
EFFECTS OF DENSITY AND PROPORTION ON SPRING
115
AN ABSTRACT OF THE THESIS OF JULIE ANN CONCANNON for the degree of MASTER OF SCIENCE in CROP SCIENCE presented on DEC. 4, 1987 Title: EFFECTS OF DENSITY AND PROPORTION ON SPRING WHEAT AND LOLIUM MULTIFLORUM LAM. Abstract approved: Redacted for Privacy Steven R. Radosevich This thesis consists of four chapters. Literature is reviewed in Chapter 1. Chapter 2 describes an addition series experiment to determine the influence of species density and proportion on vegetative and reproductive yield of spring wheat and Lolium multiflorum. Chapter 3 consists of a growth analysis experiment to determine whether proximity factors and time and temperature (incorporated into growing degree day units) affect the growth rates of species height, leaf number, tiller number, and leaf area index. The final chapter contains conclusions drawn from both experiments. A review of competition literature indicates that yield-density relationships can be studied using additive studies, replacement series, and addition series experiments. Intra- and interspecific competitive responses may be quantified best with the latter two approaches. Additive studies confound variables of species
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of EFFECTS OF DENSITY AND PROPORTION ON SPRING

AN ABSTRACT OF THE THESIS OF
JULIE ANN CONCANNON for the degree of MASTER OF SCIENCE
in CROP SCIENCE presented on DEC. 4, 1987
Title: EFFECTS OF DENSITY AND PROPORTION ON SPRING
WHEAT AND LOLIUM MULTIFLORUM LAM.
Abstract approved: Redacted for PrivacySteven R. Radosevich
This thesis consists of four chapters. Literature is
reviewed in Chapter 1. Chapter 2 describes an addition
series experiment to determine the influence of species
density and proportion on vegetative and reproductive yield
of spring wheat and Lolium multiflorum. Chapter 3 consists
of a growth analysis experiment to determine whether
proximity factors and time and temperature (incorporated
into growing degree day units) affect the growth rates of
species height, leaf number, tiller number, and leaf area
index. The final chapter contains conclusions drawn from
both experiments.
A review of competition literature indicates that
yield-density relationships can be studied using additive
studies, replacement series, and addition series
experiments. Intra- and interspecific competitive
responses may be quantified best with the latter two
approaches. Additive studies confound variables of species

density and
adequately
Spring
multiflorum
proportion and therefore, do not assess
competitive responses of weed and crop.
wheat (Triticum aestivum) (L.) and Lolium
(Lam.) were grown together in an addition
series to examine the influence of species density and
proportion on vegetative and reproductive biomass. Wheat
grain yields indicated that yield increased with wheat
monoculture density. However, the increase in grain yield
was not linear, indicating that constant final yield
occurred at high wheat monoculture densities. Grain yield
decreased with increasing ryegrass density. These
observations suggest that both intra- and interspecific
competition occurred during the study. Grain yield was
highly correlated with total biomass. Therefore mean plant
biomass was used as a competitive
predictive regression models.
Regression results indicate that wheat was a more
effective competitor than ryegrass over all species
relative and total densities. A model was developed by
regressing mean reciprocal weight as a response to final
densities of both species. Relative competitive abilities
were calculated for each species. The relative competitive
ability of wheat suggests that one wheat plant was as
competitive as 6.7 ryegrass plants. However, wheat also
was more competitive with itself than with ryegrass.
response for developing

Ryegrass relative competitive ability indicated that one
ryegrass plant was as competitive as one wheat or ryegrass
plant.
Species density and proportion and time and
temperature (incorporated in growing degree day units
(GDD)) were included in the regression model as independent
variables to describe relative rates of plant growth for
both species. Growth parameters (seed size, height, leaf
number, tiller number, and leaf area index (LAI)) were
measured throughout the growing season. Analysis of
variance and covariance on relative rates of growth and
seasonal LAI indicated that proximity factors of species
density and proportion influenced relative rates of growth
and seasonal LAI of both species. GDD was usually a
significant variable in the regression equations,
suggesting that time and temperature also influenced rates
of growth.
The regression equations describing relative rates of
growth indicate that wheat, by virtue of a large seed size
and high early season LAI, captured a large share of
resources by germinating fast and quickly overtopping
ryegrass in the mixture. The trend continued throughout
the season. However, suppressed ryegrass plants maintained
a constant rate of growth. Ryegrass eventually grew as
tall or taller than wheat plants and produced a higher LAI

than wheat in mixtures. This may be a growth strategy
which makes ryegrass a persistent weed in spring wheat
cropping systems.

THE EFFECTS OF DENSITY AND PROPORTION
ON SPRING WHEAT AND LOLIUft MULTIFLORUM LAM.
by
Julie Ann Concannon
A THESIS
submitted to
Oregon State University
in partial fulfillment ofthe requirements for the
degree of
Master of Science
Completed Dec. 4, 1987
Commencement June 1988

APPROVED:
Redacted for Privacyi-Professor of Crop Science
in charge of major
Redacted for Privacy
Head of Department of Crop Science
A I
Redacted for PrivacyDean of lraduate School
Date thesis presented Dec. 4, 1987

ACKNOWLEDGMENTS
The first question I asked Dr. Radosevich when we metwas "What is your philosophy about training graduatestudents?" He answered "I train students to be weedscientists". Over the past two years I have had a thoroughtraining from him in logic, organization, and scientificanalysis for which I am truly grateful. He has produced ascientist still filled with enthusiasm for the work whichshe has been trained to do. I consider this a greataccomplishment and thank him for all the time and energywhich he invested in me during these last two years.
Much of the impetus for the problem which Iinvestigated came from the work which Mary Lynn Roush hasdone in the last six years. She walked with me througheach step of my graduate program and helped me in areas ofprofessional as well as personal growth. For her smile,her patience and all her help on those mornings when shereally needed to get work done (but stopped to help me) Ithank her.
Special thanks go the other members of my graduatecommittee. To Dr. Arnold Appleby who taught me thatsilence and listening are truly golden, to Dr. Mark Wilsonwho taught me how to take literary criticism, and to Dr.Richard Dick who took an interest in my study even when itwas in its infant stages.
There are several areas one hopes to improve on duringtheir graduate studies. Bruce Maxwell helped me to seeboth sides of a scientific problem and beyond, by constantthought "provoking" and I thank him for that provoking! Mycomputer skills which were nil when I began, have improvedremarkably because of all the help I received from BobWagner. Tim Harrington provided much needed academic andpersonal support. For those days when I felt the leastconfident, I thank him for his boost of enthusiasm andcaring.
In each persons life there is one person whom theyrely on for the bulk of their emotional support. Myhusband Tom DeMeo supplied all the love and emotionalsupport which I required to make it through this graduateprogram. I thank him for his acceptance of the fact that I

do not do dishes.
This thesis is dedicated to two very special women.First, to Mrs. DeMoss, who stimulated my first scientificthoughts with her "imitation" of a Pronuva moth landing ona yucca plant, in my ninth grade general science course andsecondly, to my mother Margaret Concannon, who continues tomotivate me with her enthusiasm for life and encouragementof my scientific endeavors even if they do take place inthe "dirt".

TABLE OF CONTENTS
Page
CHAPTER 1: INTRODUCTION 1
Methods to Study Competition 2Expanded Reciprocal Yield Approach 5Interaction of Environmental and Biological Factors 7Growth Measurements and Their Implications 9Study Description and Objectives 10
CHAPTER 2: EFFECTS OF SPECIES DENSITY AND PROPORTIONON VEGETATIVE AND REPRODUCTIVE GROWTH OFSPRING WHEAT AND LOLIUM MULTIFLORUM 13
Introduction 13Methods and Materials 15Results and Discussion 20Conclusions 25Chapter 2 References 44
CHAPTER 3: QUANTIFYING GROWTH RESPONSES OF SPRINGWHEAT AND LOLIUM MULTIFLORUM 47
Introduction 47Methods 52Results and Discussion 56Summary 63Chapter 3 References 70
CHAPTER 4: CONCLUSIONS 73
Measuring Competition 73Importance of Competition 77Agricultural Implications 78
BIBLIOGRAPHY 80
APPENDICES 84
Tiller Number 84Individual Plot Measurements 85

LIST OF FIGURES
Figures Page
2.1 Total grain yields/ha resulting from additionseries experiments 36
2.2 Individual wheat plant biomass predicted byexpanded reciprocal approach 38
2.3 Individual ryegrass plant biomass predicted byexpanded reciprocal approach 40
2.4 Individual ryegrass plant biomass predicted byregressing both species densities and a speciesdensity interaction 42
3.1 Wheat and ryegrass height growth ratesplotted against growing degree days 68
a.1 Final mean number of wheat and ryegrasstillers plotted against planted densitiesof both species 101

LIST OF TABLES
Tables Page
2.1 Addition series design 27
2.2 Mean results of 4 replications of spring wheatLollum multiflorum addition series. Totalbiomass and wheat grain yields 28
2.3 Mean results of 4 replications of spring wheatand bolium multiflorum addition series. Speciesmean plant weight and reciprocal weights 30
2.4 Wheat harvest indices calculated for Owensand Waverly spring wheat, describing therelationship between total biomass and grainyield 32
2.5 Reciprocal yield models describing intra-specific competition between wheat andryegrass 33
2.6 Reciprocal yield models describing intra-and interspecific competition of wheat andand ryegrass as influenced by species densityand proportion 34
2.7 Reciprocal yield model predicting ryegrassreciprocal yield with independent variablesof species density and density interaction 35
3.1 Relative height growth rates of ryegrassand wheat explained by independent variablesof proximity factors, growing degree days,and interactions
3.2 Relative leaf initiation rates of ryegrass
65

and wheat explained by independent variablesof proximity factors, growing degree days,and interactions
3.3 Regression equations describing speciesleaf area index with independent variablesof proximity factors, growing degree days,and interactions
66
67
a.1 Relative tillering rates explained byindependent variables of proximity factors,growing degree days, and interactions 86
a.2 1986 spring wheat-Lolium multiflorum additionseries results. Individual plot results oftotal biomass and wheat grain yields 87
a.3 1986 spring wheat-Lolium multiflorum additionseries results. Single plot calculations formean plant weight and reciprocal plant weight....94

The Effects of Density and Proportion On SpringWheat and Lolium multiflorum Lam.
CHAPTER 1: INTRODUCTION
Weeds are unwanted plants that reduce crop yield.
Many experiments have been conducted quantifying grain
losses when weeds invade a fixed crop monoculture. The
most common experimental method to quantify grain loss is
the additive experiment. The total density of the crop and
weeds change in an additive experiment, confounding the
effects of density and proportion (Radosevich and Holt
1984). Density is the number of plants per area.
Proportion is the percentage of species in a mixture. When
these two variables are confounded it is difficult to
assess the influence of competition from the weed
(interspecific) and the crop (intraspecific). Quantifying
plant yield-density relationships between crops and weeds
should lead to better crop management strategies
(Radosevich 1987).
Most of the early work on yield-density relationships
was done with crop monocultures. Shinozaki and Kira (1956)
established a yield-density relationship between reciprocal
plant weight and monoculture density. The curvilinear
relationship that results from plotting individual plant
weight against monoculture density is a negative
rectangular hyperbola. Plant weight decreases

2
asymptotically as monoculture density increases. In order
to linearize this relationship, individual plant weight
often is transformed to its reciprocal and plotted against
species density.
On a population level, total biomass increases with an
increase in density until all available space is occupied.
Constant final yield is then achieved (Harper 1977).
Individual plants in the population often may become large
but only from independent mortality of other plants in the
population (Yoda 1962, Westoby 1981). This information has
been used to study mixed populations under the assumption
that density creates an additive response. Several methods
can be used to quantify yield-density relationships between
weeds and crops in a mixed population. The most frequently
used methods are the additive experiment, replacement
series, and addition series experiments.
Methods to Study Competition
The additive approach has been utilized in weed
control experimentation for the last twenty years.
However, confounding factors of species density and
proportion limit experimental interpretation using this
approach (Radosevich 1987). In addition, Zimdahl (1980)
reviewed many of the additive experiments and concluded
that interpretations of many studies were confounded by

3
site specificity. There has been little effort to separate
site quality factors (e.g. soil bulk density,
precipitation, humidity) from competition factors (e.g.
species density and proportion). Furthermore, in many
experiments, competition from the crop on itself or other
plants has been ignored or underestimated.
Carlson and Hill (1986) attempted to examine the
influence of interspecific (wild oat on wheat) and intra-
specific (wheat on wheat) competition on crop yield in a
comprehensive additive experiment. They observed that wild
oat always decreased the yield of wheat, but that its
influence diminished as total density of the stand
increased. Although Carlson and Hill (1986) were able to
determine a general trend, the competitive responses of the
two species could not be quantified adequately because
variables of density and proportion were varied in an
unsystematic manner (Radosevich 1987). In contrast, many
ecological studies use replacement series experiments which
do systematically vary proportions of two species at a
single total density.
The replacement series approach (de Wit 1960, Harper
1977) traditionally has been used to assess the competitive
influence of two species. Harper (1977) suggests that mean
yield of each species depends on relative proportions of
the species and the total density. Replacement series

4
experiments are interpreted qualitatively with four
possible results (models) representing competition, mutual
antagonism, mutual benefit, and no interaction (Harper
1977). It is possible to determine the relative influence
of intra- and interspecific interference using the
replacement series, but absolute effects cannot be
partitioned (Radosevich 1987). Jolliffe et al. (1984)
attempted to partition intra- and interspecific competition
by incorporating several monoculture densities of the
species proportions into the design. A no-interaction
monoculture line, an interaction monoculture line, and an
inter-specific interaction line were generated (Jolliffe et
al. 1985). By comparing each line, the effects of intra-
and inter-specific competition can be partitioned.
Jolliffe et al. (1984) quantitatively describe data by the
equation:
Rx = (Ym-Yx)/Ym
where Rx is the relative mixture response, Ym is the
experimental yield in monoculture, and Yx is the
experimental yield of at the proportion x. Rx depends on
the particular proportion used. Several replacement series
must be planted in order to assess competition over several
total densities. This requires substantial space and time
to accomplish, a major weakness in using replacement series

5
to assess competitive effects in field experiments. An
addition series can be used to generate a large number of
total and relative densities to assess competitive effects
in the most efficient amount of space.
The addition series approach (Miller and Werner
1987, Radosevich 1987, Roush and Radosevich 1987) generates
a systematically changing array of total and relative
densities in competition experiments, to increase
information which a replacement series is not able to
impart. Although adequate information can be generated
from this method, quantitatively assessing several relative
and total densities of two or more species often has been
difficult (Willey 1979). Few analyses have proven
satisfactory (Willey 1979, Spitters 1983a). Multiple
linear regression can be used to partition intra- and
interspecific competition between two species (Spitters
1983ab, Connolly 1986, Roush and Radosevich 1985).
Absolute effects of competition occurring in an addition
series experiment can be quantified by producing an
expanded reciprocal yield model based on species density
and proportion (Spitters 1983a). Mean reciprocal weights
are regressed as a response to each species density to
produce intra- and interspecific competition coefficients,
which estimate population parameters of species biomass
response to density.

6
The Expanded Reciprocal Yield Approach
The expanded reciprocal yield approach provides a
method to determine resource utilization of each species
according to its relative proportion in the mixture.
Proximity factors, the main factors manipulated by the
farmer, determine the amount of space initially available
to each plant. Space is the composite of all resources
initially available to a plant and the environmental
conditions which a plant experiences (Radosevich and Holt
1984). Total density of a population is assumed equally
additive because initially space is allocated equally by
the planting design in an addition series.
Several assumptions result from the basic premise of
additivity. An expanded reciprocal yield competition model
built from monoculture and mixture densities assumes plants
take up resources additively and equidistant spacing
provides equal allocation of initial available resources.
However, plants may not use resources or compete equally
given an equal amount of space (Watkinson 1980) because of
different germination times or initial plant size.
Incorporating a density interaction term into the model may
facilitate the description of how proportional differences
of each species are affecting the maximum resource uptake
ability of both species.
The expanded reciprocal approach assumes that both

7
species are at maximum resource uptake (Bleasdale 1967,
Spitters 1983ab). If one species is below maximum resource
uptake, the expanded reciprocal yield approach may over-
estimate the biomass response of that species to increasing
species density and proportion. The crop and weed may take
up resources at different efficiencies and the degree of
efficiency also may change with time (Firbank & Watkinson
1987).
The mean reciprocal weight also is assumed to be the
best transformation of the experimental data. Linearizing
the data for regression is one of the most important steps
in building a regression model (Neter and Wasserman 1983)
to describe the influence of intra- and interspecific
competition. Several other factors besides competition
also may influence individual plant weight. Accounting for
unexplained variation in the regression model is important.
Interactions of Environmental and Biological Factors With
Proximity Factors
Modeling individual plant biomass response involves
complex interactions of biological and environmental
factors (Roush and Radosevich 1985, Radosevich 1987) in
addition to yield-density relationships among species. To
quantify interference between crop and weed, as many of
these factors as possible should be determined and

8
controlled.
A model describing plant competition in terms of
biologically meaningful parameters includes biological and
environmental factors such as seasonal differences,
microenvironment and disease. These factors also
ultimately influence plant growth. Seed bank dynamics
including dormancy and survival also may influence initial
germination rates. Germination in turn is influenced by
temperature and degree of disturbance (tillage). All of
these factors and their interactions should be considered
when quantifying yield-density relationships between
plants. Each of these factors and their interactions with
proximity factors of species density and proportion may
contribute to the ability of a species to utilize available
resources.
Hypothetically, plants have available to them a
reservoir of soil- associated resources. The resource pool
in the soil may fall short of the consumption needs of
plants, resulting in competition.
Competition for light differs from competition for
other resources because there is no resource pool from
which the plant can draw light. Light, in the form of
photons, is available only briefly and must be used
immediately or lost (Radosevich and Holt 1984). The
physical position of the plant in the canopy, therefore,

9
becomes an important factor for light interception (Donald
1961, Radosevich and Holt 1984).
Since plants integrate their resources into plant
biomass (Roush and Radosevich 1985), monitoring rate of
plant growth may elucidate how fast resources are being
captured by each species. DeWit (1960) and Harper et
al.(1965) have suggested that a plants ability to capture
resources may impart a competitive advantage. When plant
parameters of LAI, leaf initiation, tiller number, and
height are measured in an addition series, a framework is
established for determining the influence of proximity
factors on both crop and weed growth rates.
Growth Measurements and Their Implications
Weeds, in competing for primary resources of water,
light and mineral nutrients, may cause a reduction in leaf
area of the crop plant (Patterson 1982, Zimdahl 1980,
Harper 1977). Harper et al. (1965) suggests that the
success of weeds is usually due to a weed species becoming
established earlier than crop plants and maintaining a
rapid growth rate. Leaf area, a measurement of the light
gathering capability of a plant has been studied
extensively. Patterson (1982) points out that one of the
factors contributing to the success of agronomic weeds is
the rapid partitioning of plant biomass into leaf area, and
consequently rapid development of canopy shade over the

10
crop. However, crop species also have been shown to
rapidly partition plant biomass into leaf area,
successfully suppressing a weed species. For example,
soybean and peanut were effective in suppressing Henry
crabgrass (Digitaria adscendens H.B.K.), chufa (Cyperus
microiria Steud.), and common purslane (Portulaca oleracea
L.) (Noguchi & Nakayama 1978b).
Several other plant characteristics can be influenced
by species density and proportion. Sestak (1983) suggests
that measuring height, leaf number initiation, and leaf
area index (LAI) could relate how a plant captures
resources over time. Peterson et al. (1982) indicates
that stress also can be measured by the number of tillers
that a plant produces over time. Stress is defined as
external phenomena that limit production, such as reduced
light intensity, water availability, nutrients, or sub-
optimal temperature (Grime 1979).
Study Description and Objectives
Spring wheat (Triticum aestivum) (L.) and Italian
ryegrass (Lolium multiflorum) (Lam.) were chosen to study
the influence of intra- and interspecific competition
between two species using the expanded reciprocal yield
approach. Italian ryegrass is a winter annual grass that
invades winter wheat fields of western Oregon, Washington,

11
and California (Appleby et al. 1976), and reduces wheat
yield. Appleby et al. (1976) indicate that ryegrass may
reduce nitrogen availability to the wheat crop causing
grain yield reductions. However, density and proportion of
the wheat and ryegrass varied in this additive experiment;
thus the influence of intra- and interspecific competition
was confounded. In this thesis, an addition series has
been used to systematically vary the density and proportion
of each species. This experimental approach generates
monoculture and mixture responses of each species in the
experiment. These competitive responses suggest resource
use of each species in the experiment as a response to
variation in species total and relative densities.
Environmental factors such as temperature, light,
water, and nutrients are relatively homogenous and
synchronous in an agricultural system (Roush and Radosevich
1985). Spring wheat and Italian ryegrass are similar C3
grasses that exhibit similar life form, life history, and
phenotypic responses. The species also emerge
simultaneously with cultivation in the spring. By
recognizing environmental and biotic factors that influence
plant growth, this experiment focuses on the proximity
factors of species density and proportion affecting growth
rates and production of both crop and weed.
The primary objective of this study was to quantify

12
antra- and interspecific competition between spring wheat
and Italian ryegrass. A second objective was to elucidate
mechanisms by which ryegrass and wheat might capture
environmental resources. Functional growth analysis (Hunt
1982) was used to compare growth rates of the species in
monoculture and mixture over time.

13
CHAPTER 2: EFFECTS OF SPECIES DENSITY AND PROPORTION ONVEGETATIVE AND REPRODUCTIVE YIELD OF SPRING WHEAT AND
LOLIUM MULTIFLORUM
Introduction
Italian ryegrass (Lolium multiflorum) (Lam.) is an
annual weed that often invades winter wheat fields
throughout western Washington, Oregon, and California
(Appleby et al. 1976). This weed species has been shown to
interfere with wheat, resulting in significant grain loss.
Appleby et al. (1976) demonstrated a decrease in wheat
yields by increasing nitrogen levels and ryegrass density
in an additive experiment. However, the experimental
variables of total plant density, species proportion, and
resource availability (nitrogen levels) were confounded in
this experiment (Harper 1977, Radosevich and Holt 1984,
Radosevich 1987), making the relative contributions of
intra- and interspecific competition and resource
availability difficult to assess.
Carlson and Hill (1986) examined the influence of wild
oat (Avena fatua) on wheat yields by varying the densities
of both crop and weed. They observed that wild oat reduced
wheat yield at each crop density. However, the influence
of wild oat on wheat yield declined as the total density of
the stand increased. Although the approach of Carlson and
Hill (1986) diminished the influence of two simultaneously
varying factors in their experiment, it was difficult to

14
distinguish adequately between the influence of intra- and
interspecific competition (Carlson and Hill 1986,
Radosevich 1987). The distinction between intra- and
inter-specific influence is necessary to establish
biological and economic thresholds for crop and weed
species (Radosevich 1987, Spitters 1983ab, Cousens 1986).
Multiple linear regression has been proposed as a
method to assist in the quantification of intra- and inter-
specific competition between weed and crop species
(Spitters 1983ab, Firbank and Watkinson 1987). In this
approach, the influence of competition is assessed by
producing an expanded reciprocal yield model based on final
densities and proportions of each species. Reciprocal
weights are regressed as a response to the densities of
both species to produce intra- and interspecific
competition coefficients, which indicate competitive
ability. The approach provides a means to assess the
influence of each species on crop or weed yield according
to their proportions (relative densities) in the
association (Spitters 1983ab, Roush and Radosevich 1987,
Radosevich 1987).
The primary objective of this study was to develop a
gradient of interference between spring wheat and Italian
ryegrass by systematically varying densities and
proportions of each species in monocultures and mixtures.

15
The second objective was to partition intra- and inter-
specific effects between the two species using an expanded
reciprocal yield approach and multiple linear regression.
The final objective was to describe the relative
competitive abilities of the two species given changes in
both density and proportion of the plants.
Materials and Methods
Experimental Approach
Two cultivars of soft white spring wheat (Triticum
aestivum (L.) var. Owens (Sunderman) and var. Waverly
(Konzak) were selected to determine the competitiveness of
Italian ryegrass Lolium multiflorum (Lam.) var. Marshals.
Waverly and Owens wheat cultivars have height potentials of
84 and 94 cm, respectively.
An addition series experiment was used to
systematically combine six monoculture densities of wheat
with six corresponding monocultures of ryegrass, resulting
in 36 relative and total densities (Miller and Werner 1987,
Radosevich 1987, Roush and Radosevich 1987). The
monoculture densities were 0, 9, 36, 61, 220, and 440
plants per m2 for each species. These monoculture
densities and their combinations created an array of total
densities and species proportions with 36 total densities
ranging from 0 to 880 plants/m2 (Table 2.1).

16
Two experiments were established, one for each wheat
variety. Each experiment was conducted as a split-strip
design. Species densities (treatments) were planted in
strips established in opposite directions (Little and Hills
1978). Each experiment was replicated four times.
Individual plots were 1.44 m2. The outer 0.50 m 2
of each
plot represented a border area between adjacent treatments.
One block, ie. replicate, encompassed 8.5 m2
.
The study was conducted at the Oregon State University
Hyslop Crop Science Field Laboratory, near Corvallis,
Oregon. The soil was a sandy clay loam. Annual
precipitation for the year was 108 cm. Major rains
occurred in mid-February, early May, and early July. The
mean daily minimum and maximum temperatures for the growing
season were 8.3 and 22.8 C , respectively, averaging
slightly higher than the 100-year mean for the Corvallis
area. Wheat was hand-planted using a template to maintain
equidistant spacing among individual plants (seeds). Wheat
was planted to an excess of 20 percent to obtain 100
percent germination. Ryegrass, at the appropriate density,
was hand-sprinkled uniformly over each plot containing that
species. Total germination was 99-100 percent for each
species. Following emergence, wheat was thinned to the
appropriate density. The site was fertilized with 50 kg/ha
ammonium nitrate, disked into the soil prior to planting.

17
An additional 15 kg/ha ammonium sulfate was applied at the
end of May. The experiment was irrigated to field capacity
at the "boot stage" of wheat on June 3, and one week later.
Removal of other weed species was performed by hand
throughout the growing season.
Data Collection
Plants were harvested 110 days after emergence from a
0.75 m2area in the center of each plot. Individuals of
each species were counted, and actual species densities and
proportions were determined (Table 2.2). Total plant
biomass was removed at soil level, and plants were
separated by species. Whole plants were dried to constant
weight at a temperature of 15.6 C for 48 h, and total
biomass for each plot was weighed. Wheat and ryegrass
plants were threshed, and seed weights were recorded (Table
2.2).
Total biomass for each wheat variety was regressed
against total wheat grain yield to obtain a wheat harvest
index. The wheat harvest index provided a basis for using
biomass as criteria for assessing the affect of competition
on grain yields.
Model Development
Developing a regression model to describe mean
reciprocal yield based on species density and proportion

18
required several steps. Individual plant yield should
decrease asymptotically as a population reaches constant
final yield. If individual plant biomass response forms a
negative rectangular hyperbola with density, the reciprocal
transformation of mean biomass can be used to perform
multiple linear regression (Neter and Wasserman 1983).
Both wheat and ryegrass mean weights were plotted against
final species densities in monoculture and mixture. Both
species data resembled a negative rectangular hyperbola.
However, the ryegrass data displayed a large amount of
variation at high densities. The high degree of variation
in mean plant weight occurring in the higher densities, as
suggested by the residual error terms of each regression
equation, indicated that some of the variation in mean
reciprocal ryegrass weight was not explained by the species
densities. The reciprocal transformation of mean plant
biomass was used when comparing the relative competitive
abilities of each species.
The following reciprocal yield model was used to
describe intraspecific competition when plants were grown
in monoculture:
1/w = b10 1311N1-
The mean reciprocal weight (1/W) is described by the
intercept bio, the theoretical reciprocal weight of a plant

19
growing without competition and the regression coefficient
b11 which describes the degree of intraspecific competition
resulting from additional plants (N) to the population.
The intraspecific competitive ability of each species was
calculated by the ratio b0 /b1. If the addition of plants
to a monoculture influences the reciprocal weight (1 /W)
additively then adding plants of another species (given an
equal amount of space) should affect the reciprocal weight
in a similar manner (Bleasdale 1967, Spitters 1983ab). An
expanded reciprocal model that included both species was
developed to describe the effects of intra- and
interspecific competition:
1/W = b10 b11N1 b12N2
The coefficients b10 and b11 are as described earlier. The
regression coefficient b12 describes interspecific
competition occurring with each additional plant of the
"competing" species. A value for the relative competitive
ability (RCA) of each species was obtained by dividing the
coefficient of intraspecific competition (b11) by the
coefficient of interspecific competition, b12.
An all possible R 2procedure (SAS 1986) and stepwise
regression (SAS 1986) were performed to determine the
importance of the independent variables of species density
and proportion used to describe mean reciprocal plant

20
weight for both species. Finally, analysis of covariance
was performed using the wheat varieties, experimental
variables, and interactions to describe the reciprocal
weight of both species. The wheat varieties were used as
covariants in the analyses. However, since there were no
significant reciprocal biomass differences between
varieties in monoculture or mixture, the data were pooled
to create a regression model quantifying intra- and inter-
specific competition.
Results and Discussion
Total Grain Yields
Total grain yield per plot increased with increasing
wheat monoculture density (Table 2.2 and Figure 2.1).
However, the increase in total grain produced was not
linear. These data indicate that wheat densities became
saturated at 220-440 plants/m 2when constant final yield
occurred. Mean plant yield decreased as total wheat
density increased (Table 2.3), but total wheat grain yield
(Table 2.2) continued to increase due to large increases in
monoculture density. The short wheat variety (Waverly)
yielded less than the tall wheat variety (Owens) (Figure
2.1).
As ryegrass was added to the population, grain yields
decreased (Figure 2.1 and Table 2.2). However, the

21
decrease in grain yield between that of the highest wheat
density monoculture and the highest total density (880
plants/m2) was not proportional to the species density in
mixture. Furthermore, when wheat density was low and
ryegrass density was high in mixture, a significant
decrease in wheat grain yield was observed. These data
indicate that intra- and interspecific competitive
responses change with species density and proportion.
Systematic varying of species density and proportion
demonstrates the influence of both weed and crop on each
species.
These results support the observation that wheat
plants at the high densities acted as an overstory
population that suppressed ryegrass. However, wheat plants
also competed with other wheat plants, decreasing
individual wheat yield.
Wheat Harvest Index
To develop a relationship between total biomass and
total grain yield, a harvest index was developed for each
variety. Total biomass was regressed against total grain
yield (Table 2.4). Wheat grain yield (Kg) was determined
from an intercept value, plus a regression coefficient
which calculated the percent of grain yield obtained for
each kilogram of wheat total biomass (Table 2.3). The high

22
coefficient of determination observed between wheat biomass
and grain yield (R2 = 0.92 and 0.98) provided the basis
for using biomass as a means of relating species density
and proportion to individual plant grain yield.
Describing Individual Plant Weight
As each species total density increased in
monoculture and mixture, the mean weight per plant of each
species decreased (Table 2.3). The intraspecific
competition models predicting reciprocal plant weight (1/W)
for each species (based on data from Table 2.3) are
presented in (Table 2.5). Ryegrass intraspecific
competition (b0 /b1) was 3.5 times greater than wheat
intraspecific competition when grown in monoculture.
Ryegrass competed strongly with itself, indicating that
ryegrass plant weight was strongly influenced by its own
species in a mixed population. Wheat did not compete as
strongly with itself as did the ryegrass.
The expanded reciprocal yield models for each species
predicting reciprocal plant weight are presented in Table
2.6. The relative competitive ability (intraspecific
competition coefficient/interspecific competition
coefficient) for ryegrass in mixture indicates that
interspecific competition was only slightly more important
than intra-specific competition. On a relative basis, a
ryegrass plant competed with another ryegrass plant as

23
aggressively as one wheat plant.
The model predicting reciprocal wheat yield indicates
that a single wheat plant was as competitive as 6.7
ryegrass plants. Wheat was the superior competitor against
ryegrass, reducing ryegrass biomass significantly at high
wheat densities and proportions. Predicted wheat biomass
also was reduced by wheat neighbors in mixture (Figure
2.2). The reciprocal weights of the wheat monocultures
differed slightly from those in mixtures until 200 to 400
ryegrass plants/m 2were added to the mixture. The weight
of ryegrass plants, however, was rapidly reduced when wheat
or ryegrass were added to the mixture (Figure 2.3). The
greatest reduction in predicted ryegrass weight was due to
increasing wheat density. The coefficient of determination
(R2
) for the expanded reciprocal model of wheat indicates
that 90 percent of the variation in wheat reciprocal weight
(R2=0.90) was explained by the final densities of both
species (Table 2.6). However, only 43 percent of the
variation in ryegrass reciprocal weight (R 2=0.43) was
explained by the final species densities (Table 2.6),
suggesting that interactions of experimental variables may
have affected ryegrass in the mixtures. Therefore, a more
complete model for ryegrass biomass yield was built, using
the main experimental variables and their interactions.

24
Full Model Describing Ryegrass Reciprocal Plant Weight
Fifty-nine percent of the variation (R 2=0.59) in
reciprocal ryegrass weight was described by the final
densities of both species (p=0.05) and a density
interaction (p < 0.01) (Table 2.7). The significant
interaction of ryegrass and wheat densities indicates that
species proportion was an important variable determining
ryegrass yield (partial R2=0.57). The significant density
interaction suggests that as total density increased, its
influence was multiplicative, rather than additive (Figure
2.4).
Importance of Competition Factors on Both Species
Ninety percent of the reciprocal weight of wheat
(R2=0.90) was described by the final densities of wheat and
ryegrass (Table 2.6). Because wheat was such a formidable
competitor, it appears that wheat plants were influenced
predominantly by other wheat plants in the two-species
mixture. These data suggest that increasing wheat density
to decrease the effect of ryegrass density may not have a
beneficial effect on individual wheat yield. Appleby et
al. (1976) indicated that a reduction in total grain yield
was attributed to ryegrass alone. However, our study
suggests that wheat yield is a function of the ratio of
wheat and ryegrass with a major reduction of "potential"
wheat yield resulting from intraspecific competition as

25
suggested by the grain yields which resulted in
monocultures in the experiment (Figure 2.1). Potential
yield may defined as the greatest yield possible from one
plant given optimum conditions and available resources.
The greatest reduction in total wheat grain yield from
interspecific competition of ryegrass should occur when
wheat densities are low and ryegrass densities are high
(Figure 2.1).
Conclusions
An addition series and the expanded reciprocal
approach was used to determine the relative competitive
abilities of spring wheat and Italian ryegrass. This
procedure was the first step in identifying important
factors affecting spring wheat cropping systems. This
experiment demonstrated that species density and proportion
influenced intra- and interspecific competitive responses
of both species. Therefore, species density and proportion
should be included in more extensive models to predict crop
yield losses resulting from weeds.
Economic risk is an important factor to consider in a
farming program. Thus, being able to reduce risk or
improve yields are objectives for most agricultural
experimentation. The regression models produced in this
experiment, to examine wheat and ryegrass competition can

26
be used to assess the risk associated with direct weed
control and other management tactics in that cropping
system. The competitive ability of both species over a
range of species densities and proportions was confirmed.
Such information is important to manipulate crop and weed
densities in the field. By relating the biological
information obtained from this type of experiment with
other information concerning management and economics, the
economic risk may be lowered.

Table 2.1. An addition series used to generate monocultures and mixture densities ofspring wheat (W) and Lolium multiflorum (R) resulting in systematically hanging speciesrelative and total densities. Total densities range from 0-880 plants/m , and relativedensities from 0-440 plants/m . The density of spring wheat increases along the verticalaxis. Density of Lolium multiflorum increases along the horizontal axis. 1:1 ratios of thetwo species extend along the underlined diagonal.
Addition Series Experiment
W/R W/R W/R W/R W/R W/R
0/0 9/0 36/0 61/0 220/0 441/0
0/9 9/9 36/9 61/9 220/9 441/9
0/36 9/36 36/36 61/36 220/36 441/36
0/61 9/61 36/61 61/61 220/61 441/61
0/220 9/220 36/220 61/220 220/220 441/220
0/441 9/441 36/441 61/441 220/441 441/441

28
Table 2.2. Mean results from 4 replications of 1986spring wheat-Lolium multiflorum addition series exper-ments. Wheat and ryegriss planting density/m (WTR, RTR)and final density/.75 m (FWD, FRD). Biomass (RB, WB),,and wheat reproductive yield (WGY) measured as kg/.75Two varieties of spring wheat were grown with Marshalsryegrass to determine competitive response of both species.
Wheat WTR RTR FWD FRD RB WB WGYVariety
Owens 0 0 0.0 0.00 9 0.0 9.7 0.123 0.0000 36 0.0 27.7 0.238 0.0000 61 3.7 43.2 0.277 0.0000 220 0.0 99.5 0.437 0.0000 440 0.0 189.0 0.490 0.0009 0 0.6 0.0 0.000 0.146 0.04309 9 0.7 5.2 0.153 0.094 0.02709 36 31.2 15.2 0.161 0.058 0.02029 61 1.5 30.2 0.283 0.099 0.01059 220 0.7 84.7 0.446 0.074 0.01379 440 2.0 136.5 0.514 0.074 0.0195
36 0 10.5 0.0 0.000 0.403 0.156536 9 12.7 10.5 0.042 0.521 0.222036 36 14.2 34.2 0.228 0.419 0.170036 61 14.2 28.2 0.215 0.416 0.148636 220 12.0 91.6 0.321 0.285 0.111336 440 14.5 116.0 0.357 0.285 0.104761 0 21.2 0.0 0.000 0.758 0.304661 9 24.0 5.7 0.054 0.647 0.266061 36 25.2 25.0 0.077 0.620 0.253761 61 22.0 40.5 0.164 0.495 0.171061 220 24.2 92.7 0.228 0.450 0.161261 440 88.0 101.7 0.238 0.423 0.1562
220 0 54.7 0.0 0.000 0.819 0.3440220 9 45.2 11.0 0.025 0.791 0.2940220 36 51.5 16.2 0.032 0.809 0.3417220 61 54.0 24.2 0.080 0.760 0.3067220 220 43.7 72.0 0.114 0.700 0.2710220 440 49.6 150.3 0.205 0.634 0.2433440 0 201.2 0.0 0.000 0.923 0.3707440 9 227.0 6.6 0.022 0.944 0.3743440 36 201.7 18.2 0.023 0.832 0.3485440 61 201.0 27.0 0.029 0.857 0.3490440 220 194.5 49.5 0.029 0.826 0.3267440 440 205.2 133.7 0.063 0.782 0.3015

29
Table 2.2. cont.
Wheat WTR RTR FWD FRD RB WB WGYVariety
Waverly 0 0 0.0 0.00 9 0.0 5.0 0.101 0.0000 36 0.0 17.5 0.211 0.0000 61 0.0 36.7 0.277 0.0000 220 0.0 75.7 0.351 0.0000 440 0.0 157.0 0.456 0.0709 0 1.5 0.0 0.000 0.236 0.03139 9 2.0 14.7 0.131 0.083 0.01669 36 3.0 32.2 0.256 0.147 0.05029 61 1.0 36.0 0.295 0.045 0.01509 220 1.6 81.3 0.411 0.052 0.01409 440 2.2 105.5 0.385 0.073 0.017336 0 10.0 0.0 0.000 0.477 0.186036 9 13.2 8.5 0.047 0.501 0.200536 36 10.2 21.2 0.070 0.407 0.167536 61 12.2 15.5 0.109 0.438 0.173036 220 29.2 63.7 0.240 0.242 0.106736 440 12.2 144.2 0.368 0.240 0.092761 0 26.0 0.0 0.000 0.650 0.281361 9 26.0 13.5 0.050 0.640 0.267061 36 28.2 23.0 0.072 0.559 0.208561 61 23.2 30.0 0.100 0.544 0.208761 220 21.2 93.7 0.327 0.460 0.1830
61 440 20.6 148.0 0.344 0.278 0.1176220 0 57.7 0.0 0.000 0.686 0.2843220 9 52.7 10.7 0.017 0.760 0.3215220 36 61.3 24.0 0.060 0.557 0.2700220 61 29.5 31.2 0.102 0.621 0.2220220 220 46.0 93.2 0.124 0.497 0.1817220 440 51.0 158.5 0.251 0.492 0.1860440 0 169.7 0.0 0.000 0.738 0.3020440 9 188.7 3.0 0.010 0.896 0.3375440 36 173.7 9.6 0.007 0.802 0.3032440 61 179.2 22.7 0.023 0.848 0.3355440 220 178.7 51.2 0.047 0.796 0.3030440 440 193.0 84.0 0.074 0.681 0.2512

30
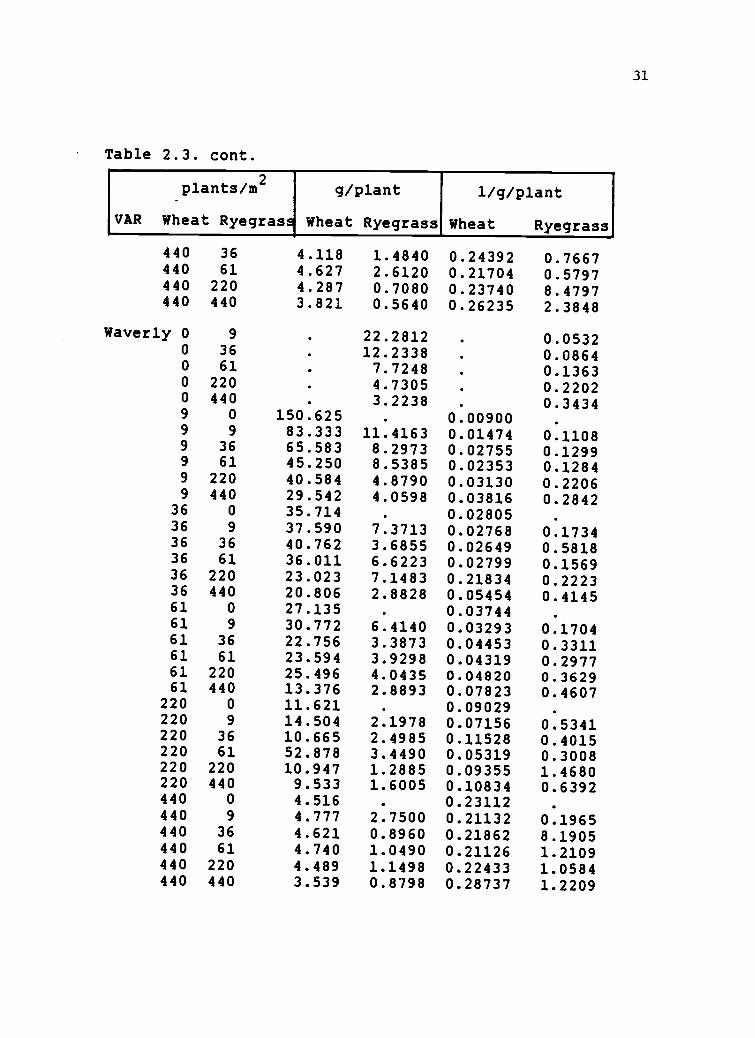
Table 2.3. Mean individual plant responses calculated asmeans from four replications of the 1986 springwheat-Lolium multiflorum addition seriesexperiments. Two wheat varieties (VAR) were grownwith Marshals ryegrass. Mean plant responses (g /plantwere calculated by dividing total species biomass/.75m bythe final species density. Mean reciprocal weight(1/giplant) was used in the multiple linear regressionmodels as a competitive response to species density andproportion. Treatments in the addition series included6 densities of wheat (WTR) combined with correspondingdensities of ryegrass (RTR).
VAR
'
plants/m2
Wheat Ryegrass
g/plant
Wheat Ryegrass
1/g/plant
Wheat Ryegrass
Owens 0 9 . 18.4260 . 0.07030 36 9.2413 . 0.12410 61 0.000 7.3228 . 0.16430 220 4.7157 . 0.23850 440 . 2.6387 . 0.44179 0 164.000 . 0.006249 9 89.667 36.5937 0.01156 0.04479 36 50.227 13.4847 0.49977 0.07769 61 88.333 10.2000 0.01519 0.10969 220 66.667 5.6902 0.01652 0.19209 440 33.000 3.8100 0.03304 0.2633
36 0 38.786 0.0262936 9 40.590 6.8570 0.02503 0.274736 36 32.520 7.5522 0.03380 0.214536 61 30.168 5.9287 0.03571 0.178936 220 23.795 4.6730 0.04227 0.292436 440 20.486 3.2210 0.05076 0.324361 0 28.563 0.03566 .
61 9 27.120 9.2160 0.03745 0.170461 36 24.719 3.3603 0.04066 0.324161 61 22.561 4.1857 0.04562 0.253561 220 18.939 2.7815 0.05402 0.432161 440 12.406 2.4213 0.21839 0.4352
220 0 15.230 0.06664 .
220 9 17.494 3.6813 0.05753 0.4058220 36 15.825 2.5328 0.06413 0.4728220 61 14.693 2.3163 0.07255 0.4636220 220 20.158 1.8175 0.06524 0.5935220 440 13.041 1.5773 0.07978 0.7785440 0 4.588 . 0.21822 .
440 9 4.301 4.2853 0.23430 0.6247
31
Table 2.3. cont.
plants/m2
VAR Wheat Ryegrass
g/plant
Wheat Ryegrass
lig/plant
Wheat Ryegrass
440 36 4.118 1.4840 0.24392 0.7667440 61 4.627 2.6120 0.21704 0.5797440 220 4.287 0.7080 0.23740 8.4797440 440 3.821 0.5640 0.26235 2.3848
Waverly 0 9 22.2812 0.05320 36 . 12.2338 0.08640 61 . 7.7248 0.13630 220 . 4.7305 0.22020 440 . 3.2238 . 0.34349 0 150.625 . 0.00900 .
9 9 83.333 11.4163 0.01474 0.11089 36 65.583 8.2973 0.02755 0.12999 61 45.250 8.5385 0.02353 0.12849 220 40.584 4.8790 0.03130 0.22069 440 29.542 4.0598 0.03816 0.2842
36 0 35.714 . 0.0280536 9 37.590 7.3713 0.02768 0.173436 36 40.762 3.6855 0.02649 0.581836 61 36.011 6.6223 0.02799 0.156936 220 23.023 7.1483 0.21834 0.222336 440 20.806 2.8828 0.05454 0.414561 0 27.135 . 0.0374461 9 30.772 6.4140 0.03293 0.170461 36 22.756 3.3873 0.04453 0.331161 61 23.594 3.9298 0.04319 0.297761 220 25.496 4.0435 0.04820 0.362961 440 13.376 2.8893 0.07823 0.4607
220 0 11.621 . 0.09029 .
220 9 14.504 2.1978 0.07156 0.5341220 36 10.665 2.4985 0.11528 0.4015220 61 52.878 3.4490 0.05319 0.3008220 220 10.947 1.2885 0.09355 1.4680220 440 9.533 1.6005 0.10834 0.6392440 0 4.516 . 0.23112 .
440 9 4.777 2.7500 0.21132 0.1965440 36 4.621 0.8960 0.21862 8.1905440 61 4.740 1.0490 0.21126 1.2109440 220 4.489 1.1498 0.22433 1.0584440 440 3.539 0.8798 0.28737 1.2209

32
Table 2.4. Wheat harvest indices for Owens and Waverlywheat varieties were calculated. Total wheat weights/0.75 m
2
were regressed against total grain weights (kg grain) (Table2.2). The indices were calculated to develop a relationshipbetween total wheat yield and reproductive yield. The coeffi-cients of determination indicate that 92 to 98 percent of thevariation in grain weight was described by wheat biomass.Biomass was therefore used as a competitive response in the1986 spring wheat-Lolium multiflorum competition experiment.
Wheat Variety b0
bl
Owens Kg grain = -0.1458 + 0.4183 (Kg biomass) 0.98
b0
b1
R2
Waverly Kg grain = 0.0005 + 0.3933 (Kg biomass) 0.92
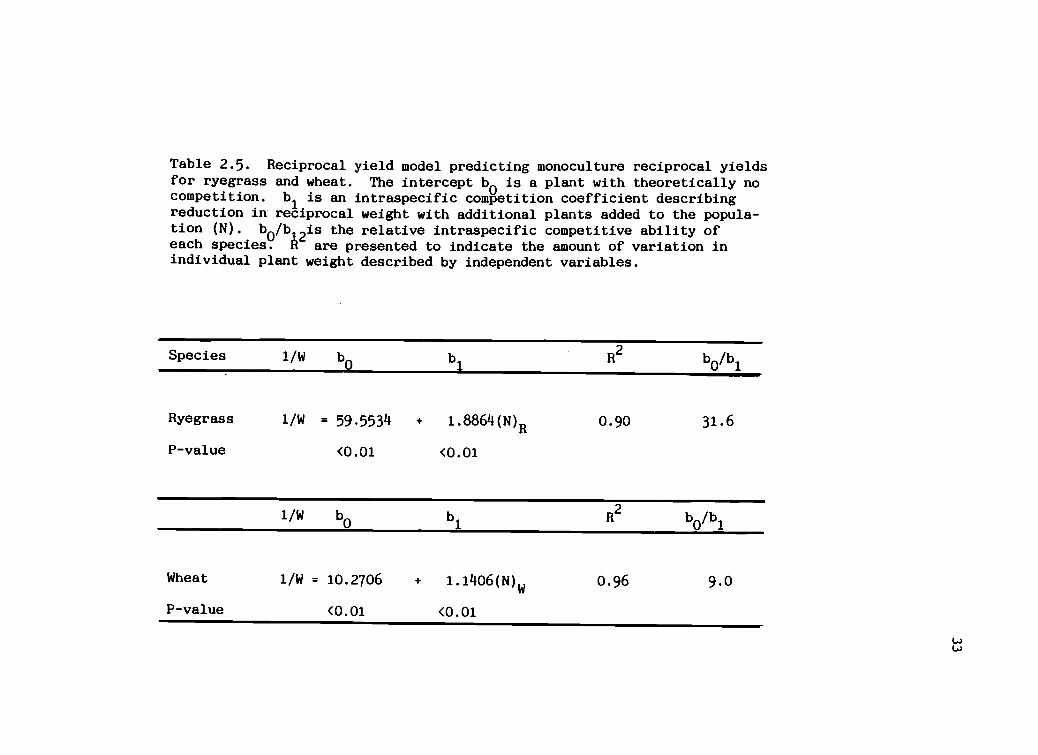
Table 2.5. Reciprocal yield model predicting monoculture reciprocal yieldsfor ryegrass and wheat. The intercept b0 is a plant with theoretically nocompetition. 1)1 is an intraspecific competition coefficient describingreduction in reciprocal weight with additional plants added to the popula-tion (N). b
0/b
12is the relative intraspecific competitive ability ofeach species. A are presented to indicate the amount of variation inindividual plant weight described by independent variables.
Species 1/W b0
b1
R2
b0/b
1
Ryegrass 1/W = 59.5534 + 1.8864(N)R 0.90 31.6
P -value <0.01 <0.01
1/W b0
b1
R2
b0/b
1
Wheat 1/W = 10.2706 1.1406(N)w 0.96 9.0
P-value <0.01 <0.01
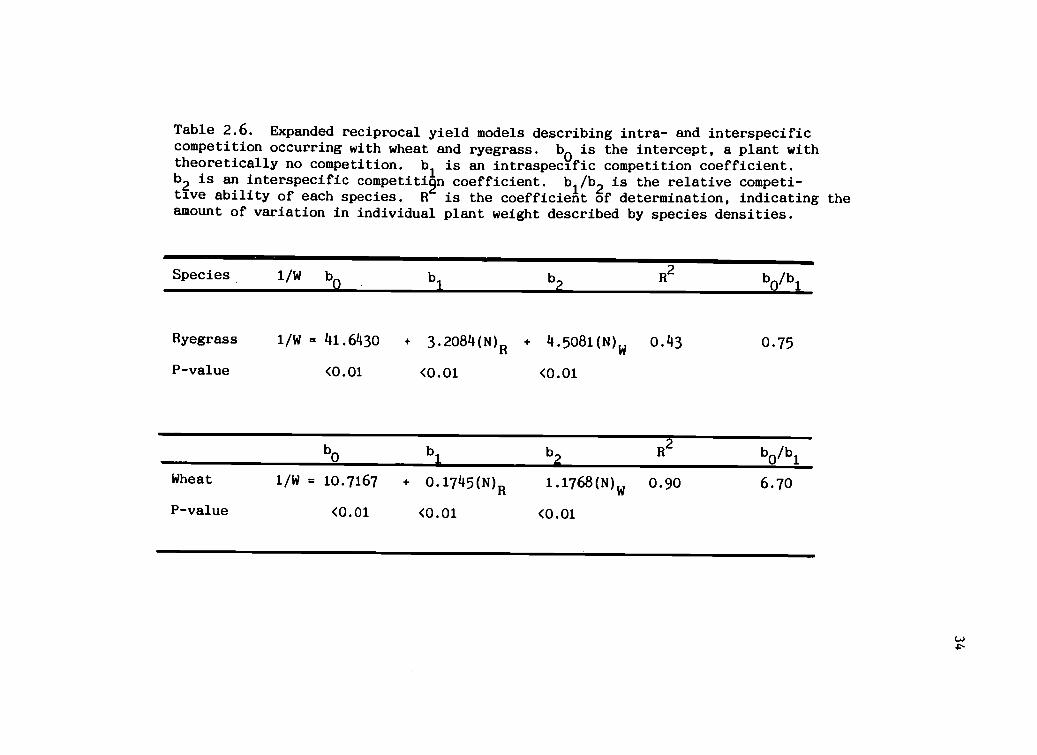
Table 2.6. Expanded reciprocal yield models describing intra- and interspecificcompetition occurring with wheat and ryegrass. bo is the intercept, a plant withtheoretically no competition. bl is an intraspecific competition coefficient.b2
is an interspecific competiti9n coefficient. b1/b
2is the relative competi-
tive ability of each species. R is the coefficient of determination, indicating theamount of variation in individual plant weight described by species densities.
Species. 1/W bo b1
b2
R2
Ryegrass 1/W = 41.6430 + 3.2084(N)R + 4.5081(N) 0.43 0.75
P-value <0.01 <0.01 <0.01
b0
b1
b2
R2
b0/b
1
Wheat 1/W = 10.7167 + 0.1745(N)R 1.1768(N)w 0.90 6.70
P-value <0.01 <0.01 <0.01

Table 2.7. Ryegrass reciprocal yield model with independent variables ofspecies densities and a density interaction (wheat density*ryegrass density).b0 is the intercept, a plant with theoretically no competition. b
1is an
intraspecific competition coefficient. 132 is an interspecific competi-tion coefficient. b is a coefficient degcribing the multiplicative effectof species prportiod (interaction). A coefficient of determination (R )
and partial R for each independent variable are presented to indicate theamount of variation in individual plant weight described by independent vari-ables.
Species b0
b1
b2
b
1/W(Ryegrass)
P-value
Partial R2
= 173.901
<0.01
+ 0.939(N)R
0.05
0.01
+ 1.360(N)w + 0.060(NR*Nw)
<0.01 <0.01
0.01 0.57
R2
= 0.59

36
Figure 2.1. Total grain yields/ha resulting from each experimentof Owens and Waverly wheat varieties planted2with Lolium multiflorumin addition series. The planted densities /m' of wheat and ryegrassare plotted along the x-axis. The resulting grain yields are plottedalong the y-axis. As wheat density increased, grain yield increased.However, the grain increase in monocultures was not linear, suggestingconstant final yield occurred at higher densities. Adding ryegrass tothe mixture slightly decreased yield. Intra- and interspecific competi-tion were responsible for grain decreases.

OWENS WHEAT GRAIN YIELDkg/hec
WAVERLY WHEAT GRAIN YIELD kg/hec

38
Figure 2.2. Individual plant biomass predicted for wheat by the expandedreciprocal yield regression equation (Table 2.6). Individual wheat biomasswas more significantly reduced by increasing wheat density than by ryegrassdensity.

PREDICTED WALLY -WHEAT G/FLANT
b1 /b2ratb2Species 1/W b0 b1
Wheat 1/W = 10.7167 + 1.1768(N)w + 0.1745(N)R .90 6.70p-value <.01 <.01 <.01

40
Figure 2.3. Individual plant biomass predicted for ryegrass by the expandedreciprocal yield regression equation. Independent variables used were speciesdensities. As total density and proportion of wheat increased, ryegrass biomasswas reduced (Table 2.6).

RYEGRASS RECITED BIOMASS YODEL
sI4-
os
G/PLANT
0411111101"
40"-011m's
Species 1/W b0 b2 ra 2
Ryegrassp-value
1/W = 41.6430 + 3.2084(N)R + 4.5081(N)w .43 0.75<.01 <.01 <.01

42
Figure 2.4. Individual plant biomass predicted for ryegrass by the expandedreciprocal yield equation, using independent variables of species density andspecies density interaction. The significant density interaction suggests thatadding plants to the mixtures has a multiplicative effect. As total density in-creased, the weight of ryegrass was reduced (Table 2.7).

PREDICTED RYEGRO BIOMNTERKIION
6.0G/PLANT
llllll
Species b0 b1 b2 b3
1/W (ryegrass)
p-value
partial r2
ra2=.59
= 173.901
<.01
+ 0.939(N)R
=.05
.01
+ 1.360(N)w
<.01
.01
+ 0.060(NR*Nw)
<.01
.57

44
Chapter 2 References
Appleby, A.P., P.D. Olson, and D.R. Colbert. 1976.Winter wheat reduction from interference by Italianryegrass. Agron. J. Vol. 68:463-466.
Bleasdale, J.K.A. 1967. Systematic designs for spacingexperiments. Exper. Agric. 3:73-85.
Carlson, H.L. and J.E. Hill. 1986. Wild oat (Avena fatua)competition with spring wheat:effects of nitrogenfertilization. Weed Sci 34:29-33.
Cousens, R.D. 1986. The use of population models in thestudy of the economics of weed control. Proc. EWRSSymposium, 1986. Economic Weed Control, 269-276.
de Wit, C.T. 1960. On Competition. No. 66.8 WageningenThe Netherlands: Center for Agricultural Pulications andDocumentation.
Donald, C.M. 1963. Competition for light in crops andpasture. La: Mechanisms in Biological Competition. Soc.Expmt. Biol., Symposium XV. Combridge Press. London. pp.283-313.
Firbank, L.G. and A.R. Watkinson. 1987. On the analysisof competition within two-species mixtures of plants. J.Appl. Ecol. 22:503-517.
Grime, J.P. 1973. Plant Strategies am Vegetationprocesses. John Wiley and Sons, N.Y. 222 p.
Harper, J.L. 1977. Population Biology fa Plants.Academic Press, New York. 892 pp.
Harper, J.L., J.T. Williams, and G.R. Sagar. 1965. Thebehavior of seeds in soil. I. The heterogeneity of soilsurfaces and its role in determining the establishment ofplants from seed. J. Ecol. 53:273-286.
Hunt, R. 1982. Plant Growth Curves. The, functionalapproach g. plant Growth analysis. Baltimore. UniversityPark Press. 248 p.
Jolliffe, P.A., A.N. MinJas, and V.C. Runeckles. 1984. Areinterpetation of yield relationships in replacementseries experiments. J. of Appl. Ecol., 21:227-243.
Jewiss, O.R. and Woledge, J. 1967. The effect of age on

45
the rate of apparent photosynthesis in leaves of tallfescue (Festyca arundinacea Schreb.). Ann. Bot. 31:661-671.
Little, T.M. and F.J. Hills. 1978. AgriculturalExperimentation:Design and Analysis. John Willey and SonsInc. Canada.
Miller, T.E. and P.A. Werner. 1987. Competitive effectsand responses in plants. Ecology (in press).
Neter, J., W. Wasserman, and M.H. Kutner, 1983.AgpliedLinear Regression Models Richard D. Irwin Inc. Illinois.
Newman, E.J., 1983. Interactions between plants. In:Encyclopedia of Plant Physiology New Series Volume 12 C.O.L. Lange, P.S. Nobel, C.B. Osmond, and H. Ziegler (Eds.).Springer-Verlag Berlin/New York. pp. 679-771.
Noguchi, K. and K. Nakayama. 1978a. Jap. J. Crop sci.47:48-55.
Patterson, D.T. 1982. Effects of light and temperature onweed/crop growth and competition. ja:Biometerology inI.P.M. Academic Press.
Peterson, C.M., B. Klepper, and R.W. Rickman, 1982.Tiller development at the coleoptilar node in winter wheat.Agronomy J.74:781-784.
Radosevich, S.R. 1987 (in press). Experimental methodsto study crop and weed interactions. In:Altieri, M.A., M.Z.Liebman (Eds). Plant Competition and Other EcologicalApproaches to Weed Control in Agriculture. CRC Press.
Radosevich, S.R. and J.S. Holt. 1984. WeedEcology:;mplications for, Management. John Wiley and SonsInc. New York. 300 pp.
Roush, M.L. and S.R. Radosevich. 1985. Relationshipsbetween growth and competitiveness of four annual weeds.J. of Appl. Ecol. 22:895-905.
Roush, M.L. and S.R. Radosevich. 1987. A weed communitymodel of germination, growth, and competition of annualweed species. WSSA abstracts. St. Louis.
SAS Institute. SAS/STAT Guide for Personal computers,Version 6 Edition, 1985. Cary, N.C.:SAS Institute Inc.

46
SAS Institute. SAS/System for Linear Models. 1986, Cary,N.C.:SAS Institute Inc.
Sestak, Z. 1983. Photosynthesis During Leaf development.Dr. W. Junk Publ., Dordrecht/Boston. 396 p.
Shinozaki,K. and T. Kira. 1956. Intraspecificcompetition among higher plants. V11. Logistic theory ofthe C-D effect. J. of the Inst. of Polytechnics, Osaka CityUniversity, 7:35-72.
Spitters, C.J.T. 1983 a. An alternative approach to theanalysis of mixed cropping experiments. I. Estimation ofcompetition effects. Netherlands J. of Agr. Sci. 31:1-11.
Spitters, C.J.T. 1983 b. An alternative approach to theanalysis of mixed cropping experiments. II. Marketableyield. Netherlands J. of Agr. Sci. 31:143-155.
Tilman, D. 1982. Re*ource competition and, communityStructure.Princeton University Press. Princeton, NewJersey.
Watkinson, A.R. 1980. Density-dependence in single-species populations of plants. J. Teor. Biol. 83:345-357.
Westoby, M. 1981. The place of the self thinning rule inpopulation dynamics. Am. Nat. 118, pp. 581-587.
Willey. R.W. 1985. Evaluation and presentation ofintercropping advantages. Expl. Agric. 21:119-133.
Yoda, K., T. Kira, H. Ogawa, and H. Hozumi. 1963. Self-thinning in overcrowded pure stands under cultivated andnatural conditions. J. Biol. Osaka City University. 14:107-129.
Zimdahl, R.H. 1980. Weed-crop Competition:A Review.International Plant Protection Center, Corvallis, OR. 196pp.

47
CHAPTER 3: QUANTIFYING GROWTH RESPONSES OF SPRING WHEATTRITICUM AESTIVUM (L.) VAR. OWENS (SUNDERMAN), VAR.
WAVERLY (KONZAK), AND LOLIUM MULTIFLORUM (LAM.)
Introduction
Relative growth rate is a measure of how successfully
a plant captures resources relative to its original size
(Roush and Radosevich 1985). Numerous studies have
demonstrated that a rapid growth rate can be advantageous
in resource capture for both weeds and crops (Patterson
1982). Relative rate of growth is dependent on
intraspecific as well as interspecific competition when
crops and weeds occur together in mixture. In experiments
containing two or more species, separating these two types
of competition has been difficult (Spitters 1983a) when
variables of species density and proportion were not
systematically varied (Radosevich 1987). Species density
and proportion along with spacing, are proximity factors
determining the nearness and identity of neighboring
plants.
Individual plant growth also is influenced by
biological traits such as genetic capability (e.g. seed
size) seed bank dynamics (e.g. seed survival) and
phenotypic factors (e.g. emergent size). Furthermore, life
history traits of the crop and weed, such as annuality
versus perenniality also may contribute to the ability of a
plant to usurp resources (Roush and Radosevich, in

48
progress). However, in annual cropping systems, the life
history traits of a weed and crop usually parallel one
another. Agricultural systems also are influenced strongly
by disturbance (Grime 1979) and resource availability. Any
or all of these factors interact with one other to
determine plant emergence and growth in mixtures.
A plant that emerges first in a mixture is likely to
usurp a disproportionate amount of the available resources
throughout the season (Radosevich and Holt 1984), growing
larger than surrounding neighbors and retaining a dominant
role in the mixture. If emergence of both species is
simultaneous (or controlled) in an experiment, then seed,
and consequently germinant size, may play an important role
in determining the relative growth rate of a plant in
mixture.
Ultimate plant size is determined by the amount of
resources which a plant obtains and uses during the growing
season. "Space" is considered to be a unit of resource
uptake, as influenced by environmental conditions, such as
temperature (Radosevich and Holt 1984). Varying the
proximity factors of species density and proportion
systematically in an addition series (Miller and Werner
1987, Roush and Radosevich 1987, Radosevich 1987), while
maintaining equidistant spacing, initially allocates
resources equally to each neighboring plant.

49
Plant size also is dependent on plant structure. Plant
structure (especially flowers and leaves) depends on a set
of morphological repeating units. The size and form of
these units is tightly controlled by genetically inherent
properties. The relative initiation rate of the
morphological units varies with different levels of
competition (Harper 1977). Harper (1977) also suggests
that the number of units and consequently, the size of the
whole plant may vary with age and environmental conditions
throughout the growing season. For instance, the relative
rate of initiation of leaves, tillers, and flowers may be
cued by temperature and light. Alternatively, these
initiation rates may vary with different levels of
competition for resources.
Plant competition occurs when resources become
limiting. Harper (1977) suggests that plants in mixtures
begin to compete early for available resources. When
stores of soil-associated resources (e.g. nutrients and
water) fall short of plant population needs, competition
for these resources begins.
Above-ground competition for light resources differs
from below-ground resource competition. Light is available
briefly and cannot be stored (Radosevich and Holt 1984).
In below-ground resource competition space capture occurs
by root expansion. In above-ground resource competition

50
the position of a plant becomes an important factor for
light interception (Donald 1963, Radosevich and Holt 1984).
Thus, the rate at which plants integrate resources may
elucidate strategies by which antra- and interspecific
plant neighbors pre-empt available resources.
Firbank and Watkinson (1987) categorize the pre-
emption of resources as either "one-sided" or "two-sided"
competition. One-sided competition occurs when plants grow
larger and faster in the population, over-topping smaller
and slower growing plants. Thus, one competitor (overstory
plants) influences its neighbor (understory plants) but is
not in turn influenced by that neighbor. This phenomenon
can occur when seed sizes, times of germination, or growth
rates differ among the individuals in a mixture. When
seeds are allowed to emerge simultaneously, large seed size
and phenotypically larger plant emergents are considered
advantageous when competing for available resources
(Radosevich and Holt 1984).
Two-sided competition occurs when plants emerge
simultaneously, pre-empting space proportional to their
size, until all space around each plant is occupied by
other plants. Further development is dependent on vertical
growth (Ross and Harper 1972, Firbank and Watkinson 1987).
In a plant mixture, two-sided competition usually occurs
between plants in the upper canopy which have emerged and

51
grown faster than the lower canopy plants. These upper
canopy plants continue to capture space in a uniform
fashion until all available upper canopy space is occupied.
The relative size of a plant at emergence may
determine the type of competition it experiences from
neighboring plants. A larger seed containing a large
endosperm supply usually grows faster and larger in the
first few days than a smaller seed requiring photosynthetic
processes (Esau 1978). Seed or emergent size is accounted
for when relative growth rate measurements begin.
Monitoring changes in plant relative rates of growth
may better define the period when competition between crops
and weeds begins. Furthermore, monitoring growth rate
changes in systematically varying levels of competition
produces a predictive method of assessing competition.
Plant allometry, the study of change in growth of various
plant parts from those more easily measured, utilizes this
method of monitoring plant growth. Plant allometry is
already widely used in forestry (Grier et al. 1984, Bold
and Radosevich 1987). For example, timber stand volume can
be predicted from basal area and height of individual
trees. Simple agronomic plant 'mAasurements such as plant
height and tiller number, measured over time can be used to
determine relative growth rates and predict desired
parameters such as grain yield. Furthermore, data taken

52
Methods
Plant material, planting methods, and cultural
practices were those described in Chapter 2. One
individual of each species was tagged in each plot. Plant
height, leaf number, tiller number, and leaf area index
(LAI) were measured.
The 100-seed weight of each wheat variety and Italian
ryegrass was weighed and recorded. Plant height was
measured from the soil line to the tip of the longest leaf.
The number of leaves was recorded, with the coleoptile
designated as the zero leaf. Subsequent leaves were
numbered 1 to 9. Numbers of tillers and subtillers were
considered equally when counted. Measurements were taken
weekly.
A point frame was constructed with eight aluminum
sliding points spaced 6.5 cm. apart for measuring LAI. LAI
was measured 5 times during the season, beginning in the
middle of the season when average LAI values for both
species exceeded 2. Species LAI was determined by
calculating the average percentage of species hits for the
eight sliding points. Individual plants measured for
height, leaf number, and tiller number were harvested
separately from the total plot harvest described in Chapter
2. Plants were dried to constant weight (15.6 C, 48 h),
and weights were included in biomass determination.

53
Growing Degree Day Units. Planting of the addition series
experiment required ten days. Plants in the first
replication were, therefore, planted ten days before those
in the fourth replication. Plants of equal GDD accumulate
similar developmental time (integrated temperature and
time) despite variation in chronological time (Peterson
1982). Therefore, GDD was used as the time variable in all
analyses of growth.
Ambient air temperatures were collected by the Oregon
State University Climatic Research Institute at Hyslop Crop
Science Field Laboratory, near Corvallis. GDD were
calculated from daily maximum and minimum temperatures
using a baseline growth temperature of 3 C (Klepper
personnel communication).
Calculations. Instantaneous relative growth rates were
calculated for plant height, leaf number and tiller number
(Hunt 1982). The log transformation of primary growth data
was regressed in a stepwise procedure (SAS 1985) against
the variables of GDD, polynomial transformations of GDD
(GDD2, GDD
3), and the log of GDD. These transformations
were used to fit the functional curve of growth data that
may have changed with each relative and total density in
the addition series. The lowest mean square error (MSE) (P
< 0.15), and appropriate residual diagrams were used for

54
determining the best independent variables to describecurves of the log of plant growth. The relative growthrates-are the slopes of the log (growth) versus the GDDcurves. Regression models describing relative rates ofgrowth were constructed for each total density treatment inthe addition series. The appropriate regression equationfor each treatment were then
differentiated, creatinginstantaneous relative growth rate equations for individualplants in each addition series treatment (Hunt 1978).Finally, the acquired GDD for each measurement time wasinput into these equations for
instantaneous plant relativegrowth rates.
Statistical Analysis. Analysis of variance was performedon the final set of plant growthmeasurements to partitionthe seasonal variation among observations into portions
associated with treatmentvariables of replication, wheatvariety, wheat density, ryegrass density, and variable
interactions.
Analyses of variance and covariance to test thehomogeneity of slopes were performed on instantaneousrelative growth rates of plant height, leaf number, andtiller number. Wheat varieties were used as covariants.Independent variables used to describe instantaneousrelative rates of growth were replication, wheat variety,

55
wheat density, ryegrass density, GDD, and interactions of
these variables. Regression variables were chosen based on
significance of independent variables (P < 0.07), largest
coefficient of determination (R 2), lowest mean square error
(MSE), appropriate residual diagrams, and lowest possible
multicolinearity. The maximum variance inflation factor
(VIF) was 6. This value occurred when GDD was selected by
the stepwise regression procedure as the most significant
variable as determined by the partial R2. Relative growth
rates were calculated from descriptive variables of GDD and
transformations of GDD. Transformations of GDD such as log
and cubic, describe accelerations and decelerations of
growth rate. The differences in changes of growth rate
were attributed to proximity factors of species density and
proportion when analyzed with analysis of variance and
covariance. Growth rates which were not affected by
proximity factors followed a linear pattern during the
season. Therefore, these growth rates were highly
correlated with GDD, the variable which was used to
calculate the growth rates and also describe them in the
analysis of variance and covariance.
Analyses of variance and covariance were performed on
species LAI. The continuous measurement of LAI was
described by independent variables of replication, wheat
variety, wheat density, ryegrass density, ODD, and all

56
interactions of these variables. Again, wheat varieties
were chosen as covariants. Criteria used to select
independent variables to explain instantaneous relative
growth rates of plant height, leaf number, and tiller
number also were used to select variables to describe
species LAI.
Results and Discussion
Wheat seed was 15-18 times larger than that of
ryegrass. Furthermore, Owens wheat seed was 16 percent
larger than that of Waverly. The large seed size of wheat
could impart an advantage over ryegrass at the beginning of
the season since both species emerged simultaneously (Esau
1978, Harper 1977). Wheat plants emerged 1-2 days earlier
than the ryegrass over all densities and were
phenotypically larger plants. Thus, wheat plants began as
larger emergents than ryegrass and had an advantage in
usurping resources early in the season.
Height. Plant height is a genetic attribute bred into
grain crops. In this experiment, both wheat varieties were
semi-dwarfs with a 10 cm potential difference. Mean
heights calculated for both varieties in each addition
series indicated that a 10 cm difference existed between
the varieties in the same treatment plots. Analysis of
variance performed on final wheat heights for both

57
varieties indicated that identical variables affected both
varieties. A significant amount of the variation in wheat
height was attributed to wheat density and replication (P <
0.01). However, analysis of variance performed on final
ryegrass heights suggested that both proximity variables of
species density and proportion influenced ryegrass height.
Variation in final ryegrass heights was attributed to wheat
density, ryegrass density, replication, and wheat variety
(P < 0.01).
To describe when and how proximity variables
influenced plant height rates of each species, analyses of
variance and covariance were performed on the instantaneous
height growth rates. Fifty-two percent of the variation in
ryegrass height growth rates (R2 = 0.52) was explained by
independent variables of wheat density (P = 0.02), GDD (P <
0.01), and the interactions of GDD*wheat density (P <
0.01), and wheat density*wheat variety (P = 0.03) (Table
3.1).
The rate of ryegrass height growth does not appear to
differ from either wheat variety. Both species grew at a
constant rate until the .boot stage of wheat when 850 GDD
were accumulated. At this point, both species growth rates
slowed but ryegrass maintained a higher rate of growth than
wheat. This general trend occurred regardless of density
or mixture (Figure 3.1). This suggests that ryegrass

58
plants, grown in mixture with tall wheat plants (10 cm
taller), are experiencing one-sided competition from taller
and phenotypically larger wheat which formed an overstory
(Firbank and Watkinson 1987) for the first half of the
growing season. However, the height growth rates of
ryegrass which increased over the wheat growth rates for
the latter half of the growing season may have compensated
for
the
the overstory competition
beginning of the season.
Analyses of variance and
from wheat for resources at
covariance on wheat height
growth rates indicated that 77 percent of the variation of
height growth rates (R2 = 0.77) was explained by wheat
variety, GOD, and the interactions of GDD*wheat density and
wheat density*ryegrass density (P < 0.01) (Table 3.1).
These interactions suggest that after emergence, the wheat
varieties achieved their overall heights through
differences in growth rates created by variations in growth
response to plant density.
The large intercept, which was significant (P < 0.01)
in both species regression equations, suggests that height
growth rate at emergence also was important in describing
how the species achieved their heights. Though wheat was a
formed an overstory above ryegrass from emergence, ryegrass
maintained a constant rate of height growth. Evidence from
Chapter 2 suggests that ryegrass plant weight was more

59
adversely affected by the overstory competition from wheat
than ryegrass height growth rates. The tall variety of
wheat (Owens) produced higher grain yields than the short
variety of wheat (Waverly) (Figure 2.1).
Leaf Number. Although height is a vertical measure of
space capture, the horizontal level is occupied by layers
of leaves (LAI). Therefore, leaf number initiation rates
and LAI were monitored for each species to elucidate
mechanisms of above-ground resource capture, with respect
to light.
Both wheat and ryegrass normally initiate 9 major
leaves, but stress may deter leaf initiation (Peterson et
al. 1982). The number of leaves initiated often indicates
the environment to which a plant has been subjected (Newman
1986). The greater the number of leaves initiated by the
end of the season, allows a larger source of carbohydrate
for translocation into grain.
Wheat plants initiated all 9 leaves regardless of
density treatment, as indicated by an analysis of variance
performed on the final number of leaves per wheat plant.
However, analysis of variance performed on the final number
of ryegrass leaves indicated that variation in ryegrass
leaf number was attributed to interactions of ryegrass
density*replication (P = 0.03) and replication*wheat
variety (P < 0.01). These interactions suggest that the

60
number of ryegrass leaves per plant resulted from different
levels of ryegrass competition that changed over
replication. Furthermore, ryegrass leaf initiation
differed depending on which variety the ryegrass was grown
with.
The rate at which leaves of each species were
initiated also was an important factor to consider in
assessing above-ground space capture during the season.
When analyses of variance and covariance were performed on
each of the species leaf initiation rates (Table 3.2), the
same independent variables were selected to describe leaf
initiation rates. Wheat density, GDD, and the interaction
of wheat density*CDD (P < 0.01) described 78 percent of the
variation (R2 = 0.78) in wheat leaf initiation rates and 72
percent of the variation (R2 = 0.72) in ryegrass leaf
initiation rates. Furthermore, a stepwise regression
performed on each species using the independent variables
described above, indicated that almost 100 percent of the
coefficient of determination (R 2) was taken up by GDD
(Table 3.2). This indicates that the proximity factors did
not strongly influence leaf initiation rates of either
species and that these rates are probably internally
controlled and cued by temperature over time (GDD).

61
Leaf Area Index. Klepper (personal communication)
hypothesized that size of a plant part (e.g. leaves or
tillers), as opposed to the number of plant parts, may be a
conclusive measure of stress. The size of a leaf is
measured by its area. Leaf area usually is a destructive
measurement; LAI, an indirect measurement of leaf area, was
therefore determined for each species.
LAI is a three-dimensional measurement of the space
captured in each plot as opposed to the actual number of
leaves initiated. LAI was most descriptive of how the
vertical layers of the plant canopy within the plot were
occupied by each species throughout the season.
At emergence, wheat was taller than ryegrass. Wheat
leaves also were wider and spanned a greater distance than
those of ryegrass, but wheat leaves also began to overlap
each other earlier at the highest wheat densities.
Analyses of variance and covariance performed on
measurements of wheat LAI indicate that 49 percent of
variation (R2 = 0.49) was explained by independent
variables of wheat density, ryegrass density, GDD, and
interactions of GDD*wheat density, GDD*ryegrass density,
and wheat density*wheat variety (P < 0.01) (Table 3.3).
LAI of each species was influenced by proximity
factors throughout the season. Wheat LAI increased with
wheat density. However, the increase in wheat LAI differed

62
between wheat varieties. Gradual seasonal decreases in
wheat LAI also varied among the wheat densities. For
instance, wheat growing in high total densities increased
LAI at a greater rate than in low densities at the
beginning of the season. However, wheat in high total
densities decreased LAI faster at the end of the season
than low total densities.
Analyses of variance and covariance performed on
measurements of ryegrass LAI indicate that 48 percent of
variation in LAI was explained by independent variables of
ryegrass density, wheat density, and interactions of
GDD*ryegrass density and wheat density*ryegrass density (P
< 0.01) (Table 3.3). Ryegrass LAI increased with
increasing ryegrass density. However, ryegrass LAI was
adversely affected by increasing density and proportion of
wheat. Also, as the season progressed, LAI of high
ryegrass densities decreased faster than low ryegrass
densities.
The most informative independent variable in both
species' predictive LAI equations was wheat density.
Ryegrass LAI decreased with increasing wheat density in the
mixtures, but the same variable in the wheat LAI equation
increased wheat LAI. This information suggests that
ryegrass was dominated by wheat for the first half of the
season. Furthermore, ryegrass experienced one-sided

63
competition from wheat in mixture, with respect to light
resources (Firbank and Watkinson 1987). Thus, competition
resulted in reduced ryegrass competitive ability (Chapter
2). However, as the season progressed and wheat plants
matured and senesced (fastest in the high total densities),
light became available to suppressed ryegrass plants. LAI
and height of ryegrass plants then increased as these
plants completed their life-cycle, and dispersed seed.
This growth pattern may be one of the strategies that
contributes to the persistence of Italian ryegrass in
spring wheat cropping systems.
Tiller Number. A small amount of variation in tiller
initiation rates was explained by proximity variables or
GDD, and this information was therefore included in the
appendices only.
Summary
Monitoring relative growth rates of plant parameters
has elucidated possible mechanisms that both species
utilize in capturing available resources. Wheat emerged
first in the mixture, growing taller and larger than
ryegrass and capturing vertical space. Wheat plants
continued to expand in LAI, also capturing the horizontal
layers of above-ground space with respect to light
resources. Wheat remained the dominant canopy plant until

64
after "boot stage" (GDD = 850-1200). Wheat then began to
mature and its LAI declined. Presumably, greater light
resources then were made available to the ryegrass.
Before the boot stage of wheat (GDD 850-1200),
ryegrass was suppressed in the mixture. Analyses indicated
that wheat height, leaf initiation, and LAI were not
adversely affected by ryegrass plants in the mixture until
the latter part of the growing season when wheat LAI
declined. However, as density and proportion of wheat
increased, all measured growth rates of ryegrass decreased.
Although rates of height, leaf number, and tiller
growth may be influenced by proximity factors of species
density and proportion, other plant traits such as biomass
and yield appear to be better indicators of competitive
ability (Chapter 2).

Table 3.1. Height growth rates (GR) described by several proximity variables of wheatdensity (N)W, growing degree days (GDD), and the interactions of wheat*density*GDD(GDW), wheat variety*wheat density (VarW), and wheat variety*GDD (VGDD). Wheatvariety did not vary significantly among the ryegrass height growth rates but wassignificantly different among wheat height growth rates. Analysis of variance andcovariance indicated that intercept values for each wheat ariety height (GR) weresignificantly different. Coefficients of determination (R ) are presented to indi-cate the amount of variation described by independent variables.
Species b0
b1
b2
b3
b4
Ryegrass Height (GR) = 0.0059 + 3.3-7 (N)w
- 2.8-6
(GDD) - 1.0-7 (GDW) + 3.3-6
(VarW)
P-value <0.01 <0.01 <0.01 <0.01 <0.01
R2
= 0.52
b0b1
b2
b3
b4
Wheat Height (GR) =
Variety 1 = 0.0050 + 3.9-6 (GDD) + 6.0-6
(VGDD) + 1.9-6
(VarW) + 2.4-8
(GDW)Variety 2 = 0.0048
P-value
R2
= 0.76
<0.01 <0.01 <0.01rn
<0.01 <0.01 Ui

Table 3.2. Leaf number initiation rates (IR)factors of wheat density (N)w, growing degreethese two variables (wheat density*GDD (GDW))strate the large amount of variation attachedtemperature on leaf initiation rate.
described by several proximitydays (GDD) 2and the interaction of
. Partial .R s are included to demon-to GDD, indicating the importance of
Species b0 b1
b2
b3
Ryegrass Leaf IR = 0.0209 - 1.5-5(N)w - 1.4-5(GDD) + 1.0-7(GDW)
P-value <0.01 <0.01 <0.01 <0.02
Partial R2
0.003 0.71 0.003
R2
= 0.72
b0 b1
b2
b3
Wheat Leaf (IR) = 0.0205 - 7.0-6
(N) - 1.5-5 (GDD) + 1.0-7 (GDR)
P-value <0.01 <0.01 <0.01 <0.01
Partial R2
0.0006 0.77 0.008
R2
= 0.78

Table 3.3. Regression equations describing species leaf area index (RLAI)(WLAI) with independent variables of wheat density (N)W, ryegrass density (N)R, andinteractions of wheat density*ryegrass density (INT), ryegrass density*GDD GDR),
wheat density*GDD (GDW), and wheat variety*wheat density (VarW). Partial R aredisplayed to indicate most influential variables determining canopy leaf area index.
Ryegrass (RLAI)P-valuePartial R
2
= 0.6734<0.01
+ 0.0309(N)R<0.010.33
- 0.0030(N)w<0.010.07
- 4.0E-4(INT) - 8.0E-5(GDR)<0.01 <0.01
0.01 0.05
R2
= 0.47
b0
b1
b2
b b4
Wheat (WLAI) = 0.7010 + 0.216(N) - 1.0E-4(GDW) - 1.0E-5(GDR) + 0.0020(VarW)P-value
2<0.01 <0.01 W <0.01 <0.01 <0.01
Partial R 0.20 0.22 0.02 0.003
R2
= 0.46

68
Figure 3.1. Height growth rates are plottedagainst growing degree days (GDD) for Owensspring wheat and Lolium multiflorum.Plants which were growing in addition seriestreatments with 1:1 proportions are representedin the diagram.
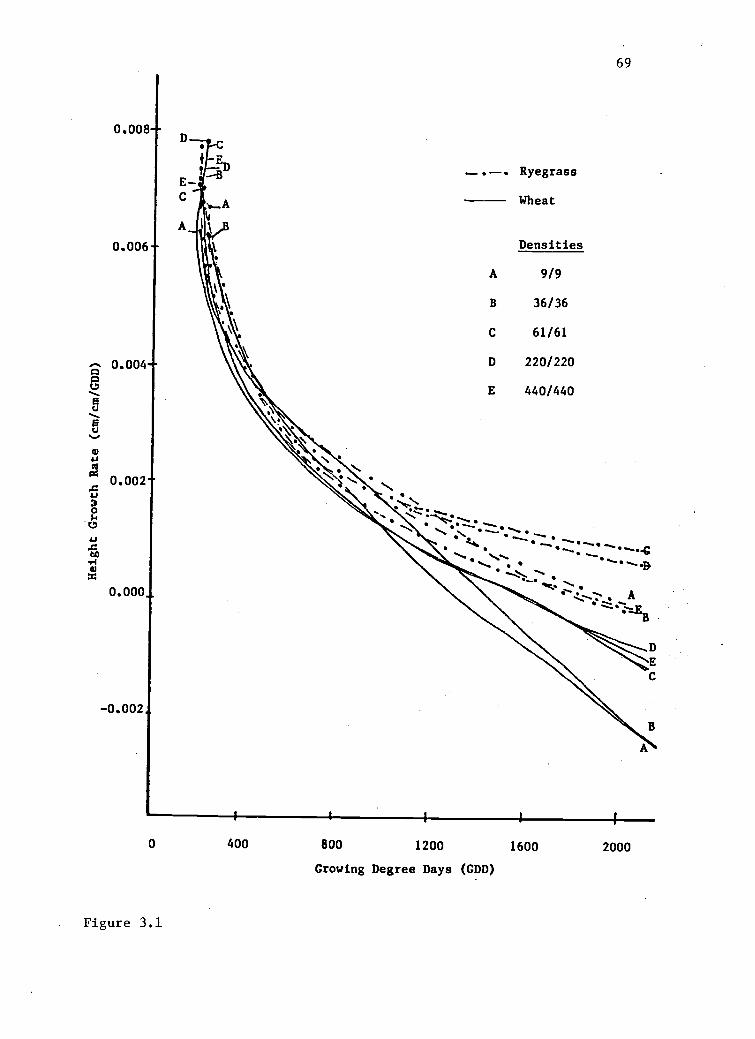
0.008--D
E-C
0.006-
a 0.004 -aLI
iiu
0u
......
'd0.002 -4
4.,
3o
04.,
X140=
0.000_
-0.002_
I E
IN. ..,
69
-_.--. Ryegrass
Wheat
Densities
A 9/9
B 36/36
C 61/61
D 220/220
E 440/440
:".. .....,
....;"'. ....op..... 0S....
-.... 44. .....-... . ...
. ..,..-.4 .V. ....s.'
\Zs:Z. , ,A
i'AEB
E
i 1
0 400 800 1200 1600 2000
Growing Degree Days (GOD)
Figure 3.1

70
Chapter 3 References
Appleby, A.P., P.D. Olson, and D.R. Colbert. 1976.Winter wheat reduction from interference by Italianryegrass. Agron. J. Vol. 68:463-466.
Baker, H.G. 1974. The Evolution of Weeds. Annual Reviewof Ecology and Systematics.
Baskerville, G.L. 1972. Dry-matter production inimmature balsam fir stands:Roots, lesser vegetation, andtotal stand. Forest Science. 12:49-53.
Black, J.N. 1958. Competition between plants ofdifferent initial sizes in swards of subterranean clover(Trifolium. subterranean L.) with particular reference toleaf area and light microclimate. Aust. J. Agric. Res9:299-318.
Cooke, P.T. and S.R. Radosevich. 1987. The Role ofDensity and Proportion In Allometric Equations of Douglas-fir and Red Alder Seedlings. M.S. Thesis. Oregon StateUniversity. Corvallis OR.
Donald, C.M. 1963. Competition for light in crops andpasture. La: Mechanisms in Biological Competition. Soc.Expmt. Biol., Symposium XV. Combridge Press. London. p.283-313.
Esau, K. 1977. Anatomy at Seed Plants. John Wiley andSons, New York, 550 pp.
Firbank, L.G. and A.R. Watkinson. 1987. On the analysisof competition within two-species mixtures of plants. J.Appl. Ecol. 22:503-517.
Grier, C., KI. Lee, and R. Archibald. 1984. Effect ofurea fertilization on allometric relations in youngDouglas-fir trees. Can. J. of For. Res. 14(6):900-904.
Grime, J.P. 1979. Plant Strategies and VegetationProcesses. John Wiley and Sons. 222 pp.
Grime, J.P., J.C. Crick, and J.E. Rincon. 1986. Theecological signifigance of plasticity. Great Britain CSoc. for Ex. Biol.
Gunst, R.F. and R.L. Mason. 1980. Regression Analysis amlLts_ application: g Data-orientated Approach. New York:Marcel Dekker Inc.

71
Harper, J.L. 1977. Population Biology Qj plants. NewYork: Academic Press, 892 pp.
Hunt, R. 1978. Plant Growth Analysis. Inst. Biol.Studies. 96. Edward Arnold. London. 63 pp.
Hunt, R. 1982. Plant Growth Curves. The, Functionalapproach ta Plant Growth Analysis. Baltimore: UniversityPark Press. 248 pp.
Miller, T.E. and P.A. Werner. 1987. Competitive effectsand responses in plants. Ecology (in press).
Newman, E.J. 1983. Interactions between plants.La:Encyclopedia of Plant Physiology New Series Volume 12 C.0.L Lange, P.S Noble, C.B. Osmond, and H. Ziegler (Eds.)Springer-Verlag Berlin/New York. pp. 679-771.
Patterson, D.T. 1982. Effects of light and temperatureon weed/crop growth and competition. _La: Biometerology inI.P.M Academic Press.
Peterson, C.M., B. Klepper, and R.W. Rickman. 1982.Tiller development at the coleoptilar node in winter wheat.Agronomy J. Vol. 74:781-784.
Radosevich, S.R. and J.S. Holt. 1984. WeedEcology:Implications for Management. John Wiley and SonsInc. New York. 300 pp.
Ross, M.A. and J.L. Harper. 1972. Occupation ofbiological space during seedling establishment. J. of Appl.Ecol. 60:77-88.
Roush, M.L. and S.R. Radosevich. 1985. Relationshipsbetween growth and competitiveness of four annual weeds.Journal of Appl. Ecol. 22:895-905.
Roush, M.L. and S.R. Radosevich. 1987. A weed communitymodel of germination, growth, and competitin of annual weedspecies. WSSA abstracs. St.Louis.
SAS Institute. SAS/STAT Guide for Personal computers,Version 6 Edition, 1985. Cary, N.C.:SAS Institute Inc.
SAS Institute. SAS/System for Linear Models. 1986, Cary,N.C.:SAS Institute Inc.
Sestak, Z. 1983. photosynthesis During Leaf Development.

72
Dr. W. Junk Publ., Dordrecht/Boston. 396 pp.
Spitters, C.J.T. 1983 a. An alternative approach to theanalysis of mixed cropping experiments. I. Estimation ofcompetition effects. Netherlands J. 0 Agr. Sci. 31:1-11.
Welden, C.W. and W.L. Slauson. 1986. The intensity ofcompetition versus its importance:an overlooked distinctionand some implications. Quart. Rev. Biol. 61:23-44.

73
CHAPTER 4: CONCLUSIONS
Measuring Competition
Intra- and interspecific competitive biomass responses
were generated in a systematic fashion by using an addition
series. These data were assessed by regressing the
reciprocal transformation of mean plant biomass against
both species densities. A regression model predicting
reciprocal plant weight was produced for each species.
The predictive model for reciprocal wheat biomass and
consequently, wheat yield, indicated that wheat was the
dominant species in mixtures with ryegrass. The relative
competitive ability of wheat suggested that one wheat plant
was as competitive as six to seven ryegrass plants.
The growth analysis experiment confirmed that wheat
was a highly competitive plant in mixture with ryegrass.
The large seed size and consequently large emergent size of
wheat allow this species to rapidly capture above ground
resources. Wheat maintained a dominant role in the
mixtures by producing a higher LAI than ryegrass for the
first half of the season. Wheat LAI also increased faster
in high wheat densities and proportions, suppressing
ryegrass plants in the mixture. However, wheat LAI also
decreased faster at the end of the season in the high wheat
densities. This release of light resources presumably
allowed ryegrass plants to grow and disperse seed

74
maintaining a persistent, though not dominant role in
spring wheat cropping systems.
The final regression model produced to predict
ryegrass reciprocal weight indicated that the interaction
of both species densities described the largest amount of
variation in ryegrass reciprocal weight (R 2= 0.57). The
significant interaction of species densities suggested that
adding plants to a mixture produced an multiplicative
affect on ryegrass reciprocal weight, as well as an
additive one. Biologically, this interaction may be caused
by differences in above-ground competition by larger wheat
plants early in the season or below-ground competition
between both species throughout the season.
If the influence of density is multiplicative, the
strict assumption of additivity on yield-density
relationships is incorrect. Furthermore, if the influence
of density is not additive, then plants in the mixture may
not take up resources or compete equally given an equal
amount of space. This violates a second assumption
(Spitters 1983a) of the expanded reciprocal approach; i.e.
that both species are at maximum resource uptake at
harvest. Thus, biomass predictions may have been over-
estimated if this assumption is not true.
The species density interaction term observed for
ryegrass also may result from a regression model generated

75
from inadequately transformed data. In this experiment,
the best transformation of mean biomass depended on the
degree of variability that each species displayed at
varying density and proportion. Ryegrass, the "loser" over
all densities and proportions, displayed the most variable
reciprocal weights as indicated by the residual plots. The
log transformation which is better able to account for
variance (Satoo and Madgwick 1982) may have been the
correct transformation. However, logarithmic equations are
not linear. Distribution of In (Y), at (X) which is normal
skews the distribution of (Y). The antilog of In (Y) used
to convert the transformed yield, resulting from the
regression equation, back to predictive yield, becomes the
median of the skewed distribution which is not the mean of
biomass (Baskerville, 1972).
Perhaps linear regression is not the most precise
method of assessing competitive biomass response. Mean
yield per plant (W) has universally been modeled as a
rectangular hyperbolic response to density (N) (Watkinson
1980, Bleasdale and Nelder 1960).
W = A + BN
The weight of an individual plant (W) raised to the
exponent (9) is a linear function of density (N). This
exponent (0) has been assumed as 1.0. The use of -1.0
for the exponent (5) led to the Reciprocal Yield Law:

76
1/W = A + BN.
8 may not approach the value of -1.0 in many situations
(Watkinson 1980, Firbank and Watkinson 1985) therefore,
linear regression may not be suitable for assessing density
responses. Non-linear regression has been suggested to
estimate the value of the exponent (9) and also the values
of other linear parameters in such equations (Watkinson
1980, Firbank and Watkinson 1985, Roush and Radosevich (in
preparation)) when one or more species in the mixture has
not attained maximum resource utilization.
Firbank and Watkinson (1987) proposed a
reparameterization of the reciprocal yield regression
equation:
W = Wm (1 + aN) b
The exponent b -1is equivalent to ( 8 of reciprocal yield
equation). The exponent b describes the curvilinear
response of species mean individual plant biomass to
species density. This may be interpreted as the resource
uptake capability of a species in monoculture or mixture
(Watkinson 1980, Firbank and Watkinson 1985, Roush and
Radosevich (in preparation)). (Wm) defines the maximum
size of a plant without competition. The parameter (a)
describes the affect of density (N) on mean plant biomass
and is an actual area necessary to avoid interaction with
other neighboring plants (Roush and Radosevich, in

77
preparation).
The Importance of Competition
Intra- and interspecific competition were important
factors influencing both species biomass yields as was
indicated by the coefficients of determination for the
final regression models (Table 2.6, Table 2.7). However,
only 59 percent of the variation in ryegrass reciprocal
weight was explained by intra- and interspecific
competitive responses. Several alternative factors could
have accounted for some of the unexplained variation. For
instance, gopher predation on wheat and ryegrass plants was
observed in the low total densities in the field (personal
observation). Downy mildew also existed in the high total
densities on both species (personal observation). A larger
amount of disturbance also seemed to play a role in
determining the competitive relationships in this
experiment. Seeds sown in the highest density of wheat
(440 plants/m2
) germinated one day earlier than in the
other densities in all cases. This early germination may
have determined competitive relationships from the
beginning of the season. Predation, disease and
disturbance are factors which may interact with proximity
factors and should also be included in predictive plant
growth models. Future experimentation should be conducted

78
to determine the role of these factors on plant growth.
Agricultural Implications
Wheat is a strong competitor against ryegrass in
mixture. However, this experiment also implicated wheat
neighbors in the populations as the strongest competitors
for resources against other wheat plants. Wheats
competitive nature can be more detrimental to "potential"
grain yield than ryegrass at high wheat densities (220-440
plants/m2).
The timing of germination also was very important in
spring wheat cropping systems. When wheat germinates with
or before ryegrass, the crop maintains an efficient
relative competitive ability. The constant rate of growth
that ryegrass displayed in most measured plant parameters
indicated that ryegrass could be a formidable competitor,
if allowed to germinate before the wheat crop.
Though this study was an empirical one, the regression
models and the growth analysis experiment revealed several
testable hypotheses concerning the mechanisms by which both
species capture available resources in time and space:
1. Light may be the limiting resources for ryegrass at the
beginning of the season, as suggested by the wheat LAI
measurements taken throughout the season. This suggests
that increasing wheat density above 440 plants/m2, which is

79
the normal planting density for Oregon farmers, may suppress
ryegrass weed growth. However, the second application of
nitrogen (applied in split-application) may be the most
limiting factor for both species during the latter half of
the season. Ryegrass may compete quite effectively for
nitrogen in the latter half of the season with the wheat
plants in the mixture, reducing wheat grain yield. The
persistence of Italian ryegrass in spring wheat cropping
systems suggests that competition in the latter half of the
growing season needs to be further examined.
2. Wheat varieties should be selected which have an
inherently larger height than ryegrass. Earlier
germinating varieties also would be advantageous for wheat
to maintain the dominant role in the mixture during the
growing season. Owens spring wheat produced higher grain
yields than Waverly throughout the addition series
experiment. The 10 cm difference appears to have made a
competitive difference.

80
BIBLIOGRAPHY
Appleby, A.P., P.D. Olson, and D.R. Colbert. 1976.Winter wheat reduction from interference by Italianryegrass. Agron. J. Vol. 68:463-466.
Baker, H.G. 1974. The Evolution of Weeds. Annual Reviewof Ecology and Systematics.
Baskerville, G.L. 1972. Dry-matter production inimmature balsam fir stands: Roots, lesser vegetation, andtotal stand. Forest Sci. 12:49-63.
Black, J.N. 1958. competition between plants ofdifferent initial sizes in swards of subterranean clover(Trifolium subterranean) with particular reference to leafarea and light microclimate. Aust. J. Agric. Res. 9:299-318.
Bleasdale, J.K.A., and J.A. Nelder. 1960. Plantpopulation and crop yield. Nature, 188, p. 342.
Bleasdale, J.K.A. 1967. Systematic designs for spacingexperiments. Experimental Agriculture, 3:73-85.
Carlson, H.L. and J.E. Hill. 1986. Wild oat (Avena fatua)competition with spring wheat:effects of nitrogenfertilization. Weed Sci 34:29-33.
Connolly, J. 1986. On difficulties with replacementseries methodology in mixture experiments. J. of AppliedEcology, 23:125-137.
Cooke, P.T. and S.R. Radosevich. 1987. The Role ofDensity and Proportion In Allometric Equations of Douglas-fir and Red Alder Seedlings. M.S. Thesis. Oregon StateUniversity. Corvallis OR.
Cousens, R.D. 1986. The use of population models in thestudy of the economics of weed control. Proc. EWRSSymposium, 1986. Economic Weed Control, 269-276.
de Wit, C.T. 1960. On Competition. No. 66.8 WageningenThe Netherlands: Center for Agricultural Publications andDocumentation.
Donald, C.M. 1963. Competition for light in crops andpasture. La: Mechanisms in Biological Competition. Soc.Expmt. Biol., Symposium XV. Combridge Press. London. pp.283-313.

81
Esau, K. 1977. Anatomy Q. Seed Plants. John Wiley andSons, New York, 550 pp.
Firbank, L.G. and A.R. Watkinson. 1985. On the analysisof competition within two-species mixture of plants. J. ofApplied Ecology, 22:503-517.
Firbank, L.G. and A.R. Watkinson. 1987. On the analysisof competition within two-species mixtures of plants. J.Appl. Ecol. 22:503-517.
Grime, J.P. 1979. Plant Strategies aaa VegetationProcesses. John Wiley and Sons, N.Y. 222 p.
Gunst, R.F. and R.L. Mason. 1980. Rearession Analysisand Itg Application: g Data- orientated Approach. New York:Marcel Dekker Inc.
Harper, J.L. 1977. Population Biology af. Plants.Academic Press, New York. 892 pp.
Harper, J.L., J.T. Williams, and G.R. Sagar. 1965. Thebehavior of seeds in soil. I. The heterogeneity of soilsurfaces and its role in determining the establishment ofplants from seed. J. Ecol. 53:273-286.
Hunt, R. 1978. Plant Growth, Analysis. Inst. Biol.Studies 96. Edward Arnold. London. 63 pp.
Hunt, R. 1982. Plant Growth Curves. The FunctionalApproach la Plant Growth Analysis. Baltimore. UniversityPark Press. 248 p.
Jolliffe, P.A., A.N. Minjas, and V.C. Runeckles. 1984. Areinterpetation of yield relationships in replacementseries experiments. J. of Appi. Ecol., 21:227-243.
Jewiss, O.R. and J. Woledge. 1967. The effect of age onthe rate of apparent photosynthesis in leaves of tallfescue (Pestucl arundinacea Schreb.). Ann. Bot. 31:661-671.
Little T.M. and F.J. Hills. 1978. AgriculturalExperimentation:Design and Analysis. John Willey and SonsInc. Canada.
Miller, T.E. and P.A. Werner. 1987. Competitive effectsand responses in plants. Ecology (in press),

82
Neter, J., W. Wasserman, and M.H. Kutner. 1983.AooliedLinear Regression Models Richard D. Irwin Inc. Illinois.
Newman, E.J. 1983. Interactions between plants. fin:
Encyclopedia of Plant Physiology New Series Volume 12 C.O.L. Lange, P.S. Nobel, C.B. Osmond, and H. Ziegler (Eds.).Springer-Verlag Berlin/New York. pp. 679-771.
Noguchi, K. and K. Nakayama. 1978a. Jap. J. Crop sci.47:48-55.
Patterson, D.T. 1982. Effects of light and temperature onweed/crop growth and competition. Ia:Biometerology inI.P.M. Academic Press.
Peterson, C.M., B. Klepper, and R.W. Rickman. 1982.Tiller development at the coleoptilar node in winter wheat.Agronomy J. 74:781-784.
Radosevich, S.R. 1987 (in press). Experimental methodsto study crop and weed interactions. Ia:Altieri, M.A., M.Z.Liebman (Eds). Plant Competition and Other EcologicalApproaches to Weed Control in Agriculture. CRC Press.
Radosevich, S.R. and J.S. Holt. 1984. WeedEcology:Imolications for. Management. John Wiley and SonsInc. New York. 300 pp.
Ross, M.A. and J.L. Harper. 1972. Occupation ofbiological space during seedling establishment. J. ofAppl. Ecol. 60:77-88.
Roush, M.L. and S.R. Radosevich. 1985. Relationshipsbetween growth and competitiveness of four annual weeds.J. of Appl. Ecol. 22:895-905.
Roush, M.L. and S.R. Radosevich. 1987. A weed communitymodel of germination, growth, and competition of annualweed species. WSSA abstracts. St. Louis.
SAS Institute. SAS/STAT Guide for Personal computers,Version 6 Edition, 1985. Cary, N.C.:SAS Institute Inc.
SAS Institute. SAS/System for Linear Models. 1986, Cary,N.C.:SAS Institute Inc.
Satoo, T. and H.A.I. Madgewick. 1982. Forest biomass.The Hague, Netherlands: Martinus Nijhoff. 152 pp.
Sestak, Z. 1983. photosynthesis During Leaf, development.

83
Dr. W. Junk Publ., Dordrecht/Boston. 396 pp.
Shinozaki, K. and T. Kira. 1956. Intraspecificcompetition among higher plants. V11. Logistic theory ofthe C-D effect. J. of the Inst. of Polytechnics, Osaka CityUniversity, 7:35-72.
Spitters, C.J.T. 1983 a. An alternative approach to theanalysis of mixed cropping experiments. I. Estimation ofcompetition effects. Netherlands J. of Aqr. Sci. 31:1-11.
Spitters, C.J.T. 1983 b. An alternative approach to theanalysis of mixed cropping experiments. II. Marketableyield. Netherlands J. of Agr. Sci. 31143-155.
Tilman, D. 1982. Resource competition and communityStructure.Princeton University Press. Princeton, NewJersey.
Watkinson, A.R. 1980. Density-dependence in single-species populations of plants. J. Theor. Biol. 83:345-357.
Welden, C.W. and W.L. Slauson. 1986. The intensity ofcompetition versus its importance:an overlooked distinctionand some implications. Quart. Rev. Biol. 61:23-44.
Westoby, M. 1981. The place of the self thinning rule inpopulation dynamics. Am. Nat. 118, pp. 581-587.
Willey. R.W. 1985. Evaluation and presentation ofintercropping advantages. Expl. Agric. 21:119-133.
Yoda, K., T. Kira, H. Ogawa, and H. Hozumi. 1963. Self-thinning in overcrowded pure stands under cultivated andnatural conditions. J. Biol. Osaka City University. 14:107-129.
Zimdahl, R.H. 1980. Weed-crop Competition:A Review.International Plant Protection Center, Corvallis, OR. 196pp.

APPENDI CES

84
APPENDICES
Tiller Number
An analysis of variance performed on final number ofryegrass and wheat tillers, in the two variety experiments,indicated that variation in species tiller number wasattributed to ryegrass density (p < 0.01),
wheat density (p< 0.01), and interaction of species densities (p < 0.01).Number of ryegrass and wheat tillers
decreased with anincrease in species relative and total density. Finalnumber of tillers produced by wheat and ryegrass plants inthe addition series (Table 2.1) were influenced byproximity factors of species density and proportion (Figurea.1).
Analyses of variance and covariance performed onrelative rates of ryegrass tillering suggests that 15percent of variation in rates (r2 = 0.15) was explained byindependent variables of wheat variety (p < 0.01), wheatdensity (p < 0.01), ryegrass density (p < 0.01), GDD (p <0.01), and
interactions of GDD*wheat variety (p < 0.01),GDD*wheat density (p < 0.01),
GDD*ryegrass density (p <0.01), and wheat
density*ryegrass density (p < 0.01) (Tablea.1).
Analyses of variance and covariance performed on wheat

85
relative tillering rates indicates that 21 percent ofvariation in rates (r
2= 0.21) was explained by independent
variables of wheat variety (p < 0.01), wheat density (p <0.01), and ryegrass density (p < 0.01), GDD (P < 0.01), andthe interactions of wheat variety*GDD (p < 0.01), GDD*wheatdensity (p < 0.01), GDD*ryegrass density (p < 0.01), andryegrass density*wheat density (p < 0.01) (Table a.1).
Final number of ryegrass and wheat tillers (Figurea.1) resemble total grain yield (Figure 2.1). These datasuggest that tiller relative growth rate (tiller biomass)would have been a better indicator of competitive response.
Individual Plot Measurements
Individual plot measurements from the addition seriesexperiments have been included in Table a.2 and Table a.3.

Table a.l. Tiller number growth rates (GR) described by experimental proximity factors and their interaction terms, including
Wheat Density (N)W
, Ryegrass Density (N)R
. Growing Degree Days (GDD), VarietyGDD (VGDD), Wheat Density*GDD (GDW), Ryegrass
Density`GDD (GD0). and Whea2 t Density`Ryegrass Density (INT). Although the variables were highly significant, the low coeffi-
ciente of determination (R ) indicated that competition was not an important process controlling the rate of this growth
parameter.
Species b b0 1
b b b b b b2 3 4 5 6 7
Ryegrass Tiller GR
Variety 1 - 0.0048 - 2.4-4
(N) - 1.9-5
(N) - 3.9-6
(GDD) 9.2-7
(VGDD) + 4.0-9
(GDw) + 1.8-4
(GDR) + 1.1-4
(INT)W R
Variety 2 . 0.0036
P-value
2B . 0.15
<0.01 <0.01 <0.01 <0.01 <0.01 <0.01 <0.01 <0.01
b b b b b0 1 2 3 4
b b b5 6 7
Wheat Tiller GR =
P-value
2R = 0.21
Variety 1 = 0.0059 - 2.9-5
(N) - 2.2-5
(N) - 4.5-6
(GOD) + 8.4-7
(VGDD) + 1.6-8
(GDw) + 1.2-8(GDR) + 7.8
-8(INT)
W RVariety 2 = 0.0048
<0.01 <0.01 <0.01 <0.01 <0.01 <0.01 <0.01 <0.01
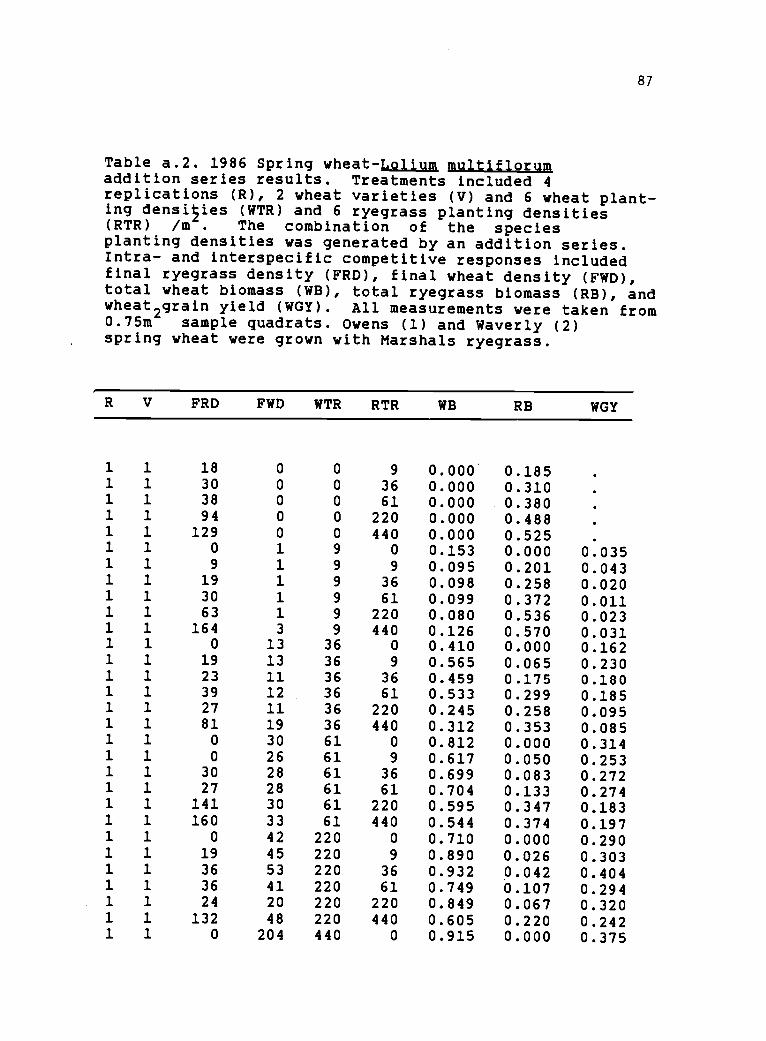
87
Table a.2. 1986 Spring wheat-Lolium multiflorumaddition series results. Treatments included 4replications (R), 2 wheat varieties (V) and 6 wheat plant-ing densiies (WTR) and 6 ryegrass planting densities(RTR) /m . The combination of the speciesplanting densities was generated by an addition series.Intra- and interspecific competitive responses includedfinal ryegrass density (FRD), final wheat density (FWD),total wheat biomass (WB), total ryegrass biomass (RB), andwheat
2grain yield (WGY). All measurements were taken from
0.75m sample quadrats. Owens (1) and Waverly (2)spring wheat were grown with Marshals ryegrass.
R V FRD FWD WTR RTR WB RB WGY
1 1 18 0 0 9 0.000' 0.1851 1 30 0 0 36 0.000 0.3101 1 38 0 0 61 0.000 0.3801 1 94 0 0 220 0.000 0.4881 1 129 0 0 440 0.000 0.5251 1 0 1 9 0 0.153 0.000 0.0351 1 9 1 9 9 0.095 0.201 0.0431 1 19 1 9 36 0.098 0.258 0.0201 1 30 1 9 61 0.099 0.372 0.0111 1 63 1 9 220 0.080 0.536 0.0231 1 164 3 9 440 0.126 0.570 0.0311 1 0 13 36 0 0.410 0.000 0.1621 1 19 13 36 9 0.565 0.065 0.2301 1 23 11 36 36 0.459 0.175 0.1801 1 39 12 36 61 0.533 0.299 0.1851 1 27 11 36 220 0.245 0.258 0.0951 1 81 19 36 440 0.312 0.353 0.0851 1 0 30 61 0 0.812 0.000 0.3141 1 0 26 61 9 0.617 0.050 0.2531 1 30 28 61 36 0.699 0.083 0.2721 1 27 28 61 61 0.704 0.133 0.2741 1 141 30 61 220 0.595 0.347 0.1831 1 160 33 61 440 0.544 0.374 0.1971 1 0 42 220 0 0.710 0.000 0.2901 1 19 45 220 9 0.890 0.026 0.3031 1 36 53 220 36 0.932 0.042 0.4041 1 36 41 220 61 0.749 0.107 0.2941 1 24 20 220 220 0.849 0.067 0.3201 1 132 48 220 440 0.605 0.220 0.2421 1 0 204 440 0 0.915 0.000 0.375

88
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
1 1 22 210 440 9 0.944 0.017 0.3821 1 32 209 440 36 0.889 0.4021 1 61 266 440 61 0.942 0.035 0.3871 1 61 176 440 220 0.891 0.002 0.3251 1 135 220 440 440 0.885 0.045 0.3432 1 2 0 0 9 0.000 0.0662 1 24 0 0 36 0.000 0.2652 1 24 0 0 61 0.000 0.2532 1 68 0 0 220 0.000 0.4892 1 124 0 0 440 . . .
2 1 0 0 9 0 . . .
2 1 3 1 9 9 0.068 0.0352 1 8 122 9 36 0.083 0.135 0.0332 1 23 2 9 61 0.096 . 0.0142 1 48 1 9 220 0.079 0.281 0.0032 1 142 1 9 440 0.020 0.549 0.0092 1 0 11 36 0 0.498 0.000 0.2282 1 20 14 36 9 0.562 0.043 0.2642 1 31 16 36 36 0.565 0.485 0.2242 1 45 13 36 61 0.398 0.190 0.1612 1 88 7 36 220 0.152 0.369 0.0612 1 148 16 36 440 0.265 0.383 0.0982 1 0 22 61 0 0.748 0.000 0.3142 1 4 24 61 9 0.718 0.072 0.3402 1 7 30 61 36 0.731 0.030 0.3102 1 52 23 61 61 0.382 0.216 0.1372 1 40 17 61 220 0.395 0.187 0.1622 1 67 271 61 440 0.395 0.225 0.1462 1 0 51 220 0 0.875 0.000 0.3552 1 2 42 220 9 0.679 0.015 0.2502 1 10 46 220 36 0.810 0.036 0.3192 1 29 51 220 61 0.765 0.070 0.3052 1 77 46 220 220 0.645 0.124 0.2372 1 98 57 220 440 0.575 0.225 0.2182 1 0 216 440 0 1.035 0.000 0.4012 1 2 274 440 9 1.038 0.020 0.4062 1 10 216 440 36 0.959 0.020 0.3742 1 20 180 440 61 0.909 0.030 0.3652 1 29 184 440 220 0.809 0.036 0.3202 1 111 196 440 440 0.745 0.085 0.3013 1 0 0 0 0 . .
3 1 9 0 0 9 . .
3 1 18 0 0 36 0.000 0.1983 1 65 15 0 61 0.000 0.2553 1 130 0 0 220 0.000 0.4603 1 224 0 0 440 0.000 0.513

89
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
3 1 0 1 9 0 0.200 0.000 0.0323 1 1 1 9 9 0.106 0.189 0.0193 1 18 1 9 36 0.052 0.180 0.0163 1 57 2 9 61 0.072 0.343 0.0113 1 130 1 9 220 0.041 0.593 0.0113 1 155 3 9 440 0.120 0.595 0.0253 1 0 9 36 0 0.380 0.000 0.1303 1 0 13 36 9 0.596 0.015 0.2393 1 56 24 36 36 0.445 0.124 0.1793 1 0 21 36 61 0.485 0.2013 1 160 18 36 220 0.460 0.338 0.1783 1 125 14 36 440 0.351 0.332 0.1383 1 0 4 61 0 .
3 1 11 22 61 9 0.682 0.069 0.2603 1 29 24 61 36 0.536 0.123 0.1973 1 37 20 61 61 0.484 0.183 0.1023 1 95 28 61 220 0.438 0.248 0.1533 1 83 21 61 440 0.355 0.184 0.1323 1 0 64 220 0 0.830 0.000 0.3373 1 7 45 220 9 0.792 0.027 0.2803 1 11 58 220 36 0.765 0.026 0.3123 1 0 70 220 61 0.655 0.093 0.2783 1 116 61 220 220 0.696 0.166 0.2903 1 . . 220 440 . . .
3 1 0 175 440 0 0.817 0.000 0.3363 1 6 215 440 9 0.908 0.021 0.3613 1 0 198 440 36 0.736 0.020 0.3173 1 5 163 440 61 0.734 0.026 0.3063 1 74 206 440 220 0.865 0.050 0.3503 1 82 178 440 440 0.697 0.075 0.2814 1 0 0 0 0 . . .4 1 10 0 0 9 0.000 0.120 .
4 1 39 0 0 36 0.000 0.179 .
4 1 46 0 0 61 0.000 0.222 .4 1 106 0 0 220 0.000 0.312 .
4 1 279 0 0 440 0.000 0.434 .
4 1 0 1 9 0 0.139 0.000 0.0514 1 8 0 9 9 0.110 0.187 0.0194 1 16 1 9 16 0.000 0.073 0.0124 1 11 1 9 61 0.130 0.134 0.0064 1 98 0 9 220 0.099 0.376 0.0184 1 85 1 9 440 0.030 0.345 0.0134 1 0 9 36 0 0.325 0.000 0.1064 1 3 11 36 9 0.362 0.045 0.1554 1 27 6 36 36 0.207 0.128 0.0974 1 29 11 36 61 0.248 0.171 0.100

90
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
4 1 89 11 36 220 0.282 0.252 0.1174 1 110 9 36 440 0.215 0.361 0.0984 1 0 29 61 0 0.714 0.000 0.2864 1 8 24 61 9 0.572 0.027 0.2114 1 34 19 61 36 0.517 0.073 0.2364 1 46 17 61 61 0.413 0.125 0.1714 1 95 22 61 220 0.375 0.131 0.1474 1 97 27 61 440 0.399 0.172 0.1504 1 0 62 220 0 0.861 0.000 0.3944 1 16 49 220 9 0.805 0.032 0.3434 1 8 49 220 36 0.731 0.024 0.3324 1 32 54 220 61 0.872 0.050 0.3504 1 71 48 220 220 0.612 0.102 0.2374 1 221 44 220 440 0.723 0.170 0.2704 1 0 210 440 0 0.925 0.000 0.3714 1 12 192 440 9. 0.887 0.025 0.3564 1 31 184 440 36 0.747 0.030 0.3014 1 22 195 440 61 0.844 0.025 0.3384 1 34 212 440 220 0.740 0.030 0.3124 1 207 227 440 440 0.804 0.050 0.2811 2 0 0 0 0 . .
1 2 4 0 0 9 0.000 0.1541 2 22 0 0 36 0.000 0.3051 2 35 0 0 61 0.000 0.3651 2 96 0 0 220 0.000 0.4091 2 190 0 0 440 0.000 0.635 .
1 2 0 2 9 0 0.195 0.000 0.0531 2 31 5 9 9 . 0.157 0.0161 2 31 3 9 36 0.181 0.261 0.0461 2 30 1 9 61 0.034 0.305 0.0071 2 82 1 9 220 0.065 0.477 0.0131 2 93 3 9 440 0.119 0.340 0.0271 2 . 0 36 0 . . .
1 2 2 15 36 9 0.725 0.030 0.2861 2 23 9 36 36 0.511 0.014 0.2201 2 37 14 36 61 0.462 0.185 0.2031 2 95 14 36 220 0.312 . 0.1141 2 85 16 36 440 0.180 0.408 0.0641 2 0 25 61 0 0.760 0.000 0.3171 2 6 29 61 9 0.765 0.050 0.2841 2 21 23 61 36 0.611 0.102 0.2221 2 40 25 61 61 0.644 0.139 0.2511 2 84 23 61 220 0.408 0.255 0.1421 2 87 . 61 440 . 0.476 .
1 2 0 98 220 0 0.819 0.000 0.3391 2 3 53 220 9 0.951 0.018 0.395

91
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
1 2 23 56 220 36 0.847 0.060 0.3441 2 44 5 220 61 0.839 0.144 0.3201 2 93 47 220 220 0.568 0.155 0.2091 2 165 59 220 440 0.702 0.298 0.2761 2 0 189 440 0 0.815 0.000 0.3521 2 5 188 440 9 0.994 0.020 0.3791 2 . 179 440 36 0.905 0.3531 2 27 192 440 61 0.910 0.013 0.3591 2 89 213 440 220 0.870 0.071 0.3361 2 72 186 440 440 0.745 0.091 0.2732 2 0 0 0 0 .
2 2 4 0 0 9 0.000 0.0872 2 12 0 0 36 0.000 0.1382 2 30 0 0 61 0.000 0.2402 2 72 0 0 220 0.000 0.441 .
2 2 163 0 0 440 0.280 0.421 .
2 2 0 2 9 0 0.490 0.0002 2 8 1 9 9 0.095 0.152 0.0222 2 29 3 9 36 0.225 0.225 0.0952 2 29 1 9 61 0.035 0.314 .
2 2 60 3 9 220 0.050 0.391 0.0202 2 64 3 9 440 0.105 0.446 .
2 2 0 14 36 0 0.525 0.000 0.2022 2 4 14 36 9 0.524 0.020 0.2032 2 12 10 36 36 0.462 0.076 0.2132 2 15 11 36 61 0.446 0.103 0.1932 2 66 9 36 220 0.290 0.210 0.1072 2 161 10 36 440 0.301 0.381 0.1332 2 0 32 61 0 . . .
2 2 11 19 61 9 0.665 0.045 0.2952 2 22 40 61 36 0.585 0.050 0.1872 2 26 24 61 61 0.597 0.084 0.2452 2 149 25 61 220 0.501 0.330 0.2012 2 100 19 61 440 0.185 0.252 0.1192 2 0 40 220 0 .
2 2 12 49 220 9 0.788 0.016 0.3042 2 20 75 220 36 0.704 0.047 0.2842 2 25 38 220 61 0.584 0.072 0.2502 2 56 47 220 220 0.413 0.015 0.1452 2 110 54 220 440 0.400 0.190 0.1352 2 0 187 440 0 0.670 0.000 0.3532 2 2 198 440 9 0.945 0.000 0.3622 2 16 165 440 36 0.843 0.001 0.3032 2 24 194 440 61 0.940 0.045 0.3822 2 39 162 440 220 0.830 0.025 0.2982 2 90 190 440 440 0.566 0.070 0.226

92
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
3 2 0 0 0 0 . . .
3 2 8 0 0 9 0.000 0.1013 2 15 0 0 36 0.000 0.230 .
3 2 37 0 0 61 0.000 0.255 .
3 2 53 0 0 220 0.000 0.265 .
3 2 71 0 0 440 0.000 0.348 .
3 2 0 1 9 0 0.200 0.000 0.0323 2 10 1 9 9 0.115 0.130 .
3 2 27 1 9 36 0.113 0.314 0.0303 2 44 1 9 61 0.050 0.359 0.0143 2 102 1 9 220 0.043 0.366 0.0093 2 152 1 9 440 0.016 0.470 0.0063 2 0 14 36 0 0.499 0.000 0.2173 2 19 14 36 9 0.385 0.100 0.1523 2 21 13 36 36 0.375 0.095 0.1133 2 10 12 36 61 0.451 0.080 0.1183 2 78 7 36 220 0.254 0.274 0.1033 2 138 11 36 440 0.247 0.400 0.0793 2 0 22 61 0 0.624 0.000 0.2813 2 11 16 61 9 0.495 0.075 0.2123 2 30 24 61 36 0.507 0.096 0.1863 2 13 18 61 61 0.455 0.088 0.1343 2 64 11 61 220 0.540 0.585 0.2153 2 142 20 61 440 0.320 0.338 0.1063 2 0 41 220 0 0.519 0.000 0.1823 2 24 53 220 9 0.698 0.035 0.3013 2 33 52 220 36 0.701 0.088 0.2633 2 41 37 220 61 0.559 0.122 0.1013 2 89 38 220 220 0.493 0.206 0.1823 2 187 40 220 440 0.342 0.307 0.1153 2 0 120 440 0 0.745 0.000 0.2723 2 3 159 440 9 0.791 0.021 0.2883 2 8 173 440 36 0.711 0.021 0.2513 2 14 151 440 61 0.740 0.015 0.2963 2 67 172 440 220 0.742 0.071 0.2693 2 114 190 440 440 0.754 0.100 0.2744 2 0 0 0 0 . . .
4 2 4 0 0 9 0.000 0.065 .
4 2 21 0 0 36 0.000 0.1734 2 45 0 0 61 0.000 0.251 .4 2 82 0 0 220 0.000 0.290 .
4 2 204 0 0 440 0.000 0.422 .
4 2 0 1 9 0 0.060 0.000 0.0094 2 10 1 9 9 0.040 0.086 0.0124 2 42 5 9 36 0.070 0.226 0.0304 2 41 1 9 61 0.062 0.205 0.024

93
Table a.2. cont.
R V FRD FWD WTR RTR WB RB WGY
4 2 69 3 9 220 0.113 0.248 0.0364 2 113 2 9 440 0.055 0.285 0.0194 2 0 12 36 0 0.408 0.000 0.1394 2 9 10 36 9 0.371 0.038 0.1614 2 29 9 36 36 0.281 0.095 0.1244 2 0 12 36 61 0.395 0.070 0.1784 2 16 87 36 220 0.113 0.236 0.1034 2 193 12 36 440 0.233 0.283 0.0954 2 0 25 61 0 0.566 0.000 0.2464 2 26 40 61 9 0.635 0.030 0.2774 2 19 26 61 36 0.535 0.040 0.2394 2 41 26 61 61 0.480 0.092 0.2054 2 78 26 61 220 0.393 0.139 0.1744 2 263 23 61 440 0.331 0.312 0.1284 2 0 52 220 0 0.720 0.000 0.3324 2 4 56 220 9 0.606 0.000 0.2864 2 19 57 220 36 0.266 0.045 0.2634 2 15 38 220 61 0.503 0.070 0.2174 2 135 52 220 220 0.517 0.122 0.1914 2 172 51 220 440 0.524 0.211 0.2184 2 0 183 440 0 0.725 0.000 0.2314 2 2 210 440 9 0.855 0.000 0.3214 2 5 178 440 36 0.749 0.000 0.3064 2 26 180 440 61 0.805 0.020 0.3054 2 10 168 440 220 0.745 0.021 0.3094 2 60 206 440 440 0.660 0.036 0.232

94
Table a.3. Single plot values resulting from the 1986spring wheat-Lolium multiflorumaddition seriesexperiment. Mean plant weight (g/plant) of wheat (MEANW)and ryegrass (MEANR) were transformed into species reci-procal plant weight (INVW (INVR). Reciprocal plant weightwas regressed against final densities (Table a.2) of wheatand ryegrass to assess intra- and interspecific competi-tive responses of both species. As total planting densityof wheat (WTR) and ryegrass (RTR) increased, mean plantweight decreased. Therefore, the plant reciprocal weightincreased. Mean and reciprocal weights calculated from0.75m plots. Experimental treatments included 4replications (R) and 2 wheat varieties (V). Variety 1= Owens and Variety 2 = Waverly.
R V WTR RTR MEANW MEANR INVW INVR
1 1 0 9 10.278 0.09731 1 0 36 10.333 0.09681 1 0 61 10.000 0.10001 1 0 220 5.191 0.19261 1 0 440 4.070 0.24571 1 9 0 153.000 0.006541 1 9 9 95.000 22.333 0.01053 0.04481 1 9 36 98.000 13.579 0.01020 0.07361 1 9 61 99.000 12.400 0.01010 0.08061 1 9 220 80.000 8.508 0.01250 0.11751 1 9 440 42.000 3.476 0.02381 0.28771 1 36 0 31.538 0.031711 1 36 9 43.462 3.421 0.02301 0.29231 1 36 36 41.727 7.609 0.02397 0.13141 1 36 61 44.417 7.667 0.02251 0.13041 1 36 220 22.273 9.556 0.04490 0.10471 1 36 440 16.421 4.358 0.06090 0.22951 1 61 0 27.067 0.036951 1 61 9 23.731 0.042141 1 61 36 24.964 2.767 0.04006 0.36141 1 61 61 25.143 4.926 0.03977 0.20301 1 61 220 19.833 2.461 0.05042 0.40631 1 61 440 16.485 2.337 0.06066 0.42781 1 220 0 16.905 0.059151 1 220 9 19.778 1.368 0.05056 0.73081 1 220 36 17.585 1.167 0.05687 0.85711 1 220 61 18.268 2.972 0.05474 0.33641 1 220 220 42.450 2.792 0.02356 0.35821 1 220 440 12.604 1.667 0.07934 0.60001 1 440 0 4.485 0.222951 1 440 9 4.495 0.773 0.22246 1.29411 1 440 36 4.254 0.23510

95
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR
1 1 440 61 3.541 0.574 0.28238 1.74291 1 440 220 5.062 0.033 0.19753 30.50001 1 440 440 4.023 0.333 0.24859 3.00001 2 0 9 38.500 0.02601 2 0 36 13.864 0.07211 2 0 61 10.429 0.09591 2 0 220 4.260 0.23471 2 0 440 3.342 0.29921 2 9 0 97.500 0.010261 2 9 9 5.065 0.19751 2 9 36 60.333 8.419 0.01657 0.11881 2 9 61 34.000 10.167 0.02941 0.09841 2 9 220 65.000 5.817 0.01538 0.17191 2 9 440 39.667 3.656 0.02521 0.27351 2 36 0
1 2 36 9 48.333 15.000 0.02069 0.06671 2 36 36 56.778 0.609 0.01761 1.64291 2 36 61 33.000 5.000 0.03030 0.20001 2 36 220 22.286 0.044871 2 36 440 11.250 4.800 0.08889 0.20831 2 61 0 30.400 0.032891 2 61 9 26.379 8.333 0.03791 0.12001 2 61 36 26.565 4.857 0.03764 0.20591 2 61 61 25.760 3.475 0.03882 0.28781 2 61 220 17.739 3.036 0.05637 0.32941 2 61 440 5.471 0.18281 2 220 0 8.357 0.119661 2 220 9 17.943 6.000 0.05573 0.16671 2 220 36 15.125 2.609 0.06612 0.38331 2 220 61 167.800 3.273 0.00596 0.30561 2 220 220 12.085 1.667 0.08275 0.60001 2 220 440 11.898 1.806 0.08405 0.55371 2 440 0 4.312 0.231901 2 440 9 5.287 4.000 0.18913 0.25001 2 440 36 5.056 0.197791 2 440 61 4.740 0.481 0.21099 2.07691 2 440 220 4.085 0.798 0.24483 1.25351 2 440 440 4.005 1.264 0.24966 0.79122 1 0 9 33.000 0.03032 1 0 36 11.042 0.09062 1 0 61 10.542 0.09492 1 0 220 7.191 0.13912 1 0 4402 1 9 02 1 9 9 68.000 11.667 0.01471 0.08572 1 9 36 0.680 16.875 1.46988 0.0593

96
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR
2 1 9 61 48.000 0.02083 .
2 1 9 220 79.000 5.854 0.01266 0.17082 1 9 440 20.000 3.866 0.05000 0.25872 1 36 0 45.273 0.02209 .
2 1 36 9 40.143 2.150 0.02491 0.46512 1 36 36 35.312 15.645 0.02832 0.06392 1 36 61 30.615 4.222 0.03266 0.23682 1 36 220 21.714 4.193 0.04605 0.23852 1 36 440 16.562 2.588 0.06038 0.38642 1 61 0 34.000 0.02941 .
2 1 61 9 29.917 18.000 0.03343 0.05562 1 61 36 24.367 4.286 0.04104 0.23332 1 61 61 16.609 4.154 0.06021 0.24072 1 61 220 23.235 4.675 0.04304 0.21392 1 61 440 1.458 3.358 0.68608 0.29782 1 220 0 17.157 0.058292 1 220 9 16.167 7.500 0.06186 0.13332 1 220 36 17.609 3.600 0.05679 0.27782 1 220 61 15.000 2.414 0.06667 0.41432 1 220 220 14.022 1.610 0.07132 0.62102 1 220 440 10.088 2.296 0.09913 0.43562 1 440 0 4.792 0.208702 1 440 9 3.788 10.000 0.26397 0.10002 1 440 36 4.440 2.000 0.22523 0.50002 1 440 61 5.050 1.500 0.19802 0.66672 1 440 220 4.397 1.241 0.22744 0.80562 1 440 440 3.801 0.766 0.26309 1.30592 2 0 9 21.750 . 0.04602 2 0 36 11.500 . 0.08702 2 0 61 8.000 . 0.12502 2 0 220 6.125 . 0.16332 2 0 440 2.583 . 0.38722 2 9 0 245.000 0.00408 .
2 2 9 9 95.000 19.000 0.01053 0.05262 2 9 36 75.000 7.759 0.01333 0.12892 2 9 61 35.000 10.828 0.02857 0.09242 2 9 220 16.667 6.517 0.06000 0.15352 2 9 440 35.000 6.969 0.02857 0.14352 2 36 0 37.500 0.026672 2 36 9 37.429 5.000 0.02672 0.20002 2 36 36 46.200 6.333 0.02165 0.15792 2 36 61 40.545 6.867 0.02466 0.14562 2 36 220 32.222 3.182 0.03103 0.31432 2 36 440 30.100 2.366 0.03322 0.42262 2 61 0 . . .
2 2 61 9 35.000 4.091 0.02857 0.2444
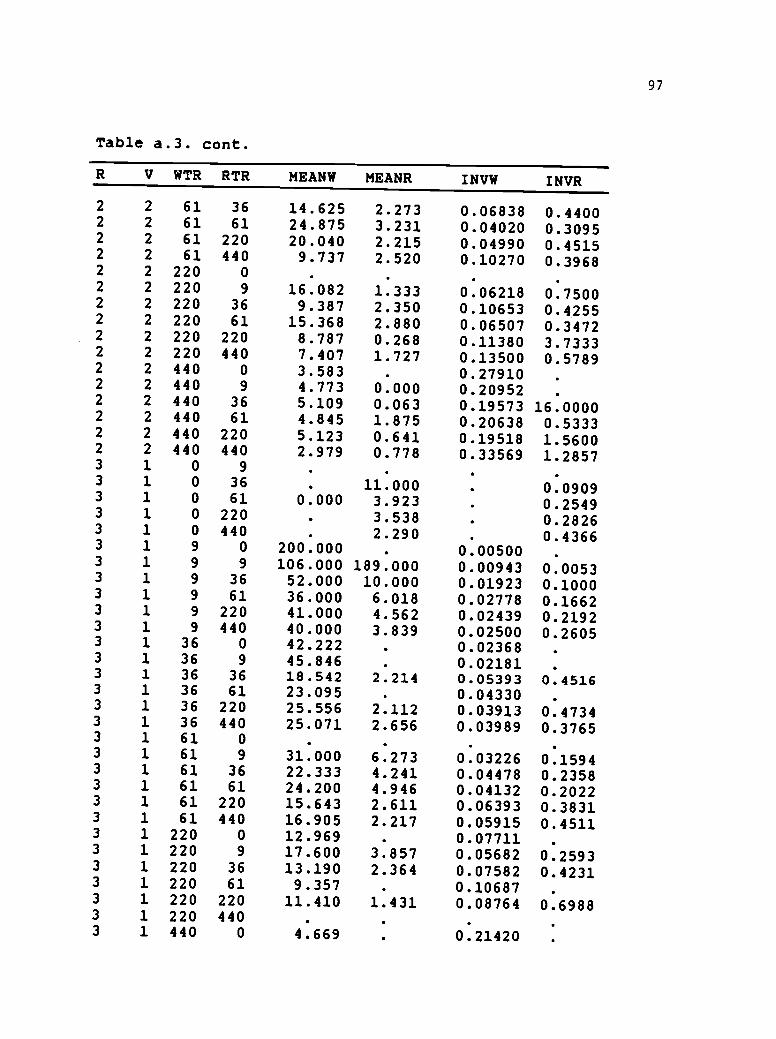
97
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR
2 2 61 36 14.625 2.273 0.06838 0.44002 2 61 61 24.875 3.231 0.04020 0.30952 2 61 220 20.040 2.215 0.04990 0.45152 2 61 440 9.737 2.520 0.10270 0.39682 2 220 02 2 220 9 16.082 1.333 0.06218 0.75002 2 220 36 9.387 2.350 0.10653 0.42552 2 220 61 15.368 2.880 0.06507 0.34722 2 220 220 8.787 0.268 0.11380 3.73332 2 220 440 7.407 1.727 0.13500 0.57892 2 440 0 3.583 0.279102 2 440 9 4.773 0.000 0.209522 2 440 36 5.109 0.063 0.19573 16.00002 2 440 61 4.845 1.875 0.20638 0.53332 2 440 220 5.123 0.641 0.19518 1.56002 2 440 440 2.979 0.778 0.33569 1.28573 1 0 93 1 0 36 11.000 0.09093 1 0 61 0.000 3.923 0.25493 1 0 220 3.538 0.28263 1 0 440 2.290 0.43663 1 9 0 200.000 0.005003 1 9 9 106.000 189.000 0.00943 0.00533 1 9 36 52.000 10.000 0.01923 0.10003 1 9 61 36.000 6.018 0.02778 0.16623 1 9 220 41.000 4.562 0.02439 0.21923 1 9 440 40.000 3.839 0.02500 0.26053 1 36 0 42.222 0.023683 1 36 9 45.846 0.021813 1 36 36 18.542 2.214 0.05393 0.45163 1 36 61 23.095 0.043303 1 36 220 25.556 2.112 0.03913 0.47343 1 36 440 25.071 2.656 0.03989 0.37653 1 61 03 1 61 9 31.000 6.273 0.03226 0.15943 1 61 36 22.333 4.241 0.04478 0.23583 1 61 61 24.200 4.946 0.04132 0.20223 1 61 220 15.643 2.611 0.06393 0.38313 1 61 440 16.905 2.217 0.05915 0.45113 1 220 0 12.969 0.077113 1 220 9 17.600 3.857 0.05682 0.25933 1 220 36 13.190 2.364 0.07582 0.42313 1 220 61 9.357 0.106873 1 220 220 11.410 1.431 0.08764 0.69883 1 220 4403 1 440 0 4.669 0.21420

98
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR3 1 440 9 4.223 3.500 0.23678 0.28573 1 440 36 3.717 0.269023 1 440 61 4.503 5.200 0.22207 0.19233 1 440 220 4.199 0.676 0.23815 1.48003 1 440 440 3.916 0.915 0.25538 1.09333 2 0 9 12.625 0.07923 2 0 36 15.333 0.06523 2 0 61 6.892 0.14513 2 0 220 5.000 0.20003 2 0 440 4.901 0.20403 2 9 0 200.000 0.005003 2 9 9 115.000 13.000 0.00870 0.07693 2 9 36 113.000 11.630 0.00885 0.08603 2 9 61 50.000 8.159 0.02000 0.12263 2 9 220 43.000 3.588 0.02326 0.27873 2 9 440 16.000 3.092 0.06250 0.32343 2 36 0 35.643 0.028063 2 36 9 27.500 5.263 0.03636 0.19003 2 36 36 28.846 4.524 0.03467 0.22113 2 36 61 37.583 8.000 0.02661 0.12503 2 36 220 36.286 3.513 0.02756 0.28473 2 36 440 22.455 2.899 0.04453 0.34503 2 61 0 28.364 0.035263 2 61 9 30.937 6.818 0.03232 0.14673 2 61 36 21.125 3.200 0.04734 0.31253 2 61 61 25.278 6.769 0.03956 0.14773 2 61 220 49.091 9.141 0.02037 0.10943 2 61 440 16.000 2.380 0.06250 0.42013 2 220 0 12.659 0.079003 2 220 9 13.170 1.458 0.07593 0.68573 2 220 36 13.481 2.667 0.07418 0.37503 2 220 61 15.108 2.976 0.06619 0.33613 2 220 220 12.974 2.315 0.07708 0.43203 2 220 440 8.550 1.642 0.11696 0.60913 2 440 0 6.208 0.161073 2 440 9 4.975 7.000 0.20101 0.14293 2 440 36 4.110 2.625 0.24332 0.38103 2 440 61 4.901 1.071 0.20405 0.93333 2 440 220 4.314 1.060 0.23181 0.94373 2 440 440 3.968 0.877 0.25199 1.14004 1 0 9 12.000 0.08334 1 0 36 4.590 0.21794 1 0 61 4.826 0.20724 1 0 220 2.943 0.33974 1 0 440 1.556 0.64294 1 9 0 139.000 0.007194 1 9 9 23.375 0.0428

99
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR
4 1 9 36 0.000 4.562 0.21924 1 9 61 130.000 12.182 0.00769 0.08214 1 9 220 3.837 0.26064 1 9 440 30.000 4.059 0.03333 0.24644 1 36 0 36.111 0.027694 1 36 9 32.909 15.000 0.03039 0.06674 1 36 36 34.500 4.741 0.02899 0.21094 1 36 61 22.545 5.897 0.04435 0.16964 1 36 220 25.636 2.831 0.03901 0.35324 1 36 440 23.889 3.282 0.04186 0.30474 1 61 0 24.621 0.040624 1 61 9 23.833 3.375 0.04196 0.29634 1 61 36 27.211 2.147 0.03675 0.46584 1 61 61 24.294 2.717 0.04116 0.36804 1 61 220 17.045 1.379 0.05867 0.72524 1 61 440 14.778 1.773 0.06767 0.56404 1 220 0 13.887 0.072014 1 220 9 16.429 2.000 0.06087 0.50004 1 220 36 14.918 3.000 0.06703 0.33334 1 220 61 16.148 1.563 0.06193 0.64004 1 220 220 12.750 1.437 0.07843 0.69614 1 220 440 16.432 0.769 0.06086 1.30004 1 440 0 4.405 0.227034 1 440 9 4.620 2.083 0.21646 0.48004 1 440 36 4.060 0.968 0.24632 1.03334 1 440 61 4.328 1.136 0.23104 0.88004 1 440 220 3.491 0.882 0.28649 1.13334 1 440 440 3.542 0.242 0.28234 4.14004 2 0 9 16.250 0.06154 2 0 36 8.238 0.12144 2 0 61 5.578 0.17934 2 0 220 3.537 0.28284 2 0 440 2.069 0.48344 2 9 0 60.000 0.016674 2 9 9 40.000 8.600 0.02500 0.11634 2 9 36 14.000 5.381 0.07143 0.18584 2 9 61 62.000 5.000 0.01613 0.20004 2 9 220 37.667 3.594 0.02655 0.27824 2 9 440 27.500 2.522 0.03636 0.39654 2 36 0 34.000 0.029414 2 36 9 37.100 4.222 0.02695 0.23684 2 36 36 31.222 3.276 0.03203 0.30534 2 36 61 32.917 0.030384 2 36 220 1.299 14.750 0.76991 0.06784 2 36 440 19.417 1.466 0.05150 0.68204 2 61 0 22.640 0.04417

100
Table a.3. cont.
R V WTR RTR MEANW MEANR INVW INVR
4 2 61 9 15.875 1.154 0.06299 0.86674 2 61 36 20.577 2.105 0.04860 0.47504 2 61 61 18.462 2.244 0.05417 0.44574 2 61 220 15.115 1.782 0.06616 0.56124 2 61 440 14.391 1.186 0.06949 0.84294 2 220 0 13.846 0.072224 2 220 9 10.821 0.000 0.092414 2 220 36 4.667 2.368 0.21429 0.42224 2 220 61 13.237 4.667 0.07555 0.21434 2 220 220 9.942 0.904 0.10058 1.10664 2 220 440 10.275 1.227 0.09733 0.81524 2 440 0 3.962 0.252414 2 440 9 4.071 0.000 0.24561 .
4 2 440 36 4.208 0.000 0.23765 .
4 2 440 61 4.472 0.769 0.22360 1.30004 2 440 220 4.435 2.100 0.22550 0.47624 2 440 440 3.204 0.600 0.31212 1.6667

FINAL NUMBEROFT TILLERS
181I- 4-120
101
WAVERLY VARIETY
eQ.
10e
C417:-1'
FINAL NUMBER OF SAT TIM
16141206
OWENS VARIETY6
1.
/aC
110u 1111, II ..1111
A01.10II 11111110
Iii1111.11
Figure a.l. Final mean number of wheat and ryegrass tillers plottedagainst the planted densities of wheat and ryegrass in addition series.Increasing species density and proportion decreased tillers of bothspecies. The effect of increasing wheat density and proportion was moreinfluential than increasing ryegrass density, however.

102
FINAL NUMBER OF REM TILLERS
0068
WAVERLY VARIETY
20e.
FINAL. NUMBER OF RYEGRASS MIMS OWENS VAR=
201 e128
Figure a.l. Cont.




















