
Electronic Supplementary Material for “Arroyo del Vizcaino, Uruguay: A fossil-rich 30 ka...
39
Electronic Supplementary Material for “Arroyo del Vizcaino, Uruguay: A fossil- rich 30 ka megafaunal locality with cut-marked bones” Richard A. Fariña 1, *, P. Sebastián Tambusso 1 , Luciano Varela 1 , Ada Czerwonogora 1 , Mariana Di Giacomo 1 , Marcos Musso 2 , Roberto Bracco 3 , Andrés Gascue 4 1 Sección Paleontología, Facultad de Ciencias, Universidad de la República, Iguá 4225, 11400 Montevideo Uruguay 2 Sección Geotécnica, Facultad de Ingeniería, Universidad de la República, Julio Herrera y Reissig 565, 11300 Montevideo, Uruguay 3 Laboratorio 14 C, Comisión Nacional de Arqueología, Cátedra de Radioquímica, Facultad de Química, Universidad de la República, Gral. Flores 2124, 11200 Montevideo, Uruguay 4 Centro Universitario Regional Este, Universidad de la República, Burnett casi M. Chiossi (Campus Municipal), 20000 Maldonado, Uruguay * e-mail: [email protected] 1. SURVEY METHODOLOGY To resume collection after the drought in January 1997, a secondary course was built to bypass the site (Figure S1) in July 2009. In February and March 2011 plastic bags were filled with the dirt removed and then piled up to work as dams on both ends of the site. The remaining water, along with that continuously supplied by the natural aquifer, was pumped out all throughout the fieldwork, which took place in late March 2011 and more recently in late January-early February 2012. In 2011, the site was uncovered and photographed and bones exposed in an area of about 30 m 2 in the streambed, at an average depth of 2 metres. Those bones regarded as too vulnerable to be left without risk of loss were collected. A metre grid system was established to map the specimens (Figure S2). In addition, samples were taken for ongoing analyses, including grain size
Transcript of Electronic Supplementary Material for “Arroyo del Vizcaino, Uruguay: A fossil-rich 30 ka...

Electronic Supplementary Material for “Arroyo del Vizcaino, Uruguay: A fossil-
rich 30 ka megafaunal locality with cut-marked bones”
Richard A. Fariña1,*, P. Sebastián Tambusso1, Luciano Varela1, Ada Czerwonogora1,
Mariana Di Giacomo1, Marcos Musso2, Roberto Bracco3, Andrés Gascue4
1Sección Paleontología, Facultad de Ciencias, Universidad de la República, Iguá 4225,
11400 Montevideo Uruguay
2Sección Geotécnica, Facultad de Ingeniería, Universidad de la República, Julio Herrera
y Reissig 565, 11300 Montevideo, Uruguay
3Laboratorio 14C, Comisión Nacional de Arqueología, Cátedra de Radioquímica,
Facultad de Química, Universidad de la República, Gral. Flores 2124, 11200
Montevideo, Uruguay
4Centro Universitario Regional Este, Universidad de la República, Burnett casi M.
Chiossi (Campus Municipal), 20000 Maldonado, Uruguay
* e-mail: [email protected]
1. SURVEY METHODOLOGY
To resume collection after the drought in January 1997, a secondary course was built to
bypass the site (Figure S1) in July 2009. In February and March 2011 plastic bags were
filled with the dirt removed and then piled up to work as dams on both ends of the site.
The remaining water, along with that continuously supplied by the natural aquifer, was
pumped out all throughout the fieldwork, which took place in late March 2011 and more
recently in late January-early February 2012. In 2011, the site was uncovered and
photographed and bones exposed in an area of about 30 m2 in the streambed, at an
average depth of 2 metres. Those bones regarded as too vulnerable to be left without
risk of loss were collected. A metre grid system was established to map the specimens
(Figure S2). In addition, samples were taken for ongoing analyses, including grain size

and pollen content. In 2012, a more exhaustive collection was performed in an area of
about 12 m2 (Figure S2). Standard techniques have been used for field and quarry
mapping, as well as for excavating and preparing the bones [1].
Figure S1. The power shovel builds a secondary course to bypass the site.
Figure S2. Metre grid system employed to map the specimens.

2. CHRONOLOGICAL, STRATIGRAPHICAL AND
PALAEOENVIRONMENTAL CONTEXT
Radiocarbon dating is based on purified and non-purified bone collagen and on wood.
Table S1 summarizes the radiocarbon dates obtained for Arroyo del Vizcaíno yielded by
samples analysed in the following laboratories: Laboratorio 14C, Comisión Nacional de
Arqueología, Cátedra de Radioquímica, Facultad de Química, Universidad de la
República, Uruguay (URU), Beta Analytic Radiocarbon Dating, Miami FL, USA (Beta)
[2], Oxford Radiocarbon Accelerator Unit, University of Oxford, UK (Ox). Samples
labelled URU 0490, 0493 and 0496 were taken from three ribs belonging to the sloth
Lestodon armatusand sample URU 0574 was taken from a scute belonging to the
glyptodont Panochthus. All of them were pre-treated as follows: the external layers
were mechanically removed and powdered to less than 500 µm. Then, the samples were
washed with distilled water at 80 ºC for 30 minutes, and ultrasonically bathed (4 times).
The inorganic fractions were removed with 8% HCl and the organic fractions were
neutralised washing with successive distilled water washes. They were treated for 24
hours with 0.5 M NaOH and the organic fractions were again neutralised with the same
procedure. Afterwards, the organic fractions of two of the samples (URU 0493 and
URU 0496) were purified as in refs. [3-5]. For control purposes, and to compare with
previous datings of material from the same site [2], the organic fraction of the sample
0490 was not purified. In all cases, the organic fraction recovered was above 8%. The
pieces of wood were pre-trated with HCl (8%)-NaOH (2%)- HCl (2%) and then the
fractions G, H and W were extracted following ref. [6]. Sample OxA-V-2474-10 was
treated following refs.[7-9].

Table S1. Samples dated with radiocarbon techniques. Bones with an asterisk (clavicle [46] and ulna) have marks that show features similar to those made by tools. Ages are given in years before present.
Material
sampled
Laboratory
and sample
number
δ13C C:N
ratio
Percentage
of extracted
collagen
Conventional
radiocarbon
age
Calendar
dates
(Oxcal 4.1)
Lestodon rib URU 0496 -18.5 N/A ≥ 5% 27,000±450 29,696±871
Lestodon rib URU 0490 -18.5 N/A ≥ 5% 27,200±900 30,433±2033
Lestodon rib Beta 204256 -18.6 N/A N/A 28,300±230 30,580±747
Lestodon
clavicle*
Beta 206660 -18.8 N/A N/A 29,150±290 31,895±745
Undetermined
wood
URU 0562 -
25.74
-- -- 29,150±320 31,872±789
Panochthus
scute
URU 0574 -20.6 2.9 ≥ 5% 29,220±300 31,939±728
Undetermined
wood
URU 0561 -
25.74
-- -- 29,350±315 32,009±715
Lestodon rib URU 0493 -18.5 -- ≥5% 30,100±600 32,886±1446
Lestodon
ulna*
OxA-V-
2474-10
-
18.72
3.2 ≥1% 28,760±210 31,561±925
Average 28,692 31,430
Pooled
average
29,000 31759
Median 29,150 31,872

Usual description methods were heeded for the geological setting [10]. The geological
model of the deposits is shown in Figure 1C. Along the stream, ponds are formed as
microbasins excavated by differential erosion on the Mercedes Fm. Cretaceous
sandstones during late Pleistocene. The prevalent climatic conditions must have been
arid to semi-arid. Sediment grain size composition in beds 1 and 2 is similar to that of
the nearby outcrops of Mercedes Fm. (Figure S3). The sediment green-greyish colour of
the bed 2 facies a is attributed to reducing environment and the brownish colour of bed
2 facies b is proposed to have developed by moderate oxidation in post-depositional
events. Still, a more complete understanding of this difference, i.e., whether those facies
correspond to separate events, will be available when larger areas of the site are more
thoroughly excavated.
The depositional environment must have included episodes of different fluvial energy,
capable of carrying pebbles, cobbles and medium to coarse sand. This corresponds to a
flow velocity of 300 to 10 cm s-1 at 1 metre depth to erode and move the particles and a
flow velocity of 100 to 1 cm s-1 to deposit them [11]. Substantial mass flow can be ruled
out as the accumulation agent due to the rather homogeneous grain size, the lack of the
typical size gradient distribution of the remains [12], the lack of predominant orientation
in the material [13] (which is also well preserved) and the very gentle relief in that
region, with just a few metres of height difference in the surrounding area. Besides, the
fossiliferous sediments seem to have been formed in situ, as evidenced by the presence
of intraclasts from the underlying bed in them.
The silicified sandstones of the Mercedes Fm. constitute the palaeorelief as an irregular
plain, with small ponds temporarily flooded. The aquifer must have been already active,
even in times of dry climate between 40 and 30 kybp, which coincides with the final
stages of the marine isotope stage 3. This must have produced reducing conditions in

the base of the fossiliferous bed 2 (facies a) and the relatively more oxidative conditions
in the upper facies b may have been due to postdepositional expositions. The presence
of coarse grains must have been due to high-energy events, and the lack of
intermediately sized grains can be explained by the lack of availability of them in the
Mercedes Fm. regolith. Those pebbles must have transported as bed load together with
the transported bones, perhaps in more than one event. Some of the bones show rather
large clasts (~ 5 cm in diameter) in their natural cavities, such as the spinal canal (Figure
S4). Incidentally, this phenomenon yielded no marks on the bone. Turbulence could
have been present since the deposition took place in an area lateral to the mainstream.
Figure S3. Granulometric analysis of the sediments of the Arroyo del Vizcaíno samples.

Figure S4. Lestodon vertebra showing pebbles in the spinal canal.
3. FAUNAL LIST
Specimens collected
Lestodon armatus: 18 astragali; 183 vertebrae; 2 talus; 15 teeth; 45 metapodials; 3
clavicles; 12 ribs and rib fragments; 14 fragments of skull; 10 scapulae; 31 phalanges;
33 complete or broken femora; 1 hyoid; 5 fibulae; 14 fragments of mandibular rami; 8
teeth; 10 radii or fragment radii; 18 pelvic girdle fragments; 23 complete or broken
tibiae.
Mylodon darwinii: 4 teeth; 1 mandibular ramus; 1 astragalus.
Glossotherium robustum: 2 teeth; 2 vertebrae
Glyptodonts: 1 atlas; 1 calcaneum; 1 fragment of vertebral column; 1 fragment of
cranium; 1 fragment of scapula; 1 femur; 1 humerus; 5 pelvic girdle fragments; 1 tibia;
1 cervical tube; 1 sacral tube; 1 ulnar fragment; 2 vertebrae; 1 trivertebral element; 5
carapace fragments and 3 isolated scutes; 1 cranium and 1 radium assigned to

Glyptodon cf. clavipes; 203 scutes of Glyptodon clavipes, 58 scutes of Doedicurus
clavicaudatus and 26 scutes of Panochthus tuberculatus.
Toxodon platensis:1 vertebra; 1 tibia; 1 cranium; 2 scapulae
Stegomastodon sp.: 1 tooth; 1 fragment of scapula; 1 vertebra; 1 pelvic girdle fragment
Cervidae indet: 1 humerus, 1 calcaneum
Hippidion principale: 1 tooth; 1 astragalum; 1 phalanx; 1 vertebra.
Smilodon populator: 1 canine, 1 cranium fragment
Fossil ground sloths: 1 rib fragment; 2 metapodial; 4 teeth; 2 fragment of scapula; 11
phalanges; 1 fragment of femur; 1 mandibular ramus fragment; 1 patella; 1 tibia
fragment; 4 vertebrae.
Mammaliaidentified specimens not assigned to any species): 110 rib fragments; 11
cranium fragments; 12 fragments of scapula; 21 fragments of femur; 2 of humeri
fragments; 1 mandibular ramus fragment; 2 metapodial; 13 fragments of pelvic girdle; 1
radius fragment; 1 patella fragment; 3 fragments of sacrum; 1 tibia fragment; 4
vertebrae.
Mammalia indet: 34 fragments
4. TAPHONOMY
The biostratinomical characterization of the vertebrate assemblage follows ref. [14]. The
number of specimens (NISP), minimum number of anatomical units (MAU), minimum
number of individuals (MNI) and minimum number of elements (MNE) were
established. Weathering and transport evidence were assessed through macroscopical
analyses. Unless otherwise stated, counts and percentages refer to a total with
glyptodont scutes excluded. The taphonomic features analysed include: number of
specimens and individuals (NISP and MNI counts), number and characterization of the

species represented (see above the faunal list), relative abundance of the species, body
size, age spectra, skeletal articulation, representation of skeletal parts, bone orientation
and bone modifications (this last feature will be detailed in the next section).
In the portion of the bonebed covered in our excavations, the bones were distributed in a
roughly oval pattern and contained more than one thousand elements (see faunal list).
The minimum number of individuals was established as follows: for each skeletal
element (a bone or set of bones, such as the cervical vertebrae), the total number of
identified specimens (NISP) was counted and the number of individual animals (NIA,
or MNI taking only one skeletal portion into account [15]) was calculated. Seventeen
right femora were collected or observed and all of them were assigned to adult or
subadult specimens of Lestodon armatus (hence, MNI=17; table S2). In addition, at
least two juvenile specimens are represented by a hyoid, a mandible, one tibia, four
femora and one rib fragment. The age estimated for the individuals preserved in the
assemblage was classified within three major groups: juveniles (derived from the
absence of the epiphyses), prime adults and old individuals (due to the presence of
osteoarthritical signs).

Table S2. Body mass, NISP (counted as the clearly identifiable specimens), abundance and MNI of the species found in Arroyo del Vizcaíno.
Taxon Body mass
[47,48] NISP
MNI
adults juveniles
Lestodon armatus 4000/1000* 452/5 15 2
Mylodon darwinii 1500 5 1 0
Glossotherium robustum 1500 3 1 0
Glyptodon cf. clavipes 1000† 23/1 0 1
Doedicurus clavicaudatus 1400 1 0
Panochthus tuberculatus 1100 1 0
Toxodon platensis 1100 5 1 0
Stegomastodon sp. 1500* 3 0 1
Hippidion principale 119* 4 0 1
Smilodon populator 400 2 1 0
Cervidae indet. 50 2 1 0
TOTAL MNI 22
(81.5%)
5
(18.5%)
* The elements assigned to these species must have belonged to a juvenile, hence this
mass estimate.
† The juvenile element of a glyptodont counted here is tentatively assigned to
Glyptodon cf. clavipes, but the other two species cannot be ruled out.
The index of anatomical representation [16]indicates the percentage of skeletal elements
preserved in relation to the sum of the total number of elements for all individuals

identified in the assemblage, providing information of the taphonomic biases affecting
the composition of the assemblage. This index (Ir=NISP x 100/MNE x MNI) was
calculated only for L. armatus and also for the glyptodonts, the best represented taxa in
the faunal association. The obtained results, 12.8% for Lestodon armatus and 3.2% for
the glyptodonts, allow the exclusion of catastrophic death or mass mortality event as the
source of the deposit. In those cases, this index would have taken its maximum value for
each represented species (100%) if an immediate burial had occurred, followed by non-
destructive diagenetic conditions [16], which is the case for our site (see below).
The degree of disarticulation and scattering of the bones was analysed in the field
during the collection of the remains. Articulation was considered as “two or more
skeletal elements being in their proper anatomical positions relative to one another, and
within a centimetre of each other if not in fact touching” [17] and scattering as
“increasing the spatial distance between anatomically related bones” [17]. In this sense,
apart from a small group of vertebra and a rib (Figure S5), most of the specimens were
found disarticulated, although in a number of cases bones of similar anatomical regions
were found scattered but in close spatial relation. This is especially true for many bones
of possibly one specimen of Lestodon armatus yet to be collected that are found not
much dispersed from their original arrangement.

Figure S5. A small group of vertebrae and a rib found articulated in the bone
assemblage.
The weathering [18]observed in more than 80% of the remains corresponds mostly to
stages 0 and 1, with the bone surfaces either smooth or cracked parallel to the fibrous
structure of the bone, which means the bones were exposed before permanent burial for
less than 3 years. Congruently, bones show a high proportion of collagen (Table S1)
[19]. Less than 20% of the bones exhibit weathering stage 2 (Fig. S6).
There is no evidence of compaction fractures during the fossildiagenesis. Fossilization
is homogeneous between elements and also most individual bones show no difference
in their regions, which is interpreted as their having undergone no reworking [20].

Figure S6. A) CAV 16, an astragalus of Mylodon darwinii, showing the usual fine
preservation in the studied site, while B) CAV 369, an astragalus of Lestodon, shows a
weathering stage 2, is cracked mainly in the sticking out regions like the odontoid
process, is rounded and shows some trampling.
To investigate transport due to fluvial action, the relative abundance of Voorhies
Groups [21] was analysed and rose diagrams for orientation of long bones [22,17] were
built up (Figure 2c). In the discrimination of the represented specimens, glyptodont
scutes were not considered in the NISP counts. We found 53% bones belonging to
Group I and 33% and 14% belonging to Groups II and III, respectively. The distribution
and orientation of these bones was quantified on images and direct observations
gathered during fieldwork. No preferential orientation was registered for the long or
elongated bones examined (Figure 2c). Further arguments against major transport [17]
are that most of the bones show no evidence of rolling (88.9%, including glyptodont
scutes in this count, NISP=988) and exhibit minimum abrasion; the state of preservation
is very good and most of the bones are included in fine sediments, composed of clay
and silt (see above: Sediments, profile, environment section). However, the existence of
more than one population of bones cannot be assessed at the present state of knowledge,

as already said. Moreover, in the collected specimens the most transportable elements of
the megamammals are present (Voorhies group I), but not the least transportable
elements of smaller individuals (Voorhies group II and III). Taking into account the
faunal composition of the assemblage, where mostly megamammals are present, it
should be noticed that the fluvial transport index (FTI) calculated from experiments
with individual skeletal elements of an Indian elephant (Elephas maximus) [23] yielded
comparable results to those obtained according to Voorhies Groups [21]
Among the identified specimens (table S3), limb bones and girdles are predominant
(41.1%, including a small number of metapodials, 4.8%), followed by vertebrae
(33.2%), rib portions (15.8%) and cranial fragments (9.2%). These proportions are also
interpreted as the accumulation being autochthonous, as already evidenced by the
Voorhies groups, although some outgoing hydraulic transport might have removed the
lighter elements (scutes, not considered in the proportions of the Voorhies groups, and
metapodials).
The observed frequencies of vertebrae, ribs, limbs and girdle bones, and phalanges,
were compared with the data of different modern and fossil assemblages summarized in
ref. [16] (Figure 3a). The values show the Arroyo del Vizcaíno site very close to the
relative frequencies of the bones exposed in Amboseli [24] and among the group of
values that points out a biogenic accumulation of the bones, in particular, of various
sites associated with human presence: the modern San hunter-gatherer camp and some
of the Plio-Pleistocene archaeological sites at Koobi Fora, Kenya (FxJj50, Olduvai
Gorge, Tanzania, and FLK Zinjantropus site [25, 26]). These African sites had been re-
evaluated from a taphonomic perspective [26], including the comparison with present-
day hunter-gatherers. For example, the Kua San in the East Central Kalahari Desert in
Botswana are known to dismember and transport very large (several hundred kg) entire

carcasses for several kilometres. At FLK Zinjanthropus, the dominant, but not the only
cause of bone accumulation at the site, is the transport and disposal of bones by humans.
However, gnawing by carnivores and small rodents in low proportions (compared to
modern animal dens), removals, and perhaps additions of bones by carnivores, among
biogenic modifications, including trampling, were also present. In the Arroyo del
Vizcaíno site a similar level of taphonomic complexity could be invoked as responsible
for the bone accumulation, excluding the action of carnivores and rodents due to the
absence of these kinds of marks on the bones (see next section).
Likewise, the alternative arguments attributing the observed patterns of skeletal
representation to post-depositional processes (i.e. biogenic accumulation) should be
strengthened with the evidence of subaerial weathering damage on surviving bones [26].
This should be extensive enough to account for the complete disintegration of less
durable (i.e., axial and epiphyseal) elements, thus implying that geological processes
altered the skeletal proportions in the bone assemblage by differential water transport of
certain elements or sediment compaction and differential destruction of buried skeletal
elements [26]. However, it does not seem likely that these processes played a dominant
role in forming the observed patterns of skeletal representation in the Arroyo del
Vizcaíno site. First, the previously-discussed weathering characteristics of the
assemblage indicate a prevalence (more than 80%) of unweathered to moderately
weathered bones, which would be inconsistent withsufficient subaerial exposure to
eliminate less resistant elements [18]. Moreover, the patterns of long bone orientation
and Voorhies groups argue against water transport being a major determinant of skeletal
composition.
In the case of Amboseli [24], the bone sample gives a more accurate representation of
the larger species due to taphonomic biases, with higher representation of animals of

more than 200 kg, as is the case observed for the Arroyo del Vizcaíno site. The
Amboseli bone assemblage reflects the relative numbers of the ten most abundant large
wild herbivores; however, the rhinoceros population show higher than expected values
due to human predation. The body size biases of the smaller bones in this assemblage
are related basically to two factors: more complete initial destruction by carnivores and
scavengers, and more rapid rates of weathering and trampling causing fragmentation
[24]. These biases could account at least partially for the observed bone concentration in
the Arroyo del Vizcaíno site, in particular for the abundance of very large species (see
below).
Table S2 shows the taxonomic composition and relative abundance of the assemblage,
in which herbivores are predominant. The relative abundance was estimated following
ref. [27] and the distributions were corrected to avoid taphonomic biases depending on
body size and differential preservation [28]. The obtained values for linear regressions
of abundance and body mass plots did not fit the expected results for an unbiased
assemblage (y= 0.094x + 2.068, r2 non significant). Most of the represented taxa are
xenarthrans (six out of eleven species), whose body size exceeds the tonne. This faunal
composition resembles the typical association described for the Lujanian (late
Pleistocene) in the current Río de la Plata area, dominated by megamammals
andxenarthrans [29, 30]. Table S3 shows the representation of the anatomical regions,
whose results, as discussed in the main text, are compatible with the removal of
consumable parts.
Finally, the faunal assemblage has little evidence of root etching. If the amount of root
etching is considered a function of the rapidity of burial [31], this evidence is congruent
with a fast burial. Additional evidence suggesting that the bones lay exposed on the
surface for a brief period comes from the absence of modifications by scavengers or

rodents (see below for a detailed characterization of surface modifications on the
bones).
Table S3. Anatomical regions of the specimens represented in the collected material.
Anatomical region Number of elements collected Observed percentage
Skull (including lower jaws and teeth) 75 9.3
Axial skeleton (vertebrae) 274 33.8
Axial skeleton (ribs) 128 11.6
Limb skeleton (including girdles) 294 36.3
Metapodials 39 4.8
Table S4 shows the values of several variables (MNI= minimum number of individuals
based on femora and taking into consideration laterality, size and epiphyses fusion;
MAU= Minimal Animal Units; %MAU= Standardized MAU; Exp. MNE (MNI)=
expected MNE based on MNI; %S= percentage of survival) of the bone representation
belonging to ground sloths, which were used as an input for Fig. 3. In table S5, the
observed frequencies of the remains of the ground sloths are compared with the
frequencies to be expected if all anatomical regions were equally represented. Both Χ2
and G-tests show that there exist significant differences between observed and expected
frequencies. The Χ2 indicates that the main elements contributing to the difference are
femora and tibiae. On the other hand, the values of the G-test show that those elements
present a positive deviation, i.e., they are overrepresented in the sample, while other
elements (in red) show a negative deviation, i.e., they are underrepresented. Those
results are congruent with the survival percentage (%S) and the %MAU (Table S4 and
Fig. 3).

Table S4. Bones assigned to ground sloths (mainly Lestodon armatus) in the bone assemblage of Arroyo del Vizcaíno. Values of %MAU are represented in Fig. 3b. Abbreviations: MNI= Minimum Number of Individuals (Based on femora and taking into consideration laterality, size and epiphyses fusion). MAU= Minimal Animal Units [49].%MAU= Standardized MAU. Exp. MNE (MNI)= expected MNE based on MNI. %S= percentage of survival [50].
Elements NISP MNE MAU %MAU Expected MNE
(MNI) %S
Skulls 25 6 6.00 48.00 17 35.29
Mandibles 20 14 7.00 56.00 17 52.94
Hyoids 2 1 0.50 4.00 34 2.94
Vertebrae (region not
determined)
19
Cervical vertebrae 25 24 3.43 27.43 119 20.17
Dorsal vertebrae 117 70 4.38 35.00 272 25.74
Lumbar vertebrae 11 6 3.00 24.00 34 17.65
Sacra 7 7 7.00 56.00 17 41.18
Caudal vertebrae 83 67 2.68 21.44 425 15.76
Ribs 119 59 2.11 16.86 476 12.39
Ventral Ribs 9 5 0.42 3.33 204 2.45
Sterna 1 1 0.14 1.14 119 0.84
Clavicles 3 3 1.5 12.00 34 8.82
Scapulae 23 8 4.00 32.00 34 23.53
Pelves 25 13 6.50 52.00 17 41.18
Humeri 20 12 6.00 48.00 34 35.29
Radii 11 10 5.00 40.00 34 29.41
Ulnae 10 10 5.00 40.00 34 29.41
Carpals 11 11 0.79 6.29 238 4.62
Metacarpals 14 14 1.40 11.20 170 8.24
Femora 61 25 12.50 100.00 34 73.53
Patellae 6 6 3.00 24.00 34 17.65
Tibiae 28 24 12.00 96.00 34 70.59
Fibulae 5 5 2.50 20.00 34 14.71
Astragali 16 16 8.00 64.00 34 47.06
Calcanei 2 2 1.00 8.00 34 5.88
Tarsals 10 10 1.25 10.00 136 7.35
Metatarsals 8 8 1.00 8.00 136 5.88
1st phalanxes 10 10 0.83 6.67 204 4.90
2nd phalanxes 22 22 2.75 22.00 136 16.18
3rd phalanxes 13 13 1.30 10.40 170 7.65
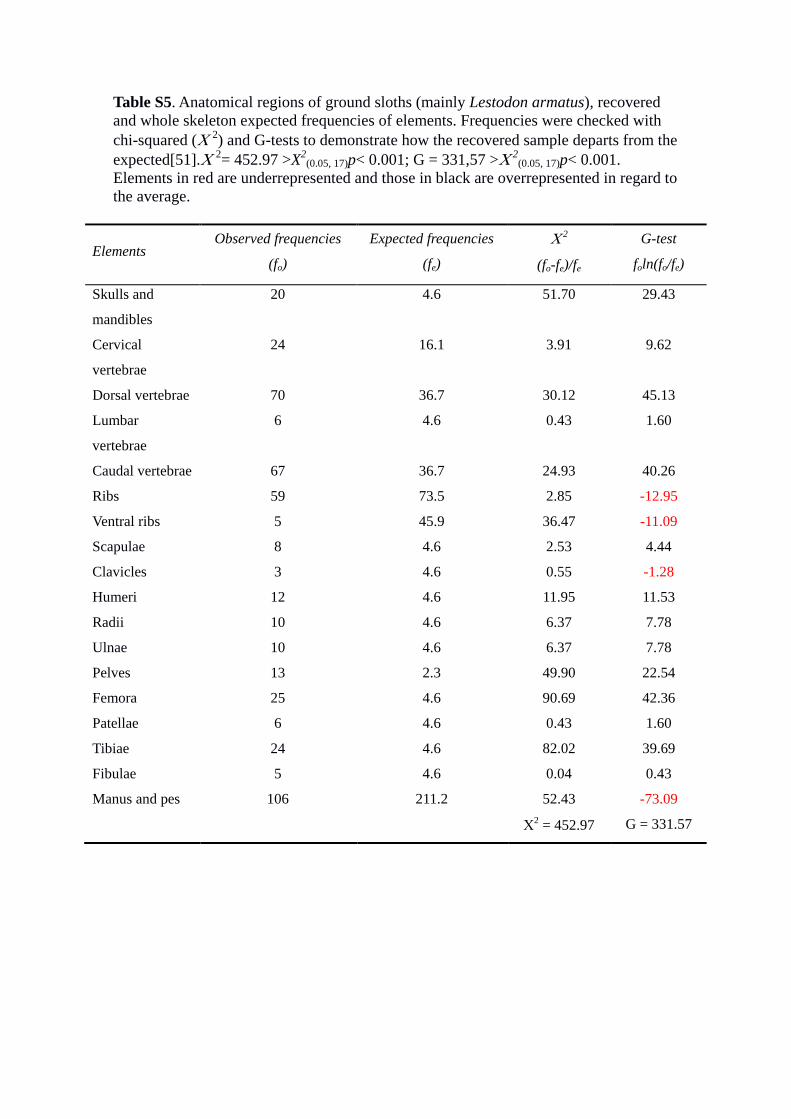
Table S5. Anatomical regions of ground sloths (mainly Lestodon armatus), recovered and whole skeleton expected frequencies of elements. Frequencies were checked with chi-squared (Χ 2) and G-tests to demonstrate how the recovered sample departs from the expected[51].Χ 2= 452.97 >X2
(0.05, 17)p< 0.001; G = 331,57 >Χ 2(0.05, 17)p< 0.001.
Elements in red are underrepresented and those in black are overrepresented in regard to the average.
Elements Observed frequencies
(fo)
Expected frequencies
(fe)
Χ 2
(fo-fe)/fe
G-test
foln(fo/fe)
Skulls and
mandibles
20 4.6 51.70 29.43
Cervical
vertebrae
24 16.1 3.91 9.62
Dorsal vertebrae 70 36.7 30.12 45.13
Lumbar
vertebrae
6 4.6 0.43 1.60
Caudal vertebrae 67 36.7 24.93 40.26
Ribs 59 73.5 2.85 -12.95
Ventral ribs 5 45.9 36.47 -11.09
Scapulae 8 4.6 2.53 4.44
Clavicles 3 4.6 0.55 -1.28
Humeri 12 4.6 11.95 11.53
Radii 10 4.6 6.37 7.78
Ulnae 10 4.6 6.37 7.78
Pelves 13 2.3 49.90 22.54
Femora 25 4.6 90.69 42.36
Patellae 6 4.6 0.43 1.60
Tibiae 24 4.6 82.02 39.69
Fibulae 5 4.6 0.04 0.43
Manus and pes 106 211.2 52.43 -73.09
Χ2 = 452.97 G = 331.57

5. SURFACE MODIFICATION ON THE FOSSIL SPECIMENS
Bones were cleaned following the usual procedures [32] and examined under low
magnification using a hand lens to determine the presence of surface
modifications.These were preliminary classified to distinguish trampling from possible
anthropogenic marks [33]. Other, larger bone modifications were analysed visually and
using virtual and plastic models.
(a) Identification and Diagnosis Methods
Further analysis of selected marks was carried out with light microscopy under
magnifications of 20x, 30x and 45x. Following methods based on extended depth-of-
field [34, 35] and, using an Olympus SZ61 stereomicroscope under a magnification of
45x, several photographs of the marks at different focus depth were taken. Helicon
Focus [34] software was used to make a complete in-focus image and a three-
dimensional model of each mark that accurately represents the micromorphology of the
modifications. These models were later processed with the software MeshLab [36] and
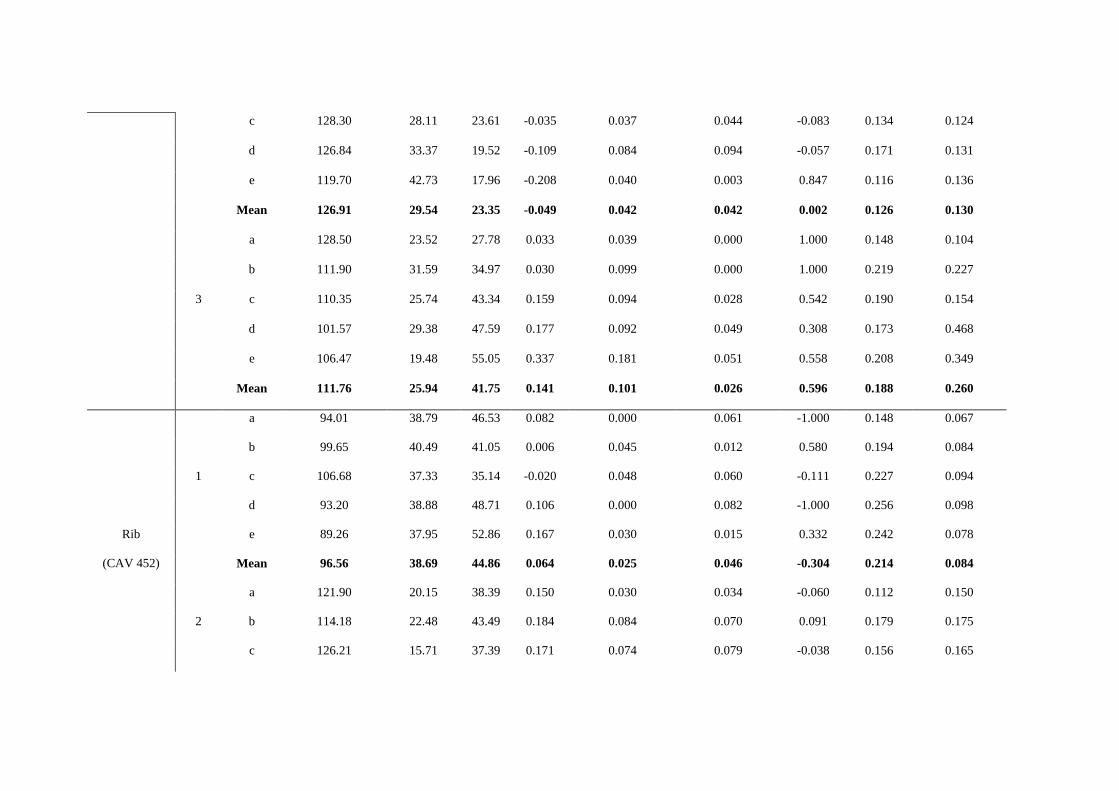
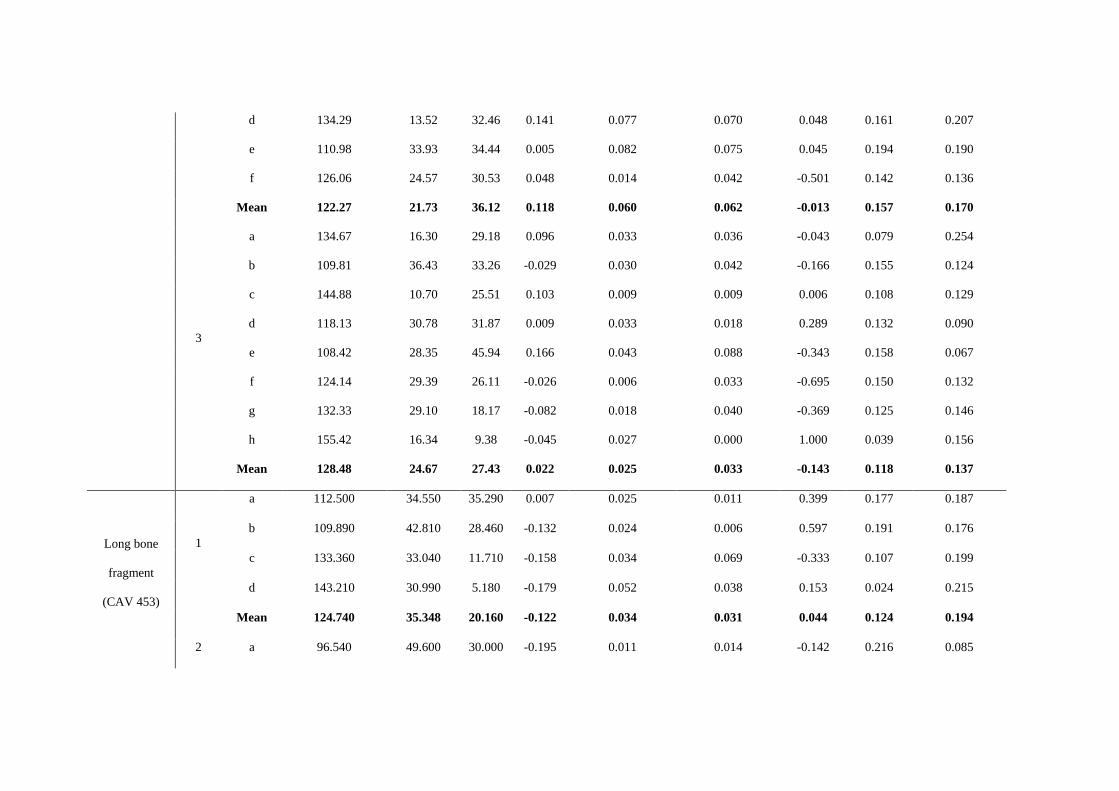
at least four perpendicular sections of the grooves were obtained for each mark. In Table
S6 we show the features observed in marks on the surface of several of the bones that
are typical of human-made marks [33, 37-38]:
• Opening angle: angle between left and right slopes at the floor of the cut mark.
• Angle of the Tool Impact Index: angles (σ1 and σ2) between the slopes S1 (left)
and S2 (right) of the cut mark and the unaffected bone surface. Because it is not
possible to determine the left and right side of the mark the ATI Index is
measured as: ((90º – σ1) – (90º – σ2)) / ((90º – σ1) + (90º – σ2)).
• Shoulder Height Index: height of the shoulders formed on either side of the cut
mark (SH1 and SH2). The SH Index is defined as (SH1 – SH2) / (SH1 + SH2).

• Depth of Cut (DC): perpendicular depth of the cut mark in relation to the
unaffected bone surface.
• Floor radius: the radius of circle fitted to the floor of the cut mark profile, with
the floor defined as lying between the two points where the profiles of the slopes
start to converge.

Table S6. Dimensions in the selected marks.
Bone Mark Section Opening Angle σ1 σ2 ATI Shoulder height
left (mm)
Shoulder height
right (mm) SH Index
Depth of
cut (mm)
Floor radius
(mm)
Tibia
(CAV 385)
1
a 100.45 41.98 38.61 -0.034 0.225 0.016 0.865 0.364 0.153
b 102.91 50.64 26.36 -0.2357 0.0114 0.0294 -0.441 0.2708 0.0748
c 91.38 39.11 49.03 0.108 0.025 0.105 -0.617 0.210 0.091
d 101.31 43.07 35.86 -0.0713 0 0.0264 -1.000 0.2458 0.1617
e 131.02 9.48 40.24 0.236 0.022 0.060 -0.467 0.112 0.099
f 129.33 24.69 25.93 0.0096 0 0.001 -1.000 0.1453 0.1407
Mean 109.4 34.83 36.01 0.0108 0.047 0.033 0.176 0.225 0.120
Rib
(CAV 451)
1
a 129.06 16.66 34.69 0.140 0.041 0.010 0.599 0.213 0.459
b 104.26 39.67 36.45 -0.031 0.040 0.030 0.143 0.336 0.473
c 114.20 36.20 29.72 -0.057 0.037 0.017 0.374 0.290 0.469
d 121.37 21.9 34.26 0.100 0.000 0.007 -1.000 0.180 0.167
e 101.88 38.99 39.34 0.003 0.017 0.060 -0.564 0.292 0.188
Mean 114.15 30.68 34.89 0.037 0.027 0.025 0.042 0.262 0.351
2 a 128.80 21.65 29.66 0.062 0.000 0.063 -1.000 0.119 0.149
b 130.90 21.85 25.99 0.031 0.051 0.007 0.763 0.088 0.112
c 128.30 28.11 23.61 -0.035 0.037 0.044 -0.083 0.134 0.124
d 126.84 33.37 19.52 -0.109 0.084 0.094 -0.057 0.171 0.131
e 119.70 42.73 17.96 -0.208 0.040 0.003 0.847 0.116 0.136
Mean 126.91 29.54 23.35 -0.049 0.042 0.042 0.002 0.126 0.130
3
a 128.50 23.52 27.78 0.033 0.039 0.000 1.000 0.148 0.104
b 111.90 31.59 34.97 0.030 0.099 0.000 1.000 0.219 0.227
c 110.35 25.74 43.34 0.159 0.094 0.028 0.542 0.190 0.154
d 101.57 29.38 47.59 0.177 0.092 0.049 0.308 0.173 0.468
e 106.47 19.48 55.05 0.337 0.181 0.051 0.558 0.208 0.349
Mean 111.76 25.94 41.75 0.141 0.101 0.026 0.596 0.188 0.260
Rib
(CAV 452)
1
a 94.01 38.79 46.53 0.082 0.000 0.061 -1.000 0.148 0.067
b 99.65 40.49 41.05 0.006 0.045 0.012 0.580 0.194 0.084
c 106.68 37.33 35.14 -0.020 0.048 0.060 -0.111 0.227 0.094
d 93.20 38.88 48.71 0.106 0.000 0.082 -1.000 0.256 0.098
e 89.26 37.95 52.86 0.167 0.030 0.015 0.332 0.242 0.078
Mean 96.56 38.69 44.86 0.064 0.025 0.046 -0.304 0.214 0.084
2
a 121.90 20.15 38.39 0.150 0.030 0.034 -0.060 0.112 0.150
b 114.18 22.48 43.49 0.184 0.084 0.070 0.091 0.179 0.175
c 126.21 15.71 37.39 0.171 0.074 0.079 -0.038 0.156 0.165
d 134.29 13.52 32.46 0.141 0.077 0.070 0.048 0.161 0.207
e 110.98 33.93 34.44 0.005 0.082 0.075 0.045 0.194 0.190
f 126.06 24.57 30.53 0.048 0.014 0.042 -0.501 0.142 0.136
Mean 122.27 21.73 36.12 0.118 0.060 0.062 -0.013 0.157 0.170
3
a 134.67 16.30 29.18 0.096 0.033 0.036 -0.043 0.079 0.254
b 109.81 36.43 33.26 -0.029 0.030 0.042 -0.166 0.155 0.124
c 144.88 10.70 25.51 0.103 0.009 0.009 0.006 0.108 0.129
d 118.13 30.78 31.87 0.009 0.033 0.018 0.289 0.132 0.090
e 108.42 28.35 45.94 0.166 0.043 0.088 -0.343 0.158 0.067
f 124.14 29.39 26.11 -0.026 0.006 0.033 -0.695 0.150 0.132
g 132.33 29.10 18.17 -0.082 0.018 0.040 -0.369 0.125 0.146
h 155.42 16.34 9.38 -0.045 0.027 0.000 1.000 0.039 0.156
Mean 128.48 24.67 27.43 0.022 0.025 0.033 -0.143 0.118 0.137
Long bone
fragment
(CAV 453)
1
a 112.500 34.550 35.290 0.007 0.025 0.011 0.399 0.177 0.187
b 109.890 42.810 28.460 -0.132 0.024 0.006 0.597 0.191 0.176
c 133.360 33.040 11.710 -0.158 0.034 0.069 -0.333 0.107 0.199
d 143.210 30.990 5.180 -0.179 0.052 0.038 0.153 0.024 0.215
Mean 124.740 35.348 20.160 -0.122 0.034 0.031 0.044 0.124 0.194
2 a 96.540 49.600 30.000 -0.195 0.011 0.014 -0.142 0.216 0.085

b 116.700 42.350 19.050 -0.196 0.069 0.041 0.250 0.083 0.172
c 101.360 47.790 30.780 -0.168 0.006 0.015 -0.426 0.205 0.110
d 102.520 50.330 27.760 -0.221 0.062 0.107 -0.265 0.121 0.228
Mean 104.280 47.518 26.898 -0.195 0.037 0.044 -0.092 0.156 0.149
Rib
(CAV 458)
1
a 129.76 21.91 28.85 0.054 0.0461 0.0049 0.808 0.0485 0.1602
b 110.52 36.92 30.61 -0.056 0.0275 0.0214 0.125 0.0673 0.0917
c 87.66 27.1 64.21 0.418 0.0329 0.1108 -0.542 0.1287 0.0898
d 161.71 11.35 8.53 -0.018 0.0256 0.0128 0.333 0.0417 0.3077
Mean 122.41 24.32 33.05 0.071 0.033 0.037 -0.063 0.072 0.162
Radii
(CAV 459)
1
a 80.580 46.280 53.040 0.084 0.018 0.000 1.000 0.367 0.172
b 112.780 13.780 54.780 0.368 0.099 0.023 0.619 0.257 0.228
c 115.890 17.690 46.770 0.252 0.067 0.015 0.643 0.386 0.231
d 116.220 33.420 27.810 -0.047 0.035 0.012 0.500 0.214 0.254
Mean 106.368 27.793 45.600 0.167 0.055 0.012 0.631 0.306 0.221
Rib
(CAV 475)
1
a 93.33 38.03 47.68 0.102 0.010 0.354 -0.945 0.169 0.071
b 86.73 45.91 47.24 0.015 0.000 0.016 -1.000 0.248 0.102
c 89.10 40.72 49.33 0.096 0.022 0.034 -0.218 0.280 0.081
d 92.80 44.58 42.42 -0.023 0.039 0.046 -0.081 0.283 0.073
Mean 90.49 42.31 46.67 0.048 0.018 0.113 -0.727 0.245 0.082

Hyoid
(CAV 476)
1
a 117.21 28.46 33.06 0.039 0.018 0.044 -0.428 0.153 0.091
b 125.77 24.3 29.67 0.043 0.021 0.050 -0.417 0.178 0.121
c 118.06 23.66 39.14 0.132 0.059 0.044 0.143 0.151 0.142
d 133.65 25.39 20.99 -0.0329 0.0368 0.0245 0.201 0.1133 0.1011
e 125.81 27.64 26.55 -0.0087 0.0421 0.0518 -0.103 0.1618 0.1035
Mean 124.10 25.89 29.88 0.0321 0.035 0.043 -0.099 0.151 0.112
Ulna
(CAV 520)
1
a 86.64 40.84 52.79 0.1384 0 0.0509 -1.000 0.2694 0.0569
b 110.17 26.35 44.26 0.164 0.071 0.000 1.000 0.276 0.100
c 140 16.8 20.43 0.0254 0.1093 0.0109 0.819 0.1011 0.1108
d 95.35 34.24 50.23 0.167 0.031 0.008 0.573 0.239 0.067
Mean 108.04 29.56 41.93 0.1140 0.053 0.018 0.501 0.221 0.084
Mandible
(CAV 897)
1
a 86.46 30.67 62.63 0.3686 0.000 0.132 -1.000 0.293 0.120
b 89.17 45.45 45.39 -0.0007 0.059 0.100 -0.259 0.294 0.091
c 106.55 28.83 44.65 0.1485 0.0134 0.0349 -0.445 0.4294 0.126
d 107.42 32.14 40.99 0.0828 0.0633 0.0495 0.122 0.2091 0.303
Mean 97.4 34.27 48.42 0.1453 0.034 0.079 -0.400 0.307 0.160

(b) Trampling
Two marks on a rib were assigned to trampling (Fig. S7). They were reconstructed using
the procedure described above and showed depths of 81.4 and 93.5 µm, opening angles
of 128º and 144º, and lacked microstriations, Herzian cones, asymmetry and shoulder
effect.
Figure S7. Trampling mark in rib CAV 120 and its section profile for comparison,
showing a smaller depth and lack of shoulders.
(c) Other marks on the bones
A few bones show peculiar indentations whose natural agency is difficult to establish: a
left maxilla (CAV 5, Figure S8) and a calcaneum (CAV 45, Figure S9), both belonging
to Lestodon and in perfect condition, with the outer bone layer well preserved on all
surfaces and lacking other types of possible human-made marks. Casts of the holes in
the bones described below were obtained by filling them with silicon rubber. A catalyser
was added to activate the elastomer. Previously, the inner surfaces of the holes were
cleaned of sediment and then protected with polyethyleneglycol to prevent detachment
of the spongy bone when the casts were extracted. The hole in the calcaneum is on the
lateral surface, while that of the maxilla is right before the first molariform. The casts

can be seen in Figure S10. The puncture in the maxilla is rather shallow, only about one
centimetre deep, and that in the calcaneum is much deeper, about 6.5 cm. The cast from
the calcaneum is interpreted to be double, with both parts at low angles to each other, as
if the object that made it had moved inside the bone after penetration. Otherwise, both
casts are very similar; in lateral view they lack symmetry, as their tips are bevelled.
Observed from the tip, both are laterally symmetrical and about 10 mm thick on average
from right to left and 2 or 3 cm from the thinner bottom to the thicker top. It is
noteworthy that this form does not correspond to that of the canine of a large
carnivoran.
Figure S8. Left maxilla of Lestodon armatus (CAV 5) showing the hole described in the
text.

Figure S9. Left calcaneum of Lestodon armatus (CAV 45) showing the indentation
described in the text.
Figure S10. Casts of the holes in the left maxilla (CAV 5, a) and the left calcaneum
(CAV 45, b) of Lestodon armatus. Black line in B indicates the boundary between each
one of the original dents.

Also, 29 elements present features that resemble those left by hammerstones, including
in some cases the anvil effect, with 9 of them showing no trampling marks, while other
8 bones are only slightly trampled. The rest are moderately to highly trampled. We
selected three elements to be further described and figured.
The dorsal vertebra CAV 305, belonging to Lestodon, shows in the ventral region of its
body a large sharp notch about 2 cm long, 4 mm wide and 4 mm deep with a partially
flaked margin (Figure S11). Such features resemble those made by chopping tools [39].
Moreover, the pelvis CAV 176, also belonging to Lestodon, shows a similar but more
flaked mark in the margin of the acetabulum (Figure S12), as well as an indentation
similar to those produced by hammerstones [39].
Finally, the fragment of long bone CAV 399 (Figure S13) shows a helicoidal fracture in
its inner side and a large tangential flake, which suggests a non-natural agent, since it is
unlikely for a bone to undergo nearly perpendicular fractures without changing its
position relative to the object that produced the impact.
Figure S11.Dorsal vertebra CAV 305, belonging to Lestodon armatus, showing a large
sharp notch with a partially flaked margin in the ventral region of its body.

Figure S12. Flaked mark in the margin of the acetabulum of the pelvis CAV 176
assigned to Lestodon armatus.
Figure S13. Fragment of long bone CAV 399 showing a helicoidal fracture in its inner
side and a large tangential flake.

6. LITHIC MATERIAL.
Lithic material was analysed with the usual techniques [40].In the fieldwork of 2011
and 2012, a total of 105 lithic elements were collected in spatial association with the
bones that precluded recent origin, i.e. they were found immediately below some of the
bones, and beside and above others. After a macroscopical analysis, 10 of them had
some features congruent with anthropic agency, such as flaking events: six possible
flakes (Figure S14), two possible cores and two possible tools on nuclei. One piece of
silcrete was mentioned in the main text to have features compatible with those of a
scraper (Fig. 4 f-g, Fig. S15). Since silcrete is present in the area and given its quality as
a raw material for human tools [41, 42], this quantity is admittedly a scarce
representation of possible lithic tools, although they have not been systematically
searched for in our excavations. The surfaces of these lithic fragments were analysed to
test the hypothesis of their anthropic origin using stereomicroscope at 20x. Nearly all of
them are made of raw material that is present in the surrounding area (quartz, silcrete,
amphibolite and sandstone), i.e. from the Mercedes Fm. [43].
Even though processing this number of giant animals would require a correspondingly
high usage of lithic material, other South American Pleistocene archaeological sites also
show a paucity of such elements [44,45] and only a part of this site has been rather
exhaustively explored.

Figure S14. Broken (above), complete (middle) and fragmentary (below) flakes [52]
found in the Arroyo del Vizcaíno site.
Fig. S15. Further images of the area a of the lithic element illustrated in Fig. 4f.
Scanning electron microphotographs at 2000 and 4300x showing micropolish. The
lighter particle is a piece of fabric left during cleaning.

References
S1. Leiggi, P., Schaff, C. R. & May, P. 1994 Field organization and specimen
collecting. In: Vertebrate Paleontological Techniques (eds. P. Leiggi, P. May), pp. 59-
76. Cambridge: Cambridge Univ. Press.
S2. Fariña, R. A. & Castilla, R. 2007Earliest evidence for human-megafauna interaction
in the Americas. In Human and Faunal Relationships Reviewed: An Archaeozoological
Approach (eds. E. Corona-M. & J. Arroyo-Cabrales), pp. 31-34. Oxford: Archaeopress.
S3. Longin, R. 1971 New method of collagen extraction for radiocarbon dating. Nature
230, 241-242.
S4. Schoeninger, M. & DeNiro, M. 1984 Nitrogen and carbon isotopic composition of
bone collagen from marine and terrestrial animals. Geochim. Cosmochim. Ac. 48, 625–
639.
S5. Higham, T. F. G., Jacobi, R. M. & Bronk Ramsey, C. 2006 AMS radiocarbon dating
of ancient bone using ultrafiltration. Radiocarbon 48, 179–95.
S6. Gupta, S. K. & Polach, H. A. 1985 Radiocarbon Dating Practices at ANU
Handbook Radiocarbon Laboratory Research School of Pacific Studies. Canberra:
ANU.
S7. Bronk Ramsey, C., Higham, T., Bowles, A. & Hedges, R. 2004 Improvements to
the pretreatment of bone at Oxford. Radiocarbon 46,155–63.
S8. Bronk Ramsey, C., Higham, T. & Leach, P. 2004 Towards high-precision AMS:
progress and limitations. Radiocarbon 46,17–24.
S9. Bronk Ramsey, C., Higham, T., Owen, D. C., Pike, W. G. &Hedges, R. 2002
Radiocarbon Dates from the Oxford Ams System: Archaeometry Datelist 31.
Archaeometry 44, 1-149.

S10. Tucker, M. 1982 The Field Description of Sedimentary Rocks. Geological Society
of London Handbook Series, No. 2. Maidenhead: Open University Press.
S11. Michols, G. 2009 Sedimentology and stratigraphy.New York: Wiley-Blackwell.
S12. Major, J. J. 2003 Debris Flow. In Encyclopedia of Sediments and Sedimentary
Rocks (eds. G. V. Middleton, M.J. Church, M. Coniglio, L. A. Hardie, & F.
J.Longstaffe), pp. 186–188. New York: Kluwer Acad. Publ.
S13. Mazza, P. P. A. & Ventra, D. 2011 Pleistocene debris-flow deposition of the
hippopotamus-bearing Collecurti bonebed (Macerata, Central Italy): Taphonomic and
paleoenvironmental analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 310, 296-314.
S14. Behrensmeyer, A. K. 1991 Terrestrial Vertebrate Accumulations. In Taphonomy:
Releasing the Data Locked in the Fossil Record (eds. P. Allison & D. E. G. Briggs), pp.
291-335. New York: Plenum.
S15. Bai, B., Wang, Y., Meng, J., Jin, X., Li, Q. & Ping L. 2011 Taphonomic analyses
of an Early Eocene Litolophus (Perissodactyla, Chalicotherioidea) assemblage from the
Erlian Basin, Inner Mongolia, China. Palaios 26, 187–196.
S16. Arribas, A. & Palmqvist, P.1998 Taphonomy and paleoecology of an assemblage
of large mammals: hyaenid activity in the lower Pleistocene site at Venta Micena (Orce,
Guadix-Baza Basin, Granada, Spain). Geobios 31, 3-47.
S17. Lyman, R. L.1994Vertebrate taphonomy. Cambridge: Cambridge Univ. Press.
S18. Behrensmeyer, A. K. 1978 Taphonomic and ecologic information from bone
weathering. Paleobiol. 4, l50-l62.
S19. Czerwonogora, A., Fariña, R. A. & Tonni, E. P. 2011 Diet and isotopes of Late
Pleistocene ground sloths: first results for Lestodon and Glossotherium (Xenarthra,
Tardigrada). Neues Jahrb. Geol. P-A 262, 257-266.

S20. Rogers, R. R. Eberth, D. A. & Fiorillo, A. R. 2007 Bonebeds: Genesis, Analysis,
and Paleobiological Significance. Chicago: Chicago Univ. Press.
S21. Voorhies, M. R. 1969 Taphonomy and population dynamics of an early Pliocene
vertebrate fauna, Knox County, Nebraska. Contrib. Geol. 1, 1-69.
S22. Kreutzer, L. A. 1988 Megafaunal butchering at Lubbock Lake, Texas: a
taphonomic reanalysis. Quaternary Research 30, 221-231.
S23. Frison, G. C. & Todd, L. C. 1986 The Colby mammoth site: Taphonomy and
archaeology of a Clovis kill in northern Wyoming. Albuquerque: Univ. New Mexico
Press.
S24. Behrensmeyer, A. K. & Dechant-Boaz, D. E. 1980 The recent bones of Amboseli
Park, Kenya. In Fossils in the Making (eds. A. K. Behrensmeyer & A. Hill), pp. 72-93.
Chicago: Chicago University Press.
S25. Bunn, H. T. 1982 Meat-eating and human evolution: studies on the diet and
subsistence patterns of Plio-Pleistocene hominids in East Africa. Ph.D. Dissertation.
Berkeley: Univ. of California.
S26. Bunn, H. T. 1986 Patterns of skeletal representation and hominid subsistence
activities at Olduvai Gorge, Tanzania and Koobi Fora, Kenya. J. Hum. Evol. 15, 673–
690.
S27. Damuth, J. 1982 Analysis of the preservation of community structure in
assemblages of fossil mammals. Paleobiol. 8, 434-446.
S28. Behrensmeyer, A. K.,Western, D. & Dechant-Boaz, D. E. New perspectives in
vertebrate paleoecology from a recent bone assemblage. Paleobiol. 5, 12-21.
S29. Tonni, E. P., Prado, J. L., Menegaz, A. N. & Salemme, M. C. 1985 La unidad
mamífero (Fauna) Lujanense. Proyección de la estratigrafía mamaliana al cuaternario de
la región pampeana. Ameghiniana 22, 255-261.

S30. Fariña, R. A., Vizcaíno, S. F. & De Iuliis, G. 2013 Megafauna: Giant Beasts of
Pleistocene South America. Bloomington: Indiana Univ. Press.
S31. Meltzer, D. J. 2006 Folsom: New Archaeological Investigations of a Classic
Paleoindian Bison Kill. Berkeley: University of California Press.
S32. May, P., Reser, P. & Leiggi, P. 1994 Macrovertebrate preparation. In: Vertebrate
Paleontological Techniques (eds. P. Leiggi, P. May), pp. 113-153. Cambridge:
Cambridge Univ. Press.
S33. Domínguez-Rodrigo, M., De Juana, S., Galán, A. B. & Rodríguez, M. 2009 A new
protocol to differentiate trampling marks from butchery cut marks. J. Archaeol. Sci. 36,
2643-2654.
S34. Berejnov, V. V. 2009 Rapid and Inexpensive Reconstruction of 3DStructures for
Micro-Objects Using Common Optical Microscopy. arXiv preprint arXiv:0904.2024.
S35. Hein, L. R. O., de Campos, K. A., Caltabiano, P. C. R de O. & Horovistiz, A. L.
2010 3-D reconstruction by extended depth-of-field in failure analysis – Case study I:
Qualitative fractographic investigation of fractured bolts in a partial valve. Eng. Fail.
Anal. 17, 515–520.
S36. MeshLab. 2009 Open source, portable, and extensible system for the processing
and editing of unstructured 3D triangular meshes. http://meshlab.sourceforge.net/.
S37. Bello, S. M. & Soligo, C. 2008 A new method for the quantitative analysis of
cutmark micromorphology. J. Archaeol. Sci. 35, 1542-1552.
S38.Bello, S. M. Parfitt, S. A.& Stringer, C. 2009 Quantitative micromorphological
analyses of cut marks produced by ancient and modern handaxes. J. Archaeol. Sci. 36,
1869-1880.

S39. Capaldo, S. D. & Blumenschein, R. J. 1994 A quantitative diagnosis of notches
made by hammerstone percussion and carnivore gnawing on bovid long bones. Am.
Antiq. 59, 724-748.
S40. Collins, M. B. 1975 Lithic Technology as a Means of Processual Inference. In
Lithic Technology: Making and Using Stone Tools (ed. E. Swanson), pp. 15-34.
Mouton: The Hague.
S41. Gascue, A. 2009 La tecnología lítica desarrollada por los habitantes prehistóricos
del Arroyo del Perdido (Soriano, Uruguay). Arqueología Prehistórica Uruguaya en el
Siglo XXI, 117-130.
S42. Gascue, A.2009 Tecnología lítica y patrones de asentamiento en la cuenca de
arroyo Grande (Soriano). Arqueología Prehistórica Uruguaya en el Siglo XXI, 133-150.
S43. Spoturno, J., Oyhantçabal, P., Aubet, N., Cazaux, S. & Morales, E. 2004 Carta
Geológica y Memoria Explicativa a Escala 1/100.000 del Departamento de Canelones,
CONICYT. Proyecto 6019. Fondo Clemente Estable. Versión I CD.
S44. Martínez, G. Gutiérrez, M. A. & Tonni, E. P. 2012 Paleoenvironments and faunal
extinctions: Analysis of the archaeological assemblages at the Paso Otero locality
(Argentina) during the Late Pleistocene–Early Holocene. Quat. Int. in press (2013).
S45. Politis, G. & Messineo, P. G. 2008 The Campo Laborde site: New evidence for the
Holocene survival of Pleistocene megafauna in the Argentine Pampas. Quat. Int. 191,
98–114.
S46. Arribas, A., Palmqvist, P., Pérez-Claros, J. A., Castilla, R., Vizcaíno, S. F. &
Fariña, R. A. 2001 New evidence on the interaction between humans and megafauna in
South America. Publ. Semin. Paleon. Zaragoza 5, 228-238.

S47. Fariña, R. A., Vizcaíno, S. F. & Bargo, M. S. 1998 Body size estimations in
Lujanian (Late Pleistocene-Early Holocene of South America) mammal megafauna.
Mastozool. Neotr. 5, 87-108.
S48. Czerwonogora, A. 2010 Morfología, sistemática y paleobiología de los perezosos
gigantes del género Lestodon (Mammalia, Xenarthra, Tardigrada). Ph.D. Dissertation.
La Plata: UNLP.
S49. Binford, L. R. 1984 Faunal Remains from Klasies River Mouth. New York:
Academic Press.
S50. Brain, C. K. 1981. The Hunters or the Hunted? Chicago: University of Chicago
Press.
S51. Sokal, R. R. & Rohlf, F. J. 1981 Biometry, 2nd ed. San Francisco: Freeman.
S52. Sullivan, A. & Rozen, K. 1985 Debitage Analysis and Archaeological
Interpretation. Am. Antiq. 50, 755-779.




















