
ecowgy and management of chromolaena odorata - horizon ird
182
ECOWGY AND MANAGEMENT OF CHROMOLAENA ODORATA BIOTROP SPECIAL PUBLICAT ION NO . 44 ISS!\· 01 25 - 975 X and SEAM EO BIOTROP Southeast Asian Regional Centre for Tropical Biology P.O. Box 17, Bogor , Indonesia 199\ [g BIOTROP
-
Upload
khangminh22 -
Category
Documents
-
view
4 -
download
0
Transcript of ecowgy and management of chromolaena odorata - horizon ird

ECOWGY AND MANAGEMENT OFCHROMOLAENA ODORATA
BIOTROP SPECIAL PUBLICATION NO. 44
ISS!\· 01 25 - 975 X
and
SEAMEO BIOTROP
Southeast Asian Regional Centre for T ropical Biology
P.O . Box 17, Bogor , Indonesia
199\
[gBIOTROP


PROCEEDINGS OFTHE SECOND INTERNATIONALWORKSHOPON BIOLOGICAL CONTROL OF CHROMOLAENA ODORATA
Bogor, Indonesia, 4 - 8 February 1991
edited byR. Muniappan
Agricultural Experiment StationUniversity of Guam .
Mangilao, Guam, USAand
Paul FerrarAustralian Centre for International Agricultural Research
Canberra, Australia
BIOTROP SPECIAL PUBLICATION NO. 44
ISSN 0125 - 975 X
Published by
CFSD1and
SEAMEO BIOTROPSoutheast Asian Regional Centre for Tropical Biology
P.O. Box 17. Bogor, Indonesia1991
[DIIO'ROP


TABLE OFCONTENTS
Preface
Welcome Address - [oko Purwanto
Opening Address - Cheng-Hwa Huang
Opening Address - Paul Ferrar
Review of the First International Workshopon Biological Control of Chromolaena OdorllUland Related Activities Since Then- R. Muniappan
The Ecology of Chromolaena odorllUl in theNeotropics - R.E. Crullwel1 McFadyen
The Ecological Role of the Exotic AsteraceousChromolaena odonuain the Bush FallowFarming System of West Africa - Barbara Herren-Gemmill
Chromolaena OdorllUl and Disturbance ofNatural Succession After Shifting CUltivation:An Example from Mayombe, Congo, CentralAfrica - H. de Foresta and D. Schwartz
Observations on Chromolaena odorllUl CL.)RM. King and H. Robinson in Indonesia -A. Sipayung, R Desmier de Chenon and P.S Sudharto
Preliminary Results oh Survey and Assessmentof Chromolaenll odorllUl (Siam weed) In Africa Sulayman S. M'Boob -
The Status of Chromolaena odorat« (L.) R.M. Kingand H. Robinson in Indonesia - S. Tjitrosoedirdjo,Sri S. Tjitrosoedirdjo and R.C Umaly
Distribution and Importance of ChromolaenllOdorllUl in Cote d'Ivoire - M.G. Zebeyou
- Preliminary Results on the Enzymatic Diversityin Chromolaenll odorllUl (L.) RM. King andH. Robinson (Asteraceae) - C Lanaud, M.P. JolivotandM. Deat.
- Mechanical, Cultural and Chemical Control ofChromolaenll odorllUl - R. Muniappan and M. Marutani
Cultural Control of feijicao (Chromolaenll odorllUl(L.) R.M. King and H. Robinson) by PlantingSignalgrass (BrllchiRriadecumbens StapO inSouthern Yunnan, People's Republic of China Wu Renrun and Xu Xuejun
v
yI
VII
IX
X
11
23 .
43
51
57 .
67
71
79
83
III

IV
Progress on Biological Control of Siam Weed,Chromolaena odorata in Thailand - BanpotNapompeth and Amporn Winotai
Status of Biological Control of the Siam Weed,Chromolaena odorata (L.) RM. King and H. Robinsonon Pohnpei, Federated States of Micronesia Nelson M'.,Esguerra, William S. William and[ustino R Smith
Highlights of Work on Chromolaena odorata IDGhana- JA Timbilla and H. Braimah
Studies on Causes of Infertility of Eggs ofPareuchaetes pseudoinsl4/ata Rego Barros .0.0. Torres, D.G. Vargas, S.M. Ritual and E.M. Alforja
The Chemistry of the Putative Female SexAttractant Pheromone of Pareuchaetespseudoinsl4/ata Rego Barros (Lep.: Arctiidae) Stefan Schulz
Arthropods and Pathogens for Biological Controlof Chromolaenll odonua - P.A.c. Ooi, A.N.G. Holdenand P.5. Baker
Guidelines for Introduction, Rearing, FieldRelease and Establishment of Pareuchaetespseudoinsl4/ata
New Approaches to the Biological Control ofChromolaena odorata in South-East Asia -RE. Cruttwell McFadyen
Succession of Vegetation After Suppression ofChromolaena odorata by Pareudiaetespseudoinsl4/ata in Guam - Mari Marutani andR.Muniappan
A Non-Nutritional Relationship of Zonocerus(Orthoptera) to Chromolaena (Asteraceae) andGeneral Implications for Weed Management Michael Boppre
Insect Induced Changes in Chromolaena odorata [ames McConneU, Mari Marutani andR.Muniappan
Country Report - Activities Carried Out byFrance - M. Deat
91
99
105
113
121
127
133
135
143
153
159
163

PREFACE
TheSiam weed. Chromolaena odorata, is considered by manyas a noxious weedo.fthe humid tropics; however, someconsiderit usefulundercertainecological conditions. Thisvolume is an outcome of the Second International Workshop on Biological Control of C.odorata held in Bogor, Indonesia, February 4 - 8, 1991. It contains articles on ecology,chemotaxonomy, cultural, mechanical and biological control and insect plant interactions,recognizing the needfor information on different aspects to tackle it effectively andeconomically without endangering the environment. We hope this publication will induce furtherresearch in multi-disciplinary areas notonly to suppress this weedbut also to tackleproblemsassociated with it
We acknowledge the support provided by the Australian Centre for InternationalAgricultural Research (ACIAR), FoodandFertilizerTechnology Centerfor the Asian PacificRegion (FFfC), U.S. Department of Agriculture, CSRS SpecialGrants Section TropicalandSubtropical Agriculture Agreement #90-34135-5182, Southeast Asian Regional Center forTropicalBiology (SEAMEO BIOTROP) andInstitutde Recherche pourle Developpement enCooperation (ORSTOM) for this workshop and preparation of this volume.
Appreciation isextended toJ. McConnell andWilson Ngforcomputertransferof files,and to RheeOralloand FrancesTorres for formating and preparing camerareadycopies.
R. MUNIAPPAN
PAULFERRAR
v

WELCOME ADDRESS
JOKOPURWANTODeputy Directorfor Administration and Finance
SEAMEO BIOTROPP.O.Box 17, Bogor,Indonesia
Welcometo BIOTROP and Bogor.On behalfof our Director,Prof.Dr.Hj. SittiSoetanni Tjitrosorno, I have the honourand pleasure of welcoming you all to BIOTROP.Our Directoris extending her regretsfor not beingable to be with you today,as she is in thePhilippines attending the SEAMEO Conference. The SEAMEO Council;consistingof theMinisters of Education from the SEAMEO member countriesis havingan annual meetingstarting todayin Manila
It is indeeda great privilegefor us to host the SecondInternational Workshop onBiological Controlof Chromolaena odorata, especially since we are undertaking the.SEAWIC project. SEAWIC,or the SoutheastAsian Weed Information Centre,was establishedin 1985 with the initial financial support from the International Development ResearchCentre (IDRC). SEAWIC,an InterProgram Projectof TAgPBand cm; has completeditsfirst phaseof operation, and starting last year itis operatingits second phase.
I am confidentthat yourdeliberation will providea very important input to SEAWICand to the development of weedsciencein this part of the world. May I also, take thisopportunity to appealfor help fromthe scientists presenthere in supporting BIOTROPSEAWIC activities.
BIOTROP has very modest facilities. I hopeyou could overlookour deficiencies. Allits staff, however, have wann hearts,and are ready tobe of service and help you. Pleasedonot hesitateto approach any of us for your needs.
Last but not least, I am wishing you all a fruitful meetingand an enjoyable stay at .BIOTROP and Indonesia.
VI

OPENING ADDRESS
CHENG-HWA HUANGDirector, Food and Fertilizer Technology Center for the Asian and Pacific Region
5th Floor, 14 Wenchow Street, TaipeiTaiwan, Republic of China
It is a great pleasure and privilege to be with you here, at the opening ceremony of theSecond International Workshop on the Biological Control of Chromolaena odorata.
On behalf of Food and Fertilizer-Technology Center (FFfC), I should like to extend acordial welcome to all participants, particularly those who have come from distant countriesoutside the ASPAC region. I should also like to express my sincere appreciation to allmembers of the eo-sponsoring organization as well as to the BIOTROP headed by Dr. JokoPurwanto, who have all been so generous with their cooperation and support during the workof preparing this workshop. In particular, I aingrateful to Prof. R. Muniappan, GuamUniversity, who has spent a great deal of time on the organization of this meeting.
The first International Workshop on the Biological Control of Chromolaena odoratawas conducted in March, 1988 at Bangkok,Thailand. In the light of the benefit derived fromthe first workshop, the participants recommended that it would be more useful to organizeanother one in the next two or three years to follow up current activities and to review the
-progress made as well as to formulate future action plans.
The Siam weed, Chromolaena odorata, is a native of South and Central America, but ithas now become a serious weed in South East Asia and Pacific Islands. It is therefore a goodcandidate for biological control by the introduction of natural enemies. In addition to this, thefact that the tribe Eupatorieae, to which C. odoratabelongs, has no crop plants or importantornamentals, is a defmite advantage for biological control of this serious weed.
I understand that among many potential biological control agents of C. odorata,anarctiid moth, Pareuchaetes pseudoinsulata, has proven effective in suppressing the weed insome parts of the ASPACregion. We consider that the current trend in research and development, which uses living species rather than manufactured inputs, is 'particular favourable tosmall farm development.
)
I am delighted that we have the opportunity here to bring together such a range ofdistinguished experts, to present recent research in this important field. I am sw:ethat thisworkshop will provide a forum for the scientists involved in research on biological control ofC, odorata to exchange views, update research activities and implementation of biologicalcontrol, and above all for active collaboration in the international area.
VII

FFI'C has always emphasizedpractical technology suitable for small farms in its workof collectingand disseminatinginformation on new agricultural technology. I hope that yourresearch willbeenrichedby your experience here,and that you will return home with freshideas on how to control the Siam Weed which is such a grave problem for farmers.
I should like to express again my thanks to the distinguishedparticipants and guestsassembled here, and to all those manypersons whose work has made this meeting possible.
VIII

OPENING ADDRESS
PAULFERRARResearch Program Co-ordinator (CropSciences),
Australian Centre for International Agricultwal ResearchG.P.O. Box 1571, Canberra, ACT 2601,Australia
I feel honoured to be here in thisdistinguished gathering to welcome youon behalfofACIAR, becausemyCentre hasonly beenable to assist thisconference in a modestway.
.ACIAR standsfor the Australian Centrefor International Agricultural Research, and becausewe are quite a youngorganisation I will tell you brieflywhatwe do.
ACIAR does not itselfundertake research. We wereestablished to arrangecollaborative research projectsbetween Australian agricultural scientists workingin State Departmentsof Agriculture, or universities, or government agenciessuchas CSIRO,and scientistsworking on similarproblems in developing countries, particularly in Asia and the Pacific..Australia has muchagricultural land in the tropics, both wetand dry, and we share manyagricultural problems in common withdeveloping countries. We are fundedas part ofAustralia'soverseasaid program, and theessenceof our mission is to help by transferringexpertise, by getting the overseas and Australian scientistsworking together in partnership tosolveagricultural problems of mutual interest
You may wonderwhy ACIAR shouldbe interested in research on Chromolaena, sincethis weedis not found in Australia It is, however, quiteclose on our doorstep,abundantly inTimor,and as I havejust learned beforetheconference started this morning, also in IrianJaya and perhapsin PapuaNewGuinea. It is probably only a matterof timebefore we sharethis problem too, and we shallbe happy to participate in appropriate controlprogramsfor thebenefitof all of us.
I myselfknowrelatively little aboutChromolaena, and I havecome here this week tolearnfrom you, the experts, whoare assembled at this workshop. On behalfof ACIARIwelcome you to this meeting, with which we are very pleasedto be associated, and 1wishyou an informative week anda fruitful outcometo all yourdiscussions on this importantproblem.
IX

REVIEW OF THE FIRST INTERNATIONAL WORKSHOPON BIOLOGICAL CONTROL OF CHROMOLAENA ODORATA
AND RELATED ACTIVITIES SINCE THEN
R. MUNIAPPANAgricultural Experiment Station
University of Guam . .Mangilao, Guam 96923
U.S.A.·
Thefirst international workshoponbiologicalcontrolofChromolaenaodoratawasheldat Bangkok, ThailandduringFebruary 29 - March4, 1988,hostedby Dr. BanpotNapompeth.Therewerenineteen participants fromeightdifferentcountries. ThecountriesrepresentedwereAustralia, France,Guam,India,Philippines, South Africa, Thailandand Vietnam.
Someof thetopicscoveredduringtheworkshop werehistory,distribution, andecologyof Chromolaena odorata. Chemical, mechanical, cultural control and different aspects ofbiological control were described, including past, present and future activities. There was apresentationoninsectplantinteractionsmostlydirected towards theinteractionbetween C.odorataand Pareuchaetes pseudoinsulata.
. RECOMMENDATIONS OF THE FIRSTINTERNATIONAL WORKSHOP
I.ThesecretariatbeestablishedandlocatedinGuamforthepresent,andtoproduce"Chromolaenaodorata Newsletter".
2. International working groupon Biological controlof Ciodorata be formally establishedandseekan affiliation with the International Organization for Biological Control (IOBC).
3.CountriesandorganizationsthatareactiveorpIaflningtoinitiatebiologicalcontro10f C.odoralaprograms should informthe Secretariat or the working group to disseminate the informationand to maximize cooperation and collaboration.
4.Encourage exchangeofnaturalenemiesofC.odorata inconformity withthelocalandregional .regulations.
x

5. Encouragevariousdonoragenciestosupportactivitiesthatwill strengthenbiologicalcontrol.of C. odorata.
6.Organizea SecondInternational Workshop as a followupof thefirstworkshopin two to threeyears time.
ACTIONS TAKEN
1. The secretariat located in Guam producedthree issuesof Chromolaena odorataNewsletterand distributed these free of cost to interested individuals and organizations. The first issuecarried a bibliography of C. odorata. The second had a list of insects and related organismsreportedon C. odorata. A mainarticleon C.odorata fromIndiaanda noteeachfromThailandand Guam were published in the third issue. The proceedingsof the first workshop was alsoproducedanddistributedfreeofcharge. Also75copiesof theNewslettersand proceedingsweresent10 IOBCsecretary,Dr.JeanPaulAeschlimann fordistribution 10 theIOBCalfiliatedglobalorganizations.
2. The recommendation to becomean international WorkingGroup of IOBC was not activelypursuedeven thoughwehadsomecommunication withtheSecretaryof IOBCpertaining10 thisrecommendation. Oneof the mainreasonswas that by becomingan affiliatedWorkingGroupthere is a commitment to organizeworkshops. As C.odorata is a problemmostly in theLDCsof the humid tropical regions, it is difficult to organize workshops without assistance fromregionaland international donororganizations. IOBCdoes not provide financial support forsuch workshops.
3. Secretariathad verygoodcooperationwith variouscountriesand organizationsin receivingand disseminating information throughNewsletters. The Secretariatassisted in securingseedsofC.odoratafromvariouscountriesforelectrophoresis studiesinMontpellier, FranceandNatal,South Africa.. Also, it providedcopies of reprints to variousresearchpersonnel upon request.
4. Pertaining to the fourthrecommendation, the University of GuamsuppliedshipmentsofP.pseudoinsulata to SouthAfrica,Ghana,Yap,Pohnpei,Kosrae,Palauand Germany. Shipmentsof P. pseudoinsulata will be sent to Indonesia upon receipt of an import permit from the .Governmentof Indonesia.
5. Since the First Workshop, the South Pacific Commission has provided funding to theUniversity ofGuam to assistYapinbiologicalcontrolofC. odorata. Recently,thisorganizationhas indicatedits willingness to assist the Governmentof Yap with the same program. Agricultural Developmentin the American Pacific (ADAP) of USDA has assisted Pohnpei in intraductionandestablishmentof P. pseudoinsulata. An EECfunded llBC and IRHOprojectonbiological control of C. odorata is in operationand you will hear more about it during thisworkshop. FAO has carried out someecologicalstudieson Ciodorata in Western Africa.IRHOand UTA havealsoconductedsomeecological studies in West Africa. The SmithsonianInstitution has approveda project for R. Muniappan to do some ecological studies in westernghatsof India. There are possibilities of CIRAD, ACIAR,GTZ and other organizations ini-
XI

tiatingbiological controlof C.odorata projects insomeof theLOC's, detailsof whichyoumaycome to know by theend of this workshop.
The first workshop wassupported by ACIAR, USOA, and NRC and NBCRC Thailand.Thepresentworkshop issupported byBIOTROP, FFrC, ACIAR, USOA, CIRAD,GTZandtheThirdWorldAcademy of Sciences. In addition, I would like to recognize the presence ofparticipants from IIBC,FAO, France,Germany, Malaysia, Thailand, Indonesia. RepublicofChina,Philippines, Pohnpei, Australia and Guam.
6. The sixthrecommendation was to organize a SecondInternational Workshopin 2 to 3 yearsafter the first workshop. I am glad to point out that we are participating in the SecondInternational Workshop on Biological Controlof C. odorata exactly at the end of the thirdyearsince the first workshop.
XII

_TIlE ECOLOGY OF CHROMOLAENA ODORATA IN THENEOTROPICS
------------
R.E. CRUTIWELL McFADYENAlanFleteherResearch Station,Queensland Department of Lands,
P.O. Box 36, Sherwood, Brisbane, Queensland, 4075, Australia
ABSTRACf
Chromolaena (Eupatorium) odorata is a perennial scrambling shrub native to theNeotropics,occurringin mostareasbelow 1000maltitudefrom southernFlorida to north-westArgentina. In the Neotropics C. odorata is alwaysfound in competition or association with acomplex of closelyrelatedspeciesnotpresentin Asiaor Africa. In theNewWorld,C. odorataiscommon in mosthabitats exceptundisturbed rainforest, but is seldomweedyand is never thetargetof specificweedcontrolmeasures. It is typically a plantof secondary succession,rapidlyinvading clearings and persisting until shaded out by the overgrowth of forest trees. Whereagricultureand human activitypreventforestregeneration. C.odorata persistsas a typicalplantofforestedgesandpaths,abandoned fieldsandpastures, buildingsites,andalongroads,railwaysand streams.
- .In the New World, C. odorata does not invade pasture nor compete successfully with
plantation crops,thoughit maybea minorproblem in theearlyestablishment stages. Nocontrolmethods other than regular slashing are required in the New World, a marked contrast to thesituation inAsia or Africa. This reducedaggressiveness is due to attackby a largecomplexofinsects,otherarthropods, anddiseases, togetherwithcompetition withrelatedplants. Soil typeis nota factor,andclimateis similarin the twoareas. The implication is that in the Neotropics,C. odorata is largelycontrolled by thesebiotic factors.
TAXONOMY OF CHROMOLAENA ODORATA
Classification and NomenclatureThe genus Chromolaena is in the family ASTERACEAE (formerly COMPOSITAE),
whichis one of the largestplantfamilies. It is a well-defined, verysuccessful family, regardedas the highesti.e. mostevolvedof the plantfamilies. The Asteraceae are found throughout theworldand are particularly abundantin the Americas. Most speciesare shrubs or herbaceous:trees arerare. Forsucha largefamily theeconomic valueis low;withrelativelyfewcropplantsthough manyornamentals (Toelken 1983).
The EUPATORIEAE is a well-defined, mostly New World tribe, with white, reddishorblueish flowers lacking ray florets (Robinson & King, 1977). There are no crop plants or
1

importantomarnentals in the Eupatorieae, but importantweed species are Mikaniamicranthaand Ageratumconyzoides.
Eupatorium, beforeitwassplitupby KingandRobinsoninthe 1970s,containedover 1200species, mostly in the Americas (King and Robinson, 1970). Several are important weeds; Ageratina altissima (E. rugosum) in the eastern USA, Ageratina adenophora andAgeratinariparia in Indomalaya to Southern China, South Africa, Hawaii and eastern Australia,Fleischmannia microstemon,a minorweedintheAmericas,andAustroeupatoriuminulaefoliumin Indomalayaand Sri Lanka (Anon. 1983). Chromolaena contains 129species,all from Southand Central Americaand theWest Indies(Kingand Robinson 1970). Of these,C. ivaefolia andC. laevigata are widespreadand occasionallyweedy in the Americas,but only C. odorata htsspreadbeyond the New World.
ECOLOGY OF CHROMOlAENA ODORATA IN THE NEOTROPICS
DistributionC. odorata is distributed naturally overa wide area of the tropical and subtropical
Americas, from southern Florida to the extreme north of Argentina. There is no evidence ofrecentspreadin thesecountries,noris theplanta significantor importantweedin theNewWorld.It is, however,one of the more widespread species in the super-genusEupatorium, most of theother specieshavingquite restricteddistributions, and even in the Americas,C. odorata showssome weedy characteristicssuch as the rapid invasionof cleared forest or abandoned pasture.
In the AmericasC.odoratais confinedto the tropicalzone (Fig. 1)and has not spread intothesub-tropicalhumidareasas it hasdone in theOldWorld,notablyinnorthernIndiaand Nepalacross to southern China and in southern Africa. This difference is presumably due tocompetitionwiththe numerousother Eupatorium speciesoccurring in the Americas,and to theeffects of attack by insectsand diseases found inthe Americas and not in Asia and Africa.
In Centraland South America,C. odoratais commonin mostcleared or secondaryforestareas below 1,500 to 1,000 metres altitude and receivingover 1500 mm annual rainfall. It isfoundas far southasParaguayand theextremenorthof Argentina,but is notpresent in southernBrazil. InGuyanaandAmazonianBrazil,it is presentbut scarce,and in Venezuelait is commonin shady sites and at higher altitudes. It is also present in most West Indian islands (Fig. 1).
Detailedstudiesof the ecologyof C. odoratahaveonly been made in Trinidad (CruttweU1972),thoughsomeobservationshavebeen madein Centraland SouthAmerica. EcologicaUy,Trinidadis an off-shoreislandin thedeltaof the riverOrinoco in north-eastSouth America,andits flora and faunaare reduced versionsof thoseof the mainland. Human populationdensity ismuchhigher than on the adjacent mainland,and more of the original forest has been cleared.
InTririidad, C.odoratais foundwhereverthereisopenwell-drainedground,includingdryand exposed slopes, though plants in such situations are stunted. It grows in young citrusplantations, but not in rainforest, nor in close-planted established citrus, coffee or cocoaplantations. C. odoratais mostcommonin abandonedfieldsandpastures,on building sites and
2

alongroads,railwaysandstreams, andis foundin thesesituations throughoutTrinidad. Allsoiltypesfoundin Trinidadappear to be suitablefor growthalthoughC. odorata is lesscommoninthe sandy soils of the east coast, and does not grow in seasonally waterlogged soils.
InTrinidad,C.odorataformsdensetangledbushes1.5to 2 minheight,veryoccasionallyreachinga maximum of 6 mas a climberon otherplants. Thestemsbranchfreely,withlaterals .developing in pairs from the axillarybuds. A large plant may have twenty or more stems ofvaryingsize,often bent over under the weightof their branches, and shadinga ground area ofapproximately 3.5m2• Theolderstemsarebrownandwoodynearthebase; tipsandyoungshootsare greenand succulent The rootsystemis fibrous and doesnot penetratebeyond20 to 30cminmostsoils. Theplantsdonotreproduce vegetatively,thougholdstemsmaydieandbereplacedby new shoots from ground level.
Life historyThereare twoseasonsinTrinidad,a dry seasonextending fromJanuary to June,anda wet
seasonstartingwithheavyrainsinJuneandJuly,andlastinguntilNovemberor December. Theseverityand durationof the seasonsvary greatlyfrom year to year (Anon. 1968).
Flowering of C. odorata, induced by shortening days, starts in late December andcontinues till the middle of February,although occasional plants, especially regrowth, mayfloweruntiltheendof March. Thecapitulaare bornein headsof20 to 60at the tipsof all stems,branches,andaxillaryshoots,and the onsetof flowering prevents furthergrowth. The flowersarewhiteorpaleblueish-lilac,andform massescoveringthewholesurfaceofthebush.C.odoratais conspicuous when in bloom in late December andJanuary, hence its West Indian name"ChristmasBush"..
When flowering is over, mostof the leaveswitherand fall. New leavesand shootsgrowfromtheold leafaxils, and thedead terminal parts of thestemsdropoff. Theextentof leaf-fall,andtherateofregrowth,dependon themoistureavailable. Bushfiresarecommonat thisseason,and frequently destroyall above-ground growth. The ripe seeds are wind-dispersed, or adhereto the fur of animalsor to clothes.
Withthestartof the rainyseasoninJune orJuly, newgrowthis rapidandextensive. Newshootsappearfrom the roots,and from all undamaged axillarybuds. Seedsgerminaterapidly,and in suitablesoil large numbers of seedlings develop. If unchecked, C. odorata plants formdense massesof lush green growthby the time the rains slacken in September. Old stemsaregradually shadedbynewgrowthabovethem,andsufferdieback. Oldbushes thus formatangledmass of old and new stems,withgreen shootsand branchesin all directions.
Status in plant communitiesIn thenaturalrainforest, C.odoratawasa plantofforestclearingsandof theedgesof rivers
and savannahs. There is, therefore, no store of C. odorata seed in the soil of undisturbed/
rainforest, andduringa visit to the mouthof the Amazon-in Brazil,it wasnoted that C. odoratawas entirelyabsent where virgin rainforesthad been cleared, but was present around an oldestablished village, andwasbeginning toinvadecultivation landwhichhadbeencleared 3yearspreviously (Cruttwell 1971). Slash-and-bum shiftingagriculture traditionally practisedin these
3

areas would have provided a succession of short-lived habitats for the plant, which wouldprobably always have been scarce.
In more densely populated areas, where the original vegetation has been completelycleared, C. odoratais typically found growing in small patches, discontinuous in time and space.Newly cleared ground is fairly quickly colonised by C. odorata together with razor grass(Paspalum virgatum) and sensitive plant (Mimosa pudica) in exposed sites, and Bidens pi/osaand other composites, and a variety of herbs, in more sheltered areas. Such land is usuallysubsequently cultivated or used for building, but if abandoned, black sage, guava or some similarshrub competes with Chromolaena and ultimately reaches a height at which C. odoratais shadedoul
In Trinidad, three other species, previously Eupatorium, occupy similar habitats and oftengrow in association with C. odorata. Austroeupatorium inulaefolium isusually found in thewetter and shadier sites; it does not occur in the more exposed and dry areas but is found undershade denser than that tolerated by C. odorata. E. iresinoides is also a plant of shady places andis often found along roads ides and forest edges. C. ivaefolia grows in similar sites to C. odoratathough again not in the more exposed places.
C. odorata is, thus, a plant ofopen land, where it supersedes the pioneer ephemeral herbs,is in turn displaced by small trees and bushes, and disappears completely when the forest canopybegins to close.
Status as a weedIn its native habitat in the Americas, C. odorata never becomes a serious weed. On
intensively cultivated land, the seedlings seldom become established. In pasture and alongroadsides, growth is usually cutlassed at 3 to 4 monthly intervals, and this is sufficient to controlC. odorata. In well-grazed pastures, trampling and competition from the pasture prevents theestablishmentof seedlings. In abandoned pasture and fallow, C. odoratamay become dominantin small areas, never exceeding O.5ha, and this dominance is short-lived, as in a few years it isdisplaced by the growth of secondary forest. Under citrus or cocoa the plants are etiolated orpresent only at the edges where they are easily controlled by annual or bi-annual CUlling. Inplantations of young trees, C. odorata establishes initially, but after four or five years, with .increasing shade from the trees the C. odorataplants become weaker and disappear completelyas the canopy closes. There are no reports ofC. odorata competing successfully with young treesin forestry plantations in Trinidad or elsewhere.
Mortality factorsIn the West Indies, many plants die each year, at all seasons, from a variety of causes, and
the maximum length of life is not known. In poor soil, cutJassing to ground level or burning maykill the plant. In good soil, regrowth is rapid, and very few plants die. Plants also die if the soilbecomes water-logged; the leaves yellow, the stems blacken and wither, and become susceptibleto fungal diseases.
Plants often die when they become shaded by the growth of bushes such as black sage(Cordia curassavica) or w~ld guava (Psidium guajava), or by rainforest regeneration. Dieback
4

of stemsbecomesincreasingly severe,and newshootslireetiolatedand feeble, Etiolatedstemsare veryliabletoinsectattack,andregrowth fromaxillarybudsisslow,untilfinallythelaststemdies.This is the normalplantsuccession as open groundgradually gives way to low bush,andthento therainforestwhichis theclimax. vegetation overmostof Trinidad. C. odoratadoes notsurviveonce trees have grownto 6 metresor higher,except in clearingsor along pathways.
Experiments with potted plants clearly demonstrated that C. odorata cannot persist inshadein Trinidad. Plants in rainforestsurvivedfor less than4 months while thosein lessdenseshadesurvived8 monthsand plantsin full sun steadilyincreasedin size throughout(Table 1).These experiments also showed that the effect of insect attack, measured as. the number ofdamagedbuds (growing points),wasgreater in shade than in sun.
Table 1. Growth & Survival of C. odorasain Sun & Shade in Trinidad
Full S\Dl Partial Shade Forest
Survival >8 8 3-4 (n=6)(in months)
Stem Length after 6 mths 160 56 0(% of initial)
Total Leaves after 6 mths 166 73 0(% of initial)
Undamaged buds after 6 mths 183 17 0
(% of initial)
- IntheNeotropics, C.odorataisattackedbya verylargenumberofinsects,over200havingbeen recordedon the plant duringa 6 year study (Cruttwell 1972). Of these,about halfwereprobably polyphagous, a quarterwere restricted19 plants in the Asteraceae, and the rest werespecificto Chromolaena. All stagesand partsof the plantswereattacked,except for the rootswhichwere not examined.
Thecumulative effectof theseinsectswasconsiderable. Regularweeklysamplesof stemtip and axillarybuds were taken at 3 sitesover 2 years,and theresultsshowed that between 2S •and 50%of all growingtips weredestroyedby insectattack (Fig. 2). Damagewas caused bydifferentinsectsat differentsitesandseasons, andingeneralwasheaviestin shadedsites. In thecoolernorthern valleys, therewasalsosignificantstemdamage bytheboringweevilRhodobaenUscarinivemris. Seed germination percentages were low, about 17% of filled seed, and manyflowerheads failedtoproduce seedsatall. Seedling survival isreducedby theattackof stemandtip feeding insects,and thegrowthand competitiveness of established plants is reducedby theinsectdamage.
Recentsurveysby theCAB International Instituteof Biological ControlhaveshownthatC. odorata in the West Indies is also attacked by diseaseorganisms,which furtherreduce thevigorand competitiveness of this plant.
5

Control MeasuresC. odorata is not a serious weed in the Neotropics and no special control methodsare
necessary. For example, of the numerous papers quoted in Weed Abstracts regarding thechemical controlof C. odorata, nonerefers to its controlin the Americas (Table2)
Table 2. Papers Published in CAB Abstracts (1972 - 1990) on the Control of Chromolaena(Eupalo"ium) odorata
(All Control Methods)
C. odoraia in Africa
C. odorata in Asia
C. odoraia in the Pacific Islands
C. odoraia in the Americas
Total
66
75
5
o
146
. Neitherclimatenorsoiltypecanexplainthisdifference, as C.odoratain Asiaand Africaoccursinclimatesexactlysimilarto thoseof SouthandCentralAmerica as wellas in moresubtropical areas,andoverthesamewiderangeof soil types. Thedifference in theaggressivenessof this weedis due to the cumulative effectof all the host-specific insectsand diseases whichattack the plant throughout its range in the Neotropics, but are largely absent from Asia andAfrica,
6

REFERENCES
Anonymous. 1968. Rainfall Reports of Ministty of Works Drainage Division. Statistical Digest,Trinidad and Tobago Govt., Port of Spain.
Anonymous; 1983. Important weeds oftheworld. 3rd Ed. January 1983. Bayer, A,O. Leverkusen,.Oennany. 711 pp.
Cruttwell, R.E. 1971. Report on a Survey of the Insectson Eupatoriumodoratum L. in South America.undertaken in April-May 1971. Cl.B.C. West Indian Station Unpubl. Report, 12 pp.
Cruttwell, R.E. 1972. The insects of EupatoriumodoratumL. in Trinidad and their potential as agentsfor biological control. Ph.D. Thesis. Univ,of West Indies.St. Augustine. Trinidad. 241 pp.
King. RM. & H. Robinson. 1970. Studies in the Eupatorieae (Compositae). 24: The genusChromo1lJena. Phytologia 20: 196-209.
Robinson. H. & RM. King. 1977. Eupatorieae - systematic review. In: Heywood, y.H.• Harbome,J.B. andTurner. B.L. (Eds) The Biology and Chemistry of the Cornpositae, Vol. I. pp. 437486.
.Academic Press. London.
Toelken, H.R. 1983. Compositae.ln: B.O. Morley and H.R. Toelken (Eds.) Flowering Plants in. Australia. pp. 300-314. Rigby Publishers. Adelaide, Sydney. .
7

8
SOUTH AMERICALEGEND
Distribution ofC.ODORATA .
Altitude (1000 m) --'
Isohyet(1000 mm). __
Figure I.Distribution of Chromolaena odorata in the Neotropics.

C.odorata in TrinidadDestruction of growing tips and buds
100
90
80"d.
~ 700
60l-o...,UJQ) 50"dUJ 40~......,
30~
20
10
0Jan
AprJul Jan
oct Aprmonths (1967-1968)
Juloct
1--- shady site -+-- open site ~ exposed site
Figure 2.
Chromolaena odorata in Trinidad: Percentage destruction of growing tips
9


THE ECOLOGICAL ROLEOF THE EXOTIC ASTERACEOUSCHROMOLAENA ODORATA IN THE BUSH FALLOWFARMING
SYSTEM OF WESTAFRICA
BARBARAHERREN-GEMMILLcloInternational Instimteof Tropical Agriculture
Benin ResearchStationB.P. 08-0932,Cotonou, Benin
ABSTRACT'
. Ecological dynamics of ChrOf1UJlaeNJ odorata dining two years of cultivation and two years offallow, under two different systems of land management, are presented. C. odorata is a noxious weedduring thecultivation period. but subsequently becomes an important component of the fallow regrowthcommunity. According to its frequency, density. average height, biomass and nutrient content, it is thepredominant species of the early fallow successional stage. It is concluded that C. odoratamay exert aconsiderable influence on today's bush fallow successional processes in West Africa, and that furtherenquiry should be made before a biological control program is launched.
INTRODUCTION
Chromolaena odorata (L.) R.M. King and H. Robinson (Family Asteraceae; TribeEupatorieae),a nativeof thetropicaland subtropicalAmericas, wasaccidentallyintroducedintoSoutheast Asia early-in this century, where it established itself as a serious weed (McFadyen1988). Introduced into WestAfrica around 1940,it has spread throughout the rain forest zoneand the borderingedges of GuineaSavannahzones fromCameroonto Cote d'Ivoire. The peststatus of C. odorata in Africa is not straightforward; in the literature it has been alternatelycondemned as a weed and praised for its nurse-plantand mineral-cycling properties (Agbim1987,CAB 1988,Ivens 1974). Pyrrolizidineallcaloids in the flowersof C. odorataattract andprobably supportthepolyphagousvariegatedgrasshopper,Zonocerusvariegatus (L.) (Acrididae).This adds another facet to the already complicateddiscussionconcerning the pest status of C.odorata and whetherand how it should be controlled.
Before a biologicalcontrol program is instituted against Chromolaena odorata in West.Africa. theecologicalrole of thisplant shouldbe betterelucidated. Whileagroforestrysystemsmaybe envisagedand promotedin which the transitionfromarable to tree cropsoccurs withoutnatural fallow, the fact remains that the vast bulk of fallow land in West Africa regeneratesnaturally. The questionof whatroleC:odorataplaysin thisregeneration,and whatwouldoccurin the absence of C. odoratashould be answeredbefore a control program is launched.
11

This submission reports on ecological dynamics of C. odorasa during two years ofcultivationand two yearsof fallow, under twodifferent systemsof land management The datareported in this paper were gathered in the frameworkof a larger study on fallow succession inWest Africa
MATE~SANDMffiTHODS
Site characterizationLocation: FieldtrialswereconductedonthegroundsoftheIntemationalInstituteforTropical
Research (DTA)'s experimental farm in southwestern Nigeria (7 30'N, 3 54'E), located thirtykilometers southof the northern limitof the lowlandrain forestzoneof theWest Africantropics. 'Elevation of the experimental farm varies from 190 to 230 m, and the rol1ingtopography isdominated by slopesthat are3 to 10%;the fieldtrialsof this study werelocatedon essentially flatbenches. .
CUmate: Rainfalldistribution is bimodal,with maximain May to June and in September,separatedby a short butcomparatively cool,dry period. The maindry periodfrom NovembertoAprilincludesat least five"arid" months, definedas thosewith less than 100mm precipitation ina month. Meanannualrainfallis 1280mm,witha strikingvariation in therainfallpatternfromyeartoyear(l983:921mm,28%belownormal;1984: 1360inm,withalmostnoAugustbreakinrainfall;1985: 1TI2 mm,thewenestyeareverrecorded at thelITA Ibadan weatherstation,withrainfall30%above the 30yearaverage; 1986: 1232mm,close to normal,althoughrains duringboth gro\W1gseasonsendedearlier than usual),
Geology and Soils: Soils in the trial fields were Ferric Luvisols of the Egbeda series,developed oncoUuvium andresidium derivedfrompre-Cambrian bandedbiotite-muscovitegneissalternating withmicaceous schist(Moonnanet al. 1975). The soil is of coarse to mediumtextureinthesurfacehorizon, withsandyclaytocJayey subsoil,andaJayerofanguJarandsubanguIMquartzgravelbelowtheA horizon. Undersecondary forest. thesurface soilhas a fairlyconstantpH valuearound5.8 to a depthof 150cm..
Land-use history: The land was used intermittently for agriculture before UTA wasestablished in 1968. Untilmattimethearea wascharacterizedbya mosaicof landuses, fromcocoaplantations (largely neglected), to cultivation, to bushregrowththicket(Smythand Montgomery1962). From 1968,whenlocalvillageswererelocatedoutsidethepremisesof lITA,until 1982,theforestwaspermittedtoregenerate. By 1982,a weU-developed fallowvegetationhadgrownaroundscatteredhigh-forest trees whichhad beenpreserved by previouscultivators.
DATA COLLECTION
In the fall of 1982,as a demonstrationduring a land-clearingconference held at IITA, aportion of this forest was completely cleared of regrowth vegetation and high forest trees bycaterpillartractors and tree pushers. From thisclearing,a 30 m x45 m plot was reserved for thisstudy. Anadjacent30 m x90 m of the regrowthforestwasclearedat thesame timeby traditional
, methods: woody.plants were removed by machete and overstory trees were thinned. Onbothplots of land, cleared brush was distributed across the ground and burned.
12

Both plots were cultivated in the 1983and 1984growing seasons. Cassava (Manihotesculenta) wasplantedindensities of I plantpersquaremeter,and theninterplanted withmaize(Zea mays). Maize was harvested at the end of the 1983growing season while cassava washarvested at the end of the 1984 growing season. To continue the traditional vs. mechanicalcomparison, thesoilof thefieldplotclearedby tractorwasalso initiallycultivatedby tractor. Inthe manually-cleared plot. workers wereasked to cultivateas they wouldtheir own farms, Inparticular,whenweeding, workers werecarefultoproteetanyresproutingorgerminating specieswhich theymightfavor in theirownfarms. Becauseclearingand cultivation effectswere thuscombined, theresultsof thisstudymustbeconsidered purelydescriptive of the implications ofdifferentmanagement techniques, and not as functional investigations.
The emergent weed and crop community during the two years of cultivation, and thesubsequent fallow regrowth community occuring in the first two years of abandonment weresampled. Sampleswere taken,once each year, during the late growing season (September toOctober), eight weeks after the last weeding. Plant species were identified according toHutehinson and Dalziel(1972). Structural profileson 20-mpermanent transects, and speciescomposition and biomass within25 squaremeterplots were recordedin each field. Nutrientstocksin soiland in thefivemostpredominant speciesfromeachfieldweresampledin the firstyearof fallow regrowth, usingtheanalytical methodology outlinedby Black (1965).
RESULTS
Speciescomposition. C. odorata was the most predominant species, throughout both the cultivation and the
fallowing period. C.odorata hadthehighestaveragedensityper squaremeter,and thegreatestfrequency of any species,throughout this study (Table I).
Lite-form analysis.C. odorata is sometimes called an "herbaceous" plant (McFadyen 1988),and in other
contextsa shrub (Akobundu and Agyakwa 1987). For the purposes of this study it has beenassigned to the shrublife form. C. odoratastemsdo becomewoody with age, and the plant isa perennial; the herbaceous category, in terms of life-form analysis, is best left to whollyherbaceous annualplants.
Although an initialherbaceous stagein fallow succession hasbeenrecordedinotherWestAfrican sites(Richards 1953,Clayton 1964),a predominantly herbaceous periodneveroccurredin thepresentstudy. Ifsucha community waspresent,it occurredon a timeframeless thanoneyear, and was not capturedby the study's late-season sampling dates. Although herbaceous
.weedsmadeup an important component in both fields, woodyor semi-woody vegetation wasequallyor more important in all structural profiles. The dominance of the "shrub" life formthroughout thecultivation landfallow periodisalsoseenindensityand meanheightdata(Table2). And as can be seen from this table, C. odorata is the most importantcontributor to thepredominance of the shrub life form, in termsof height if not alwaysin density.
13

Population dynamicsC.odorata initially occurred at muchlowerdensities in themachine-cultivated field than
in the traditionally cultivated field, the spaceinthe former being largelyfilledby glasses andherbaceous weeds duringthe cultivation period. Population densitiesof C. odorata increasedovertimeof fallowing in the machine-cultivated field.anddecreased over timein the traditionallycultivated plot.
Species interactionsThe abundance of the different life-forms in the presenceor absenceof C. odorata was
analyzed (Fig la andb). C.odoratadidnotemergeas a noxious weedretardingsuccession. Itspresence did not preclude the establishment of high forest tree individuals; in fact more treeindividuals occurred in plots where C. odorata was established than in its absence. Anexamination ofplotdiagrams showsthatinalmosteveryplotwherea highforesttree established.it wassurrounded byC.odoratawhich grewnotnecessarily abundantly. buttallerthanaverage;and thesurrounding C. odoratawas uniformly tallerthan the tree seedling.
In theabsence or reduction of C.odorata, suchaswouldbe theaimofa biological controlproject on the weed. one would hope that other woody species which also encourage treeestablishment would increase in importance. such as Lecaniodiscus cupanoides. However.negative species association analysis only showed that there were significantly more grassindividuals in the absence of C. odorata.
BiomassBiomass trends of C. odorata over time and by treatment followed basically the same
trends as thoseof population densities duringcultivation (Fig2). Asthe population densitiesofC.odoratadecreased fromthecultivation to thefallow periodinthetraditional field.thebiomass
. of the species increased. indicating that a thinning process was resulting in fewer but largerindividuals. This did not occur in the machine cultivated field. where relatively smallerindividuals continued to increase in number in thefallow.
, Interestingly. biomass ofC.odoratahadnorelation. positiveor negaiive.toabovegroundcrop biomass.
Nutrient stocks.The contribution of C. odoratato nutrient stocksin the first yearof fallowis recordedin
Table3. Again. thecontribution isveryhighin thetraditionally managed field.andconsiderablylessso in the machine-cultivated field.
DISCUSSION
C.odorata hasbeenreadily recognized asa weedinpartbecauseit is an extremely visibleplant; it occursabundantly along roadsides, and may predominate in the plant community infallow fields. Whatisneeded. however. isa morepreciseassessmentof itsdynamicsduringboththecropping and fallowing period.
14

The following is a summary of observed C. odorata dynamics emerging from thisparticularstudy.
Althoughiris generallybelievedthat weedsincreasewith thedurationof cultivation,thecontrarywas true during the shortcultivationperiod (2 yr) of this study. Cultivatedplots wereextremelyweedyinitiallyand weeddensitiesdecreased witheach successiveweeding. Duringthe cropping period, C. odorata was the most predominant weed species, especially in thetraditionally managedfield.:Itsbiomass,however, didnotappear to affecttheabovegroundcropbiomass,comparing occurrencein meter-square plots at the end of eachgrowing season. Thisof course reflects competition between established plants; C. odorata's effect on newlyestablishedsproutsof cassavamay be more deleterious. The fact that crop establishmentandbiomass.isconsiderablylower in the traditionally-managed field, with its denser weedpopulation, than in the machine-cleared andcultivatedfields,suggeststhat there is a veryreal problemof weeds in traditional bush fallowfarming systems.
. Conventional wisdom has it that one of the functions of the fallow is to reduce weedpopulations (Ewel 1986; Moody 1974). If C. odorata is consideredas a weed, this processoccurredonly in the traditionally-managed field, where fallowing sawan increase in trees andshrub-treesat the expense of dense C. odorata stands. In the heavily-disturbed mechanicallycultivated field, C. odoratacould flourish only in the fallow recoveryperiod.
The growth,establishment and even thepossiblebreakingapartof a dense woodythicketso characteristic of fallow regrowth is the most salient feamre to arise from the fallowsuccessional study from which these data are derived. The process occurs rapidly in thetraditionally-managed field,and maybe occurringat a muchmoreretardedrate in themachinecultivated field. Thickets commonly occur in tropical forests in response to disturbance(Bourgeron 1983,Brunig 1983). Unfortunately we do not havea very clear conceptionof thesignificanceof thicket formation in fallowsuccession. For instance,we do not know whethera bushythicket is a stage whichof necessitymustprecede theestablishmentof high forest treespecies,or whetherforest treessurpassthe thicketin spiteof its presence. Neither is it apparentwhethera thicketor a well-stratified formation is moreefficaciousin the restorationof nutrientreservesand soil structureover a fallowing period. What is evident is that C. odorata hasa lifeform which lends itselfextremelywell to the formation ·of thickets,and in this sense probablycontributes to, rather than deflects, the natural course of succession following disturbance infallowforests. In addition,whilewe cannotgeneralizeforall speciesin the thicket,we can saythat C. odorata appears to facilitate the establishment of high forest tree species in the fallowsuccession. The contribution of C. odorata to nutrient stocks in fallow vegetation is alsosubstantial,and exceeds its contribution of biomass.
It is hopedthat theinformation presentedherewillbeconsideredincharacterizingtheroleof C. odoraiain WestAfrica. Asa preliminary to a biologicalcontrolprogram,the questionofhow fallow ecologicaldynamicswouldbe altered in the absence of C. odorata should be answered. And since this is an inquirywhichmuch be made in the context of along-termstudy,time is of the essence.
15

REFERENCES
Agbim, NM. 1987. Carbon cycling under Chromolaena odorata (L.) K.&R. canopy. BiologicalAgricultureand Horticulture4:203-212.
Akobundu,1.0. & C.W. Agyakwa. 1987. A Handbookof West African Weeds. UTA, Ibadan, Nigeria.
Black, C.A. 1965. Methodsof Soil Analysis. Vol. 2, Agronomy.AmericanSociety of Agronomy,Madison,Wisconsin.
Bourgeron,P.S. 1983. Spatialaspectsofvegetationstrueture. pp.29-47. In: F.B.Galley, 00.Tropical RainForest Ecosystems:Structure and Function.Ecosystemsof the World Volume 14A. ElsevierScientific PublishingCo., New York, '
Brunig,E.F. 1983.Vegetationstructureandgrowth. pp. 49-75. In: F.B. Golley, 00.Tropical Rain ForestEcosystems:StructureandFunction.Ecosystemsof theWorld Volume 14A.ElsevierScientificPublishingCo., New York. '
CAB. 1988. Prospectsfor thebiologicalcontrolofChromolaena odorata (L.) R.M.KingandH. Robinson.pp. 25-33. In: R. Muniappan, (00.) Proc.,First Intl. Workshopon Biological Control ofChromolaena odorata. Ag. Expt. Station, Universityof Guam, Mangilao, Guam.
Clayton, W.O. 1964.Secondaryvegetationand the transition to savanna near Ibadan, Nigeria. J. Ecol.46:217-238.
Ewel,1. 1986. Designingagriculturalecosystemsforthe humid tropics. Ann. Rev. of Ecol. and Syst17:245-71.
Hutehinson,1.& 1. M. Oalziel. 1972. Flora of West Tropical Africa. 2nd edn. Crown Agents, London.
Ivens, G.W. 1974. The problem of Eupalorium odoraium L. in Nigeria. PANS 20:76-82.
Mcfayden, R.E. Cruttwell. 19~8. History and distributionof Chromolaena odorata (L.) R.M. King andH. Robinson.pp. 7-12. In: R. Muniappan,00.,Proc., First Intl, Workshopon Biological ControlofChromolaena odorata. Ag.Expt. Station, Mangilao,Guam.
Moody, K. 1974. Weeds and shiftingcultivation.In: ShiftingCultivation and Soil Conservationin Africa. S9ilSBull.'24:155-66.FAO, Rome.248 pp. '
Mooiman, ER., R. La! & A.S.R.Juo. 1975.The soils of TITA. UTATechnical Bulletin No. 3. UTA,Ibadan, Nigeria.
Richards,P.W. 1953. Ecologicalstudieson therain forestof southernNigeria.I. The Structureand floristiccompositionof the primary forest J. Ecol. 27:1-61.
Smyth, AJ. & R.F. Montgomery. 1962. Soils and Land use in Central Western Nigeria. Ministry ofAgricultureand Natural Resources, Ibadan.

Table I. Average density and frequency of the most abundant species.( years and fields pooled together)
Species
Chromolaena odorataSida veronicifoliaSpigeliaanthelmia
. SoltuuunverbascifoliumPhyllanJhusfloribundus
.Brachiaria deflaaPaspalumorbicular«Lecaniodiscus cupanoidesAlbizia zygiaHippocratea pallensCnestis ferrugineaMallotusoppositifoliusIndigofera hirsutaDombeyabuettneriRauvolvia vomitoraNewbouldia laevisTalinumtriangulareCombretum hispidumCluJssalia /collyDeinbollia pinnaJaAlchornealDxifloraMotandra guineensisSynedrellanixlifloraTridaxprocumbens
Life-form class
ShrubHerbHerbShrubShrub/climberGrassGrassShrub/treeTreShrub/climberShrub/climberShrubShrubShrubShrubShrub/treeHerbShrub/climberShrub/treeShrub/treeShrubShrub/climberHerbHerb
Average densityper m2 (+S.D.)
3.75 (+5.22)1.91 (+2.38)0.84 (+5.43)0.33 (to.72)0.47 (+3.24)0.70 (+2.89)0.41 (+1.47)0.27 (to.92)0.25 (to.85)0.17 (to.35)0.21 (to.98)0.24 (+1.35)0.26 (+1.65)0.21 (+1.04)0.13 (to.45)0.13 (to.33)0.37 (+2.11)0.17 (+1.09)0.14 (to.74)0.15 (+1.02)0.21 (+2.20)0.31 (+2.97)2.05 (+24.60)0.24 (+9.03)
%Frequency
59.244.8
·.20.820.019.5 .19.219.219.217.514.814.514.014.013.211.511.211.210.810.09.58.88.08.04.0
17

Table 2. Cbanges In densIty and mean belgbt by life rorm class In 2S meter-square plots In tbemachine-cleared and cultivated neld (neld 1)vs. tbe tradltIoillllly cultivated neld (ne Id2), during twoyears or cultivatIon (year 1 and 2) and two years or raUow(year 3 and 4).
Year 1 Year 2 Year 3 Year 4Field IField 2 Field IField 2 Field IField 2 Field IField 2
mean number of individuals per life form class, per square meter:
high forest trees .04 .80 .16 .75 .60 .71 .85 1.23
understory trees, .84 .16 .04 .24 .52 .20 .76 .88
shrub/trees .80 1.12 .76 1.14 .40 2.00 .80 3.45
shrubs 2.20 11.56 1.24 9.10 3.16 5.75 6.87 4.69
(C. odorala) 1.76 9.12 .36 6.20 2.04 3.04 4.56 1.93
shrub/climber 2.00 3.08 1.68 1.81 1.52 2.30 1.69 2.13
climber .36 .88 .16 .24 .24 .40 .50 .81
grass 1.20 .132 1.08 .95 1.80 1.15 2.63 .46
, herbs 3.56 5.92 4.36 '2.00 16.56 3.40 10.26 1.81
mean height, in cm., per life form class, per Square meter:
high forest trees 1.62 17.53 8.23 27.54 16.80 29.28 19.65 32.10
understory trees 18.45 13.43 4.00 20.47 10.04 12.55 12.53 11.57
shrub/trees 15.66 28.13 12.10 16.07 18.23 57.99 25.30 72.63
shrubs 47.03 93.79 23.96 46.97 122.26184.61 134.60 226.50
109.80 25.00 67.24 •(C. odorala) 96.67 183.1023,8.22 156.38 273.20
shrub/climber 19.32 56.00 34.19 22.23 31.65 90.00 65.54127.32
climber 20.80 26.26 3.00 71.43 8.20 10.87 16.52 32.31
grass 13.00 8.84 9.27 4.28 57.00 18.00 65.44 15.60
.herbs 18.99 23.00 ' 13.32 32.14 49.65 14.50 ~1.60 9.82
18

<,Table 3. Total nutrient stocks In aboveground vegetation of a year-old faUow, and percent of this
"., contributed by C. odorQtQ.
Total In g1m2 (=100%) Contribution of C. odortltiz %
Element field 1· field 2 . field 1 field 2
C 206.9 477.91 4.8 50.5
N 5.7 13.3 4.9 64.5
P .5 1.2 5.2 73.9
Ca 3.3 3.1 .3.4 24.8
Mg .9 1.3 8.0 52.8
K 9.9 14.8 4.8 74.2
(field 1 =machine cultivated field; field 2 =traditionally cultivated field).
19

. in the presence 01:
there are significantly:
more high loresttree individualsmore understorytree individuals
taller understorytree individualsmore shrubltreeindividualstaller shrubltreeindividuals
more shrubindividualstaller shrubindividuals
taller shrub/climberindividualstaller climberindividualsmore herbindividualstaller grassindividuals
more Chromolaenaindividuals
taller Chromolaenaindividuals
Figure la. Chi-square analysis of species and life form association.(*:significant at p > .01; **: p > .001; ***: p > .0001)
20

in the absence of:
thereare significantly:
more highforesttree individualsmoreshrubltreeindividualstaller shrubltreeindividualsmore shrubindividualstaller shrubindividualsmoregrassindividualsmore Chromolaenaindividuals
* **
* ***
*Figure lb. Chl-square analysis or species and life form associations. (*: slgnlftcant at p > .01 level;**: p > .001; ***: p » .0001)
21
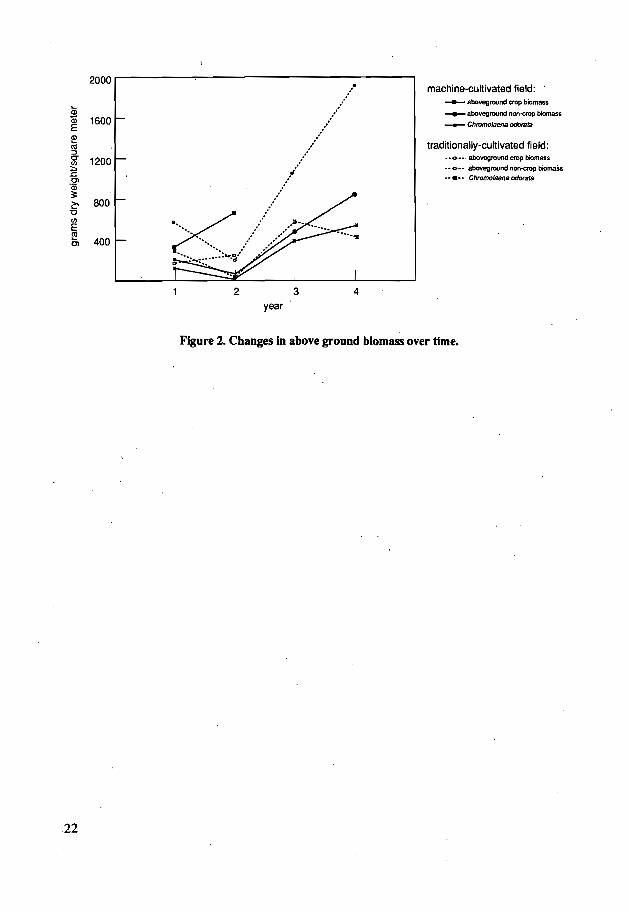
2000
QiQ) 1600E~CIl::JCT 1200(/):;:,s:Cl
"Qi3:e 800'o(/)
Ef!l 400Cl
machine-cultivated field: "___ aboveground cropbiomass
.......- aboveground non-erop biomasa
-...-- Chromolaena odorata
traditionally-cultivated field:·"0··· abovegmundcrop biomass.... 0·. aboveground non<rop biornaSl........ Chromola9flBoc:JoratB
2year
3 4
22
Figure 2. Changes In above ground blomass over time.

CHROMOlAENA ODORATA AND DISTURBANCE OF NATURALSUCCESSION AFTER SnIFfING CULTIVATION: AN EXAMPLE
FROM MAYOMBE, CONGO, CENTRAL AFRICA
H. de FORESTAORSTOM - BIOTROP,
P.O. Box 17, BOGOR, INDONESIA.
D.SCHWARTZORSTOM , B.P. 1286,
Pointe-Noire, Rep. Pop. Du Congo
ABSTRACT
In Mayombe rain forest, shifting cultivation is the only cultivation system used by localpeople in order to produce cassava, their staple food. Fields are let to natural fallow after abouttwo years, when all tubers have been harvested. Until now, abandoned fields were. rapidlyovergrown with fast growing trees like Musanga cecropioides, which within a few years madeup the infrastructure ofa real forest, 15 to 20 meter high. This pioneer forest constituted the firstphase of a succession leading to the rebuilding of the rain forest. But in fact, such a successioncould not occur, as fallow vegetation is usually cut by peasants after 5 to 15 years and enters anew phase of cultivation. Chromolaena odorata, for which local names account for its recentarrival and its striking invading power, is disturbing at an increasing rate this alternate cycle ofcultivation and tree fallow: in abandoned fields after slash and burn of pioneer forest, regrowthis now made up almost exclusively of dense thickets of C. odorata.Preventing the intrusion ofpioneer tree species, these thickets stop the natural succession for many years and are thusbecoming the main vegetation type near villages. This new expanding feature of a traditionalcultivation system is usually considered by local people as a threat; nevertheless, based uponvarious field observations, suggestions are made here about the potential positive effects of C.odorata fallows in shifting cultivation systems.
INTRODUCTION
The neotropical shrub Chromolaena odorata(L) R.M. King and H. Robinson is perceivedby agriculturists in two contradictory ways: for most of them, it is one of the worst weeds in theworld, while for a few others it is a plant with promising agronomic properties.
As in many countries of humid tropical Asia and Africa, c. odoratacan now be found inlarge quantities in the Congolese Mayombe. Its expansion may be compared to a true invasion,and in some areas C. odorata thickets already make up the larger part of the rural landscape.
23

Thepresentpaper isanattempttoanswer thefollowing mainquestionsposedby thisveryquickand intensespreadregardingtheprocessesinvolvedin C. odoraia invasionand its impacton natural and cultivated vegetation:
Which vegetation types.are replaced' by C. odorata, how are they replaced, and at whatspeed? '
What could be the future extensionof C. odoraia in Mayombe?
Whatare the presentandpotentialeffectsof this invasionon cultivation,and whatare theagronomicpotentialitiesof invadedareas ?
THE MAYOMBE MASSIF: SITUATION
The Mayombeis a lowaltitude hill range, lyingparallel to the Atlanticcoast fromGabonat theN.W.toZaireat theS.E(figures1& 2).Thehighestridgesare situatedinCongo,andreach900m. Owing to its vigorous Appalachian relief, and despite its low altitude, the Mayombemassif forms a real obstacle between coastal and inland Savannah plains. In the CongoleseMayombe,one maydistinguisha westernpart formedby a set of hillsand ridges gently slopingdown to the coastal plain, and an eastern part with a regular succession of high sandstonequartzite ridges intersected by schist valleys,which finallydips steeply to the large Niari andNyanga valleys (Schwartz,et al. 1991).
!
Therainfallincreasesfromthewest(1200mm/year)towardsthehighestridges(1980mm/year), thendecreasesto low levelsagain (1200mm/year)at theedge of theNiari plain. Rainfallisdistributedover8 months,fromOctobertoMay,butmistanddrizzleare veryabundantduringthe so-called dry season, allowing the preservation of a certain soil humidity. The amount ofsunshine is very low, especially during the dry season, totalling less than 1000 hours/year(Clairac,et al. 1989).Given thelowrainfallcitedabove,thesefeaturesof thedry seasoninsouthCongo are very important,as they allow the presence of rain forest, as well as its tendency tocolonizesavannahs(de Foresta 1990).
Soilsare mainly"ferrisols"(Duchaufour1983):soilssubjectedto ferralliticalteration,butin which still remain,owing to their relatively youngage, unalteredprimary minerals such asillites or micas (Gras 1970,Jamet 1975 • 1978;Lanfranchi& Schwartz, in press). These soilsgenerallypossessgood physicalcharacteristics but are chemicallypoor, highlydesaturatedandstronglyacid, with a very low surfacepH between3.5 to 4.5 (Misset 1989). In about 80-90 %of the soil cover in Mayombe, these soils contain a 5 cm to 2 m depth horizon showing aconsiderableenrichmentin residualroughelements,whichcan be a realobstaclefor agriculturewhen situated at low depth. This stone-line, generally located between the B horizon and thealteration horizon, can be foundat extremelyvaryingdepth, from0 to 6 m, but usuallybetween0.5 and 2 m. '
24

.vegetanoniSrfiarkedlydominatedby evergreenrainforest,witha lessertendencytowardssemideciduousrain forest(White 1983).To the west,onepassesfrom the Mayomberain forestto the coastal savannahsthrough a very progressivemosaic;to the East, the transitiontowardsthe Niari savannahsis very sharp and occurswithina few kiloineters(figure 3). Nevertheless,the forestcover is sprinkledwith numerous includedsavannahs, whosedistribution,originanddynamicsarebeginningtobebetterunderstood (deForesta1990,Schwartzet al. 1990,Schwartzet al. 1991).The Mayombeforest is at present highlydegradedby agriculturearound villages,and also in other parts,owing to an already long periodof logging (Giben & Senechal 1989).However, despite a long history of human occupation and migration, traces of which aresometimes encountered, undisturbed or almost undisturbed forest still occupies large areas(Cusset 1989).
TheCongolesepartofMayombewasverypoorlypopulateduntil theconstructionof therailway"Congo-Ocean"; after its opening to traffic in 1934, the population increased continuously,reachingabout 30, 000 inhabitants in 1987,still a meandensityof only 4 inhabitants/krn2. This .populationis irregularlydistributed, with half livinginasmallnumberof "village-gares"alongtherailwaylinesin thecentraldistrictofMvouti. This district,inwhichdensitystill reachesonly5.8 inMcm2, has seen a threefoldincreaseof its populationduring the last 50 years, and is atpresentthemostpopulatedruralareaof thewholeCongo.However, from a districtarea of 3,200km2, only 900 km2 are actuallyusedby man for plantationsor shiftingcultivation,2,300 km2of rain forest being left almost undisturbed (Boungou et al. 1989).
LES SARAS: THE ZONING OF VEGETATION AROUND A''VILLAGE-GARE''
After a comprehensive surveyalong the maintrafficaxis crossing the Mayombemassif,the "village-gare" Les Saras was chosen as a model for a study on the evolution of naturalvegetationsubjectedto localagriculture, aimingat the assessmentof both the impactof humanactivitieson natural forestregeneration and the constraintsimposedby naturalvegetationupon.thecultivation systems.Les Saras,locatedin themiddleof thecentralMvoutidistrict,alongboththe Congo Ocean railway and the NI road joining Brazzaville and Pointe-Noire(figure 3), isprobably the mostactive rural areaof Congo in termsof agriculturalproduction (Mapangui&SenechalI989).
Agriculture, whichoccupiesabout 75% of the workingpopulation,is devoted mainly tothe cultivation of sweet banana (mainly the "Gros-Michel" variety) and cassava (tubers andleaves),whichcrops are conveyedby train to be sold in Pointe-Noire and Brazzavillemarkets(Kouba1987). However, thelargerpartof cassavatuberproduction (by far the mainstaple foodin Congo) is still locally used for household consumption.
Because of the lack of aerial photographs, the zoning of land use was studied along sixtransects following roads, railway, and small tracks; the village itself is characterized by anabundanceof fruit trees, dominatedby the"safoutier" (Dacryodes edulis), whichare cultivatedby almostevery householdin welldevelopedhomegardens.Next to the village, the landscape .
25

appearsas a complexmixtureof cultivated fields and naturalfallows; the combination of twokindsof criteria,namely theproportion of each type of naturalvegetation and theproportionofeach typeof agricultural plantation, allowsthecharacterization' of four mainland usepatterns,globallyarranged in concentric rings from villagefringes to undisturbed forest (figure4):
A : a mosaic of veryold, moreor lessabandoned bananaplantations, oilpalmgroves,C. odorata bushes,and a few cultivatedfields makeup the first vegetation belt. Asin the secondbelt, fieldsare verysmallin area (about400 to 600 m2),alwaysdevotedto an'association of maize,groundnut and cassava,whichis sometimes extended toothervegetables suchas eggplantor spicessuchas chilli,andare abandoned to fallowafter one year. .
B :the secondbelt is mainlyoccupiedby C. odorata thickets,again withsomeoilpalm grovesas well as somecultivatedfields; a few scatteredprimaryforest trees(veryhard timbertrees belonging mainly to the family Irvingiaceae) recall that this areawasonce occupied by rain forest.
C: the thirdbelt is characterized mainly bya mosaic of pioneertree fallowsat variousstagesof development and moreor lesswellmaintained bananaplantations,withsomesmallpatchesof "primary" forestas wellas a few cultivatedfields and C. odoratadominated bushes.Here,fieldsare larger thanin A and B (about I ha.),.almost exclusively devoted to cassava,and gradually abandoned to fallow after one to twoyears.
D : the fourth belt formsa progressive transition zone, with new cassavafieldsand .bananaplantations, progressing at the expenseof the undisturbed forest.
Although theextension of eachvegetation beltcan varyfrom one villageto another, theglobal distribution pattern of natural and cultivated vegetation around Les Saras may beconsidered as fullyrepresentative of thesituation prevailing along theCongo-Ocean railway inthe Mvouti 'district, ..
From this schematic description of the rural landscape around Les Saras, one may.appreciate theimportantplacealready occupiedbyC. odorata, especially nearthevillage(AandB belts). However, thisdescription onlygivesan accountof thepresentsituation,and, with thehistoryof rural landscape and agricultural practicesas a reference, some information about thedynamic processes of vegetation evolution is neededto explain thispresent importance and toforecast its future.
THE HISTORICAL BACKGROUND OF C. ODORATA INVASION
The history of the Mayombe massifwas greatlyaffectedby the opening of the CongoOceanrailway. Therailwayapproximately follows thelayoutofanold foresttrack,theso-called"piste descaravanes", whichpreviously joined thecoastaland the interiorpopulations. DUring
26

thebuildingworks,from 1922to 1934,thecentralMayombe sawan intenseand tragicactivity:becauseof thefairlylownumberoflocalinhabitants, thecolonialadministration ordered"forcedwork" for local people and recruited about 40,000 people from Oubangui-Chari and Tchad(Nguembo and S~nechalI989). Sanitaryconditions were very bad and, within 13 years,about20 000 peopledied.
With the completion of the railway,mostof the foreigners left but localpeople stayed inthe newlybuilt "village-gates", whichbecameimportantimmigration centers for populationsfrom insideand outside Mayombe(Vennetier 1968).
The openingof this newcommunication axis led to a newerafor forestexploitationandcommercial cultivation. Largebananaplantations ownedbyColonists appearedasearlyas 1936,and extendeduntil 1961; with the independence of the countryin 1958,bananacultivationfortrade was resumedon old or new plantations by local people, and to the present this crop.stillrepresentstheir mainsourceof income. From 1934to 1956,as firewoodwas the main fuel forrailwayengines,largetractsof primaryandsecondary forestwerecut alongthe railway(Gibert& Senechal 1989). Moreover, the railwayopeningallowed the expansion of selective timberexploitation, whichbecame themajoreconomicactivityofcentralMayombe until 1974(Gibertand S~necIuil1989), themostimportant presenttracesof whichlie in theroadnetworkcenteredon some"village-gares".
Untiltherailwaybuilding, thevillages weresmallandfew;theywereoccupiedfora certainnumber of years and then shifted to other places following events such as the death of animportant memberof thecommunity orepidemicdiseases; agriculture wasexclusivelydevotedto shifting cultivation for household purposes, with small temporary fields (about 0.5 ha.)scattered inside the undisturbed forest and linked togetherand to the village by small pathsrunningunderthe forestcover.
Since 1934, the railway and the associated NI road, as well as the concession roadsnetwork, gradually provideda meansof forest penetration, allowinglocalpeople to graduallycolonizenewareas for cultivation.
From an ecologicalpoint of view, two main transformations have been brought by therailwayopening:
- local intensiflcations of human impact upon vegetation, with the introduction ofpermanent plantations ana the expansion of cultivated fields - fields which are no longerseparated by tractsof undisturbed forestbut only by variousstages of fallow vegetation;
- expansion of more or less wide strips supporting only a heavily disturbed and easilyinvadedvegetation, a directconsequence of the settlingof roads and railway.
These transformations obviously paved the way for the invasion of C. odorata.
According toCruuwellMcFadyen (1988),C.odorata appearedfor thefirsttimeinCentralAfricaaround 1940,withNigeriaas the firstaffectedcountry; it becamea major weed therebythe late 1960's, and since then has spread to Ghana, IvoryCoast and Cameroon.
27

Although information aboutits arrivalin Congois lacking, according to localpeople,C.odorata, which is called locally "matapa mbala" (the invader) or "Lantana Ngouabi" (acombination of the previously importantcolonizingplant and the name of.a previous Congopresident), arrived in Mayombe during the early 1970's after a rapid progression along therailway. From then on, it has been gaining moreand more importance in the rural landscape,owing to its biological properties.
C. odorata is a fast growing heliophilous shrub, reaching sexual maturity after a fewmonths; Probablyowingto the constantly highrelativehumidity characterizing the Mayombe
. climate, C. odorata shrubs never wilt on the stalk, even during the dry season; branchessupporting flowers and fruits die, but are quicklyreplacedby new ones.
-In Mayombe, C. odorata flowers and fruits during the dry season, from July to midSeptember, releasingeach yearan enormous numberof small,wind dispersedseeds; althoughrecently releasedseedscangerminate in heavy shade(Marks &Nwachuku1986),thegerminationof soil embedded seeds requiresthered/farredlight ratiocharacteristic of open vegetation(Erasmus & van Staden 1986).
As it has already been reported by some authors, C. odorata seeds can make up animportant proportion of thesurfacesoilseedbanks(Yadav&Tripathi 1982,Epp 1987).AroundLes Saras', although absent from the large areas of undisturbed forest which surrounds theagricultural landscape, viable C. odorata seeds appear in the small remnantsof undisturbedforestlocatedin the"C" belt,as wellas inold pioneerforest,althoughtheretheyare still largelyout numbered by pioneertree seeds (TableI).
In the"parcelleLikibi",the history of whichisdescribedbelow,the thicketalreadyshowsthe tremendous dominance of C.odorata withinthesoil seed bank,withmore than 1000seeds/m2 after only two years of fallow. This number still increases in the fallows that followcultivation on C. odarata thickets located in the "BOO belt, reachingabout 2000 seeds/m2 twoyearsafter the abandonment of the field.
THE LINKS BElWEEN SHIFfING CULTIVATION ANDC. ODORATA
Acomparative surveyof thevegetation associated withthetwomainagricultural systemsshowsthat theinvasion ofC.odorataiscloselylinkedtoshiftingcultivation,bananaplanrationsshowing only rare and minorsignsof disturbance.
I Six plots with different ~egetation types were tested. In each plot, 8 sites were selected 81 random and in each site. 6elementary surface soil samples were taken using a 3.2an radius and 8em long cylinder. and mixed together. Samplestaken in Les Saras on February 10. 1989, were brought to Pointe-Noire the same day. The next day they were placed ona 2cm layer of sterilized soil in germination trays. These 48 germination trays (6 plots, 8 samples perplot), placed under
slight shade and a plastic cover to protect from rain were watered every day and checked bi-monthly for new.seedlingsfor 6 months, i.e, until August 4, 1989. After each counting identified seedlings were removed and the others were
mapped.
28
i. I
/
/

Bearing in mind the main biological features linked with the settling and growth of C.odorata, four main reasons, linked to localagricultural practices, may account for the relativepreservation of bananaplantationsfrom C. odoratainvasion.
According to local people, the best soils for banana cultivation are those supportingundisturbed forest; thereforenewplantations are almostexclusively opened on primaryor oldsecondary forestlands,areaslocatedat thefringesof thecultivatedzone,whereC.odorataisstilllacking,even in the soil seed bank.
Usually, some tall trees are preserved during the forest clearing, and banana trees aretherefore planted extensively under light shade, in microclimatic conditions which are notfavorable to the growthof C. odorata.
The cleared forest is not burnt,allowing, betweenthe banana trees,a rapid recoveryof adensecover of pioneertrees,lianas,and forestherbswhichhamperan earlycolonizationby C.odorata. .
Owing to periodiccleanings (usually once a year), fast growing speciesable to supportrepeatedcuttingareprogressively favoured; theregrowth associated withbananatreesevolvesquicklyandreachesitsequilibrium assoonas thefifthyear,stayingunchanged oralmostso untilthe abandonment of plantation (the oldestproductive plantations are about 30 yearsold). Thisstablevegetation is dominated by largerhizomatous herbsbelonging to theMarantaceae family(especially Thaumatococcus daniellil) andby ferns(mainlyCyclosorus ofer); theseplantsglowvigorously aftereachcleaning,restoring withina fewweeksa verydensecover,whichprotectsplantationsfromany new settlingof C. odorata.
Incassavacultivation, on thecontrary,in everydry seasonalmostevery household opensa new field and abandons an old one, leaving every year large areas with almost bare soilsusceptible to invasionby C.odorata; in addition,moreand morepioneerforest and C. odoratathickets (with their high number of soil embeddedC. odorata seeds), are now used for thistemporary cultivation.
C.ODORATA AND THE DISTURBANCE OF NATURAL SUCCESSION
At present, the general trendsof naturalsuccession following slashand burn cultivationof fieldsopenedon undisturbed foreststill follow theclassicalschemeencountered everywherein the humidtropics(Richards 1952,Halleet al. 1978,amongothers).
The abandoned fields are invadedfirst by herbs whichachieve their cycle within a fewweeks, and are then quickly replaced<by a few pioneer tree species, almost always largelydominated in Mayombe by Musanga ceeropioides.
Two yearsafter theabandonment, the fallow makesupa real forestwithan alreadydense
29

canopy,some8 to 10m high;as thenumberof speciesincreases gradually (Table2), owing tothe settling of sciaphilous speciesunder the alreadydense canopy: the relative importanceofpioneerspecies, estimated by suchparameters as relativedensityor relativebasal area, slowlydecreases until theprogressive deathof the uppercanopy layerafter about 20 to 25 years.
The structure of thispioneerforestevolveswith time,mainlybecauseof natural thinningand individual tree growth,and maybecharacterized as earlyas thesecondyear by twolayers:an upperone madeupalmostexclusively ofMusanga treesculminating at about15mheightandquicklystabilizing at around22-25 m,and a lowerone showing large floristic variations fromsite to site,but mainly dominated by someRubiaceae andsomeMacaranga. Thisundergrowth,inside which even after 16 years of fallow the constantpresenceof cassava should be noted,densely occupiesthe space from the groundlevel to variousheights,from 3 to 8 m dependingon its localcomposition, and has its development largelyhampered by the Musangacanopy.
In thepast, 10to 15yearsoldfallows wereoftenchosenfornewcultivationowing to theirrelativelyeasyclearing,starting thena newandalmostidentical succession. Whenvillages,andwith them theircultivated areas,wereabandoned, the succession could extend:after the deathof theMusanga canopy,the pioneerforestwasreplacedby a secondary forest which itself ledaftera moreor lesslongperiod(100-150 yearsor more?) to theregeneration ofa"primary" forestfloristically and structurally similar to the initialone.
InMayombe, as wellas in mostpartsof thehumidtropics,thisclassicalsuccessioncannolongerbe achievedas fields are reopened after lessand less years, owing mainlyto permanentsettling of villagesand to increasing population densities. Fallowsof shorter duration lead tofloristic and structural transformations, favouring speciesable to reach maturityearlier and todeliverseeds in greateramounts. In Mayombe, although other tracesof past shortening in thefallow-field cycle can be' encountered, as Pteridium aquilinum or Dicranopteris linearisformations, the most obvious evidence of the increasing cultivation intensity is given by theoccurrence oflarge tractsof C. odoratadominated thickets.
At present, when a plot of pioneer forest, whatever its age and previous vegetation("primary", secondary or pioneer forest), is opened for cassava cultivation, the colonizing.vegetation following the abandonment of the field is largelydominatedby C. odorata.
This processhas beenclearlyshownon the"parcelleLikibi":half of the field was set upon a previously undisturbed forest, the other halfon a five yearsold pioneer forest succeedingdirectlyto undisturbed forest. Aftertwoyearsof fallow, theregrowth on thefirsthalf is a typical.youngpioneerforest8 to 10m high, witha treedensity of 7650indlhaanda basal area of 25 m2(Table2); on the secondhalf, thefallow is madeup of a fewscatteredpioneer treesemerging3 .to 5 m above.a2 m high thicket. Although this thicketpresentsa relatively importantspeciesrichness (47 specieswerecountedon a 25 m2 plot), C. odorata is by far the dominantspecies,as shownby itsaerialbiomass'accounting for 57 % of the totalabovegroundphytomass of thethicket(Table3).
2 Biomass was used as the more meaningful parameter to assess the relative importance of species or groups of speciesin C. odoraia thickets. In each fallow, a 25m2 representative plot was chosen, in which species or groups of species were
cut to ground level, sorted and weighed on the spot, Samples were then brought to the laboratory and dried in an oven
at 105 C for constant weight to assess the dry/fresh weight,
30

The importance of C. odoratawithin the regrowth is naturally much more pronouncedwhenthe vegetation slashedandburntforcultivation isalreadydominated by C.odorata (Table3):afteroneyearof fallow, the2 mhighthicketis mainlycomposedofC.odorata(only8specieswerecountedon25 m2),whichaccountsforabout67 %of thephytomass (treesexcluded).Afterthreeyears,thevegetation isstillverydenseandspeciespoor(only10speciescountedon25m2),reaching2.5 to 3 m high, with a very few slightlyemergingor completely included trees. C.odoratahereaccountsfor about 76 % of the biomass(treesexcluded).
In the oldest thickets found around Les Saras (7 years old), although some signs ofevolutionmay be observed, such as the differentiation of a few tree spots (about 10 % of thefallow area) and the disappearance of C. odorataunder these spots,C. odorata is still largelydominant, accounting for 84 % of the phytomass (Table3).
The comparison of the previous examplesof C.odoratathicketsshowsthatonce settledtheystayalmostunchanged.for years,and probably becauseof theirextremely densecanopybutperhaps also because of the allelopathic properties of C. odorata (Ambika and Jayachandra1980), theydo not allow the growth of other successional species,therefore makingup a kindof successional locking. This locking is obviously long lasting, but one may foresee a slowevolutionof thicketstowards impoverished kindsof pioneerforest,with the supportof the fewsurviving trees, which will after some years create many unsuitable spots for C. odorata.However, this foreseeable succession is not likely to occur in practicebecause fields are nowreopened after fallow periodsof only5 to 6 years,not longenoughto allow suchan evolution.
C.ODORATA: CONSEQUENCES OF ANtNvASION
The zoningof vegetation and land use aroundLes Saras as described above providesanaccountof theagricultural landscape and of the relativeimportance of C. odorata as it existedat the timeof study in 1988-1989. .
From the railwayopeninguntil the early 1970s, this landscape has undergoneimportantbutmainlyquantitative transformations: expansionof theareaundercultivation, and increaseof
.pioneerforest fallow areas at the expenseof the undisturbed forest
From the arrival of C. odorata until now, huge qualitative transformations began to besuperimposed on this trend: appearance of long lasting low thicketsat an increasing rate andreduction of areas supporting a forestcover,whatever its composition.
Bearingin mind the presentnatural succession trends, it.seemsobvious that C. odoratathicketsare boundto extend,and it may be assumedthat, except for a narrowstrip of pioneerforestand cassavafieldsalong the undisturbed forestborder, the wholeagricultural landscapewillbe reducedwithina few yearsto one large-area of C. odorata thickets,only locallysharedby bananaplantations and cassavafields.
This completely new process of replacement of classical pioneer forest fallows by C.
31

odorata thicketfallows is clearly visibleon the ground,and is perceived by rural people (whoindeedhavea ringside seat)asextremely frightening. Thefirst attemptsofcultivation,followingthe traditional slashand burn practices, ended in completefailure. Although they may fullybeexplained by the vigorous resprouting properties of C. odorata, manypeople attributed thesefailures to a substantial decrease in soil fertility, and thesepeoplenow believethat soils underC. odorata thicketsare lost for agriculture.
. However, thisbadreputation couldslowlychange.As reported above,C.odoraia thicketsnext to the village, in the A and B belt, are nowmoreand more used for cultivationof maize,groundnutandcassavainmixture. Suchfieldsareoftencompletely managed bywomen,butalsosometimes by menwhodo not"own" anyforestfallow or whoare too old to havetheirfield farfrom the village; they are usually very small, and only constitutesecondary fields. Providedcultivation operations arecorrectly undertaken, suchas the removing of C. odorata stumpsandcarefulweeding, these fieldscan yield successful crops.
Wehad theopportunity of following oneof thesefieldsfrom itsopeningduringthe 1988dry season to the cassava dominated stage, just before the abandonment to fallow. All crops(mainly maize, groundnut, eggplant. chilli and cassava) were vigorous and healthy, and,although it was not possible to undertake any quantitative study, their productivity could bejudgedas fairly high.
A comparative study of soil characteristics in fallows of variousage, structure, floristiccomposition, andhistory, wascarriedoutatLesSaras',Nosignificantdifferencesinsoilnutrientcomposition betweenundisturbed forestandpioneerforestfallows (HFDS 16, 10, 12, 11)were .observed. butthisstudyclearlyshowsanimprovementof soilchemical fertility underC.odoratathickets (Table4).
Two years after the abandonment of a field opened on pioneer forest fallow ("parcelleLikibi": HFDS9), the soil fertility is still similarto undisturbed forest soil fertility, but after 7years(HFDS 14)the nutrientenrichment appearsclearly,mainlywithan increasein calcium,whichis well reflected by a tremendous improvement in pH (2 points!).
Thisglobalimprovementof soilchemicalfertili tydoesnotappearto beaffectedbyfurthercyclesof cultivation/fallow, as maybededucedfrom the soilanalysisofa three yearsold fallowfollowing cultivation of a fieldopenedon C. odorata thicket (HFDS 13).
In addition, it should be noted that in the two C. odorata thickets (HFDS 13 and 14),contraryto theother vegetation types,the soil wascharacterized by intensebiological activity,shown by the important humus horizon depth (15 cm compared to about 3 or 5 cm underundisturbed or pioneerforest) and by the abundance of earthworms..
3 Surf~ soil samples (G-IOcmdepth) were taken in all vegetation types lit half slope. Each sample was made up of16 elementary samples taken within a 100m2 area (lOm x IOm) according to a systematic sampling in which the firstsample was chosen at random. and then mixed together for further analysis. In addition. soil profiles were studied in
. each vegetation type. In order to assess the evolution of soil characteristics under various treatments, each fallow type
was associated with the nearby undisturbed forest where soil samples were taken using the same methodology.
32

Thecombination of theseresultswith theaboveexamplesof successfulcultivationshowsthat a shifting cultivation system using C. odorata fallows could lead to a global and stableimprovement ot soil fertility, and that C. odorata thickets in Mayombeare not unsuitable foragriculture,providedthat peasantsadapt their cultivationpractices slightly to the new conditions:the thicketshouldbeslashed.leftto dry forsomedays,andburnt;theremainingC.odoratastumpsshouldbe uprooted. As it seems thatmostof the soil embeddedseeds are not destroyedby the high temperatures associatedwithburning' (Table5), an early weeding,followed or not,dependingof thecomposition of crop mixture,byperiodical onesare needed in order topreventthe growthof C. odorata seedlings. .
CONCLUSION
From its arrival in Mayombe in the early 1970's until now, C. odorata odorata hasdrasticallytransformed theagricultural landscape,whichwill probablybe completelyinvadedby C. odorata thickets within a few years. The analysis of natural successionafter slash and9cultivationshowsthatC.odorata invasion hasledto a heavydisturbanceof the"classical"cycleknownby localpeopleforgenerations, withthereplacement of thevariouspioneerforeststagesby stableC.odorata thickets. The incapacityof villagersto controlthis new process,associatedwiththe speedaswellas theintensity of theinvasion,hasledto anunderstandableanxietyamonglocal people, expressed by the overall opinion that C. odorata thickets are unsuitable foragriculture. .
It shouldbe stressedthat the shorteningof fallowperiods in shiftingcultivationsystemsusually leads to an increasingsoil degradation. Contrary to this very general trend, under C.odorata thicketssoilfertility seemstobefairlywellimproved, althoughmorestudiesareneededin order to confirmand to explain our data.
In the framework of slash and bum cultivation, the soil improvement exhibited by C.odorata fallowsseems to be stablewith time,a fact whichcould be of greatvalue for the futureof agriculturein humidtropicalAfrica,allowing,withinthe traditional systemsandwithoutanyinputs, a process of agricultural intensification. Assuming that field yields are similar in a ISyearscyclebasedon pioneerforestfallow and ina 2 yearscyclebasedon C. odorata fallow,theglobal production over time in the secondsystemwould be five times higher!
IndeedC.odorata shouldberecognizedas a weed,butonly fora fewgivencropsandonlyundergivencultivationsystems. In Indonesia, for instance,C. odorata has been reportedas aweed mainly for rubber, but although rubber estates seem often to be invaded, smallholders'"jungle-rubber"plantationsare never hampered.
4 Six distant surfacesoil samples, each made up of IQ elemental)' samples taken with a 3.2cm long and 8cm radiuscylinder, from a 3 years old C. odorata fallow were brought to the laboratory. They were mixed together and dividedin 6 equal pans to be subjected to various temperature oonditions. Then. they were placed in germination trays and
checked for new seedlings using the same method as in the seed bank assessment experiment
33

Considering the present distribution of C. odorata as well as its foreseeable futureextension(CruttwellMcFadyen 1988), it seemsurgentnot only to findbetterways 10eradicateand tocontrolC.odoratawhereit appearseffectively as a weed', but also to assess itspromisingagronomicpotentialities inorder 10useC.odoratain itsbeneficialaspectsfor thebenefitof ruralpeople.
ACKNOWLEDGEMENTS
This study' was in part sponsored by UNESCO, which provided financial aid andequipment(binocularmicroscopeandoven),intheframework of theDGRST-Congo,UNESCO,PNUD joint "Mayombeproject". Our grateful thanks go 10 the staff of ORSTOM center inPointe-Noire, as well as to Les Saraspopulation, especially to Mr. MassangaJoseph who wasan invaluableguide throughoutthis study. '
REFERENCES
Ambika, S. R. & Jayachandra, 1980.Influence of light on seed germination in Eupatorium. odoraium L.Indian Forester. 106:637 - 640.
Boungou, G., 1. Nguembo & 1. Senechal, 1989.Peuplement et populationdu Mayombe. Revue desconnaissances sur le Mayombe. DGRST-Congo, UNESCO et PNUD. UNESCO, Paris. 157 185.
Clairac, B., B. Cros & 1.Senechal, 1989.Le climatdu Mayombe. Revue des connaissances sur le Mayombe.DGRST-Congo, UNESCO et PNUD.UNESCO, Paris. 47 - 78.
Cruttwell McFayden, R.E. 1988.History and distribution ofChromolaena odorata (L.) RM. King and H.
Robinson. In: R. Muniappan (Ed.) Proc. First Intern. Workshop BioI. Control Chromolaena
_ odorata. pp. 7-12, Agri. Expt. Sta., Univ, Guam, Mangilao, Guam.
Cusset, G. 1989.La flore et la vegetation du Mayornbe Congolais: etat des connaissances. Revue desonnaissances sur le Mayombe. DGRST-Congo, UNESCO et PNUD: UNESCO, Paris. 103- 136.
de Foresta, H. 1990.Origine et evolution des savanes intramayombiennes (R. P. du Congo). n. Apportsde la botanique forestiere. Pays ages quaternaires de I' Afrique Centrale atlantique. R. Lanfranchiet D. Schwartz. ORSTOM, Paris. in press.
Duchaufour, P. 1983.Pedologie: classification.et pedogenese. Masson, Paris. 1: 1 - 493.
Epp, G. A. 1987.The seed bank of Eupatorium odoraium along a successional gradient in a tropical rainforest in Ghana. Journal of Tropical Ecology. 3: 139 - 149.
Erasrnus, D. 1.& 1.van Staden, 1986.Germination oi Chromolaena odorata (L.) K. & R. achenes: effectof temperature, imbibition and light. Weed Research. 26: 75 - 81.' -
34

Gibert, G. & 1.S~ochal, 1989. L'econornie forestiere. Revue des connaissances sur le Mayombe.DGRST-Congo, UNESCO et PNUD. UNESCO, Paris. 249 - 293.
Gras, F. 1970. Surfaces d'aplanissement et remaniement des sols sur la bordure orientale du Mayombe,Cahiers ORSTOM, serie Pedologie. 8: 274 - 294.
Halle, F. , R. A. A. Oldeman & P.H. Tomlinson, 1978. Tropical Trees and Forests: an architecturalanalysis. Springer-Verlag, Berlin. 1 - 441.
Jamet, R. 1975 - 1978. Pedogenese sur roches cristallophylliennes et argileuses en milieu equatorial. congolais. ORSTOM, Rapport interne.
Kouba, B. 1987. Les problernes de production et de commercialisation de la banane douce dans la regiondu Mayombe. Cas du district de Mvouti, Institut du developpement rural. Universite Marien~gouabi. Rapport de fin d'etudes,
Lanfranchi, R. & D. Schwartz, 1991. Les remaniements des sols pendant le Quatemaire superieur auCongo. IT: Evolution des paysages dans la region du Mayombe. Cahiers ORSTOM, seriePedologie, in press.
Mapangui. A. & 1.Sblechal, 1989. L'agriculture. Revue des connaissances sur le Mayombe. DGRSTCongo, UNESCO et PNUD. UNESCO, Paris. 217 - 233.
Marks, M. K. & A.~. Nwachuku, 1986. Seed bank characteristics in a group of tropical weeds. WeedResearch. 26: 151 - 157.
Misset, M. 1989. La pedologie du Mayombe. Revue des connaissances sur le Mayombe. DGRST-Congo,UNESCO et PNUD. UNESCO. Paris. 85 - 92.
Nguembo.J, & 1.Sblechal, 1989. Lestransports. Revue des connaissances sur le Mayombe. OORSTCongo, UNESCO et PNUD. UNESCO, Paris. 295 - 313.
Richards, P. W. 1952. The Tropical Rain Forest: an ecological study. Cambridge University Press,Cambridge. 1 - 450.
Schwartz, D., H. de Forests, A. Manoui, J. P. Massimba, & C. Girardin. 1991. Dynamique actuelle ducontact foret-savane dans le Mayombe Congolais. Approche pedologique, botanique etbiochimique. in press.
Schwartz, D., R. Lanfranchi, & A. Mariotti, 1990. Origine et evolution des savanes intramayombiennes(R. P. du Congo). I: apports de la pedologie et de la biogeochirnie isotopique. Paysagesquaternaires de l'Afrique Centrale atlantique. R. Lanfranchi et D. Schwartz. ORSTOM, Paris. inpress.
Vermetier, P. 1968. Pointe-Noire et la facade maritime du Congo. Memoires ORSTOM. ORSTOM, 26:1 - 458.
White, F. 1983. The vegetation of Africa. Maps & Memoirs. UNESCO AETFAT, UNESCO. 1 - 356.
Yadav, A. S. & R. S. Tripathi, 1982. A study on seed population dynamics of three weedy species ofEupatorium. Weed Research. 22: 69 - 76.
35

36
fig 1 Les grands ensembles de relief du Congo
Simplified geomorphology of Congo
1. Plarne conere - Coastal plain
2. Mayombe (massif)
3. Valtee du Nlari - Niari valley
4. Chaillu (massif)
5. Plateau et colltne - Hill and plateau
6. Cuvette conqolaise - Congo basin
[]l. 3.6
~5
IIIIO] 2. 4
Fig 2 Carte de la vegetation du Congo
Vegetation map of Congo
•Foret dense de terre fermeLowland rain forest
mm Foret dense mondee - Swamp forest
D Savane - Savannah
Source: Geoqraphre de la RepulJllque Poputaire du Congo
Q 100 20QJcm
ZAIRE

o
5°_L-_---,-- ~:i:Z. ____J
savanes cot ieres - coastal savannahs
rnosaique toret-savane de la facade maritimeoccidemal forest -savannah mosaic
[~.~:J torets - rainforests
savanes du svncl inorium Niarl- NyangaNieri- Nyanga savannahs
railway Congo-Ocean
Fig.3 Le Mayombe congolais - Congolese Mevornbe
37

N
1
--- Chemin defer/Railwav
RouteNl/N1Road
Anciennes Pistes Forestieres / Former logging Roads
A. s.C.D.E: See text for detailed composition
Legends for Figure 4:
oI
5.
Zone A: Mosaic of old banana plantations, oilpalm groves, small cassava fields and C. odorasathickets;
Zone B: mainly C. odoraia thickets, with a few small oilpalm groves and cassava fields;
Zone C: mosaic of pioneer tree fallows and banana plantations, with a few cassava fields and C.odorata thickets;
Zone D: new cassava fields and banana plantations at the fringes of the still undisturbed forest;
Zone E: "primary" forest
Fig 4: ZONATION CULTURES ET DE LA VEGETATION AUTOUR DE LES SARAS
LAND USE AND VEGETATION ZONING AROUND LES SARAS
38

Table I: Soli seed bank under various vegetation types(mean number of seedlings »er m2)
Vegetation typesSpecies or species group F1 F1(2) FF CF1 CF2
Chromolaena odorata 0 37 29 1119 2035Ruderal Herbs (20 species) 0 112 702 485 5179Pioneer Trees (16 species) 243 1039 900 506 112Forest Herbs (4 species) 0 8 24 0 0Unidentified (1 species) 83 128 83 37 651
TarAL 327 1376 1740 2147 7970
F1: undisturbed forest far from the villageF1(2): small patch of undisturbed surrounded by fields and fallowsFF: 16 year old pioneer forest fallow after cassava cultivation on undisturbed forestCF1: 2 year old C. odoraia fallow after cassava cultivation on pioneer forest fallowsCF2: 3 year old C. odoraia fallow after cassava cultivation on Crodoraia fallow
Table 2: Main florlstle and structural characteristics of pioneer forest fallows after cassavacultivation (Les Saras, MAYOMBE)
Species orgroup of species FF1 FF2 FF3
N A.B. N A.B. N A.B.% cm2 % % cm2 % % cm2 %
Musanga cecropioides 164 54 9611 96.9174 53 21070 98.136 20 16345 85.1Manihol esculenia 30 10 81 0.8 4 1 14 0.1 13 7 47 0.2Beniera racemosa 60 20 67 0.7 56 17 93 0.4 12 7 64 0.3Ponerandia cladaniha 0 0 0 0.0 22 7 27 0.1 22 12 94 0.5Macaranga spp. 0 3 69 0.7 5 2 50 0.2 7 4 153 0.8Other species 34 11 77 0.8 39 12 138 0.6 46 26 538 2.8Other species 9 3 13 0.1 30 9 88 0.4 43· 24 1960 10.2(resprouts)
TarAL (plot) 306 9918 330 21480 179 19201TarAL (per Ha.) 7650 247950 8250 537000 2983 320016
FF1 (400m2); 2 year old fallow ("parcelle Likibi")FF2 (400m2); 3 year old fallowFF3 (600m2); 16 year old fallow ("parcelle Likibi")
N: number of trees with diameter at breast height higher than 1 cmA.B.: basal area (trees with diameter at breast height higher than 1 cm)
39

Table 3: Above ground Phytomass of C. odoralil fallows (Dry weight) of varlous.C. odoratafallows
C. odorata FallowsSpecies or CFl CF3 CF2 CF4group of species .Kg/Ha % Kg/Ha % Kg/Ha % Kg/Ha %
Chromolaena 5876 36 8320 65 18052 64 13856 84Cassava 1912 12 200 ' 1 1968 7 508 3Pioneer trees 5772 36 520 4 7248 26 604 4Resprouts 604 4 0 0 0 0 0 0Vines 1444 9 0 0 880 3 1480 9Herbs 548 3 3840 30 20 0 120 0
TOTAL 16156 12880 28168 16568
CF1: 2 years old, after cassava cultivation on pioneer forestCF2: 3 years old, after cassava cultivation on C. odoraia fallowCF3: 1 year old, after cassava cultivation on C. odoraia fallowCF4: 7 years old, after cassavacultivation on pioneer forest
Table 4: Main chemical characteristics of surface soils under undisturbed forest, pioneer forestfallows and C. odoralil fallows
pH C N C/N Ca Mg K Na S C.E.C. S!CECNo.HFDS (H2O) % % me! me! me! me! me! me! %
100g 100g 100g 100g 100g 100g
16.0 4 3.62 0.197 18.4 0.81 0.21 0.27 0.23 1.52 35.15 4.310.0 4.2 2.9 0.195 14.9 0.85 0.12 0.26 0.23 1.46 33.03 4.48.1 4.6 2.03 0.169 12.0 0.84 0.27 0.18 0.1 1.39 24.43 ~.7
12.0 4.5 2.3 0.15 15.6 0.42 0.16 0.22 0.03 0.83 14.92 5.512.1 4.3 2.23 ? ? 0.560.25 0.29 0.17 1.27 33.38 3.8
11.0 4.9 1.6 ? ? 1.52 0.27 0.21 0.13 2.12 20.78 10.25.1 4.3 2.05 0.15 13.5 0.52 0.27 0.14 0.03 0.96 12.67 7.6
9.0 4.1 2.15 0.16 13.9 1.15 0.27 0.22 0.12 1.76 31.03 5.713.0 6.3 1.82 0.17 10.6 7.06 0.94 0.27 0.29 8.56 25.53 33.58.1 4.6 2.03 0.17 12.010.84 0.27 6.18 0.1 1.39 24.43 5.7
14.0 ~ 1.94 0.13 14.7 5.71 0.63 0.18 0.27 ~ 22.33 30.44.1 3.8 2.38 0.14 17.50.3 0.1 0.27 0.1 0.77 27.83 2.8
HFDS 8.1; 12.1; 5.1; 4.1; undisturbed forestHFDS 16: 2 year old pioneer forest fallowHFDS 10: 3 year old pioneer forest fallowHFDS 12: 7 year old pioneer forest fallowHFDS 11: 16 year old pioneer forest fallowHFDS 9: 2 year old C. odarata fallow (CFI, "parcelle Likibi")HFDS 13: 3 year old C. odorata fallow(CF2)HFDS 14: 7 year old C. odarata fallow (CF4)
40
Table 5: Effect.of blgb temperature on the solI seed bank under C. odorDla fallow(number of seedlings per m2, two montbs after treatment)
Species 80·c 80·c 80·c 130·c 130·c Noor species group 5 minutes 1 hour 12 hours 5 minutes 1 hour Treatment
Chromolaena odorata 1412 497 60 1431 20 1352Oldendia corymbosa . 5149 5388 2266 5050 497 5189Lindemia numuraliifolia 1869 1889 159 1431 0 2087Ageratum conyzoides 417 755 0 596 0 596Laportea aestuans 636 1491 40 517 20 457Peperomia pellucida 119 80 20 60 o . 119Musunga cecropioides 0 298 0 0 0 60Physalis angulata 179 517 99 139 20 278Paspalumspp. 258 338 99 557 0 258Cyperaceae spp. 338 437 179 179 0 239Asteraceae sp. 1 60 60 20 60 O' 0Phyllamhus sp. 1 20 60 0 99 0 40Phyllanlhus sp. 2 0 60 0 0 0 0unidentified spp. 139 40 0 99 0 40
TOTAL 10974 12565 3101 10636 557 11014
41

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j

OBSERVATIONS ON CHROMOLAENA ODORATA (L.)R.M.· KINGAND H. ROBINSON IN INDONESIA
A. SIPAYUNG, R. DESMIER DE CHENON and SUDHARTO PS.
Research Centre for Estate Crop-Marihat, P.O. Box 37, Pematang Siantar, North Surnatra,Indonesia
INTRODUCTION
The occurrence of Chromolaena (=Eupatorium) in Indonesia which was probablyChromolaena odorata(L.) R. M. King & H. Robinson was mentioned as early as 1912 at Deli,on theeast coastof North Sumatra. In 1862thecultivationof the famous Deli tobacco wasstartedin thisarea. The appearanceof Chromolaena inNorth Sumatra therefore seems tocoincide withthe advent of the first large commercial tobacco, rubber and oil palmplantations at the turn of .
.the century. The weed was either accidentally or deliberately introduced.
Furtherconfmnation of its presence in Surnatra was reported by van der Laan (1914) andBiswas (1936). C. odorata(=E. odoratum) was considered to be a very dangerous alternate hostof the vectorsof tobaccopseudomosaicdisease. InJava, C.odorata seems tohaveappeared laterthan in Surnatra.
C.odorata occurs inSumatra,Java, Bali,Kalimantanandas fareast asLornbokand Timor.(A bibliography of Chromolaena odorata (L.) R. M. King and H. Robinson in Indonesia isappended).
OBSERVATIONS IN· AREAS WHERE C. ODORATA HASESTABLISHED
1.SumatraAlthough C. odorata is widespread in Surnatra, from Aceh to Lampung, there are areas
where this plantdoes not exist due to higher alutude.Isolated regions and in unsuitable soils.
a. Effect of the higher altitudes'During a survey in the mountainsof Surnatra where observations were carried out every
20 km, in the "Bukit Barisan" fromPematang Siantar to Tarutung, Padang Sidempuan, BukitTinggi and Padang, C. odoratacompletely disappears above 1,100 m, after Balige as far asSipahulan near Tarutung. Inside Bukit Barisan, the weed does not even occur at lower altitudes(800 to 900 m) beyond Padang Sidempuan as far as Bukit Tinggi, whereas it exists in Samosirwith its special microclimate at 1,000 m.
43

Alongthe transverse road from the east coast (KotaPinang)to the westcoast (Sibolga),viaPadangSidempuan, C.odoratahasinvaded at leastall theroadsides; thesituationisthesamealong the transverse road from Pekanbaru to Bukit Tinggi - Padang,but only as far as PayaKumbuh.
b. IsolatedPlacesThereisnoC. odoratainwestcoastareasisolated bythehighaltitudeof BukitBarisanand
notlinked bythelowertransversal roadstothewestcoast;suchisthecaseof thecoastalarea southofPadang.
c. Soil typeOn sandysoilsalong the east coast,Chromolaena is wellestablishednorthand southof
MedanasfarasTanjung BalaiandBandarAcehrespectively. However,C.odorataseemsunableto developonpeatsoils,suchas in RiauProvince: Rengat,Tembilahan, SungaiGantung,PulauBurung, whereother weedssuchas Mikania totallyinvaded veryquicklyafterdeforestation forthe newoil palmand coconutplantations.
The spread of Chromolaena in Surnatra is therefore limited by altitude.of the BukitBarisan,wherethe plant is unable to developabove 1,000m or in isolatedplaces below 1,000m.
In addition,C. odorata has still not been notedon peat soils.
2. KalimantanDuring ourvisitstodifferent estatesinKalimantan, C.odorata wasseeninWest;Eastand .
SouthKalimantan, but it gradually disappears towards CentralKalimantan whereroadaccessisless.
. 3. JavaC.odorataiseverywhere exceptathighaltitudes. IndrierareassuchasEastJava ifcovers
extensive areas withdensegrowth.
A markeddry period during the year seems to favour C. odorata flowering and seedproduction. This is also the case in the SouthSurnatra (Lampung).
OBSERVATIONS IN AREAS WHERE C. ODORATA HAS NOTPREVIOUSLY BEEN REPORTED
1. SulawesiDuringoursurveyinSulawesi,Chromolaena wasfoundtobeverydenseinSouthSulawesi
Province: UjungPandang to theNorth,Parepare to Polewalion the westcoast, Watarnpone toPalopoontheeastcoast.Beyond thisontheborderofBonebayfrom Sabbang toWotu,C.odoratawas not noticed. On the otherhand, in the Jeneponto area,C. odoratagrowthis considerable,especially in the dry areasof SouthSulawesi.
44

In the Torajaland, C. odorata does not exist at high altitudes. C. odorata was only seenon the road from Palopo to Rantepao for 20 km beyond Palopo, but to the South it exists fromMakele to Kalosi - Enrekang - Rapang - Pangkajene. Near Enrekang, at lower altitude, where
. cattle farming has been developed, C. odorata has become established over a wide area. The.weed is not eaten by the cattle and forms very dense C. odorata thickets, thereby'very quicklyreducing the acreage of pastureland.
In North Sulawesi, Manado and its surroundings, C. odorata has not been observed andprobably does not exist.
2. Irian JayaIn IrianJaya C. odorata has not been reported earlier, but we found this plant particularly
well developed in Jayapura, on the site of a former second world war military base.
From the airportto the town, a distance of 33 km, C. odorata grows in very dense bushesalong the road and on the nearby hillsides. The spread of C. odorata can clearly be seen along
. the coastal roads and in the interior along the only road to the Papua New Guinea border as far. as Arso, which is now the limit of the C. odorata invasion.
There is no C. odorata in the Doberai Peninsula on the western part of Irian Jaya:Manokwariand neighbouringareas.
IV. OBSERVATIONS ON THE FLOWERING TIME OF C .ODORATAIN INDONESIA
InSumatraatPematangSiantar- Prapat,C.odorata isreallya"Christmasplant" floweringregularlyevery year at the end of Decemberand in January. In Riau Province at KotaPinang,Pekanbaru,thisweedwas floweringin March-April. InSouthSulawesithis weedwasfloweringin June-July. In Irian Jaya C. odorata was without flowersat the end of January and it was saidthat flowering occurs in March - April.
'r It seems therefore that the flowering season of C. odoratais different according to thealtitude and latitude from West to East in Indonesia.
CONCLUSION.
Our survey results indicate the recent establishmentof C. odorata in Sulawesi and Irian.Jaya. Theareaoccupiedby thisweedhasbeenconsiderablyextended to theEast morethan 1,500km, about half of the Indonesian Archipelago where it was not reported before. The existenceof C. odorata in Irian Jaya is a threat to Australia where this weed does not exist but where itsintroduction is greatly feared.
45

InrecentyearsC . odoratahasinvaded Indonesia from theWest to theEast,andprobablyisalreadypresentinMaluku. Onlyisolated placesandsomeislands, especiallyin Nusantara, arenot yet invaded by this weed.
C. odorata is now oneof the main weedsin Indonesia.
BmLIOGRAPHY OF CHROMOLAENA ODORATA (L.) R.M. KINGAND H. ROBINSON IN INDONESIA
Alif, A.F.B.M. 1982. Use of herbicides in small scale plantations in South East Asia. Biotrop SpecialPublication, 15: 83-89. .
Amypaluppy, K., A. Sudiman & Siswanto. 1988. The effect of the"turbidity of water solvent on theefficacy of Imazapyr on Alang-a1ang. Conf. Ke IX - Jilid ill, 139-143.
Ardjasa, S.W., I. G. Ismail & S. Effendi. 1981. The application ofDowpon M on Alang-alang infestedareas, in conjunction with the establishment of food crops. Prosiding Konperensi Ke-Enam,
- Himpunan Ilmu Gulma Indonesia, 15 - 23. .
Aterrado, E.D. & R. L. Talatala-Sanico. 1988. Status of Chromalaena odorata research in the Philippines. Proc. First Intern. workshop Biological Control of Chromolaena odoraia, pp. 53-55..
Agri. Expt, Sta .• Univ. of Guam, Mangilao, Guam.
Bennett, F.D. & V. P. Rao. 1968. Distribution of an introduced weed Eupatorium odoratum Linn.
(Compositae) in Asia and Africa and possibility of its biological control.
PANS (C), 14: 277 - 281.
Biswas, K. 1936. Some foreign weeds and their distribution in India and Burma. Indian Forester, 60:
861-865.
Budiman, A., M. Thamrin & S. Asikin. 1988. Some weed species growing on tidal land of different soilacidity. Prosiding Konperensi ke IX, Himpunan Ilmu Gulma Indonesia. Bogor, 22 - 24Maret 1988. Jilid IT, 81-92.
C.A.B. International Institute of Biological Control. 1988. Prospects for the Biological Control ofChromolaena odorata (L.) R.M. King and H. Robinson. In: R. Muniappan (Ed.) Proc. FirstInternational Workshop on BioI. Control of Chromolaena odoraia, pp. 25-33. Univ. ofGuam.
Cruttwell McFadyen, R.E. 1988. History and distribution of Chromolaena odorata (L.) R.M.
King and H. Robinson. In: R: Muniappan (Ed.) Proc. First Intern. Workshop BioI.Control oi Chromolaena odorata, pp. 7-12. Agri. Expt, Sta., Univ. of Guam, Mangilao,Guam.
46

Dalem, K.W.& A. Soedarsan. 1977. The effect of Bulhidazole and new Diphenyl ether onChromolaenaodorata (L.) RM. King and H. Robinson. Menara Perkebunan Bogor. 45(6),287-289.
Daryono, H. 1985. Effect of age on the composition and development of teak, Tectona grandis undergrowth flora. Bul. Penelitian Hutan. 469: 67-93.
Desmier de Chenon, R. 1988. Observationson the presence of Chromolaenaodoraia in Indonesia.
Paper presentedat theFirst International Workshop on Biological Control of Chromolaena
odoraia, Bangkok, Thailand, Country Report 3 pp.
Dove, M.R., 1986. The practical reason of weeds in Indonesia: Peasant vs. State Views ofImperala and
Chromolaena. Human Ecology 14:163-190.
Dove, M.R. & S. Martopo. 1987. Manusia dan Alang-alang di Indonesia. 286 p. Gadjah Mada University Press, 1987.
Eussen, 1.H.H.& S. Wirjahardja. 1973. Pioneer plants in the secondary forest establishment. TheSecond Indonesian Weed Science Conference, Yogyakarta, April 2 - 5,1973: 245-251.
Harnzah, Z. 1978. Forest weed problems in Indonesia. Biotrop Aniv. Publ: 75 - 83.
Holm, L.G., D. L. Plucknett, 1. V. Pancho & 1. P. Herberger. 1977. The World's Worst Weeds:Distribution and Biology. East West Center, University Press of Hawaii,USA. 609 p.
Johnstone, T. H. & H. Tryon. 1914. Report of the Prickly-Pear Travelling Commission, 1st November
1912 - 30th April 1914. Govt. Printer, Brisbane.
Karyono. 1980. Structure of homegarden in the rural area of Citarurn watershed, West Java. Diser
tation Padjadjaran University. 232 p.
. -'
Mangoendihardjo, S. 1975. Prospects of biological control of weeds in Indonesia. 3'" Indonesian Weed
Science Conf., Bandung: 33-41.
Mangoendihardjo, S.& M. Soerjani. 1978. Weed management through biological control in Indonesia.Proc. Plant Protection Conference, Malaysia: 323·337.
Muniappan, R.& M. Marutani. 1988. Ecology and distribution of Chromolaena odorata in Asia and the
Pacific. In: R. Muniappan (Ed.) First InInternational Workshop on Biological Control ofChromolaena odoraia. pp. 21-24. Agricultural Experiment Sta., Univ. Guam, Mangilao,Guam.
Naezer, H.W. & I.C. van der Meer Mohr. 1953. Insects and Eupaioriumodoraium. Trop.Natur.Bogor, 33: 59·60.
Nasution, U. 1981. Inventory of weeds in rubber estate of North Sumatra and its relation with weed
management. Prosiding Konp. ke-enam Himpunan Ilmu Gulma Indonesia, Medan 12-14Februari 1981: 193-210.
47

Nasution, U. 1986. Gulma dimpengendaliannyadi perkebunan karet Sumatra Utara dan Aceh. PusatPenelitiandan PengembanganPerkebunanTanjung Morawa (P4TM). 269 p.
Pancho,1.V.&D.L.Plucknett.1972.Chromolaena odoraia(L.)R.M.KingandH. Robinson. Anewrecordof noxious weed in the Philippines. Phil. J. Ani. Sci. 8: 143-149.
Partomihadjo,T. &Suhardjono. 1988. Analysis of weed vegetation in the field after cassava cultivationat Cibinong - Bogor. Prosiding Konp. ke IX Himpunan Ilmu Gulma Indonesia. Bogor 22 - 24Maret 1988. Jilid IT: 140 - 153.
Ramlan, A. 1988.Weed species which were used asTraditional MedicinalPlants. Prosiding Konp. Ke IXmm. Jilid ill, 1988: 276-283.
Ramlan, A.& Y. Dahyall988. Weed species in the various types of vegetation in the inundated areas ofthe Saguling and Cirata Reservoirs. Prosiding Konp. ICe IX HimpunanIlmu Gulma Indonesia.Bogor 22-24 Maret 1988, Jilid ill: 234-241.
Risdiono,B. 1975.The inter-influencebetweenChromolaena odorataL. and Brachiariabrizanlha Stapf.andthe effectof Picloramon theseplants. Proc.thirdIndonesianWeed ScienceConf., Bandung,June 24 - 28: 367-376.
Sastroutomo,S. S. & E.T. Pandegirot, 1988. Ecologicalsurvey of weeds at Ciliwung tea plantation. WestJava. Prosiding Konp. Ke IX mGL Jilid ill: 233-253.
Setiadi, D. 1988. Notes on Weeds in Teak Forests. Pros.Konp. Ke IX J:lIGL Jilidill: 74~77.
Siregar, H.H., T. L.Tobing & R. Sukarji. 1990. Dominasigulma pada daerah pengembangan perkebunankelapasawit diTapanuli Selatan(SumateraUtara)dan Riau. KongresLHimpunanPerlindunganTanaman Indonesia, Jakarta 8-10 Februari 1990. Makalah 320.11 p.
Soedarsan, A.•Noormandias & H. Santika. 1975. Glyphosate versus Dalapon for Alang-alang control.Proc.third Indonesian Weed Science Conference. Bandung, June 24-28: 428-436.
Soerjani, M.•A. Soedarsan, S. Mangoensoekarjo.T. Kuntohartono& M. Sundaru; 1975.Weed problemsand prospects for chemical control in Indonesia.5th Conference of theAsian Weed ScienceSociety: 18-22.
Soerjani, M.• A.J.G.H. Kostemans & G. Tjitrosoepomo (eds.) 1987.Weeds of rice in Indonesia. BalaiPustaka Jakarta, 716 p.
Soerohaldoko,S. 1971. On theoccurrenceof .Eupaioriumodoraiumat thegamereserve Penanjung,WestJava, Indonesia. Weeds in Indonesia. Weed Science Society of Indonesia, Bogor. 2(2).: 9.
Soerohaldoko, S. 1973.Some remarks on the seed germinationof Chromolaena odorata (L.) R.M. King. andH. Robinson. The SecondIndonesiaWeedScienceConference Yogyakarta,ApriI2-5. 1973:
167-171.
48

Soerohaldoko, S. 1975. The after effect of filtered sunlight through green leaves on the seed germination
of ChromolaeniJ odoraia (L.) R.M. King and H: Robinson compared with that of sunlight and
darkness. Proceedings of the third Indonesian Weed Science Conference, Weed Science Society
of Indonesia. Bandung, June 24 - 28.1975: 12-17.
Sudiman, A & K. Amypaluppy. 1988. The efficacy of Triclopyr against leguminous cover crops in the
planting strips of young rubber. Prosiding Konp. Ke IX HIOl. Bogor 22 - 24 Maret 1988, Jilid
ill: 286-295.
Sumardja, E.<'\.. & K. Kartawinata. 1977. Vegetation analysis of the habitat of banteng (80Sj(l\larUcus) at
the Pananjung-Pangandaran Nature Reserve. West Java. Biotrop Bulletin, 13. 1977.43 p.
Syed, R.A 1973. Some aspects ofbiological control of weeds in South East Asia. The second Indonesian
Weed Science Conference. Yogyakarta, April 2-5: 311-317.
Syed, R.A. 1977. An attempt on biological control of Eupatorium odoratumL. in Sabah, Malaysia. 6th
Conf. Asian Pacific Weed Science Society, Jakarta: 457-466.
Tjitrosemito, S., S.S. Sastroutomo & I. H. Utomo.1986. Weed management in a young rubber plantation
in Indonesia. Weedwateher.l (Supplement). 4 p.
Tobing, T.L. &C. Hutauruk. 1984. Identification ofweeds found in oil palm plantations. PedomanTeknis,
Pusat Penelitian Marihat (No. 83): 10 p.
van der Laan, P.A 1914. Motschildhuis en Eupatorium als oorjaken van pseudomozaiek. Vlugschr. Deli.Proefst. Medan, 67: 4 p.
Vayssiere, P. 1959. Agronomie Malaisie, Notes de mission. Laculture et les parasites du Tabac a Summa.
Journ. Agri. trop. Bot. appl. pp. 281-302.
Whitten, AJ., S. 1. Damanik, 1. Anwar & N. Hisyam. 1984. The ecology of Sumatra. Gadjah Mada
University Press. 583 p.
Whitten. A1.. M. Mustafa & G. S. Henderson. 1988. The ecology of Sulawesi. Gadjali Mada UniversityPress. 777 p.
WidYannoko & H. Ryanto. 1986. Pengelolaan gulma pada lahan bukaan baru pengalaman di perkebunan
tebu Gunung Madu plantations Lampung. Prosiding Konp. Ke vm HIOl, Bandung 24 - 26 Maret
1986: 143-151.
Yusron, A. 1988. Weed Inventory in Corn Crop in die Transmigration Area ofUPT A-I Pangkalan Kersik,
Regency of Musi Banyuasin, South Sumatra. Prosiding Konperensi Ke IX Himpunan IlmuGulma Indonesia. Bogor, 22 - 24 Maret 1988. Jilid ill: 59-73.
49

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
jj

PRELIMINARY RESULTS OF A SURVEY AND ASSESSMENT OF. CHROMOlAENA ODORATA (SIAM WEED) IN AFRICA
SULAYMAN S. M'BOOBSenior Crop ProtectionOfficer
FAO RegionalOffice for Africa, Accra, Ghana.
INTRODUCTION
Chromolaena odorata L. (R. M. King & H. Robinson) (Eupatorium odoratum L.) is anintroducedherbaceous,annualor perennialwoodyherbor shrub which hasbecome widespreadin West and Central Africa. It has assumed various growth habits depending on the habitat inwhich it occurs. It does not take the plant long to become established once it fmds an open,availableniche. Itgrowsand spreadsvery rapidly to cover theground. In theprocess, it chokesand preventsother residentplantspecies fromgrowing. It also prevents its own seedlings frombecomingestablished.
The perennialand vegetativereproductive habitshave madethe plant become part of andintegratedwiththeflorainmostWestAfricancountries. Inarablelands,it hasbecomea nuisanceduringthecropgrowingseason. Italso disturbsseedlingsof cashcrops in youngplantations. Inthiscontext,it hasassumeda peststatusbecauseof its interferencewithfoodproductionand theeconomy in general.
Furthermore,theextentof itsdistribution andabundanceisnotfullyknownexceptperhaps .inGhana,NigeriaandCoted'Ivoire. Asa resultof the threatposedby thisweed to theeconomyand moreespecially to foodproductionin the region, the Food and AgricultureOrganisationofthe UnitedNations (FAO)RegionalOffice for Africa is conductinga survey on Chromolaenaodorata in Africa, to assess the extent and severity of its distribution, establish the state ofknowledgeon the biology and control of the weed and make an inventoryof past and currentresearchwork. Surveys have been conducted in Ghana, Sierra Leone, Liberia, Cameroon andNigeria. The following broadgeneralizations can be made out of the survey.
BACKGROUND INFORMATION AND DISTRIBUTION
C. odorata is an introducedperennialshrub commonin the humid and semi-humidareasof thecountriesvisited Initialestablishmentis usuallyby seedwhichisan achenewitha pappusof whitebristles. Rowers occur in clustersof 10-20,in terminalcymes in axils of upperleaves.Colourof flowerscan be eitherwhite, mauveor navy blue. After establishment the weed thenachieves prodigious lateral spread by vegetative means. The origin of the weed in Africa is
51

obscure but in most cases it was believed to have occurred as a contaminant in seed lots ofGme/ina arborea, aforesttree species,whichwereimportedfromMalayaforuseinreafforestationschemesin severalWest Africancountriesduring the sixties. Within two decades after it wasnoticedin mostcountries,except in Nigeriawhereit was recordedabout four decadesago, theweedhasspreadfromeast to westandhasreplacedmostof theothernaturalbushfallowspeciesin the forest, transitional forestand parts of the wet Guinea Savannahvegetativezones.
Almost invariably, in all the countriesvisited the weed was found to occur within a beltbetweenlatitude2" and8" N stretchingfromeast towestandin areasreceivingmorethan 1000mm rainfall per annum. The weed spreadsand grows so fast that an impenetrable thicket isdeveloped withinabout two seasons.
ECONOMIC IMPORTANCE
C.odoratahasbecomethecommonweedofopenspacessuchascroplands,roadsidesandutilityrights-of-way. It is also an importantweed in youngoil palm,cocoa, rubber plantationsand other tree crops in the areas affected. As SUCh, it interfereswith both food and cash cropscausingreduction ofyieldsinthecompetition forlight.moistureandnutrientsandtosomeextent,by allelopathic effects. Extent of yield losses is yet to be determined.
Perhaps the most importanteffect of the weed on production is its high cost of controlespecially on small-holdings. A high input of labour is required for the initial clearing andsubsequentfrequentpost-planting weedings necessarytoreducecroplosses. Theslashand burnmethodof land preparation is ineffective against this weed, but rather tends to aggravate theproblem. This isbecausewhentheweedisslashed,regrowthcan occurfrombudson theaboveground portion of the stem and the root crown. Complete control can only be achieved by
" slashing,burningand uprooting the stumpsmanually.
C. odorata also constitutesa fife hazard in areas with a prolonged dry season (bimodalrainfalldistribution). Largequantitiesof combustible plantdebrisareproducedin thedryseasonand thedried vinesserve"as fire transmitting linesin theeventof bushfire outbreakscausingthefire to spreadfar and wide. Large treecrop estates havebeen destroyedthis way in Ghana andNigeria.
Still on the negativeside. the weed is known to harbourharmful insects under its densecanopy. Species of Zonocerus grasshopper breed freely under the canopy where they areprotectedfrompredators. From there they moveon to neighbouring cassava farms,feeding onthe leavesand therebyreducingcrop yields.
On the positiveside the weed may havesomepotentialvalues,chief among which is theadditionof organic matter to the soil throughcopious production of leaf litter. There are alsoindications, by wayof 'farmer discovery notes', thatC. odoraia can reduce incidenceof nematodeswhen used as a fallow crop.
52

Furthermore,theweedmay havepotentialfor soilerosioncontrolon accountof its vigourand rapid lateral spread.. In someplaces, it is being used for curative and therapeuticpurposes.
STATE OF KNOWLEDGE OF THE WEED
It is surprising that very little research work has been undertaken on such an importantweed. Mostof the researchhasbeen carriedout in NigeriaandGhanaby individualresearchersspreadinuniversitiesandinstitutesresponsibleforcashcropssuchasrobber,cocoaandoilpalm.There hasbeenno attempt to prioritizeand orchestratethese individualresearchefforts intoonecohesiveendeavourtofindsolutionstotheproblemofC.odorata. Todate, researchworkcarriedout on the weed falls into two main categories,biology and control.
BIOLOGY
In Ghana. someaspectsof reproductivebiology.seed longevity.vegetativedevelopmentand inherentgerminationinhibitorsare beingstudied. In Nigeria, theeffectof C. odorata on theperformance of establishing Iroko trees (Ch/orophora excelsa now Milicise ace/sa) wasstudied. Attention was also focusedon the possible use of the weed as a fallow crop for rapidnutrientregenerationof soils.Fieldsof maizewereimprovedby I-year C.odorata fallow in thiscontext.
MANUAL CONTROL
In general. the control measure taken to check growth of the weed in arable land andprevent its spread is slashing and burning, and to a limited extent mechanical ploughing.Ploughingto a depthof 15cmor moreisknowntodestroythe weedcompletelyand that perhapsaccounts for its-absence on mechanizedholdings.
The manual method of control. involving the initial slashing and burning'followed byfrequentpost-plantinghoeingson smallholdings,is verytediousand requireshigh labour inputThis has led 10 the spate of researchactivities in the areas of chemicaland biological control.
CHEMICAL CONTROL
Several pre-emergenceand post-emergence herbicides have been evaluated and foundusefulforcontrolof C.odorata inyoungcocoaandoil palmplantations. These includeparaquatand paraquat-basedmixtureswithdiuron(paracoland Gramuron),2,4-D, atrazine, glyphosate,ierbuthylazine, triclopyr, ioxynil, fluorodifen, etc..
53

In most cases the herbicides failed to give season-long control and repeated. costlyapplications were often necessary in young cocoa and oil plantations. For a herbicide to beeffective it must bephloem mobile so that enough of it is translocated basipetally to reach andkill the deeply seated buds on the underground portion of the stems of the weed. On this basis,only products such as glyphosate, triclopyr and to some extent, 2,4-D out of the long list ofcandidate herbicides deserve serious attention. Stage of growth of the weed at the time ofapplication tends to influence the direction of movement of synthates along which theseherbicides move and consequently is an important determinant of herbicide efficiency inperennial weeds.
BIOLOGICAL CONTROL
C. odorataappears to be a perfect target for biological control which is relatively cheapand environmentally safe. The weed is an introduced species and therefore there might existinsects or other natural enemies in its area of origin (South America) which are absent in WestAfrica It isa perennial weed, widespread and abundant, ensuring constant food supply for anyintroduced enemies. Several natural enemies, mostly insects, have been introduced .butapparentlywithlittlesuccess inAfrica. These includethearctiid mothPareuchaete~pseudoinsulata
(Rego Barros) (Syn: Amalo insulata (Walker)) introduced in Ghana and Nigeria and Apionbrunneonigrum introduced in Ghana. In all cases the insects failed to establish.
54

10
Fig. 1. Map of West Africa showing present distribution of C. odorata in countries visited.
ss


THE STATUS OF CHROMOLAENA ODORATA (L.) R.M.KING ANDH. ROBINSON IN INDONESIA
S. TJITROSOEDIRDJO 1, SRI S. TJITROSOEDIRDJO 2, andR.C. UMALy3
. 1 and 2 Scientists,Tropical AgriculturalPest BiologyProgram,3 Deputy Director for Programs,BIOTROP, P.O. Box 17, Bogor, Indonesia
ABSTRACT
Chromolaena odorata (L.) R.M.King& H.Robinsonisknownin Indonesiaas kirinyu.It isa nativeof theneotropics.It wasintroducedto Indiain the 1840sas anornamentalplant,fromwhere it spread to the SoutheastAsianregion.The first herbarium specimen identified later asC. odorata wascollectedin 1934fromLubukPakam,NorthSumatera.Sincethe SecondWarldWar it spread rapidly in Indonesiaand received the name"independenceshrub". It hasbecomean important weed in forests, pastures and in plantation crops. Manual weeding is donecommonlyand chemical control is used only in the plantations. Biologicalcontrol has not yetbeen taken up in Indonesia. Because Of its rapid spread, it is imperative that its biology andecology be studied and appropriatecontrol measures be established.
INTRODUCTION
When Beumee(1922)madean inventoryof undergrowth speciesin teakplantationsintheislandofJavahedidnotfindkirinyu(Chromolaena odorata (L.) R.M.KingandH. Robinson)at that time. Howeverabout50 yearslater,forestersin centraland east Java werestartledby theheavy infestations of C. odorata that occurred. Subsequent investigation has indicated thatinfestation of C. odorata significantly reduced the growth of young teak plants (Daryono &Hamzah, 1979).Soerohaldoko (1971)also observed the occurrenceof C. odorata at the gamereserveof Penanjung,WestJava, and blameda reductionin the grazingareaofbanteng (Bibosjavanicus) on this infestation of Codorata. Other recent reports have given more informationon the harmfuleffectsof C.odorata (Nasution1986,Tjitrosoedirdjoet al. 1984,Soerjani 1977).
TAXONOMIC ASPECTS
Kirinyubelongs to the familyAsteraceae. For a long time the Asteraeeaewas dividedinto 12 tribes. However, with recent findings in biochemistry, pollen analysis, anatomy andcytology, the family Asteraceae has undergone considerable modification (Heywood 1979).Currently, this family is divided into two sub-families Lactucoideaeand Asteroideae.Kirinyuis a memberof the tribeEupatoriae,withinthe sub-familyLactucoideae,which is characterizedby white, reddish or bluish flowers lacking ray florets.
57

Kirinyu was previously included in the genus Eupatorium but is now under the genusChromolaena. It is distinguished by the rather consistent pattern of many rows of phyllaries(bracts) that are progressively longer and give a markedly cylindrical appearance to the head, bythe three prominent veins in the leaves and by the pungent odoremitted by the crushed leaves.
The genus Chromolaena contains 129 species and all are found in the neotropics (Kingand Robinson 1970). Of these C. ivaefolia (L.) and C. larvigata (Lamarck) are widespread andoccasionally weedy in the Americas, but only C. odoratahas spread to the Old World (CruttwellMcFadyen 1989).
SPREAD OF·C ODORATA
Based on information from Voight (1845) who listed C. odorata as a garden plant inCalcutta, India, from Hooker (1882) who reported that C. odoratawas cultivated although veryrarely, and from Prain (1903, 1906) who stated that it was cultivated sparingly in gardens ofCentral and East Bengal and around Calcutta, Cruttwell McFadyen (1989) concluded that C.odorata was introduced into Calcutta as an ornamental in the 1840s from where it graduallyspread south into lower Burma and eventually reached Malaya by 1918. It was recorded as aserious and rapidly spreading weed in Burma, Assam and Bengal in 1920 (Ramachandra Rao
. 1920) and as a dangerous weed in Sri Lanka in 1944 (Grierson 1980). Once established in theBengal-Lower Burma-Malaya area, C. odorata spread rapidly throughout southeast Asia. Its
, spread to new areas was assisted by the extensive movement ofpeople, machinery and materialsduring the World War 11 (Cruttwell McFadyen 1989). In Sumatera this weed is sometimes calledas "semak merdeka" or "independence shrub" because it became abundant after the independence war..In Indonesia the first herbarium specimen of C. odorata was collected in 1934 fromLubuk Pakam, North Sumatra by van Meer Mohr and kept at the Herbarium Bogoriense (4004).This specimen was identified as Eupatorium repandum, but later was reidentified by van Steenisas Eupatorium odoratum (Tjitrosoedirdjo 1990).
The exact year of introduction of C. odorata us Indonesia is not known. It is probablethat it was introduced a few years before 1934, when the firstherbarium specimen was collected.
It is now spread all over the islands of Indonesia from Aceh to West lrian.
ECOLOGICAL STATUS
C. odorata is a perennial shrub forming dense tangled bushes up to 3 m in height.Although it burns readily. it resprouts from its stump immediately after the rains. This abilitycoupled with its fast growth rate enables it to compete well with alang-alang Umperata cylindrica(L.) Raeuschel) and shade it out. This condition will facilitate further vegetational successiontoward secondary forest.
Eussen & Wirjahardja (1979) observed 4 stages of successional development fromgrass-type alang-alang vegetation (a sign of critically degraded environment) to an earlysecondary forest stage (a more desirable ecological condition), Le. (1) Imperata formation,
58

where the vegetation is still dominated by alang-alang, an indication of acritical, degradedenvironment. (2) The Imperata - Chromolaena formation, in whichthe area is dominatedbyalang-alang and C. odorata. (3) TheChromolaena formation whereC. odorata dominates thevegetation. (4) The early young stage of secondary forest. where with the disappearance ofgrassy vegetation trees will establish: this is precededby the domination of shrubs,such as C.odorata , Melastoma malabathricum, Trema orientalisand Bridelia monoica.
A similarobservation was reponedby Tanimoto(1981)in SouthSumatera, In its roleinvegetational succession from thealang-alang community, a signof degradedenvironment, toa secondary forestcommunity, C. odorataperforms a usefulfunction. However, pastoral landinfestation of C. odorata is undesirable. .
WEED STATUS
Forestplantations
The existenceof thisweed,C. odorata, is a concernfor foresters in teakplantations.It reducesthegrowthof teakseedlings andresultsin reducedstemdiameter. The freshbiomassof C. odorata in a teak plantation per hectare was reponed to be a totalof 18.7 tons,consistingofleaf6.8 tons,stems10.2tonsand underground parts 1.7tons. Thisconstitutesapproximatelya totalof 3.7 tons of dry matter/ha. Theelementalcontentof the biomassis listed in Table 1.
. Table1.Meanpercentage of someconstituentelements of C.odorata (modified fromDaryono.& Harnzah 1979)
Plantpan
LeafStemRoot
Elemental content (%)
N P K Ca Mg Na
5.89 0.74 3.13 3.30 0.83 0.011.00 0.23 1.73 0.37 0.18 0.010.87 0.13 0.93 3.30 0.15 0.01
Whenthebiomass isconvenedtoitselemental constituents, ineach hectareC.odoratawill accumulate approximately 103.4 kg of N, 15.2 kg P, 80.9 kg K and 63.9 kg Ca. Thesenutritional elements are thus not available for the growing young teak as they are stored inC.odorata.
59

It wasalso reported thatC. odoratatranspired about 1908 mmof waterper year. Thisamountoflossmaynotbemuchofaconcern inwetterregions, butinareaswhereteak plantationsare established (especially in Centraland East Java) such water loss is certainlydamaging toyoungteak. In such situations it seemsthat the occurrence of C. odorata shouldbe controlled.
Pastoral land
In natural pasture land, as exemplified by·the reserve at Penanjung,West Java, theinfestation of C. odorata can be detrimental when left uncontrolled. Infested pastures arechanged into secondary forest succession and this reduces herbage for banteng. As a resultbantengmigrated out of the reserveanddisturbed the adjacentagricultural areas.
Prawiradiputra et al. (1986) investigated the effectof C: odorataon the production ofgrassherbageina natural pastureland,and found that it reducedthegrassherbageconsiderablyas described by this equation:
y = 178.09 - 3.51 Xl, where
y = herbage yield,andxl = summeddominance ratio of C. odorata
When C. odorata is associated with I. cylindrica, the pasture quality is reduced.However, suppression of C.odorata in suchsituations mayresultin I. cylindrica becomingthedominantvegetation.
Perennial crop plantations
The importance of C.odorataas a weedinplantation crops suchas in rubber,oilpalm,coconut, cocoa and others has been well documented (Soerjani et al. 1976,Wargadalarn &Soedarsan 1977,Nasution 1981,Tjitrosoedirdjo et al. 1984)..
Nasution (1981) showedthatit issecondonlytoalang-alang in termsof its harmfulnessin plantation crops(Table2).Theeffectof C.odorata on thegrowthof rubbertrees is showninTable 3.
. 60

Table 2. Classification of vegetation under rubber plantation for management purposes(modified from Nasution 1981)
Vegetation Group
A. Useful vegetation
. B. Harmless vegetation
e.. Harmful when excessive
D. Harmful
E. Very harmful
Botanical name
I. Calopogoniumcaeruleum(Benth.) Hemal
2. C. mucunoides Desv,3: Centrosemapubescens
Benth.4. Pueraria phaseoloides Benth.
I. Ageratum conyzoides L.. 2. Cyrtococcum spp.3. Digaaria spp .
. 4. Erechtites valerianifolia(Wolf.) D.e.
5. Phyllamhus niruri L.
I. Axonopus compressus(Swartz.) Beauv.
2. Borreria latifolia(Aubl.) K Sch.
3. CyclosorllS aridllS(Dor.) Ching
4. Cynodon dactylon (L.)Pers.
5. Cyperus spp.6. Echinochloa colonum (L.)
link7. Eleusine indica (L.)
Gaertn.8. Nephrolepis biserrata
Schou,9. Ouochloa nodosa
(Kunth.) Dandy10.Paspalum spp.
I. Brachiaria nlllica(Forsk.) Staft.
2. Chromolaena odorata (L.)R.M. King & H. Robinson3. Gleichenia linearis
Clarke,4. Lantana camara L.5. Melastomaaffine D. Don.6. Scleria sumatrensis Retz,7. Stachytarpheta indica (L.)
Vahl.8. Trema sp,9. Grewia eriocarpa Juss.
Y.Lmperata cylindrica (L.)Raeuschel
2. Mikania sp.3. Mimosa sp.
Management option
Desired cover crops; when climbingthe crop is controlled
10 sloping areas. they may act assoil binders to prevent soil erosion,however when excessive should becontrolled
10young plantations should becontrolled
Must be controlled under mostconditions
61

Table 3. The girth of rubber trees under various treatments of vegetational covers (modifiedfrom Wycherley & Chandapillai 1969).
No. Treatment
1. Ottochloa2. Creeping legume3. Axonopus4. Moghania5. Melastoma6. Brachiaria .7. Chromolaena8. Ficus9.· Mikania
Girth at 60 inch height(in inches)
20.0219.8119.5519.5518.7418.6418.5418.5118.40
NB : Treatments No. 4, 5, 7 and 8 were controlled by periodic slashing.
C. odorata reduces the girth of rubber trees when compared to those under creepinglegume or Ottochloa. Other vegetational covers which reduced the growth were Brachiariamutica, M. malabathricum (affine), and Mikania sp. In most conditions the infestation ofCodorata will be accompanied by other associated vegetation. When the infestation is verysevere and control measures are inadequate. the growth of the crop will be retarded. In someinstances the resulting trees become extremely variable in size. Such crops become worthlessto continue maintaining and the field has to be replanted again, with huge financial loss.
CONTROL AND MANAGEMENT
Once an area has become infested, the weed is difficult to control. This h; because itproduces a great number of seeds, i.e. 93,000 seeds/ plant(Weerakoon 1972). Soerohaldoko(1979) kept C. odorata seeds up to 15 weeks in about 12 hours dark and 12 under daylightandfound these seeds to be viable.
Manual controlManual control is the most commonly used method in plantation crops especially in the
island ofJava where the availability of labor is still reasonably high. Manual control is done byslashing the stem above the ground, but the plant soon resprouts from its base. A more tediousform of control is uprooting C. odorata, with almost double the cost of slashing.
A typical program of weed control in young rubber plantation when infested with C.odorata is given in Table 4. (Tjitrosemito et al. 1986).
62

Table 4. Weed control program in young rubber plantations.
No. Activity
1. Manual weedingbetween rowsagainst shrubs
2. Manual weedingin rows .
3. Weed control in'rows prior to
fertilizerapplication
Chemical control
Frequency ofcontrol per year
once in 2 months(6 x)
once in 3 months(4 x)
once in 6 months(2 x)
Mandays
48
27
6
Chemical control is.the practice usually used in estate plantations. C. odorata is controlled by herbicides such as triclopyr (3;5,5 - trichloropyridinyl-oxyacetic acid), 2,4-D (2,4dichlorophenoxyacetic acid), picloram (4-amino-3,5,6-trichloropicolinic acid). The rates givenin table 5 have been tested and found to bereasonably effective.
In an integrated system, C. odorata can becontrolled successfully in sugarcane plantations after 5-6 years. This integrated control system, which incorporates appropriate soilcultivation combined with correct timing of pre-emergence herbicide using diuron + 2,4-Damine at (1.6-2.5) + (1.0 + 2.5) kg ai/ha, is able to control C. odorataand other common weeds.When isolated specimens escape they can be treated by post emergence spraying using 2,4-Damine + ametryn together with diuron (Widyatrnoko & Riyanto 1986).
Table S. Herbicides used to control C. odorata.
No. Herbicides
1. Triclopyr2.2,4-D3. Picloram4.2,4,5-T
Rate(kg ai/ha)
0.8-1.62.52.01.0
References
Tjitrosoedirdjo et al. 1984Tjitrosoedirdjo et al..1984Prawiradiputra et al. 1986Risdiono 1975
63

Biologicalcontrol
Work on biological control ofC. odorata in Indonesia is yet to be initiated. Preliminarysurveys indicated absence ofany significant local natural enemies. An aphid has been observedto attack young shoots and Causes leaf curl.
FUTURE ACTIVITIES
C. odorata is rapidly spreading and covering new areas in Indonesia. It is necessary tounderstand its ecology to develop appropriate control methods. Research on its biology, ecology,ihfluence ofthe environment on growth, productivity, population dynamics, and interaction withother species need to be carried out.
C. odorata accumulates considerable amounts of mineral nutrients, resulting in bothuseful and harmful effects depending upon the point of view. Its ability to shade out alang-alangis considered a desirable property. However in plantation crops and in young cassava fields, theinfestation is detrimental.
Its possible uses as a source of green manure, active substances and for nematodecontrol need to be investigated. However, as a weed its population should be suppressed adoptingenvironmentally safe, economical and effective methods such as biological control.
REFERENCES
Beurnee, J.G.B. 1922. Floristic-analytische onderzoekingen van de korte flora in kunstmatingaangelegde djati plansoenen op Java in verband met de ontwikeling van de djatiopstand, Dissertasie Landbouw Hoge School, Wageningen. .
Cruttwell McFadyen, R.E. 1989. Siam Weed: a new threat to Australia's north. Plant ProtectionQuarterly 4: 3-7.
Daryono, H. & Z. Harnzah. 1979. A study of Eupatorium odoratum L. weed found in teakplantation. Lembaga Penelitian Hutan,Bogor, Laporan no. 312: 26p. (Forest ReseachInstitute, Bogor, Report no. 312: 26p.).
,Eussen, J.H.H. & S. Wirjahardja. 1979. Pioneer plants in the secondary forest establishment.Proc. 2nd Indon. Weed. Sci. Conf.: 245-251.
Grierson, AJ.G. 1980. Compositae, In FloraofCeylon Vol. I. Smithsonian Institute, USA.
Heywood, V.H. 1979. Flowering plants of the world. Oxford Univ. Press. Oxford, London.
64

Hooker, J.O. 1882. Flora of British India, Vol. Ill. Spottiswoode, London.
King, R.M. & H. Robinson. 1970. Studies in the Eupatoriae (Compositae), 24: The genusChromolaena. Phytologia 20: 196-209.
Nasution, U. 1981. Inventory of Weeds in Rubber Plantations in North Sumatra and its Relationwith Weed Management. Proc. 6th Indonesian Weed Sci. Conf. 193-210.
Nasution, U.1986. Weed and its Management in Rubber Plantation in North Sumatra andAceh(in Indonesia). 1st Ed, Pusat Penelitian & Pengembangan Perkebunan Tanjung(p4TM), Medan, Indonesia.
Prain, O. 1903. Bengal plants, Vol I. Botanical Survey of India, Calcutta.
Prain, O. 1906. Contributions to Indian Botany. West Newman & Co., London.
Prawiradiputra, BR, S. Hardjosoewignjo & S. Tjitrosoedirdjo. 1986. The effectof weed controlon the vegetational composition of-natural pasture land in Jonggol, West Java. Proc.8th Indonesian Sci. Conf. : 103-108.
Ramachandra Rao, Y. 1920. Lantana insects in India. Memoirs, Dept. of Agric. in India.Entomol. Series, Calcutta 5: 239-314.
Risdiono, B. 1975.The interinfluence between Chromolaena odorata and Brachiaria brizanthaand the effect of picloram on these plants. Proc. 3rd. Indon. Weed Sci, Conf. 1975,Bandung, Indonesia.
Soerjani, M. 1977. Weed management and weed science development in Indonesia. Proc. 6th.Conf. I: 31-41. Asian-Pacific Weed Science Society Conf. 1:31-41.
Soerjani, M., A. Soedarsan, S.Mangoensoekardjo, T. Kuntohartono & M. Sundaru. 1976. Weedproblems and prospects for chemical control in Indonesia. Proc. 5th. Asian-PacificWeed Science Society Conf. 1975, Tokyo, Japan.
Soerohaldoko, S. 1971. On the occurrence of Eupatorium odoratum at the game reserve,Pananjung, West Java. Weed in Indonesia. 2: 9.
Soerohaldoko, S. 1979. Changes in response to light by seed of Chromolaena odorata(L.) R.M-.King and H. Robinson during storage. Proc. 5th. Indonesian Sci.Conf. 1118-11/12.
Tanimoto, T. 1981. Vegetation of alang-alang grassland and its succession in the BenakatDistrict of South Surnatra, Indonesia. Bull.For, and For. Product 314: 11-18.
Tjitrosemito, S., S.S. Sastroutomo & I. H. Utomo. 1986. Weed management ina young rubberplantation. SEAWIC Weed Leaflet 1.
65

Tjitrosoedirdjo, S., I.H. Utomo& S.S. Sastroutomo. 1984: Tryclopyrfor thecontrol of woody.weeds in youngrubberplantation. Proc. 7th. Indonesian Weed Sci. Conf. 71-77.
Tjitrosoedirdjo, Sri S., 1990.Somenoteson Chromolaenaodorata. Proc. 10th.Indonesian Sci. .Conf.40-46.
Voigt. 1845. Hortussuburbanus Calcuttensis.
WargaDalem,K. & A. Soedarsan. 1977. The effectof buthidazole and new diphenylether onChromolaena odorata (L.) R.M. King and H. Robinsosn. Proc. 6th. Asian-PacificWeed Sciences Society Conf.: 506-509.
Weerakoon, L. 1972. Studieson biology andcontrolof Eupatorium odoratum L. M.Sc.Thesis,Vidyodaya Campus,Univ.ofCeylon.
Widyatmoko & H. Riyanto, 1986. Weedmanagement in newlyopenedland.(An experience ofGunungMaduSugarcanePlantation,Lampung). Proc. 8th.IndonesianWeedSci.Conf.143-151.
Wycherley, PR & M. M. Chandapillai. 1969. Effect of cover plants. J. Rubber Res. Inst.Malaya. 21: 140-157. .
66
/

DISTRmUTION AND IMPORTANCE OF CHROMOLAENA·ODORATA IN COTE D'IVOIRE
M.G.ZEBEYOUIRHO, La Me
13 BP 989, Abidjan 13 Cote d'lvoire
.INTRODUCTION
Chromolaena odorata is native to South and Central America It was introduced intoWest Africa in 1936-1937 as seeds mixed with seeds of Gmelina arborea in Enugu (Nigeria)from Sri Lanka (Odukwe 1965). In Cote d'lvoire C. odoratawas first reported in 1972 (Ivens1972). Two sources of information exist about this introduction: firstly coffee farmers fromCameroonand CentralAfricanRepublic in 1946came with the weed to Eloka where they usedit as a coverplant (Carlier-Gance -Snoeck- pers,comm.),SecondlyC.odoratawasintroducedas seeds mixed with rubber seeds: Accordingto Delabarre (1977) two written referencesalsoexist: that of Dr. Miegewho reported the introduction of the weed in 1953,and that of Dr. Akewhofound the weed in a rubber nursery in Anguededou in 1960.
DISTRmUTION
Accordingto Dr.Delabarrein 1977C. odorataoccupiedonlya small part of thesoutheast of Cote d'lvoire from Abengourou to Abidjan passing through Agboville (Fig. 1). LastDecember(1990)I madea surveyand nowonlya smallpartof theNorthEast is freeof the weed(Fig. 1).
DAMAGE
Generally this weed is a problemfor the cash and foodcrops (maize,cassava, etc.). InCoted'lvoire oureconomyis basedon agricultureand we havetwo importantcrops:coffeeandcacao. Many coffee plantations have been abandoned because they have been invaded by C.odorata in Aboissoand Abengourou (Durandeau1976).In the dry season C. odoratabecomesa fire hazard and many coffee plantations were lost due to this. C. odorata is also a problemamong youngpalm trees, and in bananaplantationsresulting in poor production.C. odorata isalso the host of the pest grasshopper Zonocerus variegatus (Atayi & Knipscheer 1980) andenhancesits development
CONTROL METHODS
Chemical treatment by herbicides is very expensive even for large plantations, sosmaller farmers cannot be expected to use it. Slashing which is usually used to eliminate C..odoratais not efficient because the weed regrows rapidly,and when one shoot is cut a further8 shootsdevelop. Burning is also used but is not efficient either.
67

So the last solutionis biologicalcontrol. So far we have tried to find natural enemieswithoutany success.Aphisgossypiiwhichcurls up the leavesbut causes the weed grow faster,and Anoplocnemis curvipes which cuts the young branches of C. odorata, do not have asignificantimpact on the weed. A. curvipes is also a pest of cotton. We plan to introduce theprovenbiologicalcontrol agent Pareuchaetes pseudoinsulata fromGuam.
REFERENCES
Atayi,E. A. & H.C. Knipscheer. 1980.SurveyofFoodCropFarmingsystemsin the"Zapi-Est".East CameroonInternationalInstitutefor Tropical Agriculture. ONAREST.p. 94
Delabarre,M. 1977.IncidenceAgronomique du Developpement de Eupatorium odoratum(Composees) en Cote d'Ivoire (These).
Durandeau,B. 1976.I.F.C.c. Communication Personnelle.
Ivens,D.0.1972. Weed ResearchProjectat the Universityof Ibadan,Nigeria PANS. 18:210.
Odukwe, D. O. 1%5. Eupatorium odoratum (Siam Weed). Bull. Nigerian Forest Dept.
68

LIBERIA
MALI
BURICINA-FASO·
LEGEND
Itinerary followed during the survey
[D]]] Zone occupied by the weed C. odoraia in 1977
r+++l Zone free of C. odorata in 1990
L..±..:t..:!:..
GHANA
Figure 1. Distribution of Chromolttell4 odoratQ in Cote d'Ivoire
69

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j

PRELIMINARY RESULTS ON THE ENZYMATIC DIVERSITY INCHROMOLAENA ODORATA (L.) R. M. KING AND H. ROBINSON
(ASTERACEAE)
C. LANAUDl ,M.P. JOLIVOTl AND M. DEAT21 AGETROP, GERDAT/CIRAD, BY. 5035, 34032Montpellier Cedex,France
2 AMATROP, IRCT/CIRAD, B.P.5035, 34032 MontpellierCedex,France
ABSTRACT
C. odorata was introduced to tropical Africa about fifty years ago and has becomeextremelyinvasive. It hascolonised the wholeof theforestzone in westandcentral Africaandis progressively spreading into the savanna zones to a minimwn isohyet of 1250 mm and amaximum altitudeof 1200m. '
A studywas undertaken on theorganisation of geneticdiversity in order10 obtainbetterknowledgeof theplantwitha viewtooptimisingcontrolmethods. Theprinciplesof descriptivetaxonomy wereusedtocompareAmerican, Asianand Africansamplesbutgaveno differentiation. Chemotaxonomy studies were therefore carried out using enzymatic markers. Electrophoresistechniques designed foryoungleavesenabledtheidentification ofsevenclearlyvisibleenzymaticsystems, five of which were polymorphic. Preliminary research showed that the .enzymeprofilesof plantsfrom America (FrenchGuiana)weredifferentfrom thoseof material
.of Asian or Africa origin, in which four enzymatic systems out of five were found to bemonomorphic.
Moredetailedand morecompleteresultsare needed. The work: is therefore continuingon a larger numberof plantsof morevariedgeographical origin.
INTRODUCTION.
ThegenusChromolaenaD.C. isoneofthemostimportantgeneraoftheEupatorieaewith165speciesdescribed todate (King& Robinson, 1987). Chromolaena odorata (L.) R.M. Kingand H.Robinson is the bestknown speciesbecauseit is nowverywidespread in the tropicsandbecauseit is important as a weedin the regionswhere it hasbeen introduced. It is often knownby its formername,Eupatorium odoratumL., or by thecommonnamesSiam Weedor Herbedu Laos. .
C. odorata originated in America, where it is found from the southernUnited States toArgentina. Seedswere introduced in Asia in the 19thcentury,probably with the earth used asballastin shipping from the Caribbean. Bennett& Rao (1968)reportedthatSingaporewasone
71

of themainentrysites.Introductions also occurred inotherAsiancountries,andthenthespecieswasspreadwidely by manandanimalsandbecamea majorweedinannualand perennialcrops.Theearliestknownintroduction toAfricawasin 1937 ineasternNigeria(Ivens1974).C.odorataseeds wereintroduced accidentally in a batchof Gmelina arboreaRoxb.seeds fromSri Lanka.Other accidental introductions occurred in the English-speaking African countries such asGhana. At the same period, it is thought that plantersfrom Indochina introduced the plant inCameroon andCentralAfrican Republic - eitherinvoluntarily or intentionally for useas a coverplant in coffee and pepper plantations (Delabarre 1977). After the Second World War, SiamWeedwasbroughtfrom theseareas toCoted'Ivoire, whereit wasobservedfor the first time in
, 1952(Miege1955). It thenspreadas in Asiaand iscontinuing to do so today.The wholeof thewestAfrican forestzoneandtheCongolese basinhasbeeninfested. C.odorataisnowbecomingestablished in the savannaregions. It so far does not seem to have colonisedareas where theannualprecipitation is lessthan1250mm,but isnevertheless foundin areaswhereprecipitationis concentrated in4 or 5 months. It is foundup to an altitudeof 1200m undertheseconditions;It can spread rapidly, for exampleit is moving at 40 km per year in the sparse savanna in theCentralAfrican Republic.
This colonising ability led us to consider that the African type may be particularlyaggressive, and we undertook a comparison, using the principles of descriptive taxonomy, ofabouta hundred samplesfromSouth America, theCaribbean, Asiaand Africa. Differentiationwasnotpossibleby thismeans, andtheworkwascontinued bychemotaxonomic researchusingenzymatic markers.
MATERIALS AND METHODS
Plant material
Chromolaena odorataseedsfrom America, AsiaandAfricawerecollectedandsentto thelaboratory for the investigation. Someweregerminated in a greenhouse and a varyingnumberof plants studiedaccording to the germination percentage of the differentbatches(Table 1).
72

Table 1: Origins of the seeds and.plants investigated
Origin of the seeds
Indonesia - West Kalimantan(Borneo)Indonesia - Sumatra- GunungBatinFrenchGuiana
FrenchGuiana - Petit SautGabon - AlovungTogo - EtoCote d'Ivoire - AngedougouBenin- SekouCentralAfrican Republic- BohongCentralAfrican Republic- M'BaikiMontpellier - AMATROP(origin: Beheke 1988)
Year collected
1988
1988
1989
198819881989198819881988
1988
1989
METHOD
Number of plants studied
2
7
23225283412310
3
6
The techniques usedat all stages from the collection and treatmentof the plant material to thevisualisation of the enzymatic systemswereselectedand refined.
Collectionand treatment of theplant material
The material was extractedfrom germinated or ungerminated seeds and from young and oldleavesfrom plantsgrown in thegreenhouse. The leaveswere usedfresh or freeze-dried. •
.73

Techniques of analysis
Four extraction buffers were tested and the extracts obtained were usedfresh or frozen in liquidnitrogen:
2 phosphate buffers:A. Potassium phosphate (0.1 M). pH 7.2 + dithiothreitol (0.005 M)B. Sodium phosphate (0.1 M). pH 7.0 + cysteine (0.005 M)
2 Tris HCI buffers:C. Tris HCI (0.1 M). pH 7.2 + cysteine (0.005 M)D. Tris HCI (0.1 M). pH 7.2 + cysteine (0.005 M) + EDTA (0.0I M) + ascorbic acid (0.3 M) +mercapto-ethanol(2%) + Triton (10%).
Six migration systems were thenused on p% starch gels or II% polyacrylamide gels (fable2).
Table 2: Different types or electrophoretic migration used for the visualisation or theenzymatic systems
Gels Migration systems Gel buffers Electrode buffers
. Starch gel Hist 6 Histidine, pH 6 Tris, citric acid, pH 6Hist 8 Histidine, pH 8 Tris, citric acid, pH 8TC7 Tris, citric acid, ,Tris, citric acid, pH 7
pH7LiOH-borate Tris, citric acid, LiOH, boric acid,
pH 8.3 pH 8.3
Acrylamide gel TBE Tris, boric acid, Tris, boric acid,EDTA,pH 8.1 EDTA, pH8.1
AOD According to
Omstein-Davis (1964)
Finally. 17 enzymatic systems were studied:•-Malate dehydrogenase (MOH)-Alcohol dehydrogenase (ADH)-Isocitrate dehydrogenase (lCD)-6 phospho-gluconate dehydrogenase (PGD)-Shikinate dehydrogenase (SKD)
74

-Phosphoglucoseisomerase (PGI)-Leucine aminopeptidase(LAP)-Phosphoglutomutase (PGM)-Esterases (ES1j .-Peroxidases(PEROX)-Acid phosphatases (PA)-Diaphorases (DIA)-Catalases (CA1j-Glutamate oxaloacetate transaminase (G01j-Endopeptidases(ENDO)-Amylases (AMYL)-Hexokinase QIEXO)
RESULTS
The choice of plant material analysed
Using extractions from germinated or ungenninated seeds is very long and theenzymaticactivitydetected is very small.The techniquewas thus not used. The legibility of thezymograms prepared from freeze-dried leaves was poor.
Old fresh leaves displayed less enzymatic activity than young leaves collected frombeneath the tip of the plant, thatgave clearer zymograms.This technique was therefore chosenin spite of the problem of theduration of the procedure,as sampling, extraction, migrationandvisualisation of the enzymatic systems had to be carried out in a single day.
The extracts were also used freshsince this gave much more distinct zymograms thanthose from the extracts frozen in liquid nitrogen.
Choice of extraction buffers
Buffer D (Tris HCl, pH 7.2) was chosen from the four buffers tested as it gave theclearest zymograms,
Choice of gels. enzymatic· systems for analysis and migrationsystems
. Someof theenzymaticsystemsdisplayedlittleor noactivity.The zymogramsof otherswere not clear and hence could not be interpreted. Only seven enzymatic systems and threemigrationsystemswerechosen(Table3) for their highlevelofenzymaticactivity andgood bandresolution.The results were all obtained by carrying out migrationon starch gels at 40 niNgelfor 6 to 7 hours. .
75

Table 3: Enzymatic systems and migration systems used
Enzyme systemsMigrationsystems MDH ICD LAP PEROX PGM PGI DIA
TC7 X XLiOHB X XH6 X X X
Study of enzymaticdiversity
Two of the sevenenzymatic systems.MDH and DIA. were still monomorphicfor the87 individualsstudied.Amongtheothersystems.LAP.PGM.PGIand ICDdisplayedenzymaticpolymorphism. They differentiate firstly the specimens from Guiana, which are all homogeneousfor the four systems.and secondlythe specimensfromAfricaand their progeny obtainedat theAMAmop laboratory.fromBorneoandSumatrawhicharealso similartoeach other withregard to the four systems (Figs. 1.2 & 3).
The PEROX system is also polymorphic and made it possible to visualise 3 genotypes (Fig.4) :
1 genotype specific to the individualsfrom Guiana, which were all similar to each other,
2 genotypes foundboth in all the samplesfrom Africancountriesand in those from BorneoandSumatra.
DISCUSSION AND CONCLUSION
This preliminarystudy shows that enzymaticpolymorphismexists in C. odorata andthat five of the sevenenzymatic systems tested were polymorphicin all the samples analysed.
The Guiana clones stand out clearly from the African and Asian clones in theses fivesystems.The secondtwogeographicalgroupsaresimilar in four of the five enzyme systemsandheterogeneous in the fifth. These results support the theory that C. odorata was introduced toAfrica from Asia.
The only American plants available were from French Guiana. These displayed adifferentenzymatic profile to that of the plantsof other origins. However. C.odorata is widelydistributed in tropical America. and in the light of its varied reproductive faculties - sexualreproduction with a high rate of polyploidyor apomixis (Ghosh 1961;Powell & King 1969) there is probably enzymaticpolymorphism within the Americanpopulations.The preliminaryapproach reported here tends to show simply that C. odorata was not introduced to Asia from
76

French Guiana.This researchwill thereforebecontinuedwitha largernumberof plants of morevaried origins to complete and refme theseresults,
REFERENCES
Bennett, ED. & V.P. Rao. 1968. Distributionof introducedweed Eupatorium odoratumLinn.(Compositae) in'Asia and Africa and possibilities of its biological control. PANS. 14 :227-281.
Delabarre, M. 1977. Incidence agronomique du developpement de Eupatorium odoratum(composee) en Cote d'lvoire. Thesis #38. Univ. Cote d'lvoire, 108p.
Ghosh, RB. 1961. Chromosome number of some flowering plants. Curr. Sci. 30: 73.
Ivens G.W. 1974.The problem of Eupatorium odoratum L. ,in Nigeria.PANS. 20: 76-82.
King, R M. & H. Robinson. 1970. Studies in the Eupatorieae (Compositae) 20. The GenusChromolaena. Phytologia 20: 196-209.
King, R M. & H.Robinson. 1987.The Generaof the Eupatorieae(Asteraceae).MonographsinSystematic Botany, vol. 22. Missouri BotanicalGarden, 581 p.
Miege, J. 1955.Les savaneset foretsclaires de Cote d'Ivoire. IV;EtudesEbumeennes. INFAN.Dakar. 67 p..
Omstein, I. & BJ. Davis.1964.DiscelectrophoresisI: Backgroundand theory.Ann.N.Y. Acad.Se. 121: 321-349,404.
PoweD, A.M. & RM. King. 1969.Chromosome number in the Compositae: West IndianSpecies. SIDA 3: 319-320.
77

TYPES OF ZYMOGRAM OBSERVED IN THE SAMPLES FROMFRENCH GUIANA, ASIA AND AFRICA
:Gulana iAsia- Afr ica
t-Leuclne aminopepti dase (LAP)
,.i
3-Phosphoglucose Isomerase (PGIl
2-Phosphoglucomutase (PGMI
78
Gulana Asia-Africa Asia - Africa
4-Peroxldase

MECHANICAL, CULTURAL AND CHEMICAL CONTROLOFCHROMOLAENA ODORATA
R. MUNIAPPAN ANDMAR! MARUTANIAgricultural ExperimentStation
University of GuamMangilao, Guam96923U.S.A.
ABSTRACT
Mechanical control is one of the common methods used for control of Chromolaenaodorata in manycountries. It is taborintensiveand providesonly short termcontrol. Culturalmethodin plantations usingmulchis effectivehowever, it is also labor intensive. Availabilityof mulch can be a problem. Cultural control utilizing various legume ground covers inplantations have been recommended in many countries. Signal grass in pastures has beenreportedto beeffectiveinreducing theincidenceofC.odorata inChina TriclopyrisaneffectiveherbicideagainstC.odorata. Otherherbicides usedare2,4-Damine,glyphosateand picloram.
C. odorata is a perennial, bushy and scrambling weed. It is neotropical in origin andintroduced to restof thehumidtropical regionsof theworldexceptsomecountriesin thePacificandAustralia It is moreofa menace in plantations of perennialcropssuchasoil palm,coconut,rubber,fruit treesand foresttrees. Pastures,vacantlandsandroadsidesare also infestedby thisweed. Differentkinds of control programshave been adopteddepending upon the resourcesavailable.
MECHANICAL CONTROL
Handweeding isa commonpracticein manycountries. Hoes,machetes and bushknivesare used for this purpose. Are and Folarin (1970)recommended slashing four times a year incocoa plantations. Komolafe (1976) suggesteda combination of slashing, ring weeding andmulching in Nigeria. Ojuederie et al. (1983) found slashing regimes in oil palm plantationsincreasedbunchyield.Liggitt(1983)andErasmus(1988)reviewedmechanical controlmethodsin South Africa Mechanical methods include labor intensive hand weeding, digging anduprooting and useof machinery suchas brushcutters,mowers,tillers,plowsand other tractordrawn equipments. Mechanical control provides short term control. A combination of othermethods suchasculturalandchemical weedcontrolprograms havebeenfoundto beeconomicaland effective.
79

CULTURAL CONTROL
Mulching: Mulching is done mostly in plantation crops around the bases of trees.Although this method effectively suppresses C;. odorata, high labor cost and problems withavailability of mulching materials limit its use.
Covercrops: Salgado(1972)foundTephrosia purpureaplantedas acovercrop incoconutplantations in Sri Lanka prevented the establishmentof C. odorata. Rai (1976) recommendedPuerariaphaseoloides as a cover crop in rubber plantations in India. Castillo et al. (1977)mentionedthatplantingLeucaena leucocephala inpasturesreducedC.odoratapopulation in thePhilippines. Puerariaphaseoloides, Calopogonium mucunoides, Centrosema pubescens and
. Yigna unguiculata were recommended cover 'crops in West African plantations (Komolafe1978).TorresandPaller(1989)suggestedplantinglegumeslikeCalopogoniummucunoidesandCentrosemapubescensandthegrass,Setariasp.immediatelyafterburningtheinfestedarea. Wuand Xu(1991)statedthatsignalgrass(Brachiaria decumbens) inpasturessuccessfullycompetesand-reducesthe incidenceof C. odorata in China.
CHEMICAL CONTROL
Most chemical control experiments on C. odorata were conducted in the Philippines,Indonesia, India, West Africa and South Africa, In the Philippines, Madrid (1974) found 2,4Dtobeeffectiveon seedlings,but for maturestands,2,4-Dhad tobecombinedwith either2,4,5T, picloramor Dicamba. Tumaliuanand Halos (1979)found Gramoxone(Paraquat20%) gavea fair control of C. odorata for a short period. According to Castillo et al. (1980) brushing orspraying the herbicide,Tordon'101 mixture reduced the incidence of C. odorata in pastures.
In Indonesia,Risdiono (1975)determinedpicloram 1kg/ha to be effective. Soerjaniet al.(1975)stated that picloramis used for C. odorata control in rubber and coconut plantationsandMCPAand2,4-D (aminesalt) in lowlandrice fieldsinJava. Tjitrosemitoetal. (1986) found2,4D, triclopyr or picloram are effective against C. odoratain rubber plantations.
In India, George (1968) mentioned that Gramoxoneat 0.5 Kg/acre provided satisfactoryresults, whereas Nair (1973) stated that Gramoxone at 0.3% concentration was not effectiveagainstC.odorata inopen fields.:Rai( 1976)suggestedGramoxone2.5litersplus 2,4-D sodiumsaltat2.5Kg sprayed3 to4 timesa yearwaseffectivein rubberplantations. Mathewetal. (1977)recommendedParaquatandFennoxone, and Borthakur(1977)suggested2,4-D forcontrolof C.odorata. .
In West Africa, 2,4-D was recommended by Sheldrick (1968), Ivens (1974), Martin(1977), Parker (1978) and Delorme (1979). Other chemicals found to be effective against C.odorata were glyphosate (Ivens 1974, Parker 1978). Dowco 290 (3-~chloropicolinic acid)(parker 1978).Tordon 101and Roundup(Dufouret al. 1979),2,4-D plus 2,4.5-T in ester (Ivens1974. Delorme 1979) and 2,4,5-T. glyphosate, picloram or 3.6-DCP (Quencez and Dufour1982).
80

Liggitt(1983)andErasmus (1988) reviewed chemicalcontrolin SouthAfrica. Erasmusand van Staden(1986, 1987) recommended triclopyrforcontrolof C. odorata.
In conclusion, mechanical control is labor intensive and it is not long lasting. Culturalcontrol is long lasting; however, either a mechanical or chemical control program has to becarriedoutinitiallyto implementthecultural control. Chemical controliseffectivebutexpensiveand posessomeenvironmental problems.
REFERENCES CITED
Are, L. A. & J. O. Folarin. 1970. Cultural weed control experiment. Ann. Rep. Cocoa Res. Inst. Nigeria1968-69. .
Botthakur, D. N. 1977. Mihmia and Eupatorium, two noxious weeds ofN.E Region. Indian Farming26:48-49.
Castillo, A. C.•F. A. Moog & C. Pineda. 1977. Introduction of ipil-ipil in "gonoy" infested pastures.Philippines J. Animal Industry 32:1-10.
Castillo, A. C.•E. M. Sena& F. A:Moog.1980. Prevalence of Chromolaena odbrala(L.) R. M.King andRobinson under different grazing intensities and methods of weed control. 17th PSAS AnnualConvention and Scientific Meeting. PICC. Manila, Nov. 21-22.
Delorme, M. 1979. Weeding ofperennial oil crops. Particular cases of young crops and control ofharmfulweeds. 01eagineux 34:4-6.
Dufour, F.•P. Quencez.& D. Boutin. 1979. Techniques for the chemical control of Eupaloriwn odoraiumin oil palm plantation. Oleagineux 34: 223-227.
Erasmus, DJ. 1988. A review ofmechnical and chemical control of Chromolaena odorata in South Africa.Proc. First International Workshop on Biological Control of Chromolaena odoraia, Bangkok.34-40. .
Erasmus, D. J. & J. van Staden. 1986. Screening of candidate herbicides in field for chemical control ofChromolaena odorata. S. Mr. J. Plant Soil 3:66-70. .
Erasmus, D. J. & J. van Staden. 1987. Chemical control ofChromolaena odorata: efficacy of triclopyr andglyphosate applied to regrowth. Applied Plant Science 1:39-42.
George, K. 1968. Herbicidal control of Eupeuorium odoratum: Indian Forester 94:817 -818.
Ivens, G. W. 1974. The problem of Eupatorium odoratum. L. in Nigeria. PANS. 20:76-82.
Komolafe, D. A. 1976. Weed problems in tree crops in Nigeria. PANS. 22:250-256.
Komolafe, D. A.1978. Weeds and their control in the humid andsubhumid tropics. Proc. Conf. Inter. Inst.Tropical Agric. 344-349.
81

Liggitt, B. 1983.The invasive alien plant Chromolaena odoraia; with regard to its status and control inNatal. Mono. 2, Inst, NaL Res., Univ. of Natal, Pietermaritzburg, Rep. South Africa.
Madrid, M. T. Ir. 1974. Evaluationof herbicides for the control of Chromolaena odoraia (L.) R.M. Kingand H. Robinson. Philippines Weed Sci. Bull. 1:25-29.
Martin, M. 1977.Theuse of herbicide products in oil palm growing. Oleagineux 32: 479-482.
Mathew, M., K. I. Punnoose & S. N. Potty. 1977. Report on the results of chemical weed controlexperiments, the rubber plantations in South India. 1. Rubber Res. Inst Sri Lanka 54:478-488.
Nair, P.N. 1973. The effect of gramoxone applicationon Eupatoriumodoraium. Indian Forester 99:4348.
Ojuederie, B. M., G. O. Iremiren & S. N. Utulu. 1983. Effects ofvarious inter-row slashing regimes and .size of weeded rings on the early growth, flowering and bunch yield of the oil palm. 1.NigerianInst, Oil ptilin Res. 6:322-334.
Parker,C. 1978.Pot experimentwithsomenew herbicideson tropicalperennial weeds. Solwna TroisiemeSymposiwn Sur le Deserbagedes Cultures Tropicales 288-296.
Quencez, P. & F. Dufour. 1982.Chemical weed control in palm plantations. n. Usu8Iherbicidal activeingredients for oil palm culture and treatment techniques. Oleagineux 37:107-113.
Rai, S. N. 1976. Eupatoriumand weedicides. Indian Forester 102:449-454.
Risdiono, B. 1975.The inter-influencebetweenChromolaena odorataL. andBrachiariabrizamha Stapfand theeffect ofPicloram on theseplants. Proc. 3rd Indonesian Weed Sci. Conf., Bandung, Iune24-28,367-376. .
Salgado, M. L. M. 1972.T'ephrosia purpurea (Pila) for the control of Eupatoriumand as a green manureon coconut estates. Ceylon Cocon. Plrs, Rev. 6:160-174.
Sheldrick, R. D. 1968.The control of Siam weed in Nigeria. 1. Nigerian Inst. Oil Palm Res. 5:7-19.
Soerjani, M., A. Soedarsan, S. Mangoensoekarja,T. Kuntohartono & M. Sundaru. 1975. Weed problemsand prospects for chemical control in Indonesia. 5th Conf. of the Asian Weed Sci. Soc. pp. 1822.
Tjitrosemito, S., S. Sastroutomo & I. H. Utorno. 1986.Weed management in a young rubber plantation inIndonesia. Weedwateher 1(supplement),4pp.
Torres, 0.0. & E.C. Paller, Ir. 1989.The devil weed (ChromolaeNJ odorata RM. King and H. Robinson)and its management. SEAWIC Weed Leaflet 4:1-5.
Twnaliuan, B. T. & S. C. Halos. 1979. Screening herbicidal preparations and mixtures for clearingreforestation areas. Sylvatrop Philippines Forest Res. 1.4:151-159.
Wu, R. & X. Xu. 1991. Effect of planting signal grass (Brachiaria decumbens Stapt) on feijicao[Chromolaena odorata King and Robinsonj-in Southern Yunnan, PRC. Second InternationalWorkshop on BiologicalControl of Chromolaena odoraia; Bogor, Indonesia, Feb. 4-8,1991.
82

CULTURAL CONTROL OF FEUICAO (CHROMOLAENA ODORATA. (L.).R. M. KiNG AND H. ROBINSON) BY PLANTING SIGNALGRASS
.(BRACHIARIA DECUMBENS STAPF) IN SOUTHERN YUNNAN,PEOPLE'S REPUBLIC OF CHINA
WURENRUNLanzhouInstitute of AnimalScience,Chinese Academy
of Agriculture Sciences, Lanzhou, Gansu730050People'sRepublic of China
XUXUFJUNYunnan Provincial Livestock and PastureResearchCenter
Xiaoshao, Kunming, Yunnan650212People's Republic of China
ABSTRACT
The noxious weed, Chromolaena odorata (1...) R. M. King & H. Robinson (known asfeijicao in China),was introduced to Yunnan Province of China via SoutheastAsiancountriesfrom theNeotropics, It has seriously affectedthe nativevegetation of the southwest, west.andcemral parts of this province for the past twenty years. It has caused loss of productivity ofrangelands and hindrance to the development of animal industry. In 1985, signalgrass(Brachiarla decumbens Stapfcv. Basilisk), wasplantedin pasturesand effectivelysuppressedfeijicao. Signalgrass has a betterdry matter yield,highcoverage,and formsa condensedtuftlayer.Feijicao infestedfieldsare burned, uprooted and allowedto dry in the sun. Signalgrassseeds arebroadcastedandfertilized foreffectiveestablishment Samplingof 10onesquaremeterquadrats within the signalgrass fieldsand one outsidearea was done to compare the incidenceof feijicao, Incidence of feijicao gradually declinedin the first three yearsafter establishmentof signalgrass. After threeyearstherewas no incidence of feijicao in the signalgrass pastures.
INTRODUCTION
Chromolaena odorata(1...) KingandH.Robinson isknownasfeijicaoinChinese,meaningairplaneweed. It occupieslargeareas in-southwest, westandcentralpartsof YunnanProvinceandseverelyinfestslocalrangelands and farmlands. It spreadfromtheneotropicsviaSoutheastAsian countries to southern China. A large part of this southern area invaded by feijicao issituatedsouthof theTropicof Cancer. Yunnanhasdifferenttopographies andclimaticregions.Thealtituderangesfrom 100to 1000m abovesea level. The weedinfestedareasare locatedupand down the hillsides from 500 to 1000 m. Generally, the rate of occurrence of feijicaogradually decreases as the elevation increases.
83

Feijicaoisa perennial plantwhichproduces thickfoliagewhenyoung. Matureplantshavemanysturdyandovergrown sterns. Theyoungsternsaresomewhatsoft,'pithy in the centerandthe uprightmain stern sometimes reaches a heightof 2 to 3 m. Leavesareopposite.rhomboidin shapeandserrated. The inflorescence headis umbelled, and theflowers crowdedand yellowcolored when mature. They produce some thousands of pentagonous achenes (seeds) with.windborne pappi. Seedsare very light and small. weighing 0.05 gm /1000.
Feijicaothrivesbest on fertileand soft soils and grows well on acid soils. If grown onalkalinesoils.theleaveswillturn yellow'andwilt, Whenthetemperature fallsto to 1-2C. chillyspotsappearon the leavesand whenitdrops to 0 C. all the above-ground partsof the plantwillbe frozen.
Signalgrass (Brachiaria decumbens Stapf) is indigenous to the rangelandsof the GreatLakePlateauin Uganda. It was introduced fromKampala. Ugandato Queensland. Australia in1930. Commercial seedsof the cv. Basilisk are derivedfrom the accessionnumberCPI 1694.It was multiplied by CSIROand approved forcommercial releaseby QueenslandDAS (Orarn1990).
This cultivar is welladapted to humid tropical areas with a dry season not exceeding45 months. It ismoredroughttolerantthanparagrass (B. muticay. Itgrowswellon steephillsidesof southernYunnan where paragrass can not establish. It forms an aggressive. high-yieldingsward, whichrespondsmarkedly to N fertilization. Itsdry matteryield ishighand it has a lowcrudeproteincontent. It is palatable exceptwhenthe stemsare mature. It standsup fairly wellto heavy stocking and trampling. Seed production is good duringthe first twelvemonthsfromplantingprovidedthegrass is not cut frequently. Germination of the seedscan be improved bytreating with sulphuric acid for 10 to 15minutes. Thiscultivarmay be plantedmixedwith thelegume.Centrosema pubescens Benth. (Orarn 1990).
In 1983.ourCenter introduced thiscultivarfrom Australia. It isa tetraploid(chromosomenumber4n =36;x =9) and an obligateaposporous apomict(Pritchard 1%7).
In 1985, we started an experiment in the southern part of Yunnan Province at theManzhongtian Ranchof SimaoCounty(geographical location: 2247' N and 101 17'E; altitude1030mabovesea level). Theyearlyaverage precipitation isabove2200mmandtheannualmeantemperature is 20.6C. Soiltype is redand brown lateriuc, pH 5.6in theupperlayer oCo-lO cm.Weestablishedimproved pastureswiththeaimof developing and increasing animalproduction.However, themainproblem encountered wastheinvasion ofail thelocalgrasslandsby feijicao.Duringtheevaluation trials of introduced grasses,wediscovered thatthiscultivar of signalgrassisveryadaptive andproduced a dry matteryieldof 11.12ton/hawith4 harvestsperyearin 1987(Xu Xuejunet al. 1989). ,It isable toforma moreaggressive sod-layer, withgoodcoverageandhigh density,and witha criss-crossed underground root system. In this paper, we present theculturalmethodof controlof feijicao by plantingsignalgrass.
84

MATERIALS AND METHODS
InFebruaryto March1985,'when feijicao wasdispersing seeds, weburnedinfestedwastelands,dugout the rootsmanually andlet them dry for a periodof twoto threeweeksin the sun.Before the advent of the rainy season. seeds of the signalgrass cv, Basilisk were broadcastedmanually at the rate of 7 kg/ha. At the sametime. fertilizerwas appliedat a rate equivalentto20 kg PIha, 9 kg S/ha. 22 kg K/haand 92 kg N/ha. No fertilizerwas appliedafterwards. Thegrazingof livestock on these newlyestablished pastures was controlled in order to develop ahealthy sward.
Ten quadratsof 1 squaremetereach were used at random in the signalgrass-establishedpasturesfor estimating feijicao infestation. The same methodwas used forestimatingfeijicaoinfestation in non-signalgrass pasturesin the neighboring areas.
Soil samples were taken at a depth of 0-10 cm and analysed for N, P and K. Kjeldahlmethodwas usedfor N-determination. P and K wereseparately determined usingspectrophotometric means with0.5 N NaHC0
3and an atomicabsorption spectrometer.
RESULTS AND DISCUSSION
Themeanvaluesof numberofplantsof feijicao persquaremeteron signalgrassstandsforsuccessive yearsindifferentdatesofsowing(August. 1985; July. 1986; August,1987;eachJunefrom 1988to 1990)weredetermined (Table 1). The resultsof samples takenon November10,1990indifferentfieldsandinfeijicao-infested rangelandaregiveninTable2. TheP andKstatusin the signalgrass pasturesand feijicao-infested area were the same, however, N was a littlehigherin the formerpastures.
Farmersin Yunnan Province spenda lotof manpower onburninganddiggingoutfeijicaowithout much success. Feijicao invasion of pastures has caused heavy losses to farmers.Establishment of signalgrass in pastures haseffectively prevented germination of the seedsandgrowthof the seedlings of feijicao. A fewseedlings of feijicao maygrow in the first two yearsafterestablishment of thesignalgrass; however, in the thirdyearvirtually no feijicao is notedinthesefields(Table 1). Plantingof signalgrass tocontrol feijicao notonly helps to get rid of thisnoxiousweed,but hasproduced theaddedbenefitof improved pasturesfor feeding livestock atManzhongtian (MZT) in southern Yunnan. Xu Xuejunet al. (1989)reponed that"Signalgrassshowed greatadaptability at MZTwhereitsaggressive competitive abilityallowedit tocontrolthe weedEupatorium odoratum (=c. odorata). It has theadditional advantageof beingeasy toestablishon steep hills."
In Yunnan, four species of Brachiaria were introduced from Australia: B. humidicola(Rendle) Schweick cv. Tully,paragrass, B. muiica (Forsk.) Stapf andB. ruzizensis R. Germ.etEvrad. Ourevaluationtrialsat MZT showedthatBr. decumbens is thebest,havingnotonlythe .
85

ability to suppress feijicao out also possessing goodseedingabilityand disease resistance(XUXuejun et al. 1989). According to WuRenrun et al. (1986), thereare4 spp,of wildBrachiariainYunnan: B. ramosaStapf,B. eruciformisGriseb.,B. subquadripara Hitehc.andB. villosaA.Camus.
The N contentshowedan increase in thefifth yearafter establishment of the signalgrass;P and K contents werethe same in signalgrass and feijicao soils. The increasing N content insignalgrass soil is probably due to the application of N fertilizers during the first year ofestablishment andadditional N cyclingfrom livestockmanure. The increasein N contentis anaddedadvantage for cultivating signalgrass. The crudeproteincontentof feijicao was27.65%,while the crude protein content of signalgrass was 3.13% (Kui Jiaxiang et al. 1990a). Tomaintain and increasethecrude proteincontentof signalgrass at MZT,we haverecommendedapplications of N fertilizer after cultivation (KuiJiaxianget al. 1990b)(which, however, is notveryeconomical owingtohighercostof fertilizer application) and mixedplantingof legumeswithsignalgrass. Thesignalgrass cultivated sitesatMZTareall steepslopesonhillsides. Hence,werecommend contourparallel planting of alternaterowsof grassanda legume. This provedbetter than planting mixedseeds of grassand a legume. This alternate planting increased thecrudeproteincontentof signalgrass from 3.13to 5.65%,(KuiJiaxianget al. 1990b). Accordingto Walsh (1959) Basilisk has performed well with the legume Centrosema pubescens inQueensland, Australia. Such a mixture is also utilized in China (XU Xuejun et al. 1989);however, at MZTRanch we usea nativelegume, Flemingia macrophylla, withthe signalgrass(WU Renrun et al. 1989).
Initialresultsusing various herbicides forcontrol offeijicao indicated that glyphosateandParaquatwerenot effective. 2,4-DA waseffectivein controlling this noxious weedand it didlittledamagetograsses. We haverecommended spotapplication of 2,4-DA tocontrolfeijicaoinsignalgrass fields, as an effectiveand economical method. Wider use of herbicides is notrecommended as it is not economical for forage production at MZT.
Table 1. Average Number of FeljicaoPlants in Signalgrass Pasture Established for Six Years and aNeighborlng Wasteland (Nov. 10, 1990).

Table 2. The Status orN, P and K In Soils or Feijlcao Waste Land and Slgnalgrass ImprovedPastures after Six Successive Years or Establishment
Ye.8lS of Establislunent Wasteland·
1 2 3 4 5 6
1990 1989 1988 1987 1986 1985
N(%) 0.19 0.17 0.19 0.20 0.13 0.20 0.15 ..P(ppm) 2.4 0.5 1.7 1.7 1.7
K(ppm) 28 14 24 12 13 18 18
• Soil samples taken on November 10, 1990 from theneighboring waste lands.
•
87

88
Figure 1. Signal grass pasture and the neigh boring feijicao infested wasteland .
Figure 2. Signal grass field a long the r iverside in the MZT ranch.

ACKNOWLEDGEMENTS
WethankMr. AlanM.Vaughan, Australian expert,forhishelpin introducing signalgrass,andmanyoftheresearchworkers atManzhongtianandYunnan ProvincialLivestockandPastureResearchCenter for their assistance in this work.
REFERENCES
Kui Jiaxiang, Deng Jufen & Zhou Ziwei. ·1990a. On the application of various herbicides to controlFeijicao (Chromolaena odoralq) in the humid-warm subtropical climate regions of YunnanProvince, China, Yunnan Provincial Livestock & Pasture Research Center (ypLPRC), (in press).
Kui Jiaxiang, Deng Jufen & Zhou Ziwei. 1990b. The preliminary report on mixed planting of legumesand swordgrass (Brachiaria decumbens Stapf) in pastures.
Dram, R.N. 1990. Register of Australian herbage plant cultivars, 3rd Edition, CSIRO, Australia,p.89-90.
Pritehard, A.I. 1967. Apomixis in Brachiaria decumbens Stapf. 1. Aust. Inst. Agric. Sci. 33: 264-265.
Vaughan, A. M. 1988. Weed problems in tropical pasture management. Training Manual for TropicalPasture Production in Yunnan, China. 95-98·.
WaIsh, S.R. 1959. Improved pastures will fatten cattle in far North. Queensland Agric. 1. 85: 576-592.
Wu Renrun, Zhang Deyin & Lu Xinshi. 1986. A preliminary report on germplasm resources of grasses,legumes' and other forage plants in Yunnan Province of China (1) Crop Pt. Germ. Res., (1): p.1-8: (2) Sichuan Grassland, 1987 (2): 1-17, With List in Germplasm Resources in Yunnan,Grasses and Legumes, p. 1-24.'
Wu Renrun, Zhang Deyin & Lu Xinshi. 1988. On the pattern of distribution of two pernicious weeds,Ageratina adenophora and Chromolaena odorata and biological control measures taken inYunnan Province, China. (Abstract). In: R. Muniappan (ed) Proc. First Intern. Workshop BioiControl Chromolaena odorata, p. 82 ., Agri. Expt. Sta., Univ. of Guam, Mangilao, Guam.
Wu Renrun, Zhang Deyin & Lu Xinshi. 1989. The study of grasses, legumes and other forage plantsgermplasm resources in Xishuangbanna of Yunnan Province, China. XVI InternationalGrassland Congress, Nice, France, 1989, p. 1541-1542.
Xu Xuejun, Wang Tu & Guo Zhenyun. 1989. Preliminary results of field trial evaluation of sometropical pasture species performance in the humid and sub-humid tropical zones of YunnanProvince, Southern China (1), XVI International Grassland Congress, Nice, France, 1989, p.1511-1512.
89


PROGRESS ON BIOLOGICAL CONTROL OF SIAM WEED,CHROMOLAENA ODORATA IN THAILAND
BANPOT NAPOMPETH AND AMPORN WINOTAINational Biological Control Research Center (NBCRC)
Kasetsart UniversityP.O. Box 9-52, Bangkok, 10900 Thailand
ABSTRACT
Both augmentative and classical biological control of Siam weed, Chromolaena odorata, were
attempted in Thailand. Of the native arthropod fauna only an aphid complex consisting of Aphis gossypii
Glover, Aphis craccivora Koch and Aphis spiraecola Patch, the amaranth weevil Hypolixus truncatulus F.,
and an eriophyid mite, Acalitus adoratus Keifer, were of some importance. The top shoot miner,
Melanagromyza eupatoriella Spencer, introduced from Trinidad in 1978, failed to survive in the quaran
tine. During 1986-1988, an arctiid moth, Pareuchaetes pseudoinsulata Rego Barros, was introduced from
Guam. The moth could not be recovered after repeated field releases. In laboratory mass-rearing, P.pseudoinsulaia lost its egg hatchability after the fourth generation necessitating reintroductions. Thispreliminary result was discouraging and there is an urgent need to formulate a new strategy on biological
control of C. odoraia in Thailand.
INTRODUCTION
The Siam weed, Chromolaena odorata (L.) R.M. King and H. Robinson (Asteraceae), isone of the target weeds of exotic origin subjected to biological control attempts in Thailand. Incontrast to other exotic weeds with satisfactory achievement and success witnessed, e.g.,alligator weed, waterhyacinth and the giant sensitive plant, biological control of C. odorata hasnot yet been very encouraging. So faran arctiid moth, PareuchaetespseudoinsulatauegcBarros(Lepidoptera:Arctiidae), has been introduced and released but its establishment is yet to beconfirmed, A failure to obtain reasonably high egg hatchability after four generations oflaboratory mass-rearing has prevented moth-rearing on a large scale for additional and continuing field releases. There exists a need to search and screen native natural enemies showingbiological control potential for augmentative use as well as to introduce additional exotic naturalenemies to the country for further utilization.
Earlier attempts at biological control of C. odorata carried out in Thailand from 1975 to1988 were described by Napompeth (1982) and Napompeth et al. (1988). No native naturalenemies were found having adequate potential as biological control agents. The top shoot miner,
91

MelanagromyzaeupatoriellaSpencer(Diptera:Agromyzidae),andanarctiidmoth,Pareuchaelespseudoinsulata Rego Barros (Lepidoptera: Arctiidae), were introduced from Trinidad and Guamin 1978 and during 1986-1987, respectively.
The present paper attempts to review further progress, although negligible, on biologicalcontrol of Codorata in Thailand.
NATIVE NATURAL ENEMIESOF C. ODORATA
Being an exotic weed species C. odoratacontains a small number ofassociated insect andother arthropod species outside of its native range ofCentral and South America. It is also lesslikely that these euryphagous species would be of any potential for further utilization inbiological control of this weed. In Thailand C. odorata was found to harbor a number of knowninsect pest species and several polyphagous insects and plant pathogens. Of all the arthropodspecies found associated with C. odoraia,those that could cause some noticeable damage were:
I.An Aphid complex consisting of Aphis gosspii Glover, aphis craccivor Koch andAphis spiraecola Patch (Homoptera: Aphididae).
2.Amaranth weevil, Hypolixus.truncatulus F. (Coleoptera: Curculionidae).
3.Eriophyid mite, Acalitus adoratus Keifer (Acarina: Eriophyidae).
The current impact of these native natural enemies as natural biological control agents of .C. odorata in Thailand is as follows.
Aphid complex, Aphis spp.
Either alone or collectively A. gossypii, A. craccivora and A. spiraecola can induceconsiderable damage to the growing top shoots of C. odorata. Curly, stunted and malformed topshoots of C. odorata, sometimes called "phyllodies", are very common and widespread in all C.odorata infested areas of the country. Such deformation is probably due to the toxin secreted byaphids in their feeding similar to the symptom of phytotoxemia. To a very large extent this typeof damage to the plant obviously obstructs flower bud formation and thus normal flowering.These aphids are widespread all over the country and have significantly reduced the spread anddensity of C. odorata in Thailand.
Amaranth weevil, Hypolixus truncatulus F.
H. truncatulus has been a major biological control agent for the spiny amaranth,AmaranthusspinosusL., in Thailand (Napompeth, 1982, 1989, 1990b). While the adult weevilscause no obvious damage, it is the grubs that form galls in the stem of A. spinosus which aredamaging. Only as recently as 1988adultsofH. truncatulushavebeenobserved to feed on young
92

branches and stems of C.odorata (Napompeth 1990a). When they occur in large numbers theweevils can causesubstantial damage to the weed. The distribution of H. truncatulusis, however,confined to the central plain areas of the country.
Eriophyid mite, Acalitus adoratus Keifer.
A. adoratus has been reported on C. odorata in Trinidad, Brazil and Bolivia since 1974 byCruttwell McFadyen (1988). It was detected in Thailand in the mid-1980s in the southernpeninsulararea ofthe country and its identity confirmed by Napompeth et al. (1988). A. adoratus .since then has spread northwards and can befound in all C.odorata infested areas. The infestedleaves, both young and mature, are not as deformed as those damaged by aphids, and floweringof the plant is apparently normal. Although widespread all over the country on C.odorata, theimpact ofA. adoratusas a biological control agent is relatively inadequate. It is also not knownhow A. adoratus reached Thailand.
EXOTIC OR INTRODUCED NATURAL ENEMIES OF C. ODORATA
Only two exotic natural enemies ofC. odorata have been introduced to Thailand. The topshoot miner, Melanagromyza eupatoriella Spencer (Diptera: Agromyzidae), was introducedfrom Trinidad in 1978. Unfortunately the culture perished under quarantine and thus no fieldrelease could bemade (Napompeth et al. 1988). The arctiid moth, PareuchaetespseudoinsulataRego Barros (Lepidoptera: Arctiidae), was introduced on several occasions from Guam during
, 1986-1988 (Table I). The firsttwo shipments received in 1986 were used in quarantine screeningand host specificity tests covering 48 species of plants in 25 families. Of all these plants, P.pseudoinsulatacould feed and survive on only two other asteraceous weeds, namely, AgeratumconyzoidesL. and Ageratina adenophora Sprengel. Biological studies and mass-rearing ofP.pseudoinsulata were made using the subsequent shipments received from Guam in 1986-1987.
Field releases were carried out at six locations in the northern highland areas ofChiang Maiand Chiang Rai provinces and at four locations at Khao Yai National Park in the central areaadjoining the rim of the northeastern plateau areas. Commencing in 1987 field releases werecarried out in.l988 (Table 2) but terminated at the end of 1988 due to failure to obtain more larvaefrom the laboratory mass rearing program.
Laboratory rearing of P. pseudoinsulata was observed to deteriorate after every fourgenerations when all the eggs laid failed to hatch. With no field population to help rejuvenatelaboratory cultures. two additional shipments of P. pseudoinsulata were made from Guam inMay and June 1988. It was suspected that sodium could play an important role in plant nutritionand subsequently egg hatchability (LW. Forno,pers. comm.). As aresult, 0.05% sodium chloride,solution was added to the soil for the potted C.odorata plants and leaves from these plants wereused for rearing the moth. Again after four generations of laboratory rearing all the eggs did nothatch (Napompeth I990a). .
93

Table 1. Introduction of natural enemies of Chromolaena odorata for biological controllnThaUand, 1978-1988. .
Natural Enemy
Melanagromyza eupatoriellaSpencer (Diptera; Agromyzidae)
Pareuchaetes pseudoinsulataRego Barros(Lepidoptera: Arctiidae)
94
Source .Date Remarks
Trinidad October 1978 No release
Guam March 1986 No release
May 1986 Quarantined
November 1986 Quarantined & released
July 1987 Mass-reared & released
November 1987 Mass-reared & released
May 1988 No release
July 1988 Mass-reared & released

Table 2. Field releases orPareucluJetes pseudoinsulata in Thailand during 1987-1988..
Date
February 1987
. January 1988
February 1988
March 1988
August1988
September 1988
October 1988
Location
Khao Yai National Park
Km #1 Chiang Mai-Phrao RoadMuang District, Chiang Mai
Km#50 ChiangMai-Phrao Road. Phrao District, Chiang Mai
Km #86 Chiang Mai-Phmo RoadPhrao District,Chiang Mai
Mae Kok River Bridge,Tha Ton, Mae AiDistrict,Chiang Mai
Pong Tarn,Ban Tha,ChaiprakamSubdistrict, Chiang Mai
Near ChiangDaoCave,Chiang DaoDistrict.ChiangMai
Khao YaiNational Park
Km #7 Prachinburi-Khao YaiRoadMuang District,Prachinburi
Khao Yai National Park
Khao Yai National Park
Khao Yai National Park
Khao Yai National Park
NumberReleased
2,000 larvae
600 larvae
200 larvae
200 larvae
200 larvae
200 larvae
200 larvae
600 pupae
200 pupae
3,000 larvae
200 larvae
500 larvae
1,000 larvae
95

"NATIVE" P. PSEUDOINSULATA • LIKE ARCTIID MOTH
Duringa fieldsurveyin 1989,aneggmassofan arctiidmothresemblingP.pseudoinsulatawas discovered on C. odorata in the central plain area. The adult moth resembles an arctiid,AmsaCla (Estigmene ) lactinea Cramer,whichis a knownphytophagous pest speciesof severaleconomiccrops. Thisarctiidmothwasable tocompleteits lifecycleonC.odorata. Additionalfieldsurveydidnotwarrantanyfurtherinvestigation: However, whentheadultmothisobservedon C.odorata plants, itcan easilybe mistaken forPcpseudoinsulaia. The larvaeare more hairy .and can easily be differentiated from those of P. pseudoinsulata.
DISCUSSION AND CONCLUSION
Preliminary workon biological controlofC.odoratainThailandcarriedout during19861988by introducingP. pseudoinsulata fromGuamdid not yieldencouraging results. Of all thearthropods surveyedand evaluatedin Thailandnoneof themcould be cost-effectively utilizedfor augmentative purposes. Being an exotic invading weed species, C. odorata should beamenable to classical biological control utilizing introduced natural enemies from its nativerange of origin. In spite of a wealthof information on phytophagous insects recorded from C.odorata in its native range and elsewhere by Cmttwell McFadyen (1988), relatively littleinformation on their potentialas biologicalcontrol agents is known except those few specieselucidated byCock(1984).Thereexistsan urgentneedtoexplorefurtherif thebiologicalcontrolprogram of C. odorata is to be realized and reasonably achieved on a world-wide basis, notmerely in a few locationsas now witnessed.
As in the case of Thailand, a new biological control strategy for C. odorata must beformulated. However, considering other C. odorata affected areas such a strategy should beequally applicable and should be devised in such a way that a more intensive regional andinternational effort isgivena highprioritytogetherwithadequatesupportprovidedtoa programsuchas theBiological Controlof Chromolaena odorataWorkingGroup underSEARS/lOBC.
96

REFERENCES
Cock. M.I.W. 1984. Possibilities for biological control of Chromolaena odoraia. Tropical Pest Manage ,ment 30(1): 7-13.
Cruttwell McFadyen, R.E. 1988. Phytophagous insects recorded from C. odoraiaiChromolaena odorataNewsletter No. 2. p. 5-23.
Napompeth, B. 1982. Biologicalcontrol research and development inThailand. Proc. Intl. Conf. PlantProt,in Tropics. p. 301-323.
Napompeth, B. 1989. Biological control of insect pests and weeds in Thailand. BIOTROP Spec. Publ,No. 36. p. 51-68.
Napompeth, B. 1990a. Contributions from Thailand. Chromolaena odoraia Newsletter No. 3. p. 6.
Napompeth, B. 1990b. Country report: Thailand. Biological control of weeds inThailand. BIOTROP Spec.Publ. No. 38. p. 23- 36.
Napompeth, B., Nguyen Thi Hai & A. Winotai. 1988. Attempts on biological control of Siam weed,Chromolaena odoraia, in Thailand. In: R. Muniappan (ed.)Proc. First Intl, Workshop on BioI.Control of Chromolaena odoraia. pp.57 -62. Agri. Expt, Sta., Univ. of Guam.
97


STATUS OF BIOLOGICAL CONTROL OF THE SIAM WEED,CHROMOLAENA ODORATA (L.) R.M. KING AND H. ROBINSON ON
POHNPEI, FEDERATED STATES OF MICRONESIA
NELSON M. ESGUERRA, WILLIAM S. WILLIAMand JUSTINO R. SMITH
Collegeof Micronesia Land GrantPrograms,P.O. Box 1179,Kolonia, Pohnpei, FSM 96941
ABSTRACT
Biological controlof theSiamweedonPohnpei,FederatedStatesof Micronesia involvedmass rearing the biological control agent, Pareuchaetes pseudoinsulata Rego Barros, in thelaboratory and mass releasing either larvaeor adults in field infestations of the weed.
Almost two years of field release did not result in establishment. It was observed thatpredatorssuchas groundlizards, spiders,birds,redand blackantspreyedheavily-on theadults,larvaeandeggs of P. pseudoinsulata.. OnOctober24, 1990,feeding injuryand thepresenceofa numberof caterpillars werenoticedin one of the releaseareas. It is planned that after furtherconfirmation of the firm establishment, fieldadaptedlarvaewillbecollectedforreleasein otherareas.About500adultswillbe releasedoneachsite to facilitate theestablishment in otherareas.
Problemsin mass rearing and mass releasing P. pseudoinsulata and techniques used toovercome the problems both in the field and in the laboratory are discussed.
INTRODUCTION
TheSiamweedhasbecomea seriousweedpest in theStatesof Pohnpei,Kosraeand YapintheFederatedStatesof Micronesia(FSM). Theweedis spreading rapidlyandis nowabundantin Madolenihmw and Kitti municipalities of thesouthernpartofPohnpei. It is spreadingslowlytoward Sokehsand isolatedpatchesof the weed are appearing on Koloniaand Nett.
In the two municipalities where they are present, Chromolaena odorata is usuallyabundantonvacantlands,pastures, roadsides, andcultivatedlands.TheweedseemstogrowwellonPohnpei,withanannualrainfallofabout500cmandtemperature rangefrom70· -92· F. Onceestablished, theweedformsa densemassofgreencoveroverland,impeding accesstoroadsandpathways. It becomesa fire hazardin dry seasons.
99

Becauseof the seriousnessof this weed in FSM, the College of MicronesiaLand GrantProgramdecided to includeC. odorataas a target for biologicalcontrol under the AgriculturaIDevelopmentin theAmericanPacific (ADAP)BiologicalControl Agent ExchangeProject. InFebruary1989,through thehelpoftheUniversity ofGuam,larvaeofPareuchaetespseudoinsulataRego Barros (Lepidoptera: Arctiidae)were brought to Pohnpei. .
This paper discusses the procedures and activities that were carried on and will beimplemented on Pohnpei aimed at controlling the Siam weed with the arctiid moth, P.pseudoinsulata.
MATERIALS AND METHODS
Laboratory rearing of P. pseudoinsulataRearing the caterpillarsof P. pseudoinsuiata to produce large numbers in the laboratory
followed the techniques used by Muniappan et al. (1989) and Seibert (1985) with somemodifications. The technique used was as follows:
Eggs of P.pseudoinsulata were laid in groupson Chromolaena leaves.Leaves witheggsweredetachedfromthe stemsand placed in ice cream cups withcovers.Eggs laid on screens inrearingcages wereallowedto hatchandnewly hatchedlarvaewerecollectedand placed in cupswithcoversand freshdetachedleavesof Chromolaena as food.Ventilationwasprovidedfor therearing cups by glueing muslin cloth on the cut portion of the lid. The first and second larvalinstars were maintained in these cups and fresh leaves were provided daily. The cups werecleanedevery2 daysand larvaefeedingon detachedleaveswere transferredto clean cups, thusavoidingaccumulation o(frass that couldbe toxic to the insect. Upon reaching the third instar,the larvaeweretransferred to25gallon-plasticcontainers.Chromolaena stemswith leaveswerekept in smallboules withwaterto preventwiltingof leavesandassuringlarvaeample supplyof freshleaves for twodays.The 25 galloncontainerswerecleanedof frass and dried leaveseverytwodaysandcaterpillarstransferredto' cleancontainerswithfresh leaves.When the caterpillars .were mature and about to pupate, fresh detached leaves were placed on the bottom of the .containersforpupation.Threedays after, the pupae werecollectedand placed inside the 30.4 x30.4 cm organdie nylon cloth and 30.4 x 30.4 x 60.9 cm fine meshed wire rearing cages. Cutshootsof Chromolaena kept in smallbottles with waterwere placed inside the rearing cages toprovide as oviposition sites for female moths. Pieces of cotton soaked in sugar solution werekept hanging inside the cages and replacedevery two days. Cotton soaked in salt solution wasalso hung inside the cages. These servedas food for the moths.
Population Density of Chromolaena at the release siteA 5 x 5 meterwoodenquadrat was thrownin the designatedrelease site and the number
of Chromolaena plantswerecounted.Tenweedswererandomlyselectedinsidethequadrat,Thesize of stems and heightof these ten weeds were measuredusing a meterstick.
100

Field Release and Monitoring of P. pseudoinsukua
Larvae:Laboratory reared third instar larvae ofP.pseudoinsulatawere placed in cups with covers
and brought to the release sites. Each release site was heavily infested with luxuriantly growingChromolaena. The larvae were individually picked from cups using a camel-hair brush andreleased on Chromolaena. Signs of feeding and presence of larvae were monitored two days andone week after their release.
Adults:Newly emerged adults ofP.pseudoinsulata from laboratory cultures were released in the
release site. One week after their release, observations were made for the presence of eggs andnewly hatched larvae on Chromolaena.
'Adult moths were also released in a 1.17 x 1.17 x1.22 m field cage with naturaUy growingChromolaena inside for oviposition. Since earlier attempts at facilitating the establishment ofadult moths inside the cagewere a failure due to predators, the cage and Chromolaena includingabout 1.2 m around the cages were sprayed with diazinon at 1 kg ai/ha to kill predaceous insectsand other organisms.
RESULTS AND DISCUSSION·
Other Insects Feeding on Chromolaena in the fieldIn the course of surveying the extent of Chromolaena infestation on the southern part of
the island, some insects were observed feeding on the weed although it is very unlikely that theycould be utilized for a biological control program for this weed. These insects were observed tofeed also on other plants and economic crops on Pohnpei.
Melon aphid, Aphis gossypii, infested the shoots and tender leaves causing curling andcupping of affected leaves. Negligible damage also occurred due to feeding by the Philippinekatydid, Phaneropterafurcifera Stal. An unidentified mealybug also infested the tender shootsof Siam weed. . -
Population And Growth of ChromolaenaPrior to releasing the larvae of P. pseudoinsulata, the release site at Kitti was sampled for
Chromolaena density in a 5 x 5 m quadrat, The total number of weeds was 116 and most of themwere in the actively growing stage. On the release site, the weed formed tangled bushes 2-3 mhigh (x= 2.35) with stem sizes ranging from 4 to 6 cm (x= 5.45). The site was chosen primarilybecause the surrounding areas were also infested with the Siam weed and therefore assured thebiocontrol agent ample food to support several generations when established.
Flowering of Chromolaena usually occurs in December through February each year onPohnpei. Since the seeds of Chromolaena are wind-borne and can easily be carried by wind tonew areas, it is expected that in 2 to 3 years, if left uncontrolled, it will be distributed throughoutthe island.
101

Rearing Problems in the Laboratory and Solutions
Rearing of larvae of P. pseudoinsukua staned from 60 larvae received from Guam.Development of larvae, pupae, adults and eggs were normal and cultures were successfullycompletedup to7 generations, andduringthattimeperiodicreleasesof the larvaeweremadeinthefield.Unfortunately, in the8thgeneration manymaturelarvaebegantodie from an unknowncause, and this almost eliminated the cultures despitecleaning of rearing cups and 25 gallonrearing containers everyother day.Only a few larvaesurvived the 8th generation. To preventfurther massmortality of larvaein25galloncontainers, theclothcoverswereremovedfrom8:00a.m. to4:30 p.m. to assureoxygenin containers for the larvae. The containerswerecoveredinthe evening to preventthe escapeof larvae.
It wasalso observed in the process of culturingthat a few eggs from the majorityof eggmasses laidon the leavesof Chromolaena andon thescreenof the rearing cages failedto hatchfor unknown reasons, despiteproviding cottonballs soaked in sugar solutionas food for theadults. Napompeth (1990) mentioned a similar problem in his laboratory cultures of P.pseudoinsulata and felt that traceamounts of sodium (Na) played ~ role in the hatchability ofeggs.Herecommended addingNaClto thedietsof bothadultsand larvae.Hence,inadditiontosugarsolution,cottonballssoakedin saltSolution werehungin rearingcages to supplementtheadult nutrition.
Antswereobservedto interfere withlaboratory culturesby preyingon eggsand larvaeofP. pseudoinsulata in rearing cups and 25 gallon containers. Pupae were collected, placed inrearing cupsandbroughttoanotherroomfor2days.Therearingroomwassprayedwithdiazinonat I kgai/lOO gallonsof watertokillants.Legsofchairsandtableswhere rearingwasbeingdonewereplaced incans with water to preventants from interfering with the culture.
Field Releases of P. pseudoinsuiata: Problems Encountered and Solutions
Larvae:TableIshowsthenumberof larvaethatwerereleased in thefield. However, fieldreleased
larvaecould notbe recovered from thereleasesiteone weekafter. It wasobservedthat spiders,redantsandblackants, birdsandgroundlizardswereattractedreadilytothereleasesites.Spiderswereseen preyingon the larvae.
Since larvae could not get established despite many releases made, possibly due topredation, releases of larvae were made inside a 1.17 x 1.17 x 1.22 m field cage withChromolaena growing naturally insidethecage.Again,thelarvaereleasedinsidethecagecouldnot be recovered after the release.
Adults:Field releasesof adult mothsof P. pseudoinsulata were madein July and August 1989.
Morethan300adultswereliberated at thereleasesite.Againone weekafter, no eggsor younglarvae could be recovered from Chromolaena. In March and April 1990 adults were againreleased, butafter a week neitheregg masses nor young larvaewereevident at the releasesite.In May 1990it wasdecidedto releaseadults in fieldcages with Chromolaena inside. A week
102

before releasing the adulls, the cages were sprayed with diazinon at 1 kg altHa, including theChromolaena inside the cagesand thesurroundingareas about 1.2maround the cages, to get ridof the existingpredatorsand topreventthembeing attracted to thecages withadult mothsinside.Again, lizards and spiders were able to enter the cages and killed those adults inside.
The arctiid moth,P.pseudoinsulata, so far is theonlybiocontrolagent that hasbeenrearedand releasedon Pohnpei for the controlof Siam weed.About 15 monthsof field release of larvaeand adults since February 1989 did not result in establishment. However, in one release sitefeeding injury on leaves of the Siam weed and few caterpillars were noticed. This fieldpopulationis now being monitoredto observe itsbuildup in the release site as well as its naturalspread.
In some parts of the world,like Indiaand West Africa, thearctiid moths failed to establishbecause these areas havea very pronounceddry season. But in Sri Lanka where the dry seasonis less severe, as in Trinidad, the insect sucessfullyestablished CAB International Institute forBiological Control (1988) suggestedthat collectionsofP.pseudoinsulata should be made fromclimatic conditions similar to the proposed released zone: in our case, similar to the wetterconditions of Pohnpei should be the most promising approach.
Seibert (1985), based on his work in Guam, mentioned that in an effort to maximize thenumber of P. pseudoinsulata individuals in one location, adult moths were released. He foundthat approximately 500 adults if released in the same location would increase the chances ofestablishment This is desirable because the high resulting density of larvae are believednecessary to overcome predatorypressure and to create an epidemic population.
Attemptsare also beingmadetoproducemoreadults in thelaboratory.Fivehundredadultswill be released each time and releases will be made two or three times in the same release siteto findout whetherP.pseudoinsulata will becomeestablishedin otherareas of Pohnpei infestedwith Chromolaena. It is also planned that the larvae from the established site be transferred tonew sites to assist in spreadingand to cover the island within a short period.
Table 1. Number of laboratory-reared P. pseudoinsu1DJa released on Slam weed Infested area.-
Date2/28/894/27/89
5/3/896/15/897/14/898/23/899/25/89
10/15/9011/21/89
1/18/903/8/90
4/12/905/2/90
LocationWenik Elem. Sch. (Kitti)Wenik Elem. Sch. (Kitti)Wenik Elem. Sch. (Kitti)Wenik Elem. Sch. (Kitti)Wenik Elem. Sch. (Kitti)Nan Mal (palikir)Nan Mal (palikir)Nan Mal (palikir)Nan Mal (palikir)Nan Mal (palikir)Palikir (Sokehs)Palikir (Sokehs)Palikir (Sokehs)
Stagelarvaelarvaelarvaelarvaeadultsadultslarvaelarvaelarvaelarvaeadults
adults(Field Cage)adults(Field Cage)
number released219
1010210502128202350751462
1503216300225
103

REFERENCES
CAB International Institute for Biocontrol Control. 1988. Prospects for the biocontrol of Chromolaena, odoraia (L.) RM. King and H. Robinson. In: R. Muniappan (00.) First Intern.Workshop on
Biol. Control Chromolaena odorata. pp.25-33. Agr. Expt. Sta., Univ. of Guam.
Muniappan, R., V.T. Sundaramurthy & CA Viraktamath. 1989. Distribution ofChromolaena odorasa. (Asteraceae) and bionomics, consumption and utilization of food by Pareuchaetes
pseudoinsulata(Lepidoptera: Arctiidae) in India. Proc. VII 1nL Sym. Biol. Contr. Weeds, 6-11March1988, Rome,Italy. Delfosse, E.S. (ed.). Ist.Sper.PathoI.Veg. (MAF), pp. 401-409
Napompeth, B. 1990. Infertility of Pareuchaetespseudoinsulasa. In: Contributions from Thailand.Chromolaena odorata Newsleuer. No. 3.p.6.
. . .Seibert, T.F. 1985. Biological Control of Chromolaena odoraia. Guam Agric, Expt. Station. Annual
Report. 12-13.
104

HIGHLIGHTS OF WORK ON CHROMOLAENA ODORATA INGHANA
J.A. TIMBILLAand H. BRAIMAHBiocontrol Division,
Crops ResearchInstitute,P.O. Box 3785,Kumasi,Ghana.
ABSTRACT
The introduction and spread of the Siam weed, Chromolaena odorata, in Ghana isreviewed,anditsweedstatus,adverseeffectsandpotential benefitsaredescribed. Recentefforts
. at ascertainingthedistribution of C.odoratainGhanaarehighlighted,andstudieson thebiologyof Pareuchaetespseudoinsulata in the laboratory are reported.
Out of 44 plant speciesof 25 families testedfor hostspecificityof P. pseudoinsulata,nonewasfoundtobeeaten. FaunaassociatedwithC.odoratawerefound to includeColeoptera,Orthoptera, Dictyoptera, Hymenoptera and Acarina. No bacterial or fungal pathogens werecollected. The possibilities of usingbiologicalagents to suppressC.odoratawhilemaintainingits benefitsare discussed.
INTRODUCTION-
Chromolaena odorata (L.) Kingand Robinson(Eupatorium odoratumL.) belongs tothe plant family Asteraceae. C. odorata occurs naturally over a wide'area in tropical- andsubtropical Americas, from southern Florida to the northern border of Argentina (CruttwellMcFadyen 1988). It was accidentally introduced into most of the other regions where it iscurrentlyfound (Biswas 1934,Simmonds1965). By the late 1960s,C. odorata had become aweedin Nigeria(sinceits introduction in the 1940s),andhassincespread toGhana, IvoryCoastandCameroon(CruttwellMcFadyen 1988). InGhana,theweedwasfrrstdiscoveredinFebruary1969infestingabandonedexperimental plotsintheLegonBotanicalGardens,andby 1972ithadbeen observedin theGreater Accra, Centraland WesternRegions (Hallet al. 1972). There aretwoschoolsofthoughtabout the introduction of C.odorata to Ghana. One holds the view thatit wasintroduced by Ghanaianresidentsin Nigeriafor its medicinal properties;theother claimsthe weed was intentionally introducedduring the construction of the AkosomboDam between1962and 1966 to suppressother weeds along the grid. It was probably a coincidence that theLegon and Central/western Region outbreaksoccurredalmost simultaneously, as C. odorataappears to beabsentfrom the intervening country(Hallet al. 1972),and it is morelikely that theWesternoutbreakmaybe the resultof spreadfrom the IvoryCoast AlthoughC. odorata is notrecorded from Cote d'Ivoire by Hutchinson & Dalziel (1963), there is in fact in the GhanaHerbarium a specimencollectednear Abidjan in 1962.
105

Locally, the weed is named after two former heads of state, who ruled around the timeof its introduction and spread, as "Busia" in the Western region and "Acheampong" elsewhere.This is probably further testimony to the different origins of the Legon and western regioncolonies. This paper reports on surveys made in Ghana on the distribution ofC. odorata and thefauna associated with it Biological studies made on Pareuchaetespseudoinsuiaia, a defoliatorof C. odorata ,since its arrival from Guam in November, 1989, are also reported.
DISTRmUTION AND WEED STATUS OF CHROMOLAENAODORATA IN GHANA
The geographical distribution ofC. odoratais known to be limited to about 30° NandS latitudes and about l000m in altitude near the equator (Muniappan & Marutani 1988). Further,its distribution is limited to areas with a rainfall of 200cm and above and where temperatureranges from 20° to 37° C.
In Ghana, C. odoratais a weed in both arable and plantation crops. Particularly affectedare cocoa, oilpalm, coffee, rubber, maize and cassava fields. Other areas include abandonedfields, roadsides, riverbanks, cleared lands and forest margins. It is usually controlled byuprooting, slashing and burning, brushing or by use of herbicides such as Gramoxone, Atrazineand Glyphosate. C. odorata is also known to suppress other weeds, and serves as an alternatehost of nymphs of the variegated grasshopper, Zonocerus variegatus, and a breeding ground forthe adults. C. odoratais suspected to be the source of the characteristic foul smell ofZ. variegaiuswhich is important in deterring predators. Its potential as a rue hazard has been reported in Asiaand the Pacific (Muniappan & Marutani, 1988), Thailand (Napompeth et al. 1988) and in India(Ambika & Jayachandra 1990). Recent extensive bush fires in Ghana are also probablyattributable to the spread of C. odorata.
On the other hand, many people claim that C. odorata has medicinal properties andcould be'used in treating wounds and abdominal disorders (pers. obs.). It is alsoclaimed to enrichsoils by recycling minerals and by litter fall. In spite of this apparent "conflict of interest",farmers welcome any effective control program because of the cost involved in its control byeither chemical or mechanical means. Secondly, the potential environmental hazards of theusage of weedicides, their unavailability at the right time and harmto operators makes their useeven if affordable unwelcome. Thus alternative less costly and safer methods are required.
Though biological control ofC. odorata has been attempted in Ghana without success,lack of sustained effort (Greathead 1989) may have been the cause. Success in control of C.odoratahas been achieved in Asia and the Pacific (Greathead 1989) and chances are that insectcontrol agents when used in a sustained effort could bring about suppression of C. odorata.Biological control of C. odorata by introduced agents will not only reduce its density to a levelwhere its benefits can be enjoyed without it being a pest, but will also reduce the possibility ofenvironmental pollution that could follow the extensive use of inorganic weedicides.
An informal survey conducted in March and April, 1990 (Haruna & Timbilla,unpublished data) revealed that C. odorata has spread over the whole of the southern part of
106

Ghanaandas far northas latitude8' 15'N. (Fig. 1).Its spread furthernorthappears tobe limitedby the amountof rainfall.
BIOLOGY OFPAREUCHAETES PSEUDOINSULATA IN GHANA
The biology of P. pseudoinsulata was studied in the laboratory to confirm reportsalready available from the Philippines (Aterrado& Talatala-Sanico 1988)and elsewhere.
MATERIALS AND METHODS
Larvae were reared in Kilner jars for the first two instars and later transferred intowooden cagescoveredwithgreybaft(30x 30x 3Ocm). FourthtoSixthinstarlarvaewerefinallyreared in45x45 x 75cmcages. In all cases,the larvaewerefedon freshC. odorata leaves. Theadultswerefedon a sucrosesolutionand allowed to mate in 30 x 30 x 30cm grey baft coveredcages.
RESULTS
Eggswerelaidin batcheson leavesand on thewallsof thecages. Egg batchsize variedfrom 2-68(Table1). Incubation periodlastedabout4.9 days. Larvalsizerangedbetween2mm22mm. The larval stage lasted 21.4 days while pupation took 9.3 days. Adults lived for anaverageof 5 days. Thus the total life cycle of P. pseudoinsulata from egg to adult emergencelasted for 35.6days.
Egg hatchability decreasedwith generation until zero hatchability was obtained afterthe4thgeneration.
HOST RANGE AND SPECIFICITY TESTS
Second instar larvae of P. pseudoinsulata were tested for their host range andspecificity. Larvaeweresuppliedwith freshsamples of the test plantsuntil theydied. So far 44plantspeciesfrom 25differentfamilieshavebeentested(Table2). Noneof theplantstestedwasfound to be eaten by P. pseudoinsulata. Larvae tested died from starvationwithin 5 days.
FAUNA ASSOCIATED WITHCHROMOLAENA ODORATAIN GHANA
A limited survey was carried out duringJuly, 1989and April, 1990to determine thefaunaassociated withC.odorata andpotentialcontrolagents. Specimenscollectedhavenotyetbeenfullyidentified; however, amongthemarearthropods belongingtotheordersHymenoptera, .Diptera, Orthoptera, Coleoptera, Dictyoptera, Homoptera and Acarina. The hymenopteraninsectswerespecifically found tobestemborerswhile theothersweregenerallyfoundon leaves .and/or flowers. No collections of fungaland bacterialagents were made.
107

DISCUSSION
C. odortuahas been a seriouspest since its introduction in Ghana both in arable andplantation crops. Its controlhas beenspecificto local needs.
There is an urgentneed to control the weed. However, physicalcontrol is expensivewhile chemicalcontrol is both expensive and potentially hazardous. The need for biologicalcontrolas a better controlalternative is very muchevident.
The studieson P.pseudoinsulata indicatethat thoughit couldbeefficientand specificas a defoliator, its ability to controlC. odoratais presently limitedby the inability to rear it inthe laboratory beyond the third generation. Although the moths appeared healthy they wereinactive, and adult size decreased from generation to generation. Given the profusegrowthofC. odortua it may be that it will require more than just a defoliator to achieve effectivesuppression.
Thefaunaassociated withC.odoratasuggeststhatotherassociatedorganismsmayalsobequitevaried,andothernatural enemies including insects,fungiandbacterialpathogens shouldbe investigated.
It isprobablethatextensive surveysin thecenteroforiginofC.odoratawilllead to thediscovery of othernaturalenemiesin thecategories of stem,rootand flowerborers/feeders thatcouldbeusedtosupplementP.pseudoinsulata. It isalsopossiblethatpathogenicfungiandothermicro-organisms couldbe found.
Itisevidentfromtheabovethatthebiological controlofC.odoratainGhana,followingits extensivespreadand establishment, will requirea specialeffort.
108

Table 1. Sizesor Egg Batches laid by 25 Females or Pareuchaetes pseudoinsulata
Sample No.
123456789
10111213
No. or EggslBatch
6323342
131035505342101013
Sample No.
141516171819202122232425
No. or EggslBatch
16112913189
1472
116819
109

Table 2. Plants Tested for Host Range and Specificity of ParellcluJeles pselldoinslllata
SER.NO. TEST PLANT BOTANIC NAME FAMILY
l. Mango Mangiferaindica Anacardiaceae2. Papaya Caricapapaya Caricaceae3. Lettuce Lactucasativa Asteraceae4. Sweet Potato Ipomoeabauuas Convolvulaceae5. WateImelon Citrullus lanoJus Cucurbitaeeae6. Cassava Manihol esculema Euphorbiaceae7. Avocado Perseaamericana Lauraceae8. Groundnut Arachishypogaea Leguminosae9. Pigeon pea Cajanuscajan10. Soybean Glycine max1l. Cocoa Theobromacacao Sterculiaceae12. Cola Cola mitida13. Jute Corchorusspp. Tiliaceae14. Onion Alliumcepa Amaryllidaceae15. Cabbage Brassicaoleracea Cruciferae16. Cocoyam Colocasiaesculenta Araceae17. Pineapple Ananas comosus Bromeliaceae18. Cowpea Vigna unguiculata Leguminosae19. Cotton Gossypiumspp. Malvaceae20. -Kenaf Hibiscuscannabinus2l. Okra/Okro H. esculentus22. Roselle H. sabdariffa23. Aramina Ilrena lobara24. Guava Psidiumguajava Myrtaceae25. Yams Dioscoreaalara Dioscoreaceae26. Sugar cane Saccharumofficinale Gramineae27. Rice Oryza Saliva28. Maize lea mays29. Common millet Panicummuliaceum30. Banana Musa sapiemum. Musaceae3l. Plantain Musa paradisiaca32. Coconut Cocos nucifera Palmae33. Oil palm Elaeisguineensis34. Arabica coffee Coffea arabica Rubiaceae35. Lemon Citrus limon Rutaceae36. Grape fruit Citrusparadisi37. Tangerine Citrus reuculasa38. Sweet orange Citrus sinensis39. Lime Citrus auranufolia40. Hot pepper Capsicumannuum Solanaceae4l. Tomato Lycopersicon esculenuan42. Tobacco Nicotiana tabacum43. Egg plant Solanummelongena44. Ginger Zingiber officinalis Zingiberaceae
110

. ;'.",._.-._ ..,.-.- ._. . GHANAB~on.o· --'( r. .
. r: "(..... "l
I()O ,I j 10'0
Co"
, 15
FP
* AREA INFESTED BY C.OOORATA
Figure 1. Distribution or Chromotaena odorata in Ghana
111

REFERENCES
Ambika,S.R. & Jayachandra. 1990.The problem of Chromolaena weed. Chromolaenaodorata Newsletter3:1-6.
Aterrado, E.D. & RL. Talatala-Sanico. 1988.Status of Chromolaena odorata. Research inthe Philippines. In: R Muniappan(00.). Proc. First InternationalWorkshop on Biol,Control of Chromolaena odorata. pp. 53-55. Agri. Expt, Sta., Univ. Guam.
Biswas, K. 1934. Some Foreign Weeds and their distribution in India and Burmah. IndianForester 60:861-865.
Cruttwell McFadyen,RE. 1988.Historyand Distribution of Chromolaena odorata. In: RMuniappan (ed.). First International workshopon Biol. Control of C. odorata.. pp7.-12. Agr.Expt. Sta., Univ. Guam.
Greathead, DJ. 1989.PresentPossibilities for BiologicalControl of InsectPests and Weeds. in Tropical Africa. Biological Control. A SustainableSolution to Crop Pest
Problems in Africa. 190 pp.
Hall, J.B., R Kumar & A.A. Enti. 1972. The obnoxiousweed Eupatoriumodoratum(Compositae)in Ghana. Ghana J. Agric.Sci. 5:75:78.
Hutehinson& J.M. Dalziel. 1963.Flora of west tropicalAfrica 2, 2nd 00. (F.N. Hepper, 00).pp. 544.
Muniappan,R & M. Marutani. 1988.Ecologyand Distributionof C. odorata in Asia and thePacific, In: R. Muniappan (ed.) Proc. First Intern.Workshop on Biol, Control ofChromolaena odorata. pp. 21-24. Agri. Expt. Sta.•Univ. ofGuam.
Napompeth,B., N. Thi Hai, & A. Winotai. 1988. Attemptson BiologicalControl of SiamWeed, Chromolaena odoratain Thailand Proceedingsof the First International.Workshopon BiologicalControl of Chromolaena odorata. pp.57-62. Agri. Expt,Sta.,Univ. Guam.
Simmonds,F. J. 1965.Report on a visit-to Sierra Leone, Ghana, Nigeria. Mauritius and EastAfrica. November 1964- March 1965.Commonw. Agric. Bureaux. 42 pp.
112

STUDIES ON CAUSES OF INFERTILITY OF EGGS OFPA/lEUCHAETES PSEUDOINSULATA REGO BARROS
0.0. TORRES.O.G. VARGAS. S.M. RI11JALand E.M. ALFORJA "UniversityResearcherand ResearchAssociates,
NationalCrop Protection Center,UPLB.College,Laguna,Philippines
The insect Pareuchaetes pseudoinsulata Rego Barros, is an important and promisingbioticagent against theSiamweed Chromolaena odorata (L.) R.M. Kingand H. Robinson. P.pseudoinsulata larvae feed voraciously on leaves and young shoots of the Siam weed. Thespecies is specific to Siam weedand a few other relatedweed species.
Oneof the majorproblemsencounteredin the utilization of thisbioticagent is laboratoryrearing. There wereinstanceswhenmost,andsometimesall. of theeggmasseslaidby the mothwere infertile. resulting in complete loss of a stock. This paper reports the results of someobservations carried out at College.Laguna,Philippines regarding the probablecauses of thisproblem.
MATERIALS AND METHODS
P.pseudoinsulata usedin thisstudywasobtainedfromPuertoPrincesa,Palawan. Larvaewerecollectedin August,1989and rearedunderlaboratory conditions. Rearingwasdone usingscreen cages and screen covered plastic trays and with fresh Siam weed leaves as diet Theobservations made wereon the insect matingbehavior,the effect of light and inbreeding.
Observation on Mating BebaviorThe matingbehaviorof theinsectwasobservedin0.5 mx0.5m x0.5 m screencages. The
top and one side of each cage were madeof glass whereasthe bottomwas.madeof galvanizedironsheet. Pupae weresexedby meansof the location of thegenitalopening. In the femalethegenitalopening is near the anteriormarginof the eighthabdominalsegmentwhile In the maleit is locatedon the ninthabdominal segment. Fivemaleandfivefemalepupaereadyforeclosionwere placed inside the cage at about 5:00 p.m. Observationswere made continuously for 24hours. takingnoteof the behaviorof the maleand femalemoths. Five such observationsweremadein differentdays in an undisturbed room. The lightingconditionof the room is describedin the results and discussion part of the paper.
Effect or LigbtAn experiment was conducted to detennineif the amount of lig~t could affect the
production of infertileeggs. This inquiry was based on the observation that adults remainedinactiveboth when fully lightedand under completedarkness.
113

Cages were covered with carbon paper to create conditions of varying light intensitiesinside. Onecage wascompletely coveredwhileanotherwascompletelyexposed to light Twootherswerepartlycovered.one thirdand two thirdsfrom the top. Ten pairsof maleand femalepupae thatwere due for eclosionwere placed insideeach cage. Thecages were left.inside thelaboratory. lighted with fluorescent lamps from the ceiling. day and night for one week. Thecages were laid out on one-meterhigh woodenstands in a randomizedcompleteblock designwith fourreplications. Thedistancebetweenthe top of thecages and the light sources was twometers.
Effects ofInbreedingFourpairs of adults were selectedfrom those thatdevelopedfrom field-collected larvae.
Eachpair wasrearedseparately and theirrespectiveeggs.representing the first generation(Fl),were labeled as A. B. C andD. Theeggs wererearedtoadultsandvariousmatingcombinationswere madeas follows:
AxA
AxB
AxC
AxD
BxB
BxC
BxD
CxC
CxD
DxD
Theresultingeggsfromthesecombinations (F2) wereagainrearedandall possiblematingcombinations suchas AAx AA. AA x AB. AA x AC•...•and DD x DD weremade. Theprocesswas repeated until the third generation when the culture had to be discontinueddue to virusdisease infection. The originalplan had been to continue up to to generations using selectedmatingcombinations.
Ineachgeneration andforeachmatingcombination. thenumberof fertileandinfertileeggmasseswas obtained and the numberof male and femalepupae producedwas recorded.
RESULTS AND DISCUSSION
Observations on MatingEclosionof adults tookplace in generalbetween5:00 and 6:30 pm. This period was not
dark at the time of this study.
Immediately aftereclosion,adultswalkedon the baseof thecage towardthe sides. Mostof themclimbedon.the sidesof the cage and thensettled in one place whilea few stayedat thebase. In somecases. the adults flew to the sidesof thecage in the directionof the light source.Very seldomdid the adults find the honey-impregnated cotton. providedas a source of food.hangingon the sides of the cage. In fact. about 50%of them did not feed on the honey within24 hours.114

Initial observationshowed that the activityof the adults is affected by light. When theobservationcage wasexposed10 light the mothsremainedmotionlessduring the day and nightand there was no mating within the 24 hour period. The eggs produced by the females wereinfertile. Succeedingobservations thereforeweremade with the room lightsoff. To make theinsectsvisible,a small flash light wasdirected to the wall of the room to provide indirect lightto the observation cage. In later observations, similar lighting conditions were obtained byleaving thedoor about 30 cm open and letting light from a sourceoutside enter indirectly.
When maintainedunder these lightingconditions,the adults also remainedstill but onlyuntil 1:45am 10 2:58 am. Withinthis range'of time,matingactivitystartedamongthe majorityof adults, althoughsome wereobservedto begin matingas late as 5:00 am. In general, a malemoth would start running around inside the cage, creating noise as if its wings were flappingrapidlyagainstthe wallof thecage. Othermaleswouldfollow, but these would walkfastwithvibratingwingsbut withoutmuchnoise. They wouldgenerallymovefromthe floor then up onthe sides of the cage. Afterreachingthe top, they woulddrop themselvesto the floor and thenmove up again, as if searchingfor a female.
Femalesnormallyremainstill,but if a malecomeswithin3 104 cm theywillpartiallyraisetheir wings.
After severalupand down movements withinthecage, the male willcome near a femaleand willmakeone, twoor severaljourneysaround her withinabouta 2 106 cm radius. He maythen move away and find anotherfemale to chase in the same way, or may approach the samefemaleand instantlycopulate. Thefemaleraisesherwingswhenthe maleapproaches,the malepositionshimselfunderherwings,and thensuddenlyturnsback withhisorganalreadyinserted.This happensin about two seconds.
Unlessdisturbed,mostpairsof adultswereobserved10remainincopulationforaboutoneanda half10three anda halfhours. A fewwouldexceedthis,andseveralcaseswerealsoobservedwhere the matingpair did not separate from each othereven up 10 death.
Effect of InbreedingThere was a wide range of percentageinfertilityof F2 egg masses among the different
mating combinations made. The lowest was 18.82% and the highest was 100%. There was,however, no statistically significantdifference obtained between crosses involving the sameparent egg massesand those involving differentparentegg masses.
Similarly, the percentage of infertileegg masses produced by F3 adults ranged widely,from25%to 100%. Whengroupedaccording10 thedegreeof inbreeding,thereweresignificantdifferences in mean percentage infertility among the different groups (Table 1).· Crossesinvolving two F2 inbreds (e.g., AA x AA or BB x CC) gave significantly lower incidence ofinfertileeggmasses(58.65% and58.05%)followed bythoseinvolving onlyoneF2 inbred. Morediversecrossesgavecorrespondingly higherpercentinfertility, suchas thoseinvolvingan inbredand itshybrid(82.44%),an inbredandan unrelatedhybrid(65.46%)thesamehybrid(orselfinga hybrid,78.05%),and completelyunrelatedhybrids(100%).
115

Table 1. Percentage of infertile egg masses produced by FJ adults
Mating No. of Percent (%) Mean*ofCombination - Egg Masses Infertile Mating Type
AAxAA 20 45.00BBxBB 28 64.28CCxCC 21 66.67 58.65 d
AAxAC 41 97.56AAxAC 25 28.00BBxBC 33 51.51BBxBD 21 100.00ABxBB 22 100.00ACxCC 9 100.00BCxCC 32 100.00 82.44 be
AAxBB 13 69.23BBxCC 32 46.87 58.05 d
AAxBC 23 73.91'AAxBD 20 25.00ABxCC 24 45.83ACxBB 10- 100.00ADxBB 25 88.00BDxCC 25 60.00 65.46 cd .
ABxAB 26 100.00.ACxAC 19 47.00BCxBC 23 95.65BDxBD 23 69.56 78.05 be
ABxAC 26 84.00ABxBC 24 54.00ABxBD 23 100.00ACxBC 44 97.73BCxBD 29 96.55 86.46 b
ACxBD 41 100.00 100.00 a
• Any two means followed by a common letter are not significantly different at 5% level(DMRn·
116

Theseresultsaresurprising because mostspeciesofplantsandanimalsbehaveotherwise.In addition, thiscannotexplain thedevelopment of infenility that isobservedin the laboratory.Continuous culture of the insect from a single stock in the laboratory is inbreeding, and yetinfertilitydevelops. The authorshaveat presentnot foundan explanation for this.
The proportion of maleand female pupaeproducedin the F2 and F3 generations(Tables2 and 3) werealso greatlyaffectedby the mating combinations used. A Chi-square test of thedata revealeda significantly lowerproportion of female pupaefromF2 inbreds(e.g.A x A) thanfromcrosses(e.g, Ax B). Thesamewastruein theF3generation. Therewasadistinctdecreasein theproportion of femalepupaeas theparentsinvolvedin thecrossesbecamemorerelated. Itis worthemphasizing thatoneof theF2 inbreds(DD)hadbeenlostdue to infertility of eggs. Inthe F3,on theother hand,one inbred(AAx AA)producedonly 29.63% femalesand two otherinbreds(BB x BB and CC x CC) produced all maleoffsprings.
Table 2. Number of male and female pupae produced by F2 adults
Mating Number of Pupae Proportion ofCombination Male Female Females"
AxA 33 35 0.5147
BxB 119 133 0.5278
CxC 238 233 0.4947
AxB 66 75 0.5319
AxC 85 112 0.5685
AxC 4 12 0.7500
BxC 117 110 0.4846
BxO 82 79 0.4907
*Proportions not significantly different.
The observedinfertility of eggs of the Siamweed mothappears to be a result of severalinterrelated factors. Lackof mating couldbea majorfactor resultingfrom theinfluenceofotherfactors likelightandinbreeding. Theeffectoflight maynotbea veryinfluentialfactorinactualrearing practice.since rearing laboratories are normally not lightedat night.
117

Table3. Number of male and female pupae produced by F3 adults
Mating No. of Pupae Proportion of FemalesCombination Male Female Per Cross Per Group
AAxAA 19 8 0.2963BBxBB 48 0 0CCxCC 5 0 0 0.1000 a
AAxAB 2 1 0.3333AAxAC 98 67 0.4061BBxBC 17 6 0.2609 0.3874 b
AAxBB 5 4 0.4444BBxCC 14 8 0.3636 0.3871 b
AAxBC 1 3 0.7500AAxBD 5 3 0.3750ABxCC 57 68 0.5440ADxBB 14 15 0.5172BDxCC 48 37 0.4343 0.5020d
ACxAC 53 39 0.4239.ACxBC 4 3 0.4286BDxBD 32 2S 0.4386 0.4295 c
ABxAC 21 30 0.5882ABxBC 36 38 0.5135ACxBC 2 2 0.5000BCxBD 8 4 0.3333 0.5248 d
Any two means having a common letter are not significantly different at 5% levels (Chi square)
118

Imbalance in theproportion of malesand females resultingfrominbreeding is perhapsthemostimportantfactorthatleadstotheabsenceofmating. Asmall proportion ofeithersexamongthepupaeproducedwillnotguaranteematingbecauseeclosionfromthis smallpopulationmaynot necessarily coincidewith that of the oppositesex. It wouldsurely help if pupaecould besubjectedtosex determination. Thoseproducedaheadof theoppositesexcould bestoredunderrefrigeration to delay eclosion.
SUMMARY
Matingin the mothP. pseudoinsulata mothwasobservedto takeplace from 1:45a.m. to5:00a.m. Adultsremained incopulation foraboutoneanda halftothree anda halfhours. Underlightedconditions, the moths remained still inside the observation cage for the wholeday andnight and did not mate, resulting in the production of infertileeggs. Results of experimentsinvolving variation in theamountsof lightby coveringthecages withcarbon paperat differentdegrees,however, failed to showany effect of such treatments on egg fertility.
Experiments on inbreeding showed that F3 inbreds hada smallerpercentageof infertileeggs compared to more diverse crosses even when reared with equal number of males andfemales. However, F3 inbredsgaveverylowproportions of females comparedto othercrosses;this femaleproportion increased to almostequalnumberof malesand females as the membersof the crosses madebecomemorediverse.
Infertility of eggs of the Siam weed moth appearedto be largely due to lack of matingresulting froman unbalanced proportion of malesand females. Amajorcauseof this imbalanceis theeffectof inbreeding. Theproblem couldbeminimized, therefore, byrenewingstocksaftera numberof generations, which is a recommended practicein laboratory rearing.
119


THE CHEMISTRY OF THE PUTATIVE FEMALE SEX-ATTRACTANTPHEROMONE OF PAREUCHAETES PSEUDOINSUlATA REGO
BARROS (LEP.: ARCTIIDAE)
STEFAN SCHUlZInstitutfur Organische Chemie,UniversiUlt Hamburg,
Martin-Luther-King-Platz 6, D-2000Hamburg 13,FederalRepublicof Germany
INTRODUCTION
ThearctiidmothPareuchaetes pseudoinsulata is a potentialbiological agentfor controllingthetropicalpestweedChromolaena odorata King& Robinson(seeSeibert1989andseveralpapers in this Proceedings). However. so far there are no meansavailable for monitoring thepresenceof thesemothsand theirpopulation dynamics, which shouldbe a routine in the courseof releasingcontrolagents(et Boppre1991).- As a basis for developing pheromone traps, thispaperreportson thechemistryoftheputativefemale sex-attraetantpheromoneofP.pseudoinsukua.
MATE~SANDMffiTHODS
PupaeofPareuchaetespseudoinsulataRegoBarroswerecollectedinthefieldinSriLankaand shippedto Germany. The pheromone glandsfromfreshly emergedfemales wereexcised.immersed into50~ pentane(Uvasol, Merck),and storedat-78° C. Massspectra(70 eV) wereobtainedwitha VarianMAT 311Amassspectrometercoupledto a Carlo-ErbaFractovap2101gas chromatograph. For analytical gas chromatographic analysesa Carlo-Erbagas chromatograph of the same type equipped with a flame ionization detector and split injection wasemployed. Separations were performed using a 50 m SE-54 glass column (inner diameter0.32 mm, film thickness 0.25 1JIll) and a 50 m WG 11 glasscolumn(inner diameter0.32 mm,film thickness 0.25 1JIll); heliumservedas carriergas. For identification of compounds,massspectraand retentiontimeswerecomparedwith those of authenticreferencesamples.
Cl
RESULTS
Gaschromatograrns ofextractsof thefemale pheromone glands(et Fig. 1)showone maincomponent accompanied by severalminorcomponents and sometracecompounds. The maincomponent exhibiteda massspectrum showing characteristic signalsat"m/z=79, m1z=108, rn/z=234and a molecular ion at rn/z=290. This massspectrumis identicalwith the spectrumof an
121

authentic sampleof (Z,Z,Z}-3,6,9-heneicosatriene; the retention times on different stationaryphases proved to be identical, too. Thus, (Z,Z,Z}-3,6,9-heneicosatriene is the main volatilecomponent of thepheromone gland.
In a similarway (Z,Z)-6,9-heneicosadiene and (Z,Z)-6,9-nonadecadiene as well as (Z)-9nonadecenewereidentifiedasminorcomponents. Smallamountsofheptadeceneandheneicosenewere also present. In addition, the corresponding saturated hydrocarbons heptadecane andnonadecane could be identified. In contrastto thesecompounds two tracecompoundsshow abranched carbon chain; they proved to be 2-methylheptadecane and 5-methylheptadecane.Figure 1 shows the relativeamountto thecompounds identified.
Blectroantennogram (EAG) recordingsdemonstrated thatmaleantennaerespondstronglyto excisedfemale glandsas well as to (Z,Z,Z}-3,6-9-heneicosatriene (for details seeSchneideret al. 1991).
DISCUSSION
(Z,Z,Z}-3,6,9-Heneicosatriene,the maincomponent of theputativefemalepheromoneofPareuehaetespseudoinsulata, is quite widespread as a constituentof sex-attractants in arctiidmoths. It hasbeen found in Halisidota leda (Descoins et al. 1989), Tyriajacobaeae (Frerot etal. 1988), Aretia villica (Einhomet al. 1984), Phragmatobta fuliginosa (Descoins & Frerot1984), and Periealliaricini (S. Schulz& D. Schneider unpubl.) and provedto bea componentof thefemale sexpheromone ofHyphantria cunea (T6thet al. 1989),CreatonotostransiensandC.gangis(Wunderer et al. 1986) and Utetheisa ornatrix(Conneret al. 1980). It also occurs inother lepidopteran families (et Am et al. 1988/1990). The minorcomponents whichlikelyaddto the specificity of thepheromonal signalare closelyrelatedto the maincomponentbut showdifferentdegreesof unsaturation along thechain,possiblyproduced from unbranched polyunsaturated fatty acids. Such acids have been identified in male pheromone glands of severalspeciesof milkweed butterflies (Danainae) (Schulz et al. 1988).
The branched hydrocarbon, 2-methylheptadecane, which appears to show a differentbiogenetic originfrom theabovementioned compounds, isalsoknowntoactasasexpheromonein some arctiid species: Holomelina lamae (Schal et al. 1987, Roelofs & Carde 1971), H.aurantiaca, H. immaculata, H. nigricans (Roelofs & Carde 1971), and Pyrrharctia isabella(Krasnoff& Roelofs1988). So far, however, Pareuehaetespseudoinsulatais the only speciesproducing both typesof compounds.
Gl
The chemical findings reported above providea first step towards developing of techniquesfor monitoring Pareuchaeteswhere it hasbeenand will be releasedas a leaf-defoliatorto control Chromolaena. However, prior to a routine application of synthetic pheromones,studies on the sexual behaviour of Pareuehaetes are required (et McNeil 1991) as well asbehavioural teststoelucidatetherole(s)of thedifferent compounds identified; verylikely,maleswill not respond to the majorcomponent alonebut to a specificblendof compounds.
122

ACKNOWLEDGEMENTS
I thankP. Kanagaratnam (Lunuwila, SriLanka) forproviding theinsectsand D.Schneider(Seewiesen), M. Boppre (Freiburg), and W. Francke (Hamburg) for helpful discussion andcriticalreview of the manuscript.
REFERENCES
-Am,H., M. T6th & E. Priesner. 1988. addendum 1990. List ofSex Pheromones ofLepidoptera and RelatedAttractants. Intern. Organs. BioI. Contr. Work Group: Pheromones. Swiss Fed. Res. StationWlldenswil
Boppre, M. 1991. A non-nutritional relationship of Zonocerus (Orthoptera) to Chromolaena (Asteraceae)and general implications for weed management In: R. Muniappan & P. Ferrer (eds.) Proc. 2ndIntern. Workshop on BioI. Control of Chromolaena odoraia: pp. 143-147. Agri. Expt. Station,Univ, Guam, Mangilao, Guam.
Conner, W. E. T. Eisner, R. K. Vander Meer, A. Guerrero, D. Ghiringelli & 1. Meinwald. 1980. Sexattractant of an arctiid moth (Utetheisa ornatrix): a pulsed chemical signal. Behav, Ecol.Sociobiol . 7: 55-63
Descoins, C. & B. FrerolI984. Taxonomic value ofthe chemical compositionofthe sex pheromone blendsin arctiid moths and related families. Abstr, of the XYll Intern.Congr. Entomol., Hamburg: 467
Descoins, C., B. Lalanne-Cassou, B. Frerot, C. Malosse & M. Renou. 1989. Comparative study ofpheromonal secretions produced by arctiid and ctenuchid moths (insects, Lepidoptera) from theneotropical area. C. R. Acad. Sci. Ser 3,309: 577-581
Einhom, J.•B. Boniface, M. Renou & M-L. Milat. 1984. Etude de la pherornone sexuelle de Arctia villicaL. (Lepidoptere, Arctiidae). C. R. Acad. Sci. Paris 298 ill: 573-576
Frerot, B.•M. Renou, C. Malosse & C. Descoins. 1988 lsolement et identification de composes aactivitepheromonale chez les femelles de I'arctiide Tyria jacobaeae: determination biologique de laconfiguration absolue du compose majoritaire. Entomol. Exp. Appl . 46: 281-289
Krasnoff, S. B. & W. L. Roelofs. 1988. Sex pheromonereleased as an aerosol by the moth Pyrrharctiaisabella. Nature 333: 263-265
McNeil. J. N. 1991. Behavioral ecology of pheromone-mediated communication in moths and its importance in the use of pheromone traps. Annu. Rev. Entomol. 36: 407 -430
Roelofs.W. L.& R. T. Carde. 1971. Hydrocarbon sex pheromone in tiger moths (Arctiidae). Science 171:684-686
Schal,C .• R. E. Charlton & R. T. Carde. 1987. The temporal patterns of sex pheromone titers and releaserates in Holomelina lamae (Lepidoptera: Arctiidae). J. Chem. Ecol. 13: 1115-1129
123

Schneider,D., S. Schulz, R. Kinmann & P. Kanagaratnam. 1991. Pheromones of both sexes of the weeddefoliator moth Pareuchaetes pseudoinsulaia (Lepidoptera: Arctiidae). J. Appl. Entomol: inpress.
Schulz,S.,W. Francke & M. Boppre. 1988. Carboxylic acids from hairpencils of male Amaurisbutterflies(Lep.: Danainae). BioI Chem Hoppe-Seyler 369: 633-638
Seibert, T. F. 1989. Biological control of the weed, Chromolaena odorata (Asteraceae), by Pareuchaetespseudoinsulata(Lepidoptera: Arctiidae) inGuam and the NorthemMariana islands. Entomophaga34: 531-539
T6th.M., H~ R. Buser, A. Pena, H. Am, K. Mori, T. Takeuchi, L. N. Nikolaeva & B. G. Kovalev. 1989.Identification of (3Z,6Z)-1.3.6,-9,1O-epoxyheneicosatriene and (3Z,6Z)-I,3,6-9,1O-epoxyeicosatriene in the sex pheromone of Hyphaniria cunea. Tetrahedron Lett 30: 3405-3408
Wunderer, H., K. Hansen, T. W. Bell, D. Schneider & J. Meinwald. 1986. Male and female pherornonesin the behaviour of two Asian moths, Creatonotos (Lepidoptera: Arctiidae). Exp Bio146: 11-27
124

11
7
82
6 5, '
[min]I
50I
40
Fig. 1 Gaschromatogramof an extract of femalepheromoneglands of Pareuchaetes pseudoinsulata Rego Barros. 1 heptadecene: 2 heptadecane; 3 5-methylheptadecane; 4 2methylheptadecane; 5 (Z,Z)-6,9-nonadecadiene; 6 (Z)-9-nonadecene; 7 nonadecane;8 hexadecanoic acid (ubiquitously occurring fatty acid); 9 (Z.Z)-6,9-heneicosadiene;10 heneicosene; 11 (Z.Z.Z)-3,6,9-heneicosatriene.
/
125


ARTHROPODS AND PATHOGENS FOR BIOLOGICAL CONTROLOFCHROMOLAENA ODORATA
P.A.C.OOP, A.N.G. HOLDEN 2 and P.S. BAKER 3
1. use, MALAYSIAN STATION2. use, SILWOOD PARK, ASCOT,U.K.
3. IIBC,CENTRAL ANDLATIN AMERICAN STATION
ABSTRACT
A study of the biological attributes of four arthropod candidates recommended forbiological control of Chromolaena odorata was made using Goeden's (1983) criteria. Theresults suggested that these agents would provide only partial control on their own. The.exception to this has been the biological control programme at Guam using Pareuchaetespseudoinsuuua. The suggestion for importation of several agents together merits seriousconsideration. Besidesarthropods, five fungalpathogensof C. odorataare being investigatedin Trinidad. Thesefungalpathogens mayprovetobe usefuleitherasclassicalbiological controlagentsor as mycoherbicides. Studiesthus far indicatethatCiononthrix praelonga and eptoriaekmaniana are promisingcandidates.
INTRODUCTION
Chromolaena odorata(L.) R. M.Kingand H.Robinson is a seriousweedin WestAfrica,Southand South-eastAsia(CruttweU McFadyen 1988a). It hasbeenthe subjectof a biologicalcontrolprogrammesince the 1970s. Two decadeslater,limitedsuccesshasbeen achievedandthereare morequestionsraisedthananswersobtainedaboutthestateofbiologicalcontrolof thisweed. Itsofteninnocuous positionintheneotropics (CruttweU McFadyen 1988b) hassuggestedthat this weedcould be similarly controlled in the Old World.
Hitherto, theonly country that has reported completesuccessin the biological controlofC.odorataisGuam(Muniappan & Marutani 1988,Seibert1989). Thiseventhasprovidedbothimpetusand frustration to proponents of theprogrammetocontrolC. odoratabiologically. Partof the impetusis the renewedinterestin otherbiological controlagentsof C.odorata in CentralAmericaand some resultsare reported. The biological attributesof four recommended agentsare analysedand discussed. In addition, an updateon the study of pathogensare given in thisreport
127

ARTHROPODS FOR BIOLOGICAL CONTROL OF C. ODORATA
Cruttwell (1974) reported that some 225 arthropods were recordedon C. odorata in itsnativerange. Of these,only four have been recommended for introduction into the Old Worldwhere the weed is a serious problem (Cock 1984). Recommendations were made followingextensivestudiesin the nativerangeof the weed. The agentsrecommended are:Pareuchaetespseudoinsulata Rego Barros(Lepidoptera: Arctiidae) (reportedby Bennett & CruttwellI973),Apionbrunneonigrum Bequin-Billecoq (Coleoptera: Apionidae) (reportedby CruttwellI973),Mescinia parvula (Zeller)(Lepidoptera: Pyralidae) (reportedby Cnittwelll977a) and kaJitusadoratus Keifer (Acari, Eriophyidae)(reportedby Cruttwell 1977b). P. pseudoinsulata wasintroduced into sevencountriesbut was reportedto be successfulonly in Guam (Seibert 1989)
.(Table I). No establishment was reported from Ghana and Nigeria. In Asia, the arctiidestablishedin Malaysia, Indiaand Sri Lankabut not in Thailand. In countries with successfulestablishment, P. pseudoinsulata did not impact significantly on the weed population. In thePhilippines, the insect fortuitously arrived in the southern islandsbut did not appear to causeseriousdamage to C. odorata (Aterrado & Talatala-Sanico 1988).
In thecase of Abrunneonigrum, the insect failed to establish in all the five countries towhich it was introduced (Table I). Longperiodswhen no flowersoccur could possiblybe thereason for the failure to establish (Syed 1979).
Considering the limited success achieved thus far, an examination of the biologicalattributesof theseagentswasattemptedusingGoeden's (1983)criteriaforselectionofarthropodagentsinbiologicalcontrolofweeds. Theagentwiththehighestscoreof 50 isP. pseudoinsulatawhichconfirmsthisinsectas theagentof firstchoiceinthebiologicalcontrolprogramme(Table2). The nextagent isM.parvulawitha scoreof40 whileA.brunneonigrum andA. adoratushada total scoreof only 29 each. Goeden(1983)suggestedthat species with scoresof between 20and 50 wouldindicatethat theagent isonlypartiallyeffective,reflectingthe needfor morethanone agent to provideadequatecontrol.
PATHOGENS OF C. ODORATA IN TRINIDAD
Five fungi attacking C. oderata were identified in Trinidadand Tobago (Table 3). Ofthese,only two were studiedextensively, viz. C. praelonga and S. ekmaniana.
Infectionof CaribbeanC.odorata fromonesite in Trinidadby C. praelonga, collectedatthat site, has beenachievedby inoculation at 15-22'Cand 100% RH for 48 hours. Symptomsappeared after 10-14days but only if the plants were protected from direct sunlight and highternperatures,
Infection studiesare continuing to establish the rust's pathogenicity to C. odorata fromotherCaribbean locations. Otherhostrangetestsconductedtodatehaveindicatedthatsymptomsdo not develop on Ageratumconyzoides, Verononia cinerea, Rolandra fructicosa, Struchiumspaganophora, Mikania micrant~ and Pseudoelephantopus spicatus.
128

These hosts havealso beenchallenged with S. ekmaniana with all but M. micrantha andP. spicatus failing to show disease symptoms.
The host range studies are continuing during which it will be necessary to confirmresistanceby microscopicexaminationof inoculated tissues.
BIOLOGICAL CONTROL POSSmILITIES
Overaperiodof twodecades,theinitialquestionof whetherC.odorata couldbecontrolledby biologicalcontrol agents has been replaced by questions pertaining to selection of types ofbiological control agents. That a biological control agent can control C. odorata has beendemonstratedbySeibert(1989). However,failuresinothercountries(Table1)suggestthatother
.factors, such as bird predation (Cruttwell McFadyen pers, comm. 1990) could affect theeffectivenessof P. pseudoinsulata. This is probablythecase in SriLanka(Dharmadhikariet al.1977) and in Sabah (Ooi et al. 1988). Cock & Holloway (1982) suggested that climatecompatibility is important and further suggested that agents should be collected from areasmatching that of the hostcountry. Theanalysis of biologicalattributesof the agents, however,suggests that more than one agent should be introduced. So far only two species have beenintroducedintoAsiaand Africa. The eriophyidmiteis believedto havearrivedfortuitouslyintoAsiaandappearedto havelittleimpacton theweed. The nextcandidateisM.parvula. However,studiesinTrinidadhaveindicatedthatthismothisdifficulttobreedin thelaboratoryandattemptsarebeing made to develop a suitablerearing technique. Countries pursuing biological controlof C. odorata should attempt to introduce all four agents. In addition, the search for newarthropodbiological controlagents should not be abandoned.
Besidesarthropods,therehasbeenadvancementin thestudyof potentialfungalpathogensto control C. odorata. It appears that C. praelonga is a host specific and recent studies of itsgerminationat IS-20°cmay make it a likelybiological control agent Studies of S. ekmanianashow encouraging results. Hence, the future of biological control of C. odorata now appearsbrighter with a wider arsenalofbiologicalcontrol agents working together to reduce the weedto an innocuousplant.
Table 1: Summary of attempts at biological control of Chromolaena odorata (after Julien1987; Napompeth et al. 1988; Seibert 1989)
Ghana Guarn India Malaysia Nigeria Sri Lanka Thailand(Sabah)
P. pseudoinsulata Not Est. S'cessful Establ'ed Establ'ed Not Est. Establ'ed Not Est.
A. brunneonigrum Not Est. Not Est. Not Est. Not Est. Not Est.
Not Est =Not establishedEstabl'ed = Agents establishedwithout much impactS'cessful = Very successfulcontrol of weed- =Not introduced
129

Table 2: Comparative score or four arthropod biologicalcontrol agents or Chromolaenaodorata using Goeden's (1983) criteria.
Category Pareuchaetes Mescinia Apion Acalituspseudoinsulata parvula brunneonigrum adoratus
1-1 4 6 1 21-2 2 2 2 01-3 4 4 2 41-4 3 3 3 31-5 0 0 0 01-6 0 0 0 01-7 3 3 3. 01-8 2 2 2 6Subtotal 18 20 13 15
11-1 6 6 6 611-2 2 0 2 211-3 6 2 2 611-4 6 6 6 6Subtotal 20 14 16 20
Ill-I 6 0 -6 0I11-2 6 6 6 6I11-3 0 0 0 -12Subtotal 12 6 0 -6
Total 50 40 29 29
Table 3: List or pathogens or Chromolaena odorata round in Trinidad and Tobago.
130
CionothrixpraelongaWint.(Arthur)
Septoria ekmaniana Petrak& Cif.
Anhellia niger (Vingas) von An.
Mycovellosiella perfoliati (Ell. &Everh.)Deighton
Phomasp.
Basidiomycotina, Uredinales
Deuterornycotina, CoeIomycetes
Ascomycotina, DothideaIes
Deuteromycotina, Hymphomycetes
Deuteromycotina, Coelomycetes

ACKNOWLEDGEMENTS
TheauthorsaregratefultoJeffWaageandDavidGreathead fororganisingthecompilationof this report. The contribution of the scientific staff at bothTrinidadand SilwoodPark, U.K.is greatlyappreciated.
REFERENCES
Aterrado, E. D. & R. L Talatala-Sanico. 1988. Status ofChromolaena odorata research in the Philippines.In: R. Muniappan (00.) Proc, First Intern. Workshop on BioI. Control of Chromolaena odoratapp. 53-55. Agr. Expt, Sta.• Unv, Guam, Mangilao, Guam
Bennett, F. D. & R. E. Cruttwell. 1973. Insects attacking Eupatorium odoraium in the Neotropics. 1.Ammalo insulata (Walk.) (Lep: Arctiidae), a potential biotic agent for the control of Eupatoriumodoraium L. (Compositae). Tech. Bul. Commonwealth Institute Bio1.Control 16: 105-115.
Cock, M. J. W. 1984. Possibilities for biological control of Chromolaena odorata. Tropical Pest Management30: 7-13.
Cock, M. J. W. & J. D. Holloway. 1982. The history of, and prospects for, the biological control ofChromolaena odorata (Compositae) by Pareuchaetes pseudoinsulaia Rego Barros and allies(Lepidoptera: Arctiidae). Bul. Entomol. Res. 72: 193-205.
Cruttwell, R. E. 1973. Insects attacking Eupatorium odoraium in the Neotropics. 2. Studies on the seedweevil Apion brunneonigrum B.B., and its potential use to control Eupatorium odoratum L.Tech. Bul. Commonw. Institute BioI. Control 16: 117-124.
Cruttwell, R. E. 1974. Insects and mites attacking Eupatorium odoratum in the Neotropics. 4. An annotatedlist of the insects and mites recorded from Eupatorium odoratum L., with a key to the types ofdamage found in Trinidad. Tech. Bul. Commonw. Institute Biol, Control 17: 87-125.
Cruttwell, R. E. 1977a Insects and mites attacking Eupatoriumodoratum L. in the Neotropics 5. Mesciniasp. nr, parvula (Zeller), Tech.Bul. Commonw.lnstitute BioI. Control 18: 49-58.
Cruttwell, R. E. 1977b. Insects and mites attacking Eupatorium odoraium L. in the Neotropics 6. Twoeriophyid mites, Acalitus adoratus Keifer and Phyllocoptes cruitwellae Keifer. Tech. Bul.Commonw. Institute BioI. Control 18: 59-63.
Cruttwell McFadyen. R. E. 1988a. History and distribution of Chromolaena odoraia (L.) R.M. King andH. Robinson. In: R. Muniappan (ed.) Proc. First Intern. Workshop BioI. Control Chromolaenaodorata pp. 7-12. Agr. Expt. Sta., Univ, Guam, Mangilao, Guam
Cruttwell McFadyen, R. E. 1988b. Ecology of Chromolaena odoroia in the Neotropics. In: R. Muniappan(ed.) Proc. First Intern. Workshop Biol. Control Chromoleana odorata pp. 13-20. Agri. Expt,Sta., Univ. Guam, Mangilao. Guam.
131

Dharmadhikari, P. R., P. A. C. R. Perera, & T. M. F. Hassen. 1977. The introduction ofAmmaIo insulasafor the control of Eupatorium odoratum in Sri Lanka. Tech. But Commonw. Institute BioI.Control 18: 129-135.
Goeden, R. D. 1983. Critique and revision of Harris' scoring system for selection of insect agents inbiological control of weeds. Protection Ecology 5: 287-301.
Julien, M. H. 1987. Biological Control of WeedS. A world catalogue of agents and their target weeds 2ndedition, CAB International Institute of Biological Control. U.K. p. 15.
Muniappan, R. & M Marutani. 1988. Rearing. release and monitoring of Pareuchaetespseudoinsulasa. In:R. Muniappan (ed.) Proc. First Intern. Workshop on BioI. Control of Chromolaena odoraie pp.43-44. Agr. Expt. Sta., Univ. Guam., Mangilao, Guam.
Napompeth, B., N.Thi Hai, & A. Winotai. 1988. Attempts on biological control of Siam weed Chromolaenaodorata in Thailand. In: R. Muniappan (ed.) Proc. First Intern. Workshop on Biol. Control ofChromolaena odorata pp. 57-62. Agri. Expt, Station" Univ, Guam, Mangilao, Guam.
Ooi, P. A. C., C. H.Sim, & E. B.Tay. 1988. Status of the arctiid moth introduced to control Siam weed inSabah, Malaysia. Planter, Kuala Lumpur 64: 298-304.
Seibert, T. F. 1989. Biological control of the weed Chromolaena odoraia (Asteraceae), by Pareuchaetespseudoinsulata (Lep.: Arctiidae) on Guam and the Northern Mariana Islands. Entomophaga 34:531-539.
Syed, R. A. 1979. An attempt on biological control of Eupatorium odoratum L.f. in Sabah, Malaysia. In:Proceedings of the sixth Asian-Pacific Weed Science Society Conference, Jakarta, Indonesia.Vol. IT pp. 459-466.
132

GUIDELINES FOR INTRODUCTION, REARING, FIELD RELEASEAND ESTABLISHMENT OF PAREUCHAETES PSEUDOINSULATA
The followingguidelineswereadoptedfollowingdiscussionat the2nd InternationalWorkshopon BiologicalControl of Chromolaena odorata.
A. Pre-importation
1. Sourcesof P. pseudoinsulaia1.1. Guam (Auention Dr. R. Muniappan)1.2. Sri Lanka or India1.3.Trinidad
2.Conduct nationwidesurvey to determinearthropodson C. odorata.
.3.Secure permission fromauthoritiesto import P.pseudoinsulala.
4. Secure funding to support programmefor at least 2 years.
5. Identify personnel to carry out the programme.(Minimum: 1 scientist/entomologist + 2 assistants).
6. Ensure adequatesupply of C. odoraia (in pots or field patches).
7. Arrangefor 1-2 weeks training for programmescientist tofamiliarize with rearing techniquesand hand carry firstconsignment to initiatecolony.
B. CoUection and Despatch
1. 1000 insects in egg stage shouldbe introduced.2. Insects preferably laboratoryreared. .. .3. Whereverpossible,handcarryconsignmenttodestinationand avoid X-ray(airport security).4. Ensure efficient communication link up betweensource and recipient countries.
C. Quarantine
P. pseudoinsulata hasbeen extensivelytestedand found to be very host specific.While furtherhost testingis unnecessary. it is prudenttoadhere to nalionalquarantinerequirementsand adoptmeasurestoensure that noarthropods other thanP.pseudoinsulata arrivewith theconsignment.
1.A minimum quarantineroom. whichis insectproof. is required toexamine theconsignment.The consignmentshould be opened only in this room.
133

2.Afterconfumingthecontent,eachbatch ofeggs IS transferred toa 500mlcupwithlid.A freshC. odorata leaf is provided.
3.Uponeclosion,removematerials from sourcecountryanddestroyinformalin/alcohol/freezer.
4.Recordand reportnumberof emergence.
D. Rearing
1. A minimum of one roomwith severalcages is required to rear P. pseudoinsulata.
2. Rear no more than 30 newly'hatched larvae in 500 ml cups togetherwith 1-2 leaves/cup
3. Change leavesevery 2 days but no more than 2 leaves/cup.
4, After2nd moult (ca. I weekafter hatching) transfer larvaeto 2 liter cups lined withgauzeor 1/4 inch hardware cloth.
5. After3rd moult(ea, to daysafter hatching) move50 caterpillars into 100liter trashcans (orsomething similar). Closethecan withfinenylonmesh clothheldbyropeorrubberband.Stemsof C. odorataare added into the can to feed the caterpillars. Clean the cans every 2-3 days.
6. Pupation occurs amongstthe frass or in leaves. Collectpupaeandplaceinoviposition cages(30 x 30 x 30 cm) for emergence and subsequent mating.
, E. Field Release
1. Select2 or 3 localities with healthy patchesof C. odorata.
2.Ateach locality release500mothsor2000larvae/releaseandcontinuetoreleaseoveraperiodof time.
3. It is preferable to release the insectat night
F. Monitoring
1.Following release,establishmentof mothshouldbemonitored monthly fora periodof 2years.
2. Astheinsect isnocturnal, suchmonitoring shouldbecarriedoutat nightorjust beforesunrise.
134

NEWAPPROACHES TO THE BIOLOGICAL CONTROL OFCHROMOLAENA ODORATA IN SOUTH-EAST ASIA
RE. CRUTIWELL McFADYENAlanFletcherResearch Station,Queensland Departmentof Lands,.P.O. Box36, Sherwood, Brisbane, Queensland, 4075, Australia
INTRODUCTION
Paperspresentedat the first International Workshop in Thailandhighlighted both theseriousness of the weed problem caused by Chromolaena and the fact that it is still activelyspreading. Successful biological control was reported from Quam and theMarianas but notelsewhere, and at that time therewere fewother biological controlprogrammes active. Threeyears later, the situation is very different; reports have been presented here of renewedinvestigations in Trinidad and South America and of agent introductions imminent in WestAfrica and Indonesia. At the same time, there, has been increasing interest in the weed inAustralia, andproposalsforactive Australian participation inbiological controlprogrammes inthe countries ID ournorth.
THE THREAT TO NORTHERN AUSTRALIA
Despite the erroneous record in Holm et al (1977), Chromolaena does not occur inAustralia, but thereare areasof northern Australia whichare climatically suitable(Fig. 1),andtherisk ID Australia is increasing. Concernovertheincreasing spreadofChromolaena in southeast Asiaand the threat that thisposed ID Australia was first raised in 1984.ID 1985, as a resultof my visit ID Manila in the Philippines, and discussions there regarding the problem ofChromolaena in thesouthern islands. The 1stInternational Workshop in 1988stimulatedfurtherinvestigations, resulting intheproduction ofa mapshowinghowtheweedhasspreadsince·1940,whichclearlydemonstrated itscontinued inexorable spreadsouthandeasttowardsAustraliaandthePacific(Fig.2). Information from thatWorkshop wasincorporated intoan articledesignedto inform and warn Australian weed scientists (McFadyen 1989), and at the same time, aQuarantine leafleton the weedwas produced and circulatedto possiblenorthern entry points(Commonwealth of Australia 1988). Anindependentstudyof weedthreatsID northernAustraliawas commissioned by the Australian Quarantine and Inspection Service, and the 1989reportidentified Chromolaena odorataas the worstweedthreat, bothon the groundsof the dangerofaccidental introduction and of the potential damage likely to be caused by its presence (p.Michael Unpublished Report 1989).
THE PROPOSED BIOLOGICAL CONTROL PROGRAMME
As earlyas 1986,suggestions had beenmadethat Australia shouldact to promote thesuccessfulbiological controlof thisweed insouth-eastAsia,thereby reducingtheamountofseedand the speed'ofcontinued spread. ACIAR wasaskedfor support,and sponsoredthree peopleto the 1stWorkshop in Thailand, twoscientistsfrom the Philippines and myself. As a resultofthat Workshop, a definite proposal was submitted to ACIAR, for a co-operative biological
135

controlprogramme whereby Australia wouldsupplythescientific expertise, andsuitableinsectswouldbe testedand releasedin Indonesia and the Philippines.
Therationale for thisapproach is thatAustralia andparticularly Queensland hasa greatfundofexpertiseandexperience inthebiological controlof introduced weeds,particularly thoseof pastureand plantation crops. The AlanFletcherResearch Stationin Brisbane is part of theQueensland Government Department of Lands, and has been actively involved in importing,host-specificity testingandfield-releasing of insectsandpathogens for thebiologicalcontrolofweedseversincethe 192Os. Wecurrently haveactiveprogrammes against9 weeds,and importonaverage3 newspeciesof insector diseaseeveryyear. In particular, we haveexperiencewiththe introduction and release of insects against two other Eupatorium species, crofton weedAgeralina (Eupatorium) adenophora and mistflower Ageratina (Eupatorium) riparia, andfortuitously we also have on the staff the entomologist (myself)who was responsible for theoriginal investigations of the insectsof Chromolaena in the West Indiesin the late 196Os.
The AFRS also hasconsiderable experience in the collection and shipmentof insects- from the Americas. We have imported insectsfrom South America for thecontrolof Harrisia
cactus, from Mexico for the controlof Parthenium weed,and currently have an entomologistbased in northern Argentina testing and shipping insects for the control of Partheniumandlantana,as wellas a secondentomologist basedin southern Texas. Boththeseare experiencedfieldbiocontrolentomologists, withconsiderableknowledge of theareaandtheplants,andeithercouldcollect the insectsrequired from Chromolaena andarrangeshipmentto Indonesiaor thePhilippines.
PROPOSED PROGRAMME
Since the original investigations were made between 1967and 1972,several papershave commented on the number of potentially useful biocontrol agents which exist in theAmericas butwhichhaveneverbeentried(Cock 1984; McFadyen 1989). Only twoagentshavebeen deliberately released; the seed-feeding weevilApionbrunneonigrum whichwas releasedin Ghana,Nigeria. India, Sri Lankaand Malaysia but failed to establish,and the arctiid mothPareuchaetespseudoinsulata whichhasbeenreleasedinGhanaandNigeriainWest AfricaandIndia, Thailand, Sri Lanka, Malaysia and the Marianas in South-east Asia, but has onlyestablished in the last 3 (Julien 1987; Muniappan et al 1988). The eriophyid mite Acalitusadoratuswas tested but nevercleared for release, but hasreachedsouth-eastAsiaaccidentallyand is now widespread through muchof the region. Partlyasaresultof the1stWorkshop,theEuropeanCommon Marketis financing a 4-yearprogramme withIRHOand IffiC to introducePareuchaetesandthestem-tipmothMesciniaparvulaintoCoted'Ivoirein westAfricaand intoIndonesia, and to investigate the possible use of pathogens as biological control agents.However, only these two insectsare to be trialled at this stage. .
136

Table 1. Potential Insect Biological Control Agents against Chromolaena
Name Damagecaused Specificity Problem Country found
LEPID. Shoot-borer: Tested;genus Mating in cages All AmericasMescinia damagingparvula
Actinote anteas Leaf-feeder untested: genus Mating in cages Trinidad,Costadamaging Rica
Bucculatrixsp. Leaf-miner, not untested: genus Mexicoverydamaging
DIPTERA Bud-galls: untested: genus rearing in cages All AmericasPerasphondylia damagingreticulata
Clinodiplosis Tip-galls: untested: genus rearing Trinidaddamaging
Procecidochares Stem-galls: untested: genus parasites Americansp. mod. damaging mainland
Melanagromyza Shoot-borer: untested: genus mating in cages WI, Southeupatorie/la damaging America
COLEOPT. Stem feeder: untested: genus All AmericasChlamisus not damaginginsularis
Aulocochlamys Stem-feeder: untested: genus Trinidadmod. damaging
Pentispa Leaf-miner: untested: genus prefers shade Trinidadexplanta mod.damaging
Rhodobaenus Stem-borer: untested: genus host range Trinidadcariniventris damaging
137

TheproposedACIAR programme. therefore. wouldconcentrateon themostpromisingof theagentspresent in Southand CentralAmerica. excludingthe 4 speciesalready trialledorincludedin the IRHOprogramme. Thereare manypotentialagents (Table 1) and the priorityassignedto thesewillvaryaccordingto the viewsofthe scientistsdoing the selectionas wellasthe country for which they are intended However. for' the proposed ACIAR programmethefollowing threefactors areof over-ridingimportance; 1)theinsectmustbe highlyhost-specific.to thegenusChromolaena at least;2) theinsectmustbeeasytokeepsafelyinquarantinewithoutthe need for a modem high-security building;3) the insect must be easy to rear. with a known,technique alreadyestablished. These three factors meanthat the ProjectLeaderswouldbe ableto assure the governments involvedthaton the one hand no risk whateverwas involved in thequarantine rearingandhost-testing of the insectselected.whileon theother that a colonycouldbeestablished andvaluableexperience in rearingandhost-testing gainedby the localentomologists involved.
In view of these factors, the first insect selected is the stem-galling tephritid flyProcecidochares sp, ThisflywasfoundinVeraCruz,MexicoandinBelem,Brazil.andprobablyoccursovermuchof mainland SouthandCentralAmerica. through not in theWest Indies. Twoother flies in the same genus. Procecidochares utilis and P. alani, have been used for thebiological controlof their hostsAgeratina adenophora and A. riparia respectively, in Hawai'iandAustralia. Bothare highlyhost-specific. attackingas faras isknownone plantspeciesonly,ie P. utilis fromA. adenophora will not attackA. riparia and vice-versa, Bothare easy to rearin cagesand, becauseonly the adult fliesare mobileand theseare quite large,3-4mmlong, arecomparatively easytomaintain safelyinquarantine. InHawaiibothspecieshavegivenexcellentcontrolof their host weeds,whichare no longerconsidered a problem. In Australiacontrol ofthe weedshas not been adequate because the galls are heavilyparasitisedby native parasites.There is no way of determining whateffect parasitesmight have on the species in the variousislandsof Indonesiaor the Philippines.
Theproposedprogramme, therefore, isCor a permitforthe introduction of thistephritidto berequestedfrom thegovernments concerned. Oncegranted.stemgallsof Procecidochareswould be collected in South America and shipped directly to the quarantine insectary' inIndonesiaor thePhilippines,wheretheywouldbereceivedby anentomologistfromthe AFRS.The galls would be kept within a secure cage for adult emergence. all parasites would bedestroyed and adultProcecidochares transferred to a rearingcage withplantsof Chromolaena.Once a parasite-free colony was established, host-testing would be carried out by the localentomologists, withassistance from theAFRS entomologistas required. Whenhost-testinghadbeen completed to the satisfaction of the government, mass-rearing and fieldreleases wouldbegin. and releases would then be planned for all islands within the archipelago. Regularmonitoring wouldbe necessary todeterminewhetherestablishmentwasoccurringand whetherparasitism wasdeveloping as a problem.
138

CURRENT STATUS
Asaresultofdiscussionsat thisWorkshop, it isclear thattheprospectsfor thisproposedprogrammehave improvedgreatlyover the last 12months. The IRHOprogrammehasreceivedthenecessarypermitfromtheIndonesiangovernment fortheintroduction ofthemothPareuchaetesfrom Guam, and the Marihat researchstation in Sumatra is well equipped to rear and host-testthis or other potential biocontrolagents. Matters are less advanced in the Philippines, but theGTZ biologicalcontrolof pestsprogrammecouldserveas a baseandliaisonfor the introductionof newagents,and thereareentomologists hereandat Los Banosinterestedin participating. Asa result,ACIARshouldnowbeabletodrawupa firm budgetand 3 yearprogramme,and allocatefunds to this, hopefully to start next year.
REFERENCES
Cock,MJ.W. 1984.Possibilitiesfor biological controlof Chromolaena odorata. Tropical PestManagement30:7-13.
Commonwealth of Austraila, 1988. ChristmasBush. Plant QuarantineLeaflet No. 56,Departmentof Primary Industriesand Energy, Canberra.
Holm, L.G., D.L. Plucknett,J.V. Pancho & J.P. Herberger. 1977. The World's WorstWeeds: Distribution and Biology. East-WestCenter, Univ. Press, Hawaii, U.S.A.
Julien, M. 1987. Biologicalcontrol of weeds: A world catalogue of agents and their targetweeds. 2nd ed. CAB. International; Wallingford.
McFadyen,R.E. Cruttwell. 1989. Siam weed: a new threat to Australia's north. PlantProtectionQuarterly,4, 3-7.
Muniappan,R., M. Marutani& G.R.W. Denton.1988. Introductionand establishment ofPareuchaetes pseudoinsulata Rego Barros (Arctiidae) against Chromolaena odoratain the Western Caroline Islands. J. BioI. Control 2: 141-142.
139

~ Potential Distribution
Lill12iJ (over 1000 mm annual rainfall) o Current Distribution
(1990)
140
Fig.I. Current & Potential Distribution of Chromoltuna odoratilin Asiaand Africa

spread 19.40-1960
Calcutta (1845)
,."". probable direction of spread
Wt,:~ present before 1940
EQuator
Tropic of Capricorn--_._..~._ _._.-..-_ _ __._-._ _ - 1····_·····_··_····1;~)j;1;lli~~1 spread 1960-1988
Fig~ 2. Spread of Chromolaean odorata in South-east Asia.
141

j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
j
J

SUCCESSION OFVEGETAnON AFTER SUPPRESSION OFCHROMOlAENA ODORATA BYPAREUCHAETES PSEUDOINSUIATA
IN GUAM
MARI MARUTANI and R. MUNIAPPANAgricultural Experiment Station,
University of Guam,UOO Station, Mangilao GU 96923 USA.
ABSTRACT
Chromolaenaodorata(L.) King & Robinson had been one of the dominant weeds in openareas and along a roadside in the northernandcentral parts ofGuam. In 1985,a biocontrol agent,Pareuchaetes pseudoinsidata Rego Barros (Lepidoptera:Arctiidae) was introduced and itsestablishment was confirmed in the north central part of Guam. Although 100 % of defoliationof C. odorata was recorded initially at release sites, the weed has recovered to some degreeafterwards. Relative importancevalueofplant species ina fivesquare meterquadratat three siteswere measured from July 1987to May 1989 to estimate dominance of plant species and to studyseasonal changes in plant competition in the field where C. odorata was dominant in 1984 andsuppressed by P. pseudoinsulata in 1985-86. Three components were included to calculaterelative importance value of a species: number of individual plants, plant height, and vegetationcover within the quadrat The names of plant species found within three 5mx5m permanentquadrats and seasonal changes of plant distribution expressed by a species importance valuearepresented. C. odorata did not become a dominant species during this study.
INTRODUCTION
The first recordof Chromolaenaodorata(L.) King & Robinson (Asteraceae) in Guam wasdocumented in 1963 as a herbarium specimen 'at the University of Guam. By early 1980, C.odorata had replaced many native and exotic plant species in fallow lands and along roadsides.Biological control of the weed was initiated at the Agricultural Experiment Station in 1984supported by a grant from thePacific Basin Tropical and Subtropical Research Program, Specialgrants, CSRS, United State Department of Agriculture. Seibert (1989) reported his work onintroductionand establishmentof Pareuchaetespseudoinsuiata Rego Barros (Lep.:Arctiidae) inGuam. Approximately 100pupae of P.pseudoinsulata were obtained from theCommonwealthInstitute of Biological Control (now named the CAB. International Institute of BiologicalControl) Bangalore, Indiaand 55 pupaeand late instar larvae from Trinidad. Mass rearing in thelaboratoryand field releasing was carried out in early 1985,and the first recovery of field-reared
143

P. pseudoinsulata was observed in June 1985in northcentralGuam. The initial spread of P.pseudoinsulata wasveryslow. Two monthsafter establishmentat the initial releasepoint therewereno signsof the insect presencebeyond30 m radius. Four monthsafterestablishment(bythe4thgeneration)defoliationwasmostlycompleteat the initialreleasepoint. After 18months,P. pseudoinsulata had defoliated over 25,000 hectares of the weed. The initial impact of P..pseudoinsulata incontrollingC.odoratawasgreat. Defoliationof theweedbythe insectreached100%in mostareas. The shoot tips wereeatenand virtuallyall buds weredestroyed, The planthad no resourcesother than to producenew shootsfrom the base which weredefoliated as theyemergedfrom theground. Whengrowingtipsandlateralbudswereeatenpriorto lateNovember,no flowerswereproduced. Asa result, thedistributionof the weedhas becomepatchyand thepopulationof P. pseudoinsulata has become smallerand localized.
It is ourconcerntodeterminethe effectivenessofP. pseudoinsulata as a biocontrolagentin the long periodof timeon Guam. For this reason, we started two types of ecologicalsurveystudies. One study was to examine plant species which had started to grow and replaced C.odorata after initialdefoliation of the weedby P. pseudoinsulata at the initial release site. Thesecondand long term study is periodical visitation to selectedsites around the island where C.odoratais present withor withoutassociationof P. pseudoinsulata. In this paper, the result ofthe first study, succession of plant species replacingC. odorata after defoliation at or near theinitial release sites using the permanentquadrats is presented. .
MATERIALS AND METHODS
Three locationswereselectedlO examinetheplant speciesreplacingC. odorata at or nearthe initial P. pseudoinsulata release site. A permanentquadrat (5mx5m) was placed at eachlocation. Within a quadrat, number of individualsof a species, average height of plants of aspecies,andcanopycoverweremeasured. Foraverageheightof thespecies,10individualplantsof the species wererandomlysampledand if less than 10plants werefound within the quadrat,the heights of all plants were measured. To measure the canopy cover of the species in thequadrat,onesquaremetergrid whichhad 100of 1Ocmx1Ocm unitswas used. The maximalareaof the species in the quadrat was25 m2• Relative importance value (RIV) (%) ofa species wascalculatedby
(Relativedensity +. Relativeheight +.Relativecanopy cover)13 where
Relativedensity (%) :;:: (no. of individualsof a species/total no. of plants in a:quadrat)xloo
Relativeheight(%):;:: (averageheightofa species/sum ofaverageheightsofall species)xloo .
Relativecanopy cover (%) :;:: (canopycover of a species/totalcanopy of all speciespresent in a quadrat)xloo..
144

Data werecollectedoncea monthin the first yearandeveryother monthduring thesecondyear. The three locations studied were NCS (Naval Communication Station), Hilaan (nearHilaan Point) and Two Lovers Point (near Two Lovers Point) (Figure 1). Climatologicaldataduring the survey were obtained from National Oceanic and Atmospheric Administration,Departmentof Commerce, USA..
RESULTS AND DISCUSSION
Monthlytotalrainfalland meanair temperaturenear the studysitesduring the experimentare presented in Figure 2. Total precipitation from July I, 1987 to June 30,1989 (24 months)was 4676 mm. During the first year (July, 1987-June1988),September 1987 was the wettestmonthandFebruarywasthedriestmonth. In thesecondyear, the heavyrainfalloccurred inJulyand October, 1988and thedriestmonthof 1989wasMarch. The monthsof February to Mayareusuallyconsideredas a dry periodof the yearon Guam;however,in 1989therewas fairly heavyrainfall in Februaryand April. Meanair temperaturewasrelativelyconsistentthroughouta yearwith the range of 24.2-27.2C. The amount of rainfall and daylength seem to be major factorsto triggerseasonal changesof plantdevelopment of manyspeciesin Guam. Thedaylengthfallsin the range of 11.3hours (December)and 12.9 hours (June).
Major species found at three study sitesduring the experimentare listed in Table 1. It isnoticeablethat the majorityof these weedsare introducedfrom tropical America (Stone 1970,Moore & McMakin 1979, Moore & Krizman 1981,Whistler 1983).
In 1985, prior to introduction of P. pseudoinsulata and its establishment,all three studysites were basically covered with C. odorata (Muniappan, unpublished data); Soon after P.pseudoinsulata was established, nearly 100%of C. odorata at the release site was defoliated(Seibert 1989). The speciesrelativeimportancevalueduring the experimentfromJuly 1987 toMay 1989indicatedthatat all three locations,C. odorata never becamea single dominantweed(Figures 3, 4 and 5). Periodically P. pseudoinsulata and aphids were observed infesting C.odorata within the quadratsor near the quadrats in the field.
Many plant species includingC.odorataproducedflowersand seedsduring the monthsof December to April in the field. The first flower bud initiationof C. odorata was noticed inNovember. Floweringoccurredsoon after the bud initiation. The floweringperiod could lastor was delayed as late as Aprilor May if therewere any biotic factors involved such as partialdefoliation of the weed by P. pseudainsulata. Under normal circumstances, seed dispersalstarted inJanuaryandat that timesproutingof newshootsanddevelopmentof newseedlingsatthe ground could be found. It was observed that after establishmentofP. pseudoinsulata inGuam, plant developmentof C. odorata in the field was not synchronizedas before.
Pennisetum polystachyon (L.) Schultes (poaceae) was one of the major species duringmonthsof July to Novemberat Two LoversPoint (Figure3) and Hilaan(Figure 5). It started toflower in September-October and the seed dispersal occurred from November. Vigorousvegetative growth of this species was observed during the monthsof July and August
145

Two other importantplant species in this study were a neotropic speciesStachytarphetajamaicensis(Verbenaceae>.andafem,Phymatodesscolopendria(polypodiaceae). Scjamaicensishada higherspeciesRIV during the first yearat Hilaan (Figure5) and P. scolopendria was thedominant species at NCS during the period of the two year study (Figure 4). Additionally,Passiflora suberosa (Passifloraceae) and Conyzacanadensis (Asteraceae) showed the higherspeciesRIVs at Two Lovers Point in early 1989.
REFERENCES
Moore, P.H. & P.D. Krizman. 1981. Field and Garden Plants of Guam. Cooperative Extension Servicepublication. College of Agriculture and Life Sciences. University of Guam. 183pp.
Moore, P.H. & P.0. McMakin. 1979. Plants ofGuam. Cooperative Extension Service publication. College. of Agriculture and Life Sciences. University of Guam. 186pp. .
Seibert, T. 1989. Biological control of the weed. Chromolaena odorata [Asteraceae]. by Pareuchaeiespseudoinsulata [Lep.: Arctiidae] on Guam and the Northern Mariana Islands. Entomophaga34:531-539.
Stone. B.C. 1970. The flora of Guam. Micronesica 6:1-659.
Whistler. W.A. 1983. Weed Handbook of Western Polynesia. Deutsehe Gesellschaft fur TechnischeZusammenarbeit (GTZ) GmbH. Eschbom. 151pp.
146

Table 1. Species found at three sites during the experiment in Guam.
Species Two Lovers Point NCS Hilaan
AsteraceaeBiden« pilosa X X XChromolaena odorata X X XConyza eanadensis X X X
ConvolvulaceaeIpomoea congesta X X XIpomoea sp. X
EuphobiaceaeEuphorbia cyathophora X X XEuphorbia geniculate X XEuphorbia hirta X X X
FabaceaeLeucaena leucocephala XMimosa pudica X
MalvaceaeSida acuta XSida rhombifolia X
PassifloraceaePassiflora foetida var.hispida X XPassiflora suberosa X X X
RhamnaceaeColubrina asiatica X
RubiaceaeSpermacoce assurgens X X X·Morinda citrifolia X
VerbenaceaeStachytarpheta jamaice1l8is X X X
Monocots:Cyperaceae
Cyperus ligularis XCyperus rotundus XCyperlf-S sp, X
PoaceaeDigitaria sp. X X? Panicum sp. XPaspalum sp. X XPennisetum polystachyon X X XStenotaphrum eecundotum X X
Ferns:Polypodiaceae
Phymatodes scolopendria X X X
X= species is present at a site.
147

HilaanTwo Lovers Point •
• •NCS
N
to 2 4 KmLLJ
Figure 1. Map of Quam. indicating three study sites. Two Lovers Point, NCS andHilaan in this experiment.
148

400
E 300.5-..J..J<.LL 200Z:;;:a:
100
-er RAINFALL (mm)
..... AIR TEMP (C)28
27
26
0'25
C.::E
24 Wt-a:
23 :;;:
22
21
O+-"'-~"'-~"'-~"'-~"""'~'T"'""~'T"'""~'T"'""~'T"'""-r-'T"'""-r--r--r--r
7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5 6'87 '88 '89
MONTH
20
Figure2. Rainfall and meanair temperature near three studysites in Guamduringthe experiment. Datawas takennear NCS by National Oceanic and AtmosphereAdministration.
149

60 -0- pe- .. ch~0- .... paCl)~ ... con
~ ... col
8 40 ... 9
c:~0a..ECl) 20>:;::caQ)~
7 8 9 10 11 12 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5
1987 1988 1989
MONTH
Figure 3. Species relative importance value of 6 major plants at Two Lovers Pointduring July 1987 - May 1969: (pc) =Pennisetum polystachyon, (ch) =Chromolaenaodorata. (pa) =Passiflora suberosa, (con) =Conyzacanadensis, (col) =Colubrinaasiatica and (e) =Euphorbia hirta. .
150
50ph
'1pe
- 40 paspG) b:J
~chle
8 30cast:8.~ 20
I:::as;! 10
7 8 9 10 1112 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5
1987 1988 1989
MONTH
Figure 4. Species relative importance value of 6 major plants at NCS during July1987 - May 1989: (Ph) = Phymatodes scolopendria, (pe) = Pennisetumpolystachyon, (pasp) =Paspalum sp., (b) =Bidenspilosa, (ch) =Chromolaenaodorata, and (le) = Leucaena leucocephala.
151

50
-e- pe- .. sta::.!! 40~ .. steG> ... pasp~
Ci ... pass> 30 ... ch8c:
·ca1:&. 20.5G>>=-ca 101)a:
7 8 9 10 1112 1 2 3 4 5 6 7 8 9 10 11 12 1 2 3 4 5
1987 1988 1989
MONTH
Figure 5. Species relative importance of major plants at Hilaan during July 1987 May 1989: (pc) =Pennisetum potystachyon, (sta) =Stachytarpheta jamaicensis,(ste) =Stenophrum secundatum, (pasp) =Paspalum sp.• (pass) =Passiflora ,suberosa, and (ch) = Chromolaena odorata.
152

A NON-NUTRITIONAL RELATIONSHIP OF ZONOCERUS (ORTHOPTERA) TO CHROMOLAENA (ASTERACEAE) AND GENERAL
IMPLICATIONS FOR WEEDMANAGEMENT
MICHAEL BOPPREForstzoologisches Institutder Universitat Freiburgi.Br.,
Fohrenbuhl 27, D-7801 Stegen-WittentaI, FederalRepublicof Germany
INTRODUCTION
For many insects,plants represent much more than food sources because they not onlyutilize the nutrients but also their "toxic" secondary compounds. These secondary substancesare thought to haveevolvedas defensive principlesunder the pressureof selective herbivory.However, fora varietyof insectsthecompounds havenowlosttheirrepellenteffectsandgaineda new significance, e.g., servinginsteadas signals for hostplant detection. Some insectshavespecialized to the extent of storing unconverted plant toxins for their own defence againstpredators, and/oreven requiring them as precursorsfor the biosynthesis of intraspecific signal .compounds (pheromones). Sequestration of secondary plant chemicalsby insects is usually :linkedwithfeeding. Thereare, however, so-called"pharmacophagous" insectsthat search forspecificsecondary compounds directly,consumethemindependently of food uptake,and usethemtoincreasetheirfitness, againeitherbystoringtheseplantmetabolites fortheirowndefenceand/or using them as pheromone precursors (Boppre 1984; for references see Boppre 1986,1990).
This-paper discusses the likelypharmacophagous relationship between the grasshopperZonocerus variegatus and the weed Chromolaena odorata in west Africa and provides anexampleof the hiddenor unpredictable effects the introduction of a foreign weedcan have onnativeinsectsand on (agro-)ecosystems. Therole of secondary plantsubstances in theecologyofexoticplantsand theirbiological controlis brieflyconsidered ingeneral,andspecifically withrespect to the use of Pareuchaetes mothsfor controllingChromolaena.
The weed Chromolaena odorata
Chromolaena odorata King& Robinson 1970(Eupatoriumodoratum L.), theSiamweed,isa perennialshrubnativeto tropical America. Following its introduction intoIndia,C.odorata;hasinvadedsouth-eastAsiafrom whereit subsequently reachedwestAfrica. In its newhabitatsitspreadsquickly,forming densethickets,andhasbecomea weedwhich seriouslyinterferes notonly with natural vegetation but also with forestry, pastures and plantationcrops. For recentsynoptic accountson Chromolaena odorata see,e.g.,Muniappan(1988),CruttwellMcFadyen(1989),Ambika& Jayachandra (1990)and severalpapers in this Proceedings.
153

The grasshopper Zonocerus variegatusZanocerus variegatus (L.) (Orthoptera: Pyrgomorphidae) is a polyphagous west African
grasshopper; its dry season generation is a pest causing serious damage to various arable crops(e.g. cassava, maize, and cotton) and plantation trees ie.g.; teak and Citrus). For synopticaccounts on Zonocerus see, e.g., Chapman et al. (1986) and Chiffaud & Mestre (1990).
Zonocerus has reached pest status only recently, and this has been assumed to be due to
increasing agriculture. Toye (1974), however, suggested that there may be a correlation betweenthe increased dry season populations of Zonocerus and the spread of Chromolaena odorata, butoffered no clue regarding any possible causal link.
Zonocerus hatehlings and early instar hoppers partly feed on Chromolaena, and thethickets are preferred roosting sites. However, this plant is not an adequate nutritional hostpermitting normal development (see, e.g., Bernays et al. 1975). Despite this, Zonocerus isstrongly attracted to flowers of Chromolaena and the grasshoppers consume the flowers in largenumbers (cl Madder 1984, 1986; Boppre & Fischer 1991). However, insects fed exclusivelyChromolaena flowers from the fourth instar failed to become adult (Chapman et al. 1986). IfSiam weed is nutritionally inadequate for Zonocerus, why do the insects only become a pestwhere it occurs?
Zonocerus, pyrrolizidine alkaloids (PAs), and Chromolaena
Zanocerus elegans(Thunb.), a species closely related to Z. variegatus and which sharesthe same aposematic coloration, is known to be attracted to withered plants containingpyrrolizidine alkaloids (PAs), as well as to pure PAs, and to ingest these secondary plantsubstances (Boppre et al. 1984). Zonocerus thus seems to exhibit phannacophagous relationships with PA-plants, as do numerous butterflies and moths (Lepidoptera), some flea beetles(Coleoptera: Chrysomelidae: Gabonia) and chloropid flies (Diptera: Chloropidae) (for reviewsee Boppre 1986, 1990). Bemays et al. (1971) reported Z. variegatus to sequester and store PAsfrom Crotalaria retusa(Fabaceae); Zanocerus also consumes other PA-containing plants, e.g.,Heliolropiwn(Boraginaceae) and Ageratum (Asteraceae) (references in Chiffaud & Mestre1990). Amongst the variety of plant genera containing PAs, species of Eupatorium have beenrecognized to possess several PAs (cl Smith & Culvenor 1980, Brown 1984, Mattocks 1986).
This knowledge suggested that Chromolaena, or at least its flowers, might serve as a PAsource for Zonocerus. Recent field studies in Benin (West Africa) revealed that larvae as wellas adults of Z. variegatus are not only attracted to Chromolaena flowers (cl Madder 1984) butdried parts of various PA-containing plants, extractsofPA-plants, and certain pure PAs were alsoeffective lures; dry leaves of Chromolaena, however, are not attractive but the roots are (Boppre& Fischer 1991). Detailed chemical analyses by Biller et al. (1991) have revealed thatChromolaena odoratacontains a mixture of five PAs (with rinderine as the major component)and proved that the highestconcentrations occur in the roots and in the inflorescences; stems andleaves, however, are very low in PAs. .
In conclusion, the puzzle of the coincidence of the spread of Chromolaena and the peststatus of Zonocerus variegatus seems to be explicable by the following hypothesis: Zonocerusenjoys a non-nutritional association with Chromolaena which provides PAs; these secondary
154

plant compounds are stored and chemically protect the grasshoppers and particularly theirdiapausingeggs from predators (e.g., larvaeof Mylabris beetles)or parasitoides,and this givesrise -to the increased fitness and population density of dry season Zonocerus. WithoutChromolaena, i.e. either before its introduction, or in areas where it is lacking, or in the wetseason when Chromolaena does not bloom, PAs seem to be a limited resource restricting thegrasshoppers' reproductive success.
Behavioural,physiologicalandchemicalstudiesinprogressstronglysupport this unlikelyhypothesis. Apartfromgatheringchemoecologicalknowledge,weaim todevelopanattraeticidebased on PAs to control Zonocerus in a way harmless to both man and environment
CONCLUSIONS AND GENERAL IMPLICATIONS
Regrettably,our findingson Chromolaena and Zonocerus do not provide good news forthebiologicalcontrolof Siam weed. Even thoughsignificantreduction in seed productionof C.odorata is caused by Zonocerus, this does not seem to affect the plant populations at all-.
. However, what this new finding does do is draw attention to the potential importance of thesecondarycompoundsof Chromolaena, an aspect whichhas largely been neglectedpreviously.Moreover, it indicates a novel and inconspicuous effect of Siam weed on (agro-)ecosystemswhich might not only affect additional insect species in Africa and Asia but may also occur ina similar way with other introduced or exotic plants.
The fundamental points arising from this story should also be considered in the generalcontextof testinghostplantspecificityof potentialcontrolagents prior to their release in foreignhabitats. The common practiceof testing only or mainly those nativeplants that are taxonomically related to the targetspecies mightbe misleadingbecause manysecondaryplant substances(and these are always, and not only in pharmacophagousspecies, essential for host specificity)have evolved polyphyletically, in unrelated taxa.
Pareuchaetes pseudoinsulata Rego Barros (Lepidoptera:Arctiidae), the only leaf-feederon Chromolaena for which potential as a biological control agent against Siam weed is beingascribedat present (see Seibert 1989,severalpapersin thisProceedings,andreferences therein),is likely to be a "PA-insect", as are several other Arctiinae which obtain PAs either from theirhost plants and/or pharmacophagously as adults, store them and/or utilize PAs as malepheromone precursors (see Boppre 1990 for references). Schneider et al. (1991) have foundhydroxydanaidal to be the major volatile of the male scent organs, and this compound is wellknown to be synthesized from PAs and play a major role as a sex pheromone in other arctiids.This does not, however, permit precise predictions based on present knowledge on PAdependentmothsbecauseall exhibit specificdifferences(Boppre 1990). Studies in progressareintended to elucidate the chemoecology of Pareuchaetes which, incidentally, eagerly feeds inour laboratoryon Heliotropium, boraginaceousplants rich in PAs and common throughout thetropics. It is not inconceivable that Pareuchaetes prefers Heliotropium (and/or other PAcontainingplants?) to Chromolaena and that introductionof the moths could cause destructionofheliotropeinsteadof controllingSiamweed. Suchanoutcomewouldcause seriousecological
155

hann. This sort of possibility is why the biology of Pareuchaetes - arid any other insectassociated with Chromolaena - needs to be investigated in detail from a chemoecologicalpointof view, together with the Siam weed itself. Furthermore, biological control agents should onlybe released in foreign habitats if their establishment can be monitored, not only by observing theassumed target plant but also generally, e.g., by using female sex attractants (cf. Schulz 1991 forsex pheromones of P. pseudoinsulatat. c
ACKNOWLEDGEMENTS
Financial support of my studies and logistic assistance during field work in Benin by theDeutsehe Gesellschaft ffir Technische Zusammenarbeit (GTZ). as well as the productivecollaboration with M. Baumgart, A. Biller, T. Hartmann and particularly O.W. Fischer isacknowledged gratefully and with pleasure. I am indebted to R.I. Vane-Wright for stylisticimprovements to my English.
REFERENCES
Ambika.S. R. & Jayachandra. 1990. The problem of Chromolaena weed. Chromolaena odorataNewsletter # 3: 1-6
Bemays, E. A., R. F. Chapman, A. G. Cook, L. 1. McVeigh & W. W. Page. (1975) Food plants in thesurvival and development of Zonocerus variegatus (L.). Acrida.4: 33-45. __
Bemays,E. A., 1. A.Edgar & M. Rothschild, 1977. Pyrrolizidine alkaloids sequestered and stored by the- aposematic grasshopper. Zonocerus variegatus. 1. Zool. (Lond) 182: 85-87
Biller, A., M. Boppre & T. Hartmann, 1991. Pyrrolizidine alkaloids in Chromolaena odoraia (Asteraceae:Eupatoriae), Manuscript
Boppre,~. 1984. Redefming "pharmacophagy". J. Chem. &01. 10: 1151~1154
Boppre, M. 1986. Insects pharmacophagously utilizing secondary plant substances (pyrrolizidine alkaloids). Naturwissenschaften 73: 17-26
Boppre, M. 1990. Lepidoptera and pyrrolizidine alkaloids: exemplification of complexity in chemicalecology. J Chem Eoo116: 165-185
Boppre, M. & O. W. Fischer. 1991. Attraction of Zonocerus variegatus (Orthoptera: Pyrgomorphidae) to
Chromolaena odoraia (Asteraceae) and other sources of pyrrolizidine alkaloids. Manuscript
Boppre,M., u. Seibt & W. Wickler. 1984. Pharmacophagy in grasshoppers? Zonocerus being attracted toand ingesting pure pyrrolizidine alkaloids. Entomol. Exp. & Appl, 35: 713-714
156

Brown. K. S. Jr. 1984. Chemical ecology of dehydropyrrolizidine alkaloids in adult Ithomiinae(Lepidoptera: Nyrnphalidae). Rev Brasil Bio 44: 435-460
Chapman, R. F., W. W. Page & A. R. McCaffery. 1986. Bionomics of the variegated grasshopper(ZimbCerus variegatus) in West and Central Africa. Annu Rev Entomo131: 479-505
Chiffaud, 1. & J. Mestre. 1990. Lecriquet puant Zonocerus variegaius (Linne, 1758). Paris: CJ.R.A.D.
Cruttwell McFadyen, R. E. 1989. Siam weed: a new threat to Australia's north. Plant Prot. Quarterly4: 3-7
Manocks, A. R. 1986. Chemistry and Toxicology'of Pyrrolizidine Alkaloids. London: Academic Press
Madder, W. W: D. 1984. The attraction of Zonocerus variegaius (L.) (Orthoptera: Pyrgomorphidae) tothe weed Chromolaena odorata and associated feeding behaviour. Bull. Ent. Res. 74: 239247
Modder, W. W. D. 1986. An integrated pest management strategy for the African grasshopperZonocerus variegatus. Nigerian Field 51: 41-52
Muniappan, R. (ed). 1988. Proc, 1st Intern. Workshop on BioI. Control of Chromolaena odoraia.Agricultural Experiment Station, Univ. of Guam, Mangilao, Guam.
Schneider, D.,S. Schulz, R. Kittrnann & P. Kanagaratnam. 1991. Pheromones of both sexes of theweed-defoliator moth Pareuchaeies pseudoinsulaia (Lepidoptera: Arctiidae). J. Appl.Entomol.. in press.
Schulz, S. 1991. The chemistry of the putative female sex attractant pheromone of Pareuchaetes __pseudoinsulata Rego Barros (Lep.: Arctiidae). In: R. Muniappan & P. Ferrar (eds). PtOc.-2ridIntern. Workshop on BioI. Control of Chromolaena odorasa pp. 113-117, ..Agri. Expt Sta.,Univ. Guarn, Mangilao, Guam.
Seibert, T. F..1989. Biological control ofthe weed, Chromolaena odoraia (Asteraceae), by Pareuchaetes pseudoinsulata (Lepidoptera: Arctiidae) in Guam and the Northern Mariana Islands._Entomophaga 34: 531-539
Smith, L. W. & C. C. J.Culvenor. 1980. Plant sources of hepatotoxic pyrrolizidine alkaloids. J. Nat.o Prod. 44: 129-152
Toye, S. A. 1974. Feeding and locomotory activities ofZonocerus variegaius (L.) (Orthoptera,Acridoidea). Rev. Zool. Mr. 88: 205-212
157


INSECT INDUCED CHANGES IN CHROMOLAENA ODORATA
JAMES McCONNELL. MARI MARUTANI. and R. MUNIAPPANAgricultural ExperimentStation
University of GuamUOO Station.Mangilao, Guam96923
ABSTRACT
Pareuchaetes pseudoinsukua was introduced to Guam to control the noxious weed.Chromolaena odorata. Biochemical and physiological studieshave been conductedto revealchangesin C.odorataleaveswhenthe larvaeof P. pseudoinsulata attack theplant. The rate ofphotosynthesis and the amountof chlorophyll werereducedin insect-infested plants. Polyacrylamide gel electrophoresis indicated a distinct change in the small subunit of ribulose-Ldbisphosphate carboxylase of insect induced yellow leaves. A preliminary experiment offlavonoid analysissuggested additional compounds present in insect inducedyellow leaves.
INTRODUCTION
Pareuchaetes pseudoinsulata Rego Barros (Lepidoptera Arctiidae) was establishedinGuamtocontrol Chromolaena odorata(L.) R.M.King& H. Robinson (Muniappan et al. 1989.Seibert 1989). When caterpillars of P. pseudoinsulata feed on C. odorata, green leaves turnyellow. Thisyellowing isa reversiblesenescence inwhichremovalofP.pseudoinsulata resultsin yellowleaves returningto a greencolor. Palatability and insect survivalare influencedbyphysiological andbiochemical changesin the plant(Marutani & Muniappan 1991). This paperpresentsthe resultsof a preliminary studyon biochemical and physiological changesin insectinducedyellowing leavesof C. odorata.
RESULTS
1. Gel electrophoresis of proteins from C. odorata.In the firstexperiment. roots.younggreenstems.olderstems.youngleaves(tipof stem).
matureleaves(middleof stem).andold leaves(baseof stem)fromC. odorataweresampledtoexamine the variation in the total protein composition in various plant parts. In the secondexperiment, mature leaves of either non-infested green C. odortua or insect-induced yellowleaves were sampled from populations in the field. In the third experiment. six plants of C.odorata, in pots. werecovered with a muslincloth. Five caterpillars of 3rd to 5th instarof P.pseudoinsulata wereplacedoneachone of thefourplants. Theplantswerecomparedover timefor physiological and biochemical changesdue to the feeding insects,
The same gel electrophoresis procedure was used in all experiments. About I g plantsamplesweremacerated in extraction buffersolutionin a plasticbag. The gels used were 12%SDSpolyacrylamidepreparedaccording to Laemmli(1970)and Ausubel et al. (l989). Samplesof 40 ul wereused per well. The gels were run with Ix SDS bufferat 30mA for 5 hours.
159

In the rust experiment, differences in the protein profilewere foundamong the variousplant parts. Rootsand older stems were found to have fewer protein bands. There were noqualitative differences in totalproteinsamongthe young.matureor old leaves.
In the second experiment, leaf tissues of all green plants in the field populations of C.odorata were identical whereas leavesof insect-induced yellow plantsdisplayedvariation ofprotein banding. There was also variation within the yellow examples. Majorchanges werenotedintheregionof thelargeandsmallsubunitsofribulose-Lf-bisphosphatecarboxylase. Thebandsbecomefaint in yellowleaves.
In the third experiment. artificial infestation of c.' odorata confirmedthe results of the'secondexperiment Among thefourplantsinfested. twoturnedyellow fasterthantheothers.Thechangein theband for the small subunitof ribulose-l,S-bisphosphate carboxylase occurredtoa greaterdegreein the fasteryellowing plants. The plantswith no insectsdisplayedthe sameprotein patternas the greenplants in experiment 2. .
.2. Rate of pbotosyntbesis, cbloropbyll content and measurement of color in insectinfested C. odorata.
The caged plants in experiment three described above were also examined for theirchangesin rateof photosynthesis, chlorophyll contentand quantitative measurement of color,Rateof photosynthesis wasestimated from therateof CO
2uptake as measured withan LI-6200
photosynthesis meter (Licor). Chlorophyll contentwas estimated with colormetric readings'usinga chlorophyll meter(SPAD MinoltaCorp.). Achromometer (Minolta Corp.)wasused to .measure color in three components. L (luminance), a (green-red) and b (blue-yellow).
Insect-infested plantsdisplayed reduced ratesof photosynthesis and reducedchlorophylllevels.
3. Leaf cbloropbylls in green and insect-induced yellow leaves by "PLC.Acetone extractsof leafchlorophylls wereanalyzed by HPLC. Leaf tissuesof 2-4gfresh
weight were immersed in deionized water for 15 min. Leaves were gently wiped dry andhomogenized in75mlacetone. Aslurrywasfiltered through glasswoolusinga Buchnerfunnelconnected toawateraspiratorandresiduewasextracted repeatedly withacetoneuntilfiltrate wascolorless. The amountof filtrate was measured and an equal amountof petroleum ether wasadded. Theentiremixturewasplacedina separatory funnel andwater(15%of mixturevolume)added. Thefunnel wasinvertedseveraltimesandthenimmobilized untiltwodistinctlayerswerepresent Mter thelowerlayerwasdrainedanddiscarded. theupperlayerwaspouredintoa roundfWk wrapped withaluminum foil. Thesolventwasevaporated andtheresiduewasresuspendedin3mlofacetoneandstoredat -20°C. Samples wereseparated inan HPLCwitha reversephasecolumn. . .
HPLCchromatogramsofthechlorophyllextractions werecompared81445 nm. Therewasa great reduction in all pigments detected at this wavelength. The pigments werecollectedasseparated and are beingidentified. '
160

4. Protein analysis of C. odorata leaves and salivary gland of P. pseudolnsulam by"PLC.
Inadditionto separating theproteinsof C.odorata andP.pseudoinsulata byelectrophoresis,asizeexclusioncolumn(SEC)hasbeenusedwithanHPLC. Thereasonforusingthismethodwas due to difficulty in obtaining the chemicals needed for preparingacrylamide gels. Thecolumnwas4.6 mmID x 25cm longandwaspackedwith5·wnparticlesizesiliconchemicallybondedwitha hydrophilic polymerlayer. Theporesizewas300A. Thiscolumnsseparatestheproteins by molecular size.
The peaks observed were similar to the results obtained by electrophoresis. Most.noteworthy, again,werethechangesof the sub unitsof ribulose-I,S-bisphosphate carboxylase.
CONCLUSIONS
The responseobservedin C. odorataappearsto be an exampleof reversiblesenescence.WhenP.pseudoinsulata larvaefeedon theleavesof C.odorata, thereis a responsein theentireplant whichresults in thedegradation. of compounds associated withthe normalfunctioning oftheplant. Specifically compounds associated withphotosynthesis havebeen foundto degrade.If the caterpillars are removed the degradation is reversedresultingin a fully recoveredplant.
REFERENCES
Ausubel, F. M., R. Brent, R. E. Kingston, D. D. Moore, 1. G. Seidman, 1. A. Smith. & K. Struhl. 1989.Short protocols in molecular biology. Greene Publishing Associates and Wiley-Interscience.New York.
Laernmli, U. K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (London) 227:680-685.
Marutani. M.&R. Muniappan. 1991. Interactions between Chromolaena odorata (Asteraceae) andPareuchaetes pseudoinsulaia (Lepidoptera: Arctiidae). Annals of Applied Biology. In Press.
Muniappan.,R., V. T. Sundaramurthy & C. A. Viraktamath. 1989. Distribution oiChromolaenaodorata (Asteraceae) and bionomics and consumption and utilization of food by Pareuchaetes pseudoinsulata (Lepidoptera:Arctiidae) in India. Proc. Vll. 1nL Symp. BioI. Contro.Weeds, Delfosse, E. S. (ed.) 1st. Sper. Pato!' Veg. (MAP) pp.401-9.
Seibert, T. F. 1989. Biological control of the weed, Chromolaena odorata (Asteraceae),byPareuchaetes pseudoinsulaia (Lepidoptera:Arctiidae) on Guam and the Northern MarianaIslands. Entomophaga.34:531-539.
161

1
111
1
11
1
1
11
1
1
1
111
1
11
1
1
11
1
1
1
111
1
11
1
1
11
1
1
11
1
11
11
1
1
11
1
1
11
1

COUNTRY REPORT
ACTIVITIES CARRIED OUT BY FRANCE
M. DeatIRCf/CIRAD
AMATROP, B.P. 5035, 34032 MontpellierCedex, France
InFrance, researchactivitieson Chromolaena odorata controlare carried out by CIRAD.CIRAD, the Intemational Center for Agricultural Research for Development, is a scientificorganization specializing in tropical and subtropical agriculture. It participates in researchprograms, rural and agriculturaldevelopment operations and training activities in cooperationwith more than 60 countries in Africa, Asia, the Pacific region and Latin America.
It employs 1900 people including 900 senior scientists. It has 11 separate departmentsincluding IRCA (Rubber Department), IRCT (Cotton and other Textile Crops Department),IRHO(OilCropsDepartment),IEMVT(LivestockProtectionand VeterinaryMedicineDepartment) and others. The headquarters is located in Paris. The research center in Montpellier, inthe southern part of France, has more than 500 scientists and technicians.
The Tropical Weed Science Laboratory (AMATROP) belongs to IRCf and developsprogramsof general interest. Eachdepartmentdevelops weedresearch in relation to its broaderprograms.
Departmentsin chargeof perennialcrops have undertakenresearch workson mechanicaland chemical corurol of C. odorata.
The Oil Crops Department (IRHO) coordinates the European Economic CommunityProject on biological control of C. odorata in cooperation with the Intemationallnstitute ofBiologicalControl, Cote d'Ivoire and Indonesia.
TheLivestockProtectionand VeterinaryMedicineDepartmentisdevelopinga methodforevaluation and mappingof C. odorata infestations in pastures of savannah regions, using highresolutiondata provided by Spot satellite,
The Weed ScienceLaboratory has undertakena research project to defme the taxonomicstatus of C. odorata from different geographic origins and coordinates the activities.of the C.odorata control program developed by the National Agencyof Livestock Production (ANDE)in the Central African Republic.
CIRAD hasdevelopedtheseactivities incooperationwith the tropicalcountries involved.
163


BIOTROP SPECIAL PUBUCATlONS
No. 1 Proceedings of the Southeast Asian Workshop on Aquatic Weeds, Malang,25-29 June 1974. BIOTROP, 1976.45 p., US$ 5.- (individual papers notincluded). _
No. 2 Proceedings of the Symposium on Forest Pests and Diseases in SoutheastASia, Bogor, 20-23 April 1976. BIOTROP, 1977. 225 p., US$ 10.-
No. 3 Proceedings of the Symposium on the Long-term Effects of Logging inSoutheast Asia, Darmaga, Bogor, Indonesia, 24-27 June 1975. BIOTROP,1978. 177 p., US$ 10.-
No. 4 Proceedings of the Symposium on Management of Forest Production inSoutheast Asia, Bangkok, 19-22 April 1977. BIOTROP, 1979. 432 p.,US$ 10.-
No. 5 Proceedings of BIOTROP Workshop on Alang-alang, Bogor, 27 - 29 April1976. BIOTROP, 1980. 277p., US$ 13.50.-
No. 6 Proceedings of the BIOTROP Symposium on Ectoparasite Biology, Bogor,21-23 June 1976. BIOTROP, 1979. 164 p., US$ 8.-
No. 7 Proceedings of the Southeast Asian Workshop on Pesticide Management,Bangkok, 14-16 March 1977. BIOTROP, 1978. 247 p., US$12.50.
No. 8 Proceedings of the BIOTROP Symposium on Animal Populations and Wildlife Management in Southeast Asia, Bogor, 15-17 March 1978. BIOTROP,1978. 236 p., US$ 12.50.-
No. 9 Proceedings of the Symposium on Pests of Stored Products, Bogor, 24 - 26April 1978. BIOTROP, 1981. 261 p., US$ 15.-
No. 10 Proceedings of the Symposium on Mangrove and Estuarine Vegetation inSoutheast Asia, Serdang, Selangor, 25 - 28 April 1978. BIOTROP, 1979.227 p., US$ 10.-
No. II Ewel, J. and L.F. Conde. Potential Ecological Impact of Increased Intensityof Tropical Forest Utilization. BIOTROP, 1980. 70 p., US$ 5.-
No. 12 Proceedings of the Symposium on Small Mammals: Problems and Control,Los Banos, Laguna, Philippines, 6 - 8 December 1977. BIOTROP, 1980.248 p., US$ 12.50.-
No. 13 Proceedings of the Symposium on Forest Regeneration in Southeast Asia,Bogor, 19-21 June 1979. BIOTROP, 1981. 233 p., US$ 15.-
NQ. 14 Biological Resources Management for Economic Development in Southeast Asia. Commemorating the Tenth Anniversary of BIOTROP 19681978. BIOTROP, 1980. 104 p., US$ 8.-
No. 15 Proceedings of the Workshop on Weed Control in Small Farms, Jakarta,Indonesia, 15-16 July 1977. BIOTROP, 1982. 162 p., US$ 8.-
No. 16 Swarbrick, J.T. et al. The Biology and Control of Hydrilla venicillata (Lf.)Royle, BIOTROP, 1982. 34 p., US$ 3.-
165

No. 17 Proceedingsof the SymposiumonMaogrove Forest Ecosystem Productivityin Southeast Asia, Bogor 20- 22 Apiil 1982. BIOTROP, 1982. 226 p.,US$ 12.-
No. 18 Proceedings of the Symposium on Pest Ecology and Pest Management,'Bogor, 18-20 September 1979. BIOTROP, 1983.286 p., US$ 10.-
No. 19 Proceedingsof BIOTROP Symposiumon Mensurational Problemsof ForestInventory in Southeast Asia, Bogor, 26-28 June 1980. BIOTROP, 1983.184 p., US$ 10.- ' '
No. 20 Proceedings of BIOTROP Symposium on Forest Pests and Diseases inSoutheast Asia, Bogor, 14-16 October 1981. BIOTROP, 1983. 243 p.,(photocopied), US$ 12.50.-
No. 21 Proceedings of BIOTROP Symposium on Wildlife Ecology in SoutheastAsia, Bogor, 7 -9 June 1983. BIOTROP, 1984. 220 p., US$ 12.-
No. 22 Proceedings of BIOTROP Symposium on Recent Research Activities onCoral Reefs in Southeast Asia, Bogor, 6-9 May, 1980. BIOTROP, 1984.83 p., US$ 4.50.- ,
No. 23 Proceedings of the DSE-BIOTROP Regional Workshop on ResearchAdvances in Agricultural Microbiology in Southeast Asia, Bogor, 24- 27November 1981. BIOTROP, 1984. 259 p., US$ 13.-
No. 24 Proceedings of the Symposium in Weed Science, Bogor, 1O-12AprilI984.BIOTROP, 1986. 376p., US$ 25.-
No. 25 Proceedings of the Symposium on Forest Regeneration in Southeast Asia,Bogor, 9-11 May 1984. BIOTROP, 1986. 227 p., US$ 17.50.-
No. 26 Proceedings of BIOTROP Symposium on the Status of Forests Pests andDiseases in Southeast Asia, Los Banos, Philippines, 3 -15 May 1985.BIOTROP, 1987. 187 p., US$ 15.-
No. 27 Proceedings of BIOTROP Seminar on Production and Exploitation of OpenWaters (With Special Emphasis on Reservoirs), Bogor, 15'-17 June 1982.BIOTROP, 1987. 125 p.', US$ 7.50.-
No. 28 Proceedingsof BIOTROP Symposiumon Practical Measures for Preventing ,and Controlling Fish Diseases, 24-26 July, 1985.BIOTROP, 1988. 155 p.,US$ 11.50.-
No. 29 Proceedings of BIOTROP Symposium on Coral Reef Management, Bogor,22-24 April, 1986. BIOTROP, 1987. 186 p., US$ 12.50.-
No. 30 Proceedings of BIOTROP Symposium on the Conservation and Management of Endangered Plants and Animals, Bogor, 18- 20 June 1986.BIOTROP, 1987.'246 p., US$ 20.-
No. 31 Proceedings of the international Conference on Tropical Plant Ecophysiology Bogor 4-6 December 1985. BIOTROP, 1988. 308 p., US$ 25.
No. 32 Proceedings of the Symposium in Pest Ecology and Pest Management,Bogor, Indonesia. 9-11 April 1985, BIOTROP, 1988. 309 p., US$ 20.-
166

No. 33 Proceedings of the Symposium on Pests of Stored Products, Bogor, Indonesia, 2-4 December 1986. BIOTROP, 1988. 161 p., US$ 15.-
No. 34 Proceedings of the Symposium on Crop Pathogens and Nematodes, Bogor,Indonesia, 7 -9 April 1987. BIOTROP, 1988, US$ 22.50.-
No. 35 Proceedings of the Symposium on the Application of Tissue CultureTechniques in Economically Important Tropical Trees, Bogor, Indonesia,7 -9 December 1987. BIOTROP, 1988. 230 p., US$ 20.-
No. 36 Proceedings of the Symposium on Biological Control of Pest in TropicalAgricultural Ecosystems, Bogor, Indonesia, 1- 3 June 1988. BIOTROP,1988. 358 p., US$ 30.-
No. 37 Proceedings of the Symposium on Mangrove Management: Its Ecologicaland Economic Considerations, Bogor, Indonesia, 9-11 August 1988.BIOTROP, 1989. 353 p., US$ 28.-
No. 38 Proceedings of the Symposium on Weed Management, Bogor, Indonesia,7 -9 June 1989. BIOTROP, 1990. 282 p., US$ 25.-
No. 39 Proceedings of the Symposium on Agroforestry Systems and Technologies,Bogor, Indonesia, 19-21 September 1989. BIOTROP, 1990. 239 p.,US$ 28.-
No. 40 Proceedings of the Symposium on Aquatic Weed Management, 'Bogor,Indonesia, 15-17 May 1990. BIOTROP, 1991. 253 p., US$ 23.-
No. 41 Proceedings of the Fourth Round-Table Conference on Dipterocarps,Bogor, Indonesia, 12-15 December 1989. BlOTROP, 1991. 574 p.,US$ 50.-
No. 42 Proceedings of the Second Asian Conference on Mycorrhizae (ACOM),Chiangmai, Thailand, 12-14 March 1991 (In press)..
No. 43 Proceeding of the Symposium on Inland Aquatic Environmental Stress. Monitoring, Bogor, Indonesia, 25 - 27 July 1990. BIOTROP, 262 p.,
US$ 25.-No. 44 Proceedings of the Second International Workshop on Ecological Control
of Chromolaena odorata Bogor, Indonesia, 4-8 February 1991.BIOTROP, 167 p., US$ 15.-
167

GAYA TEHNIK - BOGOR




















