
Diversity, Distribution and Effects on Cassava Cultivars of Cassava Brown Streak Viruses in Malawi

Ann. appl. B id . (1 997), 131:259-27 1 Printed in Great Britain
Factors determining recovery and reversion in mosaic- affected African cassava mosaic virus resistant cassava
259
By R W GIBSON* and G W OTIM-NAPE Namulonge Agricultural and Animal Research Institute, P 0 Box 7084, Kampala,
Uganda
(Accepted I August 1997)
Summary The severity and persistence of symptoms of mosaic virus disease were
monitored during the first six months of two growing seasons in cassava of the African cassava mosaic virus (ACMV)-resistant cv. TMS 30572 either inoculated by grafting with a mild or severe strain or infected from the planted cutting. Symptomless shoots developed between January and March 1995 in two field trials differing in age by c. 6 months; this recovery occurred during particularly hot weather. Recovery was often only temporary in the plants inoculated with the severe strain and occurred later compared with those inoculated with the mild. In 1996, the weather was cooler and recovery that year was delayed until flowering, c. 7 months after planting, when recovered shoots were often produced from buds in the axils of symptomless leaves produced amongst diseased leaves. Most cuttings taken from the upper parts of diseased plants produced symptomless (reverted) progenies whereas most cuttings taken from the base of diseased plants produced diseased progenies. Reversion seemed to be associated with the recovery that had already occurred in the upper stems of the parent plants.
Key words: Cassava; African cassava mosaic virus, ACMV, recovery, reversion, weather, temperature, Uganda, Africa
Introduction In Africa, mosaic disease (MD) of cassava is caused by at least two whitefly-borne
geminiviruses, namely African cassava mosaic virus (ACMV: family Geminiviridae) and, in coastal East Africa, East African cassava mosaic virus (EACMV: family Geminiviridae) (Swanson & Harrison, 1994), and is amongst the most important biotic constraints to cassava production there (Geddes, 1990). Virus isolates may differ in severity (Storey & Nichols, 1938) and the epidemic of MD currently devastating Ugandan crops of cassava (Otim-Nape, Bua & Baguma, 1994) is associated with unusually severe symptoms (Gibson, Legg & Otim- Nape, 1996) apparently caused by a geminivirus with properties of both ACMV and EACMV (Zhou et al., 1997). The whitefly now known as Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) is the only known vector (Kufferath & Ghesquiere, 1932; Dubern, 1994).
ACMV and EACMV are disseminated in cuttings taken from diseased cassava plants for use as planting material, and this can be an important means by which the viruses are
*Address for correspondence: Natural Resources Institute, University of Greenwich, Central Avenue, Chatham Maritime, Kent ME4 4TB, UK 0 1997 Association of Applied Biologists

260 R W GIBSON AND G W OTIM-NAPE
maintained in farming systems (Bock & Guthrie, 1982: Bock, 1994). A characteristic of MD- resistant cassava cultivars is that diseased plants often become symptomless (Jennings, 1957). Similarly, cuttings taken from infected plants may produce symptomless progenies (Storey & Nichols, 1938). again especially in resistant cultivars (Jennings, 1960). Such progeny plants are probably virus-free (Pacumbaba. 1985). For clarity, the term “recovery” is used to describe the production of symptomless shoots on initially diseased plants and the term “reversion“ to describe the production of symptomless progenies from cuttings derived from diseased plants. although the two terms have been used synonymously for both. Reversion is important in preventing the gradual build-up of infection in cassava stocks (Fargette, Thresh &L Otim-Nape. 1994; Fargette & Vie. 1995) which otherwise can occur in crops maintained by repeated cycles of vegetative propagation (Hollings, 1965).
Both recovery and reversion have been linked to ACMV being incompletely systemic in cassava (Rossel, Thottappilly, van Lent & Huttinga, 1987: Rossel, Changa & Atiri, 1994; Njock, Atiri, Thottappilly & Hahn, 1994: Fargette, Colon, Bouveau & Fauquet, 1996). Kecover) has also been linked to hot dry weather and simulated by a period of high temperature (Gibson. 1994). Natural recovery from virus disease has been observed in other crops in the tropics and sub-tropics: in Australia recovery of subterranean clover from subterranean clover virus (Wroth & Jones, 1992), in India recovery of potatoes from potato leafroll luteovirus (Thirumalachar, 1954) and in California recovery of strawberries from wawben-y crinkle and strawberry mottle viruses (Frazier, Voth & Bringhurst, 1965). Knowledge of the cause(s) of reversion and/or recovery in cassava may show how these phenomena can be enhanced and so improve the health of the crop. The present work investigates the effects of virus strain, seasonal changes in weather and plant development on rates of recovery and reversion in cassava.
Materials and Methods All trials were done in fields at Namulonge Agricultural and Animal Production Research
!iistitute (NAARI) Farm, Mpigi District, Uganda. NAARI is located c. 20 km north of Kampala. Maximum and minimum air temperatures were recorded daily at the Institute meteorological station using a screened tnaximum and minimum mercury thermometer. Rainfall was also recorded daily. An ACMV-resistant cassava clone bred by the Institute of Tropical Agriculture (IITA). coded by IITA as Tropical Manihot Series (TMS) 30572 (Rossel et 01.. 1994) and released in Uganda as the cultivar Migyera was used in all experiments. The kJgandan cl’. Bao was used as a susceptible check. Although some rain generally falls every month at Namulonge, there is a clear bimodal pattern of rainfall, with the first rains usually occurring mid-March to mid-May and the second rains occurring mid-September to mid- November: highest temperatures generally occur during the December to February dry season.
Scoring sxsteni for symptom severip The severity of MD was assessed only on diseased shoots (that is, any shoot to which MD
had not spread or which had recovered was ignored). Severity was assessed weekly in all experiments. Shoots were scored using a 1 4 (0 = symptomless) system, assessing diseased foliage as: 1 = mild mosaic present, no leaf distortion; 2 = slight leaf distortion; chlorosis affecting < c. 405% leaf area: 3 = severe leaf distortion: chlorosis affecting < c. 80% leaf area; 1 = leaves severely distorted and stunted. early leaf loss and 80-100% leaf area chlorotic. Recovery was considered to have occurred when at least five consecutive leaves on a shoot were symptomless.

Recovery and reversion in mosaic virus diseased cassava 26 1
Field Trial 1. The influence of severe and mild MD This trial, planted in May 1994, comprised 60 plants of symptomless TMS 30572 planted at
1 m spacing. In September 1994,20 of the plants were top-grafted with scions of the local cv. Kalitunsi from plants with the severe disease commonly found in the area of Uganda affected by the MD epidemic (Gibson et al., 1996) and obtained from farmers’ fields near Bombo, within the epidemic area. Twenty other plants were top-grafted with scions from mildly diseased plants typically found ahead of the epidemic and obtained from near Namulonge (then unaffected by the epidemic). The remaining 20 were de-topped as a mock-graft control. The grafts were applied in a random sequence. Occurrence and severity of disease on plants were recorded weekly from November 1994 to midJanuary 1995. Coloured sticky tapes were placed so as to encircle stems immediately below the first diseased leaf. Recovery was assessed on 25 January and on 28 March for every shoot which had been diseased: tapes of a different colour were used to mark where any recovery occurred. Shoots were re-examined on 23 June following the rains and the position of any further diseased leaves was again marked using tape. At the end of the trial, cuttings (c. 50 mm long) were taken from the following positions on each grafted plant which had become diseased:- (i) below the graft; (ii) above the graft but below the first diseased leaf; (iii) zones of the stem where leaves were diseased; (iv) recovered zones above the diseased zone. On de-topped controls, cuttings were taken from below the de-topping (as control for (i) above), the middle of the main stem (as control for (ii) and (iii) above) and the upper part of the stem (as control for (iv) above). Up to 50 cuttings were taken from each position on diseased and control plants depending on availability. Cuttings were planted in a whitefly-free screenhouse and the numbers of diseased and reverted progenies were recorded.
Field Trial 2. Seasonal changes in symptom expression This field trial sought to understand the effects of seasonal changes on severity of MD and
recovery. It was planted on 6 October 1994 and comprised a 4 x 4 array of square plots. Each plot was planted with 25 cuttings in a square at 1 m spacing; cuttings of either TMS 30572 or cv. Bao obtained from either diseased or symptomless plants were planted in each plot in a randomised block design. For both cultivars, cuttings from diseased plants were obtained from Nakasongola, a site affected by the epidemic. Cuttings from symptomless plants were obtained from NAARI Farm, an area then unaffected by the epidemic. The cuttings from diseased plants which produced symptomless (reverted ) plants were recorded one month after planting. Symptoms on freshly-developed foliage of one main shoot on each of the nine central plants in each plot were monitored weekly from November 1994 to May 1995, when canopies became too interlocked for records to be taken. Any reverted plant (that is; a plant that was symptomless from emergence although grown from a cutting obtained from a diseased plant) was substituted with a diseased plant selected from amongst the outer plants in that plot. The extent of recovery on every shoot of all diseased TMS 30572 plants was recorded on 5 April 1995 (towards the end of the hot dry season) and on 5 May 1995 (after the onset of the rains).
A characteristic of cassava is that the terminal shoot ultimately becomes an inflorescence, and buds in the axils of leaves immediately below the flower subsequently sprout to produce usually three or four new shoots. A further characteristic of cv. TMS 30572 is that a mixture of diseased and symptomless leaves can appear consecutively along a stem. During June when flowering was profuse, counts were made on diseased stems of the numbers of diseased and symptomless shoots which had grown from the axils of diseased and symptomless leaves. Yields of tuberous storage roots were taken from the central nine plants in each plot one year after planting. One two-node cutting was also taken from the top, the middle and the bottom

262 R W GIBSON AND G W OTIM-NAPE
Table 1 . The niirnbers of p1Liiits cf TMS 30572 bvhich developed particular rnaximuin disease severities after being grajted tvith a scion froin either a severely or u mildly diseasedplant (96
of diseased plants in each categop are in parentheses)
Seierity 0 I 2 3 4 Total plants score ( N o diFea\e 1 inoculated
Sebere scion 10 0 (0%) 3 (30%) 7 (70%) 0 (0%) 20 Mild scion I I 3 (33% I 6 (66%) 0 (0%) 0 (0%) 20 Moch control 20 0 0 0 0 20
Table 2. The total numberr of diseased arid recovered shoots 011 plants graft-infected with either n .severel! or N mildl\ diseased scion 011 different dates in 1995
Date Grafted uith seLere scion Grafted with inild scion
RecoLered but Recovered but Diseased R e c o ~ ered re-diseased Diseased Recovered re-diseased
shoot\ $hoots shoot \ \hoot\ shoots shoots
2.S Jdilu<il! 48 (92cc) 4 (8%j 0 (0%) 3 (8%) 36 (92%) 0 (0%) 28 Mxch I0 (40%) 37 (60%) 0 (0%) 3 (8%) 36 (92%) 0 (0%) 23 June 3 (3%) 74 (7Y-C) 25 (24%) 0 (0%) 51 (100%) 0 (0%)
Table 3. The nuriibers of diseused progetlies (tiurnerator) amongst batches of cuttings taken ,from different positioris on grafted diseased plants
Severe graft Mild graft Mock graft
Below the graft 25/28 (89%) 25/25 (l007c) 0/36 (0%) .Above the graft but below the first diseased leaf 37/44 (84%) 26/33 (79%) 0/37 (0%) The zone of the diseased leaves 23/27 (85%) 19/31 (61’31 The recovered zone on stems which had recovered permanently 7/39 (18%) 0/17 (0%) The ineaii severity score of progenies
1/1 I (9%)
(Standard error of difference (df I61 I = 0.104 3.5 2.5 0
section of every main stem (often 3-4/plant) of all TMS 30572 plants. These cuttings were planted outside and the numbers of diseased and reverted progenies recorded.
Field Trial 3. Seasonal changes arid effects of \catering on syzptoiii expression In field trial 2, recovery occurred during hot dry conditions in January to March but
symptoms reappeared on some recovered plants when the first rains brought wetter, cooler conditions. These observations suggested that recovery may be caused by hot weather, dry weather or a combination of the two. In order to distinguish the underlying cause, 40 cuttings of TMS 30572 taken from diseased plants in field trial 2 were planted on 12 September 1995 in four rows each of 10 cuttings at 2 m x 2 m spacing. Cuttings which reverted were replaced by extra diseased plants grown for this purpose. Twenty plants were assigned at random each to be given 10 litres of water twice a week during the December-March dry season and 20 were left unwatered. However, the dry season was unusually wet and watering was done on only 1 0 occasions:- 5, 1 1 and 18 December, 16 January, 25 February and 4, 7, 1 1 , 15 and 18 March. Severity of MD was assessed weekly on a single main shoot on each plant from November 1995 to May 1996.

Recovery and reversion in mosaic virus diseased cassava 263
Statistical analyses Frequencies of plants or shoots either symptomless or with MD and with particular severity
scores were compared by the chi-square test using Yates’ correction as appropriate. Other data were compared by analyses of variance; means are given f standard error of their means.
Results
Field Trial 1. The injluence of severe and mild MD Poor graft unions led to infection of only 19 of the 40 plants of TMS 30572 grafted in
September 1994 with scions from diseased plants of cv. Kalitunsi. Those grafted with scions derived from severely diseased plants were more severely diseased than plants grafted with scions from a mildly-diseased plants ( P < 0.05; Table 1). The mosaic on the former plants was a very pale yellowish green, clearly demarcated from disease-free areas: leaves were also often distorted. On the latter plants, the mosaic was light green and diffuse; leaf distortion was rare. All mock-grafted plants remained symptomless. By March 1995, all shoots on diseased plants grafted with mildly diseased scions, and most shoots on diseased plants grafted with severely diseased scions, became symptomless although this recovery occurred much earlier on the plants infected with the mild disease than those infected with the severe disease ( P < 0.01; Table 2). With the onset of the rains in April, symptoms reappeared on some of the previously recovered shoots of plants infected with the severe disease and by 25 June about a quarter of these shoots had again developed symptoms. In contrast, none of the recovered shoots on plants grafted with the mildly diseased scions redeveloped symptoms ( P < 0.001: Table 2). Also, no symptoms developed on mock-grafted plants so the reappearance of disease was the result of a resurgence of the severe disease rather than the result of new whitefly- borne infections.
Progeny cuttings from the plants grafted with the severely diseased scions generally had more severe symptoms than ones from plants grafted with the mildly diseased scions (Table 3: P < 0.001). Most cuttings taken from the zone of the stem where diseased leaves had been observed, or below this zone, produced diseased progenies irrespective of whether the parent plants had been grafted with the mild or the severe disease. In contrast, most cuttings taken from upper zones where the shoot had recovered produced disease-free progenies (Table 3).
Field Trial 2. Seasonal changes in symptom expression TMS 30572
Fifteen (42%) of the 36 central plants in the four plots of TMS 30572 planted in October 1994 with cuttings from infected plants were symptomless (reverted) and were replaced for monitoring purposes by outer plants. On the remaining diseased plants, symptoms were relatively severe with few shoots scoring 2 (for example, at the end of November 1994, 7S% scored 3 or more) and the mosaic was clearly demarcated and almost white, typical of the severe disease in this cultivar. Only one of the 36 monitored main shoots on diseased plants recovered during November but the number of recovered shoots increased rapidly from the beginning of February to 23 (64%) by the end of March (Table 4; Fig. 1). Disease severity on remaining affected shoots then diminished during February and March ( P < 0.001) but increased once more ( P < 0.001) during April. This was followed in early May by several of the previously recovered monitored shoots (most of which had produced disease-free foliage for at least one month) developing mosaic symptoms on new foliage (Table 4). During this period, none of the monitored main shoots on the plants grown from cuttings obtained from mosaic-free plants developed symptoms. In the assessment done on all shoots of TMS 30.572

'64
c'
13
31
29
27
25
R W GIBSON AND G W OTIM-NAPE
% recovery
A
J Max. air temperature ---f
* - , . - , I I
Yo\ Dcc Jan Feh M a April May 1994 1995
-70
50
-30
. 10
Fig. I . The mean', maximum teinperaturrs November 1991 to May 1995 (-1 and the percentage of reco\ered monitored shoots (-:+-) i n field trial 2 . " X \e\en day "running" mean obtained by calculating the mean maximum temperature of the day imcerned and the three da is before and after i y given to show trends clearly.
plants on 2 May, of 472 shoots examined on originally diseased plants, 403 (859%) had recovered but I 12 (28%) of these had again developed symptoms. In contrast, only 10 (0.9%) of the I152 shoots examined on plants grown from cuttings from mosaic-free plants were diseaxd. Thus, few of the diseased shoots developing from previously recovered shoots are likely ! P < 0.001) to have been the result of a new infection by whiteflies and the majority must have been a re-invasion from the main plant.
Flowering and the production of new shoots from leaf axils immediately below the flower followed the onset of the rains in March and April (Table 7). Recovery now largely occurred at this branching rather than along a stem. In a count taken in early June on diseased stems of TMS 30572, 46 of the 105 shoots coming from the axils of symptomless leaves (but growing amongst diseased leaves) were symptomless whereas none of 274 shoots produced in the axils of diseased leaves had recovered ( P < 0.001 ). In order to examine this phenomenon differently. short two-node cuttings were taken from stems having a mixture of symptomless and diseased leaves. Five of 45 cuttings ( 1 1 % ) taken from nodes with diseased leaves produced symptomless shoots whereas 18 of 47 cuttings (38%) taken from nodes with symptomless leaves produced symptomless shoots (P < 0.005). None of 28 shoots from fully recovered stems produced diseased shoots.
At harvest. the TMS 30572 plants in plots planted with cuttings from symptomless plants each yielded an average of 2.70 kg. only slightly greater than the 2.45 kg average yield of

Recovery and reversion in mosaic virus diseased cassava 265
Table 4. Mean monthly disease incidences and severities on monitored main shoots in plots of cv. Bao and TMS 30572 in field trial 2, planted 6 Oct 1994
cv. Bao TMS 30572
Date Plots planted with: Plots planted with: infected cuttings healthy infected cuttings healthy
Disease % % Disease % % 199411 995 severity* recovery? infection? severity? recovery? infection?
Nov 2.7 * 0.08 0 0 1.8 * 0.07 3 0 Dec 3.0 * 0.05 0 0 1.9 2 0.06 3 0 Jan 3.1 * 0.04 0 11 1.9 * 0.09 3 0 Feb 3.2 * 0.06 0 11 1.3 * 0.05 31 0 Mar 3.1 f 0.07 0 11 1.4 * 0.08 64 0 April 3.7 * 0.04 0 36 2.1 * 0.14 61 0 May 3.6 * 0.04 0 61 2.0 2 0.09 50 0 * Mean score during each month * standard error of the mean. t At assessment nearest month’s end.
Table 5. The numbers of diseased (numerator) plants (denominator = total surviving plants) grown from cuttings obtained from the top, middle and bottom of plant stems of TMS 30572 in
field trials 2 and 3
Top of Middle of Bottom of Number of stem stem stem cuttings planted
Field trial 2: planted 6 Oct 1994
diseased cuttings (24%) (48%) (86%) Parent plants from 4/70 611 11 191117 123 disease-free cuttings (6%) (5%) (16%)
Field trial 3: planted 12 Sept 1995 Watered parent 215 23/32 31/36 46 plants (40%) (72%) (86%) Unwatered parent 1 I 4 2313 1 21/27 40 plants (25%) (74%) (78%)
Parent plants from 15/62 40183 891103 110
TMS 30572 plants in plots planted with cuttings from MD-affected plants (SED = 0.100; P < 0.05).
Most cuttings (86%) taken from the basal part of the stem of plants in plots planted with diseased cuttings produced diseased progeny plants whereas about a half (P < 0.001)and about three-quarters ( P < 0.001) of cuttings obtained from respectively the middle and top of the stem produced symptomless (reverted) progenies (Table 5). About 16% of cuttings taken from the basal part of stems of plants in plots planted with disease-free cuttings produced diseased progenies. However, again only about a third as many (5-6%) of cuttings taken from the middle and top of these stems produced diseased progenies. Table 5 also shows that whereas nearly all cuttings taken from the basal part established successfully, only c. 60% of cuttings from the top of the stem survived.
Cv. Bao All plants of cv. Bao grown from cuttings taken from diseased plants were diseased and
none recovered. For all sampling dates, MD was more severe on cv. Bao than on the more
266 R W GIBSON AND G W OTIM-NAPE
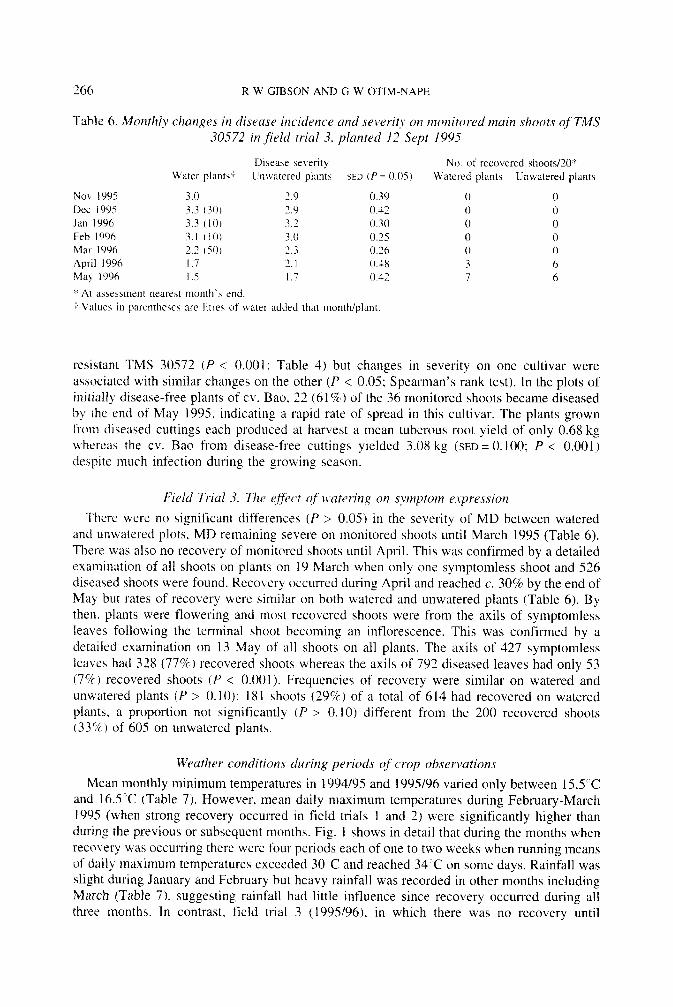
Table 6. Monthly changes in disease incidence and severih on inonitored innin shoots qf TMS 30572 in ,field trial 3, planted 12 Sept 1995
Disease sewrity No. of recovered shootsI20" Water plants? Unwatsred plants SED iP = 0.05) Watered plants Unwatered plants
NO\ 1995 3.0 2.9 0.39 0 0 Dec 1995 3.3 (30) 3.9 0.42 0 0 Jan 1996 3.3 (10) 3.3 0.30 0 0 Feb 1996 3.1 t 1 0 , 3.0 0.15 0 0 Mar I996 2.2 ( 5 0 ) 2 , 3 0.26 0 0 April 1996 I .7 2.1 0.4X 3 6 Ma) 1996 I .5 I .? 0.42 7 6
t Values in parentheses are litres of water added that month/plant. At assessment neare\t month'b end.
resistant TMS 30572 ( P < 0.001; Table 3 ) but changes in severity on one cultivar were associated with similar changes on the other ( P < 0.05; Spearman's rank test). In the plots of initially disease-free plants of cv. Bao, 22 (61%) of the 36 monitored shoots became diseased by the end of May 1995. indicating a rapid rate of spread in this cultivar. The plants grown from diseased cuttings each produced at harvest a mean tuberous root yield of only 0.68 kg urhereas the cv. Bao from disease-free cuttings yielded 3.08 kg (SED = 0.100: P < 0.001 ) despite much infection during the growing season.
Firld Trial 3. The e f k t of rtuterir?g on sjmptom expression There were no significant differences ( P > 0.05) in the severity of MD between watered
and unwatered plots, MD remaining severe on monitored shoots until March 1995 (Table 6). There was also no recovery of monitored shoots until April. This was confirmed by a detailed examination of all shoots on plants on 19 March when only one symptomless shoot and 526 diseased shoots were found. Recovery occurred during April and reached c. 30% by the end of May but rates of recovery were similar on both watered and unwatered plants (Table 6). By then, plants were flowering and most recovered shoots were from the axils of symptomless leaves following the terminal shoot becoming an inflorescence. This was confirmed by a detailed examination on 13 May of all shoots on all plants. The axils of 427 symptomless leaves had 328 (77%) recovered shoots whereas the axils of 792 diseased leaves had only 53 (7%) recovered shoots ( P < 0.001). Frequencies of recovery were similar on watered and unwatered plants ( P > 0.10); 181 shoots (29%) of a total of 614 had recovered on watered plants. a proportion not significantly ( P > 0.10) different from the 200 recovered shoots (33%) of 605 on unwatered plants.
Wecctlier conditions tizrririg periods qf crop observations Mean monthly minitnum temperatures in 1994/95 and 1995/96 varied only between 15.5"C
and 163°C (Table 7). However. mean daily maximum temperatures during February-March 1995 (when strong recovery occurred in field trials 1 and 2) were significantly higher than during the previous or subsequent months. Fig. 1 shows in detail that during the months when recovery was occurring there were four periods each of one to two weeks when iunning means of daily maximum temperatures exceeded 30-C and reached 34'C on some days. Rainfall was slight during January and February but heavy rainfall was recorded in other months including March (Table 7), suggesting rainfall had little intluence since recovery occurred during all three months. In contrast, field trial 3 (1995/96), in which there was no recovery until

Recovery and reversion in mosaic virus diseased cassava 267
Table 7. A summary of the mean (&standard error) monthly weather conditions during November to May inclusive, 1994/95 and 1995/96
Month
Nov Dec Jan Feb March April May
Mean
Mean daily minimum
temperature (“C)
16.4 * 0.16 16.0 i 0.23 15.9 i 0.15 16.4 * 0.17 16.3 i 0.14 15.9 i 0.14 15.7 k 0.17
16.1
1994195
Mean daily maximum
temperature
27.9 k 0.26 28.8 & 0.17 30.7 & 0.31 30.9 i 0.34 29.9 f 0.31 28.1 i 0.32 27.8 i 0.38
29.2
( “ C ) Rainfall
(mm) 86.3
138.0 8.9
23.9 131.7 139.7 112.2
91.5
Mean daily minimum
temperature (“C)
15.9 i 0.10 15.8 & 0.12 15.8 f 0.12 15.7 i 0.12 16.0 i 0.10 15.8 i 0.13 15.9 i 0.09
15.8
1995/96
Mean daily maximum
temperature ( “ C )
27.8 k 0.25 28.3 & 0.17 29.1 0.19 28.6 i 0.24 28.0 f 0.25 28.7 f 0.32 27.9 f 0.23
28.3
Rainfall (mm)
76.8 76.1
112.7 116.7 108.3 119.3 164.9
1 10.7
flowering began in April, had moderate temperatures, mean daily maximum temperatures for all months remained below 30°C with few days when the temperature exceeded 3 1 “C. Unlike 1995, maximum temperatures exceeded 31°C on no day in January, February or March 1996. Rainfall also fell regularly and heavily on field trial 3 during all months.
Discussion
Recovery This series of field trials confirmed that MD-resistant genotypes are more likely to recover
than susceptible ones (Jennings, 1960; Thresh, Otim-Nape & Jennings, 1994; Fargette e f al., 1994, 1996), recovery occurring in the resistant cv. TMS 30572 but not in the susceptible cv. Bao. Pathogen genotype was also shown to be important: plants of TMS 30572 graft- inoculated with the severe MD recovered later than plants inoculated with the mild MD and the recovered shoots of only severe MD-affected plants were reinvaded. The diseased plants of TMS 30572 in field trial 2 grown from cuttings from diseased plants in a farmer’s fields at Nakasongola all had symptoms resembling those of TMS 30572 grafted with the severe MD in field trial 1. A similar situation has been observed in crops of TMS 30572 in Nakasongola, Lira and Soroti (R W Gibson, unpublished data) and the absence of rapid and permanent recovery may give severe MD an advantage over the mild disease in crops of this cultivar.
Plant age Recovery occurred only after January 1995 in both field trial 1 and field trial 2, although the
two trials differed in age by about six months. Recovery was also to an extent reversed in May in both trials, contradicting the suggestion that diseased cassava “grow away from infection” (Rossel et al., 1987). Furthermore, recovery did not occur during these same months in 1996 in field trial 3 - even though plants were of a similar age to plants which recovered in field trial 2. These observations all indicate that plant age was unlikely to have been a major factor in causing recovery between January and March 1995.
Weather conditions That recovery occurred between January and March 1995 in field trials differing in age and
other factors suggest that an environmental factor(s) led to recovery. Drought was not

268 R W GIBSON AND G W OTIM-NAPE
obviously associated with recovery and Storey & Nichols ( 1938) considered temperature to be the main environmental influence on symptom expression. Recovery has been simulated by subjecting diseased TMS 30572 to 37 C for several days (Gibson, 1994) and longer periods at only 30'~C suppressed ACMV symptoms in other hosts (Robinson, Harrison, Sequeira & Duncan, 1984): the diminished recovery from the severe MD may reflect a greater tolerance to high temperature (Robinson et al., 1984). Hot weather has also caused recovery from other virus diseases in other crops elsewhere in the tropics or semi-tropics (Thirumalachar, 1954; Frazier et al.. 1965). The recovery in our trials can similarly be explained by natural thennotherapy as it was associated with prolonged periods with shade temperatures > 30°C in January, February and March 1995 (Table 7; Fig 1). Crop canopy temperatures are likely to have been even higher (Yao, Fargette & Fauquet, 1987).
Flo,t,erirzg Buds in the axils of leaves immediately below a flower naturally sprout and those shoots
from axils of symptomless leaves were mostly recovered. However, this leaves unanswered the question **What leads to the production of symptomless leaves and virus-free axillary buds amongst the diseased leaves'?''. Flowering itself appeared not to be the cause, as observation confirmed that leaves along a stem of TMS 30572 can be a mixture of diseased and symptomless (Bock & Harrison, 1985). Recovery associated with flowering even occurred in field trial 3 when prolonged periods of hot weather did not occur. However, given the significance of temperature in determining symptom expression (Storey & Nichols, 1938), perhaps a short period of high air temperature coupled with heating from the intense tropical sunlight may induce the effect on leaves and buds at a specific stage of development.
Re iiersior i
Plant gennQpe That reversion. like recovery, is a characteristic of MD-resistant genotypes (Jennings, 1957,
1960; Fargette et al., 1996) was confirmed in field trial 2, nearly half the cuttings of TMS 30572 used to plant this experiment produced disease-free plants whereas all cuttings of cv. Bao produced diseased plants.
I s i-r~vrsiniz predeterinined in the planted cutting or mused by the environment in which the cutting is grokvn?
In field trials 2 and 3. cuttings of TMS 30572 taken from the top of the stem mostly reverted. whereas cuttings taken from the bottom of the stem mostly did not, although all cuttings were sprouted together. Similar results have been obtained by Njock et al. (1994) and by Kossel et crl. ( 1 994) although the latter also showed that environmental conditions in which cuttings are grown can affect rates of reversion. Overall, they suggest that reversion is largely predetermined in the planted cutting.
Linkage M.ith recove- Cuttings taken from the top of stems, where recovery largely occurred, produced mostly
reverted progenies. Furthermore, more of the cuttings obtained from field trial 2, in which recovery was extensive, reverted than of those from field trial 3. That most cuttings taken from recovered parts of stems produced reverted progenies and that most cuttings from diseased parts did not. as also shown by Rossel et al. (1987), and that buds in the axils of symptomless leaves produced amongst diseased leaves also produced reverted progenies more

Recovery and reversion in mosaic virus diseased cassava 269
frequently than buds within the axils of diseased leaves, all link recovery in the parent plant to reversion in progeny cuttings.
Virus movement in plants Both recovery and reversion may be linked to ACMV being incompletely systemic in
cassava (Rossel et al., 1987, 1994; Njock et al., 1994). However, Jennings’ (1960) observed that virus introduced to upper leaves can move down to infect basal parts of stems even in resistant plant genotypes. This downward movement was confirmed by our top-grafting experiment, most cuttings taken from below grafts producing diseased progenies. Downward movement may occur along with photosynthates in phloem sap moving towards tuberous storage roots. Conversely, that mostly disease-free progenies were obtained from cuttings from upper, recovered shoots and the general stability of recovery imply little upward virus movement. Many recovered shoots in field trial 2 became re-diseased with the onset of the rains, and breaking off the top of a recovered plant also can result in the production of diseased shoots (Jennings, 1960; Anon., 1976); both might cause a temporary upward flow in the phloem to supply the sprouting shoot.
Exploitation of recovery and reversion In field trial 2, plants of TMS 30572 grown from cuttings from diseased plants produced c.
90% of the tuber yield of cuttings from symptomless plants. Contributors to the smallness of this loss may include:-
about 40% of the TMS 30572 cuttings obtained from diseased plants reverted; B many shoots of diseased plants recovered during the growing season; B reverted plants could compensate for reduced growth by diseased neighbours (Otim-Nape,
B diseased TMS 30572 plants had mild symptoms, so their yield may have been only slightly
This contrasts with results from plants of cv. Bao grown from cuttings taken from diseased plants. These plants, which were severely diseased and none of which reverted or recovered, produced only 20% of the tuber yield of disease-free cuttings. Other benefits specifically of reversion are that it:-
B provides an easy means of obtaining a MD-free stock (Fauquet, Fargette & Thouvenel, 1987);
B helps prevent the build-up of infection in farmers’ stocks (Fargette, Thresh & Otim-Nape, 1994; Fargette & Vie, 1995).
Reversion is associated with other aspects of disease resistance including symptom .ntensity (Fargette et al., 1996) and breeding more resistant cultivars should enhance -eversion. In the present studies, reversion occurred in only c. 20% of cuttings taken from the 3ase of TMS 30572 plants (where cuttings are preferentially obtained), suggesting that scope For such enhancement remains. Interestingly, greater reversion occurs in TMS 30572 in lowland Nigeria (Rossel et al., 1994) perhaps because temperatures can be higher there than in mid-altitude Uganda. Whatever the explanation, it indicates that reversion is malleable. rhus, planting crops prior to the hot season or in locations with hot microclimates may ncrease reversion or early recovery. Rates of reversion could also be enhanced by selecting Jlanting material from recovered parts of diseased plants either directly, or indirectly by .aking cuttings preferentially from the top part of stems. However, using cuttings from the top Tart of stems (Ilona, Akaroda & Dixon, 1995) and dry conditions associated with the hot
Thresh & Shaw, 1997);
decreased (Otim-Nape, Shaw & Thresh, 1994).

270 R W GIBSON AND G W OTIM-NAPE
seacon can lead to poor crop establishment, and in practice compromises have to be made to obtain satisfactory yields.
Acknowledgements This work was jointly undertaken under a scientific arrangement between the Ugandan
National Agricultural Research Organisation and the Natural Resources Institute, UK, funded by the Natural Resources Division of the British Department For Overseas Development. We wish to thank Mr I Mpembe for technical support and acknowledge valuable advice from Dr J M Thresh. We thank Mr A Ssali for kindly providing meteorological data.
References Anon. 1976. Pathology. Atiii~id Report .for 1976 of the Intertiatioritrl Institrite for Tropical Agriculture,
Rock K R. 1994. Control of African cassava mosaic geniinivirus by using virus-free planting material. Tropicul Sciet7ce 34: 102-109.
Rock K R, Guthrie E J. 1982. Control of c ava mosaic disease in Kenya. Tropical Pest Management 2x:219-222.
Bock K R, Harrison B D. 1985. African cassava mosaic virus. Associution of Applied Biologists, Drsc.riptioii ( f Plant Viriises No. 297. 6 pp.
Dubern J. 1994. Transmission of African cassava mosaic geniinivirus by the whitefly Bemisia tabaci. Tropical Science 34:82-9 1 .
Fargette D, Vie K. 1995. Simulation of the effects of host resistance, reversion, and cutting selection on incidence of African cassava mosaic virus and yield losses in cassava. Pbytopathology 85:370-375.
Fargette D, Thresh J M, Otim-Nape G W. 1994. The epidemiology of African cassava mosaic virus: reversion and the concept of equilibrium. Tropical Science 34: 123-1 33.
Fargette D, Colon L T, Bouveau R, Fauquet C. 1996. Components of resistance of cassava to African cassava mosaic virus. Errroperiii Jnrrniul ($ Plant Pathology 102:645-654.
Fauquet C, Fargette D, Thouvenel J-C. 1987. Selection of healthy cassava plants obtained by reversion in cassava fields. T h Intertzcrtiond Seniiiiar 017 African cassava mosaic virus and its coiztrol. Published by CTA. FAO. ORSTOM, IITA & IAPC. Yanioussoukro, CBte d’Ivoire 1987, pp. 146--149.
Frazier N W, Voth V, Bringhurst R S. 1965. Inactivation of two strawberry viruses in plants grown in a natural high-temperature environment. Pb>~topatbology 55: 1203-1 205.
Geddes A M W. 1990. The relative importance of crop pests in sub-Saharan Africa. Natrrral Resources Ii~,stiti(te Rulleriri No. 36:vi + 69 pp.
Gibson R W. 1994. Long-term absence of symptoms in heat-treated African cassava mosaic geminivirus-infected resistant cassava plants. Tropiccrl Science 34: 154-158.
Gibson R W, Legg J P, Otim-Nape G W. 1996. Severe symptoms are a feature of an epidemic of mosaic virus disease of cassava in Uganda. Annals of Applied Biology 128:479-490.
Hollings M. 1965. Disease control through virus-free stock. Annual Review of Phytopathology 3:367- 396,
Ilona P C, Akoroda M 0, Dixon A G 0. 1995. Effect of stem sections on propagation, yield and yield components of cassava. In Root Crops n i i d Po1.er-h ullei3iation. Shtli Triennial Symposium of the Iriteriinrioizcil Socieh , for- Tropicrrl Root C r o p - A,fi-icn Brcinclz (ISTRC-AB) Lilongwe, Mulawi. p. 131.
Jennings D L. 1957. Further studies in breeding cassava for virus resistance. The East African A,yridtirrul Journal 22:2 13-2 19.
Jennings D L. 1960. Observations on virus diseases of cassava in resistant and susceptible varieties I Mosaic disease. Empire Joirrnal of Eqieriitieiii~il Agricrrltzire 28:23-34.
p. 57.

Recovery and reversion in mosaic virus diseased cassava 27 1
Kufferath H, Ghesquiere J. 1932. La mosaique du manioc. Compte-Rendu de la Socie'te' de Biologie Belge 109: 1 146.
Vjock T E, Atiri G I, Thottappilly G, Hahn S K. 1994. Distribution and disease recovery of African cassava mosaic virus (ACMV) in stems of resistant and susceptible cassava. In Root Crops for Food Security, pp. 149-152. Ed. M 0 Akoroda. Published by ISTRC, CTA & IITA. Proceedings of the Fifih Symposium of the International Society for Tropical Root Crops - Africa Branch. (Kampala,
Accelerating the transfer of improved production technologies: controlling cassava mosaic virus disease epidemics in Uganda. African Crop Science Journal 2:419495.
3tim-Nape G W, Shaw M W, Thresh J M. 1994. The effects of African cassava mosaic geminivirus on the growth and yield of cassava in Uganda. Tropical Science 34:43-54.
3tim-Nape G W, Thresh J M, Shaw, M W. 1997. The effects of cassava mosaic disease on yield and compensation in mixed strands of healthy and infected cassava. Annals of Applied Biology 130503- 521.
'acumbaba R P. 1985. Virus-free shoots from cassava stem cuttings infected with cassava latent virus. Plant Disease 69:231-232.
iobinson D J, Harrison B D, Sequeira J C, Duncan G H. 1984. Detection of strains of African cassava mosaic virus by nucleic acid detection and some effects of temperature on their multiplication. Annals of Applied Biology 105:484493.
iossel H W, Changa C M, Atiri G I. 1994. Quantification of resistance to African cassava mosaic virus (ACMV) in IITA-improved, mosaic-resistant cassava breeding materials. In Root Crops for Food Security, pp. 280-281. Ed. M 0 Akoroda. Published by ISTRC, CTA & IITA. Proceedings of the Fifth Symposium of the International Society for Tropical Root Crops -Africa Branch. (Kampalu, Uganda).
tossel H W, Thottappilly G, van Lent J M W I, Huttinga H. 1987. The etiology of cassava mosaic in Nigeria. The International Seminar on African cassava mosaic disease and its control, Yamoussoukro, Cbte d'lvoire, pp 43-56. Published by CTA, FAO, ORSTOM, IITA & IAPC.
itorey H H, Nichols R F W. 1938. Studies of the mosaic diseases of cassava. Annals of Applied Biology 25:790-806.
iwanson M M, Harrison B D. 1994. Properties, relationships and distribution of cassava mosaic geminiviruses. Tropical Science 34: 15-25.
rhirumalachar M J. 1954. Inactivation of potato leafroll by high temperature storage of seed tubers in the Indian plains. Phytopathologische Zeitschrij? 23:429436.
rhresh J M, 0th-Nape G W, Jennings D L. 1994. Exploiting resistance to African cassava mosaic virus. Aspects of Applied Biology 39, The impact of genetic variation on sustainable agriculture, pp.
Wroth J M, Jones R A C. 1992. Subterranean clover mottle virus infection of subterranean clover: widespread occurrence in pastures and effects on productivity. Australian Journal of Agriculturul Research 43: 1597-1608.
lao N R, Fargette D, Fauquet C. 1987. Microclimate of a cassava canopy. The International Seminar on African cassava mosaic virus and its control, pp. 92-104. Published by CTA. FAO, ORSTOM, IITA & IAPC. Yamoussoukro, C6te d'Ivoire.
5hou X, Liu Y, Calvert L, Muiioz C, 0th-Nape G W, Robinson D J, Harrison B D. 1997. Evidence that DNA-A of a geminivirus associated with severe cassava mosaic disease in Uganda ha5 arisen by interspecific recombination. Journal of General Virology 78:2101-2111.
Uganda). 3tim-Nape G W, Bua A, Baguma Y. 1994.
5 1-60.
(Received 3 April 1997)
Copyright © 2022 FDOKUMEN




















