
Don’t Smash Those Bones! Anatomical Representation and Bone Tool Manufacture in the Pampean Region...
14
Bones for tools – tools for bones The interplay between objects and objectives Edited by Krish Seetah & Brad Gravina ISBN: 978-1-902937-59-5 ISSN: 1363-1349 (McDonald Institute) © 2012 McDonald Institute for Archaeological Research All rights reserved. No parts of this publication may be reproduced, stored in a retrieval system, or transmied, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior permission of the McDonald Institute for Archaeological Research. Publisher contact information: McDonald Institute for Archaeological Research University of Cambridge, Downing Street, Cambridge, UK, CB2 3ER (0)(1223) 333538 (0)(1223) 339336 (Production Office) (0)(1223) 333536 (FAX) [email protected] www.mcdonald.cam.ac.uk Distributed by Oxbow Books United Kingdom: Oxbow Books, 10 Hythe Bridge Street, Oxford, OX1 2EW, UK. Tel: (0)(1865) 241249; Fax: (0)(1865) 794449 USA: The David Brown Book Company, P.O. Box 511, Oakville, CT 06779, USA. Tel: 860-945-9329; Fax: 860-945-9468 www.oxbowbooks.com Chapter 10 Don’t Smash Those Bones! Anatomical Representation and Bone Tool Manufacture in the Pampean Region (Argentina, South America) Daniel Loponte and Natacha Buc How to cite this chapter: Loponte, D. & N. Buc, 2012. Don’t smash those bones! Anatomical representation and bone tool manufacture in the Pampean region (Argentina, South America), in Bones for Tools – Tools for Bones: the Interplay Between Objects and Objectives, eds. K. Seetah & B. Gravina. (McDonald Institute Monographs.) Cambridge: McDonald Institute for Archaeological Research, 117–29.
Transcript of Don’t Smash Those Bones! Anatomical Representation and Bone Tool Manufacture in the Pampean Region...

Bones for tools – tools for bonesThe interplay between objects and objectives
Edited by Krish Seetah & Brad Gravina
ISBN: 978-1-902937-59-5ISSN: 1363-1349 (McDonald Institute)
© 2012 McDonald Institute for Archaeological Research
All rights reserved. No parts of this publication may be reproduced, stored in a retrieval system, or transmitted, in any form or by any means, electronic, mechanical, photocopying, recording or otherwise, without the prior permission of the McDonald Institute for Archaeological Research.
Publisher contact information:McDonald Institute for Archaeological ResearchUniversity of Cambridge, Downing Street, Cambridge, UK, CB2 3ER(0)(1223) 333538(0)(1223) 339336 (Production Office)(0)(1223) 333536 (FAX)[email protected]
Distributed by Oxbow BooksUnited Kingdom: Oxbow Books, 10 Hythe Bridge Street, Oxford, OX1 2EW, UK.Tel: (0)(1865) 241249; Fax: (0)(1865) 794449USA: The David Brown Book Company, P.O. Box 511, Oakville, CT 06779, USA.Tel: 860-945-9329; Fax: 860-945-9468www.oxbowbooks.com
Chapter 10Don’t Smash Those Bones! Anatomical Representation and Bone Tool Manufacture in the Pampean Region (Argentina, South America)
Daniel Loponte and Natacha Buc
How to cite this chapter:Loponte, D. & N. Buc, 2012. Don’t smash those bones! Anatomical representation and bone tool manufacture in the Pampean region (Argentina, South America), in Bones for Tools – Tools for Bones: the Interplay Between Objects and Objectives, eds. K. Seetah & B. Gravina. (McDonald Institute Monographs.) Cambridge: McDonald Institute for Archaeological Research, 117–29.

117
Don’t Smash Those Bones!
Chapter 10
Don’t Smash Those Bones! Anatomical Representation and Bone Tool Manufacture in the Pampean Region
(Argentina, South America)
Daniel Loponte and Natacha Buc
nutritive processes of bone destruction due to human and post-depositional activity (Blumenschine 1986; 1989; Lyman 1994; Marean & Cleghorn 2003). Even so, less attention has been paid to SEA in relation to bone technology. Undoubtedly, in general terms the presence of bone tools is a marginal component of a bone assemblage. However, their study along with the analysis of operational sequences could be useful for reaching an alternative or complementary SEA picture. In our case, this methodological approach is necessary to understand ungulate anatomical representation patterns in the archaeological sites under investigation.
Archaeological SEA studies, despite analysis of taphonomical forces, belong to a set of optimization models known as ‘optimal-foraging theory’ (Kramer 2001; Krebs et al. 1987; Stephens 1990; Stephens & Krebs 1986) that offer a framework for interpreting zoo archaeological data. These models employ con-cepts such as ‘prey and patch selection’ and ‘central place foraging (CPF)’ to predict foraging strategy, length of time spent searching for food prey species acquired and butchering strategies. Consequently, these models also predict and explain skeletal representation of different preys in archaeofaunal assemblages (Broughton 1994a,b; Broughton & Grayson 1993; Cannon 2003; Grayson 2001; Gray-
Bone tools are one of the main features of hunter-gatherer campsites in the lower Paraná wetland (Argentina, South America) dated between 1.0 and 0.6 uncal. bp. This chapter examines how the selection of bone elements used to make tools limits the percentage of minimum animal units (%MAU) of ungulates established for archaeofaunal assemblages in the area. For that purpose, we analysed the manufacturing process involved in the creation of archaeological bone tools. This information is evaluated relative to deer anatomical representation, bone mineral density (BMD) and food utility index (FUI). Results suggest that analysis of anatomical representation of faunal resources requires a careful study of
manufacturing processes.
The analysis of bone tools has a long tradition in Western archaeology (Backwell & d’Errico 2001; Camps-Fabrer 1967; d’Errico & Backwell 2003; Lyman 1994; Newcomer 1974; Olsen 1979; Poplin 1974; Sadek Kooros 1972; Stordeur 1980). Currently, there are a number of specialized methodological approaches ranging from morphological (Aver-bouh & Catellain 2001; Campana 1989; Julien 1986; Pétillon 2001, among others) to mechanical aspects (Guthrie 1983; Knecht 1997; Scheinsohn 1997) and microwear analysis (Averbouh 2000; Campana 1989; Griffitts 2006; Legrand 2005; Maigrot 2003; Le Moine 1991; Peltier 1986; Peltier & Plisson 1989). Although archaeologists point out the importance of analysing bone tools relative to faunal structure (Hodgetts 2000; Hodgetts & Rahemtulla 2001), such studies are not abundant in the archaeological literature (Choyke 2003; Scheinsohn et al. 1992; Sidéra 1989; 1991; 2000). Furthermore, there is an emphasis in recent zooarchaeological papers on solving problems of equifinality inherent to skeletal element abundance (SEA) (Bar-Oz & Munro 2004; Cleghorn & Marean 2004; Enloe 2004; Lyman 2004; Marean & Cleghorn 2003; Munro & Bar-Oz 2005). These studies have focused on the relationships between SEA and post-encounter return rates as well as nutritive and non-
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

118
Chapter 10
son & Delpech 1998; Grayson et al. 2001; Kaplan & Hill 1992; Metcalfe & Barlow 1992; Nagaoka 2001; O’Connell 1995; Orians & Pearson 1979; Smith 1983; Stiner et al. 2000). One of the basic assumptions of CPF is that as distance of prey capture increases, transport of low-utility skeletal parts to base camps decreases, thus increasing delivery rate or the amount of energy carried back to a central place per unit foraging time (Cannon 2003; Orians & Pearson 1979). This behaviour has also been documented from theoretical approaches that are not explicitly framed in optimal-foraging studies (Binford 1978; White 1954). Nevertheless, in some contexts, specific bones used as raw material introduce another variable in hunter-gatherer decisions regarding which elements are returned to residential sites. Equally, in contexts where bone fracturing for nutrient extraction results in fragmentation (Lyman 2003; Munro & Bar-Oz
2005), bone selection for tool-making increases the probability of identification. The aim of this chapter is to focus on certain aspects linked to bone technology in order to better understand SEA in two archaeo-logical assemblages where bone tools are quantitatively significant relative to the number of identified specimens (NISP) of all taxa. We analyse representation patterns of Ozotoceros bezoarticus (pampas deer1) in the faunal assemblages and then add bone tools and manufacture by-products (dis-carded after tool manufacturing).
Faunal assemblage — general features
The assemblages analysed in this chapter come from the Anahí (1020±70 uncal. bp, Beta-147108) and La Bellaca 2 (680±80 uncal. bp, LP-1263)2 archaeological sites. Both are situated in the lower Paraná’s wetland (Pampean region, Argentina: Fig. 10.1) and have similar taphonomic conditions (Acosta 2005).
Hunter-gatherer groups in the late Holocene created both of the bone assem-blages under study. Archaeological materi-als were extracted from an approximately 50 cm-thick soil horizon. This soil had a neutral to slightly acidic pH and was affected by pooling processes and the action of ground-water. Consequently, bones are highly min-eralized by deposition of iron (Fe), strontium (Sr) and manganese (Mn) salts. This not only changed the colour of the bones, but also increased the bone mineral density
(BMD), thus augmenting their survival (Musali et al. 2003; Acosta 2005). Table 10.1 shows BMD, Sr, Fe and Mn changes in the same bone elements of fish from the Paraná system and the archaeological sites. The sites under analysis were located in a low-energy environment with significant vegetal cover. These conditions would probably have led to rapid burial of the archaeological assemblage (Acosta 1997; 2005).
In contrast, mammal bones showed weathering to stages 1–2 (cf. Behrensmeyer 1978) and a low occur-rence of carnivore and rodent gnawing (<1 per cent of NISP). Anatomical representation, both in mam-mals and fishes, is not correlated with bone mineral density (Acosta 2005; Acosta et al. 2004; 2010; Loponte & Acosta 2004; Musali 2005) hence SEA is not linked with BMD.
The archaeological record at the Anahí and La Bellaca 2 sites included incised and plain pottery
Buenos AiresBuenos Aires
A R G E N T I N AA R G E N T I N A
U R U G U AY
U R U G U AY
Paraná Urugu
ay
Lower Paraná wetlandLower Paraná wetland
58°60°
32°
34°
58°60°
32°
34°
Archaeological sites
Drawn by D. Kemp
La Bellaca 2La Bellaca 2
Anahí Anahí
Figure 10.1. Lower Paraná’s wetland, Pampean region, Argentina.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
119
Don’t Smash Those Bones!
(Pérez & Cañardo 2004). Furthermore, projectile points made from quartzite and chalcedonies were recovered (Loponte 2008; Loponte & Acosta 2003; Loponte & Sacur Silvestre 2003). Pottery refitting and artefactual techno-typological features suggest that each site represents either the deposition of refuse as a single event, or smaller deposits with similar characteristics. However, the existence of less-dense occupational events cannot be rejected (Acosta 2005; Pérez & Cañardo 2004).
The recovered fish bones were mainly from the orders Siluriformes and Characiformes. Two rodents (Myocastor coypus and Cavia aperea) and two cervids (Blastocerus dichotomus: marsh deer, and Ozotoceros bezoarticus: pampas deer) were also noted in the assemblage. Other taxa are present but with very low NISP values, represented by one or a few individuals, which could have been attributed to eto-ecological causes and/or opportunistic hunting (Acosta 2005; García Esponda et al. 2001; Loponte 2008; Loponte & Acosta 2004; Musali 2005; Teta et al. 2004) (Table 10.2).
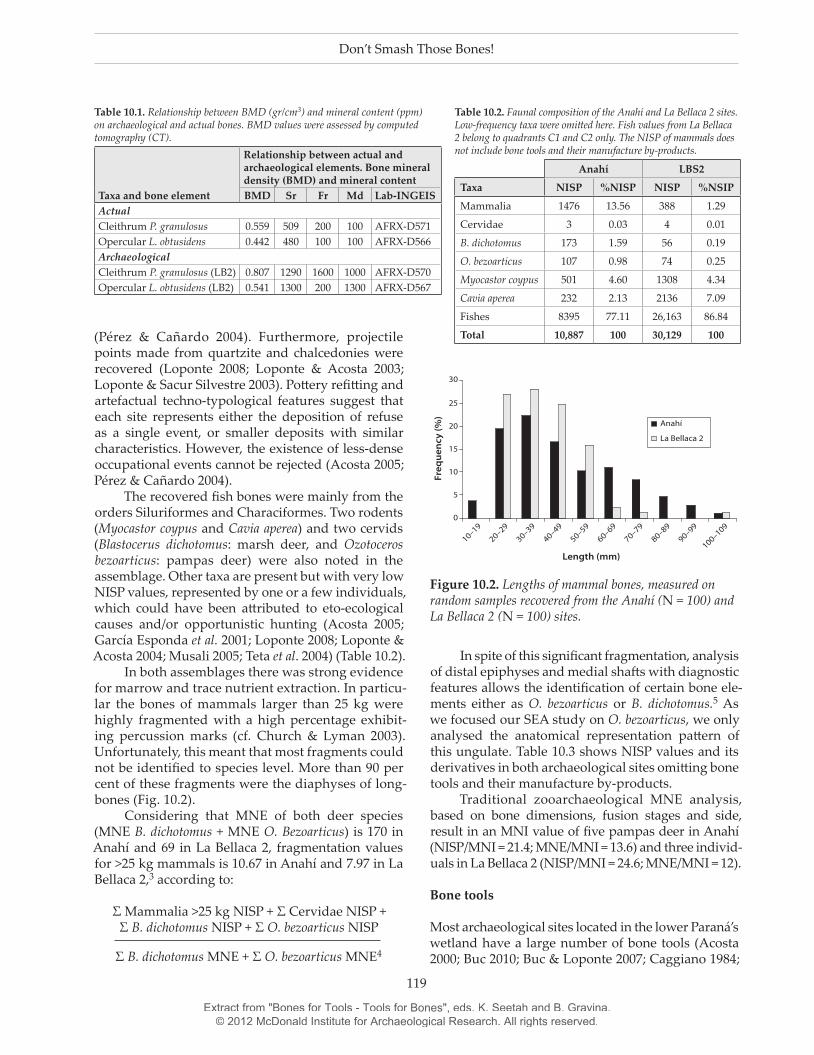
In both assemblages there was strong evidence for marrow and trace nutrient extraction. In particu-lar the bones of mammals larger than 25 kg were highly fragmented with a high percentage exhibit-ing percussion marks (cf. Church & Lyman 2003). Unfortunately, this meant that most fragments could not be identified to species level. More than 90 per cent of these fragments were the diaphyses of long-bones (Fig. 10.2).
Considering that MNE of both deer species (MNE B. dichotomus + MNE O. Bezoarticus) is 170 in Anahí and 69 in La Bellaca 2, fragmentation values for >25 kg mammals is 10.67 in Anahí and 7.97 in La Bellaca 2,3 according to:
Σ Mammalia >25 kg NISP + Σ Cervidae NISP + Σ B. dichotomus NISP + Σ O. bezoarticus NISP
Σ B. dichotomus MNE + Σ O. bezoarticus MNE4
Table 10.1. Relationship between BMD (gr/cm3) and mineral content (ppm) on archaeological and actual bones. BMD values were assessed by computed tomography (CT).
Taxa and bone element
Relationship between actual and archaeological elements. Bone mineral density (BMD) and mineral contentBMD Sr Fr Md Lab-INGEIS
ActualCleithrum P. granulosus 0.559 509 200 100 AFRX-D571Opercular L. obtusidens 0.442 480 100 100 AFRX-D566ArchaeologicalCleithrum P. granulosus (LB2) 0.807 1290 1600 1000 AFRX-D570Opercular L. obtusidens (LB2) 0.541 1300 200 1300 AFRX-D567
Table 10.2. Faunal composition of the Anahí and La Bellaca 2 sites. Low-frequency taxa were omitted here. Fish values from La Bellaca 2 belong to quadrants C1 and C2 only. The NISP of mammals does not include bone tools and their manufacture by-products.
Anahí LBS2
Taxa NISP %NISP NISP %NSIP
Mammalia 1476 13.56 388 1.29
Cervidae 3 0.03 4 0.01
B. dichotomus 173 1.59 56 0.19
O. bezoarticus 107 0.98 74 0.25
Myocastor coypus 501 4.60 1308 4.34
Cavia aperea 232 2.13 2136 7.09
Fishes 8395 77.11 26,163 86.84
Total 10,887 100 30,129 100
Length (mm)
Freq
uen
cy (%
)
0
5
10
15
20
25
30
10–19
20–29
30–39
40–49
50–59
60–69
70–79
80–89
90–99
100–109
Anahí
La Bellaca 2
Figure 10.2. Lengths of mammal bones, measured on random samples recovered from the Anahí (N = 100) and La Bellaca 2 (N = 100) sites.
In spite of this significant fragmentation, analysis of distal epiphyses and medial shafts with diagnostic features allows the identification of certain bone ele-ments either as O. bezoarticus or B. dichotomus.5 As we focused our SEA study on O. bezoarticus, we only analysed the anatomical representation pattern of this ungulate. Table 10.3 shows NISP values and its derivatives in both archaeological sites omitting bone tools and their manufacture by-products.
Traditional zooarchaeological MNE analysis, based on bone dimensions, fusion stages and side, result in an MNI value of five pampas deer in Anahí (NISP/MNI = 21.4; MNE/MNI = 13.6) and three individ-uals in La Bellaca 2 (NISP/MNI = 24.6; MNE/MNI = 12).
Bone tools
Most archaeological sites located in the lower Paraná’s wetland have a large number of bone tools (Acosta 2000; Buc 2010; Buc & Loponte 2007; Caggiano 1984;
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
120
Chapter 10
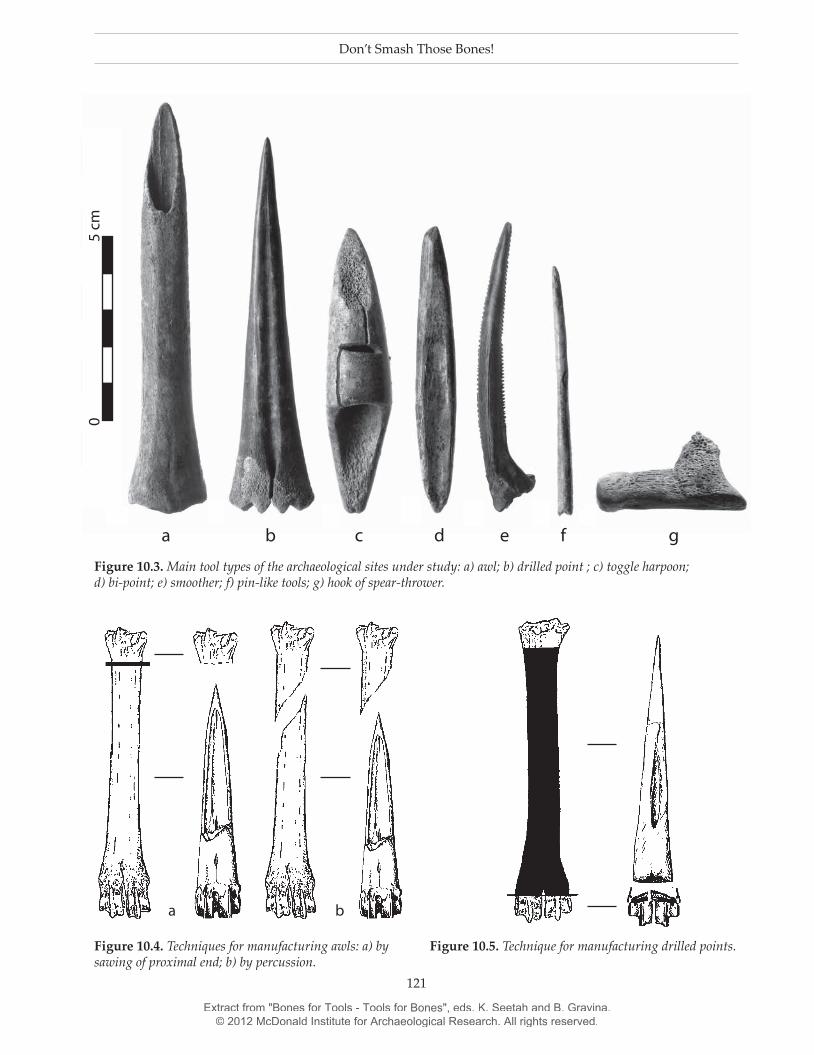
Lothrop 1932; Torres 1911). There were approximately 30 bone tools in Anahí and 90 in La Bellaca 2.7 The main morphological groups were correlated with specific bone elements (Acosta et al. 2010), for example the hooks of spear-throwers were made from astragali, while toggle harpoons were fabricated from antler. A large number of drilled points were fashioned from cervid metapodials or antler (Fig. 10.3; see Buc 2007 for a detailed discussion). Awls were made from O. bezoar-ticus metapodials and bi-points from the long-bones of mammals (almost certainly cervids) or antler. Finally, there are other tools shaped from fish spines such as smoothers, pins and needles8 (Acosta 2000; Acosta et al. 2010; Buc 2010; Buc & Loponte 2007; Fig. 10.3).
The size of spear-throwers’ hooks suggests they were made from the astragali of large mammals. A few of these hooks were recovered in their initial stages of manufacture and this allows us to identify the raw material as B. dichotomus. However, intense modification of finished hooks prevented specific or general taxonomical identification. This is also true for tools made on antler (bi-points and toggle harpoons): macroscopic taxonomic identification suggested that the material could have been sourced from O. bezoar-ticus, B. dichotomus or even Mazama guazoubira (a small
cervid species currently present in more northerly regions).
Fortunately, awls and drilled points were usually identifiable to element and taxon: they were regularly made from O. bezoarticus metapodials. The manufac-turing process for these tools resulted in not only the complete artefact but also in by-products. Figures 10.4 and 10.5 illustrate the probable manufacturing sequence for each tool type. During the manufacture of awls the proximal end of the metapodial was often discarded. Conversely, in the case of drilled points, it was the distal portion that was rejected.
Since awls were used during low-risk activities, such as piercing soft materials (mostly leather: Buc 2010; Buc & Loponte 2007), complete tools were recov-ered and, therefore, their morphological identification was straightforward. Inherent metrical variation sug-gests their apical ends had been re-touched through time. Consequently, it could be supposed that awls entered the campsite already manufactured. Because of this assumption, zooarchaeologists generally omit bone tools from archaeofaunal analysis (cf. Lyman 1994).
Awls manufactured using technique ‘a’ result in small bone tubes that have a low survival rate
Table 10.3. O. bezoarticus: skeletal element abundance in Anahí and La Bellaca 2.6
Anahí La Bellaca 2Element NISP MNE MAU %MAU Element NISP MNE MAU %MAUTeeth 22 Teeth 7Skull 1 1 1 25 Mandible 1 1 0.5 18.2Hemimandible 3 1 0.5 12.5 Axis 1 1 1 36.4Cervical vertebra* 7 7 1.4 35 Pelvis 2 1 1 36.4Thoracic vertebra 1 1 0.1 2.5 Scapula 2 2 1 36.4Lumbar vertebra 2 2 0.33 8.25 Humerus 2 2 1 36.4Sacrum 1 1 1 25 Femur 2 1 0.5 18.2Rib 3 3 0.1 2.5 Tibia 0 0 0 0Pelvis 3 1 1 25 Metapodial 40 11 2.75 100Scapula 8 8 4 100 Carpal (2nd & 3rd) 1 1 0.5 18.2Humerus 1 1 0.5 12.5 Tarsal (3rd & 4th) 3 3 1.5 54.5Radius 1 1 0.5 12.5 Calcaneus 1 1 0.5 18.2Ulna 2 2 1 25 1st phalanx 3 3 0.4 14.5Femur 1 1 0.5 12.5 2nd phalanx 3 3 0.4 14.5Patella 1 1 0.5 12.5 3rd phalanx 6 6 0.7 25.5Tibia 2 2 1 25 Total 74Metapodial 18 6 1.5 37.5Radiocarpal 2 2 1 25Tarsal (3rd & 4th) 6 6 3 75Calcaneus 5 5 2.5 62.5Astragalus 4 4 2 501st phalanx 5 4 0.5 12.52nd phalanx 2 2 0.25 6.253rd phalanx 6 6 0.75 18.75Total 107
*Excluding atlas and axis.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
121
Don’t Smash Those Bones!
0
5 cm
a b c d e f g
Figure 10.3. Main tool types of the archaeological sites under study: a) awl; b) drilled point ; c) toggle harpoon; d) bi-point; e) smoother; f) pin-like tools; g) hook of spear-thrower.
a b
Figure 10.4. Techniques for manufacturing awls: a) by sawing of proximal end; b) by percussion.
Figure 10.5. Technique for manufacturing drilled points.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

122
Chapter 10
owing to fragmentation; therefore, this method is very difficult to identify in the archaeological record. Awls created by technique ‘b’ can be identified by the presence of impact pits on the tool’s internal facets (cf. Camps-Fabrer & d’Anna 1977). Despite the risk of destroying the bones, controlled fracture may have been an expedient technique for initial reduction. Remains of this procedure are more fragmented than those of technique ‘a’ and thus identification of this activity is even harder.
The situation is different for drilled points where a segmentation of the distal end is used to drill the base. This activity produces a greater number of sawed distal condyles that are not prone to destruction by fragmen-tation and, consequently, are useful for taxonomic iden-tification. Therefore, the presence of sawed metapodial condyles in bone assemblages is direct evidence of on-site manufacture of these tools and hence, is a more accurate indicator of hunting events. Thus, the study of these artefacts makes an appropriate contribution to the zooarchaeological data.
As drilled points were possibly used for prey capture (Buc 2007; 2010; Loponte 2008), they would have been under high mechanical pressures. Conse-quently, they may have had to be frequently replaced, probably more often than harpoons made from ant-ler, and almost certainly, more than awls. Antler has structural properties more suitable to hunting activi-ties (i.e. appropriate to activities involving impact: cf. Guthrie 1983). Many broken drilled points were recovered with copious numbers of tool fragments (mostly indeterminate ‘point fragments’) likely to be parts of this type of artefact. Moreover, we must take into account the possibility of off-site breakage of drilled points during hunting, which would result in their discard. As the manufacturing technique for drilled points results in two sawed metapodial con-dyles, the high number of these bones found within the faunal assemblages indicate the existence of more drilled points than the quantity actually recovered. This situation suggests two possible explanations: 1) most of these drilled points were discarded off site; or 2) they cannot be morphologically identified because of fragmentation.
Data analysis
In Tables 10.4 and 10.5, the left-hand columns show skeletal element abundance (SEA) in Anahí and La Bellaca 2 sites respectively. Variations seen in the right-hand columns are due to adding bone tools and manufacture by-products (CSEA).
Both distributions permit the analysis of differ-ences and similarities between sites. In Anahí and La
Bellaca 2 differences are associated with the inclusion of drilled points (Anahí NISP = 1; LB2 NISP = 6) and awls (Anahí NISP = 2; LB2 NISP = 2). However, the most obvious variations are due to adding in the analysis of the sawed distal metapodial condyles of pampas deer (Anahí NISP = 23; LB2 NISP = 40).
When adding bone tools and by-products to the analysis, an increment in the representation of high-density bones can be expected. However, in our sam-ples there are no substantial changes in %MAU and BMD relationship between SEA and CSEA (Fig. 10.6).
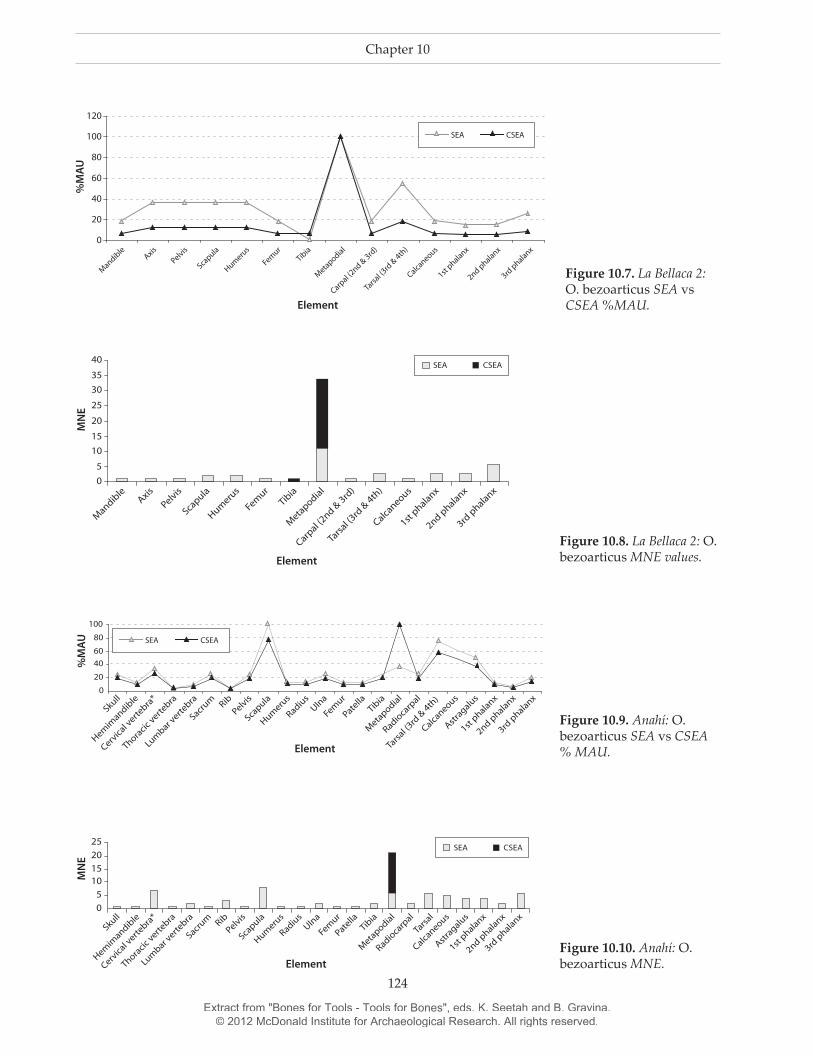
Conversely, Tables 10.4 and 10.5 show that %MAU is substantially modified when considering both bone tools and by-products. Figure 10.7 graphi-cally illustrates the variation between SEA and CSEA distributions in La Bellaca 2 site.
Figure 10.7 shows that metapodials are the best represented bones in both distributions. However, bones with medium %MAU in SEA decrease notably in CSA (axis, pelvis, scapula, humerus and tarsals). In CSEA, %MAU generates a pattern of representation smaller than 20 %MAU of the complete skeleton. This clearly indicates on-site bone-tool manufacture using metapodials as raw material. Moreover, tibiae are absent from the archaeofaunal assemblage with the exception of a single by-product fragment.10 Other important variation in CSEA analysis occurs in the relationship between MNE and MNI (Fig. 10.8).
At La Bellaca 2, MNE values imply a MNI of nine individuals versus only three identified by traditional zooarchaeological analysis. This new MNE value is almost certainly more accurate as it is unlikely that three pampas deer would offer enough raw materials to account for the numerous bone tools recovered on the site. This important MNI increment implies taking
A - %MAU (FSEA)
A - %MAU (CSEA)
LB2 - %MAU (FSEA)
LB2 - %MAU (CSEA)
0
20
40
60
80
100
0 1 2 3 4 5 6
BMD
%M
AU
Figure 10.6. Correlation between O. bezoarticus %MAU and BMD in Anahí and La Bellaca 2 for both distributions (SEA-CSEA).9
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
123
Don’t Smash Those Bones!
Table 10.4. Anahí: O. bezoarticus SEA vs CSEA.
SEA CSEA
Element NISP MNE MAU % MAU NISP MNE MAU % MAU
Teeth 22 22
Skull 1 1 1 25 1 1 1 19Hemimandible 3 1 0.5 12.5 3 1 0.5 9.5Cervical vertebra* 7 7 1.4 35 7 7 1.4 26.7Thoracic vertebra 1 1 0.1 2.5 1 1 0.1 1.9Lumbar vertebra 2 2 0.33 8.25 2 2 0.33 6.3Sacrum 1 1 1 25 1 1 1 19Rib 3 3 0.1 2.5 3 3 0.1 1.9Pelvis 3 1 1 25 3 1 1 19.0Scapula 8 8 4 100 8 8 4 76.2Humerus 1 1 0.5 12.5 1 1 0.5 9.5Radius 1 1 0.5 12.5 2 1 0.5 9.5Ulna 2 2 1 25 2 2 1 19Femur 1 1 0.5 12.5 1 1 0.5 9.5Patella 1 1 0.5 12.5 1 1 0.5 9.5Tibia 2 2 1 25 2 2 1 19
Metapodial 18 6 1.5 37.5 44 21 5.25 100Radiocarpal 2 2 1 25 2 2 1 19Tarsal (3rd & 4th) 6 6 3 75 6 6 3 57.1Calcaneus 5 5 2.5 62.5 5 5 2.5 47.6Astragalus 4 4 2 50 4 4 2 38.11st phalanx 5 4 0.5 12.5 5 4 0.5 9.52nd phalanx 2 2 0.25 6.25 2 2 0.25 4.83rd phalanx 6 6 0.75 18.75 6 6 0.75 14.3
Total 107 68 134 83
*Excluding atlas and axis. Teeth were excluded in MNE.
Table 10.5. La Bellaca 2: O. bezoarticus SEA vs CSEA.
SEA CSEA
Element NISP MNE MAU % MAU NISP MNE MAU %MAU
Teeth 7 7Mandible 1 1 0.5 18.2 1 1 0.5 5.9Axis 1 1 1 36.4 1 1 1 11.8
Pelvis 2 1 1 36.4 2 1 1 11.8
Scapula 2 2 1 36.4 2 2 1 11.8
Humerus 2 2 1 36.4 2 2 1 11.8
Femur 2 1 0.5 18.2 2 1 0.5 5.9
Tibia 0 0 0 0 1 1 0.5 5.9
Metapodial 40 11 2.75 100 88 34 8.5 100
Carpal (2nd & 3rd) 1 1 0.5 18.2 1 1 0.5 5.9
Tarsal (3rd & 4th) 3 3 1.5 54.5 3 3 1.5 17.6
Calcaneus 1 1 0.5 18.2 1 1 0.5 5.9
1st phalanx 3 3 0.4 14.5 3 3 0.4 4.7
2nd phalanx 3 3 0.4 14.5 3 3 0.4 4.7
3rd phalanx 6 6 0.7 25.5 6 6 0.7 8.2
Total 74 36 123 60
Teeth were excluded in MNE.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
124
Chapter 10
0
20
40
60
80
100
120
MandibleAxis
Pelvis
Scapula
Humerus
FemurTibia
Metapodial
Carpal (2
nd & 3rd)
Tarsa
l (3rd
& 4th)
Calcaneous
1st phalanx
2nd phalanx
3rd phalanx
SEA CSEA
Element
%M
AU
Figure 10.7. La Bellaca 2: O. bezoarticus SEA vs CSEA %MAU.
05
1015
20
25
3035
40
Mandible
AxisPelvis
Scapula
Humeru
s
Femur
Tibia
Meta
podial
Carpal (2
nd & 3rd
)
Tarsa
l (3rd
& 4th
)
Calcaneous
1st phalanx
2nd phalanx
3rd phalanx
SEA CSEA
MN
E
Element
Figure 10.8. La Bellaca 2: O. bezoarticus MNE values.
0
20
4060
80
100
Skull
Hemim
andible
Cervica
l verte
bra*
Thoracic
vertebra
Lumbar v
ertebra
Sacrum Rib
Pelvis
Scapula
Humeru
s
RadiusUlna
Femur
Patella
Tibia
Meta
podial
Radiocarp
al
Tarsa
l (3rd
& 4th
)
Calcaneous
Astragalus
1st phalanx
2nd phalanx
3rd phalanx
%M
AU SEA CSEA
Element
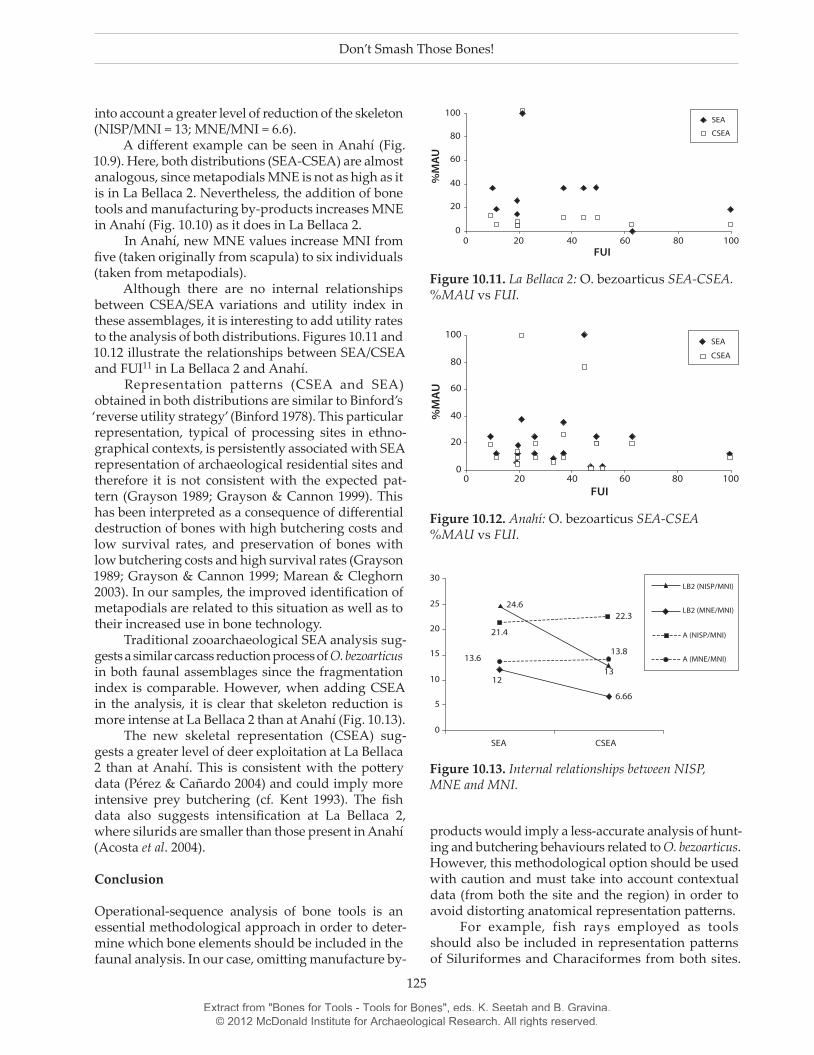
Figure 10.9. Anahí: O. bezoarticus SEA vs CSEA % MAU.
05
10152025
Skull
Hemim
andible
Cervica
l verte
bra*
Thoracic
vertebra
Lumbar v
ertebra
Sacrum Rib
Pelvis
Scapula
Humeru
s
RadiusUlna
Femur
Patella
Tibia
Meta
podial
Radiocarp
al
Tarsa
l
Calcaneous
Astragalus
1st phalanx
2nd phalanx
3rd phalanx
MN
E
Element
SEA CSEA
Figure 10.10. Anahí: O. bezoarticus MNE.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.
125
Don’t Smash Those Bones!
into account a greater level of reduction of the skeleton (NISP/MNI = 13; MNE/MNI = 6.6).
A different example can be seen in Anahí (Fig. 10.9). Here, both distributions (SEA-CSEA) are almost analogous, since metapodials MNE is not as high as it is in La Bellaca 2. Nevertheless, the addition of bone tools and manufacturing by-products increases MNE in Anahí (Fig. 10.10) as it does in La Bellaca 2.
In Anahí, new MNE values increase MNI from five (taken originally from scapula) to six individuals (taken from metapodials).
Although there are no internal relationships between CSEA/SEA variations and utility index in these assemblages, it is interesting to add utility rates to the analysis of both distributions. Figures 10.11 and 10.12 illustrate the relationships between SEA/CSEA and FUI11 in La Bellaca 2 and Anahí.
Representation patterns (CSEA and SEA) obtained in both distributions are similar to Binford’s ‘reverse utility strategy’ (Binford 1978). This particular representation, typical of processing sites in ethno-graphical contexts, is persistently associated with SEA representation of archaeological residential sites and therefore it is not consistent with the expected pat-tern (Grayson 1989; Grayson & Cannon 1999). This has been interpreted as a consequence of differential destruction of bones with high butchering costs and low survival rates, and preservation of bones with low butchering costs and high survival rates (Grayson 1989; Grayson & Cannon 1999; Marean & Cleghorn 2003). In our samples, the improved identification of metapodials are related to this situation as well as to their increased use in bone technology.
Traditional zooarchaeological SEA analysis sug-gests a similar carcass reduction process of O. bezoarticusin both faunal assemblages since the fragmentation index is comparable. However, when adding CSEA in the analysis, it is clear that skeleton reduction is more intense at La Bellaca 2 than at Anahí (Fig. 10.13).
The new skeletal representation (CSEA) sug-gests a greater level of deer exploitation at La Bellaca 2 than at Anahí. This is consistent with the pottery data (Pérez & Cañardo 2004) and could imply more intensive prey butchering (cf. Kent 1993). The fish data also suggests intensification at La Bellaca 2, where silurids are smaller than those present in Anahí (Acosta et al. 2004).
Conclusion
Operational-sequence analysis of bone tools is an essential methodological approach in order to deter-mine which bone elements should be included in the faunal analysis. In our case, omitting manufacture by-
products would imply a less-accurate analysis of hunt-ing and butchering behaviours related to O. bezoarticus. However, this methodological option should be used with caution and must take into account contextual data (from both the site and the region) in order to avoid distorting anatomical representation patterns.
For example, fish rays employed as tools should also be included in representation patterns of Siluriformes and Characiformes from both sites.
0
20
40
60
80
100
0 20 40 60 80 100FUI
%M
AU
SEA
CSEA
0
20
40
60
80
100
0 20 40 60 80 100FUI
%M
AU
SEA
CSEA
Figure 10.11. La Bellaca 2: O. bezoarticus SEA-CSEA. %MAU vs FUI.
Figure 10.12. Anahí: O. bezoarticus SEA-CSEA %MAU vs FUI.
24.6
6.66
22.3
1312
21.4
13.613.8
0
5
10
15
20
25
30
SEA CSEA
LB2 (NISP/MNI)
LB2 (MNE/MNI)
A (NISP/MNI)
A (MNE/MNI)
Figure 10.13. Internal relationships between NISP, MNE and MNI.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

126
Chapter 10
However, the analysis of toggle harpoons made on antler must follow a different protocol. In this latter case, evidence of on-site manufacture is not so clear. Moreover, it is impossible to establish whether antler from archaeological contexts is related to hunting events or was harvested from the local environment (Choyke 2003; Knecht 1997), as neither carnivore nor other taphonomic forces would have affected antler in such a way as to modify its physical and mechani-cal properties.
We must remark that the diaphysis is a portion of bone with high mineral density and survival rate (cf. Grayson & Frey 2004; Lyman 1994). If fragmented shafts from bone assemblages at both sites could be identified, it is probable that %MAU and BMD distributions would have a higher correlation, and differences between MNE and %MAU of metapodials and other long-bones would be smaller. The reductive process employed on O. bezoarticus indicates specific cultural attitudes. Undoubtedly, this technique has mainly impacted on bones with low mineral density (proximal femur, proximal humerus, scapula, verte-brae and ribs), which also represents the elements with the highest utility rates. Accordingly, in both assem-blages the causal agent of that pattern equivalent to Binford’s ‘reverse utility strategy’ (Binford 1978) has been a cultural one, interacting to a lesser extent with post-depositional taphonomic factors. Therefore, the CSEA analysis permits a more accurate determination of the intensity of this reductive process, and serves as an alternative tool for comparison of the intensifica-tion processes of environmental exploitation.
Acknowledgements
We thank Krish Seetah for his invitation to write this paper. We also thank Alejandro Acosta and Leonardo Mucciolo for stimulating discussion about deer-anatomical repre-sentation and bone tool production. Krish Seetah, Romina Silvestre, Dánae Fiore and Javier Musali helped us with the English version. Financial support and facilities were provided by CONICET and INAPL.
Notes
1. Approximate weight: between 25 and 40 kg (Loponte 2008).
2. The sample analysed here from Anahí (bone tools and faunal elements) was recovered in quadrants E9, G10, F10 and F11 by Lafón in 1969/71. Sample of LB2 was recovered in our excavations carried out in 1998–2000 in quadrants C1, C2, C3, C4, C5, C6 and C7.
3. O. bezoarticus MNE values were taken from this paper, while B. dichotomus MNE values were taken from Acosta 2005, table VII.6. A different fragmentation index can be seen in Acosta (2005). See also Mucciolo (2004).
4. This fragmentation index is derived considering all NISP mammals >25 kg, in order to obtain a more accu-rate value than the traditional MNE/NISP (e.g. Lyman 1994).
5. Identifiable bone specimens (distal epiphysis and shafts) were assigned to element, side and portion. Frequencies of element portions were used to calculate the minimum number of skeletal elements (MNE), the minimum number of animal units (MAU) and the minimum number of individuals (MNI) were taken following Klein & Cruz-Uribe (1984) and Lyman (1994).
6. Teeth were not included in the MNE analysis in this paper.
7. These values consider formal tools or fragments (exclud-ing manufacture by-products). In Anahí the sample include only those quadrants analysed in this paper.
8. ‘Smoothers’ are flat tools polished on their facets (Buc 2010; Buc & Loponte 2007), needles and pin-likes are fine-tipped objects with and without eye, respectively (Campana 1989, 51).
9. Since correlations between BMD values in camelids and cervids are consistently high, BMD values were taken from Stahl (1999) for South American camelids.
10. Final tools made of tibia were not identified in the site. 11. FUI values were taken from Metcalfe & Jones (1988) for
complete bones. Because the evident disparity of utility index between metatarsal (FUI = 37) and metacarpal (FUI = 5.2), and the difficulty of distinguishing meta-tarsal to metacarpals condyles, values were averaged resulting a metapodial FUI value of 21.1.
References
Acosta, A., 1997. Estados de conservación y problemas de contaminación de las estructuras arqueofaunísticas en el extremo nororiental de la Pcia. de Buenos Aires, in Arqueología Pampeana en la Década de los ´90, eds. M. Berón & G. Politis. San Rafael: Museo de Historia Natural de San Rafael, INCUAPA, 187–99.
Acosta, A., 2000. Huellas de corte relacionadas con la manufactura de artefactos óseos en el nordeste de la provincia de Buenos Aires. Relaciones de la Sociedad Argentina de Antropología 25, 159–78.
Acosta, A., 2005. Zooarqueología de cazadores-recolectores del extremo nororiental de la provincia de Buenos Aires (humedal del río Paraná inferior, Región Pam-peana, Argentina). Unpublished PhD thesis, Univer-sidad Nacional de La Plata.
Acosta, A., D. Loponte, J. Musali & J. Olub, 2004. Estima-ciones de tamaño de Pterodoras granulosus (armado común) recuperados en sitios arqueológicos del humedal del Paraná inferior. Unpublished paper presented at XVth Congreso Nacional de Arqueología Argentina, Río Cuarto.
Acosta, A., N. Buc & L. Mucciolo, 2010. Linking evidences: from carcass processing to bone technology. The case of the Lower Paraná wetlands (Late Holocene, Argentina), in Ancient and Modern Bone Artefacts from America to Russia: Cultural, Technological and Functional Signature, eds. A. Legrand-Pineau, I. Sidéra, N. Buc,
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

127
Don’t Smash Those Bones!
E. David & V. Scheinsohn. (British Archaeological Reports, International Series 2136.) Oxford: BAR, 303–14.
Averbouh, A., 2000. Technologie de la matière osseuse tra-vaillée et implications paleoetnologiques: l’exemple des chaînes d’exploitation du bois de cervidé chez les Magdaléniens des Pyrénées. Unpublished PhD thesis, Université de Paris I.
Averbouh, A. & P. Catellain, 2001. De l’analyse fonctionelle des propulseurs par l’économie de leur production: l’industrie osseuse pré- et protohistorique en Europe, in L´Industrie osseuse pré- et protohistorique en Europe: Approches technologiques et fonctionnelles, eds. M. Patou-Mathis, P. Catellain & P. Ramseyer. Amay: Université de Liège, 63–73.
Backwell, L. & F. d’Errico, 2001. Evidence of termite forag-ing by Swartkrans early hominids. Proceedings of the National Academy of Sciences of the USA 98(4),1358–63.
Bar-Oz, G. & N.D. Munro, 2004. Beyond cautionary tales: a multivariate taphonomic approach for resolving equifinality in zooarchaeological studies. Journal of Taphonomy 2(4), 201–20.
Behrensmeyer, A.K., 1978. Taphonomic and ecological infor-mation from bone weathering. Paleobiology 4, 150–62.
Binford, L., 1978. Nunamiut Ethnoarchaeology. New York (NY): Academic Press.
Blumenschine, R.J., 1986. Early Hominid Scavenging Opportu-nities: Implications of Carcass Availability in the Serengeti and Ngorongoro Ecosystems. (British Archaeological Reports, International Series 243.) Oxford: BAR.
Blumenschine, R.J., 1989. An experimental model of the tim-ing of hominid and carnivore influence on archaeo-logical bone assemblages. Journal of Archaeological Science 15, 483–502.
Broughton, J.M., 1994a. Declines in mammalian foraging efficiency during the late Holocene, San Francisco Bay, California. Journal of Anthropological Archaeology 13, 371–401.
Broughton, J.M., 1994b. Late Holocene resource intensi-fication in the Sacramento Valley, California: the vertebrate evidence. Journal of Archaeological Science 21, 501–14.
Broughton, J.M. & D.K. Grayson, 1993. Diet breadth, adap-tive change, and the White Mountains faunas. Journal of Archaeological Science 20, 331–6.
Buc, N., 2007. Ser o no ser: arpones y ‘arpones B’ en el humedal del Paraná inferior, in Arqueología en las Pampas, tomo I, eds. C. Bayón, A. Pupio, M.I. González, N. Flegenheimer & M. Frère. Buenos Aires: Sociedad Argentina de Antropología, 325–42.
Buc, N., 2010. Tecnología ósea de cazadores-recolectores del humedal del Paraná inferior. (Arqueología de la Cuenca del Plata.) Buenos Aires: Instituto Nacional de Antro-pología y Pensamiento Latinoamericano.
Buc, N. & D. Loponte, 2007. Bone tool types and microwear patterns: some examples from the Pampa region, South America, in Bones as Tools: Current Methods and Interpretations in Worked Bone Studies, eds. C. Gates St-Pierre & R.B. Walker. (British Archaeological Reports, International Series 1622.) Oxford: BAR, 143–57.
Caggiano, M.A., 1984. Prehistoria del NE. Argentino. Sus vinculaciones con la República Oriental del Uruguay y sur de Brasil. Pesquisas (Serie Antropología) 38, 1–109.
Campana, D., 1989. Natufian and Protoneolithic Bone Tools: the Manufacture and Use of Bone Implements in the Zagros and the Levant. (British Archaeological Reports, Inter-national Series 494.) Oxford: BAR.
Camps-Fabrer, H., 1967. Typologie de l’industrie osseuse en Afrique du Nord et au Sahara, in Congrés Panafrican de Préhistorie (Dakar), ed. H.-J. Hugot. Chambéry: Les imprimeries réunies de Chambéry, 279–83.
Camps-Fabrer, H. & A. d’Anna, 1977. Fabrication expéri-mentale d’outils à partir de métapodes de mouton et de tibias de lapin, in Deuxième Colloque International sur l’industrie de l’os dans la Préhistoire, Méthodologie appliquée a l’industrie de l’os préhistorique. Paris: CNRS, 311–23.
Cannon, M.D., 2003. A model of central place forager prey choice and an application to faunal remains from the Mimbres Valley, New Mexico. Journal of Anthropologi-cal Archaeology 22, 1–25.
Choyke, A., 2003. Backward reflections on ancient environ-ments: What can we learn from bone tools?, in People and Nature in Historical Perspective, eds. J. Laslovszky & P. Szabo. Budapest: Central European University Press, 139–56.
Church, R.R. & R.L. Lyman, 2003. Small fragments make small differences in efficiency when rendering grease from fractured artiodactyl bones by boiling. Journal of Archaeological Science 30, 1077–84.
Cleghorn, N. & C.W. Marean, 2004. Distinguishing selective transport and in situ attrition: a critical review of ana-lytical approaches. Journal of Taphonomy 2(4), 257–63.
Enloe, J.G., 2004. Equifinality, assemblage integrity and behavioral inferences at Verberie. Journal of Taphonomy 2(3), 147–64.
d’Errico, F. & L. Backwell, 2003. Possible evidence of bone tools sharping by Swarktrans early hominids. Journal of Archaeological Science 30, 1559–76.
García Esponda, C., A. Acosta, D. Loponte & L. De San-tis, 2001. Registro de Chrysocyon brachiurus (CAR-NIVORA: CANIDAE) en contextos arqueológicos en el noreste de la provincia de Buenos Aires. Mastozo-ología Neotropical 8(2), 159–63.
Grayson, D.K, 1989. Bone transport, bone destruction, and reverse utility curves. Journal of Archaeological Science 16, 643–52.
Grayson, D.K, 2001. The archaeological record of human impacts on animal populations. Journal of World Pre-history 15, 1–68.
Grayson, D.K. & M.D. Cannon, 1999. Human paleoecology and foraging theory in the Great Basin, in Models for the Millennium: Great Basin Anthropology Today, ed. C. Beck. Salt Lake City (UT): University of Utah Press, 141–51.
Grayson, D.K. & F. Delpech, 1998. Changing diet breadth in the early Upper Palaeolithic of southwestern France. Journal of Archaeological Science 25, 1119–29.
Grayson, D. & C. Frey, 2004. Measuring skeletal part rep-resentation in archaeological faunas. Journal of Tapho-nomy 2(4), 233–56.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

128
Chapter 10
Grayson, D.K., F. Delpech, J.P.H. Rigaud & J. Simek, 2001. Explaining the development of dietary dominance by a single ungulate taxon at Grotte XVI, Dordogne, France. Journal of Archaeological Science 28, 115–25.
Griffitts, J., 2006. Bone Tools and Technological Choice: Change and Stability on the Northern Plains. Unpub-lished PhD dissertation, University of Arizona.
Guthrie, D., 1983. Osseous projectile point: biological con-siderations affecting raw material selection and design among Paleolithic and paleoindian peoples, animals and archaeology, part 1, in Hunters and their Prey, eds. J. Clutton-Brock & C. Grigson. (British Archaeologi-cal Reports, International Series 163.) Oxford: BAR, 274–94.
Hodgetts, L., 2000. Hunting reindeer to support a marine rconomy. An example from Arctic Norway. Archaeo-fauna 9, 17–28.
Hodgetts, L. & F. Rahemtulla, 2001. Land and sea: use of terrestrial mammal bones in coastal hunter–gatherer communities. Antiquity 75, 56–62.
Julien, M., 1986. La fonction des outils d´os peu élabores de Tel-armachay (Pérou). Outillage peu élabore en os et en bois de cervidés II. Paris/Abidjan: Editions du C.E.D.A.
Kaplan, H. & K. Hill, 1992. The evolutionary ecology of food acquisition, in Evolutionary Ecology and Human Behavior, eds. E.A. Smith & B. Winterhalder. New York (NY): Aldine de Gruyter, 167–201.
Kent, S., 1993. Variability in faunal assemblages: the influ-ence of hunting skill, sharing, dogs and mode of cooking on faunal remains at a sedentary Kalahari community. Journal of Anthropological Archaeology 12, 323–83.
Klein, R. & C. Cruz-Uribe, 1984. The Analysis of Animal Bones from Archaeological Sites. Chicago (IL): University of Chicago Press.
Knecht, H., 1997. Projectile points of bone, antler and stone: experimental exploration of manufacture and use, in Projectile Technology, ed. H. Knecht. New York (NY): Plenum Press, 191–213.
Kramer, D., 2001. Foraging behavior, in Evolutionary Eco logy: Concept and Case Studies, eds. L.W. Fox, D.A. Roff & D.J. Fairnbairn. Oxford: Oxford University Press, 232–42.
Krebs, J.R., M.I. Avery & A.I. Houston, 1987. Delivering food to a central place: three studies of Bee-eaters (Merops apiaster), in Foraging Behavior, eds. A.C. Kamil, J.R. Krebs & H.R. Pulliam. New York (NY): Plenum Press, 173–91.
Legrand, A., 2005. Nouvelle approche méthodologique des assemblages osseux du Néolithique de Chypre. Entre technique, fonction et culture. Unpublished PhD the-sis, Université de Paris I.
Loponte, D., 2008. Arqueología del humedal del Paraná inferior (Bajios Ribereños Meridionales). (Serie Arqueología de la Cuenca del Plata.) Buenos Aires: Instituto Nacional de Antropología y Pensamiento Latinoamericano, Secretaría de Cultura de la Nación.
Loponte, D. & A. Acosta, 2003. Arqueología de cazadores-cecolectores del sector centro-oriental de la región Pampeana. RUNA, Archivo para las ciencias del hombre 24, 173–212.
Loponte, D. & A. Acosta, 2004. Late Holocene hunter-gath-erers from the Pampean wetlands, Argentina, in Zoo-archaeology of South America, ed. G. Mengoni Goñalons. (British Archaeological Reports, International Series 1298.) Oxford: BAR, 39–57.
Loponte, D. & R. Sacur Silvestre, 2003. Lejos de las can-teras: la explotación de recursos líticos en el sector centro-oriental de la región Pampeana. Unpublished manuscript.
Lothrop, S.K., 1932. Indians of the Paraná Delta river. Annals of the New York Academy of Sciences 33, 77–232.
Lyman, L., 1994. Vertebrate Taphonomy. Cambridge: Cam-bridge University Press.
Lyman, R.L., 2003. The influence of time averaging and space averaging on the application of foraging theory in zooarchaeology. Journal of Archaeological Science 30, 595–610.
Lyman, R.L., 2004. The concept of equifinality in taphonomy. Journal of Taphonomy 2(4), 221–32.
Maigrot, Y., 2003. Étude technologique et fonctionnelle de l’outillage en matières dures animales. La station 4 de Chalain (Néolithique final, Jura, France). Unpublished PhD thesis, Université de Paris I.
Marean, C.W. & N. Cleghorn, 2003. Mammal skeletal ele-ment transport: applying foraging theory in a complex taphonomic system. Journal of Taphonomy 1, 15–42.
Metcalfe, D. & K.R. Barlow, 1992. A model for exploring the optimal trade-off between field processing and transport. American Anthropologist 94, 340–56.
Metcalfe, D. & K. Jones, 1988 A reconsideration of animal body-part utility index. American Antiquity 53(3), 486–504.
Le Moine, G., 1991. Experimental Analysis of the Manu-facture and Use of Bone and Antler Tools among the Mackenzie Inuit. Unpublished PhD thesis, University of Calgary, Alberta.
Mucciolo, L., 2004. Explotación y procesamiento de cérvi-dos en el sitio La Bellaca 2, provincia de Buenos Aires. Unpublished paper presented at XV Congreso Nacional de Arqueología Argentina.
Munro, N.D. & G. Bar-Oz, 2005. Gazelle bone fat processing in the Levantine Epipalaeolithic. Journal of Archaeologi-cal Science 32, 223–39.
Musali, J., 2005. Ictioarqueología del Delta del río Paraná inferior. Unpublished Grade thesis, Universidad de Buenos Aires.
Musali, J., A. Acosta & D. Loponte, 2003. Methodological issues: an attempt to assess bone representation of Pampa’s wetlands ichthyoarchaeological record, Argentina, in Presence of the Archaeoichthyology in Mexico. Proceedings of the Meeting of the Fish Remains Working Group of the International Council for Archaeo-zoology, eds. A.F. Guzman, O.J. Polaco & F.J. Aguilar. México: ICAZ, 117–23.
Nagaoka, L., 2001. Using diversity indices to measure changes in prey choice at the Shag River Mouth site, southern New Zealand. International Journal of Osteo-archaeology 11, 101–11.
Newcomer, M., 1974. Study and replication of bone tools from Ksar Akil (Lebanon). World Archaeology 6, 138–53.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.

129
Don’t Smash Those Bones!
O’Connell, J.F., 1995. Ethnoarchaeology needs a general theory of behavior. Journal of Archaeological Research 3, 205–55.
Olsen, S., 1979. A study of bone artifacts from Grasshopper Pueblo, AZ P: 14: 1. The Kiva 44(4), 341–71.
Orians, G.H. & N.E. Pearson, 1979. On the theory of central place foraging, in Analysis of Ecological Systems, eds. D.J. Horn, R.D. Mitchell & G.R. Stairs. Athens (OH): Ohio University Press, 155–77.
Peltier, A., 1986. Etude expérimentale des surfaces osseuses façonnées et utilisées. Bulletin de la Societé Préhistorique Francaise 83(1), 5–7.
Peltier, A. & H. Plisson, 1989. Microtraceologie fonctionnelle sur l’os: quelques résultats expérimentaux. Artefacts 3, 69–79.
Pérez, M. & L. Cañardo, 2004. Producción y uso de cerámica en el norte de la provincia de Buenos Aires, in Aproxi-maciones Contemporáneas a la Arqueología Pampeana, eds. G. Martínez, M.A. Gutiérrez, R. Curtoni, M. Berón & P. Madrid. Olavarría: Facultad de Ciencias Sociales, Universidad Nacional del Centro de la provincia de Buenos Aires, 335–48.
Pétillon, J.M., 2001. Typologie et utilisation: l’exemple des pointes à base fourchue Magdaléniennes, in L´Industrie osseuse pré- et protohistorique en Europe: Approches Technologiques et Fonctionnelles, eds. M. Patou-Mathis, P. Catellain & P. Ramseyer. Amay: Université de Liège, 53–62.
Poplin, F., 1974. Principes de la détermination des matières dures animales, in Premier colloque sur l’industrie de l’os dans la préhistoire, ed. H. Camps-Fabrer. Abbaye de Sénanque: Université de Provence, 15–20.
Sadek Kooros, H., 1972. Primitive bone fracturing: a method of research. American Antiquity 37, 369–82.
Scheinsohn, V., 1997. Explotación de materias primas óseas en la Isla grande de Tierra del Fuego. Unpublished PhD thesis, Universidad de Buenos Aires.
Scheinsohn, V., A. Di Baja; M. Lanza & L. Tramaglino, 1992. El aprovechamiento de la avifauna como fuente de materia prima ósea en la Isla Grande de Tierra del Fuego: Lancha Packewaia, Shamakush y Tunel I. Arqueología 2, 135–47.
Sidéra, I., 1989. Un complément des données sur les sociétés rubanées, l’industrie osseuse à Cuirylès-Chaudardes. (Brit-ish Archaeological Reports, International Series 520.) Oxford: BAR.
Sidéra, I., 1991. Processus économiques, choix tech-nologiques et culturels dans l’exploitation des faunes protohistoriques des VIe au IVe millénaires en France septentrionale. État de la documentation. Revue Archéologique de Picardie 1–2, 3–19.
Sidéra, I., 2000. Animaux domestiques, bêtes sauvages et objets en matières animales du Rubané au Michelsberg. De l’économie aux symboles, des tech-niques à la culture. Gallia Préhistoire 42, 107–94.
Smith, E.A., 1983. Anthropological applications of optimal foraging theory: a critical review. Current Anthropo logy 24(5), 625–52.
Stahl, P.W., 1999. Structural density of domesticated South American camelid skeletal elements and the archaeo-logical investigation of prehistoric Andean Ch´arki. Journal of Archaeological Science 26, 1347–68.
Stephens, D.W., 1990. Foraging theory: up, down, and side-ways. Studies in Avian Biology 13, 444–54.
Stephens, D.W. & J.R. Krebs, 1986. Foraging Theory. Princeton (NJ): Princeton University Press.
Stiner, M.C., N.D. Munro & T.A. Surovell, 2000. The tortoise and the hare: small-game use, the broad-spectrum revolution, and Paleolithic demography. Current Anthropology 41, 39–73.
Stordeur, D., 1980. Harpons Paléo-Esquimaux de la région d´Igloulik. Recherche sur les grandes civilisations. Cahier 2, Préhistoire: enquêtes et méthodes. Paris: Editions ADPF.
Teta, P., D. Loponte & A. Acosta, 2004. Sigmodontinos (Mam-malia, Rodentia) del Holoceno tardío del nordeste de la provincia de Buenos Aires (Argentina). Masto-zoología Neotropical, Journal of Neotropical Mammalogy 11(1), 69–80.
Torres, L.M., 1911. Los primitivos habitantes del Delta del Paraná. La Plata: Universidad Nacional de La Plata, Biblioteca Centenaria.
White, T.E., 1954. Observations on the butchering technique of some aboriginals people (3–6). American Antiquity 20, 254–64.
Extract from "Bones for Tools - Tools for Bones", eds. K. Seetah and B. Gravina. © 2012 McDonald Institute for Archaeological Research. All rights reserved.




















