
Thalamic neuropeptide mediating the effects of nursing on lactation and maternal motivation
Upload
independentCategory
view
0download
0
ELSEVIER
BEHAVIOURAL BRAIN
RESEARCH Behavioural Brain Research 67 (1995) 51-58
Research report
Effects of mammillary bodies and mediodorsal thalamic lesions on the acquisition and retention of a learning set in mice: paradoxical effect of the
intersession interval
Ali Krazem”, Daniel BCracochCab7*, Robert Jtiardb
a UniversitP de Tizi-Ouzou, Tizi-Ouzou, Algeria b Laboratoire de Neurosciences Cognitives et Comportementales, URA CNRS 339, Universitk de Bordeaux 1, avenue des FacultPs. 33405 Talence Cedex,
France
Received 27 December 1993; revised 15 July 1994; accepted 18 July 1994
Abstract
The effects of ibotenic acid lesions of either the mediodorsal thalamus (MD) or the mammillary bodies (MB) on the acquisition and retention of a reversal learning set in a T-maze were studied in mice. Both the MB and MD lesions disrupted performance in this task, but only MD lesions slowed down the rate of learning, MD-lesioned subjects requiring more trials than the other two groups to master the discrimination at the end of the learning phase. Surprisingly, increasing the intersession interval from 24 h to 10 days totally al- leviated the deficit of MD-subjects and even improved their performance as compared to the last (8th day) session. The overall re- sults of the study show that both MD and MB lesions do not induce important learning deficits nor anterograde amnesia in the re- versal learning set task.
Key words: Reversal learning; Frontal cortex; T-maze; Behavioral flexibility; Rodent; Diencephalic amnesia
1. Introduction
A number of studies have attributed some of the cog- nitive impairments observed in chronic alcoholics to le- sions of the frontal cortex [ 9,14,20]. More particularly, it has been reported that chronic alcoholics were impaired in tasks involving the ability to remember a sequence of events, spatial working memory, or in reversal discrimi- nation tasks [ 9,19,20], these processes also being impaired following frontal lobe damage [ 9,11,12,20,23]. However, in alcoholics, it is not clear whether these cognitive defi- cits are specifically due to direct damage of the frontal cortex, since subcortical lesions, mainly localized into the medio-dorsal thalamus (MD), the mammillary bodies (MB) and the anterior thalamic nuclei’, have also been observed and may contribute to the memory and cognitive impairments [ 1,2,3,5,6,13,15,16]. Indeed, it has been shown that MD damage reproduce some forms of ‘frontal- like’ dysfunctions, such as impairments of behavioral flex-
* Corresponding author.
0166-4328/95/%9.50 0 1995 Elsevier Science B.V. All rights reserved SSDI 0166-4328(94)00103-O
ibility [ 11,121, and of the ability to learn a cognitive skill such as the ‘learning set’ rule [22].
At least to our knowledge, direct comparisons of sep- arate lesions of either the MD or the MB in learning tasks involving the frontal cortex activity are scarce. In view of this situation, we have decided to study the effects of separate MD and MB lesions in a ‘learning set’ procedure. Indeed, previous studies performed in our laboratory have shown that this task is sensitive to frontal damage, since lesioning either the anterior or posterior cingulate areas in the mouse led to impairments, as reflected by differential effects of the lesions on the rate of learning over succes- sive discriminations [ 181. Moreover, Meunier [ 171 has shown that, in the mouse, the MD on the one hand and the anterior thalamic nuclei (which are the major target sites of MB efferents) on the other hand send outputs to the anterior and posterior cingulate areas respectively which, as mentioned above, are differently involved in learning processes. Taking into account the anatomical connections of the MD and MB (even though indirectly via the anterior thalamus) on the anterior and posterior cingulate cortex respectively, one would expect that MB

52 A. Krazem et al. / Behavioural Brain Research 67 (1995) 51-58
and MD lesions may also lead to different cognitive defi- cits in the learning set procedure.
2. Materials and methods
2.1. Subjects
The subjects were experimentally naive male mice of the Balb/c strain obtained at the age of 5-6 weeks from IFFA- CREDO (Lyon, France). The mice were aged 12-14 weeks at the beginning of the experiment and weighed approxi- mately 28 g (range: 26-3 1 g). They were housed individu- ally in a room maintained at constant temperature (23 “C) and on a 12:12 light/dark cycle (lights on at 07.00). Be- havioral testing was carried out during the light phase of the cycle. Food and water were provided ad libitum, ex- cept during the behavioral testing in which food was re- stricted as described below.
2.2. Surgery
Surgery was conducted under sodium thiopental anes- thesia (80 mg/kg i.p.). After sedation, the mouse was placed in a stereotaxic apparatus (KOPF instruments) and an incision was made in the scalp to expose the skull. Trephine openings were then drilled in the skull at the appropriate locations.
Lesions were made by using multiple injections of a small amount of ibotenic acid. The ibotenic acid (SIGMA) was dissolved in phosphate-buffered saline (final pH: 7.4) at the concentration of 10 mg/ml. The solution was then divided into aliquots and stored frozen until use. Injec- tions were made through a glass micro-pipette glued to a l-p1 Hamilton-syringe. The Hamilton-syringe was mounted on a stereotaxic manipulation arm equipped with a micrometer screw which enabled a regular and slow-rate injection of the drug. Each injection lasted approximately 5 min and, to prevent back-diffusion of the neurotoxin, the pipette was left in place 7 additional minutes before its withdrawal. MB lesions required a single injection (0.30 ~1) centered on the posterior median mammillary nucleus, whereas mediodorsal thalamic lesions were bilateral (0.20 $/per site). Lesions were made at the following coordi- nates: MB: 2.5 mm posterior to the bregma, midline, and 5.3 mm below the skull; MD: 1.4 mm posterior to the bregma, 0.5 mm bilateral, and 3.2 mm below the skull. The controls animals were either sham-operated or unoper- ated.
On completion of the injections, the wound was closed and the animal was placed in a heated room until1 full recovery from anesthesia. Behavioral testing was initiated approximately 12 days after the surgical intervention.
The learning set. The learning set has been widely used to study frontal lobe function [4]. Basically, in the learning-set procedure in a T-maze, subjects are required to alternate, from day to day, choices of the goal-arms of the maze (left vs. right), until a given criterion was met. In this task, during each daily session, subjects learned to enter repeatedly a given arm of the maze, in order to ob- tain a reward, the other arm being non-rewarded. The number of trials required to meet the criterion (for example 5 errorless trials) decreased over days in normal subjects, reflecting thereby the ability of the subjects to learn or to apply more rapidly the procedure (‘rule learning’ or ‘learn- ing to learn’ phenomenon). However, it is not clear as to whether subjects learn to alternate from day to day choices of the goal-arm, or if they learn to apply a simple win-stay strategy regardless of the reinforced response emitted the day before. If subjects learn to alternate over days of testing (at least for the first choice), changing the ‘rule’ at the end of the learning phase (i.e. the subjects is then re- quired to re-enter the arm baited the day before) or in- creasing the intersession interval should have detrimental effects on performance. If, on the contrary, subjects learn a simple win-stay strategy which consists of re-entering the arm baited during the ongoing session (regardless of the choice emitted the day before), then the change of the rule or the increase of the intersession interval should have little effect on performance. The multiple phase paradigm described below is aimed at testing the effects of the le- sions in these various situations.
2.3. Apparatus
Behavioral testing was carried out in a T-maze con- structed of grey Plexiglas. Stem and arms were 35 cm long, 10 cm wide, and 25 cm high. The start-box (10 x 12 cm) was separated from the stem by a horizontal sliding door. Horizontal sliding doors were also placed at the entrance of each arm. A low intensity diffuse illumination (10 lux) was provided above the apparatus and background white noise (40 dB) was used during all test sessions.
2.4. Procedure
The food deprivation schedule was gradually initiated over 4 days so that, at the time of training, the mice weighed 88% of their initial free-feeding weights. Food ration was adjusted individually in order to maintain the same level of deprivation throughout the ensuing experimental pe- riod.
2.5. Preliminary training
Preliminary training was carried out over 3 consecutive days. On the first day, all animals were allowed 10 min free

A. Krazem et al. / Behavioural Brain Research 67 (1995) 51-58 53
exploration of the apparatus in order to familiarize them with the experimental conditions. No food reward was available or given during this free-exploration session. On the 2 following days, each mouse was submitted daily to a session of 6 successive trials during which both arms of the maze were baited (BIOSERV pellets, 20 mg). On each trial, the mouse was placed in the start-box, and, 30 s later, the door of the box was opened. When the animal entered one of the two arms, the door of that arm was then closed. After a 30-s confinement in the chosen arm, the mouse was removed and placed again in the start-box for the next trial. For each trial, the chosen arm and the time that elapsed between the opening of the door of the startbox and the closing of the door of the chosen arm (running time) were noted. These two 6-trial sessions were designed to ensure that each animal learned to go to the end of the maze-arms in order to obtain food reward.
2.6. Formal testing
As described in Fig. 1 (upper part), the formal testing was composed of different phases including an acquisition phase (Day 1) followed by a ‘learning phase’ constituted of a series of 7 reversal sessions (Day 2 to Day 8). How- ever, on Day 8, some subjects were reinforced for choos- ing the same arm of the maze which was rewarded dur- ing the previous (7th) session. Only the subjects having
A Learning Phase
B Memory
DAY 1 DAY 1
T__T_______-___________T retention 6min mtontbn 24 hr
Fig. 1. A: learning phase; learning sessions consisted of an initial ac- quisition session (Day 1) followed by 8 reversal sessions (Days 2-8 and 18). The first 7 reversal session (Days2-8) were given at 24-h intervals, whereas the 8th session (Day 18) was given 10 days after the last daily reversal (Day 8). Solid arrows indicate the correct response for each session. On Days 8 and 18 of testing, a number of subjects were sub- mitted to a repetition session, which consisted of re-entering the arm reinforced at the immediate preceeding session (7th and 8th respectively). B: memory; 2 single retention-test trials were given at 5 min and 24 h after the end of each learning session (Day 1 in the example). In this way, the 24-h test trial (Day 2 in the example) constituted the first trial of the following learning session. Broken arrows depict the correct response for each retention-test trial.
been submitted to the reversal sessions were then placed in the colony room and maintained under the food dep- rivation schedule during 10 consecutive days. Then (Day 18), half of the subjects were submitted to a reversal ses- sion during which the reinforced arm was opposite to the arm reinforced on the previous reversal session (i.e. Day 8 of the learning phase; Intersession interval: 10 days); the remaining half subjects were reinforced for choosing on Day 18 the arm already reinforced on the last session (i.e. Day 8 of the learning phase; intersession interval: 10 days).
The acquisition session (Day 1) consisted of a succes- sion of trials identical to those described above for the preliminary training, with the exception that only one arm of the T-maze was baited. For each mouse, the baited arm selected on Day 1 was its ‘non-preferred’ arm during the preliminary trials (i.e. the arm that the animal had less frequently chosen). The acquisition session was continued until the subject reached the criterion of 5 correct responses in 5 consecutive trials.
Following acquisition, daily reversal sessions (Fig. 1A: learning phase) took place over 7 consecutive days (Day 2 to Day 8) during which the baited arm was reversed from day to day. Each reversal session was pursued until1 the animal achieved the same criterion of 5 consecutive error- less trials.
During the first 7 days of testing (acquisition session + 6 reversal sessions), 2 additional ‘retention’ trials were given at the end of each session (Fig. 1B: memory): one trial 5 min and one trial 24 h after the criterion was met. The 24-h retention trial of each learning session consti- tuted, in fact, the first trial of the following session; thus, at the first trial of the session, the food pellet was placed in the arm opposite to the one reinforced the day before.
This behavioral paradigm enabled us to measure: (i) the rate of acquisition of the initial spatial discrimination (Day l), (ii) the performance on the first reversal session (Day 2), (iii) the performance savings over successive daily re- versal sessions (Day 1 to Day 8), (iv) the eventual change of performance as a function of the change of the rule (repetition versus reversal, Day 8), and (v) the saving of the initial learning (Day 1 to Day 8) following a long (10 days) time interval using either the same rule (reversal session) or using the opposite one (repetition session).
2.7. Histology
After completion of behavioral testing, all operated ani- mals were deeply anesthetized and then perfused with physiological saline followed by 10% formalin. The brains were removed, post-fixed in 10% formalin for several days and then soaked in a 30% sucrose-formalin solution for 24 h. They were frozen and then sectioned in a coronal plane at 40 pm. Brain sections were stained with thionin

54 A. Krazem et al. / Behavioural Brain Research 67 (I 995) 51-58
in order to evaluate, under a light microscope, the location and extent of the cell loss.
2.8. Data analysis
Performance during the learning phase was assessed by the number of errors preceding criterion trials (5 consecu- tive errorless trials); retention was expressed by the per- centage of correct responses. Two-way analysis of vari- ance with one repeated measure (either days of testing or retention intervals) were performed to assess the effects of surgical treatment on the animal’s performance. Signiti- cant differences between groups were analysed by facto- rial ANOVA. Comparisons of performance within a given group was subjected to Student’s t-test for dependant samples.
3. Results
3.1. Histology
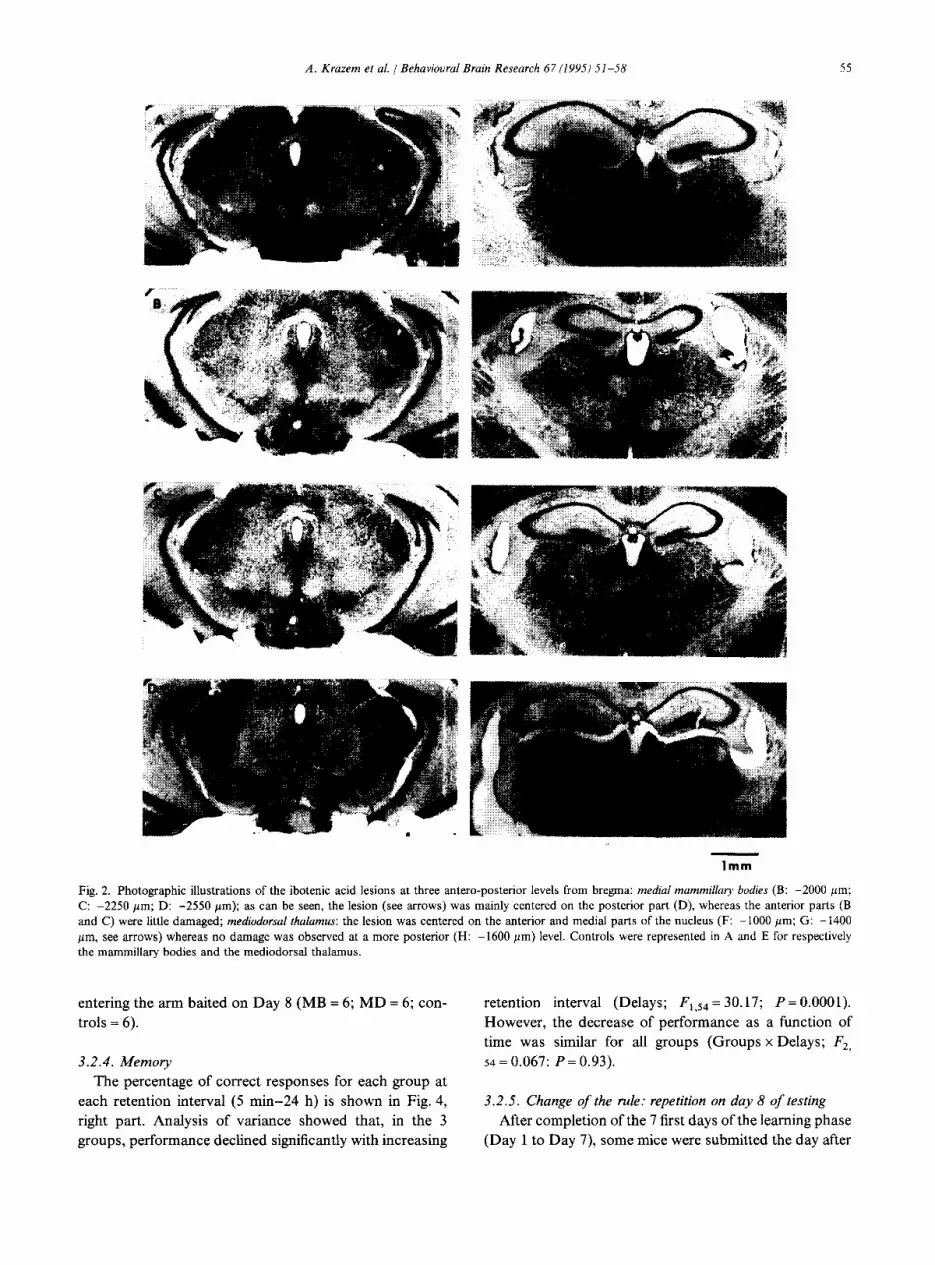
All MB-lesioned mice exhibited lesions which were mainly centered on the medial mammillary nucleus, even though damage extended as well into the adjacent lateral nuclei and, in some subjects (8 mice), into the supramam- millary nucleus. However, no obvious behavioral differ- ences were observed among lesioned animals in relation to the extent of these damage. The antero-posterior extent of the MB lesion was about 600 pm, from the antero-medial part of the MB to the posterior part, which was the area mainly damaged (see Fig. 2). Mice of the MD group exi- bited bilateral lesions mainly centered on the MD nucleus; however, in all lesioned mice, some loss of cells was also observed in the centralis medialis and in the midline tha- lamic nuclei; in all subjects, no damage was observed into the anterior thalamic nuclei. The antero-posterior extent of the MD lesion was about 400 mm per side.
3.2. Spatial reversal learning
3.2. I. Acquisition (Day 1) Results are represented in Fig. 3. During the initial spa-
tial discrimination, mice of the control group (n = 20) re- quired an average of 11.8 & 3.7 trials to reach the criterion of 5 errorless trials. Lesioned animals did not significantly differ from controls (F2,56 = 0.9; P= 0.40); thus, MD- lesioned subjects (n = 17) required an average of 11.5 + 3.0 errorless trials whereas MB-lesioned mice (n = 20) required an average of 11.7 + 6.3 trials to achieve the criterion.
3.2.2. First reversal (Day 2) Results are represented in Fig. 3. On Day 2 of testing,
animals were required to reverse choice of the goal-arm,
until 5 successive errorless trials. A significant between- groups difference was oberved (Groups; F2,56 = 11.0; P= 0.0001). This difference was mainly due to MD-lesioned subjects which required significantly more trials than the other groups to meet the criterion (16.1 k 5.2, 12.6 k 4.4 and 11.3 & 4.7 for respectively MD, MB and control subjects; paired comparisons: PC 0.01 in all cases). The evolution of performance from Day 1 to Day 2 was significantly different among the groups (Groups x Days; F2,54 = 4.12; P = 0.02). This difference was mainly due to MD subjects which signifi- cantly required more trials to meet the criterion as compared to controls (Groups x Days; F,,,, = 8.47; P= 0.006) whereas the evolution of performance was similar for MB and control mice (Groups x Days; F ,,j* = .141).
3.2.3. Successive reversals (Day I to Da,y 8) and retention The rate of learning across a succession of daily dis-
crimination reversals was evaluated by repeated analysis of variance of the animal’s performance from Day 1 to Day 8. The analysis was carried out with 13 MB subjects, 12 MD subjects and 14 controls that performed the 8 reversal sessions. Except for the first and last day of the learning phase, performance of subjects were pooled by using pairs of two successive days (i.e. Days 2 + 3, Days 4 + 5 and Days 6 + 7). The overall patterns of performance (see Fig. 4) showed that the number of trials necessary to reach the criterion decreased significantly over days of testing (Days; F4,144 = 30.2; P= 0.0001). A significant between-groups difference was observed (Groups; F2, 144 = 13.8; P = 0.0001). Specifically, MB-lesioned mice re- quired an average of 14.1 trials to reach the criterion whereas controls required an average of 12.5 trials (Fl,,6 = 10.96; P = 0.002); MD-lesioned mice also required more trials to reach the criterion as compared to control mice (15.7 trials vs 12,5 trials respectively; Groups; F ,,7,, = 11.9; P = 0.001). Moreover, the evolution of per- formance across days of testing was significantly different between groups (Groups x Days; F8,144 = 2.03: P = 0.04). This difference was mainly due to MD subjects whose rate of learning was significantly lower as compared to controls (Groups x Days; F5,120 = 3.77; P= 0.003) whereas the evolution of performance of MB-lesioned subjects was similar to that of controls (Groups x Days; F,,,, = 1.34; P = 0.24).
Following Day 8 of testing, mice which had been sub- mitted to the reversal condition were placed in the colony room for ten days and maintained under the food depri- vation schedule. They were then divided into two groups, one assigned to a reversal session, as compared to Day 8 (MB = 6; MD = 6; controls = 7) whereas the other one as- signed to a repetition condition which consisted of re-
A. Krazem et al. I Behavioural Brain Research 67 11995) 5 I-58 55
lmm
Fig. 2. Photographic illustrations of the ibotenic acid lesions at three antero-posterior levels from bregma: medial mammillary bodies (B: -2000 pm; C: -2250 pm; D: -2550 pm); as can be seen, the lesion (see arrows) was mainly centered on the posterior part (D), whereas the anterior parts (B and C) were little damaged; mediodorsal thalamus: the lesion was centered on the anterior and medial parts of the nucleus (F: -1000 pm; G: -1400 pm, see arrows) whereas no damage was observed at a more posterior (H: -1600 pm) level. Controls were represented in A and E for respectively the mammillary bodies and the mediodorsal thalamus.
entering the arm baited on Day 8 (MB = 6; MD = 6; con- trols = 6).
3.2.4. Memory The percentage of correct responses for each group at
each retention interval (5 min-24 h) is shown in Fig. 4, right part. Analysis of variance showed that, in the 3 groups, performance declined significantly with increasing
retention interval (Delays; F,,,, = 30.17; P = 0.0001). However, the decrease of performance as a function of time was similar for all groups (Groups x Delays; F2, 54 = 0.067: P = 0.93).
3.2.5. Change of the rule: repetition on day 8 of testing After completion of the 7 first days of the learning phase
(Day 1 to Day 7), some mice were submitted the day after

56 A. Krazem et al. / Behavioural Brain Research 67 (1995) 51-58
16-
16-
14 - ._ ii ‘C 0 g ‘2-
P
c
10
8
6 II Controls MB MD Controls MB MD
Acquisition First reversal Day 1 Day 2
Fig. 3. Left: mean number of trials required to meet the criterion (5 er- rorless trials) in Control, MD and MB groups, on the acquisition ses- sion (first day of testing). No significant differences were observed among the groups. Right: mean number of trials required to meet the criterion in Controls, MD and MB groups on the first reversal session. As can be seen, MD-lesioned animals make more errors than the other two groups. ***: P<O.Ol.
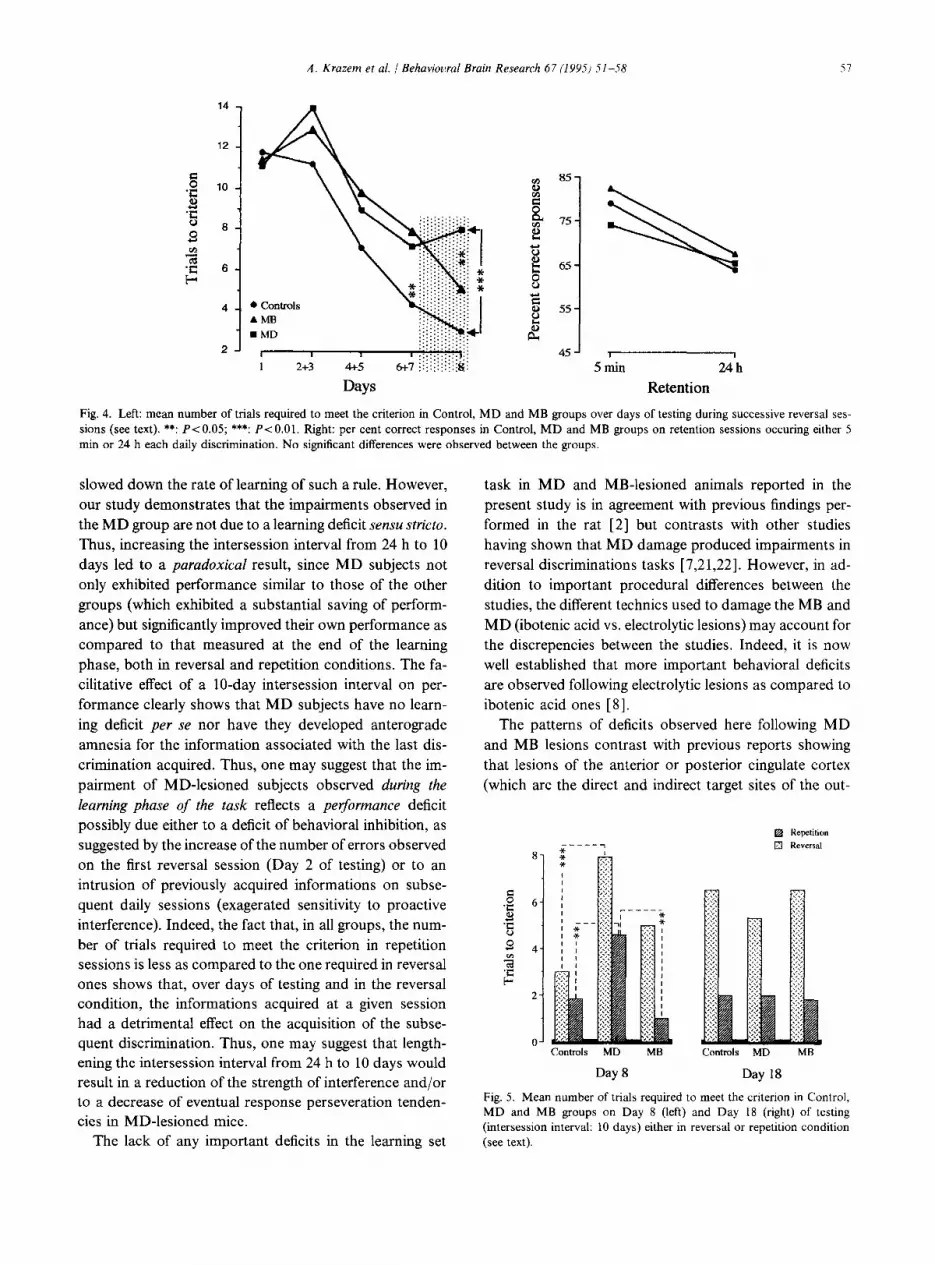
(Day 8) to a repetition session (which consisted of re- entering the arm baited on the previous day). 7 MB- lesioned mice, 5 MD-lesioned animals and 6 controls were run in the repetition condition. The performances of each of these sub-groups were similar to the ones observed in the population from which these sub-groups belonged (P> 0.05 for all statistical analyses). In the repetition con- dition, a significant between-groups difference was ob- served (Groups; F2,17 = 5.9; P= 0.01). Again, MD-le- sioned subjects required significantly more trials than both controls and MB-lesioned mice to reach the criterion (4.6 + 0.6 (MD), 1.8 ? 0.9 (Controls) and 1.0 f 0.6 (MB) P~0.05) whereas MB-lesioned mice did not differ from controls (P> 0.05) (see Fig. 5, left). One might observe that the number of trials necessary to reach the criterion was less in the repetition condition as compared to the reversal one (Conditions F,.,, = 8.96; P = 0.004).
3.2.6. Reversal and Repetition after a long{10 days) interses- sion interval
Results are shown in Fig. 5, right. The statistical analysis performed on all groups, condi-
tions, and delays showed no significant interaction be- tween the three parameters (Groups x Delays x Condi- tions; Fz,31 = 1.19; P = 0.3 1). The interaction between Groups and Conditions was not significant (F2,31 = 0.23; P= 0.79). However, the analysis showed a significant Groups x Delays interaction (F2,31 = 5.03; P = 0.013) as well as a significant Delays x Conditions interaction (F,,,, = 17.22; P = 0.0001). This lead us to perform sepa-
rate analyses of the effects of the delay as a function of the conditions.
3.2.6.1. Reversal sessions. Performances of subjects were significantly modified by the increase of the interses- sion interval (Groups x Delay; F2,17 = 6.37; P = 0.008). In- deed, on day 18 of testing, both controls and MB-lesioned subjects increased the number of errors as compared to day 8 of testing (t = 2.7; P= 0.04 and t = 3.41; P= 0.01 respectively) whereas a decrease of errors was observed in MD-lesioned animals even though differences were not statistically significant (P = 0.37). Given the different evo- lution of performances among the three groups, between- groups performances were very similar, the number of errors being 5.3, 6.5 and 6.5 for MD, MB and control groups respectively (P>O.O5 in all between-groups com- parisons).
3.2.6.2 Repetition sessions. The overall analysis of per- formances among the groups showed that, on day 18 of testing, the number of trials necessary to reach the crite- rion was significantly less in the repetition sessions, as compared to the reversal sessions (Conditions; F1,31 = 43.8; P = 0.001). No significant between-groups differ-
ences were observed on day 18 of testing (F2,17 = 0.08). As a whole, the evolution of performances among the groups were similar (Groups x Delays; (F2,L5 = 1.7; P = 0.21), all groups reducing the number of errors as compared to day 8 of testing (Delays; F1,15 = 15.0; P = 0.0001). However, these statistical levels of significance was mainly due to the modification of performances of MD subjects; indeed; only MD subjects exhibited a significant improvement of performance following the long intersession interval, as compared to Day 8 of testing (t = 10.6; P= 0.0004), whereas differences in MB and control subjects were not statistically significant (P>O.O5 in both cases).
4. Discussion
Our study mainly shows that MD and MB lesions pro- duced behavioral impairments in the learning phase of the reversal discrimination task, as compared to controls. Specifically, these two groups required, from day to day, more trials than controls to master the criterion. Indeed, during the learning phase of the task, MD-lesioned mice exhibited an important deficit on the first reversal session and on subsequent ones, in spite of a general decrease in the number of trials required to meet the criterion over days of testing. However, in contrast to MB-lesioned ani- mals, MD-lesioned mice still significantly differed from controls at the end of the learning phase. If the experiment was stopped on day 8 of testing, we could have concluded that MD-lesions but not MB ones impaired the cognitive ability to develop a learning set rule and/or significantly
A. Krazem et al. / Behaviowal Brain Research 67 (1995) 51-58
14 -
12 -
8 *g lo-
'6 s
8-
-3 s 6-
4-
2-
6 Conrrols AMB ??MD
I I I 1 2+3 4+5
Days
B5-
45 I Smin 24h
Retention
Fig. 4. Left: mean number of trials required to meet the criterion in Control, MD and MB groups over days of testing during successive reversal ses- sions (see text). **: PtO.05; ***: P< 0.01. Right: per cent correct responses in Control, MD and MB groups on retention sessions occuring either 5 min or 24 h each daily discrimination. No significant differences were observed between the groups.
slowed down the rate of learning of such a rule. However, our study demonstrates that the impairments observed in the MD group are not due to a learning deficit sensu stricto. Thus, increasing the intersession interval from 24 h to 10 days led to a paradoxical result, since MD subjects not only exhibited performance similar to those of the other groups (which exhibited a substantial saving of perform- ance) but signiticantly improved their own performance as compared to that measured at the end of the learning phase, both in reversal and repetition conditions. The fa- cilitative effect of a lo-day intersession interval on per- formance clearly shows that MD subjects have no learn- ing deficit per se nor have they developed anterograde amnesia for the information associated with the last dis- crimination acquired. Thus, one may suggest that the im- pairment of MD-lesioned subjects observed during the learning phase of the task reflects a pefirmance deficit possibly due either to a deficit of behavioral inhibition, as suggested by the increase of the number of errors observed on the first reversal session (Day 2 of testing) or to an intrusion of previously acquired informations on subse- quent daily sessions (exagerated sensitivity to proactive interference). Indeed, the fact that, in all groups, the num- ber of trials required to meet the criterion in repetition sessions is less as compared to the one required in reversal ones shows that, over days of testing and in the reversal condition, the informations acquired at a given session had a detrimental effect on the acquisition of the subse- quent discrimination. Thus, one may suggest that length- ening the intersession interval from 24 h to 10 days would result in a reduction of the strength of interference and/or to a decrease of eventual response perseveration tenden- cies in MD-lesioned mice.
The lack of any important deficits in the learning set
task in MD and MB-lesioned animals reported in the present study is in agreement with previous findings per- formed in the rat [2] but contrasts with other studies having shown that MD damage produced impairments in reversal discriminations tasks [7,21,22]. However, in ad- dition to important procedural differences between the studies, the different technics used to damage the MB and MD (ibotenic acid vs. electrolytic lesions) may account for the discrepencies between the studies. Indeed, it is now well established that more important behavioral deficits are observed following electrolytic lesions as compared to ibotenic acid ones [8].
The patterns of deficits observed here following MD and MB lesions contrast with previous reports showing that lesions of the anterior or posterior cingulate cortex (which are the direct and indirect target sites of the out-
??Kepetition
-----7 El Reversal
Controls MD MB Controls MD MB
Day 8 Day 18
Fig. 5. Mean number of trials required to meet the criterion in Control, MD and MB groups on Day 8 (left) and Day 18 (right) of testing (intersession interval: 10 days) either in reversal or repetition condition (see text).

58 A. Krazem et al. 1 Behavioural Brain Research 67 (199s) 51-58
puts originating from the MD and the MB respectively in mice) led to important and long-lasting deficits in a simi- lar task and mouse strain [ 181. Several explanations may account for this discrepancy. Firstly, it is possible that the loss of cortical afferents resulting from single diencepha- lit damage are compensated by the increase of the activ- ity of secondary pathways over the experiment, reducing thereby the eventual cognitive impairments induced by the lesion. Secondly, the information processing used to solve the task may be more particularly sensitive to a frontal dysfunction, but not to a diencephalic one. Indeed, the task may be solved by aplying from day to day a simple win-stay rule, which certainly involves the use of an ego- centric body-turn strategy, since mice are required to make five successive identical runs into the same arm to meet the criterion. It has been shown that frontal lesions disrupt learning task involving the use of egocentric informations [lo]. Thus, an impairment in the processing of egocentric informations may be superimposed to and/or causal of rule learning impairments in cortical-lesioned subjects; in contrast, the MB and MD may not be involved in pro- cessing egocentric strategy, and/or not disruptive enough of the frontal activity to induce impairments.
Acknowledgements
This study was supported by the CNRS, URA339. We thank Dr. Thomas Durkin for his help in writing this manuscript.
References
111
121
[31
[41
[51
Aggleton, J.P. and Sahgal, A. The contribution of the anterior thalamic nuclei to anterograde amnesia, Neuropsychologia, 3 1 (1993) 1001-1019. Beracochea, D., JalIard, R. and Jarrard, L.E., Effects of anterior or dorsomedial thalamic ibotenic lesions on learning and memory in rats, Behav. Neural Biol., 51 (1989) 364-376. Beracochea, D. and JalTard, R., Efects of anterior thalamic lesions on spatial memory in mice, Neuroreport, 5 (1994) 917-920. Bitterman, M.E., The evolution of intelligence, Scient. American, 212 (1965) 92-100. Brian, S., Mikol, J. and Plas, J., Neuropathologie des syndromes amnesiques chez l’homme, Rev. Neurol., 141 (1985) 627-643.
[61
I71
[81
[91
[lOI
[Ill
I121
[I31
1141
iI51
[I61
[I71
[I81
[I91
PO1
P11
t2.21
u31
Dusoir, H., Kapur, N., Bymes, D.P., MC Kinstry, S. and Hoare, R.D., The role of diencephalic pathology in human memory dis- orders, Brain, 113 (1990) 1695-1706. Holmes, E.J., Butters, N., Jacobson, S. and Stein, B.M. An ex- amination of the effects of mammillary-body lesions on reversal learning sets in monkeys, Physiol. Psychol., 11 (1983) 159-165. Jarrard, L.E., On the use of ibotenic acid to lesion selectively dif- ferent components of the hippocampal formation, J. Neurosci. Methods, 29 (1989) 251-259. Joyce, E.M. and Robbins, T.W., Frontal lobe function in Korsa- koff and non-Korsakoff alcoholics: planning and spatial working memory, Neuropsychology, 8 (1991) 709-723. Kesner, R.P., Farnsworth, G. and Di Mattia, B.V., Double disso- ciation of egocentric and allocentric space following medial pre- frontal and parietal cortex lesions in rat, Behav. Neurosci., 5 (1989) 956-961. Kolb, B., Functions of the frontal cortex of the rat: a comparative review, Brain Res. Bull., 8 (1984) 65-98. Kolb, B., Pittman, K., Sutherland, R.J. and Wishaw, I.Q., Disso- ciation of the contributions of the prefrontal cortex and dorsome- dial thalamic nucleus to spatially guided behavior in the rat, Behav. Bruin Res, 6 (1982) 365-378. Lhermitte, F. and Signoret, J.L., The amnesic syndromes and the hippocampal-mammillary system. in M.R. Rosenzweig and E.L. Bennet (Eds.), Neural Mechanisms of Learning and Memory, MIT Press, Cambridge, MA, 1976, pp 49-56. Lishman, W.A., Cerebral disorders in alcoholism: syndromes of impairments, Brain, 104 (1981) l-20. Mair, W.G.P., Warrington, E.K. and Weiskrantz, L., Memory dis- orders in Korsakoff s psychosis: a neuropathological and neurop- sychological investigations in two cases, Bruin, 102 (1979) 749- 783. Markowitsch, H.J., Thalamic dorsomedial nucleus and memory: a critical evaluation of studies in animals and man, Neurosci. Biobe- hav. Rev., 6 (1982) 351-380. Meunier, M., Le cortex cingulaire: connexions neuroanatomiques et implication dans les processus d’apprentissage et de memoire chez la souris, These, Universite de Bordeaux 1, 1988, pp. 221. Meunier, M., JaIIard, R. and Destrade, C., Differential involvement of anterior and posterior cingulate cortices in spatial discriminative learning in a T-maze in mice, Behav. Brain Res., 44 (1991) 133- 143. Oscar-Berman, M. and Zola-Morgan, S.M., Comparative neurop- sychology and Korsakoffs syndrome. I. Spatial and visual rever- sal learning, Neuropsychology, 18 ( 1980) 499- 5 12. Schacter, D.L., Memory, amnesia, and frontal lobe dysfunction, Psychobiology, 15 (1987) 21-36. Slotnick, B.M. and Kaneto, N., Role of the dorsomedial thalamic nucleus in olfactory discrimination learning in rats, Science, 214 (1981) 91-92. Staubli, U.,Schottler,F. and Nejat-Bina, D., Role of the dorsome- dial thalamic nucleus and piriform cortex in processing olfactory information, Behav. Brain Res., 25 (1987) 117-129. Winocur, G., The hippocampus and prefrontal cortex in learning and memory: an animal model approach. In Neuropsychology of Memory, 2nd edn., The Guilford Press, 1992. pp. 429-439.
Copyright © 2022 FDOKUMEN




















