1990b) and at the end ofthe final grow-out period in hapa cages (Basiao et al.1996). Inclusion of an internal reference strainincreased the sensitivity of the statistical test to detecta 7% significant difference among strains.Approximately 450 replicate families would have beenneeded to detect the significant strain differences ifthe reference strain had not been included in theexperimental design.
Genetic variation in growth was found in O. niloticusstrains held in freshwater for 2 weeks, acclimated for4 days in a salinity of 32 ppt and reared in 32 pptseawater for 2 weeks and then reared again infreshwater for another 2 weeks (Basiao, unpubl.).
Growth and survival of five Asian red tilapia strainswere compared in brackish and seawater. Statisticalanalyses on mean specific growth rate showed that aPhilippine strain grew best in seawater while a Thailandstrain performed well in brackishwater (Romana-Eguiaand Eguia 1999).
Genotype x environment interaction
Genotype x environment interactions (GXE) or thedifferential response of specific genotypes underdifferent environmental conditions (Falconer 1989) hasimportant implications for selection, strain testing andaquaculture in general. Romana-Eguia and Doyle(1992) using an internal reference fish foundindications of GXE interactions among threeO. niloticus strains reared in low (rice bran) or high(commercial diet) quality feeds. These strains alsodiffer in their growth response under restricted ornonrestricted feeding regime (Romana-Eguia andEguia 1993). However, ranking of the strains is similarin the two feeding regimes.
Resistance to heavy metal stress
Small fingerlings of two O. niloticus strains showedsignificant strain differences in terms of heavy metaltolerance when exposed to mixtures of zinc, cadmiumand mercury (Cuvin-Aralar 1993). This difference wasdiminished, however, in older and bigger fish. Themore resistant members of the population (survivors),which have the ability to adapt to the heavy metaltoxicants were able to pass on the resistance to theiroffspring (Cuvin-Aralar and Aralar 1994).

143
Selective Breeding and BroodstockManagement of Tilapias
Collimated mass selection
An important objective of aquaculture genetics is toprovide improved fish that will benefit the privatesector and also the small fish farmers. However,genetics research is an expensive endeavor for mostdeveloping countries and most results do not benefitmarginalized fish farmers. The conventional massselection on tilapia, which has not shown positiveresults in early studies, has been modified by thecollimation procedure or the early culling of large frybefore a two-step directional selection is applied(Basiao and Doyle 1999). The collimation procedurereduces the self-amplifying phenotypic variance ingrowth and increases the genetic to phenotypicvariance at the time of selection. One generation ofcollimated mass selection resulted in a significantpositive response of 3% relative to the control. Therealized heritability (h2) of approximately 16% iscomparable with other recent estimates of h2 inaquaculture populations of tilapia.
Farm-based broodstock improvement
The collimated mass selection procedure developedat SEAFDEC/AQD was pilot- tested in a small hatcheryfarm in Laguna, Philippines. The objective of thisfarmer participatory research is to help farmersdevelop their own tilapia broodstock. The farm trialshowed a 7-9% response to selection after onegeneration of selection (SEAFDEC/AQD 1999). Animportant implication of a farmer-based broodstockselection is that fish farmers would have more controlover their choice of good quality spawners.Dependence on a franchise-dealer type of seedproduction will be minimized and socioeconomicallyself-sustaining genetic conservation will be achieved.
Development of high-yield red tilapia strain
The genetic improvement of a Philippine red tilapiastrain through introgressive hybridization with theChitralada strain of O. niloticus from Thailand andsubsequent application of size-specific mass selectionis in progress. One generation of introgression resultedin a significant positive response of 2.5% as measuredby the percent differences in the lengths of introgressedred-orange offspring from the normal red tilapia(SEAFDEC/AQD 1999). As an accompanying study, a2 x 2 factorial experiment to determine the
reproductive efficiency of the introgressed red tilapiaand the normal red tilapia fed with two formulateddiets was done. The frequency of spawning and seed(fry and egg) production were highest in the normalred breeders fed a commercially formulated diet. Theintrogressed red and normal red tilapia fed theSEAFDEC-formulated feed showed comparablenumber of spawning. However, fry production washigher in the normal breeders, while egg productionwas higher in the introgressed red tilapia. The lowestspawning frequency and seed production wereobserved in the introgressed red breeders given thecommercially formulated feed.
The reproductive efficiency of three genetically diversePhilippine red tilapia strains (BFS, FAC and PF) andone Thai red tilapia strain (NIFI) in two seedproduction systems were compared with the objectiveof improving red tilapia fry production (Romana-Eguiaand Eguia 1999). Seed production in all strains wasconsiderably higher in tanks than in cages. The meandaily seed production in tanks was highest for FAC,followed by NIFI, BFS and PF. In fine-meshed netcages, mean daily seed produced per female washighest for BFS, followed by NIFI, FAC and PF.
Genetic characterization of hatchery-bredNile tilapia
Genetic variation is the basic resource of any successfulartificial selective breeding program. Hence, the levelof genetic variation should be monitored in hatcherystocks for their efficient management. This concern isaddressed by SEAFDEC/AQD by monitoring thegenetic variation of the hatchery-bred tilapiamaintained at the Binangonan Freshwater Stationusing protein and enzyme analysis. Sixteen enzymesand 24 loci were screened in three generations ofhatchery-bred O. niloticus. The six polymorphic locidetected in the base population were also found in theF1 and F2 generations (SEAFDEC/AQD 1999).
Genetic Diversity of Wild and CulturedBlack Tiger Shrimp (Penaeus monodon) inthe Philippines
The genetic variability of wild P. monodon collected fromfour geographical regions (Capiz, Negros Occidental,Palawan and Quezon) in the Philippines was comparedwith cultured samples collected from Antique andNegros Occidental, Philippines. Fifteen out of the 18commercially available primers were selected and usedto produce randomly amplified polymorphic DNAs.
Genetics Research at the Southeast Asian FisheriesDevelopment Center/Aquaculture Department

Fish Genetics Research in Member Countries and Institutions of INGA144
Initial results showed that the wild samples are morepolymorphic (66-71%) than the cultured samples(54%). Wild samples from Palawan showed the highestpolymorphism (71%).
Six microsatellites were used to establish possiblecorrelation between genetic diversity among wild P.monodon populations and status of mangroves andshrimp culture systems in the Philippines. Two culturedpopulations were examined as controls. Resultsindicated that all six microsatellite loci werepolymorphic and observed heterozygosity ranged from46.6% to 100%. Genetic differences betweenpopulations, based on genotypic and allelicfrequencies, exhibited some degree of association withthe status of the mangroves and culture system nearthe site where the samples were collected. The NegrosOccidental population, with the most severe mangroveloss and the most intensive culture systems, was themost significantly differentiated population. It alsoshowed the lowest average number of alleles per locus,suggesting a decreased diversity in the population. Thesecond most differentiated population was thepopulation from Capiz, a province with wide areaof extensive culture ponds and few remainingsecondary mangroves. The Quezon and the Palawanpopulations were not genetically different from eachother. The two cultured populations showed lessgenetic diversity and were significantly different fromthe four wild populations. The potential risk of geneticcontamination by the accidental or intentional releaseof cultured populations of shrimp into the wild shouldbe addressed in the light of these results.
Future Activities
Refinements of on-station and farmer-based researchon selective breeding of tilapia will be pursued. Geneticimprovement of the bighead carp (Aristichthys nobilis)introduced in the Philippines will be initiated. Geneticcharacterization of hatchery-bred fish and crustaceansand the population genetic structure of indigenousfreshwater fishes will be done. These futureresearch thrusts of SEAFDEC/AQD will be done incollaboration with other research institutions anduniversities in the Philippines.
References
Basiao, Z.U. and R.W. Doyle. 1990a. Interaction between test andreference populations when tilapia strains are compared bythe “internal control” technique. Aquaculture 85: 207-214.
Basiao, Z.U. and R.W. Doyle. 1990b. Use of internal referencepopulations for growth rate comparisons of tilapia strains. I.In a crowded environment, p. 503-504. In R. Hirano and I.Hanyu (eds.) The Second Asian Fisheries Forum. AsianFisheries Society, Manila, Philippines. 991 p.
Basiao, Z.U. and R.W. Doyle. 1999. Test of size-specific massselection for tilapia cage farming in the Philippines. Aquacult.Res. 30: 373-378.
Basiao, Z.U., R.W. Doyle and A.L. Arago. 1996. A statistical poweranalysis of the ‘internal reference’ technique for comparinggrowth and depensation of tilapia strains. J. Fish Biol. 49:277-286.
Cuvin-Aralar, M.L.A.1993. Survival and heavy metal accumulationof two Oreochromis niloticus (L.) strains exposed to mixturesof zinc, cadmium and mercury. Sci. Total Environ. 148: 31-38.
Cuvin-Aralar, M.L.A. and E.V. Aralar. 1994. Resistance to heavymetal mixture in Oreochromis niloticus progenies from parentschronically exposed to the same metals. Chemosphere 30(5):953-963.
Falconer, D.S. 1989. Introduction to quantitative genetics. Thirded. Longman, London, UK. 438 p.
Macaranas, J., J.R. Pante and L.V. Benitez, 1990. Heterogeneityin Philippine milkfish populations, p. 477-481. In R. Hiranoand I. Hanyu (eds.) The Second Asian Fisheries Forum. AsianFisheries Society, Manila, Philippines. 991 p.
Romana-Eguia, M.R.R. and R.V. Eguia. 1993. Growth response ofthree Oreochromis niloticus strains to feed restriction. Isr. J.Aquacult.-Bamidgeh 45: 8-17.
Romana-Eguia, M.R.R. and R.V. Eguia. 1999. Growth of five Asianred tilapia strains in saline environments. Aquaculture 173:161-170.
Romana-Eguia, M.R.R. and R.W. Doyle. 1992. Genotype-environment interaction in the response of three strains ofNile tilapia to poor nutrition. Aquaculture 108: 1-12.
SEAFDEC/AQD. 1999. Annual report. Aquaculture Department,Southeast Asian Fisheries Development Center, Tigbauan,Iloilo, Philippines.
Villegas, C.T. 1990a. Evaluation of the salinity tolerance ofOreochromis mossambicus, Oreochromis niloticus and their F1hybrids. Aquaculture 85: 281-292.
Villegas, C.T. 1990b. Growth and survival of Oreochromis niloticus,O. mossambicus and their F1 hybrids in various salinities, p.507-510. In R. Hirano and I. Hanyu (eds.) The Second AsianFisheries Forum. Asian Fisheries Society, Manila, Philippines.991 p.

145
ABSTRACT
The Genetics and Reproduction research group is one of the largest within the Institute of Aquaculture (IOA) at theUniversity of Stirling. The institute works with a broad range of species from diverse geographical areas and habitatsand has benefited from collaborations with other institutions in many countries. This overview of activities focusesparticularly on areas of relevance to the International Network on Genetics in Aquaculture (INGA) and collaborations
with INGA members.
OVERVIEW OF AQUACULTUREGENETICS RESEARCH
IN THE INSTITUTE OF AQUACULTURE,UNIVERSITY OF STIRLING
D.J. PenmanInstitute of Aquaculture, University of Stirling
Stirling FK9 4LA, Scotland
PENMAN, D.J. 2001. Overview of aquaculture genetics research in the Institute of Aquaculture, University of Stirling, p.145-149. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries and institutions of theInternational Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Aquaculture Genetics Research
Molecular taxonomy, population andconservation genetics
Allozyme, mtDNA, ribosomal DNA, minisatellite andmicrosatellite DNA variation have been used inInstitute of Aquaculture (IOA) laboratories in studieson the taxonomy and population genetics of wildorganisms of interest to aquaculture, and captiveaquaculture stocks. Examples of these studies aremolecular taxonomy and population genetics oftilapias (Sodsuk et al. 1995; Rognon et al. 1996);population genetics of Penaeus monodon (Klinbunga etal. 1996, 1998a, 1998b, 1999), Macrobrachiumnipponense (Wong and McAndrew 1994), Dicentrarchuslabrax (Castilho and McAndrew 1998; MartinezRodriguez et al. 1998), Salmo salar (Taggart et al.1995a, 1995b; Prodoehl et al. 1997; Stone et al. 1997)and Catla catla.
Overview of Aquaculture Genetics Researchin the Institute of Aquaculture, University of Stirling
IOA’s interest in conservation genetics has focused onthe use of cryopreservation and on the potential ofandrogenesis in gene banking, with tilapia andsalmonids as the main experimental subjects(McAndrew et al. 1993; Myers et al. 1995a, 1995b;Rana 1995; Penman et al. 1997). The combination ofsperm cryopreservation and diploid androgenesisdoes allow recovery of viable animals from a spermgene bank, but successful cryopreservation of eggs isstill not possible.
Broodstock management and quantitativegenetics
IOA has been involved in collaboration with theUniversity of Agricultural Sciences (UAS), Bangalore,India, and the University of Wales Swansea onbroodstock management and selective breeding ofC. catla and Cyprinus carpio in Karnataka, India(Basavaraju et al. 1998). In UK, IOA is involved in

Fish Genetics Research in Member Countries and Institutions of INGA146
developing broodstock management and selectivebreeding programs on molecular markers (principallymicrosatellites), in Atlantic salmon and other cold-water species.
Sex determination and differentiation
Research on sex determination mechanisms andtheir manipulation to produce, e.g., monosex stocks,has been a major interest for IOA for some time.Tilapia, principally Oreochromis niloticus, has beenthe major model, due to its commercial importance,ease of maintenance under laboratory conditions,amenability to genetic and hormonal manipulations,and the emergence of temperature sex determination(TSD) and autosomal genetic factors affecting sexdetermination in addition to the major (chromosomal)sex determination.
Recently, the chromosomal location of the sexdetermining region in O. niloticus has beendemonstrated (Carrasco et al. 1999a). Fully inbredclonal XX and YY lines in this species have beenproduced (Sarder et al. in press). Studies now focus onmolecular and cytogenetic analysis of the sexdetermination system and on the role of aromatase insex differentiation and TSD.
Collaborative research with the National AquacultureGenetics Research Institute, (NAGRI) Thailand andthe Bangladesh Fisheries Research Institute has led tothe development of genetically monosex femaleproduction in the silver barb (Barbodes gonionotus) foraquaculture (Pongthana et al. 1995a, 1999).
Genetic manipulations
Gynogenesis (Hussain et al. 1993, 1995; Myers et al.1995; Pongthana et al. 1995b) and androgenesis(Myers et al. 1995a, 1995b) have proven to be usefultools in the study of sex determination and thedevelopment of monosex culture, the development ofclonal lines (Hussain et al. 1998; Sarder et al. in press)and gene mapping, etc.
These tilapia clonal lines have already proved to beextremely useful in studies on nonspecific and specificimmune responses, and the institute is planning touse them in research on areas as diverse as nutritionand quantitative genetics.
Studies have also been carried out on induced triploidy
in tilapia (Hussain et al. 1995, 1996), rainbow trout(Carrasco et al. 1999a, 1999b) and halibut in the lastfew years. Although triploidy could not currently beused commercially in tilapia due to the difficulties ofcollecting adequate numbers of unfertilized eggs,studies elsewhere have shown its potential in limitingreproduction in pond culture. Triploidy is, however,used commercially in rainbow trout. A recent Ph.D.study at IOA (Carrasco et al. 1998) revealed differencesin meiotic chromosome pairing between genetic maleand female triploids which may account for thedifferences in the success of maturation of male andfemale triploids (male triploids are generally capableof producing some aneuploid sperm whereas femaletriploids are generally sterile).
IOA carried out a policy study on genetically modifiedfish for the UK Department of the Environment(Woodwark et al. 1994; Penman et al. 1995), whichled to drafting guidelines for the UK government.
Genomics
Gynogenesis has proven to be a useful tool in genemapping, in addition to the more conventional crosses.IOA initially studied gene-centromere recombinationdistances for allozymes, color loci, etc (Hussain et al.1994). More recently, highly polymorphic DNAmarkers, such as microsatellites and AFLPs or RFLPshave been developed and, in collaboration with Dr.Tom Kocher at the University of New Hampshire, alinkage map has been developed using haploidgynogenetic embryos (Kocher et al. 1998) and gene-centromere linkages are being estimated for a widervariety of loci (Markert et al. 1999). IOA has a particularinterest in mapping sex determining loci in tilapiasand is pursuing a variety of strategies for this.
The genomics capabilities at IOA have beenstrengthened in the last two years by a new initiativeknown as “Aquagene”. Prof. Alan Teale and his groupcome with a background of mammalian genetics, witha past focus on mapping disease resistance genesin cattle and mice, while Dr. John Taggart, Dr. MargaretCairney and Dr. Mike Leaver are involved in mappingsalmonid genomes and identifying ESTs in thesespecies.
IOA is currently developing strategies for the detailedanalysis of the genetic basis of a number of traits relatedto disease resistance, growth performance, nutritionand control of reproduction.

147Overview of Aquaculture Genetics Researchin the Institute of Aquaculture, University of Stirling
Aquaculture Development and CoordinationProgram
The Aquaculture Development and CoordinationProgram (AADCP) was implemented from 1990 to1994, with funding from the European Union (EU)and the Association of Southeast Asian Nations(ASEAN). Five components in different ASEANcountries were twinned with partner institutions fromEU countries (Penman 1995). In component 5 ofAADCP (aquaculture genetics), NAGRI in Thailandwas twinned with IOA. The two main areas of researchwere genetic manipulations and population genetics.
DFID Fish Genetics Research Programme
The IOA genetics group has been involved in severalcollaborative projects with INGA member countriesthrough projects funded by the Department ofInternational Development (DFID, formerly OverseasDevelopment Agency) Fish Genetics ResearchProgramme (see Mair and Beardmore, this vol., formore details). The main projects under this programare on population genetics of Penaeus sp. (withUniversity of Putra, Malaysia); monosex culture of B.gonionotus (with NAGRI, Thailand, and the BangladeshFisheries Research Institute); and aquaculture geneticsof Indian and common carps (with UAS, Bangalore,India, and University of Wales, Swansea, UK).
Ph.D. studentships, visiting scientists andothers
The IOA genetics group has been fortunate to havehad several Ph.D. students from INGA countriesin the research group over the last decade. The studentscame from Thailand, Bangladesh and Malaysia andwere funded through international programs (e.g.,AADCP) and UK or local government, etc. There werealso visiting scientists from a number of countries,including India and Thailand, under various schemes.Every year, there are M.S. Aquaculture studentscarrying out their research projects in IOA.
IOA hopes to obtain funding for new projects inthe near future, and welcomes new collaborationsthrough INGA.
Basavaraju, Y., B.S. Renuka Devi, G. Mukthayakka, L.Purushotham Reddy, G.C. Mair, E.E. Roderick and D.J.Penman. 1998. Evaluation of marking and taggingmethods for genetic studies in carp. J. Biosci. (India)23: 585-593.
Carrasco, L.A.P., D.J. Penman and N. Bromage. 1999a.Evidence for the presence of sex chromosomes in theNile tilapia (Oreochromis niloticus) from synaptonemalcomplex analysis of XX, XY and YY genotypes.Aquaculture 173: 207-218.
Carrasco, L.A.P., D.J. Penman and N. Bromage. 1999b.Morphometric parameters of commercial interest intriploid rainbow trout, Oncorhynchus mykiss. Aquatic 6:http://AquaTIC.unizar.es/ (In Spanish).
Carrasco, L.A.P., D.J. Penman, S. Doroshov and N. Bromage.1998. Long-term, quantitative analysis of gametogenesisin autotriploid rainbow trout, Oncorhynchus mykiss. J.Reprod. Fertility 113: 197-210.
Castilho, R. and B. McAndrew. 1998. Two polymorphicmicrosatellite markers in the European seabass,Dicentrarchus labrax (L.). Anim. Genet. 29: 151.
Hussain, M.G., B.J. McAndrew and D.J. Penman. 1995b.Phenotypic variation in meiotic and mitotic gynogeneticdiploids of the Nile tilapia (Oreochromis niloticus L.).Aquacult. Res. 26: 205-212.
Hussain, M.G., D.J. Penman and B.J. McAndrew. 1996.Effects of triploidy on sexual maturation andreproduction in Nile tilapia, Oreochromis niloticus L., p.320-325. In R.S.V. Pullin, J. Lazard, M. Legendre, J.B.Amon Kothias and D. Pauly (eds.) Proceedings of theThird International Symposium on Tilapia inAquaculture. ICLARM, Manila, Philippines. 575 p.
Hussain, M.G., D.J. Penman and B.J. McAndrew. 1998.Production of heterozygous and homozygous clones inthe Nile tilapia. Aquacult. Int. 6: 197-205.
Hussain, M.G., B.J. McAndrew, D.J. Penman and P. Sodsuk.1994. Estimating gene-centromere recombinationfrequencies in gynogenetic diploids of Oreochromisniloticus L., using allozymes, skin colour and a putativesex-determination locus (SDL-2), P. 502-509. In A.Beaumont (ed.) Genetics and evolution of aquaticorganisms. Chapman and Hall.
Hussain, M.G., D.J. Penman, B.J. McAndrew and R.Johnstone. 1993. Suppression of first cleavage in theNile tilapia, Oreochromis niloticus L. - a comparison ofpressure and heat shocks. Aquaculture 111: 263-270.
Hussain, M.G., G.P.S. Rao, N.M. Humayan, C.F. Randall,D.J. Penman, D. Kime, N.R. Bromage, J.M. Myers andB.J. McAndrew. 1995. Comparative performance ofgrowth, biochemical composition and endocrine profilesin diploid and triploid Oreochromis niloticus L.Aquaculture 138: 87-98.
Klinbunga, S., D.J. Penman and B.J. McAndrew. 1998b. Apreliminary study of ribosomal DNA polymorphism in
Collaborative Projects with INGAMember Countries
References

Fish Genetics Research in Member Countries and Institutions of INGA148
the tiger shrimp, Penaeus monodon. J. Mar. Biotechnol.6: 186-188.
Klinbunga, S., D.J. Penman, B.J. McAndrew and A.Tassanakajon. 1999. Mitochondrial DNA diversity inthree populations of the giant tiger shrimp Penaeusmonodon. Mar. Biotechnol. 1: 113-121.
Klinbunga, S., S. Sodsuk, D.J. Penman and B.J. McAndrew.1996. An improved protocol for total DNA isolation andvisualisation of mtDNA RFLP(s) in tiger prawn, Penaeusmonodon. Thai J. Aquat. Sci. 3: 36-41.
Klinbunga, S., D.J. Penman, B.J. McAndrew, A. Tassanakajonand P. Jarayabhand. 1998a. Genetic variation,population differentiation and gene flow of the gianttiger shrimp (Penaeus monodon) inferred from mtDNA-RFLP data, p. 51-59. In T.W. Flegel (ed.) Advances inshrimp biotechnology. National Center for GeneticEngineering and Biotechnology, Bangkok, Thailand.
Kocher, T.D., W.-J. Lee, H. Sobolewska, D. Penman and B.McAndrew. 1998. A genetic linkage map of a cichlidfish, the tilapia (Oreochromis niloticus). Genetics 148:1225-1232.
Markert, J.A., T.D. Kocher, D. Penman and I. Karayucel.1999. Gene-centromere distances in tilapia, p.33. InPlant and animal genome. Vol. 7: Abstracts. San Diego,California, USA.
Martinez Rodriguez, G., M.C. Alvarez and B.J. McAndrew.1998. Genetic variability of European sea bass,Dicentrarchus labrax L.: data from a hatchery stock.Aquacult. Res. 29: 851-853.
McAndrew, B.J., K.J. Rana and D.J. Penman. 1993.Conservation and preservation of genetic variation inaquatic organisms, p. 295-336. In J.F. Muir and R.J.Roberts (eds.) Recent advances in aquaculture. Vol. 4.Croom Helm.
Myers, J.M., S.F. Powell and B.J. McAndrew. 1995a.Induction of tetraploidy in brown trout, Salmo trutta L.,using hydrostatic pressure. Aquacult. Res. 26: 229-232.
Myers, J.M., D.J. Penman, Y. Basavaraju, S.F. Powell, P.Baoprasertkul, K.J. Rana, N. Bromage and B.J.McAndrew. 1995b. Induction of diploid androgeneticand mitotic gynogenetic Nile tilapia (Oreochromisniloticus). Theor. Appl. Genet. 90: 205-210.
Penman, D.J. 1995. The ASEAN-EC AquacultureDevelopment and Coordination Programme (AADCP).EC Fish. Coop. Bull. 8(2): 13-17.
Penman, D.J., J.M. Myers and B.J. McAndrew. 1997.Restoration of diploid genotypes by androgenesis, p.469-473. Sciences et Techniques du Froids. Actes duColloques Refrigeration and Aquaculture, BordeauxAquaculture ’96, 20-22 March 1996.
Penman, D.J., M. Woodwark and B.J. McAndrew. 1995.Genetically modified fish populations, p. 22-27. InEnvironmental impacts of aquatic biotechnology.OECD, Paris.
Pongthana, N., P. Baoprasertkul and D.J. Penman. 1995b.Hormonal feminisation in the catfish Clarias
macrocephalus. Aquaculture 137: 158-159.Pongthana, N., D.J. Penman, J. Karnasuta and B.J.
McAndrew. 1995a. Induced gynogenesis in the silverbarb (Puntius gonionotus Bleeker) and evidence forfemale homogamety. Aquaculture 135: 267-276.
Pongthana, N., D.J. Penman, P. Baoprasertkul and B.Tongmee. 1995b. Sex identification in tilapia. Ext. Pap.No. 3, 20 p. National Aquaculture Genetics ResearchInstitute, Department of Fisheries, Thailand. (In Thai/English).
Pongthana, N., D.J. Penman, P. Baoprasertkul, M.G.Hussain, M.S. Islam, S.F. Powell and B.J. McAndrew.1999. Monosex female production in the silver barb(Puntius gonionotus Bleeker). Aquaculture 173: 247-256.
Prodoehl, P.A., A.F. Walker, R. Hynes, J.B. Taggart and A.Ferguson. 1997. Genetically monomorphic brown trout(Salmo trutta L.) populations, as revealed bymitochondrial DNA, multilocus and single-locusminisatellite (VNTR) analyses. Heredity 79: 208-213.
Rana, K. 1995. Cryopreservation of aquatic gametes andembryos: recent advances and applications, p. 85-89.In F.W. Goetz and P. Thomas (eds.) Proceedings of theFifth International Symposium on the ReproductivePhysiology of Fish, 2-8 July 1995, University of Texasat Austin, USA.
Rana, K.J., B.J. McAndrew, G. Wohlfarth and I. Macgowan.1996. Observations on intergeneric hybrids in tilapias,p. 391-397. In R.S.V. Pullin, J. Lazard, M. Legendre,J.B. Amon Kothias and D. Pauly (eds.) Proceedings ofthe Third International Symposium on Tilapia inAquaculture. ICLARM, Manila, Philippines. 575 p.
Rognon, X., M. Andriamanga, B. McAndrew and R.Guyomard. 1996. Allozyme variation in natural andcultured populations in two tilapia species: Oreochromisniloticus and Tilapia zillii. Heredity 76: 640-650.
Sarder, M.R.I., D.J. Penman, J.M. Myers and B.J. McAndrew.Production and propagation of fully inbred clonal linesin the Nile tilapia (Oreochromis niloticus L.). J. Exp. Zool.(In press).
Sodsuk, P.K., B.J. McAndrew and G.F. Turner. 1995.Evolutionary relationships of the Lake MalawiOreochromis species: evidence from allozymes. J. FishBiol. 47: 321-333.
Stone, C.E., J.B. Taggart and A. Ferguson. 1997. Single locusminisatellite DNA variation in European populationsof Atlantic salmon (Salmo salar L.). Heredity 126: 269-275.
Taggart, J.B., P.A. Prodoehl and A. Ferguson. 1995a. Geneticmarkers for Atlantic salmon (Salmo salar L.): single locusinheritance and joint segregation analyses ofminisatellite (VNTR) DNA loci. Anim. Genet. 26: 13-20.
Taggart, J.B., E. Verspoor, P.T. Galvin, P. Moran and A.Ferguson. 1995b. A minisatellite DNA marker fordiscriminating between European and North AmericanAtlantic salmon (Salmo salar). Can. J. Fish. Aquat. Sci.

149
52: 2305-2311.Trillo, E.M., Z.Z. Rutczynska, B. McAndrew, V. Vincek, F.
Figueroa, H.S. Ultmann and J. Klein. 1998. Linkagerelationships and haplotype polymorphism amongcichlid mhc class ii b loci. Genetics 149:1527-1537.
Wong, J.T.Y. and B.J. McAndrew. 1994. Allozyme variationin riverine and lacustrine populations of Macrobrachiumnipponense (de Haan). Aquacult. Fish. Manage. 25: 393-400.
Woodwark, M., D.J. Penman and B.J. McAndrew. 1994.Genetic modification of fish: a UK perspective.Genetically Modified Organisms Res. Rep. No. 2, UKDepartment of the Environment. 84 p.
Overview of Aquaculture Genetics Researchin the Institute of Aquaculture, University of Stirling

Fish Genetics Research in Member Countries and Institutions of INGA150

151
ABSTRACT
The School of Biological Sciences, University of Wales Swansea (UWS), has for many years, worked on a range oftopics relating to the genetics of aquatic species, from fundamental studies on population structure, speciation andmolecular evolution to very applied projects for developing better fish for aquaculture.
The university is contracted by the Department of International Development (DFID), U.K. to manage the DFIDResearch Programme in Fish Genetics which aims to harness the wide range of genetic techniques now availableto provide productive inputs to aquaculture and enhancement fisheries in developing countries targeted at thealleviation of poverty and improvement of livelihood among rural poor. While tilapias and carps, being low down in thefood chain and appropriate for low-input aquaculture, are the species of most direct interest, recent projects haveinvolved kapenta (Limnothrissa miodon), shrimps and groupers.
Currently, five projects are running within the programme. These are focused on: (i) improved tilapia through transgenesis(Southampton and UWS); (ii) improved production in India of common carp (Cyprinus carpio) and Catla (Catla catla)(Bangalore, Stirling and UWS); (iii) Socioeconomic factors in uptake of new technology (Centre for DevelopmentStudies, UWS); (iv) development of genetic technology of monosex production in Oreochromis mossambicus forSouth Africa (Stellenbosch University and UWS); and (v) development and extension of the YY male technology in O.niloticus in Southeast Asia (Asian Institute of Technology, Thailand; Central Luzon State University, Philippines; andUWS).
_____________________a Current address: Aquaculture and Aquatic Resources Management Program of the Asian Institute of Technology,P.O. Box 4, Klong Luang, Pathumthani 12120, Thailand.
RESEARCH ON THE GENETICSOF AQUATIC ORGANISMS
AT THE SCHOOL OFBIOLOGICAL SCIENCES,
UNIVERSITY OF WALES SWANSEA
G.C. Maira and J.A. BeardmoreSchool of Biological SciencesUniversity of Wales Swansea
Singleton Park, Swansea SA2 8PP, Wales
MAIR, G.C. and J.A. BEARDMORE. 2001. Research on the genetics of aquatic organisms at the School of Biological Sciences,University of Wales Swansea, p. 151-159. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in membercountries and institutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179p.
Research on the Genetics of Aquatic Organismsat the School of Biological Sciences, University of Wales Swansea

Fish Genetics Research in Member Countries and Institutions of INGA152
Introduction
The School of Biological Sciences (SBS), University ofWales Swansea (SWS) has, for many years, worked ona wide range of topics on the genetics of aquatic species,from fundamental studies on population structure,speciation and molecular evolution to very appliedprojects developing better fish for aquaculture.Although members of the school have worked on adiverse range of species, the major species ofcommercial interest on which research has beenconducted include tilapias, carps, salmonids, bivalvemolluscs and Artemia. This paper reports progress insome of the more recent research conducted in Swanseain collaboration with associates throughout the world.The major emphasis of recent work has been conductedunder the auspices of the British GovernmentDepartment for International Development’s FishGenetics Research Programme (DFID/FGRP) whichhas been managed by UWS since its inception in 1990.
DFID Fish Genetics Programme
The overall objective of this programme is to harnessthe wide range of genetic techniques now available toprovide productive inputs to aquaculture andenhancement fisheries in developing countries.
The ultimate aim is to benefit livelihoods of poorpeople and to aid in the alleviation of poverty throughimprovement of production in enhanced andmaintained populations of fish and shellfish, especiallyby small farmers. The programme has a dominanttheme of sustainability in the widest sense (includingenvironmental impacts of all kinds), paying full regardto the provisions of the Convention on Biological
Diversity on access to genetic resources. In practicalterms, research uses the wide range of analytical,manipulative and breeding techniques available ingenetics to generate better strains of fish. This workfulfils the programme’s purpose in aquaculture andenhancement fisheries of improved performance inproductivity giving benefits of increased income, foodsecurity and food costs. While any approach toimproved production carries a risk to sustainability,genetic approaches are intrinsically more likely tosupport sustainable development than approachesbased on increased levels of inputs which frequentlyhave environmentally undesirable effects.
While the global yield from captive fisheries is static,at best, the world output from aquaculture whichalready forms about 20% of shellfish and finfishproduction, is increasing at about 10% per annum. Inpopulous developing countries, such as India, Chinaand the Philippines, the yield from aquaculture forms30-60% of total aquatic production. Aquaculture, thus,has an increasingly important role in world nutritionespecially in developing countries.
The range of researchable constraints to which geneticscan supply solutions is considerable. Major practicalimprovements in the diversion of energy fromreproduction to growth through control ofreproduction, greater resistance to disease andextending tolerance to environmental constraints liketemperature, in which genetic factors play a significantrole, are to be expected. In general, solutions generatemore and better fish for the same inputs throughincreased efficiency and thus do not impose additionalburden on scarce resources (see Table 1). In applyinggenetics-based technologies to greater efficiency of
Table 1. Examples of targets for improved products in aquaculture and how these can be addressed using genetic techniques.
Target for increasedProduction
Removal of problems associated with early sexualmaturity and unwanted reproduction
Increased growth rate
Expanded geographic utilization withoutenvironmental risks to ecosystems
Increased environmental tolerances such as coldor salinity
Improved quality of stock
Improved disease resistance
Solution using genetic techniques
Production of monosex by developing “supermales” siringonly male offspring or “neomales” siring only femaleoffspring or sterile triploids
Transgenics of growth hormone gene; geneticcharacterization and breeding programs
Transgenics which are sterile or are inviable innature; production of triploids
Selective breeding and/orcrossbreeding/hybridization
Development of good broodstock and user-friendlymaintenance protocol for this
Selective breeding; transgenics
Examples inrelevant species
Tilapia, silver barb
Tilapia, salmon,carp
Tilapia (especiallyfor Africa)
Tilapia hybrids
All species,especially carp
Shrimp

153
production in the low-input aquaculture systemscharacteristic of developing countries, the needs ofpoor families seeking to develop better livelihoods arebeing addressed in a direct and useful way. The greaterefficiency of improved strains should lead, ultimately,to significantly cheaper food for both rural and urbanpoor.
The scale of benefit to be expected from applicationsof genetic technologies is very large. The evidence fromthe many species of cultured crop plants anddomesticated farm animals, which are geneticallyimproved, is compelling. Gains in productivity arisingfrom application of techniques such as selectivebreeding frequently exceed 100%. Even in salmon, theonly aquatic species in which systematic long-termbreeding has been carried out, the gains in productivityalready amount to more than 40%.
While tilapias and carps are the species of most directinterest within the programme, as they feed low downin the food chain and are best adapted to low-inputculture or enhancement systems, recent projects haveinvolved species such as kapenta, shrimp and groupers.
In addition to the use of genetic technology, animportant need for radically improved and rationalgenetic management of broodstock in aquaculture isbecoming increasingly evident in many areas. Notinfrequently, in some societies, traditional custom mayact against best genetic practice by, for example, thechoice of the smallest individuals in a harvest to bethe parents of the next generation because these haveleast value at market. Thus, establishing protocols forthe proper genetic management of broodstock is afactor of considerable importance in both aquacultureand enhancement fisheries. The selection ofappropriate techniques chosen from the nowformidable array of methodologies available requiresthat the end product be environmentally neutral (asin use of sterile strains). However, the advent ofimproved strains takes place within communities andecosystems. It follows that suitable concurrent studiesneed to accompany the genetic technology to ensurethat environmentally deleterious effects do not ensue.Furthermore, it must be assured that acceptance of suchtechniques is assisted by good understanding ofsocioeconomic structures, that the benefits ofgenetically improved strains are available to supportlivelihoods of families with few resources and that ingeneral terms the objectives of UK InternationalDevelopment Aid are met in identifying appropriate
target beneficiaries. The availability of water is obviouslya central need but even poor families can often share inimproved aquaculture production, and benefits canaccrue to nonproducers through provision ofemployment and increasing the availability of cheapfish protein.
The dissemination of programme products is assistedby income-generating activities of a UWS companyFishgen Ltd. and its sister organization at thePhilippines’ Central Luzon State University (CLSU),Phil-Fishgen.
Genetic Projects with UWS Involvement
Table 2 lists the current and recent projects conductedunder the DFID/FGRP.
Sex determination and the YY maletechnology in the Nile tilapia
This project arose from early research work in Swanseaon the genetics of the inheritance of sex in tilapia.This work has developed toward the current consensusthat, in the commercially important Oreochromis species,sex determination is “predominantly” monofactorial,being controlled by sex chromosomes or primary sexdetermining gene(s). Research using sex reversal,progeny testing and chromosome set manipulation hasrevealed two alternative “sex chromosome” models.In O. niloticus, the female is homogametic XX, the malebeing heterogametic XY (Mair et al. 1991a) while inthe closely related O. aureus, the alternative model ofheterogametic WZ females and homogametic ZZ malesapplies (Mair et al. 1991b). There is also substantialevidence for effects of one or more autosomal geneson sex ratio together with an increasingly well-documented effect of temperature in influencing sex.Elevated temperatures (~36ºC) during the period ofsex differentiation have been shown to increase theproportion of males in putative monosex femaleO. niloticus and, to a lesser degree, to increase theproportion of females in putative all male progeny(Abucay et al. 1999). Similarly high temperatures havebeen shown to modify sex ratios in O. aureus (Mair etal. 1990). The only recorded effect of low temperatureswas to increase the proportion of males in O. mossambicus(Mair et al. 1990).
Based on the theory of predominantly monofactorialsex determination, it has proved possible to manipulatesex ratio using a combination of sex reversal and
Research on the Genetics of Aquatic Organismsat the School of Biological Sciences, University of Wales Swansea
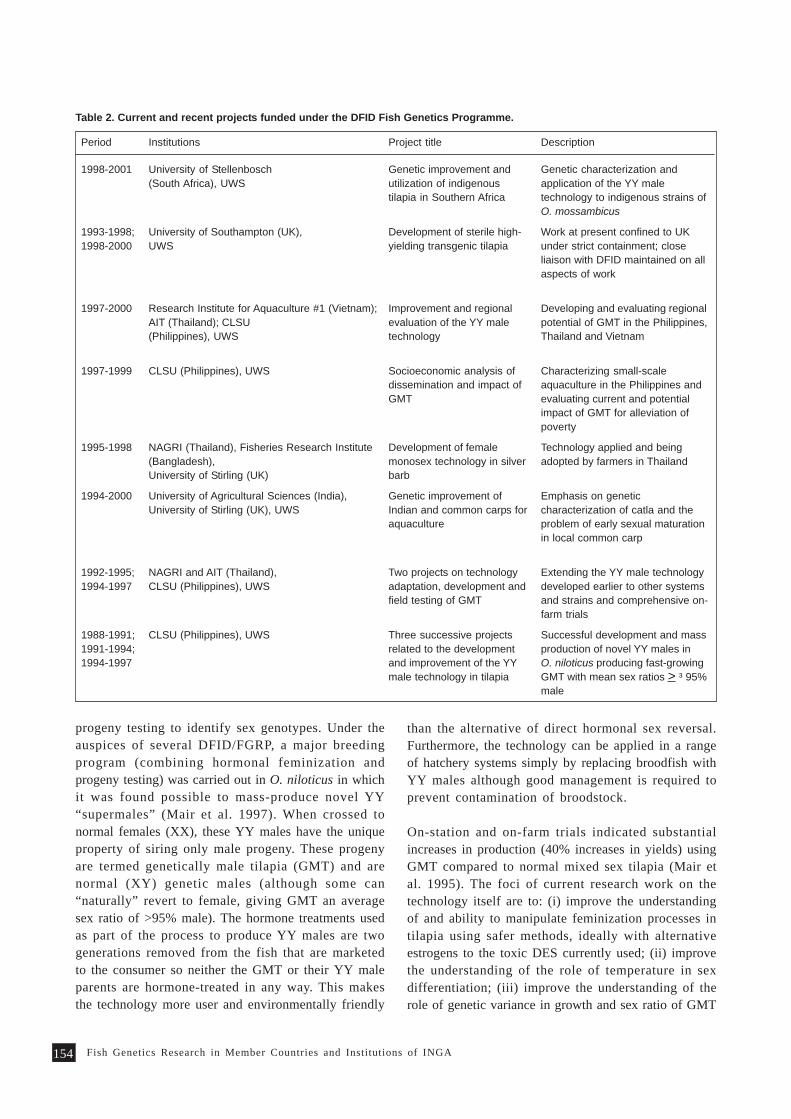
Fish Genetics Research in Member Countries and Institutions of INGA154
progeny testing to identify sex genotypes. Under theauspices of several DFID/FGRP, a major breedingprogram (combining hormonal feminization andprogeny testing) was carried out in O. niloticus in whichit was found possible to mass-produce novel YY“supermales” (Mair et al. 1997). When crossed tonormal females (XX), these YY males have the uniqueproperty of siring only male progeny. These progenyare termed genetically male tilapia (GMT) and arenormal (XY) genetic males (although some can“naturally” revert to female, giving GMT an averagesex ratio of >95% male). The hormone treatments usedas part of the process to produce YY males are twogenerations removed from the fish that are marketedto the consumer so neither the GMT or their YY maleparents are hormone-treated in any way. This makesthe technology more user and environmentally friendly
than the alternative of direct hormonal sex reversal.Furthermore, the technology can be applied in a rangeof hatchery systems simply by replacing broodfish withYY males although good management is required toprevent contamination of broodstock.
On-station and on-farm trials indicated substantialincreases in production (40% increases in yields) usingGMT compared to normal mixed sex tilapia (Mair etal. 1995). The foci of current research work on thetechnology itself are to: (i) improve the understandingof and ability to manipulate feminization processes intilapia using safer methods, ideally with alternativeestrogens to the toxic DES currently used; (ii) improvethe understanding of the role of temperature in sexdifferentiation; (iii) improve the understanding of therole of genetic variance in growth and sex ratio of GMT
Table 2. Current and recent projects funded under the DFID Fish Genetics Programme.
Period
1998-2001
1993-1998;1998-2000
1997-2000
1997-1999
1995-1998
1994-2000
1992-1995;1994-1997
1988-1991;1991-1994;1994-1997
Institutions
University of Stellenbosch(South Africa), UWS
University of Southampton (UK),UWS
Research Institute for Aquaculture #1 (Vietnam);AIT (Thailand); CLSU(Philippines), UWS
CLSU (Philippines), UWS
NAGRI (Thailand), Fisheries Research Institute(Bangladesh),University of Stirling (UK)
University of Agricultural Sciences (India),University of Stirling (UK), UWS
NAGRI and AIT (Thailand),CLSU (Philippines), UWS
CLSU (Philippines), UWS
Project title
Genetic improvement andutilization of indigenoustilapia in Southern Africa
Development of sterile high-yielding transgenic tilapia
Improvement and regionalevaluation of the YY maletechnology
Socioeconomic analysis ofdissemination and impact ofGMT
Development of femalemonosex technology in silverbarb
Genetic improvement ofIndian and common carps foraquaculture
Two projects on technologyadaptation, development andfield testing of GMT
Three successive projectsrelated to the developmentand improvement of the YYmale technology in tilapia
Description
Genetic characterization andapplication of the YY maletechnology to indigenous strains ofO. mossambicus
Work at present confined to UKunder strict containment; closeliaison with DFID maintained on allaspects of work
Developing and evaluating regionalpotential of GMT in the Philippines,Thailand and Vietnam
Characterizing small-scaleaquaculture in the Philippines andevaluating current and potentialimpact of GMT for alleviation ofpoverty
Technology applied and beingadopted by farmers in Thailand
Emphasis on geneticcharacterization of catla and theproblem of early sexual maturationin local common carp
Extending the YY male technologydeveloped earlier to other systemsand strains and comprehensive on-farm trials
Successful development and massproduction of novel YY males inO. niloticus producing fast-growingGMT with mean sex ratios > ³ 95%male

155
including relative performance of intra- and interstraincrosses, G x E interactions and response to selection;(iv) assess the relative performance of GMT and theirparental stock in Thai and Vietnamese aquaculturesystems; and (v) investigate the potential for producingearly season GMT using new species or hybrids inVietnam.
The outputs of the research work on the developmentof the YY male technology are now being widelydisseminated in the Philippines and Thailand throughthe distribution of YY male and normal femalebroodstock to accredited hatcheries. This process isbeing conducted through financially self-sustainingmechanisms coordinated by UWS and CLSU in thePhilippines and the National Aquaculture GeneticsResearch Institute (NAGRI) of the Department ofFisheries in Thailand. The impact of GMT in SoutheastAsia is likely to be considerable. For example, NAGRIestimates indicate that approximately 25% of thetilapia harvested in Thailand in 1999 are GMT. Thegains from additional income and dietary protein arenot precisely quantified but are considerable. Issuesrelated to the social and economic impact of theintroduction of GMT in the Philippines have beenaddressed in another DFID project.
Social and economic impact assessment ofGMT
This project (1997-1999) was carried out ascollaboration between the Freshwater AquacultureCenter (FAC) of CLSU and School of BiologicalSciences and the Centre for Development Studies ofUWS. The overall objective of the project was toimprove equitable uptake of the products of YY maletechnology by all sectors of the tilapia culture industryin the Philippines. There were four major activities:• Characterization of small-scale aquaculture in the
Philippines;• Assessment of the impact of three genetics-based
technologies - GMT; genetically improved farmtilapia (GIFT) and sex-reversed tilapia (SRT) - inhatcheries, and especially, grow-out farms;
• Study of interrelationships between differentsectors of the aquaculture industry; and
• Conduct trials of GMT among currentlynonbenefiting sectors of the industry.
The project concluded that the current “passive”dissemination process is achieving its originalobjectives of dissemination of research outputs (namely,improved fish) and financial sustainability of research,
development and dissemination. However, it was evidentthat hatcheries which are benefiting are relatively large-scale, semi-intensive and has commercial operations,and that small-scale hatcheries were effectively excludedfrom adopting the technology on the basis of cost andawareness. At a premium of only 12.5% over normalmixed sex fish, GMT was the cheapest of the available“improved” fish. Furthermore, there is a relativelywidespread awareness of GMT across all sectors of theindustry, to a greater extent than the alternative monosextechnology of SRT which has been promoted in thePhilippines for 20 years. However, the nature ofdissemination and lack of targeted promotion arelimiting adoption of GMT to medium-scale tilapiagrowers in the mainstream of the industry, with fullaccess to its resources.
The results provided valuable pointers with regard tothe reformulation of the dissemination program. Theresults from field trials in which two externalorganizations participated actively (an NGO and amultilateral donor funded project) indicated thatnongovernmental organizations (NGO) and people’sorganizations (PO) can be potentially valuable partnersin dissemination efforts targeted at small-scaleproducers, e.g., improved relations between FAC-CLSU and other institutions, such as NGO.
Private small-scale hatcheries operating outsidefingerling trading networks were also identified ashaving significant potential to enable small-scale grow-out producers to adopt GMT production. Dissemina-tion through state sector hatcheries appearedinefficient and failed to target small-scale producers.These and other findings have been incorporated in aseries of recommendations to guide Phil-Fishgen inits dissemination and research efforts.
According to supplementary research, tilapia has animportant role in the diet of poor in inland provincesin Luzon. Enhanced production of tilapia can thereforecontribute to better food security among the poor ofsuch provinces, as well as protect their entitlement tononmarket food fish.
Overall, the results from this project suggest that small-scale farmers benefit from adoption of GMT wheredissemination is accurately targeted and coordinatedwith agencies that can provide credit, training andextension support. These efforts can act as catalyst toattract new entrants to aquaculture and to provideextension material. Also indicated is that enhanced
Research on the Genetics of Aquatic Organismsat the School of Biological Sciences, University of Wales Swansea

Fish Genetics Research in Member Countries and Institutions of INGA156
tilapia production associated with the adoption of GMTby small, medium and large-scale producers can helpthe rural poor by increasing supply of food fish criticalto their food.
Genetic improvement of O. mossambicus inSouthern Africa
Having begun in early 1999, this project is very muchin its formative stages. The overall developmentobjective is the enhancement of livelihood of rural poorin Southern Africa through: (i) improved productionof fish from existing aquaculture facilities utilizingenhanced varieties of indigenous fish species; (ii) sus-tainable use of existing resources, infrastructure andexpertise with the settlement of small-scale farmers asindependent fish producers;(iii) building entrepre-neurial capacity and providing employmentopportunities in an environment of decreasing demandfor farm labor; and (iv) increased availability ofaffordable fish protein through expanded culture oflow- input, low-cost species.
The specific research objectives are as follows:• Develop an assembly of accessions of up to 15
strains of O. mossambicus from throughout SouthernAfrica; conduct growth performance trials of aminimum of eight strains, under communalstocking in two environments, to include farmcages and earthen ponds;
• Genetically characterize available strains ofO. mossambicus (minimum of eight) using moleculartechniques (microsatellite DNA loci, amplifiedfragment length polymorphisms and/or restrictionendonuclease analysis of mitochondrial DNAhaplotypes); strains will be characterized for strainspecific markers, levels of genetic variability andpopulation structure;
• Adapt YY male technology to strains ofO. mossambicus chosen on the basis of growth dataand species purity; YY males will be generatedusing two approaches: (a) the standard breedingprogram combining feminization and progenytesting (Mair et al. 1997) and (b) the more rapidmeans using androgenesis which may enableproduction of a few YY males in 1.5 generations(1-1.5 years) compared to 2.5 (1.5-2.5 years) inthe standard breeding program;
• Conduct on-farm trials of GMT in both cages andponds; comparisons will be made withnonimproved, locally available O. mossambicus ascontrols; and
• Determine potential social and economic impactsof the promotion of improved fish throughparticipatory research with farmers and potentialconsumers.
Production of transgenic tilapia
Transgenesis is one of the most promising technologiesfor relatively rapid genetic improvements. It involvesthe introduction, via techniques of genetic engineering,of a DNA sequence into a recipient organism to confernovel phenotypic characteristics on that organism(Beardmore 1997). Typically, the desired genes are firstidentified, sequenced and then cloned. Multiple copiesof the gene are then introduced to the fertilized eggs,commonly by microinjection. At a later stage ofdevelopment, cells of the organism are tested todetermine whether copies of the transgene havebecome incorporated into the genome and whetherthis incorporation is in all cells or only in the cells ofsome tissues (i.e., a mosaic). After incorporation isdetermined, the organism can be evaluated todetermine if the transgene product is being expressedand in what amounts; and then whether the transgeneis inherited and expressed in the next generation viathe germ cells. Inheritance of the transgene would berequired to develop true breeding lines of thetransgenic organism. Among the several attempts todevelop transgenic lines of tilapia, one of the mostsuccessful has been conducted under DFID, in acollaborative project between the University ofSouthampton and UWS.
The study, which is presently limited to stringentlycontained laboratory conditions in the UK, involvesthe introduction of a fish growth hormone gene intoO. niloticus (Rahman and Maclean 1999). The majorpart is conducted at Southampton University with therole of UWS in gene mapping for tilapia. Introductionof the transgene by microinjection incorporation wasdetected. Initial transmission rate from G0 to G1
generation was found to be less than 10% in these lines,indicating a mosaic distribution of the transgene inthe germ cells. However, transmission rates from thefirst to the second generations were found to followthe expected Mendelian ratios. The chinook salmongrowth hormone was produced in several generationsof the transgenic tilapia indicating expression of thegene, resulting in dramatic growth enhancement withthe average weight of the transgenic fish being fourtimes that of their nontransgenic siblings.

157
Transgenesis appears to offer very considerablepotential for enhancement of yield in tilapia. However,the rate of genetic change in transgenics is such thatphenotypic and behavioral properties cannot easilybe predicted and the introduction of fish forcommercial aquaculture faces many constraints. Therisks to the environment posed by the uncontrolledintroduction of transgenic fish need to be adequatelyassessed, and many governments are adopting cautiouspolicies with regard to their introductions. In addition,consumer response to these genetically modifiedorganisms (GMOs) in some countries may be verynegative to the extent that adoption by farmers mayinvolve significant economic risks. A lot of theseconstraints may be overcome if guaranteed steriletransgenic tilapia can be produced through efficientmethods of triploidization or, in the medium to long-term, through disruption of the physiological pathwaysof reproduction via the introduction of new antisensetransgenes. Very thorough and comprehensive fieldtrials will be required prior to general release of suchorganisms to aquaculture.
Genetic improvement strategies for carp inKarnataka, Southern India
This project is a collaboration among the Universityof Agricultural Sciences, Bangalore, India, UWS andthe Institute of Aquaculture, Stirling University. Theprincipal objectives of the project are to: (i) investigatethe present status of the cultured Indian major carpCatla (Catla catla) in Karnataka State, India, and designand initiate a program for the genetic improvementof the cultured strains; and (ii) investigate thephenomenon of early sexual maturation and unwantedreproduction during culture of common carp (Cyprinuscarpio) in Karnataka and develop solutions throughgenetic or hormonal manipulations.
Work on catla has slow progress due to the longgeneration time, and short and unpredictablespawning season in Southern India. Early in theproject, fry and fingerlings were collected fromdomesticated stocks in several major hatcheries inKarnataka and from wild stocks from the Rivers Gangesand Brahmaputra. Fin tissue samples were alsocollected from other wild and hatchery populationsfrom different parts of South Asia.
A survey of broodstock management practicesconcluded that the Karnataka hatchery stocks, which
had been reproductively isolated since the introductionof the species in the 1950s, were likely to be inbredwith rates ranging from 4.7 to 10% per annum.However, the results from genetic characterizationusing randomly amplified polymorphic DNA analysis(RAPD) and microsatellite DNA loci and restrictionfragment length polymorphism (RFLP) of mito-chondrial DNA were ambiguous. Results from RAPDof nuclear DNA did not enable the discrimination ofhatchery stocks and indicated that the levels of geneticvariation were similar both within and between the wildand hatchery stocks. Analysis of results from fourmicrosatellite DNA loci showed both hatchery and wildstocks to have similar levels of average heterozygositywith most populations in Hardy-Weinberg equilibrium.Furthermore, there were similar mean numbers ofalleles at each locus between the hatchery and the wildstocks, contradicting the hypothesis that the hatcherystocks are inbred.
Results from growth trials over three successive yearshave consistently indicated higher yields fromcrossbreds between the two major hatchery strainscompared to the pure strains, indicating positiveheterosis for commercial traits. Inclusion of wild caughtstocks in these trials has not yet been possible.
Studies on the feasibility of the application ofmicrosatellite markers for pedigree analysis producedpromising results but require further primers to bedeveloped to provide the necessary levels of resolutionby evaluating more loci. Work on the development ofadditional primers is well underway. If microsatellitemarkers can be used for identification of fish at thefamily level they may become useful tools incommunally stocked growth trials and in farm-basedselection programs.
The work on common carp has progressed well. Earlyon-station and on-farm trials revealed relatively earlymaturation of local stocks of common carp under arange of culture conditions although spawning did notoccur in the absence of aquatic vegetation as asubstrate. Fish became mature at less than six monthsold and as small as 150 g in weight. Furthermore, GSIwas as high as 20% in both males and females prior toattaining a harvestable size.
Three approaches to solving this problem have beenevaluated. Masculinization protocols have beendeveloped prior to identifying sex-reversed XX males,
Research on the Genetics of Aquatic Organismsat the School of Biological Sciences, University of Wales Swansea

Fish Genetics Research in Member Countries and Institutions of INGA158
which can be used to produce all-female progeny.Protocols for induction of triploidy (with triploidinduction rates up to 100%) have also been developed.Growth trials of sterile triploid fish up to the age atwhich sexual maturation occurs in diploid fish revealedno differences in growth rates although dress-outweights of triploid fish were greater than those ofdiploids, post-sexual maturation.
Recently a selected strain was introduced from Vietnamand early growth trials have indicated that this hasfaster growth rates and later sexual maturation thanthe local strains of common carp. Analysis of standardbroodstock management protocols among thedominant state hatcheries revealed potential forselection for early maturation and revised managementprotocols are presently being developed in conjunctionwith the Karnataka Department of Fisheries.
Other research
Researchers at UWS are active in many other areas ofresearch outside of those under DFID/FGRP.
Genetic diversity of silver barb
This three-year project (started in 1998) is fundedby the DFID Competitive Research Facility, incollaboration with ICLARM, developing the themeof the interrelations of aquaculture and biodiversity(Beardmore et al. 1997). The project aims to collectand collate data on genetic diversity of wildpopulations of the silver barb, Barbodes gonionotus(formerly Puntius gonionotus) throughout its naturalrange in order to document genetic resources. Thisis being achieved through the application of a suiteof biochemical and molecular techniques at thelaboratories at UWS, with emphasis on mtDNA andmicrosatellite loci. These data will be analyzed todetermine the levels of genetic variability in thenatural stocks of B. gonionotus and to identify thosepopulations that can act as valuable reserves ofgenetic diversity. It is hoped that the degree ofresolution of these techniques will be sufficient, whencombined with indigenous knowledge, to determinethe source of introductions of any stocks that maybe feral and non-native.
Samples for B. gonionotus were collected from severalsites in Indonesia and Indochina, and furthersampling trips are planned. To date the control
region of the mtDNA molecule has been surveyed infour populations, two from Java, Indonesia, and twofrom Vietnam, with a number of fixed differencesnoted between the samples from the two regions.Molecular markers are now being used to survey thepopulations collected so far. With the isolation of alarge number of positive clones from themicrosatellite libraries, a large suite of microsatellitemarkers is anticipated for use, in addition to thosealready obtained from other researchers.
Collectively, it is hoped that the techniques andapproaches used in this project would represent avaluable case study investigating the genetic diversityof a commercially important species. Furthermore, itis intended that the data gathered can be used torecommend a long-term strategy for the conservationand management of this diversity in relation to futureintroductions and the predicted expansion of thefarming of this species.
Future Directions
Many of the research themes highlighted above willcontinue. DFID/FGRP projects will focus even moretightly on potential impacts of research outputs on thealleviation of poverty. It is likely that most projects willinclude research components evaluating importantsocial and economic factors related to the role ofincreased efficiency of aquaculture productionachieved through application of genetics-basedtechnologies, on the livelihood of the poor.
Following are other areas of research currently beingconsidered (not necessarily within the framework ofthe DFID/FGRP):• In tilapia, studies on the use of clonal lines as
internal controls in growth trials, comparisons andintegration of different genetic improvementstrategies, and research on dissemination strategiesof improved fish for the benefit of the poor;
• Surveys on existing broodstock managementprotocols prevalent for commercially importantspecies in developing countries and their likelyimpact upon seed quality;
• Strain comparisons of common carp in the tropics;and
• Application of molecular markers in fisheries stockassessment in the Mekong basin.

159
References
Abucay, J.S., G.C. Mair, D.O.F. Skibinski. and J.A. Beardmore. 1999.Environmental sex determination: the effect of temperatureand salinity on sex ratio in Oreochromis niloticus L. Aquaculture173: 219-234.
Beardmore, J.A. 1997. Transgenics and allotransgenics. TransgenicRes. 6: 107-108.
Beardmore, J.A., G.C. Mair and R.I. Lewis. 1997. Biodiversity inaquatic systems in relation to aquaculture. Aquacult. Res.28(10): 829-839.
Mair, G.C., J.S. Abucay, J.A. Beardmore and D.O.F. Skibinski. 1995.Growth performance trials of genetically male tilapia (GMT)derived from YY-males in Oreochromis niloticus L.: on stationcomparisons with mixed sex and sex reversed malepopulations. Aquaculture 137: 313-322.
Mair, G.C., J.S. Abucay, D.O.F. Skibinski, T.A. Abella and J.A.Beardmore. 1997. Genetic manipulation of sex ratio for thelarge-scale production of all-male tilapia, Oreochromis niloticus.Can. J. Fish. Aquat. Sci. 54: 396-404.
Mair, G.C., J.A. Beardmore, and D.O.F. Skibinski. 1990.Experimental evidence for environmental sex determinationin Oreochromis species, p. 555-558. In R. Hirano and I. Hanyu(eds.) The Second Asian Fisheries Forum. Asian FisheriesSociety, Manila, Philippines.
Mair, G.C., A.G. Scott, D.J. Penman, J.A. Beardmore and D.O.F.Skibinski. 1991a. Sex determination in the genus Oreochromis.1. Sex reversal, gynogenesis and triploidy in O. niloticus (L).Theor. Appl. Genet. 82: 144-152.
Mair, G.C., A.G. Scott, D.J. Penman, D.O.F. Skibinski and J.A.Beardmore. 1991b. Sex determination in the genus Oreochromis.2. Sex reversal, hybridisation, gynogenesis and triploidy in O.aureus Steindachner. Theor. Appl. Genet. 82: 153-160.
Rahman, M.A. and N. Maclean. 1999. Growth performance oftransgenic tilapia containing an exogenous piscine growthhormone gene. Aquaculture 173: 333-346.
Research on the Genetics of Aquatic Organismsat the School of Biological Sciences, University of Wales Swansea

Fish Genetics Research in Member Countries and Institutions of INGA160

161
ABSTRACT
An overview of genetics research in the Fish Culture and Fisheries Group, Department of Animal Science, WageningenUniversity and Research Centre, the Netherlands, is presented. The major model species for genetics research iscommon carp, Cyprinus carpio, L. In carp, several inbred strains have been produced by gynogenesis orandrogenesis. Techniques for gynogenesis and androgenesis, hormonal sex reversal and cryopreservation, andthe consequences of inbreeding and crossbreeding in terms of genetic and phenotypic variation are discussed.Theoretical considerations for application of gynogenesis and androgenesis in selective breeding programs areillustrated with experimental results and a case study on selective breeding for stress response. The consequencesof inbreeding for sex determination and differentiation are also presented.
GENETICS RESEARCH ATWAGENINGEN UNIVERSITY
AND RESEARCH CENTRE,THE NETHERLANDS
H. Komen, M. Tanck and N. RuaneFish Culture and Fisheries Group, Department of Animal Sciences
Wageningen University and Research CentreP.O. Box 338, 6708 AH Wageningen, The Netherlands
KOMEN, H., M. TANCK and N. RUANE. 2001. Genetics research at Wageningen University and Research Centre, theNetherlands, p. 161-167. In M.V. Gupta and B.O. Acosta (eds.) Fish genetics research in member countries andinstitutions of the International Network on Genetics in Aquaculture. ICLARM Conf. Proc. 64, 179 p.
Introduction
Research by the genetics section of the Fish Cultureand Fisheries Group at the Wageningen University,Netherlands focuses on the use of induced gynogeneticand androgenetic reproduction in selective breedingprograms. The common carp (Cyprinus carpio) is thespecies mainly used in the studies, although somebreeding work with the African catfish, Clarias gariepinusis performed. The following are the main areas ofresearch since 1992:
1. Development of a toolbox for gyno/androgenesis, for sex control and cryopre-servation of gametes;
2. Development of a theoretical framework forbreeding with gyno/androgenesis in fish;
Genetics Research at Wageningen University and Research Centre, The Netherlands
3. Case studies in which the principles laid downin the theoretical framework could be tested;and
4. Development of genetic markers which couldbe used in combination with gyno/androgenesis to study quantitative trait loci(QTL) associations for traits of interest.
A brief description of the progress in each of these stagesis given below.
Common carp can be reproduced by gynogenesis orandrogenesis. In gynogenesis, eggs are fertilized with
Principles of Androgenesisand Gynogenesis

Fish Genetics Research in Member Countries and Institutions of INGA162
variance since genetic variance does not increase.Theoretically, the additive genetic relation betweenparent and offspring equals 1 (Fig.1; Bongers et al.1998). Therefore, gynogenetic reproduction could beuseful for selection purposes when one is interested toestimate breeding values of individual dams (or sires inandrogenesis) included in a breeding program.Likewise, gynogenesis can be used to estimate the geneticload of a dam (or sire) included in a breeding program.In this case, the expression of deleterious recessive genesis measured. However, all estimations based onphenotypic variances should be treated with cautionsince the phenotypic variance can also increase due totreatment effects or increased sensitivity toenvironmental influences (Bongers et al. 1997a).
Three types of environmental variance (VE) should beconsidered within homozygous offsprings:
1. “true” VE (interindividual variance);2. VE, due to developmental instability (DI,
intraindividual variance); and3. VE originating from embryonic damage (ED)
caused by the chromosome manipulationtreatment.
To investigate the relative contributions of each of thesefactors, homozygous (F = 1) androgenetic andgynogenetic families and partly heterozygous (2pb-gynogenesis: F = 0.79) gynogenetic families wereobserved. All families were produced from geneticallyidentical parents. DI and ED were determined bymeasuring fluctuating asymmetry (FA) of five bilateralsymmetric morphometric characteristics (Bongers et al.1997c).
The androgenetic groups showed highest FA andvariations caused by ED, followed by 2pb-gynogeneticand homozygous gynogenetic groups, respectively (Table1). It was concluded that increased variation withingynogenetic or androgenetic offsprings is mainly theresult of embryonic damage, caused by the chromosomemanipulation treatment.
Parthenogenetic reproduction of animals which arehomozygous, i.e., animals from a gynogenetic or
Phenotypic variation ingynogenetic and androgeneticclones
genetically inactivated sperm to produce haploidembryos. These embryos are made diploid by eitherinhibition of the extrusion of the second polar body(2pb-gynogenesis) (Komen et al. 1988) or inhibition ofthe first cellular division (endomitosis) (Komen et al.1991). In the latter case, the result is a double haploid(= diploid) homozygous individual. In androgenesis,the same principle is applied but the maternal genomeis genetically inactivated and fertilization is performedwith intact sperm (Box 1; Bongers et al. 1994). Survivalis typically low: around 5-15 % of treated eggs developas viable fry, which necessitates rigorous screening ofthe progeny for true parthenogenesis (no contributionfrom the irradiated genome) and homozygosity. Toconfirm the parthenogenetic status of the fry,phenotypic markers can be used for which the dominantalleles are carried by the “irradiated genome” and therecessive alleles by the genome which will be reproducedby gyno or androgenesis. True parthenogenonts do notexpress dominant alleles. Segregation ratios of aphenotypic marker, for which the parent isheterozygous, can also be used as a simple first checkfor homozygosity of the parthenogenetic progeny. Inthe studies, S/s (scaled/mirror) was used as dominantmarker, and Bl/bl (wild type vs. blond, orange or yellowphenotypes) as a marker for segregationa . Homozygositycan also be determined by the use of DNA markers, or,in the case of clones, simply by skin or scaletransplantations (Komen et al 1990).
After androgenetic or gynogenetic reproduction, a largeexpansion of phenotypic variance is generally observed(Komen et al. 1992a). Within one homozygous family,this expansion is the result of increased environmental
a In many carp strains, B1 is still present as a tetraploid locus with duplicated alleles b11 and b12. Only the homozygous recessivegenotype b11/b11; b12/12 is colored (Komen et al. 1991)
Genetic and phenotypicvariation in gynogenetic andandrogenetic families
Box 1:Androgenesis in cyprinids is difficult since eggs are activatedthe moment they come into contact with aquatic solutions.To overcome this problem, the centre developed a techniqueto irradiate eggs from common carp with UV, by using anartificial ovarian fluid for irradiation medium (Bongers et al.1994). Eggs are shaken in this medium which ensures aneven irradiation and proper genetic inactivation of DNA. Eggsare irradiated to a total dose of 175 mJ/cm2. Diploidization isachieved with a 40°C heat shock for 2 min, given around thetime of the first mitotic metaphase, which is between 26 and32 min after activation (at 24°C).
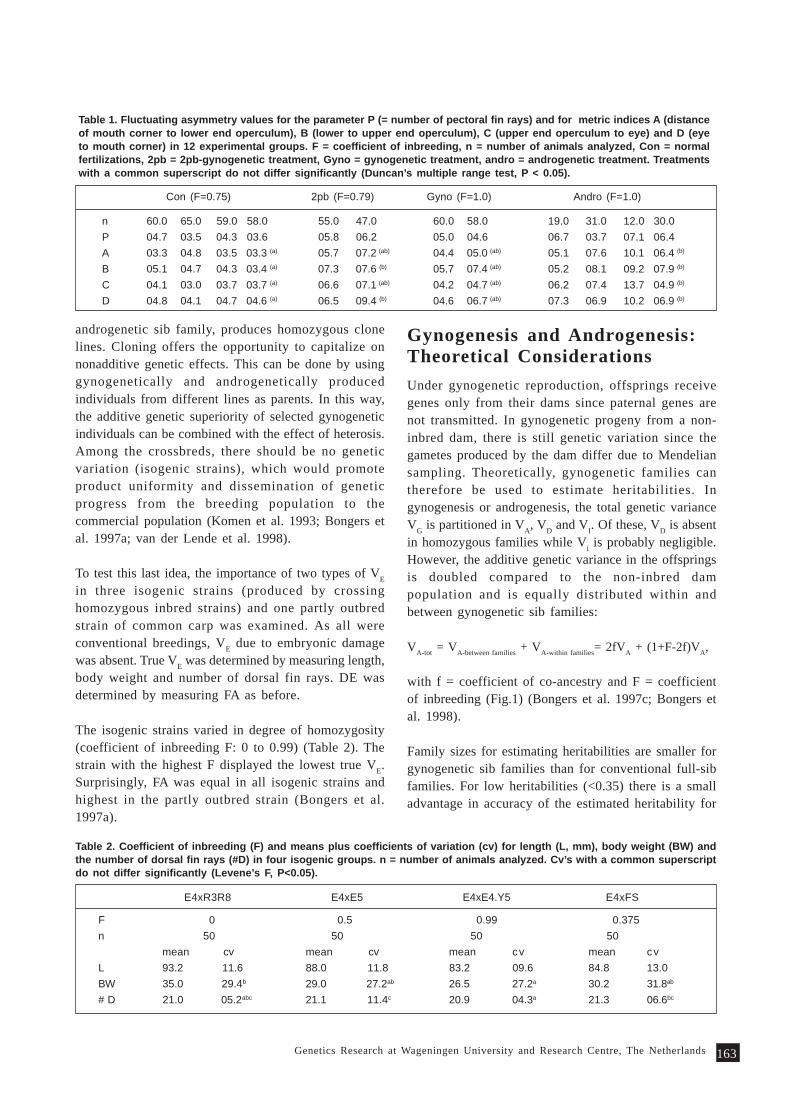
163
androgenetic sib family, produces homozygous clonelines. Cloning offers the opportunity to capitalize onnonadditive genetic effects. This can be done by usinggynogenetically and androgenetically producedindividuals from different lines as parents. In this way,the additive genetic superiority of selected gynogeneticindividuals can be combined with the effect of heterosis.Among the crossbreds, there should be no geneticvariation (isogenic strains), which would promoteproduct uniformity and dissemination of geneticprogress from the breeding population to thecommercial population (Komen et al. 1993; Bongers etal. 1997a; van der Lende et al. 1998).
To test this last idea, the importance of two types of VE
in three isogenic strains (produced by crossinghomozygous inbred strains) and one partly outbredstrain of common carp was examined. As all wereconventional breedings, VE due to embryonic damagewas absent. True VE was determined by measuring length,body weight and number of dorsal fin rays. DE wasdetermined by measuring FA as before.
The isogenic strains varied in degree of homozygosity(coefficient of inbreeding F: 0 to 0.99) (Table 2). Thestrain with the highest F displayed the lowest true VE.Surprisingly, FA was equal in all isogenic strains andhighest in the partly outbred strain (Bongers et al.1997a).
Under gynogenetic reproduction, offsprings receivegenes only from their dams since paternal genes arenot transmitted. In gynogenetic progeny from a non-inbred dam, there is still genetic variation since thegametes produced by the dam differ due to Mendeliansampling. Theoretically, gynogenetic families cantherefore be used to estimate heritabilities. Ingynogenesis or androgenesis, the total genetic varianceVG is partitioned in VA, VD and VI. Of these, VD is absentin homozygous families while Vi is probably negligible.However, the additive genetic variance in the offspringsis doubled compared to the non-inbred dampopulation and is equally distributed within andbetween gynogenetic sib families:
VA-tot = VA-between families + VA-within families= 2fVA + (1+F-2f)VA,
with f = coefficient of co-ancestry and F = coefficientof inbreeding (Fig.1) (Bongers et al. 1997c; Bongers etal. 1998).
Family sizes for estimating heritabilities are smaller forgynogenetic sib families than for conventional full-sibfamilies. For low heritabilities (<0.35) there is a smalladvantage in accuracy of the estimated heritability for
Genetics Research at Wageningen University and Research Centre, The Netherlands
Gynogenesis and Androgenesis:Theoretical Considerations
Table 1. Fluctuating asymmetry values for the parameter P (= number of pectoral fin rays) and for metric indices A (distanceof mouth corner to lower end operculum), B (lower to upper end operculum), C (upper end operculum to eye) and D (eyeto mouth corner) in 12 experimental groups. F = coefficient of inbreeding, n = number of animals analyzed, Con = normalfertilizations, 2pb = 2pb-gynogenetic treatment, Gyno = gynogenetic treatment, andro = androgenetic treatment. Treatmentswith a common superscript do not differ significantly (Duncan’s multiple range test, P < 0.05).
Con (F=0.75) 2pb (F=0.79) Gyno (F=1.0) Andro (F=1.0)
n 60.0 65.0 59.0 58.0 55.0 47.0 60.0 58.0 19.0 31.0 12.0 30.0P 04.7 03.5 04.3 03.6 05.8 06.2 05.0 04.6 06.7 03.7 07.1 06.4A 03.3 04.8 03.5 03.3 (a) 05.7 07.2 (ab) 04.4 05.0 (ab) 05.1 07.6 10.1 06.4 (b)
B 05.1 04.7 04.3 03.4 (a) 07.3 07.6 (b) 05.7 07.4 (ab) 05.2 08.1 09.2 07.9 (b)
C 04.1 03.0 03.7 03.7 (a) 06.6 07.1 (ab) 04.2 04.7 (ab) 06.2 07.4 13.7 04.9 (b)
D 04.8 04.1 04.7 04.6 (a) 06.5 09.4 (b) 04.6 06.7 (ab) 07.3 06.9 10.2 06.9 (b)
Table 2. Coefficient of inbreeding (F) and means plus coefficients of variation (cv) for length (L, mm), body weight (BW) andthe number of dorsal fin rays (#D) in four isogenic groups. n = number of animals analyzed. Cv’s with a common superscriptdo not differ significantly (Levene’s F, P<0.05).
E4xR3R8 E4xE5 E4xE4.Y5 E4xFS
F 0 0.5 0.99 0.375n 50 50 50 50
mean cv mean cv mean cv mean cvL 93.2 11.6 88.0 11.8 83.2 09.6 84.8 13.0BW 35.0 29.4b 29.0 27.2ab 26.5 27.2a 30.2 31.8ab
# D 21.0 05.2abc 21.1 11.4c 20.9 04.3a 21.3 06.6bc

Fish Genetics Research in Member Countries and Institutions of INGA164
gynogenetic sib families, but with higher heritabilitiesthere is a clear disadvantage (Bongers et al. 1997c).
In 1996, a project was started to investigate whetherselection for a stress response in common carp isfeasible, and how heritabilities can be estimated usingandrogenetic progenies. The starting population forthis study was an F1 hybrid cross between a domesticatedcarp strain from Wageningen and 6 males from a feralcarp strain caught in a Dutch lake (Fig. 2) (Vandenputteet al., in press). The stressor used within the selectionprogram was a simple cold shock of 9°C for 3 hours(Tanck et al. 2000). A first generation of 33 androgeneticfamilies was produced using the F1 hybrids and thesefamilies were subjected to a cold shock and blood wassampled 20 min after onset of the shock. Furthermore,all individual fish were tested for homozygosity using apanel of 11 microsatellite markers.
Heritability estimates on a group of 512 homozygousanimals ranged from 0.60 ± 0.16 for cortisol responseto 0.09 ± 0.05 for weight at 12 months. From these 33families, 12 potential high and low responderhomozygous males were selected for furtherandrogenetic reproduction. The progenies from thesemales are expected to be homozygous clones andgenetically identical to their respective “fathers”.
In 1998, new clones of common carp were producedfrom the 12 homozygous males which were selected for
a high or low cortisol response during a temperatureshock. From the 12 homozygous clones, four wereselected, which descended from the twosires with the lowest and the two sires with thehighest estimated breeding values. These clones weregiven a standard cold shock of 3 hours to induce acortisol response and again sampled 20 min afteronset of the shock (Komen et al. 2000). The aim was to
350
300
250
200
150
100
50
0
(ng/
ml)
Fig. 3. Plasma cortisol levels in four clones of carp, derivedfrom parents selected for a high or low cortisol responseafter a cold shock. Means (± SD) with similar superscriptsare not significantly different (Tukey’s multiple comparisonprocedure, P<0.05).
High1
a
High2
ab
Low2
bc
Low1
c
Fig. 2. Breeding strategy with androgenesis.
XFemale E4
Homozygous inbredstrain Wageningen
Males AP 1-6Wild population
Anna Paulowna Polder
33 families with a maximum of40 homozygous individuals
Heritability?
F1 panelInheritance?
12 homozygous inbred lineswith a high of low cortisol
stress responseConsistency?
Selection and androgenesis:6 families 2 males / family
AndrogenesisRandom 5 - 6 males per family
F1 : 6 Families (E4 x AP 1-6)Repeatability?
Selection for high and low stressresponders in androgeneticfamilies
Fig. 1. Determination of genetic relations in gynogeneticoffspring. Parents (P) are assumed not to be related. F1females are reproduced by homozygous gynogenesis toproduce homozygous gynogenetic offspring (G). F =coefficient of inbreeding; f = coefficient of co-ancestry; a =additive genetic relation; a= 2*f.
f=1/4 a=1/2
f=1/2 a=1
i,k j,l
ij; il; kj; kl ij; il; kj; kl
ii; ll kk; ll ii; jj kk; jj G; F=1
F1; F=0
P; not related
gynogenesis

165Genetics Research at Wageningen University and Research Centre, The Netherlands
see if, and to what extent, a cortisol response ofa homozygous clone reflects the phenotype of thesire for this trait (the genetic correlation is 1.) Indeed,high responder clones were significantly different fromlow ones (Fig. 3).
In breeding programs, clone lines (crossbred or pure)can be used to measure different traits on the samegenotype, for example, carcass quality at different ages,disease resistance, or gonadal growth (Bongers et al.1997a). Clone lines might therefore be very useful forestimating genetic correlations and genotype byenvironment interactions, because performance on thesame genotype can be observed in differentenvironments. Since clone lines are not subject togenetic change, they can be used as internal controllines in long-term breeding programs where estimationsof genetic progress are based by improvements inhusbandry (Vandenputte et al., in press). This isparticularly important for fish, where estimates ofgenotype by environment interactions can vary fromnegligible to considerable.
In 1998, a project was started to investigate the impactof various aquaculture practices on stress and health infish (van Weerd and Komen 1998). Preliminaryexperiments in which the effect of crowding at differentdensities (control 25 g l-1; crowded 50 and 100 g l-1)were examined, showed that common carp can adaptto fairly high densities of up to 100 g l-1, provided that
water quality conditions (particularly oxygen) areoptimal. An additional 1 hour net confinement stressorwas applied to both uncrowded and crowded fish as astress test.
Netting induced a high cortisol response, of amagnitude comparable to a temperature shock of 9°C(compare Figs. 3 and 4). Crowding and netting alsoinduced significant changes in blood glucose, lactateand free fatty acid levels.
The next step in this research will be to investigate theresponse to crowding and netting in carp strains whichhave been selected for high and low cortisol responseafter cold shock and strains selected for a high and lowantibody response to a synthetic antigen. The idea isto investigate whether:
• selection for one stressor (e.g., cold shock)conveys resistance to other stressors (positivecorrelation between stress responses) or
• a negative correlation between antibodyresponse as a trait and cortisol response, as atrait exists.
The common carp is a gonochorist species with XX/XY sex determination. Androgenetic progenies usuallyconsist of varying proportions of males, females andsterile fish. The males produce all male offspring whencrossed with normal females, and are therefore YY(Bongers et al. 1997b; Bongers et al. 1998). All maleprogenies are currently used in tests to screenenvironments on the presence of industrial pollutantswith estrogenic activity (Gimeno et al. 1996).
In gynogenetic progenies, males are often detected at
Determining genetic correlationsbetween traits in crosses betweenhomozygous clones of commoncarp
Clones, correlations and genotypeenvironment interaction
Sex determination and sexdifferentiation in common carp
Fig. 4(a). Changes in plasma cortisol levels during crowding in common carp over an 87-hour period. (b) Plasma cortisollevels in control and crowded common carp after a 1-hour net confinement, during the crowding period. *(p<0.05);***(p<0.001) denote significant differences to the controls.
crowd 50
control
crowd 100
con1
corti
sol (
ng/m
l)
15 39 con2 87
350
300
250
200
150
100
50
0
***
*
***
con1
corti
sol (
ng/m
l)
15 39 con2 87
9080706050403020100
***(a) (b)

Fish Genetics Research in Member Countries and Institutions of INGA166
high frequencies. The analysis of one such case showedthat XX animals, which are homozygous for a recessivemutation in a putative sex determining genemas(-culinization), undergo female to male sex reversal(Komen et al. 1992b; Komen et al. 1992c).
In order to identify the function of mas in sex deter-mination, the sex reversal of XX (mas/mas) common carpwas investigated by comparing clones of XY males,XX(mas/mas) males and XX(mas/+) females. Fish in eachof these clones are genetically uniform and of the same
sex. From this study, it was concluded that sex reversalin XX (mas/mas) animals might becaused by a precociousproduction of 11-oxygenated androgens, whichgradually overrule female sex differentiation by malesex reversal of female steroid producing cells. Basedon these findings, a new female to male sex reversalprotocol for common carp was designed, using 11 keto-androstenedion (Box 2).
Between 1996 and 1998, some 83 highly polymorphicpoly(CA) type microsatellite markers have beendeveloped in common carp (Crooijmans et al. 1997).Clones containing a (CA) repeat were isolated from acommon carp genomic library and sequenced. Thenumber of repeats found was high compared to othermammals, but comparable with other teleost fishes. Mostmicrosatellites are highly polymorph (average of 4.7alleles) with a few markers giving additional polymorphicamplification products. It is suspected that these lociare tetraploid. Of the 83 markers, 11 are currently usedas a panel to screen homozygous clones and to perform
pedigree analyses. The developed microsatellites appearto be species-specific. Cross hybridization with DNA ofIndian carp Catla catla gave no amplification.
One of the centre’s main activities for the near futurewill be the search for markers in common carp whichare linked to QTL. For this, an F1 hybrid cross wasproduced between an ornamental (koi) carp and adomesticated common carp, as these strains appear tobe genetically most distant. Koi and edible commoncarps differ in many traits, due to their domesticationhistory. Androgenetic and gynogenetic progenies willbe produced for direct AFLP mapping. The progeniesare expected to differ in a large number of single andmultilocus traits, i.e., scalation, color, sex determination,onset of maturity, fecundity, growth, etc., and it isexpected that markers will segregate with major genesaffecting these traits. In the end, such markers will bevery valuable for selective breeding programs withcommon carp anywhere in the world.
References
Bongers, A.B.J., M. Sukkel, G. Gort, J. Komen and C.J.J. Richter.1998. Clones of common carp in experimental animalresearch. Lab. Anim. 32: 1-15.
Bongers, A.B.J., M.Z. Ben-Ayed, B. Zandieh Doulabi, J. Komenand C.J.J. Richter. 1997a. Origin of variation in isogenic,gynogenetic and androgenetic strains of common carp,Cyprinus carpio, L. J. Exp. Zool. 277: 72-79.
Bongers, A.B.J., B. Zandieh Doulabi, P.K. Voorthuis, H. Bovenhuis,J. Komen and C.J.J. Richter. 1997b. Genetic analysis of testisdevelopment in all-male F1 hybrid srains of common carp,Cyprinus carpio, L. Aquaculture 158: 33-41.
Bongers, A.B.J., E in het Veld, K. Abo-hashema, I. Bremmer, E.Eding, J. Komen and C.J.J. Richter. 1994. Androgenesis incommon carp using UV irradiation in a synthetic ovarianfluid and heat shocks. Aquaculture 122: 119-132.
Bongers, A.B.J., H. Bovenhuis, A.C. van Stokkom, G.F. Wiegertjes,B. Zandieh Doulabi, J. Komen and C.J.J. Richter. 1997c.Distribution of genetic variance in gynogenetic orandrogenetic families. Aquaculture 153: 225-238.
Crooijmans, R.P.M.A., V.A.F. Bierbooms, J. Komen, J.J. van derPoel and M.A.M. Groenen. 1997. Microsatellite markers incommon carp (Cyprinus carpio, L.). Anim. Genet. 28: 129-134.
Gimeno, S., A. Gerritsen, T. Bowmer and J. Komen, Feminizationof male carp. 1996. Nature 384: 221-222.
Komen, J. and C.J.J. Richter. 1993. Sex control in common carp(Cyprinus carpio L.), p. 78-84. In Roberts and Muir (eds.)Recent advances in aquaculture. IV. Blackwell Science, UK.
Komen, J., M. Yamashita and Y. Nagahama. 1992a. Testiculardevelopment induced by a recessive mutation during gonadaldifferentiation of female common carp. Dev. Growth Differ.34: 535-544.
Komen, J., P. de Boer and C.J.J. Richter. 1992b. Male sex reversalin gynogenetic XX females of common carp (Cyprinus carpio,L.) by a recessive mutation in a sex determining gene. J.Hered. 83: 431-434.
Komen, J., E.H. Eding, A.B.J. Bongers and C.J.J. Richter. 1993.
Box 2:Methyltestosteron (MT) is the most widely used hormoneto induce sex reversal in cyprinids. However, its efficiencyis variable, and high incidences of sterility have beenreported (Komen et al. 1989; Komen and Richter 1993). Incommon carp, 11-keto androstenedion (KA) is the mostimportant steroid in male sex differentiation. The efficiencyof KA to induce functional sex reversal in common carpwas compared with 17a-MT. Two homozygous clones andfour F1 hybrids were treated with either 50 ppm 11 KA or 50ppm MT, incorporated in the pellet, for a period of 5 weeks,starting from 6 weeks post-hatch. While results with MTwere typically variable, with high numbers of intersexes insome clones and of sterile fish in others, KA induced a100% female to male sex reversal in all strains. The protocolfor sex reversal with KA is currently being tested in fieldconditions in cooperation with AIT, Thailand.
The Future: Microsatellite Markers andthe Search for QTL

167Genetics Research at Wageningen University and Research Centre, the Netherlands
Gynogenesis in common carp. IV: growth, phenotypicvariation and gonad differentiation in normal andmethyltestosterone treated homozygous clones and F1hybrids. Aquaculture 111: 271-280.
Komen, J., G. Wiegertjes, V. van Ginneken and C.J.J. Richter.1992c. Gynogenesis in common carp (Cyprinus carpio L.). III:the effects of inbreeding on gonad development inhomozygous and heterozygous gynogenetic offspring.Aquaculture 104: 51-66.
Komen, J., J. Duynhouwer, C.J.J. Richter and E.A. Huisman.1988. Gynogenesis in common carp (Cyprinus carpio L.). I:the effects of genetic manipulation of sexual products andincubation conditions of eggs. Aquaculture 69: 227-239.
Komen, J., G. Bongers, C.J.J. Richter, W.B. van Muiswinkel andE.A. Huisman. 1991. Gynogenesis in common carp (Cyprinuscarpio L.). II: the production of homozygous gynogeneticclones and F1 hybrids. Aquaculture 92: 127-142.
Komen, J., P.A.J. Lodder, F. Huskens, C.J.J. Richter and E.A.Huisman. 1989. Effects of oral administration of 17a-methyltestosterone and 17ß-estradiol on gonadaldevelopment in common carp (Cyprinus carpio L).Aquaculture 78: 349-363.
Tanck, M.W.T., G.H.R. Booms, E.H. Eding, S.E. Wendelaar Bongaand J. Komen. 2000. Cold shocks: a stressor for commoncarp. J. Fish Biol. 56.
van der Lende, J. Komen and E.W. Brascamp. 1998. The role ofreproductive techniques in genetic improvement anddissemination of genetic gain in livestock production andaquaculture. Tr. Acta Agric. Scand. Anim. Sci. (Suppl.) 29:90-97.
Vandenputte, E., Peignon, D. Vallod, P. Haffray, J. Komen and B.Chevassus. Use of an internal control in the comparison ofgrowth performances of three French strains of commoncarp at 5 weeks of age. (In press).
van Weerd, J.H. and J. Komen. 1998. The influence ofenvironmental stressors on fish growth. Comp. Physiol.Biochem. 120: 107-112.

Fish Genetics Research in Member Countries and Institutions of INGA168

169Species Index
AAcipenseridae 21 see Acipenser dabryanus,
Acipenser sinensisAcipenser dabryanus 21Acipenser sinensis 21Aeromonas hydrophila 81, 85Anabantidae 8, 87Anabas testudineus 8, 73, 87 see climbing perchAnguilla bicolor 56 see also eelAristichthys nobilis 9, 15-18, 21, 44, 81-82, 119, 144 see
bighead carpArtemia 152Artemia salina 33
BBarbs 8-9, 13barb, Java 52, 83barb, Silver 3, 8-9, 77, 80, 84, 100, 131, 146, 152, 154, 158
see Barbodes gonionotusbarb, Tiger 68-69Balantiocheilos melanopterus 66Bangus 72 see milkfishBarbodes altus 101Barbodes belleroides 101Barbodes daruphani 87Barbodes gonionotus 3-4, 7-12, 14, 52, 67,69, 77, 80-82, 84,
87, 91, 93, 95-96, 100-101, 130-131, 146-147, 158Barbodes orphoides 87Barbodes schwanenfeldii 87, 101Barbodes stoliczkanus 87bass, Striped 104Bata 8, 13 see Labeo bataBelontiidae 87 see Trichogaster PectoralisBetta splendens 84Bhangan 13Bighead carp 9, 15-18, 23, 44, 66, 119, 144 see
Aristichthys nobilisBlack porgy 19 see Sparus MacrocephalusBlunt snout bream 17-23, 100, 103 see Megalobrama
amblycephalaBream 100 see seabream and Megalobrama
amblycephalaBrigidae 8 see Mystus cavasius
CCalbasu 8 see Labeo calbasuCarassius auratus 15, 17, 20-21, 91, 93 see goldfishCarassius auratus gibeleo 17, 19, 191Carassius auratus var. pengzenensis 17Carassius carassius 104Carcharinus spp. 26Carps 1, 3, 7-8, 13, 20-21, 23, 29, 43-44, 49, 51, 58, 65-66,
91, 97, 100-101, 103, 105-107, 119, 151-153, 157, 161, 164see Cyprinidae
carps, Asian 119carps, Chinese 8, 15-18, 21-22, 29, 43-44carp, Black 9, 15-18, 23, 105 see also Mylopharyngodon
piceuscarp, Croatian Nasic 119carp, Common 3-4, 9, 15-22, 29, 43-45, 51-52, 66, 100, 103-
106, 119-121, 131, 141, 147, 151, 154, 157-158, 161-166see Cyprinus carpio
SPECIES INDEXcarp, Crucian 15, 17-20, 22, 104carp, Penzhe crucian 17, 22carp, Silver crucian 23carp, Xiangyuan crucian 19carp, Domai 51carp, Feng 18carp, Furong 18carp, Glass red 16, 18carp, Grass see 9, 15-20, 23, 43-44, 66,105
Ctenopharyngodon idellacarp, Heyuan 18carps, Indian major 43-45, 46-49, 147,154, 157 see
Catla catla Labeo rohita, Cirrhinus mrigalacarp, Japanese 51, 68carp, Javanese 66carp, Jian 18carp, Jing 15carp, Kaca 51carp, Kancra-domas 51carp, Kumpai 51carp, Majalaya 51, 58carp, Merah 51carp, Mirror 18-19, 20, 23, 51, 120
carp, Mirror Czech 119carp, Mud 22carp, Nilem 51carp, Ornamental 65-66, 68-69, 103-105, 107, 119, 166 see
koicarp, Poliana 119carp, Punten 51carp, Rajadanu 51, 58carp, Red common 15, 20, 22
carp, Red crucian 20carp, Red purse 16, 18, 22carp, Russian Ropsha 119carp, Scaly 119, 120carp, Scattered mirror 18carp, Sinyonya 51carp, Silver 9, 15-19, 23, 43-44, 119 see
Hypopthalmichthys molitrixcarp, Silver crucian 17, 19, 22 see
Carassius auratus gibeliocarp, Sutisna Kunginga 51, 58carp, Taiwan 51carp, Wildan Cianjur 51, 58carp, Wuyan red purse 22carp, Xingguo red 16, 18-20, 22-23carp, Yin common 18carp, Yue 18Catla 8, 13, 43-47, 100, 151, 154, 157 see also Catla catlaCatla catla 4, 7-8, 13, 43-49, 100, 145, 151, 157, 166Catfish see 7-8, 12, 33, 51-52, 55, 58, 65-67, 77, 80-81, 83-
86, 93, 109-115 Heteropneustes fossilis, Pangasius sp. andClarias sp.
catfish, African 9, 29, 33, 68, 161 see Clarias gariepinuscatfish, Asian 8 see Clarias batrachuscatfish, Blue 110-113 see Ictalurus furcatuscatfish, Channel 2, 30, 80, 110-114 see also Ictalurus
punctatuscatfish, Indian 49catfish, Walking 67catfish, River 69

Fish Genetics Research in Member Countries and Institutions of INGA170
Catlocarpio siamensis 87Ceratoglanis scleronema 66Cestraeus plicatilis 72Channa striata 8Chanos chanos 3, 53, 56, 59, 74, 130-131, 141Cherax 137Cherax destructor 137, 139Cherax quadricarinatus 137Cherax tenuimanos 137Chromileptes altivelis 56Chrysichthys nigrodigitatus 25-26Cichlidae (Cichlids) 21, 26, 64, 72 see also tilapiasCirrhinus 44Cirrhinus mrigala 3-4, 8, 44-46, 48, 130-131Cirrhinus reba 8, 13Clarias anguillaris 25-26Clarias batrachus 8, 49, 55, 67-68, 87Clarias gariepinus 9, 25-26, 33, 55, 67-68, 80, 83,-84, 161Clarias lazera 33Clarias leather 19Clarias macrocephalus 67-68, 73, 80-87, 91, 93-94, 96Clarias maurus 26Clarias mossambicus 33Clarias senagalensis 33Clarias sp. 33, 52, 55, 68, 81-83Clariidae 8, 58, 80, 87 see catfishClimbing perch 8 see Anabas testudineusclupeid, Anadromous 7Clupeidae 21Cockles 66crab, River 22Crassostrea belcheri 77, 79, 81, 85Crassostrea lugubris 81, 85crayfish, Freshwater 41, 133, 137-139Ctenopharyngodon idella 9, 15-18, 21, 44-45, 105 see
grass carpCyprinidae (Cyprinids) see Indian carps,Chinese carps 8-9, 13, 17, 21, 62, 72, 87, 162, 166Cyprinus carpio 3-4, 9, 15-16, 18-21, 44-45, 48, 51-52, 57-58,
91, 94-95, 100, 103-104, 107, 119, 145, 151, 157, 161 seecommon carpsCyprinus carpio var. Jian 18 see Jian carpCyprinus carpio var. singnonensis 16, 18 see
Xiangguo red common carpCyprinus carpio var. specularis 9 see mirror carpCyprinus carpio var. wananensis 16, 18 see glass red
carpCyprinus carpio var. wuyuanensis 16, 18 see redpurse common carpCyprinus carpio var. yuankiang 18
DDicentrarchus labrax 104-107, 145Discherondontus halei 66Distichodus rostratus 28Dreissena polymorpha 105 see also zebra mussel
EEel 52, 56Epihephelus aeneus 105-107Epihephelus bonthoides 56Epinephelus coioides 56Epinephelus fuscogutatus 56Epinephelus microdon 56Ephinephelus spp. 56
Eriocher sinensis 21Ethmalosa fimbriata 26
FFrog 77 see Rana rugulosa
GGhonia 13Ghora maach 13gilthead sea bream 33, 103-107 see Sparus auratusGonious 8 see Labeo goniusgourami, Snakeskin 77, 80, 83Grapsidae 21Groupers 19, 52, 56, 66, 107, 151, 153 see
Epinephelus spp.Goldfish 20-21, 48, 105grouper, White 105-106Gulsha 8Guppy 66, 68, 105
HHalibut, Atlantic 129, 146Helicophagus wandersii 66Helostomatidae 87Helostoma teminckii 87Heterobranchus bidorsalis 25-26Heterobranchus longifilis 25-26Heteropneustidae 8Heteropneustes fossilis 7-8, 12, 14, 45, 49Horabagus brachysoma 48Hilsa shad see Tenualosa ilisha 7-8, 13Hypopthalmichthys molitrix 9, 15-18, 21, 44, 119
IIctaluriidae (Ictalurids) 80, 112Ictalurus furcatus 110Ictalurus punctatus see Channel catfish 80, 110, 112
KKalbaus 13Kapenta 151, 153Kelp 22 see Laminaria, PorphyraKoi 51, 66, 68, 105, 107, 119, 166 see ornamental carpKrytopterus bicirrhis 87Krytopterus bleekeri 87Kryptopterus cryptopterus 87
LLaachu, bata 13Labeo sp. 44Labeo bata 8, 13Labeo bicolor 87Labeo boga 13Labeo calbasu 8, 13, 44-45Labeo coubie 28Labeo dussumieri 48Labeo erythrurus 87Labeo fimbriatus 44, 48Labeo gonius 8, 13Labeo mesops 62Labeo nandina 13Labeo pangusia 13Labeo rohita see Rohu 3-4, 7-9, 13, 43-49, 81-82, 87,100,Lates niloticus 28

171
Limnothrissa miodon 151Lithognathus mormyrus 33Loach 19Luciosoma trinema 66Ludong 72 see Cestraeus plicatilis
Valamugil seheliLutjanus argentimaculatus 86
MMacrobrachium rosenbergii 52, 56-57, 66, 68, 77, 79, 83, 91,
93, 96Macrognathus aculeatus 87Macrobrachium australiense 139Macrobrachium nipponense 145Macrognathus cicumcinctus 87Macrognathus siamensis 87Mahashol 13Mahseer see Tor putitora 9, 48Mahseer, Deccan 48Marcusenius furcidens 26Macrura reevesi 21Marcusenius ussheri 26Marron 137 see Cherax tenuimanosMastacembelidae 87Mastacembelus favus 87Mastamcembelus erythrotaemia 87Mastamcembelus armatus 87Megalobrama amblycephala 3-4, 17, 20-21, 100 see bluntsnout bream
Mesopristes cancellatus 72Milkfish 3, 52, 56, 72, 74-75, 131,141 see Chanos chanosMisgurnus anguillicaudatus 19-20, 73, 91, 93-95Moina sp. 84Molluscs 152Morone chrysops 105Morone saxatilis 104Morone sp. 105Moruluis chrysophekadion 87Mrigal 3, 8, 44, 47, 131 see Cirrhinus mrigalaMugilidae 33Mugil cephalus 33, 107Mullet 29, 107 see Mugil cephalusMussels 66mussel, Zebra see Dreissena polymorpha 105Mylopharyngodon piceus see black carp 9, 15-18, 105Mystus cavasius 8Mystus numerus 69Myxocyprinus Asiaticus 21
NNandina 13Nile tilapia 1-3, 7-9, 13-14, 17-19, 22, 29-30, 33, 35, 51-52, 55-
56, 58, 71, 83, 97-98, 114, 131, 142-143, 153 seeOreochromis niloticus
Notopteridae 87Notopterus blanci 87Notopterus chitala 87Notopterus notopterus 87Nyasalapia sp. 61-62
OOmpok bimaculatus 87Ompok pabda see Pabda 8Oncorhynchus mykiss 19, 48 see rainbow trout
Opsaridium microleps 62Oreochromis aureus 4, 17-19, 21, 25-26, 29, 31, 66-67, 81, 91-
92, 99, 100, 104-107, 153 see blue tilapiaOrnamental carp 65-66, 68-69, 103-105, 107, 119, 166 see
koiOsteochilus hasseltiOsteochilus melanopleuraSarotherodon galileausOreochromis lidole 61Oreochromis karongae 61-63Oreochromis macrocephalusOreochromis macrochir 62Oreochromis mossambicus 9, 19, 31, 35, 52, 62-63, 65-66,
67-68, 74-75, 83, 89, 98, 104, 106, 133-137, 139, 142, 151,153-154, 156
Oreochromis niloticus see Nile tilapia 2-4, 7-9, 13-14, 17-19,21, 25-29, 31-40, 51-52, 55-59, 61, 66-69, 71-75, 81, 83-84,91-92, 95-100, 103-106, 114, 130-131, 133-137, 139, 142-143, 146, 151, 153-154, 156-157
Oreochromis Nyasalapia sp. 61-63Oreochromis Ny. karongae 61-63Oreochromis Ny. Lowuwiala 62Oreochromis Ny.lidole 61-63Oreochromis Ny. saka 62Oreochromis Ny.squamipinnis 61-63
Oreochromis Oreochromis 62Oreochromis placidus 62-63Oreochromis rovumae 62Oreochromis spp. 61-62, 66, 68, 103, 153Oreochromis rukwaensis 62Oreochromis shiranus sp. 4, 62-64Oreochromis shiranus chilwae 62-63Oreochromis shiranus shiranus 62-63Oreochromis spilurus 81Osteochilus hasselti 52, 87Osphronemidae 87Osphronemus goramy 87Osteochilus melanopleura 87Oysters 77, 79, 81, 85-86, 129 see Saccostrea cucullata Crassostrea belcheri and Crassostrea lugubrisoyster, Mangrove 85
PPabda 8 see Ompok pabdaPagrus major 104, 106Pangasiidae (Pangasiids) 55, 58, 80, 87 see Pangasius
larnaudiiPangasius gigas 80-81Pangasius hypopthalmus 55, 80-81Pangasius jambal 59Pangasius larnaudii 87Pangasius nasutus 59Pangasius sp. 52, 55, 59, 82-83Pangasius sutchi 9, 81-82, 84-85, 87 see Thai pangasParachana obscura 28Parachela maculicauda 66Paralaubuca riveroi 87Paramisgurnus dabryanus 20Paramisgurnus sp. 20Penaeids 137Penaeus sp. 147Penaeus indicus 84Penaeus merguiensis 66, 77, 79Penaeus monodon 57, 59, 66, 77-79, 141, 143-145

Fish Genetics Research in Member Countries and Institutions of INGA172
Penaeus schwanenfeldii 67Penaeus semisulcatus 107perch, Climbing 8Phallotethus dunckeri 66Pigek 72 see Mesopristes cancellatusPoecilia reticulata 105Pomacea sp. 105 see golden snailPrawns 72, 79, 93prawn, Freshwater 56, 68-69, 77, 79, 81, 83, 139 see
Macrobrachium rosenbergiiprawn, Tiger 66, 72, 141Probarbus jullieni 65-66, 87Puntioplites proctozysron 87Puntius gonionotus 52, 158Puntius sarana 8-9, 13Puntius sirang 72Puntius tetrazona 68Puntius ticto 13
RRainbow trout 2, 19-20, 30, 48, 129, 131, 141, 146 see
Oncorhynchus mykissRana rugulosa 77Rasbora sumatrana 87Redclaw crayfish 133, 137-138 see crayfish, Freshwater
and Cherax quadricarinatusRhodeus spp 20Rohu 3-4, 7-9, 13, 43-44, 47, 100, 131 see Labeo rohita
SSaccostrea cucullata 81-82, 85Salmonids 130, 145-146, 152Salmon 2, 20, 129, 131, 152-153
Salmon, Chinook 156Salmon, Atlantic 2, 129, 131, 146Salmon, Coho 2, 131
Salmo salar 145Salmo trutta 48Sardinella aurita 26Sardinella maderensis 26Sardinella tawilis 72Sarotherodon galilaeus 29, 62, 99, 104Sarotherodon hornorum 67Sarotherodon melanotheron 25-26, 74, 101Sarotherodon mossambica 67Sarpunti 13Scallops 129Scleropages formosus 66Seabass 66, 74, 103-104, 106 see Dicentrarchus labraxseabass, European 104-106seabass, Striped 104 see Morone saxatilisseabass, White 105Seabream 103seabream, Gilthead 103-107seabream, Red 104Seaweeds 72Siamese fighting fish 84Scleropages formosus 65-66shad, Hilsa 7-8, 13 see Tenualosa ilishashrimp 22, 52, 57, 66, 72, 75, 78, 91, 103, 107, 125-126, 131,
137, 143-144, 151-153
shrimp, Banana 77, 79shrimp, Black tiger 77, 143 see Penaeus monodonshrimp, Marine 84, 107Siluridae 8, 87 see SiluriformesSiluriformes (Siluroidei) 25-26, 55 see Clarias gariepinus
Clarias anguillaris, Heterobranchuslongifilis, Heterobranchus bidorsalisand Chrysichthys nigrodigitatus
Silver barb see 3, 8-9, 77, 80, 84, 100, 131, 146, 152, 154,158 barb, Silver and Barbodes gonionotus
Silver carp see 9, 15-19, 23, 43-44, 119 carp, Silver andHypopthalmichthys Molitrix
snail, Golden 105 see Pomacea sp.Snapper 66snapper, Red 66, 86Sparidae 33, 104 see Sparus auratus Lithognathus
mormyrusSparus aurata 103-107Sparus auratus 33, 103 see gilthead seabreamSparus macrocephalus 19 see black porgySunsetplaty 84 see Xiphophorus variatus
TTawilis 72 see Sardinella tawilisTenualosa ilisha 7-8, 13-14, 48 see Hilsa shadThai pangas 9 see Pangasius sutchiTilapias 2, 4, 17-18, 25-26, 28-29, 31, 35-36, 56, 59-63, 64,
67-68, 73-75, 81, 83, 98-100, 103-105, 133, 136, 152-153see Sarotherodon melanotheron, Oreochromis niloticus,and Oreochromis aureus
tilapia, Black-chinned 101tilapia, Blue 17-19, 21, 29 see Oreochromis aureustilapia, Chitralada 35-40, 55-56, 66, 92, 133-134tilapia, Green 29 see Tilapia zilliitilapia, Mozambique 52, 98tilapia, Red 4, 9, 19, 35, 56, 66, 68, 104, 142-143tilapia, White 29 see Sarotherodon galilaeusTilapia rendallii 62-63Tilapia spp. 2-4, 28-30, 33, 43, 59, 61-63, 65-68, 71-73, 74-75,
77, 81, 84, 92, 95, 97-101, 103, 104-107, 109-110, 114-115,124, 130-135, 138-139, 141-146, 151-158, 158
Tilapia zilii 29, 99Tit punti 13Trichogaster pectoralis 66, 73, 77, 80, 83, 87Trichogaster trichopterus 73Trionyx sinensis 21Tor khudree 48Tor masher 8 see Tor putitoraTor putitora 8-9, 13, 48Tor soro 87Tor tor 13trout, Rainbow 2, 19-20, 30, 48, 129, 131, 141, 146turtle, Soft-shelled 22Turtle, soft-shelled 22
VValamugil sehelis 72 see Ludong
WWallago attu 87Wallago dinema 87Wallago miostoma 87

173
XXiphophorus variatus 84
YYabby 137 see Cherax destructor
ZZebrafish 115Zicai 22

Fish Genetics Research in Member Countries and Institutions of INGA174

175
List of Participantsa
Austral iaDr. P. MatherSchool of Natural Resources SciencesFaculty of Science, Queensland University of TechnologyGPO Box 2434, Brisbane Q 4001Tel.: (61-7) 3864-3142Fax: (61-7) 3864-3529E-mail: [email protected]
B a n g l a d e s hDr. M. G. HussainBangladesh Fisheries Research InstituteMymensingh 2201Tel.: (880-91) 54874; 54410Fax: (880-91) 55259E-mail: [email protected]
C h i n aProf. Li SifaLaboratory of Aquatic Genetic ResourcesShanghai Fisheries University334 Jungong RoadShanghai, 200090Tel.: (86-21) 6571-0333Fax: (86-21) 6568-4153E-mail: [email protected]
Côte d’IvoireDr. V. Yapi-GnaorePisciculture StationCentre National de Recherche Agronomique01 B.P., 633 Bouake 01Tel.: (225) 633-526; 633-139Fax: (225) 633-126
E g y p tDr. H. ElghobashyCentral Laboratory of Aquaculture ResearchNational Aquaculture CenterAbbassa, Abou Hammad, SharkiaTel.: (20-55) 400-497; 401-027Fax: (20-55) 400-498
FijiMr. S. Nand LalFisheries DivisionMinistry of Agriculture, Fisheries and ForestryNaduruloulou Research StationP.O. Box 819, NausoriTel.: (679) 477-781Fax: (679) 361-184
H u n g a r yDr. L. VaradiFish Culture Research InstituteP.O. Box 47, H-5541, SzarvasTel.: (36-66) 312-311Fax: (36-66) 312-142E-mail: [email protected]
Mr. S. Gorda
Carp Breeding ProgrammeFish Culture Research InstituteP.O. Box 47, H-5541, Szarvas
Mrs. T. PederiCarp Breeding ProgrammeFish Culture Research InstituteP.O. Box 47, H-5541, Szarvas
I n d i aDr. S. AyyappanCentral Institute for Freshwater AquacultureP.O., Kausalyaganga, BubhaneswarOrissa 751 002Tel.: (91-674) 465-421; 465-446Fax: (91-674) 465-407E-mail: [email protected]
Dr. P.V.G.K. ReddyCentral Institute of Freshwater AquacultureKausalyanganga, Bhubaneswar-751 002Orissa 751 002Tel.: (91-674) 465-421; 465-446Fax: (91-674) 465-407E-mail: [email protected]
I n d o n e s i aDr. A. HardjamuliaResearch Institute for Freshwater FisheriesJln. Sempur No. 1 P.O. Box 150Bogor 16154, Jawa BaratTel.: (62-251) 313200, 327890Fax: (62-251) 327890E-mail: riff@indonet,id
Ita lyDr. D. BartleyFisheries DepartmentFood and Agriculture Organizationof the United NationsViale delle Terme di Caracalla00100, RomeTel.: (39-06) 5705-4376Fax (39-06) 5705-3020E-mail: [email protected]
MalawiDr. A. AmbaliBiology DepartmentChancellor CollegeUniversity of MalawiPO Box 280, ZombaTel.: (265) 522-222Fax: (265) 522-046; 522-523E-mail: [email protected]
List of Participants
a Affiliations of participants as of March 1999, at the time of the meeting.

Fish Genetics Research in Member Countries and Institutions of INGA176
MalaysiaProf. Dr. T.K. MukherjeeProfessor of Genetics and Deputy DeanInstitute of Advanced StudiesUniversity of Malaya50603 Kuala LumpurTel.: (03) 759-4602/4603/4604Fax: (03) 756-8940/759-4606E-mail: [email protected]
Dr. O.S. SelvarajInstitute of Postgraduate Studies andResearch, University of Malaya50603, Kuala LumpurTel.: (60-3) 759-4513Fax: (60-3) 756-8940E-mail: [email protected]
Dr. K. EmbongInstitute of Biological SciencesUniversity of Malaya50603 Kuala LumpurTel.: (60-3) 759-5834Fax: (60-3) 759-5908E-mail: [email protected]
Mr. Hambal Haji HanafiDirectorFreshwater Fisheries Research CentreBatu Berendam, 75350 MelakaTel.: (60-3) 317-2485Fax: (60-3) 317-5705E-mail: [email protected]
Mrs. N. SahidinResearch Officer - BiotechnologyFreshwater Fisheries Research CenterBatu Berendam, 75350 MelakaTel.: (60-6) 317-1131Fax: (60-6) 317-5705E-mail: [email protected]
Prof. Tan Soon GuanProfessor of GeneticsBiology DepartmentUniversiti Putra Malaysia43400 SerdangTel.: (60-3) 948-6101 ext. 3909Fax: (60-3) 943-2508E-mail: [email protected]
Dr. Siraj Siti ShaporDepartment of Biology and EnvironmentUniversiti Putra Malaysia43400 SerdangTel.: (60-3) 948-6101 ext. 3724E-mail: [email protected]
N e t h e r l a n d sDr. J.H. KomenFish Culture and Fisheries GroupWageningen Agricultural UniversityPO Box 338, NL-6700 AH WageningenThe NetherlandsTel.: (31-317) 482465Fax: (31-317) 483937E-mail: [email protected]
N o r w a yDr. T. SvealvInstitute of Aquaculture Research Ltd.(AKVAFORSK)Box 5010, N-11432, AasTel.: (47-64) 949-500Fax: (47-64) 949 502E-mail: [email protected]
Dr. T. GjedremInstitute of Aquaculture Research Ltd. AKVAFORSKP.O. Box 5010, N-1432 AasTel.: (47-64) 949-500Fax: (47-64) 949 502E-mail: [email protected]
P h i l i p p i n e sDr. A. CamachoBureau of Fisheries and Aquatic ResourcesArcadia Building, 860 Quezon AvenueQuezon City 3008Tel.: (63-2) 372 5057Fax: (63-2) 372 5048
Dr. Z. BasiaoBinangonan Freshwater StationSoutheast Asian Fisheries Development CenterBinangonan, RizalTel.: (63-912) 353-3906Fax: (63-912) 652-0077E-mail: [email protected]
T h a i l a n dDr. N. PongthanaNational Aquaculture Genetics Research InstitutePathumthani 12120Tel.: (66-2) 577-5058 to 59Fax: (66-2) 577-5062E-mail: [email protected]
U KProf. J.A. BeardmoreSchool of Biological SciencesUniversity of Wales, SwanseaSingleton Park, Swansea SA2 8PP WalesTel.: (44-1792) 295-382Fax: (44-1792) 513-030E-mail: [email protected]
Dr. G. C. Mairc/o Aquaculture and Aquatic Resources ManagementAsian Institute of TechnologyP.O. Box 4, Klong LuangPathumthani 12120Tel./fax: (66-2) 524-5463E-mail: [email protected]
U S ADr. Zhanjiang LiuDepartment of Fisheries and Allied SciencesAuburn UniversityAlabama 36849-5419Tel.: (1-334) 844-4054Fax: (1-334) 844-9208E-mail: [email protected]

177
V i e t n a mDr. Tran Mai ThienResearch Institute for Aquaculture No. 1Dinh Bang, Tien SonBac NinhTel.: (84-48) 273-072Fax: (84-48) 273-070E-mail: [email protected]
ICLARM-the World Fish CenterDr. P. R. GardinerP. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
Dr. M. V. GuptaP. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
Dr. R. Dunhama
P. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
Dr. R. S.V. Pullinb
P. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
Dr. M. M. DeyP. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
Ms. B. O. AcostaP. O. Box 500, GPO10670, Penang, Malaysia.E-mail: [email protected]
List of Participants
a Current address:Department of Fisheries and Allied Aquaculture,Auburn University, Alabama 36848, USA
b Unit 7A, Legaspi Park View134 Legaspi Street, Makati, Metro ManilaPhilippines

Fish Genetics Research in Member Countries and Institutions of INGA178

179 Participants of the Fifth INGA Steering Committee Meeting, 3-5 March 1999, Kuala Lumpur, Malaysia.





















