
Evolution of the ovenbird-woodcreeper assemblage (Aves: Furnariidae) - major shifts in nest...
13
Evolution of the ovenbird-woodcreeper assemblage (Aves: Furnariidae) / major shifts in nest architecture and adaptive radiation Martin Irestedt, Jon Fjeldsa ˚ and Per G. P. Ericson Irestedt, M., Fjeldsa ˚, J. and Ericson, P. G. P. 2006. Evolution of the ovenbird- woodcreeper assemblage (Aves: Furnariidae) / major shifts in nest architecture and adaptive radiation. / J. Avian Biol. 37: 260 /272 The Neotropical ovenbirds (Furnariidae) form an extraordinary morphologically and ecologically diverse passerine radiation, which includes many examples of species that are superficially similar to other passerine birds as a resulting from their adaptations to similar lifestyles. The ovenbirds further exhibits a truly remarkable variation in nest types, arguablyapproaching that found in the entire passerine clade. Herein we present a genus-level phylogeny of ovenbirds based on both mitochondrial and nuclear DNA including a more complete taxon sampling than in previous molecular studies of the group. The phylogenetic results are in good agreement with earlier molecular studies of ovenbirds, and supports the suggestion that Geositta and Sclerurus form the sister clade to both core-ovenbirds and woodcreepers. Within the core-ovenbirds several relationships that are incongruent with traditional classifications are suggested. Among other things, the philydorine ovenbirds are found to be non-monophyletic. The mapping of principal nesting strategies onto the molecular phylogeny suggests cavity nesting to be plesiomorphic within the ovenbird /woodcreeper radiation. It is also suggested that the shift from cavity nesting to building vegetative nests is likely to have happened at least three times during the evolution of the group. We suggest that the shifts in nest architecture within the furnariine and synallaxine ovenbirds have served as an ecological release that has facilitated diversification into new habitats and new morphological specializations. M. Irestedt (correspondence) and P. G. P. Ericson, Department of Vertebrate Zoology and Molecular Systematics Laboratory, Swedish Museum of Natural History, P.O. Box 50007, SE-104 05 Stockholm, Sweden. E-mail: [email protected]. J. Fjeldsa ˚, Vertebrate Department, Zoological Museum, University of Copenhagen, Universitetsparken 15, DK /2100 Copenhagen Ø, Denmark. The Neotropical ovenbirds (Furnariidae) exhibit a unique morphological and ecological variation among passerine birds (Leisler 1977, Vaurie 1980, Remsen 2003). The group includes many examples of species that exhibit remarkable superficial morphological similarities with other passerine birds that only are distantly related to ovenbirds, for instance the cinclodes species (Cinclodes ) resembling muscicapine thrushes (Turdidae) or dippers (Cinclidae), miners (Geositta ) resembling larks (Alaudidae), earthcreepers (Upucerthia ) resembling thrashers (Toxostoma , Mimidae) and various synallaxine genera resembling various Old World grass warblers (‘‘Sylviidae’’) or creepers (Certhiidae). Also in their placement and structure of the nest the ovenbirds exhibit an extraordinary diversity. In fact, the variation in nest types in this family has been suggested to approach that found in the entire passerine clade (Sick 1993, Collias 1997, Zyskowski and Prum 1999, Remsen 2003). Nevertheless, breeding in ‘‘closed’’ nest chambers, whether placed inside a cavity or in the vegetation, seems to unite all ovenbirds (and woodcreepers). # JOURNAL OF AVIAN BIOLOGY JOURNAL OF AVIAN BIOLOGY 37: 260 /272, 2006 260 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Evolution of the ovenbird-woodcreeper assemblage (Aves: Furnariidae) - major shifts in nest...

Evolution of the ovenbird-woodcreeper assemblage
(Aves: Furnariidae) �/ major shifts in nest architecture and
adaptive radiation
Martin Irestedt, Jon Fjeldsa and Per G. P. Ericson
Irestedt, M., Fjeldsa, J. and Ericson, P. G. P. 2006. Evolution of the ovenbird-woodcreeper assemblage (Aves: Furnariidae) �/ major shifts in nest architecture andadaptive radiation. �/ J. Avian Biol. 37: 260�/272
The Neotropical ovenbirds (Furnariidae) form an extraordinary morphologically andecologically diverse passerine radiation, which includes many examples of species thatare superficially similar to other passerine birds as a resulting from their adaptations tosimilar lifestyles. The ovenbirds further exhibits a truly remarkable variation in nesttypes, arguably approaching that found in the entire passerine clade. Herein we presenta genus-level phylogeny of ovenbirds based on both mitochondrial and nuclear DNAincluding a more complete taxon sampling than in previous molecular studies of thegroup. The phylogenetic results are in good agreement with earlier molecular studies ofovenbirds, and supports the suggestion that Geositta and Sclerurus form the sisterclade to both core-ovenbirds and woodcreepers. Within the core-ovenbirds severalrelationships that are incongruent with traditional classifications are suggested. Amongother things, the philydorine ovenbirds are found to be non-monophyletic. Themapping of principal nesting strategies onto the molecular phylogeny suggests cavitynesting to be plesiomorphic within the ovenbird�/woodcreeper radiation. It is alsosuggested that the shift from cavity nesting to building vegetative nests is likely to havehappened at least three times during the evolution of the group. We suggest that theshifts in nest architecture within the furnariine and synallaxine ovenbirds have served asan ecological release that has facilitated diversification into new habitats and newmorphological specializations.
M. Irestedt (correspondence) and P. G. P. Ericson, Department of VertebrateZoology and Molecular Systematics Laboratory, Swedish Museum of Natural History,P.O. Box 50007, SE-104 05 Stockholm, Sweden. E-mail: [email protected]. Fjeldsa, Vertebrate Department, Zoological Museum, University of Copenhagen,Universitetsparken 15, DK�/2100 Copenhagen Ø, Denmark.
The Neotropical ovenbirds (Furnariidae) exhibit a
unique morphological and ecological variation among
passerine birds (Leisler 1977, Vaurie 1980, Remsen
2003). The group includes many examples of species
that exhibit remarkable superficial morphological
similarities with other passerine birds that only are
distantly related to ovenbirds, for instance the cinclodes
species (Cinclodes ) resembling muscicapine thrushes
(Turdidae) or dippers (Cinclidae), miners (Geositta )
resembling larks (Alaudidae), earthcreepers (Upucerthia )
resembling thrashers (Toxostoma , Mimidae) and various
synallaxine genera resembling various Old World
grass warblers (‘‘Sylviidae’’) or creepers (Certhiidae).
Also in their placement and structure of the nest the
ovenbirds exhibit an extraordinary diversity. In fact,
the variation in nest types in this family has been
suggested to approach that found in the entire passerine
clade (Sick 1993, Collias 1997, Zyskowski and Prum
1999, Remsen 2003). Nevertheless, breeding in ‘‘closed’’
nest chambers, whether placed inside a cavity or in
the vegetation, seems to unite all ovenbirds (and
woodcreepers).
# JOURNAL OF AVIAN BIOLOGY
JOURNAL OF AVIAN BIOLOGY 37: 260�/272, 2006
260 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)

Clades of passerine birds differ greatly in their degree
of morphological divergence (Ricklefs 2003). Remark-
able radiations on small and species-poor landmasses,
such as islands, are often interpreted as a consequence of
competition for a limited range of food types (e.g., Grant
1986). The variation in the ovenbird clade may represent
another model, given their vast distribution in South and
Central America and the great diversity of other kinds of
birds. ‘‘Key innovations’’ may represent another possible
explanation for non-random variation in diversity
among clades. The relative success of passerine birds
has for example been postulated to be promoted by the
complexity of the passerine foot (Raikow and Bledsoe
2000) or by the ability to build nests from a wide variety
of materials (Olson 2001). The ability of ovenbirds to
construct nests of a tremendous architectural diversity
could thus be a possible explanation for their successful
colonization of a wide range of habitats, along with a
special modification of the kinetic properties of the bill,
which allowed them to extract food that was otherwise
hidden in complex vegetation structures (Fjeldsa et al.
2005).
Monophyly of the ovenbirds and woodcreepers is
inferred from their shared, unique syrinx morphology
(Ames 1971), and has also been supported by molecular
data (Sibley and Ahlquist 1990, Irestedt et al. 2002,
Chesser et al. 2004, Fjeldsa et al. 2005). Mostly based on
overall similarity ovenbirds have traditionally been
divided into three major groups, Furnariinae, Synallax-
inae and Philydorinae (i.e. Hellmayr 1925, Vaurie 1971,
1980; see Sibley and Ahlquist 1990 for a historical
review), with the woodcreepers as their closest relatives.
However, Feduccia (1973) noted cranial similarities
between certain woodcreepers and the philydorine oven-
birds, and suggested that the woodcreepers had evolved
from a philydorine ovenbird. Several recent DNA
analyses strongly support Feduccia?s hypothesis that
woodcreepers are nested within the ovenbirds, but reject
a close affinity to the philydorines. Instead, it is
suggested that the leaftossers (Sclerurus ) and miners
(Geositta ) are basal to both woodcreepers and core
ovenbirds, and that these latter two groups are recipro-
cally monophyletic (Irestedt et al. 2002, Chesser 2004,
Fjeldsa et al. 2005). The molecular studies also suggest
other relationships at odds with traditional classifica-
tions of ovenbirds: the placement of Pseudoseisura
among the synallaxines and of Lochmias among the
furnariines (as opposed to their traditional placement
among philydorine ovenbirds), and Xenops in a basal
position in the woodcreeper radiation (instead of among
philydorines) (Fjeldsa et al. 2005). These studies ob-
viously raise doubts about the monophyly of the
‘‘philydorine’’ ovenbirds.
Herein, we present a molecular phylogenetic hypoth-
esis of ovenbird relationships based on an expanded
taxonomic sample and data from two nuclear introns,
myoglobin intron 2 and G3PDH intron 11, and the
mitochondrial cytochrome b gene. Based on these
phylogenetic results we will reassess the evolution of
nest-building strategies of ovenbirds.
Materials and methods
Taxon sampling, amplification and sequencing
The ingroup in this study includes 48 ovenbirds (repre-
senting 40 out of 55 genera recognized by Remsen 2003)
and 9 woodcreepers. The ovenbirds were chosen to
represent all major subgroups previously suggested
(Vauri 1971, 1980, Feduccia 1973, Raikow 1994, Ridgely
and Tudor 1994, Zimmer and Isler 2003). In addition,
we included several species that have proven difficult to
place in either of these subgroups. A selection of
woodcreepers was also included, as recent molecular
data have shown this group to be nested within oven-
birds (Irestedt et al. 2002, Chesser 2004, Fjeldsa et al.
2005). As outgroups we used two representatives of the
family Rhinocryptidae (Pteroptochos tarnii and Scyta-
lopus spillmanni ) and one of the family Formicariidae
(Chamaeza meruloides ), which form the sister clade to
the Furnariidae (Irestedt et al. 2002, Chesser et al. 2004).
Sample identifications and GenBank accession numbers
are given in Table 1.
The complete myoglobin intron 2, the complete
glyceraldehydes-3-phosphodehydrogenase (G3PDH) in-
tron 11, and 999 bp from the cytochrome b gene have
been sequenced (see Fjeldsa et al. 2003, and Irestedt et
al. 2002 for sequencing procedures). For each gene and
taxon, multiple sequence fragments were obtained by
sequencing with different primers. These sequences were
assembled to complete sequences with SeqMan IITM
(DNASTAR Inc.). Positions where the nucleotide could
not be determined with certainty were coded with the
appropriate IUPAC code. Due to the low number of
insertions in the introns the combined sequences could
easily be aligned by eye. All gaps have been treated as
missing data in the analyses. No insertions, deletions,
stop or nonsense codons were observed in any of the
cytochrome b sequences.
Certhiaxis cinnamomea produced a partly unreadable
sequence for the G3PDH intron 11, and the PCR
product from this taxon was cloned. The cloning,
amplification and sequencing were done with the
TOPO TA Cloning† Kit (Invitrogen Life Technologies),
using the manufacturer’s primers and protocol. In one of
the two clones studied, an autapomorphic insertion was
found at the positions where the sequences from the
uncloned PCR-products became unreadable. Thus, this
insertion likely explains the reading problems in the
latter. In the phylogenetic analysis of the G3PDH intron
11 both clones were included, but as these two clones
grouped together, the G3PDH partition in Certhiaxis
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 261

Table 1. Samples used in the study. Family and subfamily names follow the classification of Remsen (2003). Abbreviations: AHMN�/American Museum of Natural History, New York;ANSP�/Academy of Natural Sciences of Philadelphia; NRM�/Swedish Museum of Natural History; ZMUC�/Zoological Museum of the University of Copenhagen. References: (1)Irestedt et al. (2001), (2) Irestedt et al. (2004b), (3) Ericson et al. (2002), (4) Fjeldsa et al. (2005), and Olson et al. (2005).
Species Subfamily/family Sample No. Cytochrome b Myoglobin G3PDH
Geositta rufipennis Furnariidae: Furnariinae ZMUC S290 AY590042 (ref. 4) AY590052 (ref. 4) AY590062 (ref. 4)Geositta tenuirostris Furnariidae: Furnariinae ZMUC S292 AY590043 (ref. 4) AY590053 (ref. 4) AY590063 (ref. 4)Upucerthia jelskii Furnariidae: Furnariinae ZMUC S439 AY065700 (ref. 1) AY065756 (ref. 1) AY590064 (ref. 4)Cinclodes fuscus Furnariidae: Furnariinae ZMUC S220 AY590044 (ref. 4) AY590054 (ref. 4) AY590065 (ref. 4)Furnarius leucopus Furnariidae: Furnariinae ZMUC 125590 AY996351 (ref. 5) AY996345 (ref. 5) AY996357 (ref. 5)Furnarius cristatus Furnariidae: Furnariinae NRM 966772 AY064279 (ref. 3) AY064255 (ref. 3) AY590066 (ref. 4)Limnornis curvirostris Furnariidae: Synallaxinae USNM B2735 AY996353 (ref. 5) AY996346 (ref. 5) AY996359 (ref. 5)Limnoctites rectirostris Furnariidae: Synallaxinae USNM B14895 AY996352 (ref. 5) AY996347 (ref. 5) AY996358 (ref. 5)Phleocryptes melanops Furnariidae: Synallaxinae USNM B2734 AY996354 (ref. 5) AY996348 (ref. 5) AY996360 (ref. 5)Aphrastura spinicauda Furnariidae: Synallaxinae ZMUC 134531 AY998188 AY998225 AY998206Leptasthenura pileata Furnariidae: Synallaxinae ZMUC S338 AY590045 (ref. 4) AY590055 (ref. 4) AY590067 (ref. 4)Schizoeaca harterti Furnariidae: Synallaxinae ZMUC S2398 AY998189 AY998226 AY998207Oreophylax moreirae Furnariidae: Synallaxinae ZMUC S128816 AY998190 AY998227 AY998208Schoeniophylax phryganophilus Furnariidae: Synallaxinae NRM 947184 AY998191 AY998228 AY998209Synallaxis ruficapilla Furnariidae: Synallaxinae NRM 956643 AY065707 (ref. 1) AY065763 (ref. 1) AY590068 (ref. 4)Synallaxis scutata Furnariidae: Synallaxinae ZMUC 125635 AY998192 AY998229 AY998210Hellmayrea gularis Furnariidae: Synallaxinae ZMUC 124846 AY998193 AY998230 AY998211Cranioleuca sulphurifera Furnariidae: Synallaxinae USNM B17199 AY996350 (ref. 5) AY996344 (ref. 5) AY996356 (ref. 5)Cranioleuca pyrrhophia Furnariidae: Synallaxinae NRM 966821 AY065708 (ref. 1) AY065764 (ref. 1) AY590069 (ref. 4)Cranioleuca albicapilla Furnariidae: Synallaxinae ZMUC 124797 AY996349 (ref. 5) AY996343 (ref. 5) AY996355 (ref. 5)Certhiaxis cinnamomeus Furnariidae: Synallaxinae NRM 937358 AY998194 AY998231 AY998212, AY998213Asthenes cactorum Furnariidae: Synallaxinae ZMUC S150 AY065705 (ref. 1) AY065761 (ref. 1) AY590070 (ref. 4)Asthenes urubambensis Furnariidae: Synallaxinae ZMUC S172 AY998195 AY998232 AY998214Phacellodomus ruber Furnariidae: Synallaxinae NRM 947206 AY590046 (ref. 4) AY590056 (ref. 4) AY590071 (ref. 4)Anumbius annumbi Furnariidae: Synallaxinae NRM 966903 AY065709 (ref. 1) AY065765 (ref. 1) AY590072 (ref. 4)Coryphistera alaudina Furnariidae: Synallaxinae NRM 966910 AY065710 (ref. 1) AY065766 (ref. 1) AY590073 (ref. 4)Metopothrix aurantiaca Furnariidae: Synallaxinae NRM 569302 AY998224Xenerpestes singularis Furnariidae: Synallaxinae ANSP 4370 AY998205Premnornis guttuligera Furnariidae: Philydorinae ZMUC 128014 AY998196 AY998233 AY998215Premnoplex brunnescens Furnariidae: Philydorinae ZMUC 124927 AY998197 AY998234 AY998216Margarornis squamiger Furnariidae: Philydorinae ZMUC S1112 AY065703 (ref. 1) AY065759 (ref. 1) AY590074 (ref. 4)Pseudoseisura lophotes Furnariidae: Philydorinae NRM 976799 AY998199 AY998236 AY998218Pseudocolaptes boissonneautii Furnariidae: Philydorinae ZMUC 124935 AY998198 AY998235 AY998217Berlepschia rikeri Furnariidae: Philydorinae ZMUC S1214 AY590047 (ref. 4) AY590057 (ref. 4) AY590075 (ref. 4)Anabacerthia striaticollis Furnariidae: Philydorinae ZMUC 124673 AY998200 AY998237 AY998219Syndactyla rufosuperciliata Furnariidae: Philydorinae ZMUC124972 AY998201 AY998238 AY998220Philydor atricapillus Furnariidae: Philydorinae NRM 937334 AY065702 (ref. 1) AY065758 (ref. 1) AY590076 (ref. 4)Thripadectes flammulatus Furnariidae: Philydorinae ZMUC S428 AY065701 (ref. 1) AY065757 (ref. 1) AY590077 (ref. 4)Automolus leucophthalmus Furnariidae: Philydorinae NRM 937251 AY590048 (ref. 4) AY590058 (ref. 4) AY590078 (ref. 4)Hylocryptus erythrocephalus Furnariidae: Philydorinae ZMUC 124862 AY998202 AY998239 AY998221Sclerurus mexicanus Furnariidae: Philydorinae ZMUC S1443 AY590049 (ref. 4) AY590059 (ref. 4) AY590079 (ref. 4)Sclerurus scansor Furnariidae: Philydorinae NRM 937258 AY065715 (ref. 1) AY065772 (ref. 1) AY590080 (ref. 4)Lochmias nematura Furnariidae: Philydorinae ZMUC S2577 AY065699 (ref. 1) AY065755 (ref. 1) AY590081 (ref. 4)Heliobletus contaminatus Furnariidae: Philydorinae ZMUC 127191 AY998203 AY998240 AY998222Xenops minutus Furnariidae: Philydorinae ZMUC S451 AY590050 (ref. 4) AY590060 (ref. 4) AY590082 (ref. 4)Xenops rutilans Furnariidae: Philydorinae ZMUC S452 AY590051 (ref. 4) AY590061 (ref. 4) AY590083 (ref. 4)Megaxenops parnaguae Furnariidae: Philydorinae ZMUC 125605 AY998204 AY998241 AY998223Pygarrhichas albogularis Furnariidae: Philydorinae AMNH PRS1128 AY065704 (ref. 1) AY065760 (ref. 1) AY590084 (ref. 4)
26
2JO
UR
NA
LO
FA
VIA
NB
IOL
OG
Y3
7:3
(20
06
)

cinnamomea in the combined analysis is represented by
the consensus sequence from both clones.
Phylogenetic inference and model selection
Bayesian inference and Markov chain Monte Carlo
(MCMC) were used for estimating phylogenetic hypoth-
esis from our DNA data (see recent reviews by Huel-
senbeck et al. 2001, Holder and Lewis 2003). The models
for nucleotide substitutions used in the analyses were
selected for each gene individually by using Akaike
Information Criterion (AIC, Akaike, 1973) and the
program MrModeltest (Nylander 2002) in conjunction
with PAUP* (Swofford 1998).
The posterior probabilities of trees and parameters in
the substitution models were approximated with MCMC
and Metropolis coupling using the program MrBayes
(Ronquist and Huelsenbeck 2003). Analyses were per-
formed for both the individual gene partitions and the
combined data set. In the analysis of the combined data
set the models selected for the individual gene partition
were used, but the topology was constrained to be the
same. One cold and three incrementally heated chains
were run for 2.5 million generations, with a random
starting tree. Trees were sampled every 100th genera-
tions, and the trees sampled during the burn-in phase
(i.e., before the chain had reached its apparent target
distribution) were discarded. Two runs, starting from
different, randomly chosen trees, were made to ensure
that the individual runs had converged on the same
target distribution (Huelsenbeck et al. 2002). After
checking for convergence, final inference was made
from the concatenated output from the two runs.
Results
Variation in the molecular data set
The sequences obtained ranged from 667 bp (Helioble-
tus ) to 716bp (Chamaeza ) in the myoglobin intron 2,
and from 287 bp (Chamaeza ) to 410 bp (Margarornis ) in
the G3PDH intron 11. Most indels observed in the two
introns were autapomorphic and mainly found in
particularly variable and repetitive regions. However,
some synapomorphic indels were observed when map-
ping the data onto the tree topologies obtained from the
Bayesian analyses of the combined data set. In the
myoglobin intron 2, all ovenbirds and woodcreepers lack
28 bp present in the outgroup (Scytalopus also lack 12 of
these bp but shares the remaining 16 bp with the other
outgroup taxon); the woodcreeper representatives share
a deletion of 12 bp; Pteroptochos tarnii and Scytalopus
spillmanni share a deletion of 13 bp and an insertion of
10 bp; Sclerurus mexicanus and Sclerurus scansor share
a deletion of 4 bp; and Philydor atricapillus andTab
le1
(Co
nti
nued
)
Sp
ecie
sS
ub
fam
ily
/fa
mil
yS
am
ple
No
.C
yto
chro
me
bM
yog
lob
inG
3P
DH
Ca
mpy
lorh
am
phu
str
och
ilir
ost
ris
Den
dro
cola
pti
da
eN
RM
94
71
83
AY
44
29
87
(ref
.2
)A
Y4
429
61
(ref
.2
)A
Y5
90
08
5(r
ef.
4)
Dec
onych
ura
lon
gic
aud
aD
end
roco
lap
tid
ae
ZM
UC
S1
24
9A
Y4
429
89
(ref
.2
)A
Y4
429
63
(ref
.2
)A
Y5
90
08
6(r
ef.
4)
Den
dro
cin
cla
tyra
nn
ina
Den
dro
cola
pti
da
eN
RM
97
66
62
AY
06
57
13
(ref
.2
)A
Y0
657
70
(ref
.2
)A
Y5
90
08
7(r
ef.
4)
Dry
mo
rnis
bri
dge
sii
Den
dro
cola
pti
da
eN
RM
96
69
30
AY
06
57
11
(ref
.1
)A
Y0
657
68
(ref
.1
)A
Y5
90
08
8(r
ef.
4)
Gly
ph
ory
nch
us
spir
uru
sD
end
roco
lap
tid
ae
ZM
UC
S1
52
1A
Y4
429
92
(ref
.2
)A
Y4
429
66
(ref
.2
)A
Y5
90
08
9(r
ef.
4)
Na
sica
lon
gir
ost
ris
Den
dro
cola
pti
da
eZ
MU
CS
183
1A
Y4
429
95
(ref
.2
)A
Y4
429
69
(ref
.2
)A
Y5
90
09
1(r
ef.
4)
Sit
taso
mus
gri
seic
apil
lus
Den
dro
cola
pti
da
eN
RM
96
70
31
AY
06
57
14
(ref
.1
)A
Y0
657
71
(ref
.1
)A
Y5
90
09
2(r
ef.
4)
Xip
ho
cola
pte
sm
ajo
rD
end
roco
lap
tid
ae
NR
M9
66
84
7A
Y0
657
12
(ref
.1
)A
Y0
657
69
(ref
.1
)A
Y5
90
09
3(r
ef.
4)
Xip
ho
rhy
nch
us
ery
thro
pyg
ius
Den
dro
cola
pti
da
eZ
MU
CS
161
6A
Y4
429
97
(ref
.2
)A
Y4
429
71
(ref
.2
)A
Y5
90
09
4(r
ef.
4)
Ch
am
aez
am
eru
loid
esF
orm
ica
riid
ae
ZM
UC
S2
05
3A
Y0
657
18
(ref
.1
)A
Y0
657
76
(ref
.1
)A
Y5
90
09
5(r
ef.
4)
Pte
rop
toch
os
tarn
iiR
hin
ocr
yp
tid
ae
AM
NH
RT
C4
67
AY
06
57
17
(ref
.1
)A
Y0
657
74
(ref
.1
)A
Y5
90
09
6(r
ef.
4)
Scy
talo
pu
ssp
illm
an
ni
Rh
ino
cryp
tid
ae
ZM
UC
S5
40
AY
06
57
16
(ref
.1
)A
Y0
657
73
(ref
.1
)A
Y5
90
09
7(r
ef.
4)
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 263

Heliobletus contaminatus share a deletion of 14 bp. In
the relatively more indel-rich G3PDH intron 11 indels
with different lengths sometimes overlap. However, if
these are treated as independent events the following
synapomorphic indels are found; Geositta rufipennis and
tenuirostris share a deletion of 23 bp; Deconychura ,
Dendrocincla and Sittasomus share two deletions of 34
and 1 bp, respectively; Furnarius cristatus and leucopus
share a deletion of 19 bp; Xenops minutus and rutilans
share a deletion of 1 bp; Pteroptochos tarnii and
Scytalopus spillmanni share a deletion of 3 bp;
all ovenbirds and woodcreepers share an extra basepair
lacking in the outgroup and in the representatives
of the genera Geositta and Sclerurus ; Anabacerthia ,
Automolus, Heliobletus, Hylocryptus, Megaxenops,
Philydor and Syndactyla share an insertion of 1 bp
(Thripadectes has an autapomorphic deletion in
that region); and Heliobletus and Philydor share an
insertion of 9 bp.
A few indels were also found to be incongruent
with the phylogenetic tree obtained from the analysis
of the combined data set. These were generally found
in the most variable regions and some of the single
basepair insertions actually consist of different bases.
However, one of these indels in myoglobin consists of
a 4 bp deletion shared between Certhiaxis cinnamomea
and Schoeniophylax phryganophila . The remaining,
phylogenetically incongruent indels were found to in
the G3PDH intron 11 and includes a 1 bp insertion in
Cinclodes, Deconychura and Premnoplex ; a 1 bp inser-
tion in Asthenes cactorum and Furnarius cristatus ; a 1 bp
deletion in Automolus and Thripadectes ; a 1 bp deletion
in Pteroptochos and Glyphorynchus ; a 10 bp deletion
in Drymornis and Xiphorhynchus ; and a 3 bp insertion in
Glyphorynchus, Campylorhamphus, Drymornis, Nasica ,
Xiphocolaptes and Xiphorhynchus.
The observed, pairwise distances (p-distances) range
between 0.3% (Limnoctites rectirostris and Cranioleuca
sulphurifera , Synallaxis scutata and Synallaxis rufica-
pilla , Cranioleuca pyrrhophia and Cranioleuca albica-
pilla ) and 8.8% (Scytalopus spillmanni and Synallaxis
scutata ) in myoglobin, 0.0% (Xenops minutus and
rutilans ) and 13.2% (Berlepschia rikeri and Scytalopus
spillmanni , Berlepschia rikeri and Chamaeza meruloides )
in G3PDH, and between 2.4% (Lochmias nematura and
Upucerthia jelskii ) and 20.3% (Chamaeza meruloides and
Synallaxis ruficapilla , Pteroptochos tarnii and Nasica
longirostris ) in cytochrome b.
Model selection and phylogenetic relationships
The priori selection of substitution models supported
that the GTR�/I�/G model had the best fit for the
cytochrome b , and the myoglobin partitions, while the
GTR�/G model was selected for the G3PDH intron 11.
These models were used in the Bayesian analyses of
the individual genes as well as in the combined analysis.
After discarding the burn-in phase the inference for
the cytochrome b gene was based on a total of 48,400
samples from the posterior, while the inference
for myoglobin and G3PDH introns were based on
47,800 samples and 47,500 samples for the combined
data set. For the phylogenetic inference, the mode of
the posterior distribution of topologies was presented
as a majority-rule consensus tree from each analysis
(Figs. 1 and 2).
The trees obtained from the Bayesian analyses of
the individual gene partitions (Fig. 1) shows a similar
degree of resolution (although the tree obtained from
the cytochrome b partition is slightly more resolved).
The gene trees are also overall topologically similar
and the non-congruent relationships are mostly at short
nodes with rather modest posterior probabilities. Phylo-
genetic relationships congruently supported by all
three genes includes a strong support for a sister
relationship between the genera Geositta and Sclerurus ;
a clade consisting of the Cranioleuca representatives
and Limnoctites rectirostris ; a core foliage-gleaners
clade (Anabacerthia , Syndactyla , Megaxenops, Helioble-
tus, Philydor, Automolus, Hylocryptus, and Thripa-
dectes ); and a clade consisting of traditional
Furnariinae ovenbirds (Furnarius, Cinclodes and Upu-
certhia ) plus the genera Lochmias, Phleocryptes and
Limnornis.
A few conflicting topologies between the individual
gene trees are supported by rather solid posterior
probabilities, but there is no indication that one gene
tree should be more different than the others. Examples
of strong topological conflicts include the relative
position of Limnoctites rectirostris. While the G3PDH
tree gives a strong support placing Limnoctites recitros-
tris with Cranioleuca albicapilla , both the myoglobin
and cytochrome b trees placed it with C. sulphurifera .
Another example is in the relative positions of Certhiaxis
cinnamomea and Synallaxis scutata , where the topology
of the G3PDH tree differs from those in the myoglobin
and cytochrome b trees. There are also examples where
the cytochrome b tree or the myoglobin tree supports
relationships different from what suggested by the
G3PDH tree. For example, the cytochrome b tree
supports a basal position of Xenops minutus and Xenops
rutilans relative to all other ovenbirds except the genera
Geositta and Sclerurus, while both the G3PDH and
myoglobin trees suggest that Xenops instead is basal
within the woodcreeper clade. The analysis of the
combined data set results in a tree (Fig. 2) that is better
resolved than the individual gene trees, although topo-
logically overall similar to them. Overall the combined
tree tends to support relationships that are supported by
a majority of the gene partitions.
264 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)

Discussion
Phylogenetic resolution
The overall topological similarity between the individual
gene trees (Fig. 1) support that the tree obtained in the
analysis of the combined data set (Fig. 2) is an overall
good estimate of the generic relationships among oven-
birds. Nevertheless, the topological disagreements that
do exist between the gene trees might cause some
concern about the accuracy of certain parts of the
combined tree.
It has been postulated that trees based on the
mitochondrial genome has a better chance to correctly
estimate avian phylogenies than single nuclear genes
(Moore 1995, Moore and DeFilippis 1997), but exam-
ples exist where mitochondrial trees yield phylogenetic
estimates that are at odd with both nuclear data and the
relationships that could be expected based on biogeo-
graphy or morphological data (Degnan 1993, Alstrom
and Odeen 2002). Furthermore, even though the ob-
served topological conflicts could be genuine, i.e., that
the different genomic partitions used herein have differ-
ent phylogenies (reviewed in e.g., Moore 1995, Maddison
1997, Mindell 1997), the conflicts might also be due to
inaccuracies of the substitution models, or in the
methods used to infer the trees. Without further evidence
from, for example, DNA sequence data obtained from
other linkage groups, morphological or behavioral
characters, it seems virtually impossible to discriminate
between individual gene trees. As the combined tree is
based on all gene partitions it tends to exhibit relation-
ships supported by a majority of these. Consequently, we
believe that the combined tree overall is a better
Fig. 1. The 50% majority ruleconsensus trees obtained from theBayesian analyses of the individualgenes. A) the tree obtained from theanalyses of G3PDH (glyceraldehydes-3-phosphodehydrogenase) intron 11data set, B) the tree obtained from theanalyses of the myoglobin intron 2 dataset, and C) tree obtained from theBayesian analyses of the cytochrome bdata set. Posterior probability valuesare indicated to the right of the nodes.
Cranioleuca albicapillaLimnoctites rectirostris
Cranioleuca pyrrhophia
Cranioleuca sulphurifera
Metopothrix aurantiaca
Certhiaxis cinnamomeus cl 2Certhiaxis cinnamomeus cl 1
Schoeniophylax phryganophilusSynallaxis ruficapilla
Asthenes cactorumPseudoseisura lophotes
Synallaxis scutataOreophylax moreirae
Schizoeaca hartertiAsthenes urubambensis
Aphrastura spinicaudaLeptasthenura pileata
Anumbius annumbiCoryphistera alaudina
Hellmayrea gularisPhacellodomus ruber
Premnoplex brunnescensLimnornis curvirostris
Phleocryptes melanopsLochmias nematura
Cinclodes fuscusUpucerthia jelskii
Furnarius cristatusFurnarius leucopus
Premnornis guttuligeraPseudocolaptes boissonneautiiMargarornis squamiger
Pygarrhichas albogularisAnabacerthia striaticollisSyndactyla rufosuperciliata
Megaxenops parnaguaeHeliobletus contaminatus
Philydor atricapillusAutomolus leucophthalmus
Hylocryptus erythrocephalusThripadectes flammulatus
Berlepschia rikeri
Nasica longirostrisXiphocolaptes major
Campylorhamphus trochilirostrisDrymornis bridgesii
Xiphorhynchus erythropygius
Glyphorynchus spirurus
Dendrocincla tyranninaSittasomus griseicapillus
Deconychura longicauda
Xenops minutusXenops rutilans
Sclerurus mexicanusSclerurus scansor
Geositta rufipennisGeositta tenuirostris
Pteroptochos tarniiScytalopus spillmanni
Chamaeza meruloides
1.000.99
1.00
0.72
0.991.00
1.00
0.520.63
0.501.00
0.90
0.69
0.95
0.960.79
0.78
0.70
0.77
0.961.00
0.96
1.000.98
0.97
1.00
1.00
0.98
1.000.99
1.00
0.98
1.00
0.78
0.790.72
0.74
0.59
0.66
1.00
0.86
0.78
0.99
1.00
1.00
1.00
A)
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 265

phylogenetic estimate than any of the individual gene
trees. In fact, we found relationships based on the
individual genes to be in conflict with those of the
combined tree mainly where nodes are short and poster-
ior probabilities modest (B/0.95).
The combined phylogeny is in many respects incon-
gruent with the traditional division of ovenbirds into
three major groups, Furnariinae, Synallaxinae and
Philydorinae (i.e. Hellmayr 1925, Vaurie 1971, 1980).
Many recent molecular studies of other passerine bird
groups (i.e. Madagascan songbirds, see Cibois et al.
2001; antbirds, see Irestedt et al. 2004a) have shown that
traditional classifications, based on overall similarities,
often overlooked natural relationships, and the present
result is therefore no great surprise. The overall good
phylogenetic agreement between the results presented
herein and other studies of ovenbirds based on DNA-
sequences partly obtained from other genes (Irestedt
et al. 2002, Chesser 2004) are, on the other hand, a
strong indication that our combined phylogeny is a good
estimate of the generic relationships of ovenbirds.
The finding that Geositta and Sclerurus are basal
positioned to both the woodcreepers and remaining
ovenbirds are, for example, also conclusively supported
by the b-fibrinogen intron 7 (Chesser 2004). In addition,
recently revealed morphological characters also support
several of these novel molecular hypotheses. Fjeldsa
et al. (2005) demonstrate that Geositta and Sclerurus
share similar skull morphologies with the so called
‘‘transitory woodcreepers’’ (Feduccia 1973), and differ
from the skull morphologies found in other ovenbirds.
An even more unexpected example, exposed by Fjeldsa
et al. (2005), is that Xenops shares with Glyphorhynchus
a unique functional system for hammering in wood. Also
a different interpretation of Clench’s (1995) study of the
pterylosis patterns of woodcreepers and ovenbirds sup-
port this relationship, as Xenops and woodcreepers
shares a lower number of feathers in the pars dorsalis
Fig. 1 (Continued )Cranioleuca sulphuriferaLimnoctites rectirostris
Cranioleuca albicapillaCranioleuca pyrrhophia
Synallaxis ruficapillaSynallaxis scutata
Schoeniophylax phryganophilusCerthiaxis cinnamomeus
Pseudoseisura lophotesOreophylax moreirae
Schizoeaca hartertiAsthenes urubambensis
Anumbius annumbiPremnornis guttuligera
Hellmayrea gularis
Limnornis curvirostrisPhleocryptes melanops
Cinclodes fuscusUpucerthia jelskii
Furnarius cristatusFurnarius leucopus
Lochmias nematura
Aphrastura spinicaudaCoryphistera alaudina
Asthenes cactorumPhacellodomus ruber
Leptasthenura pileata
Pseudocolaptes boissonneautii
Automolus leucophthalmus
Thripadectes flammulatusHylocryptus erythrocephalus
Anabacerthia striaticollisSyndactyla rufosuperciliataMegaxenops parnaguae
Heliobletus contaminatusPhilydor atricapillus
Margarornis squamigerPremnoplex brunnescens
Pygarrhichas albogularis
Berlepschia rikeri
Drymornis bridgesiiXiphorhynchus erythropygius
Campylorhamphus trochilirostris
Xenops minutusXenops rutilans
Deconychura longicaudaSittasomus griseicapillus
Dendrocincla tyranninaNasica longirostris
Xiphocolaptes majorGlyphorynchus spirurus
Geositta rufipennisGeositta tenuirostris
Sclerurus mexicanusSclerurus scansor
Pteroptochos tarniiScytalopus spillmanni
Chamaeza meruloides
1.00
1.00
0.990.90
1.00
1.00
1.00
0.73
1.000.83
0.99
1.00
1.00
1.000.93
1.00
1.00
0.90
1.00
0.950.89
0.780.96
0.860.68
0.76
1.00
0.99
1.00
1.00
0.56
0.781.00
0.590.73
0.941.00
1.001.00B)
266 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)

than found in ovenbirds. Nevertheless, other morpholo-
gical characters are sometimes in conflict with the results
presented herein. Thus, the ‘‘Margarornis assemblage’’,
as defined from characters from the hindlimb muscu-
lature (Rudge and Raikow 1992), is suggested to be non-
monophyletic by our molecular data. However, Irestedt
et al. (2004b) and McCracken et al. (1999) have found
similar conflicts between molecular data and morpho-
logical characters from the hindlimb, suggesting that
hindlimb characters may often be under adaptive selec-
tion and less useful in phylogenetic reconstructions. This
might also be case for several other morphological
characters (that have been used to infer relationships
among ovenbirds and woodcreepers) that are in conflict
with the present molecular phylogeny.
As a consequence of our much denser taxon sampling
than in previous DNA-based phylogenies of ovenbirds,
several taxa with contested affinity can now be placed,
some traditional groupings can for the fist time be
corroborated by molecular data, while other groups
could be rejected or modified. While the Furnariinae
and Synallaxinae in large parts remains intact, although
with somewhat different generic compositions, the
philydorine ovenbirds are unlikely to be monophyletic
(see also Fjeldsa et al. 2005): four ‘‘philydorine’’ clades
could be recognized; (1) a core foliage-gleaners clade
(Anabacerthia , Syndactyla , Megaxenops, Heliobletus,
Philydor, Automolus, Hylocryptus, and Thripadectes ),
(2) a clade with Margarornis, Premnoplex and Pygar-
rhichas, (3) a clade with Premnornis and Pseudocolaptes,
and (4) a clade for the aberrant Berlepschia . Except for
the Premnornis and Pseudocolaptes clade, these foliage-
gleaner clades are positioned basal to traditional furnar-
iines and synallaxines, but the combined phylogeny
suggests that they are separated by fairly long inter-
nodes. Most noticeable is the position of Premnornis and
Pseudocolaptes, which strongly suggests that the Phily-
dorinae is an unnatural group.
Fig. 1 (Continued )
Cranioleuca albicapillaCranioleuca pyrrhophia
Cranioleuca sulphuriferaLimnoctites rectirostris
Xenerpestes singularisAsthenes cactorum
Schoeniophylax phryganophilusSynallaxis scutata
Synallaxis ruficapilla
Certhiaxis cinnamomeusPseudoseisura lophotes
Oreophylax moreiraeSchizoeaca harterti
Asthenes urubambensisAnumbius annumbi
Coryphistera alaudinaHellmayrea gularis
Phacellodomus ruberLeptasthenura pileata
Premnornis guttuligeraPseudocolaptes boissonneautii
Limnornis curvirostrisPhleocryptes melanops
Lochmias nematuraFurnarius cristatus
Furnarius leucopus
Cinclodes fuscusUpucerthia jelskii
Aphrastura spinicauda
Berlepschia rikeri
Anabacerthia striaticollisSyndactyla rufosuperciliata
Philydor atricapillus
Megaxenops parnaguaeHeliobletus contaminatus
Automolus leucophthalmusThripadectes flammulatus
Hylocryptus erythrocephalus
Premnoplex brunnescensPygarrhichas albogularis
Margarornis squamiger
Xenops minutusXenops rutilans
Campylorhamphus trochilirostrisDrymornis bridgesii
Xiphorhynchus erythropygiusNasica longirostris
Xiphocolaptes majorDeconychura longicauda
Sittasomus griseicapillusDendrocincla tyrannina
Glyphorynchus spirurusSclerurus mexicanus
Sclerurus scansorGeositta rufipennis
Geositta tenuirostrisPteroptochos tarnii
Scytalopus spillmanniChamaeza meruloides
1.00
0.76
1.001.00
1.00
0.84
0.96 1.00
0.64
1.000.80
0.59
0.66
1.00
0.95
1.00
0.94
0.69
1.00
0.53
1.000.74
0.620.90
1.000.56
0.830.64
1.000.801.00
0.61
0.78
0.96
0.93
0.640.99
1.001.00
1.00
1.00
0.64
1.00
0.97
0.62 0.64
1.00
1.00
1.00C)
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 267

Ecological shifts and morphology
The new phylogenetic groupings might seem surprising
in view of the general similarities in external morphology
between ‘‘philydorine’’ ovenbirds (even though certain
genera such as Margarornis and Pygarrhichas are quite
divergent from the typical philydorines). However, based
on our combined phylogeny (Fig. 2), it is obvious that
deeper branches (except for Geositta ) comprise forest
birds of a fairly uniform appearance.
The most parsimonious solution of the ovenbird
radiation is thus an early forest diversification, leading
to divergence of woodcreepers, a large radiation of
foliage-gleaners, most of which obtained their food by
probing and prying in internodes and among masses of
dead leaves and debris suspended among vines and
branches (Fjeldsa et al. 2005), and lineages that specia-
lized to search food in epiphyte-clad cloud-forest and
shrubbery of the rising Andean mountains. Thus the
terminal radiations of Furnariinae and Synallaxinae
ovenbirds (generally adapted to more xeric life zones
and more opened landscapes) would represent more
drastic shifts of environment.
Although the Furnariinae and Synallaxinae clades
remain largely intact in the combined molecular tree,
some of the suggested relationships are novel. Most
noticeable is the placement of reedhaunters, with Lim-
nornis forming a clade with Phleocryptes, close to
Lochmias, within the Furnariinae ovenbirds, and Lim-
noctites being placed among the Synallaxinae ovenbirds.
The relationships of all these have been obscure, but the
suggested relationship between Phleocryptes and Lim-
nornis is supported by their similar nest architecture
(Zyskowski and Prum 1999). These relationships are
discussed in further detail by Olson et al. (2005).
Other forest taxa that have been difficult to place are
Megaxenops and Heliobletus, which now can be placed,
with high confidence, among the philydorine foliage-
gleaners. Cytochrome b and G3PDH data, respectively,
also provide good evidence for placing Xenerpestes and
Fig. 2. The 50% majority ruleconsensus tree obtained from theanalyses of the combined data set(G3PDH intron 11, the myoglobinintron 2 and the cytochrome b datasets). Posterior probability values areindicated to the right of the nodes.
Cranioleuca sulphuriferaLimnoctites rectirostris
1.00
Cranioleuca albicapillaCranioleuca pyrrhophia1.00
1.00
Synallaxis ruficapillaSynallaxis scutata
1.00
Schoeniophylax phryganophilus0.97
Certhiaxis cinnamomeus1.00
0.98
Asthenes cactorumPseudoseisura lophotes
0.93
1.00
Oreophylax moreiraeSchizoeaca harterti
1.00
Asthenes urubambensis1.00
0.99
Anumbius annumbiCoryphistera alaudina
1.00
Hellmayrea gularis0.70
1.00
Phacellodomus ruber
0.89
Leptasthenura pileata0.98
Aphrastura spinicauda0.96
Limnornis curvirostrisPhleocryptes melanops
1.00
Lochmias nematura0.99
Cinclodes fuscusUpucerthia jelskii
1.000.68
Furnarius cristatusFurnarius leucopus
1.001.00
Premnornis guttuligeraPseudocolaptes boissonneautii
1.00
0.90
1.00
Margarornis squamigerPremnoplex brunnescens
0.69
Pygarrhichas albogularis0.99
0.74
Anabacerthia striaticollisSyndactyla rufosuperciliata
0.98
Megaxenops parnaguae1.00
Heliobletus contaminatusPhilydor atricapillus
1.001.00
Automolus leucophthalmusHylocryptus erythrocephalus
0.97
Thripadectes flammulatus1.00
1.00
Berlepschia rikeri
1.00
Campylorhamphus trochilirostrisDrymornis bridgesii
0.95
Xiphorhynchus erythropygius1.00
Nasica longirostrisXiphocolaptes major
1.00
1.00
Deconychura longicaudaSittasomus griseicapillus
0.95
Dendrocincla tyrannina1.00
1.00
Glyphorynchus spirurus1.00
Xenops minutusXenops rutilans
1.00
1.00
1.00
Sclerurus mexicanusSclerurus scansor
1.00
Geositta rufipennisGeositta tenuirostris
1.00
1.00
1.00
Pteroptochos tarniiScytalopus spillmanni
1.00
Chamaeza meruloides
268 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)

Metopothrix among the synallaxines, near the Cranio-
leuca group, and it is therefore likely that Siptornis and
the odd, new-described Acrobatornis (Pacheco et al.
1996) also belong here.
It is finally interesting to see that the large genus
Asthenes, which is mainly associated with open montane
habitats, may not be a natural taxon. Further studies are
needed to see how many Asthenes species (and Thripo-
phaga softtails) that fall outside the main group
(represented here by A. cactorum ).
Environmental factors clearly put morphological
constraints on birds. Certain morphologies are repeat-
edly observed in obviously unrelated passerine species
that occupy forested habitats, while other morphologies
are shared by species living in open habitats. Not
surprisingly, the large and ecologically diverse
ovenbird�/woodcreeper radiation includes many exam-
ples of morphological parallels to distantly related
passerine birds with similar adaptations (Remsen
2003). Given two independent cases of cavity nesters
colonizing open habitats in the ovenbird�/woodcreeper
radiation (Geositta and the Furnariinae clade) and that
shifts from cavity nesting to building vegetative nests
have independently happened at least three times within
the ovenbird�/woodcreeper radiation (see below), it is
not surprising to also find several examples of parallel
morphological evolution within this radiation. However,
other instances of similar-looking taxa, for example the
‘‘philydorine’’ Premnoplex and Premnornis, may not be
explained by convergent evolution. Instead, the phylo-
geny suggests that the deep nodes in the ovenbird�/
woodcreeper radiation mainly consisted of forest birds
that nested in cavities and that often resemble each other
in size, plumage and shape. The superficial similarities
between various ‘‘philydorine’’ lineages may thus be due
to their shared retention of a plesiomorphic, ‘‘philydor-
ine’’ appearance.
Shifts in nest architecture as innovations in the
diversification of ovenbirds
As mentioned, ovenbirds exhibit an extraordinary di-
versity in nest placement and structure, including the
construction of various types of nests in cavities and
crevices, massive ‘‘houses’’ in clay, and vegetative
structure, which in some cases attain gigantic dimensions
and also serve as dormitories for family groups (Zys-
kowski and Prum 1999, Remsen 2003). Mapping of the
general nesting strategies (as coded by Zyskowski and
Prum 1999) onto the combined phylogeny indicates that
cavity nesting is plesiomorphic within ovenbirds (Fig. 3),
a hypothesis that accords with the frequent use of cavity-
nesting among the closest relatives of Furnariidae
(Ridgely and Tudor 1994, Krabbe and Schulenberg
2003) and with the phylogenetic results by Zyskowski
and Prum (1999). The woodcreepers often use cavities
rather low in the trees, or even subterranean (as
Sclerurinae), and have simple nest-cups, and we suggest
that this was the ancestral condition in the Furnariidae.
The shift from cavity nesting to building vegetative
nests is a rather rare evolutionary event in birds, but our
data suggest that it has happened at least three times in
the Furnariidae: One time early in the evolution of the
synallaxine clade, another time in the ancestor of some
genera within the enlarged furnariine clade, and a
third time within the ‘‘philydorine’’ Pygarrhichas �/
Premnoplex �/Margarornis clade (Fig. 3). However, note
that the synallaxine, furnariine, and Pygarrhichas �/Pre-
mnoplex �/Margarornis clades are rather terminal
lineages that share a common ancestor in our phylogeny.
An alternative explanation might thus be that this
common ancestor sometimes placed a reduced vegetative
nest in cavities and built a more advanced vegetative nest
placed in the open at other times, as do Premnoplex and
Leptasthenura . The pre-disposed ability to vary nest
architecture may then have been the major source for the
nest patterns we find within these clades today. Note also
that our phylogeny lacks Eremobius, a genus close to
Upucerthia based on morphology that builds a stick-nest
placed in the open.
It is noticeable that species that built enclosed
vegetative nests within cavities are basal in all the
lineages where this major shift in nests architecture
took places, suggesting that the shift from cavity nesting
to building enclosed vegetative nests is a gradual process.
Lochmias, which is positioned basally in the phylogeny
to the vegetative nesters Limnornis and Phleocryptes
within the furnariine clade, builds a domed vegetative
nest within a cavity. Within the ‘‘philydorine’’
Pygarrhichas �/Premnoplex �/Margarornis clade, Premno-
plex often places its domed moss nest in a cavity, and
that of Margarornis is often placed under a limb or a
rock. Aphrastura , which constitutes the most basal
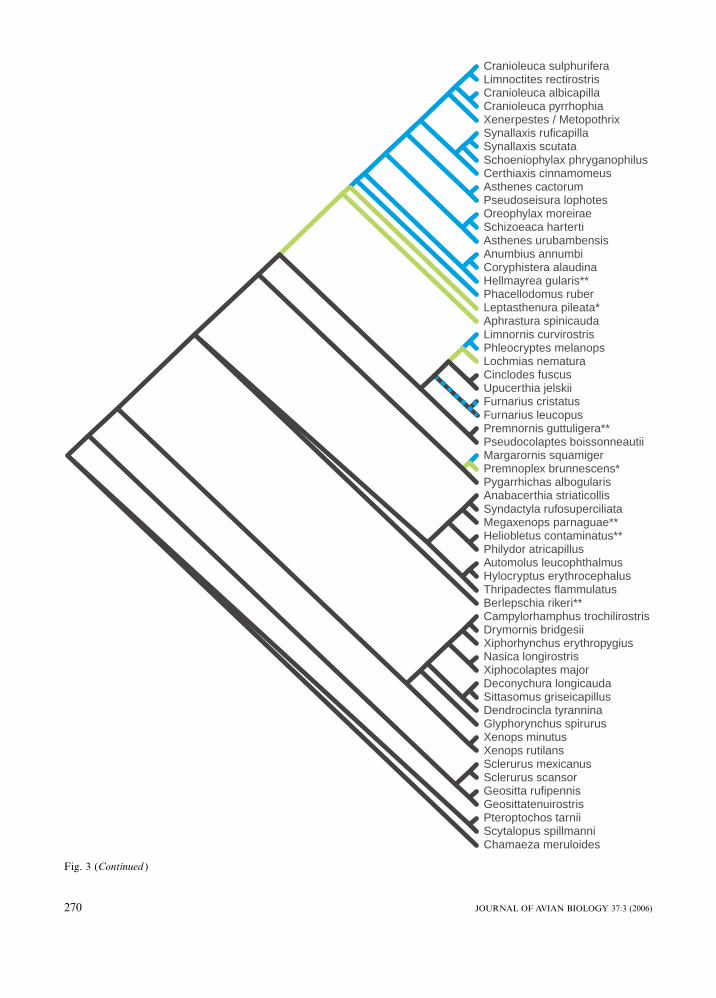
Fig. 3. Distribution of nest habits within the ovenbird-woodcreeper radiation. Principal nest strategies (nest in cavity and/or domedvegetative structure) have been mapped on the tree obtained from the analysis of the combined molecular data set. Data on nesthabits are mainly from Zyskowski and Prum (1999), and for a few taxa from Remsen (2003). Black branches indicate cavity nesting,blue branches represent the habit of building enclosed vegetative nests placed in the open, and green branches indicate that enclosedvegetative nests are placed within cavities. Note also that the black branches to the representatives of Furnarius have been markedwith blue stripes as it is difficult to tell whether the complex, domed Furnarius nest, built of clay and dung with admixed hairs andplant fibres, should be regarded as a case of cave-nesting (as coded by Zyskowski and Prum 1999) or as homologous with a domedvegetative nest. The asterisk after the names Premnoplex brunnescens and Leptasthenura pileata indicates that these taxa sometimesplace their vegetative nests in cavities and sometimes in the open. Two asterisks mark taxa for which the nest habits are insufficientlyknown. Note that Xenerpestes singularis and Metopothrix aurantiaca have tentatively been added to the tree based on their relativeposition in the cytochrome b and G3PDH trees, respectively.
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 269
Cranioleuca sulphuriferaLimnoctites rectirostrisCranioleuca albicapillaCranioleuca pyrrhophiaXenerpestes / MetopothrixSynallaxis ruficapillaSynallaxis scutataSchoeniophylax phryganophilusCerthiaxis cinnamomeusAsthenes cactorumPseudoseisura lophotesOreophylax moreiraeSchizoeaca hartertiAsthenes urubambensisAnumbius annumbiCoryphistera alaudinaHellmayrea gularis**Phacellodomus ruberLeptasthenura pileata*Aphrastura spinicaudaLimnornis curvirostrisPhleocryptes melanopsLochmias nematuraCinclodes fuscusUpucerthia jelskiiFurnarius cristatusFurnarius leucopusPremnornis guttuligera**Pseudocolaptes boissonneautiiMargarornis squamigerPremnoplex brunnescens*Pygarrhichas albogularisAnabacerthia striaticollisSyndactyla rufosuperciliataMegaxenops parnaguae**Heliobletus contaminatus**Philydor atricapillusAutomolus leucophthalmusHylocryptus erythrocephalusThripadectes flammulatusBerlepschia rikeri**Campylorhamphus trochilirostrisDrymornis bridgesiiXiphorhynchus erythropygiusNasica longirostrisXiphocolaptes majorDeconychura longicaudaSittasomus griseicapillusDendrocincla tyranninaGlyphorynchus spirurusXenops minutusXenops rutilansSclerurus mexicanusSclerurus scansorGeositta rufipennisGeosittatenuirostrisPteroptochos tarniiScytalopus spillmanniChamaeza meruloides
Fig. 3 (Continued )
270 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)

branch in the synallaxine clade, builds a cup-shaped nest
that partially covers the wall of the cavity. Leptasthe-
nura , the next branch in the synallaxine clade, builds a
domed vegetative nest that often is placed within a cavity
(Zyskowski and Prum 1999).
Building a closed nest structure inside a cavity would
immediately seem to represent unnecessary extra labour,
but may have served a function of securing less humid
conditions inside the nest. It is noteworthy that the
change in nest construction happened in lineages in-
habiting montane environments, and it is worth noting in
this respect that also Andean tapaculos (Scytalopus )
often build a closed nest of moss inside cavities. It is
unfortunate, for this interpretation, that the nest of
Hellmayrea , which inhabits humid Andean forest, is still
undescribed. Lochmias breeds near streams in montane
forests, thus in a humid environment, and Leptasthenura
spp. inhabit a broad habitat spectrum from humid tree-
lines to semiarid highlands. However, other taxa repre-
senting deeper synallaxine branches, Phacellodomus,
Coryphistera and Anumbius, mainly inhabit drier scrub
and savanna, where they assemble enormous masses of
interwoven thorny twigs, containing a system of tunnels,
and a nest chamber. It is therefore not fully clear under
what environmental conditions the shifts in nest-con-
struction happened.
The fact that the terminal synallaxine and furnariine
radiations comprise no less than 115 species and 38
species, respectively, compared with 119 species in all the
remaining lineages of Furnariidae (incl. Dendrocolapti-
nae), suggests that the shift in nest architecture may have
served as an innovation spurring further diversification.
This includes adaptations to new environments. Natural
cavities or ground patches suitable for excavating tunnels
are often in limited supply in open landscapes (Remsen
2003), and the ability to build own vegetative nest
obviously gives a competitive advantage in such environ-
ments. In case of marsh habitats, home of Phleocryptes,
Limnornis and Limnoctites, there is no possibility to find
a cavity or excavate a tunnel. In the Andean cloud-
forests a small closed nest structure can easily be hidden
in the enormous masses of epiphytes that are often
found, which may have played a role among the
synallaxines, and could also explain why Margarornis
and Premnoplex build domed moss nests.
It is noticeable that several of the nest characteristics
of ovenbirds, as coded by Zyskowski and Prum (1999),
become synapomorphies for subclades of taxa when
mapped onto our combined phylogeny. Examples are the
Sphagnum nests of Schizoeaca and Oreophylax , the
thatch placed over the camber in Certhiaxis, Schoenio-
phylax and Synallaxis, the roof adornments in Anum-
bius and Coryphistera , and the pensile nest-type of
Cranioleuca and possibly Xenerpestes (Remsen 2003, p.
319). Other nest characteristics exhibit a large degree of
convergent evolution. However, a phylogeny based on a
denser taxon sampling, as well as a more information
about nest architecture in critical groups, is needed to get
a full understanding of the evolution of nesting strategies
in ovenbirds.
Acknowledgements �/ Tissue and blood samples were mainlyobtained from the Zoological Museum of Copenhagen (withdata collecting supported by the Danish Research Councils) andthe Swedish Museum of Natural History (collected incollaboration with the Museo Nacional de Historia Naturaldel Paraguay, San Lorenzo). Important samples have also beenobtained from the American Museum of Natural History, NewYork, Academy of Natural Sciences of Philadelphia, and theNational Museum of Natural History, Smithsonian Institution.Mari Kallersjo provided logistic support and advice for thework at the Molecular Systematics Laboratory at the SwedishMuseum of Natural History, and Pia Eldenas and DarioZuccon is thanked for practical support at the laboratory.Bernd Leisler and an anonymous reviewer are thanked forvaluable comments on the manuscript. The Swedish ResearchCouncil (grant no. 621�/2004�/2913 to P.E.) funded thelaboratory work.
References
Akaike, H. 1973. Information theory as an extension of themaximum likelihood principle. �/ In: Petrov, B. N., Csaki, F.(eds). Second International Symposium on InformationTheory. Akademiai Kiado, Budapest, pp. 267�/281.
Ames, P. L. 1971. The morphology of the syrinx in passerinebirds. �/ Bull. Peabody Mus. Nat. Hist. 37: 1�/194.
Alstrom, P. and Odeen, A. 2002. Incongruence betweenmitochondrial DNA, nuclear DNA and non-moleculardata in the avian genus Motacilla : implications for estimatesof species phylogenies. �/ In: Alstrom, P. Species limits andSystematics in Some Passerine Birds. Ph. D.-thesis, UppsalaUniversity.
Chesser, R. T. 2004. Molecular systematics of New WorldSuboscine birds. �/ Mol. Phyl. Evol. 32: 11�/24.
Cibois, A., Slikas, B., Schulenberg, T. S. and Pasquet, E. 2001.An endemic radiation of Malagasy songbirds is revealed bymitochondrial DNA sequence data. �/ Evolution 55: 1198�/
1206.Clench, H. M. 1995. Body pterylosis of woodcreepers and
ovenbirds (Dendrocolaptidae and Furnariidae). �/ Auk 112:800�/804.
Collias, N. E. 1997. On the origin and evolution of nest buildingby passerine bird. �/ Condor 99: 253�/270.
Degnan, S. M. 1993. The perils of single gene trees �/
mitochondrial versus single-copy nuclear DNA variation inwhite-eyes (Aves: Zosteropidae). �/ Mol. Ecol. 2: 219�/225.
Ericson, P. G. P., Christidis, L., Irestedt, M. and Norman, J. A.2002. Systematic affinities of the lyrebirds (Passeriformes:Menura ), with a novel classification of the major groups ofpasserine birds. �/ Mol. Phyl. Evol. 25: 53�/62.
Feduccia, A. 1973. Evolutionary trends in the Neotropicalovenbirds and woodhewers. �/ Ornithol. Monogr. 13: 1�/69.
Fjeldsa, J., Zuccon, D., Irestedt, M., Johansson, U. S. andEricson, P. G. P. 2003. Sapayoa aenigma : a New Worldrepresentative of ‘Old World suboscines’. �/ Proc. R. Soc.Lond. B. (Suppl.) 270: 238�/241.
Fjeldsa, J., Irestedt, M. and Ericson, P. G. P. 2005. Moleculardata reveal some major adaptational shifts in the earlyevolution of the most diverse avian family, the Furnariidae.�/ J. Ornithol. 146: 1�/13.
Grant, P. R. 1986. Ecology and evolution of Darwin’d finches.�/ Princeton Univ. Press, Princeton N.J.
Hellmayr, C. E. 1925. Catalogue of birds of the Americas.�/ Field Mus. Nat. Hist., Zool. Ser., vol. 13.
JOURNAL OF AVIAN BIOLOGY 37:3 (2006) 271

Holder, M. and Lewis, P. O. 2003. Phylogeny estimation:Traditional and Bayesian approaches. �/ Nature Genetics4: 275�/284.
Huelsenbeck, J. P., Larget, B., Miller, R. E. and Ronquist, F.2002. Potential applications and pitfalls of Bayesian infer-ence of phylogeny. �/ Syst. Biol. 51: 673�/688.
Huelsenbeck, J. P., Ronquist, F., Nielsen, R. and Bollback, J. P.2001. Reverend Bayes meets Darwin: Bayesian inference ofphylogeny and its impact on evolutionary biology. �/ Science288: 2349�/2350.
Irestedt, M., Fjeldsa, J., Johansson, U. S. and Ericson, P. G. P.2002. Systematic relationships and biogeography of thetracheophone suboscines (Aves: Passeriformes). �/ Mol.Phyl. Evol. 23: 499�/512.
Irestedt, M., Fjeldsa, J., Nylander, J. A. A. and Ericson, P. G. P.2004a. Phylogenetic relationships of typical antbirds (Tham-nophilidae) and test of incongruence based on Bayesfactors. �/ BMC Evolutionary Biology 4: 23.
Irestedt, M., Fjeldsa, J. and Ericson, P. G. P. 2004b. Phyloge-netic relationships of woodcreepers (Aves: Dendrocolapti-nae)-incongruence between molecular and morphologicaldata. �/ J. Avian Biol. 35: 280�/288.
Krabbe, N. K. and Schulenberg, T. S. 2003. Family Formicar-iidae (Ground-antbirds). �/ In: del Hoyo, J., Elliot, A. andChristie, D. (eds). Handbook of the Birds of the World.Broadbills to Tapaculos. BirdLife International and LynxEdicions; Cambridge U.K. �/ and Barcelona 8: 682�/731.
Leisler, B. 1977. komorphologische Aspekte von Speziation undadaptiver Radiation bei Vogeln. �/ Vogelwarte 29: 136�/153.
Maddison, W. P. 1997. Gene trees in species trees. �/ Syst.�/ Biol. 46: 523�/536.
McCracken, K. G., Harshman, J., McClellan, D. A. and Afton,A. D. 1999. Data set incongruence and correlated characterevolution: An Example of functional convergence in thehind-limbs of stifftail diving ducks. �/ Syst. Biol. 48: 683�/
714.Mindell D.P. 1997. Avian molecular evolution and systematics.
�/ Academic press, San Diego, California.Moore, W. S. 1995. Inferring phylogenies from mtDNA varia-
tion: Mitochondrial-gene trees versus nuclear-gene trees.�/ Evolution 49: 718�/726.
Moore W. S. and DeFillips V. R. 1997. The window oftaxonomic resolution for phylogenies based on mitochon-drial cytochrome b. �/ In: Mindell, D. P. (ed.). AvianMolecular Evolution and Systematics. San Diego andLondon: Academic press, pp. 83�/119.
Nylander, J. A. A. 2002. MrModeltest v.1.0. Program distrib-uted by the author. �/ Department of Systematic Zoology,Uppsala University, Uppsala.
Olson, S. R. 2001. Why so many kinds of passerine birds? �/
BioScience 51: 268�/269.
Olson, S. R., Irestedt, M., Ericson, P. G. P and Fjeldsa, J. 2005.Independent evolution of two Darwinian marsh-dwellingovenbirds (Furnariidae: Limnornis, Limnoctites). �/ Orn.Neotropical 16: 347�/359.
Pacheco, J. F., Whitney, B. M. and Gonzaga, L. P. 1996. A newgenus and species of furnariid (Aves: Furnariidae) from thecocoa-growing southeastern Bahia, Brazil. �/ Wilson Bull.108: 397�/606.
Raikow, R. J. 1994. A phylogeny of the woodcreepers (Den-drocolaptinae). �/ Auk 111: 104�/114.
Raikow, R. J. and Bledsoe, A. H. 2000. Phylogeny and evolutionof the Passerine birds. �/ BioScience 50: 487�/499.
Remsen, J. V. 2003. Family Furnariidae (Ovenbirds). �/ In: delHoyo, J., Elliot, A. and Christie, D. (eds). Handbook of theBirds of the World. Broadbills to Tapaculos. BirdLifeInternational and Lynx Edicions; Cambridge U.K. �/ andBarcelona 8: 162�/357.
Ricklefs, R. E. 2003. Global diversification rates of passerinebirds. �/ Proc. R. Soc. B 270: 2285�/2291.
Ridgely, R.S. and Tudor, G. 1994. The birds of South America.�/ University of Texas Press, Austin, vol. II.
Ronquist, F. and Huelsenbeck, J. P. 2003. MRBAYES 3:Bayesian phylogenetic inference under mixed models.�/ Bioinformatics 19: 1572�/1574.
Rudge, D. W. and Raikow, R. J. 1992. The phylogeneticrelationship of the Margarornis assemblage (Furnariidae).�/ Condor 94: 760�/766.
Sibley, C. G. and Ahlquist, J. E. 1990. Phylogeny andclassification of the birds of the World. �/ Yale UniversityPress, New Haven, CT.
Sick, H. 1993. Birds in Brazil. A natural history. �/ PrincetonUniversity Press, Princeton, New Jersey.
Swofford, D. L. 1998. Paup*. Phylogenetic analysis usingparsimony (* and other methods), v.4. �/ Sinauer, Sunder-land.
Vaurie, C. 1971. Classification of the ovenbirds (Furnariidae).�/ London.
Vaurie, C. 1980. Taxonomy and geographical distribution of theFurnariidae (Aves, Passeriformes). �/ Bull. Am. Mus. Nat.Hist. 166: 1�/357.
Zimmer, K.J. and Isler. K.J. 2003. Family Thamnophilidae(typical antbirds). �/ In: del Hoyo, J., Elliot, H. and Christie,D. (eds). Handbook of the birds of the World, vol. 8,broadbills to tapaculos, Lynx Ed., Barcelona, pp. 448�/681.
Zyskowski, K. and Prum, R. O. 1999. Phylogenetic analysis ofthe nest architecture of neotropical ovenbirds (Furnariidae).�/ Auk 116: 891�/911.
(Received 3 January 2005, revised 8 April 2005, accepted9 April 2005.)
272 JOURNAL OF AVIAN BIOLOGY 37:3 (2006)




















