
Executive and mnemonic functions in early Huntington's disease
13
Brain (1996), 119, 1633-1645 Executive and mnemonic functions in early Huntington's disease Andrew D. Lawrence, 12 Barbara J. Sahakian, 2 John R. Hodges, 3 Anne E. Rosser, 34 Klaus W. Lange 5 and Trevor W. Robbins 14 Departments of 'Experimental Psychology, 2 Psychiatry and ^Neurology and the *MRC Centre for Brain Repair, University of Cambridge, Cambridge and the ^Department of Clinical Neurology, Institute of Neurology, University of London, London, UK Correspondence to: Dr Barbara J. Sahakian, Department of Psychiatry, University of Cambridge School of Clinical Medicine, Addenbrooke's Hospital, Cambridge CB2 2QQ, UK Summary Eighteen patients with early Huntington's disease were compared with age- and IQ-matched control volunteers on tests of executive and mnemonic function taken from the Cambridge Neuropsychological Test Automated Battery. Tests of pattern and spatial recognition memory, spatial span, spatial working memory, spatial planning and visual discrimination learning/attentional set shifting were employed. These tests have previously been found to be sensitive to the later stages of Huntington 's disease. Patients with early Huntington's disease were found to have a wide range of cognitive impairments encompassing both visuo- spatial memory and executive functions, a pattern distinct from those seen in other basal ganglia disorders. In contrast to patients with more advanced Huntington's disease, early Huntington's disease patients were not impaired at simple reversal learning, but were impaired at performing an extradimensional shift (EDS). The results will be discussed in relation to the hypothesized neuropathological staging of Huntington's disease and to the anatomical connectivity of the striatum. Keywords: Huntington's disease; striatum; subcortical dementia; executive functions; memory Abbreviation: EDS = extradimensional shift Introduction Huntington's disease is characterized clinically by a triad of progressive motor, psychiatric and cognitive symptoms and neuropathologically by the loss of GABA-containing medium-size spiny neurons in the neostriatum (Folstein, 1989). Since the primary pathology in Huntington's disease lies within subcortical regions of the brain, Huntington's disease has been characterized as a 'subcortical' dementia, in contrast with the 'cortical' dementia of Alzheimer type where the primary site of pathology lies in cortical regions (Albert et al., 1974). The cardinal features of 'subcortical' dementias are considered to be problems in retrieving memories, slowed information processing, cognitive inflexibility and mood and personality changes, in contrast to the aphasia, agnosia and amnesia of dementia of Alzheimer type (Cummings, 1990). It has been noted by many investigators that the cognitive disturbances seen in 'subcortical dementia' resemble those seen following lesions of the prefrontal cortex (Cummings, 1993). In order to explain © Oxford University Press 1996 these findings, recourse has been made to recent anatomical evidence for the existence of a series of five parallel and (at least partially) segregated circuits, each of which leads from a discrete frontal region, via separate areas of the basal ganglia and thalamus, back to the frontal region from which it originates (Alexander et al., 1986; Parent and Hazrati, 1995). In the light of these findings one might well expect the cognitive symptoms of Huntington's disease to be similar to those seen following focal lesions to the prefrontal cortex and if so, the term 'fronto-striatal' dementia (Robbins et al., 1994) might be more appropriate for Huntington's disease. Lesions of the prefrontal cortex result in impairments in 'executive' functions. The term executive function refers to a variety of cognitive activities considered to be at the pinnacle of human cognition, e.g. planning and decision- making. These are functions which rely upon, but cannot be reduced to, other cognitive faculties, such as memory and perception (Tranel et al., 1994). Several investigators have by guest on October 20, 2016 http://brain.oxfordjournals.org/ Downloaded from
Transcript of Executive and mnemonic functions in early Huntington's disease

Brain (1996), 119, 1633-1645
Executive and mnemonic functions in earlyHuntington's diseaseAndrew D. Lawrence,12 Barbara J. Sahakian,2 John R. Hodges,3 Anne E. Rosser,34 Klaus W. Lange5
and Trevor W. Robbins14
Departments of 'Experimental Psychology, 2Psychiatry and^Neurology and the *MRC Centre for Brain Repair,University of Cambridge, Cambridge and the ^Departmentof Clinical Neurology, Institute of Neurology, University ofLondon, London, UK
Correspondence to: Dr Barbara J. Sahakian,Department of Psychiatry, University of Cambridge Schoolof Clinical Medicine, Addenbrooke's Hospital, CambridgeCB2 2QQ, UK
SummaryEighteen patients with early Huntington's disease werecompared with age- and IQ-matched control volunteers ontests of executive and mnemonic function taken from theCambridge Neuropsychological Test Automated Battery. Testsof pattern and spatial recognition memory, spatial span,spatial working memory, spatial planning and visualdiscrimination learning/attentional set shifting wereemployed. These tests have previously been found to besensitive to the later stages of Huntington 's disease. Patientswith early Huntington's disease were found to have a wide
range of cognitive impairments encompassing both visuo-spatial memory and executive functions, a pattern distinctfrom those seen in other basal ganglia disorders. In contrastto patients with more advanced Huntington's disease, earlyHuntington's disease patients were not impaired at simplereversal learning, but were impaired at performing anextradimensional shift (EDS). The results will be discussedin relation to the hypothesized neuropathological staging ofHuntington's disease and to the anatomical connectivity ofthe striatum.
Keywords: Huntington's disease; striatum; subcortical dementia; executive functions; memory
Abbreviation: EDS = extradimensional shift
IntroductionHuntington's disease is characterized clinically by a triad ofprogressive motor, psychiatric and cognitive symptoms andneuropathologically by the loss of GABA-containingmedium-size spiny neurons in the neostriatum (Folstein,1989). Since the primary pathology in Huntington's diseaselies within subcortical regions of the brain, Huntington'sdisease has been characterized as a 'subcortical' dementia,in contrast with the 'cortical' dementia of Alzheimer typewhere the primary site of pathology lies in cortical regions(Albert et al., 1974). The cardinal features of 'subcortical'dementias are considered to be problems in retrievingmemories, slowed information processing, cognitiveinflexibility and mood and personality changes, in contrastto the aphasia, agnosia and amnesia of dementia ofAlzheimer type (Cummings, 1990). It has been noted bymany investigators that the cognitive disturbances seen in'subcortical dementia' resemble those seen following lesionsof the prefrontal cortex (Cummings, 1993). In order to explain
© Oxford University Press 1996
these findings, recourse has been made to recent anatomicalevidence for the existence of a series of five parallel and (atleast partially) segregated circuits, each of which leads froma discrete frontal region, via separate areas of the basalganglia and thalamus, back to the frontal region from whichit originates (Alexander et al., 1986; Parent and Hazrati,1995). In the light of these findings one might well expectthe cognitive symptoms of Huntington's disease to be similarto those seen following focal lesions to the prefrontal cortexand if so, the term 'fronto-striatal' dementia (Robbins et al.,1994) might be more appropriate for Huntington's disease.
Lesions of the prefrontal cortex result in impairments in'executive' functions. The term executive function refers toa variety of cognitive activities considered to be at thepinnacle of human cognition, e.g. planning and decision-making. These are functions which rely upon, but cannot bereduced to, other cognitive faculties, such as memory andperception (Tranel et al., 1994). Several investigators have
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

1634 A. D. Lawrence et al.
claimed that patients with Huntington's disease show anexecutive impairment as they perform poorly on tasks suchas the Wisconsin Card Sorting Test (Josiassen et al., 1983),the Stroop task (Swerdlow et al., 1995), verbal fluency (Rosserand Hodges, 1994) and Tower of Hanoi-style problems (Saint-Cyr et al., 1988; but see Butters et al., 1985), tasks which areall sensitive to prefrontal cortical lesions (Tranel et al., 1994).
In a recent study, the performance of patients withHuntington's disease relatively late in the course of thedisease and patients with dementia of the Alzheimer type,matched for degree of dementia has been examined by uson tasks of executive and mnemonic function (Lange et al.,1995). In keeping with the proposed distinction betweenfronto-striatal and cortical dementias, the patients withHuntington's disease were significantly more impaired thanpatients with dementia of Alzheimer type on tasks ofexecutive function which have been shown previously to beimpaired by focal lesions to the prefrontal cortex (for review,see Robbins et al., 1996). These included the Tower ofLondon test of planning, a self-ordered spatial workingmemory task and an attentional set shifting paradigm whichdeconstructs the Wisconsin Card Sorting Test into itsconstituent elements. The former two of these tasks havebeen shown to activate distinct regions of the prefrontal andparietal cortices using PET (Baker et al., 1996; Owen et al.,1996). Performance on the spatial working memory task andthe Tower of London is quite highly correlated (Owen et al.,1992), suggesting that spatial working memory is an importantcomponent of planning ability. Additional tests were alsoadministered in the investigation of late Huntington's diseasepatients by Lange et al. (1995), further to specify the natureof possible contributory spatial memory impairments toimpaired executive functions. These included a test of spatialrecognition memory, sensitive to frontal lobe lesions (Owenet al., 1995) and a test of spatial span, which althoughinsensitive to frontal lobe damage (Owen et al., 1995),activates a region of ventral prefrontal cortex (area 47) aswell as a region of parietal Cortex (Owen et al., 1996).
In the Lange et al. (1995) study, patients with Huntington'sdisease also showed a greater impairment than patients withdementia of Alzheimer type on a test of pattern recognitionmemory, sensitive to temporal lobe lesions and amygdalo-hippocampectomy but not to frontal lobe lesions (Owenet al., 1995). This result suggests that not all the cognitivedeficits of Huntington's disease can be explained by a 'fronto-striatal' hypothesis. However, the Huntington's diseasepatients were in the advanced stages of the disease, whererather widespread cortical atrophy is known to occur (e.g.Mann et al., 1993) and it is likely that differences betweenthe dementias will be at their greatest in the earlier stages ofdisease, when pathological changes are relatively restricted.Thus, in order to test the fronto-striatal hypothesis moreclearly we examined the performance, on the same testbattery, of patients in the relatively early stages ofHuntington's disease where damage is thought to be restrictedprimarily to the caudate nucleus and putamen (Vonsattel
et al., 1985). In addition, although there have been a smallnumber of studies of the progression of cognitive decline inHuntington's disease (e.g. Moses et al., 1981; Josiassen et al.,1983; Bamford et al., 1995), there is still relatively littleknowledge about the exact staging of executive functionimpairments in Huntington's disease. A cross-sectionalcomparison with the data of Lange et al. (1995) allowed usto examine this issue in detail.
MethodsSubjectsPatient groupEighteen out-patients with symptomatic Huntington's disease(Huntington's disease), six women and 12 men, mean age44.1 years (SD 12.2 years), took part in this study. Care wastaken to select patients early in the course of the disease.Eight of the patients attended the Department of ClinicalNeurology at the National Hospital, London and werediagnosed on the basis of clinical signs and a positive familyhistory (Folstein et al., 1986). The remaining 10 patientsattended the Huntington's disease clinic at the Departmentof Clinical Genetics, Addenbrooke's Hospital, Cambridgeand had tested positive for the abnormal expansion ofthe huntingtin gene (Huntington's Disease CollaborativeResearch Group, 1993; Warner et al, 1993).
The mean age at onset for the group as a whole was40.1 years (SD 10.7 years) and the mean duration of choreawas 4.0 years (SD 2.3). Three patients were receivingmedication at time of testing. One patient was receivingdiazepam (2 mg per day), one sulpiride (50 mg per day) andone temazepam and haloperidol (3 mg twice daily). Twoof the Huntington's disease patients were suffering fromdepression at the time of testing.
The eight London patients were administered the choreascale of the Quantified Neurological Examination (Davidet al., 1987). This scale ranges from 0 to 25 (a high scoredenoting severe impairment) and the patient group scored amean of 6.1 (SD 1.4). In addition, the London group wereadministered the Kendrick Object Learning Test, a dementiascreening test taken from Kendrick's cognitive tests for theelderly (Kendrick, 1985), scoring a mean of 35.9 (SD 2.5),which is above the cut-off score for dementia of 22.
The 10 Cambridge patients were administered thefunctional assessment scale of the unified Huntington'sdisease rating scale (Huntington Study Group, 1996). This isa 25-item scale designed to assess functional impairment inHuntington's disease, a score of 25 indicating no impairmentin activities of daily living and 0 representing maximalimpairment. The patients' mean score was 21.9 (SD 3.1),indicating very mild disability. In addition, the Cambridgegroup were administered the Mini-Mental State Examination(Folstein et al., 1975), a validated measure of dementia,scoring a mean of 28.0 (SD 1.3) out of a possible maximumof 30. None of the patients scored below the cut-off score
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from
Cognition in early Huntington's disease 1635
for dementia of 24. Unfortunately, the London group werenot administered an activities of daily living scale. However,functional capacity correlates highly with scores on dementiarating scales (Folstein, 1989, p. 36) and none were consideredto be more than mildly functionally impaired. CT scans werenot available for either group.
A one-way ANOVA revealed that the London groupwere significantly younger than the Cambridge group[F(l,16) = 5.40, P = 0.03]. The mean age of the Londongroup was 37.4 years (SD = 9.1 years) compared with 49.4years (SD 12.1 years) for the Cambridge group. Furtheranalyses, using age as a covariate, revealed that the twopatient groups did not differ significantly on any of thecognitive measures.
Control subjectsEighteen control subjects were chosen to match the patientgroup as closely as possible with respect to both age andpremorbid verbal IQ as measured by the National AdultReading Test (Nelson, 1982). These subjects were drawn froma pool of control volunteers recruited through advertisementsplaced in the Cambridge employment centre. Unpaired t testsrevealed that the two groups did not differ in terms of age[f(34) = 0.45, P > 0.6] or estimated premorbid verbal IQ[f(34) = 0.35, P > 0.7].
For the visual discrimination learning and attentional setshifting task, the patient group was compared with a separategroup of control volunteers [n = 30; mean age = 42.4 years(SD = 19.9); mean National Adult Reading Test = 114.7(SD = 7.5)] drawn from the North East Age Researchpanel in Newcastle upon Tyne and from the Newcastle,Cambridgeshire and London areas. This group did not differsignificantly from the Huntington's disease patients in age[f(46) = 0.32, P = 0.8] or National Adult Reading Testscores [f(46) = \.2,P = 0.2]. A summary of the characteristicsof the three groups is presented in Table 1. Informed consentwas obtained from all patients and control volunteers.
ProcedureAll tests were taken from the Cambridge NeuropsychologicalTest Automated Battery, a suite of computerized cognitivetasks run on either an Acorn BBC Master microcomputerwith a high resolution Microvitec colour monitor and aMicrovitec (TouchTec 501) touch sensitive screen or on aportable 'Carry I' 486 microcomputer, fitted with a dataluxtouch-sensitive screen. Subjects were seated comfortably,-0.5 m from the touchscreen and it was explained to themthat they would have to respond to stimuli presented on themonitor screen by touching them. In order to introducesubjects to the apparatus, they were initially given a 'motorscreening task' in which they were asked to respond to aseries of 10 flashing crosses presented at varying locationson the monitor screen by placing the index finger of theirpreferred hand on the centre of each cross. After completion
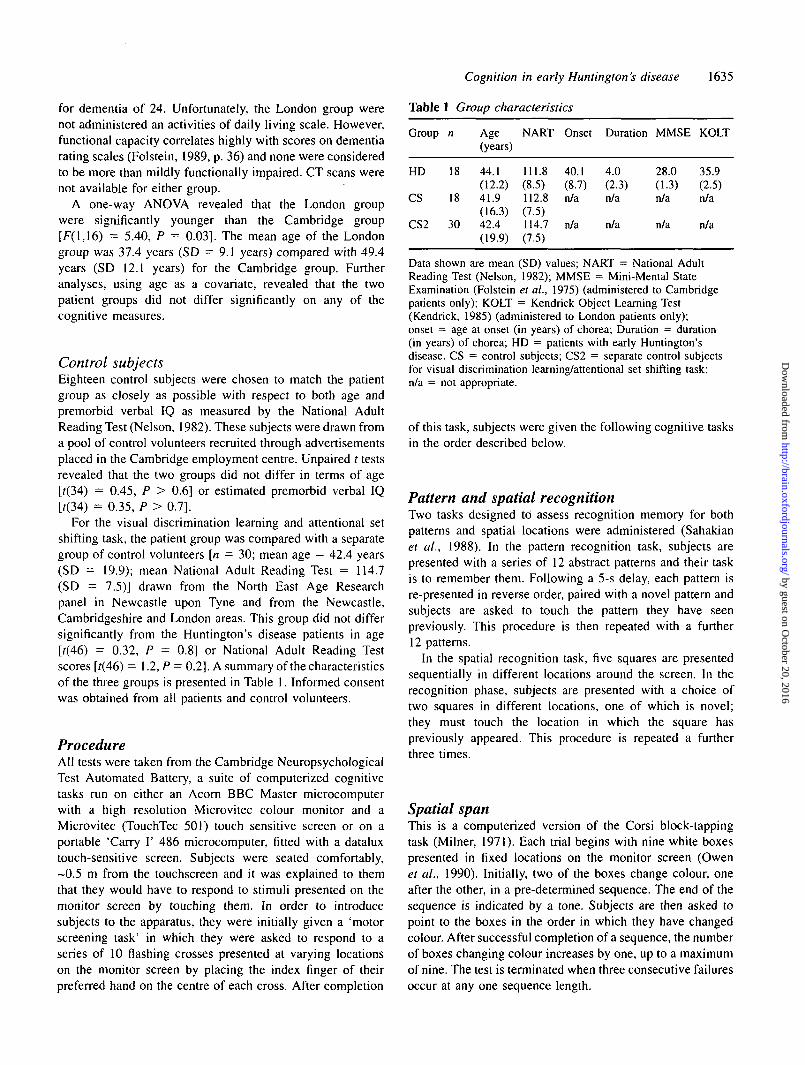
Table 1 Group characteristics
Group
HD
CS
CS2
n
18
18
30
Age(years)
44.1(12.2)41.9(16.3)42.4(19.9)
NART
111.8(8.5)112.8(7.5)114.7(7.5)
Onset
40.1(8.7)n/a
n/a
Duration
4.0(2.3)n/a
n/a
MMSE
28.0(1.3)n/a
n/a
KOLT
35.9(2.5)n/a
n/a
Data shown are mean (SD) values; NART = National AdultReading Test (Nelson, 1982); MMSE = Mini-Mental StateExamination (Folstein et al., 1975) (administered to Cambridgepatients only); KOLT = Kendrick Object Learning Test(Kendrick, 1985) (administered to London patients only);onset = age at onset (in years) of chorea; Duration = duration(in years) of chorea; HD = patients with early Huntington'sdisease. CS = control subjects; CS2 = separate control subjectsfor visual discrimination learning/attentional set shifting task;n/a = not appropriate.
of this task, subjects were given the following cognitive tasksin the order described below.
Pattern and spatial recognitionTwo tasks designed to assess recognition memory for bothpatterns and spatial locations were administered (Sahakianet al., 1988). In the pattern recognition task, subjects arepresented with a series of 12 abstract patterns and their taskis to remember them. Following a 5-s delay, each pattern isre-presented in reverse order, paired with a novel pattern andsubjects are asked to touch the pattern they have seenpreviously. This procedure is then repeated with a further12 patterns.
In the spatial recognition task, five squares are presentedsequentially in different locations around the screen. In therecognition phase, subjects are presented with a choice oftwo squares in different locations, one of which is novel;they must touch the location in which the square haspreviously appeared. This procedure is repeated a furtherthree times.
Spatial spanThis is a computerized version of the Corsi block-tappingtask (Milner, 1971). Each trial begins with nine white boxespresented in fixed locations on the monitor screen (Owenet al., 1990). Initially, two of the boxes change colour, oneafter the other, in a pre-determined sequence. The end of thesequence is indicated by a tone. Subjects are then asked topoint to the boxes in the order in which they have changedcolour. After successful completion of a sequence, the numberof boxes changing colour increases by one, up to a maximumof nine. The test is terminated when three consecutive failuresoccur at any one sequence length.
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

1636 A. D. Lawrence et al.
Spatial working memoryThis is a test of spatial working memory for humans (Owenet al., 1990), which is formally analogous to the Olton radialarm maze (Olton and Samuelson, 1976), a task of spatialworking memory for the rat. Both tasks require memory forlocations already visited. In this task, subjects are requiredto 'search through' a number of coloured boxes presentedon the monitor screen (by touching each one), in order tofind blue 'tokens' which are hidden inside. On any one trial,only a single token is hidden in one of the boxes. Oncefound, the next token is hidden. The key instruction is thatonce a token has been found within a particular box, thenthat box will not be used again to hide a token. Two error-types are possible. First, a subject may return to open a boxin which a token has already been found (a 'between-search'error). Secondly, a subject may return to a box alreadyopened and shown to be empty earlier in the same trial (a'within-search' error). There are four trials with each of four,six and eight boxes. The task is scored according to thenumber of between- and within-search errors at each levelof difficulty and also for the use of an efficient search strategy(Owen et al., 1990). A particularly efficient strategy forcompleting this task is to follow a predetermined searchsequence, starting with a particular box and then returningto start each new sequence with that same box as soon as atoken is found (editing the sequence when a token is foundin that box). The extent to which such a strategy is used isestimated from the number of search sequences starting witha novel box for just the more difficult six- and eight-boxproblems. The total of theses scores provides a measure ofstrategy for each subject, with a high score (many sequencesstarting with a novel box) representing poor use of a strategyand vice versa.
Tower of LondonThe Cambridge Neuropsychological Test Automated Batterycontains a modified version of the Tower of London spatialplanning task (Shallice, 1982), in which it is possible tomeasure the speed and accuracy of thinking. In the problem-solving condition, subjects have to move coloured ballsbetween suspended vertical 'stockings' represented on themonitor screen in order to establish a goal position in aspecified number of moves (Owen et al., 1990). In a yokedcontrol condition, the computer 'plays back' the solution toeach problem using the sequence of moves actually employedby the subject, a single move at a time. The subject has tocopy each individual move without having to .plan them, aspart of a sequence. Subtraction of the latencies to move theballs in the yoked control condition from that in the problem-solving condition provides a measure of thinking time. Theaccuracy of thinking can be measured in a number of ways,the most stringent being the proportion of problems solvedin the minimum specified number of moves (the number of'perfect' solutions). A less stringent measure is the number
of moves taken in excess of the specified minimum number,but within the maximum allowed.
Visual discrimination learning!attentional setshiftingThis task of discrimination learning has been described ingreater detail elsewhere (Roberts et al., 1988; Downes et al.,1989). The task requires subjects to learn a series of two-alternative forced-choice discriminations using feedbackprovided automatically by the computer. The task iscomposed of nine stages presented in the same fixed order,starting with a simple discrimination and its reversal forstimuli varying in just one dimension (e.g. two differentwhite line configurations). A second, alternative dimensionis then introduced (purple filled shapes) and compounddiscrimination and reversal are tested. To succeed, subjectsmust continue to respond to the previously relevant dimensionwhilst ignoring the presence of the new, irrelevant dimension.At the intradimensional shift stage, novel exemplars of eachof the two dimensions are introduced and subjects mustcontinue to respond to one of the two exemplars from thepreviously relevant dimension. Following another reversal,the EDS and its reversal are presented, again using novelexemplars of each stimulus dimension. In order to succeedat this stage, the subject must shift 'response set' to thepreviously irrelevant stimulus dimension, whilst ignoring thepreviously relevant dimension. The EDS stage is akin to achange in category in the Wisconsin Card Sorting Test.
Data analysisThe majority of the experimental data were analysed usingthe statistical packages STATWORKS (Cricket Software Inc.,Philadelphia, Penn., USA), CLR ANOVA (Clear LakeResearch Inc., USA) and SPSS V4.0 (SPSS Inc., Chicago,111., USA) running on an Apple Macintosh SE/30 computer.Data were transformed where appropriate (Tukey, 1977). Testvariables were compared using ANOVA. The ANOVA modelwas a two-factor design with group and difficulty level asfactors. Student's t tests were applied to test the significanceof differences between means where factorial ANOVA wasnot required. Where appropriate, Pearson product moment andSpearman rank order correlation coefficients were calculated.
For the attentional set shifting task, the data for the numberof subjects passing or failing at each stage of the task werecast into contingency tables and analysed using the likelihoodratio method (Kullback, 1968; Robbins, 1977), which isuseful for analysing data with small cell frequencies and forpartitioning inhomogeneities in the contingency table byadditive, orthogonal contrasts. The resulting 'informationstatistic' (2i) is distributed as %2.
ResultsPattern and spatial recognition memoryThe mean percentage correct scores for both the pattern andspatial recognition memory tests are presented in Table 2.
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from
Cognition in early Huntington's disease 1637
Table 2
Group
HDCS
Group comparisons
Patternrecognition
74.5 (15.4)90.5 (8.9)
of visuospatial
Spatialrecognition
71.9(15.4)86.4(6.1)
memory tasks
Spatial span
4.7 (0.7)6.2 (1.3)
Data shown are mean (SD) values. HD = patients with earlyHuntington's disease; CS = control subjects.
Unpaired / tests revealed that the Huntington's disease patientsperformed significantly worse than control subjects on bothpattern [?(34) = 4.05, P < 0.001] and spatial [/(34) = 3.69,P < 0.002] recognition. A paired t test revealed that controlsubjects found spatial recognition more difficult than patternrecognition [f(17) = 2.36, P < 0.03], whereas this was notthe case for the Huntington's disease patients [/(17) = 0.74,P > 0.4].
The mean response latencies for both tasks were alsocompared, following logarithmic transformation to reducepositive skew in the distribution. Unpaired t tests revealedthat the Huntington's disease group were significantly slowerthan controls on the pattern recognition task [f(34) = 4.29,P < 0.001] and also the spatial recognition task [f(34) = 2.02,P = 0.05].
Spatial spanMean spatial spans for the two groups are presented in Table2. Huntington's disease patients had a significantly shorterspatial span than did their control subjects [f(34) = 4.20,P < 0.001].
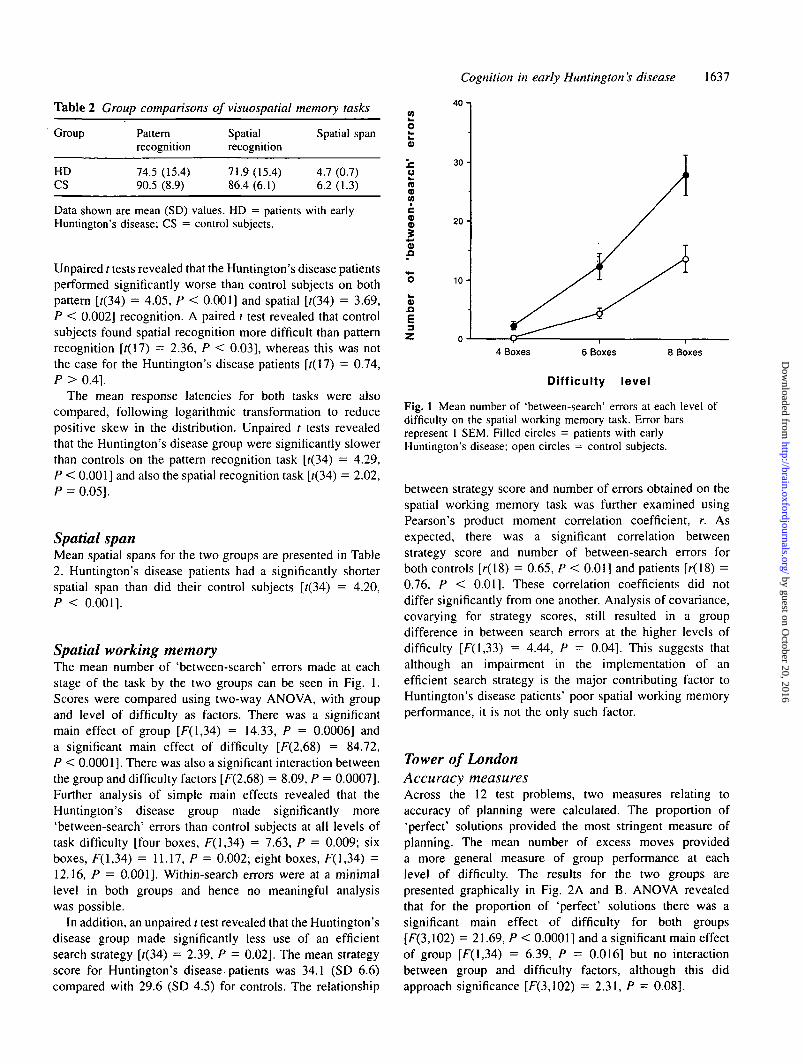
Spatial working memoryThe mean number of 'between-search' errors made at eachstage of the task by the two groups can be seen in Fig. 1.Scores were compared using two-way ANOVA, with groupand level of difficulty as factors. There was a significantmain effect of group [F(l,34) = 14.33, P = 0.0006] anda significant main effect of difficulty [F(2,68) = 84.72,P < 0.0001]. There was also a significant interaction betweenthe group and difficulty factors [F(2,68) = 8.09, P = 0.0007].Further analysis of simple main effects revealed that theHuntington's disease group made significantly more'between-search' errors than control subjects at all levels oftask difficulty [four boxes, F(l,34) = 7.63, P = 0.009; sixboxes, F(l,34) = 11.17, P = 0.002; eight boxes, F(l,34) =12.16, P = 0.001]. Within-search errors were at a minimallevel in both groups and hence no meaningful analysiswas possible.
In addition, an unpaired / test revealed that the Huntington'sdisease group made significantly less use of an efficientsearch strategy [f(34) = 2.39, P = 0.02]. The mean strategyscore for Huntington's disease-patients was 34.1 (SD 6.6)compared with 29.6 (SD 4.5) for controls. The relationship
(0
o
ora
McoV
o
.aE3
40 -i
30-
20-
10-
4 Boxes 6 Boxes 8 Boxes
Difficulty level
Fig. 1 Mean number of 'between-search' errors at each level ofdifficulty on the spatial working memory task. Error barsrepresent 1 SEM. Filled circles = patients with earlyHuntington's disease; open circles = control subjects.
between strategy score and number of errors obtained on thespatial working memory task was further examined usingPearson's product moment correlation coefficient, r. Asexpected, there was a significant correlation betweenstrategy score and number of between-search errors forboth controls [r(18) = 0.65, P < 0.01] and patients [r(18) =0.76, P < 0.01]. These correlation coefficients did notdiffer significantly from one another. Analysis of covariance,covarying for strategy scores, still resulted in a groupdifference in between search errors at the higher levels ofdifficulty [F(l,33) = 4.44, P = 0.04]. This suggests thatalthough an impairment in the implementation of anefficient search strategy is the major contributing factor toHuntington's disease patients' poor spatial working memoryperformance, it is not the only such factor.
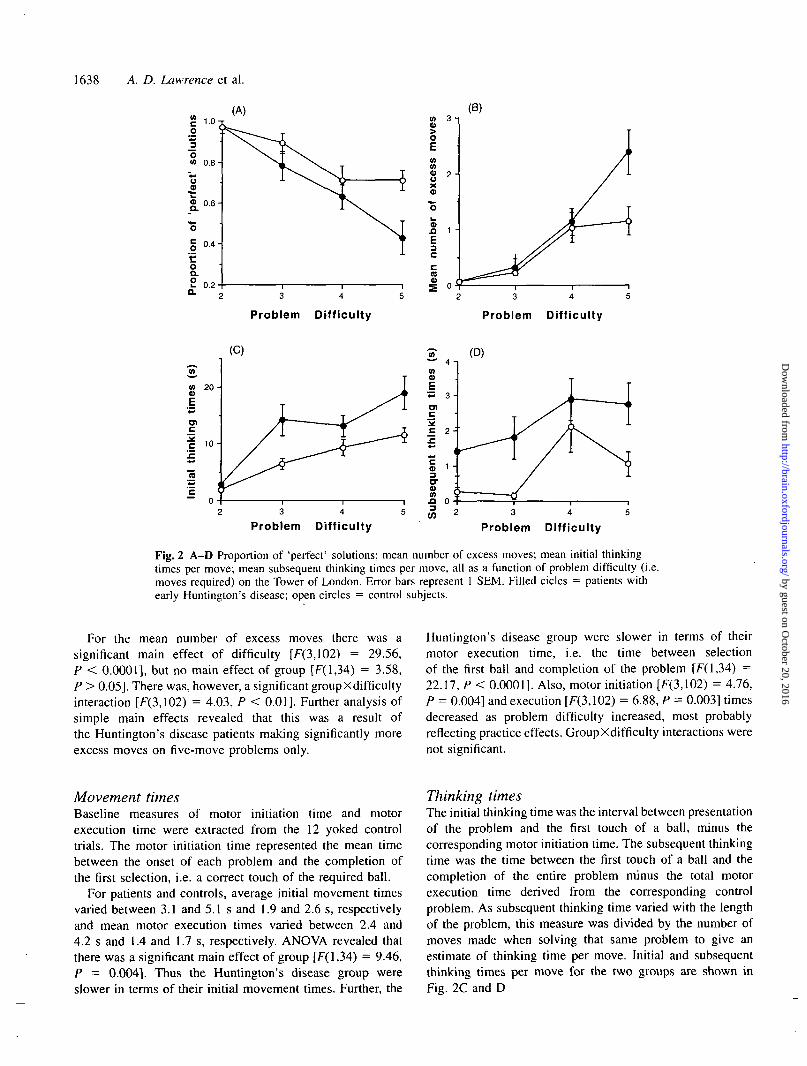
Tower of LondonAccuracy measuresAcross the 12 test problems, two measures relating toaccuracy of planning were calculated. The proportion of'perfect' solutions provided the most stringent measure ofplanning. The mean number of excess moves provideda more general measure of group performance at eachlevel of difficulty. The results for the two groups arepresented graphically in Fig. 2A and B. ANOVA revealedthat for the proportion of 'perfect' solutions there was asignificant main effect of difficulty for both groups[F(3,102) = 21.69, P < 0.0001] and a significant main effectof group [F(l,34) = 6.39, P = 0.016] but no interactionbetween group and difficulty factors, although this didapproach significance [F(3,102) = 2.31, P = 0.08].
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from
1638 A. D. Lawrence et al.
3 4
Problem Difficulty Problem Difficulty
3 4
Problem Difficulty Problem Difficulty
Fig. 2 A-D Proportion of 'perfect' solutions; mean number of excess moves; mean initial thinkingtimes per move; mean subsequent thinking times per move, all as a function of problem difficulty (i.e.moves required) on the Tower of London. Error bars represent 1 SEM. Filled cicles = patients withearly Huntington's disease; open circles = control subjects.
For the mean number of excess moves there was asignificant main effect of difficulty [F(3,102) = 29.56,P < 0.0001], but no main effect of group [F(l,34) = 3.58,P > 0.05]. There was, however, a significant groupXdifficultyinteraction [F(3,102) = 4.03, P < 0.01]. Further analysis ofsimple main effects revealed that this was a result ofthe Huntington's disease patients making significantly moreexcess moves on five-move problems only.
Huntington's disease group were slower in terms of theirmotor execution time, i.e. the time between selectionof the first ball and completion of the problem [F(l,34) =22.17, P < 0.0001]. Also, motor initiation [F(3,1O2) = 4.76,P = 0.004] and execution [F(3,102) = 6.88, P = 0.003] timesdecreased as problem difficulty increased, most probablyreflecting practice effects. GroupXdifficulty interactions werenot significant.
Movement timesBaseline measures of motor initiation time and motorexecution time were extracted from the 12 yoked controltrials. The motor initiation time represented the mean timebetween the onset of each problem and the completion ofthe first selection, i.e. a correct touch of the required ball.
For patients and controls, average initial movement timesvaried between 3.1 and 5.1 s and 1.9 and 2.6 s, respectivelyand mean motor execution times varied between 2.4 and4.2 s and 1.4 and 1.7 s, respectively. ANOVA revealed thatthere was a significant main effect of group [F( 1,34) = 9.46,P = 0.004]. Thus the Huntington's disease group wereslower in terms of their initial movement times. Further, the
Thinking timesThe initial thinking time was the interval between presentationof the problem and the first touch of a ball, minus thecorresponding motor initiation time. The subsequent thinkingtime was the time between the first touch of a ball and thecompletion of the entire problem minus the total motorexecution time derived from the corresponding controlproblem. As subsequent thinking time varied with the lengthof the problem, this measure was divided by the number ofmoves made when solving that same problem to give anestimate of thinking time per move. Initial and subsequentthinking times per move for the two groups are shown inFig. 2C and D
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from
Cognition in early Huntington's disease 1639
o
c!Eo
£co
oQ.O
1.0-
0 .9 -
0 .8 -
0 .7-
0 .6-
0.5
0.4
0 .3 -
0 .2 -
0.1 -
0.0SD SDR C_D CD COR IDS IDR EDS ECR
Stage
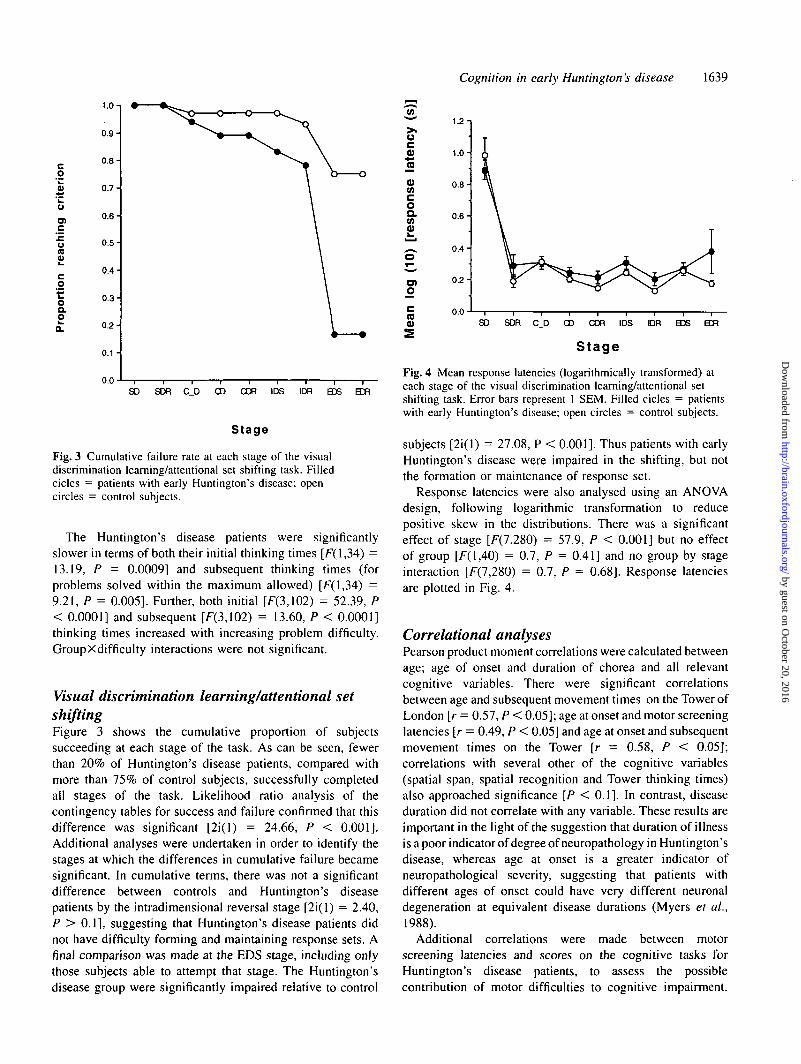
Fig. 3 Cumulative failure rate at each stage of the visualdiscrimination learning/attentional set shifting task. Filledcicles = patients with early Huntington's disease; opencircles = control subjects.
The Huntington's disease patients were significantlyslower in terms of both their initial thinking times [F(l,34) =13.19, P = 0.0009] and subsequent thinking times (forproblems solved within the maximum allowed) [F(l,34) =9.21, P = 0.005]. Further, both initial [F(3,102) = 52.39, P< 0.0001] and subsequent [F(3,102) = 13.60, P < 0.0001]thinking times increased with increasing problem difficulty.GroupXdifficulty interactions were not significant.
Visual discrimination learning/attentional setshiftingFigure 3 shows the cumulative proportion of subjectssucceeding at each stage of the task. As can be seen, fewerthan 20% of Huntington's disease patients, compared withmore than 75% of control subjects, successfully completedail stages of the task. Likelihood ratio analysis of thecontingency tables for success and failure confirmed that thisdifference was significant [2i(l) = 24.66, P < 0.001].Additional analyses were undertaken in order to identify thestages at which the differences in cumulative failure becamesignificant. In cumulative terms, there was not a significantdifference between controls and Huntington's diseasepatients by the intradimensional reversal stage [2i(l) = 2.40,P > 0.1], suggesting that Huntington's disease patients didnot have difficulty forming and maintaining response sets. Afinal comparison was made at the EDS stage, including onlythose subjects able to attempt that stage. The Huntington'sdisease group were significantly impaired relative to control
42-
50)JO0)COcoaCO0)
1.2 -i
1.0-
0 . 8 -
0 . 6 -
^ 0 . 4 -
o
n
0 . 2 -
0.0SD SDR C_D CD COR IDS IDR EDS ECR
Stage
Fig. 4 Mean response latencies (logarithmically transformed) ateach stage of the visual discrimination learning/attentional setshifting task. Error bars represent 1 SEM. Filled cicles = patientswith early Huntington's disease; open circles = control subjects.
subjects [2i(l) = 27.08, P < 0.001]. Thus patients with earlyHuntington's disease were impaired in the shifting, but notthe formation or maintenance of response set.
Response latencies were also analysed using an ANOVAdesign, following logarithmic transformation to reducepositive skew in the distributions. There was a significanteffect of stage [F(7,280) = 57.9, P < 0.001] but no effectof group [F(l,40) = 0.7, P = 0.41] and no group by stageinteraction [F(7,280) = 0.7, P = 0.68]. Response latenciesare plotted in Fig. 4.
Correlational analysesPearson product moment correlations were calculated betweenage; age of onset and duration of chorea and all relevantcognitive variables. There were significant correlationsbetween age and subsequent movement times on the Tower ofLondon [r = 0.57, P < 0.05]; age at onset and motor screeninglatencies [r = 0.49, P < 0.05] and age at onset and subsequentmovement times on the Tower [r = 0.58, P < 0.05];correlations with several other of the cognitive variables(spatial span, spatial recognition and Tower thinking times)also approached significance [P < 0.1]. In contrast, diseaseduration did not correlate with any variable. These results areimportant in the light of the suggestion that duration of illnessis a poor indicator of degree of neuropathology in Huntington'sdisease, whereas age at onset is a greater indicator ofneuropathological severity, suggesting that patients withdifferent ages of onset could have very different neuronaldegeneration at equivalent disease durations (Myers et al.,1988).
Additional correlations were made between motorscreening latencies and scores on the cognitive tasks forHuntington's disease patients, to assess the possiblecontribution of motor difficulties to cognitive impairment.
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

1640 A. D. Lawrence et al.
Only spatial span correlated significantly with motorscreening latencies [r = -0.54, P < 0.05], suggesting thatonly spatial span was affected by motor difficulties.
DiscussionIn this study we examined the cognitive performance ofpatients in the early stages of Huntington's disease on arange of tests of executive and visuospatial mnemonicfunction. These tests have already been shown to revealdeficits in late Huntington's disease which are significantlygreater than those observed in patients with Alzheimer'sdisease, matched for degree of dementia (Lange et al., 1995).The results show that even early in the course of the diseasewhen they are largely unimpaired in activities of daily living,Huntington's disease patients show a wide range of cognitiveimpairments encompassing both executive and visuospatialmnemonic functions, although the magnitude of the deficitsis smaller than in late Huntington's disease. These deficits,however, cannot be attributed to global intellectual or motordifficulties. For instance, the planning deficits on the Towerof London are only apparent at the highest levels of difficulty;there is a sparing of set formation and maintenance; and theimpairment in pattern recognition is much greater than thatin spatial recognition, as evidenced by response latencies,suggesting particular difficulties in visual object recognitionmemory. In addition, only spatial span correlated significantlywith motor screening latencies in Huntington's diseasepatients.
By comparing the performance of the early and lateHuntington's disease groups on the same tasks we are ableto obtain a profile of the progression of cognitive deficits inHuntington's disease, similar to that already reported forParkinson's disease (Owen et al., 1992). The use of a commonbattery, including tests differentially sensitive to frontal andtemporal lobe dysfunction, also allows a direct comparisonbetween these two forms of basal ganglia disorder that canbe related to the pattern of neuropathology in the striatum andits cortico-striatal circuitry. The comparison with Parkinson'sdisease shows that while the pattern of deficits in earlyHuntington's disease is somewhat similar to that of patientswith clinically severe Parkinson's disease, importantdifferences are present, notably in pattern recognitionmemory.
Staging of set-shifting deficits in Huntington 'sdiseaseThe most striking deficit seen in the Huntington's diseasegroup was in their performance on the visual discriminationlearning/attentional set shifting task. They were impairedspecifically at the EDS stage, with fewer than 20% of theHuntington's disease patients able to reach the criterion ofsix consecutive correct responses within 50 trials. This is afar greater impairment than that seen in patients with frontal
lobe lesions of a similar age (Owen et al., 1991) and iscomparable to that found in patients with clinically severeprogressive supranuclear palsy (Robbins et al., 1994). Thedeficit is also greater than that seen in unmedicated, early-in-the course patients with Parkinson's disease where, however,there is typically greater attrition in the earlier, set-formingstages of the task (Owen et al., 1992). Furthermore, patientswith mild dementia of Alzheimer type with similar scoreson the Mini-Mental State Examination are unimpaired on theEDS stage of this task, but show marked visuospatial memorydeficits, thus ruling out an explanation in terms of a globalcognitive impairment (Sahakian et al., 1990). Moreover, thedeficit was specific to the EDS stage of the task: Huntington'sdisease patients were not impaired on the earlier stages ofthe task, showing that this shifting impairment is confined tothe level of shifting between perceptual dimensions and thatpatients with early Huntington's disease do not have difficultyin the formation and maintenance of response set. Thesefindings are in agreement with Josiassen et al. (1983) whofound an increase in perseverative, but not non-perseverativeerrors on the Wisconsin Card Sorting Test in earlyHuntington's disease.
The increases in perseverative responding at the EDS stagein the early Huntington's disease patients are not observedin the reversal learning phases of this paradigm. However,in late Huntington's disease, we have shown that patientsexhibit dramatic increases in perseveration in the reversalphases of the task, preventing them from reaching the EDSstage. These deficits are truly perseverative because thepatients continue to select the formerly reinforced stimulusto a significant degree. The results from the two studiessuggest that perseverative behaviour, presumably arising froman inability to switch between response sets, is a cardinalfeature of Huntington's disease, but that its expression variesaccording to the course of the disease. It can be argued thatreversal learning and extra-dimensional shifting representtwo distinct levels of an hierarchy of inhibitory associativeand attentional mechanisms by which responding is normallycontrolled (Sutherland and Mackintosh, 1971). It appears thatin Huntington's disease this hierarchy of inhibitory processescollapses, presumably at least partly via neuropathologicalchanges at the level of the striatum, especially the head ofthe caudate nucleus. The data from Huntington's diseasepatients thus agree with the hypothesis that a major functionof the striatum is to mediate aspects. of 'response set'(Alexander etal., 1986; Chevalier and Deniau, 1990; Robbinsand Brown, 1990), including cognitive as well as motorfunctions.
Anatomical and neuropathological implicationsof set shifting deficitThe results with the attentional set-shifting task are ofadditional interest as almost exactly the same task has beenused with non-human primates. Recent findings show doubly
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

Cognition in early Huntington's disease 1641
Table 3 Tower of London
Perfect solutionsInitial thinking timesSubsequent thinking times
performance
Early HD
XX
X
in patients
NMED
V
V
with fronto-striatal dysfunction
PD MED PD(mild)
X
MEDPD(severe)
XX
V
PSP
XX
V
FLE
X
X
V = unimpaired; X = impaired. Early HD = patients with early Huntington's disease; NMED PD = unmedicated, early-in-the-eourse patientswith Parkinson's disease; MED PD (mild) = medicated Parkinson's disease patients with mild clinical disability; MED PD (severe) = medicatedParkinson's disease patients with severe clinical disability; PSP = patients with progressive supranuclear palsy; FLE = frontal lobe excisionpatients. Data from Owen et al. (1990, 1992) arid Robbins et al. (1994a).
dissociable effects of lesions to different portions of theprefrontal cortex on the extra-dimensional shifting andreversal phases of the visual discrimination learning/attentional set shifting paradigm in non-human primates.Marmosets with lesions of Brodmann area 9 exhibit deficitsin the extra-dimensional shift, whereas lesions of theorbitofrontal cortex selectively impair reversal learning (Diaset al., 1996). Anatomical evidence in monkeys suggests thatthese regions of the prefrontal cortex project to dorsal aspectsof the caudate nucleus in the case of the dorsolateral prefrontalcortex and to ventromedial regions in the case of orbitofrontalprefrontal cortex (Alexander et al., 1986; Yeterian andPandya, 1994), thus possibly constituting distinct cortico-striatal 'loops'.
These anatomical results, which suggest that the reversaldeficits might be related to the ventromedial head of thecaudate nucleus, whereas the shifting deficits may be relatedto more dorsal regions of the head of the caudate, thus fitwith what is known about the neuropathological progressionof Huntington's disease in which neuronal loss begins withthe striosome compartment of the head of the caudate andprogresses in a dorsal-to-ventral direction (Hedreen andFolstein, 1995). Striosomes in the dorsal regions of thecaudate receive input primarily from dorsal prefrontal cortex,whereas those in more ventral regions of the caudate receiveinput from limbic-related areas (Gerfen, 1992). Therefore,the dorsal-to-ventral striatal pathology in Huntington's diseaseeffectively leads to disruption of the output of thecomputations of the prefrontal cortex in a dorsal-to-ventraldirection, with those functibns associated with dorsolateralprefrontal cortex being disrupted before those associated withorbitofrontal regions.
Planning impairments in Huntington's diseaseand their cognitive componentsThe ability to plan or 'look ahead' is another centralcomponent of executive function (Tranel et al., 1994). Inprevious studies of planning in Huntington's disease, variousversions of the Tower of Hanoi task have been used (Butterset al., 1985; Saint-Cyr et al., 1988). Using a five-disc versionof the Tower of Hanoi, Butters et al. (1985) failed to findsignificant deficits in a group of early Huntington's diseasepatients. Saint-Cyr et al. (1988) did find impairments in a
small subset of four Huntington's disease patients on thefour-disc 'tower of Toronto'. However, the methods usedin both of these studies may not have provided a trueindication of the ability to plan. Goel and Grafman (1995)have argued that completion of these tasks does not necessitatethe formulation and implementation of a plan because thesolutions can be 'edited' on-line and the repetitive trial anderror learning is more akin to procedural learning than to'look ahead' planning. They agree that the present, singletrial, Tower of London task provides a purer measure ofthis 'look ahead' function in planning. The importance ofthis point is shown by the fact that across all problems, theHuntington's disease patients in this study were only impairedon the most stringent index of planning efficiency, the numberof 'perfect' solutions, and only made more excess movesthan control subjects on the most difficult five-move problems.
Patients with early Huntington's disease were also impairedin terms of their initial and subsequent thinking times, aftercorrection for motor slowing. These results are thus alsoconsistent with findings of- severe bradyphrenia inHuntington's disease (Sprengelmeyer et al., 1995). Thispattern of results is qualitatively different from that seen ineither frontal lobe excision patients (Owen et al., 1990) orin patients with other forms of basal ganglia disorder suchas Parkinson's disease, progressive supranuclear palsy ormultiple system atrophy {see Table 3). The Huntington'sdisease patients in this study can be seen to show deficits inthinking time consistent with both striatal and frontal lobedysfunction. It is unlikely that these qualitative differenceswith the other disorders simply reflect the overall severity ofthe impairment, because the impairment in producing'perfect' solutions is of a similar level to that of frontal lobeexcision patients (Owen et al., 1990). These specific deficitsin performance on the Tower of London in early Huntington'sdisease are also present in late Huntington's disease to agreater extent than in patients with dementia of Alzheimertype matched for level of dementia (Lange et al., 1995).However, there is evidence for considerable progression inthe nature of the impairment in late Huntington's disease,with markedly impaired accuracy even at two and three moveproblems. Such deficits probably reflect a breakdown in theelementary perceptual functions that contribute to higherorder planning functions.
As the planning deficit on the Tower of London task may
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

1642 A. D. Lawrence et al.
arise from any number of its required component abilities,we included separate tests of the capacity to sequenceresponses spatially, and to manipulate them in workingmemory; tests of externally ordered spatial span and self-ordered spatial working memory, as well as a test of spatialrecognition which resembles that used by Owen et al. (1996)in a PET study to activate mid-dorsolateral prefrontal cortex.We found that patients with early Huntington's disease, as isthe case for late Huntington's disease (Orsini et al., 1987;Lange et al., 1995) do indeed have impaired spatial spans.Also, in agreement with the findings of Moss et al. (1986),the Huntington's disease patients showed impaired spatialrecognition memory, as do patients with frontal but nottemporal lobe lesions (Owen et al., 1995). In contrast, onlypatients with clinically severe Parkinson's disease showedany deficits on these tasks (Owen et al., 1993).
On the spatial working memory task, the Huntington'sdisease patients made significantly more 'between-search'errors than controls but were only mildly impaired in theiruse of an effective search strategy, as in clinically severeParkinson's disease (Owen et al., 1992). Although patientswith frontal lobe lesions show a significant impairment inthe use of a search strategy, they also show more 'withinsearch' errors than control subjects, which was not the casefor the present Huntington's disease group. In contrast,amygdalo-hippocampectomized patients only showimpairments on this task at the highest levels of difficultyand are unimpaired in their use of a search strategy (Owenetai, 1995).
The advanced Huntington's disease patients of Lange et al.(1995) also showed a profound spatial working memoryimpairment, with a very high degree of perseverativeresponding which included repetition of responses withinthe same sequence ('within-search' errors). Thus, again thenature of the deficit in Huntington's disease shows adiscernible progression, in which mechanisms that exert ahierarchical control over responding, first in monitoringresponses with respect to previous self-ordered searches andthen in monitoring responses even within the same sequence,show a graded breakdown.
Anatomical and neuropathological implicationsof planning and memory deficits inHuntington's diseaseThe deficits in spatial memory in early Huntington's diseaseare perhaps not surprising in the light of primate studies (e.g.Friedman and Goldman-Rakic, 1994), which have revealedthat spatial working memory is subserved by a distributednetwork encompassing the dorsolateral prefrontal cortex andposterior parietal cortex. The dorsolateral prefrontal cortexis implicated in working memory processes (Petrides, 1994),whilst the parietal cortex is known to play a crucial rolein visuospatial processing (Ungerleider and Haxby, 1994).Moreover, a recent PET study (Owen et al., 1996) has shown
a role for these regions in the performance of spatial spanand spatial working memory tasks analogous to the onesused in this study. There are heavy projections from posteriorparietal cortex to the dorsal regions of the caudate nucleus(Baizer et al., 1993; Yeterian and Pandya, 1993). The dorsalcaudate nucleus may thus form part of the 'where' or parietalstream involved in visuospatial functions (Ungerleider andHaxby, 1994). The dorsolateral prefrontal cortex and posteriorparietal areas have common projections to the dorsal caudatenucleus (Selemon and Goldman-Rakic, 1988) and hence itis not surprising that damage to the dorsal portions of thecaudate nucleus in early Huntington's disease results inimpaired spatial working memory, again consistent with thedisruption of distinct cortico-striatal 'loops'.
The impairments in visual pattern recognition memoryseen in the current Huntington's disease patients are consistentwith the results of other studies (Moss et al., 1986; Lundervoldet al., 1994), but contrast with our findings that patients withParkinson's disease generally show no deficits on this taskat even severe levels of clinical disability, althoughimpairments are apparent in Parkinson's disease patients whoare close to meeting the clinical criteria of dementia (Owenet al., 1993). The present pattern recognition memoryimpairment has also been observed in patients with temporallobe lesions or amygdalo-hippocampectomy (Owen et al.,1995). Such deficits are associated with damage to a circuitconnecting inferotemporal cortex with the perirhinal andentorhinal cortices (Mishkin and Murray, 1994).
Connections from inferotemporal cortex project heavily tothe ventrocaudal neostriatum (ventral putamen and tail of thecaudate) (Yeterian and Pandya, 1995). Thus, the ventrocaudalstriatum may form part of an inferotemporal (or 'what')stream, processing visual objects (Ungerleider and Haxby,1994). Whilst there have been relatively few functionalstudies of these visual pathways, lesion (Divac et al., 1967;Buerger et al, 1974) and electrophysiological (Rolls, 1994;Brown et al, 1995) studies confirm they have importantfunctions in visual processing, which bear on the presentdeficit seen in Huntington's disease, as some of the earliestpathological changes in Huntington's disease have beenreported to occur in the tail of the caudate nucleus (Vonsattelet al., 1985). Although some authors have found temporallobe damage in post-mortem Huntington's disease brains (dela Monte et al., 1988; Mann et al., 1993), PET imagingstudies of early Huntington's disease patients show normaltemporal lobe metabolism (Martin et al., 1992). Hence, itseems likely that the impaired pattern recognition memoryof early Huntington's disease patients is a result of damageto the ventrocaudal striatum. A similar conclusion has recentlybeen reached by Jacobs et al. (1995), who found impairedfacial recognition in early Huntington's disease. However, inthe absence of neuroimaging data for the patients, we cannotdefinitively exclude the possibility of cortical atrophy thatmight produce a similar deficit.
In conclusion, the range of cognitive deficits seen in earlyHuntington's disease is consistent with a distinct striatal
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

Cognition in early Huntington's disease 1643
pathology which leads to deficits in tests of executive functionprobably dependent upon intact fronto-striatal function, butalso to impairment of basic cognitive functions, some ofwhich are qualitatively distinct from those in the early stagesof other basal ganglia disorders. By means of a cross-sectionalcomparison with patients with advanced Huntington's disease(Lange et al., 1995) we have shown that major features ofthe cognitive decline seen in Huntington's disease areinabilities in switching response set and failures in monitoringresponses in self-ordered memory tasks. These deficits maylead to cognitive impairments at different stages of thedisease that may appear qualitatively distinct (e.g. set shiftingversus simple reversal). However, such impairments mayreflect a gradual deterioration of mechanisms of responseinhibition operating at different levels of processing, butunder the control of fronto-striatal circuitry.
AcknowledgementsWe wish to thank Dr Niall Quinn, Institute of Neurology,London for referring patients under his care; the staff ofthe Addenbrooke's Hospital Huntington's Disease Clinic,particularly Ann Kershaw, for their invaluable assistance andall the patients for their participation in this study. Wewould also like to thank two anonymous reviewers for theircomments on the manuscript. This work was supported by aprogramme grant from the Wellcome Trust to Drs T. W.Robbins, B. J. Sahakian, B. J. Everitt and A. C. Roberts.A.D.L. is supported by an MRC studentship. K.W.L. wassupported by the Deutsche Forschungsgemeineschaft.
ReferencesAlbert ML, Feldman RG, Willis AL. The 'subcortical dementia' ofprogressive supranuclear palsy. J Neural Neurosurg Psychiatry 1974;37: 121-30.
Alexander GE, Delong MR, Strick PL. Parallel organization offunctionally segregated circuits linking basal ganglia and cortex.[Review]. Annu Rev Neurosci 1986; 9: 357-81.
Baizer JS, Desimone R, Ungerleider LG. Comparison of subcorticalconnections of inferior temporal and posterior parietal cortex inmonkeys. Vis Neurosci 1993; 10: 59-72.
Baker SC, Rogers RD, Owen AM, Frith CD, Dolan RJ, FrackowiakRSJ, et al. Neural systems engaged by planning: a PET study ofthe Tower of London task. Neuropsychologia, 1996; 34: 515-26.
Bamford KA, Caine ED, Kido DK, Cox C, Shoulson I. A prospectiveevaluation of cognitive decline in early Huntington's disease:functional and radiographic correlates. Neurology 1995; 45:1867-73.
Brown VJ, Desimone R, Mishkin M. Responses of cells in the tailof the caudate nucleus during visual discrimination learning. JNeurophysiol 1995; 74: 1083-94.
Buerger AA, Gross CG, Rocha-Miranda CE. Effects of ventralputamen lesions on discrimination learning by monkeys. J CompPhysio! Psychol 1974; 86: 440-6.
Butters N, Wolfe J, Martone M, Granholm E, Cermak LS. Memorydisorders associated with Huntington's disease: verbal recall, verbalrecognition and procedural memory. Neuropsychologia 1985; 23:729^3.
Chevalier G, Deniau JM. Disinhibition as a basic process in theexpression of striatal functions. [Review]. Trends Neurosci 1990;13: 277-80.
Cummings JL. Subcortical dementia. New York: Oxford UniversityPress, 1990.
Cummings JL. Frontal-subcortical circuits and human behavior.[Review]. Arch Neurol 1993; 50: 873-80.
David AS, Jeste DV, Folstein MF, Folstein SE. Voluntary movementdysfunction in Huntington's disease and tardive dyskinesia. ActaNeurol Scand 1987; 75: 130-9.
de la Monte SM, Vonsattel JP, Richardson EP Jr. Morphometricdemonstration of atrophic changes in the cerebral cortex, whitematter, and neostriatum in Huntington's disease. J Neuropathol ExpNeurol 1988; 47: 516-25.
Dias R, Robbins TW, Roberts AC. Dissociation in prefrontal cortexof affective and attentional shifts. Nature 1996; 380: 69-72.
Divac I, Rosfvold HE, Szwarcbart MK. Behavioral effects ofselective ablation of the caudate nucleus. J Comp Physiol Psychol1967; 63: 184-90.
Downes JJ, Roberts AC, Sahakian BJ, Evenden JL, Morris RG,Robbins TW. Impaired extra-dimensional shift performance inmedicated and unmedicated Parkinson's disease: evidence for aspecific attentional dysfunction. Neuropsychologia 1989; 29:1329^3.
Folstein SE. Huntington's disease: a disorder of families. Baltimore:Johns Hopkins University Press, 1989.
Folstein MF, Folstein SE, McHugh PR. 'Mini-mental state'. Apractical method for grading the cognitive state of patients for theclinician. J Psychiatr Res 1975; 12: 189-98.
Folstein SE, Leigh RJ, Parhad IM, Folstein MF. The diagnosis ofHuntington's disease. Neurology 1986; 36: 1279-83.
Friedman HR, Goldman-Rakic PS. Coactivation of prefrontal cortexand inferior parietal cortex in working memory tasks revealed by2DG functional mapping in the rhesus monkey. J Neurosci 1994;14: 2775-88.
Gerfen CR. The neostriatal mosaic: multiple levels of compartmentalorganization in the basal ganglia. [Review]. Annu Rev Neurosci1992; 15: 285-320.
Goel V, Grafman J. Are the frontal lobes implicated in 'planning'functions? Interpreting data from the Tower of Hanoi.Neuropsychologia 1995; 33: 623-42.
Hedreen JC, Folstein SE. Early loss of neostriatal striosome neuronsin Huntington's disease. J Neuropathol Exp Neurol 1995; 54: 105-20.
Huntington's Disease Collaborative Research Group. A novel genecontaining a trinucleotide repeat that is expanded and unstable onHuntington's disease chromosomes [see comments]. Cell 1993; 72:971-83. Comment in: Cell 1993; 72: 817-18.
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

1644 A. D. Lawrence et al.
Huntington Study Group. The Unified Huntington's Disease RatingScale. Mov Disord 1996; 11: 136-42.
Jacobs DH, Shuren J, Heilman KM. Impaired perception of facialidentity and facial affect in Huntington's disease. Neurology 1995;45: 1217-18.
Josiassen RC, Curry LM, Mancall EL. Development ofneuropsychological deficits in Huntington's disease. Arch Neurol1983; 40: 791-6.
Kendrick DC. Kendrick cognitive tests for the elderly. Windsor:NFER-Nelson, 1985.
Kullback S. Information theory and statistics. New York: Dover,1968.
Lange KW, Sahakian BJ, Quinn NP, Marsden CD, Robbins TW.Comparison of executive and visuospatial memory function inHuntington's disease and dementia of Alzheimer type matched fordegree of dementia. J Neurol Neurosurg Psychiatry 1995; 58:598-606.
Lundervold AJ, Karlsen NR, Reinvang I. Assessment of 'subcorticaldementia' in patients with Huntington's disease, Parkinson's disease,multiple sclerosis and AIDS by a neuropsychological screeningbattery. Scand J Psychol 1994; 35: 48-55.
Mann DMA, Oliver R, Snowden JS. The topographic distribution ofbrain atrophy in Huntington's disease and progressive supranuclearpalsy. Acta Neuropathol (Berl) 1993; 85: 553-9.
Martin WRW, Clark C, Ammann W, Stoessl AJ, Shtybel W,Hayden MR. Cortical glucose metabolism in Huntington's disease.Neurology 1992; 42: 223-9.
Milner B. Interhemispheric differences in the localisation ofpsychological processes in man. Brit Med Bull 1971; 27: 272-77.
Mishkin M, Murray EA. Stimulus recognition. [Review]. CurrOpinNeurobiol 1994; 4: 200-6.
Moses JA Jr, Golden CJ, Berger PA, Wisniewski AM.Neuropsychological deficits in early, middle, and late stageHuntington's disease as measured by the Luria-NebraskaNeuropsychological Battery. Int J Neurosci 1981; 14: 95-100.
Moss MB, Albert MS, Butters N, Payne M. Differential patterns ofmemory loss among patients with Alzheimer's disease, Huntington'sdisease, and alcoholic Korsakoff's syndrome. Arch Neurol 1986;43: 239^16.
Myers RH, Vonsattel JP, Stevens TJ, Cupples LA, Richardson EP,Martin JB, et al. Clinical and neuropathologic assessment of severityin Huntington's disease. Neurology 1988; 38: 341-7.
Nelson HE. National Adult Reading Test Manual. Windsor: NFER-Nelson, 1982.
Olton DS, Samuelson RJ. Remembrance of places passed: spatialmemory in rats. J Exp Psychol Anim Behav Proc 1976; 2: 97-116.
Orsini A, Fragassi NA, Chiacchio L, Falanga AM, CocchiaroC, Grossi D. Verbal and spatial memory span in patients withextrapyramidal diseases. Percept Mot Skills 1987; 65: 555-8.
Owen AM, Downes JJ, Sahakian BJ, Polkey CE, Robbins TW.Planning and spatial working memory following frontal lobe lesionsin man. Neuropsychologia 1990; 28: 1021-34.
Owen AM, Roberts AC, Polkey CE, Sahakian BJ and Robbins TW.Extradimensional versus intradimensional set shifting performancefollowing frontal lobe excisions, temporal lobe excisions oramygdalo-hippocampectomy in man. Neuropsychologia 1991; 29:993-1006.
Owen AM, James M, Leigh PN, Summers BA, Marsden CD, QuinnNP, et al. Fronto-striatal cognitive deficits at different stages ofParkinson's disease. Brain 1992; 115: 1727-51.
Owen AM, Beksinska M, James M, Leigh PN, Summers BA,Marsden CD, et al. Visuospatial memory deficits at different stagesof Parkinson's disease. Neuropsychologia 1993; 31: 627—44.
Owen AM, Sahakian BJ, Semple J, Polkey CE, Robbins TW. Visuo-spatial short-term recognition memory and learning after temporallobe excisions, frontal lobe excisions or amygdalo-hippocampectomy in man. Neuropsychologia 1995; 33: 1-24.
Owen AM, Evans AC, Petrides M. Evidence for a two-stage modelof spatial working memory processing within the lateral frontalcortex: a positron emission tomography study. Cereb Cortex 1996;6: 31-8.
Parent A, Hazrati L-N. Functional anatomy of the basal ganglia. I.The cortico-basal ganglia-thalamo-cortical loop. [Review]. BrainRes Brain Res Rev 1995; 20: 91-127.
Petrides M. Frontal lobes and working memory: evidence frominvestigations of the effects of cortical excisions in nonhumanprimates. In: Boiler F, Grafman J, editors. Handbook ofneuropsychology, Vol. 9. Amsterdam: Elsevier, 1994: 59-82.
Robbins TW. A critique of the methods available for themeasurement of spontaneous motor activity. In: Iversen LL, IversenSD, Snyder SH, editors. Handbook of psychopharmacology, Vol. 7.New York: Plenum Press, 1977: 37-82.
Robbins TW, Brown VJ. The role of the striatum in the mentalchronometry of action: a theoretical review. Rev Neurosci 1990; 2:181-213.
Robbins TW, James M, Owen AM, Lange KW, Lees AJ, LeighPN, et al. Cognitive deficits in progressive supranuclear palsy,Parkinson's disease, and multiple system atrophy in tests sensitiveto frontal lobe dysfunction. J Neurol Neurosurg Psychiatry 1994;57: 79-88.
Robbins TW, Owen AM, Sahakian BJ. The neuropsychology ofbasal ganglia disorders; an integrative cognitive and comparativeapproach. In: Ron M, David A, editors. Disorders of brain andmind. Cambridge: Cambridge University Press, 1996. In press.
Roberts AC, Robbins TW, Everitt BJ. The effects of intradimensionaland extradimensional shifts on visual discrimination learning inhumans and non-human primates. Q J Exp Psychol 1988; 40B:321^1 .
Rolls ET. Neurophysiology and cognitive functions of the striatum.Rev Neurol (Paris) 1994; 150: 648-60.
Rosser A, Hodges JR. Initial letter and semantic category fluencyin Alzheimer's disease, Huntington's disease, and progressivesupranuclear palsy [see comments]. J Neurol Neurosurg Psychiatry1994; 57: 1389-94. Comment in: J Neurol Neurosurg Psychiatry1995; 58: 520-1.
Sahakian BJ, Morris RG, Evenden JL, Heald A, Levy R, Philpot
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from

Cognition in early Huntington's disease 1645
M, et al. A comparative study of visuospatial memory and learningin Alzheimer-type dementia and Parkinson's disease. Brain 1988;111: 695-718.
Sahakian BJ, Downes JJ, Eagger S, Evenden JL, Levy R, PhilpotM, et al. Sparing of attentional relative to mnemonic function ina subgroup of patients with dementia of the Alzheimer type.Neuropsychologia 1990; 28: 1197-213.
Saint-Cyr JA, Taylor AE, Lang AE. Procedural learning andneostriatal dysfunction in man. Brain 1988; 111: 941-59.
Selemon LD, Goldman-Rakic PS. Common cortical and subcorticaltargets of the dorsolateral prefrontal and posterior parietal corticesin the rhesus monkey: evidence for a distributed neural networksubserving spatially guided behavior. J Neurosci 1988; 8: 4049-68.
Shallice T. Specific impairments of planning. Philos Trans R SocLond B Biol Sci 1982; 298: 199-209.
Sprengelmeyer R, Canavan AGM, Lange HW, Homberg V.Associative learning in degenerative neostriatal disorders: contrastsin explicit and implicit remembering between Parkinson's andHuntington's diseases. Mov Disord 1995; 10: 51-65.
Sutherland NS, Mackintosh NJ. Mechanisms of animaldiscrimination learning. New York: Academic Press, 1971.
Swerdlow NR, Paulsen J, Braff DL, Butters N, Geyer MA, SwensonMR. Impaired prepulse inhibition of acoustic and tactile startleresponse in patients with Huntington's disease. J Neurol NeurosurgPsychiatry 1995; 58: 192-200.
Tranel D, Anderson SW, Benton A. Development of the concept of'executive function' and its relationship to the frontal lobes. In:Boiler F, Grafman J, editors. Handbook of neuropsychology, Vol.9. Amsterdam: Elsevier, 1994: 125^18.
Tukey JW. Exploratory data analysis. Reading (MA): Addison-Wesley, 1977.
Ungerleider LG, Haxby JV. 'What' and 'where' in the human brain.[Review]. Curr Opin Neurobiol 1994; 4: 157-65.
Vonsattel JP, Myers RH, Stevens TJ, Ferrante RJ, Bird ED,Richardson EP Jr. Neuropathological classification of Huntington'sdisease. J Neuropathol Exp Neurol 1985; 44: 559-77.
Warner JP, Barron LH, Brock DJP. A new polymerase chain reaction(PCR) assay for the trinucleotide repeat that is unstable and expandedon Huntington's disease chromosomes. Mol Cell Probes 1993; 7:235-9.
Yeterian EH, Pandya DN. Striatal connections of the parietalassociation cortices in rhesus monkeys. J Comp Neurol 1993; 332:175-97.
Yeterian EH, Pandya DN. Laminar origin of striatal and thalamicprojections of the prefrontal cortex in rhesus monkeys. Exp BrainRes 1994; 99: 383-98.
Yeterian EH, Pandya DN. Corticostriatal connections of extrastriatevisual areas in rhesus monkeys. J Comp Neurol 1995; 352: 436-57.
Received January 18, 1996. Revised February 28, 1996.Accepted May 13, 1996
by guest on October 20, 2016
http://brain.oxfordjournals.org/D
ownloaded from




















