
Effect of Drought on Growth Attributes of Tomato (LycopersicoN Esculentum Mill.) Genotypes

Postharvest Biology and Technology 15 (1999) 13–23
Effect of photochemical treatment in the preservation of freshtomato (Lycopersicon esculentum cv. Capello) by delaying
senescence
Rohanie Maharaj a,*, Joseph Arul a, Paul Nadeau b
a Department of Food Science and Technology (STA), Uni!ersite La!al, Ste Foy, G1K 7P4, Quebec, Canadab Station de Recherches, Agriculture and Agri-Food Canada, 2560 Boul. Hochelaga, Ste Foy, Quebec, G1V 2H4, Canada
Accepted 13 September 1998
Abstract
The effect of hormic dosage of ultraviolet radiation in delaying the senescence of tomato was investigated.Mature-green tomato fruit (var. Capello) were irradiated with ultraviolet light (UV-C, 200–280 nm) corresponding to0, UV–3.7!103 J m"2 and UV-24.4!103 J m"2, and were stored at 16°C, under high relative humidity for a periodof 35 days. Attributes of senescence such as weight loss, color, texture, respiration rate, ethylene production andputrescine were monitored periodically throughout the storage period. A dose of UV-3.7!103 J m"2 was found tobe beneficial (hormic) in delaying ripening and senescence, while the higher dose impaired ripening and causedabnormal browning, manifested as sun-scalding of the fruit’s surface. The development of color and softening oftissue were significantly retarded during storage in response to the treatment with the hormic dose (UV-3.7!103 Jm"2). In addition to a delay in the climacteric response by at least 7 days, the respiration rate and ethyleneproduction of the treated fruit were also reduced. The delay in senescence was attributed in part, to the maintenanceof a high level of putrescine (antisenescence agents exerting opposite physiological effect to ethylene). The resultssuggest that photochemical treatment may have potential for preservation of fresh fruit and vegetables. © 1999Elsevier Science B.V. All rights reserved.
Keywords: Ultraviolet radiation; Tomato; Senescence; Color; Texture; Carbon dioxide; Ethylene; Putrescine
1. Introduction
The major limiting factors in the storage offresh fruit and vegetables are senescence, fungalinfection and transpiration. Senescence is the pro-
* Corresponding author. Present address: c/o Dr C.K.Sankat, Department of Mechanical Engineering, U.W.I, St.Augustine, Trinidad, West Indies; fax: +1 868 6624414.
0925-5214/99/$ - see front matter © 1999 Elsevier Science B.V. All rights reserved.PII S0925-5214(98)00064-7

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–2314
cess where anabolic reactions are suppressed bydegradative changes leading to death and decay ofthe plant (Brady, 1987). The production of ethyleneaccelerates the biochemical and physiologicalchanges that occur during ripening leading tosenescence (Leshem et al., 1986). Under ambientconditions, tomato (a climacteric fruit) ripensrapidly before becoming excessively soft and nolonger marketable (Davies et al., 1981). Low tem-perature is effective in delaying the onset of senes-cence and decay while high humidity conditionsreduce transpiration losses. However, in the case oftomato, it is not possible to exploit low tempera-tures because of chilling injury (Morris, 1982).
A postharvest treatment which can be adjunct torefrigeration for preserving fruit and vegetables isthe use of non-ionizing, artificial ultraviolet (UV-C)radiation. Literature on artificial UV-C is primarilyrelated to its germicidal activity in the range of200–280 nm (with peak emission at 254 nm) as wellas to bacterial mutation (Kleczkowski, 1968; WorldHealth Organization, 1979). UV-C radiation ismainly used as a surface treatment because itpenetrates only 50–300 nm into the tissue (Jagger,1965). Studies have shown that UV-C elicits phy-toalexins, compounds important in disease resis-tance in many systems: including resveratol ingrape vines (Langcake and Pryce, 1977) and hy-droxyphaseollin in soybean hypocotyls (Bridge andKlarman, 1973). Induced resistance was also ob-served with walla walla onions (Lu et al., 1987),potato tuber slices (Cheema and Haard, 1978),sweet potatoes (Stevens et al., 1990) and carrotslices (Mercier et al., 1993a,b). Exposure of horti-cultural crops to UV-C light has thus been consid-ered as a possible alternative to chemical fungicidesfor the control of postharvest diseases. UV-Cradiation has also been reported to delay ripeningof climacteric fruit such as apples and peaches (Luet al., 1991), but the biological mechanisms bywhich such changes take place have not beendescribed.
DNA is one of the most important targetmolecules for UV-C effects (Mitchell and Clarkson,1984). When DNA absorbs UV radiation, a num-ber of photochemical lesions result, e.g. the dimer-ization of thymine, with a resulting loss inbiological activity of DNA (Caldwell, 1981). UV
light can also be involved in photooxidation reac-tions in plants via free radical production withdeleterious effects. Free radicals and superoxidescan target cell membranes, nucleic acids, cell wallsand enzymes, resulting in the acceleration of senes-cence (Kon and Schwimmer, 1977; Leshem et al.,1986).
Changes in polyamine (PA) biosynthesis in planttissues have been correlated with various stressessuch as K+ deficiency (Smith, 1963), cold acclima-tization (Nadeau et al., 1987), chemical stress(Bertrand et al., 1990) and controlled atmospheres(Kramer et al., 1989). Polyamines are apparentlyubiquitous, but are present in relatively low concen-trations (10 !M–10 mM) in mature plant tissues(Slocom et al., 1984). Multifunctional PAs areantisenescence agents, exerting an opposite physio-logical effect to ethylene while sharing a commonprecursor S-adenosylmethionine (SAM) (Galstonand Kaur-Sawhney, 1990). The biochemical path-way from methionine to ethylene involves SAMand 1-aminocyclopropane-1-carboxylic acid(ACC). SAM is a substrate for ACC synthase in thesynthesis of ethylene via ACC (Adams and Yang,1977). SAM is also a substrate for SAM decarboxy-lase in a pathway which leads to the synthesis ofPAs (Galston and Kaur-Sawhney, 1990).Polyamines have shown inhibitory effects onethylene synthesis in a variety of plant tissues(Apelbaum et al., 1981; Dibble et al., 1988; Pon-nampalam et al., 1993). Polyamines inhibit ethylenebiosynthesis probably by inhibiting the activities ofACC synthase and ACC as well as suppressing denovo synthesis of ACC synthase (Apelbaum et al.,1982). The antisenescent activity of PAs may alsobe related to their ability to be effective free radicalscavengers (Drolet et al., 1986) as well as stabilizingDNA and membranes because of their cationicnature (Slocom et al., 1984). Although there isconsiderable literature on the effects of polyaminesin plant growth and development, there is very littleinformation on their mode of action at themolecular level. The work of Li et al. (1992) usingtomato have demonstrated that wounding thefruit resulted in the accumulation of an ACCsynthase transcript, and the induction of thistranscript andACC synthase activity were blockedin the presence of polyamines. The resultsfurther indicate that polyamines may specifically

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–23 15
regulate ethylene biosynthesis at the level of ACCsynthase transcript accumulation and thus influ-ence ripening.
UV-C is generally harmful but can producebeneficial effects on horticultural crops at lowdoses, a phenomenon known as hormesis (stimu-lation of beneficial responses by low levels ofstressors which are otherwise detrimental); (Cal-abrese et al., 1987). The hormic (beneficial) effectdecreases with age of the plant and in the case ofcertain crops, they are more susceptible to reduc-tions in disease resistance as they mature (Mercieret al., 1993b). Plants are equipped with bothnon-enzymic antioxidants (e.g. carotenoids andphenols) as well as enzymic scavengers (superox-ide disutase) in response to the damaging effectsof free radicals (Wellman, 1976; Leshem et al.,1986). One can hypothesize that these antioxidantcompounds including polyamines, which are in-duced by various stresses, can also be induced byUV-C irradiation. Stimulation of such com-pounds (depending on the stress level) in youngfruit, can probably counteract the effect of DNAand free radical damage which becomes importantwith age. Such mechanisms can lead to a delay insenescence and help maintain longevity of har-vested crops.
The objectives of this study were to: (1) evalu-ate the quality of irradiated fruit by measuringcertain senescence parameters such as weight loss,color and texture periodically throughout storage,when fruit were stored at 16°C, under high rela-tive humidity (95%) and for a maximum period of35 days; and (2) gain some understanding of thephysiological basis of delayed senescence.
2. Materials and methods
2.1. Plant material
Mature-green tomatoes (Lycopersicon esculen-tum Mill cv. Capello) were harvested manuallyfrom plants grown in a greenhouse in Quebecduring the summer. Fruit of uniform shape andsize and free from fungal infection were selected.After harvest, fruit were washed in tap water,air-dried at 25°C using a fan and individuallylabelled and weighed.
2.2. Irradiation and storage conditions
UV-C radiation was provided by fluorescentgermicidal lamps (GE 30 W) with a peak emissionat 254 nm. Irradiation was carried out on thesame day of harvest and under ambient condi-tions. Radiation doses were varied according tothe intensity of radiation (20.3 W m"2), as mea-sured by a portable digital radiometer (UVX Dig-ital Radiometer UVP Inc. San Gabriel, CA).Tomato were placed at approximately 30 cm fromthe lamps in a UV chamber and rotated so thattheir blossom and calyx ends faced the lamp toensure uniform irradiation. After irradiation, fruitwere placed in plastic humidified (about 95% RH)containers (5-l capacity and four fruit per con-tainer) and stored at 16°C in the dark for amaximum of 35 days.
Three separate experiments were conducted ontomato to establish: (1) general senescenceparameters, (2) carbon dioxide (CO2) andethylene (C2H4) changes and (3) putrescine(polyamine) levels. Irradiated fruit correspondingto 0, UV-3.7!103 and UV-24.4!103 J m"2 wereused. In the first experiment, a total of 108 fruitwere analyzed with 36 fruit per treatment (four–six replicates) and destructive quality evaluationswere carried out at weekly intervals over a periodof 35 days. In the second experiment, non-de-structive CO2 and C2H4 analyses were made dailyusing six fruit (placed in individual 500-ml plasticcontainers) per treatment for up to 30 days. In thethird experiment, destructive analyses of pu-trescine were carried out at weekly intervals overa period of 21 days, using 12 fruit per treatment(three replicates).
2.3. Measurement of senescence parameters
The loss in weight of individual fruit was deter-mined at weekly intervals as a percentage of theoriginal weight. A mean of four fruit was used foreach sampling period and for each irradiationtreatment. Fruit color was monitored using arating scale from 1 to 6, with: 1, mature green; 2,breaker; 3, turning; 4, pink; 5, light red; and 6, red(Big Red Color Chart, FL, USA). A mean of sixfruit was used for each sampling period and for

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–2316
each irradiation treatment. Surface colors oftomato were also objectively determined using acolorimeter (Gardner Colorgard 1000/05; PacificScientific, Silver Spring, MD) and Hunter ‘a’ val-ues ("green to +red) were recorded for eachfruit (Clydesdale, 1978). A mean of four fruit wasused for each sampling period and for each irradi-ation treatment. The force required to puncturethe skin of whole fruit (rupture of skin followedby deformation) was determined from a modifiedmethod of Holt (1970), using the Instron Univer-sal Testing Instrument (Model TMS, InstronCanada Ltd; Burlington, Ont.). A cylindricalplunger with a flat end (1 mm dia.) was attachedto the crosshead and driven at a constant speed of5 cm min"1, in a vertical direction into thetomato placed on a flat aluminium plate. Threedeterminations were made on each fruit awayfrom the locular ridge. The force transducer andstrip chart recorder were adjusted to provide fullscale sensitivity of 9.8 N. Calibration was per-formed before each set of measurements and thefirmness was expressed as peak force and recordedin Newtons. Four fruit were used for each sam-pling period and for each irradiation treatment.
2.4. Carbon dioxide and ethylene determination
The respiration rate was determined by measur-ing the head space carbon dioxide concentrationof individual fruit in gas-tight, 500-ml plastic con-tainers fitted with sampling ports. A 1.0 ml gassample was withdrawn within 1 h of containerclosure. The container was flushed with air afterremoving the gas sample. Analyses were per-formed on a gas chromatograph (GC–PerkinElmer, Model 8500; Mississauga, Ont.) with athermal conductivity detector maintained at 60°C.The carrier gas was helium at a flow rate of 30 mlmin"1 and column temperature was maintainedat 35°C. The respiration rate was calculated tak-ing into account the weight of the fruit, residualvolume of the container and the incubation time(1 h). CO2 evolution rates were expressed as mlkg"1 h"1. Ethylene (C2H4) was measured byplacing individual fruit in sealed containers andwithdrawing 1.0 ml gas samples within 1 h of
closure with a Hamilton gas tight syringe. Thecontainer was flushed with air after removing thegas sample. Analyses were performed on a gaschromatograph fitted with a flame ionization de-tector (FID) and 2 m!4 mm stainless steelcolumn packed with 80–100 mesh Poropak-Q.The temperature of the column was maintained at100°C, and helium was used as the carrier gaswith a flow rate of 35 ml min"1. Nitrogen, hydro-gen and oxygen flow rates were 35, 30 and 300 mlmin"1 respectively. Ethylene identification wasbased on the retention time compared to C2H4
purity standards (99.9%). C2H4 evolution rateswere expressed as ! l kg"1 h"1.
2.5. Putrescine determination
Endogenous soluble free putrescine was ex-tracted from tomato pericarp (exocarp) of individ-ual fruit with 5% perchloric acid (w/v),benzoylated and separated by HPLC on a re-versed phase C18 Radial Pak column (Nova-Pak,8 mm!100 mm; Waters Assoc.) using themethod of Slocom et al. (1989). The mobile phaseconsisted of methanol:water as solvent with a flowrate of 2.0 ml min"1 in a gradient program asfollows: running gradient, initial conditions 50%MeOH for min 0–10, 50–70% MeOH for min10–15, 70–85% MeOH for min 15–22, 85%MeOH for min 22–25, 85–50% for min 27 andreturn to equilibration during min 27–33.Polyamines were separated in 33 min using thiselution gradient system and spectrometric detec-tion was at 254 nm. Quantification of putrescinelevels was made by comparing the results withthose of benzoylated standards. Putrescine evolu-tion was expressed on a fresh weight basis withunits of !mol kg"1. Three fruit were used foreach sampling period and for each irradiationtreatment.
Experiments were laid out in a completely ran-domized factorial design (storage time! treat-ments) and analysis of variance was performedwith the General Linear Model procedure of theStatistical Analysis Systems (SAS Institute, 1990).Significant differences were established at 5, 1 and0.1% levels of significance.

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–23 17
3. Results and discussion
3.1. General senescence parameters
With increasing storage time, both control andirradiated fruit lost weight almost linearly withtime (p!0.001) and averaged a loss of 0.16 (r2=0.999), 0.19 (r2=0.997) and 0.21 (r2=0.998) %day"1 respectively for control, UV-3.7!103 and24.4!103 J m"2 fruit. Weight loss was also afunction of the dose used (p!0.05) and presentedmaximum values of 5.5, 6.7 and 7.2% for control,UV-3.7!103 and UV-24.4!103 J m"2 fruit re-spectively (Table 1). Control fruit showed leastweight losses.
Initially all fruit were mature green (rating 1) incolor. However, as storage progressed, controlfruit lost their green color faster than UV-treatedfruit and after 4 weeks in storage average colorrating values were 6.0, 5.0 and 4.0 for control,UV-3.7!103 and 24.4!103 J m"2 fruit respec-tively. Color changes in fruit as measured by the
Hunter tristimulus ‘a’ values were significantlyaffected by irradiation treatment (p!0.05) andstorage time (p!0.001). Highly negative ‘a’ valuedenotes a high degree of greenness of the fruit.For the fresh tomato (day 0), the ‘a’ value aver-aged "9.4 as shown in Table 1. After 28 days instorage, ‘a’ values averaged 33.7, 19.3 and 17.8 fordoses 0, UV-3.7!103 and 24.4!103 J m"2 re-spectively indicating a significant loss in greencolor. Control fruit developed their red colorfaster than the UV-3.7!103 and UV-24.4!103 Jm"2 irradiated fruit. However, UV-24.4!103 Jm"2 fruit showed increased surface browning af-ter 21 days in storage and did not ripen normally.The delay in color development was more evidentfor the UV-3.7!103 J m"2 (optimal dose) fruitas compared to the UV-24.4!103 J m"2 irradi-ated fruit. Although the color development andripening of tomato fruit treated with 3.7!103 Jm"2 (optimal dose) was delayed, they ripenednormally toward the end of the storage period.
Table 1Quality attributes of control and UV-C irradiated tomato stored at 16°C
Storage time (days) Control UV-3.7!103 J m"2 UV-24.4!103 J m"2
Weight Loss (%)0.0"0.00 0.0"0.0 0.0"0.0
7 1.0"0.0 1.1"0.0 1.2"0.014 2.3"0.1 2.7"0.1 2.9"0.2
4.3"0.33.9"0.221 3.5"0.14.5"0.2 5.5"0.328 5.0"0.2
7.2"0.35.6"0.2 6.6"0.235
Tristimulus Hunter ‘a’ !alues0 "9.4"0.0 "9.4"0.0 "9.4"0.0
"6.4"0.07 "8.5"0.0 "6.4"0.014 "1.1"0.0"3.4"0.06.4"0.0
2.4"0.6 14.2"0.615.7"0.42128 33.7"1.0 19.3"1.3 17.8"1.1
22.1"0.835.4"0.5 24.7"0.135
Texture measured by peak puncture force (N)39.3"1.00 39.1"2.0 39.3"1.032.0"2.07 39.6"1.0 30.0"1.224.0"1.0 30.3"2.027.3"0.21414.2"1.121 19.5"0.7 18.9"0.6
28 9.9"1.8 15.6"0.3 17.1"1.235 9.7"1.3 11.4"1.1 13.3"0.5
Standard errors of the mean for n=4 are indicated.

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–2318
Storage time (p!0.001) and irradiation doses(p!0.001) significantly affected fruit firmness.With increasing storage, there was a loss in firm-ness as shown in Table 1. Control fruit exhibitedincreased softening with storage when comparedto UV-treated fruit. After 28 days of storage, theaverage texture of control, UV-3.7!103 and24.4!103 J m"2 fruit as measured by their peakdeformation/puncture forces were 9.9, 15.6 and17.1 N respectively. The UV-3.7!103 and 24.4!103 J m"2 irradiated fruit were firmer than con-trol fruit (Table 1).
The softening of tissues during ripening resultsin part from progressive changes in cell wall com-position as well as cell wall separation. Cell wallseparation is brought about by the solubilizationof the pectin-rich lamella (Brady, 1987). A directcorrelation was found between the rapid increasein cell wall depolymerizing enzymes and the cli-macteric rise in respiration and ethylene produc-tion in avocado (Awad and Young, 1979). Themost important softening enzyme in tomato ispolygalacturonase. This enzyme has been shownto increase in activity with ripening and senes-cence of tomato (Grierson and Kader, 1986). It isunlikely that the differential dehydration (asreflected by weight loss) shown by the three treat-ments had any effect on the texture. During weeks4 and 5, weight losses in the range of 5–7.2% werenoted and such a range is normally quite accept-able for fresh produce. Texture changes as ob-served in Table 1 would be more the result ofphysiological ripening as previously noted ratherthan any losses in moisture and subsequentdehydration.
The softening of control fruit could also berelated to the production of free radicals as aresult of senescence (Leshem et al., 1986) and itseffect on the cell wall. By breaking hydrogenbonds, free radicals are presumably responsiblefor structural modifications in cell wall polysac-charides (Kon and Schwimmer, 1977). The in-creased firmness of UV-treated fruit whencompared to the control could be associated withincreased levels of PAs. Kramer et al. (1989)reported that exogenous PAs effectively sup-pressed cell wall softening and activity of poly-galacturonase. The PAs were assumed to function
via a mechanism similar to that of calcium, in-volving the formation of cation cross-links withpectic acid and other polysaccharides, thus limit-ing accessibility of the cell wall to degradativeenzymes.
Furthermore, Knee and Bartley (1981) pro-posed that the loss in cohesion of cell walls mayresult from S-adenosylmethionine (SAM) methy-lation of the free carboxylic groups in the pectinmaterial and the disruption of calcium cross link-ages of adjacent polyuronides. The negativelycharged carboxyl groups of uronic acids in pec-tinic material are normally available for cross-linkages through multivalent cations, mainlycalcium, one of the mechanisms proposed for cellwall cohesion. Since SAM was probably routed topolyamine synthesis, it was not readily availablefor disruption of calcium bridges that could leadto increased firmness of the UV-treated fruit.
3.2. Carbon dioxide, ethylene and putrescineproduction
In order to gain some understanding of thephenomenon of delayed senescence in UV-treatedfruit, the respiration rate, ethylene production ofthe whole fruit as well as the putrescine levels inthe pericarp tissue were examined. Controltomato exhibited a climacteric rise in CO2 (Fig. 1)and C2H4 (Fig. 2) production while irradiatedfruit exhibited a significantly (p!0.001) reducedrespiratory and ethylene climacteric activity.There were however, initial transient rises in CO2
and C2H4 production following irradiation whichwere proportional to the dose used. Initial CO2
production for untreated fruit was 5.8 ml kg"1
h"1 compared to 7.7 and 9.3 ml kg"1 h"1 for theUV-3.7!103 and 24.4!103 J m"2 irradiatedfruit respectively. This is presumably a stress re-sponse to the radiation as has been reported forgamma-irradiated fruit (Frylinck et al., 1987). Byday 10, CO2 production peaked to 11.7 ml kg"1
h"1 for untreated fruit while for irradiated fruitCO2 production peaked to 10.2 and 9.2 ml kg"1
h"1 at days 17 and 18 for the UV-3.7!103 and24.4!103 J m"2 fruit respectively. Overall CO2
production rate increased 2-fold from pre-climac-teric phase for untreated fruit. Additionally, there
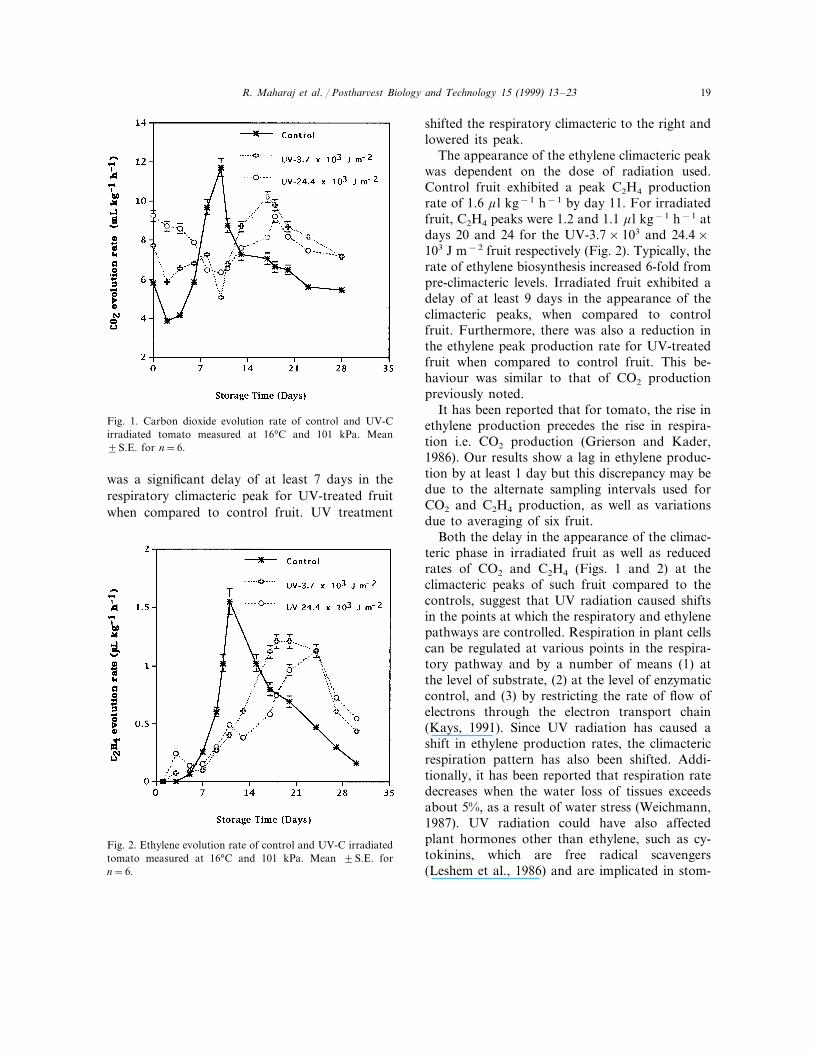
R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–23 19
Fig. 1. Carbon dioxide evolution rate of control and UV-Cirradiated tomato measured at 16°C and 101 kPa. Mean"S.E. for n=6.
shifted the respiratory climacteric to the right andlowered its peak.
The appearance of the ethylene climacteric peakwas dependent on the dose of radiation used.Control fruit exhibited a peak C2H4 productionrate of 1.6 ! l kg"1 h"1 by day 11. For irradiatedfruit, C2H4 peaks were 1.2 and 1.1 ! l kg"1 h"1 atdays 20 and 24 for the UV-3.7!103 and 24.4!103 J m"2 fruit respectively (Fig. 2). Typically, therate of ethylene biosynthesis increased 6-fold frompre-climacteric levels. Irradiated fruit exhibited adelay of at least 9 days in the appearance of theclimacteric peaks, when compared to controlfruit. Furthermore, there was also a reduction inthe ethylene peak production rate for UV-treatedfruit when compared to control fruit. This be-haviour was similar to that of CO2 productionpreviously noted.
It has been reported that for tomato, the rise inethylene production precedes the rise in respira-tion i.e. CO2 production (Grierson and Kader,1986). Our results show a lag in ethylene produc-tion by at least 1 day but this discrepancy may bedue to the alternate sampling intervals used forCO2 and C2H4 production, as well as variationsdue to averaging of six fruit.
Both the delay in the appearance of the climac-teric phase in irradiated fruit as well as reducedrates of CO2 and C2H4 (Figs. 1 and 2) at theclimacteric peaks of such fruit compared to thecontrols, suggest that UV radiation caused shiftsin the points at which the respiratory and ethylenepathways are controlled. Respiration in plant cellscan be regulated at various points in the respira-tory pathway and by a number of means (1) atthe level of substrate, (2) at the level of enzymaticcontrol, and (3) by restricting the rate of flow ofelectrons through the electron transport chain(Kays, 1991). Since UV radiation has caused ashift in ethylene production rates, the climactericrespiration pattern has also been shifted. Addi-tionally, it has been reported that respiration ratedecreases when the water loss of tissues exceedsabout 5%, as a result of water stress (Weichmann,1987). UV radiation could have also affectedplant hormones other than ethylene, such as cy-tokinins, which are free radical scavengers(Leshem et al., 1986) and are implicated in stom-
was a significant delay of at least 7 days in therespiratory climacteric peak for UV-treated fruitwhen compared to control fruit. UV treatment
Fig. 2. Ethylene evolution rate of control and UV-C irradiatedtomato measured at 16°C and 101 kPa. Mean "S.E. forn=6.

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–2320
atal opening (Weichmann, 1987). Since UV-treated fruit exhibited higher weight losses whencompared to control fruit, the involvement ofcytokinins could account for the delayed senes-cence associated with UV-treated fruit.
The shift in ethylene peak production associ-ated with UV-treated fruit suggests that SAM(ethylene precursor) was probably a limiting fac-tor with UV-treated fruit. At the higher dose used(UV-24.4!103 J m"2), UV could have alsocaused irreversible membrane deterioration, whichcould have affected the activity of ACC oxidase(loosely bound to membrane), and thus explainthe reduced ethylene production for such fruit.Similar results were reported for high gamma-irradiated cherry tomato fruit (Larrigaudiere etal., 1991). Ethylene production pattern (totalethylene) was similar at both doses of UV radia-tion, although it was reduced at the higher dose.
Polyamines are of interest because they candelay senescence in plant tissue (Galston andKaur-Sawhney, 1990), and perform multiple func-tions: stabilize DNA and cell membranes, scav-enge free radicals, suppress ethylene productionand inhibit RNAse and proteases (Drolet et al.,1986; Galston and Kaur-Sawhney, 1990). Therewas a significant difference in the free putrescinelevels among treatments and among storage times,as well as a significant time/treatment interaction(p!0.001). Generally, UV-treated fruit main-tained higher putrescine levels compared to thecontrol fruit throughout the storage period (Fig.3). In both cases, only toward the post-climactericphase was there a significant decrease in freeputrescine levels, but this was markedly greater incontrol fruit than treated fruit. By day 21, controlfruit had strikingly reduced putrescine levels (39!mole kg"1) compared to 452 and 293 !molekg"1 for the UV-3.7!103 and 24.4!103 J m"2
fruit respectively.The ethylene-forming enzyme (EFE) ACC oxi-
dase has been shown to be a soluble enzyme(Ververidis and John, 1991) although in vivo itmay be loosely bound to a membrane (Amrhein etal., 1987). The anti-senescent activity ofpolyamines is generally attributed to their abilityto stabilize and protect membranes by associatingwith negatively charged phospholipids (Galston
Fig. 3. Variation in levels of free putrescine in pericarp ofcontrol and UV-C irradiated tomato during storage at 16°C.Mean "S.E. for n=4.
and Kaur-Sawhney, 1990). However, the suppres-sion of ethylene production by polyamines ap-pears to be a result of interference with the levelsof ACC synthase and ACC (Apelbaum et al.,1982) rather than with the activity of the EFE(Hyodo and Tanaka, 1986). Additionally the con-version from ACC to ethylene is a superoxide-mediated reaction (Legge et al., 1982). Polyaminesmay inhibit conversion of ACC to ethylene byreducing synthesis of ACC synthase and scaveng-ing oxygen free radicals involved in catalytic con-version of ACC to ethylene. As plants age,putrescine is no longer synthesized, making moreSAM available as a precursor for ACC. Moresuperoxide is available because free-radical scav-enging is probably reduced in the absence ofputrescine. Consequently, ACC conversion toethylene can progress (Pell and Dann, 1991).
Rastogi and Davies (1991), reported that thehigh putrescine levels in Alcobaca tomato wererelated to the slow ripening and thus prolongedkeeping qualities. Additionally Kramer et al.(1989) have shown that PAs inhibit softening byreducing the activity of cell wall degrading en-zymes such as polygalacturonase. It is known that

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–23 21
endogenous levels of PAs decrease with age inboth plants and animals (Slocom et al., 1984), butUV treatment contributed to the maintenance ofhigher levels of putrescine even after the respira-tory climacteric phase. Correlated with this eventwas a retardation in the appearance of theethylene and respiratory climacteric peaks,ethylene production was suppressed, softening ofthe tissues was reduced and a delay in ripeningand senescence of tomato was observed.
4. Conclusion
UV-C irradiation of tomato with a hormic doseof UV-3.7!103 J m"2 together with moderatetemperature (16°C) and high humidity storage(95%) significantly retarded the progress of senes-cence in tomato fruit. As a result, color develop-ment as well as softening of fruit were delayedduring the 35 day storage period. Additionally,the climacteric peaks of respiration and ethyleneproduction were delayed by 7 days for UV-3.7!103 J m"2 and 9 days for UV-24.4!103 J m"2
irradiated fruit. One possible mode of UV actionmaybe attributable to the maintenance of higherlevels of putrescine.
An abnormal superficial browning was associ-ated with the high dose of UV-24.4!103 J m"2.Surface color of tomato is the most importantcriterion of quality from a consumers’ point ofview and thus, this high dose is not recommendedfor improving the postharvest storage life oftomato. The UV-3.7!103 J m"2 treatment whichcorresponded to very high putrescine levels couldbe used as a hormic dose for delaying senescence.Our results suggest that photochemical treatment(UV-C-irradiation) appears to be a promising newtechnique for improving the postharvest storagelife of climacteric tomato and possibly other fruitand vegetables.
Acknowledgements
The authors are grateful to the Department ofFood Science and Technology (STA), UniversiteLaval, and Station de Recherches, Agriculture
and Agri-Food Canada, Ste Foy for providingmuch additional support. Special thanks to Do-minique Roussel, Ronan Corcuff (STA) andLucette Chouinard (Agriculture Canada) for theirtechnical assistance.
References
Adams, D.O., Yang, S.F., 1977. Methionine metabolism inapple tissue. Implication of S-adenosylmethionine as anintermediate in the conversion of methionine to ethylene.Plant Physiol. 60, 892–896.
Amrhein, N., Forreiter, C., Kionka, C., Skorupka, H.,Tophof, S., 1987. Metabolism, and its compartmentationof 1-aminocyclopropane-1-carboxylic acid in plant cells.In: Scheiber, K., Schutte, H.R., Sembnder, G. (Eds.),Proceedings of the International Symposium on Conju-gated Plant Hormones—Structures, Metabolism andFunction. Deutscher Verlag der Wissenschaften, Berlin,pp. 102–109.
Apelbaum, A., Burgoon, A.C., Anderson, J.D., Lieberman,M., Ben-Arie, R., Mattoo, A.K., 1981. Polyamines inhibitbiosynthesis of ethylene in higher plant tissue and fruitprotoplasts. Plant Physiol. 68, 453–456.
Apelbaum, A., Icekson, R., Burgoon, A.C., Lieberman, M.,1982. Inhibition by polyamines of macromolecular synthe-sis and its implications for ethylene production and senes-cence processes. Plant Physiol. 70, 1221–1223.
Awad, M., Young, R.E., 1979. Postharvest variation in cellu-lase, polygalacturonase and pectinmethylesterase in ava-cado (Persea americana Mill cv. Fuerte) fruit in relation torespiration and ethylene production. Plant Physiol. 64,306–308.
Bertrand, S., Nadeau, P., Dostaler, D., Gosselin, A., 1990.Variations in the polyamine content of control and stressedtomato leaves: induction by chemical stress. In: Flores,H.E., Arteca, R.N., Shannon, J.C. (Eds.), Polyamines andEthylene: Biochemistry, Physiology and Interactions.American Society of Plant Physiology, Rockville, MD, pp.410–414.
Brady, C.J., 1987. Fruit ripening. Annu. Rev. Plant Physiol.38, 155–178.
Bridge, M.A., Klarman, W.L., 1973. Soybean phytoalexin,hydroxy-phaseolin, induced by ultraviolet irradiation. Phy-topathology 63, 606–609.
Calabrese, E.J., McCarthy, M.E., Kenyon, E., 1987. Theoccurrence of chemically induced hormesis. Health Phys.52, 531–541.
Caldwell, M.M., 1981. Plant response to solar ultravioletradiation. In: Lange, O.L., Nobel, P.S., Osmond, C.B.,Ziegler, H. (Eds.), Physiological Plant Ecology 1. Springer,New York, pp. 170–194.
Cheema, A.S., Haard, N.F., 1978. Induction of rishitin andlubimin in potato tuber discs by non-specific elicitors and

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–2322
the influence of storage conditions. Physiol. Plant Pathol.13, 233–240.
Clydesdale, F.M., 1978. Colorimetry—methodology and ap-plications. CRC Crit. Rev. Food Sci. Nutr. 243–262.
Davies, J.N., Hobson, G.E., McGlasson, W.B., 1981. Theconstituents of tomato fruit—the influence of environ-ment, nutrition and genotype. CRC Crit. Rev. Food Sci.Nutr. 15, 205–280.
Dibble, A.R.G., Davies, P.J., Mutschler, M.A., 1988.Polyamine content of long-keeping Alcobaca tomato fruit.Plant Physiol. 86, 338–340.
Drolet, G., Dumbroff, E.B., Legge, R.L., Thompson, J.E.,1986. Free radical scavenging properties of polyamines.Phytochemistry 25, 367–371.
Frylinck, L., Dubery, I.A., Schabort, J.C., 1987. Biochemicalchanges involved in stress response and ripening behaviourof gamma irradiated mango fruit. Phytochemistry 26, 681–686.
Galston, A.W., Kaur-Sawhney, R., 1990. Polyamines in plantphysiology. Plant Physiol. 94, 406–410.
Grierson, D., Kader, A.A., 1986. Fruit ripening and quality.In: Atherton, J.G., Rudich, J. (Eds.), The Tomato Crop—A Scientific Basis for Improvement. Chapman and Hall,London, pp. 241–280.
Holt, C.B., 1970. Measurement of tomato firmness with auniversal testing machine. Texture Stud. 1, 491–501.
Hyodo, H., Tanaka, K., 1986. Inhibition of 1-aminocyclo-propane-1-carboxylic acid synthase activity by polyamine,their related compounds and metabolites of S-adenosylme-thionine. Plant Cell Physiol. 27, 391–398.
Jagger, J., 1965. Photoprotection from far ultraviolet effects incells. In: Duchesne, J. (Ed.), Advances in ChemicalPhysics, vol. VII, The Structure and Properties ofBiomolecules in Biological Systems. Interscience, NewYork, pp. 548–601.
Kays, S.J., 1991. Postharvest Physiology of Perishable PlantProducts. AVI/Van Nostrand Reinhold, New York.
Kleczkowski, A., 1968. Methods of inactivation by UV radia-tion. Methods Virol. 4, 93–138.
Knee, M., Bartley, I.M., 1981. In: Friend, J., Rhodes, M.J.C.(Eds.), Recent Advances in the Biochemistry of Fruit andVegetables. Academic Press, New York, pp. 131–148.
Kon, S., Schwimmer, S., 1977. Depolymerization of polysac-charides by active oxygen species derived from a xanthineoxidase system. J. Food Biochem. 1, 141–144.
Kramer, G.F., Wang, C.Y., Conway, W.S., 1989. Correlationof reduced softening and increased polyamine levels duringlow-oxygen storage of ‘McIntosh’ apples. J. Am. Soc.Hort. Sci. 114, 942–946.
Langcake, P., Pryce, P., 1977. The production of resveratroland viniferins by grapevines in response to ultravioletirradiation. Phytochemistry 16, 1193–1196.
Larrigaudiere, C., Latche, A., Pech, J.C., Triantaphylides, C.,1991. Relationship between stress ethylene production in-duced by gamma irradiation and ripening of cherry toma-toes. J. Am. Soc. Hort. Sci. 116, 1000–1003.
Legge, R.L., Thomas, J.E., Baker, J.E., Lieberman, M., 1982.The effect of calcium on the fluidity and phase propertiesof microsomal membranes isolated from postclimactericgolden delicious apples. Plant Cell Physiol. 23, 161–169.
Leshem, Y.Y., Halevy, A.H., Frenkel, C., 1986. Processes andControl of Plant Senescence. Elsevier, Amsterdam.
Li, N., Parsons, B.L., Liu, D., Mattoo, A.K., 1992. Accumula-tion of wound-inducible ACC synthase transcript intomato fruit is inhibited by salicylic acid and polyamines.Plant Mol Biol. 18, 477–487.
Lu, Y.J., Stevens, C., Yakubu, P., Loretan, P.A., Eakin, D.,1987. Gamma, electronbeam and ultraviolet radiation onthe control of storage rots and quality of walla wallaonions. J. Food Proc. Pres. 12, 53–62.
Lu, Y.J., Stevens, C., Khan, V.A., Kabwe, M., 1991. Theeffect of ultraviolet irradiation on shelf-life and ripening ofpeaches and apples. Food Qual. 14, 299–305.
Mercier, J., Arul, J., Ponnampalam, R., Boulet, M., 1993a.Induction of 6-methoxymellin and resistance to storagepathogens in carrot slices by UV-C. Phytopathology 137,44–54.
Mercier, J., Arul, J., Julien, C., 1993b. Effect of UV-C onphytoalexin accumulation and resistance to Botrytis cinereain stored carrots. Phytopathology 139, 17–25.
Mitchell, D.L., Clarkson, J.M., 1984. Induction of photoprod-ucts in synthetic polynucleotides by far and near ultravioletradiation. Photochem. Photobiol. 40, 735–741.
Morris, L.L., 1982. Chilling injury of horticultural crops: anoverview. Hortscience 17, 161–162.
Nadeau, P., Delaney, S., Chouinard, L., 1987. Effects of coldhardening on the regulation of polyamine levels in wheat(Triticum aesti!um L.) and alfalfa (Medicago sati!a L.).Plant Physiol. 84, 73–77.
Pell, E.J., Dann, M.S., 1991. Multiple stress-induced foliarsenescence and implications for whole-plant longevity. In:Mooney, H.A., Winner, W.E., Pell, E.J., Chu, E. (Eds.),Response of Plants to Multiple Stresses. Academic Press,New York, pp. 189–204.
Ponnampalam, R., Arul, J., Makhlouf, J., Castaigne, F., 1993.Inhibition of 1-aminocyclopropane-1-carboxylic acid-de-pendent ethylene production in tomato-tissue discs by bio-chemical treatments. Postharvest Biol. Technol. 2,291–300.
Rastogi, R., Davies, P.J., 1991. Polyamine metabolism inripening tomato fruit. II—Polyamine metabolism and syn-thesis in relation to enhanced putrescine content and stor-age life of alc tomato fruit. Plant Physiol. 95, 41–45.
SAS Institute, 1990. SAS User’s Guide: Basics, Version 5 ed.SAS Institute, Cary, NC.
Slocom, R.D., Kaur-Sawnhey, R., Galston, A.W., 1984. Thephysiology and biochemistry of polyamines in plants.Arch. Biochem. Biophys. 225, 283–303.
Slocom, R.D., Flores, H.E., Galston, A.W., Weinstein, L.H.,1989. Improved method for HPLC analysis of polyamines,agmatine and aromatic monoamines in plant tissue. PlantPhysiol. 89, 512–517.

R. Maharaj et al. / Posthar!est Biology and Technology 15 (1999) 13–23 23
Smith, T.A., 1963. L-Arginine carboxylase of higher plants andits relation to potassium nutrition. Phytochemistry 2, 241–243.
Stevens, C., Khan, Y.A., Tang, A.Y., Lu, Y.J., 1990. Theeffect of ultraviolet radiation on mold rots and nutrients ofstored sweet potatoes. J. Food Protect. 53, 223–226.
Ververidis, P., John, P., 1991. Complete recovery in vitro ofethylene forming enzyme activity. Phytochemistry 30, 725–
727.Wellman, E., 1976. Specific ultraviolet effects in plant morpho-
genesis. Yearly Rev. Photochem. Photobiol. 24, 659–660.Weichmann, J., 1987. Postharvest Physiology of Vegetables.
Marcel Dekker, New York.World Health Organization, 1979. Ultraviolet Radiation. En-
vironmental Health Criteria, vol. 14. WHO, Geneva, pp.1–20.
..
Copyright © 2022 FDOKUMEN




















