
Download this - Gravitational and Space Research
171
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Download this - Gravitational and Space Research


Gravitational and Space Biology
Volume 19, Number 2
August 2006
Publication of the American Society for Gravitational and Space Biology ISSN 1089-988X
ASGSB EDITORIAL BOARD
Augusto Cogoli Zero-G LifeTec GmbH
Zürich, Switzerland
Luis Cubano Univ. Central del Caribe
Camuy, Puerto Rico
Emily Holton
NASA Ames Research Center
Moffett Field, CA
John Kiss Miami University
Oxford, OH
Patrick Masson University of Wisconsin
Madison, WI
Gloria Muday Wake Forest University
Winston Salem, CT
Anna-Lisa Paul University of Florida
Gainesville, FL
April Ronca Wake Forest University
Winston Salem, CT
Gerry Sonnenfeld SUNY Binghamton
Binghamton, NY
Paul Todd SHOT, Inc.
Greenville, IN
Sarah Wyatt Ohio University
Athens, OH
William Landis NE Ohio Univ College of Medicine
Rootstown, OH
PUBLISHING STAFF
Stan Roux Editor-in Chief
University of Texas
Austin, TX
Mary E. Musgrave Publishing Editor
University of Connecticut
Storrs, CT
Robert Blasiak Assistant Editor
Albert-Ludwigs-Universität Freiburg
Freiburg, Germany
John Kiss Symposium I Editor Miami University
Oxford, OH
Charles Wade Symposium II Editor US Army Inst. of Surgical Res.
Fort Sam Houston, TX
Paul Todd Symposium III Editor SHOT, Inc.
Greenville, IN

ii Gravitational and Space Biology 19(2) August 2006
GENERAL INFORMATION
Gravitational and Space Biology (ISSN 1089-988X) is a journal devoted to research in gravitational and space biology. It
is published by the American Society for Gravitational and Space Biology, a non-profit organization whose members share a
common goal of furthering the understanding of the biological effects of gravity and the use of the unique environment of
spaceflight for biological research. Gravitational and Space Biology is overseen by a steering committee consisting of the
Publications Committee, the Editor, the President, and the Secretary-Treasurer of the ASGSB.
The American Society for Gravitational and Space Biology was created in 1984 to provide an avenue for scientists
interested in gravitational and space biology to share information and join together to speak with a united voice in support of
this field of science. The biological effects of gravity have been acknowledged since Galileo’s time, but only since the 1970s
has gravitational biology begun to attract attention. With the birth of the space age, the opportunity for experimentation over
the full spectrum of gravity finally became a reality, and a new environment and research tool became available to probe
biological phenomena and expand scientific knowledge. Space and spaceflight introduced new questions about space
radiation and the physiological and psychological effects of the artificial environment of spacecraft.
The objectives of ASGSB are:
• To promote research, education, training, and development in the areas of gravitational and space biology and
to apply the knowledge gained to a better understanding of the effect of gravity and space environmental
factors on the flora and fauna of Earth.
• To disseminate information on gravitational and space biology research and the application of this research to
the solution of terrestrial and space biological problems.
• To provide a forum for communication among professionals in academia, government, business, and other
segments of society involved in gravitational and space biological research and application.
• To promote the study of concepts and the implementation of programs that can achieve these ends and further
the advancement and welfare of humankind.
A Collaborative Production: This issue of the Gravitational and Space Biology was produced through collaboration with
the Professional Writing and Technical Communication Program at the University of Massachusetts Amherst. Under the
direction of Dr. John Nelson, the PWTC Program has trained students for a variety of professions that demand excellent
technical writing and editing skills. Since its inception in 1990, the program has placed nearly 100% of its graduates. The
American Society for Gravitational and Space Biology is pleased to sponsor an editorial fellowship for students in the PWTC
program at the University of Massachusetts, and gratefully acknowledges the contributions of its students and directors to the
production of our journal.
MEMBERSHIP: The American Society for Gravitational and Space Biology welcomes individual, organizational, and
corporate members in all of the basic and applied fields of the space and gravitational life sciences. Members are active in the
fields of space medicine, plant and animal gravitational physiology, cell and developmental biology, biophysics, and space
hardware and life support system development. Membership is open to nationals of all countries. Members must have
education or research or applied experience in areas related to the Society’s purposes: i.e., Doctorate, Masters with 2 years
experience, Bachelors with 4 years experience (student members must be actively enrolled in an academic curriculum leading
toward a career related to the Society’s purposes), or special appointment by the Board of Directors. Membership
applications may be obtained by writing the American Society for Gravitational and Space Biology, P.O. Box 2581, Chapel
Hill, NC 27515, or at the society website (http://www.asgsb.org).
Gravitational and Space Biology is sent to all members of the American Society for Gravitational and Space Biology.
Requests for copies, information about subscriptions and membership, changes of address, questions on permission to
reproduce parts of this volume, and other correspondence should be sent to the American Society for Gravitational and Space
Biology P.O. Box 2581, Chapel Hill, NC 27515.
Copyright © 2006 by the American Society for Gravitational and Space Biology

Gravitational and Space Biology 19(2) August 2006 iii
American Society for
Gravitational and Space Biology
Proceedings of the 21st Annual Meeting of the
American Society for Gravitational and Space
Biology (Reno, NV November 1-4, 2005)
Featuring:
Symposium I: Biological Advanced Life Support Systems
Symposium II: Astronaut Health: From the Bench to Flight Across the Gravity
Continuum
Symposium III: Planetary Biology and Terraforming
Additional Short Papers

iv Gravitational and Space Biology 19(2) August 2006
Table of Contents
Symposium I: Biological Advanced Life Support Systems ………………………..……...…………..1
THE BIOLOGY OF LOW ATMOSPHERIC PRESSURE – IMPLICATIONS FOR EXPLORATION MISSION
DESIGN AND ADVANCED LIFE SUPPORT - A-L. Paul and R. J. Ferl ……………...………………………….…….3
PLANT-GROWTH LIGHTING FOR SPACE LIFE SUPPORT: A REVIEW – G.D. Massa, J.C. Emmerich, R.C.
Morrow, C.M. Bourget and C.A. Mitchell…………………………………………………………………….…………….…19
INCREASED BACTERIAL RESISTANCE AND VIRULENCE IN SIMULATED MICROGRAVITY AND ITS
MOLECULAR BASIS - A. Matin, S.V. Lynch and M.R. Benoit………………………………………….………..…31
Symposium II: Astronaut Health: From the Bench to Flight Across the Gravity Continuum…. 43
EXPLORATION CLASS MISSIONS AND RETURN: EFFECTS ON THE IMMUNE SYSTEM -
G. Sonnenfeld ………………………………………………………………………………………………………………….…..45
NUTRITION, METABOLISM AND THE CRITICAL PATHS: A CRITICAL REVIEW - T.P. Stein ………..….49
TESTING EXERCISE COUNTERMEASURES DURING 30 DAYS OF SIMULATED MICROGRAVITY:
LESSONS LEARNED FROM STUDIES OF IDENTICAL TWINS – A.R. Hargens, B.R. Maclas, C.M. Echon, E.
Brzezinski, A. Hawkins, K. Hawkins, and R.S. Meyer……………………………………………….....................……...…53
TRANSLATIONAL MEDICINE: FROM GROUND-BASED STUDIES OF TRAUMATIC INJURIES TO
ASTRONAUT HEALTH AND EARTH BENEFITS. – C.E. Wade...................................................................….....65
Symposium III: Planetary Biology and Terraforming………………………………………...…….77
PLANETARY BIOLOGY AND TERRAFORMING – P. Todd.................................……................................….....79
LAST PLACE TO BOIL AWAY, FIRST PLACE TO LOOK: THE HUNT FOR WATER AND LIFE ON MARS –
L.H. Kuznetz ……………………………………………………………………………………………………………………….85
EXTREMOPHILES FOR ECOPOIESIS: DESIRABLE TRAITS FOR AND SURVIVABILITY OF PIONEER
MARTIAN ORGANISMS – D.J. Thomas, J. Boling, P.J. Boston, K.A. Campbell, T. McSpadden, L. McWilliams,
and P. Todd ……………………………………………………………………………………………………………….………91
PLANETARY ECOSYNTHESIS AS ECOLOGICAL SUCCESSION – J.M. Graham………………………...…….105
Short Papers…….…………………………………………………………………………………….121
Advanced Life Support and Biotechnology:
DEVELOPMENT OF A MICROFLUIDIC ION SENSOR ARRAY (MISA) TO MONITOR GRAVITY-
DEPENDENT CALCIUM FLUXES IN CERATOPTERIS SPORES - A.R. De Carlo, M. Rokkam, A. ul Haque, S.T.
Wereley, P.P. Irazoqui, H.W. Wells, W.T. McLamb, S.J. Roux, D.M. Porterfield…………………………..…………..123
USE OF AN INTEGRATED FLOW-CHAMBER ADHESION ASSAY FOR MEASURING LEUKOCYTE
ADHESION PROPERTIES IN SIMULATED AND ACTUAL MICROGRAVITY – D.F. Kucik, R.L. Rouleau,
L.W. Smith, X. Wu and K.B. Gupta……………………………………………………………………………………..……..125
DEVELOPMENT OF THE EMCS HARDWARE FOR MULTIGENERATIONAL GROWTH OF DROSOPHILA
MELANOGASTER IN SPACE - M. E. Sanchez, M. Shenasa, A. Maldonado, A. Kakavand, D. Leskovsky, E.
Houston, A. Howard, M. K. Steele, and S. Bhattacharya…………………………………………………….……..……...127

Gravitational and Space Biology 19(2) August 2006 v
INVESTIGATING LOCAL IMPACTS OF HEAT-PULSE SENSORS FOR MEDIA MOISTURE CONTENT -
M.A. Ask, J.J. Prenger, D. Rouzan-Wheeldon, V. Rygalov, J. Norikane and H.G. Levine………………..……………129
PERFORMANCE EVALUATION OF A LABORATORY TEST BED FOR PLANETARY BIOLOGY – N.A.
Thomas, P. Todd, G.W. Metz, M.A. Kurk, D.J. Thomas……………………………………………………….…….…….131
Animal Development, Physiology and Gravity Response:
A STUDY OF THE EFFECTS OF SPACE FLIGHT ON THE IMMUNE RESPONSE IN DROSOPHILA
MELANOGASTER – T.F. Fahlen, M. Sanchez, M. Lera, E. Blazevic, J. Chang, and S. Bhattacharya……..……....133
EFFECTS OF ALTERED GRAVITY ON IDENTIFIED PEPTIDERGIC NEURONS OF THE CRICKET ACHETA
DOMESTICUS – U. Kirschnick, H-J. Agricola and E.R. Horn ………………………………………………..…………135
COUNTERMEASURES TO THE EFFECTS OF GRAVITY ON THE SKULLS OF HUMAN INFANTS - R. Lee,
J. English, J. Duke , and J. Teichgraeber…………………………………………………………………………………….137
NUTRIENT DIFFUSION THROUGH ARTICULAR CARTILAGE: DEVELOPMENT AND USE OF A MODEL
SYSTEM – C. Marshall, R. Flowers, N. Goli, M. Vandromme, D. Paulsen and B. Klement ……………….………..139
HYPERGRAVITY INDUCES DAMAGE TO ROD PHOTORECEPTORS - A.J. Barnstable, A.R. Tink, S. Viviano,
L. Baer, C. Wade, C.J. Barnstable and J. Tombran-Tink……………………………………………………….………….141
Cell Biology:
A GLOBAL TRANSCRIPTIONAL ANALYSIS OF STREPTOCOCCUS PNEUMONIAE IN RESPONSE TO
LOW-SHEAR MODELED MICROGRAVITY – C. Allen, C. Galindo, N. Williams, U. Pandya, A. Chopra, and D.
Niesel ……………………………………………………………………………………………………………..……………….143
PROTEOMIC RETRIEVAL FROM NUCLEIC ACID DEPLETED SPACE-FLOWN HUMAN CELLS - D.K.
Hammond, T.F. Elliott, K. Holubec, T.L. Baker, P.L. Allen, T.G. Hammond and J.E. Love…………….……….……145
THE EFFECT OF CHAGES OF GRAVITY ON HUMAN MONOCYTE CELL (TUR) PHAGOCYTOSIS – C.B.
Johnson, L.S. Waldbeser ………………………………………………………………………………………………….....…147
MICROTUBULE DISRUPTIONS AND REPAIR PHENOMENA IN CULTURED GLIAL CELLS UNDER
MICROGRAVITY – M.A. Masini, F. Strollo, F. Ricci, M. Pastorino, B. M. Uva, ………………………..…………..149
MICROGRAVITY-INDUCED CHANGES IN GENE EXPRESSION IN ACTIVATED T-LYMPHOCYTES
INVOLVE MULTIPLE REGULATORY PATHWAYS – N.E. Ward, N.R. Pellis, S.A. Risin and D. Risin...……..151
POSSIBLE INVOLVEMENT OF FLOW DETECTION IN THE ACTIVATON OF OSTEOBLASTS – M. Takaoki,
H. Park, N. Murakami, D. Shiba, and J-I. Gyotoku ……………………………………………………………………...…153
Plant Development and Gravity Response:
TRANSCRIPTIONAL PROFILING OF THE gps1 MUTANT OF ARABIDOPSIS – V. Nadella, C.D. Hildenbrand
and S.E. Wyatt………………………………………………………………………………..…………………………………..155
NITRIC OXIDE AND CGMP DEPENDENT SIGNALING IN ARABIDOPSIS ROOT GROWTH – J. Jacobi, J.
Elmer, K. Russell, R. Soundur, and D.M. Porterfield.………………………………………………………………………157
REGULATION OF TRANSCRIPTION IN ROOTS OF ARABIDOPSIS GRAVITY MUTANTS – J.W. Yester,
J.M. Kimbrough, R. Salinas-Mondragón, P.H. Masson, C.S. Brown, and H. Winter Sederoff………………………..159
MODIFICATION OF RESERVE DEPOSITION IN WHEAT AND BRASSICA SEEDS BY SYNTHETIC
ATMOSPHERES AND MICROGRAVITY - A. Kuang, J. Blasiak, S. Chen, G. Bingham, M.E. Musgrave..……..161
Index ………………………..……...………………………………………………………………….A-1

vi Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 1
Symposium I: Biological Advanced Life
Support Systems
John Kiss, Editor

2 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 3
THE BIOLOGY OF LOW ATMOSPHERIC PRESSURE – IMPLICATIONS FOR EXPLORATION
MISSION DESIGN AND ADVANCED LIFE SUPPORT
Anna-Lisa Paul and Robert J. Ferl
University of Florida, Gainesville, FL
ABSTRACT
Atmospheric pressure is a variable that has been often
manipulated in the trade space surrounding the design and
engineering of space exploration vehicles and extraterrestrial
habitats. Low pressures were used to reduce structural
engineering and launch mass throughout the early human space
program; moreover, low pressures will certainly be considered
in future concepts for the same reasons. Fundamental
understanding of the biological impact of low pressure
environments is therefore critical for the successful
consideration of this variable, being particularly important when
considering future, potentially complex bioregenerative life
support systems. However, low pressure biological effects are
also critical considerations that should be incorporated into near
term vehicle designs, designs that may set hardware and
operations criteria that would carry over into far-term future
designs.
In order to begin to define the fundamental biological responses
to low atmospheric pressure, we have identified the molecular
genetic responses central to the initial exposure of the model
plant Arabidopsis to hypobaric stress. Less than half of the
genes induced by hypobaria are induced by hypoxia,
establishing that response to hypobaria is a unique biological
response and is more complex than just an adaptation to low
partial pressures of oxygen. In addition, the suites of genes
induced by hypobaria confirm that water movement is a
paramount issue in plants. Current experiments examine gene
expression profiles in response to a wide variety of pressures,
ranging from slight to extreme hypobaria. Results indicate that
even small changes in atmospheric pressure have attendant
biological consequences deserving consideration during the
concept and design of vehicles and habitats. Moreover, the range
of pressures to which plants can adapt suggests that very low
pressures can be considered for plant-specific habitats.
The choices of atmospheric pressure within spaceflight and
extraterrestrial habitats are not merely engineering
considerations but are biological considerations of the highest
order, and modern molecular tools can be employed to increase
understanding of the biological consequences of pressure
engineering decisions.
INTRODUCTION
An almost bewildering myriad of environmental
parameters have been presented to Terran life forms
during the process of evolutionary change. Life has
therefore adapted and colonized a vast variety of
environments and many of those environments contain
extremes of one parameter or another; however, extreme
terrestrial altitude has escaped colonization by higher life
forms because of a suite of parameters that together
prevent habitability. Atmospheric pressure is one of the
major parameters that limit life to lower earth altitudes.
Figure 1. The relationship between altitude and atmospheric
pressure. As the elevation increases from sea level the
atmospheric pressure decreases. The range of the earth’s
atmosphere is 101 kPa at sea level to near 0 kPa at 30,000 m.
The current atmospheric attributes of the Earth’s surface
are the culmination of the physics of our planet and the
impact of over a billion years of biology and geology.
Biology has evolved and expanded into a wide range of
environments, including those that press the limits of
terrestrial altitudes, where physiology is limited by the
extremes of temperature, moisture and atmospheric
pressure that are the intrinsic components of terrestrial
high altitudes (Figure 1). Indeed, it is only where all three
of these extremes converge that we see an absence of life
on our planet. On tropical mountains mammals and large
plants (e.g. hyraxes and giant lobelias of Kilimanjaro) are
found no higher than altitudes of 5000 m (Njiro, 2005)
and herbaceous plants no higher than 5600 m (Körner,
2003). Yet even these altitude limits are dependent upon
other environmental variables. For example, the upper
limit of forests in tropical mountains may be 4000 m, yet
that same limit can be less than 2000 m in more temperate
latitudes (Körner and Paulsen, 2004). In addition, for
mammals and birds, the limits of typical habitation appear
to be delimitated as much by easy access to food and
available oxygen as by temperature, moisture and
atmospheric pressure. For humans, this habitation limit is
around 4200 m in the latitude of the Himalayas, where the
village of Kibber, India, is located (Table 1). In general,
human excursions to higher altitudes and lower pressures
____________________
* Correspondence to: Robert J. Ferl Horticultural Sciences Department
University of Florida
Gainesville, FL 32611-0690
Email: [email protected]
Phone: 352-192-1928x301; Fax: 352-392-4072

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
4 Gravitational and Space Biology 19(2) August 2006
requires supplemental oxygen – though oxygen alone
cannot alleviate all difficulties for humans at low
pressures and high altitudes (Maggiorini et al., 2001;
Bartsch et al., 2005).
In natural environments, plant growth at high altitudes is
more limited by temperature than pressure; the 5600 m
limit on Kilimanjaro is not imposed by the atmospheric
pressure but rather by the fact the ground freezes every
night. Laboratory experiments indicate that if plants are
kept from freezing and are provided with adequate water,
they can be maintained at pressures far less than that
present at the summit of Kilimanjaro (e.g. Mansell et al.,
1968; Gale, 1973; Boston, 1981; Rule and Staby, 1981;
Andre and Richaux, 1986; Musgrave et al., 1988a; Andre
and Massimino, 1992; Daunicht and Brinkjans, 1992;
Ohta et al., 1993; Corey et al., 1996; Iwabuchi et al.,
1996; Corey et al., 2000; Ferl et al., 2002; Goto et al.,
2002; He et al., 2003; Paul et al., 2004). It is only in
artificial environments, such as those of the human
spaceflight program, that atmospheric pressure becomes a
variable independent of the temperature, moisture and gas
composition concerns that accompany terrestrial altitudes.
In the contained, closed, and engineered volumes of
extraterrestrial habitats and vehicles, challenges are
created by the need to contain atmospheric pressure
against the vacuum of space. Hence, atmospheric pressure
has been independently manipulated to levels well beyond
the limits imposed by altitude conditions on Earth.
The idea that plants can be successfully cultured in very
low atmospheric pressures for the purposes of advanced
life support in non-terrestrial environments has serious
implications for attaining the goal of taking humans to
new planetary surfaces. A primary and long-term goal of
sustaining life in remote space locations is to minimize
the amount of mass, and therefore energy, required to
launch and maintain life support systems. Furthermore, if
one considers maximizing the use of local resources, then
it would be desirable to make use of ambient light, which
makes it necessary to have a structure with maximum
transparency. This then leads to the question of what
materials would be both sufficiently transparent and
sufficiently strong to contain a plant growth atmosphere
that would sustain a higher pressure than the near vacuum
present on the Moon or the low pressure atmosphere on
the Martian surface. At present there are no materials that
would be generally accepted as sufficiently transparent,
lightweight, and strong enough to meet all of these criteria
at a full earth normal pressure. However, reducing the
pressure within a plant habitat would consequently reduce
the intrinsic strength required for such structures and
materials, reducing the mass of material that must be
lifted from the Earth’s surface, and potentially allowing
the capture of ambient light as a resource.
In this review we present a brief history of the various
atmospheric pressures and gas compositions that have
been used within the human-habitable vehicles of the
space programs, with an eye toward the possible
atmospheric configurations that might be used in future
vehicles and habitats. This narration is followed by a
general discussion of the uses of low pressure
atmospheres in plant biology applications. These two
threads will be integrated with a discussion of
experiments focused specifically on low pressure
atmospheres in plant space biology applications. We will
then develop an argument that low atmospheric pressures
present a serious environmental challenge to plants, a
challenge that requires an adaptive response and
redirection of metabolic resources. Understanding of this
response is enhanced by analysis of the gene expression
Table 1. Atmospheric pressure relative to altitude – a biological perspective. These data provide a perspective of the habitation of humans,
flora and fauna at increasing altitude and reduced atmospheric pressure.

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 5
changes that take place as plants respond and adapt. Much
like response and adaptation to other environmental
stresses, such understanding can lead to both a definition
of the current limits of terrestrial plants as well as a path
for producing plants with enhanced capacity for growth
and production at low pressures. Such information is
critical for the space program, as atmospheric pressure
directly impacts the designs and operational procedures
that are being considered for future space vehicles and
extraterrestrial habitats, including greenhouses.
ATMOSPHERIC PRESSURES IN SPACE
EXPLORATION
It is a generally accepted fact that reduction in the
pressure differential between an internal and external
environment correlates with reduction in engineering
costs, especially in the form of structural mass. Given that
mass is such a crucial cost variable in launch
considerations, low atmospheric pressure environments
have been utilized throughout the human space programs.
Lower atmospheric pressures simply reduce the masses of
structural components of space vehicles and have the
associated effect of reducing the amount of atmospheric
consumables required for the mission. Historically such
reductions in mass allowed for increased mission lengths
and increased payload masses.
The Mercury, Gemini, and Apollo environments were
operated at 34 kPa. In order to compensate for the
hypoxia attendant with such a low pressure, the gas
composition of the atmosphere was maintained at 100%
oxygen (Baker, 1981; Martin and McCormick, 1992)
(Table 2). While the pure oxygen environment provided a
suitable mitigation of hypoxia for the astronauts, that
atmosphere carried with it the risk of fire. It is worth
noting that recent studies have also begun to elucidate the
negative effects of prolonged exposure to pure oxygen
environments; however, hyperoxia appears to be toxic
only at partial pressures above 30 kPa. It should also be
noted that all Extravehicular Vehicle Activity (EVA)
space suit environments, beginning in the Gemini
program, have been maintained at 26 kPa in order to
minimize the physical effort required for manipulations of
the suit components. This need to operate EVAs at
reduced atmospheric pressure imposes constraints on
atmospheric management that continue to the present day
(see Table 2). The Skylab environment was also operated
at 34 kPa; however, the composition of the atmosphere
was maintained at 70% O2 : 30% N2. This pressure and
composition was a compromise that reduced the
engineering costs as well as the fire risk, yet maintained
astronaut health with regard to hypoxia. EVAs during
Skylab were conducted with minimal transition to the 26
kPa, pure oxygen environment of the EVA suits, so that
no pressure changes occurred within Skylab during EVA
activities or EVA preparations. Anecdotal evidence
indicates that the Skylab astronauts noticed no obvious ill
effects of living for prolonged periods at hypobaric
pressures, save for the attendant cooling effects of rapid
evaporation of water and sweat and the difficulty of
hearing due to poor sound propagation.
Table 2. US Space vehicles and atmospheric pressure. These data provide descriptions of various NASA orbital and transit vehicles with
respect to their internal cabin atmospheres.

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
6 Gravitational and Space Biology 19(2) August 2006
The Space Shuttle, Mir and the International Space
Station environments were and are operated at Earth-
normal pressures near 101 kPa with an Earth-normal gas
mixture of 21% oxygen. Operation at 101 kPa solves
many of the issues of atmospheric pressure and
composition effects on biology. However, there remain
two pressure issues in recent and current operations;
unintentional excursions to low pressures during leaks,
and intentional reductions in pressure to accommodate
EVA activities. Mir suffered several leak events that
seriously impacted operations (Holliman and Aaron,
1997; Zak, 2000), most notably after collision with a
Progress resupply vehicle. While the ISS maintains
pressure in preparation for and during EVAs (requiring
astronauts to do pressure / atmosphere accommodation in
the air lock), the pressure in the Shuttle is lowered to 70
kPa for 24 hrs or more (Figure 2) to accommodate the
acclimation-deacclimation activities that are necessary for
the human transition between 101 kPa / 21% oxygen and
26 kPa / 100% oxygen (Winkler, 1992; Wieland, 1998).
The reduced pressures are employed to facilitate both the
pre-breathing requirements as well as the actual EVA
events.
Figure 2. Pressure profile from the orbiter Endeavour during
the day of the spacewalk EVA on December 10, 2001. The
actual time of the EVA itself was 4 hr 12 min, yet the shuttle
pressure was altered for essentially a full day to accommodate
all of the acclimation for the astronauts to move to and from the
lower pressure, pure oxygen environment of the EVA suits. Data
courtesy of Joe Benjamin, Dynamac, KSC.
PLANTS IN SPACE VEHICLES
Plants have experienced virtually all of the spaceflight
pressure scenarios mentioned above. Seeds were carried
aboard Gemini and Apollo missions, most notably in the
Biostack series of experiments of Apollo and in the Moon
Trees, which were personal effects of astronaut Stu Russa
on Apollo 14. Apparently seeds were also carried by
Astronaut Ed White in his space suit during the first EVA
space walk on Gemini 4. However, in these instances the
exposure was of quiescent seeds, with no studies of those
seeds performed until their return to earth and subsequent
growth at earth normal pressures. Nonetheless, the corn
and Arabidopsis seeds of Biostack germinated and grew,
though with abnormalities generally correlated with hits
of heavy ion radiation. On Skylab, rice plants were
sprouted and grown in a study of tropisms, inadvertently
demonstrating plant germination and development at
reduced atmospheric pressure, within the enhanced
oxygen levels of Skylab (see Table 2 and see also
http://history.nasa.gov/SP-401/ch5.htm). On Mir, plants
were being grown during the Progress collision and
subsequent loss of atmospheric pressure. Perhaps most
importantly, plant experiments have been conducted on
the Space Shuttle on flights where EVA activities
occurred. While many shuttle environmental variables are
replicated in simulation chambers at KSC, atmospheric
pressure is simply not addressed in the simulator ground
controls. Therefore plants have experienced a number of
exposures to spaceflight relevant atmospheric pressures,
but not always in situations where the effects of that
atmospheric pressure might have been noted or
accounted. This concept of unrecognized consequences of
changes in atmospheric pressure extend to several
relevant but non-spaceflight environments, such as the
KC-135 parabolic flight aircraft, which experiences
pressure deltas during each parabola as the plane’s
pressurization system attempts to maintain cabin pressure
during the extreme changes in altitude associated with
parabolic flight (Figure 3).
Figure 3. Pressure profile of a typical KC-135 parabolic flight.
These data were collected by a HOBO datalogger during a
typical life sciences KC-135 flight in January of 2000. These
data are not calibrated and are presented for demonstration
only. Note the pressure drop as the plane takes off and the
pressure stabilizes at a lower pressure characteristic of airliners
at cruise altitude. Note also that each parabola is characterized
by a further transient drop in pressure as the aircraft cabin
pressure system struggles to maintain a constant pressure. The
parabolas occur usually in groups of ten, characterized by the
closely spaced deviations in cabin pressure. The stable areas of
the graph between the groups of parabola pressure deviations
denote the times of constant altitude turns during which the KC-
135 changes direction to orient for the next series of parabolas.

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 7
PLANT PHYSIOLOGY IN HYPOBARIC
ENVIRONMENTS
Although plants are able to survive and even thrive at
pressures well below the threshold of typical human
habitation, it is very likely that most plants would
perceive such hypobaria as a stressful environment
requiring response and adaptation. The likelihood of a
hypobaria stress response is derived from knowledge of
plant responses to other forms of environmental stress,
including hypoxia, cold and dehydration (Ferl et al.,
2002). Yet, since hypobaria, alone, is not a native
terrestrial environment, the nature of the hypobaria stress
response could be entirely unique. Thus, fundamental
understanding of plant responses and adaptability to low
atmospheric pressures is a requisite part of developing
insights into pressure effects on terrestrial life-forms in
general as well as maximizing plant growth under
hypobaric advanced life support conditions. There is a
rich history of low atmospheric pressure research within
the plant biology community, a history that explores the
responses of plants to hypobaria and the adaptations of
plants to related environmental stresses such as hypoxia.
Most early and many recent hypobaria observations were
focused on direct physiological changes and metabolic
impact of low atmospheric pressures on plants (reviewed:
Daunicht and Brinkjans, 1996; Salisbury, 1999; Wheeler
et al., 2001; Corey et al., 2002; Ferl et al., 2002). While
many of the early investigations had a connection to the
international Advanced Life Support (ALS) community,
others explored the effects of low pressure environments
on the post-harvest physiology of fruits, vegetables and
flowers in commercial applications (e.g. Burg, 2004 and
references therein).
One of the earliest hypobaria studies attempted to recreate
a Mars-like atmosphere in bell jars and observed the
effect of that environment on rye seed germination.
Although seeds would germinate in an approximation of
martian atmospheric composition (0.24% CO2, 0.09% O2,
1.39% argon, balance of N2) at normal and slightly
reduced atmospheric pressures (101 kPa and 50 kPa),
lower ranges of pressure (10 kPa and 3 kPa) would not
support germination (Siegel et al., 1962, 1963). Another
early study was conducted by the Air force in 1968 to
explore the utility of using higher plants for food and
atmospheric regeneration in extended human space
missions (Mansell et al., 1968). Since the atmospheres of
the Mercury, Gemini and Apollo spacecraft were kept at
lower atmospheric pressures with elevated partial
pressures of oxygen, it was important to determine
whether suitable plants could also thrive in similar and
related hypobaric conditions. The growth and
development of turnip plants (Brassica rapa) were
evaluated at 50 kPa compared against a control of 93 kPa.
In these experiments, the partial pressure of oxygen was
kept at normoxic conditions of 21 kPa. After 21 days of
growth, it was concluded that there were no adverse
effects seen in the 50 kPa plants, although it was noted
that transpiration rates were elevated compared to the 93
kPa control (Mansell et al., 1968). It is interesting to note
in a historical context how early the spaceflight
community was actively considering the utility of plants
as part of advanced life support systems, under space-
vehicle relevant pressures and gas compositions. These
plant experiments were performed shortly after a series of
animal experiments were conducted in 1965 to determine
the toxicology of the 34 kPa low pressure, pure oxygen
environment being employed for the space capsule
atmosphere (Thomas, 1965). A few years later plants
were also subjected to germination and growth
experiments in a simulated space capsule environment
(Lind, 1971). It can be concluded that these early studies
clearly established the viability of both plants and animals
in low atmospheric pressures and within gas compositions
relevant to space exploration.
In the 1960’s it was also discovered that hypobaric
pressures may have an impact on plant hormone related
physiology, and this realization expanded the interest in
hypobaric research to the commercial fruit industry.
Holding fruit at sub-atmospheric pressures delayed
ripening (Burg and Burg, 1965) and subsequent studies
related this phenomenon to the depletion of ethylene from
fruit tissues that was accelerated by the hypobaric
conditions. Tomatoes, bananas, mangos, cherries, limes
and guavas were incubated in atmospheric pressures
ranging from 48 kPa to 20 kPa and compared to similar
treatments in normal atmospheric pressure (Burg and
Burg, 1966a, 1966b). Later studies used hypobaric
conditions to enable the discrimination between the
effects of ethylene and abscissic acid in the formation of
abscission zones. Bell jars with atmospheres reduced to
20 kPa were used to determine that ethylene is the
primary effector of abscission in citrus fruits (Cooper and
Horanic, 1973). Since these early studies there has been a
wide application of these principles (e.g. Dilley et al.,
1975; Spalding and Reeder, 1976; Lougheed et al., 1978;
Nilsen and Hodges, 1983; Jardine et al., 1984; Kirk et al.,
1986) and this field has been reviewed recently (Burg,
2004).
Investigations into the underlying physiology of growing
plants in hypobaric environments continued to be
explored with both basic physiology and spaceflight
applications in mind, although early experiments
sometimes produced mixed results. For instance, in one
study tomato seedlings grown at 17 kPa were stunted in
growth, while plants grown at 33 kPa were more robust
when compared to the 100 kPa control (Rule and Staby,
1981). Yet in another, tomato plants were uniformly
stunted in reduced atmospheric environments of 40 and
70 kPa compared to 100 kPa control (Daunicht and
Brinkjans, 1992) while the negative effects of extreme
hypobaria (3 kPa) on rye seed germination could be
mitigated with added oxygen (Andre and Richaux, 1986).
Thus, as plant growth experiments explored low pressure
environments, it was found that the composition of the
air, especially with respect to O2 and CO2 can have a
profound effect on the physiological response of the
plants growing in a reduced overall pressure,
complicating the interpretation of the underlying effects

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
8 Gravitational and Space Biology 19(2) August 2006
of low pressure that are independent of gas composition.
The two atmospheric components that contribute most to
complicating a purely hypobaric response are CO2 and O2.
For CO2 related processes, the increase in molecular
diffusion rates of CO2 at lower atmospheric pressures
enhances the ability of plants to take up the gas in
hypobaric environments (Gale, 1972). Thus the decrease
in absolute CO2 concentrations at lower pressures is
counterbalanced by the increase in CO2 diffusion rates. In
natural environments this positive effect could in turn be
counteracted by the decreases in temperature that
accompany reduced atmospheric pressures at high
altitudes (Gale, 1973), as well as by the concomitant
decreases in stomatal aperture, increases in transpiration
rates, and the increases in water exchange demands that
accompany higher elevations and lower pressures (Smith
and Geller, 1979; Kirk et al., 1986; Mott and Parkhurst,
1991; Gale, 2004). It has been suggested that such
demands might impact stomatal distribution and density
in plants adapted to high altitudes (Körner et al., 1986)
and this correlation has even been used to predict
probable attitudes of habitation for fossilized leaves
(McElwain, 2004). However, there is some dispute in this
latter application (Johnson et al., 2005) as stomatal
distribution can be associated with changes in CO2
concentrations independent of altitude (Lake et al., 2001).
In experimental scenarios where temperature and pressure
can be controlled, it appears that, at least for hypobaric
pressures of about 50 kPa and above, the increase in CO2
uptake facilitated by increased diffusion rates of CO2
counterbalances the relative scarcity of the gas (Smith and
Donahue, 1991).
As mentioned in earlier sections, the amendment of
oxygen to hypoxic atmospheres can have a profound
effect on the ability of animals to cope with hypoxic
environments. Oxygen also ameliorates some of the
effects of hypobaric stress in plants. One such study
demonstrated that wheat was capable of germinating and
growing at total atmospheric pressures of 10 kPa, and that
pressures of 20 kPa even enhanced growth when the
partial pressure of nitrogen was kept low by using oxygen
to displace much of the nitrogen in the maintenance of the
total atmospheric pressure. The effect of this replacement
created an atmosphere with a total pressure of 20 kPa, and
partial pressures of O2 at 14 kPa, N2 at 3.4 kPa and CO2 at
3.4 kPa. Interestingly, the positive effects of this
atmosphere on growth were couched in terms of a
reduction of nitrogen rather than an increase of oxygen
(Andre and Massimino, 1992). Indeed, if a pure oxygen
atmosphere is used, rye seeds are capable of germinating
at pressures of 3 kPa (Andre and Richaux, 1986).
However, oxygen does not ameliorate all issues of growth
and development at low pressures. In a study with mung
bean seedlings that compared mitochondrial respiration
rates with overall growth, it was found that while
respiration responded to oxygen concentration
independently of the overall atmospheric pressure down
to 21 kPa, quite the opposite was found for seedling
growth. In this case, growth (as assayed by mass
accumulation and length of seedlings) was negatively
correlated with a decrease in pressure and was
independent of the partial pressure of oxygen (Musgrave
et al., 1988a). Arabidopsis and rice also appear to be
impacted by oxygen concentrations in their ability to cope
with hypobaric conditions. Although both species could
grow at atmospheric conditions of 25 or 50 kPa of total
pressures, the addition of oxygen to a partial pressure of
at least 10 kPa appeared to compensate for any
disadvantageous of growing in a hypobaric environment
(Goto et al., 2002).
Independent of the gas composition in a hypobaric
environment is the more direct effect that low
atmospheric pressures have physically on gas relations in
plant physiology; all other things remaining constant, if
the atmospheric pressure is reduced, the rate of gas
exchange will increase. Evaporation, for example,
increases as pressure is reduced, an effect that can account
at least in part for increased plant transpiration at low
pressures (Rygalov et al., 2002). However, it is not clear
whether all things do remain constant in a plant adapting
to hypobaric conditions. In a short term experiment CO2
assimilation rates and transpiration rates were enhanced
for spinach in 25 kPa environments compared to 101 kPa
controls. However, in long term experiments it was
demonstrated that the gas exchange rates for plants in 25
kPa did not vary from those grown at normal pressures.
One apparent reason for this result was that over time, the
stomatal openings in these plants became correspondingly
smaller, thereby reducing the rate of gas exchange
through these pores (Iwabuchi and Kurata, 2003). Long
term experiments with lettuce at slightly reduced
atmospheric pressure (70 kPa) demonstrated that plants
could adapt with no adverse effects, and even tended to be
slightly more robust than their 101 kPa counterparts
(Spanarkel and Drew, 2002).
These findings suggest that plants appear to respond to
low atmospheric pressures through a fairly complex set of
adaptations, and further, there are situations where the
benefits of hypobaric environments may outweigh the
metabolic cost, in part through effects on the gaseous
plant hormone ethylene. Where plants are cultivated in
closed containers, hypobaric environments of 30 kPa total
pressure appear to offset the detrimental effects of
ethylene and other volatile biological compounds that
normally accumulate in closed systems (He et al., 2003).
Some of these effects appeared to be due to an inhibition
of ethylene production by the low partial pressure of
oxygen, as hypoxia alone was not as uniformly effective
in ethylene management. Although ethylene production
could be inhibited in lettuce grown at 101 kPa with a
partial pressure of oxygen equivalent to that found at 30
kPa of total pressure, the same was not true of wheat.
Ethylene production in wheat could only be controlled
when the overall atmospheric pressure was reduced to 30
kPa (He et al., 2003).

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 9
LOW ATMOSPHERIC PRESSURE HABITATS
From a physiological point of view, plants cope well with
hypobaric atmospheres, and conditions can be engineered
for closed system environments where plants may even
benefit from atmospheric pressures as low as 30 kPa. This
set of circumstances lends itself very well to the physical
engineering needs of vehicles and habitats that
exploration may take to other planetary surfaces (Wheeler
et al., 2001; Corey et al., 2002) and sets the stage for the
genetic engineering to further enhance the production of
plants that may populate such habitats (Ferl et al., 2002).
Low atmospheric environments were proposed for
Martian greenhouses in early concepts that examined
human exploration on Mars (Boston, 1981). It was
thought that low pressure greenhouses would be the most
effective means to grow plants in support of a mission on
a planet that had a vast differential between the external
atmospheres (ca. 0.7 kPa) and the internal environment.
Early experiments to support the concept employed plants
grown in largely CO2 atmospheres at a total pressure of 5
kPa. Radish, alfalfa and mung beans were germinated and
grown for 2 to 4 days with only minimal mortality (10%)
and much of the mortality was due to secondary effects,
such as fungal contamination (Boston, 1981). The idea of
an inflatable structure within a man-made or natural (e.g.
lava tube) rigid structure captured many imaginations and
has endured as a valid model for more than 20 years. The
inflatable greenhouse concept supports the notion that on
a long duration mission to Mars resupply is not an easy
operational option, and any decrease in initial supply
mass enhances launch capabilities (McKay and Toon,
1991; Schwartzkopf and Mancinelli, 1991; Mitchell,
1994; Schwartzkopf, 1997; Kennedy, 1999; Salisbury,
1999; Clawson, 2000; Fowler et al., 2000; Alling et al.,
2002; Corey et al., 2002; Sadler and Giacomelli, 2002;
Hublitz et al., 2004). Habitats have been envisioned that
range from significantly reduced atmospheric pressure
that would require pressure suits and robotic interventions
(Boston, 1981; Clawson, 2000; Corey et al., 2002) to
habitats that are on the edge of human comfort (50 kPa)
that would permit short excursions by crew with minimal
support (Hublitz et al., 2004). Many of these designs
employ transparent inflatable structures to make maximal
use of ambient light (Clawson et al., 2005).
PLANT GENE EXPRESSION IN HYPOBARIC
ENVIRONMENTS
Plants are able to cope with a vast variety of
environmental conditions that differ from a well hydrated
temperate meadow. Plants can engage metabolisms that
enable them to thrive in niches characterized by extremes
in temperature, humidity and oxygen availability. The
responses associated with these stress environments (cold,
desiccation, flooding, heat shock) have been well
characterized; yet until recently, it was not known
whether the response strategies elicited by hypobaria
engaged unique pathways or similar pathways as the
known environmental stresses (Ferl et al., 2002). To
address this question, experiments were conducted with
Arabidopsis to examine the patterns of gene expression on
a genome wide scale as plants were introduced to
hypobaric and comparable hypoxic environments (Paul et
al 2004). Affymetrix GeneChip
8K arrays were used to
characterize the effects of 24 hour exposure of
Arabidopsis seedlings to an atmospheric environment of
10 kPa. The gene expression patterns were then compared
to those of plants exposed to a normal pressure hypoxic
environment of 2% oxygen (the partial pressure of O2 at
10 kPa) and both patterns compared to 24 hour exposure
to Earth-normal air at 101 kPa.
The hypobaric and hypoxic environments were created
within a Low Pressure Growth Chamber (LPGC). The
LPGC controlled lighting, temperature, CO2 partial
pressure and humidity across all experiments such that the
only variables were either atmospheric pressure or oxygen
content. The experimental plants were nine day old
Arabidopsis seedlings grown on nutrient agar plates, a
configuration that ensured that both roots and shoot
received identical atmospheric conditions throughout the
treatment, and that the entire surface of the plant received
the full impact of the environment in which it was placed.
It is also important to note that the relative humidity of
the plant’s microenvironment inside the culture plate
remained at or above 95% in every treatment. Further, all
plants remained turgid and showed no signs of wilting or
desiccation at the end of the 24 hours of treatment.
Of the 8,000 genes represented on the array, more than
200 were differentially expressed in the shoots of plants
exposed to a hypobaric environment of 10 kPa for 24
hours. A comparable number of genes were similarly
differentially expressed in hypoxic treatment - yet only a
fraction of the two sets of differentially expressed genes
overlapped in patterns of expression (Figure 4). Many of
the hallmarks of hypoxic stress were found in both the 10
kPa and 2% O2 treatment, as oxygen is limiting in both
scenarios (groups 3 and 4, as well as members of groups 2
and 5, Figure 4). Genes required to support fermentative
pathways and those involving oxygen transport are
required under low oxygen conditions (e.g. Klok et al.,
2002) so it is not surprising to find representatives of
these metabolisms in both treatments. The genes
repressed by both hypoxia and hypobaria included
examples from a wide range of metabolic pathways that
are globally down-regulated in response to hypoxic stress
(Sachs et al., 1980; Sachs et al., 1996; Vartapetian and
Jackson, 1997; Klok et al., 2002). The hypobaria response
therefore included many of the gene activity changes
typical for a hypoxic response, but the hypobaric response
was not limited to genes involved in hypoxia.
The group of genes that was most highly induced by
hypobaria, yet unaffected by hypoxia, were ones involved
with desiccation-related metabolic pathways. Many of the
genes represented in this group displayed as much as a
30-fold difference in expression between 2% O2 and 10
kPa (see group 1, containing 30 genes, Figure 4). Among
the most abundant genes in this category were cold

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
10 Gravitational and Space Biology 19(2) August 2006
induced genes (e.g. Cor28), dehydrins, which are
typically associated with ABA regulated seed storage
proteins (e.g. LEA) as well as many other examples of
desiccation and ABA regulated genes (Yamaguchi-
Shinozaki and Shinozaki, 1993a; Siddiqui et al., 1998;
Thomashow, 1999; Finkelstein et al., 2002; Soulages et
al., 2003). Also found in this category were genes that are
associated with the regulation and distribution of stomates
in Arabidopsis leaves (Berger and Altmann, 2000) and
many genes associated with the mediation of signal
transduction, especially those related to kinases (Guo et
al., 2002; Yoshida et al., 2002), calcium binding
(Takahashi et al., 2000; Sadiqov et al., 2002) and a
cytochrome p450 (Reddy et al., 2002). There were two
smaller groups of genes that exhibited interestingly
unique patterns of gene expression among the hypoxic
and hypobaric treatments. First, there were a few genes
that were repressed in hypoxia, but were unaffected by
hypobaria. Representatives from this group encoded
proteins similar to those involved in oxygen sensing
processes that are mediated through a heme- or iron-based
sensor (Aravind and Koonin, 2001; Zhu and Bunn, 2001;
Quinn et al., 2002). Second, there were those that were
induced in hypoxic environments, while remaining
unaffected by hypobaria (some of the genes in group 6 of
Figure 4). Genes encoding heme-related proteins were
also found in this group, as well as genes typical of a
hypoxic stress response. The dissimilarity in gene
expression patterns in what might be considered related
oxygen sensing systems, suggests that that hypobaria and
hypoxia may differentially activate and repress certain
alternative oxygen sensing and transport pathways.
Figure 4. Differential expression of Arabidopsis genes in response to hypobaria (10 kPa) and hypoxia (2% O2). Over 200 genes of the
8,000 of the Affymetrix Arabidopsis Gene Chip© were differentially expressed in the shoots of plants exposed to a hypobaric environment of
10 kPa or a hypoxic treatment of 2% O2 for 24 hours. The panel on the left contains three columns of stacked, colored lines. Each line
represents one gene that exhibits differential expression with regard to the Earth Normal (101 kPa / 21% O2) control. When a gene is
induced relative to the control it is indicated in red, when it is repressed it is indicated in green. The reference column of normal sea level
pressure looks entirely black as all expression is calculated relative to the values therein. The two columns to the right (hypoxia and
hypobaria) display patterns of differential gene expression and are clustered into 7 groups of gene exhibiting similar response. The cluster
number and the number of genes represented in that cluster are indicated between the two panels. The graph on the right displays the
relative normalized gene expression averages (Paul et al., 2004) for the groups of genes within the group for hypoxic (open bars) and
hypobaric (closed bars) treatments. The color version of this figure can be found on line: http://asgsb.org/publications.html ). In the
grayscale print version, induced appears as a darker gray and repressed as a lighter gray.
A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 11
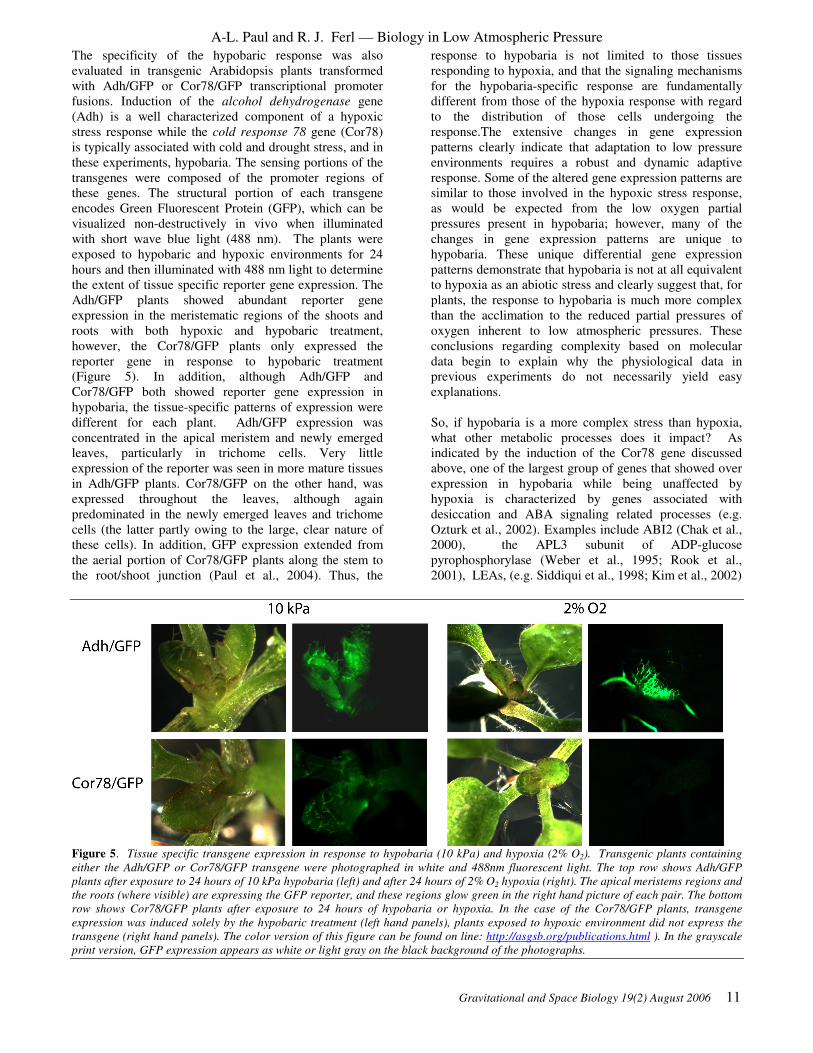
The specificity of the hypobaric response was also
evaluated in transgenic Arabidopsis plants transformed
with Adh/GFP or Cor78/GFP transcriptional promoter
fusions. Induction of the alcohol dehydrogenase gene
(Adh) is a well characterized component of a hypoxic
stress response while the cold response 78 gene (Cor78)
is typically associated with cold and drought stress, and in
these experiments, hypobaria. The sensing portions of the
transgenes were composed of the promoter regions of
these genes. The structural portion of each transgene
encodes Green Fluorescent Protein (GFP), which can be
visualized non-destructively in vivo when illuminated
with short wave blue light (488 nm). The plants were
exposed to hypobaric and hypoxic environments for 24
hours and then illuminated with 488 nm light to determine
the extent of tissue specific reporter gene expression. The
Adh/GFP plants showed abundant reporter gene
expression in the meristematic regions of the shoots and
roots with both hypoxic and hypobaric treatment,
however, the Cor78/GFP plants only expressed the
reporter gene in response to hypobaric treatment
(Figure 5). In addition, although Adh/GFP and
Cor78/GFP both showed reporter gene expression in
hypobaria, the tissue-specific patterns of expression were
different for each plant. Adh/GFP expression was
concentrated in the apical meristem and newly emerged
leaves, particularly in trichome cells. Very little
expression of the reporter was seen in more mature tissues
in Adh/GFP plants. Cor78/GFP on the other hand, was
expressed throughout the leaves, although again
predominated in the newly emerged leaves and trichome
cells (the latter partly owing to the large, clear nature of
these cells). In addition, GFP expression extended from
the aerial portion of Cor78/GFP plants along the stem to
the root/shoot junction (Paul et al., 2004). Thus, the
response to hypobaria is not limited to those tissues
responding to hypoxia, and that the signaling mechanisms
for the hypobaria-specific response are fundamentally
different from those of the hypoxia response with regard
to the distribution of those cells undergoing the
response.The extensive changes in gene expression
patterns clearly indicate that adaptation to low pressure
environments requires a robust and dynamic adaptive
response. Some of the altered gene expression patterns are
similar to those involved in the hypoxic stress response,
as would be expected from the low oxygen partial
pressures present in hypobaria; however, many of the
changes in gene expression patterns are unique to
hypobaria. These unique differential gene expression
patterns demonstrate that hypobaria is not at all equivalent
to hypoxia as an abiotic stress and clearly suggest that, for
plants, the response to hypobaria is much more complex
than the acclimation to the reduced partial pressures of
oxygen inherent to low atmospheric pressures. These
conclusions regarding complexity based on molecular
data begin to explain why the physiological data in
previous experiments do not necessarily yield easy
explanations.
So, if hypobaria is a more complex stress than hypoxia,
what other metabolic processes does it impact? As
indicated by the induction of the Cor78 gene discussed
above, one of the largest group of genes that showed over
expression in hypobaria while being unaffected by
hypoxia is characterized by genes associated with
desiccation and ABA signaling related processes (e.g.
Ozturk et al., 2002). Examples include ABI2 (Chak et al.,
2000), the APL3 subunit of ADP-glucose
pyrophosphorylase (Weber et al., 1995; Rook et al.,
2001), LEAs, (e.g. Siddiqui et al., 1998; Kim et al., 2002)
Figure 5. Tissue specific transgene expression in response to hypobaria (10 kPa) and hypoxia (2% O2). Transgenic plants containing
either the Adh/GFP or Cor78/GFP transgene were photographed in white and 488nm fluorescent light. The top row shows Adh/GFP
plants after exposure to 24 hours of 10 kPa hypobaria (left) and after 24 hours of 2% O2 hypoxia (right). The apical meristems regions and
the roots (where visible) are expressing the GFP reporter, and these regions glow green in the right hand picture of each pair. The bottom
row shows Cor78/GFP plants after exposure to 24 hours of hypobaria or hypoxia. In the case of the Cor78/GFP plants, transgene
expression was induced solely by the hypobaric treatment (left hand panels), plants exposed to hypoxic environment did not express the
transgene (right hand panels). The color version of this figure can be found on line: http://asgsb.org/publications.html ). In the grayscale
print version, GFP expression appears as white or light gray on the black background of the photographs.

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
12 Gravitational and Space Biology 19(2) August 2006
dehydrins and rab-like proteins, (e.g. Mantyla et al., 1995;
Nylander et al., 2001; Kizis and Pages, 2002) rd29B and
cor15B (Wilhelm and Thomashow, 1993; Yamaguchi-
Shinozaki and Shinozaki, 1993b) and aldehyde
dehydrogenase (Kirch et al., 2001). Thus, it appears that a
primary impact of a hypobaric environment, independent
of oxygen stress, is on the mechanisms by which a plant
perceives and responds to water movement or desiccation.
It is important to emphasize that it is the perception
mechanism that is being impacted, not the actual
desiccation process. The plants in these experiments were
grown in a humid environment (>95% rh within the
plates), showed no loss of fresh weight or turgor, and yet
still responded to 10 kPa as if they were dehydrated or in
the process of dehydration. This pivotal result indicates
that the desiccation response is likely due to the
perception of increased water flux caused by the low
pressure environment rather than actual loss of water
content within the plants.
Thus the analysis of the differential gene expression
patterns from these experiments lead to two fundamental
conclusions: first, that hypobaria does not equal hypoxia,
and second, that the primary metabolic pathways that are
engaged by hypobaric stress include those that encompass
the desiccation response or otherwise touch ABA
mediated metabolisms. The response to hypobaria is
much more complex than the acclimation to low partial
pressures of oxygen. Therefore, it would be predicted that
complete compensation for hypobaria in plants would not
be accomplished by increasing the oxygen content, as was
done for low pressure environments of the Mercury,
Gemini and Apollo vehicles. The desiccation-related
response of Arabidopsis plants at 10 kPa in the absence of
real desiccation or wilting also raises the question as to
whether adaptation to a low atmospheric pressure actually
requires the activation of desiccation related pathways, or
if the induction of desiccation-related metabolism may
not be necessary for survival. If the desiccation response
is not necessary for adaptation to low pressure, then the
inappropriate induction of these pathways could represent
costs to production and genetic engineering to remove the
response may be beneficial. If the desiccation response is
indeed required, then enhancing the desiccation response
may increase production at low atmospheric pressure.
CONCLUSION
The experiments and conclusions discussed here expose
the need for continued evaluation of the effects of low
pressure on biological systems. In some ways, the gene
expression data suggest that we actually have only a small
fraction of the data necessary to make informed choices
on the gas composition and pressures within vessels that
support plant growth. By extension, these data also beg
the questions of what sort of undetected responses and
adaptations to low pressures occur in other organisms,
including humans, and whether low pressure
environments have contributed to any of the existing
spaceflight data on biology and biological responses in
spaceflight environments. There is little data available on
the response of any animal system to hypobaric
environments at the level of gene expression, but there are
studies that explore the physiological responses of
animals to hypobaric environments. Of particular interest
are those that indicate hypobaric environments could
contribute to compromised health and resistance to
disease. Early ground-based studies in mice demonstrated
that hypobaric environments, even when supplemented
with oxygen, can lead to stimulated growth of
gastrointestinal bacteria (Gillmore and Gordon, 1975),
retard healing of staphylococcus skin infections (Schmidt
et al., 1967), increase mortality in animals with
intraperitoneal infections (Ball and Schmidt, 1968) and
depress interferon levels (Huang and Gordon, 1968).
More recently, studies have linked hypobaric
environments with an impaired immune response in rats
(SaiRam et al., 1998) mice (Biselli et al., 1991) and
humans (Meehan et al., 1988; Facco et al., 2005). Rarely
has hypobaria been studied independently from hypoxia,
but a recent study examining the effects of mild hypobaria
on fluid balance issues conducted hypobaric treatments in
normoxic atmospheres to control for the effects of
hypoxia. They concluded that although there is a
synergistic effect of hypoxic and hypobaric stress at
altitude, hypobaria alone can adversely impact the fluid
and ionic balance of humans (Loeppky et al., 2005).
So the short term questions surrounding low pressure
spaceflight and extraterrestrial environments involve
deepening our understanding of responses and adaptation
to hypobaria for all organisms and biological systems that
may be a part of spaceflight and extraterrestrial habitats.
Indeed, unpublished molecular data indicate that even the
relatively small pressure drops to 75 kPa that occur in the
space shuttle effect the expression of hundreds of genes in
Arabidopsis. That being the case, even seemingly small
choices about operations and procedures conducted within
habitat environments can have measurable and dramatic
impacts on the biology of those habitats. Therefore very
near term decisions involving, for example, the choices of
atmospheric composition and pressure in the Crew
Exploration Vehicle will have long term implications for
biology. In the longer term, the use of low pressure
environments may enable life support systems that
otherwise would be impossible to construct or maintain
under higher atmospheric pressures, keeping viable the
concepts of inflatable greenhouses. For example,
transparent greenhouses could be erected on Mars using
currently available materials, but only if the internal
pressure of the greenhouse could be maintained below 7.5
kPa (Boston, 1981). The rather dramatic changes in gene
expression that occur at 10 kPa suggest that response and
adaptation to lower pressures will be even more
extensive, but the study of such pressures are well within
the capacity of current experiment chamber design and
molecular expression technology.

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 13
ACKNOWLEDGEMENTS
The spaceflight and low pressure experiments of the Ferl
laboratory were supported by NASA grants NAG 10-316,
NAG 10-291 and AO-99-HEDS-01-032. The authors
wish to recognize the tremendous contributions of the
many scientists whose work is summarized in this review.
In particular, we thank Andrew Schuerger, Jeff Richards,
Ken Corey, Ray Bucklin, Ray Wheeler, Vadim Rygolov,
Phil Fowler and other folks involved in developing the
low pressure facilities at KSC. We thank Mike Dixon and
his associates at the University of Guelph and the
Controlled Environment Systems Research Facility
(CESRF). We thank Bill Wells, Joe Benjamin and Kelly
Norwood from KSC for providing source material
regarding shuttle and KC-135 atmospheric pressures. We
also thank our colleagues at UF who participated in low
pressure and recent KC-135 experiments, including
Jordan Barney, Matt Reyes, Bill Gurley, Mike Manak and
John Mayfield.
The authors also wish to acknowledge the difficulty of
deriving original references for some of the discussions
presented in this paper. Any mistakes or
misrepresentations or misattributions are regretted and we
would welcome any comments, corrections or any
additional references at [email protected] or [email protected].
This paper is dedicated to the memory of Guy Etheridge.
His appreciation of science and love of space research had
such a wonderful influence on all.
REFERENCES
Alling, A., Nelson, M., Silverstone, S., and Van Thillo,
M. 2002. Human factor observations of the Biosphere 2,
1991-1993, closed life support human experiment and its
application to a long-term manned mission to Mars. Life
Support Biosph Sci 8: 71-82
Andre, M., and Massimino, D. 1992. Growth of plants at
reduced pressures: Experiments in Wheat -Technological
advantages and constraints. Adv. Space Res. 12: 97-106
Andre, M., and Richaux, C. 1986. Can plants grow in
quasi-vacuum? In: CELSS 1985 Workshop, Ames
Research Center. NASA Publication TM 88215: 395-404
Aravind, L., and Koonin, E.V. 2001. The DNA-repair
protein AlkB, EGL-9, and leprecan define new families of
2-oxoglutarate- and iron-dependent dioxygenases.
Genome Biol 2: 007
Baker, D. 1981. The History of Spaceflight. Crown
Publishers, Inc., New York
Ball, R.J., and Schmidt, J.P. 1968. Mortality of altitude-
exposed mice infected with Pasteurella tularensis. Appl
Microbiol 16: 1451-1453
Bartsch, P., Mairbaurl, H., Maggiorini, M., and Swenson,
E.R. 2005. Physiological aspects of high-altitude
pulmonary edema. J Appl Physiol 98: 1101-1110
Berger, D., and Altmann, T. 2000. A subtilisin-like serine
protease involved in the regulation of stomatal density
and distribution in Arabidopsis thaliana. Genes Dev. 14:
1119-1131
Biselli, R., Le Moli, S., Matricardi, P.M., Farrace, S.,
Fattorossi, A., Nisini, R., and D'Amelio, R. 1991. The
effects of hypobaric hypoxia on specific B cell responses
following immunization in mice and humans. Aviat Space
Environ Med 62: 870-874
Boston, P.J. 1981. Low-pressure greenhouses and plants
for a manned research station on Mars. J. British
Interplanetary Soc. 54: 189-192
Burg, S.P. 2004. Postharvest Physiology and Hypobaric
Storage of Fresh Produce, Ed first edition. Oxford
University Press
Burg, S.P., and Burg, E.A. 1965. Ethylene Action and the
Ripening of Fruits. Science 148: 1190-1196
Burg, S.P., and Burg, E.A. 1966a. Fruit storage at
subatmospheric pressures. Science 153: 314-315
Burg, S.P., and Burg, E.A. 1966b. The interaction
between auxin and ethylene and its role in plant growth.
Proc Natl Acad Sci U S A 55: 262-269
Chak, R.K., Thomas, T.L., Quatrano, R.S., and Rock,
C.D. 2000. The genes ABI1 and ABI2 are involved in
abscisic acid- and drought-inducible expression of the
Daucus carota L. Dc3 promoter in guard cells of
transgenic Arabidopsis thaliana (L.) Heynh. Planta 210:
875-883
Clawson, J.M., ed (2000) Development of an inflatable
greenhouse for a modular-crop production system., Vol
NASA TM 2000-208577.
Clawson, J.M., Hoehn, A., and Wheeler, R. 2005.
Inflatable Transparent Structures for Mars Greenhouse
Applications. SAE International. 2005-01-2845
Cooper, W.C., and Horanic, G. 1973. Induction of
abcission at hypobaric pressures. Plant Physiol 51: 1002-
1004
Corey, K.A., Barta, D.J., and Wheeler, R.M. 2002.
Toward Martian agriculture: responses of plants to
hypobaria. Life Support Biosph Sci 8: 103-114
Corey, K.A., Bates, M.E., and Adams, S.L. 1996. Carbon
dioxide exchange of lettuce plants under hypobaric
conditions. Adv. Space Res. 18: 265-272

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
14 Gravitational and Space Biology 19(2) August 2006
Corey, K.A., Fowler, P.A., and Wheeler, R.M. 2000.
Water flux from lettuce plants at reduced atmospheric
pressure. HortScience 35
Daunicht, H., and Brinkjans, H. 1996. Plant responses to
reduced air pressure: advanced techniques and results.
Adv Space Res 18: 273-281
Daunicht, H.J., and Brinkjans, H.J. 1992. Gas exchange
and growth of plants under reduced air pressure. Adv.
Space Res. 12: 107-114
Dilley, D.R., Carpenter, W.J., and Burg, S.P. 1975.
Principles and application of hypobaric storage of cut
flowers. Acta Hort. 41: 249-268
Facco, M., Zilli, C., Siviero, M., Ermolao, A., Travain,
G., Baesso, I., Bonamico, S., Cabrelle, A., Zaccaria, M.,
and Agostini, C. 2005. Modulation of immune response
by the acute and chronic exposure to high altitude. Med
Sci Sports Exerc 37: 768-774
Ferl, R.J., Schuerger, A.C., Paul, A.L., Gurley, W.B.,
Corey, K., and Bucklin, R. 2002. Plant adaptation to low
atmospheric pressures: potential molecular responses.
Life Support Biosph Sci 8: 93-101
Finkelstein, R.R., Gampala, S.S., and Rock, C.D. 2002.
Abscisic acid signaling in seeds and seedlings. Plant Cell
14 Suppl: S15-45
Fowler, P.A., Wheeler, R.M., Bucklin, R.A., and Corey,
K.A. 2000. Low pressure greenhouse concepts for Mars.
Inflatable Greenhouse Workshop. NASA TM 2000-
208577: 116-123
Gale, J. 1972. Availability of carbon dioxide for
photosynthesis at high altitudes: theoretical
considerations. Ecology 53: 494-497
Gale, J. 1973. Experimental evidence for the effect of
barometric pressure on photosynthesis and transpiration.
In: Plant Responses to Climatic Factors. Proceedings of
the Uppsala Symposium, UNESCO, Paris, : 289-294
Gale, J. 2004. Plants and altitude--revisited. Ann Bot
(Lond) 94: 199
Gillmore, J.D., and Gordon, F.B. 1975. Effect of
exposure to hyperoxic, hypobaric, and hyperbaric
environments on concentrations of selected and aerobic
and anaerobic fecal flora of mice. Appl Microbiol 29:
358-367
Goto, E., Arai, Y., and Omasa, K. 2002. Growth and
development of higher plants under hypobaric conditions.
SAE Technical Paper 2002-01-2439
Guo, Y., Xiong, L., Song, C.P., Gong, D., Halfter, U.,
and Zhu, J.K. 2002. A calcium sensor and its interacting
protein kinase are global regulators of abscisic acid
signaling in Arabidopsis. Dev Cell 3: 233-244
He, C., Davies, F.T., Jr., Lacey, R.E., Drew, M.C., and
Brown, D.L. 2003. Effect of hypobaric conditions on
ethylene evolution and growth of lettuce and wheat. J
Plant Physiol 160: 1341-1350
Holliman, J., and Aaron, B. 1997. Mir at half power after
collision. In CNN Sci-Tech,
Huang, K.Y., and Gordon, F.B. 1968. Production of
interferon in mice: effect of altered gaseous environments.
Appl Microbiol 16: 1551-1556
Hublitz, I., Henninger, D.L., Drake, B.G., and Eckart, P.
2004. Engineering concepts for inflatable Mars surface
greenhouses. Advances in Space Research 34: 1546-1551
Iwabuchi, K., and Kurata, K. 2003. Short-term and long-
term effects of low total pressure on gas exchange rates of
spinach. Advances in Space Research 31: 241-244
Iwabuchi, K., Saito, G., Goto, E., and Takakura, T. 1996.
Effect of vapor pressure deficit on spinach growth under
hypobaric conditions. Acta Hortic 440: 60-63
Jardine, D.J., Dilley, D.R., and Price, H.C. 1984.
Hypobaric storage of celery transplants. HortScience 19:
869-870
Johnson, D.M., Smith, W.K., and Silman, M.R. 2005.
Climate-independent paleoaltimetry using stomatal
density in fossil leaves as a proxy for CO2 partial
pressure: Comment and Reply. Geology on-line Forum
http://www.gsajournals.org/i0091-7613-31-6-e82.pdf
Kennedy, K.J. 1999. Inflatable habitats and greenhouse
design: technology and development for Mars
implementation. In Inflatable Greenhouse Workshop,
Kennedy Space Center, Florida
Kim, S.Y., Ma, J., Perret, P., Li, Z., and Thomas, T.L.
2002. Arabidopsis ABI5 Subfamily Members Have
Distinct DNA-Binding and Transcriptional Activities.
Plant Physiol. 130: 688-697
Kirch, H.-H., Nair, A., and Bartels, D. 2001. Novel
ABA- and dehydration-inducible aldehyde dehydrogenase
genes isolated from the resurrection plant Craterostigma
plantagineum and Arabidopsis thaliana. Plant J 28: 555-
567
Kirk, H.G., Andersen, A.S., Veierskov, B., Johansen, E.,
and Aabrandt, Z. 1986. Low-pressure storage of hibiscus
cuttings - effect on stomatal opening and rooting. Annals
of Botany 58: 389-396
Kizis, D., and Pages, M. 2002. Maize DRE-binding
proteins DBF1 and DBF2 are involved in rab17 regulation

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 15
through the drought-responsive element in an ABA-
dependent pathway. Plant J 30: 679-689
Klok, E.J., Wilson, I.W., Wilson, D., Chapman, S.C.,
Ewing, R.M., Somerville, S.C., Peacock, W.J., Dolferus,
R., and Dennis, E.S. 2002. Expression profile analysis of
the low-oxygen response in Arabidopsis root cultures.
Plant Cell 14: 2481-2494
Körner, C. 2003. Alpine plant life, Ed 2nd edition,
Heidelberg
Körner, C., Bannister, P., and Mark, A.F. 1986.
Altitudinal variation in stomatal conductance, nitrogen
content and leaf anatomy in different plant life forms in
New Zealand. Oecologia 69: 577–588.
Körner, C., and Paulsen, J. 2004. A world-wide study of
high altitude treeline temperatures. Journal of
Biogeography 31: 713-732
Lake, J.A., Quick, W.P., Beerling, D.J., and Woodward,
F.I. 2001. Plant development. Signals from mature to new
leaves. Nature 411: 154
Lind, C.T. 1971. Germination and growth of selected
higher plants in a simulated space cabin environment.
Wright-Patterson AFB, Ohio AMRL-TR-70-121
Loeppky, J.A., Roach, R.C., Maes, D., Hinghofer-
Szalkay, H., Roessler, A., Gates, L., Fletcher, E.R., and
Icenogle, M.V. 2005. Role of hypobaria in fluid balance
response to hypoxia. High Alt Med Biol 6: 60-71
Lougheed, E.C., Murr, D.P., and Berard, L. 1978. Low
pressure storage for horticultural crops. HortScience 13:
21-27
Maggiorini, M., Melot, C., Pierre, S., Pfeiffer, F., Greve,
I., Sartori, C., Lepori, M., Hauser, M., Scherrer, U., and
Naeije, R. 2001. High-altitude pulmonary edema is
initially caused by an increase in capillary pressure.
Circulation 103: 2078-2083
Mansell, R.L., Rose, G.W., Richardson, B., and Miller,
R.L. 1968. Effects of prolonged reduced pressure on the
growth and nitrogen content of turnip (Brassica rapa L.).
SAM-TR-68-100. Tech Rep SAM-TR: 1-13
Mantyla, E., Lang, V., and Palva, E. 1995. Role of
Abscisic Acid in Drought-Induced Freezing Tolerance,
Cold Acclimation, and Accumulation of LT178 and
RAB18 Proteins in Arabidopsis thaliana. Plant Physiol
107: 141-148
Martin, C.E., and McCormick, A.K. 1992. Air handling
and atmosphere conditioning systems for manned
spacecraft. ICES Paper No. 921350. SAE International.
McElwain, J.C. 2004. Climate-independent paleoaltimetry
using stomatal density in fossil
leaves as a proxy for CO2 partial pressure. Geology 32:
1017-1020
McKay, C.P., and Toon, O.B. 1991. Making Mars
habitable. Nature 352: 489-496
Meehan, R., Duncan, U., Neale, L., Taylor, G.,
Muchmore, H., Scott, N., Ramsey, K., Smith, E., Rock,
P., Goldblum, R., and et al. 1988. Operation Everest II:
alterations in the immune system at high altitudes. J Clin
Immunol 8: 397-406
Mitchell, C.A. 1994. Bioregenerative life-support
systems. Am J Clin Nutr 60: 820S-824S
Mott, K.A., and Parkhurst, D.F. 1991. Stomatal responses
to humidity in air and helox. Plant Cell Environ. 14: 509-
515
Musgrave, M.E., Gerth, W.A., Scheld, H.W., and Strain,
B.R. 1988a. Growth and mitochondrial respiration of
mungbeans (Phaseolus aureus Roxb.) germinated at low
pressure. Plant Physiol 86: 19-22
Nilsen, K.N., and Hodges, C.F. 1983. Hypobaric Control
of Ethylene-Induced Leaf senescence in Intact Plants of
Phaseolus vulgaris L. Plant Physiol 71: 96-101
Njiro, E.I. 2005. The Montane Ecosystems:
Characteristics and conservation. In Mtn-Forum On-Line
Library Document, Vol
http://www.mtnforum.org/resources/library/njire97a.htm.
University of Nairobi, Institute of African Studies,
Nairobi-Kenya
Nylander, M., Svensson, J., Palva, E.T., and Welin, B.V.
2001. Stress-induced accumulation and tissue-specific
localization of dehydrins in Arabidopsis thaliana. Plant
Mol Biol 45: 263-279
Ohta, H., Goto, E., Takakura, T., Takagi, F., Hirosawa,
Y., and Takagi, K. 1993. Measurement of photosynthetic
and transpiration rates under low total pressures.
American Society of Agricultural Engineering Paper No.
934009
Ozturk, Z.N., Talame, V., Deyholos, M., Michalowski,
C.B., Galbraith, D.W., Gozukirmizi, N., Tuberosa, R.,
and Bohnert, H.J. 2002. Monitoring large-scale changes
in transcript abundance in drought- and salt-stressed
barley. Plant Mol Biol 48: 551-573
Paul, A.L., Schuerger, A.C., Popp, M.P., Richards, J.T.,
Manak, M.S., and Ferl, R.J. 2004. Hypobaric biology:
Arabidopsis gene expression at low atmospheric pressure.
Plant Physiol 134: 215-223
Quinn, J.M., Eriksson, M., Moseley, J.L., and Merchant,
S. 2002. Oxygen deficiency responsive gene expression in
Chlamydomonas reinhardtii through a copper-sensing
signal transduction pathway. Plant Physiol. 128: 463-471

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
16 Gravitational and Space Biology 19(2) August 2006
Reddy, A.R., Ramakrishna, W., Sekhar, A.C., Ithal, N.,
Babu, P.R., Bonaldo, M.F., Soares, M.B., and Bennetzen,
J.L. 2002. Novel genes are enriched in normalized cDNA
libraries from drought-stressed seedlings of rice (Oryza
sativa L. subsp. indica cv. Nagina 22). Genome 45: 204-
211
Rook, F., Corke, F., Card, R., Munz, G., Smith, C., and
Bevan, M.W. 2001. Impaired sucrose-induction mutants
reveal the modulation of sugar-induced starch
biosynthetic gene expression by abscisic acid signalling.
Plant J 26: 421-433
Rule, D.E., and Staby, G.L. 1981. Growth of tomato
seedlings at sub-atmospheric pressures. HortScience 16:
331-332
Rygalov, V.Y., Fowler, P.A., Metz, J.M., Wheeler, R.,
and Bucklin, R. 2002. Water cycles in closed ecological
systems: effects of atmospheric pressure. Life Support
Biosph Sci 8
Sachs, M.M., Freeling, M., and Okimoto, R. 1980. The
anaerobic proteins of maize. Cell 20: 761-767
Sachs, M.M., Subbaiah, C.C., and Saab, I.N. 1996.
Anaerobic gene expression and flooding tolerance in
maize. J. Exp Bot 47: 1-15
Sadiqov, S.T., Akbulut, M., and Ehmedov, V. 2002. Role
of Ca2+ in drought stress signaling in wheat seedlings.
Biochemistry (Mosc) 67: 491-497
Sadler, P.D., and Giacomelli, G.A. 2002. Mars inflatable
greenhouse analog. Life Support Biosph Sci 8: 115-123
SaiRam, M., Sharma, S.K., Dipti, P., Pauline, T., Kain,
A.K., Mongia, S.S., Bansal, A., Patra, B.D., Ilavazhagan,
G., Devendra, K., and Selvamurthy, W. 1998. Effect of
hypobaric hypoxia on immune function in albino rats. Int
J Biometeorol 42: 55-59
Salisbury, F.B. 1999. Growing crops for space explorers
on the moon, Mars, or in space. Adv Space Biol Med 7:
131-162
Schmidt, J.P., Cordaro, J.T., and Ball, R.J. 1967. Effect
of Environment on Staphylococcal Lesions in Mice. Appl
Microbiol 15: 1465-1467
Schwartzkopf, S.H. 1997. Human life support for
advanced space exploration. Adv Space Biol Med 6: 231-
253
Schwartzkopf, S.H., and Mancinelli, R.L. 1991.
Germination and growth of wheat in simulated Martian
atmospheres. Acta Astronaut. 25: 245-247
Siddiqui, N.U., Chung, H.J., Thomas, T.L., and Drew,
M.C. 1998. Abscisic acid-dependent and -independent
expression of the carrot late-embryogenesis-abundant-
class gene Dc3 in transgenic tobacco seedlings. Plant
Physiol 118: 1181-1190
Siegel, S.M., Rosen, L.A., and Giumarro, C. 1962.
Effects of reduced oxygen tension on vascular plants -
Winter Rye germination under near-martian conditions
and in other non-terrestrial environments. Proc Natl Acad
Sci U S A 48: 725-728
Siegel, S.M., Rosen, L.A., and Giumarro, C. 1963. Plants
at sub-atmospheric oxygen levels. Nature 198: 1288-1289
Smith, W.K., and Donahue, R. 1991. Simulated effect of
altitude on photosynthetic CO2 uptake potential in plants.
Plant, Cell & Environment 14
Smith, W.K., and Geller, G.N. 1979. Plant transpiration
at high altitudes: theory, field measurements and
comparisons with desert plants. Oecologia 41: 109-122
Soulages, J.L., Kim, K., Arrese, E.L., Walters, C., and
Cushman, J.C. 2003. Conformation of a Group 2 Late
Embryogenesis Abundant Protein from Soybean.
Evidence of Poly (L-Proline)-type II Structure. Plant
Physiol. 131: 963-975
Spalding, D.H., and Reeder, W.F. 1976. Low pressure
storage of avocados. HortScience 11: 491-492
Spanarkel, R., and Drew, M.C. 2002. Germination and
growth of lettuce (Lactuca sativa) at low atmospheric
pressure. Physiol Plant 116: 468-477
Takahashi, S., Katagiri, T., Yamaguchi-Shinozaki, K.,
and Shinozaki, K. 2000. An Arabidopsis gene encoding a
Ca2+-binding protein is induced by abscisic acid during
dehydration. Plant Cell Physiol 41: 898-903
Thomas, A.A. 1965. Low ambient pressure environments
and toxicity - a new approach to space cabin toxicology.
Arch. Environ. Health 11: 316-322
Thomashow, M.F. 1999. Plant cold acclimation: freezing
tolerance genes and regulatory mechanisms. Ann. Rev.
Plant Physiol. Mol. Biol. 50: 571-599
Vartapetian, B.B., and Jackson, M.B. 1997. Plant
adaptations to anaerobic stress. Anals of Botany 79: 3-20
Weber, H., Heim, U., Borisjuk, L., and Wobus, U. 1995.
Cell-type specific, coordinate expression of two ADP-
glucose pyrophosphorylase genes in relation to starch
biosynthesis during seed development of Vicia faba L.
Planta 195: 352-361
Wheeler, R., Stutte, G., Subbarao, G.V., and Yorio, N.C.
2001. Plant growth and human life support for space
travel, Ed 2nd. Marcel Dekker, New York

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
Gravitational and Space Biology 19(2) August 2006 17
Wieland, P.O. 1998. Living together in space: The design
and operation of the life support systems on the
International Space Station. NASA TM 1998-206956. 1
Wilhelm, K.S., and Thomashow, M.F. 1993. Arabidopsis
thaliana cor15b, an apparent homologue of cor15a, is
strongly responsive to cold and ABA, but not drought.
Plant Mol Biol 23: 1073-1077
Winkler, H.E. 1992. Shuttle Orbiter ECLSS Flight
Experience. ICES Paper No. 921348. SAE International.
Yamaguchi-Shinozaki, K., and Shinozaki, K. 1993a.
Arabidopsis DNA encoding two desiccation-responsive
rd29 genes. Plant Physiol 101: 1119-1120
Yamaguchi-Shinozaki, K., and Shinozaki, K. 1993b.
Characterization of the expression of a desiccation-
responsive rd29 gene of Arabidopsis thaliana and analysis
of its promoter in transgenic plants. Mol Gen Genet 236:
331-340
Yoshida, R., Hobo, T., Ichimura, K., Mizoguchi, T.,
Takahashi, F., Aronso, J., Ecker, J.R., and Shinozaki, K.
2002. ABA-activated SnRK2 protein kinase is required
for dehydration stress signaling in Arabidopsis. Plant Cell
Physiol 43: 1473-1483
Zak, A. 2000. How Mir Will Come Back to Life. In
Space.com, Ed
http://www.space.com/news/mir_update_000201.html
Zhu, H., and Bunn, H.F. 2001. Signal transduction. How
do cells sense oxygen? Science 292: 449-451

A-L. Paul and R. J. Ferl — Biology in Low Atmospheric Pressure
18 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 19
PLANT-GROWTH LIGHTING FOR SPACE LIFE SUPPORT: A REVIEW Gioia D. Massa
1, Jeffrey C. Emmerich
2, Robert C. Morrow
2, C. Mike Bourget
2, and Cary A. Mitchell
1
1 Department of Horticulture and Landscape Architecture and NASA Specialized Center of Research and Training
in Advanced Life Support, Purdue University, West Lafayette, IN 47907 2 Orbital Technologies Corporation, Madison, WI 53717
ABSTRACT
The lighting system is one of the most important components of
a greenhouse or chamber that will be used for plant growth in an
Advanced Life-support System (ALSS). Several designs of
such lighting systems have been proposed including the use of
natural sunlight with supplemental electric lighting. Although
electric lighting is energy intensive, it is necessary when
balanced against the hazards and limitations of plant growth
under natural sunlight on the surface of the moon or Mars. The
characteristics of different electric light types are reviewed, and
functionality for space application is compared. Different
lighting systems used in Earth-based advanced life-support
(ALS) simulations are highlighted. A new design using light-
emitting diodes (LEDs) for intracanopy lighting, i.e. lighting
entire crop canopies for energy savings and crop enhancement,
is discussed. When the benefits of LEDs are combined with the
advantages of intracanopy lighting, electric lighting for ALS
becomes increasingly feasible.
INTRODUCTION
Food crops will have to be grown when it is necessary to
support human life off Earth for an extended time period.
NASA’s (former) Advanced Life Support (ALS) program
included research associated with water purification, air
revitalization, waste processing, food production and
preparation, and system integration using a combination
of bioregenerative and physico-chemical technologies.
The ultimate goal of life-support research is to support
human habitation off Earth for an indefinite period by
creating a sustainable life-support system that is open
with respect to energy but closed with respect to mass.
NASA’s ALS program evolved from an earlier Closed
Ecological Life-Support System, or CELSS, program.
Several other countries or groups have worked on ALS-
related topics, including Russia, Japan, Canada, the
European Space Agency, and most recently, China. Early
work on human life support emphasized synergisms
between plants and humans in closed systems, with
particular focus on future crop-production systems in
space.
The early ALS work in the US, the USSR, and elsewhere
focused on use of microalgae such as Chlorella sp. for
food production as well as for oxygen and water
revitalization in an ALSS. The lighting systems used for
algae will not be discussed here, as most research moved
early on to higher plants, but for an excellent review of
those experiments, refer to Gitelson, Lisovsky, and
MacElroy (2003).
More recent ALS research includes the design and testing
of habitats for plant growth that will be used on the
surface or subsurface of the moon or Mars. One such
design proposes the use of an inflatable greenhouse that
relies on solar photosynthetically active radiation (PAR)
to directly irradiate crops. The design of such
greenhouses requires transparent films that would have to
withstand low external atmospheric pressure, large
external temperature gradients and swings, strong UV-C
fluxes, micrometeorite impacts, solar particle events,
cosmic-galactic radiation, and must address issues
associated with the widely varying intensity of solar PAR
available to support plant growth (Cockell and Andrady,
1999; Cockell, 2001; Rontó et al., 2003; Horneck et al.,
2003). On the moon, the latter relates to extended periods
of darkness; on Mars, to global dust storms of
unpredictable duration, to diurnal and seasonal light
cycles, and to the planet’s variable distance from the sun.
To deliver solar PAR to plants growing underground or in
containment on the moon or Mars, there not only will be
the same solar-availability constraints, but additional
losses of light as it travels through fiber optics or light
pipes (Cuello et al., 1998; Jack et al., 2002). Jack et al.
(2002) recently evaluated the efficiency of Fresnel-lens
Himawari and parabolic-mirror optical waveguide solar
collection and distribution systems for plant growth. The
first of these devices consists of a collection of Fresnel
lenses that track the sun and focus collected irradiance
onto a cable of fiber optics that can be directed into the
plant-growth area (Jack et al., 2002). The second device
has primary concentrators in the form of rotating
parabolic mirrors that focus the light onto a solid quartz
secondary concentrator that is then linked to a fiber-optic
cable (Jack et al., 2002). Losses of 2.5% - 6.7% per meter
were seen as the light was transmitted through the optical
fibers (Jack et al., 2002; Nakamura et al., 1998).
Although intense light levels over 800 µmol m-2
s-1
could
be obtained in areas of the plant-growth chamber with the
optical waveguide concentrator, lack of uniformity and
variability of light levels was extremely high (Jack et al.,
2002). Cloudy or rainy days reduce the available light
and can cause crop loss (Nakamura et al., 1998). On
Mars, dust storms of great intensity and long duration
occur and limit the already reduced solar radiation that is
available. On the moon, night length is about 2 weeks in
length at most locations except for the south polar region.
Direct or indirect, solar radiation is not reliable enough to
____________________
* Correspondence to: Gioia D. Massa
Department of Horticulture and Landscape Architecture
Purdue University
625 Agriculture Mall Drive
West Lafayette, IN 47907-2010
Email: [email protected]
Phone: 765-496-2124; Fax: 765-494-0391

G.D. Massa— Plant-growth Lighting for Space Life Support
20 Gravitational and Space Biology 19(2) August 2006
be the primary source of photosynthetic energy for human
life support on the moon or Mars.
Since solar energy is not a viable primary option to
support crop production, electric lighting must be
considered. Characteristics to consider when adapting an
electric lighting source for a life-support system away
from Earth include the following:
● Conversion efficiency of electrical power to
photons of photosynthetically active radiation
(PAR)
● Thermal burden (amount and location relative to
crop canopy)
● Lifetime of light source
● Durability of light source
● Mass of light source and associated hardware
● Volume (both storage and usage)
● Light output
-- Quantity
-- Quality
--Tunability/dimmability (versatility for a variety
of crops)
Electrical conversion efficiency refers to the wattage of
electric power that is converted to wattage of light
formed, specifically PAR (400 - 700 nm, Sager and
McFarlane, 1997). Electrical energy that is not converted
to light energy is given off as sensible heat as well as
long-wave radiation, both from ballasts (transformers) as
well as from light sources (lamps) themselves. That part
of the thermal burden generated by lamp surfaces needs to
be removed from the plant-growth area and either
redirected to some heat-requiring region of an ALSS or be
rejected from the habitat. The electrical power required to
run the lamps and to remove the heat burden becomes the
single most expensive cost factor for crop production in
an ALSS. Thus, a primary goal of plant research for ALS
is to generate the largest amount of edible biomass
possible for the least amount of electrical energy used
(i.e., optimize for energy-use efficiency).
Another consideration in adapting an electric lighting
source for ALS is the lifetime of the source, which
impacts the number, volume, and mass of replacement
lamps that must be launched as well as the crew time
needed to maintain a lighting system. Durability of the
source refers to its structural integrity when subjected to
the rigors of launch from Earth to space, including routine
handling, and the conditions encountered in microgravity
for a duration that might be fairly short (Lunar transit,
ISS) or long (Mars Transit). Improved launch, transit,
and landing technologies may reduce such stresses, but
the nature and composition of the light source will
continue to be a factor both for functionality as well as for
issues of astronaut safety.
The mass and volume of a light source, including all
necessary mounting infrastructure, and accessory
equipment, become important in terms of the lift capacity
needed to send those materials to the desired location in
space. Current estimates of launch costs from the Earth’s
surface vary depending on vehicle, but can be
approximated at $10,000 per Kg to low earth orbit and
$300,000 per Kg to the surface of Mars in 2000/2002
dollars, with costs to the lunar surface falling between the
two (Futron Corporation, 2002; A. Drysdale, 2005 Pers.
Comm.). Related issues include the transport and storage
volume of replacement lamps, both in the spacecraft and
at the outpost. Another issue in the design is volume of
space that the lighting infrastructure will occupy within
and above a plant-growth compartment. Larger lighting
systems will limit the space available to accommodate
plants. Therefore, a design with smaller, less massive
lighting systems is desirable.
Other design considerations include a need for flexibility
in the output of light quality, quantity, intensity, and
positioning of the lighting system. Specifically, design
questions to be addressed regarding light quantity include
how many plants each light source will support, and how
flexible a particular light source is with respect to planting
designs? Also, how will photon flux influence lighting
configuration and uniformity? The quality factor of light
output can be very important as well. Some species have
special wavelength requirements for flower initiation in
long-day plants, such as barley requiring far-red
wavelengths (Deitzer et al., 1979). Intumescence in
tomato is eliminated by the presence of UV (Lang and
Tibbitts, 1983) or far-red light (Morrow and Tibbitts,
1988). Certain crops, like lettuce and wheat, grow under
red light alone (Hoenecke et al., 1992, Goins et al., 1997),
but these and other crops have improved biomass
production with the addition of small amounts of blue
light (Hoenecke et al., 1992, Goins et al., 1997; Brown et
al., 1995). Is it better to have one standard white-light
source including all wavelengths, allowing increased
cropping flexibility? Is it better to have custom light
sources designed for each crop, allowing energy
conservation? Or, is some middle ground preferable? The
issues of tunability (e.g., altering color output by shifting
red : blue ratio) and dimmability (altering intensity of
emission) also are light-output factors. Some types of
crops thrive under light with high blue fluxes, while
others grow better under red-enriched light with minimum
blue. Certain crops prefer dim light, while others require
bright light, or may require it only during certain periods
of development. Fluorescent lamp dimming is linear, with
little efficiency drop-off as input power is reduced to a
lower limit (Osram Sylvania, 2004). High-intensity
discharge (HID) lamps have an issue of spectral shift as
input power is decreased (Bubenheim et al., 1995).
Additionally, minimum power levels required for an arc
to fire are still high for both metal halide (MH) and high-
pressure sodium (HPS) lamps (Bubenheim et al., 1995).
For LEDs, which are current controlled, dimming is linear
and continuous, with a correlation between PAR and
current that can be greater than 0.99 (Massa et al., 2005b).
It would be best to maximize edible biomass output per
unit energy input by tuning the light source to suit the
crop at different stages of its development, but features

G.D. Massa— Plant-growth Lighting for Space Life Support
Gravitational and Space Biology 19(2) August 2006 21
such as dimming typically come with associated energy,
mass, and volume costs for traditional lamp types.
Another consideration in reducing the energy requirement
for crop production in a closed system involves reducing
the transpiration rate of crops and thus the amount of
energy that will be required to condense transpired water
vapor. If HID lamps are used, the area of a closed-canopy
crop stand necessary to continuously support the food
requirement of one person in an ALSS (~50m2) will
transpire approximately 4 L/ m2/dayof water vapor, which
amounts to 200 L of water transpired per person per day
(Wheeler et al., 2003). While this at first may seem like a
benefit, in terms of water reclamation, an average person
requires only 2.5 L per day of water to satisfy their
drinking-water requirement (Hopkins, 1993), and at most
40 L of water per day in an ALSS for all purposes
(Mitchell et al., 1996). This leaves an excess of 160 L of
water per crewmember, which adds substantially to the
energy burden of crop production. The change of phase
from liquid to vapor requires 582 cal/g of water at 25˚C,
and the reverse phase change back to liquid is similar.
Each latent heat draw will add 0.68 KW-h for every liter
of water condensed from transpiration (Nobel, 1974),
adding to the thermal burden of crop production in an
ALS system. If a crop-lighting technology can be
developed resulting in a lower thermal load on plants,
then this transpiration rate and the concomitant water-
condensation burden also can be reduced.
Several excellent articles have been written reviewing the
efficiency and usefulness of different types of electric
light sources for plant-growth applications. Bickford and
Dunn (1972) wrote a definitive reference book on this
subject characterizing not only available light sources, but
also addressed the specialized lighting requirements of
numerous crops. An international workshop organized in
1994 compiled a number of papers on plant and animal
lighting in controlled environments with intent for use in
a life-support system (Tibbitts et al., 1994a). Sager and
McFarlane (1997) reviewed radiation impacts on plant
growth and evaluated a wide range of lighting sources for
use in controlled environments. The light sources used in
terrestrial plant growth chambers, however, are not
necessarily a good indicator of what would work best in
an ALSS. Traditional light sources, for example, can be
hot, fragile, and require large, heavy ballasts. Table 1
compares some characteristics important to an ALSS
between different electric light sources.
Table 1. A comparison of several characteristics of light sources important for an ALSS. These values are averages and may not reflect
recent advances in some lamp types.
High Intensity Discharge (HID) Lamps
Parameter
Cool White
Fluorescent
High Pressure
Sodium
Metal
Halide
Microwave Light-
Emitting
Diodes
% Power to
PAR
22%1 35%
1 29%
1 38%
2 Red, 21.5%
Blue, 11%3
Lifetime 20,000 hr1 70%
or less output at
6,000 hr4
24,000 hr1
12,000-
20,000hr4
10,000 hr or
more1
~100,000 hr5
Composition Glass tubes, Hg
vapor, phosphor
coating4
Ceramic
alumina arc
tube contains
Hg, Xe, and
Na, glass outer
bulb1, 6
Quartz tube
in glass bulb,
metal halides
+ pressurized
Hg4
Electrodeless
quartz bulb
filled with S1
Solid state,
materials vary
with LED
color, discrete,
or multiple3, 5
Light quality 400-700nm4
Red shifted,
Peak 550-
650nm4
Blue-biased
400-700nm4
400-700nm1
Various, use
multiple LEDs
for desired
spectrum
Availability available available available not available available
1. MacLennan et al., 1994, 2. Ciolkosz et al., 1998, 3. Mike Bourget, Personal Communication, 2005, 4. Sager and MacFarlane, 1997, 5.
Barta et al., 1992, 6. Bickford and Dunn, 1972

G.D. Massa— Plant-growth Lighting for Space Life Support
22 Gravitational and Space Biology 19(2) August 2006
LIGHTING IN ALS RESEARCH
Fluorescent light was a standard for controlled plant
growth research for many years, and numerous baseline
studies were done using this lighting system. With
fluorescent light, especially cool-white fluorescent, and
often with additional incandescent light, plants grow
“normally” and in approximate proportion to growth of
plants outdoors (Downs and Hellmers, 1978). However,
fluorescent lamps decline gradually in output, do not
typically emit high intensities, and have a limited lifetime
(Sager and McFarlane, 1997). For physiological studies
this may not be an issue, but for crop production,
fluorescent lamps lack the sustained photosynthetic
photon flux (PPF) capability necessary for high
productivity. Most of the lighting sources previously
considered for ALS are in the (extremely fragile) HID
category of lamps. In the Russian Bios–2 facility, a 4.5
m2 phytotron was irradiated with four water-cooled 6-kW
xenon lamps with an average irradiance of 90-115 W/m2
of PAR (Gitelson et al, 2003). Bios-3 was an impressive
closed life-support system containing two plant-growth
compartments, an algae cultivation compartment, and a
habitation unit for three crewmembers. Bios-3 was
unique in that it was designed for control entirely by
crewmembers within the system, as in an off-Earth life-
support scenario, and had teams of people enclosed for up
to 6 months. In the plant chamber, Bios-3 used the same
lamp type as Bios-2, with 20 lamps placed in each of two
31.5 m2 compartments (Gitelson et al, 2003). To reduce
the IR radiation coming from the xenon lamps, the light
sources were installed in a quartz jacket. This was then
enclosed by a glass jacket, and water for cooling flowed
between the two layers to remove heat. In Bios-3, lamp-
cooling water was pumped directly from the nearby
Siberian Yenisei River since it is both cold and pure
(Gitelson et al, 2003). In Bios-3, plant productivity
produced oxygen levels greater than required for a three-
person crew. Food production from crops ranged from
30% of their food requirements in the first three-person
trial to 77.5% in the final two-person, five-month closed
trial. Other foods, predominantly animal products, were
stored in lyophilized or canned forms. Roughly 21% of
crew time for a three-person crew and 19% of crew time
for a two-person crew was spent maintaining the
operation of plant and algal growth areas. Significant
amounts of time were also spent in food preparation,
water purification, and other life support-related
maintenance tasks. Overall, Bios-3 achieved 78-95%
closure of the system and reduction of pre-stored
substances that would need to be supplied for human life
support (Gitelson et al., 2003).
At Kennedy Space Center (KSC), a different kind of HID
lighting was utilized within the closed Biomass
Production Chamber (BPC). This large, closed plant-
growth chamber was a converted hypobaric test chamber
for the Mercury program. Generally, this chamber was
lighted by ninety-six 400-W HPS lamps with remote,
dimmable ballasts, but occasionally MH lamps were used
when crops required it (Wheeler, 1992; Wheeler et al.,
2003). The BPC had 20 m2 of plant growth area within a
volume of 113 m3, and more than twenty tests were
performed. Several crops were grown, including
potatoes, wheat, soybean, lettuce, tomatoes, and rice, and
one test even ran for longer than a year with four
successive potato generations grown in the same nutrient
solution. Numerous important conclusions were reached
during the course of those experiments, and research
continues using conventional growth chambers (Wheeler
et al., 2003). Such studies demonstrated that, if crops
required wide spacing at maturity, productivity and
radiation-conversion efficiency could be improved if they
are transplanted from smaller areas. Also, a nearly linear
response was found between daily PAR and productivity
across a range from 15-60 mol m-2
day-1
for a variety of
unrelated crops (Wheeler et al., 2003). Another key
result that complements other published research indicates
that, in nearly all plant species tested, those grown in a
controlled environment have higher protein, and ash
levels and lower carbohydrate levels than crops grown in
the field (Wheeler et al., 2003).
The Lunar-Mars Life Support Test Project (LMLSTP) at
Johnson Space Center (JSC) incorporated an 11.2 m2
Variable Pressure Growth Chamber (VPGC) to grow a
wheat crop for air revitalization (Gitelson et al., 2003).
Eight banks of 400 W HPS lamps were used in this
chamber and it was demonstrated that wheat growth in
this area revitalized 25% of the carbon dioxide produced
by a crew of four over the 91 days of Phase III testing
(Lawson, 2004). Physico-chemical systems regenerated
the remaining 75%. In addition, some food was provided
from the wheat as bread, and from a separate small plant
growth chamber that used LEDs to produce lettuce
(Gitelson et al., 2003; Lawson, 2004). Following these
tests, the BIO-Plex facility at JSC was designed to use
water-jacketed HPS lamps for crop growth (Tri, 1999).
However, the current status and plans for this integrated
plant growth and human habitation facility remain unclear
(Gitelson et al, 2003).
Lastly, the Closed Ecology Experimental facility, or
CEEF, on the northern island of Honshu, Japan, utilizes
HPS lamps in four plantation chambers, as the sole PAR
source in three of those chambers, and as a supplement to
sunlight in the other (Masuda et al., 2005; Nitta, 2005).
CEEF has a total cultivation area of 150 m2, and early
experimental results indicated that, without increased
productivity, they would need almost 255 m2 crop growth
area per person for a balanced diet. Peanut cultivation for
oil accounted for 78% of this estimate, due to the low
level of fats available in a typical vegetarian diet (Masuda
et al., 2005). Although rice and soybean showed high
productivity within the facility, several other crops require
further optimization to produce optimum biomass.
Currently, only short-term closure experiments have been
performed in CEEF, although one-month habitation
experiments are scheduled to start in the summer of 2006
(Nitta, 2005).

G.D. Massa— Plant-growth Lighting for Space Life Support
Gravitational and Space Biology 19(2) August 2006 23
LED RESEARCH
Light-emitting diodes, LEDs, are a comparatively new
light source for plant growth and are being actively
investigated for numerous applications. Every week new
articles appear in the popular press about advances in
LED technology and the potential of this solid-state light
source for automotive and home lighting, computing,
public works light sources, etc. Red LEDs originally had
15-18% efficiency, but now are up to almost 22%,
whereas blue LEDs were only 3-4% efficient and are now
at 11%. This increase in efficiency makes LEDs
competitive with other sources for plant-growth lighting
(Tennessen and Ciolkosz, 1998; M. Bourget, 2005 Pers.
Comm.). Another important advance in LED research is
the commercial availability of “chip-on-board” LED light
engines. Unlike discrete LEDs with plastic lenses, these
light engines are small printed-circuit wafers that pack
large numbers of small LEDs of selectable emission
colors into close proximity. For example, the ORBITEC
light engine can array 132 LEDs of five colors in a 6.25
cm2 square (Massa et al., 2005a). This allows for
unprecedented color blending and very bright light levels.
LED emissions are current-controlled, and the light output
is directly proportional to input current within their
operating range, so unlike other types of dimming
systems for lighting, dimming of LEDs directly reduces
power usage. LEDs have solid-state construction, are
extremely durable, and resistant to shock. Transparent
coatings on the chips protect them against high humidity
and allow for cleaning without reducing light levels. LED
chips, like discrete LEDs, have low mass and volume.
LEDs generally emit light in a narrow region of the color
spectrum. The number of available colors is extremely
large, with one of the most efficient being red LEDs
emitting at 640 nm, where light has a relative quantum
efficiency for photosynthesis of ~96% (Sager and
McFarlane, 1997). Experimentation has demonstrated
that different species can be grown successfully under
LEDs, including spinach (Goins and Yorio, 2000), lettuce
(Goins et al., 2001; Kim et al., 2004), radish (Goins et al.,
2001), wheat (Goins et al., 1997), and micropropagated
potato plantlets (Miyashuta et al., 1995). Generally, about
15% blue light is required for normal growth, and yields
have been achieved that are comparable to growth under
white light (Yorio et al., 1998). Research has
demonstrated that green light also can have beneficial
effects for growth and plant assessment, especially within
dense foliar canopies (Kim et al., 2004; 2005).
INTRACANOPY LIGHTING FOR CROP GROWTH
Intracanopy (IC) lighting aims to improve lighting
efficiency by providing light distribution throughout the
canopy of a crop. In planophile crops, where leaves
present themselves perpendicular to overhead light and
eventually close off their inner canopy to light, mutual
shading of lower leaves by those above leads to net
carbon loss via respiration, premature leaf drop, and often
flower bud and fruit abortion inside the canopy (Ohler et
al., 1996). Thus, unshaded top and side leaves end up
doing all photosynthetic work for the entire crop stand. If
the light sources could instead irradiate from within the
canopy, a much greater percentage of available leaf
surface could be utilized for photosynthetic work. This
should increase biomass output per energy input
efficiency. Additionally, light intensity drops off
exponentially from a point irradiation source according to
the inverse square law, where
I = E / d 2
with I being the irradiation on a surface at a distance d
from the light source emitting radiant energy E (Bickford
and Dunn, 1972). Thus, light levels drop off rapidly with
increasing increments of distance between lamp and plant,
so that with the necessary separation of hot light sources
above a crop stand the amount of light incident upon the
leaves is highly attenuated, further requiring that the hot
source be high-emitting and high power. If a much cooler
light source can be maintained in close proximity to or
even touching leaves, more light will be available at leaf
level for lower power cost. This will lead to a greater
energy-use efficiency of the biomass-production system.
IC lighting has been previously examined, either as a
supplement to traditional overhead lighting, or as a sole
lighting source. Stasiak and colleagues tested soybean
grown under microwave lamps and supplemented with
side-mounted lighting that was piped into the canopy via
glass tubes lined with optical lighting film to levels of at
least 150 µmol m-2
s-1
PAR at 100 mm from the tube
surface. When overhead light of 400-1200 µmol m-2
s-1
PAR was supplemented with inner canopy lighting,
productivity increased 23-87% (Stasiak et el., 1998).
Also, Tibbitts and Wheeler found that using fluorescent
side lights or MH light pipes with overhead-lighted potato
crops gave increases in tuber dry weights of 12-16%
(Tibbitts et al., 1994b). Sideward lighting systems for
production of plants from cuttings was developed to
reduce the vertical PAR gradient found in overhead-
lighted propagation chambers (Hayashi et al., 1992; Kozai
et al.,1992). One system used fluorescent lamps and it
was demonstrated that sideward lighting reduced the
electricity cost per potato plantlet produced from cuttings
(Hayashi et al., 1992). Fluorescent lamps, however, take
up a large volume of space, and they release heat that then
has to be removed. To counteract these issues, Kozai and
others (1992) used diffusive optical fibers as a light
source for side lighting. This allows plant containers to
be stacked, and also allows placement of containers near
the light source, thereby increasing the efficiency of light
capture and the vigor of biomass accumulation by
plantlets (Kozai et al, 1992).
If low-intensity IC lighting is used as a sole source of
PAR starting from the seedling stage, Frantz and others
demonstrated that expanding cowpea leaves adapted
physiologically to become shade leaves, with lower light-
saturation levels and light-compensation points than
plants lit with more intense light from above (Frantz et al.,
1998). They used short, 15-watt fluorescent tubes

G.D. Massa— Plant-growth Lighting for Space Life Support
24 Gravitational and Space Biology 19(2) August 2006
suspended within the crop canopy by monofilament and
surrounded by transparent Mylar sleeves to prevent leaf
scorch (Frantz et al. 1998; 2000; 2001). With IC lighting
as a sole source, they found twice as much edible biomass
production per unit energy input as in overhead-lit
canopies (Frantz et al., 2000). The two lighting
architectures combined, however, did not increase overall
yield relative to input wattage, probably because the
fixed-position overhead lights were underutilized until the
plants grew to sufficient height (Frantz et al., 2000).
Frantz and colleagues demonstrated that increasing lamp
number within the canopy by 38% raised stand
productivity by 45%, and that the highest energy-use
efficiencies could be obtained by switching lights on
higher up in a canopy as the plants increased in height
(Frantz et al., 2001). When the data were normalized,
plants grown under low-intensity IC lighting produced
50% of the edible biomass of those grown under high-
intensity overhead lighting but with only 10% of the total
electrical energy input (Frantz et al., 1998). Further
increases could not be accomplished, however, due to the
volume occupied by the heat-shielded lamps – if more
lamps were added to the canopy, the available planting
space decreased. Those proof-of-concept studies with
fluorescent lamps illustrated the need for a cool, small-
volume light source that will allow switching on of lights
to keep pace with plant growth. Vertical, linear-arrayed
LEDs were found to fit those requirements.
LED RECONFIGURABLE LIGHTING ARRAYS
The NASA Specialized Center of Research and Training
in Advanced Life Support (ALS NSCORT) was created to
develop technologies to lower the equivalent system mass
(ESM) of an advanced life-support system (Drysdale,
1997). The crops focus area of the ALS NSCORT has
entered into a collaboration with Orbital Technologies
Corporation (ORBITEC, Madison, WI) to develop a
reconfigurable LED lighting array that will significantly
reduce the power and energy required to grow plants
using electric lights. The development and preliminary
testing of this lighting-array system has been described in
previous publications (Massa et al., 2005a; 2005b).
Briefly, the prototype system uses ORBITEC’s
proprietary light engine, consisting of 100 chip-on-board
LEDs set into a 6.25 cm2 square chip. There are sixty-
four 640-nm-emitting (red) LEDs, sixteen 440-nm-
emitting (blue) LEDs, and twenty 540-nm-emitting
(green) LEDs on each chip. Additionally, there are two
photodiodes. The green LEDs and the photodiodes are in
place to accommodate future system-upgrade capabilities.
The small size and close proximity of the LEDs allows for
uniform spectral blending of photon emissions. Since the
LEDs are current controlled, with the colors controlled
separately, both the red-blue ratio and the light intensity
output can be adjusted continuously. Twenty each of
these light engines are mounted along a hollow linear
support (approximately 3 cm wide x 1.5 cm thick x 65 cm
long) that is attached to an electronics enclosure
(approximately 5 cm x 12 cm x 10 cm) also containing
two fans. Figure 1 shows such a “lightsicle”. The hollow
design of the lightsicles allows air to be drawn through
them from the bottom of the canopy past the circuitry
controlling the LEDs and out the top of the enclosure,
thus removing electrically generated heat from the
vicinity of the plants. Each lighting array presently
consists of 16 such lightsicles sized to light a growth area
~0.25 m2 and a growth volume ~0.15 m
3. These arrays
are currently configured to energize light engines from the
bottom up, so that the lights can be switched on
incrementally to keep pace with changing plant height.
Fig. 1 A lightsicle for the reconfigurable lighting array with the major external components labeled. Each lightsicle consists of 20 LED
light engines mounted to a tubular support, with associated electronics. Cooling air is pulled through the support past the internal
electronics by fans mounted on the electronics enclosure.

G.D. Massa— Plant-growth Lighting for Space Life Support
Gravitational and Space Biology 19(2) August 2006 25
The electronics systems within the lightsicles
communicate with a control enclosure via a Controller
Area Network (CAN) communication system. The
control enclosure allows the user to select LED power
levels as well as the number of light engines energized to
allow manual control that keep pace with plant growth.
Red and blue LEDs have independent controls.
Photoperiod is controlled by a programmable timer. The
control enclosure also houses the system power supply
and plugs directly into grounded 110 V power sources.
Each lightsicle can be hung independently in a variety of
configurations, allowing for flexibility in the intracanopy
plant-growth arrangement. In addition, the lightsicle
array can be reconfigured into a rectangular planar array
consisting of 20 x 16 light engines for close-canopy,
overhead lighting. Figure 2A and B shows lightsicles in
intracanopy and overhead configurations, respectively.
This close-canopy configuration is ideal for lighting
rosette (e.g., lettuce) or erectophile (e.g. dwarf wheat)
crops. The light array can be brought in close proximity
to crop surfaces without scorching them, and after the
development and integration of automated switching
protocols, the energized engines will be able to track and
mirror plant growth. Light engines positioned directly
above each seedling will switch on automatically, and
then adjacent engines will illuminate as the leaves of the
seedlings are produced and expand until all engines are
on. As with the IC lighting, greater efficiency will be
achieved by not lighting empty space, but rather targeting
lighting only where photosynthesis can occur.
Fig. 2 A. Intracanopy LED lightsicle array with LEDs off. B. Overhead LED array with LEDs energized. C. Intracanopy array with closed
canopy of cowpea plants. LEDs are not energized. D. Overhead array with closed canopy of cowpea while LEDs are energized. Note
senescence of lower leaves. Plants in C. and D. are both 32 days old.

G.D. Massa— Plant-growth Lighting for Space Life Support
26 Gravitational and Space Biology 19(2) August 2006
Five hardware tests were performed with the first
prototype lighting array using cowpea crop stands.
Modifications to the experimental design were made
between trials 1 through 4 with incremental
improvements in crop productivity achieved at each
successive trial (Massa et al., 2005a). Figure 2C shows
an example of an intracanopy-lighted plant canopy prior
to harvest. Following intracanopy trial 4, the lighting
system was reconfigured into a planar array, and a fifth
trial was run using conditions identical to trial 4 but
mounting the lights overhead (Fig. 2B.). In the overhead
trial, all light engines were energized throughout the trial,
while in the intracanopy trials, lights were switched on
incrementally to keep pace with plant growth. To
normalize the total power usage, the overhead lights were
run at a current that was identical to the average daily
current of intracanopy trial 4 so that the same total
amount of electrical energy (99kW-h) was used during the
month-long trial. In the overhead trial, we observed
mutual shading and drop of the lowermost leaves, so that
11% of the total biomass senesced prior to the end of the
trial. Figure 2D shows the overhead-lighted canopy prior
to harvest. Overall, plants grown under overhead lights
produced less biomass and had a reduced energy
conversion rate than plants grown with intracanopy lights,
with overhead-lighted plants averaging 75% of the
productivity of intracanopy-lighted plants (data not
shown).
These trials were conducted prior to the correction of
certain electronic anomalies reported by Massa et al.
(2005a). In the first hardware tests, PAR output from the
light engines was very low when only lower engines were
energized, with a maximum of 100 µmol m-2
s-1
emitted
from a lower engine with 5 engines energized. As more
engines on each lightsicle were energized, the light output
from each engine increased until more than 700 µmol m-2
s-1
of PAR were detected from the same lower engine
when all 20 engines were energized at the same power
level. Thus, when plants were young and only a few light
engines were energized, the light emission from that
engine was still very low, causing elongate growth and
spindly stems of seedlings. This light-output issue has
been rectified through a modification of the software
controlling the light-engine drivers, and now we are able
to obtain uniform irradiation from a given engine
regardless of the number of light engines energized along
the array. A second set of lightsicles has been constructed
by ORBITEC, and tests are underway to examine IC vs.
OH irradiation in a side-by-side experiment. An
additional feature added to the second prototype array is
an extension so that the control box is raised on 8 of the
16 lightsicles. This allows the shorter lightsicle
electronics enclosures to nestle under the longer
lightsicles, giving a much wider range of possible lighting
configurations. Light-engine positions in the longer
lightsicles are the same as in the shorter ones.
FUTURE DIRECTIONS
A canopy gas-exchange-measurement system is being
developed specifically for IC LED lighting. A custom-
made, whole-canopy cuvette will allow real-time
photosynthesis and transpiration rate measurements of an
entire crop stand growing among the IC lights. Gas
exchange will be measured as a function of environmental
parameters such as light level, red-blue ratio, CO2, and
temperature. This powerful tool will permit rapid
optimization of IC lighting and growth conditions for a
variety of ALS candidate crop species.
A second research focus is being developed at ORBITEC
under a Phase II SBIR from NASA for “High Efficient
Lighting with Integrated Adaptive Control (HELIAC)”.
This project focuses on the development of automated
plant detection and light-engine switching using green
LEDs and photodiodes embedded on individual light
engines. Automation of the switching system to energize
LEDs only when leaves are in front of light engines will
conserve considerable energy by not lighting empty
space, will maximize biomass production by keeping pace
with plant growth, and will significantly reduce the
personnel time involved with light operation.
Additionally, these added capabilities will allow
development of a close-canopy lighting system for
targeted overhead lighting of erectophile and rosette
crops.
CONCLUSIONS When considering a light source for ALS, several
important characteristics must be kept in mind: A variety
of light sources have been evaluated from this
perspective. LEDs, especially the relatively new chip-on-
board LED light engines, appear to be optimal lighting
systems for ALS crop growth for a variety of reasons. As
a rapidly developing technology, electrical efficiency of
these light sources continues to increase. In addition, the
ability to precisely select a spectrum that is efficient for
photosynthesis, growth, and flowering, the durable solid-
state nature of LEDs, the relatively cool emitter surface,
their long lifetime, tunability of the spectrum and
irradiation levels, and ability to easily remove heat all
combine to make this lighting type the best contender for
ALS crop production. When the benefits of LEDs are
coupled with techniques that apply light only where there
is photosynthetic capability, the increased lighting
efficiency will result in a significant reduction in the
power required to maintain desired levels of biomass
production, reducing the cost of growing plants in an
ALSS, and bringing crop growth on Luna and Mars that
much closer to reality.
ACKNOWLEDGEMENTS
The authors wish to acknowledge Mercedes Mick for help
with lighting installation, plant growth, and data
collection and analysis. We also wish to thank Bruce
Bugbee and Ray Wheeler for helpful discussions, and
Thomas Crabb and Charles Barnes for support and

G.D. Massa— Plant-growth Lighting for Space Life Support
Gravitational and Space Biology 19(2) August 2006 27
encouragement. This work was supported in part by
NASA grant NAG5-12686.
REFERENCES
Barta, D.J., Tibbitts, T.W., Bula, R.J., Morrow, R.C.
1992. Evaluation of Light Emitting Diode characteristics
for a space-based plant irradiation source. Adv. Space
Res. 12: 141-149.
Bickford, E.D., Dunn, S. 1972. Lighting for Plant
Growth. The Kent State University Press, pp. 1-221.
Brown, C.S., Schuerger, A.C., Sager, J.C. 1995. Growth
and photomorphogenesis of pepper plants under red light-
emitting diodes with supplemental blue or far-red
lighting. J. Amer. Soc. Hort. Sci. 120: 808-813.
Bubenheim, D.L., Sargis, R., Wilson, D. 1995. Spectral
changes in metal halide and high-pressure sodium lamps
equipped with electronic dimming. HortSci. 30: 1086-
1089.
Ciolkosz, D.E., Albright, L.D., Sager, J.C. 1998.
Microwave Lamp Characterization. Life Support
Biosphere Sci. 5: 167-174.
Cockell, C.S., Andrady, A.L. 1999. The Martian and
extraterrestrial UV radiation environment – 1. Biological
and closed-loop ecosystem considerations. Acta
Astronaut. 44: 53-62.
Cockell, C.S. 2001. The Martian and extraterrestrial UV
radiation environment. Part II: further considerations on
materials and design criteria for artificial ecosystems.
Acta Astronaut. 49: 631-640.
Cuello, J.L., Sadler, P., Jack, D., Ono, E., Jordan, K.A.
1998. Evaluation of light transmission and distribution
materials for lunar and Martian bioregenerative life
support. Life Support Biosphere Sci. 5: 389-402.
Deitzer, G.F., Hayes, R., Jabben, M. 1979. Kinetics and
time dependence of the effect of far red light on the
photoperiodic induction of flowering in wintex barley.
Plant Physiol. 64: 1015-1021.
Downs, R.J., Hellmers, H. 1978. Environment and the
Experimental Control of Plant Growth. London:
Academic Press, pp. 31-82.
Drysdale, A.1997. Computer modeling for advanced life
support system analysis. Life Support Biosphere Sci. 4:
21-29.
Frantz, J.M., Chun, C., Joly, R.J., Mitchell, C.A. 1998.
Intracanopy lighting of cowpea canopies in controlled
environments. Life Support Biosphere Sci. 5: 183-189.
Frantz, J.M., Joly, R.J., Mitchell, C.A. 2000. Intracanopy
lighting influences radiation capture, productivity, and
leaf senescence in cowpea canopies. J. Amer. Soc. Hort.
Sci. 125: 694-701.
Frantz, J.M., Joly, R.J., Mitchell, C.A. 2001. Intracanopy
lighting reduces electrical energy utilization by closed
cowpea stands. Life Support Biosphere Sci. 7: 283-290.
Futron Corporation. 2002. Space transportation costs:
trends in price per pound to orbit 1990-2000.
http://www.futron.com/spaceandtelecom/src/whitepapers.
htm, pp. 1-7.
Gitelson, I.I., Lisovsky, G.M., MacElroy, R.D. 2003.
Manmade Closed Ecological Systems. New York: Taylor
and Francis, pp. 31-309.
Goins, G.D., Sager, J.C., Wheeler, R.M., Ruffe, L.M.,
Yorio, N.C. 2001. Salad crop production under different
wavelengths of red light-emitting diodes (LEDs). SAE
Technical Paper # 2001-01-2422.
Goins, G.D., Yorio, N.C. 2000. Spinach growth and
development under innovative narrow- and broad-
spectrum lighting sources. SAE Technical Paper # 2000-
01-2290.
Goins, G.D., Yorio, N.C., Sanwo, M.M., Brown, C.S.
1997. Photomorphogenesis, photosynthesis, and seed
yield of wheat plants grown under red light-emitting
diodes (LEDs) with and without supplemental blue
lighting. J. Exper. Bot. 48: 1407-1413.
Hayashi, M., Fujita, N., Kitaya, Y., Kozai, T. 1992. Effect
of sideward lighting on the growth of potato plantlets in
vitro. Acta Hort. 319: 163-166.
Hoenecke, M.E., Bula, R.J., Tibbitts, T.W. 1992.
Importance of ‘blue’ photon levels for lettuce seedlings
grown under red-light-emitting diodes. HortSci. 27: 427-
430.
Hopkins, B. 1993. In: Nutrition Support Dietetics, 2nd
Edition (Gattschlich, M., Matarese, L., Shronts, E., Eds.)
Silver Spring, MD: American Society for Parenteral and
Enteral Nutrition, p. 63.
Horneck, G., Facius, R., Reitz, G., Rettberg, P.,
Baumstark-Khan, C., Gerzer, R. 2003. Critical issues in
connection with human missions to Mars: protection of
and from the Martian Environment. Adv. Space Res. 31:
87-95.
Jack, D.A., Nakamura, T., Sadler, P., Cuello, J.L. 2002.
Evaluation of two fiber optic-based solar collection and
distribution systems for advanced space life support.
Trans. ASAE 45: 1547-1558.
Kim, H.H., Goins, G.D., Wheeler, R.M., Sager, J.C. 2004.
Green-light supplementation for enhanced lettuce growth
under red- and blue-light-emitting diodes. HortSci. 39:
1617-1622.

G.D. Massa— Plant-growth Lighting for Space Life Support
28 Gravitational and Space Biology 19(2) August 2006
Kim, H.H., Wheeler, R.M., Sager, J.C., Yorio, N.C.,
Goins, G.D. 2005. Light-emitting diodes as an
illumination source for plants: a review of research at
Kennedy Space Center. Habitation10: 71-78.
Kozai, T. Kino, S. Jeong, B.R., Kinowaki, M., Ochiai, M.,
Hayashi, M., Mori, K. 1992. A sideward lighting system
using diffusive optical fibers for production of vigorous
micropropagated plantlets. Acta Hort. 319: 237-242.
Lang, S.P., Tibbitts, T.W. 1983. Factors controlling
intumescence development on Tomato plants. J. Amer.
Soc. Hort. Sci. 108: 93-98.
Lawson, M. 2004. Lunar-Mars Life Support Test Project.
http://advlifesupport.jsc.nasa.gov/ehti3/index.html
MacLennan, D.A., Turner, B.P., Dolan, J.T., Ury, M.G.,
Gustafson, P. 1994. Efficient, full-spectrum, long-lived,
non-toxic microwave lamp for plant growth. In:
Proceedings, International Lighting in Controlled
Environments Workshop. (Tibbitts, T.W., Chairman)
NASA Kennedy Space Center, FL.: NASA Conference
Publication (CP)-3309, pp. 243-254.
Massa, G.D., Emmerich, J.C., Morrow, R.C., Mitchell,
C.A. 2005a. Development of a Reconfigurable LED
Plant-growth Lighting System for Equivalent System
Mass Reduction in an ALS. SAE Technical Paper #
2005-01-2955.
Massa, G.D., Emmerich, J.C., Mick, M.E., Kennedy, R.J.,
Morrow, R.C., Mitchell, C.A. 2005b. Development and
testing of an efficient LED intracanopy lighting design for
minimizing equivalent system mass in an advanced life-
support system. Gravit. Space Biol. Bull. 18: 87-88.
Masuda, T., Arai, R., Komatsubara, O., Tako, Y.,
Harashima, E., Nitta, K. 2005. Development of a 1-week
cycle menu for an advanced life support system (ALSS)
utilizing practical biomass production data from the
closed ecology experiment facilities (CEEF). Habitation
10:87-97.
Mitchell, C.A., Dougher, T.A.O., Nielsen, S.S., Belury,
M.A., Wheeler, R.M. 1996. Costs of providing edible
biomass for a balanced vegetarian diet in a controlled
ecological life-support system. In: Plants in Space
Biology (Suge, H. Ed.) Sendai, Japan: Tohoku University
Press, pp. 245-254.
Miyashita, Y., Kitaya, Y., Kozai, T., Kimura, T. 1995.
Effects of red and far-red light on the growth and
morphology of potato plantlets in vitro: using light
emitting diode as a light source for micropropagation.
Acta Hort. 393: 189-194.
Morrow, R.C., Tibbitts, T.W. 1988. Evidence for
involvement of phytochrome in tumor development on
plants. Plant Physiol. 88: 1110-1114.
Nakamura, T., Case, J.A., Mankamyer, M. 1998.
Development of the optical waveguide solar lighting
system for space-based plant growing. Life Support
Biosphere Sci. 5: 205-215.
Nitta, K. 2005. The Mini-Earth facility and present status
of habitation experiment program. Adv. Space Res. 35:
1531-1538.
Nobel, P.S. 1974. Introduction to Biophysical Plant
Physiology. San Francisco: W. H. Freeman and Company,
pp. 420-424.
Ohler, T.A., Nielsen, S.S., Mitchell, C.A. 1996. Varying
plant density and harvest time to optimize cowpea leaf
yield and nutrient content. HortSci. 31: 193-197.
Osram Sylvania. 2004. Quicktronic Helios T8 dimming.
http://www.sylvania.com/BusinessProducts/LightingForB
usiness/Products/Systems/DimmingandDaliBallasts/
Rontó, G., Bérces, A., Lammer, H., Cockell, C.S.,
Molina-Cuberos, G.J., Patel, M.R., Selsis, F. 2003. Solar
UV irradiation conditions on the surface of Mars.
Photochem. Photobiol. 77: 34-40.
Sager, J.C., McFarlane, J.C. 1997. Radiation. In: Plant
Growth Chamber Handbook. (Langhans, R.W. and
Tibbitts, T.W., Eds.) Iowa State University: North
Central Regional Research Publication No. 340, Iowa
Agriculture and Home Economics Experiment Station
Special Report no. 99, pp. 1-29.
Stasiak, M.A., Côté, R., Dixon, M., Grodzinski, B. 1998.
Increasing plant productivity in closed environments with
inner canopy illumination. Life Support Biosphere Sci. 5:
175-181.
Tennessen, D.J., Ciolkosz, D.E. 1998. Towards efficient
conversion of electricity into edible biomass in crop
production systems: a transgenic approach. Life Support
Biosphere Sci. 5: 217-233.
Tibbitts, T.W., chairman, and others. 1994a. Proceedings,
International Lighting in Controlled Environments
Workshop. NASA Kennedy Space Center, FL.: NASA
Conference Publication (CP)-3309, pp. 1-393.
Tibbitts, T.W., Cao, W., Wheeler, R.M. 1994b. Growth of
potatoes for CELSS. NASA Contractor Report 177646.
Ames Research Center, Moffett Field, CA.
Tri, T.O. 1999. Bioregenerative planetary life support
system test complex (BIO-Plex): test mission objectives
and facility development. SAE Technical Paper # 1999-
01-2186.
Wheeler, R.M. 1992. Gas-exchange measurements using
a large, closed plant growth chamber. HortSci. 27: 777-
780.

G.D. Massa— Plant-growth Lighting for Space Life Support
Gravitational and Space Biology 19(2) August 2006 29
Wheeler, R.M., Sager, J.C., Prince, R.P., Knott, W.M.,
Mackowiak, C.L., Stutte, G.W., Yorio, N.C., Ruffe, L.M.,
Peterson, B.V., Goins, G.D., Hinkle, C.R., Berry, W.L.
2003. Crop Production for Advanced Life Support
Systems – Observations from the Kennedy Space Center
Breadboard Project. NASA Kennedy Space Center: NASA
Technical Memorandum TM-2003-211184, pp. 1-25.
Yorio, N.C., Wheeler, R.M., Goins, G.D., Sanwo-
Lewandowski, M.M., Mackowiak, C.L., Brown, C.S.,
Sager, J.C., Stutte, G.W. 1998. Blue light requirements
for crop plants used in bioregenerative life support
systems. Life Support Biosphere Sci. 5: 119-128.

G.D. Massa— Plant-growth Lighting for Space Life Support
30 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 31
INCREASED BACTERIAL RESISTANCE AND VIRULENCE IN SIMULATED MICROGRAVITY
AND ITS MOLECULAR BASIS
A. Matin1, S.V. Lynch
2 and M.R. Benoit
1
1 Department of Microbiology & Immunology, Stanford University School of Medicine
2 Department of Anesthesia and Perioperative care, UCSF Medical School
ABSTRACT
Experiments conducted during space flights showed that space
conditions affect basic aspects of bacterial physiology, such as
growth, conjugation efficiency, and secondary metabolite
production. Since logistical constraints hamper research during
space flights, use of ground based systems has greatly stimulated
progress in this field. A commonly used system is the rotating
bioreactor which simulates some aspects of microgravity.
Studies using this system have shown that bacteria grown under
diminished gravity become more virulent and resistant to a
variety of antimicrobial treatments [referred to as
“comprehensive cellular resistance (CCR)]. In this respect, the
diminished gravity effect resembles the well studied bacterial
general stress response. The latter is centrally controlled by an
alternate sigma factor, called σs. However, the diminished
gravity-conferred CCR is not controlled by this sigma factor in
rapidly growing cells. Moreover, the proteins known to confer
CCR under the Earth conditions do not appear to have a role in
this resistance under the diminished gravity conditions. Growth
in the HARV system altered the of σs regulation pattern at
transcriptional, translational and posttranslational levels. Since
macromolecular folding patterns play a role in this regulation,
the findings raise the possibility that diminished gravity may
alter the folding pattern of macromolecules. The effect of
diminished gravity on bacterial biofilms, which are responsible
for serious and recalcitrant diseases, has recently become
possible on earth by the designing of a novel HARV adaptation.
The results show that diminished gravity stimulates biofilm
formation and makes them highly resistant to antimicrobial
agents. Given that the human immune response is compromised
in space, these results highlight the serious danger that microbes
can potentially pose in space.
INTRODUCTION
Space travel, although tantalizing, is fraught with hazards.
There are first the purely human factors, such as the
psychological stress originating from the fear of the
unknown, isolation, and the claustrophobic atmosphere of
a confined environment; and upsetting of the normal life
rhythm that results in sleep deprivation and
malnourishment. Then there are the peculiarities of space:
unusually strong radiation and diminished gravity. Life on
Earth evolved in the presence of gravity and thus the latter
aspect is an unprecedented experience for the earthlings.
A good deal of research in recent years has concerned
with determining the effect of diminished gravity on
human physiology as well as on bacterial characteristics.
And the answer seems to be that diminished gravity or
‘microgravity’ as it is termed, weakens humans and
makes bacteria stronger, more difficult to kill, and more
accomplished at causing disease. In this review, we will
discuss the findings underlying these conclusions.
Bone decalcification and loss are well documented in
astronauts during space travel and residence. This
predisposes them to bone fracture as well as kidney stones
from resorbed bone material. In microgravity, muscles
atrophy and blood production decreases. The latter results
in diminished pumping by the heart and, combined with
the concomitant blood shift to the upper torso, can
damage heart muscles. In addition, microgravity
compromises the human immune response. This appears
to result from several factors: the proportion and number
of circulating lymphocytes and their cytokine production
are detrimentally altered, lymphocytes produce lower
levels of human leukocyte antigen, there is increased
apoptosis of peripheral blood mononuclear cells, and
dendritic cells become defective in phagocytosis
(Sonnenfeld and Shearer, 2002). Additionally,
pharmacodynamics and pharmacokinetics are expected to
be altered during spaceflight (Graebe et al., 2004).
The study of spaceflight effects on bacteria originated
during the advent of manned spaceflight programs in the
1960’s. Since that time, many bacterial studies have been
conducted onboard spacecraft, and several published
reviews of these studies exist (Cioletti, 1991; Dickson,
1991; Moore, 1996; Pierson and Mishra, 2000; Klaus,
2002). In general, these studies have shown that
spaceflight causes increased final cell populations,
shortened lag phase duration, increased conjugation
efficiency, and increased production of secondary
metabolites. Other findings, which pose a more direct
threat to human health, include decreased antibiotic
effectiveness during spaceflight experiments (Tixador et
al., 1985; Lapchine et al., 1988; Tixador et al., 1994) and
increased spacecraft contamination. Observations of
spacecraft contamination include the discovery of
biofilms in the Space Shuttle water system (Koenig and
Pierson, 1997), free-floating condensate containing
microbes onboard the International Space Station (Ott et
al., 2004), and microbial contamination onboard the Mir
Space Station (Novikova, 2004). The combination of
potentially less effective antibiotic treatment, increased
____________________
* Correspondence to: A. Matin
Department of Microbiology & Immunology
Stanford University School of Medicine
Stanford, CA 94305
Email: [email protected]
Phone: 650-725-4745; Fax: 650-725-6757

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
32 Gravitational and Space Biology 19(2) August 2006
Fig. 1A HARV system used to simulate conditions of microgravity in Earth-based investigations. B. HARV vessel components. (Synthecon,
Inc.)
bacterial growth, contaminated spacecraft, and
suppressed astronaut immune systems presents a serious
threat to crew health. This threat will become
exacerbated with increased mission duration (Pierson,
2001), and future missions planned for travel outside of
low Earth orbit will not be capable of a swift return home
in the case of an untreatable infectious disease.
Studies on the effect of diminished gravity on bacteria as
well as mammalian cells have been greatly facilitated by
the invention of earth based systems that reproduce some
aspects of microgravity. The clinostat, which was
developed over a century ago for studying gravitropic
plant responses (Dedolph and Dipert, 1971), rotates a
container of suspension cultures about an axis that is
perpendicular to the gravity vector. Thorough descriptions
of this device and its limitations have been published
(Albrecht-Buehler, 1992; Klaus, 2001). More recently,
rotating systems such as the high aspect ratio vessel
(HARV) bioreactor (Fig. 1a), have been designed for
studying mammalian tissue culture (Wolf and Schwarz,
1991; Schwarz et al., 1992; Wolf and Schwarz, 1992).
The HARV has front and back faces that are separable;
the front face contains two sampling ports, and the back
face is equipped with a semi-permeable membrane for
aeration (Fig. 1b). The vessel is completely filled with
the medium and inoculum so that no head space remains.
Air bubbles are removed, to prevent fluid mixing. Two
such vessels are used per experiment, of which one is
rotated about a vertical axis, perpendicular to the
gravitational vector, and the other about a horizontal axis,
parallel to this vector.
In both the clinostat and the HARV, virtually no relative
fluid motion exists and after an initial start-up time, solid
body rotation of the fluid ensues. Since mixing occurs by
gentle rotation in the absence of stirring vanes, and
aeration is affected by dissolved gasses with no headspace
or bubbles, there is no shear-field inhomogeneity;vessels
rotated around either axis thus possess a low-shear
environment. In the vessel rotated about the horizontal
axis, particles quickly reach a terminal settling velocity,
but cumulative sedimentation is avoided due to
hydrodynamic forces, viz., shear, centrifugal and Coriolis
(Hammond and Hammond, 2001). The gravity vector is
continually re-oriented as the cells and fluid rotate
together, which dramatically influences large cells, e.g.,
those of plants with gravity sensing mechanisms
(Dedolph and Dipert, 1971).
Both the clinostat, and more recently the HARV, have
been used to study bacteria. Although planned
experiments to demonstrate the effectiveness of the
HARV as a spaceflight analog have yet to be conducted
(Niesel et al., 2005), ground-based clinostat experiments
have been correlated with spaceflight experiments
yielding similar results, at least in terms of increased final
cell populations (Kacena et al., 1999; Brown et al., 2002).
Due to the small size of bacteria, rotating culture systems
operated at the correct rotation rate can reduce
gravitational cell motion to less than Brownian motion
(random diffusion) for the duration of the experiment
(Todd and Klaus, 1996), thereby making a suitable
simulation of the reduced fluid motion provided by
weightlessness. This condition has been referred to as
Low Shear Modeled MicroGravity (LSMMG) (Nickerson
et al., 2003; Nickerson et al., 2004) or simply Simulated
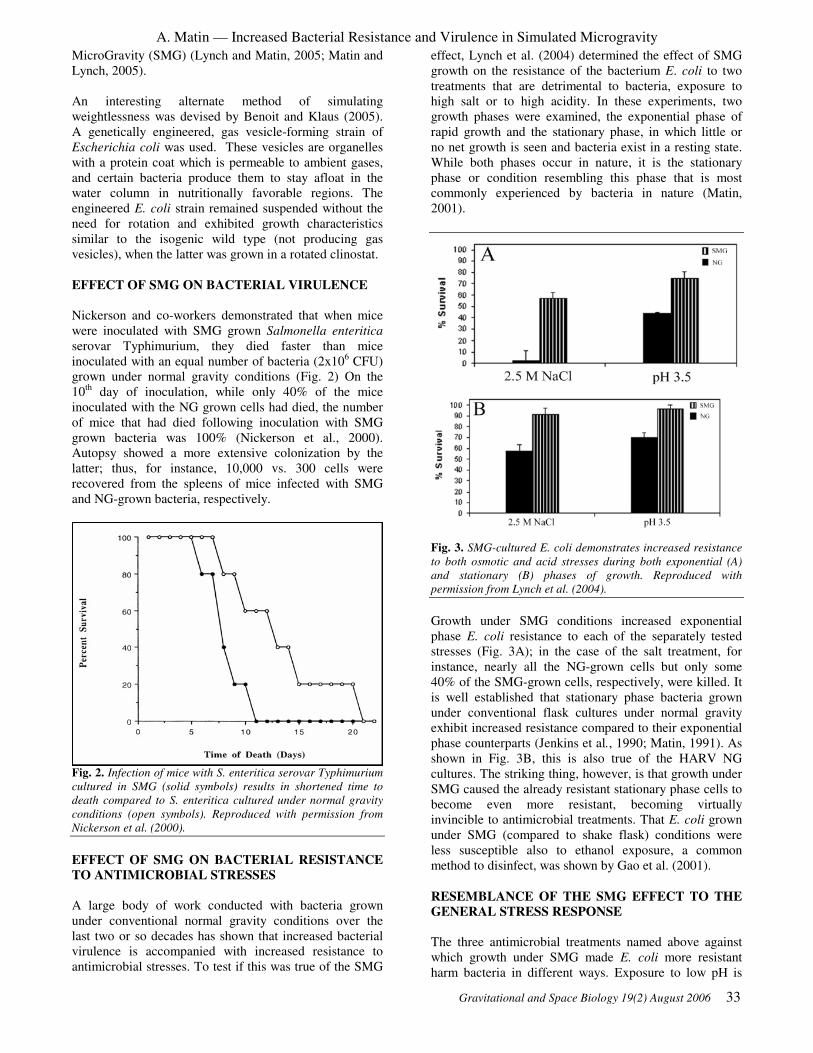
A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 33
MicroGravity (SMG) (Lynch and Matin, 2005; Matin and
Lynch, 2005).
An interesting alternate method of simulating
weightlessness was devised by Benoit and Klaus (2005).
A genetically engineered, gas vesicle-forming strain of
Escherichia coli was used. These vesicles are organelles
with a protein coat which is permeable to ambient gases,
and certain bacteria produce them to stay afloat in the
water column in nutritionally favorable regions. The
engineered E. coli strain remained suspended without the
need for rotation and exhibited growth characteristics
similar to the isogenic wild type (not producing gas
vesicles), when the latter was grown in a rotated clinostat.
EFFECT OF SMG ON BACTERIAL VIRULENCE
Nickerson and co-workers demonstrated that when mice
were inoculated with SMG grown Salmonella enteritica
serovar Typhimurium, they died faster than mice
inoculated with an equal number of bacteria (2x106 CFU)
grown under normal gravity conditions (Fig. 2) On the
10th
day of inoculation, while only 40% of the mice
inoculated with the NG grown cells had died, the number
of mice that had died following inoculation with SMG
grown bacteria was 100% (Nickerson et al., 2000).
Autopsy showed a more extensive colonization by the
latter; thus, for instance, 10,000 vs. 300 cells were
recovered from the spleens of mice infected with SMG
and NG-grown bacteria, respectively.
Fig. 2. Infection of mice with S. enteritica serovar Typhimurium
cultured in SMG (solid symbols) results in shortened time to
death compared to S. enteritica cultured under normal gravity
conditions (open symbols). Reproduced with permission from
Nickerson et al. (2000).
EFFECT OF SMG ON BACTERIAL RESISTANCE
TO ANTIMICROBIAL STRESSES
A large body of work conducted with bacteria grown
under conventional normal gravity conditions over the
last two or so decades has shown that increased bacterial
virulence is accompanied with increased resistance to
antimicrobial stresses. To test if this was true of the SMG
effect, Lynch et al. (2004) determined the effect of SMG
growth on the resistance of the bacterium E. coli to two
treatments that are detrimental to bacteria, exposure to
high salt or to high acidity. In these experiments, two
growth phases were examined, the exponential phase of
rapid growth and the stationary phase, in which little or
no net growth is seen and bacteria exist in a resting state.
While both phases occur in nature, it is the stationary
phase or condition resembling this phase that is most
commonly experienced by bacteria in nature (Matin,
2001).
Fig. 3. SMG-cultured E. coli demonstrates increased resistance
to both osmotic and acid stresses during both exponential (A)
and stationary (B) phases of growth. Reproduced with
permission from Lynch et al. (2004).
Growth under SMG conditions increased exponential
phase E. coli resistance to each of the separately tested
stresses (Fig. 3A); in the case of the salt treatment, for
instance, nearly all the NG-grown cells but only some
40% of the SMG-grown cells, respectively, were killed. It
is well established that stationary phase bacteria grown
under conventional flask cultures under normal gravity
exhibit increased resistance compared to their exponential
phase counterparts (Jenkins et al., 1990; Matin, 1991). As
shown in Fig. 3B, this is also true of the HARV NG
cultures. The striking thing, however, is that growth under
SMG caused the already resistant stationary phase cells to
become even more resistant, becoming virtually
invincible to antimicrobial treatments. That E. coli grown
under SMG (compared to shake flask) conditions were
less susceptible also to ethanol exposure, a common
method to disinfect, was shown by Gao et al. (2001).
RESEMBLANCE OF THE SMG EFFECT TO THE
GENERAL STRESS RESPONSE
The three antimicrobial treatments named above against
which growth under SMG made E. coli more resistant
harm bacteria in different ways. Exposure to low pH is

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
34 Gravitational and Space Biology 19(2) August 2006
detrimental because it acidifies the cytoplasm and leads to
decomposition or denaturation of vital cell constituents.
High salt dehydrates the cell, and exposure to ethanol
damages the cell envelope. That SMG made cells resistant
to these three disparate stresses, which kill bacteria by
causing different kinds of injuries, indicates that the
mechanism of resistance activated by SMG encompasses
preventing and/or repairing injury to many different cell
constituents.
Fig. 4. Comparison of the H2O2 resistance of exponential phase
(○), or glucose-starved (□) E. coli cultures to growing cultures
stressed by heat (∆), H2O2 (▲), or ethanol (●). Reproduced
with permission from Matin (2001).
In this respect, the SMG effect resembles the general
stress response (GSR) that has been extensively studied
under conventional culture conditions of normal gravity
(Matin, 2001). This phenomenon is illustrated in Fig. 4.
When bacteria are exposed to small doses of H2O2, they
acquire the capacity to resist much higher and normally
lethal doses of this toxic agent. But the striking thing is
that protection against lethal doses of H2O2 results from
prior exposure of cells to a large number of separate
unrelated antimicrobial treatments, such as starvation,
heat, or ethanol. This sort of cross protection operates
against almost any stress so that, for instance, H2O2-
treated cells become more resistant to the other
deleterious agents as demonstrated in Fig. 4; and heat or
starvation-stressed cells acquire cross protection to
unrelated insults such as osmotic shock, or exposure to
low pH. Thus, the essence of GSR is that exposure of
bacteria to small sub-lethal dose of an antimicrobial
treatment makes them resistant not only to subsequent
exposure of lethal doses of the same agent but also to
others that injure bacteria by different mechanisms.
MOLECULAR AND BIOCHEMICAL BASIS OF
GSR AND SMG EFFECTS
What is the molecular basis that renders cells
comprehensively resistant after exposure to small doses of
a deleterious agent? Bacterial cells exposed to a large
variety of harmful agents respond by increasing the
cellular concentration of a “stress” sigma factor, σs. This
sigma factor replaces the “house keeping” sigma factor
(σ70
) on the RNA polymerase enzyme, thereby changing
its regulatory properties. σs-RNA polymerase recognizes
a different promoter sequence compared to σ70
-RNA
polymerase; promoter regions consist of sequences
present in front of the coding region of genes. The σs-
RNA polymerase recognized promoters are present in
front of genes that encode proteins, which can protect
against different types of cell injuries. Examples of
proteins thus induced are molecular chaperones, such as
DnaK which prevents damage to and can repair denatured
proteins; the SOS response proteins, e.g., PexB that can
insulate the cell against DNA damage, and proteins such
as D-alanine carboxypeptidase and PexAB that are
concerned with cell envelope integrity (Matin, 2001).
Consequently, bacteria subjected to sub-lethal dose of one
harmful treatment become capable of withstanding
multiple types of damage to their cell constituents.
Fig. 5. Quantification of σs protein levels in exponential (A) and
stationary (B) phase NG and SMG cultures. Reproduced with
permission from Matin and Lynch (2005).
As we have seen (Fig. 3), growth under SMG also makes
cells resistant to mechanistically different killing agents,
and since it resembles GSR in this respect, it was logical
to suspect that the underlying molecular and biochemical
mechanisms may be similar, and that SMG acted in the
same way as do other deleterious agents. However,
exponential phase cells grown under SMG conditions

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 35
possessed lower σs levels than their NG-grown
counterparts (Fig. 5A; (Lynch et al., 2004)). This is the
first instance in which increased cellular resistance to
deleterious treatments is found accompanied with lowered
levels of σs. In contrast, the stationary phase SMG-grown
cells that exhibit super resistance (Fig. 3), conformed to
the normal paradigm: their increased resistance was
accompanied by increased σs levels compared to the NG-
grown cells (Fig. 5B). The conclusion suggested by these
findings, namely, that the SMG-conferred resistance is
independent of σs in exponential phase, but dependent on
it in the stationary phase was confirmed by repeating
these experiments with a mutant of E. coli missing this
sigma factor. The mutant developed resistance normally
in the exponential phase (Fig. 6A), when grown under
SMG conditions, but was severely weakened under both
NG and SMG conditions in stationary phase (Fig. 6B).
Fig. 6. Stress resistance of an rpoS mutant strain grown under
NG and SMG conditions in exponential (A) and stationary (B)
phases of growth. Reproduced with permission from Lynch et al.
(2004).
The Nickerson group has examined the genes that are
influenced in exponential phase-cells grown under SMG
conditions (Wilson et al., 2002). Expression of over 160
genes was affected by growth under SMG conditions in S.
enterica typhimurium serovar Typhimurium. Surprisingly,
none of the up-regulated genes included those known to
encode protective proteins of the type mentioned above.
Furthermore, many of the genes involved in determining
virulence under the conventional flask culture conditions
were in fact down-regulated by SMG growth. The ferric
iron uptake protein, Fur, which has been shown to have a
role in SMG acid resistance, could have a larger role in
regulating gene expression under these conditions, since
many of the SMG-induced genes apparently possess Fur-
binding sites (Wilson et al., 2002). How gene expression
is regulated by SMG growth in the stationary phase has
not yet been investigated. It is clear however, that the
SMG and conventional normal gravity stress resistance
and virulence mechanisms differ in at least two respects:
in the exponential phase, it is independent of σs and does
not involve induction of genes known to be responsible
for conferring protection on the bacterial cell; and in that
the relationship of this mechanism to σs, and possibly also
to the known stress genes, reverses during transition from
exponential to stationary growth phase.
MOLECULAR REGULATION OF σσσσS
These unexpected findings led Lynch et al. (2004) to
explore if fundamental cell processes, such as regulation
of protein synthesis, were affected by growth under SMG.
They addressed this question by looking at σs for two
reasons, one that under SMG it is up or down regulated
depending on the growth phase; and two that its synthesis
has been thoroughly studied in conventional gravity
conditions, providing guidelines on aspects to study.
These prior conventional investigations have established
that σs synthesis is regulated by a combination of
regulatory mechanisms that operate at transcriptional,
translational efficiency, and protein stability levels.
Fig. 7. Western analysis of σs protein degradation over time in
exponential (A) and stationary (B) phases of growth under NG
and SMG conditions. N, Normal gravity time point; S, Simulated
microgravity time point. Reproduced with permission from
Lynch et al. (2004).
σs is encoded by the rpoS gene. To study the effect of
SMG on the transcription rate of this gene, the steady-
state levels of the rpoS messenger RNA (mRNA) were
quantified as well as the half-life of this mRNA in cells
grown under NG and SMG conditions, using quantitative
PCR. The transcription rate was not altered in exponential
phase but was decreased by one-half in the stationary
phase in SMG-grown cells (Table 1). A much more
pronounced effect of SMG was seen on the stability of σs
protein (Fig. 7). In the exponential phase, growth under
SMG rendered this protein over three-fold less stable. A
similar, less pronounced, but reproducible effect occurred
also in the stationary phase cells. Translational efficiency
of the rpoS mRNA, i.e., the amount of σs protein
synthesized per molecule of the rpoS mRNA, was
calculated from the rate of σs synthesis (calculated in turn
from steady state σs
levels and from the half-life of this
protein) and that of rpoS mRNA synthesis (Table 2). It
increased over two-fold under SMG regardless of the
growth phase.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
36 Gravitational and Space Biology 19(2) August 2006
Table 1. Copies of rpoS mRNA synthesized per minute under NG and SMG conditions in exponential or stationary phases of growth
(Lynch et al., 2004).
NG SMG
Exponential phase 1.6 x 107
1.6 x 107
Stationary phase 2.8 x 105 1.2 x 10
5
Table 2. Translational efficiency of σs protein per copy of rpoS mRNA per minute (Lynch et al., 2004).
NG SMG
Exponential phase 3.6 x 1011
8.4 x 1011
Stationary phase 3.3 x 1011
8.4 x 1011
It is not known how SMG affects these parameters of σs
synthesis. It is possible that the levels of molecules that
regulate these processes are affected by growth under
SMG. Another possibility is suggested by the fact that
regulation at all the levels mentioned above involves
higher order structures of the concerned molecules.
Stability of the σs protein depends on whether it is subject
to proteolysis by a specific protease, called the ClpXP
protease (Schweder et al., 1996; Becker et al., 1999;
Hengge-Aronis, 2002). Why it is attacked more readily by
the ClpXP protease under certain conditions and not
others is not fully understood. Inactivation of another
protein, called RssB (or SprE), appears to have a role
under carbon starvation conditions. In addition, the
folding pattern of σs could also be involved. The activity
of ClpXP protease is markedly affected by the folding
pattern of its substrate (Kenniston et al., 2004).
Experiments conducted in space have shown that proteins
form crystals more readily under these conditions. Taking
these findings together, it has been hypothesized that
diminished gravity and low shear may influence protein
folding pattern (Fig. 8; (Lynch et al., 2004)). In the
present case, this may mean that growth under SMG
causes the σs protein to acquire a configuration that is
more amenable to attack by the ClpXP protease.
Fig. 8. Schematic representation of post-translational degradation of σs protein involving RssB, the degradation tag (termed SsrA) and the
ClpXP protease complex.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 37
Fig. 9. rprA a small non-translated regulatory RNA possesses stretches of sequence homology with the untranslated region of rpoS,
permitting binding and relief of translation inhibition of the rpoS mRNA. Reproduced with permission from Matin and Lynch (2005).
Secondary structure formation also plays a central role in
the translational efficiency of rpoS mRNA. This
messenger molecule possesses internal homologies in its
untranslated region that result in the formation of a
hairpin structure, which block its translational start site,
making it unavailable to the ribosomes. Studies under
conventional conditions of normal gravity strongly
suggest that this structure is relaxed when cells are
exposed to antimicrobial treatments. This is brought about
by interaction between the rpoS mRNA, a protein called
Hfq, and one of several small RNAs, for example, rprA.
The latter possesses homologies to the interfering
sequence responsible for the hairpin structure formation
in the rpoS mRNA and, in association with Hfq and
perhaps other factors, can disrupt this structure. As a
result the translational start site becomes untangled and
available to the ribosomes, promoting synthesis of σs
(Fig. 9). SMG could act in stimulating translation by
affecting any of the participants in this mechanism,
including a direct effect in minimizing secondary
structure formation within the rpoS mRNA. Only future
work will reveal what factors are involved in these
phenomena, but if diminished gravity and low shear can
influence folding patterns of molecules, implications of
space conditions on life processes would be wide ranging
indeed.
EFFECT OF SMG ON BACTERIAL BIOFILM
RESISTANCE
Biofilms are bacterial communities that are surrounded by
a polysaccharide matrix. Different cells within such
communities may perform different functions, and
biofilms may have a primitive circulatory system to
ensure efficient distribution of nutrients and removal of
waste products. Bacterial biofilms have a considerable
medical importance because they exhibit much greater
resistance to antibiotics and other antimicrobial treatments
than their constituent individual cells when they exist
outside the community. Consequently, diseases in which
bacterial biofilms have a major role, such as cystitis,
endocarditis, and cystic fibrosis, are chronic and difficult
to treat (Fux et al., 2005).
During experiments onboard the Space Shuttle,
Burkholderia cepacia formed biofilms (Pyle et al., 1999)
and Pseudomonas aeruginosa biofilms developed on
polycarbonate membranes (McLean et al., 2001).
Furthermore, biofilms were found in the Space Shuttle
water system (Koenig and Pierson, 1997) and on surfaces
and equipment onboard the Mir Space Station (Novikova,
2004). These findings point to increased danger to
astronaut health. Conversely, however, biofilms are
commonly used for nitrification and organic carbon
removal of wastewater, making them suitable for

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
38 Gravitational and Space Biology 19(2) August 2006
wastewater treatment devices onboard future long-
duration space missions (Sharvelle et al., 2002). Because
biofilms will be present in the spacecraft environment,
understanding the effects of microgravity on biofilms will
be important for their control and exploitation.
How SMG growth affects biofilm resistance is therefore
of interest. However, this question has not been
adequately addressed because of a lack of earth-based
system to generate SMG-grown biofilms. Recently, a
modification of the HARV apparatus, involving the use of
appropriate beads, permitted facile bacterial biofilm
cultivation under SMG conditions on Earth (Lynch,
Mukundakrishnan, Ayyaswami and Matin, submitted).
SMG stimulated more copious biofilm formation and
these were much more resistant than their NG grown
counterparts to two antibiotics, penicillin and
chloramphenicol, as well as to the antimicrobial
treatments, such as exposure to high salt and ethanol. The
SMG-mediated enhanced resistance to salt and ethanol,
but not to the antibiotics, required the presence of σs in
the cells since an rpoS mutant failed to exhibit this
phenomenon for the former but not for the latter agents.
What aspect of the biofilms is controlled by σs that
enhances their resistance to various agents in a differential
manner has not as yet been determined. Lynch, Dixon and
Matin. (submitted) have, however, recently identified
another gene in uropathogenic E. coli which controls
biofilm resistance and whose mode of action has become
partially known. Comparison between biofilms of wild
type uropathogenic E. coli and a rapA mutant shows that
the rapA gene contributes to the increased resistance of
biofilms by promoting matrix exopolysaccharide
formation. When penetration of a fluorescent probe of
penicillin was visualized, it became clear that the
antibiotic penetrates the biofilm of the mutant much more
rapidly than that of the wild type. In addition, the rapA
gene is required for the expression of two genes that
appear to encode multidrug resistance pumps. Thus, this
gene appears to enable biofilms to utilize a dual strategy
for resistance to antimicrobials, retarded penetration and
more effective means of effluxing the antimicrobial that
does manage to finds its way into the biofilm cells. The
studies on the rapA gene have so far been done only
under the conventional conditions of normal gravity. How
SMG may affect these parameters is under investigation.
SMG AND SPACEFLIGHT EFFECTS ON
SECONDARY METABOLITE AND SPORE
PRODUCTION
In addition to the general stress response and increased
virulence, other survival strategies of bacteria include the
production of secondary metabolites, such as antibiotics,
which kill off competing organisms, and spore
production, which allows cells to wait out poor
environmental conditions. A few reports exist of
increased antibiotic production by bacteria on spacecrafts
(Lam et al., 1998; Luo et al., 1998; Lam et al., 2002;
Benoit et al., 2005). Following 15 days of Streptomyces
ansochromogenus growth onboard a Chinese satellite,
Luo et al. (1998) found that some mutants produced
increased concentrations of Nikomycin compared to
ground controls. A 17-day experiment conducted on the
Space Shuttle (STS-80) showed increased production of
Actinomycin D by Streptomyces plicatus. Due to
spaceflight hardware constraints, however, the absolute
production was low compared to conventional culture
methods. In a follow-up study, a newly designed space
bioreactor was used allowing antibiotic production at
similar concentrations to conventional methods. This
study involved a 72-day experiment conducted onboard
the International Space Station. Samples taken at days
eight and twelve showed increased spaceflight
production, but subsequent production was lower than the
matched ground controls.
Several ground-based studies of bacterial secondary
metabolite production have been conducted with HARV
bioreactors. In four of these experiments, secondary
metabolite production (of β-lactam antibiotics and
rapamycin) was inhibited by SMG compared to normal
gravity mode of the HARV (Fang et al., 1997a; Fang et
al., 1997c; Fang et al., 2000; Gao et al., 2001). In the fifth
experiment, gramicidin synthesis by Bacillus brevis was
unaffected by SMG (Fang et al., 1997b). However, the
site of antibiotic accumulation shifted from inside to the
outside of the cell, which is beneficial as it facilitates
purification. In addition, secondary metabolite production
in the RWV bioreactors was much lower than production
in shake flasks for all five studies. Demain and Fang
(2001) hypothesized that reduced shear stress in the RWV
bioreactors caused the reduced secondary metabolite
production. In support of this hypothesis, beads were
added to RWV bioreactors to increase shear stress,
resulting in increased production of secondary metabolites
(Fang et al., 2000; Gao et al., 2001). However, bacterial
production of a polymer has been shown to increase for
bacteria grown in a HARV compared to shake flask
cultures (Thiruvenkatam and Scholz, 2000).
In terms of spore production, Mennigmann and Lange
(1986) reported a lower number of spores from B. subtilis
cultures in spaceflight (5×104 spores ml
-1) compared to
ground controls (8×105 spores ml
-1). In a follow on
experiment, Mennigmann and Heise (1994) reported the
opposite trend: the ratio of spores from spaceflight to
ground cultures was 12.64 and the ratio of spores per cell
was higher for flight cultures (0.64) compared to ground
cultures (0.17). A centrifuge was used onboard the Space
Shuttle during this experiment to provide 1 g flight
control, which showed no significant difference in spore
production from 1 g ground control. Launch
accelerations and space radiation were therefore ruled out
as possible causes of altered spore production. Lam et al.
(2002) reported that cultures of Streptomyces plicatus
flown in space for 17 days sporulated profusely when
plated post-flight, but the ground controls lost their ability
to sporulate when similarly plated. Finally, Benoit et al.
(2005) reported that significantly less spores were
recovered from the spaceflight residual viable cultures.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 39
However, the post-flight sporulating ability of the flight
cultures was found to be approximately 8 times that of the
corresponding ground controls. In addition, scanning
electron micrograph images showed that spore
morphology was altered by spaceflight.
CONCLUSION
Spaceflight alters bacteria in several ways, but the
potential threat to humans is not well understood.
Rotating bioreactors such as the clinostat and HARV are
effective at simulating some aspects of microgravity for
bacterial cell cultures. While much remains to be learned,
it is clear already that SMG growth affects bacterial
resistance and virulence both in planktonic and biofilm
mode of growth in ways as to constitute a serious
potential threat to astronaut health. The molecular
mechanisms underlying this phenomenon remain largely
unexplored but initial indications are that novel, as yet
unknown, strategies may be involved. Besides being
potentially harmful, bacteria and their biofilms also have
beneficial uses, for example, in microbial-based
regenerative systems of waste water treatment. These
systems, commonly used on Earth, represent a viable
alternative to physical and chemical methods in use today
on the International Space Station. Long duration
missions planned for travel outside of low Earth orbit may
demand regenerative systems, which are intended to
minimize consumable mass. Thus, continued
investigation of molecular physiology of bacteria under
SMG is necessary to better control their harmful effects as
well as exploit their beneficial roles.
ACKNOWLEDMENT
Work reported from this laboratory was
supported by NASA grant NNA04CC51G.
REFERENCES
Albrecht-Buehler, G. 1992. The Simulation of
Microgravity Conditions on the Ground. ASGSB Bulletin
5(2): 3-10.
Becker, G., Klauck, E., and Hengge-Aronis, R. 1999.
Regulation of RpoS proteolysis in Escherichia coli: the
response regulator RssB is a recognition factor that
interacts with the turnover element in RpoS. Proceedings
of the National Academy of Sciences USA 96(11): 6439-
44.
Benoit, M. and Klaus, D. 2005. Can genetically modified
Escherichia coli with neutral buoyancy induced by gas
vesicles be used as an alternative method to clinorotation
for microgravity studies? Microbiology 151(1): 69-74.
Benoit, M. R., Li, W., Roane, T., Stodieck, L. S., Lam, K.
S., and Klaus, D. M. 2005. Microbial Antibiotic
Production Aboard the International Space Station.
Applied Microbiology and Biotechnology DOI:
10.1007/s00253-005-0098-3.
Brown, R. B., Klaus, D., and Todd, P. 2002. Effects of
Space Flight, Clinorotation, and Centrifugation on the
Substrate Utilization Efficiency of E. coli. Microgravity
Science and Technology XIII(4): 24-29.
Cioletti, L. A., Pierson, D. L., Mishra, S. K. 1991.
Microbial Growth and Physiology in Space: A Review.
In: International Conference on Environmental Systems,
SAE: 911512: 1-11.
Dedolph, R. R. and Dipert, M. H. 1971. The Physical
Basis of Gravity Stimulus Nullification by Clinostat
Rotation. Plant Physiology 47: 756-764.
Dickson, K. J. 1991. Summary of biological spaceflight
experiments with cells. ASGSB Bulletin 4(2): 151-260.
Fux, C. A., Costerton, J. W., Stewart, P. S., and Stoodley,
P. 2005. Survival strategies of infectious biofilms. Trends
in Microbiology 13(1): 34-40.
Gao, Q., Fang, A., Pierson, D. L., Mishra, S. K., and
Demain, A. L. 2001. Shear stress enhances microcin B17
production in a rotating wall bioreactor, but ethanol stress
does not. Applied Microbiology and Biotechnology 56(3-
4): 384-7.
Graebe, A., Schuck, E. L., Lensing, P., Putcha, L., and
Derendorf, H. 2004. Physiological, pharmacokinetic, and
pharmacodynamic changes in space. Journal of Clinical
Pharmacology 44(8): 837-853.
Hammond, T. G. and Hammond, J. M. 2001. Optimized
suspension culture: the rotating-wall vessel. American
Journal of Physiology. Renal Physiology 281(1): F12-25.
Hengge-Aronis, R. 2002. Signal transduction and
regulatory mechanisms involved in control of the
sigma(S) (RpoS) subunit of RNA polymerase.
Microbiology and Molecular Biology Reviews 66(3): 373-
95.
Jenkins, D. E., Chaisson, S. A., and Matin, A. 1990.
Starvation-induced cross protection against osmotic
challenge in Escherichia coli. Journal of Bacteriology
172(5): 2779-81.
Kacena, M. A., Manfredi, B., and Todd, P. 1999. Effects
of space flight and mixing on bacterial growth in low
volume cultures. Microgravity Science and Technology
12(2): 74-77.
Kenniston, J. A., Burton, R. E., Siddiqui, S. M., Baker, T.
A., and Sauer, R. T. 2004. Effects of local protein stability
and the geometric position of the substrate degradation
tag on the efficiency of ClpXP denaturation and
degradation. Journal of Structural Biology 146(1-2): 130-
40.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
40 Gravitational and Space Biology 19(2) August 2006
Klaus, D. M. 2001. Clinostats and bioreactors.
Gravitational and Space Biology Bulletin 14(2): 55-64.
Klaus, D. M. 2002. Space Microbiology: Microgravity
and Microorganisms. In G. Britton, (Ed.) The
Encyclopedia of Environmental Microbiology, NY: John
Wiley & Sons: 2996-3004.
Koenig, D. W. and Pierson, D. L. 1997. Microbiology of
the space shuttle water system. Water Science and
Technology 35(11-12): 59-64.
Lam, K. S., Gustavson, D. R., Pirnik, D., Pack, E.,
Bulanhagui, C., Mamber, S. W., Forenza, S., Stodieck, L.
S., and Klaus, D. M. 2002. The Effect of Space Flight on
the Production of Actinomycin D by Streptomyces
plicatus. Journal of Industrial Microbiology and
Biotechnology 29: 299-302.
Lam, K. S., Mamber, S., Pack, E., Forenza, S., Fernandes,
P., and Klaus, D. 1998. The Effects of Space Flight on the
Production of Monorden by Humicola fuscoatra WC5157
in Solid State Fermentation. Applied Microbiology and
Biotechnology 49: 579-583.
Lapchine, I., Moatti, N., Richoilley, G., Templier, J.,
Gasset, G., and Tixador, R. 1988. The Antibio
Experiment. In: Biorack on Spacelab D-1, ESA SP-1091:
45-51.
Luo, A., Gao, C., Song, Y., Tan, H., and Liu, Z. 1998.
Biological responses of a Streptomyces strain producing
Nikkomycin to space flight. Space Medicine & Medical
Engineering 11(6): 411-4.
Lynch, S. V., Brodie, E. L., and Matin, A. 2004. Role and
regulation of sigma(s) in general resistance conferred by
low-shear simulated microgravity in Escherichia coli.
Journal of Bacteriology 186(24): 8207-8212.
Lynch, S. V. and Matin, A. 2005. Travails of
microgravity: man and microbes in space. Biologist 52(2):
80-87.
Matin, A. 1991. The molecular basis of carbon-starvation-
induced general resistance in Escherichia coli. Molecular
Microbiology 5(1): 3-10.
Matin, A. 2001. Stress Response in Bacteria. In G.
Britton, (Ed.) Encyclopedia of Environmental
Microbiology, NY: John Wiley & Sons: 3034-3046.
Matin, A. and Lynch, S. V. 2005. Investigating the Threat
of Bacteria Grown in Space. ASM News 71(5): 235-240.
McLean, R. J. C., Cassanto, J. M., Barnes, M. B., and
Koo, J. H. 2001. Bacterial biofilm formation under
microgravity conditions. FEMS Microbiology Letters
195(2): 115-119.
Moore, D., Cogoli, A. 1996. Gravitational and Space
Biology at the Cellular Level. In D. Moore, P. Bie, & H.
Oser, (Eds.) Biological and Medical Research in Space,
Berlin, Germany: Springer: 1-106.
Nickerson, C. A., Ott, C. M., Mister, S. J., Morrow, B. J.,
Burns-Keliher, L., and Pierson, D. L. 2000. Microgravity
as a novel environmental signal affecting Salmonella
enterica serovar Typhimurium virulence. Infection and
Immunity 68(6): 3147-52.
Nickerson, C. A., Ott, C. M., Wilson, J. W., Ramamurthy,
R., LeBlanc, C. L., Honer zu Bentrup, K., Hammond, T.,
and Pierson, D. L. 2003. Low-shear modeled
microgravity: a global environmental regulatory signal
affecting bacterial gene expression, physiology, and
pathogenesis. Journal of Microbiological Methods 54(1):
1-11.
Nickerson, C. A., Ott, C. M., Wilson, J. W., Ramamurthy,
R., and Pierson, D. L. 2004. Microbial responses to
microgravity and other low-shear environments.
Microbiology and Molecular Biology Reviews 68(2): 345-
361.
Niesel, D. W., Williams, N., Pandya, U., Rask, J.,
Kakavand, K., Kirven-Brooks, M., and Aquilina, R. 2005.
SPEGIS: Streptococcus pneumoniae gene expression and
virulence potential in the space environment.
Gravitational and Space Biology 19(1): 36.
Novikova, N. D. 2004. Review of the knowledge of
microbial contamination of the Russian manned
spacecraft. Microbial Ecology 47(2): 127-132.
Ott, C. M., Bruce, R. J., and Pierson, D. L. 2004.
Microbial characterization of free floating condensate
aboard the Mir space station. Microbial Ecology 47(2):
133-136.
Pierson, D. L. 2001. Microbial contamination of
spacecraft. Gravitational and Space Biology Bulletin
14(2): 1-6.
Pierson, D. L. and Mishra, S. K. 2000. Space Flight,
Effects on Microorganisms. In J. Lederberg, (Ed.)
Encyclopedia of Microbiology, Second ed. San Diego.
Academic Press: 344-352.
Pyle, B. H., McFeters, G. A., Broadaway, S. C., Johnsrud,
C. K., Storfa, R. T., and Borkowski, J. 1999. Bacterial
Growth on Surfaces and in Suspensions. In M. Perry (Ed.)
Biorack on Spacelab, European Space Agency, SP-1222:
95-99
Schwarz, R. P., Goodwin, T. J., and Wolf, D. A. 1992.
Cell Culture for Three-Dimensional Modeling in
Rotating-Wall Vessels: An Application of Simulated
Microgravity. Journal of Tissue Culture Methods 14: 51-
58.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 41
Schweder, T., Lee, K. H., Lomovskaya, O., and Matin, A.
1996. Regulation of Escherichia coli starvation sigma
factor (sigma s) by ClpXP protease. Journal of
Bacteriology 178(2): 470-6.
Sharvelle, S., Silverstein, J., Klaus, D., and Supra, L.
2002. Nitrogen Removal from a Urine-Soap Wastewater
Using a Bioprocessor System: Process Monitoring and
Control. In: International Conference on Environmental
Systems, SAE: 2002-01-2353.
Sonnenfeld, G. and Shearer, W. T. 2002. Immune
function during space flight. Nutrition 18(10): 899-903.
Thiruvenkatam, R. and Scholz, C. 2000. Synthesis of
Poly(ß-hydroxybutyrate) in Simulated Microgravity: An
Investigation of Aeration Profiles in Shake Flask and
Bioreactor. Journal of Polymers and the Environment
8(4): 155-159.
Tixador, R., Gasset, G., Eche, B., Moatti, N., Lapchine,
L., Woldringh, C., Toorop, P., Moatti, J. P., Delmotte, F.,
and Tap, G. 1994. Behavior of Bacteria and Antibiotics
under Space Conditions. Aviation Space and
Environmental Medicine 65(6): 551-556.
Tixador, R., Richoilley, G., Gasset, G., Templier, J., Bes,
J. C., Moatti, N., and Lapchine, I. 1985. Study of Minimal
Inhibitory Concentration of Antibiotics on Bacteria
Cultivated Invitro in Space (Cytos-2 Experiment).
Aviation Space and Environmental Medicine 56(8): 748-
751.
Todd, P. and Klaus, D. M. 1996. Theories and Models on
the Biology of Cells in Space. Advances in Space
Research 17(6/7): 3-10.
Wilson, J. W., Ramamurthy, R., Porwollik, S.,
McClelland, M., Hammond, T., Allen, P., Ott, C. M.,
Pierson, D. L., and Nickerson, C. A. 2002. Microarray
analysis identifies Salmonella genes belonging to the low-
shear modeled microgravity regulon. Proceedings of the
National Academy of Sciences 99(21): 13807-13812.
Wolf, D. A. and Schwarz, R. P. 1991. Analysis of
Gravity-Induced Particle Motion and Fluid Perfusion
Flow in the NASA-Designed Rotating Zero-Head-Space
Tissue Culture Vessel. NASA Technical Paper 3143.
Wolf, D. A. and Schwarz, R. P. 1992. Experimental
Measurement of the Orbital Paths of Particles
Sedimenting Within a Rotating Viscous Fluid as
Influenced by Gravity. NASA Technical Paper 3200.

A. Matin — Increased Bacterial Resistance and Virulence in Simulated Microgravity
42 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 43
Symposium II: Astronaut Health: From
the Bench to Flight Across the Gravity
Continuum
Charles Wade, Editor

44 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 45
EXPLORATION CLASS MISSIONS AND RETURN: EFFECTS ON THE IMMUNE SYSTEM Gerald Sonnenfeld
Division of Research and Department of Biological Sciences
Binghamton University, State University of New York
Binghamton, NY 13902-6000, USA
ABSTRACT
Immune responses have been shown to be altered when humans,
animals and cell cultures are exposed to space flight conditions.
It is clear that, although there have been some issues regarding
infection on short-term space flights, alterations in the immune
system have not been a “show-stopper” for space flight missions
to date. However, as very long-term multi-year exploration
class missions are contemplated, the potential for space flight-
induced alterations of the immune system to have major impact
on crew health and success of the mission increases. The
increased concern is due to exposure to new conditions such as
novel forms of radiation, and the isolation of the crew from the
ability to obtain rapid relief and rescue due to health problems.
If, during space flight, immune capacity is diminished while
bacterial growth and virulence is enhanced, and antibody
efficacy is altered, a difficult situation could arise. If an
ineffective antibiotic is transported for use by the crew, there
will be no replacement readily available during an exploration
class mission. Additionally, radiation exposure combined with a
suppressed immune system could facilitate tumor development
after return to earth. In order to protect crews and assure the
success of exploration class missions, studies must be
undertaken in space, as well as on earth using ground-based
analog models, to fully establish the risk and to develop
countermeasures to prevent or minimized unwanted outcomes.
INTRODUCTION
It is clear that immune responses are dramatically altered
after space flight (Space Studies Board, 1998; Sonnenfeld
et al., 2003). Multiple experiments, including human,
animal and tissues cultures studies, have demonstrated
that a variety of immune responses are altered after
exposure to space flight conditions during actual flight
experiments or during experiments using ground-based
analog models that re-create some of the conditions of
space flight (Space Studies Board, 1998; Sonnenfeld et
al., 2003). Among the immune responses that have been
shown to be affected by space flight conditions are:
leukocyte blastogenesis (division of white blood cells
required to start immune responses), cytokine production,
leukocyte subset analysis, natural killer cell activity and
delayed hypersensitivity skin tests (Space Studies Board,
1998; Sonnenfeld et al., 2003).
Although it is clear that the immune response changes
during and after space flight, the biomedical significance
of these changes is still not fully established (Space
Studies Board, 1998; Sonnenfeld et al., 2003). There
have been reports of changes in resistance to infection
during space flight, with the development of a urinary
tract infection in one case (Space Studies Board, 1998;
Sonnenfeld et al., 2003). In the Apollo program, problems
with development of respiratory tract infections in crews
led to the development of the Crew Health Stabilization
program countermeasure of decreased interaction of
crews with other people on the ground still in use today
(Space Studies Board, 1998; Sonnenfeld et al., 2003).
Additionally, it is now clear that latent viruses, such as
Epstein-Barr virus, are reactivated in space flight crews
(Pierson et al, 2005; Shearer et al., 2005; Stowe et al.,
2001). Studies using the hindlimb unloading model of
rodents some space flight conditions have shown that
hindlimb unloading of mice has resulted in decreased
resistance to infection with bacteria and viruses
(Sonnenfeld et al., 2003). Therefore, the potential for
space flight conditions inducing changes in the immune
systems yielding altered resistance to infection and/or
tumors certainly exists. However, it is also clear that
decreased resistance to infection or tumors has not been a
“show-stopper” to date for the space flight program
(Space Studies Board, 1998; Sonnenfeld et al., 2003). In
almost every case, crews have been able to function
normally without serious incident.
However, we are now facing a new era of space flight, i.e.
the development of exploration class missions.
Exploration of space beyond low earth orbit has the
possibility of changing the conditions of the flight
environment to create greater risk.
THE EXPLORATION CLASS MISSION ISSUES
As we now move towards exploration class missions
beyond low earth orbit, we face new scenarios then what
has been seen before during space flight missions (Table
1). We will face new types of radiation that have never b
been encountered before by humans in space or, if they
have been encountered, humans have never been exposed
to them in space for the length of time they will face in
the future (Space Studies Board, 1996).
Additionally, crews will be away from earth for a more
extended period of time, with limited chances for very
rapid rescue, return or re-supply (Board on Health
Science Policy, 2001). This means a scenario could
develop where if an infection developed and an antibiotic
____________________
* Correspondence to: Gerald Sonnenfeld, PhD
Vice President for Research
Binghamton University
State University of New York
Post Office Box 6000
Binghamton, NY 13902-6000
Email: [email protected]
Phone: 607-777-4818; Fax: 607-777-2188

G. Sonnenfeld — Exploration Class Missions and Return: Effects on the Immune System
46 Gravitational and Space Biology 19(2) August 2006
was taken to which the organisms was resistant, there
would be limited opportunity to provide a different
antibiotic or to rapidly return the individual to earth for
additional treatment. Additionally there have been
unconfirmed reports of antibiotics not working as
effectively in space as on the ground (Space Studies
Board, 1998).
An additional challenge that may be faced was made clear
by some modeling experiments on earth (Nickerson et al.,
2001, 2004; Wilson et al., 2002). In these experiments,
bacteria were grown in clinostat-derived tissue culture
models that replicated some space flight conditions such
as low shear. These experiments clearly demonstrated
that growth in those conditions altered both the growth
rates of the bacteria and the expression of virulence
factors (Nickerson et al., 2001, 2004; Wilson et al., 2002).
Therefore, a possible scenario could develop where a
bacteria or virus that is part of the normal microbial
components of human microbial flora (that normally does
not cause a problem for the host) could have enhanced
growth and virulence in space, not respond well to the
available antibiotics, and the astronaut host for this
bacteria could have a space-flight impaired immune
response (that would not have a negative effects under
normal conditions). The possibility for a very serious
infectious disease problem that could threaten both the
health of the astronaut and the success of the mission is
obvious!
We must be prepared to both detect and deal with this
problem. There are several steps that should be taken
(Table 2). We must carry out experimentation in
appropriate space flight models on the ground and in
space to fully identify the risk. We must know what
changes happen to infectious agents and to the host
immune system under the conditions we will encounter in
exploration class missions.
We must know how to test whether the host is
compromised. This involves development of standards
for immunological parameters that need to be tested for
astronauts on a routine basis. This testing needs to be
carried out on the ground before flight to establish base
lines and on a regular basis before and after the mission.
We must assure that the standard testing can actually be
carried out in space both on the exploration vehicles and
on the ground on the Moon or Mars. Only then can we
see if the host’s immune defenses are compromised.
We must also assure that we are prepared to deal with
violations of standards induced by space flight conditions.
This would involve understanding the growth and
virulence characteristics of microorganisms that could be
potential pathogens under space flight conditions.
Additionally, there is a great need for testing of antibiotics
and antivirals to ensure that they actually function as they
are designed under space flight conditions. We must also
assure that countermeasure are developed that will
ameliorate or eliminate any negative effects of the space
flight environment on the immune system. In case of
antimicrobials or lack of provision of the appropriate
antimicrobials, be must be sure that the crews are as
immuncompetent as possible throughout their space flight
missions.

G. Sonnenfeld — Exploration Class Missions and Return: Effects on the Immune System
Gravitational and Space Biology 19(2) August 2006 47
CONCLUSION
Only by vigilance and preparedness can we avoid
unexpected deleterious effects of exploration class space
flights on the immune system and, therefore, the health of
the crews. A high level of preparedness will be required
to assure the health and survivals of exploration class
mission crews and the successful completion of
exploration class missions.
AKNOWLEDGEMENTS
Studies funded in the author’s laboratory were funded, in
part, by the National Aeronautics and Space
Administration through NASA Cooperative Agreement
NCC9-58 with the National Space Biomedical Research
Institute.
REFERENCES
Board on Health Sciences Policy, Institute of Medicine.
(2001). Safe passage:Astronaut care for exploratory
missions. National Academies Press, Washington, DC.
Nickerson, C.A., Goodwin, T.J., Terlonge, J., Ott, C.M.,
Buchanan, K.L., Uicker, W.C., Emami, K., LeBlanc,
C.L., Ramamurthy, R., Clarke, M.S., Vanderburg, C.R.,
Hammond, T., Pierson, D.L (2001)Three-dimensional
tissue assemblies: novel models for the study of
Salmonella enterica serovar Typhimurium pathogenesis.
Infect. Immun. 11:7106-7120.
Nickerson, C.A., Ott ,C.M., Wilson, J.W., Ramamurthy,
R., Pierson, D.L. (2004).
Microbial responses to microgravity and other low-shear
environments.
Microbiol. Mol. Biol. Rev. 68:345-361.
Pierson, D.L., Stowe, R.P., Phillips, T.M., Lugg, D.J.,
Mehta, S.K. (2005) Epstein-Barr virus shedding by
astronauts during space flight. Brain Behav. Immun.
19:235-242.
Shearer, W.T., Zhang, S., Reuben, J.M., Lee, B.N., Butel,
J.S.. (2005). Effects of radiation and latent virus on
immune responses in a space flight model.
J. Allergy Clin. Immunol.. 115:1297-1303.
Sonnenfeld, G., Butel, J.S., Shearer, W.T. (2003). Effect
of the space flight environment on the immune system.
Rev. Environ. Health 18:1-18.
Space Studies Board, National Research Council. (1996).
Radiation hazards to crews of interplanetary missions:
Biological issues and Research strategies. National
Academies Press, Washington, DC.
Space Studies Board, National Research Council. (1998).
A strategy for research in space biology and medicine in
the new century. National Academis Press, Washington,
DC.
Stowe, R.P., Mehta, S.K., Ferrando, A.A., Feeback, D.L.,
Pierson, D.L.. (2001). Immune responses and latent
herpesvirus reactivation in spaceflight. Aviat. Space
Environ. Med..72:884-891.
Wilson, J.W., Ott, C.M., Ramamurthy, R., Porwollik, S.,
McClelland, M., Pierson ,D.L., Nickerson, C.A. (2002).
Low-Shear modeled microgravity alters the Salmonella
enterica serovar typhimurium stress response in an RpoS-
independent manner. Appl. Environ. Microbiol. 68:5408-
5416.

G. Sonnenfeld — Exploration Class Missions and Return: Effects on the Immune System
48 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 49
NUTRITION, METABOLISM AND THE CRITICAL PATHS: A CRITICAL REVIEW T.P. Stein
Department of Surgery, University of Medicine and Dentistry of New Jersey - SOM, Stratford, NJ, 08084
ABSTRACT
A mission to Mars is estimated to take ~2.5 years. However
there remain some problems which until they are resolved will
limit how long humans can remain away from earth. The
biomedical counter-measures part of the critical paths has
evaluated these risks. They include, bone loss, cardiovascular
effects, muscle loss, nutrition and radiation exposure. Three of
the risks are related to metabolism, namely bone loss, muscle
atrophy and nutrition. Probably the most encouraging aspect of
these problems is that they are known, and because they are
important for human health on the ground there are reasonably
effective treatments available for the ground analogs. However
it still remains to confirm that the actual proposed flight
protocols are effective, first on the ground (with analogs of
flight hardware) and then inflight.
INTRODUCTION
Forty years have elapsed since the first humans first
ventured into space. By any standards the venture has
been successful; humans survive well enough that
manned missions to Mars are being planned. A mission
to Mars is estimated to take ~2.5 years. However there
remain some problems which until they are resolved will
limit how long humans can remain away from earth.
Probably the most encouraging aspect of these problems
is that they are known, and because they are important for
human health on the ground there are reasonably effective
treatments available for the ground analogs.
During the last few years NASA has systematized its
approach to developing a counter-measures program. The
process is the bioastronautics critical paths road map. The
road map lists 50 risks. The document is a living object,
undergoing frequent updates. (For the latest update, see
www.criticalpath.jsc.nasa.gov). The risks have been
divided into three color coded categories; potential show
stoppers (red, R), serious risk of mission impairment
(yellow, Y), and probably controllable (green, G). The
risks have been further sub-divided into 551 ‘critical
questions’. As with the risks, some questions are more
critical than others.
Risks in the biomedical counter-measures part of the
critical paths include, bone loss, cardiovascular effects,
muscle loss, nutrition and radiation exposure. Three of
the risks are related to metabolism, namely bone loss,
muscle atrophy and nutrition. They are the topic of this
review. Table 1 shows an abbreviated version of the
critical paths for the bone, muscle and nutrition
disciplines.
Table 1: Critical paths with metabolic/nutritional aspects.
The objectives of the critical paths road map is to assess
risks for human space exploration and to use this
information for prioritizing research and technology
programs. Defining the questions and targets provides a
means assessing progress towards the reduction and
deciding when the level of risk is low enough to be
acceptable.
BONE The losses are primarily from the weight bearing bones.
Figure 1 shows a summary of some of the available bone
data compiled by A.D. LeBlanc. Note the high degree of
individual variation. There is also much variation in loss
sites. Bone loss is unevenly distributed.
There are two counter measures for bone loss. Firstly
the use of anti-resorptive drugs (e.g. bisphosphonates).
Millions of people already take bisphosphonates as a
treatment for osteoporosis. Bisphosphonates have also
been found to be successful in long duration bed rest
studies (Shackelford et al., 2004). A second treatment for
bone loss is to place load on bone by resistance exercise.
This too has been found to be effective in attenuating the
bone loss during bed rest (Cavanagh, Licate and Rice,
2005; Shackelford et al., 2004). Thus two independent
counter-measures for the bone loss have been found to be
effective in the best ground based model for space flight
(Alkner and Tesch, 2004; Iwase et al., 2004; Shackelford
et al., 2004).
DISCIPLINE ISS RISK (1 yr) ISS
(1 yr)
MOON
(30d)
MARS
(2.5 yr)
Bone Bbone loss and
fracture risk
Y G Y
Bone Impaired fracture
healing
G G R
Bone Injury to joints and
interveterbral
structures
Y Y Y
Bone Renal stone
formation
G G G
Muscle Skeletal muscle
atrophy with
reduced strength
and endurance
G G Y
Muscle Increased
susceptibility to
muscle damage
G G Y
Nutrition Inadequate
nutritional
requirements
G G Y
____________________
* Correspondence to: T.P. Stein, PhD
Department of Surgery
University of Medicine and Dentistry of New Jersey - SOM
Stratford, NJ, 08084
Email: [email protected]
Phone: 609-566-6036

T.P. Stein — Nutrition, Metabolism and the Critical Paths: A Critical Review
50 Gravitational and Space Biology 19(2) August 2006
% C
HA
NG
E I
N B
ON
E D
EN
SIT
Y
-20
-15
-10
-5
0
5
10
COSMONAUTS ASTRONAUTS BEDREST ALENDRONATE RES. EXERCISE
Figure 1: Bone loss with space flight, bed rest and treatments.
Data supplied by Dr. A.D. LeBlanc, USRA)
The reduction of the bone loss may not be complete or
equally effective for all bones, but judging by the
published data, it is better than 80%. Reducing the bone
loss by a minimum of 80% makes the problem
manageable. Consider a worst case scenario. According
to recent flight data from the ISS, the highest bone loss
rate observed for an astronaut/cosmonaut was ~2% per
month (Lang et al., 2004). Reducing this by 80% reduces
the rate to 0.4% per month. For a 2.5 year round trip this
translates into a maximum loss of 12%. For 6 months ISS
stay the corresponding value is 2.4% which for a worst
case is acceptable. NASA’s management seems to have
bought into this argument. They have decided to give US
astronauts a bisphosphonate (Alendronate) starting in
2007.
Since the number of future US astronauts is going to be
somewhere between 20 and 40 before the shuttle is retired
means there will be little further opportunities to evaluate
any other in-flight measures for long duration missions
between now and 2010. It is not likely that there will be
other long duration opportunities between 2010 and the
start of the Mars expeditions 30 years from now.
Between shuttle and Mars there will be lunar missions.
The lunar missions will be only about 30 days in duration.
It would therefore seem that NASA has made the decision
that the bone ‘counter-measure’ problem is well on the
way to being solved. However it still remains to confirm
that the actual proposed flight protocols are effective, first
on the ground (with analogs of flight hardware) and then
in-flight. If the studies are successful, it would be
appropriate for NASA to descope ‘bone’ and its
associated questions from the critical paths and redirect
resources to other high priority risks.
MUSCLE
Skeletal muscle is plastic, responding to external
workloads. Less work is required during space flight and
bed rest from the anti-gravity muscles because there is no
longer the need to support the body mass. The result is a
reductive remodeling of the anti-gravity muscles to adapt
to the reduced work-load (Grigoriev and Egorov, 1992;
Thornton and Rummel, 1977).
In addition to a net loss of protein, there is a shift in
myosin isoforms from slow to fast isoforms (Fitts, Riley
and Widrick, 2000). Fast twitch fibers are primarily
glycolytic and prone to fatigue with endurance. This
metabolic shift towards increased reliance on glycolysis is
found with space flight (Baldwin, Herrick, and McCue,
1993), the rat hind limb suspension model (Fitts, Riley
and Widrick, 2000; Henriksen and Tischler, 1988;
Langfort et al., 1997) and bed rest (Acheson, et al., 1995).
Glycolysis is very effective for high intensity short
duration acute activities, but if sustained output is needed,
an energy profile where fat use is favored is desirable. An
inability to sustain work output is a major cause for
concern. Astronauts need to maintain as much functional
capacity as possible during space flight for extra-
vehicular activities (space station construction and
maintenance, emergency egress and eventually exploring
Mars. These are all functions that require sustained work
output. Less protein to do the work and increased
fatigability in this context are counter-productive.
A recent Soyuz landing clearly illustrated the seriousness
of this problem. After a 5½-month flight, the Soyuz
landed somewhere in Kazakhstan. It took more than 5 h
to recover the crew. For safety reasons the crew had to
get out of the landing capsule unassisted. It took them
five hours to accomplish what should have been a half
hour task. The misadventure provides strong evidence for
impaired muscle functionality after Mars-like transit,
demonstrating serious weaknesses in crew performance.
All three crew members exhibited reduced capability, up
to voluntary immobility. More work is needed on the
muscle atrophy problem. Because of the limitations of
the space craft environment it may not be possible to
totally prevent the reductive remodeling of skeletal
muscle, but certain functional capacities must be retained.
NASA has defined what these capabilities are for a
successful Mars mission. They include the need for
dexterity, essential for manipulating tools and performing
complex actions; examples given by NASA are deploying
solar arrays and erecting a habit; good hand eye
coordination is needed for driving the mission exploration
vehicle and for teleoperating robotic aides. There is of
course a need for maintaining strength, flexibility, agility
for such essential tasks as putting on and taking off the
pressure suits and most importantly the ability to move
away from danger should the situation merit. All of these
functions are well met by maintaining a similar level of
physical fitness as pre-flight.
There is no longer any doubt that exercise is effective in
ground based models for preventing disuse muscle
atrophy (Bamman et al., 1998; Ferrando, et al., 1997;
Shackelford et al., 2004). However the results have not
been replicated in flight – beginning with Skylab and
continuing through shuttle, MIR and the ISS. Indeed,

T.P. Stein — Nutrition, Metabolism and the Critical Paths: A Critical Review
Gravitational and Space Biology 19(2) August 2006 51
exercise has been part of US and Russian missions for
more than 30 years; yet the problem persists (Cena,
Sculati and Roggi, 2003; Smith, et al., 2005; Stein, 2001).
In the first 10 ISS missions full functionality of the
exercise equipment has been less than 50% and this
together with the inadequate dietary intake accounts for
the unsatisfactory results with the current in-flight
exercise regimens (Smith, et al., 2005, Stein, 2001).
ENERGY BALANCE
Energy deficits have been found on most, but not all short
duration shuttle missions and has been a consistent
finding with long duration missions (Rambaut, Leach and
Leonard, 1977; Rambaut, et al., 1977). The negative
energy balance is a mission related effect on dietary
intake and not a specific response to the absence of
gravity. Dietary intake is mission dependent rather than
subject dependent (figure 2, (Stein, 2001, Stein, 2000))
Astronauts on the same mission appear to eat about the
same amount of food (figure 2).
DAYS
EN
ER
GY
IN
TA
KE
(kca
l. k
g-1
.d-1
)
0
10
20
30
40
50
LMS
SLS1/2
SKYLAB 2
SKYLAB 3
SKYLAB 4
5 10 15
Figure 2: Comparison of energy intake during the first two
weeks of space flight for the 3 Skylab missions, shuttle SLS1/2
and shuttle LMS (Stein, 2001).
Elsewhere, we have suggested that the inverse
relationship between exercise and energy intake is due to
problems in disposing of the metabolic by-products from
exercise, namely heat and CO2 (Stein, 2000). Thermo-
regulatory mechanisms are less efficient during space
flight and this persists into the immediate post flight
period (Acheson, et al., 1995; Fortney et al., 1998). Heat
disposal has been shown to depress intake in both rats
(Llamas-Lamas and Combs, 1990) and humans (Edholn,
Fox, Goldsmith et al., 1964).
A negative energy balance is not an appropriate
physiologic state when the need is to counter-act a net
muscle catabolic state. Proteins are the 'machinery' of the
body. All of the metabolic functions in the body, from
cell division to obtaining energy from foodstuffs to host
defense mechanisms to doing muscle work involve
proteins. So it is important to prevent the breakdown of
body protein for use as an energy source.
A similar degree of weight loss together with poor intake
has been found with ISS astronauts. The same degree of
poor intake and weight loss (4.4 + 0.1 kg, n=6) that was
found on MIR (Stein et al., 1999) is now being found with
ISS crews (3.8 + 0.1 kg, n=24, personal communication,
S.M. Smith, NASA and (Smith, et al., 2005)).
DAYS IN ORBIT
0 50 100 150 200%
BO
DY
WT
LO
SS
-14
-12
-10
-8
-6
-4
-2
0
2
4
6
Figure 3: Weight change with flight on the ISS (Smith, 2005).
There is great variation on the magnitude of the weight
loss. It is not known whether the weight loss is
continuous or whether an adaptation/accommodative state
is achieved. Figure 3 shows data from the ISS (Smith, et
al., 2005). The lines represent individual weight changes.
The two heavy black lines represent least squares
regression analysis of the data with either a linear or
curvilinear fit. The curve fits suggests that a steady state
is reached after about 100 days. The linear regression
suggests that the weight loss is continuous. Both
regressions are statistically significant. Which is correct?
If the weight loss is continuous, then the problem is very
serious, chronic weight loss is unsustainable over the 2.5
yr period it will take to go to Mars. On the other hand,
establishment of an adapted state after ~100 days may be
acceptable. This question needs to be addressed.
As yet there is no counter-measure being evaluated for the
inadequate intake. We would like to suggest one.
Weekly measurement of body mass with dietary
consultation with the ground support team. In contrast to
dietary records, measurement of body mass is a real
measurement and can be directly related to energy
balance status. Astronauts losing weight inflight should
be encouraged to eat more. Body mass devices have been
flown on many prior missions beginning with Skylab and
including shuttle.

T.P. Stein — Nutrition, Metabolism and the Critical Paths: A Critical Review
52 Gravitational and Space Biology 19(2) August 2006
REFERENCES
Acheson KJ, Decombaz J, Piguet-Welsch C, Montigon F,
Decarli B, Bartholdi I, and Fern EB. Energy, protein, and
substrate metabolism in simu-lated microgravity. Am J
Physiol (Regul and Integ) 269: R252-260, 1995.
Alkner BA and Tesch PA. Efficacy of a gravity-
independent resistance exercise device as a
countermeasure to muscle atrophy during 29-day bed rest.
Acta Physiol Scand 181: 345-357, 2004.
Baldwin KM, Herrick RE, and McCue SA. Substrate
oxidation capacity in rodent skeletal muscle: effects of
exposure to zero gravity. J Appl Physiol 75: 2466-2470,
1993.
Bamman MM, Clarke MS, Feeback DL, Talmadge RJ,
Stevens BR, Lieberman SA, and Greenisen MC. Impact
of resistance exercise during bed rest on skeletal muscle
sarcopenia and myosin isoform distribution. J Appl
Physiol 84: 157-163., 1998.
Cavanagh PR, Licate AA, and Rice AJ. Exercise and
pharmacological countermeasures for bone loss during
long duration space flight. Gravitational and Space
Biology Bulletin 18: 39-58, 2005.
Cena H, Sculati M, and Roggi C. Nutritional concerns and
possible countermeasures to nutritional issues related to
space flight. Eur J Nutr 42: 99-110, 2003.
Edholn O, Fox GRH, Goldsmith IFG, and al. e. Report to
the MRC, No. APRc64/C65. London, Army personnel
research Committee, 1964. London: HMSO, 1964.
Ferrando AA, Tipton KD, Bamman MM, and Wolfe RR.
Resistance exercise maintains skeletal muscle protein
synthesis during bed rest. J Appl Physiol 82: 807-810,
1997.
Fitts RH, Riley DR, and Widrick JJ. Physiology of a
microgravity environment. J Appl Physiol 89: 823-839.,
2000.
Fortney SM, Mikhaylov V, Lee SM, Kobzev Y, Gonzalez
RR, and Greenleaf JE. Body temperature and
thermoregulation during submaximal exercise after 115-
day spaceflight. Aviat Space Environ Med 69: 137-141,
1998.
Grigoriev AI and Egorov AD. Physiological aspects of
adaptation of main human body systems during and after
spaceflights. Advances in Space Biology & Medicine 2:
43-82, 1992.
Henriksen EJ and Tischler ME. Glucose uptake in rat
soleus: effect of acute unloading and subsequent
reloading. J Appl Physiol 64: 1428-1432., 1988.
Iwase S, Takada H, Watanabe Y, Ishida K, Akima H,
Katayama K, Iwase M, Hirayanagi K, Shiozawa T,
Hamaoka T, Masuo Y, and Custaud MA. Effect of
centrifuge-induced artificial gravity and ergo-metric
exercise on cardiovascular deconditioning, myatrophy, &
osteoporosis induced by a -6 degrees head-down bedrest.
J. Grav. Physiol: 11: P243-244, 2004.
Lang T, LeBlanc A, Evans H, Lu Y, Genant H, and Yu A.
Cortical and trabecular bone mineral loss from the spine
and hip in long-duration spaceflight. Journal of Bone &
Mineral Research 19: 1006-1012, 2004.
Langfort J, Zernicka E, Mayet-Sornay MH, Dubaniewicz
A, and Desplanches D. Effects of acute and chronic
hindlimb suspension on sensitivity and responsiveness to
insulin in the rat soleus muscle. Biochem Cell Biol 75:
41-44, 1997.
Llamas-Lamas G and Combs DK. Effects of
environmental temperature and ammoniation on
utilization of straw by sheep. J Anim Sci 68: 1719-1725,
1990.
Rambaut PC, Leach CS, and Leonard JI. Observations in
energy balance in man during spaceflight. Am J Physiol
(Endo and Metab) 233: R208-212, 1977.
Rambaut PC, Smith MC, Jr., Leach CS, Whedon GD, and
Reid J. Nutrition and responses to zero gravity. Fed Proc
36: 1678-1682, 1977.
Shackelford LC, LeBlanc AD, Driscoll TB, Evans HJ,
Rianon NJ, Smith SM, Spector E, Feeback DL, and Lai
D. Resistance exercise as a countermeasure to disuse-
induced bone loss. J Appl Physiol 97: 119-129, 2004.
Smith SM, Zwart SR, Block G, Rice BL, and Davis-Street
JE. The nutritional status of astronauts is altered after
long-term space flight aboard the International Space
Station. J Nutr 135: 437-443, 2005.
Stein TP. Nutrition in the space station era. Nutr Res Rev
14: 87-114, 2001.
Stein TP. The relationship between dietary intake,
exercise, energy balance and the space craft environment.
Pflugers Arch 441: R21-31., 2000.
Stein TP, Leskiw MJ, Schluter MD, Donaldson MR, and
Larina I. Protein kinetics during and after long term space
flight on MIR. Am J Physiol (Endo and Metab) 276:
E1014-1012, 1999.
Thornton W and Rummel J. Muscular deconditioning and
its prevention in space flight. Biomedical results from
Skylab (NASA SP-377) US Government Printing Office,
Washington, DC: 191-197, 1977.

Gravitational and Space Biology 19(2) August 2006 53
TESTING EXERCISE COUNTERMEASURES DURING 30 DAYS OF SIMULATED
MICROGRAVITY: LESSONS LEARNED FROM STUDIES OF IDENTICAL TWINS
Alan R. Hargens, Brandon R. Macias, Cece M. Echon, Eva Brzezinski, Andrea Hawkins, Kristina Hawkins, and
R. Scott Meyer
Department of Orthopaedic Surgery, University of California, San Diego, San Diego, CA 92103-8894
ABSTRACT
This paper reviews our experience with 30-day head-down tilt
(HDT) bed rest studies of supine lower body negative pressure
(LBNP) treadmill exercise countermeasure in identical twins.
We review our experience with 16 sets of identical male and
females twins in order to improve future countermeasure studies
for long-duration space flight. All male sets of twins and all but
one female set tolerated the study well and no medical
complications occurred. Our experimental design and use of
identical twins may represent an innovative method for
evaluating the physiologic efficacy of a treatment when
comparing an exercise versus control group. Also, our
nutritional procedures maintained body weight in both groups.
Pre- and post-bed rest tests were scheduled to avoid
confounding effects due to test order and test volume. As
reported elsewhere, our treadmill exercise protocol within LBNP
maintained plasma volume, orthostatic responses, upright
exercise capacity, muscle strength and endurance during bed
rest. Future bed rest studies should focus on the recruitment of
healthy, non-sedentary subjects and may want to follow our
recommendations in terms of screening, testing sequence,
nutrition, medical care and subject support. Identical twins offer
unique insights into countermeasure development and
heritability of physiologic traits.
KEY WORDS
Bed rest, recruitment, testing sequence, nutrition, identical
twins
INTRODUCTION
Microgravity leads to cardiovascular deconditioning in
humans, which is manifested by post-flight reduction of
orthostatic tolerance and upright exercise capacity
(Watenpaugh and Hargens, 1996). During upright posture
on Earth, blood pressures are greater in the feet than at
heart or head levels due to gravity’s effects on columns of
blood in the body (Hargens et al., 1992). Upon exposure
to microgravity, all gravitational blood pressures
disappear. Presently, there is no exercise hardware
available for space flight to provide this gravitational
blood pressure to tissues of the lower body.
Lower-body musculoskeletal loss is experienced by crew
exposed to long-term space flight (Arnaud and Morey-
Holton, 1990; LeBlanc et al., 1996; Morey-Holton et al.,
1996; Schneider et al., 1989; Thornton and Rummel,
1977; Whedon et al., 1977). Although physical activity is
considered the best way to protect bones, muscles and the
cardiovascular system, exercise protocols and equipment
for simulated and actual microgravity are still unresolved
(Convertino, 1990). For example, studies of simulated
microgravity by Vernikos and associates (1996) indicate
that standing for four hours completely prevents and
standing for two hours partially prevents decreases in
post-bed rest orthostatic intolerance. Walking for four
hours prevents and walking for two hours attenuates
decreases in peak oxygen uptake compared to non-
exercise controls. They conclude that various
physiological systems benefit differentially from passive
gravity/activity in one g. Moreover, in addition to the
duration of the stimulus, the number of exposures to
upright posture are important as well (Vernikos et al.,
1996).
Recent calculations suggest that all exercise in space to
date has lacked sufficient loads to maintain pre-flight
bone mass (Cavanaugh et al., 1992, Whalen et al., 1988).
Although treadmill exercise with bungee cords (about 2 hr
per day) was the most common exercise for cosmonauts
during long-duration Mir missions, biomechanical loads
on musculoskeletal tissues of the lower body are only
about 60-70% of those present on Earth (Whalen, 1993).
One example of the seeming lack of exercise
effectiveness is that three Mir 18 crewmembers varied
widely in their amounts of exercise performed, while their
post-flight muscle volumes (LeBlanc et al., 1996) and
bone resorption (Smith et al., 1999) were nearly identical.
Theoretically, an integrated countermeasure for extended
exposure to microgravity should combine high loads on
the musculoskeletal system (Whalen et al., 1988) with
normal regional distributions of transmural pressure
across blood vessels (Hargens et al., 1992) and
stimulation of normal neuromuscular locomotor patterns.
We have postulated that LBNP treadmill exercise may
prevent bed rest- and microgravity-induced
deconditioning by simulating gravity effects. Static
ground reaction force (GRF) in a LBNP chamber is a
product of the body cross-sectional area at the waist seal
(Axy) and the pressure differential between the external
ambient and internal chamber environments (∆P), where
∆P = LBNP: GRF = Axy × ∆P. For the average male
subject, an additional GRF of about one equivalent body
weight (BW) is generated for each 100 mmHg of LBNP
when the negative pressure acts only through the cross-
sectional area of the subject’s waist (Hargens et al., 1991).
____________________
* Correspondence to: Alan R. Hargens, PhD
Department of Orthopaedic Surgery
University of California, San Diego
350 Dickinson St. San Diego, CA 92103-8894
Email: [email protected]
Phone: 619-543-6805; Fax: 619-543-2540

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
54 Gravitational and Space Biology 19(2) August 2006
The LBNP exercise concept avoids the discomfort of
localized high pressures typical of bungee cord harness
systems by distributing the net force of the air pressure
uniformly over the entire upper surface of the body. By
expanding the area through which the pressure produces
force, we found that the amount of negative pressure
required to generate one BW could be decreased. If the
waist seal area equals twice the subject’s waist cross-
sectional area, the negative pressure necessary to produce
one BW decreases from 100 mmHg to 50-55 mm Hg
(Watenpaugh et al., 1994). The reduced negative pressure
required to generate one (BW) of force lowers the risk of
excessive heart rate, footward fluid redistribution,
syncope, hernia, and petechiae associated with higher
levels of LBNP (Wolthius et al., 1974).
In our recent studies of identical twins exposed to 30 days
of bed rest and reported elsewhere (Cao et al., 2005;
Hargens et al., 2003; Macias et al., 2005; Monga et al.,
2006a,b; Smith et al., 2003), we hypothesized that 40
minutes of supine treadmill running per day in a LBNP
chamber at 1.0 to 1.2 BW (approximately 50-60 mmHg
LBNP) with a 5-minute resting, non-exercise LBNP
exposure at 50 mmHg after the exercise session maintains
aerobic fitness, orthostatic tolerance, and musculoskeletal
structure and function during 30 days of bed rest
(simulated microgravity). The purpose of the present
paper is to review our experience with 15 sets of identical
male and females twins in order to improve future
countermeasure studies for long-duration space flight.
EXPERIMENTAL DESIGN
To initiate the project, we proposed to study 16 sets of
identical, female and male twins following a thorough
medical examination to ensure their suitability for a safe
and well-controlled study. Acceptable subjects were
thoroughly briefed by the investigating team and provided
informed, written consent before participation in this
study.
Identical twin sets were randomly divided into two groups
to investigate the mechanism of action and efficacy of our
partial vacuum exerciser concept. These 30-day bed rest
studies were chosen to approximate longer-term
microgravity exposures. Our previous shorter-term bed
rest studies (5 and 15 days) achieved significant results
with seven subjects (Lee et al., 1997, Watenpaugh et al.,
2000). By coin toss selection, one twin exercised while
their sibling was a non-exercise control subject. The
exercising twin ran in the supine LBNP chamber for 40
minutes plus a 5-minute static LBNP period. They
exercised once a day at 1.0 to 1.2 BW of footward force,
while their sibling was a non-exercise “control” subject.
The interval exercise protocol was similar to that
employed by us in our previous two week studies: 7 min
warm-up at 40% peak oxygen uptake, followed by 3 min
at 60%, 2 min at 40%, 3 min at 70%, 2 min at 50%, 3 min
at 80%, 2 min at 60%, 3 min at 80%, 2 min at 50%, 3 min
at 70%, 2 min at 40%, 3 min at 60%, and 5 min cool-
down at 40% peak oxygen uptake (40 min total) with an
additional 5 min of supine, stationary exposure to 50
mmHg LBNP (Greenleaf et al., 1989; Lee et al., 1997).
Exercise bouts occurred at the same time of day for each
subject in the exercise group. During bed rest, the LBNP
treadmill exercise was undertaken six days per week with
Sundays allowed for rest. All exercise subjects were able
to carry out this program unless exercise was too
uncomfortable due to menstrual cramps, illness, muscle
soreness, knee pain, or initial adaptation to the suspension
system. In these cases, slower treadmill speeds or as a last
resort, lower levels of LBNP, were instituted.
A 3-day orientation period occurred prior to the start of
bed rest to acquaint subjects with our facilities, personnel,
procedures and experiments, including orthostatic
tolerance, muscle strength and exercise capacity tests. The
first phase of the bed rest study (including 6 days of
baseline testing, orientation, familiarization, 30 days of
bed rest, and 3 days of recovery) began in mid-1999. All
subjects reported to the UCSD General Clinical Research
Center (GCRC) on their designated start day. Subjects
were usually discharged from the GCRC 39 days later.
In the baseline control period, activity levels were
maintained by each subject, and ambulatory levels of
plasma and urinary markers of bone loss were measured
on the three days prior to bed rest. All physiologic tests
took place at the same time of day for a given subject.
These tests were staggered so that sufficient time was
allowed to complete each procedure and to prevent
confounding effects of one test on another test. While the
subjects lived at the GCRC, their daily sodium intake was
controlled at approximately 170 mEq (3.5 g per day) and
their diet also was controlled (approximately 2500-3000
kcal per day, depending on exercise level). Their body
weight, fluid intake, and urine output were monitored on a
daily basis. We expected that subjects would maintain a
neutral or positive fluid balance and neutral energy
balance so that they would not lose weight during 30 days
HDT. During the entire period of bed rest, all subjects
were required to remain in 6° HDT except during periods
for bath/shower and exercise (0.5-1.5 h/day), when they
were horizontal (0°). Compliance to this strict bed rest
protocol was required of all subjects so that the scientific
integrity of the results could be maintained.
In the baseline control period, the Hamilton Depression
Rating Scale and Beck Depression Inventory were
administered to each subject to assess any pre-study
depressive symptoms. These measures were re-
administered during the last week of the bed rest period.
Prior to initiation of HDT, plasma volume, leg and spinal
muscle strength, Dual Energy X-ray Absorptiometry
(DXA), spine Magnetic Resonance Imaging (MRI), heart
muscle mass, arm venous pressure, and plasma and
urinary bone markers were measured in all subjects
(Table 1). On the day before initiation of HDT, all
subjects were tested chronologically for orthostatic
tolerance, sprint speed, postural stability, gait parameters,
and upright peak oxygen uptake. By using this order of

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 55
tests, we think we were able to avoid possible
confounding effects of one test on a subsequent test. For
example, the plasma volume and orthostatic tolerance
tests were scheduled before peak upright exercise
capacity to eliminate the effect of elevated plasma volume
due to exercise on peak upright exercise capacity. During
the recovery period these tests were repeated in the exact
same sequence within 4 hr of return to upright posture to
maintain comparability. We were usually able to test
subjects at a rate of four per day in alphabetic order with
tests on one set of twins undertaken in the morning and on
a second set of twins in the afternoon. This was important
so that subjects had time to rest, eat and digest their
meals, adequate time for setup and troubleshooting
equipment was allowed and we could finish testing of
subjects before dinner.
An attempt was made to stagger the subjects with respect
to time of HDT initiation and return to upright posture to
allow investigators to perform all tests without delays.
Subjects were active and took controlled walks during
pre-bed rest and recovery days to avoid the well-known
deconditioning due to confinement and inactivity.
Experimental stations for pre- and post-bed rest tests
included: 1) bedside plasma volume early in morning, 2)
orthostatic tolerance, 3) sprint speed, 4) posture and
balance time, and 5) posture/gait analysis and peak
upright exercise capacity. There also was a room
specifically set aside for LBNP exercise. We provided
TVs, VCR players, DVD players, a computer room with
internet access, bed-side phone, and we allowed cell
phones in order to help prevent boredom. We found the
volume and frequency of tests represented an optimized
compromise between an insufficient number of
physiologic tests and too many tests that would
overburden and psychologically stress our volunteers. For
example, none of our subjects quit before or after any of
our tests, even though our testing protocol was time
consuming and physically demanding over the three day
periods before and after bed rest.
Typically, three full days were needed to set-up
equipment. We had three separate testing rooms: an
exercise room, an orthostatic tolerance testing room and
the upright treadmill testing room for peak V02. We
performed sprint testing in a hallway of the hospital that
was manned by many research and GCRC personnel to
maintain safety. However, some twins fell down during
their sprint test but none suffered injury. All subjects were
able to sprint a second time.
Two staff members were needed to operate the LBNP
exerciser. It took about 15-20 minutes to setup the subject
for exercise. Each exercising subject was transferred from
their bed to an HDT gurney and from the gurney to the
LBNP exercise apparatus. Subjects brought along their
drinking bottles and urinals to the exercise session. One
staff member applied leg straps to the subject, while the
other staff member prepared the waist seal, shoulder
straps, treadmill and heart rate monitor. One staff member
controlled the negative pressure in the chamber and
treadmill speed, while the second staff member recorded
time, HR and perceived exertion data. The second staff
member was always in immediate contact with the
subject. Also, we provided water in a container with a
flexible straw so they could drink in supine position. It
was also required to have two people in the room at all
times in case of an emergency, one to attend to the subject
and the other to call for help. An orthopaedic surgery
resident from our Department at UCSD was on call for
the duration of the study.
Scheduling tests and making final equipment check-ups
occurred one month in advance. Some of our testing was
offsite. We rented a UCSD vehicle to transport subjects so
that all subjects were covered by UCSD insurance. Our
bed rest project involved collaboration with many US and
international investigators and required data management
and communication to occur in a timely manner.
Frequently, for example, exercise logs of HR were
transformed into a .pdf file and transmitted electronically
to a co-investigator reviewing exercise work loads. Also,
scanning documents into .pdf files allowed for immediate
review of data by national and international collaborators,
and it allowed for a back-up system of data storage.
Scheduling of tests supported by the UCSD Medical
Center (plasma volume, MRI, echocardiography) required
an appointment at least one month in advance. This was
an issue because we needed to test four subjects
concurrently which required large blocks of clinical time
to be set aside.
LESSONS LEARNED FROM RECRUITMENT OF
TWINS
Twinsworld.com, an internet newsletter, was the best
source for recruiting twins into our study. Another source
of twins existed in the lab of an investigator at UCSD
who performs research on twins. This investigator agreed
to mail out a flyer about our study to his previous
subjects. This was helpful in recruiting two sets of local
twins, in case twins outside the San Diego area failed to
report or were subsequently disqualified. Recruitment was
a time consuming and challenging effort. To save time we
asked twins to respond to some questions for screening
purposes; for example, age, history of smoking and
exercise, fitness, weight and height to calculate BMI, and
previous absence from home for 40 days. If twins passed
these initial screening questions, we then explained our
study in detail. If they were still interested, we obtained
all of their contact information (phone number, cell phone
number, mail address, email address, parent’s phone
number) and mailed them our consent form and a video of
previous subjects exercising in the LBNP chamber. A
week later we called to follow up if they were still
interested in our study. Also, we arranged a
teleconference with both twins to answer specific
questions about bed rest, testing and exercise. Most twins
were concerned about gaining weight, using the bed pan
and monetary payment. Many twins could not or did not
want to participate because of job or family commitments.
The older twins expressed interest, but their established

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
56 Gravitational and Space Biology 19(2) August 2006
Table 1. Daily order of events
Day Procedure Time required
Familiarization day 1 Report to GCRC, check in 15 min
Orthostatic tolerance 90 min
Peak upright exercise capacity 60 min
Noninvasive Intracranial Pressure (ICP) 30 min
Familiarization day 2 Muscle strength 90 min
LBNP exercise for selected twin 90 min
Familiarization day 3 Plasma volumes and hematocrit 30 min
Regional arterial compliance 15 min
Echocardiography 60 min
Control day 1 Serum and 24 hr urine collection 30 min
Discomfort questionnaire 15 min
Orthostatic tolerance 90 min
Sprint speed 10 min
Posture and balance analysis 20 min
Peak upright exercise capacity 60 min
Control day 2 Serum and 24 hr urine collection 15 min
Discomfort questionnaire 15 min
DXA 60 min
Muscle strength 90 min
Regional arterial compliance 15 mm
Sleep quality 9 hr
Control day 3 Serum and 24 hr urine collection 30 min
Discomfort questionnaire 15 min
Spinal compression and heart mass 60 min
Hamilton and Beck tests 30 min
Sleep quality 9 hr
Bed rest day 1 Begin bed rest
Discomfort questionnaire 15 min
Bed rest (all days) Discomfort questionnaire 15 min
Bed rest Exercise for one twin 90 min
(all days except 7, 14, 21, 28, and 30)
Bed rest 4,11,18,25 Noninvasive ICP 30 min
Transcranial Doppler (TCD) 30 min
Bed rest 5,12,19,26 Serum and 24 hr urine collection 30 min
Bed rest 17/18, 26/27 Sleep quality 9 hr during sleep
Bed rest 14 Regional arterial compliance 15 min
Bed rest 24 Hamilton and Beck test 30 min
Bed rest 28 Spinal compression and heart mass 60 min
Bed rest 30 Plasma volume and hematocrit 30 min
Echocardiography 60 min
Regional arterial compliance 15 min
Recovery day 1 Serum and 24 hr urine collection 30 min
Discomfort questionnaire 15 min
Orthostatic tolerance 90 min
Sprint speed 10 min
Posture and balance analysis 20 min
Peak upright exercise capacity 60 min
Recovery day 2 Serum and 24 hr urine collection 15 min
Discomfort questionnaire 15 min
DXA 60 min
Muscle strength 90 min
Recovery day 3 Discomfort questionnaire 15 min
Ambulation, discharge from GCRC 60 min

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 57
careers and family obligations usually prevented us from
recruiting them. However, younger twins, especially those
in college, had the time during the summer and were
usually without family and work commitments. We relied
heavily on the twins’ answers to our questions because we
could not support their travel to UCSD for selection
purposes. Only one set of twins could not participate
because of their extremely low fitness level, i.e., they
were unable to jog. Otherwise, all the other twins who
traveled to UCSD for the study were healthy and
reasonably fit candidates. Typically we flew twins to San
Diego one day before familiarization day 1. We had a list
of items we asked the twins to bring: black exercise shorts
for gait studies, comfortable running shoes and socks,
running shorts and t-shirts, toiletries, clothes for 2 weeks,
bathroom air freshener, videotapes and DVDs, music,
books, and a camera. All subjects were encouraged to
have a personal goal during their stay at UCSD. Affiliated
Genetics performed zygosity tests to verify that the twin
sets were monozygotic. The test kit was sent by mail and
the company provided results in a timely manner.
We had an orientation the afternoon all twins arrived in
order to review the consent form, all protocol risks and so
they could meet key research, medical and GCRC
personnel. Then twins were asked to sign both the UCSD
and NASA Johnson Space Center (and initially NASA
ARC) consent forms. After consenting to the study, the
twins were admitted to the UCSD GCRC and history and
physical exams were performed. It typically took one
week for the exercising subject to adjust and attain their
prescribed exercise protocol speeds. The twins liked to
watch TV and/or listen to music during the LBNP
exercise session. It was important to make sure that the
twins had eaten and used the bed pan and/or urinal prior
to exercise. Also, the twins brought their graduated water
container to each exercise session.
LESSONS LEARNED FROM CARE OF TWINS IN
THE UCSD GCRC
Conducting a clinical research study on the effects of
microgravity in the GCRC was interesting and
challenging from the Nurse Manager’s standpoint. It
required weeks of planning and preparation that consisted
of the following: 1) assessment of physical and staff
resources, 2) procedural preparation, including label and
tube set-up, 3) coordination of activities that required
accurate and frequent communication, 4) collection and
management of data, 5) management of subject care, and
6) management of staff, investigators’ and subjects’ issues
and concerns.
Physical Resources:
For the nurse manager, identifying a space to house large,
heavy equipment such as the LBNP exercise chamber and
treadmill apparatus required departmental and
administrative approvals from the Medical Center CEO
and from the Director of Patient Care Services. In
addition, locating a comfortable and functioning bed scale
was also a challenge. An accurate bed scale is essential to
measure daily weights of all twins so that diets can be
modified to maintain constant body weight. Usually, bed
scales were assigned to critical areas in the hospital such
as the ICUs. Careful negotiation with our Clinical
Engineering Department provided the four bed scales
needed upon admission of each two sets of twins. The bed
scales required fine-tuning and calibrations at repeated
intervals prior to use.
The GCRC is often not equipped with resources suitable
to conduct bed rest studies for an extended period of forty
days. One example is the shower room. It is located on
the 5th
floor of the hospital in the Burn Unit. The
availability of the shower room depended on the schedule
for the burn patients. The shower was not readily
accessible for our twins after the 40-minute daily exercise
Monday through Saturday. The twins were hot and
sweaty and this situation sometimes resulted in
inconvenience and annoyance. Twins were transported
within the hospital via gurney with HDT to maintain the
head-down posture for all subjects. Other resources
needed for the study and located outside the hospital were
the MRI Facility and the bone density (DXA) scanner.
The transport of patients to these outside facilities became
an issue and required the involvement of our Risk
Management Office due to liability issues. A waiver
allowed use of a UCSD van by the research personnel
versus a more expensive ambulance. A research-
designated MRI facility and DXA scanner would be
optimal to prevent scheduling conflicts and liability
issues.
Staffing Resources:
The staffing patterns were adjusted to meet the needs of
the study. Experimental volunteers on bed rest are
considered “total care” subjects; hence contract labor was
utilized to augment the staffing for the evening and night
shifts.
Procedural Preparation:
Preparation of computerized labels, tube set-up (185
tubes/per twin), charts, log sheets, pain questionnaires,
physician’s orders for the entire 40-day admission for two
to three sets of twins required two full-time nurses. These
two research nurses worked together and coordinated the
daily care and schedule of the GCRC staff. They updated
the entire staff including the dietitians and sleep
technicians on a daily basis. Communication among the
staff and research team was extremely critical in
preventing scheduling dilemmas. Communication with
the twins was equally critical to maintain compliance and
positive attitude with the study. Often the twins wanted a
set schedule and to know at least a day in advance when a
particular test would occur during the bed rest period.
Therefore, the subjects were provided summarized testing
schedules. For example, our twins preferred to watch TV
programs and if they were involved in a test, they
preferred to record the TV show on video.
Data Collection and Management:
Data collection and management required extreme

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
58 Gravitational and Space Biology 19(2) August 2006
attention to detail due to the volume generated during
each procedure time. Correct tube labeling with the twin’s
identifier, date, time, and type of sample was checked
carefully and matched with the log sheet. The
computerized labeling of sample tubes was efficient and
clear, but became problematic when it was discovered
that the label used was not intended for long-term storage.
Dry ice delivery and supply was increased by two-fold to
maintain the quality and integrity of the samples.
Frequent auditing and monitoring of the data sheets
helped achieve accuracy, quality and completeness. Dry
ice was needed to maintain urine and serum samples at
−79° Celsius for sample storage within the hospital and
for shipment via overnight air mail. Additionally, labeling
the samples at times was an issue because the ink faded.
This required the GCRC to purchase a labeling system
with special ink for dry ice storage samples.
Management of Care for Our Twin Subjects:
Primary care was the model used for patient care. Ideally
two RNs were in charge of the total care for the twins.
However, with the GCRC being small and understaffed,
the care was divided among the staff on the day shift
during the 40-day admission period. The first day began
with the orientation of the twins to the unit’s routine
which included the following:
• Meal time
• Bedtime
• Phone system
• Entertainment
• Massage therapist
• Volunteer services (haircut, manicure, and pedicure)
• Chaplain services
• Open door policy to monitor subject well-being and
compliance to strict HDT
• Visiting hours
• Sleep studies
• Protocol for obtaining daily metabolic weight by the
night nurse
• Daily strict intake and output measurements
• Availability of the internet access
• Laundry services
The maintenance of strict HDT bed rest was emphasized
and all orientation sessions with the volunteers,
investigators and staff ended with a question and answer
period. With this orientation process, many of the twins
had difficulty remembering the open door policy. This
was a challenge with which the nurses were faced with
quite frequently. As one staff member stated, “I have an
issue with trust in caring for the twins.” There were a few
sets of twins who wanted their door closed at all times. A
one-way window or a camera in the room may be a good
addition in designing a bed rest study room in the future
to be able to monitor subject well-being and compliance
to HDT more effectively.
During the first week of the bed rest study, the biggest
dilemma for the female twins was adapting to the female
urinal and bedpan, while remaining in the required HDT
posture. Some twins had so much difficulty that they
would end up in tears out of frustration. Some would even
“hold it” for as long as they could to avoid using the
bedpan which sometimes resulted in being constipated.
This was probably the biggest complaint of the twins.
There is a need to design a practical “female urinal” and a
bed that can convert into a commode while maintaining
the HDT position. One possible solution may be a bed
with a hole in the middle such a “cholera cot.”
Because each of the twin pairs was in the same room for
almost 24 hours a day for 40 days, some twins developed
“cabin fever” during which they would constantly bicker
and sometimes throw objects at each other. In one
instance, one twin fell off the bed during a heated
argument with their sibling. The personality of the twins
played a major role in the degree of ease for their care.
Those who were young, gregarious, and college students
were easy to care for and had an easier time fulfilling the
requirements of the study. The staff spent more time in
the room with them compared to those who had a litany
of complaints whenever they were visited by the staff.
Unfortunately, because of being on bed rest, the twins
spent a majority of the time in their rooms, so trying to
keep them entertained was also difficult. On some days,
twins became so bored with the monotonous daily routine
that they appeared depressed and withdrawn. The staff
would try to encourage conversation, a new activity or
take them outside for some “fresh air” and sunlight to
boost their spirits. Thus, it is very important for bed rest
subjects to keep themselves busy with a goal or interest
throughout their entire period of confinement. On the
other hand, we found that twin pairs often supported each
other in completing their full period of bed rest.
Management of Staff, Research Team and Twins
Complaints:
Working with a diverse group of people is exciting,
interesting and requires tact. Issues and concerns need to
be dealt with in a fair manner and also need to be brought
to the table and investigated in an objective fashion. The
skills and knowledge of the Principal Investigator were
very important to complete the project successfully.
Issues and concerns were dealt with on a professional
level. The GCRC staff (nurses, dieticians and sleep
technicians) worked hard to perform a meaningful study
and the last female set of twins made the completion of
the bed rest study a great pleasure.
LESSONS LEARNED FROM NUTRITION OF
TWINS
Prior to initiation of the study, an 8-day menu cycle was
developed using Nutritionist IV analysis program. These
menus had a consistent macronutrient composition of
15% protein, 55% carbohydrate, and 30% fat. Sodium
was kept constant at 3.5 grams/day. Other nutrients that
were controlled included calcium (between 800 to 1000
mg/day) and fiber (1 gram per 100 calories or at least 20

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 59
grams for subjects whose menus were less than 2000
calories). Menus were randomly assigned throughout the
study.
Foods that were used to develop the menus were “typical
American foods” that are easily obtained through the
years and easy to store and prepare. Some of the foods
included dry cereal, low fat or non fat milk, yogurt,
American cheese, Swiss cheese, chicken breast, canned
tuna in water, whole wheat bread, white rice, pasta,
tomatoes, romaine lettuce, salad dressing, and tapioca
pudding. In order to control sodium, we used distilled
water to cook all the foods as well as for ice and drinking
water. Once the menus were calculated, salt was added to
meet the sodium requirement of the study. After the
menus were developed and tested, a food preference sheet
was developed and used for each participant. Once food
preferences were known, menus were individualized.
Calories were determined using the Harris-Benedict
equations plus an activity factor. Weight goals for each
subject were determined by using the average of the
initial two through four days of HDT bed rest. This
allowed time for the cephalad fluid shift and loss of body
weight during the first day of bed rest due to diuresis.
Calories were adjusted to maintain the constant body
weight within 1 kg. Importantly, we were able to maintain
body weight in exercise and control groups, thus avoiding
confounding effects of variable weight changes on the
post-bed rest test parameters. A detailed record of all
foods and fluids consumed was kept and all such data
were later provided to the investigators.
The following are key points/issues for the success of
future bed rest studies from a nutritionist’s point of view:
1) use foods that are easy to obtain all year long, 2)
develop menus with variety to avoid food boredom, 3)
explain with as much detail as possible the diet the
subjects will be following, 4) no matter how much you
explain, subjects will have issues with food, 5) use the
same nutrition analysis program for the entire study, 6)
keep in mind that food formulations change overtime, so
make sure to update values as needed, and 7) be flexible
and have fun with the food preparation. Twins were very
aware of the menu and that items repeated day to day. A
creative chef to develop meals that can be easily adjusted
in composition is recommended.
Non-select menus offered to the subjects became more
streamlined and labor-intensive menu items were omitted.
Subjects were contacted a month before the start of the
study to obtain a list of foods that they do not eat. Menus
were then created in advance that would work for all the
subjects. Nutrition staff communicated several times a
day with the subjects. They then communicated with the
nutrition manager (who was in charge of menu updates
and changes) regarding changes in food preferences so the
diets could be altered and adjusted accordingly.
As the menus became more streamlined, the food
ordering and preparation became more accurate. Storage
problems and over-ordering of food were prevented. At
the inception of the study we were weighing out raw
foods before cooking, and then reweighing the cooked
food before serving the subject. We found this to be an
unnecessary and time-consuming step. The diets
gradually incorporated more pre-packaged cold foods
(juices, granola bars) that helped streamline food
preparation time. As a result, volume of food presented to
the subjects gradually decreased. We found that the
volume of food presented to the subjects in the beginning
was too great. Foods were too bulky and overwhelmed the
subjects (salads, vegetables for example), especially with
the requirement that all subjects eat all food. We made
diet modifications to balance food volume (adding more
juices for example) and thus alleviated volume problems.
The subjects’ tight schedules because of testing and
exercise sessions sometimes resulted in dinner served
very late or breakfast very early. The problem was
compounded if they were slow eaters. Very early and very
late eating schedules were more labor-intensive for the
Nutrition and GCRC staff. The more we were able to
streamline the subjects’ schedules so they had
overlapping meal times, the more efficient our nutrition
team became with meal preparation and delivery.
Subjects’ satisfaction also increased.
Lastly, data entry was important to maintain on a
consistent basis in order to assess the subjects’ energy
balance. Towards the end of the study, we found it
beneficial to enter nutrition data (on Excel spreadsheets)
on a daily, versus weekly basis.
LESSONS LEARNED FROM MEDICAL CARE OF
TWINS
The identical twins used in this study were healthy
volunteers between 21 and 48 years old and provided
informed, written consent prior to arriving at the UCSD
GCRC. On the first day in the GCRC, a complete history
and physical exam were performed on each subject to
ensure that all inclusion/exclusion criteria were met. All
male subjects were free from medications. If female
subjects were taking prescribed oral contraceptives, these
medications were continued for the duration of the study
as the risk of deep vein thrombosis during bed rest was
considered low in this normal population. A urine
pregnancy test was administered on each female subject
and all results were negative. Two of our female subjects
were using inhalers for mild asthma and were allowed to
use this medication as needed and one female subject had
a history of gastroesophageal reflux disease and was
placed on famotidine, histamine-2 receptor antagonist.
These subjects developed no problems related to their
disease or medications. The physical examinations
revealed no significant findings and all subjects were
allowed to participate in the study.
During their 3-day familiarization period, the subjects
underwent orthostatic tolerance and exercise capacity
tests. These tests were then administered again two days
later to obtain baseline, pre-bed rest data and immediately
after 30 days of bed rest. All subjects performed well

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
60 Gravitational and Space Biology 19(2) August 2006
during orthostatic tolerance testing and no subject
experienced syncope or any other medical problem. All
subjects gave excellent efforts during exercise capacity
testing and no medical problems were encountered during
this test either. One set of female twins, however, had
significantly limited exercise tolerance and, after
determining that neither of them could complete the
LBNP exercise familiarization session, they were
disqualified from the study prior to the start of bed rest.
Despite their lack of admission of smoking during initial
screening, both of these twin subjects were found to be
smokers with relatively sedentary lifestyles during the
baseline testing period. In future bed rest studies, it is
important to screen out subjects who have this type of
lifestyle. However, in the past we trusted that all subjects
were honest in reporting their qualifying habits and
activity levels prior to arrival at the UCSD GCRC.
Shortly after being placed into HDT bed rest, all subjects
reported mild back pain. This pain typically lasted a few
days and was never associated with any neurologic
complaint or deficit. Several exercising subjects
developed mild discomfort in their lower extremities and
one female subject developed tendonitis of her knee. Her
physical examination and an MRI of the knee failed to
show any intra-articular derangement. She was able to
work through her discomfort and complete the study
without any problems. No exercising subject developed a
specific problem, such as hernia, from the lower body
negative pressure exercise.
During the 30 days of HDT bed rest, there were no
unexpected events in any of the male subjects and all, but
one, of the female subjects. When some subjects
developed minor problems related to HDT bed rest, they
sometimes tried to self-diagnose and exaggerate the
problem. This occurrence was frustrating to the attending
physician and nursing staff, especially if similar problems
occurred frequently for a given subject. One female
subject developed constipation and abdominal pain and,
despite a normal medical workup and physical
examination, she and her twin sister disqualified
themselves from the study several days into the bed rest
phase. An overriding issue was their inability to comply
with the strict HDT regulation during use of the bed pan.
These twin sisters were very close to the 50 year age limit
for our protocol and were relatively less physically fit
than the other subjects. It is likely that older subjects are
less able to adapt to the rigors of HDT bed rest compared
to younger volunteers. This may be important to consider
when recruiting subjects for other bed rest studies. All
other subjects completed the entire 30 days of bed rest
without difficulty. In fact, a common finding was that
each twin helped support their sibling to complete the
entire study.
In summary, all male sets of twin and all but one female
set tolerated the study well and no medical complications
occurred. Future bed rest studies should focus on the
recruitment of healthy, young, non-sedentary subjects.
Mild controlled medical conditions such as asthma or acid
reflux are not contraindications to participating in bed rest
studies successfully.
PERSPECTIVES FROM ONE SET OF TWIN
VOLUNTEERS
Participating in the NASA Twin Study was a great
experience for both of us (Andrea Hawkins and Kristina
Hawkins). Like anything in life, there were challenges
that we inevitably encountered. We entered this study
expecting to encounter many challenges, but it was
surprising that we had more positive experiences than
negative experiences. We used each day as a new way of
making the next day more enjoyable. It was an
opportunity that many people aren’t given and we’re both
glad we seized it.
Using the “female urinal” was by far the most challenging
aspect of the study. It was such a foreign object to us and
we both went into bed rest with the mind-set of “we’ll
worry about it when the time comes.” This was a bad
idea. It just caused a lot of anxiety and frustration. A
week later, we became more at ease with the whole
situation. We highly recommend getting comfortable with
the urinal prior to bed rest. After my twin sister told me
that the handle had to be facing upward, I tried it again
and it worked fine. That is why we recommend that other
female subjects try to get comfortable with the urinal
prior to bed rest. Once we mastered the urinal, drinking
water became more enjoyable and in the long run was
probably better to maintain fluid balance and digestive
processes.
The bedpan was also a very unpleasant issue for us.
Similar to the urinal, it just took time to adjust to a new
way of doing business. The entire process was never
anticipated. It was awkward in the beginning because we
did not quite understand how we could do it without
making a mess. Once we used it the first time, it was fine.
We just made it part of our routine to use it immediately
after being weighed in the morning. The nurses did their
best to make us feel comfortable. We felt guilty that they
had to come to collect our “stuff,” but they were very
good about it. It was more of an issue for us than it was
them. After a week or so, it was no big deal. It was
actually better having my sister in the room with me
because she would make me laugh every time I had to use
the urinal or bed pan. We think it is unlikely that we
would be able to take care of these normal body functions
in a room of other subjects without significantly more
privacy.
As far as physical discomfort, the most significant pain
we encountered was an intense headache. Day one of bed
rest was the most severe. Within hours of bed rest the
pressure built up in our heads from the HDT and caused
an intense headache. By the next day we had adjusted to
the change. We discovered that every time we went
outside for a short period of fresh air and sun, we’d
always develop a headache an hour after re-entering our
room. We were given 400 mg of Motrin pain reliever for

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 61
our headaches, which really did not take the pain away
completely, so we found that putting an ice pack over our
eyes and turning the lights off helped ease the pain.
Minor challenges we encountered on a daily basis were
morning weigh-ins and late showers. We were awakened
every morning at 6am and weighed. The routine entailed
our first morning urination, putting on a hospital gown,
transferring to a gurney, and transferred back to the bed
for weighing, which was not a challenge, but more of a
minor inconvenience. Another adjustment was the
evening shower. We were always accustomed to
showering first thing in the morning, so it took some
adjusting to showering at night. Every morning we would
“bathe” with a tub of water and washcloths. It was not the
same, but it helped. It was a slight challenge to shower
horizontally because the shower bed was also tilted down.
The slippery surface and soapsuds caused us to slide
down. Laying a towel beneath us prevented the sliding
problem.
Challenges for control, non-exercise twin
We were fortunate to have the support of each other
throughout this journey. We were always able to keep
each other’s spirits up by making each other laugh. There
were brief moments when we’d had time to ourselves.
These were times when one of us was showering and the
exercise twin was running. Those moments were often
used to write in journals. However, after a while, boredom
set in and in a situation like this, boredom is the enemy.
We would find things to do such as e-mailing friends on
the computer or socializing with the nurses. The nurses
played a crucial role in making this a great experience for
us. They kept us entertained throughout the days. We
tried not to watch TV throughout the day because it made
us feel like we were “sick patients” in bed. We found it
more enjoyable to watch movies at night.
Challenges for exercise twin
The first exercise on the treadmill was the most difficult
of all. Running on a vertical LBNP treadmill in supine
posture is different from any other form of running. The
first problem I encountered was developing a rash from
wearing shorts during the run. Because of the rubbing
from the shorts, I developed a painful rash during a few
more exercises. After the first week, the exercise started
to become more familiar. Having a visual print out of my
exercise protocol allowed me a greater chance of meeting
my prescribed exercise goals and knowing in advance
what was expected prior to my run made me more
comfortable. I discovered during the first run how
important it was to use the urinal before each run. Even
though I didn’t have the urge to urinate, I made sure I
always tried because once I was inside the chamber the
negative pressure seemed to expand my bladder. A V02
test was performed once a week during the study. This
posed a challenge because I ran with a mouthpiece in
place that restricted heavy breathing. To help eliminate
outside stressors, it was helpful to listen to my Walkman
and run with my eyes closed. This gave me the chance to
listen to my favorite songs and daydream about the
outside world.
Useful Tips
A significant amount of time is needed to become familiar
with everything and become comfortable with the new
environment. We have compiled some beneficial
instructions that could help future subjects have a more
enjoyable experience. The most valuable suggestion we
can offer is to familiarize oneself with the urinal prior to
bed rest. The first week is somewhat overwhelming so if
you can eliminate the stress of getting comfortable using
the urinal ahead of time, it will make the transition more
comforting. It would be useful for subjects who are
dependant on caffeine to eliminate it at least a week prior
to bed rest. This is something that might prevent
headaches during the initial phase of bed rest. Also,
waking up at 6am every morning is an adjustment. We
suggest that the subjects go to sleep early and avoid
napping during the day.
Finding things to do during the day may pose a challenge
for some subjects. We found it very useful to incorporate
activities of daily living into our bed rest routine. This
included: grooming, eating, correspondence, and reading.
Showering was always an anticipated activity during the
day. To avoid sliding during the shower we recommend
lying on top of a towel. Also, we suggest keeping a towel
near bedside to avoid getting the pillow wet after a
shower.
Being confined to the same room for a long period of time
can lower your spirits. We found it uplifting to have our
room decorated with familiar items from home such as
pictures, flowers, blankets, and other memorable items.
No matter how unmotivated we were feeling, we always
kept our room bright by having the curtains opened. It
always cheered us up and helped to maintain a positive
attitude.
QUANTITATIVE ADVANTAGES OF USING
IDENTICAL TWINS
In the present review, we compared paired analysis
against an unpaired analysis to determine if we needed
more subjects to detect significant differences in select
physiologic tests. As previously stated, our experimental
design and subsequent analyses were intended to use a
paired model with identical twins.
Paired Model
Eight male and seven female identical twin sets were
recruited. One twin was randomly assigned to the control
group (CON) and his or her twin was assigned to the
exercise group (EX) via a coin toss. The CON and EX
groups were compared before bed rest using a paired t-
test to determine if the measured parameters were
significantly different. Paired t-test revealed that the

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
62 Gravitational and Space Biology 19(2) August 2006
groups were not significantly different. Based on these
results and the assumption that the identical twins have
identical genetics, paired t-tests were preformed
comparing the orthostatic tolerance test time, treadmill
exercise capacity test time, maximal oxygen consumption,
and urine markers in the CON and EX group. After bed
rest, the paired t-test yielded significant differences
between the CON and EX groups (Table 2).
Unpaired Model
Sixteen males and fourteen females were recruited. The
subjects were randomly assigned to the CON and EX
group. The CON and EX groups were compared using an
unpaired t-test to determine if the measured parameters
were significantly different before bed rest. Unpaired t-
test revealed that the groups were not significantly
different before bed rest. The following parameters,
orthostatic tolerance test time, treadmill exercise capacity
test time, maximal oxygen consumption, muscle back
strength, and urine markers were compared using an
unpaired t-test after bed rest between the CON and EX
group. The unpaired t-test p-values for orthostatic
tolerance test time, maximal oxygen consumption, muscle
back strength, and urine markers were not significantly
different between the CON and EX groups (Table 2).
Table 2: P Values Obtained Using Paired and Unpaired Models
Paired p-value Unpaired p-value
VO2pk 0.001 0.2
VORTTT <0.0005 0.0005
TUVol. <0.05 >0.25
Ca 0.01 0.15
Oxalate 0.055 >.25
OT Time 0.0025 0.1
Back Strength 0.005 0.2
Key: Orthostatic tolerance (OT); Time Treadmill exercise
capacity test time (VOKTT); maximal oxygen
consumption (VO2pk); urine markers (TUVol., Ca,
Oxalate)
When using an unpaired model, p-values increase,
suggesting that we would need to study more subjects in
order to detect a significant difference with bed rest. The
paired model using identical twins as used in a crossover
study decreases the group variation. Some of the loss in
statistical power using an unpaired t-test may be due to
pooling male and female data. For example, extensive
previous research has documented that orthostatic
tolerance time, exercise capacity and muscle strength are
less in females than males. Therefore, when pooling male
and female data, a larger range and variance occurs,
reducing statistical power when using an unpaired t-test.
SUMMARY
Our experimental design and use of identical twins may
represent an innovative method for evaluating the
physiologic efficacy of a treatment when comparing
experimental and control groups. As reported elsewhere
(Cao et al., 2005; Hargens et al., 2003; Macias et al.,
2005; Monga et al., 2006b; Smith et al., 2003), our
treadmill exercise protocol within LBNP maintained
plasma volume, orthostatic responses, upright exercise
capacity, muscle strength and endurance during bed rest.
Future bed rest studies should focus on the recruitment of
healthy, non-sedentary subjects and may want to follow
our recommendations in terms of screening, testing
sequence, nutrition, medical care and subject support.
Identical twins offer unique insights into countermeasure
development and heritability of physiologic traits (Monga
et al., 2006 a).
ACKNOWLEDGMENTS
The authors gratefully acknowledge the enthusiastic
participation of all of our outstanding twins, excellent
support from staff at the UCSD GCRC, MRI and DXA
facilities. This study was supported by NASA grants 199-
26-12-34 and NAG 9-1425 as well as by an NIH grant to
the UCSD GCRC M01RR00827.
REFERENCES
Arnaud SB, Morey-Holton E. 1990. Gravity, calcium, and
bone. Physiologist 33:S65-S68.
Cao P, Kimura S, Macias BR, Ueno T, Watenpaugh DE,
Hargens, AR. 2005. Exercise within lower body negative
pressure partially counteracts lumbar spine
deconditioning associated with 28 day bed rest. Journal
of Applied Physiology 99:39-44.
Cavanagh PR, Davis BL, Miller TA. 1992. A
biomechanical perspective on exercise countermeasures
for long term spaceflight. Aviation, Space, and
Environmental Medicine 63:482-485.
Convertino VA. 1990. Physiological adaptations to
weightlessness: effects on exercise and work
performance. Exercise Sports Science Review 18:119-66.
Greenleaf JE, Bulbulian R, Bernauer EM, Haskell WL,
Moore T. 1989. Exercise-training protocols for astronauts
in microgravity. Journal of Applied Physiology 67: 2191-
2204.
Hargens AR, Whalen RT, Watenpaugh DE, Schwandt
DF, Krock LP. 1991. Lower body negative pressure to
provide load bearing in space. Aviation, Space, and
Environmental Medicine 62:934-937.
Hargens AR, Watenpaugh DE, Lee SMC, Boda WL,
Smith SM, Macias B, Groppo E, Schneider S, O’Leary D,
Meyer RS, Kawai Y. 2003. Physiologic countermeasures
for long-duration space flight: review of treadmill
exercise within lower body negative pressure. Journal of
Adaptation Medicine 7:2-6.
Hargens AR, Watenpaugh DE, Breit GA. 1992. Control

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
Gravitational and Space Biology 19(2) August 2006 63
of circulatory function in altered gravitational fields.
Physiologist 35:S80-S83.
LeBlanc A, Lin C, Rowe R, Belichenko O, Sinitsyn V,
Shenkman B, Oganov V, Shackelford L and Feeback D.
1996. Muscle Loss After Long Duration Spaceflight on
Mir 18/STS-71. AIAA Life Sciences and Space Medicine
Conference, (Abstract 96-LS-71): 53-54.
Lee SMC, Bennett BS, Hargens AR, Watenpaugh DE,
Ballard RE, Murthy G, Ford SR, Fortney SM. 1997.
Upright exercise or supine lower body negative pressure
exercise maintains exercise responses after bed rest.
Medicine and Science in Sports and Exercise 29:892-900.
Macias BR, Groppo ER, Eastlack RK, Watenpaugh DE,
Lee SM, Schneider SM, Boda WL, Smith SM, Cutuk A,
Pedowitz RA, Meyer RS, and Hargens AR. 2005. Space
exercise and Earth benefits. Current Pharmaceutical
Biotechnology 6:305-317.
Monga M, Macias B, Groppo E, and Hargens A. 2006a.
Genetic heritability of urinary stone risk in identical
twins. Journal of Urology 175: 2125-2128.
Monga M, Macias B, Groppo E, Kostelec M, and Hargens
AR. 2006b. Renal stone risk in a simulated microgravity
environment: impact of treadmill exercise with lower
body negative pressure. Journal of Urology 176: 127-131.
Morey-Holton ER, Whalen RT, Arnaud SB, Van der
Meulen, MC. 1996. The skeleton and its adaptation to
gravity. In: Handbook of physiology: section 4,
environmental physiology, Ill: The gravitational
environment, volume 1: microgravity. (Fregly MJ and
Blatteis CM, eds.) Oxford University Press, pp. 691-719.
Schneider VS, LeBlanc A, Rambaut PC. 1989. Bone and
mineral metabolism. In: Space physiology and medicine,
2nd
ed. (Nicogossian AE, Leach-Huntoon C, Pool SL,
eds.) Lea and Febiger, pp. 214-221.
Smith SM, JE Davis-Street, JV Fesperman, DS Calkins,
M Bawa, BR Macias, RS Meyer and AR Hargens.
Evaluation of treadmill exercise in a lower body negative
pressure chamber as a countermeasure for weightlessness-
induced bone loss: a bed rest study with identical twins.
Journal of Bone and Mineral Research 18: 2223-2230,
2003.
Smith SM, ME Wastney, BV Morukov, IM Larina, LE
Nyquist, SA Abrams, EN Tanan, C-Y Shih, JL Nillen, JE
Davis-Street, BL Rice, HW Lane. 1999. Calcium
metabolism before, during, and after a 3-month space
flight: kinetic and biochemical changes. American
Journal of Physiology 277(1 Pt 2): R 1-10.
Thornton WE, Rummel JA. 1977. Muscle deconditioning
and its prevention in space flight. In: Biomedical results
from Skylab. (Johnston RS, Dietlein LF, eds.) Washington
DC: NASA SP-377, 191-197.
Vernikos J, Ludwig DA, Ertl AC, Wade C, Keil L,
O’Hara D. 1996. Effect of standing or walking on
physiological changes induced by head down bed rest:
implications for spaceflight. Aviation, Space, and
Environmental Medicine 67:1069-1079.
Watenpaugh DE, Ballard RE, Fortney SM, Hargens AR.
1994. Larger waist seal area decreases the lower body
negative pressure required to produce a given level of foot
wand force. Aviation, Space, and Environmental
Medicine 65:A25.
Watenpaugh DE, RE Ballard, SM Schneider, SMC Lee,
AC ErtI, JM William, WL Boda, KJ Hutchinson and AR
Hargens. 2000. Supine lower body negative pressure
exercise during bed rest maintain upright exercise
capacity. Journal of Applied Physiology 89:218-227.
Watenpaugh DE, Hargens AR. 1996. The cardiovascular
system in microgravity. In: Handbook of physiology:
section 4, environmental physiology, Ill: The gravitational
environment, 1: microgravity. (Fregly MJ and Blatteis
CM, eds.) Oxford University Press, pp. 631-674.
Whalen R. 1993. Musculoskeletal adaptation to
mechanical forces on Earth and in space. Physiologist
36:S127-S130.
Whalen R T, Carter DR, Steele CR. 1988. Influence of
physical activity on the regulation of bone density.
Journal of Biomechanics 21:825-837.
Whedon GD, Lutwak L, Rambaut PC, Whittle MW,
Smith MC, Reid J, Leach C, Stadler CR, Sanford DD.
1977. Mineral and nitrogen metabolic studies, experiment
M071. In: Biomedical Results from Skylab. (Johnston RS,
Dietlein LF, eds.) Washington, DC: NASA SP-377, 164-
174.
Wolthuis RA, Bergman SA, Nicogossian AE. 1974.
Physiological effects of locally applied reduced pressure
in man. Physiological Review 54:566-595.

A. Hargens — Testing Exercise Countermeasures during 30 Days of Simulated Microgravity
64 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 65
TRANSLATIONAL MEDICINE: FROM GROUND-BASED STUDIES OF TRAUMATIC INJURIES TO
ASTRONAUT HEALTH AND EARTH BENEFITS.
Charles E. Wade
US Army Institute of Surgical Research, Fort Sam Houston TX 78234
ABSTRACT
Traumatic injuries are an everyday occurrence, from minor
scratches to the massive injuries of auto accidents. Based on this
reality NASA has prepared for a range of contingencies to care
for injured astronauts. These preparations have been based upon
the clinical incidence of traumatic injuries in environmental
analogs such as Arctic habitats. However, information is needed
as to the effect of acclimation to the space environment on
responses to injury, subsequent treatment and rehabilitation.
Deconditioning of a subject leads to significant alterations in
responses to hemorrhage, susceptibility to infection, and the
healing process. The treatment of the injured astronaut may have
to be altered to compensate for the effects of acclimation in light
of these findings. Furthermore, upon return to Earth's gravity
therapeutic interventions, such as administration of anesthetics,
may have to be adjusted. The response of patients to the
deconditioning of acute bedrest associated with the care of
traumatic injuries provides insight into care of injured
astronauts. In order to assure the mission success of long
duration space flights the care of even minor injuries should be
extensively evaluated.
Traumatic injuries are often the result of a single acute
event. However, the consequences of traumatic events can
be far reaching, extending from the patient to families,
communities, nations, and the world as a whole (Kauvar
& Wade 2005). Spaceflight programs have not been
immune to traumatic injuries. While most of these injuries
have been lethal, space agencies have focused extensive
resources on consideration of the traumatic injuries that
may occur during the course of space flights. As the
duration and planned scope of space flight missions is
expanded the probability of traumatic injuries is
increased. Of concern is the health and well being of the
injured astronaut as well as the impact on mission
success.
Epidemiology of Emergencies
Traumatic injuries take many forms, from the very simple
scratch to the massive crush injury with broken bones and
hemorrhage. The space agencies have focused on the
injuries serious enough to be classified as emergencies,
where the life of an individual or the success of a mission
is threatened (Safe Passage: Astronaut Care for
Exploration Missions 2005; Billica et al. 1996;
Kirkpatrick et al. 2005, 1997; McCuaig 1994; McCuaig
& Houtchens 1992; Summers et al. 2005). In the 44 years
of manned spaceflight 5 major accidents have resulted in
21 deaths (Safe Passage: Astronaut Care for Exploration
Missions 2005). Other accidents have resulted in
traumatic injuries, but most of these have occurred on
Earth where traditional emergency advanced life support
measures were applied (Summers et al. 2005). During
flight in the microgravity environment few medical
emergencies have been documented, with only two
reported incidents of traumatic injuries (Safe
Passage:Astronaut Care for Exploration Missions 2005;
Billica et al. 1996; Summers et al. 2005).
In the general population, the incidence of medical
emergencies is 0.06 events per person year (Safe Passage:
Astronaut Care for Exploration Missions 2005; Billica et
al. 1996; Summers et al. 2005). Using this model, a ten
member crew on a 2.4 year mission to Mars would have
at least one medical emergency (10 people x 2.4 years x
0.06 incidence per person-year = 1.4 emergent medical
events). It could be expected that the incidence in the
extreme environment of space would be higher than the
rate observed in the general population. However, studies
conducted in different injury settings have helped to
formulate the direction and policies of the NASA
emergency medical program. These environments include
the US Navy experience in nuclear submarines and the
Antarctic McMurdo Station (Thomas et al. 2003; Lugg
2000). Estimates of the occurrence of medical
emergencies during spaceflight are based on the data from
these environments. At McMurdo Station the incidence of
medical emergencies requiring medical evacuation is
reported as 0.036 events per person-year (Lugg 2000).
Review of the Russian program found four medical
evacuations over 72 person-years in space with an
incidence of 0.056, very similar to that of the general
population (Summers et al. 2005). Thus both the data
from spaceflight and simulation environments yield rates
of 0.04 to 0.05 per person-year for a medical emergency
requiring evacuation. In planning for long-duration
spaceflight the probability of an emergent event must be
considered and appropriate measures to deal with the
situation must be in place.
There have been 17 medical emergencies during
spaceflight in the NASA program, two of which were
traumatic injuries (Safe Passage:Astronaut Care for
Exploration Missions 2005; Summers et al. 2005). In the
NASA program the incidence of emergencies related to
trauma (12%) falls far below that observed in the
McMurdo Station (48%) (Lugg 2000; Safe Passage:
Astronaut Care for Exploration Missions 2005).
Irrespective of the present rate of injury, traumatic
injuries are of great concern to the NASA program as they
____________________
* Correspondence to: Dr. Charles Wade
US Army Institute of Surgical Research
3400 Rawley E. Chamber Dr.
Fort Sam Houston, TX 78234
Email: [email protected]
Phone: 210-916-3698; Fax: 210-916-2942

C.E.Wade — Translational Medicine
66 Gravitational and Space Biology 19(2) August 2006
are mentioned in 44% of the 45 risk areas in the
Bioastronautics Critical Path Road Map
(http://bioastroroadmap.nasa.gov/index.jsp). Further, five
questions under the heading “Major Illness and Trauma”
present the following:
18d) What resources are required for telemedical
consultation, diagnosis, and management of major
trauma?
18f) What are the resources and procedures needed to
perform basic and advanced management of trauma?
18I) What procedures and protocols are necessary for
rehabilitation after an acute medical illness or trauma?
18AH) What resources and procedures are needed for the
surgical management of major illness, injury, and trauma?
20E) What are the resources and procedures required for
the treatment of minor trauma, emphasizing autonomous
decision-making, based on known space flight illnesses,
injuries, and expedition analogs? How might they be
adapted to reduced-G operations?
NASA has focused its efforts on applying the standard
medical treatments used on Earth to spaceflight (Safe
Passage:Astronaut Care for Exploration Missions 2005;
Kirkpatrick et al. 2005, 1997; Summers et al. 2005). The
agency has contributed significantly to the medical field
by making equipment/devices simpler, smaller, and
autonomous. Furthermore, efforts have been made to take
into account the physical environment (microgravity,
closed-loop environmental systems, etc.) when caring for
the spaceflight patient. For example, alternatives to the
use of volatile anesthetics and the means of restraint of
the surgical patient have been evaluated (Campbell 2002,
1996, 2002, 2004, 2001). However, the physiological
changes of the patient due to acclimation to the
microgravity environment have not been adequately
considered. In formulating and reviewing NASA’s
policies regarding the care of the astronaut with an
emergency due to traumatic injuries, the altered
physiological status of the patient is recognized and the
need for additional information noted. Though the
problem is clearly defined it has not been adequately
addressed.
The study of the care of traumatic injuries is conducted in
animal models that provide evidence for the formulation
of guidelines for treatment of human patients. The
efficacy of the guidelines is then evaluated over the
course of patient care. An example of this process is the
study that led to the classification of astronauts as at risk
when undergoing anesthesia. On the Bion 11 mission
anesthetic complications occurred in two primates who
had been in spaceflight for 14 days (Ilyin et al. 2000).
This project was a multinational effort to study the effects
of spaceflight on bone, muscle and the vestibular system.
Anesthetic was administered to the animals by a
competent team of veterinarians on the first day post-
flight for minor surgery, muscle biopsies. One animal
died during the procedure and the other experienced
serious complications. Previously, surgeries had not been
performed until the seventh day of recovery following
Bion flights, and no complications were observed.
Furthermore, the procedure done multiple times on
control animals produced no adverse events. This
experience pointed out that the knowledge of risk
imposed by anesthesia following acclimation to the
spaceflight environment is limited (Safe
Passage:Astronaut Care for Exploration Missions 2005;
Norfleet 2000). This incident led to the reclassification of
astronauts by the American Society of Anesthesiologists
(ASA) 4: A patient with severe systemic disease that is a
constant threat to life. The change in classification means
increased vigilance when administering anesthetic to
patients and during recovery. Another example of the
value of animal studies in space was the series of surgical
procedures performed on rats during Neurolab mission
which confirmed the expectation that surgery can be
conducted in microgravity (Safe Passage: Astronaut Care
for Exploration Missions 2005; Campbell et al. 2005;
Kirkpatrick et al. 2005). These examples of the findings
derived from the study of animals subsequently impacted
patient care. While these are anecdotal observations there
is increased interest in establishing requirements for the
acute care of astronauts by performing classically
designed studies, as noted in a recent National Academy
of Science (Safe Passage: Astronaut Care for Exploration
Missions 2005) report: “Although it may not be realistic
to replicate these myriad physiological studies during the
postoperative period, carefully designed experiments with
simple outcome measures (supplemented by highly
selected physiological, histological, or metabolic studies)
performed in a microgravity environment such as the
International Space Station (ISS) after surgery in animals
might yield significant information of value.”
Presented below is an example of how animal
experimentation provides information to predict the
possible impact of the microgravity environment on the
care of astronauts with traumatic injuries. This example,
fluid resuscitation in response to hemorrhage, was
selected as it is part of the initial response to injury and
was called out in the National Academy Report (Safe
Passage: Astronaut Care for Exploration Missions 2005).
Fluid Resuscitation in Response to Hemorrhage On Earth the care of the patient with severe traumatic
injuries is compartmentalized into a series of care
responses. The first responder, in most developed
countries is a paramedic, who renders first aid assuring
that the airway is open, there is adequate ventilation, and
bleeding is controlled. Once this is accomplished,
resuscitation is initiated, often entailing the administration
of fluids. It is expected that in the course of a continued
human presence in space a number of situations will
require the use of resuscitation solutions. These include
traumatic injuries, blood loss during surgery and a
number of circumstances resulting in dehydration. At
present the requirement for resuscitation fluids has not
been documented in the space programs. However, one
could assume that the need would be similar to those
observed in other extreme environments such as
submarines or Antarctica.

C.E.Wade — Translational Medicine
Gravitational and Space Biology 19(2) August 2006 67
The primary use of resuscitation fluids in spaceflight
would be associated with emergency and surgical
procedures (Kirkpatrick et al. 2005, 1997). Emergency
surgery might have to be performed in a variety of
scenarios. While resuscitation fluids are mostly used as
volume support in the course of a conventional surgery,
the rate of blood loss could be greater during spaceflight
as blood clotting may be compromised, and a trained
surgeon may not perform the procedure. Thus, a greater
amount of blood loss would be expected.
Attempts to evaluate the performance of surgery in
microgravity generally concluded that operative
procedures may be conducted (Safe Passage: Astronaut
Care for Exploration Missions 2005; Campbell et al.
2005; Kirkpatrick et al. 2005). However, the limitations
of the procedures were not defined. Fluid would also be
required for the care of an astronaut requiring surgery
immediately upon return to Earth for an emergency
procedure. These two surgical environments may
necessitate different types of fluids or different procedural
approaches to fluid administration than those normally
used.
Traumatic injuries may also occur in space. For example
the ISS is a construction zone with astronauts often
moving large objects during extravehicular activity (EVA
or “space walk”). These activities are believed to be the
riskiest for traumatic injuries. Though the utmost safety
precautions are in place, the probability of injury is still
high. These injuries could be both blunt and penetrating
and could result in bleeding from an open wound or into a
body cavity. On Earth hemorrhage is the leading cause of
death due to traumatic injuries and accounts for 80% of
intraoperative trauma deaths (Champion 2004; Sauaia et
al. 1995). Fluids for the patient with traumatic injuries
would stabilize the individual prior to definitive care,
such as a surgical procedure, which would in turn
necessitate further fluids.
Resuscitation solutions would also be used for the
treatment of hypovolemia induced by dehydration.
Astronauts spend extended periods of time doing work in
space suits and in exercise with significant dehydration. It
is conceivable that extreme dehydration could require the
administration of fluids. Another likely cause of fluid loss
would be food poisoning or gastroenteritis resulting in
excessive diarrhea and vomiting, though oral rehydration
has been demonstrated as the ideal means of replacement
for gastrointestinal losses.
The goal of fluid administration for these conditions is
hemostasis (correcting physiological parameters to
normal levels). On Earth, accomplishing this goal is often
difficult, time consuming and requires extensive
resources. In spaceflight hemostasis for a victim of
hypovolemia by fluid resuscitation may be insufficient or
impossible to attain due to limited resources and
expertise. A comprehensive review of resuscitation fluids
for spaceflight by Kirkpatrick et al. (Kirkpatrick et al.
2005) reports that, “Present medical care in space has
been developed on the basis of proven conventional
terrestrial strategies of proven health care.” There has
been no attempt to systematically evaluate resuscitation
procedures or solutions to meet the unique requirements
of the altered physiology of an astronaut acclimated to
spaceflight.
Presently, normal saline (NS; 0.9% NaCl) is the primary
solution carried aboard the Shuttle and the ISS. The total
volume available is 4 L in the Advanced Life Support
Pack (ALSP) and 7.5 L in the Health Maintenance System
Ancillary Support Pack. The ALSP carries an additional
0.5 L of dextrose (D5W) (information provided by the
NASA Crew Office). Therefore, the total available fluid is
12 L. For the care of a major traumatic injury this would
equate to a blood volume expansion capability of only 4 L
(American College of Surgeons, C. O. T. Advanced
Trauma Life Support Student Manual (ATLS) 1997;
Fluid Resucitation: State of the Science for Treating
Combat Casualties and Civilian Injuries. 1999). In a
recent study of penetrating trauma we found the fluid
requirements to be 12.5 L over the first 24 hours (Wade et
al. 2003). Therefore, the volume and type of fluid
presently stored on space vehicles appears insufficient.
In the administration of normal saline to critically injured
patients it is recommended that the patient receive three
times the volume of blood loss (American College of
Surgeons, C. O. T. Advanced Trauma Life Support
Student Manual (ATLS) 1997; Fluid Resucitation: State
of the Science for Treating Combat Casualties and
Civilian Injuries.). However, the Advanced Trauma Life
Support recommends the limitation of fluid administration
to maintain systolic pressure to reduce uncontrolled
bleeding and the use of normal saline has been
questioned, as large amounts of fluid collect in the
extravascular space (American College of Surgeons, C. O.
T. Advanced Trauma Life Support Student Manual
(ATLS) 1997). This “third spacing” has been implicated
in subsequent morbidity, specifically, an increased
incidence of multi-organ dysfunction and acute
respiratory distress. Another risk factor in the use of large
volumes of normal saline is the hemodilution of red cell
mass that decreases oxygen carrying capacity and dilution
of clotting factors that would result in secondary re-
bleeding.
At present the various space programs recognize the need
for resuscitation but specific requirements are not clearly
defined. What is not understood is the impact of the space
environment on the response of the patient to
hypovolemia and the ability to adequately resuscitate the
individual. The spaceflight environment presents a
number of other challenges in the treatment of the patient
with hypovolemia. These include alterations in the
cardiovascular, endocrine, immune, and hemopoietic
systems as well as metabolic changes coupled with the
closed environment of the spacecraft. Below we present
possible modulators to the response of hypovolemia and
subsequent resuscitation. In addition we discuss the use of
a ground-based animal model in light of these changes.

C.E.Wade — Translational Medicine
68 Gravitational and Space Biology 19(2) August 2006
Based on the plethora of changes in spaceflight any
studies on the ground should employ a model emulating
as many of these alterations as possible. In human
subjects this has been accomplished by extended periods
of bed rest. While this model is appropriate for studying
countermeasures and provides some information as to
acute fluid shifts, the use of humans to evaluate
hemorrhagic hypovolemia due to injury is not warranted.
An animal model is the method of choice, especially if the
influence of long-term acclimation to the spaceflight
environment is to be evaluated.
The animal model we have focused on is the hind limb
suspended (HLS) rat (Morey-Holton & Globus 2002).
This model reproduces many of the characteristics
observed in humans and rodents in response to
spaceflight. In addition, if efficacy of resuscitation
solutions is to be evaluated in the microgravity
environment of spaceflight the rodent is the model
specimen of choice. Rodents have an extensive flight
history, and have been manipulated during spaceflight and
have undergone surgical procedures. Thus, rats provide
the basis for subsequent studies evaluating fluid
resuscitation procedures and techniques in actual
spaceflight.
Response to Spaceflight: human vs. hind limb
suspended (HLS) rat model
Spaceflight results in pronounced changes in the
physiology of astronauts and rodents. We will focus on
the changes observed in the acclimated individual and do
not discuss the acute responses before acclimation upon
insertion into orbit. These changes have been extensively
reviewed. We will use the reviews as a basis for the
discussion of the impact of these changes on the ability to
compensate for blood loss due to hemorrhage and the
subsequent ability to resuscitate the patient. We will then
address the response of the animal model and its viability
as a surrogate to study the responses in astronauts.
Cardiovascular changes Hemodynamics: The hemodynamic status of the astronaut
once acclimated to space flight is relatively normal
(Convertino 1996; Hargens & Watenpaugh 1996; Sawin
1998; Watenpaugh 2001). Blood pressure and heart rate
are within normal limits. Some reports note decreases in
resting heart rate, as well as a trend toward reduction in
diastolic pressure. These changes have been highly
dependent upon the flight mission and are within normal
clinical variability. Rodents exposed to HLS have an
initial period of acclimation similar to that of astronauts in
spaceflight (Overton et al. 1989; Ray et al. 2001).
Following this period mean arterial pressure (MAP), heart
rate (HR), systolic arterial blood pressure (SBP) and
diastolic blood pressure (DBP) seldom differ from those
of control animals.
Cardiac output following acclimation to spaceflight is
normal in astronauts (Hargens & Watenpaugh 1996). If
there is a reduced heart rate there is an accompanying
increase in stroke volume to sustain cardiac output. In the
rat HLS model there is no difference in cardiac output at
rest compared to control animals (Overton et al. 1989;
Woodman et al. 1995). As with the astronauts, reduced
responsiveness in cardiac output paralleled increasing
exercise loads. Failure to increase cardiac output in the
presence of increased metabolic demands as noted during
exercise would not bode well for correction of the loss of
oxygen-carrying capacity, and thus delivery, during
hemorrhage or the subsequent hemodilution and volume
expansion of resuscitation.
Blood Volume: Cardiovascular changes include a
reduction in blood volume on the order of 15% in
astronauts (Convertino 1996; Hargens & Watenpaugh
1996; Leach et al. 1996; Strollo 1999; Watenpaugh 2001).
The decrease in blood volume occurs by reductions in
both plasma volume and red cell mass, such that
hematocrit is not changed. This decrease in blood volume
could be detrimental upon return to Earth in the
hypovolemic patient. With re-exposure to Earth’s gravity
a shift in fluids has led to orthostatic hypotension
necessitating countermeasures. It would be logical that the
reduced blood volume would adversely impact a patient
brought back to Earth exacerbating the hypovolemic state.
In flight the reduced volume appears to be defended (see
Fluid Homeostasis below). The blood volume of HLS
animals appears to be reduced, but not to the same
magnitude as observed in astronauts. The reduction in
HLS animals is on the order of 6% and appears to be
dependent upon the duration of the HLS. The majority of
the literature reports the change in blood volume during
HLS not to be significant (Bouzeghrane et al. 1996; Chew
& Segal 1997; Tipton et al. 1998). A reduction in blood
volume prior to hemorrhage drastically reduces survival
and the success of resuscitation (Ho et al. 1996;Wade et
al. 1992).
Transcapillary fluid flux: Spaceflight appears to
redistribute fluid between body compartments in
astronauts (Convertino 1996; Hargens & Watenpaugh
1996; Leach et al. 1996; Watenpaugh 2001). There is the
decrease in the extracellur fluid volume comprised of a
contraction of blood volume previously mentioned and a
reduction in interstitial fluid volume. Simultaneously,
intracellular fluid volume increases on the order of 10%.
A major compensatory response to hemorrhage is the flux
of fluid from the interstitial space into the vascular
compartment (Hannon et al. 1990; Wade et al. 1992). In
time there is movement from the intracellular space. This
response is referred to as “auto-transfusion”. The lack of
adequate movement from the intracellular compartment
during hemorrhage would reduce survival. In the HLS
model this has yet to be addressed.
Fluid homeostasis
The reduction in blood volume of astronauts during
spaceflight suggests a new set-point for the maintenance
of fluid homeostasis. The regulation of fluid balance in
astronauts is altered with longer retention of a fluid load

C.E.Wade — Translational Medicine
Gravitational and Space Biology 19(2) August 2006 69
as a result of a reduced rate of urine excretion due to
alteration in the response of regulatory hormones
(Convertino 1996; Watenpaugh 2001). In the HLS rat
model though blood volume is slightly reduced, fluid
intake and urinary excretion are normal, suggesting a
shifted fluid homeostasis similar to that observed in
astronauts. This finding has led to the supposition that
there is attenuation of the responses to changes in blood
volume (Bouzeghrane et al. 1996; Deavers et al. 1980;
Steffen et al. 1984; Tucker & Mendonca 1995). Upon
return to Earth both astronauts and rats have a pronounced
diuresis (Wade & Morey-Holton 1998; Wade et al. 2000).
This increase in urine output is not immediately
compensated for by an increase in fluid intake and results
in hypovolemia. This response to landing, coupled with
hemorrhagic hypovolemia, would put the patient at risk
for a poor outcome. Thus, in the case of hypovolemia due
to hemorrhage the compensatory responses may be
attenuated in flight and after landing.
Endocrine System Pressor hormones: In response to hemorrhage there is a
progressive recruitment of pressor hormones. These
hormones, norepinephrine, epinephrine, angiotensin II
and vasopressin, increase peripheral resistance, leading to
maintenance of blood pressure and blood flow to essential
organs. In addition, the sympathetic hormones play a
major role in substrate mobilization. Reports as to
alteration of the sympathetic tone of astronauts are
variable, appearing to be highly mission dependent
(Leach et al. 1996, 1983; Macho et al. 1996; Stein &
Wade 2001; Tipton et al. 1996). Changes in sympathetic
tone and responsiveness are essential compensatory
responses to hemorrhage. With HLS animals, as with
astronauts, the reported plasma norepinephrine levels are
variable(Hasser & Moffitt 2001). Furthermore, changes in
sympathetic output in responses to manipulation are not
definitive, but appear to be reduced. The renin-
angiotensin system of astronauts is reduced (Leach et al.
1996, 1983). In addition, the response of this system as
determined by response to fluid load was attenuated. In
the HLS rodent plasma renin activity (PRA) decreased
after 7 days of suspension (Bouzeghrane et al. 1996).
However, no difference in PRA from control was noted
after 14 days of acclimation to HLS.
Astronauts experience reduced vasopressin (Leach et al.
1996,. 1983). In response to HLS the response of
vasopressin in animals is variable. An acute increase in
the rate of urinary excretion of vasopressin is found in
response to HLS, however, 14 days of suspension
produced no difference in plasma levels (Bouzeghrane et
al. 1996). Alterations in astronauts and HLS rats of most
of the pressor hormones occur during the period of
acclimation with control values attained after a period of
time. However, the alteration in responsiveness of the
hormones to volume manipulations suggests
compensatory changes due to hemorrhagic hypotension
would be altered, contributing to an inadequate
compensatory response.
Reproductive hormones: Spaceflight consistently reduces
testosterone levels in astronauts and rats (Ortiz et al.
1999; Strollo 1999; Tou et al. 2002). The cause of this
change is not well defined. There is no information as to
the change of reproductive hormones in female astronauts
(Ortiz et al. 1999; Strollo 1999; Tou et al. 2002). The
level of reproductive hormones has been demonstrated to
account for the pronounced gender differences in the
response to hemorrhage (Jarrar et al. 2000; Remmers et
al. 1998a; 1997, 1998b). A reduction in testosterone in
male rats is beneficial in the presence of hemorrhage. The
addition of estrogen contributes to an increase in survival.
With the HLS model male rats have a significant
reduction in plasma testosterone levels (Ortiz et al. 1999;
Tou et al. 2002). In female rats HLS reduces estrogen and
disrupts the reproductive cycle (Tou et al. 2005, 2004).
Therefore, changes in reproductive hormones during
spaceflight may be of benefit to male astronauts whose
injuries result in hemorrhage, and a decrement in female
astronauts.
Insulin/Glucagon: Investigations during spaceflight, in
astronauts and rats, have demonstrated changes in glucose
and insulin metabolism suggestive of a diabetic-like state
(Macho et al. 1996; Stein et al. 2000; Tobin et al. 2002).
There is an increase in circulating glucose and a reduction
in insulin concentrations for the associated plasma
glucose level (Macho et al. 1996). Furthermore, there
appears to be a decrease in insulin sensitivity. Plasma
glucagon levels are not changed. Hemorrhage decreases
sensitivity to insulin in liver and muscle tissues (Carter
1998; Custalow et al. 2001). This leads to a catabolic
state. Muscle atrophy and reduced insulin sensitivity as a
result of spaceflight would theoretically adversely affect
the normal compensatory response to hemorrhage. HLS
rats exhibit enhanced sensitivity to insulin-induced
glucose uptake when compared with normal control rats,
and resistance to the actions of insulin when contrasted
with rats similarly matched for the reduction in body mass
gain (Mondon et al. 1992). Insulin binding is also
reduced. A number of studies have produced conflicting
results in this area (Koebel et al. 1993; Stump et al. 1992,
1993), but the ability to mobilize and transport glucose in
response to hemorrhage is important to survival (Barton
& Passingham 1982; Carter 1998; Chang et al. 2000; Ma
et al. 2003). The diabetogenic state of space flight would
adversely affect this compensatory response and alter
outcome from hemorrhage.
Metabolic Changes
Negative energy balance: Astronauts experience a
negative energy balance that is highly mission dependent
and related to work load requirements (Stein 2000; Wade
et al. 2002). Over the course of a Shuttle mission body
mass is reduced (Sawin 1998; Stein 2000). A negative
energy balance adversely impacts survival following
hemorrhage (Nettelbladt et al. 1996). Bacterial
translocation from the gut may also increase after a
normally non-lethal hemorrhage if the patient is food
deprived (Bark et al. 1995). A consistent finding in the
HLS rat is a decrease in body mass on the order of 10%

C.E.Wade — Translational Medicine
70 Gravitational and Space Biology 19(2) August 2006
(Morey-Holton & Globus 2002; Thomason & Booth
1990). Attempts to determine energy balance have been
limited. We reported that food intake adjusted for body
mass was similar for HLS and control rats (Wang &
Wade 2000). Further, Blanc et al. (Blanc 2001, 2000)
reported no change in energy expenditure during 7 days
of suspension, and a net reduction in energy balance as
intake was reduced. We have reported no change in the
body mass of space flown rats (Wade et al. 2002). In this
case the HLS model may be more representative of the
astronaut than an animal on board the space craft.
Substrate metabolism: There are numerous reports of
changes in substrate metabolism of astronauts and rats
flown in space (Fitts et al. 2001; Stein 2000; Thomason,
1990). Many of the changes are associated with muscle
disuse (Fitts et al. 2001; Picquet et al. 2000; Thomason &
Booth 1990). There appears to be a preferential shift in
large normally weight-bearing muscle from fat
metabolism to glucose. In rats flown in space or HLS the
changes in fiber type are similar to those observed in
astronauts. This change in fiber type is associated with a
reduction in the ability of the muscle to oxidize long-
chain fatty acids, and an increased reliance on
carbohydrate metabolism (Stein et al. 2000; Thomason &
Booth 1990). In response to exercise these changes have
led to a reduction in exercise performance due to limited
metabolic reserves. Reliance on carbohydrates as a
metabolic substrate is high in the presence of major
hemorrhage (Boija et al. 1988; Custalow et al. 2001).
Thus, increased reliance on this metabolic pathway for
normal metabolism may limit the ability of this energy
pool to serve as a reserve during hemorrhage. The end
product of the use of carbohydrates in the presence of
limited oxygen is lactate. Lactate increase is indicative of
the magnitude of the oxygen debt incurred and is related
to subsequent outcome (Wade et al. 1989). In the HLS
model Fitts and colleagues (Fitts et al. 2001) have
performed studies showing there is an increase in muscle
glycogen, but in response to stimulation this reserve is
rapidly used. Further, at rest there was an increase in
tissue lactate that rapidly increased during stimulation. As
blood flow is shunted away from muscle during
hemorrhage the lack of shift in the metabolic substrate
would lead to rapid depletion in energy reserves and a
greater production of lactate and acidosis. These shifts in
metabolism would have a negative impact on survival
from hemorrhage and may preclude some commonly used
resuscitation solutions.
Changes in liver metabolism during spaceflight will also
influence responses to hemorrhage (Merrill et al. 1992;
Stein et al. 1994). Flight alters a variety of metabolic
pathways in liver tissue, reducing P-450 pathway
enzymes and cholesterol biosynthesis. There appears to be
a shift favoring gluconeogenesis as glycogen stores are
increased. The liver is one of the first organs to show
signs of injury during hemorrhage (Boija et al. 1988). The
pathway changes during spaceflight may impact the
ability of the liver to provide adequate substrate during
hemorrhage, leading to a reduction in ATP stores. A
reduction in liver ATP levels is associated with an
increase in apoptosis after hemorrhagic hypotension
(Mongan et al. 2002; Paxian et al. 2003). Alteration of
substrate metabolism during spaceflight may adversely
impact outcome following hemorrhagic hypotension due
to traumatic injuries. The metabolic imbalance may
persist during resuscitation as normal compensatory
pathways diminish. The capability to stabilize the patient
for an extended period of time following resuscitation is
in jeopardy as well.
Immune system:
The immune system of astronauts and rats is dramatically
altered during space flight (Sonnenfeld 2005b, 2005a,
2005c). The HLS model emulates many of the changes in
the immune system reported during spaceflight. HLS has
been used to assess possible countermeasures to immune
dysfunction associated with space flight. The response of
the immune system to hemorrhage plays a major role in
outcome of the patient and subsequent morbidity. The
dysfunction of this system could lead to a variety of
differences in the response to hemorrhage and
resuscitation.
Environmental:
The spaceflight environment is often thought of as only
involving responses to microgravity. However, there are
numerous other environmental factors that could
adversely affect the ability of a patient to survive
hemorrhage. A major environmental factor may be the
transition from the microgravity to the normal gravity of
Earth and the experience of hypergravity during landing.
An example of the influence of the return to Earth’s
gravity is the high incidence of astronauts experiencing
orthostatic intolerance (Hargens & Watenpaugh 1996;
Sawin 1998). Brizzee and Walker (Brizzee & Walker
1990) examined the response of reloading on blood
pressure, heart rate and cardiac output following hind
limb suspension for 7 days. They noted no difference in
any of the measured hemodynamic parameters. With 10
minutes reloading, mean arterial pressure did not change,
with trends towards an increase in heart rate and total
peripheral resistance and a fall in cardiac output. Others
have noted a significant reduction in blood pressure with
reloading (Bayorh et al. 2002). These responses are
similar to those observed in astronauts and do not bode
well for an individual injured in space who is returned to
Earth in a hypovolemic state.
Response of the HLS Model to Hypotension:
Studies of the HLS model response to hypotension are
limited and only one examined the response to
hemorrhage. Studies of hypotension have been directed at
the baroreflex to assess the impact of return to the gravity
of Earth on orthostatic intolerance, not the study of
responses to hemorrhagic hypovolemia. In studies of the
baroreflex the animals were administered a vasodilator at
increasing doses. The heart rate response to a given fall in
blood pressure provides an index of the responsiveness of
the system. This compensatory response is mediated by
the sympathetic nervous system. The response of heart

C.E.Wade — Translational Medicine
Gravitational and Space Biology 19(2) August 2006 71
rate to administration of the vasodilator was not affected
by HLS (Brizzee & Walker 1990; Hasser & Moffitt 2001;
Moffitt et al. 1998). However, the response of
sympathetic nerve activity decreased (Foley et al. 2005;
Hasser & Moffitt 2001; Moffitt et al. 1998; Mueller et al.
2005; Mueller & Hasser 2003). The authors concluded
that, as previously reported in astronauts (Hargens &
Watenpaugh 1996; Sawin 1998), HLS alters the arterial
baroreflex, possibly contributing to orthostatic intolerance
(Foley et al. 2005; Hasser & Moffitt 2001; Moffitt et al.
1998; Mueller et al. 2005; Mueller & Hasser 2003). The
inability to increase sympathetic tone in response to a
decrease in arterial pressure would also suggest a negative
impact of HLS on the response to hemorrhage, adversely
affecting outcome. The same authors have conducted
preliminary experiments of the response to hemorrhage
(Hasser & Moffitt 2001). The rats were exposed to HLS
for 12 days. Mean arterial pressure, heart rate and
sympathetic nerve activity were recorded. In response to
HLS mean arterial pressure did not change, but heart rate
increased. Rats were hemorrhaged at 3 ml/kg/min until
attaining a mean arterial pressure of less than 40 mmHg.
The amount of blood removed to reach the desired blood
pressure was similar (control 20.2 ± 2.1 ml/kg and HLS
19.0 ± 1.8 ml/kg). However, the increase in sympathetic
nerve activity in response to a reduction in pressure was
attenuated in HLS animals. This preliminary work on
hemorrhage, coupled with a body of literature that
addresses the modification of the cardiovascular
responsiveness of the HLS model, supports use of this
method. Further, as the HLS model produces many of the
metabolic, cardiovascular, immunologic and endocrine
changes observed in astronauts during spaceflight we
propose it as an appropriate means for the initial
evaluation of resuscitation procedures to be used to treat
hemorrhagic hypovolemia in astronauts.
Hemorrhage Models: As noted previously, the traumatic injuries that may
induce loss of blood volume necessitating fluid
replacement in an astronaut are quite diverse. Fortunately,
a number of experimental hemorrhage models can mimic
a range of clinical scenarios, such as fixed pressure, fixed
volume, uncontrolled and crush hemorrhage models. The
first model, fixed pressure, is one of the most common.
This method developed by Wiggers and colleagues bleeds
the animal to a fixed pressure and sustains that pressure
by removal of additional blood or replacement of the shed
blood volume (Wiggers 1950). This model contributed to
the hypothesis of irreversible hemorrhage in that at some
point, no matter what resuscitation interventions are used,
survival is not possible (Shah et al. 1998). This model
provides insights into cardiovascular compensation and
other endogenous factors mediating restitution. The
second is the fixed volume hemorrhage model with the set
removal of volume of blood per kilogram of body mass
(Hannon et al. 1990; Shah et al. 1998). This approach
allows differences between initial conditions to be
assessed. It also allows the condition of the animal to be
evaluated in terms of the net response to hypervolemia.
We have used this approach with a slight modification,
extending the period of bleeding at a slower rate to mimic
the drop in the drive pressure observed clinically. The
third model is of uncontrolled hemorrhage to simulate a
major injury in which the control of bleeding is not easily
attained (Bickell et al. 1991, 1989, 1994; Krausz et al.
2000). This simulates major traumatic injuries to the
torso. The model has been used extensively to assess
initial resuscitation in the pre-hospital setting. The fourth
model includes major tissue damage in the presence of a
fixed volume of bleeding (Jarrar et al. 2000; Remmers et
al. 1998al., 1997, 1998b). This model allows the study of
responses to tissue injury which modifies compensatory
adjustment. In evaluating approaches to resuscitation the
use of a number of the animal models of hemorrhage is
highly recommended (Fluid Resucitation:State of the
Science for Treating Combat Casualties and Civilian
Injuries. 1999). Although there are other approaches to
the study of hemorrhage however, the above methods are
widely used to assess the efficacy of resuscitation
procedures. We have deliberately not addressed burn or
septic shock. However, these forms of hypovolemic shock
would also be modified by acclimation to the spaceflight
environment and should be evaluated.
Resuscitation Parameters:
The approach to resuscitation following traumatic injuries
and the appropriate endpoints are controversial. Recent
studies using physiological endpoints have been effective
in increasing survival and reducing morbidity. However,
they require extensive physiological monitoring and
staffing. Thus, they have not been used outside the
hospital. This has led to the approach of stabilizing the
patient until definitive care can be administered, which
would be the optimal situation in the spaceflight
environment. These methods include delayed fluid
administration, hypotensive resuscitation, modifications
of the fluids administered and mechanisms for lowering
metabolism (Fluid Resucitation:State of the Science for
Treating Combat Casualties and Civilian Injuries. 1999;
Bickell et al. 1994; Dutton et al. 2002; Knudson et al.
2003; Kramer 2003).
Another area of controversy is the endpoint for evaluation
of resuscitation efficacy (Fluid Resucitation: State of the
Science for Treating Combat Casualties and Civilian
Injuries. 1999; Revell et al. 2003; Rhee et al. 1998;
Shoemaker et al. 1996; Wade & Holcomb 2005; Zhao et
al. 2002). The ultimate endpoint is survival. However,
these studies are expensive and involve large numbers of
subjects and thus are not appropriate for the initial
assessment of an intervention. Later in the development
of a process they are necessary. Many have felt that
restoration of blood volume and cardiovascular function
are adequate indices of resuscitation. This is not always
true. For example, the use of crystalloids may replace
blood volume and improve cardiac output in the short
term. However, due to hemodilution, oxygen delivery is
reduced, leading to death. Therefore, long-term survival
must be assessed. This is especially true in the spaceflight
environment where the extraction of the patient will be
days, if not weeks, and the acclimation process may

C.E.Wade — Translational Medicine
72 Gravitational and Space Biology 19(2) August 2006
necessitate alteration of the standard of care approaches to
resuscitation.
SUMMARY
The question is not if a traumatic injury requiring
resuscitation during spaceflight will occur, but what
systems and clinical guidelines will be in place to handle
it when it does occur, without loss of life or compromise
of the mission. The need for the care of traumatic injuries
has been recognized by the space agencies. Extensive
efforts and resources have been expended to meet these
contingencies by applying successful Earth-based clinical
practices. However, acclimation to the spaceflight
environment results in alterations in the cardiovascular,
endocrine, immune, and hemopoietic systems as well as
metabolic changes. These changes will drastically alter
the response of the patient to injury. These alterations
have to be considered in clinical guidelines to care for the
injured space traveler, and must be validated on the
ground and in flight in clinically relevant animal models.
REFERENCES
American College of Surgeons, C. O. T. Advanced
Trauma Life Support Student Manual (ATLS). 1997.
Bark T, Katouli M, Svenberg T, Ljungqvist O. 1995.
Food deprivation increases bacterial translocation after
non-lethal haemorrhage in rats. Eur J Surg 161. 67-71.
Barton RN, Passingham BJ. 1982. Early responses to
hemorrhage in the conscious rat: effects of corticosterone.
Am J Physiol 243: R416-23.
Bayorh MA, Eatman D, Walton M, Socci RR, Emmett N.
2002. Post-suspension hypotension is attenuated in
Sprague-Dawley rats by prostacyclin synthase inhibition.
Prostaglandins Leukot Essent Fatty Acids 66: 511-7.
Bickell WH, Bruttig SP, Millnamow GA, O'Benar J,
Wade CE. 1991. The detrimental effects of intravenous
crystalloid after aortotomy in swine. Surgery 110: 529-36.
Bickell WH, Bruttig SP, Wade CE. 1989. Hemodynamic
response to abdominal aortotomy in the anesthetized
swine. Circ Shock 28: 321-32.
Bickell WH, Wall MJ, Jr., Pepe PE, Martin RR, Ginger
VF, et al.. 1994. Immediate versus delayed fluid
resuscitation for hypotensive patients with penetrating
torso injuries. N Engl J Med 331: 1105-9.
Billica RD, Simmons SC, Mathes KL, McKinley BA,
Chuang CC, et al.. 1996. Perception of the medical risk
of spaceflight. Aviat Space Environ Med 67: 467-73.
Boija PO, Nylander G, Suhaili A, Ware J. 1988.
Posthemorrhage glycogen and lactate metabolism in the
liver: an experimental study with postprandial rats. J
Trauma 28: 779-83.
Bouzeghrane F, Fagette S, Somody L, Allevard AM,
Gharib C, Gauquelin G. 1996. Restraint vs. hindlimb
suspension on fluid and electrolyte balance in rats. J Appl
Physiol 80: 1993-2001.
Brizzee BL, Walker BR. 1990. Altered baroreflex
function after tail suspension in the conscious rat. J Appl
Physiol 69: 2091-6.
Campbell MR. 2002. A review of surgical care in space.
J Am Coll Surg 194: 802-12.
Campbell MR, Billica RD, Jennings R, Johnston S, 3rd.
1996. Laparoscopic surgery in weightlessness. Surg
Endosc 10: 111-7.
Campbell MR, Billica RD, Johnston SL, 3rd, Muller MS.
2002. Performance of advanced trauma life support
procedures in microgravity. Aviat Space Environ Med 73:
907-12.
Campbell MR, Johnston SL, 3rd, Marshburn T, Kane J,
Lugg D. 2004. Nonoperative treatment of suspected
appendicitis in remote medical care environments:
implications for future spaceflight medical care. J Am
Coll Surg 198: 822-30.
Campbell MR, Kirkpatrick AW, Billica RD, Johnston SL,
Jennings R, et al.. 2001. Endoscopic surgery in
weightlessness: the investigation of basic principles for
surgery in space. Surg Endosc 15: 1413-8.
Campbell MR, Williams DR, Buckey JC, Jr., Kirkpatrick
AW. 2005. Animal surgery during spaceflight on the
Neurolab Shuttle mission. Aviat Space Environ Med 76:
589-93.
Carter EA. 1998. Insulin resistance in burns and trauma.
Nutr Rev 56: S170-6.
Champion H. 2004. Epidemiological basis for future
improvements in trauma care. Semin Hematol 41: 173.
Chang CG, Van Way CW, 3rd, Dhar A, Helling T, Jr.,
Hahn Y. 2000. The use of insulin and glucose during
resuscitation from hemorrhagic shock increases hepatic
ATP. J Surg Res 92: 171-6.
Chew HG, Jr., Segal SS. 1997. Arterial morphology and
blood volumes of rats following 10-14 weeks of tail
suspension. Med Sci Sports Exerc 29: 1304-10.
Convertino VA. 1996. Clinical aspects of the control of
plasma volume at microgravity and during return to one
gravity. Med Sci Sports Exerc 28: S45-52.
Custalow CB, Watts JA, Thornton L, O'Malley P, Barbee
RW, et al.. 2001. Role of fatty acids in the recovery of
cardiac function during resuscitation from hemorrhagic
shock. Shock 15: 231-8.

C.E.Wade — Translational Medicine
Gravitational and Space Biology 19(2) August 2006 73
Deavers DR, Musacchia XJ, Meininger GA. 1980. Model
for antiorthostatic hypokinesia: head-down tilt effects on
water and salt excretion. J Appl Physiol 49: 576-82.
Dutton RP, Mackenzie CF, Scalea TM. 2002.
Hypotensive resuscitation during active hemorrhage:
impact on in-hospital mortality. J Trauma 52: 1141-6.
Fitts RH, Riley DR, Widrick JJ. 2001. Functional and
structural adaptations of skeletal muscle to microgravity.
J Exp Biol 204: 3201-8.
Fluid Resucitation:State of the Science for Treating
Combat Casualties and Civilian Injuries. 1999. ed.
Medicine: National Academy Press
Foley CM, Mueller PJ, Hasser EM, Heesch CM. 2005.
Hindlimb unloading and female gender attenuate
baroreflex-mediated sympathoexcitation. Am J Physiol
Regul Integr Comp Physiol 289: R1440-7.
Hannon JP, Wade CE, Bossone CA, Hunt MM, Coppes
RI, Loveday JA. 1990. Blood gas and acid-base status of
conscious pigs subjected to fixed-volume hemorrhage and
resuscitated with hypertonic saline dextran. Circ Shock
32: 19-29.
Hargens AR, Watenpaugh DE. 1996. Cardiovascular
adaptation to spaceflight. Med Sci Sports Exerc 28: 977-
82.
Hasser EM, Moffitt JA. 2001. Regulation of sympathetic
nervous system function after cardiovascular
deconditioning. Ann N Y Acad Sci 940: 454-68.
Ho HS, Sondeen JL, Dubick MA, Wade CE, Gunther RA.
1996. The renal effects of 7.5% NaCl-6% dextran-70
versus lactated Ringer's resuscitation of hemorrhage in
dehydrated sheep. Shock 5: 289-97.
Ilyin EA, Korolkov VI, Skidmore MG, Viso M,
Kozlovskaya IB, et al.. 2000. Bion 11 mission: primate
experiments. J Gravit Physiol 7: S9-17.
Jarrar D, Wang P, Cioffi WG, Bland KI, Chaudry IH.
2000. The female reproductive cycle is an important
variable in the response to trauma-hemorrhage. Am J
Physiol Heart Circ Physiol 279: H1015-21.
Kauvar DS, Wade CE. 2005. The epidemiology and
modern management of traumatic hemorrhage: US and
international perspectives. Crit Care 9 Suppl 5: S1-9.
Kirkpatrick AW, Campbell MR, Jones JA, Broderick TJ,
Ball CG, et al.. 2005. Extraterrestrial hemorrhage control:
terrestrial developments in technique, technology, and
philosophy with applicability to traumatic hemorrhage
control in long-duration spaceflight. J Am Coll Surg 200:
64-76.
Kirkpatrick AW, Campbell MR, Novinkov OL,
Goncharov IB, Kovachevich IV. 1997. Blunt trauma and
operative care in microgravity: a review of microgravity
physiology and surgical investigations with implications
for critical care and operative treatment in space. J Am
Coll Surg 184: 441-53.
Knudson MM, Lee S, Erickson V, Morabito D, Derugin
N, Manley GT. 2003. Tissue oxygen monitoring during
hemorrhagic shock and resuscitation: a comparison of
lactated Ringer's solution, hypertonic saline dextran, and
HBOC-201. J Trauma 54: 242-52.
Koebel DA, Fell RD, Steffen JM. 1993. Increased
glucose and 2-deoxy-D-glucose uptake in skeletal muscle
of suspended rats. Aviat Space Environ Med 64: 1016-22.
Kramer GC. 2003. Hypertonic resuscitation: physiologic
mechanisms and recommendations for trauma care. J
Trauma 54: S89-99.
Krausz MM, Bashenko Y, Hirsh M. 2000. Crystalloid or
colloid resuscitation of uncontrolled hemorrhagic shock
after moderate splenic injury. Shock 13: 230-5.
Leach CS, Alfrey CP, Suki WN, Leonard JI, Rambaut PC,
et al.. 1996. Regulation of body fluid compartments
during short-term spaceflight. J Appl Physiol 81: 105-16.
Leach CS, Altchuler SI, Cintron-Trevino NM. 1983. The
endocrine and metabolic responses to space flight. Med
Sci Sports Exerc 15: 432-40.
Lugg DJ. 2000. Antarctic medicine. Jama 283: 2082-4.
Ma Y,Wang P,Kuebler JF,Chaudry IH,Messina JL: 2003.
Hemorrhage induces the rapid development of hepatic
insulin resistance. 284: G107-15.
Macho L, Kvetnansky R, Nemeth S, Fickova M, Popova
I, et al.. 1996. Effects of space flight on endocrine system
function in experimental animals. Environ Med 40: 95-
111.
McCuaig K. 1994. Surgical problems in space: an
overview. J Clin Pharmacol 34: 513-7.
McCuaig KE, Houtchens BA. 1992. Management of
trauma and emergency surgery in space. J Trauma 33:
610-25, discussion 25-6.
Merrill AH, Jr., Wang E, Mullins RE, Grindeland RE,
Popova IA. 1992. Analyses of plasma for metabolic and
hormonal changes in rats flown aboard COSMOS 2044. J
Appl Physiol 73: 132S-5S.
Moffitt JA, Foley CM, Schadt JC, Laughlin MH, Hasser
EM. 1998. Attenuated baroreflex control of sympathetic
nerve activity after cardiovascular deconditioning in rats.
Am J Physiol 274: R1397-405.

C.E.Wade — Translational Medicine
74 Gravitational and Space Biology 19(2) August 2006
Mondon CE, Rodnick KJ, Dolkas CB, Azhar S, Reaven
GM. 1992. Alterations in glucose and protein metabolism
in animals subjected to simulated microgravity. Adv
Space Res 12: 169-77.
Mongan PD, Capacchione J, West S, Karaian J, Dubois
D, et al.. 2002. Pyruvate improves redox status and
decreases indicators of hepatic apoptosis during
hemorrhagic shock in swine. Am J Physiol Heart Circ
Physiol 283: H1634-44.
Morey-Holton ER, Globus RK. 2002. Hindlimb
unloading rodent model: technical aspects. J Appl Physiol
92: 1367-77.
Mueller PJ, Foley CM, Hasser EM. 2005. Hindlimb
unloading alters nitric oxide and autonomic control of
resting arterial pressure in conscious rats. Am J Physiol
Regul Integr Comp Physiol 289: R140-7.
Mueller PJ, Hasser EM. 2003. Enhanced
sympathoinhibitory response to volume expansion in
conscious hindlimb-unloaded rats. J Appl Physiol 94:
1806-12.
Nettelbladt CG, Alibergovic A, Ljungqvist O. 1996. Pre-
stress carbohydrate solution prevents fatal outcome after
hemorrhage in 24-hour food-deprived rats. Nutrition 12:
696-9.
Norfleet WT. 2000. Anesthetic concerns of spaceflight.
Anesthesiology 92: 1219-22.
Ortiz RM, Wang TJ, Wade CE. 1999. Influence of
centrifugation and hindlimb suspension on testosterone
and corticosterone excretion in rats. Aviat Space Environ
Med 70: 499-504.
Overton JM, Woodman CR, Tipton CM. 1989. Effect of
hindlimb suspension on VO2 max and regional blood
flow responses to exercise. J Appl Physiol 66: 653-9.
Paxian M, Bauer I, Rensing H, Jaeschke H, Mautes AE, et
al.. 2003. Recovery of hepatocellular ATP and
"pericentral apoptosis" after hemorrhage and
resuscitation. Faseb J 17: 993-1002.
Picquet F, Canu MH, Falempin M. 2000. Phenotypic
changes in the composition of muscular fibres in rat
soleus motor units after 14 days of hindlimb unloading.
Pflugers Arch 440: 229-35.
Ray CA, Vasques M, Miller TA, Wilkerson MK, Delp
MD. 2001. Effect of short-term microgravity and long-
term hindlimb unloading on rat cardiac mass and
function. J Appl Physiol 91: 1207-13.
Remmers DE, Cioffi WG, Bland KI, Wang P, Angele
MK, Chaudry IH. 1998a. Testosterone: the crucial
hormone responsible for depressing myocardial function
in males after trauma-hemorrhage. Ann Surg 227: 790-9.
Remmers DE, Wang P, Cioffi WG, Bland KI, Chaudry
IH. 1997. Testosterone receptor blockade after trauma-
hemorrhage improves cardiac and hepatic functions in
males. Am J Physiol 273: H2919-25.
Remmers DE, Wang P, Cioffi WG, Bland KI, Chaudry
IH. 1998b. Chronic resuscitation after trauma-
hemorrhage and acute fluid replacement improves
hepatocellular function and cardiac output. Ann Surg 227:
112-9.
Revell M, Greaves I, Porter K. 2003. Endpoints for fluid
resuscitation in hemorrhagic shock. J Trauma 54: S63-7.
Rhee P, Burris D, Kaufmann C, Pikoulis M, Austin B, et
al.. 1998. Lactated Ringer's solution resuscitation causes
neutrophil activation after hemorrhagic shock. J Trauma
44: 313-9.
Safe Passage:Astronaut Care for Exploration Missions.
2005. Presented at Committe on Creating a Vision for
Space Medicine During Travel Beyond Earth Orbit,
Board on Health Sciences Policy, Institute of Medicine,
Washington DC
Sauaia A, Moore FA, Moore EE, Moser KS, Brennan R,
et al.. 1995. Epidemiology of trauma deaths: a
reassessment. J Trauma 38: 185-93.
Sawin CF. 1998. Biomedical investigations conducted in
support of the Extended Duration Orbiter Medical Project.
Tex Med 94: 56-68.
Shah NS, Kelly E, Billiar TR, Marshall HM, Harbrecht
BG, et al.. 1998. Utility of clinical parameters of tissue
oxygenation in a quantitative model of irreversible
hemorrhagic shock. Shock 10: 343-6.
Shoemaker WC, Peitzman AB, Bellamy R, Bellomo R,
Bruttig SP, et al.. 1996. Resuscitation from severe
hemorrhage. Crit Care Med 24: S12-23.
Sonnenfeld G. 2005a. Experimentation with animal
models in space. Introduction. Adv Space Biol Med 10: 1-
5.
Sonnenfeld G. 2005b. The immune system in space,
including Earth-based benefits of space-based research.
Curr Pharm Biotechnol 6: 343-9.
Sonnenfeld G. 2005c. Use of animal models for space
flight physiology studies, with special focus on the
immune system. Gravit Space Biol Bull 18: 31-5.
Steffen JM, Robb R, Dombrowski MJ, Musacchia XJ,
Mandel AD, Sonnenfeld G. 1984. A suspension model
for hypokinetic/hypodynamic and antiorthostatic
responses in the mouse. Aviat Space Environ Med 55:
612-6.

C.E.Wade — Translational Medicine
Gravitational and Space Biology 19(2) August 2006 75
Stein T, Schluter MD, Galante P, Soteropoulos P, Tolias
P, et al.. 2000. Energy metabolism pathways in rat
muscle under conditions of simulated microgravity. J
Nutr Biochem 13: 471.
Stein TP. 2000. The relationship between dietary intake,
exercise, energy balance and the space craft environment.
Pflugers Arch 441: R21-31.
Stein TP, Schulter MD, Boden G. 1994. Development of
insulin resistance by astronauts during spaceflight. Aviat
Space Environ Med 65: 1091-6.
Stein TP, Wade CE. 2001. The catecholamine response
to spaceflight: role of diet and gender. Am J Physiol
Endocrinol Metab 281: E500-6.
Strollo F. 1999. Hormonal changes in humans during
spaceflight. Adv Space Biol Med 7: 99-129.
Stump CS, Balon TW, Tipton CM. 1992. Effects of
insulin and exercise on rat hindlimb muscles after
simulated microgravity. J Appl Physiol 73: 2044-53.
Stump CS, Woodman CR, Fregosi RF, Tipton CM. 1993.
Muscle glucose uptake in the rat after suspension with
single hindlimb weight bearing. J Appl Physiol 74: 2072-
8.
Summers RL, Johnston SL, Marshburn TH, Williams DR.
2005. Emergencies in space. Ann Emerg Med 46: 177-84.
Thomas TL, Garland FC, Mole D, Cohen BA, Gudewicz
TM, et al.. 2003. Health of U.S. Navy submarine crew
during periods of isolation. Aviat Space Environ Med 74:
260-5.
Thomason DB, Booth FW. 1990. Atrophy of the soleus
muscle by hindlimb unweighting. J Appl Physiol 68: 1-12.
Tipton CM, Greenleaf JE, Jackson CG. 1996.
Neuroendocrine and immune system responses with
spaceflights. Med Sci Sports Exerc 28: 988-98.
Tipton CM, Monnin KA, Hall MC, Sebastian LA. 1998.
The hematological responses of rats exposed to conditions
of simulated microgravity and acute exercise. Aviat Space
Environ Med 69: 267-76.
Tobin BW, Uchakin PN, Leeper-Woodford SK. 2002.
Insulin secretion and sensitivity in space flight:
diabetogenic effects. Nutrition 18: 842-8.
Tou J, Ronca A, Grindeland R, Wade C. 2002. Models to
study gravitational biology of Mammalian reproduction.
Biol Reprod 67: 1681-7.
Tou JC, Arnaud SB, Grindeland R, Wade C. 2005. The
effect of purified compared with nonpurified diet on bone
changes induced by hindlimb suspension of female rats.
Exp Biol Med (Maywood) 230: 31-9.
Tou JC, Grindeland RE, Wade CE. 2004. Effects of diet
and exposure to hindlimb suspension on estrous cycling in
Sprague-Dawley rats. Am J Physiol Endocrinol Metab
286: E425-33.
Tucker BJ, Mendonca MM. 1995. Effects of 30 day
simulated microgravity and recovery on fluid homeostasis
and renal function in the rat. J Gravit Physiol 2: P33-4.
Wade CE, Grady JJ, Kramer GC. 2003. Efficacy of
hypertonic saline dextran fluid resuscitation for patients
with hypotension from penetrating trauma. J Trauma 54:
S144-8.
Wade CE, Holcomb JB. 2005. Endpoints in clinical trials
of fluid resuscitation of patients with traumatic injuries.
Transfusion 45: 4S-8S.
Wade CE, Miller MM, Baer LA, Moran MM, Steele MK,
Stein TP. 2002. Body mass, energy intake, and water
consumption of rats and humans during space flight.
Nutrition 18: 829-36.
Wade CE, Morey-Holton E. 1998. Alteration of renal
function of rats following spaceflight. Am J Physiol 275:
R1058-65.
Wade CE, Ortiz RM, Baer LA. 2000. Increases in body
mass of rats during spaceflight: models and
measurements. Aviat Space Environ Med 71: 1126-30.
Wade CE, Tillman FJ, Loveday JA, Blackmon A,
Potanko E, et al.. 1992. Effect of dehydration on
cardiovascular responses and electrolytes after hypertonic
saline/dextran treatment for moderate hemorrhage. Ann
Emerg Med 21: 113-9.
Wade CE, Trail DS, Gildengorin VL, Hannon JP. 1989.
Blood lactate as a prognosticator of survival following
hemorrhage in conscious swine. Lab Anim Sci 39: 44-6.
Wang TJ, Wade CE. 2000. Effects of hindlimb
suspension and elevated ambient CO2 on rat growth and
renal function. Aviat Space Environ Med 71: 610-8.
Watenpaugh DE. 2001. Fluid volume control during
short-term space flight and implications for human
performance. J Exp Biol 204: 3209-15.
Wiggers CJ. 1950. The problem of functional coronary
collaterals. Exp Med Surg 8: 402-21.
Woodman CR, Sebastian LA, Tipton CM. 1995.
Influence of simulated microgravity on cardiac output and
blood flow distribution during exercise. Am J Physiol
Gastrointest Liver Physiol Am J Physiol Gastrointest
Liver Physiol Shock 18: 189-92.

C.E.Wade — Translational Medicine
76 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 77
Symposium III: Planetary Biology and
Terraforming
Paul Todd and David Thomas, Editors

78 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 79
PLANETARY BIOLOGY AND TERRAFORMING
Paul Todd
SHOT, Inc., 7200 Highway 150, Greenville, IN 47124 ABSTRACT
Planetary biology can be considered in terms of four
components: (1) planetary protection, (2) the search for life, (3)
human life support and (4) ecopoiesis and terraforming.
Initially, contamination of a planet with Earth life is to be
minimized in order to facilitate a search for planetary life.
Meanwhile, humans, if present, must be sustained.
Subsequently, large-scale modifications of a planetary
environment can be considered. “Ecopoiesis” is a term
introduced by McKay and Haynes to describe the initiation of a
living, self-sustaining ecosystem in a planetary (Mars)
environment. “Ecosynthesis” refers to the development of an
ecosystem that includes succession (ecosystem maturation by
the replacement of organisms). “Terraforming” refers to
creating an Earth-like world and includes planetary engineering.
A considerable amount of debate, discussion and publication has
been devoted to these subjects, but, at least in the case of
ecopoiesis, there has been very little, if any, experimental
research. The speakers in this session address, in order, issues
of planetary environments and habitability with reference to
Earth analogues, the role of extremophiles as pioneer organisms
in ecopoiesis, and the concept of succession as it applies to
terrestrial ecology and ecosynthesis.
KEYWORDS planetary biology, ecopoiesis, ecosynthesis, terraforming,
Mars, extremophiles
INTRODUCTION
Mars is considered the ultimate target for terraforming.
There are at least four levels of inquiry concerning biology
and Mars:
(1) Planetary protection, contamination and quarantine
issues (NRC, 1992; Rummel, 2001),
(2) The search for life on Mars (Banin and Mancinelli,
1995; Ivanov & Lein, 1995; Koike et al., 1995),
(3) Human expeditions to Mars and biological life support
(Meyer & McKay, 1984, 1989) and
(4) The terraforming of Mars starting with ecopoiesis
(McKay, 1982; Fogg, 1995a, b; Haynes, 1990; McKay and
Haynes, 1990; Haynes and McKay, 1992; McKay et al.,
1991; Hiscox, 1995) and continuing with ecosynthesis
(Graham, 2005),
The discussions that follow are concerned with the fourth
subject, terraforming.
A variety of claims have been made concerning the future
of ecopoiesis (starting an ecosystem) and terraforming
(creating an earth-like environment) – highly
controversial subjects. Planetary protection is a
significant component of the ecopoiesis debate. One of
the significant outcomes of performing research in this
area is expected to be the informing of a scientific
community consensus concerning these subjects.
Today’s thinkers hold varying opinions concerning
ecopoiesis, the process of evolving a physical and
biological environment that can lead to “terraforming” –
the deliberate introduction of terrestrial-type ecosystems
on remote celestial bodies such as planets, moons and
asteroids (Haynes, 1990). On one extreme, principles of
planetary protection prohibit the introduction of any
living organisms onto Earth’s neighbors in the solar
system while, on the other extreme advocates of
terraforming propose the early modification of Mars (for
example) to initiate processes that will make the planet a
more hospitable place for humans should they decide to
go there (Zubrin and Wagner, 1996). Either way,
knowledge is lacking, and experimental testing is non-
existent, related to the basic understanding of the
ecopoiesis process. Since the implementation of any
concepts related to ecopoiesis would occur several
decades in the future, the experimental study of
ecopoiesis now would seem an ideal objective. While
ethical considerations are outside the scope of this
presentation, such a research program could inform this
debate.
It is of great value to cause the visions of the prophets of
the field to be extended an entire century forward.
Meanwhile, a series of benign, but difficult experiments
(laboratory, field and theory) can and should be
performed. The undertaking of ecopoiesis/terraforming
research can be considered a direct response to the challenge
originally put forth by Christopher McKay and the late
Robert Haynes: “ . . if it is decided to implement such a
program of planetary engineering, a slow and conservative
approach is essential. Sufficient time must be allowed for a
wide range of studies of Mars as it exists at present, and for
careful planning, modeling and ‘pilot-plant’ trials (where
possible) of all successive steps in the enterprise.” (Haynes
& McKay, 1995). This challenge calls for research that
includes biological experiments and theoretical modeling
directed at the implementation of the enabling notions in
terraforming offered by Fogg (1995), McKay et al.(1991),
and Boston and Thompson (1991) among others. To date,
only individual components of ecopoietic systems have
been investigated – a few species of microorganisms and
certain primitive and vascular plants.
____________________
* Correspondence to: Paul Todd
SHOT, Inc.
7200 Highway 150
Greenville, IN 47124
Email: [email protected]
Phone: 812-923-9591x242; Fax: 812-923-9598

P. Todd — Planetary Biology and Terraforming
80 Gravitational and Space Biology 19(2) August 2006
THE MARTIAN ENVIRONMENT AND
PLANETARY ENGINEERING
A set of ecopoiesis research principles is offered, and they
are listed in the order in which they would logically be
implemented in an integrated ecopoiesis research program
(Boston et al., 2004):
1. Assume the ultimate problem to be addressed is
the terraforming of the surface of Mars.
Mars lies within the habitable zone of the solar system
(Fogg, 1998, McKay et al., 1991) and is second only to
Earth in biocompatibility. If the laws of physics and
chemistry are universal laws, and if the sequence of
events that led to the origin of life on Earth occurs
elsewhere, Mars may thus be considered a prime
candidate for both the existence of life and human
settlement (Meyer and McKay, 1995, 1984, 1989; Zubrin
and Wagner, 1996). The chemical-elemental inventory
of Mars' surface (both absolute and relative
concentrations, Table I) is quite similar to the Earth's crust
with the exception of a potentially low nitrogen content
(Banin, 1989; Banin et al., 1992; Banin and Mancinelli,
1995).
The primary physical properties of the Martian
environment that are a challenge for the survival and
Table I. Physical properties of Mars______________________________________
Pressure 6 – 15 millibars (0.6 – 1.5 kPa)
Global temperature range -133oC -- +23
oC
Mean Solar irradiance 589 W m-2
(~50% of Earth's value)
Solar UV flux 6500 ergs cm-2
(190 – 300 nm)
(<190 nm absorbed by CO2)
Water Vapor 0.03% of atmosphere mass (saturated at 8.1 mbar)
Gravitational constant 0.387g
Escape velocity 5km s-1
Day length 24h 37min 22s
Year length 687 Earth days
Equatorial diameter 6788 km (~1/2 Earth)
Mass 6.4 X 1023
kg (1/10 Earth)
Water ice North polar ice cap
Atmospheric composition
CO2 95% (<0.05% on Earth)
N2 2.7% (78 % on Earth)
O2 0.13% (21% on Earth)
CO 0.07%
Argon 1.6%
Neon ppm
Krypton ppm
Xenon ppm
Lithosphere elemental composition excluding oxygen (most elements are in oxides)
SULFUR 3.1%
MAGNESIUM 5%
CALCIUM 4%
IRON 12.7%
POTASSIUM 0.08%
CHLORINE 0.7%
SILICON 21%
ALUMINUM 3%
SODIUM 2.3%
PHOSPHOROUS 0.3%
Mn, Co, Cu, I, Zn, B, V, Mo, Trace
F, Cr, Se, Tl, Br
CARBON, NITROGEN Low levels (Abundant on Earth)

P. Todd — Planetary Biology and Terraforming
Gravitational and Space Biology 19(2) August 2006 81
growth of terrestrial organisms include; low atmospheric
pressure (~ 7 millibar), low temperature ranges (-100°C to
- 5°C, with a rare high of 26°C), no observed liquid water
to date (Soderblom, 1992), unfiltered ultra violet (UV)
radiation, no significant ozone layer, no organic material,
and high concentrations of surface oxidants (Kieffer et al.,
1973; Rothschild, 1990; Banin & Mancinelli, 1995). The
atmospheric pressure on Mars, mostly due to CO2, varies
from approximately 7.4 to 10 millibar (mbar). Extremely
low pressure can damage organisms and affect efficient
DNA repair (Ito, 1991; Koike et al., 1991).
2. Starting conditions to be studied on Earth should
resemble those expected to exist at the best possible
location on the surface of Mars (a temperate location at
low altitude or near potential water sources at the north
polar cap).
The physical and chemical properties of the planet are
summarized in Table I.
On Mars there are difficult trade-offs between water
availability and temperature. In general, water is
available only at or near polar caps because the
temperature is sufficiently low to prevent sublimation at
such low pressure. Liquid water may exist transiently in
the Martian lowlands at mid-day in the summer months
on occasions when the pressure (15- 20 mbar) and
temperature (2-20oC) are both above the triple point of
water – that unique combination of pressure and
temperature at which all three phases co-exist, 0.006oC
and 6.1 mbar (Kuznetz and Gan, 2002).
3. The regolith must be heated to a temperature
compatible with the most robust form of terrestrial life at
low temperature.
Shortly after the Viking missions, Averner and MacElroy
(1976) and Kuhn, Rogers and MacElroy (1979) launched
a program to study the habitability and planetary
ecosynthesis of Mars by designing an energy-balance
model. The initial studies evaluated the effects of a
Martian atmosphere on photosynthetic microorganisms by
approximating equatorial equinox parameters for solar
and thermal radiation, convective and conductive energy
transport and evaporative cooling. These early
investigations took into consideration the effect of diurnal
variation on the organisms’ temperature, transpiration and
photosynthetic rates, all of which are equally applicable to
current ecosystem models. Initial heating of the regolith
is to be achieved by the addition of black material (soot,
soil or microorganisms) to the near-polar ice to initiate
melting of water and CO2 to increase atmospheric
pressure and solar heat retention, and heating of the
atmosphere is achieved by the addition of fluorocarbon
gas into the planetary atmosphere, a very effective
greenhouse gas. Laboratory investigations of heating
strategies will require a combination of experiment and
modeling, since the laboratory scale differs significantly
from the planetary scale.
4. The first organisms must derive energy from the
mineral content of the regolith and/or sunlight, and their
metabolism must produce a net increase in greenhouse
gas(es).
Since the advent of MacElroy's program in 1975, several
living systems on Earth have been identified as potential
pioneer organisms for the Martian environment. These
biota may be classified as "extremophiles" and include
classes of anhydrobiotes (Dose et al., 1995), cryophiles,
thermophiles and halophiles, to name a few. Examples
include cryophiles found in the ice-covered lakes in the
Bunger Hills Oasis of Antarctica (Andersen et al., 1995;
McKay, 1991, 1993) and in Arctic permafrost in the
Kolyma lowland of Siberia, Russia (Ostroumov, 1995;
Soina et al., 1995). Thermo- and halophiles including
endolithic bacteria found in extreme arid deserts, high
Alpine rocks, and hot springs (Boston et al., 1992;
Friedmann, 1982, 1986; Friedmann & Weed, 1987),
evaporite biota found in the hypersaline habitats
(Rothschild, 1990; Rothschild et al., 1994) and cryptic
microbial mats in the Guerrero Negro, Baja California
Sur, Mexico.
An effective ecopoietic strategy requires knowing the limits
of habitability for Earth organisms as we understand them,
especially under conditions of limited water and oxygen
(Banin and Mancinelli, 1995; Carr, 1987; Fogg, 1995;
Hoffert et. al., 1981). To understand these limits, the
broadest possible range of genetically encoded capabilities
must first be considered, then, in a narrower way, conditions
that are and are not destructive to Earth's most extremophilic
organisms are to be identified (Hiscox, 1995; Hiscox and
Thomas, 1995). Using Mars regolith simulant (Allton et.
al., 1985) and planetary environment simulation chambers
(Sagan and Pollack, 1974), the vigor of such organisms can
be tested (Koike et. al., 1991; Kuhn et. al., 1979;
Hawrylewicz et. al., 1962; Thomas et al., 2006).
5. The first organisms must be capable of
withstanding, or be protected from, the ultraviolet and
ionizing radiation present on the Martian surface and the
high CO2 of the atmosphere.
The Martian atmosphere, devoid of an ozone layer, allows
solar ultraviolet (UV) radiation to penetrate to the Martian
surface in wavelengths ranging from 190 to 300 nm. In
living cells UV radiation is absorbed by nucleic acids to
activate the chemical formation of various adjuncts that
inhibit replication and transcription of DNA and can be
lethal (Lindberg & Horneck, 1991). The high
concentration of atmospheric carbon dioxide (~95.3%)
would, in most terrestrial organisms cause a low
intracellular pH, resulting in damage to cellular
components, proteins and metabolism.
6. Organisms early in succession should produce
significant amounts of O2.
At some point in the early stages of succession
(ecosystem maturation by the replacement of organisms),
O2 will be essential to drive the dark reactions of
photosynthetic organisms, which are O2 concentration
dependent and to support the metabolism of heterotrophic
aerobes (Graham, 2005).

P. Todd — Planetary Biology and Terraforming
82 Gravitational and Space Biology 19(2) August 2006
7. Higher plants late in succession will need to be
studied under conditions that pioneer organisms
preceding them can create.
Cold-weather species that do not require pollination or
can pollinate without insect assistance, that can function
in low O2 and high CO2 and extreme drought and that are
useful to humans are among those to be considered and
investigated (Graham, 2005).
THE PRESENTATIONS IN THIS SERIES
The subsequent presentations in this series address, in a
similar sequence, the importance of and evidence for
liquid water on Mars, the habitability of extraterrestrial
and extreme terrestrial venues for Earth life, the capability
of extremophiles to serve as pioneer organisms in
ecopoiesis and the succession of organisms in
ecosynthesis required to fulfill the ultimate goals of
terraforming.
ACKNOWLEDGMENTS
Assistance with literature review efforts by Penelope
Boston and Kelli McMillen are gratefully acknowledged.
Material for this presentation was developed under the
support of subcontract agreement 07605-003-026 with
NASA's Institute for Advanced Concepts (NIAC), a
program of the Universities Space Research Association
(USRA) funded by NASA contract NAS5-03110 and also
with the support of SHOT, Inc.
REFERENCES
Allton, J.H., Galindo, C. Jr. and Watts, L.A. 1985. Guide
to using lunar soil and simulants for experimentation. In,
Lunar Bases and Space Activities of the 21st Century, ed.
W.W. Mendell, Lunar and Planetary Institute, Houston,
TX. 497-506.
Andersen, D. T., Doran, P., Bolshiyanov, D., Rice, J.,
Galchenko, V., Cherych, N., Wharton, R. A., McKay, C.
P., Meyer, M. and Garshnek V. 1995. A preliminary
comparison of two perennially ice-covered lakes in
Antarctica: Analogs of past Martian lacustrine
environments. Advances in Space Research 15:199-202.
1995.
Averner, M, M. and MacElroy R. D. 1976. On the
habitability of Mars: An approach to planetary
ecosynthesis. NASA SP-414.
Banin, A. 1989. Mars Soil: A sterile regolith or a
medium for plant growth? In, The Case for Mars III:
Strategies for Exploration, ed. C.R. Stoker. American
Astronautical Society Vol. 74, AAS 87-215, 559-571.
Banin A., Clark B. C. and Wänke H. 1992. Surface
chemistry and mineralogy. In Mars (eds. Kieffer et al.),
pp. 594-625. Univ. Arizona Press, Tucson.
Banin, A. and Mancinelli, R.L. 1995. Life on Mars? I.
The chemical environment. Advances in Space Research
15:163-170.
Boston, P.J. and Thompson, S.L. 1991. Theoretical
microbial and vegetation control of planetary
environments. In, Scientists on Gaia. pp. 99-117. MIT
Press, Cambridge, MA.
Boston, P. J., Ivanov, M. V. and McKay, C. P. 1992. On
the possibility of chemosynthetic ecosystems in
subsurface habitats on Mars,” Icarus 95: 300-308.
Boston, P. J., Todd, P. and McMillen, K. R. 2004.
Robotic Lunar Ecopoiesis Test Bed: Bringing the
experimental method to terraforming. Space Technology
and Applications International Forum – STAIF 2004, Ed.
M. S. El-Genk, American Institute of Physics,
Washington, DC (2004).
Carr, M. H. 1987. Mars: a water-rich planet. In: MECA
Symposium On Mars: Evolution Of Its Climate and
Atmosphere, Baker V., Cart, M. H., Fanale-F., P.,
Greeley, R., Haeberli, R. M., Leovy, C., and Maxwell, T.
A. (eds.), Pp. 23-25.
Carr, M. H. 1987. Water on Mars. Nature 326: 30-35.
1987.
Dose, K., Stridde, C., R., Dillmann, R., Risi, S. and
Bieger-Dose A. 1995. Biochemical constraints for
survival under Martian conditions,” Advances in Space
Research 15: 203-210.
Fogg, M. J. 1995a. Terraforming: Engineering planetary
environments. SAE International, Warrendale, PA.
Fogg, M. J. 1995b. Terraforming Mars: Conceptual
solutions to the problem of plant growth in low
concentrations of oxygen. Journal of the British
Interplanetary Society 48, 427-434.
Fogg, M.J. 1998. Terraforming Mars: A review of
ongoing research. In: The Case for Mars VI: Enabling A
Human Presence on Mars, Ed. K.R. McMillen.
American. Astronautical Society, Univelt, Inc., San
Diego, CA.
Friedmann, E. I. 1982. Endolithic microorganisms in the
cold Antarctic desert. Science 215: 1045-1053.
Friedmann, E. I. 1986. The Antarctic cold desert and the
search for traces of life on Mars. Advances in Space
Research 6: 265-268.
Friedmann, E. I. and Weed, R. 1987. Microbial trace-
fossil formation, biogenous, and abiotic weathering in the
Antarctic cold desert. Science 236: 703-705.

P. Todd — Planetary Biology and Terraforming
Gravitational and Space Biology 19(2) August 2006 83
Graham, J. M. 2005. The biological terraforming of
Mars: Planetary ecosynthesis as ecological succession on
a global scale. Astrobiology 4: 168-195.
Hawrylewicz, C., Gowdy, B., and Erlich, R. 1962.
“Microorganisms under a simulated Martian
environment.” Nature 193:497.
Haynes, R. H. 1990. Ethics and planetary engineering. 1.
Ecce ecopoiesis: Playing God on Mars. In Macniven, D.
(Ed). Moral Expertise:161-183. Routledge, London and
New York.
Haynes, R. H., and McKay, C. P. 1992. The implantation
of life on Mars: Feasibility and motivation. Advances in
Space Research 12, 133-140.
Haynes, R. H., and McKay, C. P. 1995. Designing a
biosphere for Mars. In: Chemical Evolution: Self-
Organization of the Macromolecules of Life, Ed. J. Chela-
Fiores, M. Chadha, A. Negron-Mendoza, Tairo Oshima.
A. Deepak Publishing, Hampton, VA. Pp. 201-211.
Hiscox, J. A. 1995. Modification of microorganisms for
Mars. The Terraforming Report 2: 136-150.
Hiscox, J. A. and D. J. Thomas. 1995. Modification and
selection of microorganisms for growth on Mars. Journal
of the British Interplanetary Society 48: 419-426.
Hoffert , M.I., Callegari, A. J., Hsieh, C. T., Ziegler, W.
1981. Liquid water on Mars: An energy balance climate
model for CO2/H2O atmospheres. Icarus. 47:112-149.
Ito, T. 1991. The effects of vacuum-UV radiation (50-190
nm) on microorganisms and DNA. Advances in Space
Research 12: 249-253.
Ivanov, M.V. and Lein, A.Yu. 1995. Biogeochemical
Evidence of Microbial Activity on Mars. COSPAR,
Advances in Space Research 15, (3)215-(3)221.
Kieffer , H.H., Chase, S. C. Jr., Miner, E., Munch, G. and
Neugebauer, G. 1973. Preliminary Report on Infrared
Radiometric Measurements from the Mariner 9
Spacecraft. Journal of Geophysical Research 78:4291.
Koike, J. T., Oshima, K.A., Koike, H., Taguchi, R.,
Tanaka, K., Nishimura, M., and Myaji, M. 1991. Survival
rates of some terrestrial microorganisms under simulated
space conditions.” Advances in Space Research 12: 271-
274 (1991).
Koike, J., Oshima, T., Kobayashi, K. and Kawasaki, Y.
1995. Studies in the Search for Life on Mars. Advances
in Space Research 15, (3)211-(3)214.
Kuhn, W.R., Rogers, S.R., and MacElroy, R.D. 1979.
“The response of selected terrestrial organisms to the
Martian environment.” Icarus 37:336-346.
Kuznetz, L. and Gan, D. 2002. On the existence and
stability of liquid water on the surface of Mars today.
Astrobiology 2 (2): 183-195.
Lindberg, C. and Horneck, G. 1991. Action spectra for
survival and spore photoproduct formation in Bacillus
subtilis irradiated with short wavelength (200-300 nm)
UV at atmospheric pressure and in vivo. Journal of
Photochemistry and Photobiology 11: 69-80.
McKay, C. P. 1982. Terraforming Mars. Journal of the
British Interplanetary Society 35: 427-433.
McKay, C. P. 1993. Relevance of Antarctic microbial
ecosystems to exobiology. Antarctic Microbiology: 593-
601. Wiley-Liss Inc., New York.
McKay, C. P. and Haynes, R. H. 1990. Should we implant
life on Mars? Scientific American, December, 144.
McKay, C. P., Toon, O. B. and Kasting, J. F. 1991.
Making Mars habitable. Nature 352: 489-496.
Meyer, T. R. and McKay, C. P. 1984. The atmosphere of
Mars-resources for the exploration and settlement of
Mars. The Case for Mars. (Ed. Boston, P. J.). American
Astronautical Society Science and Technology Series 57:
209-232.
Meyer, T. R. and C. P. McKay, C. P. 1989. The resources
of Mars for human settlement. Journal of the British
Interplanetary Society 42: 147-160.
Meyer, T.R. and McKay, C. P. 1995. Using the
resources of Mars for human settlement. In, Strategies
for Mars: A Guide to Human Exploration, C.R. Stoker,
Ed. 86, 393-442.
NRC --National Research Council. 1992. Biological
Contamination of Mars. Issues and Recommendations.
National Academy Press, Washington.
Ostroumov, V. 1995. A physical and chemical
characterization of Martian permafrost as a possible
habitat for viable microoganisms. Advances in Space
Research 15(3):229-236 (1995).
Rothschild, L. J. 1990. Earth analogs for Martian life.
Microbes in evaporites, a new model system for life on
Mars. Icarus 88: 246.
Rothschild, L. J., Mancinelli, R., White, M. R. and Giver,
L. P. 1994. Metabolic activity of microorganisms in
gypsum-halite crusts. Journal of Phycology 30: 431-438.
Rummel, J. D. 2001. Planetary exploration in the time of
astrobiology: Protecting against biological contamination.
Proceedings of the National Academy of Sceinces 98 (5):
2128-2131.

P. Todd — Planetary Biology and Terraforming
84 Gravitational and Space Biology 19(2) August 2006
Sagan, C. and Pollack, J. B. 1974. Differential
transmission of sunlight on Mars: Bilogical implications.
Icarus 21, 490-495.
Soderblom, L.. 1992 The composition and mineralogy of
the Martian surface from spectroscopic observations: 0.3
µm to 50 µm. In, Mars, Ed. H.H. Kieffer, B.M. Jakosky,
C. W. Snyder, M.S. Matthews. Univ. Arizona Press,
Tucson, AZ.557-595.
Soina, V.S., Vorobiova, E.A., Zvyagintsev, D. G. and
Gilichinksy, D. A.. 1995. Preservation of Cell Structures
in Permafrost: A Model for Exobiology. Advances in
Space Research 15(3):237-242.
Thomas, D. J., Boling, J., Boston, P. J., Campbell, K. A.,
McSpadden, T., McWilliams, L. and Todd, P. 2006.
Extremophiles for ecopoiesis: Desirable traits for and
survivability of pioneer Martian organisms. Gravitational
and Space Biology 19: this issue.
Zubrin, R. and Wagner, R. 1996. The Case for Mars. The
Plan to Settle the Red Planet and Why We Must. Simon
and Schuster, New York.

Gravitational and Space Biology 19(2) August 2006 85
LAST PLACE TO BOIL AWAY, FIRST PLACE TO LOOK: THE HUNT FOR WATER
AND LIFE ON MARS
Lawrence H. Kuznetz
Space Spinoffs, Inc., Universities Space Research Associates (USRA) NASA Johnson Space Center, Houston,
Texas 77058
ABSTRACT
The primary goals of the robotic exploration of Mars are to
characterize the geology and climate of the planet; determine
whether or not life ever arose and establish the planet’s
suitability for human exploration. Inseparable in this quest is the
hunt for water and ancient seas. To date, the landing sites
associated with the quest have been driven in equal measures by
geology and conservatism (ie, a safe place in which to land).
This study uses principles of physics and thermodynamics to
target alternative landing sites that might maximize the
probability of finding water, past and present and extant life as
well. The phase diagram of water together with predictions of
temperature and pressure at specific locations is used with
elevation data from MOLA (Mars Orbiter Laser Altimeter) to
predict locations where water in liquid form last existed on the
surface and may, with accompanying extremophiles, exist today.
Calculations at Candor Chasma and Hellas are presented as
examples of how the principles of physics may be applied to
choose landing sites of the future that would optimize the goals
of robotic exploration.
KEYWORDS Mars, liquid water, landing sites, water phase diagram,
atmospheric pressure, Candor Chasma, Hellas, Mars probes
ABBREVIATIONS
MGS: Mars Global Surveyor, MOLA: Mars Orbiting
Laser Altimeter, RS: Radio Science Instrument on MGS,
VL1: Viking Lander 1, VL2 Viking Lander 2
INTRODUCTION
Follow the water: long the mantra of NASA’s robotic
program to explore Mars. Follow the water to discover
life, extinct or extant, and the other things. To date,
geology, and the safety net of relatively flat landing sites
have played the major role in looking for water, ancient
or otherwise. But thermodynamics has a role to play, too.
Hence the title, Last Place to Boil Away, First Place to
Look. Consider the following: If you were a liquid water
droplet millions or billions of years ago, and realized the
atmosphere above you was disappearing, and oceans were
boiling away, where would you rather be, in a high place
or low place, in a warm place or cold place? Physics
dictates low and warm, since that’s what it takes to stay
liquid. The lower the altitude, the higher the pressure,
and only the combination of higher pressures and warmer
temperatures will keep you and any life forms that you
support from freezing, boiling or sublimating away.
Granted, such places are rare on Mars today but they were
plentiful in the past. And they may exist today. Finding
them is the subject of this presentation.
TARGETS OF OPPORTUNITY, PAST AND
PRESENT
Logically, if you were an alien from Earth landing on
Mars, such as Viking, Pathfinder, Spirit or Opportunity,
and your task was to “follow the water,” you would likely
take the low, warm road to find it because that’s where
liquid water and any life forms within would have
survived the longest. This road has not been taken for
two reasons. First, low warm places are not as easy land
in; and second, we have assumed there is no liquid water
left on the surface today anyway and decided to look for
signs of past water only. Past or present, physics can help.
It leads us, in fact, to an entirely different set of landing
sites from the ones we have visited thus far. To
understand why, we must start with the water phase
diagram.
THE PHASE DIAGRAM OF WATER AND
CONDITIONS ON MARS
1. There is no liquid water on the surface of Mars
today; the pressure is so low that ice sublimates to water
vapor without ever passing through a liquid phase. This
statement, popularized by Norman Horowitz, the Cal
Tech Nobel laureate, reflected the thinking in the years
immediately prior and subsequent to the Viking missions.
The recent article in Nature, Evidence from HRSC Mars
Express for a Frozen Sea (Murray, et. al., 2005}, is a case
in point. The authors surmised a frozen sea at southern
Elyssium (+5o south, 150
o east) close to the Martian
equator. While adding to the growing body of knowledge
about recent geologic activities that lend credence to
active hydrothermal processes (with tantalizing hints of
microbial life), it repeats an oft-stated misconception that
detracts from its central theme. Specifically, the
statement, "Ice is unstable at the surface of Mars due to
sublimation in the 6-mbar atmosphere," is misleading.
While a frozen sea certainly could exist at temperatures
below freezing, ice is definitely not unstable. A close look
at the water phase diagram of Figure 1 reveals why. The
triangular region marked off shows the window of
opportunity in which water can exist as a liquid under
Martian datum level (mean surface altitude) conditions
today. At a 10mbar surface pressure, the liquid state exists
between 0 and 7oC. This window will be narrower at
lower pressures and wider at higher pressures but as long
as the pressure remains between 6.1 and 10 mbar, it exists
____________________
* Correspondence to: Lawrence H. Kuznetz
Space Spinoffs, Inc.
c/o NASA Johnson Space Center
Houston, TX 77058
Email: [email protected]
Phone: 510-205-8056

L. Kuznetz — The Hunt for Water and Life on Mars
86 Gravitational and Space Biology 19(2) August 2006
Figure 1. A selected portion of the water phase diagram (Weast, 1973-74). The phase diagram illustrates when pure water exists as a
solid, liquid, or vapor as a function of pressure and temperature. Triangle at the left end of the liquid region indicates conditions for liquid
water at Mars datum level.
nonetheless. Surface pressure data from the Viking and
Pathfinder landers have always been in this window, with
a 6.7 to 10 mbar pressure range. The above authors’ claim
of instability and sublimation is feasible only under two
conditions:
1. At higher elevations (hundreds or thousands of
kilometers above datum level) where the total pressure
will always be below the triple point of 6.1 mbar or
2. If the partial pressure of water vapor cannot build to
the total pressure within the boundary layer above the ice
source.
Neither of these conditions likely exists at Elyssium, the
site of the above authors’ proposed frozen lake. Firstly,
Elyssium is located approximately 2 km below datum
level, where total pressure should always be above the
triple point and secondly, unless strong winds exist to
disrupt the boundary layer, the partial water vapor
pressure should always build to levels approaching total
pressure. In other words, the surface of this frozen lake
should pass through a liquid phase as temperatures climb
above freezing. If this happens, the fundamental question
then becomes the fate of this liquid phase. Will it boil,
evaporate, or pool after it melts, and for how long?
This question is crucial because it speaks to nothing less
than the stability of liquid water and its link to life. If
liquid water can still form and pool on Mars then the
presence of such standing water increases the possibility
of extant life. If it is the evaporation (boiling) rate, not the
sublimation rate that determines the fate of the standing
water, one must ask the next question: Assuming frost,
snow or ice is available at or near the surface (an entirely
different question not addressed here), then under what
conditions will it form into a liquid and remain that way
for more than a few seconds?
EVAPORATION RATE AND THE LIKELIHOOD
OF STANDING WATER ON MARS TODAY
Studies by Kuznetz and Gan (2000, 2002); Quinn and
McKay (2000); Hecht (2002) and most recently by Moore
and Sears (2005) exposed ice to simulated Mars datum
level conditions and demonstrated conclusively that it will
pass through a liquid phase as long as the temperature
rises above freezing and remains below the boiling point.
Typical results are shown in Figure 2, which shows water
drops falling from an ice cube above 0oC in a Mars
vacuum jar (desiccator). Kuznetz and Gan (2002) also
photographed the melting sequence of frost on a
simulated Martian rock as it first forms a ring, then
coalesces to stable water droplets after it melts. In these
studies, water was not only observed to form after the ice
or frost melted, but once formed, lingered for substantial
time periods due to slow evaporation rates. How slow is
this evaporation rate? Kuznetz and Gan (2002) found
evaporation rates averaging 0.023 g/cm2-h in a Mars
simulation vacuum jar controlled to temperatures between
-2 to +20oC, and pressures of 6.3-14 mbar with and
without advection. Quinn and McKay (2000), Ingersol
(1970) and Hecht (2002) found similar scales of
evaporation, and Moore and Sears (2005) reported rates
around 1 mm/hr. These rates are slow enough to permit
standing water to remain liquid as long as it does on
Earth, assuming ice is present and a heat source above the
freezing point is there to melt it. Climate modeling studies
by Haberle (2000) supported these lab findings, predicting
in excess of 135 days per year (668 Earth days) during

L. Kuznetz — The Hunt for Water and Life on Mars
Gravitational and Space Biology 19(2) August 2006 87
which conditions along Mars’ equatorial region could
support liquid water (innermost contour areas of Figure 3,
surrounded by lines labeled “135”). These are not
"unstable," conditions as the Nature authors maintain, but
"metastable," as Hecht (2002) pointed out. (A glass of
water on a table top on Earth is similarly metastable).
Figure 2. Liquid water droplet formation in a vacuum
desiccator jar under simulated Martian conditions. The vacuum
desiccator is evacuated to 10 mbar and cooled with solid CO2,
which also provides Mars atmosphere stimulant. (Kuznetz and
Gan, 2002).
Examining the liquid-phase area in the phase diagram of
Figure 1 provides further insight. If standing water drops
can linger between 0 and 7oC at 10 mbar, they should
linger longer at the higher pressures that occur at lower
elevations on Mars. The window of opportunity at 15
mbar, for example, is 7 times greater than at 6.3 mbar (0
to 14.9oC, vs 0 to 2
oC on the phase diagram before
boiling occurs) and 12 times greater at 25 mbar. By
inference, the odds of standing water in the northern
lowlands should be substantially greater than in the
southern highlands, an intuitively obvious finding since
water flows toward the lowest level. This is predicated by
the temperature cooperating, i.e., staying between the
freezing and boiling points for extended periods of time at
the lower elevations.
THE INFLUENCE OF ELEVATION
Returning to our central thesis, if the window of
opportunity for liquid water increases at higher pressures,
where then are the locations associated with these
pressures? These would be the places where water would
have boiled away last, leaving its most recent signature,
as well as fossil traces if they exist. These would also be
the places where standing water and extant life, if it exists
at all, may reside. Insight to such locations is provided by
a combination of data from Viking Lander 1 ; MGS’
MOLA- Mars Global Surveyor’s Mars Orbital Laser
Altimeter, which shows the depth of Candor Chasm in
Figure 4; and by the MGS’ Radio Science instrument,
which shows the depth-pressure profile over Hellas in
Figure 5.
Figure 3. Predicted durations (given in Mars sols per year on
each contour line) suitable for liquid water brine on Mars. From
the Mars Global Climate model (Haberle, 2002).
Three approaches can be used to employ these data sets in
the search for optimum locations. In the first, Viking
Lander 1 (VL1) pressure and elevation data at Chryse
Planitia are referenced against elevations at other
locations derived from the Mars Orbital Laser Altimeter
(MOLA). The difference can be used to extrapolate the
pressure at these other elevations. In the second approach,
pressures measured by the MGS radio occultation
instrument at these same surface locations can be
compared to the pressures deduced by Approach 1. In the
third, the logarithmic relationship between altitude and
pressure, as shown in Figure 5, can be extrapolated to
lower depths below datum level and compared to the
other approaches. Examples of how this might be done for
Candor Chasma and Hellas follow.
CANDOR CHASM, -5 KM
Figure 4. The topography of Mars from MOLA, with a closeup
of Candor Chasm, showing elevation on a graded scale.
(http://ltpwww.gsfc.nasa.gov/tharsis/Mars_topography_from_M
OLA/M_-80_-70_-10_0.html)
In Approach 1, the elevation of Viking Lander 1 (VL1) is
known from MOLA to be -3683 meters, while the peak
pressure recorded by VL1 was 9.0 mbar (near Ls=270,

L. Kuznetz — The Hunt for Water and Life on Mars
88 Gravitational and Space Biology 19(2) August 2006
northern winter solstice). Extrapolations of pressure at
lower elevations were made assuming an atmospheric
scale height of 13 km (corresponding to a temperature of
250o K). The pressure calculated this way at Candor
Chasma, located at –5000 meters (about 1.3 km below
VL1), is roughly 10 mbar (~9 x exp(1.3/13) = 9 x 1.11).
The pressure at the low point in Hellas, on the other hand,
between –7500 and –8000 meters, could be as high as 14
mbar (using a low point for Hellas of 4.5 km below VL1
yields ~9 x exp(4.5/13) = 9 x 1.414, or 12.7 mbar.
-10
0
10
20
30
40
50
1 10 100 1000 10000
PRESSURE, PASCALS
KM
AB
OV
E S
UR
FA
CE
Figure 5. Pressure as a function of altitude at Hellas from MGS
Radio Occultation. Pressure extrapolated to lowest altitude of
about -8 km is about 1400 Pascals, equal to 14 mbar.
(http://nova.stanford.edu/projects/mgs/late.html)
Approach 2 uses independent pressure measurements
from MGS radio occultation data. Radio occultation
employs microwave radiation transmitted through the
Martian atmosphere by MGS prior to entry and just after
exiting the backside orbit where loss of signal (LOS)
occurs (hence the name occultation or hidden). Analyzing
the phase shift of these waves provides pressure for
specific longitudes, latitudes and time of day. Data for
Hellas at a depth of –6.86 km below datum was recorded
at 11.4 mbar, for example (see Table 1 and Figure 5).
Approach 3 simply uses the trend line of Figure 5 to
predict pressures at lower elevations by extrapolating the
trend line, in this case, 8 km below datum at Hellas as
shown in the Figure.
Pressures deduced from the above analyses predict ~10
mbar 5 km below datum level at Candor Chasma and ~13
to 14 mbar at the low point of Hellas (-8 km). It should be
noted that pressures in Approach 1 have a 10% inherent
error due to uncertainties in scale height and atmospheric
mass distribution versus latitude and longitude in a given
season. Errors in Approach 2 associated with MGS Radio
Occultation near the surface are ~ 1% for large areas and
increase at abrupt topographical features such as Candor
due to vertical resolution of the MGS oscillator. Errors in
Approach 3 result from assumptions of constant density
and curve extrapolation.
Table I. Martian Weather Observations at Hellas
(http://nova.stanford.edu/projects/mgs/late.html)
Date of Measurement: 04-01-2004
Time of Measurement: 09:23 GMT
Local Time on Mars: 03:41
Latitude: 39.9 degrees S
Longitude: 66.9 degrees E
Elevation: -6860 meters
Surface Temperature: -77.4 Celsius (-107.3
Fahrenheit)
Surface Pressure: 11.40 millibars
Martian Season: Early Fall
DISCUSSION AND SUMMARY
Mars is a fascinating planet in many respects, one of them
thermodynamically. A glass of water there could
sublimate, freeze, boil or remain liquid for hours, perhaps
days on end. Depending on location, all four processes
could occur at the same place over the course of a single
day. MGS Radio Science Weather Reports have observed
a preponderance of such locations from orbit, typically at
higher elevations above datum level. The Landers, on the
other hand, (VL1, VL2 and Pathfinder) have only seen
pressures above the triple point, probably owing to the
lower elevations at their landing sites.
This study has focused on using the water phase diagram
and data from surface probes and orbiters to identify
landing sites that could maximize the probability of liquid
water, past and present. Sites such as Candor Chasma and
Hellas have been shown to be locations where pressures
of 10-14 mbar could be routinely expected. The heat trap
effect at depressions such as these will also drive up the
temperature. The Inner Gorge of the Grand Canyon, for
example (elevation 670 meters), is substantially warmer
than the 2300-m high South Rim. In mid-April the
daytime temperature in the Inner Gorge exceeds 85oF and
cools only slightly at night, while night-time temperature
on the South Rim fall below the freezing point of water.
By inference, this suggests daily temperature excursions
at Candor and Hellas, taken together with higher
pressures should translate to increased residency time in
the liquid water envelope between freezing and boiling.
The appeal of Hellas and Candor increases with
hypotheses such as Hynek’s (2004), that vast ancient seas
may have covered Hellas in the distant past, and Malin
and Edgett’s (2000a,b) statement that MGS MOC
observations strongly indicate that the deepest parts of
Vallis Marineris were not brecciated but layered. If true,
the bottom of these locales would have been the last
refuge for large amounts of water on Mars; any life that

L. Kuznetz — The Hunt for Water and Life on Mars
Gravitational and Space Biology 19(2) August 2006 89
may have existed at the time, and their fossilized remains
that may exist today. All of this begs the question asked
by physics: Might not these and other places like them be
the most ideal landing sites to look for liquid water and
life on Mars today? If so, identifying and targeting these
areas should be a priority. Equipping future rovers with a
suite of instruments that can “sniff” liquid water should
also be considered. After landing, such a rover might
“predict-correct” over a wide area like a “water witch”,
fine tuning the location of surface or subsurface water
with a suite of instruments such as an alpha X-ray proton
spectrometer, temperature, pressure and water vapor
sensors.
The human exploration program would also benefit from
a high probability water site. Other than the obvious need
for a reliable, low cost water source, the design of a blue
collar space suit may well hinge on higher pressure,
warmer temperature locales. Current suits function by
sublimation, a process that will not work at datum level or
below. They are also far too heavy. A key to cutting mass
and complexity is to design a suit to its climate of intent,
as protective clothing does on Earth. Studies indicate that
a suit designed for Candor or Hellas might be more like
an Arctic coverall than the thermos bottle-like designs
that exist today (Kuznetz, 1989). Pressures above the
triple point and warmer temperatures may not only be the
key to finding environments that support liquid water and
life, but maximize human exploration potential as well.
ACKNOWLEDGEMENTS
The author is grateful to Drs. David Hinson and David E.
Smith of the MGS Radio Science Team for suggesting
approaches to use the MGS Radio Occultation and VL1
data to correlate longitude, latitude and elevation with
atmospheric pressure, and for providing error estimates in
MGS MOLA and RS data.
REFERENCES
Haberle, R.. 2000. On the stability of liquid water on
present day Mars. Proceedings of the First NASA Ames
Astrobiology Conference, NASA Ames Res Center,
Moffett Field, CA.
Hecht, H. 2002. Metastability of liquid water on Mars.
Icarus 156: 373-376.
Holman, J.P. 1990. Heat Transfer, 7th
ed. McGraw Hill,
San Francisco.
Hynek, B. 2004. Opportunities Great Lakes. University of
Colorado report, Astrobiology Magazine.
Ingersoll A.P. 1970. Mars: Occurrence of liquid water.
Science 168: 972-973.
Kuznetz, L. 1991. Space suits and life support systems for
the exploration of Mars. National Science Foundation
Fellowship, NASA Ames Research Center, Moffett Field,
CA.
Kuznetz, L.H. and Gan D. 1999. The hunt for liquid water
on Mars today. Third Mars Society Conference, Mars
Society, Boulder, Colorado.
Kuznetz, L. H. and Gan, D. 2002. On the existence and
stability of liquid water on the surface of Mars today.
Astrobiology 2 (2): 183-195.
Malin, M. and Edgett, K., 2000a. Evidence for recent
groundwater seepage and surface runoff on Mars. Science
288: 2330.
Malin, M. and Edgett, K., 2000b. Sedimentary rocks on
early Mars. Science 290: 5498. 1927
McKay, C. and Stoker, C. 1989. The early environment
and its evolution on Mars: Implications of Mars. Reviews
for Geophysics 27 (2): 189-214.
Moore, S. and Sears, D. 2005. Stability of contemporary
liquid water on Mars. Journal of Geophysical Research
Planets, in review.
Murray, J., Muller, J., Neukum, G. Werner, S., Gassest, S.
2005. Evidence from HRSC Mars Express for a frozen
sea. Nature 434: 352-356.
Quinn, R. and McKay, C. 2000. Mars Analog
Experiment, Space Science Division, NASA Ames
Research Center, Moffett Field, CA.
Weast, R. (Ed.) 1973-74. Water Phase Diagram. Chemical
Rubber Handbook, 54th
ed., pp DD 158-160, Cleveland.

L. Kuznetz — The Hunt for Water and Life on Mars
90 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 91
EXTREMOPHILES FOR ECOPOIESIS: DESIRABLE TRAITS FOR AND SURVIVABILITY OF
PIONEER MARTIAN ORGANISMS
David J. Thomas1, John Boling
1, Penelope J. Boston
2, 3, Kathy A. Campbell
1, 4, Tiffany McSpadden
1, Laura
McWilliams1, and Paul Todd
5
1Lyon College, Science Division, 2300 Highland Road, Batesville, AR 72501
2Complex Systems Research, Inc., P.O. Box 11320, Boulder, CO 80301.
3Department of Earth and Environmental Science, MSEC 208, New Mexico Institute of Mining and Technology,
801 Leroy Place, Socorro, New Mexico 87801. 4Newark High School, 1500 N. Hill Street, Newark, AR 72562.
5SHOT, Inc., 7200 Highway 150, Greenville, IN 47124.
ABSTRACT
Humanity is on the verge of having the capability of
constructively directing environmental changes on a planetary
scale. Within the foreseeable future, we will have the
technology to modify Mars' environment, and make it a
habitable planet. However, we do not have enough information
to determine the course of such an event. To our knowledge, no
known terrestrial organism has the capability of living on Mars'
surface under present conditions. However, with some
modification, Mars' environment could be brought into the
survival and growth range of currently known microorganisms.
Using the SHOT Ecopoesis Testbed, we performed
survival/growth experiments to determine the suitability of
potential pioneering life forms for Mars. Included among the
potential pioneers were five genera of cyanobacteria (Anabaena,
Chroococcidiopsis, Plectonema, Synechococcus and
Synechocystis), three partially-characterized Atacama Desert
heterotrophic eubacterial strains, and several desert varnish
isolates. Microorganisms were exposed to a present-day mix of
martian atmospheric gases, but at a pressure of 100 mbar (10
times Mars' current atmospheric pressure). Cultures were
inoculated into samples of JSC Mars-1 soil stimulant and
exposed to full-spectrum simulated martian sunlight. Day/night
temperature cycled from 26°C to -80°C and back. Preliminary
results indicate that both autotrophic and heterotrophic bacteria
can survive in the simulated engineered martian environment.
INTRODUCTION
In relation to terrestrial organisms, Mars' environment is
harsher than any on Earth. The combination of low
temperature, low atmospheric pressure, low moisture,
high atmospheric CO2 fraction and high UV flux is not
experienced by any known Earth organism. However,
Earth organisms may be found in environments that
include at least one of the martian environmental factors
(although Mars is still harsher in most respects). The dry
valleys of Antarctica provide a low temperature, low
moisture, high UV environment. Chile's Atacama Desert
is arguably the driest place on Earth. Some hot springs
and volcanic vents produce anoxic environments with
very high CO2 levels. Alpine and stratospheric
environments possess low pressure, low temperature and
high UV.
Each of these terrestrial environments harbors some form
of life. Organisms living there have adapted to the
extreme conditions, and those adaptations are genetically
encoded. In theory, genes encoding favorable
characteristics from one organism could be transferred to
another organism, which already possesses some other
characteristics for extreme environment survival. With
enough genetic alterations, we could conceivably "build"
a microorganism—a "marsbug"—that could survive,
reproduce and grow in a somewhat modified martian
environment.
Ten years ago, Hiscox and Thomas (Hiscox and Thomas,
1995) detailed some of the characteristics that an ideal
marsbug might possess. Here, we elaborate upon and
update these characteristics, and summarize some initial
experiments in which organisms with some of these
characteristics were tested under approximated martian
conditions.
DESIRABLE TRAITS FOR PIONEER
MICROORGANISMS
For the purposes of this paper, we assume that initial,
non-biological, modification of Mars' environment would
occur before pioneer microorganisms are introduced. At
minimum, Mars would have an atmospheric pressure of at
least 25 mbar, significant periods of above-freezing
temperatures, and significant bodies of liquid water
(Thomas, 1995). At 25 mbar, water boils at 9°C,
providing a narrow temperature window at which water
remains liquid. Also, recent studies suggest that Earth
bacteria do not grow at pressures below 25 mbar
(Schuerger et al., 2006a; Schuerger and Nicholson, 2005).
Even with initial modification, the martian environment
would be at least as harsh as the least hospitable
environments on Earth. Accordingly, pioneer
microorganisms would require many, if not all, of the
following characteristics (summarized in Table I).
Autotrophy. Mars has no known reserves of organic
materials. Pioneer organisms would need to make their
____________________
* Correspondence to: David J. Thomas
Lyon College, Science Division
2300 Highland Road
Batesville, AR 72501
Email: [email protected]
Phone: 870-698-4269

D.J. Thomas — Extremophiles for Ecopoiesis
92 Gravitational and Space Biology 19(2) August 2006
Table I. Desirable characteristics for pioneer martian microorganisms. The lists of microorganisms and references are examples
only, and are not meant to be exhaustive.
Characteristic Example Microorganisms References
Autotrophy: oxygenic
photosynthesis
Cyanobacteria, algae (Blankenship and Hartman, 1998; Burger-Wiersma
and Matthijs, 1990; Dismukes et al., 2001; Fay, 1983;
Goodwin, 1980; Ho and Krogman, 1982; Kaplan et
al., 1988; Tabita, 1994; Xiong and Bauer, 2002)
Autotrophy: chemosynthesis Nitrosamonas, Nitrobacter,
Thiobacillus, methanogens
(Atlas, 1997; Boston et al., 1992; Chyba and Hand,
2001; Northup et al., 2003; Stevens and McKinley,
1995)
Psychrophily Pyramimonas, Fragilariopsis,
Pseudo-nitzschia, Porosira,
Entomoneis, Nitzschia.
(Chen and Berns, 1978; Christner et al., 2003;
Daugbjerg, 2000; Gaidos et al., 2004; Gilichinsky et
al., 2003; McMinn et al., 2005; Morita, 1975)
CO2 tolerance Anabaena, Plectonema, Cyanidium,
Nannochloris
(Negoro et al., 1991; Seckbach et al., 1970; Seckbach
and Libby, 1970; Thomas et al., 2005)
Hypoxia tolerance Anabaena, Plectonema, Cyanidium (Seckbach et al., 1970; Thomas et al., 2005; Zehnder
and Svensson, 1986)
Carbonate dissolution Matteia, other cyanobacteria and
algae
(Crispim et al., 2003; Friedmann et al., 1993)
Denitrification Pseudomonas, Paracoccus,
Streptomyces
(Baker et al., 1998; Hart et al., 2000; Kumon et al.,
2002; Zumft, 1997)
Nitrogen fixation Anabaena, Chroococcidiopsis,
Cyanothece, Synechococcus,
Trichodesmium
(Almon and Böger, 1988; Berman-Frank et al., 2001;
Böhme, 1998; Fay, 1983; Friedmann and Kibler,
1980; Golden and Yoon, 1998; Klingler et al., 1989;
Mitsui and Cao, 1988; Olson et al., 1998; Potts et al.,
1983; Schneegurt et al., 2000; Wolk, 1988)
Osmotic tolerance,
desiccation resisistance
Chroococcidiopsis, Halomonas,
Klebsormidium, Oscillatoria,
Phormidium, endospore-forming
bacteria, extreme halophiles
(Abyzov et al., 1967; Billi et al., 2001; Billi et al.,
2000; Cockell et al., 2005; de Winder et al., 1989;
Frankenberg-Schwager et al., 1975; Imshenetsky and
Lysenko, 1965; Kraegeloh and Kunte, 2002; Olson et
al., 1998)
UV/ionizing radiation
resistance
Aspergillus, Bacillus,
Chroococcidiopsis, Deinococcus,
Mycobacterium, Rubrobacter
(Battista et al., 1999; Bauche and Laval, 1999; Billi et
al., 2000; Cockell et al., 2005; Ferreira et al., 1999;
Imshenetsky et al., 1967; Imshenetsky et al., 1977)
Hypobaric tolerance Aspergillus, Mycobacterium,
Pseudomonas, Staphylococcus,
Streptococcus
(Frankenberg-Schwager et al., 1975; Hawrylewicz et
al., 1967; Imshenetsky et al., 1977; Imshenetsky et al.,
1970; Silverman and Beecher, 1967)
Switchable genes and
pathways
Cyanobacteria, facultative
autotrophs, genetically modified
agricultural plants
(Garlick et al., 1977; Guay and Silver, 1975; Sarhan
and Danyluk, 1998; Sorokin et al., 2000)
own biomolecules from inorganic constituents via
photosynthesis or chemosynthesis. Photosynthesis would
replace the CO2 in Mars' atmosphere with O2—a key
process in planetary engineering (Graham, 2004; McKay,
1998; McKay et al., 1991; Thomas, 1995). Iron and
manganese-oxidizing chemolithotrophs produce dark-
colored byproducts (Dorn and Oberlander, 1981), which
decrease the albedo of rock surfaces, and would aid in the
heating of Mars (Boston et al., 2004).
Antioxidants. Oxygenic photosynthesis produces a
variety of reactive oxygen species (ROS) as normal
byproducts of electron transfer. In addition, Mars’
surface is currently a highly oxidizing environment,
presumably due to UV-produced ROS (Hunten, 1979;
Plumb et al., 1989; Yen et al., 2000). Currently, we do
not know whether initial engineering efforts would
change the oxidizing nature of Mars' surface. However,
other environmental stresses, such as chilling, increase the
amount of ROS produced by photosynthesis (Clare et al.,
1984; Hodgson and Raison, 1991a; Hodgson and Raison,
1991b; Thomas et al., 1999; Wise, 1995; Wise and
Naylor, 1987). Pioneer microorganisms would require
robust antioxidant systems in order to detoxify both
internally- and externally-generated ROS.
Psychrophily. Even under engineered conditions, Mars
will be cold. Truly psychrophilic organisms have
optimum growth temperatures below 20°C (Atlas, 1997).
The ability to grow at low temperature requires an entire
suite of enzymes that operate well at low temperature as
well as highly unsaturated membrane lipids
(Chattopadhyay and Jagannadham, 2001; Gerday et al.,
2000; Morita, 1975; Russell, 2000; Zecchinon et al.,
2001). Thus, psychrophilic autotrophs may be good
starting organisms to which other characteristics could be
added.
CO2 tolerance. Mars' atmosphere currently contains 95%
CO2, albeit at low pressure. We expect initial engineering

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 93
efforts to increase the total atmospheric pressure primarily
through the release of additional CO2 from the polar caps
and from possible carbonate rocks, which would further
increase the proportion of CO2. At high CO2 levels,
decreased pH becomes a factor as well. When CO2
dissolves in water, it forms carbonic acid with a pH of 5.5
- 6.0 at 1000 mbar. Fortunately, several cyanobacteria
and at least one alga survive and grow in >20% CO2 at
1000 mbar, and many microorganisms grow within the
pH range of 5 - 6 (Negoro et al., 1991; Seckbach et al.,
1970; Seckbach and Libby, 1970; Thomas et al., 2005).
Hypoxia tolerance. Mars' atmosphere contains very little
O2. Pioneer microorganisms will have to grow during
extended periods of hypoxia or anoxia. Even though
photosynthesis produces O2, plants require at least 50
mbar pO2 for proper development (due in part to the large
proportion of non-photosynthetic tissues) (Alpi and
Beevers, 1983; Atwell et al., 1982; Barclay and Crawford,
1982; Jackson and Drew, 1984). Most chemoautotrophs
also require O2 in order to oxidize inorganic molecules for
energy production (Atlas, 1997), thus limiting their initial
growth on Mars. Fortunately, most of the CO2-tolerant
photoautotrophic microbes, cited previously, also grow
anaerobically.
Carbonate dissolution. As mentioned previously,
carbonates are a potential source of atmospheric CO2.
Many cyanobacteria and algae cause damage to
limestone-based structures due to carbonate dissolution—
the conversion of carbonate ions into CO2 (Crispim et al.,
2003). If Mars has extensive carbonate deposits, such
organisms could speed the process of thickening Mars'
CO2 atmosphere while also providing oxygen. The
Antarctic cyanobacterium, Matteia, has been specifically
suggested for this purpose (Friedmann et al., 1993).
Denitrification. Although Mars' atmosphere could be
thickened by the addition of CO2, photosynthesis will
remove the CO2 and replace it with O2. An atmosphere of
pure O2 would result in catastrophic combustion. On
Earth, the atmosphere is buffered with N2, which is
largely nonreactive. If Mars possesses significant nitrate
deposits, these can be converted to N2 via the process of
denitrification (Thomas, 1995). Denitrifying bacteria use
nitrate in place of O2 for respiration, and thus are usually
hypoxia tolerant as well. Pseudomonas aeruginosa, a
heterotrophic facultative anaerobe, grows well and
denitrifies under conditions similar to those expected
during ecopoesis (Hart et al., 2000). Additionally, some
chemoautotrophs utilize nitrate in place of oxygen
(Oremland et al., 2002), thus avoiding the problem of
hypoxia and chemosynthesis mentioned previously.
Nitrogen fixation. Later in the process of planetary
engineering, biologically-usable nitrogen will need to be
recycled from the atmosphere. Nitrogen fixation
(Mancinelli, 1996)—the opposite of denitrification—
would be detrimental during the initial stages, but would
be required later in order to stabilize biological
communities (Thomas, 1995). Azotobacter vinelandii and
Azomonas agilis have the capability of fixing nitrogen at
pN2 as low as 5 mbar (Klingler et al., 1989). Many
genera of cyanobacteria also fix nitrogen (Böhme, 1998;
Fay, 1983; Friedmann and Kibler, 1980; Golden and
Yoon, 1998; Mitsui and Cao, 1988; Mulholland and
Capone, 2000; Olson et al., 1998; Potts et al., 1983;
Schneegurt et al., 2000; Zehr et al., 2001).
Osmotic tolerance. The initial bodies of water that form
on Mars probably will contain significant amounts of
dissolved salts; they may also be transient. Pioneer
microorganisms will need to tolerate osmotic stress, and
survive periods of desiccation. A large number of Gram-
positive bacteria form endospores that allow long periods
of dormancy under adverse conditions. Many non-spore-
forming bacteria also survive periods of desiccation. The
period between dormancy and the resumption of
metabolism should be as short as possible for pioneering
marsbugs so that they can take full advantage of periods
of favorable conditions. Some cyanobacteria undergo
seasonal desiccation and quickly become active during
wet periods (de Winder et al., 1989; Hershovitz et al.,
1991a; Hershovitz et al., 1991b; Scherer et al., 1984).
Ultraviolet and ionizing radiation resistance. Mars has
very little free O2 and only a very thin ozone layer that
changes with season and latitude (Barth et al., 1974;
Lefèvre et al., 2004), which would offer little or no
protection to potential marsbugs. Even though Mars
receives about half as much total solar radiation of Earth,
the amount of UV at the surface is much higher. Sun-
exposed surfaces on Mars receive sterilizing doses of UV
radiation. However, presumed planetary engineering
processes would release additional CO2 into the
atmosphere, and reduce the UV flux at the surface. For
example, Mars currently receives approximately 3.5
Watts m-2
UVC at the equator during the Vernal Equinox,
but with an atmosphere of 500 mbar CO2, the UVC flux
drops to 0.9 Watts m-2
(Cockell et al., 2000). Also, Mars
doesn't internally generate a magnetic field like Earth's,
and possesses only patchy remnant crustal magnetic fields
(Acuna et al., 1999; Acuna et al., 1998; Connerney et al.,
2004; Connerney et al., 2001). Although some localized
remnant magnetic fields are up to 30 times stronger than
those of Earth (Connerney et al., 2001), the lack of a
global magnetic field allows the surface of Mars to
receives more cosmic radiation than Earth – 20-30
centiSieverts/year (Cucinotta et al., 2001). A marsbug
would need mechanisms to resist radiation damage, and
repair any damage that occurs. Endolithic cyanobacteria,
which have been suggested as models for martian
microorganisms (Friedmann and Ocampo-Friedman,
1994; Friedmann and Ocampo-Friedmann, 1984; Thomas
and Schimel, 1991), protect themselves from UV by their
habitats (porous rocks) and by the production of
photoprotective pigments (Villar et al., 2005). The
bacterium, Deinococcus radiodurans, survives very high
doses of ionizing radiation due to its efficient DNA repair
mechanisms (Battista et al., 1999; Bauche and Laval,
1999; Levin-Zaidman et al., 2003; Venkateswaran et al.,
2000). Desiccation resistant strains of the

D.J. Thomas — Extremophiles for Ecopoiesis
94 Gravitational and Space Biology 19(2) August 2006
cyanobacterium Chroococcidiopsis also exhibit resistance
to ionizing radiation, presumably due to efficient DNA
repair as well (Billi et al., 2000). Protective pigments and
repair enzymes potentially could be genetically added to
other microorganisms, increasing their abilities to live in
the martian environment.
Hypobaric tolerance. Even after initial engineering
efforts, the atmospheric pressure of Mars would be far
lower than that of Earth. While many bacteria can
survive the desiccation that may occur at low pressure, a
marsbug would have to actively metabolize and grow. As
mentioned previously, bacteria may have a 25 mbar lower
limit for active growth (Schuerger et al., 2006a; Schuerger
and Nicholson, 2005). Bacteria have been isolated from
the upper atmosphere; however, their metabolic state
remains controversial (Imshenetsky et al., 1977;
Wainwright et al., 2003; Wainwright et al., 2004). They
may be actively growing without ever "touching ground,"
or they may be dormant while being transported through
the air. If these bacteria do indeed remain active while in
the upper atmosphere, they may have physiological
characteristics that could be useful on Mars.
"Switchable" genes. While the introduction of new
genes into organisms is relatively easy, their regulation is
more problematic. Many of the characteristics described
here are metabolic opposites (e.g., respiration and
photosynthesis, denitrification and nitrogen fixation). The
ability to control pioneer microorganisms over large areas
with the application of a very dilute controlling agent
would be highly desirable. Large numbers of genes are
switched on and off in organisms that have multiple
energetic pathways (Garlick et al., 1977; Guay and Silver,
1975; Sorokin et al., 2000). "Switchable" genes are also
desirable in genetically engineered agricultural crops to
make them resistant to environmental stresses (Sarhan
and Danyluk, 1998); research in this area may also benefit
planetary engineering.
A relatively small number of experiments have examined
the effects of parts of Mars' environment on potential
pioneer organisms, including UV radiation (Cockell et al.,
2005; Hansen et al., 2005; Nicholson and Schuerger,
2005; Schuerger et al., 2003), high CO2 (Hart et al., 2000;
Kanervo et al., 2005; Thomas et al., 2005), low pressure
and hypoxia (Boston, 1981; Kanervo et al., 2005; Paul et
al., 2004). However, these experiments only integrated 2-
5 of the variable conditions that would occur during
ecopoeisis on Mars. To address this problem, SHOT, Inc.
built a simulator capable of specifically replicating most
of the environmental parameters (with the notable
exceptions of gravity and cosmic radiation) on Mars
during all phases of planetary engineering (Thomas et al.,
in review).
ECOPOESIS SIMULATIONS
The SHOT Martian Environment Simulator has been
operational since May 2005. As of October 2005, six
experiments have been performed with durations of 24
hours to six weeks (Thomas et al., accepted for
publication). The initial experiments were primarily to
determine logistical and analytical needs for longer-
duration research, but we also determined short-term
survival and growth characteristics for a variety of
heterotrophic and autotrophic bacteria.
A 24-hour day was used in all experiments. Illumination
was provided by a xenon arc lamp (Sylvania 69263-0
Short Arc Lamp, XBO, 1000 W/HS OFR) fitted with a
solar filter that provided a close approximation of solar
radiation. Photosynthetically active radiation at sample
level ranged from 15 µmol photons m-2
s-1
in the shaded
region to 1000 µmol photons m-2
s-1
in direct light. Total
ultraviolet radiation (250-400 nm) was 1.7 µmol photons
m-2
s-1
in the shaded region, and 50 µmol photons m-2
s-1
in direct light. The temperature regime cycled from a low
of -80°C to a high of 26°C, diurnally. This approximated
the lower latitudes of Mars during the vernal equinox
(Carr, 1996), but probably is more extreme than what
would occur after the initial stages of planetary
engineering. As of October 2005, all of the experiments
were performed at an atmospheric pressure of 100 mbar—
10 times Mars' current highest pressure, but only 10% of
Earth's atmospheric pressure. For experiments of 14 days
or less, we used simulated atmosphere of 95% CO2, 2.7%
N2, 1.6% Ar and 0.13% O2 (Owen et al., 1977). For
longer-duration experiments, we used an atmosphere of
100% CO2. In most of the experiments, water was added
(1 mL per day or less) in order to maintain atmospheric
water saturation.
Cyanobacteria stock cultures were grown in liquid BG-11
medium, and diluted to A720 = 0.25 with fresh BG-11.
Heterotrophic bacteria stock cultures were grown in
trypticase soy broth (TSB), and diluted to A720 = 0.25 with
fresh TSB. Several sample configurations were tested,
including open trays, multi-well tissue culture plates and
arrays of individual sample containers. Individual liquid
cultures or mixtures of cultures were added to sterile JSC
Mars-1 simulant (5-15 g, depending on the container) to
the point of saturation. Desert varnish and cave
microorganisms were grown on BG-11 agar. Agar
cultures were than macerated and mixed with JSC Mars-
1. While samples were in the simulator, parallel control
samples were kept at 4°C in darkness. In cases where
samples could not be analyzed at the SHOT facility, the
samples were placed on ice and transported by car or by
overnight courier.
Samples were analyzed for esterase activity via an assay
of fluorescein diacetate (FDA) hydrolysis (Adam and
Duncan, 2001; Schnürer and Rosswall, 1982) at the
beginning and end of each experiment. The FDA
hydrolysis assay indicates microbial metabolism across a
wide variety of taxa, and correlates well with assays of
respiration. Subsamples of 0.5 - 1.0 g were taken from
each sample before and after each experiment and
transferred into 15 mL centrifuge tubes. 5 mL of 60 mM
K2PO4 buffer (pH 7.6) was added to each tube and briskly
shaken for 10-20 seconds. Ten µL FDA in ethanol (5 mg

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 95
mL-1
) was added to each tube, and then all tubes were
incubated for 3-5 hours at 25°C on a rocker table.
Following incubation, the samples were extracted by
adding 5 mL 2:1 chloroform:methanol. The samples were
centrifuged for 10 minutes at 1000 x g, and the
supernatant was measured spectrophotometrically at 490
nm.
Chlorophyll a extractions were also used to determine the
relative abundance of photosynthetic organisms (Bowles
et al., 1985; Myers et al., 1980). Subsamples of 0.5 - 1.0
g were taken from each sample before and after each
experiment and transferred into 15 mL centrifuge tubes. 5
mL of 80% ethanol was added to each tube. Tubes were
placed in a -20°C freezer overnight, and then were
centrifuged for 10 minutes at 1000 x g. The supernatant
was measured spectrophotometrically at 664 nm.
For the first two experiments (24 hours and 14 days),
plate counts (Atlas, 1997) and trypan blue live-dead stains
(Sigma-Aldrich Co., 2005) were also used to determine
survival. However, the tests were very time-consuming
with ambiguous results, and were discontinued in favor of
the FDA and chlorophyll assays.
The full results of the initial experiments are reported
elsewhere (Thomas et al., accepted for publication).
Here, we summarize the most relevant results.
In experiments of seven days or longer duration, a "water
cycle" became evident within the test chamber as water
tended to condense at the ends as the chamber cooled at
"night," and the samples at the ends of the chamber
tended to be moister than samples in the middle. While
this was disconcerting at first, it probably approximates
the conditions on a planet-wide scale—some areas of
Mars will be wetter than others, and life will become
distributed according to moisture levels.
Live-dead microscopic assays and plate counts showed
survival of all of the organisms tested in the overnight and
14-day trials. However, because of the small sizes of the
cells tested, the live-dead assays were prone to large
errors. Non-cellular material was sometimes counted as
cells, and some cells were incorrectly counted as debris.
Plate counts of cyanobacteria took 1-2 weeks in order to
grow countable colonies. In addition, Plectonema and
Anabaena, being filamentous, gave underestimated results
from plate counts. Further use of these assays was
discontinued.
Figure 1. FDA hydrolysis by eubacteria and cyanobacteria after a 14-day ecopoesis trial. Hydrolysis rates are per gram of total soil
simulant. Samples were placed in two 24-well tissue culture plates—one in direct light, the other in shadow. "Atacama," "Rock Garden,"
and "Yungay" refer to partially-characterized, heterotrophic Atacama Desert isolates. "Atacama 2002" is a strain of Klebsiella oxytoca;
"Rock Garden-2" is a strain of Bacillus licheniformis; and "Yungay-2" is another strain of Bacillus. The other "Rock Garden" strains
appear to be species of Staphylococcus (but not S. aureus). All heterotrophic bacteria were exposed in JSC Mars-1 simulant amended with
trypticase soy broth. The cyanobacteria (five organisms on the right side of the graph) were grown in JSC Mars-1 amended with BG-11
medium. Each bar represents a single sample.
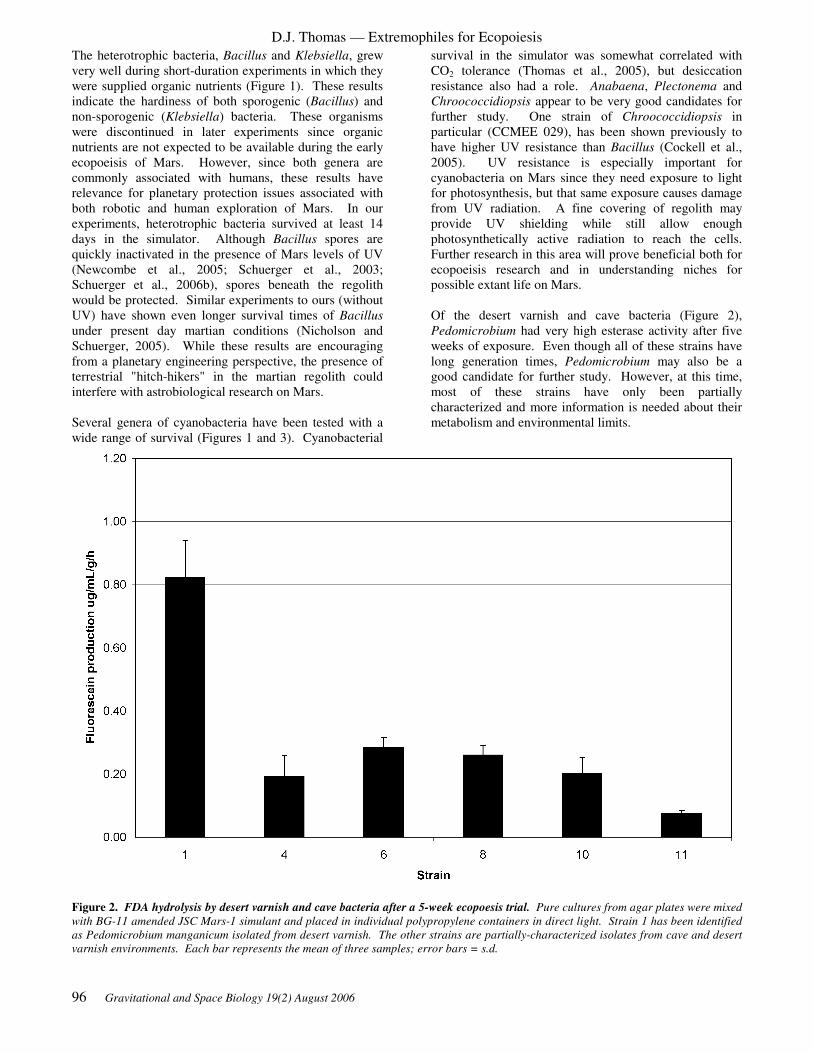
D.J. Thomas — Extremophiles for Ecopoiesis
96 Gravitational and Space Biology 19(2) August 2006
The heterotrophic bacteria, Bacillus and Klebsiella, grew
very well during short-duration experiments in which they
were supplied organic nutrients (Figure 1). These results
indicate the hardiness of both sporogenic (Bacillus) and
non-sporogenic (Klebsiella) bacteria. These organisms
were discontinued in later experiments since organic
nutrients are not expected to be available during the early
ecopoeisis of Mars. However, since both genera are
commonly associated with humans, these results have
relevance for planetary protection issues associated with
both robotic and human exploration of Mars. In our
experiments, heterotrophic bacteria survived at least 14
days in the simulator. Although Bacillus spores are
quickly inactivated in the presence of Mars levels of UV
(Newcombe et al., 2005; Schuerger et al., 2003;
Schuerger et al., 2006b), spores beneath the regolith
would be protected. Similar experiments to ours (without
UV) have shown even longer survival times of Bacillus
under present day martian conditions (Nicholson and
Schuerger, 2005). While these results are encouraging
from a planetary engineering perspective, the presence of
terrestrial "hitch-hikers" in the martian regolith could
interfere with astrobiological research on Mars.
Several genera of cyanobacteria have been tested with a
wide range of survival (Figures 1 and 3). Cyanobacterial
survival in the simulator was somewhat correlated with
CO2 tolerance (Thomas et al., 2005), but desiccation
resistance also had a role. Anabaena, Plectonema and
Chroococcidiopsis appear to be very good candidates for
further study. One strain of Chroococcidiopsis in
particular (CCMEE 029), has been shown previously to
have higher UV resistance than Bacillus (Cockell et al.,
2005). UV resistance is especially important for
cyanobacteria on Mars since they need exposure to light
for photosynthesis, but that same exposure causes damage
from UV radiation. A fine covering of regolith may
provide UV shielding while still allow enough
photosynthetically active radiation to reach the cells.
Further research in this area will prove beneficial both for
ecopoeisis research and in understanding niches for
possible extant life on Mars.
Of the desert varnish and cave bacteria (Figure 2),
Pedomicrobium had very high esterase activity after five
weeks of exposure. Even though all of these strains have
long generation times, Pedomicrobium may also be a
good candidate for further study. However, at this time,
most of these strains have only been partially
characterized and more information is needed about their
metabolism and environmental limits.
Figure 2. FDA hydrolysis by desert varnish and cave bacteria after a 5-week ecopoesis trial. Pure cultures from agar plates were mixed
with BG-11 amended JSC Mars-1 simulant and placed in individual polypropylene containers in direct light. Strain 1 has been identified
as Pedomicrobium manganicum isolated from desert varnish. The other strains are partially-characterized isolates from cave and desert
varnish environments. Each bar represents the mean of three samples; error bars = s.d.

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 97
Figure 3. FDA hydrolysis within a simulated soil community after a 5-week ecopoesis trial. A simulated soil community was formed by
mixing cultures of Anabaena sp., Chroococcidiopisis CCMEE171 and CCMEE662, Plectonema boryanum, Klebsiella oxytoca, Bacillus
licheniformis and Bacillus sp. in JSC Mars-1 simulant amended with BG-11 medium. The soil was spread over a polystyrene tray, which
was placed half in light and half in shadow. After the experiment, the soil was divided into three regions: direct light, transition, and
indirect light. Composite samples were mixtures of soil from all three regions. Three samples were obtained from each region and
averaged; error bars = s.d.
CONCLUSIONS
With the increasing availability of martian environment
simulators, the science of planetary engineering is moving
from the theoretical to the experimental realm. Although
planetary-scale ecopoesis is still in the distant future,
population and community level experiments are possible
and are currently in progress. In addition to providing
insight about possible future life on Mars, these
experiments also tell us about the survivability of Earth
organisms in the present martian environment.
As additional simulators become available, voluntary
standardization of environmental parameters will be
desirable. We envision a consortium that would set
guidelines for planetary simulators so that experiments
undertaken with different simulators would still be
directly comparable. Efforts to form such a consortium
are currently underway.
Ecopoeisis also provides an exciting opportunity to
engage students in science. While many students have
heard of terraforming through Star Trek and other science
fiction stories, they usually do not realize that serious
science underlies the fiction. Ecopoesis can be used as a
focal point for discussions of geology, environmental
science, microbiology, ecology and other disciplines to
show the interdisciplinary nature of planetary science—
whether that planet is Earth, Mars or one that has not been
discovered yet.
ACKNOWLEDGEMENTS
This research was funded under subcontract agreement
07605-003-026 with NASA's Institute for Advanced
Concepts (NIAC). Additional support was provided by
the Arkansas Space Grant Consortium. KAC's
participation was supported in part by the Arkansas
Supporting Teachers Research Involvement for Vital
Education (STRIVE) Program.
REFERENCES
Abyzov, S. S., Zavarzin, G. A., Ivanov, M. V., Seregin,
V. I., and Shirokov, O. G. 1967. On the possibility of
protracted anabiosis in micro-organisms. Life Sciences
and Space Research 5: 261-267.
Acuna, M. H., Connerney, J. E. P., Ness, N. F., Lin, R. P.,
Mitchell, D., Carlson, C. W., McFadden, J., Anderson, K.
A., Reme, H., Mazelle, C., Vignes, D., Wasilewski, P.,
and Cloutier, P. 1999. Global distribution of crustal

D.J. Thomas — Extremophiles for Ecopoiesis
98 Gravitational and Space Biology 19(2) August 2006
magnetization discovered by the Mars Global Surveyor
MAG/ER experiment. Science 284: 790-793.
Acuna, M. H., Connerney, J. E. P., Wasilewski, P., Lin,
R. P., Anderson, K. A., Carlson, C. W., McFadden, J.,
Curtis, D. W., Mitchell, D., Reme, H., Mazelle, C.,
Sauvaud, J. A., d'Uston, C., Cros, A., Medale, J. L.,
Bauer, S. J., Cloutier, P., Mayhew, M., Winterhalter, D.,
and Ness, N. F. 1998. Magnetic field and plasma
observations at Mars: initial results of the Mars Global
Surveyor mission. Science 279: 1676-1680.
Adam, G., and Duncan, H. 2001. Development of a
sensitive and rapid method for the measurement of total
microbial activity using fluorescein diacetate (FDA) in a
range of soils. Soil Biology and Biochemistry 33: 943-
951.
Almon, H., and Böger, P. 1988. Hydrogen metabolism of
the unicellular cyanobacterium Chroococcidiopsis
thermalis ATCC29380. FEBS Letters 49: 445-449.
Alpi, A., and Beevers, H. 1983. Effects of O2
concentration on rice seedlings. Plant Physiology 71: 30-
34.
Atlas, R. M. 1997. Principles of Microbiology. Wm. C.
Brown Publishers, Dubuque, IA.
Atwell, B. J., Waters, I., and Greenway, H. 1982. The
effect of oxygen and turbulence on elongation of
coleoptiles of submergence-tolerant and -intolerant rice
cultivars. Journal of Experimental Botany 33: 1030-1044.
Baker, S. C., Ferguson, S. J., Ludwig, B., Page, M. D.,
Richter, O. M. H., and van Spanning, R. J. M. 1998.
Molecular genetics of the genus Paracoccus:
metabolically versatile bacteria with bioenergetic
flexibility. Microbiology and Molecular Biology Reviews
62: 1046-1078.
Barclay, A. M., and Crawford, R. M. M. 1982. Plant
growth and survival under strict anaerobiosis. Journal of
Experimental Botany 33: 541-549.
Barth, C. A., Hord, C. W., Stewart, A. I., Lane, A. L.,
Dick, M. L., and Anderson, G. P. 1974. Mariner 9
Ultraviolet Spectrometer Experiment: seasonal variation
of ozone on Mars. Science 179: 795-796.
Battista, J. R., Earl, A. M., and Park, M. J. 1999. Why is
Deinococcus radiodurans so resistant to ionizing
radiation? Trends in Microbiology 7: 362-365.
Bauche, C., and Laval, J. 1999. Repair of oxidized bases
in the extremely radiation-resistant bacterium
Deinococcus radiodurans. Journal of Bacteriology 181:
262-269.
Berman-Frank, I., Lundgren, P., Chen, Y. B., Kupper, H.,
Kolber, Z., Bergman, B., and Falkowski, P. 2001.
Segregation of nitrogen fixation and oxygenic
photosynthesis in the marine cyanobacterium
Trichodesmium. Science 294: 1534-1537.
Billi, D., Friedmann, E. I., Helm, R. F., and Potts, M.
2001. Gene transfer to the desiccation-tolerant
cyanobacterium Chroococcidiopsis. Journal of
Bacteriology 183: 2298-2305.
Billi, D., Friedmann, E. I., Hofer, K. G., Caiola, M. G.,
and Ocampo-Friedmann, R. 2000. Ionizing-radiation
resistance in the desiccation-tolerant cyanobacterium
Chroococcidiopsis. Applied and Environmental
Microbiology 66: 1489-1492.
Blankenship, R. E., and Hartman, H. 1998. The origin and
evolution of oxygenic photosynthesis. Trends in the
Biochemical Sciences 23: 94-97.
Böhme, H. 1998. Regulation of nitrogen fixation in
heterocyst-forming cyanobacteria. Trends in Plant
Science 3: 346-351.
Boston, P. J. 1981. Low-pressure greenhouses and plants
for a manned research station on Mars. Journal of the
British Interplanetary Society 34: 189-192.
Boston, P. J., Ivanov, M. V., and McKay, C. P. 1992. On
the possibility of chemosynthetic ecosystems in
subsurface habitats on Mars. Icarus 95: 300-8.
Boston, P. J., Todd, P., and McMillen, K. 2004. Robotic
lunar ecopoiesis test bed: bringing the experimental
method to terraforming, AIP #654. Space Technology and
Applications Forum 2003 Proceedings. American
Institute of Physics, College Park, MD.
Bowles, N. D., Paerl, H. W., and Tucker, J. 1985.
Effective solvents and extraction periods employed in
phytoplankton carotenoid and chlorophyll determinations.
Canadian Journal of Fishery and Aquatic Science 42:
1127-1131.
Burger-Wiersma, T., and Matthijs, H. C. P. 1990. The
biology of the Prochlorales. Pages 1-24 in Codd, G. A.,
Dijkhuizen, L., and Tabita, F. R., eds. Autotrophic
Microbiology and One-Carbon Metabolism. Kluwer
Academic Publishers, Dordrecht.
Carr, M. H. 1996. Water on Mars. Oxford University
Press, New York.
Chattopadhyay, M., and Jagannadham, M. 2001.
Maintenance of membrane fluidity in Antarctic bacteria.
Polar Biology 24: 386-388.
Chen, C. H., and Berns, D. S. 1978. Comparison of the
stability of phycocyanins from thermophilic, mesophilic,
psychrophilic and halophilic algae. Biophysical Chemistry
8: 203-213.

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 99
Christner, B. C., Kvitko, B. H., 2nd, and Reeve, J. N.
2003. Molecular identification of Bacteria and Eukarya
inhabiting an Antarctic cryoconite hole. Extremophiles 7:
177-83.
Chyba, C. F., and Hand, K. P. 2001. Life without
photosynthesis. Science 292: 2026-2027.
Clare, D. A., Rabinowitch, H. D., and Fridovich, I. 1984.
Superoxide dismutase and chilling injury in Chlorella
ellipsoidea. Archives of Biochemistry and Biophysics 231:
158-163.
Cockell, C. S., Catling, D. C., Davis, W. L., Snook, K.,
Kepner, R. L., Lee, P., and McKay, C. P. 2000. The
ultraviolet environment of Mars: biological implications
past, present, and future. Icarus 146: 343-359.
Cockell, C. S., Schuerger, A. C., Billi, D., Friedmann, E.
I., and Panitz, C. 2005. Effects of a simulated martian UV
flux on the cyanobacterium, Chroococcidiopsis sp. 029.
Astrobiology 5: 127-140.
Connerney, J. E. P., Acuña, M. H., Ness, N. F., Spohn, T.,
and Schubert, G. 2004. Mars crustal magnetism. Space
Science Reviews 111: 1-32.
Connerney, J. E. P., Acuña, M. H., Wasilewski, P. J.,
Kletetschka, G., Ness, N. F., Rème, H., Lin, R. P., and
Mitchell, D. L. 2001. The global magnetic field of Mars
and implications for crustal evolution. Geophysical
Research Letters 28: 4015-4018.
Crispim, C. A., Gaylarde, P. M., and Gaylarde, C. C.
2003. Algal and cyanobacterial biofilms on calcareous
historic buildings. Current Microbiology 46: 79-82.
Daugbjerg, N. 2000. Pyramimonas tychotreta, sp. nov.
(Prasinophyceae), a new marine species from Antarctica:
light and electron microscopy of the motile state and
notes on growth rates. Journal of Phycology 36: 160-171.
de Winder, B., Matthijs, H. C. P., and Mur, L. R. 1989.
The role of water-retaining substrata on the
photosynthetic response of three drought tolerant
phototrophic micro-organisms isolated from a terrestrial
habitat. Archives of Microbiology 152: 458-462.
Dismukes, G. C., Klimov, V. V., Baranov, S. V., Kozlov,
Y. N., DasGupta, J., and Tyryshkin, A. 2001. The origin
of atmospheric oxygen on Earth: the innovation of
oxygenic photosynthesis. Proceedings of the National
Academy of Sciences USA 98: 2170-2175.
Dorn, R. I., and Oberlander, T. M. 1981. Microbial origin
of desert varnish. Science 213: 1245-1247.
Fay, P. 1983. The Blue-Greens. Edward Arnold
(Publishers) Ltd., London.
Ferreira, A. C., Nobre, M. F., Moore, E., Rainey, F. A.,
Battista, J. R., and da Costa, M. S. 1999. Characterization
and radiation resistance of new isolates of Rubrobacter
radiotolerans and Rubrobacter xylanophilus.
Extremophiles 3: 235-8.
Frankenberg-Schwager, M., Turcu, G., Thomas, C.,
Wollenhaupt, H., and Bucker, H. 1975. Membrane
damage in dehydrated bacteria and its repair. Life
Sciences and Space Research 13: 83-88.
Friedmann, E. I., Hua, M., and Ocampo-Friedmann, R.
1993. Terraforming Mars: dissolution of carbonate rocks
by cyanobacteria. Journal of the British Interplanetary
Society 46: 291-292.
Friedmann, E. I., and Kibler, A. P. 1980. Nitrogen
economy of endolithic microbial communities in hot and
cold deserts. Microbial Ecology 6: 95-108.
Friedmann, E. I., and Ocampo-Friedman, R. 1994. A
primitive cyanobacterium as pioneer organism for
terraforming Mars. Advances in Space Research.
Friedmann, E. I., and Ocampo-Friedmann, R. 1984. The
antarctic cryptoendolithic ecosystem: relevance to
exobiology. Origins of Life 14: 771-776.
Gaidos, E., Lanoil, B., Thorsteinsson, T., Graham, A.,
Skidmore, M., Han, S. K., Rust, T., and Popp, B. 2004. A
viable microbial community in a subglacial volcanic
crater lake, iceland. Astrobiology 4: 327-44.
Garlick, S., Oren, A., and Padan, E. 1977. Occurrence of
facultative anoxygenic photosynthesis among filamentous
and unicellular cyanobacteria. Journal of Bacteriology
129: 623-629.
Gerday, C., Aittaleb, M., Bentahir, M., Chessa, J. P.,
Claverie, P., Collins, T., D'Amico, S., Dumont, J.,
Garsoux, G., Georlette, D., Hoyoux, A., Lonhienne, T.,
Meuwis, M. A., and Feller, G. 2000. Cold-adapted
enzymes: from fundamentals to biotechnology. Trends in
Biotechnology 18: 103-107.
Gilichinsky, D., Rivkina, E., Shcherbakova, V.,
Laurinavichuis, K., and Tiedje, J. 2003. Supercooled
water brines within permafrost-an unknown ecological
niche for microorganisms: a model for astrobiology.
Astrobiology 3: 331-41.
Golden, J. W., and Yoon, H. S. 1998. Heterocyst
formation in Anabaena. Current Opinion in Microbiology
1: 623-629.
Goodwin, T. W. 1980. Algae. Pages 207-256. The
Biochemistry of the Carotenoids: Plants. Richard Clay
(The Chaucer Press), Ltd., Suffolk.

D.J. Thomas — Extremophiles for Ecopoiesis
100 Gravitational and Space Biology 19(2) August 2006
Graham, J. M. 2004. The biological terraforming of Mars:
planetary ecosynthesis as ecological succession on a
global scale. Astrobiology 4: 168-95.
Guay, R., and Silver, M. 1975. Thiobacillus acidophilus
sp. nov.; isolation and some physiological characteristics.
Canadian Journal of Microbiology 21: 281-288.
Hansen, A. A., Merrison, J., Nørnberg, P., Lomstein, B.
A., and Finster, K. 2005. Activity and stability of a
complex bacterial soil community under simulated
Martian conditions. International Journal of Astrobiology
4: 135-144.
Hart, S. D., Currier, P. A., and Thomas, D. J. 2000.
Denitrification by Pseudomonas aeruginosa under
simulated engineered martian conditions. Journal of the
British Interplanetary Society 53: 357-359.
Hawrylewicz, E. J., Hagen, C., Tolkacz, V., and Ehrlich,
R. 1967. Effect of reduced barometric pressure on water
availability related to microbial growth. Life Sciences and
Space Research 5: 174-186.
Hershovitz, N., Oren, A., and Cohen, Y. 1991a.
Accumulation of trehalose and sucrose in cyanobacteria
exposed to matric water stress. Applied and
Environmental Microbiology 57: 645-648.
Hershovitz, N., Oren, A., Post, A., and Cohen, Y. 1991b.
Induction of water-stress proteins in cyanobacteria
exposed to matric- or osmotic-water stress. FEMS
Microbiology Letters 83: 169-172.
Hiscox, J. A., and Thomas, D. J. 1995. Genetic
modification and selection of microorganisms for growth
on Mars. Journal of the British Interplanetary Society 48:
419-426.
Ho, K. K., and Krogman, D. W. 1982. Photosynthesis.
Pages 191-214 in Carr, N. G. and Whitton, B. A., eds. The
Biology of Cyanobacteria. University of California Press,
Berkeley.
Hodgson, R. A. J., and Raison, J. K. 1991a. Lipid
peroxidation and superoxide dismutase activity in relation
to photoinhibition induced by chilling in moderate light.
Planta 185: 215-219.
Hodgson, R. A. J., and Raison, J. K. 1991b. Superoxide
production by thylakoids during chilling and its
implication in the susceptibility of plants to chilling-
induced photoinhibition. Planta 183: 222-228.
Hunten, D. M. 1979. Possible oxidant sources in the
atmosphere and surface of Mars. Journal of Molecular
Evolution 14: 71-8.
Imshenetsky, A. A., Abyzov, S. S., Voronov, G. T.,
Kuzjurina, L. A., Lysenko, S. V., Sotnikov, G. G., and
Fedorova, R. I. 1967. Exobiology and the effect of
physical factors on micro-organisms. Life Sciences and
Space Research 5: 250-260.
Imshenetsky, A. A., and Lysenko, S. V. 1965. Ultra-high
vacuum and microorganisms. Life Sciences and Space
Research 3: 142-148.
Imshenetsky, A. A., Lysenko, S. V., Kasakov, G. A., and
Ramkova, N. V. 1977. Resistance of stratospheric and
mesospheric micro-organisms to extreme factors. Life
Sciences and Space Research 15: 37-39.
Imshenetsky, A. A., Lysenko, S. V., and Komolova, G. S.
1970. The effect of high vacuum on oxidative reactions in
bacteria and the activity of certain enzymes. Life Sciences
and Space Research 8: 77-81.
Jackson, M. B., and Drew, M. C. 1984. Effects of
flooding on growth and metabolism of herbaceous plants.
Pages 47-128 in Kozlowski, T. T., ed. Flooding and Plant
Growth. Academic Press, Orlando.
Kanervo, E., Lehto, K., Ståhle, K., Lehto, H., and
Mäenpää, P. 2005. Characterization of growth and
photosynthesis of Synechocystis sp. PCC 6803 cultures
under reduced atmospheric pressures and enhanced CO2
levels. International Journal of Astrobiology 4: 97-100.
Kaplan, A., Marcus, Y., and Reinhold, L. 1988. Inorganic
carbon uptake by cyanobacteria. Pages 534-539. Methods
in Enzymology. Academic Press, New York.
Klingler, J. M., Mancinelli, R. L., and White, M. R. 1989.
Biological nitrogen fixation under primordial Martian
partial pressures of dinitrogen. Advances in Space
Research 9: 173-6.
Kraegeloh, A., and Kunte, H. J. 2002. Novel insights into
the role of potassium for osmoregulation in Halomonas
elongata. Extremophiles 6: 453-62.
Kumon, Y., Sasaki, Y., Kato, I., Takaya, N., Shoun, H.,
and Beppu, T. 2002. Codenitrification and denitrification
are dual metabolic pathways through which dinitrogen
evolves from nitrate in Streptomyces antibioticus. Journal
of Bacteriology 184: 2963-2968.
Lefèvre, F., Lebonnois, S., Montmessin, F., and Forget, F.
2004. Three-dimensional modeling of ozone on Mars.
Journal of Geophysical Research 109: 1-20.
Levin-Zaidman, S., Englander, J., Shimoni, E., Sharma,
A. K., Minton, K. W., and Minsky, A. 2003. Ringlike
structure of the Deinococcus radiodurans genome: a key
to radioresistance? Science 299: 254-256.
Mancinelli, R. L. 1996. The nature of nitrogen: an
overview. Life Support and Biosphere Science 3: 17-24.
McKay, C. P. 1998. Past, present, and future life on Mars.
Gravitational and Space Biology Bulletin 11: 41-50.

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 101
McKay, C. P., Toon, O. B., and Kasting, J. F. 1991.
Making Mars habitable. Nature 352: 489-496.
McMinn, A., Pankowski, A., and Delfatti, T. 2005. Effect
of hyperoxia on the growth and photosynthesis of polar
sea ice algae. Journal of Phycology 41: 732-741.
Mitsui, A., and Cao, S. 1988. Isolation and culture of
marine nitrogen-fixing unicellular cyanobacteria
Synechococcus. Pages 105-113 in Packer, L. and Glazer,
A. N., eds. Cyanobacteria. Academic Press, Inc., San
Diego.
Morita, R. Y. 1975. Psychrophilic bacteria.
Bacteriological Reviews 39: 144-67.
Mulholland, M. R., and Capone, D. G. 2000. The nitrogen
physiology of the marine N2-fixing cyanobacteria
Trichodesmium spp. Trends in Plant Science 5: 148-153.
Myers, J., Graham, J.-R., and Wang, R. T. 1980. Light
harvesting in Anacystis nidulans studied in pigment
mutants. Plant Physiology 66: 1144-1149.
Negoro, M., Shioji, N., Miyamoto, K., and Miura, Y.
1991. Growth of microalgae in high CO2 gas and effects
of SOX and NOX. Applied Biochemistry and
Biotechnology 28-29: 877-886.
Newcombe, D. A., Schuerger, A. C., Benardini, J. N.,
Dickinson, D., Tanner, R., and Venkateswaran, K. 2005.
Survival of spacecraft-associated microorganisms under
simulated martian UV irradiation. Applied and
Environmental Microbiology 71: 8147-8156.
Nicholson, W. L., and Schuerger, A. C. 2005. Bacillus
subtilis spore survival and expression of germination-
induced bioluminescence after prolonged incubation
under simulated Mars atmospheric pressure and
composition: implications for planetary protection and
lithopanspermia. Astrobiology 5: 536-44.
Northup, D. E., Barns, S. M., Yu, L. E., Spilde, M. N.,
Schelble, R. T., Dano, K. E., Crossey, L. J., Connolly, C.
A., Boston, P. J., Natvig, D. O., and Dahm, C. N. 2003.
Diverse microbial communities inhabiting
ferromanganese deposits in Lechuguilla and Spider
Caves. Environmental Microbiology 5: 1071-86.
Olson, J. B., Steppe, T. F., Litaker, R. W., and Paerl, H.
W. 1998. N2-fixing microbial consortia associated with
the ice cover of Lake Bonney, Antarctica. Microbial
Ecology 36: 231-238.
Oremland, R. S., Hoeft, S. E., Santini, J. M., Bano, N.,
Hollibaugh, R. A., and Hollibaugh, J. T. 2002. Anaerobic
oxidation of arsenite in Mono Lake water and by a
facultative, arsenite-oxidizing chemoautotroph, strain
MLHE-1. Applied and Environmental Microbiology 68:
4795-802.
Owen, T., Biemann, K., Rushneck, D. R., Biller, J. E.,
Howarth, D. W., and Lafleur, A. L. 1977. The
composition of the atmosphere at the surface of Mars.
Journal of Geophysical Research 82: 4635-4639.
Paul, A. L., Schuerger, A. C., Popp, M. P., Richards, J. T.,
Manak, M. S., and Ferl, R. J. 2004. Hypobaric biology:
Arabidopsis gene expression at low atmospheric pressure.
Plant Physiology 134: 215-23.
Plumb, R. C., Tantayanon, R., Libby, M., and Xu, W. W.
1989. Chemical model for Viking biology experiments:
implications for the composition of the martian regolith.
Nature 338: 633-635.
Potts, M., Ocampo-Friedmann, R., Bowman, M. A., and
Tözún, B. 1983. Chroococcus S24 and Chroococcus N41
(cyanobacteria): morphological, biochemical and genetic
characterization and effects of water stress on
ultrastructure. Archives of Microbiology 135: 81-90.
Russell, N. J. 2000. Toward a molecular understanding of
cold activity of enzymes from psychrophiles.
Extremophiles 4: 83-90.
Sarhan, F., and Danyluk, J. 1998. Engineering cold-
tolerant crops--throwing the master switch. Trends in
Plant Science 3: 289-290.
Scherer, S., Ernst, A., Chen, T.-W., and Böger, P. 1984.
Rewetting of drought-resistant blue-green algae: time
course of water uptake and reappearance of respiration,
photosynthesis, and nitrogen fixation. Oecologia 62: 418-
423.
Schneegurt, M. A., Tucker, D. L., Ondr, J. K., Sherman,
D. M., and Sherman, L. A. 2000. Metabolic rhythms of a
diazotrophic cyanobacterium, Cyanothece sp. strain
ATCC 51142, heterotrophically grown in continuous
dark. Journal of Phycology 36: 107-117.
Schnürer, J., and Rosswall, T. 1982. Fluorescein diacetate
hydrolysis as a measure of total microbial activity in soil
and litter. Applied and Environmental Microbiology 43:
1256-1261.
Schuerger, A. C., Berry, B., and Nicholson, W. L. 2006a.
Terrestrial bacteria typically recovered from Mars
spacecraft do not appear able to grow under simulated
martian conditions. Pages 1397. 37th Lunar and
Planetary Science Conference, Houston, TX.
Schuerger, A. C., Mancinelli, R. L., Kern, R. G.,
Rothschild, L. J., and McKay, C. P. 2003. Survival of
endospores of Bacillus subtilis on spacecraft surfaces
under simulated martian environments: implications for
the forward contamination of Mars. Icarus 165: 253-276.
Schuerger, A. C., and Nicholson, W. L. 2005. Synergistic
effects of low pressure, low temperature, and CO2

D.J. Thomas — Extremophiles for Ecopoiesis
102 Gravitational and Space Biology 19(2) August 2006
atmospheres inhibit the growth of terrestrial bacteria
under simulated martian conditions. Pages 1366. 36th
Lunar and Planetary Science Conference, Houston, TX.
Schuerger, A. C., Richards, J. T., Newcombe, D. A., and
Venkateswaren, K. 2006b. Rapid inactivation of seven
Bacillus spp. under simulated Mars UV irradiation. Icarus
181: 52-62.
Seckbach, J., Baker, F. A., and Shugarman, P. M. 1970.
Algae thrive under pure CO2. Nature 227: 744-745.
Seckbach, J., and Libby, W. F. 1970. Vegetative life on
Venus? Or investigations with algae which grow under
pure CO2 in hot acid media at elevated pressures. Space
Life Sciences 2: 121-143.
Sigma-Aldrich Co. 2005. Use of trypan blue stain and the
hemocytometer to determine total cell counts and viable
cell number. Sigma-Aldrich Co.
Silverman, G. J., and Beecher, N. 1967. Survival of cocci
after exposure to ultrahigh vacuum at different
temperatures. Applied Microbiology 15: 665-667.
Sorokin, D. Y., Tourova, T. P., and Kuenen, J. G. 2000. A
new facultatively autotrophic hydrogen- and sulfur-
oxidizing bacterium from an alkaline environment.
Extremophiles 4: 237-45.
Stevens, T. O., and McKinley, J. P. 1995.
Lithoautotrophic microbial ecosystems in deep basalt
aquifers. Science 270: 450-454.
Tabita, F. R. 1994. The biochemistry and molecular
regulation of carbon dioxide metabolism in cyanobacteria.
Pages 437-467 in Bryant, D. A., ed. The Molecular
Biology of Cyanobacteria. Kluwer Academic Publishers,
Dordrecht.
Thomas, D. J. 1995. Biological aspects of the ecopoeisis
and terraformation of Mars: current perspectives and
research. Journal of the British Interplanetary Society 48:
415-418.
Thomas, D. J., Boston, P. J., Todd, P., Boling, J.,
Campbell, K., Gregerson, R. G., Holt, A., III,
McSpadden, T., and McWilliams, L. accepted for
publication. Early results of ecopoeisis experiments in the
SHOT martian environment simulator in Crossman, F.,
ed. Proceedings of the 8th International Mars Society
Convention.
Thomas, D. J., and Schimel, J. P. 1991. Mars after the
Viking missions: is life still possible? Icarus 91: 199-206.
Thomas, D. J., Sullivan, S. L., Price, A. L., and
Zimmerman, S. M. 2005. Common freshwater
cyanobacteria grow in 100% CO2. Astrobiology 5: 66-74.
Thomas, D. J., Thomas, J. B., Prier, S. D., Nasso, N. E.,
and Herbert, S. K. 1999. Iron superoxide dismutase
protects against chilling damage in the cyanobacterium
Synechococcus species PCC7942. Plant Physiology 120:
275-282.
Thomas, N. A., Todd, P., Metz, G. W., Platt, H., Kurk, A.,
and Thomas, D. J. in review. Performance evaluation of a
laboratory test bed for planetary biology. Gravitational
and Space Biology.
Venkateswaran, A., McFarlan, S. C., Ghosal, D., Minton,
K. W., Vasilenko, A., Makarova, K., Wackett, L. P., and
Daly, M. J. 2000. Physiologic determinants of radiation
resistance in Deinococcus radiodurans. Applied and
Environmental Microbiology 66: 2620-2626.
Villar, S. E., Edwards, H. G., and Cockell, C. S. 2005.
Raman spectroscopy of endoliths from Antarctic cold
desert environments. Analyst 130: 156-62.
Wainwright, M., Wickramasinghe, N. C., Narlikar, J. V.,
and Rajaratnam, P. 2003. Microorganisms cultured from
stratospheric air samples obtained at 41 km. FEMS
Microbiology Letters 218: 161-165.
Wainwright, M., Wickramasinghe, N. C., Narlikar, J. V.,
and Rajaratnam, P. 2004. Are these stratospheric
nanoparticles bacteria? Microbiology 150: 756-8.
Wise, R. R. 1995. Chilling-enhanced photooxidation: the
production, action and study of reactive oxygen species
produced during chilling in the light. Photosynthesis
Research 45: 79-97.
Wise, R. R., and Naylor, A. W. 1987. Chilling-enhanced
photooxidation. Evidence for the role of singlet oxygen
and superoxide in the breakdown of pigments and
endogenous antioxidants. Plant Physiology 83: 278-282.
Wolk, C. P. 1988. Purification and storage of nitrogen-
fixing filamentous cyanobacteria. Pages 93-95 in Packer,
L. and Glazer, A. N., eds. Cyanobacteria. Academic
Press, Inc., San Diego.
Xiong, J., and Bauer, C. E. 2002. Complex evolution of
photosynthesis. Annual Review of Plant Biology 53: 503-
521.
Yen, A. S., Kim, S. S., Hecht, M. H., Frant, M. S., and
Murray, B. 2000. Evidence that the reactivity of the
martian soil is due to superoxide ions. Science 289: 1909-
1912.
Zecchinon, L., Claverie, P., Collins, T., D'Amico, S.,
Delille, D., Feller, G., Georlette, D., Gratia, E., Hoyoux,
A., Meuwis, M. A., Sonan, G., and Gerday, C. 2001. Did
psychrophilic enzymes really win the challenge?
Extremophiles 5: 313-21.

D.J. Thomas — Extremophiles for Ecopoiesis
Gravitational and Space Biology 19(2) August 2006 103
Zehnder, A. J., and Svensson, B. H. 1986. Life without
oxygen: what can and what cannot? Experientia 42: 1197-
205.
Zehr, J. P., Waterbury, J. B., Turner, P. J., Montoya, J. P.,
Omoregie, E., Steward, G. F., Hansen, A., and Karl, D.
M. 2001. Unicellular cyanobacteria fix N2 in the
subtropical North Pacific Ocean. Nature 412: 635-638.
Zumft, W. G. 1997. Cell biology and molecular basis of
denitrification. Microbiology and Molecular Biology
Reviews 61: 533-616.

D.J. Thomas — Extremophiles for Ecopoiesis
104 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 105
PLANETARY ECOSYNTHESIS AS ECOLOGICAL SUCCESSION. James M. Graham
Department of Botany, University of Wisconsin-Madison, Madison, WI. 53706
ABSTRACT
Terraforming is the process of applying global
engineering techniques to transform the climate of a
planet into one that is habitable for terrestrial organisms.
Ecosynthesis is the process of introducing a succession of
ecosystems to such a terraformed planet. The process of
introducing terrestrial ecosystems to Mars can be
compared to a descent down a terrestrial mountain. Each
drop in elevation results in a warmer, wetter climate and
more diverse biological community. Beginning with a
polar desert, the sequence of ecosystems passes through
tundra, boreal forest, and temperate ecosystems where
moisture determines the presence of desert, grassland, or
forest. Mars is like a very high terrestrial mountain. The
goal of planetary engineering is to bring the climate of
Mars down that mountain to the point where some areas
of the planet have a climate similar to a polar desert At
that point a microbial ecosystem containing bacteria,
cyanobacteria, green algae and lichens can be introduced.
Subsequent engineering and biological feedbacks will
move the climate of Mars further down the mountain
through successively more moderate climates and more
diverse ecosystems. At each stage the organisms will alter
the environment to prepare the way for subsequent stages.
This model provides a logical framework for the sequence
of ecosystems to be established on Mars. The physical
requirements for the introduction of each system on Mars
can be extrapolated from the known limitations on Earth.
The ecosystems define the communities to search for
potential colonizing species based on their physiological
properties.
INTRODUCTION
Mars is a bitterly cold planet. Its average surface
temperature is only –60oC compared to +15
oC for the
Earth. The martian atmosphere has a surface pressure of
only 0.5-1 kilopascals (kPa) or 5–10 mbar, a pressure so
low that liquid water is not stable. Intense ultraviolet
(UV) radiation bathes the surface, and the regolith is
thought to contain oxidants, which would break down any
organic material. Terrestrial organisms cannot survive
under these surface conditions (Clark, 1998). Spacecraft,
however, have returned images of valley systems
resembling dried-up river systems and wide outflow
channels indicating the passage of vast quantities of water
in the past. Mars Global Surveyor (MGS) returned
altimeter data indicating shorelines consistent with the
past existence of a northern ocean (Head et al., 1999). If
water was once stable on the surface and flowed in such
vast quantities, the atmosphere must have been denser and
the climate warmer in the past.
These observations are the basis for the current search for
evidence of past or present life on Mars (McKay, 1997).
They are also the source of speculation that Mars might
be returned to its former warmer climate by some sort of
global engineering techniques (Sagan, 1971; Oberg, 1981;
McKay et al., 1991). The new science of planetary
engineering became known as “terraforming” in which an
extraterrestrial body is modified to make an environment
suitable for habitation by terrestrial organisms. The term
“planetary ecosynthesis” emphasizes the fact that the
result of planetary engineering is not a carbon copy of
Earth but a habitable world with its own unique
properties. Fogg gathered this early work in
Terraforming: Engineering Planetary Environments
(1995), and review articles by Fogg (1998), McKay
(1999), and McKay and Marinova (2001) have
established terraforming as a branch of the field of
astrobiology.
As established by NASA the Astrobiology program
includes the study of planetary ecosynthesis. One of the
six key questions addressed by the Astrobiology Institute
is: What is the potential for survival and biological
evolution beyond the planet of origin? Biological
evolution is a phenomenon of populations of organisms
interacting and genetically changing over long periods of
time. To address this question entire ecosystems
(communities of organisms plus their physical and
chemical environment) would have to be placed on an
extraterrestrial planetary body, and Mars is the only
planet in the Solar System capable of receiving terrestrial
ecosystems after modification with existing technology.
The purpose of this paper is to present a model for the
process of the introduction of terrestrial ecosystems to
Mars. The model provides a framework for the sequence
of ecosystems to be established and the physical
requirements for each system. The individual ecosystems
define the communities from which potential colonists
may be selected based on their biological characteristics.
The potential colonists then define areas for research to be
performed before the actual transplantation to Mars.
A MODEL FOR ECOSYNTHESIS ON MARS
A model for the process of ecosynthesis on Mars can be
found in a diagram often found in introductory biology
texts. As one climbs up a mountain, each 150 m rise in
____________________
* Correspondence to: Dr. James M. Graham
Department of Botany
430 Lincoln Drive
University of Wisconsin-Madison
Madison, WI 53706
Email: [email protected]
Phone: 608-262-0657

J.M. Graham — Ecosynthesis on Mars
106 Gravitational and Space Biology 19(2) August 2006
Figure 1. The mountain model for ecosynthesis on Mars. The present climate of Mars can be compared to a very high mountain on Earth,
one higher than any real mountain that exists in fact. The prebiotic stage of ecosynthesis employs planetary engineering techniques to
bring the climate of Mars at some latitudes into the range of that in the terrestrial Dry Valleys of Antarctica. At that point the introduction
of the first microbial ecosystems can begin. Continued modification of the planetary climate will permit the introduction of a sequence of
ecosystems from tundra through boreal forests to temperate ecosystems.
elevation produces a decrease in average annual
temperature of about 1oC. Moisture and evaporation also
change with altitude. Therefore anyone climbing up a
mountain passes through a series of ecosystems from base
to top. Depending on the latitude of the mountain, the
base may be surrounded in tropical forest or temperate
forest, a prairie or sagebrush area above that, followed by
a montane or coniferous forest, an alpine tundra at higher
elevations, and finally a polar desert ending in permanent
snow and ice. Since the effect is similar at all latitudes,
mountains in the tropics can be capped in permanent ice if
the mountain is high enough. This is the model of the
ecosynthesis process for Mars (Figure 1). The present
climate of Mars is like that of a very high mountain on
Earth. The average atmospheric surface pressure on Mars
(6-7 mbar) occurs in the atmosphere of Earth at an
altitude of about 37,000 m (121,000 ft), far higher than
any terrestrial mountain.
The mountain model of ecosynthesis on Mars provides an
order to the sequence of ecosystems that will be
implanted. The initial engineering stage will employ
global techniques to move the climate of Mars from its
present state to one similar to a high arctic barren or polar
desert. The first ecosystem will then be a microbial one
dominated by the kinds of microorganisms found in the
Dry Valleys of Antarctica or polar deserts in the Arctic.
The second ecosystem in the sequence will be similar to
arctic tundra and dominated by plants called bryophytes
(mosses and liverworts) and lichens (organisms formed
by symbioses between algae and fungi). The third
ecosystem will be characterized by the arrival of
flowering plants, initially as small numbers of herbs but
with an increasing role as climate and moisture improve
and ending with the first few stunted trees in favorable
sites. Boreal forest will mark the fourth stage in the
process, which will likely end with the establishment of
temperate ecosystems on Mars. The nature of the
temperate ecosystem will depend on the amount of
moisture available. If moisture is low, a desert may
develop, but if moisture is higher, grasslands and
temperate forests may be possible. The climate of
temperate ecosystems can support a martian agriculture
on the surface.
Each of these ecosystems is a stage in the process of
planetary ecosynthesis. Each stage may be characterized
by certain physical parameters, such as oxygen levels and
UV radiation, and by the dominant organisms, such as
microorganisms or flowering plants. Each stage is part of
a process that may extend over 1,000 years or more. The
entire process of ecosynthesis can be compared to the
terrestrial process of ecological succession. Consider the
retreat of a glacier in a high latitude environment on
Earth. As the glacier melts back within its valley, new
land is exposed. The first pioneer organisms on this
barren land are microorganisms, lichens, and liverworts.
Their activities add organics to the rocky detritus left by

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 107
TABLE 1. The physical parameters of Mars and Earth
Parameter Mars Earth
Mean distance from Sun 2.28 x 108 1.49 x 10
8
Orbital eccentricity 0.0934 0.0167
Rotation rate 24.62 h 24.0 h
Year 668.59 sols1
365.25 days
Obliquity 25.19° 23.45°
Surface gravity 0.38 g 1.00 g
Mean surface temperature
-60°C
+15°C
Surface temperature range -145°C to 20°C -60°C to 50°C
Insolation (PAR) 860 µmol quanta m
-2 s
-1 2000 µmol quanta m
-2 s
-1
UV radiation >190 nm >300 nm
Atmospheric pressure 5 – 10 mbar 1013 mbar
Atmospheric composition N2 0.189 mbar (2.7%) N2 780 mbar (78%)
O2 0.009 mbar (0.13%) O2 210 mbar (21%)
CO2 6.67 mbar (95.3%) CO2 0.38 mbar (0.038%)
Ar 0.112 mbar (1.6%) Ar 10.13 mbar (1%)
1686.98 Earth days
Data from Carr (1981), Kieffer et al., (1992), and McKay and Marinova, (2001).
the glacier and begin to erode exposed rocks. Soon
grasses and herbs colonize the area and generate a true
soil. Alders add nitrogen to the soil, and finally spruce
trees form a dense forest. Each stage in the succession
prepares the way for the next stage by altering the
environment. Similarly, on Mars each stage in
ecosynthesis alters the environment such that the next
stage becomes possible. The microbial stage will
transform the martian regolith into a true soil and
facilitate the arrival of bryophytes in the second stage. As
climate improves under global engineering, the bryophyte
stage will replace a pure microbial stage in favorable
areas while the microbial stage will spread into areas that
were not previously habitable. The microbial ecosystem
will persist even on a fully terraformed Mars at high
latitudes and elevations. Planetary geography always
plays a strong role in the locations of ecosystems.

J.M. Graham — Ecosynthesis on Mars
108 Gravitational and Space Biology 19(2) August 2006
The rest of this paper will examine each of the stages in
the ecosynthesis process. The initial planetary engineering
stage will be presented in less detail because the
engineering processes and their effects on martian climate
have been given previously (McKay and Marinova,
2001). The physical characteristics of each stage will be
considered, and the organisms present will be presented in
relation to their ability to adapt and thrive under those
physical conditions. Where predictable, the ways that
those organisms will alter the environment of Mars to
prepare for later stages will also be discussed.
THE INITIAL ENGINEERING STAGE
The physical parameters of Mars and Earth are given in
Table 1. The rotation rate (and therefore the day length)
and the obliquity of the two planets are very similar.
Surface gravity and solar radiation are lower on Mars than
on Earth, but the year length and orbital eccentricity are
longer. None of these differences creates a problem for
planetary ecosynthesis. Mars can be transformed into a
habitable planet by altering its atmospheric composition
and density. These alterations will raise the mean surface
temperature, reduce the surface temperature range, and
reduce the level of UV radiation at the surface.
The main planetary engineering techniques for Mars are
the manufacture and release of greenhouse gases such as
perfluorocarbons from materials in the regolith and the
potential use of orbiting solar mirrors to raise the amount
of solar radiation at the planet’s surface. If both methods
are used, planetary engineering will raise the surface
temperature of Mars by increasing the amount of solar
radiation at the surface and the retention of that radiation
through the greenhouse effect. The immediate goal of
both methods is the release into the atmosphere of carbon
dioxide frozen at the poles or absorbed in the regolith.
The released carbon dioxide will increase the atmospheric
pressure and further warm the planet by creating a
runaway greenhouse effect. Estimates of the amount of
this carbon dioxide vary from 2 to 200 kPa (20 to 2000
mbar), but for purposes of planetary ecosynthesis an
intermediate amount of 10 to 40 kPa (100 to 400 mbar)
would be adequate for the early stages. In addition to
raising the surface temperature of Mars, a thicker
atmosphere of carbon dioxide would also retain heat
better over the day-night cycle and therefore reduce the
diurnal temperature range. One objective of the
engineering phase will be to raise the mean surface
temperature into the range of the Dry Valleys of
Antarctica (-20°C).
A denser atmosphere around a warmer planet will also
permit liquid water to be stable on the surface. That Mars
once had abundant liquid water running on its surface has
long been established (Carr, 1981; Squyres, 1989). That it
still has abundant water in its regolith has been recently
shown by the Gamma Ray Spectrometer (GRS) aboard
the Mars Odyssey spacecraft. The GRS found evidence of
abundant hydrogen as water ice within one meter of the
surface below 60° south latitude. The percentages of ice
by mass within this one-meter depth varied from 20 to 50
percent. Abundant water ice was also detected above 60°
north latitude. Water is clearly available for ecosynthesis.
Mars has relatively little oxygen in its atmosphere (0.009
mbar), but the surface is highly oxidized. Many of these
oxidants will likely release molecular oxygen when a
warmer wetter climate is established. The net amount of
this oxygen released to the atmosphere would be slight,
probably less than 2 mbar, but the fact that the surface is
oxidized rather than reduced simplifies the ecosynthesis
process. Oxygen produced by photosynthetic
microorganisms can enter the atmosphere to raise the
levels of oxygen and ozone rather than rusting reduced
metals as occurred on the early Earth.
As the initial planetary engineering stage draws to a close,
the atmospheric pressure on Mars will reach 10 to 40 kPa.
The atmosphere is largely composed of carbon dioxide
with small amounts of nitrogen, argon, water vapor, and
oxygen. Average surface temperatures over some areas of
the planet reach –20°C and the daily temperature range is
reduced. The same greenhouse gases that have warmed
the planet also provide a shield from UV radiation. Liquid
water is now stable on the surface. Water will be cycling
through the atmosphere and returning to the surface as
some form of precipitation. It will likely collect in basins
and craters and may resume flowing down ancient
channels during summers. The stage is set for the
introduction of the first ecosystem.
Figure 2. A Dry Valley in Antarctica. The Antarctic Dry Valleys
are the terrestrial analogue of the first ecosystem on Mars.
Photo by Joby Chesnick.
THE MICROBIAL ECOSYSTEM
It is not possible to discuss all possible microorganisms
that would play some role in the first ecosystem on Mars.
This discussion will therefore focus on four groups of
organisms: the bacteria involved in a nitrogen cycle,
cyanobacteria, green algae, and lichens, which are
symbiotic associations between fungi and algae. These
four groups of microorganisms occur in polar deserts such
as the Dry Valleys of Antarctic (Figure 2) and will play a
major role in the first ecosystem on Mars. Other groups of
algae may occur, but they will be restricted to aquatic
habitats and therefore less widespread initially.

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 109
Figure 3. The nitrogen cycle on Mars. Initially the nitrogen cycle will be dominated by denitrification of regolith nitrate and nitrite into
molecular nitrogen (N2) and the assimilation of nitrate and nitrite by microbial reductases into ammonia and amino acids. Nitrogen will
cycle among microorganisms through ammonia and amino acids. As the nitrogen level of the atmosphere increases, nitrogen fixation will
begin at a pN2 of about 5 mbar.
Bacteria and the nitrogen cycle
In the present atmosphere of Mars, nitrogen is present at
about 2.7% (0.16 mbar), but Mars should have outgassed
some 300 mbar of nitrogen. If this nitrogen gas were
present initially, it was probably oxidized to nitrate by
lightning and volcanic electric discharges and is now in
the regolith. If nitrate is in the regolith, it is possible to
propose an order in which a nitrogen cycle might be
established. The initial processes will be essentially
anaerobic and will utilize nitrate and nitrite in the regolith
via two main pathways (Figure 3). Denitrification is
carried out by a variety of bacteria that convert nitrate to
nitrite, nitrite to nitric oxide, nitric oxide to nitrous oxide
and finally nitrous oxide to nitrogen gas. This process
could raise the partial pressure of N2 (pN2) in the
atmosphere to 60 to 300 mbar. Experiments with
denitrifying bacteria in a CO2 atmosphere indicate that
nitrate could be rapidly converted to N2 if liquid water
and organic substrates were present (Hart et al., 2000).
The microorganisms performing the second pathway
(assimilatory nitrate reduction) would provide the organic
substrates. Many microorganisms, including algae and
cyanobacteria, can perform assimilatory nitrate reduction
using nitrate and nitrite reductases. Nitrate is reduced to
ammonia that is incorporated into amino acids within
living organisms. When these organisms die and decay,
they release ammonia and amino acids into the
environment where other microbes can take them up by
ammonia assimilation. Ammonia thus cycles through a
closed loop. These two pathways will reduce nitrate in the
regolith as the pN2 in the atmosphere rises.
The remaining process shown in Figure 3 is nitrogen
fixation. Prokaryotic microorganisms such as archaeans,
bacteria, and cyanobacteria can fix nitrogen, but the
present atmospheric levels on Mars (0.16 mbar) are too
low for fixation. Klinger et al. (1989) examined
Azotobacter and Azomonas over a range of pressures of
N2. No growth occurred at a pN2 less than 1 mbar, but
both bacteria grew and fixed nitrogen at a pN2 of 5 mbar.
Limited data suggest that nitrogen fixation could begin on
Mars once the pN2 reaches at least 5 mbar. The
establishment of a nitrogen cycle on Mars will profoundly
change the composition of the atmosphere from one
composed of mostly CO2 at a pressure of around 90-400
mbar to one with an additional 60-300 mbar of N2.

J.M. Graham — Ecosynthesis on Mars
110 Gravitational and Space Biology 19(2) August 2006
TABLE 2. Mechanisms of ultraviolet radiation resistance in cyanobacteria
Category Mechanism
Avoidance Growth in fissures, under the surfaces of porous rocks (endolithic), beneath translucent
rocks, under water, and under the ice on ice-covered lakes.
Screening Scytonemin, yellow-brown sheath pigment
Phycoerythrin, red intracellular pigment
Mycosporine-like amino acids (MAA), colorless
Quenching Carotenoids, orange pigments
Xanthophylls, yellow pigments
Superoxide dismutase
Repair Enzyme-based DNA repair of UV damage
Polyploid genomes, up to 10 copies
Based on data in Vincent and Quesada (1994) and D. Wynn-Williams (1994)
Cyanobacteria
Cyanobacteria are widespread members of the lake,
stream, soil and lithic communities in Antarctica,
especially in the Dry Valleys (Vincent, 1988). They do
not require oxygen and can carry out both oxygenic
photosynthesis and anoxygenic photosynthesis using
hydrogen sulfide as proton donor. They are highly
resistant to freezing and drying. Filamentous
cyanobacteria in Dry Valley streams remain freeze-dried
and exposed for months until the few brief weeks in
austral summer when the streams flow with water
(Vincent and Howard-Williams, 1986).
Their resistance to UV radiation is varied and versatile
(Table 2). They occupy a number of habitats that permit
them to avoid exposure to full UV radiation. Some
cyanobacteria also occur as epilithic crusts, directly on
rock surfaces, at high latitudes and altitudes. Screening
pigments and quenching agents heavily protect these
filaments.
Phycoerythrin and mycosporine-like amino acids (MAA)
are both intracellular compounds that act to screen out
UV radiation in cyanobacteria. Scytonemin is an UV
screening pigment found in the outer sheaths that
surround cyanobacteria (Vincent and Quesada, 1994). The
level of scytonemin in cyanobacteria increases with
increasing light levels (Garcia-Pichel and Castenholz,
1991). Carotenoids and xanthophylls are quenching
agents that dissipate excess solar energy absorbed by
chlorophyll that would otherwise result in damaging free
oxygen radicals and hydrogen peroxide within cells.
Superoxide dismutase scavenges free oxygen radicals (O2-
) and converts them to hydrogen peroxide. The hydrogen
peroxide is then converted to water and oxygen. In
Antarctica mats of cyanobacteria typically contain high
concentrations of these substances as photochemical
protection (Vincent and Quesada, 1994).
As a final
defense against UV radiation, cyanobacteria have
mechanisms for repair of damage to their DNA (Levine
and Thiel, 1987) and are polyploids, meaning they have
multiple copies (as high as 10) of their genome. Damage
to a single strand of DNA is therefore less likely to
disrupt the cell. All these UV radiation adaptations to the
harsh environment of Antarctica preadapt cyanobacteria
to fill a wide range of habitats in the first stage of the
ecosynthesis.
Green algae
Many green algae are not obligate aerobes, including
members of such common genera as Chlorella,
Chlamydomonas, Scenedesmus and Selenastrum (Spruit,
1962). There is one genus of green algae called
Pyrobotrys found only in anaerobic soils (Nozaki, 1986).
In the dark these algae will initially carry out fermentation
and produce carbon dioxide and organic acids, as do

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 111
many plants. But after some time they will evolve
hydrogen gas. If anaerobic conditions persist in the light
and levels of carbon dioxide and hydrogen are adequate,
these algae may reduce carbon dioxide to simple
carbohydrates using molecular hydrogen as electron and
proton donor. Thus green algae possess a more diverse
range of metabolic processes than just photosynthesis and
aerobic respiration. Certain species have the ability to
grow under low pH levels (Sheath et al., 1982), low
temperatures, and high salinity (Vincent, 1988). The
major adaptation of Antarctic green algae is the ability to
survive repeated freezing and thawing (Holm-Hansen,
1963).
The ability to withstand repeated freeze-thaw
cycles would have considerable value during early
planetary ecosynthesis on Mars.
Green algae may be as resistant as cyanobacteria to UV
radiation. Green algae share the same range of habitats as
cyanobacteria in Antarctica (Vincent, 1988). In the Dry
Valleys of Antarctica, cryptoendolithic lichens often
occur as a layer beneath the surfaces of translucent porous
rocks, and the green alga Trebouxia is often the algal
symbiont in the lichen. Beneath this lichen layer the green
alga Hemichloris, which is endemic to Antarctica, may
form another distinct layer. Flavonoids and phenolics are
UV screening agents known to be present in some green
algae. The level of quenching agents such as carotenoids
in green algae is related to the level of exposure to direct
sunlight and UV radiation (Thomas and Duval, 1995).
Lichens
Lichens are generally slow growing, but they grow faster
under elevated carbon dioxide levels, at least up to about
2 mbar of carbon dioxide (Nash et al., 1983). It is
possible therefore that lichens would perform better on
Mars during early stages of ecosynthesis than they do on
Earth because the atmosphere would be largely carbon
dioxide. Antarctic lichens are extremely tolerant of
desiccation and cold. Lange and Kappen (1972) found that
several Antarctic lichens could survive and recover even
after being cooled to –196°C, a temperature below levels
that currently occur at the martian poles. Some lichens
were able to assimilate carbon dioxide at temperatures as
low as –12.5°C to –18°C.
Lichens are extremely resistant to UV radiation. Siegel
and Daly (1968) exposed the arctic lichen Cladonia
rangiferina to UV radiation of 3.6 x 109 ergs
.cm
-2 over 24
hours. This is equivalent to 4.16 x 104 ergs
.cm
-2.s
-1. By
comparison the UV radiation at the equator of Mars
averages 7 x 103 ergs
.cm
-2.s
-1. Cladonia rangiferina covers
large areas of the arctic and presumably could also do so
on Mars. Table 3 lists some of the UV radiation
resistance mechanisms in lichens. Endolithic lichens lie
under the surfaces of translucent porous rocks in the
Antarctic Dry Valleys. The overlying rock layers shield
the lichens from direct solar and UV radiation. Many
lichens, however, grow as crusts on rocks where they may
be colored black, brown, orange or yellow. The black and
brown colors are due to melanins that screen the lichens
from excess solar and UV radiation and also absorb heat
to raise the temperature of the lichen above that of the
surrounding air. The orange and yellow colors are due to
carotenoids that are quenching agents.
TABLE 3. Mechanisms of UV resistance in lichens and mosses
Category Mechanism
Avoidance Endolithic lichens
Screening Lichens: black melanins in endolithic and surface lichens
Mosses: black and brown pigments and flavonoids.
Phenolics in the walls of the moss Andreaea.
Quenching Lichens: carotenoids
Mosses: carotenoids such as violaxanthin (purple)
Repair Mosses: polyploid genomes
Based on data in Post (1990) and D. Wynn-Williams (1994).

J.M. Graham — Ecosynthesis on Mars
112 Gravitational and Space Biology 19(2) August 2006
Although lichens are slow growing on Earth, their role in
the ecological process of succession makes them
important to include in the early stages of ecosynthesis on
Mars. On Earth lichens are the first pioneer species to
colonize bare rock. They excrete organic acids that slowly
dissolve rock and free minerals to join the accumulation
of organic material that slowly creates a true soil. Their
growth opens the way for the establishment of other
plants.
The microbial stage of ecosynthesis will begin the process
of adding organic matter to the martian regolith, binding
together regolith particles, and transforming the
atmosphere by increasing the amount of free oxygen and
converting nitrates to free nitrogen. The climate will
begin as frigid-polar with average temperatures in the
warmest “month” less than 0°C (Longton, 1988). On
Earth a frigid-polar climate is found only in continental
Antarctica. Toward the end of the first stage, the climate
should warm to cold-polar where the average temperature
in the warmest month falls in the range of 0° to 2°C.
Maritime Antarctica and the arctic polar deserts have a
terrestrial cold-polar climate. Microbial life will thrive in
the regolith, on the surfaces of rocks, under the surfaces
of rocks in the pores between mineral grains, and in
gathering surface waters as plankton and as filaments
attached to surfaces.
THE BRYOPHYTE ECOSYSTEM
Bryophytes consist of relatively simple, usually small,
green plants called mosses, liverworts, and hornworts.
They lack roots but form cellular extensions called
rhizoids for attachment. Bryophytes are extremely
important in terrestrial polar and alpine ecosystems where
the severe climate prohibits most flowering plants (Figure
4).
Figure 4. Alpine tundra. The terrestrial equivalent of the second
stage in ecosynthesis on Mars. Photo by Lee Wilcox.
In the bryophyte stage of ecosynthesis the atmosphere
will be mainly carbon dioxide and nitrogen with small but
increasing amounts of oxygen. Mosses show an unusual
capacity to utilize carbon dioxide at high external levels,
far more so than flowering plants. Silvola (1990) reported
that Sphagnum fuscum, a widespread moss in northern
peatlands, exhibited increased rates of photosynthesis up
to a carbon dioxide pressure of 9 mbar, the highest level
tested. Grimmia antarctici, a common carpet-forming
moss in East Antarctica, reached its maximum rates of
photosynthesis at CO2 levels of 10-20 mbar (Tarnawski
et al., 1992). Mosses photosynthesize more slowly than
flowering plants at terrestrial ambient carbon dioxide
levels of 0.38 mbar. At higher levels of CO2 the
photosynthetic rates of mosses approach those of
flowering plants. Mosses are also capable of carrying out
photosynthesis at lower light levels and lower
temperatures, even under snow, than flowering plants
(Oechel and Sveinbjornsson, 1978; Collins and
Callaghan, 1980). Mosses do require oxygen for aerobic
respiration, but how much oxygen they require is
unknown. The moss Hypnum cupressiforme showed an
increase in photosynthetic rate as oxygen was reduced to
3% (30 mbar) (Aro et al., 1984).
Mosses are not thought to have a high degree of resistance
to UV radiation, but this may be due to lack of
experimental data. Mosses occur in the same habitats as
lichens and cyanobacteria, including on the surfaces of
rocks exposed to full solar radiation at polar latitudes and
alpine locations. Like lichens, mosses in polar and alpine
environments may be dark brown or black in color. These
pigments screen out harmful UV radiation and excess
visible light while warming the body of the moss (Table
3). Phenolics such as flavonoids occur in the walls of a
number of mosses, and these phenolics act to screen out
UV radiation and possibly also reduce desiccation
(Graham et al., 2003). Markham
et al. (1990) reported
UV protective flavonoids in the Antarctic moss Bryum
argenteum. The lower the level of ozone over the field
sites the higher the level of protective flavonoids. Mosses
also possess quenching agents such as carotenoids. The
Antarctic moss Ceratodon purpureus produces the
carotenoid violaxanthin as a photoprotective pigment
(Post, 1990). Mosses are generally polyploid.
Mosses will likely play an important role in planetary
ecosynthesis by sequestering large amounts of carbon
dioxide in the form of decay resistant organic compounds
in martian peatlands. At the same time these peatlands
will add large quantities of oxygen to the atmosphere of
Mars. Martyn Fogg (1995) estimated that a cover of
peatlands on Mars could produce 20 mbar of atmospheric
oxygen in about 700 years. Since mosses photosynthesize
faster in a CO2 atmosphere than on Earth, martian
peatlands might be able to produce this amount of oxygen
in less time, depending on the degree of surface cover and
moisture availability. For bryophytes to sequester large
amounts of carbon in peatlands, planetary engineering
must raise the climate of significant areas on Mars to the
level of mild-polar environments on Earth (Longton,
1988). Mild-polar habitats have average monthly
temperatures in the warmest month of 7° to 12°C. On
Earth they occur in Alaska and northern mainland
Canada. These conditions permit Sphagnum moss to form
the extensive peatlands that sequester so much carbon on
Earth.

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 113
With the full development of the bryophyte ecosystem,
the surface of Mars will appear vastly different from that
shown in the cameras of Spirit and Opportunity. Rocks
will be dotted with lichens and mosses. In areas where
water collects, Sphagnum moss may form extensive
peatlands. Mars will assume a distinctly green aspect
interrupted by bright spots of yellow, orange, and red
lichens. At some point in this stage the partial pressure of
oxygen will surpass 20 mbar, a value that is significant
for the next stage in planetary ecosynthesis.
THE FLOWERING HERBS ECOSYSTEM
The crucial factor in determining the transition to the third
stage in martian ecosynthesis, which is characterized by
the arrival of flowering plants, is the level of atmospheric
oxygen. In most terrestrial environments only two types
of flowering plant structures risk low oxygen levels: seeds
and underground organs such as roots, rhizomes and
tubers (Crawford, 1992). Rhizomes and tubers are
underground stems. Many seeds show reduced
germination if oxygen falls below 10 kPa or 100 mbar.
On Earth oxygen always limits to some extent the depth
to which roots can penetrate. Armstrong and Gaynard
(1976) found that 20 to 25 mbar was the critical oxygen
pressure to maintain root respiration in rice (Oryza sativa)
and the aquatic sedge Eriophorum augustifolium. Fogg
(1995) used this data when he proposed that the minimum
pO2 for plants would lie around 20 mbar or 2% O2 in the
martian atmosphere. Higher values, however, have been
reported for other plant species. Corn (Zea mays) had a
critical oxygen pressure of 6kPa (6% or 60 mbar) (Saglio
et al., 1984). A third way in which low oxygen can
constrain flowering plants is through restricting animal
pollinators. Many plants require animal pollinators to
produce seeds. Hard data are lacking but a pO2 of 20 mbar
is unlikely to support pollinators such as flying insects
and certainly not bats and birds. Cockell et al. (1999)
examined a number of insects from different orders and
found that all species seemed to function normally down
to a pressure of 200 mbar (about 40 mbar of oxygen) for
up to 24 h. Low oxygen levels will likely prohibit certain
plants from colonizing Mars until oxygen levels rise
enough to permit their animal pollinators to thrive also.
The first flowering plants on Mars will have anoxia-
tolerant seeds and underground organs and mechanisms
for reproduction that do not require animals.
Seeds, rhizomes, and whole plants under low oxygen
Seeds may store food reserves as lipids, proteins or starch.
The germination rate of seeds that store lipids or proteins
begins to decline if external oxygen levels fall below
10kPa or 100 mbar. Starch seeds, however, can maintain
50% germination rates when the pO2 drops below 1kPa or
10 mbar and can germinate at about 20% even at 0.1kPa
(1 mbar). Rice is an aquatic monocot, and its seeds can
still germinate when pO2 falls below 0.01kPa (0.1 mbar)
and even extend their shoots above water (Al-Ani et al.,
1985). Starch fermentation powers germination.
Although roots cannot survive more than a few days of
anoxia, the underground rhizomes of a number of aquatic
plants can survive under total anoxia for two to three
months (Crawford 1992). Survival depends on a large
reserve of carbohydrates that the rhizome ferments into
ethanol to generate energy in the form of adenosine
triphosphate (ATP). The ATP from fermentation
maintains the rhizome in winter and also powers the
extension of a new shoot in spring (Braendle and
Crawford, 1987). Once the shoot has elongated and the
leaves expanded, aquatic plants develop another
mechanism to thrive in an oxygen-poor environment–a
special tissue called arenchyma. Aerenchyma consists of
interconnected gas-filled channels through stems and
roots that provide a pathway for the movement of oxygen,
carbon dioxide, and ethanol. Aquatic plants are also
tolerant of high levels of carbon dioxide. A number of
aquatic plants should be good candidates for colonizing
Mars early in the third stage.
One other group of flowering plants possesses
considerable tolerance of anoxia. A number of vascular
plants from Spitsbergen in the High Arctic have an
exceptionally high level of anoxia tolerance (Crawford et
al., 1994). This anoxia tolerance extends to the entire
plant including the leaves. The anoxia tolerance of these
high arctic species appears to be an adaptation to ice-
encasement, which may last as long as 8 to 10 months.
The extraordinary anoxia tolerance of these plants makes
them prime candidates for colonization on Mars.
Reproduction without animals at low oxygen
Flowering plants have a number of reproductive
mechanisms that do not require animal pollinators. These
mechanisms include vegetative reproduction, apomixis,
autogamy, and anemophily. In vegetative reproduction
certain somatic (body) cells divide to produce structures
called runners, rhizomes, or bulbils. Runners are
aboveground stems that spread out from the parent plant
and establish new roots and leaves. Strawberry plants are
a familiar example of a plant that spreads by runners.
Rhizomes are underground stems that spread out from the
parent plant. The crab grass in your lawn and the Iris in
your garden both spread by rhizomes. The final type of
vegetative reproduction is less familiar. Bulbils are small,
bud-like plants born on the parent plant. The common
houseplant Kalanchoae produces bulbils along the
margins of its leaves, from which the bulbils drop off and
root in the ground.
Apomixis is asexual seed formation, but there are a
number of different types of apomixis (Asker and Jerling,
1992). The type most suitable for colonizing Mars is
called autonomous apomixis. Autonomous apomictic
plants reproduce independently of pollination. The
familiar lawn and garden dandelion (Taraxacum
officinale) is an example of a common autonomous
apomictic flowering plant. The flowering plant family
Asteraceae, which contains daisies, sunflowers, and

J.M. Graham — Ecosynthesis on Mars
114 Gravitational and Space Biology 19(2) August 2006
TABLE 4. Flowering plants growing in the cool-polar regions of Earth that reproduce
without animal pollinators.
Genera and species Common name Reproductive mechanism(s)
Cassiope tetragona Arctic bell heather Autogamous
Dryas integrifolia Mountain avens Autogamous
Epilobium latifolium Dwarf fireweed Autogamous
Erysimum pallasi Apomictic
Lesquerella arctica Autogamous
Papaver radicatum Arctic poppy Autogamous
Pedicularis arctica
P. hirsute
P. lapponica
P. sudetica
Arctic lousewort
Hairy lousewort
Lapland lousewort
Sudetan lousewort
Autogamous
Autogamous
Autogamous/vegetative
Autogamous
Potentilla nivea Snow rose Autogamous/apomictic
Saxifraga tricuspidata
S. flagellaris
S. cernua
Three-pointed saxifrage
Whip saxifrage
Nodding saxifrage
Autogamous
Apomictic/vegetative
Apomictic/vegetative
Silene acaulis Cushion pink Autogamous
Taraxacum arctogenum Arctic dandelion Apomictic
Some may use insects if available. Based on data from Kevan (1972) and Williams
and Batzli (1982).
dandelions, has many autonomous apomictic species and
therefore many potential martian colonists (Graham,
2003).
Autogamy means literally self-marriage. An autogamous
plant acquires pollen for fertilization of its egg cells from
itself. Autogamy is a form of sexual reproduction, and it
is widespread among flowering plant families.
Anemophily (wind loving) or wind pollination is normal
sexual cross-pollination where the agent transporting the
pollen is wind rather than animals. Wind pollination
dominates the gymnosperms and prevails in the flowering
plant families Poaceae (grasses), Cyperaceae (sedges),
and Juncaceae (reeds)(Faegri and van der Pijl, 1979).
These three families dominate terrestrial wetlands. Wind-
pollinated plants form a fourth group of potential martian
colonists.
These alternative mechanisms of reproduction occur in
many different groups of flowering plants and in many
environments, but the flowering plants of polar and alpine
environments utilize them to an extreme degree. In these
harsh environments populations of animal pollinators are
often severely restricted by low temperatures, and
population size may vary widely from year to year. If
animal pollinators are scarce and unreliable, then
reproduction is best insured by having some mechanism
that is not dependent on them. The maritime Antarctic is a
cold-polar environment where the mean temperature in
the warmest month is only 0-2°C (Longton, 1988). There
are only two native species of flowering plants: the grass
Deschampsia antarctica and the carnation Colobanthus
quitensis. Colobanthus is autogamous and Deschampsia
is apomictic.
Cool-polar environments, such as occur in Greenland and
the Canadian Islands in the High Arctic, and isolated

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 115
islands such as South Georgia and Macquarie around
Antarctica, are the first polar environments with
significant flowering plant communities. In a cool-polar
environment the average temperature of the warmest
month is 3° to 7°C. Table 4 lists a number of cool-polar
plants that are known to be capable of reproducing
without animal pollinators. Dryas integrifolia has roots
that harbor nitrogen-fixing bacteria. The whip saxifrage
(Saxifraga flagellaris) has yellow apomictic flowers,
which form seeds for long-distance dispersal, and long
whip-like runners to spread locally. During the stage of
flowering herbs the level of atmospheric oxygen should
continue to rise and exceed 60 mbar. Beyond this point
the level of oxygen should no longer be a significant
constraint on the subsequent ecosystems on Mars. The
remaining constraints will be the same as those that
operate on Earth, namely temperature and moisture. The
next stage begins when the climate of parts of the martian
surface reaches that of a boreal ecosystem.
THE BOREAL FOREST ECOSYSTEM
During the later part of the third stage of ecosynthesis,
sheltered sites insulated by martian winter snows could
harbor dwarf trees of birch (Betula) and willow (Salix) up
to one meter tall and stands of open, stunted trees
consisting of birches, larches (Larix) and spruces (Abies)
could be common in sheltered valleys where water flows.
But the development of true forests on Mars will first
occur when the climate of some regions reaches that of
the terrestrial taiga or boreal coniferous forest (Figure 5).
The climate of the boreal forest can be defined by the
average temperature of the warmest month of the year,
which is 13 to 18°C in July (Elliott-Fisk, 2000).
Alternatively, the boreal climate can be defined by the
length of its growing season, which on Earth is 30 to 120
days with a daily mean temperature above 10°C (Walter,
1973). The lower figure represents the northern boundary
of the boreal forest and the higher figure the southern
boundary. On Mars these numbers should be revised
upward to 54 and 218 sols to account for the longer
seasons. On Earth annual precipitation in a boreal forest
varies from 30 to 50 cm (Elliott-Fisk, 2000). The revised
values for Mars would be 55 to 90 cm per martian year.
These temperature and precipitation values define the
requirements for establishing a boreal coniferous forest on
Mars.
On Mars as on Earth a boreal coniferous forest is a
spruce–fir forest. White spruce (Picea glauca), black
spruce (Picea mariana) and balsam fir (Abies balsamea)
are the most widespread North American dominants.
Black spruce and tamarack (Larix laricina), a deciduous
conifer, are common in bogs. Jack pine (Pinus banksiana)
and two deciduous flowering plants, paper birch (Betula
papyrifera) and quaking aspen (Populus tremuloides)
dominate disturbed areas (Vankat, 1979). The deciduous
flowering trees are wind pollinated. The seeds of all three
disturbance-site species are adapted for wind dispersal, a
feature that will aid their spread across the surface of
Mars. Willows, dwarf birch (Betula glandulosa), green
alder (Alnus crisps), and members of the heath family
(Ericaceae) make up most of the shrubs in the boreal
coniferous forest. The understory layer is dominated
either by mosses in wet areas or lichens in drier
environments. Bogs dominated by Sphagnum moss are
the most common form of wetland within the boreal
coniferous forest zone.
Figure 5. Boreal forest. The terrestrial analogue of the fourth
stage in martian ecosynthesis. Photo by the author.
The North American boreal coniferous forest is the most
diverse boreal forest on Earth, but its diversity is still the
lowest of any forest in North America. For additional
conifer species to use in ecosynthesis, the best sources are
montane coniferous forests. Montane coniferous forests
occur in the Appalachian Mountains, the Rocky
Mountains, and the Sierra Nevada and Cascade
Mountains of North America. Another potential source of
martian forests might be the high altitude montane forests
of the tropics. With a timberline at or above 4000 m in
Mexico (Beaman, 1962; Lauer, 1978), these montane
forests are adapted to grow and reproduce at lower
atmospheric pressures, higher levels of radiation, and
lower levels of moisture than sea-level forest species.
TEMPERATE ECOSYSTEMS
Temperate ecosystems represent the culmination of
planetary ecosynthesis on Mars. On Earth the growing
season ranges from 120 days to 250 days with mean daily
temperatures above 10°C. The amount of annual

J.M. Graham — Ecosynthesis on Mars
116 Gravitational and Space Biology 19(2) August 2006
precipitation determines whether a temperate ecosystem is
a forest, grassland, desert grassland, or desert. The eastern
temperate deciduous forest of North America varies in
annual precipitation from a maximum of 150 cm along
the Gulf Coast to a minimum of 75 cm around the Great
Lakes (Vankat, 1979). Annual precipitation in temperate
grasslands varies from less than 75 cm to 25 cm, where
desert grasslands may occur. Deserts generally receive
less than 20 cm in precipitation per year. On Mars these
temperate zone figures would be a growing season of 220
to 455 sols. A martian deciduous forest would receive
from 135 to 275 cm in annual precipitation, grassland
from 45 to 135 cm, and desert less than 45 cm in
precipitation per year. Temperate ecosystems would
permit a widespread open surface agriculture on Mars,
just as they do on Earth.
Because the type of temperate ecosystem depends so
strongly on the amount of annual moisture and on Mars
moisture levels are likely to be lower initially in a stage,
the first temperate systems to consider are arid ones,
deserts and desert grasslands. A good terrestrial analogue
of a martian temperate desert would be the Great Basin
Desert. The soil is low in organic matter and high in
salinity. A sagebrush (Artemisia tridentata) community
dominates lower salinity soils, and a shadscale (Atriplex
tridentata) community occurs on higher salinity soils.
Sagebrush is apomictic, and shadscale is autogamous.
Grasses such as grama grass (Bouteloua spp.) and wire
grass (Aristidia spp.) dominate desert grasslands. The
grasses are all wind-pollinated. Both arid systems should
be capable of colonizing Mars.
TABLE 5. Genera of wind-pollinated flowering trees north of Mexico.
Genus Common name Genus Common name
Ulmus Elm Lithocarpus Tanoak
Fagus Beech Celtis Hackberry
Quercus Oak Morus Mulberry
Corylus Hazel Maclura Osage orange
Alnus Alder Planera Planertree
Betula Birch Trema Trema
Juglans Walnut Platanus Sycamore
Carya Hickory Liquidambar Sweetgum
Fraxinus Ash Populus Poplar
Castanea Chesnut Ostrya Hophornbeam
Castanopsis Chinkapin Carpinus Hornbeam
When the genera of conifers are added to the above list, the groups represent 240 species
or 30% of 787 native trees in the region. Data from Regal (1982).
Grasslands are especially important to establish on Mars
because they generate some of the most fertile soils for
agriculture on Earth. Grasses are wind-pollinated but they
also reproduce by vegetative growth, spreading by
underground stems (rhizomes) or above ground stems
(runners) to form mats. Grasslands also include many
species from the sunflower (Asteraceae) and pea families
(Fabaceae) that contain numerous apomictic and
autogamous species.
Temperate deciduous forests can also generate good to
excellent soils for agriculture as well as timber for wood
and paper products. Many of the genera of trees that
dominate the temperate deciduous forest are wind-
pollinated (Table 5). The genera listed include some 240
species or about 30% of the 787 native species of trees in
the region (Regal, 1982). A temperate deciduous forest
contains five vertical strata: two tree layers, a shrub
stratum, herb layer and surface layer. The surface layer

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 117
includes lichens, mosses and liverworts. The herb layer in
cool, wet forests may have a number of ferns and species
of Lycopodium (ground pine). None of these plants
requires animal pollinators. The flowering plants of the
herb layer contain many species that are autogamous or
apomictic. The rose family (Rosaceae) contains a number
of genera and species of shrubs that are apomictic,
including Crataegus (hawthorn), Cotoneaster, Rubus
(blackberries) and Sorbus. Some species of Sorbus are
subcanopy trees. It should be possible to assemble a
temperate deciduous forest on Mars using species that do
not require animal pollinators.
The establishment of temperate ecosystems on Mars
permits the eventual development of agriculture on the
open surface, even if animal pollinators are still restricted
by low levels of oxygen. All our major cereal crops such
as wheat, oats, barley, corn, rice, and rye are wind-
pollinated cereal grasses. Many other crops, such as
tomatoes, some potatoes, beans, lettuce, and strawberries
are autogamous. Agriculture on the open surface would
permit a large, self-sufficient martian civilization.
DISCUSSION
The data used to develop this manuscript came from basic
research on biological phenomena that were not directly
related to astrobiology. The parameters of temperature
and moisture that define different types of ecosystems and
the zonation of ecosystems along an altitudinal gradient
are a long established part of the ecological literature. The
discovery of an ozone hole over Antarctica prompted
research on the effects of UV radiation on various
organisms, particularly microorganisms. This research
was very important in assessing organisms for planetary
ecosynthesis. The effects of elevated levels of carbon
dioxide on growth of photosynthetic microbes and plants
had been largely unexamined until rising levels of CO2
were detected in the terrestrial atmosphere. Most studies,
however, focused on levels of CO2 less than 5 mbar. It is
still not known how well plants can adapt to higher levels
of carbon dioxide, as occurred on the early Earth or will
occur during planetary ecosynthesis. Relatively little is
known about the minimum oxygen requirements of many
terrestrial plants, especially bryophytes, yet this
information is crucial to the transitions to both the second
and third stages in the ecosynthesis process. The
discovery of anoxia tolerant plants on the island of
Spitsbergen was especially important to the arrival of
flowering plants on Mars (Crawford et al., 1994). Even
less is known about the oxygen requirements of terrestrial
insects that might act as pollinators in transplanted
ecosystems. If some insects can function as pollinators at
low ambient oxygen levels, then the dynamic process of
planetary ecosynthesis would be altered considerably.
New discoveries will continue to be made, but until there
is actual funding for ecosynthesis research, these
discoveries will be made for other reasons than their
significance to astrobiology.
This manuscript has attempted to describe the process of
establishing functional ecosystems containing
communities of terrestrial organisms on the surface of
Mars. The process is likely to take from 500 to 1000 years
to approach the final stage. Given the vast scope of the
process, it has been necessary to describe it in somewhat
broad terms. Although the data upon which this
description is based are incomplete, the broad picture
should be correct. Planetary ecosynthesis on Mars will be
a long and great adventure. It will yield a vast amount of
new knowledge and become one of the great acts of
positive creative energy by the human species.
ACKNOWLEDGMENT
I wish to thank Kandis Elliot, the staff artist at the
Department of Botany, UW-Madison, for preparing the
illustrations for this manuscript and for her interest in
astrobiology and planetary ecosynthesis.
REFERENCES
Al-Ani, A., Bruzau, F., Raymond, P., Saint-Ges, V.,
Leblanc, J.M. and Pradet, A. (1985) Germination,
respiration, and adenylate energy charge of seeds at
various oxygen partial pressures. Plant Physiol. 79, 885-
890.
Armstrong, W. and Gaynard, T.J. (1976) The critical
oxygen partial pressures for respiration in intact plants.
Physiol. Plant. 37, 200-206.
Aro, E.-M., Gerbaud, A. and André, M. (1984) CO2 and
O2 exchange in two mosses, Hypnum cupressiforme and
Dicranum scoparium. Plant Physiol. 76, 431-435.
Asker, S.E. and Jerling, L. (1992) Apomixis in Plants.
CRC Press, Boca Raton, pp. 49-79.
Beaman, J. H. (1962) The timberlines of Iztaccihuatl and
Popocatepetl, Mexico. Ecology 43, 377-385.
Braendle, R. and Crawford, R.M.M. (1987) Rhizome
anoxia tolerance and habitat specializations in wetland
plants. In Plant Life in Aquatic and Amphibious Habitats,
edited by R.M.M. Crawford, Blackwell, Oxford, pp. 397-
410.
Carr, M.H. (1981) The Surface of Mars. Yale University
Press, New Haven.
Clark, B.C. (1998) Surviving the limits to life at the
surface of Mars. J. Geophys. Res. 103, 28545-28556.
Cockell, C., Catling, D. and Waites, H. (1999) Insects at
low pressure: applications to artificial ecosystems and
implications for global windborne distribution. Life
Support & Biosphere Sci. 6, 161-167.

J.M. Graham — Ecosynthesis on Mars
118 Gravitational and Space Biology 19(2) August 2006
Collins, N.J. and Callaghan, T.V. (1980) Predicted
patterns of photosynthetic production in maritime
Antarctic mosses. Annals of Bot. 45, 601-620.
Crawford, R.M.M. (1992) Oxygen availability as an
ecological limit to plant distribution. Adv. Ecol. Res. 23,
93-185.
Crawford, R.M.M., Chapman, H.M. and Hodge, H.
(1994) Anoxia tolerance in high arctic vegetation. Arctic
and Alpine Res. 26, 308-312.
Elliott-Fisk, D.L. 2000. The taiga and boreal forest. In
North American Terrestrial Vegetation, edited by M.
Barbour and W. Billings, Cambridge University Press,
Cambridge, pp. 41-74.
Faegri, K. and van der Pijl, L. (1979) The Principles of
Pollination Ecology, Pergamon Press, Oxford.
Fogg, M.J. (1995a) Terraforming: Engineering Planetary
Environments, Society of Automotive Engineers,
Warrendale.
Fogg, M.J. (1998) Terraforming Mars: a review of current
research. Adv. Space Res. 22, 415-420.
Garcia-Pichel, F. and Castenholz, R.W. (1991)
Characterization and biological implications of
scytonemin, a cyanobacterial sheath pigment. J. Phycol.
27, 395-409.
Graham, J.M. (2003) Stages in the terraforming of Mars:
the transition to flowering plants. Amer. Instit. Physics
Conf. Proceed. 654, 1284-1291.
Hart, S.D., Currier, P.A. and Thomas, D.J. (2000)
Denitrification by Pseudomonas aeruginosa under
simulated engineered Martian conditions. J.Br.
Interplanet. Soc. 53, 357-359.
Head, J.W. III, Hiesinger, H., Ivanov, M.A., Kreslavsky,
M.A., Pratt, S., and Thomson, B.J. (1999) Possible oceans
on Mars: evidence from Mars Orbiter Laser Altimeter
data. Science 286: 2134-2137.
Holm-Hansen, O. (1963) Viability of blue-green and
green algae after freezing. Physiologia Plantarum 16,
530-539.
Kevan, P.G. (1972) Insect pollination of high arctic
flowers. J. Ecol. 60, 831-847.
Kieffer, H.H., Jakosky, B.M. and Snyder, C.W. (1992)
The planet Mars from antiquity to the present. In Mars,
edited by H.H. Kieffer et al., University of Arizona Press,
Tucson, pp. 1-33.
Klinger, J.M., Mancinelli, R.L. and White, M.R. (1989)
Biological nitrogen fixation under primordial martian
partial pressures of dinitrogen. Adv. Space Res. 9, 173-
176.
Lange, O.L. and Kappen, L. (1972) Photosynthesis of
lichens from Antarctica. In Antarctic Terrestrial Biology,
edited by G.A. Llano, Antarctic Research Series 20, 83-
95.
Lauer, W. (1978) Timberline studies in central Mexico.
Arct. Alp. Res. 10, 383-396.
Levine, F. and Thiel, T. 1987. UV-inducible DNA repair
in the cyanobacteria Anabaena spp. J. Bacteriol. 169,
3988-3993.
Longton, R.E. (1988) The Biology of Polar Bryophytes
and Lichens. Cambridge University Press, Cambridge.
Markham, K.R., Franke, A., Given, D.R. and Brownsey,
P. (1990) Historical Antarctic ozone level trends from
herbarium specimen flavonoids. Bull. Liason Groupe
Polyphenols 15, 230-235.
McKay, C.P. (1997) The search for life on Mars. Origins
Life Evolution Biosphere 27, 263-289.
McKay, C.P. (1999) Bringing life to Mars. Scientific
American Presents 10: 52-57.
McKay, C.P. and Marinova, M.M. (2001). The physics,
biology and environmental ethics of making Mars
habitable. Astrobiology 1, 89-109.
McKay, C.P., Toon, O.B., and Kasting, J.F. (1991)
Making Mars habitable. Nature 352, 489-496.
Nash, T.H., Moser, T.J., Link, S.O., Ross, L.J., Olafsen,
A. and Matthes, U. (1983) Lichen photosynthesis in
relation to CO2 concentration. Oecologia 58, 52-56.
Nozaki, H. (1986) A taxonomic study of Pyrobotrys
(Volvocales, Chlorophyta) in anaerobic pure culture.
Phycologia 25, 455-468.
Oberg, J.E. 1981. New Earths: Restructuring Earth and
Other Planets, The New American Library, New York.
Oechel, W.C. and Sveinbjörnsson, B. (1978) Primary
production processes in arctic bryophytes at Barrow,
Alaska. In Vegetation and Production Ecology of an
Alaskan Arctic Tundra, edited by L.L. Tieszen, Springer,
New York, pp. 269-298.
Post, A. (1990) Photoprotective pigment as an adaptive
strategy in the Antarctic moss Ceratodon purpureus.
Polar Biol. 10, 241-245.
Regal, P.J. (1982) Pollination by wind and animals:
ecology of geographic patterns. Ann. Rev. Ecol. Syst. 13,
497-524.

J.M. Graham — Ecosynthesis on Mars
Gravitational and Space Biology 19(2) August 2006 119
Sagan, C. (1971) The long winter model of Martian
biology: a speculation. Icarus 15, 511-514.
Saglio, P.H., Rancillac, M., Brazan, F. and Pradet, A.
(1984) Critical oxygen pressure for growth and
respiration of excised and intact roots. Plant Physiol. 76,
151-154.
Sheath, R.G., Havas, M., Hellebust, J.A. and Hutchinson,
T.C. (1982) Effects of long-term acidification on the algal
communities of tundra ponds at the Smoking Hills,
N.W.T., Canada. Can. J. Bot. 60, 58-72.
Siegel, S.M. and Daly, O. (1968) Responses of Cladonia
rangiferina to experimental stress factors. Bot. Gaz.
129,339-345.
Silvola, J. (1990) Combined effects of varying water
content and CO2 concentration on photosynthesis in
Sphagnum fuscum. Holarctic Ecol. 13, 224-228.
Spruit, C.J.P. (1962) Photoreduction and anaerobiosis. In
Physiology and Biochemistry of Algae, edited by R.A.
Lewin, Academic Press, New York, pp. 47-60.
Squyres, S.W. (1989) Water on Mars. Icarus 79, 229-288.
Tarnawski, M., Melick, D., Roser, D., Adamson, E.,
Adamson, H. and Seppelt, R. (1992) In situ carbon
dioxide levels in cushion and turf forms of Grimmia
antarctici at Casey Station, East Antarctica. J. Bryol. 17,
241-249.
Thomas, W.H. and Duval, B. (1995) Sierra Nevada,
California, U.S.A., snow algae: snow albedo changes,
algal-bacterial interrelationships, and ultraviolet radiation
effects. Arctic and Alpine Res. 27, 389-399.
Vankat, J.L. (1979) The Natural Vegetation of North
America. John Wiley & Sons, New York.
Vincent, W.F. (1988) Microbial Ecosystems in
Antarctica. Cambridge University Press, Cambridge.
Vincent, W.F. and Howard-Williams, C. (1986) Antarctic
stream ecosystems: physiological ecology of a blue-green
algal epilithon. Freshwater Biology 16, 219-233.
Vincent, W.F. and Quesada, A. (1994) Ultraviolet
radiation effects on cyanobacteria: implications for
Antarctic microbial ecosystems. In Ultraviolet Radiation
in Antarctica: Measurements and Biological Effects.
Antarctic Research Series 62, 111-124.
Walter, H. (1973) Vegetation of the Earth. Springer-
Verlag, New York.
Williams, J.B. and Batzli, G.O. (1982) Pollination and
dispersal of five species of lousewort (Pedicularis) near
Atkasook, Alaska, U.S.A. Arctic and Alpine Res. 14, 59-
74.
Wynn-Williams, D.D. (1994) Potential effects of
ultraviolet radiation on Antarctic primary terrestrial
colonizers: cyanobacteria, algae, and cryptogams. In
Ultraviolet Radiation in Antarctica: Measurements and
Biological Effects. Antarctic Research Series 62, 243-257.

J.M. Graham — Ecosynthesis on Mars
120 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology 19(2) August 2006 121
Short Papers
Advanced Life Support and Biotechnology (page 123)
Animal Development, Physiology and Gravity Response (page 133)
Cell Biology (page 143)
Plant Development and Gravity Response (page 155)

122 Gravitational and Space Biology 19(2) August 2006

Gravitational and Space Biology Bulletin 19(2) August 2006 123
DEVELOPMENT OF A MICROFLUIDIC ION SENSOR ARRAY (MISA) TO MONITOR GRAVITY-
DEPENDENT CALCIUM FLUXES IN CERATOPTERIS SPORES
A.R. De Carlo1, M. Rokkam
2, A. ul Haque
3, S.T. Wereley
3, P.P. Irazoqui
4, H.W. Wells
5, W.T. McLamb
6, S.J.
Roux7, D.M. Porterfield
1,8
1Dept. of Ag. & Bio. Eng.,
2Dept. of Elec. & Comp. Eng.,
3Dept. of Mech. Eng.,
4Weldon School of Biomed. Eng.,
Purdue University, West Lafayette, IN, 5Bionetics Corporation, Kennedy Space Center, FL,
6Dynamac
Corporation, Kennedy Space Center, FL., 7Molecular, Cellular & Devel. Bio., Univ. of Texas, Austin, TX,
8Dept.
of Hort. & Landscape Arch., Purdue Univ., West Lafayette, IN.
Early development of the germinating Ceratopteris fern
spore involves the establishment of cellular polarity based
on gravity sensing in the single cell system (Edwards and
Roux; 1994). This process of cell polarization culminates
in directional nuclear migration which determines the
plane of the first cell division. The product of this first
mitotic division is a prothalial cell (top) and a rhizoid
(bottom).
Previous experiments have shown that polar Ca2+
currents are involved with gravimorphogenic
development in Ceratopteris fern spores (Chatterjee et al.,
2000). In a series of experiments that covered the time
period of polarity development that leads up to spore
germination, it was possible to measure calcium flux at
the top, bottom, and sides of the spore using a self-
referencing Ca2+
electrode (Porterfield, 2002). Using the
single sensor we were able to determine that there is a
transcellular Ca2+
current (into the bottom and out of the
top) that correlates with responsiveness of the cell to
gravity. More recently we have attempted to identify the
differential role of ion channels and pumps in this process
of cell polarity development using both molecular and
electrophysiological methods. We used a microsystem to
rotate individual cells 180° and measure changes in Ca2+
ion currents after reorientation using a single Ca2+
-
selective microelectrode operated as a self-referencing
sensor. This approach was limited as we could only
effectively measure before and after signals from a single
position, and was only able to do these experiments in a
40-second time frame (Stout et al., 2003). These
experiments were not able to measure how fast the
transcellular calcium current reorients, because of the
basic limitations in the sensor technology used.
In order to overcome the technical limitations to
studying this system associated with the current sensor
technology we have been working towards developing
new sensor systems based on adaptation of MEMS
fabrication techniques. The goal is to fabricate and test
dynamic sensing systems for advanced-throughput
physiological measurements of calcium signaling events
at the cellular level. Using silicon micromachining we
have constructed and tested prototype versions of the
Microfluidic Ionic Sensor Array (MISA) lab-on-a-chip
device for conducting real-time multidimensional calcium
flux measurements around 16 fern spores arranged in a
vertically positioned 4x4 matrix. A microfluidic pore
sustains each fern spore in basic growth media while four
solid-state calcium-selective electrodes measure the
calcium concentration differential pairwise between four
positions around the cell (1 top, 1 bottom, and 2 sides).
The microfluidic system and sensor array are fabricated
on a silicon substrate. The pyramidal pores which hold the
fern spores in place are etched at an angle of 54° using
KOH etching, and the electrodes and bonding pads are
defined by depositing silver and gold on top of photoresist
and lifting the photoresist off. Chloride plating of the
electrodes is accomplished through immersion in 6%
(w/w) NaClO. SU-8 photoresist (MicroChem, USA)is
applied to the chip as a structural and insulating layer, and
Ca++
-selective PVC membrane made with ETH-5234
ionophore (Sigma-Aldrich, St. Louis MO) is spin coated
onto the electrode surfaces. The dimensions of the whole
chip are 9.7mm x 11mm, with 250µm x 1000µm
silver/gold composite bonding pads on all sides and 20µm
x 20µm electrodes. For a single wafer there are 41 MISA
chips (Figure 1), of which the current process yields
approximately 80-90%.
Figure 1. Picture of the 9x11 mm silicon fabricated MISA chip.
The MISA chips are mounted and wire bonded
in the center of a printed circuit board (PCB) that contains
an array of 64 non-inverting amplifiers, using low-noise
(~3nV/√Hz), low-drift (~0.8µV/°C) operational
amplifiers, corresponding to the 64 electrodes. Each of the
amplifiers provides a signal gain of 30 and filters out
noise above 50Hz. The PCB is connected to four data
cables through 16-pin connectors with 2mm spacing. The
total size of the board is 30.25in2. The output from the
PCB is then multiplexed by a 512-point crosspoint matrix,
then processed and stored using a 32-input 18-bit data
acquisition (DAQ) card in differential mode. MISAPlot
2.0 software was written in LabVIEW (National
Instruments, Austin TX) for the DAQ. There are four
main modules in the software to facilitae experimental
functionality. The software also provides accesss
tomoothing and compression functions are written in, as
well as functions for logging acquired data, switching
data and notes from the user. Data extraction and rapid
calibration (Figure 2) are also possible.

AR DeCarlo et al. – Microfluidic Ion Sensor Array
124 Gravitational and Space Biology Bulletin 19(2) August 2006
We have also developed a novel coupling
method, dual-electrode differential coupling (DEDC), so
that the differentials between two working electrodes
could be amplified and digitized directly without the use
of reference electrode. This method was validated in tests
(Figure 3) that involved placing the two ion-selective
probes in separate Ca++ solutions, connected by a 3M
KCl salt bridge. One probe was calibrated in 100µM,
1mM and 10mM solution while the second electrode
remained in a standard solution. Each calibration was
done three times in each of the standard solutions.
Calcium Concentration (mM)
0.1 1 10
Raw
Ele
ctr
ode O
utp
ut (m
V)
-160
-140
-120
-100
-80
-60
Figure 2. Calibration (Average +/- SD) of a planar array of 64
separate calcium sensors using the ETH-5234 calcium-selective
membrane (Anker et al. 1981, Bühlmann et al. 1998). After spin
coating to form the membrane, the sensor array was left to sit
overnight in a dust-free container, then soaked in a solution of
10µM CaCl2 + 10mM NaNO3 (Konopka et al., 2004). Potentials
for the electrodes were measured in calibration solutions of
100µM, 1mM and 10mM CaCl2. The potentials were normally
distributed around a slope of 27mV which is in good agreement
with the theoretical slope predicted by the Nernst equation.
Calcium Concentration (mM)
0.01 0.1 1 10 100
Raw
Ele
ctr
ode o
utp
ut (m
V)
-60
-40
-20
0
20
40
60
10.0mM
1.0mM
0.1mM
Figure 3. The DEDC (dual-electrode differential coupling)
method was developed so that transcellular ions concentration
differentials could be measured without need for system offset
settings. Two glass-bodied probes were placed in separate
Ca++ solutions, connected by a salt bridge. Each series was
replicated using each of three standard solutions as a base
solution (figure legend). For each series the Nernst slope was
approximately 27mV, with the potential equal to zero when both
electrodes are in the same solution.
These basic tests have validated the functionality
of the technology. The MISA chip will be able to measure
the calcium concentration differentials associated with
transcellular currents from individual fern spores in real
time. The array of 64 electrodes is organized in quads
within 16 fluidic pores, enabling simultaneous
measurement of calcium concentrations differentials
around sixteen spores. The MISA system will be used in
parabolic flight/microgravity based physiological
experimentation, to study the role of polar calcium
currents in gravity-dependent cellular development.
Future uses of this in silico cell physiology lab-on-a-chip
technology include biomedical applications such as
advanced through-put cell viability and cancer detection.
Possible pharmacological applications include testing the
effectiveness and dose-response curves of drugs. We
anticipate adapting this base system by development and
adaptation of ionophores for other ions. We are also
developing different versions of this system for
amperometric electroanalytical chemistry, which will be
used for analytes like oxygen, nitric oxide, and various
neurotransmitters to name a few. This will also provide
the technical foundation for adapting enzyme based
biosensors for use on the system.
REFERENCES
Anker P, Wieland E, Ammann D, Dohner R, Asper R,
Simon W. 1981. Neutral carrier based ion-selective
electrode for the determination of total calcium in blood
serum. Analytical Chemistry 53: 1970-1974.
Bühlmann, P., Pretsch, E., Bakker, E. 1998. Carrier based
ion-selective electrodes and bulk optodes. 2.
ionophores for potentiometric and optical sensors.
Chemical Reviews. 98: 1593-1687.
Chatterjee, A., Porterfield, D.M., Smith, P.J.S., Roux, S.J.
2000. Gravity-directed calcium current in germinating
spores of Ceratopteris richardii. Planta 210: 607-610.
Edwards, E.S., Roux, S.J. 1994. Limited period of
graviresponsiveness in germinating spores of Ceratopteris
richardii. Planta 195: 150-152.
Konopka, A., Sokalski, T., Michalska, A., Lewenstam,
A., Maj-Zurawska, M. 2004. Factors affecting the
potentiometric response of all-solid-state solvent
polymeric membrane calcium-selective electrode for low-
level measurements. Analytical Chemistry 76: 6410-6418.
Porterfield, D.M. 2002. The use of microsensors for
studying the physiological activity of plant roots. In
Waisel, Y., Eshel, A., Kafkafi, U. (eds) Plant roots the
hidden half, 3rd Edition. Marcel Decker, New York. 333-
347.
Puglisi, J., Bers, D. 2001. LabHEART: An interactive
computer model of rabbit ventricular myocyte ion
channels and Ca transport. American Journal of
Physiology – Cell Physiology 281: C2049-C2060.
Stout, S.C., Porterfield, D.M., Roux, S.J. 2003. Calcium
signaling during polarity development. Annual Meeting
of the American Society for Gravitational and Space
Biology Abstract #39.

Gravitational and Space Biology Bulletin 19(2) August 2006 125
USE OF AN INTEGRATED FLOW-CHAMBER ADHESION ASSAY FOR MEASURING LEUKOCYTE
ADHESION PROPERTIES IN SIMULATED AND ACTUAL MICROGRAVITY
Dennis F. Kucik1,2,3,4
, Robyn L. Rouleau3, Lisa W. Smith
3, Xing Wu
1 and Kiran B. Gupta
1
Departments of 1Pathology and
2Biomedical Engineering, and
3Center for Biophysical Science and Engineering,
University of Alabama at Birmingham; and Birmingham VA Medical Center, Birmingham AL, 35233, USA
It has been known for many years that space flight
results in altered immune performance. After long-
duration missions, astronauts have increased leukocyte
counts, decreased T cell counts, and decreased production
of certain inflammatory mediators. Even after short-
duration flights, astronauts have minor immune-system
abnormalities. Although potential causes of space flight-
related immune dysfunction include physical stress and
cosmic radiation, microgravity alone has been shown in
both ground-based and flight experiments to have an
independent effect on white blood cells (Cogoli, 1993)
A key component of leukocyte function that may be
affected by microgravity is leukocyte adhesion to blood
vessel walls. This has been difficult to approach
experimentally, however, since adhesion is a functional
assay requiring live cells, skilled personnel and
sophisticated equipment, and cannot be readily performed
in flight.
To simulate microgravity on Earth, rotating wall
vessels (RWV) are commonly used. However, adhesion
assays as currently conducted require that rotation be
stopped, a sample removed, and cells washed and labeled
before an assay can be performed. The entire process
from cell sampling to completion of an adhesion assay
can take hours, giving the cells time to recover at 1g and
complicating interpretation of results.
To solve this problem, we have designed and built a
highly automated, integrated system that consists of a
modified RWV coupled to an adhesion assay capable of
real-time bio-imaging and recording. Our integrated
RWV/adhesion assay system can measure adhesion of
cells experiencing simulated microgravity within seconds
of returning to 1g, without stopping rotation of the
chamber.
One of the major challenges to overcome in designing
such a system is efficient, reliable extraction of cells from
a RWV without interrupting its rotation. A sample of
uniform cell concentration is essential for cell adhesion
assays, where adhesion is typically expressed as cells
adhering/minute (which depends on the concentration of
cells entering the chamber per minute). Theory predicts
that the distribution of the cells in the vessel may not be
uniform, but will depend on cell density (buoyancy) and
rotation speed, and that the cells will be subject to forces
of gravity, centrifugal force, and Coriolis-induced motion
(spiraling) (Hammond et al, 2001). Our current system is
apable of delivering leukocyte samples of constant
concentration for prolonged periods (Table 1).
Table 1. Uniformity of concentration of cell sample
extracts. 90 µl samples were withdrawn from the
integrated RWV (without stopping rotation) at the time
points indicated (minutes). These aliquots, which would
normally be perfused through the flow adhesion assay
chamber, were saved and cell concentration (X 106
cells/ml) was determined. Concentrations are the mean
of three measurements + SEM. The system reaches a
steady state within 10 minutes of initiation of rotation,
and delivers samples of uniform concentration thereafter.
Time 1 10 20 30 40 50 60
Conc.
(SEM)
0.7
+.1
1.3
+.1
1.3
+.2
1.3
+.1
1.4
+.1
1.3
+.1
1.3
+.1
The adhesion assay in this system is a laminar flow
chamber, which mimics the conditions in the blood vessel
under which leukocytes actually adhere to vascular
endothelial cells in vivo. Besides being more
physiological, there are additional advantages to using a
flow adhesion assay (as opposed to static assays) to
measure the effects of microgravity. First, the assay can
be completed in seconds, minimizing the recovery time at
1g. Second, a flow chamber assay is more sensitive to
weak adhesion forces than static assays In addition,
some adhesive events (such as adhesion mediated by
selectins, an important class of adhesion molecule) occur
only in the presence of shear, and, thus, cannot be
characterized under static conditions (Finger et al, 1996).
Finally, adhesion to blood vessel walls takes place in a
series of steps, or sub-processes. The flow cell assay
allows visualization of these sub-processes of adhesion,
including rolling, firm arrest, adhesion strengthening,
spreading, and migration.
In order to minimize the time between extraction of
cells and delivery to a microscope-based assay (which
depends on the length of the intervening tubing), an
integrated bio-imaging module was designed that was
small enough to be permanently mounted a few inches
from the RWV. A custom optical train was constructed,
with integrated illumination designed to efficiently
visualize unlabeled, live leukocytes as they were perfused
through a laminar flow chamber, and this was coupled to
a videocamera. Images of sufficient quality to permit
detailed computerized motion analysis were obtained with
this system, as illustrated in figure 1.

D.F. Kucik et al – Integrated RWV adhesion assay
126 Gravitational and Space Biology Bulletin 19(2) August 2006
0.0 0.3 1.0 3.00.0
2.5
5.0
7.5
10.0
Cyto D (µµµµg/ml)
Cell
s a
dh
ere
ing
/min
ute
Figure 1. Leukocyte adhesion to vascular endothelial
cells as visualized in the integrated system. Endothelial
cells were cultured for 6 days and incorporated into a
flow-chamber assay system as previously described
(Kevil, Chidlow, Bullard, and Kucik, 2003). Adhesion of
HL-60 monocytic cells to the endothelial cell monolayer
was then observed and analyzed. Arrows indicate
monocytic cells adhering to a monolayer of vascular
endothelium.
Both integrin- and selectin-mediated adhesion were
assayed with this system and compared to results obtained
using a conventional microscope-mounted shear-flow
adhesion assay system. Our data show that the new
integrated assay can detect defects in both rolling and
firm adhesion with sensitivity equal to that of large,
microscope-based flow chamber adhesion assays. Figure
2 shows the dose-dependent effect of cytochalasin D on
HL-60 monocytic cells rolling on purified E-selectin and
ICAM-1 as assayed with the integrated system. This is a
test for modulation of β2-integrin-mediated adhesion that
occurs in response to rearrangement (clustering) of the
integrins. As demonstrated earlier (Kucik et al, 1996),
low-dose cytochalasin D releases β2 integrins from
cytoskeletal constraints, and the resulting rearrangement
into clusters (van Kooyk et al, 1999) is pro-adhesive. At
higher cytochalasin D concentrations, the morphology of
the cell is altered, and adhesion becomes less effective (Ni
et al, 2003). The integrated system was sensitive enough
to detect both the increase in adhesion and its dose
dependence, in good agreement with previous studies
using more conventional, commercial microscope-based
assay systems (Ni et al, 2003). Thus, the integrated
system will be suitable for detecting even subtle effects of
simulated microgravity on adhesivity.
This system has now been adapted to measure acute
effects of actual microgravity in parabolic flight
experiments. Lessons learned from development of this
Earth-based system can form the basis for a future space-
flight culture and assay system to allow NASA to monitor
microgravity-inducible physiological events during space
flights.
Figure 2. Detection of integrin rearrangement-induced
effects on adhesion. HL-60 cells were pre-treated with
cytochalasin D at the concentration indicated and
perfused over a substrate of 25 ng/ml ICAM-1 and 0.5
ng/ml E-selectin (SOURCE) at a shear stress of 0.5
dynes/cm2. HL-60 cells/minute adhering to the substrate
during the period of observation (3 min.) were then
quantified. Data are the mean (+SEM) of at least 5
observations for each concentration.
REFERENCES
Cogoli,A. 1993. The effect of hypogravity and
hypergravity on cells of the immune system. Journal Of
Leukocyte Biology 54:259-268.
Finger,E.B., K.D.Puri, R.Alon, M.B.Lawrence, U.H.Von
Andrian, and T.A.Springer. 1996. Adhesion through L-
selectin requires a threshold hydrodynamic shear. Nature
379:266-269.
Hammond,T.G. and J.M.Hammond. 2001. Optimized
suspension culture: the rotating-wall vessel. American.
Journal of Physiology. - Renal Physiology. 281. (1):F12. -
25. , 2001. Jul.F12-F25.
Kevil,C.G., J.H.Chidlow, D.C.Bullard, and D.F.Kucik.
2003. High-temporal-resolution analysis demonstrates
that ICAM-1 stabilizes WEHI 274.1 monocytic cell
rolling on endothelium. American Journal of Physiology -
Cell Physiology. 285:C112-C118.
Kucik,D.F., M.L.Dustin, J.M.Miller, and E.J.Brown.
1996. Adhesion activating phorbol ester increases the
mobility of leukocyte integrin LFA-1 in cultured
lymphocytes. J. Clin. Invest. 97:2139-2144.
Ni,N., C.G.Kevil, D.C.Bullard, and D.F.Kucik. 2003.
Avidity modulation activates adhesion under flow and
requires cooperativity among adhesion receptors.
Biophysical Journal. 85:4122-4133.
van Kooyk Y., van Vliet SJ, and C.G.Figdor. 1999. The
actin cytoskeleton regulates LFA-1 ligand binding
through avidity rather than affinity changes. J. Biol.
Chem. 274:26869-26877.

Gravitational and Space Biology 19(2) August 2006 127
DEVELOPMENT OF THE EMCS HARDWARE FOR MULTIGENERATIONAL GROWTH OF
DROSOPHILA MELANOGASTER IN SPACE
M. E. Sanchez1, M. Shenasa
2, A. Maldonado
1, A. Kakavand
1, D. Leskovsky
1, E. Houston
3, A. Howard
3, M. K.
Steele1, and S. Bhattacharya
4.
1Lockheed Martin Space Operations,
2 NASA/Ames Internship Program,
3H&H Engineering, and
4NASA
Ames Research Center, Life Sciences Division, Moffett Field, CA 94035
Understanding the changes that occur in living
organisms to bring about adaptation to the space
environment is essential to supporting future plans for
long-term missions to the Moon and Mars. The European
Modular Cultivation System (EMCS), a life science
research facility developed by the European Space
Agency (ESA), can serve as a habitat for culturing
multiple generations of Drosophila melanogaster. Based
on the results obtained from previously tested prototype
hardware, a new Prototype Container III (PIII) was
developed (Figure 1-A) and tested. Data from Prototype
containers I and II were published previously (M.E.
Sanchez, et al., 2004).
Figure 1. Figure 1-A depicts the EMCS Drosophila
Prototype Container III and its food compartment. Figure
1-B is a close-up of an Experiment Container containing
two Drosophila Prototype Containers and the staggered
configuration of the prototype containers within the
Experiment Container allow optimal lighting and imaging
of the flies.
The primary differences between the new design and
the previous models were: A. Increase in growth chamber
volume, B. Modification of air holes in order to fit the
holding frame for the Experiment Container or EC
(Figure 1-B), C. Addition of stainless steel membranes to
cover air holes, and D. Addition of a rubber band
covering the junction between the growth chamber and
food cylinder in order to prevent larval escape.
The goal of these experiments was to optimize growth
of Drosophila cultures and to ensure biocompatibility of
the cultures within the containers. During the
development phase, modifications to the containers were
made as required to resolve any suboptimal performance
issues. Fly behavior, humidity levels, and hardware
mechanics were assessed. Flies were grown in the
containers using standard techniques for fly handling and
videotaped using the video capabilities of the EMCS
ERM (Experiment Reference Module) camera system.
Assessing optimal humidity levels within the growth
chamber is critical for culture health. Adults could adhere
to container surfaces and die if the humidity levels
dramatically increase, while larvae might dessicate if
humidity levels decrease. In order to find an optimal
material that allows sufficient airflow and maintains
normal levels of humidity, we grew flies in the containers
using three different types of breathable membranes:
stainless steel mesh, Nytex™ (nylon mesh), and a paper
membrane. Growth in these conditions was compared to
standard culture vials covered with the three different
types of membranes (Figure 2).
Figure 2. Population densities of flies in containers that
were covered with stainless steel, Nytex, and paper
membranes. All three membranes allowed air exchange.
Due to shortage of hardware, results of the containers are
representative of a single experiment with duplicates for
each condition. Results of the control-covered vials were
representative of three independent experiments.
While all three membranes allowed air exchange
(Figure 2), only the Nytex membrane covered prototype
containers showed a reduction in progeny size (Figure 3).
Since the integrity of the paper and Nytex membranes are
affected by the humidity levels, the stainless steel
membrane was selected for further experiments.
Selecting a food media that can withstand long-term
storage without losing its nutritional value is also critical
for space flight experiments. Making fresh media on orbit
is impractical and would require significant crew time,
therefore pre-prepared fly media will need to be stored for
several weeks prior to conducting multi-generation
experiments in space. In this test, four media recipes
were made and stored in Mylar™ bags (Impak
Corporation PAKVF 3.5M Silver, available from
www.sorbentsystems.com). We tested the following four
media recipes: A. Semidefined (1% Agar, 8% Brewer’s
Yeast, 2% yeast extract, 2% peptone, 3% sucrose, 6%
glucose, 0.05% magnesium sulfate, 0.05% calcium
chloride, 0.6% propionic acid, 1% of 10% p-Hydroxy-
benzoic acid methyl ester in 95% ethanol) B. Semi-
Biocompatibility of Membranes
0
50
100
150
200
250
300
350
400
450
500
PIII Control-covered
Container Type
Stainless steel
Nytex
Paper
A B

M.E. Sanchez et al. – Development of EMCS Hardware
128 Gravitational and Space Biology 19(2) August 2006
defined with cornmeal (same as semidefined with the
addition of 60g of cornmeal) C. EMCS Fab Feast 1:1
(6.47% dextrose 6.47% molasses, 0.93% agar, 6.12%
cornmeal, 3.24% yeast, and 2.0% Tegosept) D. EMCS
Fab Feast 3:1 (Same as EMCS Fab Feast 1:1, with the
exception of 9.7% dextrose and 3.23% molasses).
The media was prepared and packed under sterile
conditions. In addition, we used proton beams at a range
of 25-33 kgrays to sterilize a number of our samples. This
was done through the company Nutek at Hayward
California. The additional irradiation was conducted in
order to assess if additional sterilization was necessary for
samples that will be stored for long periods of time prior
to use.
Figure 3. Figures A and B show the results from non-
proton sterilized and proton sterilized media respectively.
Overall, the non-proton sterilized media performed better
than the sterilized media with fly cultures. Data shown
here represent four independent studies at room
temperature.
All four media recipes were able to support fly growth
(Figure 3-A and 3-B). Non-proton sterilized media
performed better than proton sterilized media, by
supporting larger fly populations while still showing no
bacterial or fungal contamination. The reason for the
decrease in performance of the proton sterilized food is
unclear, but it may be due to the degradation of
constituents in the media as a result of proton irradiation.
For purposes of supporting growth in the prototype
containers for 2-3 day egg-lays, we found that the
standard Semidefined media performed the best. The
progeny yields from the Semidefined with cornmeal
media were too large and caused overcrowding of the
containers. However, during space flight experiments, if
longer egg lay periods need to be used to accommodate
crew schedules, then Fab Feast media can be used or for
shorter egg lays, Semidefined with cornmeal can be used.
The data shown in Figure 3 shows that the food is usable
for 2 months. We have also tested these media for longer
periods of time, and find that the food can be stored in
this way at room temperature for periods of over a year
(data not shown) and still sustain fly growth.
Having completed a considerable amount of testing
to optimize the Prototype Fly Containers, we proceeded to
test the Prototype Containers within the Experiment
Reference Model (ERM) hardware in order to optimize
performance within the full flight hardware configuration.
The ERM hardware provides the interfaces and
environmental controls similar to the flight EMCS
research facility that will eventually be used for
conducting Drosophila research in space. Temperature
levels were set at 23°C during the light cycle and 25°C
during the dark cycle. This delta of 2°C was to neutralize
the heat generated from the LED board immediately
above the EC. Growth under these conditions was
compared to standard culture vials that were maintained
in our laboratory incubators.
The ERM was able to support fly growth (Figure 4).
Both locations of the EC, top and bottom, produced
nearly the same numbers of flies. These positions of the
hardware were described earlier in Fig 1B. Temperature
and humidity levels were monitored throughout the
experiment (data not shown). About three days before the
1st generation emerged, we removed the parental
population. Figure 4 shows the counts from the 1st
generations that emerged from each container.
Figure 4. Testing of Prototype containers within the
Experiment Reference Model hardware. Both positions in
the holding frame of the EC generated a comparable
numbers of flies. The controls were conducted using
standard culture vials in our laboratory incubators.
Summary A prototype container has been developed for conducting
experiments with Drosophila melanogaster in the EMCS
hardware on the International Space Station. This
container is capable of supporting larger populations of
flies than standard laboratory vials, as depicted in figures
2 and 4. Our studies have helped optimize humidity
levels, food formulations, and container materials in order
to effectively support fly populations in this hardware.
Tests that should continue in the future include
multigenerational growth testing and the development of
sampling devices.
A Non-Proton Sterilized
0
100
200
300
400
500
600
0 1 2
Age (month)
Semidefined
Semidefined-cornmeal
EMCS Fab 1:1
EMCS Fab 3:1
B Proton Sterilized
0
100
200
300
400
500
600
0 1 2
Age (month)
Semidefined
Semidefined-cornmeal
EMCS Fab 1:1
EMCS Fab 3:1
0
50
100
150
200
250
300
350
400
Bottom Container Top Container Controls
Container Location

Gravitational and Space Biology 19(2) August 2006 129
INVESTIGATING LOCAL IMPACTS OF HEAT-PULSE SENSORS FOR MEDIA MOISTURE
CONTENT
M.A. Ask1, J.J. Prenger
2, D. Rouzan-Wheeldon
2,
V. Rygalov3, J. Norikane
4 and H.G. Levine
5.
1Kennesaw State University, Kennesaw, GA
2Dynamac Corporation, Kennedy Space Center, FL
3University of
North Dakota, Grand Forks, ND 4Universtiy of Kentucky, Lexington, KY
5NASA Biology Division, Kennedy Space Center, FL
Introduction Cultivating plants in space will require automated
water provision systems to both maximize plant
performance and minimize crew operations. We
report here on studies evaluating small (1 cm
diameter) heat-pulse sensors designed to measure
media moisture content (TMAS sensors produced by
Orbital Technologies Inc., Madison, WI).
Calibrating the sensors’ response to the moisture
content of the media is a critical step towards their
effective and accurate use. The sensors were
designed to perform two functions: heat up and take
temperature readings. When heat is applied to an
object, the local environment of that object, relative
to the heat source, is affected. Analyzing the sensor
areas after an extended period of use was important
to determine the sensors’ lasting effects on their
environment. Thermal imaging was used to observe
the heat dissipation during the heating and cooling
processes. In this way we have addressed the
question of whether the sensors’ thermal input could
change their local environment with respect to the
bulk media. Studying these local effects will guide
procedures for calibration and operation.
Materials and Methods Six moisture sensors were prepared for testing.
Pairs of felt capillary mat squares were cut and
stitched together to hold the sensors in place (Figure
1). The mats were designed to fasten to small,
horizontally oriented fixtures. Relative water
content (RWC) of the capillary mats was calculated
as the ratio of the mass of water in the capillary mat
(the medium) to the mass of water capable of being
held within the mats at 100% saturation (measured
gravimetrically). Different RWC levels were
achieved by wetting the capillary mats to 100%
RWC and then allowing them to dry in open air until
the desired RWC was reached. The TMAS sensors
were programmed to provide thermal pulses of 90
second (s) durations, and temperature data were
acquired at 10 s intervals during each heat pulse and
for a 90 s cooling interval.
7 cm
7 cm
1 cm
Sensor
Software Sensor
Board
Capillary Mats
Legend
TMAS Sensors
Stitching
Drawing not to scale
Top View
IR Camera
Capillary Mats
Plastic bases
Standoffs
Side View
7 cm
7 cm
1 cm
7 cm
7 cm
1 cm
Sensor
Software Sensor
Board
Capillary Mats
Legend
TMAS Sensors
Stitching
Drawing not to scale
Top View
IR Camera
Capillary Mats
Plastic bases
Standoffs
Side View
Figure 1. Experimental setup for the TMAS sensor
testing.
Measurement frequency treatments were
administered at TMAS thermal measurement
(sampling) intervals of 10 and 20 minutes (with 3
sensors in each treatment) for 1 week. Mats were
stored in closed containers during the test to reduce
evaporation and control RWC levels. After one
week, thermal imaging was used to observe heat
dissipation. Heat was applied to each mat internally
via TMAS sensors and externally using a heat gun.
Mathematical Modeling
TMAS readings were used to develop models
relating change in temperature and the slope of the
cooling curve with RWC based upon temperature
measurements recorded every 10 s during heating
and cooling intervals. Five sets of TMAS
measurements were taken at similar RWC levels
during mat drying. Three tests were used to create a
correlation between the reference RWC and TMAS
response for two calibration equation methods.
Independent data sets for TMAS response were used
to validate the calibration methods:
Method 1: ( )iif TcTTccRWC 321 +−+=
Method 2: coolingdt
dTccRWC 54 +=
where: RWC is Relative Water Content (%), Tf is
final temperature after heat pulse (°C), Ti is initial
temperature before heat pulse (°C), dT/dt is the slope
of the cooling curve for the first 20 s after heat pulse
(°C/s), and c1 - 5 are empirical constants.

M.A.Ask, et al. – Local Impacts of Heat-Pulse Sensors
130 Gravitational and Space Biology 19(2) August 2006
Thermal Imaging
Heat dissipation patterns from TMAS sensor thermal
input and from the external (heat gun) source were
observed in the mats using time-lapsed thermal
photography. Spatial temperatures (as shown by
color) were compared before and after thermal
inputs, internal and external, to determine if there
was a lasting difference attributable to sensor
location compared to the rest of the capillary mat.
Results The heating and cooling phases of the treatment
presented two unique mathematical problems. The
correlations between sensor responses and RWC
were used to create calibration equations using two
mathematic approaches, while independent data
were used to validate the calibrations. Table 1
shows the range of error for all sensors with average
errors of 6.3% and 6.1% for Methods 1 and 2,
respectively. There were data points for which the
prediction error was significantly greater than the
average error. Though there was a lower degree of
variation in Method 2, there was still enough error to
warrant further investigation.
Table 1. The root mean square errors (RMSE) of
validation data sets (N=8) are given for two separate
methods used to calibrate the sensors.
Method 1 Method 2
Sensor 1 8.1% 5.5%
Sensor 2 5.1% 7.1%
Sensor 3 5.6% 5.9%
Sensor 4 6.4% 5.2%
Sensor 5 6.1% 6.3%
Sensor 6 6.3% 6.3%
Average RMSE 6.3% 6.1%
Figure 2 shows typical heating/cooling profiles for
one TMAS sensor (colorized version available at
publication website). After 210 s, observable heat
had dissipated across the mat and temperatures had
returned to pre-heating levels for all but the lowest
RWC level. Measurement frequency during week-
long operation prior to thermal imaging did not
affect the results.
Summary and Conclusions Thermal heat-pulse moisture sensors operate by
emitting heat that could alter the sensor’s
environment and possibly bias their accuracy.
Experiments were done to determine the localized
effects of frequent sensor heating. Two methods
were devised to model the sensor’s response with the
RWC of the media. While one method more
accurately modeled the relationship, significant
errors warrant further investigation. Time-lapsed
thermal photography was used to monitor the
media’s response to internal (TMAS) and external
(heat gun) heating. Infrared images showed no
significant long-term (>3 min) localized changes in
thermal characteristics within the substrate from
either internal or external heat sources.
Figure 2. Thermographic images of the experimental
mats with sensors embedded. A. For internal heat
pulses, faster heat dissipation was evident at higher RWC
levels, but for all RWC levels there was nearly complete
heat dissipation after 210 s. NA – image Not Available.
B. For the external heat applications (white represents
the highest temperature) recovery was again nearly
complete at 210 s. The color version of this figure may be
viewed in the online publication of the journal at
http://asgsb.org/publications.html, link to Gravitational
and Space Biology.
Acknowledgments
This research was supported by NASA contract
NCC10-52. We thank the SLSTP program and
Bionetics Corp. for their participation in this
research.
NA
Temp ¡C
0
15
30
45
60
75
90
120
150
180
210
Time (s)
Relative Water Content
60% 75% 90% 100%
0
15
30
45
60
75
90
120
150
180
210
Time (s)
Relative Water Content
60% 75% 90% 100%
Heating Cooling0 s 210 sB.
A.
NA

Gravitational and Space Biology Bulletin 19(2) August 2006 131
PERFORMANCE EVALUATION OF A LABORATORY TEST BED FOR PLANETARY
BIOLOGY
Nathan A. Thomas1, Paul Todd
1*, G. William Metz
1, Michael A. Kurk
1, David J. Thomas
2,
1SHOT, Inc.,
Greenville, IN 47124, and 2Lyon College, Batesville, AR.*Corresponding author [email protected]
The major issues of planetary biology consist of
searching for life, planetary protection, environmental life
support for human visitors, and terraforming. A
definition of terraforming is the conversion of a planetary
surface to earth-like conditions hospitable to Earth life. A
subset of terraforming is ecopoiesis, the early
development of a living ecosystem (Haynes and McKay,
1992). Despite approximately two decades of public
discussion there has been little or no experimental
research focusing on ecopoiesis. Initially, small
experiments with terrestrial organisms in a planetary
(Mars-like) environment are required. Such experiments
simultaneously address issues of ecopoiesis and planetary
protection. To facilitate such experiments in research
laboratories a laboratory test bed was constructed for the
simulation of planetary environments for biological
experimentation.
The surface of Mars was chosen as the principal
environment to simulate. The requirements for high-
fidelity simulation of the Mars environment are based on
current knowledge or understanding of each of the
following features (Banin and Mancinelli, 1995):
Atmosphere: The atmospheric pressure on the surface of
Mars varies between 7 and 15 mbar, depending on
altitude (mountains vs. valleys) and season. This is about
1% of the pressure on the surface of the Earth. The
atmosphere contains 95% CO2, 2.7% N2, 1.6% Ar, 0.13%
O2, 0.03% H2O and trace amounts of Ne, Kr, Xe and O3.
Since the trace O3 does not contribute to the UV
absorbance of the atmosphere it and the minor inert gases
are not essential components of a simulated Mars
atmosphere. Temperature: The maximum temperature on
the surface of Mars on a cloudless summer day at
equatorial and mid-latitudes is about 26oC. The minimum
night-time temperature at higher latitudes is -135oC. An
average daily cycle under consideration for simulation
purposes is -80oC minimum night-time temperature and
+26oC maximum mid-day temperature (Carr, 1996). This
day-night temperature cycle must be repeated every 24.6
h, the length of the Martian sol (day). Sunlight: The total
intensity of sunlight on the surface of Mars is about 40%
of that at the top of Earth’s atmosphere, so that the
Martian surface receives 590 W-m-2
of the essentially
unattenuated solar spectrum. This spectrum includes
ultraviolet light with wavelength below 300 nm, including
down to 190 nm. This spectral range is photochemically
damaging to nucleic acids and known to kill cells and is
included in the spectral output of xenon arc lamps.
Regolith: The dust on the surface of Mars has a high
content of oxides of iron, some of which were found to be
oxidizing agents in the experiments on the Viking
missions. In elemental composition the regolith is 12.5%
Fe, 21% Si, 5% Mg, 4% Ca, 3.1% S, 3% Al, 2.3% Na,
0.7% Cl, 0.3% P, and contains traces of Mn, Co, Cu, I,
Zn, B, etc. Thus, in addition to the aluminosilicates there
is an ample supply of components required by living
organisms, except nitrogen and carbon, which are
atmospheric (Banin and Mancinelli, 1995).
In order to match these requirements the test bed
consists of the following subsystems: a 1,000 W xenon
arc lamp with an “AM0” filter to simulate solar irradiance
in space, an 8 ft3 cryogenic thermal cabinet capable of
cooling to -135oC and heating to +50
oC using evaporating
nitrogen and resistive heaters, a fused silica cylinder that
transmits the full solar spectrum and can be evacuated to
<10 mbar through stainless steel end caps, a 500-gal
liquid nitrogen storage tank, a cylinder of pressurized
support gas, and a control system that automatically
cycles the interior of the cylinder through a Mars daily
cycle, for example. Biological samples placed in the
quartz cylinder are typically mixed with simulated
planetary regolith. A rendering of the simulator design is
shown in Figure 1.
During initial operation the test bed was used for
preliminary biological experiments designed to test
concepts of ecopoiesis. The light and temperature profiles
matched those of Mars near the equator at the vernal
equinox. The test bed was cycled daily between -80oC
and +26oC following a published temperature curve (Carr,
1996) shown in Figure 2, which includes a demonstration
of maximum heating and cooling rates achievable in the
test bed.
The atmospheric pressure on Mars is 7-15 mbar, and the
day length is 24.6 hours, but internal pressure of 100 mbar
and day length of 24.0 hours were used for early
biological experiments. The gas composition used was
0.139% O2, 1.615% Ar, 2.760% N2 with the balance CO2
or, in some cases, pure CO2. The atmosphere was
saturated with water by the daily injection of a few grams
of (liquid) distilled water into the atmosphere (water boils
at 49oC at 100 mbar). The measured photosynthetically
active photon flux was 1100 µmoles-m-2
-s-1
in direct light
and 12 µmoles-m-2
-s-1
in the shaded interior of the
cylinder. This corresponds closely to values determined
for the Martian surface and is consistent with the 590 W-
m-2
total energy flux. Pressure, illumination, cabinet
temperature and regolith temperature were recorded
during operation of the test bed for 1, 8, 14 and 35-day
periods. An example of a 14-day data set is shown in
Figure 3. Under daily cycling liquid nitrogen was
consumed at approximately 1600 liters/week and was
exhausted safely to the exterior of the laboratory.

N. Thomas et al. – Planetary Biology Test Bed
132 Gravitational and Space Biology Bulletin 19(2) August 2006
Figure 1. Laboratory Test Bed design showing components for illumination, temperature control, atmosphere supply, and
specimen chamber (“Mars Jar”).
Figure 2. Temperature profile chosen for simulation in
early test-bed experiments. Left (triangles) and right
(squares) vertical curves represent maximum heating and
cooling rates, respectively, in the environmental chamber.
This research was funded under subcontract agreement
07605-003-026 with NASA's Institute for Advanced
Concepts (NIAC), a program of the Universities Space
Research Association (USRA) funded by NASA contract
NAS5-03110.
Figure 3. A record of physical data for a 14-day
experiment with daily cycling of temperature between -
80oC and +26
oC, 12 h of light and 12 h of darkness,
pressure of 100 mbar. Lower trace is cabinet
temperature, and upper trace is regolith temperature.
Horizontal bar at 0 indicates illuminator on, bar at 10,
off.
REFERENCES
Banin, A. and Mancinelli, R.L. 1995. Life on Mars? I.
The chemical environment. Advances in Space Research
15:163-170.
Carr, M. H. 1996. Water on Mars. Oxford University
Press, New York.
Haynes, R. H., and McKay, C. P. 1992. “The
implantation of life on Mars: Feasibility and motivation.”
Advances in Space Research 12: 133-140.
ENVIRONMENTAL
CONTROL UNIT
1.5kW SOLAR
SPECTRUM
GENERATOR LAPTOP
COMPUTER
SPECIMEN
CHAMBER
FULL SPECTRUM
LIGHT DISTRIBUTION
MIRROR
2 STAGE REGULATOR
LIQUID
NITROGEN
SIPHON TANK
VACUUM
PUMP
LOW PRESSURE
GAS RESERVOIR
MARS ATMOSHPERE
HIGH PRESSUREGAS TANK
STAND
MARS GAS SUPPLY/
VACUUM SYSTEM
MULTI-PANE
WINDOW (UV SAFE)
WITH CURTAIN
MARS
ATMOSPHERE
GAS LINE IN
VACUUM
LINE OUT
INSULATED
DOOR

Gravitational and Space Biology 19(2) August 2006 133
A STUDY OF THE EFFECTS OF SPACE FLIGHT ON THE IMMUNE RESPONSE IN DROSOPHILA
MELANOGASTER
Thomas F. Fahlen1*
, Max Sanchez1*
, Matthew Lera2, Edina Blazevic
2, Jennifer Chang
1, and Sharmila
Bhattacharya3
1Lockheed Martin Space Operations, NASA Ames Research Center, Moffett Field, CA 94035
2NASA/Ames Internship Program, NASA Ames Research Center, Moffett Field, CA 94035
3NASA Ames Research Center, Life Sciences Division, Moffett Field, CA 94035. *Both authors contributed
equally to this work and should be considered as joint first authors.
Drosophila melanogaster (fruit fly) is a small, highly
tractable and well-characterized organism that is ideal for
the study of molecular, cellular, developmental, and
physiological processes. In Drosophila, many of these
processes, including certain aspects of innate immunity,
are very similar to those in humans (1). There is evidence
that space flight affects the immune system of Astronauts
such that the Astronauts may become
immunocompromised during and after flight (2, 3). In
order to study those possible effects, our laboratory will
be supporting a study to be carried in the mid-deck of the
Space Shuttle to examine the effects of space flight on the
immune system using D. melanogaster as a model.
Our experiment configuration consists of 10 sample
containers (divided into 5 primary and 5 secondary
containers) that support adult Drosophila and allow egg
laying and larval development during flight. During the
scheduled flight, virgin female and male adult flies will be
loaded into the primary containers. The flies will then be
allowed to lay eggs on the food for three days. At the end
of the third day, the food tray in the primary containers
will be switched to the secondary containers. This will
move eggs and larvae produced during pre-flight and
launch to the secondary cassettes. A new food tray will be
placed in the primary container. The sample containers
consist of ventilated aluminum cassettes enclosed by
ventilated type I containers (Fig. 1 A-B). In order to
execute this experiment in space, these aluminum
containers were built specifically to allow Drosophila
growth under the ambient conditions of the shuttle
middeck, without requiring power. This paper describes
biocompatibility tests to show the suitability of the
materials to specimen welfare, and should facilitate the
use of these containers by other investigators in the future.
Figure 1. Figure 1-A depicts one fully loaded Drosophila
container. Figure 1-B depicts the Type I container in
which the cassette will be enclosed.
The individual containers will be housed in one of the
middeck drawers of the Space Shuttle. This consists of a
foam tray holding change-out platform. The foam used in
this drawer is made out of polyurethane foam Pyrell®.
This material is frequently used in the packaging of flight
payloads because it is a low flammability foam. In order
to check if the foam used in the mid-deck drawer had any
adverse effect on the viability of the flies, 1cm cubes of
flight foam were placed in vials containing fly media. 10
virgin females and 10 male flies were added to each of
these vials. The flies were allowed to mate and lay eggs
for two days at 25°C. At the end of this period, the
parental population was removed from the vials. The vials
containing eggs were allowed to develop for 12 days at
25°C. The progeny were counted and the male/female
ratios were scored (Figure 2).
Figure 2. Total progeny produced from eggs laid in the
presence or absence of polyurethane foam Pyrell®. N=5
The results on Figure 2 suggest no significant difference
in the number of progeny produced in the presence or
absence of the flight foam material. Additionally, no
significant death of parental flies was observed (data not
shown). It was concluded, therefore, that Pyrell® foam
was biocompatible with the specimen in this experiment
configuration.
The next study was used to evaluate the effects of
different starting population densities on progeny
production. In order to find the optimal number of starting
parental population, we tested two different
configurations: A. 5 virgin females and 5 males (5F+ 5M)
and B. 10 virgin females and 5 males (10F + 5M). It is
important to optimize the size of the starting population in
order to maximize the number of progeny developed in
space for each experiment. As shown on Figure 3, a
A B

T. Fahlen et al. – A Study of the Effects of Space Flight on the Immune Response in Drosophila
melanogaster
134 Gravitational and Space Biology 19(2) August 2006
starting population of 10 virgin females and 5 virgin
males is optimal for our 12 day experiment in space. The
total progeny size from 10 females and 5 males was
bigger than that produced by 5 virgin females and 5
males, and yet these containers did not show signs of
overcrowding.
When the samples return from space, a considerable
amount of work will be conducted with blood obtained
from third instar larval samples. An experiment was
performed with the flight containers for the full flight
duration and the number of resulting embryos and larvae
were counted as shown in Figure 4. Large numbers of
third instar larvae were obtained which should be more
than adequate for the hemocyte analyses to be conducted
from the space-flown samples. The embryos and earlier
instar larvae are also present in sufficient numbers for
adequate morphology and gene expression studies
planned for our flight experiment.
Figure 4. Number of animals at each developmental
phase.
Our experiment is designed to assess the immune function
of flies following a shuttle mission. As an assay for
immune function, we have adapted a bacterial clearance
assay, previously described by Brandt et al. (4), to our
post-flight processing. Briefly, this assay involves
infecting the flies with a known quantity of bacteria and
assessing the ability of the flies to clear the infection.
Ground testing in our laboratory shows that we are able to
infect flies with 10,000 E. coli HB101 bacterial cells with
greater than 95% survival of the flies (data not shown).
Following 3 days of infection we find that the flies reduce
their bacterial load by two orders of magnitude (Figure 5).
Our flight experiment will utilize this assay to assess the
function of the immune system of flies that have
developed in space.
Figure 5. Bacterial challenge assay. Flies were injected
with 50 nl of a 2 x 108 cells/ml suspension of E. coli
HB101 in PBS (10,000 cells/fly). A subset of the flies
(Day 0) were immediately assayed for bacterial load by
dilution plating of homogenates prepared in Luria Broth
+ 1% triton X-100 with 3 flies per homogenate. A second
subset of flies were incubated at 29 °C for 3 days and then
assessed for bacterial load in the same manner. Only live
flies were assayed. Error bars show the standard
deviation from the mean. N = 3.
We have developed and tested containers that will sustain
our specimen for an experiment designed to address the
effects of spaceflight on the innate immune system of
Drosophila. These containers should also be appropriate
for use by other investigators needing to use Drosophila
melanogaster as a model specimen in space.
ACKNOWLEDGEMENT
We gratefully acknowledge helpful discussions with Dr.
David Schneider concerning the bacterial infection assay.
REFERENCES
1. Hoffman, J.A., F.C. Kafatos, Ç.A. Janeway Jr.,
R.A.B. Ezekowitz. 1999. Phlogenetic Perspectives in
Innate Immunity. Science. 284:1313-1318
2. Borchers, A.T., C.L. Keen, M.E. Gershwin. 2002.
Microgravity and immune responsiveness:
implications for space travel. Nutrition. 18:889-898.
3. Sonnenfeld, G. and W.T. Shearer. 2002. Immune
function during space flight. Nutrition. 18:899-903.
4. Brandt, S.M., M.S. Dionne, R.S. Khush, L.N. Pham,
T.J. Vigdal, D. S. Schneider. 2004. Secreted Bacterial
Effectors and Host-Produced Eiger/TNF Drive Death
in a Salmonella-Infected Fruit Fly. PLOS. 2:e418.
100
1000
10000
100000
DAY 0 Day 3
Condition
Figure 3. Comparison of starting parental density on
progeny production in flight cassettes.
0
100
200
300
400
500
600
700
800
900
1000
Males Females Males Females
Primary Containers Secondary Containers
To
tal
Nu
mb
er o
f A
du
lt F
lies 5F + 5M
10F + 5M
0
100
200
300
400
500
600
700
800
900
Embryos 1st Instar 2nd Instar 3rd InstarDevelopmental Phase
5F + 5M
10F + 5M

Gravitational and Space Biology Bulletin 19(2) August 2006 135
EFFECTS OF ALTERED GRAVITY ON IDENTIFIED PEPTIDERGIC NEURONS OF THE CRICKET
ACHETA DOMESTICUS.
Uta Kirschnick1, Hans-Jürgen Agricola
1 and Eberhard R. Horn
2
1 University of Jena, Department of Cell Biology, Philosophenweg 12 , 07743 Jena, Germany
2 University of Ulm, Gravitational Physiology, Albert-Einstein-Allee 11, 89081 Ulm, Germany
The development of organisms requires a permanent
interplay between genes and environment. Identified
neurons that are common in insects are excellent models
to determine the specific contribution of these factors for
the development of the nervous system. The aim of the
study was to investigate effects of microgravity (µg) on
identified peptidergic neurons in a cricket (Acheta
domesticus) (1) if eggs were fertilized in space and (2) if
the period of neuronal proliferation took place in µg.
Proliferation is mainly completed after 50% of embryonic
development. Thus, an 8 days lasting µg-exposure was
sufficient to answer the question. The 10 days lasting
Italian Soyuz taxi flight ENEIDE to ISS in April 2005
was used.
We chose neurons immunoreactive (ir) to allatostatin
(AST), crustacean cardioactive peptide (CCAP), and
perisulfakinin (PSK) because these neuropeptides are
involved in developmental and neuronal processes
(Homberg 1994). AST inhibits the synthesis of juvenile
hormone; AST neurons are usually equipped with short
axons; rarely do they extend to more than 4 segments
(Fig. 1). CCAP has myotrope function with strong effect
on heart activity; it probably contributes also to the
regulation of molting. PSK is a myotrope neuropeptide; in
addition, effects on the central nervous system were
described. Due to its wide projections (Fig. 2), P-PC1
neurons probably exert a modulating effect on neuronal
activity.
We considered these peptidergic neurons as model
structures to study the sensitivity of the topological
organization within the central nervous system to µg
because it is very unlikely that gravity effects on the
development of topological organization depends on
specific neurotransmitters. This assumption might not be
correct if the investigated neurons have mono- or
polysynaptic input from gravity receptors such as the
position sensitive interneuron PSI (cf. Sakaguchi and
Murphey, 1983).
The experiment CRISP-2/ENEIDE was a follow-up
study of the Neurolab experiment CRISP (Crickets in
Space). Then post-flight observations on cricket larvae
included neuroanatomical, neurophysiological, and
behavioral studies of different developmental stages. All
stages as well as embryos used in CRISP/Neurolab had a
fully developed set of neurons at onset of the mission.
The main observations were absence of significant
behavioral modifications and anatomical changes of
cerebral peptidergic neurons, but a significant depression
of the sensitivity of the PSI (Horn, 2003; Horn et al.,
2002).
A special experimental container was developed for
inflight fertilization on ENEIDE. It consisted of an adult
chamber and larval chambers, separated by egg collectors
(Fig. 3). If females inseminated on ground had access to
Figure 1. Allatostatin A-PC2-ir neuron and its primary neurite
within the right hemisphere from a space (middle) and ground
(right) embryo. Post-flight fixation at 60-65% of embryonic
development. The mature A-PC2-ir group consists of 6 neurons
(left). At stage 60-65% embryos, only 4 out of the 6 neurons are
visible in both, flight and ground embryos.
Figure 2. Basic projection pattern of a PSK PC1,2-ir neuron.
Figure 3. Cricket container for in-flight fertilization. On top the
adult chamber (CC-AC), on bottom 2 larval chambers (CC-
LC1,2); in-between 3 egg collectors (CC-EC1,2,3). They were
filled with vermiculite; 2.5 days after launch, females had access
to the vermiculite for 20 hrs to deposite eggs. Inset: individual
egg collector with deposition area on top.
the vermiculite within the egg collectors, they deposited
their eggs. During this process, the eggs were fertilized.
After termination of the 8 days lasting µg-exposure, the
nervous system was taken from embryos or, after hatch-

U. Kirschnick, H. Agricola and E.R. Horn – Effects of altered gravity on identified peptidergic neurons
136 Gravitational and Space Biology Bulletin 19(2) August 2006
ing, from the 1st instars for immunocytochemical staining
of neurons. The 1st fixation was done 6 hours after landing
in Kazakhstan, a 2nd
fixation 34 hours later in Moscow,
and a 3rd
fixation after hatching and testing of the geotaxis
of the 1st instars. A total number of 112 embryos and 1
st
larvae from µg-fertilization were available, and 103 from
on-ground fertilization.
The ENEIDE flight revealed that after in-µg
fertilization, AST-ir-, PSK-ir- and CCAP-ir-neurons
developed as after on-ground fertilization with respect to
the location of somata and arborizations. This held not
only for cerebral (Fig. 1), thoracic or abdominal (Figs.
4,5) neurons with short neurites, but also for PSK-ir
neurons that project throughout the whole nervous
system, with cell somata lying in the protocerebrum and
arborizations within the cerebral, thoracic and abdominal
ganglia (Figs. 2,6).
Figure 4. Somata of CCAP-ir neurons C-BM1 in the abdominal
ganglia (AG) from 55% embryos (top and middle) and 1st
instars (bottom). Left: Basic C-BM1-ir projection patterns.
Figure 5. Location of 2 types of CCAP-ir neuron in the
terminal (last) ganglion of 1st instars. Neurons at the rostral
corner of the ganglion: C-BM1+2-ir neuron. - Neurons at the
median caudal location: C-TG neuron. Note the clear similarity
of the arborization and the projection pattern of the flight and
ground embryos. - Calibration 100 µm.
Thirty-three space embryos were reared up until
hatching. They hatched about 1.5 days earlier than the
ground reared embryos. The space larvae revealed no
abnormal geotactic behaviour compared to ground larvae;
however, the quantitative analysis concerning walking
velocity and
curvature of walking trajectories is not yet completed.
These studies demonstrate that developmental
processes within the nervous system of insects are
obviously under a dominant genetic control. This does not
exclude an influence of epigenetic factors on other
developmental processes in insects including the nervous,
motor and sensory systems. The earlier hatching of the 1st
larvae after inflight fertilization makes this hypothesis
very likely.
Supported by DLR: grant 50WB0323 to Horn.
Figure 6. PSK PC1,2-ir neurons from two 1st larvae, after
fertilization in µg (left) or on ground (right). Top: Somata of
PC1,2-ir neurons in the brain. Middle: Projections of the
neurons at the level of the 6th abdominal ganglion. Bottom:
Projections of the neurons within the terminal ganglion (cf. Fig.
5a). Calibration 50 µm (top, middle), 100 µm (bottom).
REFERENCES
Homberg, U. 1994. Distribution of neurotransmitters in
the insect brain. Gustav Fischer, Stuttgart Jena New York.
Horn, E.R. 2003. The development of gravity sensory
systems during periods of altered gravity dependent
sensory input. Adv. Space Med. Biol. 9:133-171.
Horn, E., Agricola, H., Böser, S., Förster, S., Kämper, G.,
Riewe, P., and Sebastian, C. 2002. Crickets in Space:
Morphological, physiological and behavioral alterations
induced by space flight and hypergravity. Adv. Space
Res. 30:819-828.
Sakaguchi, D.S., and Murphey, R.K. 1983. The
equilibrium detecting system of the cricket: physiology
and morphology of an identified neuron. J. Comp.
Physiol. 150:141-152.

Gravitational and Space Biology Bulletin 17(2) June 2004 137
COUNTERMEASURES TO THE EFFECTS OF GRAVITY ON THE SKULLS OF HUMAN INFANTS. R. Lee
1, J. English
1, J. Duke
1 , and J. Teichgraeber
2.
1Dept. of Orthodontics, Dental Branch,
2 Division of Pediatric Surgery, Department of Surgery, Medical School,
University of Texas Health Science Center at Houston, Houston, TX.
The effects of altered gravity on the skeleton have
been studied mainly in weight-bearing bones, so few
gravitational biologists know how Earth’s gravity shapes
the skulls of human infants. Skull deformations due to
prolonged exposure to the g vector in one direction are
well-known to craniofacial experts, and include brachy-
cephaly, a flattening of the back of the skull, and posterior
deformational plagiocephaly (PDP), in which the infant’s
skull, viewed from the top, has the shape of an oblique
oval, which can be enclosed in a parallelogram (Teich-
graeber et al., 2002; Fig. 1A, 1B). This shape results from
a flattening of either the left or right occipito-parietal re-
gion of the skull, causing bossing, or bulging, of the con-
tralateral occipital region and the ipsilateral frontal bone.
The ipsilateral ear and malar eminence, or cheekbone, are
displaced anteriorly as are the eye and the temporo-
mandibular joint (TMJ).
PDP is associated with a number of factors related to
intrauterine constraint: loss of amniotic fluid, premature
and multiple births, the birthing process itself and post-
natal positioning. Some infants have a torticollis, a short-
ened muscle affecting head positioning. Recalcitrant
cases require surgery, and increased levels of TGF β2 and
β3 have been found in affected sutures (Lin et al., 1997).
An infant’s skull is designed to be deformed, being
composed of plates of intramembranous bone connected
by tough fibrous membranes or sutures, a construction
allowing for compression of the infant’s head during birth
and postnatal growth of the bones. In craniosynotosis,
sutures fuse prematurely, resulting in a malformed skull
requiring surgery. Fusion of the lambdoid suture between
the occipital and parietal bones, like PDP, results in
bossing of the contralateral occipital region. The frontal
bossing, however, is on the contralateral side, and the ear
and malar eminence are not displaced (Habal et al., 2003).
PDP includes a facial asymmetry, continuing into
adulthood, due to rotation of the cranial base and the re-
sulting anterior displacement of the TMJ and consequent
asymmetric mandibular growth (St. John et al., 2002).
A. B. C.
Figure 1: A. Normal head shape. B. Parallelogram
shape of head of infant with PDP. C. Thick line indicates
how DOC helmet therapy changes growth of regions of
the skull.
Disturbingly, a number of developmental anomalies are
associated with PDP (Habal, et al., 2003). Delays in both
cognitive and psychomotor development have been noted
and auditory processing disorders affecting speech and
cognition have been found. CT scans have revealed an
accumulation of fluid in the frontal region of the cranium.
In light of these problems in infants with PDP, the
increase in incidence from 1 in 300 to 1 in 10 since 1992
is alarming (Teichgraeber et al., 2002). In 1992, the
American Academy of Pediatrics urged parents to place
sleeping infants on their backs to prevent Sudden Infant
Death Syndrome. The message was reiterated in 1994 in
the AAP’s “Back to Sleep” campaign. As a result, the
incidence of SIDS decreased by 45%, but craniofacial
centers were inundated with parents seeking treatment for
children with misshapen skulls. In 1995, the Division of
Pediatric Surgery at the University of Texas Medical
School in Houson treated two infants with PDP; in 1996,
they treated 54 (Teichgraber et al., 2002).
Some pediatricians advocate a wait and see attitude, but
craniofacial specialists prefer early intervention, while the
skull still has sufficient growth potential to correct the
deformations (Teichgraber et al., 2002). In some cases,
the problem can be corrected with repositioning the infant
during sleep, so that the head does not rest on the
flattened portion of the skull. Increased tummy time, in
which the infant is placed prone under parental
supervision, also alleviates pressure on the occipital
region, and encourages head raising, thus strengthening
the neck muscles. If a torticollis is involved, physical
therapy is required to stretch the infant’s muscles in order
to reposition the head. Time spent in car seats, carriers or
swings should be limited.
Simple countermeasures are not always successful in
skull reshaping, and in these cases a molding helmet such
as Dynamic Orthotic Cranioplasty (DOC) is used (Fig.
1C). The custom molded DOC helmet consists of a semi-
rigid styrene shell and inner polyurethane liner, and
relieves pressure on the flattened part of the skull
allowing growth in that region. Parts of the skull that
developed bossing or bulging are constrained by the
helmet, and growth in those areas thus restricted. DOC
helmet treatment has disadvantages; the helmet must be
worn 23 hrs/day,requires weekly monitoring/adjustment,
and is costly. Sometimes, the helmet does not correct the
deformation, and surgery is required.
A study of 125 patients treated with DOC helmet
therapy (Cranial Technologies, Inc., Tempe, AZ) at the
UT School of Medicine found that asymmetries of the
cranial vault (CV) and the cranial base (CB) were signif-
icantly reduced (41.6% and 40.2%) by treatment (Teich-
graber et al., 2002). Orbitotragial (OT) asymmetry was
improved by 18.7%. Recently, the question of retention
of improvement was addressed by re-examining these
children 5 years after their treatment.

R. Lee et al. – Countermeasures to the Effects of Gravity on the Skulls Of Human Infants
138 Gravitational and Space Biology 19(2) August 2006
MATERIALS AND METHODS
14 male and 14 female patients from the previous study
were re-examined. 79% had right side flattening. Mean
start age for treatment was 6.7 months, and mean treat-
ment time was 6.2 months. 7.1% were born prematurely,
3.6% were from breech births, and 3.6% from multiple
births. 7.1% had a torticollis. These percentages are
similar to those in the general population. Skull
measurements used anthropometric landmarks (Figure 2):
frontozygomatic point (FZ)-suture of frontal and zygo-
matic bones, midpoint of outer edge of eye orbit; euryon
(EU)-most lateral point on the side of the cranium-from
EU to EU is the widest portion of the skull; exocanthion
(EX)-where eyelids meet in outer corner of eye; tragus
point (TR)-notch in ear cartilage in front of ear; inion
point (IN)- the most prominent point on the external
occipital protuberance in the median sagittal plane. CV
asymmetry was the distance from left FZ point to right
EU point minus the distance from the right FZ point to the
left EU point. Orbitotragial depth asymmetry was the
distance from right EX point to right TR point minus the
left EX point to the left TR point. CB asymmetry was IN
point to right TR minus IN to left TR. Asymmetries were
compared to those immediately post treatment. Facial
asymmetry and malocclusions were also tabulated.
Figure 2: Points for anthropometric measurements.
RESULTS Measurements of asymmetry immediately after treatment
and 5 years later are shown in Table 1. Only the %
change in CB asymmetry was significant.
Table 1: Asymmetry changes 5 years posttreatment.
Measurement Asymmetry (mm) % Change
Cranial Vault 5.57 6.89 +23.7% (n.s.)
Orbitotragical
Depth
2.89 2.25 -22.2% (n.s.)
Cranial Base 3.64 8.11 +122.5%
(p< .004)
In terms of facial asymmetry, 19 of 28 (67.86%) had chin
point deviation to the unaffected side. Deviation of the midline was observed in 67.86%, with upper and lower
jaws not necessarily congruent. Posterior dental crossbites
were noted in 17.89% of the sample. Molar relationships
were examined in 17 patients, and the incidence of Class
II (lower molar posterior to upper) and Class III (lower
molar anterior to upper) relationships was greater than
normal.
DISCUSSION
DOC helmet therapy effectively corrects cranial a-
symmetry, especially CV and CB, but after 5 years,
significant relapse in CB occurs, and may include jaw
asymmetries leading to misaligned teeth, TMJ problems,
and esthetic concerns. Reasons for relapse are unknown,
but likely involve skull growth patterns. CB asymmetry is
based on IN, the middle of the back of the head, and
improves with DOC helmet treatment because cranial
flattening improves. The ear (TR) remains forward due to
the rotated CB, the anterior portion of which continues
growing, increasing the posttreatment asymmetry. Less
than 5% of the general population has noticeable facial
asymmetry.
Since CB asymmetry recurs, producing facial asym-
metries and malocclusions, greater efforts at prevention of
PDP are needed, including increased awareness on the
part of pediatricians, family members and other care-
givers. Parents must realize that the shape problem of
their baby’s skull may not resolve with time, and needs to
be addressed with repositioning from an early age.
The AAP 1992 recommendations on sleep position
allowed side sleeping, but in October, 2005, side sleeping
was recommended against, a change that is likely to cause
a further increase in PDP and makes the need for ed-
ucation even more urgent. The cure for PDP is preven-
tion, and the key to prevention of PDP is education.
ACKNOWLEDGEMENTS
Supported by UTDB Dept of Orthodontics.
REFERENCES
Habal MB. Leimkuehler T. Chambers C. Scheuerle J.
Guilford AM. Avoiding the sequela associated with defor-
mational plagiocephaly. Journal of Craniofacial Surgery
14: 430-7, 2003.
Lin KY. Nolen AA. Gampper TJ. Jane JA. Opperman LA.
Ogle RC. 1997. Elevated levels of transforming growth
factors beta 2 and beta 3 in lambdoid sutures from
children with persistent plagiocephaly. Cleft Palate-
Craniofacial Journal. 34:331-7.
St John D. Mulliken JB. Kaban LB. Padwa BL. 2002.
Anthropometric analysis of mandibular asymmetry in
infants with deformational posterior plagiocephaly.
Journal of Oral & Maxillofacial Surgery. 60: 873-7.
Teichgraeber, JF, Ault, JK, Baumgartner J, Waller, A,
Messersmith M, Gateno J, Bravenec B, Xia, J.
Deformational posterior plagiocephaly: diagnosis and
treatment. Cleft Palate Craniofacial J. 2002: 39: 582-586.
eu eu
tr tr
fz fzex ex ex
tr
in

Gravitational and Space Biology Bulletin 19(2) August 2006 139
NUTRIENT DIFFUSION THROUGH ARTICULAR CARTILAGE: DEVELOPMENT AND USE OF A
MODEL SYSTEM.*
Candace Marshall, Ruby Flowers, Neeta Goli, Marianne Vandromme, Douglas Paulsen and Brenda Klement
Dept. of Anatomy and Neurobiology, Morehouse School of Medicine, Atlanta, GA 30310
Cartilage is an avascular connective tissue composed of
cells (chondrocytes) suspended in a gel-like matrix that
gives cartilage its ability to bear mechanical stresses
without distortion. The main function of cartilage is to
provide a shock-absorbing and sliding area for joints thus
facilitating bone movements. Components of cartilage
extracellular matrix (ECM) include type II collagen,
glycosaminoglycans (GAGs) and other molecules cross-
linked to give firmness. There is also a substantial
amount of water bound to the negative charges of GAGs
which give the tissue a firm, gel-like consistency allowing
it to act as a biomechanical spring and shock absorber
(Paulsen, 2000).
To maintain structure and function, the chondrocytes
must obtain needed nutrients and molecules via diffusion
through the abundant matrix (Pelletier et al,. 2000). The
composition of the ECM is known to change with age and
with diseases such as arthritis (Bayliss et al., 2000). A
change in composition of the ECM would result in a
change in the arrangement or architecture of the ECM
components, and potentially result in a change in nutrient
diffusion. Disruptions in nutrient flow and availability
have been associated with articular disorders such as
arthritis (Kuettner 1992).
Astronauts experience a dramatic headward fluid shift
in microgravity, which would be expected to decrease the
hydration level of cartilage in the joints of the lower
extremities, and subsequently compromise mobility. To
our knowledge, nutrient flow has not been evaluated in
animal or human articular cartilage during spaceflight.
The goal of this project was to develop a model system
that could be used to assess nutrient diffusion through
articular cartilage matrix.
In this model system, fluorescent labeled glucose (2-(N-
(7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino)-2-
deoxyglucose), referred to as 2-NBDG, was used. These
initial studies examined only the non-metabolized 2-
NBDG. Future studies will explore the 2-NBDG
metabolized by chondrocytes and the 2-NBDG that
remains associated with the ECM. The model can be
used to assess 1) the 2-NBDG that diffuses into cartilage
within a certain time period or 2) the 2-NBDG that
diffuses out of cartilage after saturation with 2-NBDG.
The ultimate objective is to use this model to determine if
changes in diffusion reflect changes in the composition
and architecture of the ECM occurring with disease, age,
and the fluid shifts associated with spaceflight.
In the original model system, 50µm slices of bovine
articular cartilage were used. The slices were incubated
in 50µM 2-NBDG at room temperature. The longer the
tissue was incubated, the more 2-NBDG diffused into the
tissue, and reached a maximum by 1 hour (figure 1). To
* presented at 2004 meeting
quantitate the amount of 2-NBDG that diffused out of
each cartilage section, the sections were soaked in 2-
NBDG for one-hour then incubated for one-minute
intervals in phosphate buffered saline (PBS). The
quantity of fluorescent 2-NBDG in each PBS aliquot was
measured with a Cytofluor 96-well plate
spectrophotometer. The sum of the fluorescence was
determined for each cartilage section. Cartilage sections
were weighed (wet weight) then dried overnight at 60°C
and re-weighed (dry weight). The amount of non-
metabolized 2-NBDG that diffused out of the cartilage
sections was normalized to section wet and dry weights.
The location of 2-NBDG can be seen in cartilage
sections using confocal microscopy. After soaking in 2-
NBDG the entire tissue fluoresces (Figure 2A). After the
non-bound 2-NBDG has diffused out of the ECM,
metabolized 2-NBDG is seen within the chondrocytes and
some appears to remain associated with the matrix (figure
2B). Quantitation of the metabolized and matrix
associated 2-NBDG has not yet been conducted and is not
part of the current model system. However, this appears
to be an exceptionally interesting aspect and future assays
will be conducted to assess this component as well.
Figure 2. Confocal images of a cartilage section soaked in 2-
NBDG (A) and approximately 1 minute after it had been rinsed
(B). The lighter regions represent areas where the fluorescent
2-NBDG is located.
This model system was used to assess 2-NBDG
diffusion in cartilage obtained from hindlimb unloaded
(HLU) rats. Rat HLU has been used extensively to model
some of the aspects of microgravity such as bone and
muscle loss and headward fluid shift. The tails of HLU
rats were anchored above the cage so that the hindlimbs
were elevated off the cage floor. The tails of the control
rats were also anchored but all four feet remained on the
0 30 60 90 120 150 180 2100
1
2
3
4
5
6
Soak time in 2-NBDG (minutes)
2-N
BD
G (
µµ µµM
)
Figure 1. Amount of 2-
NBDG that diffused into
cartilage sections after
different incubation
times. N≥7, error bars
represent standard error.
A B

C. Marshall et al. – Nutrient Diffusion through Cartilage
140 Gravitational and Space Biology Bulletin 19(2) August 2006
cage floor, tethering (TH) them within the cage. The tails
of normally caged rats (NC) were not anchored, and their
movement was unrestricted. Articular cartilage was
collected from NC rats and rats that had been HLU and
TH for 7 or 21 days. Rat tibias were cleaned of soft tissue
and the cartilage from the tibial plateaus were cut in
40µm sections. After incubating in 2-NBDG for 1-hour,
the amount that diffused from the sections was collected
and quantitated as described above (figure 3).
Figure 3. Amount of 2-NBDG that diffused from cartilage
sections obtained from hindlimb unloaded, tethered and
normally caged rats. The amount of 2-NBDG that diffused from
cartilage sections collected from each rat were averaged. The
mean was based on the number of rats within an experiment.
(A) The total amount of 2-NBDG that diffused from the sections.
(B) The amount of 2-NBDG diffusion normalized to wet weight
of the tissue section. N=4 for normally caged rats, N=3 rats for
each group in the 7 day experiment, N=8 rats for each group in
the 21 day experiment. The 21 day data is a combination of
data from 2 experiments. Error bars represent standard error.
There was no significant difference in 2-NBDG
diffusion between cartilage obtained from HLU and TH
rats at 7 days. Cartilage from both 7-day groups showed
less 2-NBDG diffusion than the NC and 21-day
experiment rat cartilage, but was not significant. This
decrease may be due to the initial stress associated with
the experiment, but cannot be determined until additional
parameters are evaluated. The percent of the sections
composed of fluid was the same for the 7-day and NC
groups (figure 4), indicating that the difference in
diffusion was not due to a difference in hydration levels.
In the 21-day experiments the amount of 2-NBDG
diffusion was 42% less in the cartilage sections obtained
from HLU rats than from the TH rats, although this was
not statistically significant (figure 3A). Multiple
experiments were conducted and each showed the same
trend. Combined data from two of the experiments are
shown in figure 3. A significant difference is seen in the
percent of the cartilage sections composed of water in the
HLU and TH rats, indicating that the hydration level of
the cartilage decreased during extended HLU. The
decrease in fluid content may explain the reason for the
reduced 2-NBDG diffusion in the cartilage from the HLU
rats. The amount of 2-NBDG normalized to wet weight
(figure 3B) or dry weight (data not shown), shows the
same pattern as the total 2-NBDG, in all groups.
The headward fluid shift during HLU reduces the fluid
level in the tibial plateau cartilage by 21 days. Diffusion
of 2-NBDG appears to be dependent on the hydration of
the cartilage tissue and may effect the availability of
nutrients within the ECM. This could significantly
impact the structure and function of articular cartilage.
Future studies will determine if the reduction in nutrient
availability has an impact on the health of the
chondrocytes. Expanding the scope of the model system
to include evaluation of metabolized 2-NBDG and 2-
NBDG associated with the ECM will also enable a more
comprehensive evaluation of diffusion in arthritic, and
aged cartilage as well as that from animals subjected to
microgravity, hypergravity and their respective model
systems.
REFERENCES Bayliss, M., Howat, S., Davidson, C., Dudhia, J., 2000.
The organization of aggrecan in human articular cartilage.
Evidence for age-related changes in the rate of
aggregation of newly synthesized molecules. J Biol Chem
275:6321-7.
Kuettner, K.E. 1992. Biochemistry of articular cartilage in
health and disease. Review, Clinical Biochemistry.
25(3):155-63.
Paulsen, D.F. 2000. Histology & Cell Biology:
Examination and Board Review, 4th ed., Lange/McGraw-
Hill, New York, pp.376.
Pelletier, J.P., Jovanovic, D.V., Lascau-Coman, V.,
Fernandes, J.C., Manning, P.T., Connor .JR., Currie
M.G., and Martel-Pelletier, J. 2000. Selective inhibition
of inducible nitric oxide synthase reduces progression of
experimental osteoarthritis in vivo: possible link with the
reduction in chondrocyte apoptosis and caspase 3 level.
Arthritis & Rheumatism. 43(6):1290-9.
ACKNOWLEDGEMENTS We thank Drs. Bayorh and Eatman and their laboratory
assistants for providing the tissue from the unloaded and
control rats. This work was supported by NASA grants
NAG3-2611 and NCC9-112 as well as RR-03034.
0
25
50
75
0 7 21
Unloaded TetheredNormal
*
Hindlimb Unloading (days)
wate
r as %
of
tota
l w
eig
ht Figure 4. Water content
of the cartilage sections
used in figure 3 (wet
weight – dry weight). *
significantly different from
tethered (p=0.0307).
Error bars represent
standard error.
0
1
2
3
4
5
6
7
8
Unloaded
Tethered
NormalA
2-N
BD
G (
µµ µµM
)
0
2
4
6
8
10
12
0 7 21
B
Hindlimb Unloading (days)
2-N
BD
G/w
et
weig
ht
( µµ µµM
/mg
)

Gravitational and Space Biology Bulletin 19(2) August 2006 141
HYPERGRAVITY INDUCES DAMAGE TO ROD PHOTORECEPTORS* A.J. Barnstable
1, A.R. Tink
1,2, S. Viviano
1, L. Baer
3, C. Wade
3, C.J. Barnstable
1 and J. Tombran-Tink
1,2.
1Dept. Ophthalmology, Yale University School of Medicine, New Haven, CT,
2Pharmaceutical Sciences, UMKC,
Kansas City, MO and 3Ames Research Center, Moffett Field, CA.
The retina is an outgrowth of the embryonic forebrain
neuroepithelium and provides an excellent model of brain
development and function. The neurons of the retina,
particularly those in the photoreceptor layer, are very
sensitive to environmental perturbations, and is thus a
good sensor for adverse conditions.
The adult retina has a relatively simple laminar
structure containing six neuronal and two glial cell
classes. Light is detected by rod (dim light) or cone
(bright light) photoreceptor neurons in the outer layer of
the retina. The light sensing portion of these cells contain
opsin visual pigment proteins that initiate the visual
transduction cascade that amplifies photon signals into
changes in membrane potential and neurotransmitter
release. The photoreceptor outer segments depend on
intimate contact with both glial cells and the adjacent
retinal pigment epithelial cells for both metabolic support
and regeneration of molecules needed for the visual
transduction cascade.
Retinal rod photoreceptor degeneration can result from
a variety of environmental and genetic factors. Retinitis
Pigmentosa (commonly known as night-blindness) is an
inherited condition in which rod photoreceptors
degenerate. As a secondary effect, cone photoreceptors
are then lost. At least 40 genes have been identified as
causing RP, but no effective therapy has yet been
approved. A number of animal models of RP exist but
there is a strong need for other models that can be
produced without complex genetic or surgical
intervention.
We have shown that the environment encountered in
space shuttle flight can disrupt normal retinal
development and mimic stimuli that induce similar retinal
degenerations (Tombran-Tink and Barnstable, this
volume). We have now examined eyes from adult
animals maintained in a hypergravity environment and
find that these show signs of rod degeneration as well.
Groups of pregnant rats were maintained at 2.0 xg for
approximately 14 days by centrifugation with a normal
lighting schedule. Age- and weight-matched stationary
controls were housed in the centrifuge rotunda, which
allowed them to be exposed to the same environmental
conditions as the hypergravity animals. Eyes from the
adult rats were fixed in formalin, cryoprotected, frozen
and cryosectioned. Sections were labeled with a variety
of cell type-specific antibodies including those
recognizing rod photoreceptor cell bodies, inner segments
and outer segments.
Eyes from rats maintained at 2.0 xg showed loss of rod
photoreceptors with varying degrees of severity. This is
most clearly noted in Figure 1 that shows substantial loss
of rod photoreceptor bodies in the outer nuclear layer. In
* presented at 2004 meeting
the most extreme cases the cell body layer was reduced to
approximately 20% of its normal thickness.
The outer segments were shorter and much more
disorganized although inner segments showed less
change. Stationary control retinas showed normal
photoreceptor morphology, suggesting that the damage
seen was due to the increased g-force.
We measured the thickness of various layers from
cryostat sections of a series of experimental and control
animals. Attempts were made to measure the same areas
in each animal but some variation remained.
Nevertheless, comparisons of relative layer thicknesses
are not affected by this variation. Figure 2 shows the
decrease in ONL thickness in a series of hypergravity
treated and control animals. In all the hypergravity
treated animals the outer nuclear layer is thinner than the
inner nuclear layer, a situation never seen in normal adult
rats. Other cell layers across the retina showed essentially
no change.
The selective loss of rod photoreceptors is similar to
that seen in a number of mutant strains of mice that serve
as models for the human disease of retinitis pigmentosa
(Figure 3). Rd1 mice show an early and rapid loss of rod
photoreceptors such that by 5-6 weeks of age they have
lost essentially all of these cells.
How does hypergravity cause rod photoreceptor
degeneration? One obvious factor to be considered is that
the increase in g-force causes a damaging increase in
Figure 1. After exposure to a 2X g environment for
2 weeks the retinas showed severe damage in the
outer retina. The outer nuclear layer normally
contains about 10 rows of cell bodies (SC215).
After hypergravity exposure this is reduced to
approximately 3 rows of cell bodies (HG203).
ONL, Outer Nuclear Layer; OPL, Outer Plexiform
Layer; INL, Inner Nuclear Layer; IPL, Inner
Plexiform Layer; GCL, Ganglion Cell Layer.

A. Barnstable et al. – Hypergravity damages rod photoreceptors
142 Gravitational and Space Biology Bulletin 19(2) August 2006
Figure 2. Summary data for a series of hypergravity
treated (upper histogram) and control (lower histogram)
eyes. Sets of data are for individual animals. All
measurements of layer thicknesses are in microns. Layer
naming is as given in the legend to figure 1.
intraocular pressure. This is unlikely to be the whole
explanation, however, because we observed no loss of
retinal ganglion cells. Increased intraocular pressure is
frequently associated with ganglion cell death in
glaucoma, a disease with little if any effect on rod
photoreceptors. It remains possible, however, that
increased pressure in some way affects the choroidal
blood supply that provides oxygen and nutrients to the rod
photoreceptors but not the retinal blood supply that
provides oxygen and nutrients to the ganglion cells. At
present we have no information about similar
degenerative changes that might occur in other brain
regions.
Further studies are needed to understand the time
course and mechanism of the rod photoreceptor cell death
we have observed. The non-invasive treatment of these
animals provides a valuable model with which to test
novel therapeutic strategies to combat blinding diseases.
One advantage over existing genetic models is that the
degeneration may be induced at different ages, something
that might better mimic a number of human blinding
diseases. Specifically, this model will be valuable for
testing the role of novel neuroprotective factors that can
block rod photoreceptor cell death.
Although short term increases in g-force are
experienced in both jet flight and space flight launches,
there are few studies of the effects of long term increased
g-force on the nervous system in general and the visual
system in particular. Knowledge of how increased g-
force affects cell metabolism and survival in the visual
system, and elsewhere in the nervous system, is important
as we think about exploration of different space
environments.
We thank Louis Ostrach (NASA, Washington, DC) for
advice and help with procuring tissue samples. Supported
by NIH, RPB Inc., and the Connecticut Lions.
wt rd
1
Figure 3. Sections of postnatal day 35 wild-type
(wt) and mutant (rd1) retinas showing the loss of
rod photoreceptors in the mutant. Rd1 mice carry a
mutations in the beta subunit of the rod
photoreceptor-specific cGMP phosphodiesterase.

Gravitational and Space Biology 19(2) August 2006 143
A GLOBAL TRANSCRIPTIONAL ANALYSIS OF STREPTOCOCCUS PNEUMONIAE IN RESPONSE
TO LOW-SHEAR MODELED MICROGRAVITY
Christopher Allen1, Cristi Galindo
1, Nina Williams
1, Utpal Pandya
1, Ashok Chopra
1, and David Niesel
1
1
University of Texas Medical Branch, Galveston, TX 77555 USA
Long-term habitation in space exposes humans to a
variety of environmental stresses not encountered on earth such as ionizing radiation and microgravity. Decreases in
immune function and increases in microbial growth and
antibiotic resistance during space flight could create an
increased health risk to crewmembers (Sonnenfeld et al.,
1990; Mishra et al., 1992). Streptococcus pneumoniae is
an upper respiratory pathogen which causes pneumonia,
meningitis, and bacteremia. S. pneumoniae has
previously been isolated from crewmembers and could
present a potential health hazard during extended
spaceflight (Cioletti et al., 1991). To facilitate ground-
based microgravity analogue studies, NASA developed
high aspect ratio vessels (HARVs) which model low-
shear microgravity conditions (LSMMG). The LSMMG
environment is generated through the continual rotation of
cell cultures in fluid-filled vessels which minimize
prolonged exposure to gravity through the randomization
of gravity vectors. Recently, these ground-based models
have been used to study the effects of LSMMG on
bacteria to understand the physiological and molecular
changes they undergo in the presence of microgravity as
encountered during spaceflight.
This study utilized S. pneumoniae as a Gram-positive
bacterial model to investigate how LSMMG affects
environmental stress resistance and global transcriptional
activity in opportunistic bacterial pathogens. DNA
microarray analysis was carried out on S. pneumoniae
strain TIGR4 after cultivation in HARVs under LSMMG
and compared to 1 x g and static conditions (Figure 1).
Figure 1. HARV Vessel Orientations for Growth.
Microarrays were acquired from The Institute for
Genomic Research (TIGR) Pathogen Functional
Genomics Resource Center (PFGRC). They were
designed based on the genomic sequence of the TIGR4
strain and contain amplicons representing 2131 open-
reading frames.
Stress analysis was carried out on TIGR4 cultures
grown to mid-logarithmic phase under LSMMG and
controls (1 x g rotating) followed by exposure to thermal
(55°C) and acid (pH 3.5) stress conditions (Figure 2).
Significant decreases (p ≤ 0.05) in both thermal and acid
stress resistance were found in LSMMG-grown cultures
after exposure for 30 minutes using a Student’s t test.
Similar results were found after exposure when stress
conditions were increased up to 1 hour (data not shown).
These results were in contrast to previous reports which
have shown increases in environmental stress resistance
by Salmonella enterica serovar Typhimurium and
Escherichia coli (aWilson et al., 2002; Lynch et al., 2004)
in response to LSMMG. These results may be indicative
of the different responses by Gram-positive and Gram-
negative bacteria to LSMMG. Alternatively, the results
may reflect the different phenotypes exhibited by enteric
and respiratory pathogens which colonize and persist in
distinct niches within the host.
Thermal Stress Resistance
Acid Stress Resistance
Figure 2. Thermal and Acid Stress Resistance in Response to
LSMMG.
In order to understand what genes may be involved in
bacterial LSMMG-mediated responses at the
transcriptional level, global transcriptional analysis was
carried out using mid-logarithmic phase cultures grown
under LSMMG and control cultures (1 x g rotating, 1 x g
static). A stringent analysis of the array data was carried
out using multiple analysis methods (Genepix Pro 6.0,
Spotfire 7.3, SAM, and ANOVA) and a threshold of ≥
1.5-fold was used to identify differentially expressed
genes. Results were confirmed by quantitative real time

C. Allen et al. – A Global Transcriptional Analysis of Streptococcus pneumoniae in Response to Low-Shear
Modeled Microgravity
144 Gravitational and Space Biology 19(2) August 2006
RT-PCR of six representative genes differentially
expressed under LSMMG. Genes were chosen which
were differentially expressed ≥ 1.5-fold in response to
LSMMG, represented different functional categories, and
located within different regions within the genome.
Pneumococcal genes (81 total) which were found to be
responsive to LSMMG represented different functional
groups located throughout the genome (Table 1). The
statistical analysis program, Significance Analysis of
Microarrays (SAM), confirmed the statistical significance
of these differentially expressed genes based on three
independent experiments.
Functional Category LSMMG-Responsive Genes
Antibiotic Resistance 1
Cell Envelope 11
DNA Repair/Recombination 4
Metabolism 13
Signaling 3
Stress Response 1
Transcriptional Regulation 4
Transporters/Ion Channels 8
Hypothetical/Unknown 36
Table 1. Functional Gene Groups Significantly Altered by
LSMMG.
Interestingly, LSMMG-responsive genes were found to be
predominantly down regulated in response to LSMMG.
Previous studies have shown that global transcriptional
profiles of Gram-negative enteric bacteria contain both
up- and down-regulated genes after growth under
LSMMG (bWilson et al., 2002). This consistant pattern of
down-regulation seen in S. pneumoniae represents a
distinct response by this Gram-positive respiratory
pathogen compared with the more extensively
characterized Gram-negative enteric pathogens.
Hierarchical clustering analysis (Cluster/Treeview,
CLUSFAVOR 6.0, and Spotfire 7.3) revealed several
gene clusters displaying similar expression patterns.
These clusters consisted of different genes from diverse
functional groups. Currently, how these genes function in
response to LSMMG remains unknown. Likewise, a
common mechanism to explain these regulatory changes
in response to LSMMG remains uncharacterized. Further
studies will need to be carried out to identify LSMMG-
responsive regulators.
Controls for the LSMMG experiments included both a
1 x g rotating control and a 1 x g static control (Figure 1)
as a secondary filter to identify LSMMG-responsive
genes. SAM analysis of genes differentially expressed
between static and rotating controls revealed 147
differentially expressed genes. Among these genes, 46
were also altered between 1 x g and LSMMG conditions
while 9 were altered between static and LSMMG
conditions. The results from this analysis clearly indicate
that the choice of control has a significant impact on the
assignment of LSMMG-responsive genes and possibly
physiological properties. Further, the presence of other
external forces such as shear and rotation exert individual
effects on cultured cells independent of LSMMG.
Overall, these results indicate that LSMMG represents
a unique environment that can alter a bacterium’s
transcriptional profile and physiological state.
Investigating how this unique environment can lead to
new properties will allow us to better understand the
impact of LSMMG on microbial virulence and the health
risks these changes may create during long-term space
habitation.
REFERENCES
Sonnenfeld, G., Mandel A.D., Konstantinova I.V., Taylor
G.R., Berry W.D., Wellhausen S.R., Lesnyak A.T., Fuchs
B.B. 1990. Effects of Space Flight on Levels and Activity
of Immune Cells. Aviat. Space Env. Med. 61: 648.
Mishra, S., Pierson, D. 1992. Space Flight, Effects on
Microorgansims. In: Encyclopedia of Microbiology.Vol.
4. Academic Press, Inc. pp. 53-60.
Cioletti, L., Pierson, D., Mishra, S. 1991. Microbial
Growth and Physiology in Space: A Review. SAE
Technical Paper Series No.911512.
aWilson, J., Ott, C.M., Ramamurthy, R., Porwollik, S.,
McClelland, M., Pierson, D.L., Nickerson, C.A. 2002.
Low-Shear Modeled Microgravity Alters the Salmonella
enterica Serovar Typhimurium Stress Response in an
RpoS-Independent Manner. Appl Environ Microbiol
68(11): 5408-5416.
Lynch, S.V., Brodie, E.L., Matin, A. 2004. Role and
Regulation of sigma S in General Resistance Conferred
by Low-Shear Simulated Microgravity in Escherichia
coli. J Bacteriol 186(24): 8207-8212.
bWilson, J.W., Ramamurthy, R., Porwollik, S.,
McClelland, M., Hammond, T., Allen, P., Ott, C.M.,
Pierson, D.L., Nickerson, C.A. 2002. Microarray Analysis
Identifies Salmonella Genes Belonging to the Low-Shear
Modeled Microgravity Regulon. Proc Natl Acad Sci USA
99(21): 13807-13812.

Gravitational and Space Biology Bulletin 19(2) August 2006 145
PROTEOMIC RETRIEVAL FROM NUCLEIC ACID DEPLETED SPACE-FLOWN HUMAN
CELLS
D. K. Hammond1, T.F. Elliott
2, K. Holubec
2, T.L. Baker
2, P.L. Allen
3, T.G. Hammond
3 and J.E. Love
4
1Enterprise Advisory Services, Inc., Houston, TX; 2 Wyle Life Sciences, Houston, TX; 3V.A. Medical
Center and Tulane University Health Sciences Center, New Orleans; 4Human Adaptation and
Countermeasures Office, NASA, Johnson Space Center, Houston, TX.
Cells grown in space or in any hostile environment
with a necessity for preservation are subject to
decomposition and loss of valuable scientific
information. To maximize the science return from
flight samples, an optimized method was developed
to recover protein from samples that had been stored
for long periods of time at refrigerated temperatures.
This technique also allows multiple analyses on a
single cellular sample when frozen. Our group
specifically designed this for use with samples from
the International Space Station (ISS) (Love, et al.,
2003) at the same time that a different technique was
established by others (Rodrigo, et al., 2002). This
work extends our initial work, using additional
antibodies and cell lines, as well as corroborating
with histological staining.
Cell cultures were grown in American
Fluoroseal bags (TCM) either on the ISS or in flight
hardware on the ground, and treated at regular
intervals with an RNA stabilizing agent (RNAlater®,
Ambion, Austin, TX) or a formalin solution. The
samples were refrigerated for 3 months. RNA was
purified using an RNAqueous® kit (Ambion) and the
remaining RNA free supernatant was precipitated
with 5% trichloroacetic acid. The precipitate was
dissolved in SDS running buffer and tested for
protein content using a bicinchoninic acid assay
(Sigma, Dallas). Equal loads of protein were placed
on SDS-PAGE gels and transferred using Western
Blotting techniques (Towbin, et al., 1979). Protein
on the blots from preserved cells were analyzed using
horseradish peroxidase antibodies, with an ECL+
fluorescent stain (Amersham, Piscataway, NJ). The
Storm imager and Imagequant software (Amersham)
quantified the pixel volumes for rectangles of equal
size. Human renal cortical epithelial (HRCE) cells
(Cubano, et al., 2004; Cowger,et al., 2002) grown
onboard the ISS during Increment 3 and in ground
control cultures exhibited similar immunoreactivity
profiles for antibodies to the Vitamin D receptor
(VDR) (Fig 1), the beta isoform of protein kinase C
(PKCß) (Fig 2), and glyceraldehyde-3-phosphate
dehydrogenase (GAPDH) (Fig 3). The ground (Grd)
and flight (Flt) samples are presented on the graphs
using an untreated control to normalize the data. A
Vitamin D Receptor
0
20
40
60
80
100
120
140
Grd 3 Grd 6 Grd 9 Flt 3 Flt 6 Flt 9 Control
Perc
ent of C
ontr
ol
Figure 1 - Vitamin D Receptor (VDR) antibody stained
blots, prepared with equal loading of protein (5 µg/lane),
measured a protein with a molecular weight of 60-64kD.
Samples are from ISS Flt samples from days 3, 6 and 9 and
the Grd matched controls from the same days. An example
of one of the blots is shown below the graph, which is an
average of four blots.
PKC-beta
0
20
40
60
80
100
120
140
Grd 3 Grd 6 Grd 9 Flt 3 Flt 6 Flt 9 Control
Perc
ent
of
Contr
ol
Figure 2 - Protein Kinase CβII (PKCβII) antibody stained
blot, prepared with 6 µg/lane of protein, measured a
protein with a molecular weight of 80 kD. The graph is an
average of four blots with an example below the graph.

D. Hammond et al. – Antigenic Protein in LN1 Cells Grown in Space
146 Gravitational and Space Biology Bulletin 19(2) August 2006
GAPDH
0
20
40
60
80
100
120
Grd 3 Grd 6 Grd 9 Flt 3 Flt 6 Flt 9 Control
Perc
ent
of
Contr
ol
Figure 3 - Glyceraldehyde-3-phosphate dehydrogenase
(GAPDH) antibody stained blot, prepared with 6 µg/lane of
protein per lane, measured a protein with a molecular
weight of 42-44 kD. The graph is the average of three blots
with an example below the graph.
Student's t-test was used to compare the ground and
flight samples at each time point, days 3, 6 and 9.
There was no significant difference between Flt and
Grd samples at any time point (p > 0.05).
Parallel immunohistochemical studies on
formalin-fixed flight and ground cultures also
showed positive immunostaining for VDR (Fig 4) as
well as other biomarkers (not shown here). These
results are consistent with data from antigenic
recovery experiments performed on human mixed
Müllerian tumor cells cultured in microgravity
(Hammond, et al., 2005) as well as data from cells
grown using different culture methods (Cubano,et al.,
2004). Although on a protein per protein basis, the
preserved cells demonstrated slightly less antigenic
protein than untreated cells (control), there was a
high percentage of recovery of antigenic protein
using three different antibodies in both the ground
and flight samples.
These studies demonstrate that quantitative
proteomic information can be acquired from samples
stored in less than optimum circumstances, such as
long term storage at refrigerated temperatures with
different cell lines, using multiple antibodies. Since
space flight experiments are often executed under
resource constraints, this is a valuable addition to our
knowledge of how to best utilize our space samples
and to other work done in extreme environments such
as in undeveloped countries where biospecimens are
kept in suboptimal conditions.
(Supported by NAS9-02078 and NRA NAG8-1362).
Figure 4 - Light photomicrograph (600x) of Vitamin D
receptor immunoreactivity in sectioned formalin fixed
HRCE cells cultured on Cytodex-3 microcarrier beads in
microgravity for 12 days during ISS Increment 4.
Cubano, L., Allen, P.L., Stodieck, L., Genova, J.
Klassen, R.B.B, Love, J., Hammond, T.G. 2004.
Transcription factor translocation during changes in
renal cell mechanical culture. Gravitational and
Space Biology Bulletin 17(2): 75-82
Cowger, N.L., Benes, E., Allen, P., Hammond T.G.
2002. Expression of renal cell protein markers is
dependent on initial mechanical culture conditions. J
Appl Physiol 92:691
Hammond, D.K., Becker, J., Elliott, T.F., Holubeck,
K, Baker, T.L., Love, J.E. 2005. Antigenic protein in
microgravity-grown human mixed Müllerian ovarian
tumor (LN1) cells preserved in RNA stabilizing agent
Gravitational and Space Biology 18(2): 99-100
Love, J.E., Hammond, D.K., Elliott, T.F., Holubec,
K., Baker, T.L., Allen, P.L. and Hammond, T.G.
2003. Antigenic protein from RNALater™ treated
cell cultures grown on the international space station
(ISS), using the cellular biotechnology operations
support system (CBOSS). Gravitational and Space
Biology Bulletin. 17:39
Rodrigo, M.C., Martin D.S.K., Redtzke, R.A., Eyster,
K.M. 2002. A method for the extraction of high-
quality RNA and protein from single small samples
of arteries and veins preserved in RNAlater. J.
Pharm. Tox. Methods. 47:87.
Towbin, H., Staehelin, T., Gordon, J. 1979.
Electrophoretic transfer of proteins from
polyacrylamide gels to nitrocellulose sheets:
procedure and some applications. Proc. Natl. Acad.
Sci. USA. 76:4350.

Gravitational and Space Biology 19(2) August 2006 147
THE EFFECT OF CHAGES OF GRAVITY ON HUMAN MONOCYTE CELL (TUR) PHAGOCYTOSIS
Cassidy B. Johnson1*
, Lillian S. Waldbeser2
1Dept. of Biochemistry and Cell Biology, PO Box 1892, Rice University, Houston, Texas 77251
2
Dept. of Physical and Life Sciences, Texas A&M University-Corpus Christi, Corpus Christi, TX 78412 USA
The function of the immune system is compromised
during spaceflight (Cogoli, 1996). Many cellular activities
crucial for an effective immune response are diminished
in microgravity conditions. Phagocytosis is a vital,
cytoskeleton-mediated immune process necessary for the
clearance of pathogenic microorganisms; therefore
alterations to this important process may leave astronauts
vulnerable to infection. We sought to determine if
modeled microgravity or hypergravity conditions alter the
phagocytic capacity of a human monocyte-like phagocytic
immune cell (TUR; ATCC, Manassas, V.A.).
Microgravity and hypergravity conditions were
modeled using ground-based systems. A rotary cell-
culture system (RCCS-4D, Synthecon, Houston, TX)
was used to was used to simulate the free-fall conditions
that occur in microgravity, and a novel device called the
Hypergravity 3000 (HG3K), was designed and built for
the analysis of hypergravity. This machine was designed
to fit inside a tissue culture incubator and to concurrently
simulate 4,6, and 8 g.
The TUR cells were cultured under standard tissue
culture conditions until they reached confluence. Cells
were then seeded into the RCCS-4D or the HG3K at a
concentration of 1 x 106. The cells were cultured under
microgravity or hypergravity conditions for 10 days, and
100 µL aliquots of cells were removed from the machines
at 1, 4, 12, 24, 48, 120, and 240 hours, counted and
assayed for phagocytosis. For the phagocytosis assay,
fluorescein-conjugated Escherichia coli bacteria
(BioParticles, Molecular Probes, Eugene, OR) were
added to the experimental cells at a 1:10 cell:particle
ratio. The cells were allowed to incubate with the
BioParticles for 45 minutes, the optimal time allotment
for phagocytosis that minimized re-exposure to 1 g
conditions. After this 45-minute period, the cells were
briefly spun down and washed at 4°C to terminate
phagocytosis and remove the majority of non-
phagocytosed particles. Ethidium bromide was
additionally added to the cell samples to distinguish dead
or apoptotic cells from non-phagocytic cells. An average
of 500 cells were counted and scored as phagocytic or
non-phagocytic at each time point for every level of g and
a linear regression analysis was used to compare the
probability of phagocytosis occurring at each level of g
over time.
* Correspondence to: Cassidy B. Johnson
Dept. of Biochemistry and Cell Biology
Houston, PO Box 1892, TX 77251
Email: [email protected]
Phone: 713-348-2715
Overall, exposure to modeled microgravity increased
the incidence of phagocytosis in TUR cells over the 10-
day exposure period (Fig.1). The most dramatic increase
in phagocytic probability occurred within the first 48
hours of exposure where greater than 60% of the TUR
cells underwent phagocytosis compared to ~42%
observed in the 1 g control cells. This increase diminished
over the next 5 to 10 days, however phagocytosis never
returned completely to 1 g levels. Conversely, 10-day
hypergravity exposure decreased phagocytosis overall in
TUR cells (Fig. 1). Similar to the trend observed in
microgravity, the greatest decrease in phagocytosis
occurred within the first 48 hours, however a ‘peak’ of
phagocytic activity was observed at 24 hours. To further
examine this phenomenon, three additional 5-day
hypergravity experiments were performed to establish if
this peak was due to experimental conditions or sampling
error (Fig.2). Interestingly, after an initial decrease in
activity, phagocytosis increased during all three of these
experiments, the highest proportion of phagocytosis
occurring at 24 hours at all hypergravity levels. The peak
noted at 24 hours during the first 10-day experiment was
also replicated in all three of these concurrent
experiments, suggesting that this observation was not due
to sampling error.
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0 40 80 120 160 200 240
Time (h)
0 g 1 g
4.1 g 6 g
8 g
Figure 1. Probability of phagocytosis occurring in TUR
cells during a 10-day exposure to modeled microgravity or
hypergravity conditions. One hour of exposure to modeled
microgravity (0 g) or hypergravity (4.1, 6, and 8 g)
increases phagocytosis compared to control cells (1 g). The
percentage of cells undergoing phagocytosis remains
elevated in microgravity but reduced in hypergravity over a
48-hour period, after which phagocytosis in both conditions
slowly returns to almost 1 g levels at the end of the 10-day
period.

C. Johnson and L. Waldbeser – Effect of Changes of Gravity on Human Monocyte (TUR) Phagocytosis
148 Gravitational and Space Biology 19(2) August 2006
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0 20 40 60 80 100 120
Time (h)
0 g 1 g
4 g 6 g
8 g
The results of these experiments suggest that that
microgravity and hypergravity conditions modify
phagocytic function in human TUR cells. The
discrepancies between the first 10-day and the concurrent
5-day hypergravity experiments may be explained by the
age of the TUR cells. The cells utilized for the 10-day
experiment were cultured for less than a week at 1 g in
tissue culture flasks before use, while the cells utilized for
the 5-day experiments had been cultured for 10 to 18
days. The synchronously cultured 1 g control cells
exhibited the same phagocytic probability regardless of
their age, indicating that change in gravity coupled with
age may modify phagocytosis. The cells cultured at 8 g
exhibited the lowest phagocytic probability during the
first 10-day trial and the highest during the following 5-
day experiments, whereas the cells exposed to 4 g
remained very close to the “base-line” values of 1 g. This
data suggests that the hypergravitational force acts in a
“dose-dependant” manner. Changes in the levels of
protein kinase C (PKC) may explain these modifications
in phagocytosis, including the ‘peak’ phenomenon that
occurred at 24 hours in all three hypergravity conditions
and the less distinct peak at 24 hours observed in
microgravity. Several PKC isoforms, including PKC α,
PKC β PKC δ and PKC ε are involved in antibody-
receptor mediated phagocytosis (Allen and Aderem,
1996). The translocation and concentration of PKC is
modified in both microgravity and hypergravity
conditions (Schmitt et al., 1996). In microgravity,
translocation of PKC occurs rapidly over a 1-hour period,
then steadily declines over time whereas translocation of
PKC in hypergravity steadily increases over time and with
increased g-level (Hatton et al., 1999). Translocation of
PKC plays an important role in phagocytosis (Larsen et
al., 2000), therefore the additional movement of this
kinase to the cytosol may be causing the enhanced
phagocytic activity of TUR cells, a known PKC deficient
cell (Hass et al., 1993), cultured in altered gravity
conditions. Additional experiments utilizing other cell
lines along with assessment of PKC gene expression are
warranted to fully investigate this phenomenon.
The percent of TUR cells undergoing phagocytosis
started to return to 1 g levels after the 5th
day of
microgravity or hypergravity exposure. The delayed
hypersensitivity reaction of astronauts was additionally
found to normalize after the fifth day of microgravity
exposure (Taylor and Janney, 1992). These observations
suggest that immune cells may have the capacity to adapt
to gravitationally altered environments. This ability to
‘adapt’ may involve the cell’s cytoskeleton, including
actin which is the major cytoskeletal component involved
in phagocytosis. Disorganization and reorganization of
the cytoskeleton in microgravity has been observed in
several cell types, with much of the reorganization
occurring after 24 hours of exposure (Lewis et al., 1998).
This reorganization corresponds to the large phagocytic
peak observed at 24 hours, suggesting a connection
between these two events.
(Supported by: Texas Space Grant Consortium “New Principle
Investigator Grant 2004-2005 ” and Texas Excellence Fund Grant
2005.)
REFERENCES Cogoli, A. 1996. Gravitational physiology of human immune cells: A
review of in vivo, ex vivo, and in vitro studies. J. Grav. Physiology. 3:
1-9.
Allen, L.H., and Aderem, A. 1996. Molecular definition of distinct
cytoskeletal structures involved in compliment and Fc receptor-mediated
phagocytosis in macrophages. J. Exp. Med. 184: 627-637.
Schmitt, D.A., Hatton, J.P., Emond, C., Chaput, D., Paris, H., Levade,
T., Cazenave, J.P., and Schaffar, L. 1996. The distribution of protein
kinase C in human leukocytes is altered n microgravity. FASEB J. 10:
1627-1634.
Hatton, J.P., Gaubert, F., Lewis, M.L., Darsel, Y., Ohlmann, P.,
Cazenave, J.P., and Schmitt, D. 1999. The kinetics of translocation and
cellular quantity of protein kinase C in human leukocytes are modified
during spaceflight. FASEB J. 13: S23-S33.
Larsen, E.C., DiGennaro, J.A., Saito, N., Mehta, S., Loegering, D.J.,
Maxurkiewiez, J.E., and Lennartz, M.R. 2000. Differential requirements
for classic and novel PKC isoforms in respiratory burst and phagocytosis
in RAW 264.7 cells. J. Immuno. 165: 2809-2817.
Hass, R., Hirano, M., Kharbanda, S., Rubin, E., Meinhardt, G., and
Kufe, D. 1993. Resistance to phorbol-ester induced differentiation of a
U937 myeloid leukaemia cell variant with a signalling defect upstream
to Raf-1 kinase. Cell Growth &Diff. 4: 657-663.
Taylor, G.R., and Janney, R.P. 1992. In vivo testing confirms a blunting
of the human cell-mediated immune mechanism during space flight. J.
Leukocyte Biol. 51: 129-132.
Lewis, M.L., Reynolds, J.L., Cubano, L.A., Hatton, J.A., Lawless, B.D.,
and Piepmeier, E.H. 1998. Spaceflight alters microtubules and increases
apoptosis in human lymphocytes (Jurkat). FASEB J. 12: 1007-1018.
Figure 2. Average phagocytic probability for three successive 5-
day hypergravity experiment compared to modeled microgravity
(0 g) and controls (1 g). Hypergravity initially decreases
phagocytosis, however phagocytic activity increases at 12 hours
and the percent of phagocytic cells remains elevated over the 5-
day period compared to1 g controls. Note the peak of phagocytic
activity at 24 hours observed during the 10-day trial at 24 hours
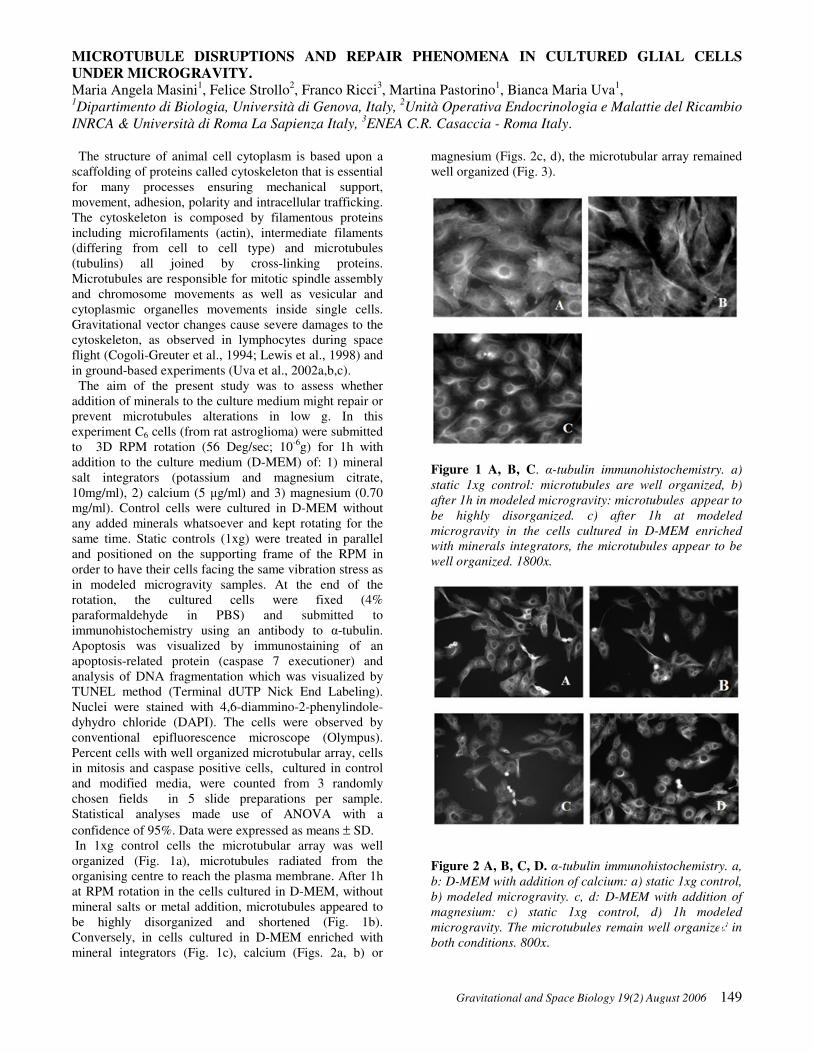
Gravitational and Space Biology 19(2) August 2006 149
MICROTUBULE DISRUPTIONS AND REPAIR PHENOMENA IN CULTURED GLIAL CELLS
UNDER MICROGRAVITY.
Maria Angela Masini1, Felice Strollo
2, Franco Ricci
3, Martina Pastorino
1, Bianca Maria Uva
1,
1Dipartimento di Biologia, Università di Genova, Italy,
2Unità Operativa Endocrinologia e Malattie del Ricambio
INRCA & Università di Roma La Sapienza Italy, 3ENEA C.R. Casaccia - Roma Italy.
The structure of animal cell cytoplasm is based upon a
scaffolding of proteins called cytoskeleton that is essential
for many processes ensuring mechanical support,
movement, adhesion, polarity and intracellular trafficking.
The cytoskeleton is composed by filamentous proteins
including microfilaments (actin), intermediate filaments
(differing from cell to cell type) and microtubules
(tubulins) all joined by cross-linking proteins.
Microtubules are responsible for mitotic spindle assembly
and chromosome movements as well as vesicular and
cytoplasmic organelles movements inside single cells.
Gravitational vector changes cause severe damages to the
cytoskeleton, as observed in lymphocytes during space
flight (Cogoli-Greuter et al., 1994; Lewis et al., 1998) and
in ground-based experiments (Uva et al., 2002a,b,c).
The aim of the present study was to assess whether
addition of minerals to the culture medium might repair or
prevent microtubules alterations in low g. In this
experiment C6 cells (from rat astroglioma) were submitted
to 3D RPM rotation (56 Deg/sec; 10-6
g) for 1h with
addition to the culture medium (D-MEM) of: 1) mineral
salt integrators (potassium and magnesium citrate,
10mg/ml), 2) calcium (5 µg/ml) and 3) magnesium (0.70
mg/ml). Control cells were cultured in D-MEM without
any added minerals whatsoever and kept rotating for the
same time. Static controls (1xg) were treated in parallel
and positioned on the supporting frame of the RPM in
order to have their cells facing the same vibration stress as
in modeled microgravity samples. At the end of the
rotation, the cultured cells were fixed (4%
paraformaldehyde in PBS) and submitted to
immunohistochemistry using an antibody to α-tubulin.
Apoptosis was visualized by immunostaining of an
apoptosis-related protein (caspase 7 executioner) and
analysis of DNA fragmentation which was visualized by
TUNEL method (Terminal dUTP Nick End Labeling).
Nuclei were stained with 4,6-diammino-2-phenylindole-
dyhydro chloride (DAPI). The cells were observed by
conventional epifluorescence microscope (Olympus).
Percent cells with well organized microtubular array, cells
in mitosis and caspase positive cells, cultured in control
and modified media, were counted from 3 randomly
chosen fields in 5 slide preparations per sample.
Statistical analyses made use of ANOVA with a
confidence of 95%. Data were expressed as means ± SD.
In 1xg control cells the microtubular array was well
organized (Fig. 1a), microtubules radiated from the
organising centre to reach the plasma membrane. After 1h
at RPM rotation in the cells cultured in D-MEM, without
mineral salts or metal addition, microtubules appeared to
be highly disorganized and shortened (Fig. 1b).
Conversely, in cells cultured in D-MEM enriched with
mineral integrators (Fig. 1c), calcium (Figs. 2a, b) or
magnesium (Figs. 2c, d), the microtubular array remained
well organized (Fig. 3).
Figure 1 A, B, C. α-tubulin immunohistochemistry. a)
static 1xg control: microtubules are well organized, b)
after 1h in modeled microgravity: microtubules appear to
be highly disorganized. c) after 1h at modeled
microgravity in the cells cultured in D-MEM enriched
with minerals integrators, the microtubules appear to be
well organized. 1800x.
Figure 2 A, B, C, D. α-tubulin immunohistochemistry. a,
b: D-MEM with addition of calcium: a) static 1xg control,
b) modeled microgravity. c, d: D-MEM with addition of
magnesium: c) static 1xg control, d) 1h modeled
microgravity. The microtubules remain well organized in
both conditions. 800x.
B D

M.A. Masini, et al. - Microtubule Disruptions and Repair Phenomena in Cultured Glial Cells under Microgravity
150 Gravitational and Space Biology 19(2) August 2006
Figure 3. Number of cells with well organized
microtubules. Means ± SD.
The number of apoptotic cells was very high in the
cultures containing only D-MEM, but got back to control
levels (11±4.2% versus 70.62±11.06%) after magnesium
addition, while the number of cell division increased both
with calcium and magnesium (Fig.4A, B).
Figure 4 A, B. Number of cell divisions (A), and number
of apoptic cells (B) after 1 h at low g in controls and in D-
MEM modified by addition of calcium and magnesium.
Means ± SD
Our data confirm previously reported observations that
severe microtubular changes occur in low g and there is a
lack of microtubular self organization under modeled or
real microgravity conditions (Papaseit et al.; 2000;
Tabony, 1994). However divalent ions, as calcium and
magnesium, are known to play a major role in the
regulation of microtubular polymerization in vivo under
earth gravitational force. In fact a high magnesium
concentration is needed to assemble tubulin dimers into
microtubules in vitro (Gaskin, 1981).
The present results show that the addition of calcium or
magnesium to the culture medium results in protection of
microtubular integrity and chromatin, preventing
apoptosis and enhancing cell divisions, probably due to
the pivotal role of divalent ions in building the mitotic
spindle and the cleavage furrow.
In conclusion these data show that an enriched metal
concencentration in the extracellular compartments may
prevent low g alterations at the cellular level.
REFERENCES
Cogoli-Greuter, M., Pippia, P., Siola, I., and Cogoli, A.
1994. Lymphocytes on sounding rocket flights. J. Gravit.
Physiol. 1: 90-91.
Gaskin, F. 1981. In vitro microtubule assembly regulation
by divalent cations and nucleotides. Biochemistry 20:
1318-1322.
Lewis, M., Reynolds, J.L., Cubano, I.A., Hatton, J.P.,
Lowless, B.D., and Piepmeier, E.H. 1998. Spaceflight
alter microtubules and increases apoptosis in human
lymphocytes (Jurkat). Faseb J. 1007-1018.
Papaseit, C., Pochon, N., and Tabony J. 1998.
Microtubule self-organization is gravity-dependent. Proc.
Natl. Acad. Sci. USA 97: 83634-8368.
Tabony, J. 1994. Morphological bifurcations involving
reaction-diffusion processes during microtubule
formation. Science 264: 245-248.
Uva, B.M., Masini M.A., Sturla, M., Tagliaferro, G., and
Strollo, F. 2002a Microgravity-induced programmed cell
death in astrocites. Proceedings of “Life in Space for Life
on Earth” 23rd
Annual International Gravitational
Physiology Meeting, Stockholm, Sweden.
Uva, B.M., Masini, M.A., Sturla, M., Prato, P.,
Passalacqua, M., Giuliani, M., Tagliaferro, G., and
Strollo, F. 2002b. Clinorotation - induced weightlessness
influences the cytoskeleton of glial cells in culture. Brain
Res. 934: 132-139.
Uva, B.M., Masini M.A., Sturla, M., Buzzone, F.,
Giuliani, M., Tagliaferro, G., and Strollo, F. 2002c.
Microgravity-induced apoptosis in cultured glial cells.
Eur. J. Histochem. 46:209-214.
Control
70.62±1.06
+Ca2+
15±3.6 +Mg2+
11±4.2
A
B

Gravitational and Space Biology 19(2) August 2006 151
MICROGRAVITY-INDUCED CHANGES IN GENE EXPRESSION IN ACTIVATED T-LYMPHOCYTES INVOLVE MULTIPLE REGULATORY PATHWAYS.
Nancy E. Ward1, Neal R. Pellis
2, Semyon A. Risin
3 and Diana Risin
2
1Wyle Life Sciences, Houston, TX,
2NASA, Johnson Space Center, Houston, TX and
3University of Texas-Houston
Medical School.
We and others have shown that exposure to microgravity
(MG) results in a decline in cellular immune function. To
further investigate this phenomenon, microarray analysis
was employed to identify associated gene expression
changes and possible gravity sensitive genes. CD3- and
IL2-activated human T-cells were seeded at a density of
1 x 106cells/ml and maintained either in static (1g) or
modeled MG conditions for 24 hours at 37°C in an
atmosphere of 95% air and 5% CO2. To model MG
conditions we used the Rotating wall vessel (RWV)
culture system developed at NASA-Johnson Space Center
and commercially available from Synthecon, Inc. This
very low shear culture system randomizes gravitational
vectors and approximates the microgravity environment
by sustaining cells in continuous free fall. RWV
bioreactors were continuously rotated at a speed of
22 rpm.
To decrease biological variation and aid in detecting
microgravity-associated changes, experiments were
performed in triplicate utilizing T-cells obtained from
different blood donors. Total RNA was isolated using the
RNeasy isolation kit (Qiagen, Valencia, CA). RNA was
then analyzed for quality, using the Agilent 2100
Bioanalyzer, and the concentration was determined using
the NanoDrop® ND-1000 Spectrophotometer. Microarray
analysis was performed using the Affymetrix GeneChip®
Human U133A array as described in the Affymetrix
Analysis Technical Manual. The Human U133A array
contains >22,000 probe sets corresponding to 18,400
genes. A cutoff was set at >1.5 fold for analysis and
identification of genes of interest with the criterion for
significance <0.01.
Exposure to modeled microgravity resulted in
alteration of 89 genes, 10 of which were up-regulated and
79 down-regulated. Genes with altered expression were
categorized according to their function and their structural
role as well as association with certain metabolic and
regulatory pathways. A large proportion of them were
found to be involved in fundamental cellular processes:
signal transduction, DNA repair, apoptosis, and multiple
metabolic pathways (Table 1). A group of genes directly
related to immune and inflammatory responses was also
identified.
Up-regulated genes included IL7-receptor which plays
a role in lymphocyte development, and several heat shock
protein family members which function together as
molecular chaperones, stabilize existing proteins from
aggregation and mediate folding of newly translated
proteins.
Down-regulated genes included granzyme B, beta-3-
endonexin, and Apo-2 ligand, all three of which have
various roles in the induction of apoptosis. Among
down-regulated genes directly related to immune and
inflammatory responses we identified granulysin,
proteasome activator subunit 2, peroxiredoxin 4 gene,
HLA-DRA, lymphocyte antigen 75, IL18 receptor gene
and DOCK2 gene. Granulysin is a cytolytic molecule
demonstrating broad antimicrobial activity (Clayberger
and Krensky, 2003). Proteasome activator subunit 2
encodes the beta subunit of the 11S regulator of the
immunoproteasome which is important in the processing
of class I MHC peptides and induced by gamma
interferon (Zhang et al., 1998; Sijts et al., 2002; Kloetzel,
2004). The peroxiredoxin 4 gene encodes a cytosolic
antioxidant enzyme shown to play a regulatory role in the
activation of NF-KB (Kang et al., 1998; Fujii and Ikeda,
2002). HLA-DRA functions in presenting peptides
derived from extracellular proteins (Germain, 1994).
Lymphocyte antigen 75 is involved in the endocytosis of
antigen by dendritic cells (Rajagopal et al., 2004). The
protein encoded by the IL18 receptor gene specifically
binds IL18 and is essential for IL18 mediated signal
transduction (Wu et al., 2003; Subramaniam et al., 2004).
DOCK2 gene product is a protein indispensable for
lymphocyte chemotaxis (Reif and Cyster, 2002).
Also down-regulated were three proteasome beta
family members (10, 9 and 8) making up part of the 20S
proteasome core structure; several genes potentially
affecting the integrity of the cellular cytoskeleton, such as
CD2 associated protein, tubulin, and pericentrin 1; genes
affecting DNA repair, such as nudix and FEN1, and genes
affecting several metabolic pathways.
These data provide base information and a means to
evaluate the possible roles of genes demonstrating altered
expression in lymphocyte response to microgravity at the
molecular level.
Table 1. Functional clustering of genes responding to modeled
microgravity.
Number of genes
down-
regulated
up-
regulated
Tissue /growth regulation 1 1
Protein folding / degradation 3 3
Nucleotide / purine-pyrimidine 19 1
Metabolic pathways / metabolism 5 4
Structure / transport / binding
protein
11
Immune / inflammatory / defense 10 1
Apoptosis 3
Transcription factor 3
Signal transduction 17
Unknown function 7

N.E. Ward et al. – MICROGRAVITY-INDUCED CHANGES
152 Gravitational and Space Biology 19(2) August 2006
REFERENCES
Clayberger, C. and Krensky, A.M. 2003. Granulysin.
Current Opinion in Immunology 15: 560-565.
Fujii, J. and Ikeda, Y. 2002. Advances in our
understanding of peroxiredoxin, a multifunctional,
mammalian redox protein. Redox Rep. 7(3): 123-30.
Germain, R.N. 1994. MHC-dependent antigen processing
and peptide presentation: providing ligands for T
lymphocyte activation. Cell 76(2): 287-299.
Kang, S.W., Chae, H.Z., Seo, M.S., Kim, K., Baines, I.C.
and Rhee, S.G. 1998. Mammalian peroxoredoxin
isoforms can reduce hydrogen peroxide generated in
response to growth factors and tumor necrosis factor-
alpha. Journal of Biological Chemistry 273(11): 6297-
6302.
Kloetzel, P.M. 2004. The proteasome and MHC class I
antigen processing. Biochim Biophys Acta 1695(1-3):
225-233.
Rajagopal, D., Bal, V., George, A. and Rath, S. 2004.
Diversity and overlap in the mechanisms of processing
protein antigens for presentation to T cells. Indian J Med
Res 120: 75-85.
Reif, K. and Cyster, J. 2002. The CDM protein DOCK2
in lymphocyte migration. Trends Cell Biol 12(8): 368-
373.
Sijts A., Sun, Y., Janek, K., Kral, S., Paschen, A.,
Schadendorf, D. and Kloetzel, P.M. 2002. The role of
proteasome activator PA28 in MHC class I antigen
processing. Molecular Immunology 39(3-4): 165-169.
Subramaniam, S. Stansberg, C. and Cunningham, C.
2004. The interleukin 1 receptor family. Dev Comp
Immunol 28(5): 415-428.
Wu, C., Sakorafas, P., Miller, R., McCarthy, D., Scesney,
S., Dixon, R. and Ghayur, T. 2003. IL-18 receptor
induced changes in the presentation of IL-18 binding sites
affect ligand binding and signal transduction. Journal of
Immunology 170: 5571-5577.
Zhang, Z., Clawson, A. and Rechsteiner, M. 1998. The
proteasome activator 11S regulator or PA28. Journal of
Biological Chemistry 273(46): 30660-30668.

Gravitational and Space Biology Bulletin 19(2) August 2006 153
POSSIBLE INVOLVEMENT OF FLOW DETECTION IN THE ACTIVATON OF OSTEOBLASTS*
Muneo Takaoki1
, Hyekeyong Park2
, Naoko Murakami3
, Dai Shiba2
, and Jun-Ichiro Gyotoku2
1
Space Environment Utilization Center, OSFO, JAXA; 2
ISS Science Project Office, ISAS, JAXA, 3Advanced
Engineering Services Co. Ltd.; Tsukuba 305-8505, Japan
One of the most potent mechanical stimulators for bone
cells may be the interstitial fluid flow. Fluid flow induces
various events such as Ca2+
signaling, c-fos expression,
actin fiber reorganization, and appearances of bone
markers in cultured cells (Chen et al., 2000). Fluid shear
stress may distort cellular membrane and other structures,
inducing cellular response. However, the nature of
molecules that convert the mechanical strain into
biological signals remains unclear.
The difficulty in identifying the mechanosensor
molecules seems to lie, at least in part, in the lack of
reliable methods for analysis. In contrast to regular cell
culture experiments, that compare the effects of dilute
substances, cellular responses to mechanical stimulations
are often difficult to interpret. The mechanical stimulation
may alter culture conditions drastically. It is extremely
difficult to apply only mechanical stimulations, leaving all
other conditions uniform. Excluding the effects of such
environmental changes, induced secondarily by the
operational procedure, is almost impossible. Thus, this
study seeks to develop a simple method for applying
mechanical stimulation with minimum secondary
influences, especially from temperature changes during
laboratory procedures.
Figure 1. C-fos expression induced either by rocking (A) or
cooling (B) of the culture flask. MC3T3-E1 cells were pre-
cultured for 3 days at 37°C in T-25 flasks. Culture flasks were
placed in small Styrofoam boxes with heat accumulators (small,
refreezable pack equilibrated to 37°C). This minimized the
cooling during incubator door opening and rocking. Gently
replacing heat accumulators with cold (4°C) packs for 1 min.
achieved cooling with the least possible fluid agitation (B).
After replacing with warm packs again, all flasks were
incubated at 37°C for additional 10 min. Total RNA was
extracted and c-fos gene expression was evaluated with RT-
PCR, and the values were normalized to 18S rRNA as described
previously (Takaoki et al., 2004). Values are means and
standard errors for 3 flasks.
We observed that conventional laboratory handling of
culture flasks could stimulate the murine osteoblastic cell
line MC3T3-E1. Fluid flow, generated either by manual
rocking or by unplanned agitation of culture flasks,
possibly induced c-fos expression to a far greater extent
than that caused by intended stimulation such as
centrifugation (Takaoki et al., 2004). Moreover, cooling
of the culture fluid, due to handling at room temperature,
induced similar response. To distinguish the effect of
rocking from the influence of cooling, we successfully
utilized small Styrofoam boxes and heat accumulators.
With this simple system, it was shown that either fluid
flow or cooling independently induced the responses
(Figure 1). Although flow-cells are normally used for the
purpose (Stevens and Frangos, 2003), this rocking method
was suitable for the parallel analyses of responses in
multiple flasks.
St i l l at 37Åé
10 minafte r 1stRocking
3
2. 5
2
1. 5
1
0. 5
0
Rela
tiv
e
E
xp
ress
ion
c-f
os
60 minafte r 1stRocking
10 minafte r 2ndRockingat 60min
70 minafte r 1stRocking
Figure 2. C-fos expression response to the second stimulation.
Culture flasks were rocked and then kept undisturbed for one
hour before the second stimulation. Values are means and
standard errors for 3 flasks.
The response reached a peak 10 to 15 min. after the
stimulation and settled to the background level within an
hour. The second stimulation given 60 min. later resulted
in a comparable response (Figure 2). In this study, we kept
all cultures inside the incubator without any disturbance
for at least 2 hours before rocking.
There was some possible mediation of the c-fos
expression response by Ca2+
and the ERK signaling
pathway. The addition of an ERK inhibitor, U0126
suppressed the response (Figure 3). Fluid flow can be more efficiently detected when seen
as a drag force against a projected structure than as a
shear stress to a smooth surface. One such projection,
primary cilium, served as a flow meter in kidney epithelial
cells. A transient receptor potential (TRP) channel family
protein, polycystin-2 (PC2) with polycystin-1, is one of
the key
* correction of article that was published in 2005
(volume 18)

M. Takaoki et al. –Flow detection by Osteoblasts
154 Gravitational and Space Biology Bulletin 19(2) August 2006
molecules for this function. PC2 localized at the primary
cilia and functioned as mechanosensitive Ca2+
ion
channel. Ca2+
signalling initiated from primary cilium
distorted by the drag of flowing fluid are sent to
neighbouring cells through gap junctions (Nauli et al.,
2003). Whitfield (2003) discussed the possible
involvement of similar mechanisms in bone cells.
Interstitial fluid flow could be caused by the slightest
distortion of bone tissue by a mechanical load. This flow
could be easily detected by sensitive flow detecting
primary cilia.
Primary cilia were found in the osteoblastic cell line
MC3T3-E1. The cells also expressed PC2. These
observations suggested that bone cells and kidney cells
detected fluid flow through similar mechanisms. However,
the localization of PC2 to the primary cilia was not as
preferential as had been reported in kidney cells (Figure
4). This weak localization of PC2 in the primary cilia and
the difference in reported flow sensitivities (Chen et al.,
2000, Nauli et al., 2003) suggested the involvement of
molecules other than those in kidney cells.
VehicleStill
U0126Still
3
2. 5
2
1. 5
1
0. 5
0
Rel
ativ
e E
xpre
ssio
nc-
fos
VehicleRocked
U0126Rocked
Figure 3. Suppression of c-fos response to fluid flow by U0126
37°C. Values are means and standard errors for 4 flasks.
Figure 4. Immunofluorescent staining of primary cilium
(arrow) of an MC3T3-E1 cell. Immunostainings with anti-
acetylated α-tubulin (AcTub) or anti-PC2 antibodies (right) are
shown.
We observed that MC3T3-E1 cells expressed c-fos in
cooling as well as in rocking of the culture flasks. This
response to the cooling would not seem physiological,
since we could find virtually no literature describing
temperature dependencies in bone metabolism.
Nevertheless, the response could be a factor observable
only under culture conditions with a cell line. Detection
of lowered temperature by PC2 or other TRP channel
superfamily proteins could also be possible. Some TRP
channel superfamily proteins have been shown to sense
either cold, cool or hot temperatures (Clapham, 2003),
while another member was assumed to sense mechanical
stimuli (Corey et al., 2004). Most of these proteins are
expressed in the respective neurons in a highly specific
manner, but not in bone or kidney cells. However, we
have not entirely excluded the possibilities of
mechanosensing and temperature sensing by PC2, or
contributions of other bone-specific TRP channel
superfamily proteins.
The transient c-fos expression response could be
induced by a variety of stimulations. These stimulations
could be applied, either intentionally or unintentionally, to
the cell culture during laboratory procedures. However,
the effects of these stimulations might be considered
trivial in regular experiments, where both reference
controls and experimental groups undergo virtually
identical environmental changes, and are treated side by
side in a similar manner. On the other hand, cultures must
usually be placed in separate locations when applying
different mechanical stimulations. Therefore, it is
extremely important to keep the environmental changes,
such as temperature, humidity, and CO2 concentration
uniform among the groups. This is particularly true in
space experiments, which must be precisely controlled to
enable comparisons between flight and ground specimens.
REFERENCES Chen, N.X., Ryder, K.D., Pavalko, F.M., Turner, C.H., Burr,
D.B., Qui, J., and Duncan, RL. 2000. Ca2+ regulates fluid shear-
induced cytoskeletal reorganization and gene expression in
osteoblasts. Am. J. Physiol. Cell Physiol. 278:C989-C997.
Clapham, D.E. 2003. TRP channel as cellular sensors. Nature
426:517-524
Corey, D.P., Garcia-Anoveros, J.. Holt, J.R., Kwan, K.Y., Shuh-
Yow, L., Vollrath, M.A., Amalfitano, A., Cheung, E.L.-M.,
Derfler, B., Duggan A., Geleoc, G.G., Gray P.A., Hoffman,
M.P., Rehm, H.L., Tamasauskas, D., Zhang D.-S. 2004. TRPA1
is a candidate for a mechanosensitive transduction channel
vertebrate hair cells. Nature 432:723-730.
Nauli, SM, Alenghat, F.J., Luo, Y., Williams, E., Vassilev, P.,
Li, X., Elia A.E.H., Lu, W., Brown, E.M., Quinn, S.J., Ingbar
D.E., and Zhou, J. 2003. Polycystins 1 and 2 mediate
mechanosensation in the primary cililum of kidney cells. Nature
Genetics. 33:129-137.
Stevens H.Y., and Frangos, J.A., 2003. Bone cell responses to
fluid flow. In Helfrich, M.H. and Ralston, S.H. eds. “Bone
Research Protocols”, Humana Press (Totowa, NJ) pp381-398.
Takaoki, M., Murakami, N. Gyotoku, J.-I. 2004. C-fos
expression of osteoblast-like MC3T3-E1 cells induced either by
cooling or by fluid flow Biological Science in Space 18:181-
182
Whitfield, J.F. , 2003. Primary cilium--is it an osteocyte's
strain-sensing flowmeter? J. Cell. Biochem. 89:233-237.

Gravitational and Space Biology 19(2) August 2006 155
TRANSCRIPTIONAL PROFILING OF THE gps1 MUTANT OF ARABIDOPSIS
Vijayanand Nadella1, 2
, C. Dustin Hildenbrand1 and Sarah E. Wyatt
1*
1
Department of Environmental and Plant Biology, Ohio University, Athens, Ohio, USA; 2Current Address:
Department of Biology, Purdue University, West Lafayette, IN, USA; *corresponding author [email protected]
A plant’s response to gravity is a complex mechanism
involving stimulus perception, signal transduction and a
differential growth response. Disturbance of any of these
steps would affect a plant’s ability to respond to gravity.
The gravity persistence signal (gps) mutants were
identified using a cold treatment to select for mutants
specifically defective in the signal transduction events
prior to auxin redistribution. Plants perceive gravity at
4oC but auxin transport is abolished; transport is restored
upon return to room temperature (RT) (Wyatt et al. 2002).
Although auxin transport is restored in gps1 after return to
RT, auxin is not asymmetrically redistributed, and the
inflorescence stems fail to bend in response to the cold
gravistimulation (Nadella et al., 2006). To identify
additional genes in the pathway, especially those affected
by the gps1 mutation, we analyzed differential transcript
abundance between gps1 and wild-type (WT).
gps1 and wild-type plants were grown to maturity,
gravistimulated at 4oC for 1 h and returned to RT (GPS
treatment) (Wyatt et al. 2002). Inflorescence stems were
sprayed with RNAlater (Ambion) 5 min after return to
RT, and tissue from the region of elongation was
collected and analyzed for relative changes in transcript
levels. As a control for the cold treatment, additional WT
and gps1 plants were placed vertically at 4°C for 1h,
returned to RT and tissue collected as above. Total RNA
was extracted and purified (RNAeasy, Qiagen) from 10-
12 plants for each treatment and sent to the Microarray
Facility at University of California, Irvine for
amplification and hybridization to Arabidopsis ATH1
GeneChips (Affymetrix). Expression values were
normalized across each chip using the Cross Gene Error
Model (GeneSpring, Silicon Genetics, CA) and
significance determined by a Student T-test (p value=0.1).
From the 24,000 transcripts analyzed, 258 genes
showed differential expression: 128 up-regulated and 129
down-regulated in gps1 as compared to wild-type after
cold gravistimulation (Fig. 1). Those genes that were
similarly expressed in response to the cold treatment
alone were eliminated from further consideration.
Ten genes were selected, based on level of differential
expression, for analysis of transcript abundance
throughout the GPS treatment (Table 1). Both WT and
gps1 plants were grown to maturity, cold gravistimulated
for 60 min then returned to vertical at RT (Wyatt et al.
2002). RNA was extracted from the elongation zone of
the inflorescence stems at 0, 30, and 60 min after
gravistimulation and at 70 min after gravistimulation (10
min after return to vertical at RT). Transcript abundance
was determined by real time PCR using an ABI7900HT
sequence detection system according to Kimbrough et al.
(2004). Actin8 expression was measured separately for
Figure 1. Biological function of genes down-regulated (top) and
up-regulated (bottom) in gps1 as compared to wild-type under
the GPS treatment. Numbers indicate the number of genes in
each functional group. Genes selected showed at least a 2-fold
change in expression.
Table 1. Selected genes differentially expressed in gps1
as compared to WT after GPS treatment.
Protein Name Gene
Identifier
Fold
Change
putative 3-methyl crotonyl-
CoA carboxylase
At1g03090 4
putative protein kinase At1g30640 3
Spot 3 protein/vacuolar
sorting receptor homolog
At1g30900 3
hypothetical protein At1g50040 4
F-box protein At1g68050 5
CONSTANS B-box
zinc finger family protein
At1g78600 4
bZIP transcription factor At2g22850 5
cyclobilin like protein At2g39400 6
putative phospholipase At2g39400 6
myb transcription factor At5g12870 5
each time point, and the values used to normalize
transcript abundance of the selected genes.
electron transport/ energy pathways, 3
other cellular processes, 3
transport, 3
signal transduction, 2
cell organization &
biogenesis, 1
other metabolic
processes, 32
biological process
unknown, 31
other physiological
processes, 14 protein metabolism, 9
other biological
processes, 9
transcription,
7
developmental
processes, 5
abiotic / biotic
stimulus response, 5
Stress response , 4
other physiological
processes, 24
biological process
unknown, 7
other cellular
processes, 10
transcription, 9
protein metabolism, 17
other biological
processes, 7
electron transport/
energy pathways, 7
transport, 24
other metabolic
processes, 24

V Nadella et al-Transcriptional profiling of the gps1 mutant of Arabidopsis
156 Gravitational and Space Biology 19(2) August 2006
Based on expression pattern, most genes fell into two
categories. Transcripts of the Spot 3 vacuolar sorting
protein, 3-methylcrotonyl-CoA carboxylase and
hypothetical protein increased dramatically in WT within
30 min after gravistimulation but were virtually absent in
gps1 (Fig. 2) suggesting that these genes may be regulated
either directly or indirectly by GPS1. Transcript level of
the phospholipase, CONSTANS B box protein, bZIP
transcription factor and cyclophilin were either unaffected
or down-regulated in WT but spiked in gps1 between 30
and 60 min after gravistimulation (Fig. 3). These genes
appear to be negatively regulated by GPS1 or its products.
Figure 2. Time course of transcript intensity of genes up-
regulated in WT (solid line) but absent in gps1 (dashed line).
Time after gravistimulation is indicated on the X-axes and
relative intensity of gene expression (as normalized to Actin8) is
shown on the Y-axes. Two biological replicates, with three
technical replicates each, were performed for each time point.
Bars indicate standard error.
Figure 3. Time course of transcript intensity of genes up-
regulated in gps1 (dashed line) compared to WT (solid line).
Time after gravistimulation is indicated on the X-axes and
relative intensity of gene expression (as normalized to Actin8) is
shown on the Y-axes. Two biological replicates, with three
technical replicates each, were performed for each time point.
Bars indicate standard error.
Although several of these genes are novel to
gravitational biology, potential roles for a vacuolar sorting
protein or a phospholipase could been imaged. Morita et
al. (2002) suggested that vacuole formation and
functioning plays a key role in the early stages of shoot
gravitropism. Phospholipase A2 has also been implicated
in shoot gravitropism of Arabidopsis by auxin mediated
cell elongation (Lee et al., 2003). An increase in the
At2g39400 transcript in gps1 suggests that a
phospholipase, regulated by the GPS1 protein, might be
directly or indirectly involved in lateral auxin
redistribution thereby affecting cell elongation and
contributing to the gps1 no response phenotype. In any
event, we have identified genes that appear to be
regulated early in the gravity signal transduction pathway
and provide interesting new candidates for study.
ACKNOWLEDGEMENTS
Our sincere thanks to the members of the Plant
Gravitational Genomic Center at North Carolina State
University for their assistance with the quantitative RT-
PCR experiments and helpful discussions and to Heather
Sanders for her help in preparing the manuscript. This
project was partially supported by NASA: NAG2-1608 to
SEW and a Summer Undergraduate Research Fellowship
from the American Society of Plant Biologists to CDH.
REFERENCES
Kimbrough, J.M., Salinas-Mondragon, R., Boss, W.F.,
Brown, C.S., Sederoff, H.W. 2004. The fast and transient
transcriptional network of gravity and mechanical
stimulation in the Arabidopsis root apex. Plant
Physiology 136:2790-805.
Lee, H.Y., Bahn, S.C., Kang, Y.M., Lee, K.H., Kim, H.J.,
Noh, E.K., Palt, J.P., Shin, J.S., Ryu, S.B. 2003. Secretory
low molecular weight phospholipase A2 plays important
roles in cell elongation and shoot gravitropism in
Arabidopsis. Plant Cell 15: 1990-2002.
Morita, M.T., Kato, T., Nagafusa, K., Saito, C., Ueda, T.,
Nakano, A., Tasaka, M. 2002. Involvement of the
vacuoles of the endodermis in the early process of shoot
gravitropism in Arabidopsis. Plant Cell 14: 47-56.
Nadella ,V., Shipp, M.J., Muday, G.K., Wyatt, S.E. 2006.
Evidence for altered polar and lateral auxin transport in
the gravity persistent signal (gps) mutants of Arabidopsis.
Plant Cell and Environment 29: 682-690.
Wyatt, S.E., Rashotte, A.M., Shipp, M.J., Robertson, D.,
Muday, G.K. 2002. Mutations in the gravity persistence
signal loci in Arabidopsis disrupt the perception and/ or
signal transduction of gravitropic stimuli. Plant
Physiology 130: 1426-1435.
At1g03090
(putative 3-methylcrotonyl-CoA
carboxylase)
-4
-2
0
2
4
6
8
10
12
14
16
0 min 30 min 60 min 70 min
At1g50040
(hypothetical protein)
-4
-2
0
2
4
6
8
10
12
14
16
18
0 min 30 min 60 min 70 min
At1g30900
(Spot3 vacuolar sorting protein)
-5
0
5
10
15
20
0 min 30 min 60 min 70 min
At1g78600
(CONSTANS B box Zinc finger family
protein)
-4
-2
0
2
4
6
8
10
0 min 30 min 60 min 70 min
At2g39400
(putative phospholipase)
-10
-5
0
5
10
15
20
25
30
0 min 30 min 60 min 70 min
At2g22850
(bZIP family transcription factor)
-3
-2
-1
0
1
2
3
4
5
0 min 30 min 60 min 70 min
At2g36130 (cyclophilin like protein)
-3
-2
-1
0
1
2
3
0 min 30 min 60 min 70 min

Gravitational and Space Biology 19(2) August 2006 157
NITRIC OXIDE AND CGMP DEPENDENT SIGNALING IN ARABIDOPSIS ROOT GROWTH
Jennifer Jacobi1
, Jacob Elmer1
, Kristin Russell1
, Raj Soundur2
, and D. Marshall Porterfield2,3,4
1Department of Biological Sciences, University of Missouri Rolla, Rolla, MO;
2
Department of Agriculture and
Biological Engineering, 3Department of Horticulture and Landscape Architecture,
4Bindley Bioscience Center,
Purdue University, West Lafayette, IN.
Nitric oxide (NO) and cGMP are established
components of many calcium-dependent signaling
pathways in animal systems. Discovering this has been
fundamental to understanding some of the most basic
physiological processes (Lamattina, et al. 2003;
Wendehenne et al. 2001). In animals NO can act with or
without cGMP to activate protein kinases and cellular
responses. Studies of NO activity in plants have suggested
that the basic NO/cGMP mediated signaling pathway,
which operates in signaling in animals (Figure 1), may
also be active in plant cells. For example, recent work
has shown that NO and cGMP are involved in the Ca++
dependent cellular development in pollen (Prado et al.,
2004)). Here our work focuses on identifying potential
involvement of NO and cGMP in controlling cell
elongation and root extension, with an emphasis on
gravitropism in Arabidopsis roots.
We hypothesized that a calcium-dependent
NO/cGMP dependent signaling pathway may be active in
root gravitropism, either as a primary signal, driving
differential auxin transport in the root cap, or downstream
of an IAA receptor to elicit differential changes in cell
elongation. In these early studies we adopted a
pharmacological approach to investigate the pathway. We
exposed Arabidopsis thaliana seedlings to NO donor
compounds (DEA and Spermine NONOates), dibuturyl
cGMP, L-Name, IBMX, Viagra, and cPTIO. The
seedlings were grown for about a week, then transferred
to agar plates containing varying doses (0–700 uM) of the
drugs, and root growth was monitored for 24 hours.
We tested direct effects of NO on root elongation
(Figure 2) using NO donors (n=6) and found that low
concentrations of DEA (3.0-100 µM) and Spermine (5.0,
10.0 µM) significantly increased root elongation, while
high concentrations of DEA (700 µM) and Spermine
(300-700 µM) inhibited root elongation (t-test: p≤ 0.05).
Two well known phosphodiesterase inhibitors were also
used (n=6). Low concentrations of Viagra™ (1.0-10 µM)
and IBMX (1.0-150 µM) also significantly increased root
elongation (t-test: p≤ 0.05) but this effect was attenuated
as the concentration increased (Figure 2). These two sets
of experiments thereby suggest that there is a connection
between NO and cGMP in A. thaliana root elongation.
A common approach in NO research is to apply
NO using various donor molecules, but to understand the
resulting NO responses we have to account for the
L-Arginine + O2
X L-NamecPTIO –NO
DEA +NO
Spermine +NO
SNP +NO +CN-
NO
GTP
cGMP
PDE
X ODQ
X Viagra, IBMX
Cellular Response
+ Dibutryl cGMP
NO
Synthase
NO/cGMP Signaling Pathway
Guanylate
Cyclase
NO
cGMP
Hydrolysis
Auto-oxidation
L-Arginine + O2
X L-NamecPTIO –NO
DEA +NO
Spermine +NO
SNP +NO +CN-
NO
GTP
cGMP
PDE
X ODQ
X Viagra, IBMX
Cellular Response
+ Dibutryl cGMP
NO
Synthase
NO/cGMP Signaling Pathway
Guanylate
Cyclase
NOGuanylate
Cyclase
NO
cGMP
Hydrolysis
Auto-oxidation
L-Arginine + O2
X L-NamecPTIO –NO
PTIO –NO
Spermine +NONO
GTP
cGMP
PDE
X ODQ
X Viagra IBMX
Cellular Response
+ Dibutryl cGMP
NO
Synthase
NO/cGMP Signaling Pathway
Guanylate
Cyclase
NOGuanylate
Cyclase
NO
cGMP
Hydrolysis
Auto-oxidation
Calcium/Calmodulin
cPTIO X
Figure 1. NO signaling cascade. Nitric Oxide Synthase (NOS)
is activated by the Ca++ calmodulin complex. NOS converts L-
Arginine and oxygen to citruline and NO. L-Name is an analog
of arginine that inhibits NO production. cPTIO inhibits NOS by
reacting with NO to form carboxy-PTI derivatives. cPTIO also
scavenges NO. DEA, Spermine, and SNP are NO donors. SNP
also produces cyanide. ODQ is a selective inhibitor of the NO
sensitive Guanylyl Cyclase. ViagraTM and IBMX affect the
pathway in similar ways. Both are phospho-diesterase
inhibitors, although Viagra™ is a more specific inhibitor.
Dibutryl cGMP is a cGMP analog that activates protein kinase
G.
NO Donor Concentration (µM)
0 200 400 600
Root E
longation (
rela
tive g
row
th r
ate
)
0.0
0.2
0.4
0.6
0.8
1.0
1.2
1.4
1.6
DEA
Spermine
Phosphodiesterase inhibitor Concentration (µM)
0 50 100 150 200 250
Root E
longation (
rela
tive g
row
th r
ate
)
0.8
1.0
1.2
1.4
1.6
1.8
ViagraIBMX
Figure 2. Vertically grown Arabidopsis plants on agar media
that contained experimental concentrations of NO donors (DEA,
Spermine) and phosphodiesterase inhibitors (Viagra™ and
IBMX). The growth after 24 hours was measured and
normalized to the control giving relative growth rate.
Time (minutes)
0 20 40 60 80 100 120
NO
co
nce
ntr
atio
n (
µM
)
0
5
10
15
20
120 140 160 180 200 220 240
0.00
0.05
0.10
0.15
0.20
0.25
0.30
Time (minutes)
0 20 40 60 80 100 120
NO
co
nce
ntr
atio
n (
µM
)
0
5
10
15
20
120 140 160 180 200 220 240
0
1
2
3
4
5
Figure 3. DEA (left) and Spermine (right) drug kinetics. Both
DEA and Spermine modeling is plotted based on NO
concentration vs. time. NO decomposition was modeledfor eack
concentration (700, 500, 300,100 ,50, 30, 20, 10, 7, 5, 3, and 1
µM) as first order and the formation of nitrite was considered to
be second order. The rate constants used were K1-DEA=
1.4x10-3 s-1 (Schmidt et al., 1997), K1-Spermine = 3.2x10-4 s-1
(Ramamurthi et al. 1997) K2 = 9.2x106 M-2 s-1 (Schmidt et al.,
1997) was the same for both drugs.

J. Jacobi et al. – Nitric Oxide and cGMP Dependent Signaling in Arabidopsis Root Growth
158 Gravitational and Space Biology 19(2) August 2006
balance between NO production, and auto-oxidation in
the system. A key is to understand that NO levels
dynamically change, and the amount of NO available to
the plants at a given time is significantly less than the
actual NO donor drug concentration (Figure 3). Spermine
is a more consistent, sustainable NO source, and as a
result, the pharmacological effects of Spermine are more
dramatic than DEA (Figure 2). Sodium nitroprusside
(SNP) has become a common NO donor in plant
literature, despite being almost completely abandoned by
animal research. We did not use SNP as it is inconsistent,
requiring a reducing agent or light for activation, and is
also produces more cyanide (6:1) than NO (Yamato and
Bing 2000). While SNP has been reported to alter root
physiology (Cassimiro et al. 200l, Correa-Aragunde et al.
2004, Gouvea et al. 1997, Hu et al. 2005, Pagnussat et al.
2003) we found that SNP concentrations ranging from 5
to 300mM completely inhibited growth and killed the A.
thaliana plants.
These experiments suggest that altering NO and
cGMP using NONOate donors (Spermine and DEA),
phospohodiesterase inhibitors (Viagra ™ and IBMX), and
NO synthase inhibitors (L-Name, an arginine/citruline
intermediate) alters root elongation (Figures 2 and 4,) and
gravitropism (Figure 5). In general low doses of NO cause
growth to increase, while high doses of NO cause growth
to slow down and in some instances stop. Both cPTIO and
L-Name (Figure 4) reduce the amount of endogenous NO
production, and enhanced root elongation in a dose
dependent manner (Hilland Plot analysis)
These data are consistent with the effects of
auxin on root growth (Cassimiro et al. 200l, Correa-
Aragunde et al. 2004, Gouvea et al. 1997, Hu et al. 2005,
Pagnussat et al. 2003, Taiz et al. 2002), where high levels
of auxin (10-6
M) reduce/inhibit root growth while low
concentrations (10-10
or 10-9
M) increase elongation. We
believe that these results suggest an auxin-mediated
NO/cGMP signalling cascade may control cell elongation
at the cellular level. Auxin might also play a direct role in
graviperception signalling but further experiments are
required to answer that question.
REFERENCES Correa-Aragunde, N., Graziano, M., Lamattina, L. 2004. Nitric
oxide plays a central role in determining lateral root
development in tomato. Planta. 218: 900-905.
Casimiro, I., Marchant, A., et al. 2001. Auxin transport
promotes lateral rot iitiation. The Plant Cell. 13: 843-852.
Gouvea, C.M.P., Souza, J. F., et al. 1997. NO-releasing
substances that induce growth elongation in maize root
segments. Plant Growth Regulation. 21: 183 – 187.
Hu, X., Neill, S. J., et al. 2005. Nitric oxide mediates gravitropic
bending in soybean roots. Plant Physiology Preview.
Lamattina, L., Garcia-Mata, C., et al. 2003. Nitric Oxide: the
versitality of an extensive signal molecule. Annu. Rev. Plant
Biol. 54: 109-136.
Pagnussat, G.C., Lanteri, M.L., Lamattina, L. 2003. Nitric oxide
and cyclic GMP are messengers in the indole acetic acid –
induced adventitious rooting process. Plant Physiology. July,
132: 1241-1248.
Prado, A. M., Porterfield, D. M., Feijo, J. A. 2004. Nitric oxide
is involved in growth and regulation and reorientation of pollen
tubes. Development. 131: 2707-2714.
Ramamurthi, A., Lewis, R. S. 1997. Measurement and modeling
of nitric oxide release rates for nitric oxide donors. Chemical
Research in Toxicology. 10: 408-413.
Schmidt, K., Desch, W., K, et al. 1997. Release of nitric oxide
from donors with known half life: a mathematical model for
calculating nitric oxide concentrations in aerobic solutions.
Naunyn-Schmiedeberg’s Arch Pharmacol. 355: 457-462.
Taiz, L., Zeiger, E.. 2002. Auxin: the growth hormone. Murphy,
Angus. Plant Physiology,Tthird Addition. 423-460.
Wendehenne, D., Durner, J., Klessig, D. F. 2004. Nitric oxide: a
new player in plant signaling and defense responses. Current
Opinion in Plant Biology. 7:449-455.
Yamato, Tadahiko, Bing, Richard. 2000. P.S.E.B.M. Vol 225:
200-206.
Drug Concentration
0 100 200 300 400
Gra
vitro
pic
Re
ori
en
tatio
n (
Re
lative
to c
ontr
ol)
-1.0
-0.5
0.0
0.5
1.0
1.5
LNAME
Viagra
Figure 5. L-Name and Viagra disrupt the gravitropic response
of horizontally-oriented, gravistimulated roots. These plants
were grown on agar media containing experimental drug
concentrations that were turned so the plants were horizontal.
Reorientation angle was measured after 24 hours of
gravistimulation and compared with controls.
cPTIO Concentration (µM)
0.1 1 10 100 1000
Roo
t E
lon
gation
(R
ea
ltiv
e g
row
th r
ate
)
1.1
1.2
1.3
1.4
1.5
1.6
1.7
1.8
Measured
Hilland Plot
LNAME µM
0 100 200 300
Ro
ot
Elo
ng
ation
(R
ela
tive G
row
th R
ate
)
0.9
1.0
1.1
1.2
1.3
1.4
1.5
1.6
Figure 4: Pharmacological dose response analysis (Hilland
plot) of cPTIO and LNAME. Arabidopsis root growth increased
along with the concentration of cPTIO in the experiment. Low
concentrations of L-Name slowed the growth of Arabidopsis
compared to the control and high concentrations increased
growth. The Arabidopsis plants were grown vertically on agar
plates containing the drugs. Growth rate was measured after 24
hours and compared to the control.

Gravitational and Space Biology 19(2) August 2006 159
REGULATION OF TRANSCRIPTION IN ROOTS OF ARABIDOPSIS GRAVITY MUTANTS.
Jessie W. Yester1, Jeffery M. Kimbrough
1, Raul Salinas-Mondragón
1
, Patrick H. Masson3, Christopher S.
Brown1,2
, and Heike Winter Sederoff1*
1
Dept. of Botany, and 2
Kenan Institute for Science and Engineering, North Carolina State University, Raleigh
27695; 3Laboratory of Genetics, University of Wisconsin, Madison, WI, USA. * corresponding author:
Plant growth is guided by gravity. The mechanism by
which plants sense gravity is not understood.
Sedimentation of starch-containing amyloplasts (statolith)
in the columella cells is involved in the perception of
changes in the orientation towards the vector of gravity.
Inositol-1,4,5,-triphosphate (InsP3), Ca2+
, and proton
transport, as well as phytohormones (auxin, cytokinin,
ethylene, brassinosteroids), reactive oxygen species, and
nitric oxide have been shown to mediate the gravity
induced signal transduction. Several factors involved in
gravitropism have been identified by mutants defective in
their response to changes in their orientation towards
gravity. In order to elucidate the role of some of these
signal transduction elements in their regulation of gravity
specific transcription, we analyzed temporal changes in
gravity specific up-regulated genes in mutant and their
respective wildtype backgrounds (ecotypes) after
reorientation.
Increases in transcript abundances of some genes occur
within 1 minute after reorientation in the Arabidopsis root
apex, and are regulated specifically by gravity, not
mechanical stimulation (Kimbrough et al. 2004).
Focusing on three of these gravity-specific genes, we
present evidence that gravity-induced transcript
abundance changes of these genes are not strictly
conserved between ecotypes. In the root gravity response
mutants arg1 (defective in a DNAJ protein; Sedbrook et
al. 1999) and arl2 (defective in DNAJ protein similar to
ARG1, Guan et al. 2003), transcript abundance changes in
those selected gravity specific genes were not initiated.
This indicates that the DNAJ protein is required for
gravity-induced transcriptional regulation. In the gravity
response mutant adk1 (defective in adenosine kinase,
Young et al., unpublished) transcript abundance of one
gene was up-regulated, while two other genes did not
show an increase in transcript abundance after
reorientation. This indicates that transcriptional regulation
of those genes requires different signal transduction
elements in response to gravity stimulation.
Methods
We harvested Arabidopsis root apices (7 day old
etiolated seedlings) in order to analyze their relative
changes in transcript level in response to gravistimulation.
For our time course experiment, approximately 150 root
tips were harvested before (0 time point) and 2, 5, 15 min
after 135◦ reorientation by pouring RNAlater (Ambion)
onto the plates and cutting off the root apex (~7.5 mm).
Total RNA was extracted, purified with RNeasy (Qiagen),
and cDNA was synthesized using gene specific primers.
Arabidopsis gravity signaling mutants arg, arl, and adk
along with wild type controls were grown for seven days
in the dark, reoriented (135◦) and time-dependent changes
in transcript abundance measured for the gravity specific
genes (At4g23670; At5g38020; At5g48010).
Results
The gravity-response mutant plants used in this study
were derived from the Arabidopsis ecotypes
Wassilewskja (mutants arg, arl) and Landsberg (mutant
adk). We measured gravity-induced transcript abundance
changes in these ecotypes in comparison to Columbia-O
which was used in the microarray study (Kimbrough et al.
2004). We found that there are differences in the gravity-
induced transcriptional regulation of specific genes
between different Arabidopsis ecotypes in both temporal
profile and magnitude of regulation. (Figures 1 A,B,C).
Figure 1: Transcript abundance changes of At4g23670,
At5g38020, and At5g48010 in response to reorientation
in Arabiodopsis ecotype Columbia (A), Wassilewskija (B)
and Landsberg (C).
0
1
2
3
0 5 10 15
At4g23670
At5g38020
At5g48010
Columbia (Col)
min after reorientation
tra
nsc
rip
t a
bu
nd
an
ce f
old
ch
an
ge
A
0
1
2
3
0 5 10 15
At4g23670
At5g38020
At5g48010
tran
scrip
t ab
un
dan
ce f
old
ch
an
ge
min after reorientation
Wassilewskija (WS)B
0
1
2
3
0 5 10 15
At4g23670
At5g38020
At5g48010
tra
nsc
rip
t a
bu
nd
an
ce f
old
ch
an
ge
min after reorientation
Landsberg (Ler)C

J.W. Yester et al. – Regulation of Transcription in Roots of Arabidopsis Gravity Mutants.
160 Gravitational and Space Biology Bulletin 19(2) August 2006
DNAJ protein is essential for gravity induced
transcription
ARG and ARL code for DNA-J proteins which are
essential for gravity-induced regulation of transcript
abundance changes for the genes we tested. Reorientation
of these gravity mutants did not induce any significant
increases in transcript abundance of the genes tested
compared to their respective ecotype (Figures 2 A,B).
0
1
2
3
0 2 4 6 8 10 12 14min after reorientation
At4g23670
At5g38020
At5g48010
arl(ecotype Ws)
A
tra
nsc
rip
t a
bu
nd
an
ce f
old
ch
an
ge
0
1
2
3
0 5 10 15min after reorientation
At4g23670
At5g38020
At5g48010
arg(ecotype Ws)
B
tra
nsc
rip
t a
bu
nd
an
ce f
old
ch
an
ge
Figure 2: Transcript abundance changes of At4g23670,
At5g38020, and At5g48010 in response to reorientation
in Arabiodopsis mutants arl (A), and agr (B)
Adenosine kinase (ADK) regulates transcription of some
genes probably involved in hormone synthesis
ADK is involved in modulating root-tip architecture
and gravitropism in the Arabidopsis root tip. Mutation in
this gene alters gravitropic sensitivity (Masson et al.,
unpublished). Transcript levels of At5g38020 (SamT-
functions in Ado-Met pathway as methylating enzyme)
and At5g48010 (AtPEN1 – involved in brassinosteroid
synthesis) do not increase in the root apex after gravity
stimulation, while transcriptional regulation of At4g23670
(Major latex protein-like) is enhanced or at least not
repressed. This indicates that ADK is part of a signal
transduction pathway that activates transcriptional
regulation for At5g38020 and At5g48010. It is therefore
“downstream” of arg and arl in the signal transduction
pathway.
0
1
2
3
4
5
6
0 5 10 15min after reorientation
At4g23670
At5g38020
At5g48010
adk(ecotype Ler)
transc
rip
t abu
ndance
fold
chan
ge
Figure 3: Transcript abundance changes of At4g23670,
At5g38020, and At5g48010 in response to reorientation
in Arabiodopsis mutant adk.
Conclusions
� We measured variation in the gravity-induced
transcriptional regulation of specific genes between
different Arabidopsis ecotypes.
� ARG and ARL code for proteins which are essential for
gravity-induced regulation of transcript abundance
changes for at least those genes tested.
� ADK expression is required for gravity-induced
transcriptional regulation of some specific genes
(At5g38020, At5g48010) possibly involved in hormonal
regulation.
At4g23670
GS ARG, ARL
ADK
At5g38020
At5g48010
Figure 4: The transcriptional regulation induced by
gravity stimulation (GS) requires the DNAJ proteins ARG
and ARL. The transcription of the Major-latex like gene
(At4g23670) does not require functional expression of
ADK.
(Supported by NASA grant NAG2-1566 to C.S.B. and
North Carolina Space Consortium grant to J.W.Y.)
REFERENCES
Guan, C., Rosen E., Boonsirichai, K., Poff, K., Masson, P. 2003.
The ARG1-LIKE2 gene of Arabidopsis functions in a gravity
signal transduction pathway that is genetically distinct from
the PGM pathway. Plant Physiol. 133: 100-112.
Kimbrough, J., Salinas-Mondragon, R., Boss, W., Brown, C.,
Sederoff H. 2004. The fast and transient transcriptional
network of gravity and mechanical stimulation in the
Arabidopsis root apex. Plant Physiol. 136: 2790-2805.
Sedbrook, J., Chen, R., Masson, P. 1999. Arg1 (altered response
to gravity) encodes a DNAJ-like protein that potentially
interacts with the cytoskeleton. Proc. Natl. Acad. Sci. USA 96:
1140–1145.

Gravitational and Space Biology Bulletin 19(2) August 2006 161
MODIFICATION OF RESERVE DEPOSITION IN WHEAT AND BRASSICA SEEDS BY SYNTHETIC
ATMOSPHERES AND MICROGRAVITY
Anxiu Kuang1, John Blasiak2, S. Chen1, Gail Bingham3, Mary E. Musgrave2
1Dept. of Biology, Univ. of Texas Pan American, Edinburg, TX 78541; 2Dept. of Plant Science, Univ. of
Connecticut, Storrs, CT 06269; 3Space Dynamics Lab, Utah State Univ., Logan, UT 84341
Movement of gases in microgravity may be reduced due
to a lack of buoyancy-driven convection (Musgrave, et
al., 1997). We are investigating how seed development in
microgravity could be modulated through changes in
metabolic gas bioavailability (Blasiak et al., 2006). To do
this we have grown wheat (Triticum aestivum L. cv.
Apogee) and Brassica (Brassica rapa L.) in synthetic
atmospheres in which the bulk gas (normally N2 in
ambient air) is replaced by either Ar (slowing diffusion)
or He (speeding diffusion). The seeds produced in the
synthetic atmospheres were then compared with seeds
grown in microgravity on the Mir space station in the Svet
greenhouse. Since starch is the major component of
wheat seeds, while in Brassica protein and lipid are the
major storage reserves, we have compared how these
storage reserves changed in microgravity, ground control,
and synthetic atmosphere environments.
Wheat (Triticum aestivum L. cv. Apogee) and Brassica
(Brassica rapa L.) plants were grown from seed-to-seed
on the Mir space station in the Svet greenhouse. Mature
dry seeds were collected and subsequently examined by
light and electron microscopy. In ground-based studies,
wheat and Brassica plants were grown in synthetic
atmospheres differing in the bulk carrier gas (He, N2 or
Ar), but containing earthnormal concentrations of O2
(21%) and CO2 (400 ppm), to produce treatments in
which the metabolic gases would diffuse at different rates.
The plants were grown under fluorescent lights
(continuous light, 120 mmol/m2/sec, PAR) using a wick-
fed hydroponic system. Gas-flow through the chambers
was approximately 350 ml/min. Dry seeds of wheat and
Brassica were colleted and immature fresh seeds (18 d
post-pollination) of Brassica were fixed and processed for
microscopy using procedures as in Kuang et al. (2000).
Analysis of the wheat seeds produced on the Mir space
station by scanning electron microscopy revealed that
starch grains were deformed, compared to the ground
seeds (Fig. 1). Those starch grains had a rough surface
with microholes that were evenly distributed on the
surface (Fig. 1). In wheat seeds from the synthetic
atmospheres, starch grains revealed comparable
morphology and structures in the He and air (N2)
treatments, however the Ar treatment had smaller,
rounder grains compared to the air (N2) control treatment
(Fig. 2).
Brassica seeds produced on Mir showed retention of
starch grains while lipid and protein were the major
storage reserves in mature ground seeds. Protein bodies
were also a major reserve in Mir seeds but their size was
diminished (Musgrave et al, 2000). Brassica seeds
produced in all of the synthetic atmosphere treatments had
lipid and protein as the major storage reserves, however,
protein bodies in the helium treatment were larger
compared to those in the nitrogen treatment (Fig. 3),
which might have contributed to the greater weight of the
seeds (Table 1). In immature seeds, protein bodies were
smaller and starch grains persisted in embryo cells in the
Ar treatment, while protein and lipid were major reserves
in the seeds produced in the N2 chamber, indicating that
development of Brassica seeds was retarded in the Ar
atmosphere (Fig. 4).
Table 1. Mean seed weight, number of protein bodies per
cell, and mean diameter (µm) of protein bodies in Brassica
dry seeds from the second helium and argon experiments.
Treatment Seed dry Protein bodies Diameter of
weight (mg) (n/cell) protein (µm)
Air 1.84 2.7 6.02
Helium 2.05 2.7 7.54
P-value 0.04 ns 0.002
Air 1.54 2.3 6.80
Argon 1.43 3.0 6.08
P-value ns 0.06 0.09
The results show that small differences in metabolic gas
diffusion rates lead to ultrastructural changes in seed
storage reserves. Starch grains of wheat from the He and
N2 treatments were comparable. While the starch grains
from the Ar treatment did not exhibit the abnormalities
found in the flown material (Fig. 2), the grains were
smaller and rounder than in the N2 treatment. Changes in
starch grain morphology in the spaceflight environment
have been reported by others (Kuznetsov et al., 2001).
Synthetic atmosphere experiments show that diffusion
rate affects development of Brassica seed-storage
components. Development in a He atmosphere enhanced
the size of protein storage bodies and mature seed dry
weight. In contrast, the reduced-diffusion rate of the Ar
atmosphere retarded development: protein bodies were
smaller than in the control treatment, and starch grains
were retained later in seed development. The development
of Brassica seed-storage components under the reduced-
diffusion rates of the argon atmosphere was similar to the
pattern observed under microgravity conditions (Kuang et
al., 2000; Musgrave at al., 2000)
REFERENCES
Blasiak, J, A Kuang, CS Farhangi, and M. E. Musgrave. 2006.
Roles of intra-fruit oxygen and carbon dioxide in controlling
pepper (Capsicum annuum L.) seed development and storage

A. Kuang et al. – Simulation of Changes in Seed Storage Reserves in Microgravity
162 Gravitational and Space Biology Bulletin 19(2) August 2006
reserve deposition. J. Amer. Society of Horticultural Science,
131(1):164-173.
Kuang, A, Y Xiao, G. McClure, and ME Musgrave (2000).
Influence of Microgravity on ultrastructure and storage reserves
in seeds of Brassica rapa L. Annals of Botany, 85:851-859.
Kuznetsov, OA, CS Brown, HG Levine, WC Piastuch, MM
Sanwo-Lewandowski, and KH Hasenstein. Composition and
physical properties of starch in microgravity-grown plants.
Advances in Space Research (2001), 28 (4): 651-658
Musgrave, ME, A. Kuang, and SW Matthews (1997). Plant
reproduction during spaceflight: importance of the gaseous
environment. Planta, 203: s177-s184.
Musgrave, ME, A. Kuang, Y Xiao, GE Bingham, L. G. Briarty,
M.A Levinski, VN. Sychev, amd IG Podolski (2000). Gravity
independence of seed-to-seed cycling in Brasicca rapa. Planta,
210(3): 440-406.
Funded by NASA grants NAG2-1375 and NAG10-329 to MEM
and AK.
Figure 1. SEM micrographs showing starch grains in wheat
seeds produced on the Mir space station (A & B) and in the
ground control (C & D). Note that microholes (arrowheads)
are present on the surface of starch grains in the spaceflight
seeds. Bars = 10 µm
Figure 2. SEM micrograph of starch grains in wheat seeds
produced in the synthesized atmosphere experiments. A), air
control for helium treatment; B), helium treatment; C), air
control for argon treatment; D), argon treatment. Bars = 10 µm
Figure 3. Ultrastructure of cotyledon cells of dry Brassica
seeds produced in the air chambers (A & C), helium chamber
(B), and argon chamber (D). Note that both protein bodies and
lipid are major storage reserves but protein bodies (pb) in seeds
produced in the helium chamber (B) are lager in size (see Table
1) compared to seeds produced in air the chamber (A). Bars =
4.2 µm
Figure 4. Ultrastructure of cotyledon cells of Brassica seeds
(18 d old) produced in the argon chamber (A & B) and in the air
chamber (C & D). Starch grains (arrows) were present (A) and
protein bodies (pb) were relatively smaller (B) in seeds from the
argon chamber, compared to the seeds from the air chamber (C
& D). The nucleus (nu) was obvious, whereas the nucleus in
seeds produced in the air chamber became shrunken (D)
indicating that dehydration was in progress. Bars = 3.9 µm.
A
B
C
D
pb
pb
pb
pb
pb pb
pb
pb
pb
A
B
C
D
nu
nu
A B
C D
A B
CD

Gravitational and Space Biology 19(2) August 2006 A - 1
Author Index
Agricola, H-J., 135
Allen, C., 143
Allen, P.L., 145
Ask, M.A., 129
Baer, L., 141
Baker, T.L., 145
Barnstable, A.J., 141
Barnstable, C.J., 141
Benoit, M.R., 31
Bhattacharya, S., 127, 133
Bingham, G., 161
Blasiak, J., 161
Blazevic, E., 133
Boling, J., 91
Boston, P.J., 91
Bourget, C.M., 19
Brown, C.S., 159
Brzezinski, E., 53
Campbell, K.A., 91
Chang, J., 133
Chen, S., 161
Chopra, A., 143
De Carlo, A.R., 123
Duke, J., 137
Echon, C.M., 53
Elliott, T.F., 145
Elmer, J., 157
Emmerich, J.C., 19
English, J., 137
Fahlen, T.F., 133
Ferl, R.J., 3
Flowers, R., 139
Galindo, C., 143
Goli, N., 139
Graham, J.M., 105
Gupta, K.B., 125
Gyotoku, J-I., 153
Hammond, D.K., 145
Hammond, T.G., 145
Hargens, A.R., 53
Hawkins, A., 53
Hawkins, K., 53
Hildenbrand, C.D., 155
Holubec, K., 145
Horn, E.R., 135
Houston, E., 127
Howard, A., 127
Irazoqui, P.P., 123
Jacobi, J., 157
Johnson, C.B., 147
Kakavand, A., 127
Kimbrough, J.M., 159
Kirschnick, U., 135
Klement, B., 139
Kuang, A., 161
Kucik, D.F., 125
Kurk, M.A., 131
Kuznetz, L.H., 85
Lee, R., 137
Lera, M., 133
Leskovsky, D., 127
Levine, H.G., 129
Love, J.E., 145
Lynch, S.V., 31
Maclas, B.R., 53
Maldonado, A., 127
Marshall, C., 139
Masini, M.A., 149
Massa, G.D., 19
Masson, P.H., 159
Matin, A., 31
McLamb, W.T., 123
McSpadden, T., 91
McWilliams, L., 91
Metz, G.W., 131
Meyer, R.S., 53
Mitchell, C.A., 19
Morrow, R.C., 19
Murakami, N., 153
Musgrave, M.E., 161
Nadella, V., 155
Niesel, D., 143
Norikane, J., 129

Author Index
A - 2 Gravitational and Space Biology 19(2) August 2006
Pandya, U., 143
Park, H., 153
Pastorino, M., 149
Paul, A-L., 3
Paulsen, D., 139
Pellis, N.R., 151
Porterfield, D.M., 123, 157
Prenger, J.J., 129
Ricci, F., 149
Risin, D., 151
Risin, S.A., 151
Rokkam, M., 123
Rouleau, R.L., 125
Roux, S.J., 123
Rouzan-Wheeldon, D., 129
Russell, K., 157
Rygalov, V., 129
Salinas-Mondragón, R., 159
Sanchez, M.E., 127, 133
Shenasa, M., 127
Shiba, D., 153
Smith, L.W., 125
Sonnenfeld, G., 45
Soundur, R., 157
Steele, M.K., 127
Stein, T.P., 49
Strollo, F., 149
Takaoki, M., 153
Teichgraeber, J., 137
Thomas, D.J., 91, 131
Thomas, N.A., 131
Tink, A.R., 141
Todd, P., 79, 91, 131
Tombran-Tink, J., 141
ul Haque, A., 123
Uva, B.M., 149
Vandromme, M., 139
Viviano, S., 141
Wade, C.E., 65, 141
Waldbeser, L.S., 147
Ward, N.E., 151
Wells, H.W., 123
Wereley, S.T., 123
Williams, N., 143
Winter Sederoff, H., 159
Wu, X., 125
Wyatt, S.E., 155
Yester, J.W., 159




















