
Dirty teeth and ancient trade: Evidence of cotton fibres in human dental calculus from Late...
10
Dirty Teeth and Ancient Trade: Evidence of Cotton Fibres in Human Dental Calculus from Late Woodland, Ohio S. H. BLATT, a * B. G. REDMOND, b V. CASSMAN c AND P. W. SCIULLI a a Department of Anthropology, The Ohio State University, OH, USA b Department of Archaeology, The Cleveland Museum of Natural History, OH, USA c Department of Art Conservation, University of Delaware, DE, USA ABSTRACT Analysis of ancient human dental calculus for the presence of inclusions related to diet and dental health has been overlooked in anthropological literature. Small particles of archaeobotanical debris, which would otherwise not be preserved in the archaeological record, can become incorporated into unmineralised plaque on teeth during mastication and oral manipulation. When plaque mineralises into calculus, debris is preserved in situ. Samples of dental calculus (n ¼ 18) were collected from the Danbury site (33OT16) in Ottawa County, Ohio and viewed under a scanning electron microscope for inclusions. Analysis yielded a variety of noticeable inclusions, including mineralised bacteria, calcium-phosphate crystalline structures and numerous phytoliths. Here we report the first evidence of fibres consistent with cotton (Gossypium spp.) embedded in the dental calculus from the Late Woodland component (900–1100 AD) of the Danbury site. Prehistoric cotton has not been previously documented in Ohio. The distinct morphology of the Danbury cotton and its presence in the Late Woodland component at Danbury suggests long-distance interaction at a time in Ohio when movement of exotic goods appeared to have diminished. These microscopic remains provide insight into paleoethnobotanical history of ancient Ohioans and attest to how analysis of dental calculus could be used to supplement other paleodietary and archaeological analyses. Copyright ß 2010 John Wiley & Sons, Ltd. Key words: dental calculus; cotton; SEM; Late Woodland Introduction Dental calculus is an overlooked source of data in anthropological studies. Calculus provides a unique calcified reservoir of mineralised bacteria (Vander- meersch et al., 1994; Arensburg, 1996; Hershkovitz et al., 1997) and dietary debris (Armitage, 1975; Dobney & Brothwell, 1986; Middleton, 1990; Mid- dleton, 1992a; Middleton, 1992b; Cortello & Pochet- tino, 1994; Middleton & Rovner, 1994; Fox et al., 1996; Gobetz & Bozarth, 2001; Boyadjian et al., 2007; Blondiaux & Charlier, 2008) seldom preserved else- where in the archaeological record. Small particles of archaeobotanical debris can adhere to and become incorporated into the unmineralised plaque on teeth during mastication and oral manipulation. When plaque mineralises into calculus, debris and bacteria are preserved in situ. The physical inclusion of bacteria and archaeobotanical debris in the matrix of dental calculus is direct evidence for the contemporaneity of both structures, because the mineralisation of dental plaque occurs only in the presence of saliva during the life of an individual (MacPhee & Cowley, 1975). Such microscopic remains provide insight into the dental health and paleoethnobotanical history of ancient populations. Here we report the first findings of preserved cellulose fibres consistent with the characteristics of cotton (Gossypium spp.) embedded within the matrix of the dental calculus of four Late Woodland individuals from the Danbury site (33OT16), Ohio. The genus Gossypium is composed of a diverse group of species (more than 50) with a global distribution and marked morphological and cytogenetic diversity. Gossypium constitutes a monophylytic taxon belonging to the family Malvaceae. Wendel et al. (1992) and Wendel (1995) provide ample evidence to establish that New World tetraploid cottons are allopolyploids International Journal of Osteoarchaeology Int. J. Osteoarchaeol. (2010) Published online in Wiley InterScience (www.interscience.wiley.com) DOI: 10.1002/oa.1173 * Correspondence to: Department of Anthropology, The Ohio State University, OH, USA. e-mail: [email protected] Copyright # 2010 John Wiley & Sons, Ltd. Received 31 July 2009 Revised 25 February 2010 Accepted 17 March 2010
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Dirty teeth and ancient trade: Evidence of cotton fibres in human dental calculus from Late...

International Journal of Osteoarchaeology
Int. J. Osteoarchaeol. (2010)Published online in Wiley InterScience(www.interscience.wiley.com) DOI: 10.1002/oa.1173
* Correspondence to: DepaUniversity, OH, USA.e-mail: [email protected]
Copyright # 2010 Joh
Dirty Teeth and Ancient Trade: Evidenceof Cotton Fibres in Human DentalCalculus from Late Woodland, Ohio
S. H. BLATT,a* B. G. REDMOND,b V. CASSMANc AND P. W. SCIULLIa
a Department of Anthropology, The Ohio State University, OH, USAb Department of Archaeology, The Cleveland Museum of Natural History, OH, USAc Department of Art Conservation, University of Delaware, DE, USA
ABSTRACT Analysis of ancient human dental calculus for the presence of inclusions related to diet and dental health hasbeen overlooked in anthropological literature. Small particles of archaeobotanical debris, which wouldotherwise not be preserved in the archaeological record, can become incorporated into unmineralisedplaque on teeth during mastication and oral manipulation. When plaque mineralises into calculus, debrisis preserved in situ. Samples of dental calculus (n¼ 18) were collected from the Danbury site (33OT16) inOttawa County, Ohio and viewed under a scanning electron microscope for inclusions. Analysis yielded avariety of noticeable inclusions, including mineralised bacteria, calcium-phosphate crystalline structures andnumerous phytoliths. Here we report the first evidence of fibres consistent with cotton (Gossypium spp.)embedded in the dental calculus from the Late Woodland component (900–1100 AD) of the Danbury site.Prehistoric cotton has not been previously documented in Ohio. The distinct morphology of the Danbury cottonand its presence in the Late Woodland component at Danbury suggests long-distance interaction at a time inOhio when movement of exotic goods appeared to have diminished. These microscopic remains provideinsight into paleoethnobotanical history of ancient Ohioans and attest to how analysis of dental calculus couldbe used to supplement other paleodietary and archaeological analyses. Copyright� 2010 JohnWiley & Sons,Ltd.
Key words: dental calculus; cotton; SEM; Late Woodland
Introduction
Dental calculus is an overlooked source of data inanthropological studies. Calculus provides a uniquecalcified reservoir of mineralised bacteria (Vander-meersch et al., 1994; Arensburg, 1996; Hershkovitzet al., 1997) and dietary debris (Armitage, 1975;Dobney & Brothwell, 1986; Middleton, 1990; Mid-dleton, 1992a; Middleton, 1992b; Cortello & Pochet-tino, 1994; Middleton & Rovner, 1994; Fox et al., 1996;Gobetz & Bozarth, 2001; Boyadjian et al., 2007;Blondiaux & Charlier, 2008) seldom preserved else-where in the archaeological record. Small particles ofarchaeobotanical debris can adhere to and becomeincorporated into the unmineralised plaque on teethduring mastication and oral manipulation. Whenplaque mineralises into calculus, debris and bacteria
rtment of Anthropology, The Ohio State
n Wiley & Sons, Ltd.
are preserved in situ. The physical inclusion of bacteriaand archaeobotanical debris in the matrix of dentalcalculus is direct evidence for the contemporaneity ofboth structures, because the mineralisation of dentalplaque occurs only in the presence of saliva duringthe life of an individual (MacPhee & Cowley, 1975).Such microscopic remains provide insight into thedental health and paleoethnobotanical history ofancient populations. Here we report the first findingsof preserved cellulose fibres consistent with thecharacteristics of cotton (Gossypium spp.) embeddedwithin the matrix of the dental calculus of fourLate Woodland individuals from the Danbury site(33OT16), Ohio.The genus Gossypium is composed of a diverse group
of species (more than 50) with a global distribution andmarked morphological and cytogenetic diversity.Gossypium constitutes a monophylytic taxon belongingto the family Malvaceae. Wendel et al. (1992) andWendel (1995) provide ample evidence to establishthat New World tetraploid cottons are allopolyploids
Received 31 July 2009Revised 25 February 2010Accepted 17 March 2010

S. H. Blatt et al.
containing one genome similar to those found in theOld World. Of the four domesticated cotton speciesfound worldwide, two have been domesticated in theNew World. Gossypium hirsutum is widely distributed inCentral and South America, the Caribbean and evenreaches distant islands in the Pacific (Solomon Islandsand Marquesas). Gossypium barbadense has a moresoutherly indigenous range, centred in northwesternSouth America but overlaps with the range ofG. hirsutum in the Caribbean. It is widely acceptedthat, of the two varieties of pre-Columbian domesti-cated cotton in the Americas, G. barbadense (commonlyknown as Pima cotton or Egyptian cotton) was firstcultivated in Peru as early as 2500–1750 BC and thenintroduced into the Caribbean, whereas G. hirsutum (orUpland cotton) was first cultivated in Mesoamericabetween 300 BC and 500 AD and is first seen in the Gilaand Salt River regions of Arizona around 500 AD (Kent,1983; Lee, 1984; Teague, 1998). Successful cultivationof cotton requires a long frost-free period (160 days)and moderate rainfall, usually from 600 to 1200mm. Ingeneral, these conditions are met within the seasonallydry tropics and subtropics in the Northern andSouthern hemispheres (Wendel, 1995).While there is some archaeological evidence of
contact between the Caribbean islands and southernFlorida during the Late Prehistoric period (1000–1650AD), there is no evidence of pre-Columbian cotton ineastern North America (Bullen, 1974; Seidemann,2006). Knowledge of the role and extent of cottonexchange between South-western Pueblo and Plainsgroups and others has been limited due to a lack ofcomprehensive archaeobotanical evidence. In fact,there is no evidence of prehistoric cotton cultivation orexchange east of the Spiro Mound site in LaFloreCounty, Oklahoma (King & Gardener, 1981). There isa similar lack of evidence for the movement of cottonnorthward from known Preclassic through Postclassicsource areas in the Gulf lowlands of northeasternMexico (northern Veracruz and Tamaulpas) (Starket al., 1998; Evans, 2004).Though societies in Ohio were producing yarns
made from twined native plant fibres at least as early asthe Middle Woodland (100 BC–500 AD), it wasunexpected to find multiple fibres with the physicalattributes of mature and immature Gossypium spp.embedded in the dental calculus of four LateWoodland(900–1100 AD) individuals from the Danbury site. Thecultural implications of this discovery are far-reachingand could suggest long-range interaction betweeneither South-western Pueblo groups, or northernMesoamerican societies, and groups living in northernOhio during the LateWoodland period, a time in Ohio
Copyright # 2010 John Wiley & Sons, Ltd.
when trade of exotic goods is generally regarded ashaving been attenuated.
Materials and methods
Dental calculus samples were obtained from a total of18 teeth from 8 Danbury site individuals (05-02A, 05-02B, 05-02C, 05-06, 05-07, 06-01B, 06-02A, 06-03A)(see Table 1). The Danbury site is a multicomponent,pre-Columbian habitation site located on the southshore of Lake Erie in Ottawa County, Ohio (Red-mond 2005, 2006a, 2006b, 2007) (Figure 1). Inves-tigation of the site was first supervised by the OhioValley Archaeological Consultants (OVAC) in 2003during the construction of a housing developmentand then from 2004 to 2007 by the Department ofArchaeology of The Cleveland Museum of NaturalHistory (Redmond, 2005, 2006a, 2007). Forty-twoburial features (BF), containing 152 individuals, havebeen excavated from the Danbury site since 2003.Accelerator mass spectrometry (AMS) dating ofcarbon and bone collagen samples indicate habi-tation at the site during the Late Archaic (2900–2500BC), Early Woodland (800–1000 AD), Late Woodland(900–1100 AD), and Late Prehistoric (1450–1550 AD)periods. Burial treatments at the site includedprimary flexed, semi-flexed, extended and bundled;however, many of the burials appear to be secondaryinterments combining multiple individuals (Red-mond, 2006b).A wide range of non-perishable burial goods were
found at the Danbury site, the majority being marineshell beads, primarily disk beads and marginella shellbeads (Redmond, 2006a, 2006b). Prominent amongthe marine shell artefacts and associated with the threeindividuals from BF 05-02, are portions of two drilledpendants made from a lightning whelk shell (Busyconsinistrum) (Figure 2), a species whose ranges extend fromthe Gulf of Mexico to the southern Atlantic (Trubitt,2003; Redmond, 2006b). One half of a whelk pendantwas found in the mouth of individual A from BF 05-02,and the matching half of this specimen was foundadjacent to the upper lumbar vertebrae of individual B.The two halves of this first pendant tightly fit together,suggesting they were broken only shortly beforeinterment. There are also three drill holes in thespecimen which display slight wear (Redmond,2006b). The second pendant (Figure 2), found betweenthe left humerus and sternum of individual C, was madefrom the outer whorl of large whelk had ground edges,a single drill hole, and two lateral notches (Redmond,2006a).
Int. J. Osteoarchaeol. (2010)

Figure 1. Location of the Danbury site (33OT16) in northern Ohio.
Table 1. Specimen data including BF (burial feature) number, sex (P¼pelvis; Pb¼pubis; O¼ other) and age (P¼pelvis; Pb¼pubis;A¼ auricular; O¼ other; D¼dental stage; F¼ epiphyseal fusion), stature in cm, body mass in kg, calculus score (0¼ absent; 1¼mild,covering less than one third of the crown; 2¼moderate, covering from one third to two thirds of the crown; 3¼ severe, covering morethan two thirds of the crown), location of the calculus on each tooth (L¼ lingual surface; B¼buccal surface) and the presence (indicatedby ‘x’) of inclusions such as bacteria, calcium phosphate (CaP) crystals, phytoliths and/or starch grains, unidentified fibres andGossypium sp. fibres within the calculus specimens
BF No. Sex Age Stature(cm)
Mass(kg)
Tooth Score Loc. Inclusions
Bacteria CaPCrystals
Phytoliths/Starch Unid.Fibre
GossypiumFibre
05-02A F (P) 42–52 (A) 156.8 63.4 LRP4 1 L x x05-02B M (P) 29–33 (A) 172.9 63.1 URM1 1 B
URM2 2 B x xURP3 1 B x xURI2 1 BURC 1 B x
05-02C M (P, Pb) 35–40(Pb, A)
170.7 64.1 ULI2 1 L x x
URM2 1 B05-06 F (P, Pb) 25–30
(Pb, A)159.3 59.0 LLM2 1 B x
ULM1 1 BLRC 1 L xLRI2 1 L xLLC 1 L x
05-07 M (O) Adult (O) — — LRP3 1 L1 L x
LM1 2 L x x x06-01B F (P) 12–14
(D, F)— — ULP4 1 B
06-02A M (P) 13–14(D, F)
— — URI1 1 B
06-03A M (P, Pb) 50þ (Pb, A) 163.2 64.1 LI2 1 L x x
Copyright # 2010 John Wiley & Sons, Ltd. Int. J. Osteoarchaeol. (2010)
Dirty Teeth and Ancient Trade
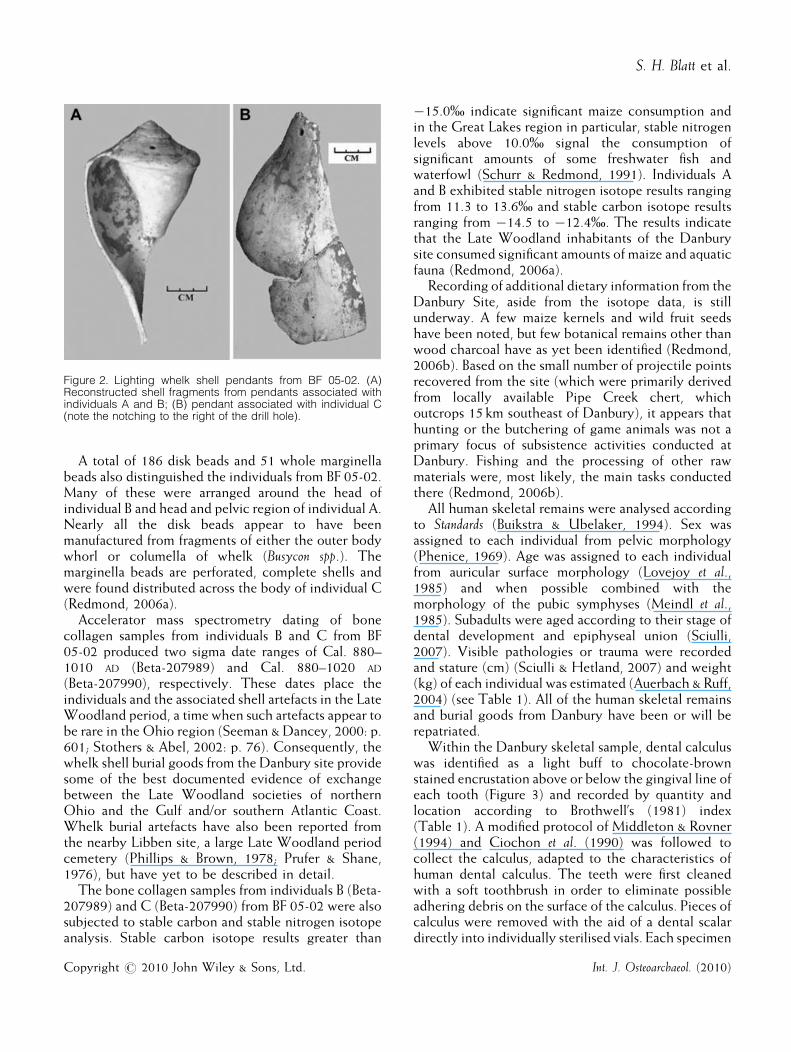
Figure 2. Lighting whelk shell pendants from BF 05-02. (A)Reconstructed shell fragments from pendants associated withindividuals A and B; (B) pendant associated with individual C(note the notching to the right of the drill hole).
S. H. Blatt et al.
A total of 186 disk beads and 51 whole marginellabeads also distinguished the individuals from BF 05-02.Many of these were arranged around the head ofindividual B and head and pelvic region of individual A.Nearly all the disk beads appear to have beenmanufactured from fragments of either the outer bodywhorl or columella of whelk (Busycon spp.). Themarginella beads are perforated, complete shells andwere found distributed across the body of individual C(Redmond, 2006a).Accelerator mass spectrometry dating of bone
collagen samples from individuals B and C from BF05-02 produced two sigma date ranges of Cal. 880–1010 AD (Beta-207989) and Cal. 880–1020 AD
(Beta-207990), respectively. These dates place theindividuals and the associated shell artefacts in the LateWoodland period, a time when such artefacts appear tobe rare in the Ohio region (Seeman & Dancey, 2000: p.601; Stothers & Abel, 2002: p. 76). Consequently, thewhelk shell burial goods from the Danbury site providesome of the best documented evidence of exchangebetween the Late Woodland societies of northernOhio and the Gulf and/or southern Atlantic Coast.Whelk burial artefacts have also been reported fromthe nearby Libben site, a large Late Woodland periodcemetery (Phillips & Brown, 1978; Prufer & Shane,1976), but have yet to be described in detail.The bone collagen samples from individuals B (Beta-
207989) and C (Beta-207990) from BF 05-02 were alsosubjected to stable carbon and stable nitrogen isotopeanalysis. Stable carbon isotope results greater than
Copyright # 2010 John Wiley & Sons, Ltd.
�15.0% indicate significant maize consumption andin the Great Lakes region in particular, stable nitrogenlevels above 10.0% signal the consumption ofsignificant amounts of some freshwater fish andwaterfowl (Schurr & Redmond, 1991). Individuals Aand B exhibited stable nitrogen isotope results rangingfrom 11.3 to 13.6% and stable carbon isotope resultsranging from �14.5 to �12.4%. The results indicatethat the Late Woodland inhabitants of the Danburysite consumed significant amounts of maize and aquaticfauna (Redmond, 2006a).Recording of additional dietary information from the
Danbury Site, aside from the isotope data, is stillunderway. A few maize kernels and wild fruit seedshave been noted, but few botanical remains other thanwood charcoal have as yet been identified (Redmond,2006b). Based on the small number of projectile pointsrecovered from the site (which were primarily derivedfrom locally available Pipe Creek chert, whichoutcrops 15 km southeast of Danbury), it appears thathunting or the butchering of game animals was not aprimary focus of subsistence activities conducted atDanbury. Fishing and the processing of other rawmaterials were, most likely, the main tasks conductedthere (Redmond, 2006b).All human skeletal remains were analysed according
to Standards (Buikstra & Ubelaker, 1994). Sex wasassigned to each individual from pelvic morphology(Phenice, 1969). Age was assigned to each individualfrom auricular surface morphology (Lovejoy et al.,1985) and when possible combined with themorphology of the pubic symphyses (Meindl et al.,1985). Subadults were aged according to their stage ofdental development and epiphyseal union (Sciulli,2007). Visible pathologies or trauma were recordedand stature (cm) (Sciulli & Hetland, 2007) and weight(kg) of each individual was estimated (Auerbach & Ruff,2004) (see Table 1). All of the human skeletal remainsand burial goods from Danbury have been or will berepatriated.Within the Danbury skeletal sample, dental calculus
was identified as a light buff to chocolate-brownstained encrustation above or below the gingival line ofeach tooth (Figure 3) and recorded by quantity andlocation according to Brothwell’s (1981) index(Table 1). A modified protocol of Middleton & Rovner(1994) and Ciochon et al. (1990) was followed tocollect the calculus, adapted to the characteristics ofhuman dental calculus. The teeth were first cleanedwith a soft toothbrush in order to eliminate possibleadhering debris on the surface of the calculus. Pieces ofcalculus were removed with the aid of a dental scalardirectly into individually sterilised vials. Each specimen
Int. J. Osteoarchaeol. (2010)

Figure 3. Molars from BF 05-02B with varying degrees of cal-culus encrustations on the lateral aspect of the posterior man-dibular dentition.
Dirty Teeth and Ancient Trade
was diluted with 5% hydrochloric acid (HCl) toremove surface carbonates and postmortem contami-nation. Specimens were allowed to digest for 2 h ingamma-sterilised microcentrifuge vials stored within adesicator cabinet.After drying, specimens were affixed to aluminium
stubs with sterile tweezers and double-sided carbontape. Specimens were coated with two layers of carbonand then stored in a humidity-controlled desiccatorcabinet until examination. Each specimen wasexamined using a JEOL JSM 820 scanning electronmicroscope (SEM) under 15Kv according to thecriteria suggested for biological specimens (Goldsteinet al., 2003). Only elements embedded within or clearlycoated with calculus were considered in situ, originatingfrom calculus calcification during the life of theindividual. Specimens were scanned at both low andhigh magnification, with most inclusions being visiblebeginning at 300�. Micrographs were obtained usingOxford Instrument’s INCA platform for SEM imaging.SHB collected and prepared the calculus samples andproduced SEM images of all specimens.Fibres were identified via SEM images according to
their distinct characteristics. Cotton seed fibres beginas elongated cells that grow on seed coats. As the fibresmature, the cell walls collapse inward to formconvolutions, giving cotton fibres their unique,ribbon-like appearance with raised edges and minimalconvolutions (Jakes et al., 1994; Teague, 1998;D’Orazio et al., 2000; Muller et al., 2004). Maturecotton fibres display well-developed convolutions,however, as the result of disease or pests, unfavourableclimatic conditions, or early harvest, the fibre in thecotton boll (a segmented pod containing seeds from
Copyright # 2010 John Wiley & Sons, Ltd.
which cotton fibres grow) may be terminated before itis fully developed, resulting in immature or ‘dead’ fibres.Immature fibres are characterised as being devoid of orhaving minimal convolutions, having little rigidity,easily creasing and causing uneven dyeing problems(David & Pailthrope, 1999).
Results
The SEM analysis of the calculus specimens fromthe entire sample yielded a variety of noticeableinclusions. These included the presence of bothmineralised bacteria (streptococci) and the outline ofdegenerated bacteria (bacillus-type), calcium-phos-phate crystalline structures, numerous identifiable andunidentifiable phytoliths (amorphous, grass and non-grass types), as well as several starch grains, a fewunidentified fibres that appear to be baste fibres, andthe presence of fibres consistent with cotton (Blatt,2007 unpublished master’s thesis). The presence ofcrystal structures and their lack of involvement withany adjacent bacteria suggests that many of thecalculus samples represent immature calculus, andthus their in situ inclusions, were mineralised less than6 months prior to the deaths of these individuals(Mandel, 1990). The presence of these inclusions aswell as the age and sex of the associated individuals,degree of calculus and calculus location are providedin Table 1. Due to the disaggregated nature of thecalculus matrix, not all specimens yielded identifiableinclusions.Evidence of cotton was recorded in the dental
calculus samples of four individuals from two burialfeatures (BF) 05-02 and BF 05-07 (Figure 4). The first,BF 05-02 was a primary burial containing three adultsextended side by side. The eastern-most individual A(BF 05-02A) was a female, approximately 45–52 yearsold, stature of 156.8 cm and body mass of 63.4 kg, withextensive degenerative joint disease of the scapulaglenoid and vertebrae (Auerbach & Ruff, 2004; Sciulli &Hetland, 2007). Individual B (BF 05-02B), a male,approximately 29–33 years old with a stature of172.9 cm and body mass of 76.3 kg, lay to her left(Auerbach & Ruff, 2004; Sciulli & Hetland, 2007).Individual C (BF 05-02C) was also a male, approxi-mately 35–45 years old at death with a stature of170.7 cm and body mass of 64.1 kg (Auerbach & Ruff,2004; Sciulli & Hetland, 2007). The human remainsfrom BF 05-07 were poorly preserved, but representedthose of an adult male; age, stature and body mass wereindeterminate due to the high degree of fragmentation(Redmond, 2006a).
Int. J. Osteoarchaeol. (2010)
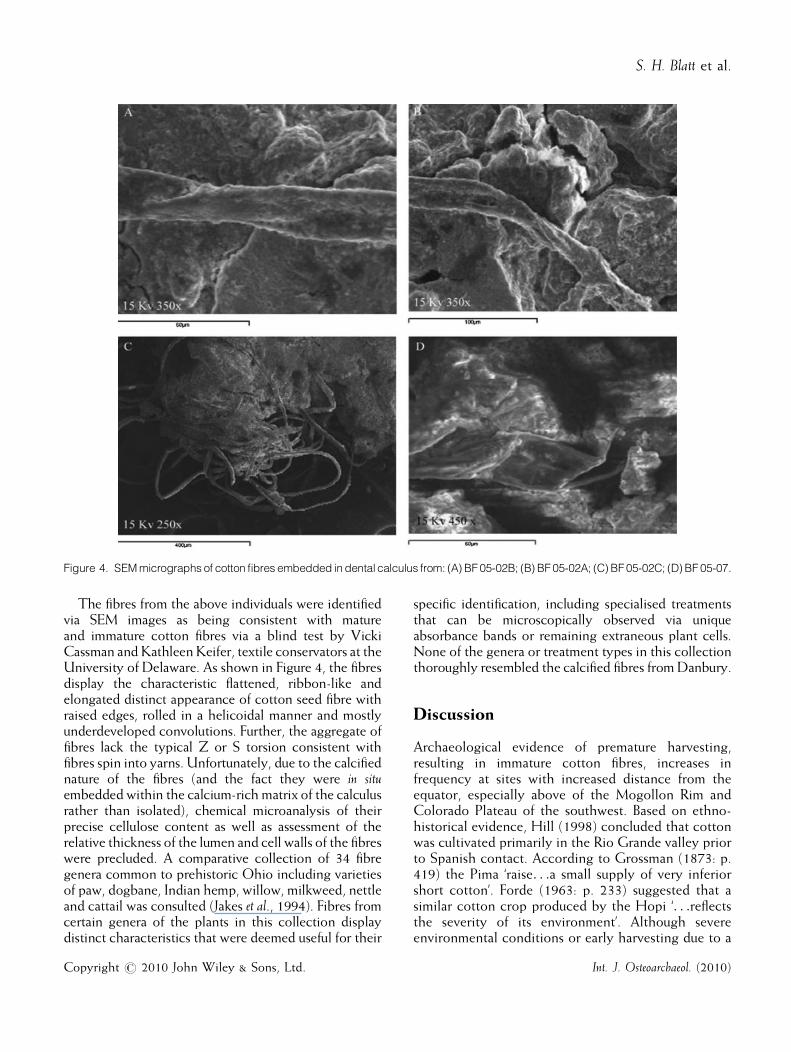
Figure 4. SEMmicrographs of cotton fibres embedded in dental calculus from: (A) BF05-02B; (B) BF05-02A; (C) BF05-02C; (D) BF 05-07.
S. H. Blatt et al.
The fibres from the above individuals were identifiedvia SEM images as being consistent with matureand immature cotton fibres via a blind test by VickiCassman andKathleenKeifer, textile conservators at theUniversity of Delaware. As shown in Figure 4, the fibresdisplay the characteristic flattened, ribbon-like andelongated distinct appearance of cotton seed fibre withraised edges, rolled in a helicoidal manner and mostlyunderdeveloped convolutions. Further, the aggregate offibres lack the typical Z or S torsion consistent withfibres spin into yarns. Unfortunately, due to the calcifiednature of the fibres (and the fact they were in situembedded within the calcium-rich matrix of the calculusrather than isolated), chemical microanalysis of theirprecise cellulose content as well as assessment of therelative thickness of the lumen and cell walls of the fibreswere precluded. A comparative collection of 34 fibregenera common to prehistoric Ohio including varietiesof paw, dogbane, Indian hemp, willow, milkweed, nettleand cattail was consulted (Jakes et al., 1994). Fibres fromcertain genera of the plants in this collection displaydistinct characteristics that were deemed useful for their
Copyright # 2010 John Wiley & Sons, Ltd.
specific identification, including specialised treatmentsthat can be microscopically observed via uniqueabsorbance bands or remaining extraneous plant cells.None of the genera or treatment types in this collectionthoroughly resembled the calcified fibres fromDanbury.
Discussion
Archaeological evidence of premature harvesting,resulting in immature cotton fibres, increases infrequency at sites with increased distance from theequator, especially above of the Mogollon Rim andColorado Plateau of the southwest. Based on ethno-historical evidence, Hill (1998) concluded that cottonwas cultivated primarily in the Rio Grande valley priorto Spanish contact. According to Grossman (1873: p.419) the Pima ‘raise. . .a small supply of very inferiorshort cotton’. Forde (1963: p. 233) suggested that asimilar cotton crop produced by the Hopi ‘. . .reflectsthe severity of its environment’. Although severeenvironmental conditions or early harvesting due to a
Int. J. Osteoarchaeol. (2010)

Dirty Teeth and Ancient Trade
short growing season could retard development,leading to immature convolution, it is difficult toassess if this trait is an effect of environment or theparticular variety of cotton (Bohrer, 1962; Kent, 1983;Teague, 1998).The Hohokam of the Sonoran Desert had a well-
established cotton industry before 1000 AD, which couldbe attributed to their construction of intricate irrigationcanals about 1500 years ago, preventing some of thedifficulties others had when growing cotton in theregion. Salinity is a concern for irrigated fields, butFryxell (1979) disregards this, describing native cottonsas having higher salt tolerance than modern cottons.This suggests that prehistoric cotton from the South-western United States could be a unique strain withimmature qualities (or that there is cotton experimen-tation within the culture that is growing this cotton).The immature length and underdeveloped convolutionsof the cotton found at the Danbury site is similar to thatof the Southwest and regions north of the equator.Historically, cotton in the Southwest was traded as rawmaterial, yarn and finished fabric, however, archae-ologically it appears that trade focused on the rawmaterial (Teague, 1998). There is evidence, based onregional fabric patterns, that long distance trade existedbetween groups in the Southwest (Teague, 1998);however, no previous evidence exists of the pre-Columbian cotton trade extending into the Midwest.As noted earlier, the nearest presence of prehistoric
cotton to the Danbury site appears to be one textilefragment tentatively identified as cotton from the SpiroMound sites in Leflore County, Oklahoma, occupiedfrom 850 to 1450 AD (King & Gardener, 1981). TheSpiro site is considered one of the most importantlocations through which exotic goods were funnelled,and there have been many artefacts from Spiroindicating long-distance trade. Most of the shell cupsfound at Spiro were made from lightning whelks(Busycon sinistrum) from the Gulf of Mexico. Also, therecent identification of Olivella beads (O. dama nativeto the California coast and found at sites throughoutthe Great Basin and southwest) from the Spiro Moundssite similarly suggests the intensification of tradebetween Puebloan and Plains peoples during the LateWoodland and Late Prehistoric periods (Kozuch,2002). The lithics from the Spiro Mounds site weremade of chert from the Cahokia, Illinois region, andobsidian from Hidalgo, Mexico; and copper from theGreat Lakes region was also among the exotic artefacts(Barker et al., 2002; Kozuch, 2002). It is possible thenthat the cotton and lightning whelk shells found at theDanbury site were obtained from a large exchangecomplex like that of Spiro.
Copyright # 2010 John Wiley & Sons, Ltd.
Although the exact origins of the cotton fibres fromDanbury are uncertain, their presence suggests longdistance trade into northern Ohio during the LateWoodland period. Both the whelk pendants (Busyconsinistrum) and cotton fibres seem to share westernGulf origins. The fact that these exogenous goods werefound together amongst a few select burials at Danburysuggests the importance of the site as a semi-permanentsettlement of rotating and interacting populations.How the cotton fibres became embedded into the
dental calculus of the Danbury individuals is unclear,but several explanations are possible. Althoughevidence of interproximal dental wear from westernGreat Basin populations suggests the use of teeth astools for weaving (Larsen, 1985), there is no suchevidence observed from the Danbury teeth, nor arethere any southwestern ethnohistoric accounts of usingteeth while spinning cotton fibres (Teague, 1998). Inthe southwest there are various spindle whorls,hooked-sticks and looms for spinning yarns andweaving fabrics, but no such evidence has been foundat the Danbury site for producing threads (Teague,1998). However, the absence of these artefacts doesnot wholly eliminate the possibility of inclusion offibres in dental calculus during spinning or fibrepreparation, which is not visible in the archaeologicalrecord (Teague, 1998). Fluffing (separating the fibres)is an important part of spinning preparation andwithout modern carding tools, pulling fibres whileanchoring with ones teeth is an easy solution. It isequally possible that while splitting plied cotton fibresby hand the fibres were anchored in the mouth andbecame incorporated into calculus. Most likely theirpresence represents the reprocessing of cotton fibresfor the production of textiles or fishing line or nets.The association of such exotic marine pendants as
the lightning whelk shells coupled with evidence ofcotton use also generates questions as to the relativeprestige of individuals A, B and C of BF 05-02 and BF05-07, since such goods were absent in other burials.Certainly the link between cotton as an exotic fibre, itsuse as a tool for improved fishing technologies and animportant source of protein might be a link to socialstatus, though further investigations are needed todocument the extent to which these objects reflectstatus within this population.
Conclusions
The analysis of human dental calculus provides aninnovative and direct assessment of ancient diet andgives additional evidence when combined with stable
Int. J. Osteoarchaeol. (2010)

S. H. Blatt et al.
isotope studies. Calculus analysis has advantages overstable isotopes of being non-destructive, less expensiveand generally more precise with regard to identifyingspecific plant materials.Though in practice it would be ideal to assess both
the physical and chemical nature of all archaeologicalfibres, we hope that the data presented here will alert theanthropological community to the value of microscopicanalysis of dental calculus in supplementing ourunderstanding of paleoethnobotanical histories. Weanticipate that additional studies of this kind will resultin the identification of cotton at sites outside theSouthwest, and yield more detailed results beyond thoseof this report. In light of these new data, further researchis definitely needed to gain a more comprehensive viewof the spheres of interaction and trade of exotic goods inprehistoric North America. While the identification ofplant fibres from archaeological contexts is essential forunderstanding ethnobotanical history, identification islimited by the wide range of plant material potentiallyin use, the absence of plant parts used for standardidentification and the alteration of fibres from proces-sing, use and degradation within the burial environment.The methods used in this paper, however, provide
the potential for analysis of plant materials that wouldotherwise be digested or lost from employing themultiple washes or higher hydrochloric acid concen-trations suggested by other authors. Analysis of dentalcalculus is an ideal complementary method to stableisotopic reconstruction, because it provides infor-mation about foods eaten a short time before death(from days to months), as opposed to the long-termperspective on diet yielded by isotope analysis. Dentalcalculus analysis offers an inexpensive and relativelyaccurate means of assessing the herbaceous portion ofancient diet, but like most paleodietary techniques, itshould be combined with other methods.What is clear is that the cotton fibres discovered
within dental calculus from the Danbury site are notindigenous to Ohio and were introduced to the LateWoodland component of this site from an unknownsouth-western source. The interaction between north-ern Ohio and southern coastal regions is supported bythe evidence of both cotton and marine shell exchangebetween these groups. The whelk shells do not point tothe Southwest as a source per se, rather they point toanother avenue of long distance exchange (from lowerGreat Lakes to the Gulf of Mexico) which was mostlikely channelled, along with cotton, through anintermediary like Spiro. Although the precise variety ofcotton found at the Danbury site and the circumstancesbehind such an undertaking are unclear, the ramifica-tions of such broad-based exchange in prehistoric
Copyright # 2010 John Wiley & Sons, Ltd.
North America could be far-reaching and is deservingof further examination.
Acknowledgements
Partial funding for this project was provided by theCleveland Chapter of the Archaeological Institute ofAmerica. Scanning electron microscopy was performedat the Microscopic and Chemical Analysis ResearchCenter (MARC), Department of Earth Sciences at TheOhio State University. The authors express gratitudeto those who offered their comments, suggestions andsupport of this project: Kristen Gremillion, DebbieGuatelli-Steinberg, Kathleen Keifer, Greg Pellam,Gabriela Jakubowska, Ana M. Casado, Gary Blattand Larry T. Boston. Appreciation is also offered tothe editor and two anonymous reviewers whose sug-gestions and criticisms have improved this work. Theauthors also thank the numerous dedicated volunteersat the Cleveland Museum of Natural History, both inthe field and in the lab, without whom this work wouldnot have been possible. For the remaining imperfec-tions, the first author is alone responsible.
References
Arensburg B. 1996. Ancient dental calculus and diet. HumanEvolution 11(2): 139–145.
Armitage PL. 1975. The extraction and identification of opalphytoliths from the teeth of ungulates. Journal of Arche-aological Science 2(3): 187–197.
Auerbach BM, Ruff CB. 2004. Human body mass estimation:a comparison ‘‘morphometric’’ and ‘‘mechanical’’ methods.American Jornal of Physical Anthropology 125(4): 331–342.
Barker FC, Griffen JB, Morgan RG, Neumann GK, TaylorLJB. 2002. Mesoamerican origin for an obsidian scraperfrom the Pre-Columbian southeastern United States.American Antiquity 67: 103–108.
Blondiaux J, Charlier P. 2008. Palaeocytology in skeletalremains: microscopic examination of putrefication fluidsand dental calculus of skeletal remains from Frencharchaeological sites. International Journal of Osteoarcheology18: 1–10.
Bohrer VL. 1962. Ethnobotanical materials from TontoNational Monument. In Archeological Studies at TontoNational Monument, Arizona, Caywood LR (ed.). SouthwestMonuments Association Technical Series 2. SouthwestMonuments Association: Globe.
Boyadjian HCC, Eggers S, Reinhard K. 2007. Dental wash: aproblematic method for extracting microfossils fromteeth. Journal of Archaeological Science 34: 1622–1628.
Brothwell DR. 1981. Digging Up Bones. Cornell UniversityPress: New York; 159–160.
Int. J. Osteoarchaeol. (2010)

Dirty Teeth and Ancient Trade
Buikstra JE, Ubelaker DH. 1994. Standards for the Data Collec-tion from Human Skeletal Remains: Proceedings of a Seminar at theField Museum of Natural History, organized by JonathanHaas. Field Museum of Natural History, Fayetteville,AR: Arkansas Archeological Survey.
Bullen RP. 1974. Were there Pre-Columbian cultural con-tacts between Florida and the West Indies: the archae-ological evidence. Florida Anthropology 27: 149–160.
Ciochon RL, Piperno DR, Thompson RG. 1990.Opal phytoliths found on the teeth of the extinct apeGigantopithecus blacki: implications for paleodietarystudies. Proceedings of National Academy of Science 87:8120–8124.
Cortello AR, Pochettino MI. 1994. Starch grain analysis as amicroscopic identification feature in the identification ofplant material. Economic Botany 48: 171–181.
David SK, Pailthrope MT. 1999. Classification of textilefibers: production, structure, and properties. In ForensicExamination of Fibres (2nd edn), Robertson J, Grieve M(eds). Taylor & Francis, Ltd.: London; 1–31.
Dobney K, Brothwell D. 1986. Dental calculus: its relevanceto ancient diet and oral ecology. In Teeth and Anthropology,Cruwys E, Foley RA (eds). BAR International Series 291:Oxford; 55–81.
D’Orazio L, Martuscelli E, Oresello G, Riva F, Scala G,Taglialetela A. 2000. Nature, origin, and technology ofnatural fibers of textile artifacts recovered in the ancientcities around Vesuvius. J Arch Sci 27: 745–754.
Evans ST. 2004. Ancient Mexico and Central America, Archaeologyand Culture History. Thames and Hudson Press: London.
Forde CD. 1963. Habitat, Economy, and Society. E.P. Dutton:New York.
Fox CL, Juan J, Albert RM. 1996. Phytolith analysis ondental calculus, enamel surface, and burial soil: infor-mation about diet and paleoenvironment. American Journalof Physical Anthropology 101: 101–113.
Fryxell PA. 1979. The Natural History of the Cotton Tribe:Malvaceae, Tribe Gossypiae. University Press: CollegeStation: Texas A&M.
Gobetz KE, Bozarth SR. 2001. Implications for Late Pleis-tocene mastodon diet from opal phytoliths in toothcalculus. Quaternary Research 55: 115–122.
Goldstein J, Newbury D, Joy D, Lyman C, Echlin P, LifshinE, Sawyer L, Michael J. 2003. Scanning Electron Microscopyand X-ray Microanalysis, (3rd edn). Springer: New York.
Grossman FE. 1873. The Pima Indians of Arizona. In AnnualReport of the Board of Regents of the Smithsonian Institution Showingthe Operations, Expenditures, and Condition of the Institution for theYear 1871. The Smithsonian Institute: Washington, D.C.
Hershkovitz I, Kelly J, Latimer B, Rothschild BM, SimpsonS, Polak J, Rosenberg M. 1997. Oral bacteria in MioceneSivapithecus. Journal of Human Evolution 33: 507–512.
Hill JB. 1998. Ecological variability and agricultural special-ization among the Protohistoric Pueblos of Central NewMexico. Journal of Field Archaeology 25(3): 275–294.
Jakes KA, Sibley LR, Yerkes R. 1994. A comparative collec-tion for the study of fibres used in prehistoric textiles from
Copyright # 2010 John Wiley & Sons, Ltd.
Eastern North America. Journal of Archaeological Science 21:641–650.
Kent KP. 1983. Prehistoric Textiles of the Southwest. School ofAmerican Research: Sante Fe.
King, M.E., Gardener, J.S., 1981. The analysis of textilesfrom Spiro Mound, Oklahoma. Annals of the New YorkAcademy of Sciences 376(1): 123–139.
Kozuch L 2002 Olivella beads from Spiro and the PlainsAmerican Antiquity 67(4): 697–709.
Larsen CS. 1985. Dental modifications and tool use in thewestern Great Basin. American Journal of Physical Anthropol-ogy 67(4): 393–402.
Lee, J.A., 1984. Cotton as a world crop. In Cotton, Kohl , RJLewis , CF (eds). Agronomu 24. American Society ofAgronomy: Madison.
Lovejoy CO,Meindl RS, Pryzbeck TR, Mensforth RP. 1985.Chronological metamorphosis of the auricular surface ofthe ilium: a new method for the determination of adultskeletal age at death. American Journal of Physical Anthro-pology 68(1): 15–28.
MacPhee T, Cowley G. 1975. Essentials of Periodontology andPeriodontics, (2nd edn). Blackwell Scientific Publications:Oxford.
Mandel ID. 1990. Calculus formation and prevention: anoverview. Compendium of Continuing Education in Dentistry,Supplement 8: 235–241.
Meindl RS, Lovejoy CO,Mensforth RP,Walker RA. 1985. Arevised method of age determination using the os pubis,with a review and tests of accuracy of other currentmethods of pubic symphyseal aging. American Journal ofPhysical Anthropology 68: 29–45.
Middleton WD. 1990. An improved method for extractionof opal phytoliths from tartar residues on herbivore teeth.Phytolitherian Newsletter 6: 2–5.
Middleton WD. 1992a. Preliminary report on the extractionof opal phytoliths from tartar residues on herbivore teeth.Phytolitherian Newsletter 7: 5.
MiddletonWD. 1992b. Preliminary report on the extractionof phytoliths from cervid dental tartar. Phytolitherian News-letter 7(1).
Middleton WD, Rovner I. 1994. Extraction of opal phyto-liths from herbivore dental calculus. Journal of ArchaeologicalScience 21: 469–473.
Muller M, Papiz MZ, Clarke DT, Roberts MA, Murphy BM,Burghammer M, Riekel C, Pantos E, Gunneweg J. 2004.Identification of textiles using microscopy and synchro-tron radiation X-ray fibre defraction. Spectrohima Acta 59:175–186.
Phenice TW. 1969. A newly developed visual method ofsexing the os pubis. American Journal of Physical Anthropology30(2): 297–301.
Phillips P, Brown JA. 1978. Pre-Columbian Shell Engravings fromthe Craig Mound at Spiro, Oklahoma, Part 1. Peabody MuseumPress: Cambridge, Massachusetts.
Prufer OH, Shane OC. 1976. The Portage-Sandusky-Vermilion River Region in Ohio. In The Late Prehistory ofthe Lake Erie Drainage Basin, A 1972 Symposium Revised,
Int. J. Osteoarchaeol. (2010)

S. H. Blatt et al.
Brose DS (ed.), Scientific Publications of the ClevelandMuseum of Natural History: Cleveland, Ohio; 283–304.
Redmond, B.G., 2005. A report of the archaeological inves-tigations at the Danbury site (33OT16): 2004 season.Cleveland Museum of Natural History, Archaeological ResearchReports, No. 146.
Redmond, B.G., 2006a A report of the archaeological inves-tigations at the Danbury site (33OT16): 2005 season.Cleveland Museum of Natural History, Archaeological ResearchReports, No. 147.
Redmond, B.G., 2006b Saving the Danbury site (33OT16):investigation of Woodland to Late Prehistoric settlementand mortuary behavior along the Lake Erie shore. OhioArchaeological Council. [HTLM title] Available online:March 15, 2006. Ohioarchaeology.org.
Redmond, B.G., 2007. A report of the archaeological inves-tigations at the Danbury site (33OT16): 2006 season.Cleveland Museum of Natural History, Archaeological ResearchReports, No. 147.
Schurr MR, Redmond BG. 1991. Stable isotope analysis ofincipient maize horticulturalists from the Gard Island 2Site. Midcontinent Journal of Archaeology 16(1): 69–84.
Sciulli PW. 2007. Relative dental maturity and associatedskeletal maturity in prehistoric Native Americans of theOhio valley area. American Journal of Physical Anthropology132: 545–557.
Sciulli PW, Hetland BM. 2007. Stature estimation for pre-historic Ohio Valley native American populations basedon revisions of the fully technique. Archaeology of EasternNorth America 35: 105–113.
Seeman MF, Dancey WS. 2000. The Late Woodland Periodin Southern Ohio: basic issues and prospects. In Late
Copyright # 2010 John Wiley & Sons, Ltd.
Woodland Societies, Tradition and Transformation Across theMidcontinent, TE, Emerson DL, McElrath AC Fortier(eds). University of Nebraska Press: Lincoln, Nebraska;583–611.
Seidemann R. 2006. The Bahamian problem in Floridaarchaeology: oceanographic perspectives on the issueof Pre-Columbian contact. KACIKE. Journal of CaribbeanAmerindian History and Anthropology 7(1): 33–61.
Stothers DM, Abel TJ. 2002. The Early Late Woodlandin the Southwestern Lake Erie Littoral Region. InNortheast Subsistence-Settlement Change A.D. 700–1300, HartJP, Rieth CB (eds). State Museum Bulletin 496: New York;73–96.
Stark BL, Heller L, Ohnersorgen MA. 1998. People withcloth: Mesoamerican economic change from theperspective of cotton in South-Central Veracruz. LatinAmerican Antiquity 9(1): 7–36.
Teague LS. 1998. Textiles in Southwestern Prehistory. Universityof New Mexico Press: Albuquerque.
Trubitt, MBD. 2003. The Production and Exchange ofMarine Shell Prestige Goods. Journal of ArchaeologicalResearch 11(3): 243–277.
Vandermeersch B, Arensburg B, Tillier A, Rak Y, Weiner S,Spiers M, Aspillaga E. 1994. Middle Paleolithic dentalbacteria from Kebara, Israel. C.R. Academy of Science Paris319(2): 727–731.
Wendel JF, Brubaker CL, Percival AE. 1992. Genetic diver-sity in Gossypium hirsutum and the origin of Upland cotton.American Journal of Botany 79: 1291–1310.
Wendel JF. 1995. Cotton. In Evolution of Crop Plants,Simmonds N, Smartt J (eds). Longman: London; 358–366.
Int. J. Osteoarchaeol. (2010)




















