
Early Upper Paleolithic Man and Late Middle Paleolithic Tools
39
Early Upper Paleolithic Man and Late Middle Paleolithic Tools DAVID S. BROSE Case Western Reserve University MILFORD H. WOLPOFF University of Michigan The appearance of anatomically modern Homo sapiens in Europe, the Near East, and Africa must represent either an in situ evolution o f Neandertals or a migration. Those who suggest the latter claim a sudden replacement of Neandertals by anatomically modern Homo sapiens. However, the “evidence” actually cited claims only the sudden replacement of Middle by Upper Paleolithic industries. We criticize the migration explanation on two grounds. (1) There is no “sudden replacement” of Middle Paleolithic by Upper Paleolithic industries, but rather a gradual change in the frequencies of already present tools. Numerous sites in these areas exhibit transitional industries. (2) Concomitantly, there is no morphological euidence indicating a “sudden replacement” of hominids. There is no absolute association between anatomically modern Homo sapiens and Upper Paleolithic industries. Instead, the evidence clearly shows that early anatomically modern Homo sapiens is a late Middle Paleolithic local phenomenon. THE RELATIONSHIP of Neandertals to Homo sapiens has always been of great interest to anthropologists. This work seeks to examine one hypothesis concerning the origin of anatomically modern H. sapiens, and to test the implications of this hypothe- sis against the archaeological and palaeon- tological record. By “Neandertals,” we refer to all hominid specimens dated within the time span from the end of Riss to the appearance of ana- tomically modern H. sapiens. Applying the term in this way is not unique to this work. Numerous authors have referred to this set of specimens as “Neandertaloid,” “Neander- thalian,” “Paleoanthropinae,” and so on. In fact, this usage is synonomous in content with the “Neandertal Peoples” referred to by Howell (1957:342 and Fig. 4). The speci- mens are identical, and include the often disputed remains from Fontechevadeagain following Howell (Ibid. 341-342). Grouping the specimens in this way can be justified on two grounds. First, there is a gap between this group and H. erectus. This gap is tem- poral, rather than morphological, and is probably the result of sampling accidents. Nevertheless, there are only a very few speci- mens spanning the time from the well- represented H. erectus populations at ter- minal Mindel to the beginning of Riss. With the discovery of additional specimens from this period, the temporal gap will close, as the morphological and archaeological gap has already done. Second, there is a general unity of grade among the specimens so referred. Neandertals evince crania expanded to modern size and posterior teeth reduced to modern size, along with anterior teeth and supporting facial architecture main- tained in the very robust H. erectus condi- tion (Brace 1967a, 1968; Wolpoff 1969, 1971). In most cases, this morphological pattern separates Neandertals from groups occupying the same areas before as well as after them. We distinguish this usage from the more restricted term: “classic” Neandertals. The later refers only to the people who inhabited Western and Southern Europe from the be- ginning of Wurm to the appearance of ana- tomically modern H. sapiens (Howell 1952, 1957; Breitinger 1955; Howells 1967; Le Gros Clark 1964; Hooton 1947; and many others). Thus, an important distinction is maintained throughout this work. The total group referred to as Neandertal consists of two smaller and mutually exclusive groups: 1156
Transcript of Early Upper Paleolithic Man and Late Middle Paleolithic Tools

Early Upper Paleolithic Man and Late Middle Paleolithic Tools
DAVID S. BROSE Case Western Reserve University
MILFORD H. WOLPOFF University of Michigan
The appearance of anatomically modern Homo sapiens in Europe, the Near East, and Africa must represent either an in situ evolution o f Neandertals or a migration. Those who suggest the latter claim a sudden replacement of Neandertals by anatomically modern Homo sapiens. However, the “evidence” actually cited claims only the sudden replacement of Middle by Upper Paleolithic industries. We criticize the migration explanation on two grounds. ( 1 ) There is no “sudden replacement” of Middle Paleolithic by Upper Paleolithic industries, but rather a gradual change in the frequencies of already present tools. Numerous sites in these areas exhibit transitional industries. (2) Concomitantly, there is no morphological euidence indicating a “sudden replacement” of hominids. There is no absolute association between anatomically modern Homo sapiens and Upper Paleolithic industries. Instead, the evidence clearly shows that early anatomically modern Homo sapiens is a late Middle Paleolithic local phenomenon.
THE RELATIONSHIP of Neandertals to Homo sapiens has always been of great interest to anthropologists. This work seeks to examine one hypothesis concerning the origin of anatomically modern H. sapiens, and to test the implications of this hypothe- sis against the archaeological and palaeon- tological record.
By “Neandertals,” we refer to all hominid specimens dated within the time span from the end of Riss to the appearance of ana- tomically modern H. sapiens. Applying the term in this way is not unique to this work. Numerous authors have referred to this set of specimens as “Neandertaloid,” “Neander- thalian,” “Paleoanthropinae,” and so on. In fact, this usage is synonomous in content with the “Neandertal Peoples” referred to by Howell (1957:342 and Fig. 4). The speci- mens are identical, and include the often disputed remains from Fontechevadeagain following Howell (Ibid. 341-342). Grouping the specimens in this way can be justified on two grounds. First, there is a gap between this group and H. erectus. This gap is tem- poral, rather than morphological, and is probably the result of sampling accidents. Nevertheless, there are only a very few speci- mens spanning the time from the well-
represented H. erectus populations at ter- minal Mindel to the beginning of Riss. With the discovery of additional specimens from this period, the temporal gap will close, as the morphological and archaeological gap has already done. Second, there is a general unity of grade among the specimens so referred. Neandertals evince crania expanded to modern size and posterior teeth reduced to modern size, along with anterior teeth and supporting facial architecture main- tained in the very robust H. erectus condi- tion (Brace 1967a, 1968; Wolpoff 1969, 1971). In most cases, this morphological pattern separates Neandertals from groups occupying the same areas before as well as after them.
We distinguish this usage from the more restricted term: “classic” Neandertals. The later refers only to the people who inhabited Western and Southern Europe from the be- ginning of Wurm to the appearance of ana- tomically modern H. sapiens (Howell 1952, 1957; Breitinger 1955; Howells 1967; Le Gros Clark 1964; Hooton 1947; and many others). Thus, an important distinction is maintained throughout this work. The total group referred to as Neandertal consists of two smaller and mutually exclusive groups:
1156

Brose & Wolpoff] TOOLS A N D M O D E R N H U M A N ORIGINS 1157
“classic,” and “non-classic.” The distinction is crucial to the case we make, because almost every statement made concerning Neandertal morphology, variation, archae- ology, and evolution will, when applicable, be shown to apply to both “classic” and “non-classic” groups. Following a technique used by Howells (1967:190, footnote), the two Neandertal subgroups are also explicitly defined by listing included specimens.
Evidence of hominid morphology, cul- ture, and stratigraphy has been used to substantiate or reject any number of hy- potheses concerned with the relationship of Neandertals to anatomically modern H. sapiens. One of the earliest hypotheses sug- gests the sudden replacement of Neandertals by a more modern taxon: anatomically modern H. sapiens. A close examination of the “sudden replacement” hypothesis indi- cates that the claim was established for tools, and not for hominids. For instance, as recently as 1966 Le Gros Clark stated:
At the end of the Mousterian phase of paleolithic culture, the Neandertal in- habitants of Europe were abruptly re- placed by people of completely modern European type. There is reason to sup- pose that this new population, the Aurig- nacians, having developed their distinctive culture elsewhere, probably in Asia, mi- grated into Europe and, with their su- perior social organization, quickly dis- placed Mousterian man and occupied his territory [1966:116-1171.
ABSOLUTE DATES OF LATE NEANDER- TALS AND EARLY ANATOMICALLY MODERN HOMO SAPIENS
In actuality, the extent of the hiatus measured between the radiocarbon dates of the youngest clearly defined Neandertal and the oldest clearly defined anatomically modern H. supiens specimens is not known with any certainty. Radiocarbon dates for this period of human evolution are neither extensive (Brace 1964; Oakley 1966) nor exceptionally accurate (Butzer 1964). The faunal connections between levels at the same site, let alone between sites, are more often than not incorrect (Kurten 1968). For
these reasons, only dates taken from the actual strata where hominid specimens have been found can be used. Even so, the methodology of many earlier investigators, and the possibility of burial, leave even this association of dates with specimens subject to doubt.
The youngest dates directly associated with Neandertals (A2 dates according to Oakley 1966:7) come from three sources: Haua Fteah Layer XXXIII (McBumey 1967) with mandibles I and I1 has a date of 40,000+500 BP; Tabun Layer C is dated at 40,900+1000 BP and Layer B at 39,700*800 BP (Oakley 1966); La Quina Level H1 is dated at 35,250+530 BP (Oakley 1966). There is some question concerning the validity of the La Quina date as repre- senting a late Neandertal. The hominid from Level H1 at La Quina is #14. This subject is represented by the posterior-inferior section of a left parietal (Henri-Martin 1923:242). The individual represented was a juvenile, and the level has a “final Mousterian” in- dustry. A fragment of this size and age cannot be clearly categorized as either Ne- andertal or anatomically modern H. sapiens (Piveteau 1967). Even if the association with the date is good, we do not know what type of hominid is thus dated.
The earliest dated specimens of ana- tomically modern H. sapiens come from the Pavlovian burials at Dolni Vestonice (25,820+180 BP) and Pavlov (26,620+260), according to Oakley (1966). An earlier date has been claimed for the Niah cave specimen (Harrison 1959). However, this specimen represents a burial (Harrison 1964:526), and the date does not even come from the level where the specimen was found, but rather was taken in a corresponding^' area (Ibid.: 526). Without further substantiation, the association of this crucial date with the specimen cannot be unequivocally accepted (see Fitting 1969:351).
With so few dates from this important period, it is difficult to draw well substan- tiated conclusions from the evidence available. It is interesting that even these few dates from widely scattered areas indicate

1158 AMERICAN ANTHROPOLOGIST [ 73,1971
Neandertals precede anatomically modern H. supiens. In no instance do either relative stratigraphies or absolute dates indicate ana- tomically modern H. supiens contemporary with or preceding Neandertals in any area as would have to be the case if the former evolved somewhere separately and then “suddenly replaced” Neandertals all over the world.
MIDDLE PALEOLITHIC INDUSTRIES
Not only do the hominids show a specific sequence and temporal separation, but in- dustries do also. Classically, the Wurm 1/11 interstadial is said to separate Middle and Upper Paleolithic industries. Wurm 1/11 is here used in the most standard sense rather than as in the more restricted French ter- minology where this period is known as Wurm II/III. This general time period has been called Gottweiger (although the type site seems to be Eemian): Paudorf; Arcy; Laufen; Rixdorf; Stillfried B; Molga- Sheksna; Hengelo-Denekamp; Bryansk; Odderade; or Aurignacian intersladial. While Pleistocene sequences in Europe have not been completely worked out, and while stratigraphic correlations are still uncertain, we might quote from one of the most recent geological monographs to the effect that
(of the last glaciation) the Middle Wurm which covers the time between 53,000 BP and 25,000 BP is the part of which not very much is known. While some authors assume the existence of the “Gottweig Interstadial” others deny the possibility of a notable interstadial (in central Europe). Certainly the Middle Wurm was not a continuous cold time without any temperature oscillation, but one has to suppose the existence of cooler and war- mer periods and of corresponding ad- vances and retreats of the ice sheets, [Woldstedt:1967; italics added]. Most prehistoric archaeologists and qua-
ternary geologists have found evidence for a warmer period of interstadial magnitude, during the Middle Wurm (Flint 1957:381-412; Alimen 1967:211; de Jong 1967:359; Bordes 1968:147; Kurten 1968:20ff; Basten 1969:3-11; Grazzini and
Rosen berg 1 9 6 9 : 2 7 9-92; Zubakov 1969:Table Z). While the exact dates of these warm periods are not precisely fixed all seem to have occurred some time after 38,000 BP and were concluded by the outset of extremely cold glacial conditions some time after 28,000 BP. It is this “interstadial” which we are using as a temporal horizon in this paper.
Wherever continuous archaeological se- quences spanning this period are clear there seems to be no overlap or obviously rapid replacement, but rather a gradual transition from late Middle Paleolithic to early Upper Paleolithic industries. This can be seen in a number of ways. The relative frequencies of various types of stone tools which charac- terize the Middle and Upper Paleolithic do not display sudden or dramatic changes during the Wurm I to Wurm I1 period or its geographical equivalent.
Most of the tool types which are con- sidered characteristic of the Upper Paleo- lithic are present (albeit in lower fre- quencies) in late Middle Paleolithic as- sem blages .
The evidence on which the validity of this statement rests is presented by geographic area in the body of this paper. Numerous examples of supposedly characteristic Upper Paleolithic tool types (described by exca- vators intimately acquainted with the ma- terial) are seen in what can only be regarded as Middle Paleolithic assemblages. One way of visualizing this evidence is simply to list the common Upper Paleolithic tool types (as described by DeSonneville-Bordes and Perrot 1953) and note the extensive and repeated occurrences in a Middle Paleolithic context (see Table I). The occurrences identified in Table I are based on both functional and morphological analogies.
In other words, the variation between these tool types occurring at Middle Paleo- lithic and Upper Paleolithic sites is no greater than the variation among the tool types at Upper Paleolithic sites alone. Purely stylistic variation in functionally similar chipped stone tools has only rarely been identified by Paleolithic archaeologists.

Brose & Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS 1159
TABLE 1. PARTI.41, LIST O F UPPER PALEOLITHIC TOOLS ( S T A N D A R D LIST) IN MIDDLE PALEOLITHIC CONTEXTS*
Number Standard Type Middle Paleolithic Occurrences
1.
2. 3. 4. 5.
6 . 7 . 8. 9.
10. 11.
12. 13.
14. 15. 16. 17. 18. 19. 20. 21. 22. 23. 24. 25. 26. 27. 28. 29. 30.
31. 32. 33. 34. 35. 36. 37. 38.
39. 40. 41. 42. 43. 44. 45. 46. 47. 48.
Simple end scraper
Atypical end scraper Double end scraper Ogival end scraper End scraper on retouched blade
End scraper on Aurignacian blade Fan shaped end scraper Scraper on flake Round or circular scraper Thumbnail scraper Carinated scraper
Atypical carinated scraper Thick-nosed scraper
Flat-nosed or shouldered scraper Core scraper Plane (rabot) Scraper-graver Scraper-truncated blade Graver-truncated blade Borer-truncated blade Borer-scra per Borer-graver Borer Atypical borer or beak Multiple borer or beak Micro-borer Straight dihedral graver Lopsided dihedral graver Angle dihedral graver Angle dihedral graver on broken
Multiple dihedral graver Busked graver Parrot-beak graver Graver on straight truncation Graver on oblique truncation Graver on concave truncation Graver on convex truncation Transverse graver o n lateral re-
Transverse graver o n notch Multiple graver on truncation Mixed multiple graver Noailles graver Graver on core Flat graver Audi knife Chatelperron point Atypical Chatelperron point Gravette point
or flake
blade or flake
touch
Y
La Quina; Arcy-sur-Cure; Sipka Cave; Kiilna Cave; Haua Fteah; Ordos
Repolust Cave; Weimar; Skhul Peche de I’Aze (lower layers) qepolust Cave; Skhul; Ordos Sipka Cave; Kiilna Cave; Haua Fteah
La Chapelle-aux-Saints; Haua Fteah Ordos; Kokkinopilos All sites Y
Arcy-sur-Cure; Sipka Cave; Weimar Peche de I’Aze La Chapelle-aux-Saints; Repolust Cave; Haua
La Chapelle-aux-Saints; Haua Fteah; Ordos Krapina; Repolust Cave; Weimar; Haua Fteah;
Fteah
Quafzeh ( E ) Y
Sipka Cave; Kiilna Cave; Weimar; Ordos
El Wad ( F ) gkhul; Ordos Sipka Cave; Weimar; Peche de I’Aze I1
Kiilna Cave; gipka Cave; Weimar; El Wad; Ordos Arcy-sur-Cure; Ordos
Haua Fteah; Peche de I’Aze Arcy-sur-Cure; Krapina; Peche de I’Aze I1 Krapina Ordos Skhul; Abou-Sif
Peche de 1’Aze I1 Quafzeh ( E ) Krapina; Weimar; Haua Fteah Arcy-sur-Cure Repolust Cave; Ordos
Krapina Tabun (B)
Arcy-sur-Cure; Repolust Cave; Weimar; Ordos Ko kkinopilos Abri Audi; La Quina; Regourdou Haua Fteah; Quafzeh; Ksar Akil; Peche de I’Aze Haua Fteah
*References by geographical area in text.

1160 AMERICAN ANTHROPOLOGIST [73,1971
TABLE I. PARTIAL LIST OF UPPER PALEOLITHIC TOOLS (STANDARD LIST) IN MIDDLE PALEOLITHIC CONTEXTS* (Continued)
Number Standard Type Middle Paleolithic Occurrences
49. 50. 51. 52. 53. 54. 55. 56.
57. 58.
59.
60. 61. 62. 63. 64. 65.
66.
67. 68. 69.
70.
-
Atypical Gravette point Micro gravette Vachoas point Fort-Yves point Humped pieces Flechette (dart) Tanged point Atypical shouldered Perigoidean
Shouldered piece Blade with continuous retouch
point
Blade with non-continuous re-
Piece with straight truncation Piece with oblique truncation Piece with concave truncation Piece with convex truncation Piece with double truncation Blade with continuous retouch
on one edge Blade with continuous retouch
o n both edges Aurignacian blade Notched or strangled blade Point with plane face (Solutrean)
Laurel Leaf Point (Solutrean)
touch
Haua Fteah
All sites Ksar Akil Krapina; Haua Fteah; Emireh; Quafzeh
Haua Fteah Arcv-sur-Cure: Abou-Sif; La Chapelle-aux-
Siints; Peche de 1’Aze’; Kiik-koba; Molodova; Haua Fteah
Fteah; Abou-Sif Arcy-sur-Cure; Starosele; Molodova; Haua
Krapina; Weimar; Ordos Ordos Haua Fteah Haua Fteah; Ordos
Nietoperzowa Cave; Molodova; Haua Fteah;
Starosel’e; Ksar Akil; Ordos; Peche de 1’Aze
La Chapelle-aux-Saints; Kiik-koba; Tabun (B)
Ordos; Quafzeh (E); Tabun
gaua Fieah Sipka Cave; Kulna Cave; Lovas Cave; Krapina;
NietoDerzowa Cave; Weimar; Salzgitter Lovas Cave; Krapina; Weimar; Salzgiiter ;
Mauren Cave
*References by geographical area in text.
There is, however, considerable variation among the various Middle Paleolithic as- semblages in the morphology of what are regarded as functionally equivalent tools, just as there is variation among the same “type” of tool found in the numerous Upper Paleolithic industries. Indeed there is a sur- prising amount of variation within any Middle or Upper Paleolithic assemblage in terms of the morphology of any “func- tional” tool type (viz. Bordes 1969). The definition of Paleolithic tool types generally depends on their function, as inferred from their morphology, rather than on their method of production (Bordes 1969:l-4).
The gradual frequency changes between Middle and Upper Paleolithic industries de- monstrated here are based on such tool type identifications, made by those workers per-
sonally familiar with the material; in ad- dition, we have also attempted to document the gradual changes in production tech- niques, an important, but separate problem. If we were unwilling to classify morpho- logically and functionally similar tools as members of the same general category, we would face the reductio that follows when we recognize that every individual chipped- stone tool is in some way different than every other chipped-stone tool in the world.
These considerations are readily apparent in the works of many authors with first hand knowledge of the technologies. For instance, Movius (1969:122) has noted for central France, the “Maginot Line” of the pur- ported invasion, that:
The Chatelperronian of the Grotte du Renne a t Arcy-sur-Cure gives the impres-

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1161
sion of an assemblage which basically remains Middle Paleolithic on evidence of the majority of the flint tools, but is Upper Paleolithic in terms of the tech- nology and style of the pieces.
The variations in relative frequency which do occur in the period from Wurm I to Wurm I1 seem no greater than differences between what Bordes has demonstrated (1955, 1961, 1963) to be several contem- poraneous Mousterian facies in a single geo- graphic area. The same situation seems also to exist in Eastern Europe (Klein
At the same time artifacts considered “typical” of the Middle Paleolithic continue with diminished frequency well into late Upper Paleolithic assemblages. Because of this fact industries or assemblages which can be considered “transitional” between Middle and Upper Paleolithic may or may not be segregated from the continuum, as the in- vestigator’s theoretical viewpoint dictates. This situation seems to occur in all areas of the Old World.
Throughout the following discussion of lithic assemblages problems of taxonomic nomenclature occur. In attempting to de- monstrate the variability of these industries, the question constantly arises as to whether some particular manifestation should be properly considered Middle Paleolithic. Several solutions present themselves. First, all the assemblages discussed have been called Middle Paleolithic, or the equivalent, by the original investigators; an assignment which never has been seriously questioned. Second, these assemblages occur in a stratigraphic context indicating deposition no earlier than late Riss or Riss/Wurm, and no later than the end of those Middle Wurm “interstadial” oscillations which began about 38,000 BP.
Finally, most if not all of these lithic assemblages, although displaying great in- ternal variability, display some degree of uniformity in terms of some proportion of the manufacturing techniques employed, or some of the tools produced. That is, typo- logically, as well as chronologically and authoritatiuely , the industries discussed here-
1969: 262-264).
in may all be justifiably considered Middle Paleolithic.
Western Europe
The common Middle Paleolithic in- dustries of Western Europe are termed Mousterian. Bordes (1953, 1961) has clearly shown that this is not a homogeneous lithic assemblage but rather a number of facies each of which contains more or less the same types of tools but in quite different fre- quencies. The differences between these Mousterian facies are not chronological but rather seem to be functional (Binford and Binford 1966, 1969). In all of these as- semblages numerous tool types occur which are generally considered characteristic of the early Upper Paleolithic (DeSonneville-Bordes 1963). In conjunction with Table I, it is significant that out of the standard list of sixty-two Lower and Middle Paleolithic tool types (Bordes 1953, 1955, 1961), over twenty-five percent (nos. 30-39, 42, 44, 52-54, 57, 63) are also characteristic of the Upper Paleolithic assemblages where they appear in relatively high frequency. Even in good Mousterian contexts these “Upper Paleolithic” end scrapers, burins, gravers, and backed blades may comprise a signifi- cant proportion of the tools. At Le Moustier they comprise twelve percent of the total lithic assemblage. At Pech-de-1’Aze (Level 4) they approach twenty percent of the total assemblage (Vandermeersch 1965). At La Chappelle-aux-Saints the industry is charac- terized as LaQuina Mousterian within which there existed a number of “Aurignacian” elements such as thin retouched blades, carinated end scrapers, and long, possibly pressure-flaked blades (Bardon and Bouys- sonie 1908).
Bordes (1961:810) has noted that “in some Mousterian assemblages blades com- prise up to forty percent of the debitage.” In this quotation, Bordes did not specify the type of blades referred to. “Blade” is gener- ally defined as any flake the length of which is greater than twice its width or as a long, thin, parallel-sided flake. Blades can be more

1162 AMERICAN ANTHROPOLOGIST [73,1971
precisely defined by their method of manu- facture: either soft-hammer direct percus- sion, hard-hammer direct percussion, in- direct percussion (punched), or pressure- flaked. Any of these may or may not have been struck from prepared cores of various types (Jelinek 1965). All of these produc- tion techniques have been employed with differing frequency in both Middle and Upper Paleolithic industries (Bordes 1968:242). No single archaeological as- semblage regularly displays all of these tech- niques. While the presence of some sorts of blades, or tools made on blades, is quite common at Upper Paleolithic sites, there are numerous Upper Paleolithic examples where functionally similar tools do not appear to have been produced by any particular “blade” techniques. Conversely, there are Middle Paleolithic sites which show the presence of blades produced by all of these techniques. If confusion is to be avoided, terms like “Mousterian Blades” and “Upper Paleolithic Blades” must be replaced by the well-defined descriptive terms based on methods of production or by detailed metric analyses of blade morphology.
Bone tools, long considered the sine qua non of the Upper Paleolithic also occur with some consistency at numerous Mousterian sites. In some cases these Middle Paleolithic bone tools appear morphologically and func- tionally indistinguishable from some Upper Paleolithic bone tool types (e.g., the bevel- base points from the Perigordian levels at Arcy-sur-Cure are identical to bevel-base bone points from the lower Middle Paleo- lithic layers [ LeRoi-Gourhan 1961; Bordes 19681 ; numerous other examples are cited in the Eastern Europe section). More im- portant is the fact that no two Upper Paleo- lithic “cultures” or “traditions” have the same kinds of bone tools. Indeed the sequent stages of a single Upper Paleolithic “culture” are frequently defined by morphological changes in bone tools. Because of the stylistic dissimilarity of bone artifacts throughout the Upper Paleolithic the tradi- tional viewpoint had considered the presence of any type of well-made bone tool in any
frequency as an indication of Upper Paleo- lithic affinity (Oakley 1962:27, 93-98; Burkitt 1963:79; Braidwood 1966:80). While Collie’s statement (1928:50) that bone tools, as such, first appear in the Aurignacian, is not the latest word on the subject, it still appears reasonable to many. It is to this hypothesis we have addressed our arguments.
At Gibraltar Dorothy Garrod recovered two fragmentary bone artifacts either of which may have been points (1928:50, Fig. 6). Henri-Martin (1923:Figs. 4, 8) noted the presence of a number of bone implements in the upper levels of A1 Quina. In England the Pin Hole cave contained a large number of ulnar bones awls and polished split bone points from both Middle Paleolithic and Aurignacian levels (Kitching 1963). At Arcy-sur-Cure LeRoi-Gourhan (196l:l-16) has excavated a “post-Mousterian” Mouster- ian level containing bevel-base bone points associated with a denticulate-looking Mous- terian industry containing backed blades and burins. Movius (1969:112) has recently characterized the deposits from the Grotte du Renne at Arcysur-Cure as definitive evidence for the continuity between Mousterian and Chatelperronian.
In addition to the Chatelperronian at Arcy-sur-Cure, numerous other western European sites evince this transitional indus- try. A partial list of these sites includes Chatelperron, Combe Capelle, Les Cottes, La Ferrassie (E), Cueva Morin, Trou de la Chevre, Roc de Combe (Lot), Grotte d’Fees, and Reclau-Viver (De Sonneville-Bordes 1963; Freeman and Echegaray 1970; Bordes 1968; Pradel 1966). As Bordes has recently observed, “it becomes more and more diffi- cult to avoid the conclusion that this lower Perigordian is derived from a local develop- ment of an Acheulean-tradition Mousterian” (1968:148).
As early as 1956 Bordes noted the lack of any real hiatus between the Middle and Upper Paleolithic and stated that the Lower Perigordian represented a typologically tradi- tional industry in western Europe. He also claimed that the hiatus between Mousterian

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1163
and Aurignacian was less evident than imag- ined (1958:176-179). Indeed, Pradel’s exca- vations at Les Cottes (Bordes 1968:150-153) and Le Fontenioux (Bordes, personal com- munication) seem to demonstrate the devel- opment of the Upper Perigordian from the Lower Perigordian, and Bordes has seen characteristic Aurignacian tool types in the La Quina Mousterian (1968:155-156) al- though he feels the centers of the Aurigna- cian development were not in southwestern France. More recently Movius (1969:117) has discussed this period and noted clear stylistic continuities between the Lower Per- igordian and the Early Aurignacian.
Eastern Europe
The Middle Paleolithic industries of East- ern Europe cover a much larger geographical area than do those of Western Europe and correspondingly greater differences exist among them. Valoch (1968) has recently characterized a number of more-or-less chro- nological and regional varieties not all of which can be correlated with Bordes’ Mous- terian facies. The closest parallels seem to occur in south-central Europe (Valoch 1968) where the Middle Paleolithic industries ap- proach the Denticulate and Charentian facies of France. At the Kulna cave (Level 7a) in Moravia (Jelinek 1970:479) there is an in- dustry associated with a hominid jaw which seems to be like Mousterian of Acheulean Tradition B where “quartzite implements of the Mousterian type prevail, however, here and there were also found Acheulean imple- ments as well as a few approaching the type of tools found in Upper Paleolithic cultures” (Jelinek 1966:701). At the Sipka cave in Moravia, Maska reported an early Wurm industry with a high frequency of denticu- late scrapers containing a large number of end scrapers, burins, and borers (Valoch 1968: 356). The apparently contemporary Szletian industries of Hungary and circum- Carpathian central Europe contain a good proportion of true blades but many more flake tools, some showing the Levallois tech- nique. Associated with these are “Upper
Paleolithic” forms such as dorsally re- touched end scrapers, carinated (Aurigna- cian) scrapers, simple burins and borers, and a large number of leaf-shaped points ranging from crude unifacial to fine bifacial retouch. At some Szletian sites (such as Lovas in Hungary) bone tools also occur (Valoch 1968:358). The latest of these manifesta- tions seem to be stratigraphically dated to a moist cool period around 38,000 B.P. In this area the Lower Aurignacian (Gravettian) is similar to the early Aurignacian of France but contains up to twenty-five percent of Mousterian forms such as side-scrapers and points. The upper stages of these industries (dated to 31,840L-250 B.P. at Willendorf 11) are lithically quite similar but contain great- er frequencies of split base and lozenge section bone points and awls (Valoch 1968:359). As Jelinek noted (1969:484) the human
burials at Predmosti are clearly E. Gravettian (e.g., red ocher burials, fired clay figures, mammoth ivory carving), but they are asso- ciated with “a stone industry in which nu- merous tools belong typologically to the traditions of the Mousterian, Acheulean, and even pebble tool cultures. This variability is reflected at the same time in the many Mousterian sites, both in Czechoslovakia and elsewhere in central and eastern Europe, that are regarded as atypical in containing indica- tions of Upper Paleolithic blade tool types.” From the Croatian site of Krapina, Gorjan- ovic-Kramberger (191 3) has illustrated plano-convex bifacially flaked points (Plate V, Figs. 11-13), piercers (Plate VI, Figs. 6-8), graves (Plate IX, Figs. 1-7; Plate X, Figs. 1-5), natural and backed-blade knives (Plate X, Figs. 7-10), basally thinned bifa- cially worked points (Plate VIII, Figs. 4, 5) along with a large number of discs, scrapers and flake tools. The site also yielded several bone tools among which were two odd split-base bone points (Gorjanovic- Kramberger 1913; Plate XIII, Figs. 1, 2).
According to Wobst (1970:455) “a good case can be made for in situ evolution from Middle to Upper Paleolithic” in Slovakia. He reasons that the increased percentage of

1164 AMERICAN ANTHROPOLOGIST [ 73,1971
Upper Paleolithic tools in late Mousterian and the decreasing frequencies of Middle Paleolithic tools in the early Upper Paleo- lithic preclude the possibility of a “sudden immigration.” The excellent evidence for a gradual change in activities, underlying the transition from Middle to Upper Paleolithic is based on the good preservation of inter- stadia1 open-air sites in numerous places.
Elsewhere in central Europe, the Repo- lust cave in Austria has a Middle Paleolithic industry which (on the basis of faunal evi- dence),may be as early as late Riss/Wurm. This industry is characterized by a high frequency of denticulates but also distinct end scrapers and oblique burins (Mottl 1951). From Thuringia in middle Germany there are several examples of a Middle Paleo- lithic industry containing Mousterian side scrapers and limaces, numerous unifacially and bifacially retouched points, end scrapers, burins, and borers (Ekhm-Blanke 1960). Valoch has noted that the Nieto- perzowa cave in Galicia contained a Middle Paleolithic industry with leaf-shaped points mostly on blades which is as early as 38,000 BP (1968:358). In northern Germany at the site of Salzgitter-Lebenstedt an industry characterized as Levalloisian Mousterian of Acheulean tradition contained bifacially re- touched points, “Clactonian flakes,” and a large number of bone and antler tools in- cluding “barbed” or split-base points. The site dates by radiocarbon to a slightly warmer period in early Wurm at 55,000 BP (Tode, Prenl, et al. 1953).
In his summary of the Mousterian of European Russia, Klein (1969) has indicated both the sophistication of the cultural adap- tations involved in the exploitation of this ecologically diverse area and the high degree of lithic variability which these Middle Pale- olithic industries exhibit. Sites such as Starosel’e Kiik-Koba and Molodova yield burins, end scrapers, and backed blades, although in low frequencies (Ibid.:261, Fig. 4). Klein also indicates the occurrence of some amount of worked -bone in these Mousterian assemblages (Ibid.:264).
Within central and eastern Europe the Szletian is sometimes recognized as an ex- ample of an industry typologically transi- tional from Middle to Upper Paleolithic (Valoch 1968:358-359), but even in other traditions most of the tools characteristic of the Upper Paleolithic are clearly present in lower frequencies in the Middle Paleolithic.
North Africa
In north Africa, numerous sites have pro- duced evidence for Middle Paleolithic Levalloiso-Mousterian industrial facies closely akin to those of France (McBurney 1960:129, 135; Bordes 1968:121-122). Many of these contain implements charac- teristic of the Aterian (equated with early Upper Paleolithic of Europe). The most thoroughly reported site in North Africa is the Haua Fteah of Cyrenacian Lybia. Here the levels deposited during early Wurm (Layers XXXV-XXIX) contain an industry typical of the local Levalloiso-Mousterian (called “evolved Hybrid-Mousterian,” McBumey 1967: 108-131). The succeeding levels are quite similar to this Levalloiso- Mousterian but include end scrapers on blades, several varieties of small retouched blades, flake awls, carinated scrapers, Chat- elperron knives, and an “unexpectedly high lamellar element. . .approaching a true blade industry” (McBurney 1967:113). McBumey goes on to add “as a whole, this assemblage is characterized by a curious mixture of evolved Levalloiso-Mousterian elements (especially in the technique of primary flaking) and traits which are frankly Upper Paleolithic in their affinities” (1967: 11 3). From the interface of Layers XXXIV/ XXXIII two Neandertal mandibles were re- covered and have been radiocarbon dated to 40,700*500 BP. Overlying these hominid remains are layers which seem to be rather normal Levalloiso-Mousterian (XXXII/ XXXI), and above these, a series of layers (XXVII/XXXI) deposited at the end of early Wurm and containing again a number of assemblages with “Aterian” and “Upper Paleolithic” elements. The conclusion we

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1165
reach, opposed to that given by McBumey, would clearly seem to be that in North Africa many of the late Levalloiso- Mousterian assemblages contain large num- bers of tools characteristic of the local early Upper Paleolithic and of the later Emiran of the Near East (Garrod 1962).
The Near East
In the Levant, Perrot recognizes several “phyla” of Mousterian (1968:342), most of which are similar to the Levalloisian Mousterian facies of France but which have a relatively high frequency of blades, knives, gravers, and burins. Most common, however, is the Levalloiso-Mousterian (which is seen in the early Wurm equivalent in the Levant) which is characterized by a few denticu- late tools (1968:346-349). The early Levalloiso-Mousterian levels at many interior sites in the Jordan River-Mount Carmel area shows clear affinities to the coastal sites such as Rasel-kelb and Chekka where early Mousterian is correlated by beaches to the period just post-Riss/Wurm (Howell 1959:18-19; Perrot 1968:352-356). In the mountain wadi shelters these Mousterian deposits show industrial facies possibly re- flecting functional differences (Howell 1959:19; Binford and Binford 1969). At Skhul the hominid remains from Level B were associated with a Levalloiso-Mousterian containing a few burins (Howell 1959:20). Similar assemblages occur mixed within the four meters of Mousterian deposits above Level L (with associated hominid remains) at Quafzeh (Howell 1959:13, 20-21). Perrot notes that it is difficult to determine just when the change from this Middle Paleo- lithic to the Upper Paleolithic occurs. Numerous sites yield transitional industrial assemblages throughout the area. At Quafzeh (Level E) and Ksar Akil above a number of good Levalloiso-Mousterian layers are levels which contain a large proportion of elongated Mousterian points and side scrapers but which also contain blade tools and “Chatelperron knives’’ associated with Levallois cores, Mousterian disc cores and
prismatic blade co res (Perrot 1968: 354-355). The “type-fossil” of this horizon (Garrod 1962) is the Emirah point. Both Bordes and Perrot feel these points are similar to the thinned Levalloisian points found as early as Level D at Shanidar in a late Levalloiso-Mousterian context (Perrot 1968; Bordes 1968). At any rate, it is clear that at sites such as Quafzeh (Level E), Ksar Akil, Tabun (Level B), Emirah, and El Wad (Level F) an evolved Levalloiso-Mousterian with thinned points, blade tools, prismatic cores, and backed knives exists.
Following these components are a num- ber of levels with fewer of the “typical Mousterian” scrapers and points and with a higher frequency of gravers, retouched blades, and Chatelperron knives. The Emirah point is absent but burins (especially dihedral and busked burins), gravers on thick flakes, end-of-blade scrapers and carinated scrapers appear in great numbers “an- nouncing the following ‘Aurignacian’ which has an even greater increase in retouched blades and points, burins, and gravers, and split-base bone points” (Perrot 1968:355). South of Shanidar in the Khorrambad valley the Kunji cave yields a Mousterian as- semblage with Mousterian points and scrapers as well as blades but no evidence of Levalloisian technique (senso stricto). These levels are dated greater than 40,000 BP (Hole and Flannery 1968).
The earliest true Upper Paleolithic in- dustrial assemblages (Neuville’s stage 111) oc- cur at Quafzeh (Level D). b a r Akil (Levels from seven to twelve meters with the twelve- meter level dating at 28,000 BP), Erg et Akmar (Levels E and F), Jabrud Shelter I1 (Levels 6 and 7), and Shanidar (Level C, dated at 29,000-26,000 BP). These clearly postdate transitional hominid populations. In the Near East it Seems that the archaeo- logical evidence, even when based only upon lithic assemblages, will not support any view of a rapid replacement of Middle by Upper Paleolithic either in terms of the morpho- logical attributes of the tools themselves or their method of manufacture (cf. Binford 1968:707-708, 715). As Howell (1959:40)

1166 AMERICAN ANTHROPOLOGIST 173,1971
concluded in an intensive review of the area, “The first recognized stage (1) of the Upper Paleolithic of Southwestern Asia had its roots in the local Mousterian (of Levallois facies).”
Asia
From Shanidar east the Middle Paleolithic is poorly understood (Ivanova 1969). At the site of Dara-i-kure a rock shelter in Afghan- istan (Dupree, Lattman, and Davis 1970) Dupree (1970:492) describes a Middle Paleo- lithic assemblage with Levallois flakes, Mousterian points, large side scrapers, cleavers, flake hand axes, and tortoise cores, as well as flake blades and possible combina- tion tools such as burin-points and burin- end-scrapers. At Teshik-Tash in Uzbekistan a rather generalized Mousterian is associated with an adolescent burial (Okladnikov et al. 1949), while at Kiik-Koba in the Crimea a Mousterian industry contained Levallois flakes, disc cores, and bone points (Mongait 1961: 82). Mousterian assemblages have been found at numerous localities in China but are undated for the most part. A late site from the Ordos area which yielded several hominid cranial fragments contained an in- dustry which was reanalyzed by the Bordes and is described by F. Bordes (1968:130) as a “Levallois-Technique Mousterian, with blades well represented (31%), 27% scrapers, 16.6% denticulates, 28% implements of Upper Paleolithic type (end-scrapers, burins, borers, occasional backed knives flakes and truncated blades) a few poor hand axes.. . discs, bladelets . . . . The impression given is in fact that of a very evolved Mousterian in the process of transition to an Upper Paleolithic stage. . . .”
Archaeological Summary
This brief review of the archaeological sequences clearly indicates several important facts. First and most evident, is the con- tinuity which exists throughout the Old
World between Middle and Upper Paleolithic industries.
Given the presence of local transitional industries throughout the Old World, in- dicated by the data discussed, the interpreta- tion of “sudden replacement” anywhere in the Old World becomes both logically im- probable and increasingly difficult to verify.
An Upper Paleolithic assemblage directly overlying a Middle Paleolithic assemblage is a common occurrence throughout the Old World, and indicates the relative chrono- logical position of the two occupations. While the Middle Paleolithic is obviously not the same as the Upper Paleolithic, we have described numerous archaeological as- semblages which can be considered typo- logically transitional between the two on a morphological-functional basis. We have also tried to indicate the artificiality of the tradi- tional distinction between Middle and Upper Paleolithic archaeological assemblages, resting as it does upon arbitrary cut-off points in relative frequencies of particular types of stone tools present in both, or upon the purported presence or absence of worked bone tools.
In any area, demonstrating the total and rapid replacement of one tradition or archae- ological complex by another requires that no transitional industries exist locally. We have found overwhelming evidence for local tran- sitional industries everywhere Middle Paleo- lithic industries occur. While we have no doubt been guilty of overlooking numerous relevant publications, it is difficult to imagine what kinds of sources could be used to argue against the interpretation of gradual transition.
Finally, we would note that the Middle Paleolithic is characterized for the first time, by significant numbers of special purpose tools. Many of these-borers, gravers, burins, and numerous others-indicate that these functional types were not neglected until the Upper Paleolithic. In addition, a large number of characteristic types of bone tools so often diagnostic of the early Upper Paleo lithic facies, have a clear prototype in thc Middle Paleolithic.

Brose C Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS 1167
THE ORIGIN OF ANATOMICALLY MODERN HOMO SAPIENS
Morphological evidence, too, indicates the continuity between Neandertals and ana- tomically modem H. sapiens. This con- tinuity has been established for all Nean- dertal populations-even the “classic” Nean- dertals of glaciated Western Europe-by numerous authors in a series of publications spanning almost a century (Arambourg 1958; Brace 1962b, 1964, 1967c, 1968; Coon 1963; Fraipont and Lohest 1887; Gorjanovic-Kramberger 1906; Hrdlicka 1927, 1930; Jelinek 1969; Pouliancs 1969; Schwalbe 1901,1914; Tobias 1964; Verneau 1924; Weidenreich 1940, 1947; Weinert 1925, 1936, 1944; Yakimov 1969). We pro- pose that transitions in hominid morphology precede the established changes in hominid industries.
While Upper Paleolithic industries are always associated with anatomically modern H. sapiens, the converse is not necessarily true. Anatomically modern H. sapiens ap- parently arises from a Neandertal ancestor still associated with Middle Paleolithic in- dustries. Thus, the earliest anatomically modern H. sapiens should be found with Middle Paleolithic cultural material. The ear- liest clearly dated specimens of anatomically modern H. sapiens are associated with an Upper Paleolithic industry, and not a par- ticularly early one. However, there are two lines of evidence indicating the in situ evolu- tion of anatomically modern H. sapiens from Neandertal populations associated with Middle Paleolithic industries.
NEANDERTAL MORPHOLOGICAL VARIATION AND VARIABILITY
The first line of evidence stems from the variability within the Neandertal populations themselves. Neandertals, contrary to the opinion of many authors (Sergi 1958a; Coon 1963; Howell 1952, 1957; Breitinger 1955), had a high degree of both inter and intra- population variability (Brace 1962b). Wit- ness the extent of difference among the
so-called “classic” specimens from Le Moustier (Weinert 1925), La Ferrassie 1 (Hrdlicka 1930; Boule 1913), La Ferrassie 2 (Captain and Peyrony 1912a; Boule 1913; Heim 1968), and Monte Circeo (Sergi 1940), let alone among other Neandertals such as Mapa (Woo and Peng 1959), Broken Hill (Morant 1928), and Omo 2 (Leakey, Butzer, and Day 1969). Indeed, the normal vari- ability within Neandertal populations is so great that almost every time more than one specimen is found at a single site, “hybridi- zation” is the only explanation deemed suf- ficient by many to explain the extensive variation among the specimens. Such an explanation has been suggested for the material from Krapina (Gorjanovic- Kramberger 1910; Klaatsch 1923), Quafzeh (Thoma 1965:139), and Skhul (Hooton 1947: 336-338; Weckler 1954:1014-1015; Thoma 1958): three of the four Neandertal sites with the remains of more than ten individuals.
A brief perusal of the literature shows that at almost every site with more than one Neandertal specimen, whether “classic” Ne- andertals or “Neandertals” in the broader sense used here, an extensive amount of variation has been recognized by, among others, the authors who worked with the original material. In addition to those already discussed, skeletal variation which is at least as great as the variation found in small samples of extant populations, hominid or pongid, has been described at the following sites: La Ferrassie (Hrdlicka 1930; Brace 19628; Captain and Peyrony 1909, 1912a, 1912b, 1921; Boule 1913; Piveteau 1969; Heim 1968), Ehringsdorf (Behm- Blancke 1960; Jelinek 1969; Virchow 1920; Weidenreich 1928; Kleinschmidt 1931), Spy (Morant 1927a; Fraipont and Lohest 1887; Brace 1968), Broken Hill (Wells 1947), Arcy-Sur-Cure (Leroi-Gourhan 1958), Solo (von Koenigswald 1958; Weidenreich 1951) Saccopastore (Howell 1960), Shanidar (Stewart 1958, 1961, 1963), Jebel Irhound (Ennouchi 1962, 1968, 1969), Subaluk (Kadic, Bartucz, Hillebrand, and Suzbo 1933), and others.

1168 AMERICAN ANTHROPOLOGIST [ 73,1971
The idea that “classic” Neandertals were particularly lacking in variability has im- plied, to many, that they were under the influence of extremely strong selection. In the words of Coon (1963:509), “They are in fact so homogenous that a strong selective agency must have been pruning off deviant individuals.”
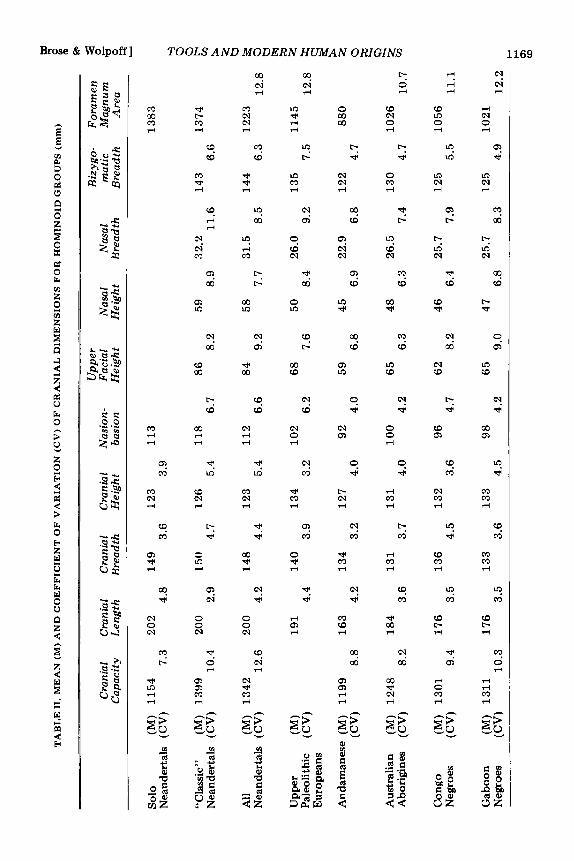
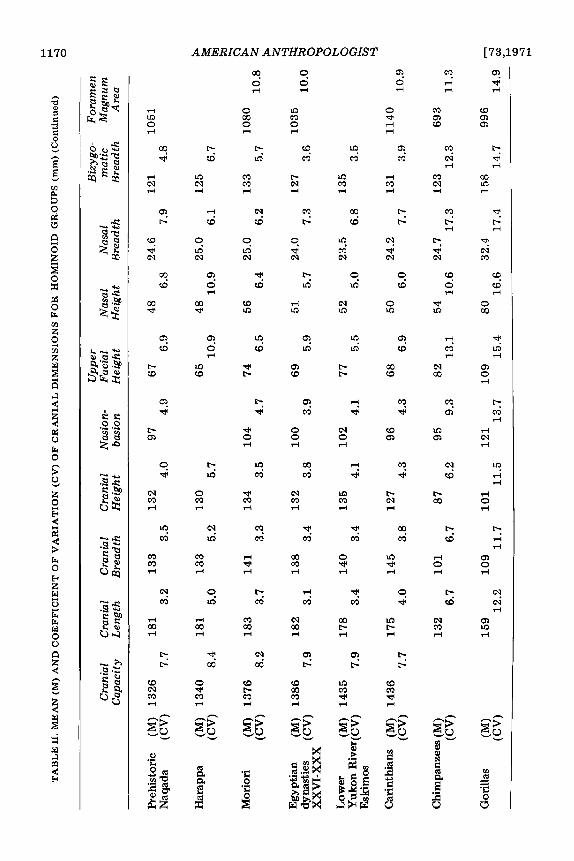
The evidence simply does not support this statement for either ‘‘classic’’ Neandertal specimens or all Neandertal specimens taken together. The coefficient of variation (CV), an index of the standard deviation divided by the mean, provides a convenient way of comparing variability between samples with different means. Coefficients of variation were calculated for the ten cranial measure- ments given in Table 11. A number of samples were utilized in this comparison of Neander- tals to chimpanzees, gorillas, and extant populations. The extant H. sapiens popula- tions include Andamanese (Sullivan 1921), Australian aborigines (Morant 1927b), Negroes from both Congo and Gaboon areas (Benington 1911), prehistoric Naquada (Morant 1924), a sample from Harappa in India (Gupta, Dutta, and Basu 1962),Moriori (Thomson 1916), a sample from Egyptian dynasties XXVI through XXX (Pearson and Davin 1924), Lower Yukon River Eskimos (Hrdlicka 1942), and Carinthians (Shapiro 1929).
We have sorted the Neandertal sample into “classic” and “non-classic” groups in as conservative a manner as possible. That is, we have only identified as “classic” those specimens to which all other authors would agree on the basis of geographic associations, stratigraphic position, and skeletal mor- phology (see Howell 1952:378-379). Dis- covering extensive variability in this re- stricted sample becomes all-the-more im- portant. Including a wider geographic or temporal range of specimens, or using individuals which workers in disagreement with our hypothesis would question, would increase the observed variation even further but at the same time cover up the meaning and importance of the variation-its range and its
form-in the restricted “classic” Neandertal sample.
Measurements for the Solo sample, a “tropical” Neandertal group, were taken from publications by Weidenreich (1951) and von Koenigswald (1958). The site may be as old as the Eemian interglacial (Howell 1967:489). The “classic” Neandertals uti- lized in these calculations (following Howell 1967), include Spy I and I1 (Morant 1927a; Fraiport and Lohest 1887), Gibraltar (Sollas 1908), La Quina 5 (Henri-Martin 1923), Le Moustier (Weinert 1925), Petralona (Kok- koros and Kannelis 1960; Poulianos 1966; Kannelis and Savas 1964), La Chapelle (Boule 1913; Morant 1927a), La Ferrassie 1 (Boule 1913; Coon 1963), Neandertal (Morant 1927a), and Monte Circeo (Sergi 1940). Other Neandertals, included with all of the above in the “total” sample, consist of both Wurm specimens from outside of Southern and Western Europe and pre-Wurm specimens from all areas. These are as fol- lows: Broken Hill (Morant 1928), Saldahna (Singer 1954), Omo 2 (Leakey, Butzer, and Day 1969), Djebel Irhound 1 (Ennouchi 1962) and 2 (Ennouchi 1968), Galilee (Morant 1927a), Shanidar I (Stewart 1958), Sala (Vleck 1965), Tabun 1 (McCown and Keith 1939), Saccopastore 1 (Sergi 1944) and 2 (Sergi 1948), Ehringsdorf H (Klein- Schmidt 1931), Fontechavade 2 (Vallois 1958), Ganovce (Vlcek 1955), Krapina crania C and E (Gorjanovic-Kramberger 1906) as well as D (Schaefer 1964), Djebel Quafzeh 6 (Coon 1963; Bode and Vallois 1957), Amud (Suzuki 1965 and 1968), and Skhul 4, 5, 6, and 9 (McCown and Keith 1939). While additional comparisons could have been made with Steinheim (Weinert 1936; Howell 1960) and Swanscomb (Morant 1938), both the morphological posi- tion (Brace 196213, 1964; Sergi 1958a; Stewart 1960; Weinert 1936) and the strati- graphic position (Howell 1960, 1967) of these specimens are clearly transitional be- tween H. erectus and the Neandertals. The Upper Paleolithic sample was taken largely from Morant (1925, 1930), Matiegka
m
TA
BL
E 1
1. M
EA
N (M) A
ND
CO
EF
FIC
IEN
T O
F V
AR
IAT
ION
(C
V)
OF
CR
AN
IAL
DIM
EN
SIO
NS
FO
Q H
OM
INO
ID G
RO
UP
S (
mm
) H m
~~
Upp
er
Biz
ygo-
F
oram
en
Cra
nial
C
rani
al
Cra
nial
C
rani
al
Nas
ion-
F
acia
l N
asal
N
asal
m
atic
M
agnu
m
Cap
acity
L
engt
h B
read
th
Hei
ght
basi
on
Hei
ght
Hei
ght
Bre
adth
B
read
th
Are
a
Solo
(M
) 11
54
202
14
9
12
3
11
3
Nea
nder
tals
(C
V)
7.3
4.8
3.6
3.9
1383
“Cla
ssic
” (M
) 13
99
200
150
126
118
86
59
32
.2
14
3
1374
N
eand
erta
ls
(CV
) 10
.4
2.9
4.7
5.4
6.7
8.2
8.9
11.6
6.
6
All
(M)
1342
20
0 1
48
1
23
1
12
8
4
58
31.5
14
4 12
23
Nea
nder
tals
(C
V)
12.6
4.
2 4.
4 5.
4 6.
6 9.
2 7.
7 8.
5 6.
3 12
.8
191
14
0
134
102
68
50
26.0
13
5 11
45
4.4
3.9
3.2
6.2
7.6
8.4
9.2
7.5
12.8
U
pper
. (M
I
And
aman
ese
(M)
1199
1
63
13
4 12
7 9
2
59
45
22.9
12
2 88
0
Pale
ohth
ic
(CV
) E
urop
eans
(CV
) 8.
8 4.
2 3.
2 4.
0 4.
0 6.
8 6.
9 6.
8 4.
7
Aus
tral
ian
(M)
1248
18
4 1
31
131
10
0
65
48
26
.5
130
1026
A
bori
gine
s (C
V)
8.2
3.6
3.7
4.0
4.2
6.3
6.3
7.4
4.7
10.7
Con
go
(M)
1301
17
6 13
6 1
32
96
62
46
25
.7
125
1056
N
egro
es
(CV
) 9.
4 3.
5 4.
5 3.
6 4.
7 8.
2 6.
4 7.
9 5.
5 11.1
3 B
0
b $
Gab
oon
(M)
1311
17
6 1
33
1
33
9
8
65
47
25.7
1
25
10
21
Neg
roes
(C
V)
10.3
3.
5 3.
6 4.
5 4.
2 9.
0 6.
8 8.
3 4.
9 12
.2
CL
CL
4 0
TA
BL
E 1
1. M
EA
N (
M)
AN
D C
OE
FF
ICIE
NT
OF
VA
RIA
TIO
N (
CV
) O
F C
RA
NIA
L D
IME
NSI
ON
S F
OR
HO
MIN
OID
GR
OU
PS
(mm
) (C
onti
nued
)
upp
er
Biz
ygo-
F
oram
en
Cra
nial
C
rani
al
Cra
nial
C
rani
al
Nas
ion-
F
acia
l N
asal
N
asal
m
atic
M
agnu
m
Cap
acit
y L
engt
h B
read
th
Hei
ght
basi
on
Hei
ght
Hei
ght
Bre
adth
B
read
th
Are
a
Pre
hist
oric
(M
) 1326
181
133
132
97
67
48
24.6
121
1051
Naq
ada
(CV
) 7.7
3.2
3.5
4.0
4.9
6.9
6.8
7.9
4.8
Har
appa
(M
) 1340
181
133
130
65
48
25.0
125
b i?
(CV
) 8.4
5.0
5.2
5.7
10.9
10.9
6.1
6.7
s M
orio
ri
(M) 1376
183
141
134
104
74
56
25.0
133
1080
c 10.8
rr: $. 2
(CV
) 8.2
3.7
3.3
3.5
4.7
6.5
6.4
6.2
5.7
Y
Egy
ptia
n (M
) 1386
182
138
132
100
69
51
24.0
127
1035
XX
VI-
XX
X
dyna
stie
s (c
v)
7.9
3.1
3.4
3.8
3.9
5.9
5.7
7.3
3.6
10.0
a $ L
ower
(M
) 1435
178
140
135
102
77
52
23.5
135
0
Yuk
on R
iver
(CV
) 7.9
3.4
3.4
4.1
4.1
5.5
5.0
6.8
3.5
Q
Esk
imos
2 Y
C
arin
thia
ns
(M) 1436
175
145
127
96
68
50
24.2
131
1140
7.7
4.0
3.8
4.3
4.3
6.9
6.0
7.7
3.9
10.9
(CV)
Chi
mpa
nzee
s (M
) 132
101
87
95
82
54
24.7
123
69 3
(CV
) 6.7
6.7
6.2
9.3
13.1
10.6
17.3
12.3
11.3
Gor
illas
(M
) 159
109
101
121
109
80
32.4
158
996
- 12.2
11.7
11.5
13.7
15.4
16.6
17.4
14.7
14.9
2 +
(D 4
w
(CV
)

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1171
(1938), Maska (1889), Werth (1928), Jelenik (1964) and von Bonin (1935).
Finally, the pongid samples were mea- sured by Wolpoff. Specimens were obtained at the Field Museum in Chicago, the Ameri- can Museum in New York, and from the Hamann-Todd collection at Case Western Reserve University and the Cleveland Mu- seum of Natural Science. Length, breadth, and height measurements of the vault, disre- garding crests and tori, were taken using techniques and measuring points suggested by Randall (1943-1944). In addition, nasion was defined on these primates according to Randall (1943-44). These procedures were followed in order to make the pongid- hominid measurements as comparable as possible.
In six of the nine comparable measure- ments the “classic” Neandertals are either as variable or more variable than the total Nean- dertal sample, although the latter includes a far wider geographic area, and spans a greater time. Such extensive variation in a sample all authors recognize as “classic” renders “hybridization” explanations of “non- classic” Neandertal samples unnecessary. I t denies support to the idea that “classic” Neandertals were under the influence of strong selective pressures which ultimately lead to their extinction. Finally, the ex- tensive variation evinced by the “classic” sample specifically suggests climatic adapta- tion. This sample, spread across the climatic extremes of partially glaciated Europe, shows the greatest variation, and the greatest difference in variation compared with all Neandertals, in nasal breadth-a feature closely tied with respiratory adaptations to cold climates (Wolpoff 1968).
The “classic” Neandertals are more vari- able in most features than any extant group. The most similar sample of anatomically modem H. sapiens is, as one could well expect, the Upper Paleolithic Europeans (Schlaginhaufen 1946). Concomitantly, co- efficients of variation for the latter sample and all Neandertals are almost identical. Variation in the fossil hominid samples is not, however, unusual for primates. Com-
parison with the two pongid samples evinces greater variation among the pongids. While high coefficients of variation for gorillas may result from the extensive sexual dimorphism, such an explanation cannot account for the nearly as high coefficients shown for chim- panzees. The same relation occurs for cranial capacities. The coefficient of variation for thirty Neandertals is 12.6. This compares favorably with the coefficient for 200 ana- tomically modern H. sapiens specimens of 11.7 (Ashton and Spence 1958) and of 14.1 for twelve specimens of H. erectus (Wolpoff 1969). Further, 144 chimpanzees have a cranial capacity coefficient of variation of 6.8, while the coefficient for 653 gorillas is 13.6 (Ashton and Spence 1958).
With one exception, the average dimen- sions for the “classic” Neandertals are al- most identical with both those for the Solo sample and for the total sample. The excep- tion is cranial base length or nasion-basion diameter. The significantly greater diameters for the “classic” specimens is a direct mea- sure of the total facial prognathism separa- ting nasal passages from brain which appears to be part of the Neandertal cold adaptation (Coon 1964). While all Neandertals have a longer cranial base than do any group of anatomically modern H. sapiens the “clas- sic” sample dimensions are far greater yet.
Finally, Table I1 provides an excellent indication that the Upper Paleolithic Europeans are metrically transitional between succeeding Neandertal populations and ex- tant Europeans.
After sorting out “classic” Neandertals on extremely conservative grounds, using among other things morphological criteria for specimens lacking adequate provenience such as Le Moustier, Neandertal, and Gibral- tar, (Howell 1952:379), the same morpho- logical criteria applied to all of the speci- mens available at this’ time indicate the largely arbitrary and anatomically un- justified basis of the initial sorting. Use of the t test evinces no significant differences between the “classic” and the “non-classic” samples for any of the measurements in Table I1 except nasion-basion; the only

1172 AMERICAN ANTHROPOLOGIST [73,1971
consistent differences occur in features reflecting climatic adaptation (Coon 1963). All of the differences between “classic” and “non-classic” groups discussed here, as well as differences discussed in the following sections, are far less than those which regularly occur among extant groups living side-by-side (see Hrdlicka 1928, 1942; DeVilliers 1968; Benington 1911; Larnach and Macintosh 1966). These extensive metric, variational, and morphologic similarities and the complete overlap of ranges occur between a group very restricted in time and space, thought by some to represent a genetic isolate and a much larger group representing a greater time span and occupying all of the Old World. If under these conditions the similarities do not indicate the phylogenic unity of the two groups, what further evidence could possibly be required to demonstrate this point?
The pattern of metric variation overlap is evinced by other morphological features presumably associated with the “classic” Neandertals. This is made particularly clear when one reviews the evidence underlying a compiled list of distinguishing “classic” Neandertal features such as that presented by Boule and Vallois (1957:251-2). The “simian” characters claimed for the vertebral column and limb bones will be ignored in the light of work by numerous authors demonstrating the completely modern anatomy of “classic” Neandertal post-cranial skeletons (Schwalbe 1914; Patte 1955; Straus and Cave 1957). Extensive lists of presumably distinctive “classic” Neandertal cranial and facial features have been prepared by a number of authors (Howell 1951, 1952, 1957; Boule and Vallois 1957; Hooton 1947). We will show both that some “classic” specimens as well as some “non-classic” specimens have almost all of these features. On the other hand, many o the r specimens without these characteristics can be found in both groups. The latter fact is of greater potential importance, because no feature distinguishes all “non-classic” or all “classic” specimens from anatomically modern H. sapiens.
Conversely, all of the presumed “ distinguishing characteristics” for anatomically modern H. sapiens occur, with regular frequency, in both “classic” and “non-classic” Neandertals. For instance, La Chapelle (Boule 1913), Ehringsdorf H (Kleinschmidt 1931), Cova Negra (de Lumley 1970), and Amud (Suzuki 1965, 1968) are large headed, while La Ferrassie 2 (Hrdlicka 1930; Heim 1968), Petralona (Kannelis and Savas 1964), Ganovce (Vlcek 1955), La Chaise (Krukoff 1970), Gibraltar (Sollas 1908), and Tabun (McCown and Keith 1939) are not. The forehead is high in Petralona (Jelinek 1969), La Ferrassie 1 (Coon 1963), Djebel Irhound 1 (Ennouchi 1962), Krapina E (Schaefer 1964) and Shanidar 1 (Stewart 1958), but is quite low in the Neandertal calvarium (Hrdlicka 1930), Broken Hill (Morant 1928), Sacco- pastore 1 (Sergi 1944), Krapina D (Gorjanovic-Kramberger 1906), and Tabun 1 (McCown and Keith 1939). Parietal bones of Krapina E (Schaefer 1964), Ehringsdorf H (Kleinschmidt 1931) and C (Behm-Blancke 1960), La Quina 5 (Henri-Martin 1923), and Saldanha (Singer 1954) are flat and evenly rounded, while Omo 2 (Leakey, Butzer, and Day 1969), Krapina K (Jelinek 1969), Bro- ken Hill (Morant 1928), Ehringsdorf B and D (Behm-Blancke 1960), and Spy 2 (Hrdlicka 1930) have high, rooflike parietals with a pronounced boss. Heinz (1967) has shown that Neandertal parietal heights over- lap extensively with those of anatomically modern H. sapiens. Thus, while La Ferrassie 1 (Boule 1913), Ganovce (Vlcek 1955), Fontechevade 2 (Vallois 1958), Sacco- pastore 1 (Sergi 1944), and Amud (Suzuki 1965, 1968) show rounded contours in norma occipitalis. Spy 2 (Hrdlicka 1930), Le Moustier (Weinert 1925), Ehringsdorf D (Behm-Blancke 1960), Broken Hill (Morant 1928), and Djebel Irhound 1 (Ennouchi 1962) have parallel sided contours.
The supraorbital torus is heavy with glabellar and lateral elements merged and the supraorbital sulcus obliterated in Mapa (Woo and Peng 1959), Broken Hill (Morant 1928), Neandertal (Boule 1913; Schwalbe

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1173
1914), Krapina D (Gorjanovic-Kramberger 1906), Galilee (Kurth 1965), Saccopastore 2 (Sergi 1948), Saldanha (Singer 1954), Mount Circeo (Sergi 1940), and Ehringsdorf H (Weidenreich 1928). It is lightly developed in Le Moustier (Weinert 1925), Gibraltar (Sollas 1908), Amud (Suzuki 1965, 1968), Krapina C (Gorjanovic-Kramberger 1906), Omo 2 (Leakey, Butzer, and Day 1969), and Djebel Irhound 2 (Ennouchi 1968). In ad- dition, both Sala (Vlcek 1965) and Galilee (McCown and Keith 1939) have a sulcus separating glabellar and lateral portions. The occipital area is low, flattened, heavily mus- cled, and clearly displays the bunned condi- tion in La Quina 5 (Henri-Martin 1923), Ganovce (Vlcek 1955), and the Spy crania (Hrdlicka 1930). Gibraltar (Sollas 1908) is bunned but lightly muscled. In other speci- mens, such as Broken Hill (Morant 1930), Amud (Suzuki 1965), Omo 2 (Leakey, Butzer, and Day 1969), and Djebel Irhound 2 (Ennouchi 1968), the bunning is not as apparent, although vertical compression of the occipital region is retained. Saldanha (Singer 1954) has a clear occipital torus but no bunning. Finally the occipital region of the cranium is not at all bunned, but rather is small and rounded with light muscle attachments in Le Moustier (Weinert 1925), Saccopastore 1 (Sergi 1944), Tabun 1 (McCown and Keith 1939), and Djebel Irhound 1 (Ennouchi 1962). The mastoid process is very small in La Quina 10 and 27 (Vallois 1969), Djebel Irhound 2 (Ennouchi 1968), Saccopastore 2 (Sergi 1948), Spy 1 and 2 (Hrdlicka 1930), Petralona (Poulianos 1966), and Broken Hill (Morant 1928). However, in Saccopastore 1 (Sergi 1944), Amud (Suzuki 1965, 1968), La Quina 5 (Henri-Martin 1923), Ehringsdorf H (Weidenreich 1928), and Djebel Irhound 1 (Ennouchi 1962) the process is quite large. According to data recently published by Vallois (1969:396 and Table 7) mastoid process size for La Chapelle and La Ferrassie 1 fit well within the range of variation for modern man. There is very little, if any, mid-facial prognathism in La Ferrassie 1 (Brace 1962b), Djebel Irhound 1 (Ennouchi
1962), and Broken Hill (Morant 1928), but a significant amount of prognathism in the nasal-maxillary region of Le Moustier (Weinert 1925), Petralona (Kokkoros and Kannelis 1960), and Tabun 1 (McCown and Keith 1939). The amount of mid-facial prognathism is sufficient to obliterate the canine fossa in most Neandertals. However, there is a definite canine fossa in Djebel Irhound 1 (Ennouchi 1962), Krapina C (Gorjanovic-Kramberger 1906), Kulna (Jeli- nek 1969), and to a lesser extent Broken Hill (Hrdlicka 1930).
The face is relatively long compared with the calvarium in Petralona (Kokkoros and Kannelis 1960; Kannelis and Savas 1964), Saccopastore 1 (Sergi 1944), and La Ferras- sie 1 (Coon 1963), but it is significantly smaller in Le Moustier (Weinert 1925), Gibraltar (Sollas 1908), and Amud (Suzuki 1968). As with all of the other features, the claim of “large rounded orbits” is neither consistent within nor distinctive for the “classic” Neandertals. Orbit shape can be expressed in terms of the orbit index (height/breadth). For five “classic” Neander- tals (Gibraltar, La Chapelle, La Ferrassie, Monte Circeo, and Le Moustier), the average index is 84, ranging from 75 to 100. Eight “non-classic” specimens average 85, and range from 76 to 100. As a size measure, the orbit areas of these two samples are, respec- tively, 1685 mm2 and 1686 mm’.
Shelving of the maxilla into the malar occurs in Saccopastore I (Sergi 1944), La Ferrassie (Boule 1913), and Shanidar I (Stewart 1958), but is completely absent (a 90 degree angle) occurs in Djebel Irhound I (Ennouchi 1962), Broken Hill (Morant 1928), and La Quina H5 (Henri-Martin 1923). With teeth, La Chapelle is inter- mediate.
There is a definite mental eminence in the mandibles of La Ferrassie 1 (Boule and Vallois 1957), La Quina 9 (Piveteau 1964), Monte Circeo 3 (Sergi 1958b), Tabun 2 (McCown and Keith 1939), Amud (Suzuki 1968), the Djebel Irhound juvenile (Ennouchi 1969), Shanidar 1 and 2 (Stewart 1958, 1961), and Sipka (Kadic, Bartucz,

1174 AMERICAN ANTHROPOLOGIST [73,1971
Hillebrand and Szabo 1933). This region is receding in Arcy-sur-Cure 2 (Leroi-Gourhan 1958), Regardou (Piveteau 1964), Le Moustier (Weinert 1925), Tabun 1 (McCown and Keith 1939), La Quina 5 (Henri-Martin 1923), Ehringsdorf adult (Virchow 1920), Krapina H (Gorjanovic-Kramberger 1906) and Spy 1 (Hrdlicka 1930). The teeth have taurodont pulp cavities in the Krapina jaws (Gorjanovic-Kramberger 1907), Ochoz (Jelinek 1969), Spy 2 (Hrdlicka 1930).and Amud (Suzuki 1965). However, the pulp cavities are non-taurodont in the mandible from Abri Bourgeouis-Delaunay (Debenath and Piveteau 1969), Dire Dawa (Vallois 1951), and Tabun 2 (McCown and Keith 1939).
One could continue this demonstration indefinitely, but there seems little point in doing so. Two conclusions are apparent. First, the distinction long maintained be- tween “classic” and “non-classic” Neander- tals is without substantive basis. The few features which distinguish the group means (although not the individual specimens) show a tendency in the Western European Wurm group for heavier bunning in the occipital region and more extensive mid- facial prognathism. The trends do not have an elaborate basis, and are slight enough so that statistical significance usually cannot be verified. Second, it is quite apparent that Neandertals overlap with anatomically modern H. sapiens in almost every morpho- logical feature, as well as almost every metric one.
The form, as well as the range, of variation within Neandertal populations clearly indicates their ancestral relationship to the succeeding populations of anatomical- ly modern H. supiens (Brace 1964) found occupying the same caves and hunting the Same game at a later time (Hrdlicka 1927, 1930). Average Neandertal morphological and metric parameters are not always iden- tical with those of modern man: Neandertals are not, after all, anatomically modern H. supiens. On the other hand, there is ex- tensive overlap between the range of almost every Neandertal characteristic and the range
of the corresponding characteristic in modern man. It is not surprising that the group of anatomically modern H. supiens represented in Table I1 showing the greatest metric similarity to the “classic” Neandertals are the Upper Paleolithic Europeans!
The close relationship of Neandertals and succeeding anatomically modern H. supiens populations can be seen on an individual basis, in comparisons of La Ferrassie 1 with Predmost 3, let alone in the often discussed comparison of Skhul 5 with Predmost 3 (Brace 1967a, 1968). Predmost 3, the object of these comparisons, is gracile compared with some specimens associated with Upper Paleolithic industries such as Brux, Pod- kumok, and Brunn 1 (Weinert 1944:153-161). We believe that this simi- larity is the result of the in situ evolution of European Neandertals (see Jelinek 1969). In this respect, the hominids and the industries display the same relationship.
Finally, the clearly ancestral position of Neandertals is also evinced by the morpholo- gy, and postulated ontogeny, of Neandertal adolescents and children (Vlcek1964; Fenart 1969; Carbonell 1965; Senyurek 1959; Piveteau, de Lumley, and Mme de Lumley 1963; Piveteau 1969; Thoma 1963). In a recent study of Neandertal characteristics in the ontogeny of anatomically modern H. supiens Vlcek concludes “the development and presence of some morphological features typical for a Neandertal skeleton can be established on the skeleton of present man in his ontogenetic development” (1964:81).
These data suggest a re-examination of the conclusions based on Morant’s metric analysis (1927a:374-375). First, Neandertals (“classic” or otherwise) are not “remarkably homogeneous.” This conclusion is adequate- ly demonstrated in Table 11. If anything, they are remarkably variable. Second, the distinct hiatus reported between Neandertak and modern groups is no longer as distinct as first believed. While differences are demon- strable with modern population averages, numerous individuals overlap. In Europe, the Upper Paleolithic populations are clearly transitional (Jelinek 1969; Schwalbe 1906).

Brose & Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS 1175
Third, while no living group seems more closely related to Neandertals than any other, Upper Paleolithic Europeans are clearly transitional between European Nean- dertals and living European populations in both metric and morphological features. The final point is not only apparent in the distributions discussed here but has been convincingly demonstrated by Jelinek (1969), Maska (1889), Mateigka (1938), Jelinek, Palisek, and Valoch (1959), Brace (1964), Coon (1963), Schwalbe (1906), and others.
TRANSITIONAL SPECIMENS: THEIR IDENTIFICATION AND
ARCHAEOLOGICAL ASSOCIATIONS
The second line of evidence indicating worldwide evolution of Neandertals into anatomically modern H. sapiens stems from the cultural associations of the truly tran- sitional specimens.
Transitional specimens can best be seen in contrast to a general picture of the ways in which Neandertals differ from extant popu- lations. While there is overlap in almost every feature, differences in averages can still be found.
Most distinctive Neandertal character- istics were maintained as the result of selection for technological, and sometimes also climatic adaptation. Indeed, it was the appearance of the greatest climatic extremes
in sub-glacial Western Europe which resulted in both increased con tinen t-wide variability and highest frequencies of characteristics associated with the so-called “classic” Neandertals.
Dental Adaptation
The most important features dis- tinguishing Neandertal dentitions from those of extant modem groups are metric, rather than morphological (Dahlberg 1963). The major differences occur in the incisors and canines: the anterior teeth (Brace 1962a, 1964, 1968; Wolpoff 1970). The posterior dentition is not particularly distinctive. Table I11 shows the summed areas of the posterior teeth, mandibular and maxillary, for a number of hominid taxa and extant groups. The table was prepared by summing the individual averages for specimens in each group. Data for the fossil groups was taken from Wolpoff (1971), as were measurements for “Australoids,” “Caucasoids,” New Britain Islanders, and Dickson Mound Indians. The latter two were measured by Wolpoff. The Japanese data was published by Miyabara (1916), Lapp data by Selmer-Olsen (1949), and Teso data by Barnes (1969). It is apparent that the Neandertal group average falls within the range of variation of extant group averages with respect to summed posterior areas.
TABLE 111. S U M M E D AREAS O F POSTERIOR TOOTH ROW FOR MAXILLA AND MANDIBLE (rnm’)
Maxilla Mandible
Australopithecines 935 906 H. erectus 630 638 Neandertals 543 533 Anatomically modern H. sapiens 490 487
“ Australoid” 581 539 New Britain 537 520 Dickson Mound Indian 495 495 Teso 48 3 482 “Caucasoid” 466 457 Japanese 449 449 Lapps 388 391

1176 AMERICAN ANTHROPOLOGIST [ 73,1971
The primary function of the anterior teeth is in gripping, holding, exerting torsion, and other manipulations. These uses have been established for extant groups (Campbell 1925, 1939; Van Reenen 1966; Brothwell 1959, 1963; Barnes 1969; Leigh 1928, 1937; Turner and Cadien 1969; Brace and Molnar 1967; Waugh 1937; Taylor 1963; Bailit, DeWitt, and Leigh 1968; Gessain 1959; Noble 1926). Other uses observed include such diverse functions as leather treating (Pedersen 1938), pulling (Noble 1926), fashioning thong, reed and thread (Dahlberg 1963), straightening wooden shafts (Sollas 1924), and prying off rusted gasoline drum covers (de Poncine 1941). Indications of similar, although more extensive, use of the anterior dentition is characteristic of Neandertals. For instance, Dahlberg (1963) shows extensive differential incisor wear for the right maxillary teeth of La Ferrassie 1. The right side is considerably more worn than the left resulting in an uneven and undulating occlusal plane. Brace (1962a:347-348) shows a similar pattern of wear in Shanidar 1, Ternifine 3 (an H. erectus specimen), and Krapina J. Wear of this nature clearly could not result from mastica- tion alone (Brace and Molnar 1967).
The anterior teeth of Neandertals were apparently important tools in the manipula- tion of the environment. As such, they required robust roots and supporting struc- tures. These features are characteristic of Neandertal jaws and faces. The anterior displacement and robustness of the maxilla in part causes the region in the vicinity of the canine fossa to become convex rather than concave, thus eliminating the fossa. In addition, these features are characteristic of H. erectus faces. These also exhibit robust anterior dentitions.
Tables IV and V give data for averaged individual mandibular (Table IV) and maxil- lary (Table V) anterior summed tooth lengths (L), breadths (B), and areas (L*B). Data for Illinois Indians and Sub-Saharan Africans have been published by Wolpoff (1971). In addition to average values, the ranges, sample sizes (N), and coefficients of
variation (CV) are listed. Because of the presence of small sample sizes t tests were calculated to determine the significance of differences between H. erectus and Neander- tals, as well as between Neandertals and anatomically modern H. sapiens. Comparing H. erectus with Neandertals at the five percent significance level, mandibular summed length is significantly greater in the former. All other dimensional comparisons for both jaws reveal no significant dif- ference. On the one percent level no com- parison of H. erectus with Neandertal reveals a significant difference. Comparing the Neandertals with anatomically modern H. sapiens, on the other hand, there is a significant difference at the five percent level for every dimension in both jaws. At the one percent level, the length comparisons for both jaws are not significant, but all other comparisons are. It appears that while the anterior dentitions of Neandertals and H. erectus are not statistically distinguishable for length, breadth, and area (for all intents and purposes), there is a very significant dimensional decrease for anatomically mod- ern H. sapiens. The breadth, and subsequent- ly area, decreases are slightly more sig- nificant. Therefore, the Middle Pleistocene anterior dentition is demonstrably larger than that of anatomically modern H. sapiens, with particular emphasis on in- creased breadth. The size difference has both statistical and morphological significance.
We feel that the large Neandertal anterior teeth are a specific adaptation to the type of extensive use implied by the pattern of anterior occlusion seen in Neandertal jaws, as well as the general muscularity evinced by Neandertal skeletal rugosity. This implica- tion is substantiated by observation of tooth use in non-agricultural peoples. The in- creased breadth is an effective means of structural reinforcement. Indeed even slight- ly worn Neandertal incisors tend to be almost square.
Climatic Adaptation
Climate must also be considered in the analysis of Neandertal morphology, for the
TA
BL
E I
V.
AV
ER
AG
ED
MA
ND
IBU
LA
R A
NT
ER
IOR
TO
OT
H R
OW
SU
MM
ED
LE
NG
TH
S (L
), B
RE
AD
TH
S (
B).
AN
D A
RE
AS
(L
*B).
N
IS T
HE
SA
MP
LE
SIZ
E A
ND
CV
TH
E C
OE
FF
ICIE
NT
OF
VA
RIA
TIO
N (
mm
)
L(M
-D)
B(B
-L)
L *
B
Mea
n R
ange
N
C
V
Mea
n R
ange
N
C
V
Mea
n R
ange
N
C
V
Aus
tral
opith
ecin
es
H. e
rect
us
Nea
nder
tals
A
nato
mic
ally
mod
ern
H. s
apie
ns
“ A
ustr
aloi
d”
Sub-
Saha
ran
Afr
ican
Il
linoi
s In
dian
D
icks
on M
ound
Ind
ian
New
Bri
tain
“C
auca
soid
”
20.3
21
.1
20.0
19.2
21.1
19
.9
19.5
19
.4
19.4
17
.4
17.2
-23.
0 18
.2-2
3.8
17.5
-22.
8
14.2
-22.
4
20.0
-21.
8 16
.5-2
2.4
18.6
-20.
5 17
.4-2
2.2
17.8
-21.
2 14
.2-2
1.3
14
7.
8 23
.0
12
7.6
23.8
31
6.
7 24
.3
18
0
7.8
20.4
6 3.
0 22
.3
25
7.5
20.9
1
0
2.5
20.4
8
5
5.2
20.0
25
4.
6 20
.6
28
11.4
21
.0
20.0
-26.
5 21
.4-2
7.8
21.4
-28.
0
14.5
-25.
0
21.0
-24.
3 14
.5-2
4.2
18.9
-22.
7 17
.1-2
3.0
18.8
-23.
9 18
.4-2
5.0
13
8.
6 1
60
1
2
7.5
170
29
6.8
166
170
7.8
13
3
4 6.
8 16
0 21
11
.3
14
1
9 7.
0 13
4 84
6.
6 13
2 24
6.
0 13
4 25
7.
8 12
4
129.
6-19
7.1
145.
6-20
1.8
130.
3-20
0.8
82.0
-178
.0
148.
0-17
6.0
82.0
-172
.0
124.
0-14
9.0
101.
0-15
8.0
114.
0-15
3.0
94.0
-178
.0
13
13
.3
12
10
.0
29
9.9
167
12.9
4 7.
7 21
17
.4
9 6.
6 84
9.
7 23
7.
8 25
18
.3
TA
BL
E V
. A
VE
RA
GE
D M
AX
ILL
AR
Y A
NT
ER
IOR
TO
OT
H R
OW
SU
MM
ED
LE
NG
TH
S (L), B
RE
AD
TH
S (B
), A
ND
AR
EA
S (
L*B
).
N IS
TH
E S
AM
PL
E S
IZE
AN
D C
V T
HE
CO
EF
FIC
IEN
T O
F V
AR
IAT
ION
(m
m)
L(M
-D)
B(B
-L)
L *
B
Mea
n R
ange
N
C
V
Mea
n R
ange
N
C
V
Mea
n R
ange
N
C
V
Aus
tral
opith
ecin
es
26.3
22
.7-2
9.0
10
7.
1 24
.7
21.4
-28.
6 20
8.
1 21
6 18
3.4-
238.
8 8
9.1
H. e
rect
us
28.0
24
.2-2
9.5
4 8.
3 27
.6
25.7
-32.
6 4
10.5
26
0 20
8.7-
320.
2 4
14.3
N
eand
erta
ls
26.1
24
.0-3
0.0
14
7.
9 25
.9
22.9
-29.
2 1
4
6.3
226
196.
8-26
8.2
14
11
.5
H. S
apie
ns
24.5
18
.7-2
9.4
164
7.5
23.0
18
.8-3
0.3
160
7.7
190
131.
0-25
8.0
16
0
13.0
Sub-
Saha
ran
Afr
ican
I 25
.8
21.0
-29.
4 1
4
9.7
23.8
18
.8-2
6.7
14
10.3
20
7 13
1.0-
250.
0 14
17
.7
New
Bri
tain
24
.8
23.2
-26.
5 22
4.
3 23
.0
21.6
-25.
9 20
4.
5 19
2 16
8.0-
228.
0 20
7.
1 “ A
ustr
aloi
d”
24.4
21
.8-2
6.9
8 8.
0 25
.2
19.8
-30.
3 8
15.2
20
8 14
5.0-
258.
0 8
20
.1
Illin
ois
Indi
an
24.2
20
.9-2
5.2
8
5.9
22.4
20
.0-2
3.8
8 5.
5 18
2 14
6.0-
195.
0 8
9.0
“Cau
caso
id”
22.7
18
.7-2
7.4
27
10.7
23
.3
20.8
-28.
2 25
8.
7 17
8 13
1.0-
258.
0 25
18
.7
Ana
tom
ical
ly m
oder
n
Dic
kson
Mou
ndIn
dian
s 24
.9
21.9
-27.
6 8
5
4.8
22.7
19
.1-2
5.7
85
5.4
190
140.
0-22
7.0
85
8.9
Ro
Y
Y
0
0 5 c r
-a
-a

1178 AMERICAN ANTHROPOLOGIST [ 73,1971
so-called “classic” Neandertals were clearly adapted to a cold climate (Coon 1963, 1964). The anterior displacement of the entire face from nasion downward, so characteristic of Western European Neander- tals, is much reduced in specimens such as Broken Hill (Morant 1928), Djebel Irhound 1 (Ennouchi 1962), and others. This anterior displacement helps separate the nasal passage from the relatively low positioned brain (Coon 1963:533). Thus the brain is maxi- mally separated from inspired cold air. The necessity of warming such inspired air ac- counts for the great breadth of the Neander- tal nose (Wolpoff 1968). The resulting mas- sive face is balanced with a relatively long and low cranium, with expanded nuchal area for muscle attachments. As a result, the Wurm adapted Neandertal crania have the highest incidence of bunning and greatest occipital breadths. The increased facial prognathism completely eliminates the canine fossa in this group, a feature which regularly appears in the “non-classic” Nean- dertals. The resulting total morphological pattern for the “classic” group is one of cold adaptation built upon the morphology of the Neandertal grade of evolutionary de- velopment. The postcranial morphology, particularly as reflected in limb to trunk proportions (Patte 1955; Vallois 1958), similarly displays a cold-adapted pattern.
This total morphological pattern includes the frontal sinus complex. While the form and size of the Neandertal frontal sinus is directly intermediate between that of H. erectus and that of anatomically modern H. supiens (Vlcek 1967:188), the supraorbital torus is especially well developed in some of the Western European glacial Neandertals (Vlcek 1964). The size and form of this sinus is directly related to both the supra- orbital torus and the extent of mid-facial prognathism. The intermediacy of the sinus form helps corroborate the hypothesis of direct Neandertal ancestry, while the exces- sive development of the sinus, and hence of mid-facial prognathism, in glacial adapted Neandertals substantiates the hypothesis of climatic selection.
Most Neandertal features are not too different from those of H. erectus. Neander- tals differ from the latter mainly in the reduction of the posterior dentition and the expansion of the brain to modern size. Table 2 identifies many of the metric features characterizing the difference between Neandertal means and the means of extant groups. These data characterize Neandertal crania-“classic,” “tropical” (Solo), and all referred specimens taken together, as long, low, and relatively broad. The great cranial length and nasion-basion diameter can be taken as an indication of the degree of toal facial prognathism. The latter dimension is maximized in the Western European Nean- dertals. While Neandertal nasal height is great, the breadth of Neandertal noses is almost one third again as large as the maximum for extant groups. Total facial size for the Neandertals is large. It is only partially approached by the Eskimo sample. With the exception of cranial capacity and cranial height the average Neandertal di- mensions exceed those of all extant group averages. However, in cranial breadth, upper facial and nasal heights, and foramen mag- num area the Neandertal sample is closely approached by some of the extant group averages. The specific pattern of the most significant differences results from a com- bination of climatic and dental adaptations. The general massiveness of Neandertal cranial dimensions and rugosity of muscle attachments attests to the major function of individual body strength in Neandertal pat- tern of life.
This complex of features is maintained by both climatic selection and selection main- taining the large anterior dentition. Nean- dertal populations maintain these characteristics at their highest frequencies in Europe where climatic selection is greatest. A large number of specimens come from outside of Europe. These retain features resulting from the massive anterior dentition and supporting structures, such as a long and low calvarium, large face, and so on. However, the additional effects of maxillary protrusion, nasal size, and other cold-

Brose & Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS 1179
weather adaptations, are not present. As a result, many of the features common to both dental and climatic adaptation appear in a less extreme form. Therefore a distinctive morphology resulting from the interaction of both climatic and dental adaptation is associated with the so-called “classic” Neandertals from Western Europe, forming an adaptive complex. Rather than indicating the isolation of Western European Neandertal populations from populations in other areas, this continuity emphasizes the presence of consistent gene flow.
The Transitional Specimens
A “transitional” Neandertal population can be defined with respect to these fea- tures. There are a number of specimens representing such transitional populations. The fact that they all are associated with Mousterian type industries constitutes the second line of evidence indicating the in situ evolution of Neandertals into anatomically modern H. sapiens.
The first specimen representing a transi- tional population comes from the Kulna cave in Czechoslovakia. The maxilla, de- scribed by Jelinek (1966, 1969) has a canine fossa indicating a decrease in anterior maxil- lary expansion. According to Jelinek (1966:701) other progressive features in- clude the specific morphology of the canine and premolars, as well as the deep palate. These features are mixed with others more commonly distinctive of Neandertals. With PM4 not fully in occlusion, the specimen can be aged at fourteen to fifteen. Even for an adult the lower facial height is unusually large so that for an adolescent the very great size of the lower face is a Neandertal feature. Judging from the occlusal view (Jelinek
1969: Figure 4a), the anterior incisor sockets and the canine which is present indicate a great breadth for the anterior dentition. In addition, the maxilla has a prenasal fossa coupled with a very weakly developed anterior nasal spine.
The second transitional group comes from Arcysur-Cure in Western Europe. Here, along with the excellent sequence of industrial evolution from Middle to Upper Paleolithic (Movius 1969), there is a cor- responding sequence of hominid evolution (Leroi-Gourhan 1958). Hominids are found at almost every level, although most of the material recovered is dental. Teeth dis- covered in the highest Mousterian levels (12 to 15) are indistinguishable from those of anatomically modern H. sapiens. Some an- terior teeth are represented (Ibid.: 113).
According to Jelinek (1969:477,499), the Sipka mandible is dated in the Wurm 1/11 interstadial by both the geology and the fauna of the site. Because of this date and the association with a Mousterian industry (Maska 1882, 1886), one would expect the mandible to evince transitional charac- teristics, if the model proposed here is correct .
The dentition of the Sipka mandible suggests a transitional status (Vlcek 1969). Neandertals differ from anatomically modern H. sapiens in the size (breadth, area) of the anterior dentitions. Table VI gives anterior lower incisor breadth and area sums for fifteen European Neandertals. When an individual had both sides present, one was chosen at random. Four Skhul specimens are separately represented. The data for Sipka were published by Virchow (1882). Ref- erences for most of the other specimens are given in another publication (Wolpoff 1971). The three Krapina mandibles (E, H, I) were
TABLE VI. SUMMED BREADTHS A N D AREAS OF MANDIBULAR INCISORS
Breadth A rea( rn rn )
Mean Range N C V Mean Range N CV
European 15.5 14.2-16.8 1 5 4.6 96.4 85.5-112.6 12 8.3 Skhul 14.2 13.2-15.7 4 7.8 83.9 71.9-107.7 4 19.3
Neandertals
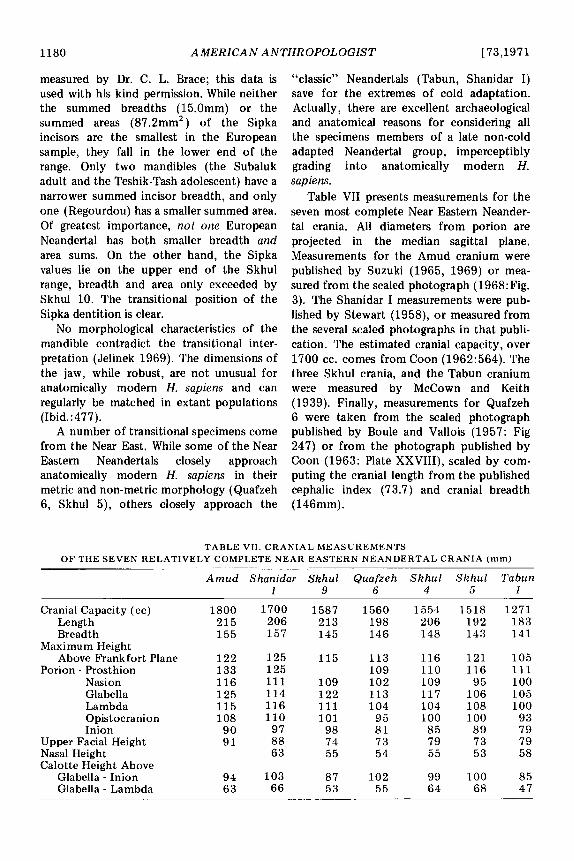
1180 AMERICAN ANTHROPOLOGIST [73,1971
measured by Dr. C. L. Brace; this data is used with his kind permission. While neither the summed breadths (15.0mm) or the summed areas (87.2mm’) of the Sipka incisors are the smallest in the European sample, they fall in the lower end of the range. Only two mandibles (the Subaluk adult and the Teshik-Tash adolescent) have a narrower summed incisor breadth, and only one (Regourdou) has a smaller summed area. Of greatest importance, no! one European Neandertal has both smaller breadth and area sums. On the other hand, the Sipka values lie on the upper end of the Skhul range, breadth and area only exceeded by Skhul 10. The transitional position of the Sipka dentition is clear.
No morphological characteristics of the mandible contradict the transitional inter- pretation (Jelinek 1969). The dimensions of the jaw, while robust, are not unusual for anatomically modern H. supiens and can regularly be matched in extant populations (Ibid. : 477).
A number of transitional specimens come from the Near East. While some of the Near Eastern Neandertals closely approach anatomically modern H. supiens in their metric and non-metric morphology (Quafzeh 6, Skhul 5), others closely approach the
“classic” Neandertals (Tabun, Shanidar I) save for the extremes of cold adaptation. Actually, there are excellent archaeological and anatomical reasons for considering all the specimens members of a late non-cold adapted Neandertal group, imperceptibly grading into anatomically modern H. supiens.
Table VII presents measurements for the seven most complete Near Eastern Neander- tal crania. All diameters from porion are projected in the median sagittal plane. Measurements for the Amud cranium were published by Suzuki (1965, 1969) or mea- sured from the scaled photograph (1968:Fig. 3). The Shanidar I measurements were pub- lished by Stewart (1958), or measured from the several scaled photographs in that publi- cation. The estimated cranial capacity, over 1700 cc. comes from Coon (1962:564). The three Skhul crania, and the Tabun cranium were measured by McCown and Keith (1939). Finally, measurements for Quafzeh 6 were taken from the scaled photograph published by Boule and Vallois (1957: Fig 247) or from the photograph published by Coon (1963: Plate XXVIII), scaled by com- puting the cranial length from the published cephalic index (73.7) and cranial breadth (146mm).
TABLE VII. CRANIAL MEASUREMENTS O F THE SEVEN RELATIVELY COMPLETE NEAR EASTERN NEANDERTAL CRANIA (mm)
A m u d Shanidar Skhul Quafzeh Skhul Skhul Tabun 1 9 6 4 5 1
Cranial Capacity (cc) 1800 1700 1587 1560 1554 1518 1271 Length 215 206 213 198 206 192 183 Breadth 155 157 145 146 148 143 141
Maximum Height Above Frankfort Plane 122 125 115 113 116 121 105
Porion - Prosthion 133 125 109 110 116 111 Nasion 116 111 109 102 109 95 100 Glabella 125 114 122 113 117 106 105 Lambda 115 116 111 104 104 108 100 Opistocranion 108 110 101 95 100 100 93 Inion 90 97 98 81 85 89 79
Upper Facial Height 91 88 74 73 79 73 79 Nasal Height 63 55 54 55 53 58 Calotte Height Above
Glabella - Inion 94 103 87 102 99 100 85 Glabella - Lambda 63 66 53 55 64 68 47

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1181
Skhul cave a t Mount Carmel is the origin of a large number of fossil hominids (Mc- Cown and Keith 1939) which taken to- gether, represent a sample of a truly transi- tional group (Brace 1962b, 1964, 1967a; McCown and Keith 1939; Boule and Vallois 1957:376-378; Howell 1958). The morpho- logical characteristics of these specimens are well known, so that repetition is not neces- sary. The direct association with a Levalloiso-Mousterian industry is un- questionable (Garrod and Bate 1937). As an individual specimen, Skhul 5 fits completely within the range of variation for anatomical- ly modern H. supiens.
At Djebel Quafzeh more than thirteen hominids have been recovered; although not one has been even partially described (Vandermeersch 1966, 1969b, 1970), the crania of one of the first six individuals has been pictured (Boule and Vallois 1957, Fig. 247; Coon 1963: Plate XXVIII). There has been a relatively great amount of recon- struction in the vault form, although the face is largely unreconstructed. The form, morphology, and shape of the cranial vault fi t equally well within both Neandertal and anatomically modern H. sapiens ranges of variation. It is most similar to crania from Mount Carmel (Boule and Vallois 1957). The “Neandertalian” characteristics which it possesses, according to Boule and Vallois (1957:377), include a relatively broad nose and a very large palate. On the other hand, the index of upper facial height t o cranial length-37, falls within the Skhul range (35-38, n=3) and far below the range of “classic” Neandertals (41-50, n=6). Similarly, the ratio is significantly smaller than the ratios for the three large-faced Near Eastern specimens: Amud (42), Shanidar I (43) and Tabun (43).
Table VII reveals a mixture of modern and archaic features in the metric dimen- sions and proportions. For instance, the distance from porion to glabella is greater than that of Skhul 5. However, the porion- prosthion measure is far less, while the porion-nasion measure is again greater. Quafzeh 6 shows the total facial prog-
nathism common to Neandertals, rather than the alveolar prognathism evinced by Skhul 5. Another archaic feature of the specimen is a very low calvarium, as indicated by the small auricular height value. However, while they are of Neandertal proportion, the facial dimensions of the cranium are absolutely small: most distances from porion (ex- cepting height) are best matched by Tabun 1, a far less transitional specimen with a much smaller cranial capacity. The calvarium is similar t o many extant crania. Like Skhul 5 it bears some resemblances to crania of Australian aborigines. Comparison with a particularly rugged Australian aborigine cranium pictured by Larnach and Macintosh (1966: Plate I) is striking. The index of facial height to cranial length in this Australian aborigine specimen is 35. As we have already noted, the Quafzeh skeletons are unquestionably associated with a Levalloiso-Mousterian industry. Vander- meersch, who has recovered a number of the skeletons from the Quafzeh Mousterian levels, observes “les hommes de Quafzeh sont a ranger parmi les neanthropiens bien qu’ils soient associes a une industrie mousterienne” (1969a: 17).
Another transitional specimen comes from the Amud cave (Suzuki 1965, 1968) associated with a Levalloiso-Mousterian in- dustry (Binford 1968:709). The specimen combines a number of transitional character- istics (Vallois 1962) and is clearly related to other Near Eastern hominids from Skhul and Quafzeh on the one hand, but also to hominids from Shanidar. The resemblance to Shanidar 1 (Stewart 1958) is marked. The anterior dentition, supraorbital torus, and upper third molars of this specimen are each relatively small. The mastoid process is well developed, the forehead high, and a mental eminence is present (Suzuki 1965). The calvarium is absolutely high. Maximum height above the Frankfort Horizontal, mea- sured from the scaled photograph (Suzuki 1968) measures 122mm . This is higher than any “classic” Neandertal, but is exceeded by Shanidar 1. These, then, are features reminis- cent of anatomically modern H. supiens. On
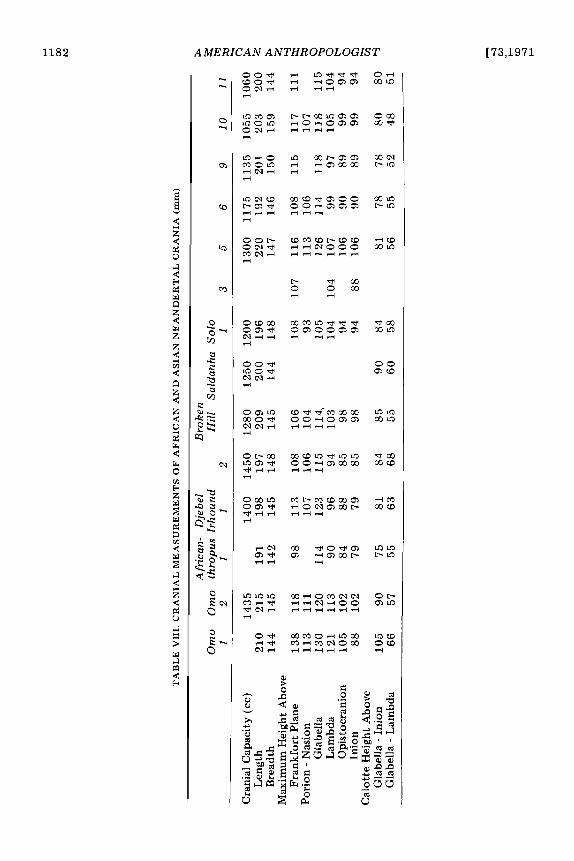
TA
BL
E V
III.
CR
AN
IAL
ME
AS
UR
EM
EN
TS
OF
AF
RIC
AN
AN
D A
SIA
N N
EA
ND
ER
TA
L C
RA
NIA
(m
m)
Afr
ican
- D
jebe
l B
roke
n b
6 % 3 11
7 ll
1 s
Om
o O
mo
thro
pus
Irho
und
Hill
Sald
anha
Sol
o 1
0
I1
1 2
1 1
2 1
3 5
6 9
Cra
nial
Cap
acity
(c
c)
1435
14
00
1450
12
80
1250
12
00
1300
11
75
1135
10
55
1060
b
L
engt
h 21
0 21
5 1
91
19
8 19
7 20
9 20
0 1
96
22
0 19
2 20
1 20
3 20
0 2
B
read
th
14
4
14
5
14
2
14
5
148
14
5
144
148
147
146
150
159
144
b
Fran
kfor
t Pl
ane
13
8
118
98
11
3
108
10
6
108
107
116
108
11
5
Max
imum
Hei
ght A
bove
Pori
on -
Nas
ion
11
3
111
107
10
6
104
93
11
3 1
06
10
7 G
labe
lla
130
120
114
123
11
5
114,
1
05
12
6 11
4 11
8 11
8 11
5 L
ambd
a 1
21
11
3 90
9
6
94
10
3
104
104
107
99
97
105
104
8 O
pist
ocra
nion
1
05
10
2 84
88
8
5
98
94
106
90
8
9
99
Inio
n 88
10
2 79
79
8
5
98
94
88
106
90
8
9
99
Et $
Cal
otte
Hei
ght
Abo
ve
2 G
labe
lla -
Inio
n 1
05
9
0
75
81
84
85
9
0
84
81
78
78
80
80
G
labe
lla -
Lam
bda
66
57
55
63
68
55
6
0
58
56
55
52
48
51

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1183
the other hand, the calvarium is extremely long (215mm) and broad (155mm); the face is long and broad with an upper facial height of 91mm. This is one of the largest Neander- tal faces matched or exceeded only by Broken Hill, La Ferrassie I, and Petralona. The occipital contour of the skull is round, the occipital region is vertically compressed in a manner similar to that of Djebel Irhound 2, and the ratio of facial height t o cranial length is relatively great (41), indi- cating a Neandertal sized and proportioned face. Measurements from porion to pros- thion and porion to nasion (Table VII) indi- cate extensive mid-facial prognathism.
The remaining transitional specimens come from Africa. There, the recently dis- covered crania from the Kibish formation of the Orno basin evince a mixture of features found both in Neandertals and in anatom- ically modern H. supiens (Leakey, Butzer, and Day 1969). No features occur which are not found in other Neandertals. However, the combinations are unique. The three skeletons have not been precisely dated. They are reported t o come from Member I of the formation, although only some re- mains of Orno 1 were recovered in si tu: Orno 2 and 3 were collected from the surface. A radiocarbon date of 37,000 BP was reported for an Eutheriu bank in Member 111, over- lying Member I. The skeletal material is probably older, although some uncertainty stems from the fact that shell-based dates have a tendency t o appear older than they actually are (Crane 1956). Corroborating dates from other sources would be useful. For the time being, the Orno skeletons appear t o be roughly contemporary with European and Near Eastern Neandertals.
Table VIII gives metric data for the two better preserved Orno crania. For purposes of comparison the other reasonably complete African and Asian Neandertal crania are tabulated. The Orno measurements were taken from Day (Leakey, Butzer, and Day 1969), and measured from the photographs in this publication. The scale indicated for the photographs is a t variance with the measurements published in the text. The
text was assumed correct (Day, personal communication), and t h e photographs re- scaled accordingly in order t o obtain the data in Table VIII. Sources for most of the other material have been discussed. The “Africanthropus” calvarium was published by Weinert (1940).
Orno 1 , like Skhul 5, would be identified as anatomically modern H. supiens if found in another context. Modern features include the large mastoid process, high cal- varium (the auricular height is 138 mm. and is the Neandertal maximum), rounded oc- cipital region and low inion, and mandibular mental eminence. The breadth of the heavily worn upper canine, 8.1 mm., is a t the low end of the Neandertal range of variation. Of 35 individuals, only Spy 1 and Skhul 1 are narrower. The length of 8.9 mm., on the other hand, is greater than that of twenty- five individuals, and is within 0.1 mm. of an additional five. Following Day (Leakey, Butzer, and Day 1969:1142), the tooth is robust by modern standards. Other features falling within t h e Neandertal range of varia- tion include t h e heavy supraorbital torus coupled with the absence of a supraorbital sulcus (similar t o “Africanthropus”), robust facial bones, low position of maximum cranial breadth, and great cranial length. In both metric and non-metric morphology, the cranium is extremely similar to Amud. Both share the heavy supraorbitals without a supraorbital sulcus, the mental eminence of the mandible, the rounded occipital contour and low inion position, the large mastoid process, and the evenly rounded lateral contour from glabella t o inion. In dimen- sions the two are almost identical. The9 are clearly more similar to each other than either is to any other Neandertal cranium. This relation is even true of the relatively great distance from porion t o glabella, char- acteristic only of these two crania. The only difference lies in the fact that the Amud cranium is about one centimeter lower and one centimeter broader.
Orno 2 differs from Orno 1 in a number of ways. Yet there is an apparent similarity between the two calvaria. They share a

1184 AMERICAN ANTHROPOLOGIST [ 73,1971
heavy supraorbital torus associated with a receding forehead and absence of a supra- orbital sulcus. However, Omo 2 shows exten- sive and detailed similarities with Solo 5. Both are long and moderately high, with identical morphology of the supraorbital region as well as the region of inion. In both, inion and opistocranion are identical. The nuchal tori are massive, and the nuchal planes flat. Inion is somewhat higher in Solo 5: while the auricular heights are the same, height above the glabella-inion line is greater in Omo 2. Most dimensions of the two are nearly identical. Indeed, the resemblance of Solo 5 to Omo 2 is greater than to any other of the Solo calvaria.
Omo 1 clearly meets the criteria of a transitional specimen. It should not be sur- prising that in East Africa, in a sense geographically between Java and the Near East, two crania show detailed and extensive similarities with specimens from these other areas. The associated lithic materials (if any) have not been described.
In sum, where appropriate archaeological associations exist, unquestionable transi- tional populations are clearly associated with Middle Paleolithic industries in Eastern and Western Europe, Africa, and the Near East. The evolution of local populations associ- ated with Middle Paleolithic industries is thus indicated.
AN ADAPTIVE MODEL
The hypothesis critically addressed by this work was recently summed up by Howell (1969:xxi): “Beginning some 35,000 years ago new peoples with new ideas and new designs for living displaced and even- tually replaced antecedent Neanderthal peo- ples and their Mousterian way of life.”
To the contrary, we suggest that in situ transitions of both hominids and their in- dustries took place throughout the Old World within the period of the last glacia- tion. These transitions did not occur at the same time. The evolution of anatomically modern H. Supiens clearly precedes that of Upper Paleolithic industries. Thus, the
selective factors leading to the evolution of anatomically modern populations are not to be sought in the Upper Paleolithic. Rather, we must seek their origins in the Middle Paleolithic.
We must ask what factors throughout the Old World undergo significant change within the Middle Paleolithic, and, of course, what factors do not. There are several indications that significant changes in human ecology do not occur a t this time. The age distributions of “classic” Neandertals and an Upper Paleo- lithic sample, both published by Vallois (1961), are almost identical to each other and to the age distribution of Indians from Indian Knoll (Snow 1948). Similar adaptive effectiveness for groups spanning this time range is thus indicated (Birch 1948; Cole 1954; Cannon 1968). The extensive hunting of megafauna has already been demonstrated at numerous Acheulean sites (see Howell 1965; Hemmer 1965). The specialized and intensive hunting of particular species can be seen as early as locality 1 at Choukoutian (Howell 1964). These two hunting behaviors do not necessarily occur together, although they often do. The areas exploited by the Acheulean hunter-gatherers included both ecologically homogeneous areas such as Ternifine (Arambourg 1963) and edge areas or ecotones such as Toralba (Butzer 1964:366-371). For these reasons, a recent suggestion concerning Neandertal evolution (Binford 1968) can be dismissed. It was suggested that the Neandertal exploitation of ecotones containing large game on a regular seasonal basis led to the formation of larger groups for more efficient exploita- tion. Clearly, such conditions were met long before the recent date Binford suggests. In any event, the formation of larger and more exogamous groups would slow, rather than hasten, the effects of selection, all other factors remaining equal (Wright 1938).
We see the major behavioral changes within the Middle Paleolithic in industrial factors. These do not relate to any single specific tool type, but rather to a general increase in the numbers of different types of

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1185
tools. A single purpose tool is generally more effective for the purpose for which it was made than any general purpose tool. With respect to the action of selection upon hominid morphology, the major implication of changes from general to specific tools concerns the use of the jaws as a vice. The importance of this usage is indicated by the maintenance of large anterior teeth (actually increasing in breadth for early Neandertals) throughout the Lower and Middle Pleisto- cene, while the posterior dentition progres- sively reduces. To put a hole in a piece of wood with a knife requires more use of a vice than if one were using a drill, That is, the development of special purpose tools reduces selection for both force and power (force over time) in the anterior dentition through a combination of less strenuous use and use over shorter periods of time. Indeed, the reduction of the anterior dentition is part of a general trend reducing skeletal and muscular rugosity, replacing them with more efficient technology.
With the reduction of selection acting to maintain a large anterior dentition, and later the increasing efficiency of cultural adapta- tions to cold, there is a concomitant reduc- tion of robustness and size of Neandertal anterior teeth, and ultimately of the s u p porting facial morphology. This reduction allows the redistribution of cranial mass in a more spherical array. Weidenreich was aware of this redistribution and of its importance in the evolution of anatomically modern H. supiens from Neandertal ancestors, although he was unable to account for it by recourse to evolutionary principles (1940, 1941, 1945, 1947).
Evidence for a selective advantage to cranial mass distribution is considerable. Schultz (1942) was able to show that in both man and apes the weight of the head is always greater anterior to the occipital con- dyles than it is posterior to them. Because the occipital condyles act as a fulcrum about which the nuchal musculature works, the force which must be exerted by these muscles depends upon both the moment of
inertia of the weight distribution in front of the condyles and the distance of the muscles behind the condyles. The counterbalancing effect of Neandertal bunning is not so much to bring additional weight behind the con- dyles, but is rather to lengthen the lever arm for the nuchal musculature as well as to provide a horizontal orientation of the nuchal plane, increasing its leverage. The changes furthest anterior to the condyles have the greatest effect upon the length and orientation of the nuchal lever arm. The anterior dentition, and its supporting facial architecture, are the most anterior features of the cranium. The reduction of this com- plex would, by necessity, result in selection acting to shorten the nuchal lever arm and make it more vertical. The resulting crania (anatomically modern H. supiens) are shorter and narrower but higher, and thus maintain the same cranial capacities.
In this respect, Schultz’s work (1942) was particularly important. His empirical deter- minations evince a surprisingly constant balance ratio for modern crania, associated with a consistant occipital condyle position. Apparently, the conditions for balance in H. supiens represent a selective optimum for erect hominids. A similar selective optimum, with longer anterior and posterior lever arms, was reached with Neandertals evincing larger faces and anterior dentitions.
Other studies of cranial balance yield much the same results (Schultz 1918; Fischer and Mollison 1923). The position of the occipital condyles is fairly constant, and the force exerted by the nuchal musculature in both static and dynamic functioning is sensitive to small mass changes maximally anterior to the condyles.
Therefore, the loss of the distinctive Neandertal cranial form is a direct conse- quence of selection relaxation for the an- terior dentition and supporting facial archi- tecture, and resulting change in selection acting on the static and dynamic properties of the nuchal musculature. That succeeding cold-adapted populations did not develop mid-facial prognathism follows from the

1186 A MERlCA N ANTHROPOLOGIST [ 73,1971
raised position of the calvarium relative to the nasal passages in anatomically modern H. sapiens.
We submit that these considerations are sufficient to account for the association of Early Upper Paleolithic man and Late Mid- dle Paleolithic tools.
Alimen, Marie-Henriette 1967 The Quarternary of France. In The
Quarternary, Vol. 2. Kalervo Ran- kama, ed. New York: Interscience.
1958 Les Stades Evolutifs de 1’Hu- manit6. The Leech 28:106-111.
1963 Le Gisement de Ternifine. Ar- chives de L’Institute de PalBontologie Humaine 32.
Arambourg, C.
Ashton, E. H., and T. F. Spence 1958 Age Changes in the Cranial Capac-
i ty a n d Foramen Magnum of Hominoids. Proceedings of the Zoo- l o g i c a l S o c i e t y of L o n d o n 130 :169-181.
Bailit, H. L., S. J. Dewitt, and R. A. Leigh 1968 The Size and Morphology of the
Nasioi Dentition. American Journal of Physical Anthropology 28 : 271-288.
1908 DBcouverte d’un Squelette Hu- maine MoustBrien & la Bouffia de la Chapelle-Aux-Saints. L’Anthropologie
1969 Tooth Morphology and Other As- pects of the Teso Dentition. American Journal of Physical Anthropology
Bardon, L. J., and A. Bouyssonie
19: 513-518. Barnes, D. S.
30:183-194. Bastin, Bruno
1969 Premiers RBsultats de 1’Analyse Pollinique des Loess en Belgique. Bul- letin de 1’Association pour 1’Etude du Quarternaire 1 :3-12.
Behm-Blanke, G. 1 9 6 0 Altsteinzeitliche Rastplatze im
Traveringebiet Taubach, Weimar: Ehringsdorf. Alt-Thiiringen 4 :1-245.
1911 A Study of the Negro Skull with Special Reference to the Congo and G a b o o n C r a n i a . B i o m e t r i k a
Benington, R. C.
8 ~292-337. Binford, L., and S. Binford
1966 A Preliminary Analysis of Func- tional Variability in the Mousterian of Levallois Facies. American Anthropol- ogist 68 : 238-29 5.
1969 , Stone Tools and Human Behavior. Scientific American 220(4):70-84.
Binford, S. 1968 Early Upper Pleistocene Adapta-
tions in the Levant. American Anthro- pologist 70 :707-7 17.
Birch, L. C. 1948 The Intrinsic Rate of Natural In-
crease of an Insect Population. Journal of Animal Ecology 1 7 :15-26.
Von Bonin 1935 The Magdelenian Skeleton from
CapBlanc in the Field Museum of Natural History. Illinois Medical and Dental Monographs 1 :1-76.
Bordes, F. 1953 Essai de Classification des In-
dustries “Moustbriennes. ” Bulletin de la Socibtb Prkhistorique Franqaise
1955 Le Palkolithique Infkrieur e t Moyen de Jabrud. L’Anthropologie
1958 Le Passage du Paleolithique Moyen au Superieur. In Hundert Jahre Nean- derthaler. G. H. R. von Konigswald, ed. Utrecht: Kemink en &on. pp.
1961 Mousterian Cultures in France. Science 134:803-810.
1968 The Old Stone Age. New York: McGraw-Hill.
1969 Reflections on Typology and Techniques in the Paleolithic. Arctic Anthropology 6 :1-29.
50 :226-235.
59:486-507.
17 5-81.
Boule, M. 1913
Boule, M., and H. V. Vallois 1957
Brace, C. L. 1962a Cultural Factors in the Evolution
of the Human Dentition. In Culture and the Evolution of Man. M. F. Ashley Montagu, ed. New York: Ox- ford. pp. 343-354.
1962b Refocusing on the Neanderthal Problem. American Anthropologist
1964 The Fate of the “Classic” Nean- derthals: A Consideration of Hominid Catastrophism. Current Anthropology 5:3-43.
1967a The Stages of Human Evolution. Englewood Cliffs: Prentice Hall.
1967b Environment, Tooth Form, and Size in the Pleistocene. Journal of Dental Research 46:809-816.
1967c More on the Fate of the “Classic” Neanderthals. Current Anthropology
L’homme Fossile de la Chapelle- aux-Saints. Paris: Mason.
Fossil Man. New York: Dryden.
64:729-741.
7:204-214.

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1187
1 9 6 8 Neanderthal. Natural History 77:38-45.
Brace, C. L., and S. Molnar 1967 Experimental Studies in Human
Tooth Wear. American Journal of Physical Anthropology 27:213-222.
1966 Prehistoric Men. New York: Scott Braidwood, Robert
Foresman. Brettinger, E.
1955 Das Schadelfragment von Swans- comb und das “Praesapiens” Problem. Mitteilungen der Anthropologischen Gesellschaft in Wien 84/85:1-45.
Brothwell, D. R. 1959 Teeth in Earlier Human Popula-
tions. Proceedings of the Nutrition Society 18:54-64.
1963 The Macroscopic Dental Pathology of Some Earlier Human Populations. In Dental Anthropology. D. R. Broth- well, ed. New York: Pergamon. pp. 271-288.
Burkitt, Miles 1963 The Old Stone Age. New York:
Atheneum. Butzer, K. W.
1963 Environment and Archaeology. London: Methuen.
Butzer, K. W. and D. L. Thurber 1969 Some Late Cenozoic Sedimentary
Formations of the Lower Omo Basin. Nature 222: 1138-1143.
Campbell, T. D. 1925 Dentition and Palate of the
Australian Aboriginal. Adelaide: Has- sell Press.
1939 Food, Food Values, and Food Habits of the Australian Aborigines. Australian Dental Journal 43 :1-15, 45-55, 73-87, 141-156, 177-198.
Cannon, K. D. Homo Neanderthalensis: His role in Hu-
man Evolution. Unpublished manu- script.
Captain, L., and D. Peyrony 1909 Deux Squelettes Humains au
Mi l ieu des Foyers de l’epoque Mouster ienne . Revue de 1’Ecole d’Anthropologie 19:402-409.
1912a Le Station Prehistorique de La Ferrassie, Commune de Savignac- du-Bugue (Dordogne). Revue Anthro- pologique 22:29-50, 76-99.
1912b Trois Nouveaux Squelettes Hu- mains Fossiles. Revue Anthropolo- gique 22:439-442.
1921 Decouverte d’un Sixieme Sque- lette Moustkrien 1 La Ferrassie. Revue Anthropologique 31 :382-388.
Carbonnell, V. M. 1965 The Teeth of the Neandertal Child
f r o m Gibraltar: A Re-evaluation. American Journal of Physical Anthro- pology 23:41-50.
Cole, L. C. 1954 The Population Consequences of
Life History Phenomena. Quarterly Review of Biology 29 :103-137.
Collie, George 1928 The Aurignacians and their Cul-
ture. Logan Museum Bulletin l. Coon, C. S.
1963 The Origin of Races. New York: Knopf.
1964 Comment on The Fate of the “Classic” Neanderthals: A Considera- tion of Hominid Catastrophism, by Brace (1964). Current Anthropology 5:21-22.
Crane, H. R. 1956 University of Michigan Radio-
carbon Dates I. Science 124:664. Dahlberg, A. A.
1963 Dental Evolution and Culture. Hu- man Biology 35:237-249.
Debenath, A., and J. Piveteau 1969 Nouvelles Decouvertes de Restes
H u m a i n s Fossiles B La Chaise- d e - V o u t h o n (Charente). Position Stratigraphique de Restes Humains de La Chaise (Abri Bourgeouis-Delaunay). Comptes Rendue de L’AcadBmie des Sciences, SBrie D 269:24-28.
De Jong, Jan 1967 The Quarternary of the Nether-
lands. In The Quarternary, Vol. 2. Kalervo Rankama, ed. New York: In- terscience.
De Lumley, M. 1970 Le Parietal Humain Anteneander-
talien de Cova Negra (Jativa, Espagne). Comptes Rendus de L’Academie des Sciences, Sdrie D 270: 39-41.
De Sonneville-Bordes, D. 1963 Upper Paleolithic Cultures in
Western Europe. Science 142:347-355.
1953 Essai d’Adaptation des Mdthodes S t a t i s t i q u e s a u P a l e o l i t h i q u e Superieur-Premiers Resultats. Bulletin de la Socidtd Prehistorique Francais
De Sonneville-Bordes, D. and J. Perrot
NO. 5-6. De Villiers, H.
1958 The Skull of the South African Negro. Johannesburg: Witwatersrand University.
Dupree, L. 1969 .Comment on Neandertal Man and
Homo sapiens in Central and Eastern Europe. Current Anthropology 10:492.

1188 AMERICAN ANTHROPOLOGIST [73,1971
Dupree, L., L. J. Lattman, and R. S. Davis 1970 Ghar-i-mordeh Gusfand (Cave of
the Dead Sheep): A New Mousterian Locality in North Afghanistan. Science 167 :1610-16 12.
Ennouchi, E. 1962 Un Neandertalien: 1’Homme der
L ’ Anthropologie
1968 Le Deuxi6me Crane de 1’Homme d’Irhound. Annales de Paleontologie Vertsbres 54 :117-128.
1969 Presence d’un Enfant NBander- thalien au Jebel Irhound. (Maroc). Annales de Paleontologie Vert6bres
J e be 1 I r h o u n d . 66:279-299.
55 : 251-265. Fenart, M. R.
1969 Le Crane de 1’Enfant de 1’Aze. E tudih dans les Axes Vestibulaires d’orientation. Comptes Rendu de L’AcadBmie des Sciences, Serie D 268 : 2042-2045.
Fischer, E., and T. Mollison 1923 Kultur der Gegenwart, part 111.
Leipzig: Teubner. Fitting, J. E.
1969 Comment on Early Man in America, by A. L. Bryan. Current Anthropology 10:339-365.
New York: John Wiley & Sons.
Flint, Richard F. 1957 Glacial and Pleistocene Geology.
Fraipont, J., and M. Lohest 1887 La Race Humaine de NBanderthal
ou de Canstadt en Belgique. Archives de Biologie 7:587-758.
Freeman, L. G., and J. Gonzalez Echegaray 1970 Aurignacian Structural Features
and Burials at Cueva Morin ( S a n t a n d e r , Spa in ) . Na tu re 226 ~722-726.
Garrod, D. A. E. 1928 Excavations of a Mousterian Rock
Shelter at Devil’s Tower, Gibraltar. Journal of the Royal Anthropological Institute 58:33-56.
1962 An Outline of Pleistocene Pre- history in Palestine. Quarternaria 6 : 541- 546.
Garrod, D. A. E., and D. M. A. Bate 1937 The Stone Age of Mount Carmel.
Volume 1: Excavations at the Wady el-Mughara. Oxford: Clarendon.
Gessain, P. 1959 La Dentition des Eskimo
d’angmassalik Genetique Croissance et Pathologie. SociBtB d’Anthropologie de Paris, Bulletin et Memoires, SBrie 10:364-396.
Gor janovic- Kramberger, D. 1906 Der Diluviale Mensch von Krapina
in Kroatien: ein Beitrag zur Palao- anthropologie. Wiesbaden: Kriedel.
1907 Die Kronen und Wurzeln der Mahlzahne des Homo Primigenius und ihre Genetische Bedeutung. Ana- tomischer Anzeiger 31 :97-134.
1910 Homo Aurignacensis Hauseri in Krapina? Verhandlungen der Geo- logische Reichsanstalt 1 4 :312-317.
Zivot i Kulture Diluvijalonga Cov- jeka iz Krapine u Hrvatsko (Hominis Diluvialis e Krapina in Croatia Vita e t Cu l t u r a ) . D j ela Jugoslavenske Akademije Znanosti i Umjetnosti
1913
23:l-54. Gupta, P., P. C. Dutta, and A. Basu
1962 Human Remains from Harappa. Anthropological Survey of India, Memoir 9:3-186.
Harrison, T. 1959 Radio-Carbon Dates from Niah.
Sarawak Museum Journal 9:136-138. 1964 50,000 Years of Stone Age Cul-
ture in Borneo. Annual Report of the S m i t h s o n i a n I n s t i t u t i o n 1964:521-530.
Heim, J-L 1968 Les Restes NBandertaliens de La ~
Ferrassie. Nouvelles DonnBes sur la Stratigraphie et Inventaire des Sque- l e t t e s . Comptes Rendus de L’AcadBmie des Sciences, SBrie D 266 : 576-578.
Heinz, N. 1967 Evolution de la Hauteur Maximale
du Frontal, du Parihtal, e t du l’occipi- tal Chez les Hominides. Annales de PalBontologie, Vertebres 53:50-75.
Hemmer, H. 1965 Die Aussage der Saugetierfauna fur
die Okologie Pleistozaner Hominiden. Homo: Zeitschrift fur die Verg- leichende Forschung am Menschen 16:95-109.
Henri-Martin, G. 1923 L’Homme Fossile de La Quina.
Archives de Morphologie GBnBrale et Experimentale 15:l-253.
Hole, F., and K. V. Flannery 1968 Prehistory of Southwestern Iran:
A Preliminary Report. Proceedings of the Prehistoric Society 33:147-206.
Hooten, E. A. 1947 Up From the Ape. New York:
Macmillan. Howell, F. C.
1951 The Place of the Neanderthal Man in Human Evolution. American Jour-

1189 Brose & Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS
nal of Physical Anthropology
1951 Pleistocene Glacial Ecology and the Evolution of “Classic Neandertal” Man. Southwest Journal of Anthropol- ogy 8:377-410.
1957 The Evolutionary Significance of Variation and Varieties of Neanderthal Man. Quarterly Review of Biology
1958 Upper Pleistocene Men of the Southwest Asian Mousterian. In Hundert Jahre Neanderthaler. G. H. R. von Koenigswald, ed. Utrecht: Kemink in Zoon. pp. 185-198.
1959 Upper Pleistocene Stratigraphy and Early Man in the Levant. Pro- ceedings of the American Philosoph- ical Society 103:l-65.
1960 European and Northwest African Middle Pleistocene Hominids. Current Anthropology 1 :195-232.
1964 The Hominization Process. In Horizons of Anthropology. Sol Tax, ed. Chicago: Aldine, pp. 49-59.
9:379-416.
32 :330-347.
1965 Early Man. New York: Time 1967 Recent Advances in Human Evolu-
tionary Studies. Quarterly Review of Biology 42:471-513.
1969 Foreward to Man and Culture in the Late Pleistocene, by R. H. Klein. San Francisco: Chandler. pp. xxi-xxvi.
1967 Mankind in the Making. (Revised
Hrdlicka, A.
Howells, W. W.
edition.) Garden City: Doubleday.
191 1 Human Dentition and Teeth from the Evolutionary and Racial Stand- point. Dominion Dental Journal (Ontario) 21 :1-15.
1927 The Neanderthal Phase of Man. Journal of the Royal Anthropological Institute 57 : 249-274.
1928 Catalog of Human Crania in the United States National Museum: Australians, Tasmanians, South African Bushmen, Hottentots, and Ne- groes. Proceedings of the United States National Museum 7 :1-140.
1930 The Skeletal Remains of Early Man. Smithsonian Miscellaneous Col- lections 83.
1942 Catalog of Human Crania in the United States National Museum col- lections: Eskimo in General. Pro- ceedings of the United States National Museum 91 :169-429.
Hvanova, I. K. 1969 Etude Geologique des Gisements
Pa lbol i th iques d e L’U.R.S.S. L’Anthropologie 73 :5-48.
Jelinek, A. 1965 Lithic Technology Conference,
Les Eyzies, France. American Antiquity 31 :277-278.
Jelinek, J. 1964 Betrachtungen uber die Verwand-
schaft der Anthropologischen Funde Dolni Vestonice, Abri Pataud, und Markina Gora. Zeitschrift fur Morpho- logie und Anthropologie 56:19-23.
1966 Jaw of an Intermediate Type of Neandertal Man from Czechoslovakia. Nature 21 2 : 701 -70 2.
Neandertal Man and Homo saptens in Central and Eastern Europe. Cur- rent Anthropology 10:475-503.
1969
Jelinek, J., J. Palisek, and K. Valoch 1959 Der Fossil Mensch Brno 11.
Anthropos 9:17-22. Kadic, O., L. Bartucz, E. Hillebrand, and J. Szabo
1933 Preliminary Report on the Results of Excavations in the Subalyuk Cave Near Cserepfalu, Hungary. Inter- national Geological Congress : Report of the XVI Session (United States). 2 :783-788.
Kannelis, A., and Savas 1964 Kraniometriki Meleti ton Homo
N eanderthalenis ton Petralonen Epistimeniki. Epetiris tis Physiko- mathemati kis Sholos 9 :6 5-9 2.
Keith, A. 1915 The Antiquity of Man. London:
Williams and Norgate. Kitching, J. W.
1963 Bone, Tooth, and Horn Tools of Paleolithic Man. Manchester: Man- Chester University.
Klaatsch, H. 1923 The Evolution and Progress of
Mankind. New York: Stokes. Klein, R. G.
1969 Mousterian Cultures in European Russia. Science 165:257-265.
Kleinschmidt, 0.
und Meyer. 1931 Der Urmensch. Leipzig: Quelle
Kokkoros, M. M., and A. Kannelis 1960 Dbcouverte d’un Crane d’Homme
Paleolithique dans la Pbninsule Chal- cidique (Petralona). L’Anthropologie 64:438-446.
Krukoff, S. 1970 L’occipital de La Chaise (Suard),
Caracteres Mbtriques, Distances de Forme et de Format. Comptes Rendus de L’Acadbmie des Sciences, Sbrie D, 170 42-45.

1190 AMERICAN ANTHROPOLOGIST [73,1971
Kurten, B. 1968 Pleistocene Mammals of Europe.
Chicago : Aldine. Kurth, G.
1965 Die (Eu) Hominiden: Ein Jeweils- bild nach dem Kenntnisstand von 1964. In Menschliche Abstammtungs- lehre. G. Heberer, ed. Stuttgart: Fischer. pp. 357-425.
Larnach, S. L., and N. W. G. Macintosh 1966 The Craniology of the Aborigines
of Coastal New South Wales. The Oceania Monographs 13.
Leakey, R.E.F., K.W. Butzer, and M.X. Day 1969 Early Homo supiens Remains from
the Omo River Region of South-west Ethiopia. Nature 222 :1132-1138.
Le Gros Clark, W. E. 1964 The Fossil Evidence for Human
Evolution. Revised edition. Chicago: University of Chicago,
1966 , History of the Primates, Fifth edi- tion. Chicago: Phoenix.
Leigh, R. W. 1928 Dental Pathology of Aboriginal
California. University of California publications in American Archaeology and Ethnology 23:399-440.
1937 Dental Morphology and Pathology of Pre-Spanish Peru. American Journal of Physical Anthropology 22:267-295.
1958 Etude des Restes Humains Fossiles Prouvenant des Grottes d’Arcy- sur-Cure. Annales de PaMontologie
1961 Les Fouilles d’Archy-Sur-Cure
Leroi-Gourhan, A.
44:87-148.
(Yonne). Gallia 4:l-16.
McBurney, C. B. M. 1960 The Stone Age of North Africa.
New York: Penguin. 1967 The Haua Fteah (Cyrenaica) and
the Stone Age of the South-East Mediterranean. London : Cambridge University.
McCown, T., and A. Keith 1939 The Stone Age of Mount Carmel.
Volume 11: The Fossil Human Re- mains from the Levalloiso-Mousterian. Oxford: Clarendon.
Mash , K. J. 1882 Uber den Diluvialen Menschen in
Stramberg. Mitteilungen der Anthro- pologischen Gesellschaft in Wien 1 2 : 32-38.
1886 Uber den Menschlichen Unter- keifers in der Sipkahahle. Verhand- lungen der Berliner Anthropologischen Gesellschaft 18 :341-344.
1889 Die Lossfunde bei Brunn und der diluviale Mensch. Mitteilungen der Anthropologischen Gesellschaft in Wien 19:46-64.
Matiegka, J. 1938 Homo predmostensis, Fosilni Clo-
vek Z Predmosti N o Morave. Czechoslo- vakian Academy of Science and Art Publication. Prague.
I I
Miyabara, T. 1916 An AnthroDoloeical Studv of the . -
Masticatory System in the japanese. Dental Cosmos 16:739-749.
Mongait, A. L. 1961 Archaeology in the USSR. New
York: Pelican. Morant, G. M.
1924 A Study of Certain Oriental Series of Crania Including the Nepalese and Tibetan Series in the British Museum ( N a t u r a l History). Biometrika . .
i6:1-105. 1925 Studies of Paleolithic Man I. The
Chancelade Skull and its Relation to the Modern Eskimo Skull. Annals of Eugenics 1 :257-276.
1927a Studies of Paleolithic Man 11. A Biometric Study of Neanderthal Skulls and their Relationships to Modern Racial Types. Annals of Eugenics
192713 A Study of the Australian and Tasmanian Skulls, Based on Previously Published Measurements. Biometrika
2: 318-381.
19 : 41 7-440. 1928 Studies of Paleolithic Man 111. The
Rhodesian Skull and its Relationships to Neanderthal and Modern Types. Annals of Eugenics 3:277-336.
1930 Studies of Paleolithic Man IV. A Biometric Study of the Upper Paleo- lithic Skulls of Europe and their Rela- tionships to Earlier and Later Forms. Annals of Eugenics 4:109-214.
1938 The Form of the Swanscombe Skull. Journal of the Royal Anthropo- logical Institute 68:67-97.
Mottl, M. 1951 Die Repolusthole bei Peggau und
ihre Eiszeitlichen Bewohner. Archaeo- logia Austriaca 8:l-78.
Movius, H. L. 1969 The Chatelperronian in French
Archaeology: The Evidence of Arcy- sur-Cure. Antiquity 43:111-123.
Neuville, R. 1951 Le Palholithique et la Mdsolithique
de , Desert de Judee. Archives de 1’Institute Paleontologie Humaine 24:l-270.

Brose & Wolpoff] TOOLS A N D MODERN HUMAN ORIGINS 1191
Noble, H. E. 1926 The Teeth of the Richmond River
Blacks. Australian Journal of Dentistry 30 :4-12.
Oakley, K. A. 1962 Man the Tool-Maker. Chicago:
1966 Frameworks for Dating Fossil University of Chicago Press.
Man. Chicago : Aldine. Okladnikov, A. P. et al.
1 9 4 9 Teshik-Tash: Paleolithicheskii Chelovek. Moscow: Trudy Nauchne Issledovatelskogo Instituta Antro- pologii.
Pattee, E. 1955 Les NBanderthaliens. Anatomie,
Physiologie, Comparaisons. Paris: Mas- son et Cie.
Pearson, K., and A. G. Davin 1926 On the Biometric Constants of the
Human Skull. Biometrika 16:328-363.
1938 Investigation into the Dental Con- ditions of about 3000 Ancient and Modern Greenlanders. Dental Record
Pedersen, P. 0.
58:191-198. Perrot, J.
1968 La Pr6histoire Paldstinienne. Supplement au Dictionnaire de la Bible 8:286-446. Paris: Letouzey and Ane.
Piveteau, J. 1964 La Grotte de Regourdou 11.
Annales de Pal6ontologie Vertebres
1967 Un Pari6tal de la Grotte du Lazaret ( Alpes-Maritimes). Annales de PalBontologie Vertebres 53 :166-199.
1969 La Pal6ontologie Humaine en France. Etudes Francaises sur Le Quarternaire. INQUA, Paris.
50: 23-62.
Piveteau, E., H. De Lumley, and M. De Lumley 1963 DBcouverte de Restes NBander-
thaliens dans la Grotte de L’Hortus. Comptes Rendus de L’AcadBmie des Sciences, SBrie D 256:40-44.
Poulianos, A. N. 1966 The Place of the Petralonian Man
among the Palaeoanthropoids. Anthro-
1969 Comment on Neandertal Man and Homo sapiens in Central and Eastern Europe. Current Anthropol- ogy 10:496.
POS 19:216-221.
Pradel, L. 1966 Transition from Mousterian to
Perigordian: Skeletal and Industrial. Current Anthropology 7:33-50.
Randall, F. E. 1943-1944 The Skeletal and Dental De-
velopment and Variability of the Gorilla. Human Biology 15:236-254, 307-337; 16:23-76.
Schaefer, U. 1964 Homo Neanderthalensis (King) 11.
E-Schadel-fragment, frontale F-1, and torus-fragment 37.2 von Krapina. Zeit- schrift fur Morphologie und Anthro- pologie 54:260-271.
Schlaginhaufen, 0. 191 8 Anthropologische Untersuchungen
an der Schadelbasis. Archiv fur Anthropologie 16 :1-103.
1942 Conditions for Balancing the Head in Primates. American Journal of Phys.ica1 Anthropology 29:483-497.
1946 Uber die Menschformen des Jungeren Palaolithikums in Europa. Experientia 2:303-306.
Schwalbe, G. 1901 Der Neanderthalschadel. Bonner
Jahrbuch 106:l-72. 1906 Studien zur Vorgeschichte des
Menschen. Stuttgart, Schweizer- bartsche.
1914 Kritische Besprechung von Boule’s Werk: “L’homme Fossile de la Chapelle-aux-Saints.” Zeitschrift fur Morphologie und Anthropologie 16 : 527-610.
Selmer-Olsen, R. 1949 An Odontometric Study of the
Norwegian Lapps, Skifter Det Norske Videnskaps-Akademi I (Oslo), Mathematisk-Naturvidenkapelig Klasse 65:l-168.
Senyurek, M. 1959 A Study of the Deciduous Teeth
of the Fossil Shanidar Infant. Division of Paleoanthropology, University of Ankara, Publication No. 128.
Sergi, S. 1940 Der Neandertalschadel vom Monte
Circeo. Anthropologischer Anzeiger
1944 Craniometria e Craniografia del F’rimo Paleanthropo di Saccopastore. Richerche di Morfologia 21 :733-791.
1948 I1 Cranio des Seconco Palean- thropo di Saccopastore. Craniografia e Craniometria. Palaeontographia Italica
1958a The Neanderthal Palaeanthropi in Italy. In Ideas on Human Evolution. W. W. Howells, ed. (1962). Cambridge: Harvard University. pp. 500-506.
1 9 5 8 b D i e N e a n d e r t a l i s c h e n Palaeanthropen in Italien. In Hundert
16~203-217.
42:4-164.

1192 AMERICAN ANTHROPOLOGIST [73,1971
Jahre Neanderthaler. G. H. R. von Koenigswald, ed. Utrecht: Kemink en Zoon. pp. 38-51.
Shapiro, H. L. 1929 Contributions to the Craniology of
Central Europe 1: Crania from Greifenberg in Carinthia. Anthropolog- ical Papers of the American Museum of Natural History 31:l-120.
Singer, R. 1954 The Saldanha Skull from Hope-
field, South Africa. American Journal of Phvsical Anthropologv 12:345-362. - --
Snow, C. E.“ 1948 Indian Knoll Skeletons of Site Oh
2. Ohio Countv. Kentuckv. University of Kentucky Re’ports in Anthropology 4:371-554.
Sollas, W. J. 1908 On the Cranial and Facial Char-
acters of the Neanderthal Race. Philo- sophical Transactions of the Royal Society of London, Series B 199: 281-339.
Stewart, T. D. 1958 First Views of the Restored Shani-
dar I Skull. Sumer 14:90-96. 1960 Indirect Evidence of the Primitive-
ness of the Swanscombe Skull. American Journal of Physical Anthro- pology 18:363.
1961 The Skull of Shanidar 11. Sumer 17 :97-106.
1963 Shanidar Skeletons IV and VI. Sumer 1923-25.
Straus, W. L., Jr., and A. J. E. Cave 1957 Pathology and the Posture of
Neanderthal Man. Quarterly Review of Biology 32:348-363.
Sullivan, L. R. 1921 A Few Adamanese Skulls with
Comparative Notes on Negrito Crani- ometry. Anthropological Papers of the American Museum of Natural History 23 :i75-200.
Suzuki, H. 1965 A Paleoanthropic Man from the
Amud Cave, Israel. Communications to the VII International Congress of Anthropological and Ethnological Sci- ences, Moscow: 1-8.
1968 A Paleoanthropic Fossil Man from Amud Cave, Israel. Congres Inter- national des Sciences Anthro- pologiques et Ethnologiques (Moscou, 1964) 3:305-316.
Taylor, R. M. S. 1934 Maori Foods and Methods of Prep-
aration. New Zealand Dental Journal 30, No. 147.
1963 Cause and Effect of Wear of Teeth. Acta Anatomica 53:97-157.
1958 Metissage ou Transformation? Essai sur les Hommes Fossiles de Palestine. L’Anthropologie 6 2 : 30-52.
1963 The Dentition of the Subalyuk Neanderthal Child. Zeitschrift fur Morphologie und Anthropologie
1965 The Definition of the Neander- thals and the Position of the Fossil Men of Palestine. Yearbook of Physi- cal Anthropology 1 3 :137-145.
Thoma, A.
54:127-150.
Thompson, E. Y. 1916 A Study of the Crania of the
Moriori or Aborigines, of the Chatam Islands, Now in the Museum of the Royal College of Surgeons. Biometrika 11 : 82-1 3 5.
Tobias, P. V. 1964 Comment on The Fate of the
“Classic” Neandertals: A Considera- tion of Hominid Catastrophism. Cur- rent Anthropology 5:30-31.
Tode, A., F. Preul, and A. Kleinschmidt 1953 Die Untersuchung der Palao-
litischen Freilandstation von Salz- gitter-Levenstedt. Eiszeitalter und Gegenwart 3:144-220.
Turner, C. G., and J. D. Cadien 1969 Dental Chipping in Aleuts,
Eskimos, and Indians. American Jour- nal of Physical Anthropology 31 :303-310.
Vallois, H. V. 1951 La Mandible Humaine Fossile de la
Grotte du Porc-Epic Pr6s Dire-Dawa. L’Anthropologie 5 5 : 23 1-238.
1958 La Grotte de Fontechevade: Anthropologie (deuxi6me partie). Archives de 1’Institut de PalBontologie Humaine, MBmoire 29:5-156.
The Social Life of Early Man: The Evidence of Skeletons. In The Social Life of Early Man. S. L. Washburn, ed. Chicago: Aldine. pp. 214-235.
1962 Un Nouveau NBanderthaloide en P a l e s t i n e . L ’ A n t h r o p o l o g i e
1969 Le Temporal Ndandertalien H 27 de La Quina. L’Anthropologie
1961
66:405-407.
73:365-400. Valoch, K.
1968 Evolution of the Paleolithic in Central and Eastern Europe. Current Anthropology 9:351-390.
Vandermeersch, B. 1 9 6 5 Position Stratigraphique et
Chronologie RBlative des Restes Hu-

Brose & Wolpoff] TOOLS AND MODERN HUMAN ORIGINS 1193
mains du PalBolithique Moyen du Sud-Ouest de la France. Annales de PalContologie VertCbres 51 :69-126.
1966 Nouvelles Dbcouvertes de Restes Humains dans les Couches Levalloiso- MoustBriennes du Gisement de Quaf- zeh (Israel) Comptes Rendus de L’AcadBmie des Sciences, SBrie D.
1969a Quoted in: Rapport d’Activite 1968-1969. Centre National de la RB- cherche Scientifique. Report on the Prehistoric and Protohistoric Civiliza- tions of the Asiatic Near East. D. Ferembach, ed. No. 50.
1 9 6 9 b Les Nouveaux Squelettes MoustBriennes DBcouverte a’Qafzeh (Israel) e t Leur Signification. Comptes Rendus de L’AcadBmie des Sciences, SBrie D, 268:2562-2565.
1970 Une Sepulture Moustdrienne avec Offrandes DBcouverte dans la Grotte de Qafzeh. Comptes Rendus de L’AcadBmie des Sciences, SBrie D,
262:1434-1436.
270 : 298-301. Van Reenen, J. F.
1966 Dental Features of a Low-Caries Primitive Population. Journal of Den- tal Research 47:703-713.
Vergnaud-Grazzini, C., and Y. Herman-Rosenberg
1969 Etude Paleoclimatique d’une Carotte de MBditerrance Orientale. RB- vue de GBographie Physique et de Geologie Dynamique 11 :279-292. - .
Verneau,R. 1924 Le Race de Neanderthal et la Race
de Grimaldi: Leurs Roles dans 1‘Hu- manitB. Journal of the Royal Anthro- pological Institute 54:211-230.
Virchow, H. 1920 Die Menschlichen Skelettreste aus
dem Kampfeschen Bruch im Travertin von Ehringsdorf bei Weimar. Jena: Fischer.
Virchow, R. 1882 Der Keifer aus Sipka-huhle und
der Keifer la Naulete. Zeitschrift fur Ethnologie 14:277-310.
Vlcek, E. 1955 The Fossil Man of Ganovce,
Czechoslovakia. Journal of the Royal Anthropological Institute 8 5 :163-171.
1964 Einige in der Ontogenese des Modernen Menschen untersuchte Neandertalmerkmale. Zeitschrift fur MorDhologie und Anthropologie 56 :63-83.-
1965 Neuer Fund eines Neandertalers in ~~
der Tschechoslowakei. Anthro- pologischer Anzeiger 27:162-166.
1 9 6 7 Die Sinus Frontales bei Europaischen Neandertalern. Anthro- pologisc her Anzeiger 30 : 166-1 69.
1969 Neandertaler der Tschechoslo- wakei. Prague : Tschechoslowakischen Akademie die Wissenschaften.
Waugh, L. M. 1937 Influence of Diet on the Jaws and
Face of the the American Eskimo. Journal of the American Dental Association 24 :1640-1647.
Weckler, J.E. 1954 The Relationships between Nean-
dertal Man and Homo sapiens. A m e r i c a n A n t h r o p o l o g i s t 56:1003-1025.
Weidenreich, F. 1928 Der Schadelfund von Weimar-
Ehringsdorf. Jena: Fischer. 1940 The “Neanderthal Man” and the
Ancestors of Homo sapiens. American Anthropologist 42 :375-383.
1941 The Brain and its Role in the Phylogenetic Transformation of the Human Skull. Transactions of the American Philosophical Society
1945 The Brachycephalization of Re- cent Mankind. Southwestern Journal
1947 Facts and Speculations Concerning the Origin of Homo sapiens. American Anthropologist 49 : 187-203.
Morphology of Solo Man. Anthro- pological Papers of the American Museum of Natural History
31:321-442.
of Anthropology 1 :1-54.
1951
43( 3):205-290. Weinert, H.
1926 Der Schadel des Eiszeitlichen _ _ _ _ Menschen von Le Moustier. Berlin: Springer.
1936 Der Urmenschenschadel von Stein- heim. Zeitschrift fur Morphologie und Anthropologie 3 5 :413-518.
1940 Africanthropus Njarasensis. Zeit- schrift fur Morphologie und Anthropologie 38:252-308.
1944 Ursprung der Menschenheit. Stutt- gart: Enke.
Wells, L. H. 1947 A Note on the Maxillary Fragment
from the Broken Hill Cave. In New Studies on Rhodesian Man. J. D. Clark, K. P. Oakley, L. H. Wells, and J. A. C. McClelland. Journal of the Royal Anthropological Institute 77 : 7-3 2.
1928 Der Fossile Mensch. Berlin: Born- Werth, E.
traeger.

1194 AMERICAN ANTHROPOLOGIST [ 73,1971
West, R. G. 1967 The Quarternary of the British
Isles. In The Quarternary, Vol. 2. Kalervo Rankama, ed. New York: Interscience.
1970 Review o f Slovensko v Starsej a Strednej Dobe Kamannej, by J. Barta. Bratislava, Slovenska Akademia Vied, 1965. American Anthropologist 72 :454-456.
Woldstedt, Paul 1967 The Quarternary of Germany. In
The Quarternary, Vol. 2. Kalervo Rankama, ed. New York: Interscience.
1968 Climatic Influence on the Skeletal Nasal Aperture. American Journal of Physical Anthropology 29 : 40 5-4 24.
1969 Cranial Capacity and Taxonomy of Olduvai Hominid 7. Nature
1971 Metric Trends in Hominid Dental Evolution. Case Western Reserve Uni- versity Studies in Anthropology 2.
Wolpoff, M. H.
223:182-183.
Woo, Ju-Kang, and Ru-Ce Peng 1959 Human Fossil Skull of Early Paleo-
anthropic Stage Found a t Mapa, Shaoquan, Kwangtung Province. Vertebrata Palasiatica 3 :176-182.
Wright, S. 1938 Size of Population and Breeding
Structure in Relation to Evolution. Science 87:430-431.
Yakimov, V. P. 1969 Comment on Neandertal Man and
Homo sapiens in Central and Eastern Europe. Current Anthropology 10:498.
Zubakov, V. A. 1969 La Chronologie des Variations
Climatiques au Cours du Pleistocene en Siberie Occidental. RBvue de GBo- graphie Physique et de GBologie Dynamique 11:315-324.




















