
HLA-E expression by gynecological cancers restrains tumor-infiltrating CD8⁺ T lymphocytes
Upload
independentCategory
view
8download
0
Differential Cell Cycle Progression Patterns of InfiltratingLeukocytes and Resident Cells After Balloon Injury of the
Rat Carotid ArteryCarl Hay, Connie Micko, Margaret Forney Prescott, Gene Liau, Keith Robinson, Hector De Leon
Abstract—The heterogeneous nature of the cell populations involved in vascular repair remains a major hurdle for theassessment of the cellular events that take place in injured arteries. The present experiments were designed to estimatethe proportions and cell cycle progression of infiltrating leukocytes versus resident vascular cells after balloon injury ofthe rat common carotid artery. After tissue disaggregation, cell suspension samples from each artery were analyzed byflow cytometry. Cells were stained with anti-CD45 or anti–�-smooth muscle actin antibodies to identify leukocytes andsmooth muscle cells, respectively. A day after injury, a 12-fold increase in CD45� leukocytes was found. Doublelabeling with CD45 and CD-3, ED-1, or granulocyte markers revealed that most infiltrating cells were monocytes andgranulocytes. Approximately 14% of infiltrating leukocytes were found to enter apoptosis at day 1, and 17% entered Sphase at day 3. In contrast, the highest proliferation rate of resident �-smooth muscle actin–positive cells was observedat day 7 (19%). The present results demonstrate that infiltrating leukocytes and resident vascular smooth muscle cellshave dissimilar cell cycle profiles. Furthermore, our study demonstrates the feasibility of using flow cytometry toquantitatively determine the cell types and their relative activation state in injured arteries. (Arterioscler Thromb VascBiol. 2001;21:1948-1954.)
Key Words: restenosis � vascular remodeling � inflammation � flow cytometry � apoptosis
Quiescent vascular smooth muscle cells (SMCs) respondto acute vascular injury with an array of phenotypic
changes, including proliferation, migration, programmed celldeath, and modulation to a less differentiated state.1–3 Deen-dothelialization of the carotid artery is followed by prolifer-ation of medial SMCs, with subsequent migration of thesecells to the intima, where they proliferate and secrete extra-cellular matrix.4
Inflammation is associated with the development of ath-erosclerotic lesions and is thought to play a critical role invascular SMC proliferation, migration, and matrix productionin response to injury.5 Interleukin (IL)-1� and tumor necrosisfactor-� may induce the expression of monocyte chemoat-tractant protein (MCP)-1, IL-8, intercellular adhesionmolecule-1, and vascular adhesion molecule (VCAM)-1 toassist the recruitment of leukocytes into a mechanicallyinjured tissue or an atherosclerotic lesion.6–9 MCP-1 andMCP-3 have been shown to be upregulated in the neointi-ma,10,11 and inhibition of intimal proliferation by antibodies(Abs) against MCP-1 supports the role of these molecules invascular lesion formation.12
To further evaluate the contribution of proliferating leuko-cytes to the vascular repair process, we examined the cellcycle status of infiltrating leukocytes and vascular SMCs in
vivo by using a well-characterized model of balloon injury.By use of a double-labeling technique, the proliferative andDNA content profiles of vascular SMCs and infiltratingleukocytes were analyzed by flow cytometry.
MethodsCarotid InjuryBalloon injury of the common carotid artery was performed asdescribed by Clowes et al.13 To identify proliferating cells, bromode-oxyuridine (BrdU, 50 mg/kg IP, Sigma Chemical Co) was adminis-tered 18 hours before tissue harvesting. For the time-course flowcytometric analysis, rats (n�4 per time point) were euthanized atdays 1, 3, 7, and 15. Carotid arteries from 4 naive non–balloon-injured animals were used as controls. Two additional groups of rats(n�3 each) were balloon-injured and euthanized 1 day after injury toestimate the subpopulations of infiltrating leukocytes by flow cy-tometry and immunohistochemistry. Contralateral uninjured arterieswere used as controls in these experiments. No attempt was made toremove the adventitia or perivascular connective tissue. Animalstudies were approved by the Genetic Therapy, Inc and AtlantaCardiovascular Research Institute Institutional Committees for theCare and Use of Animals and were in accordance with federalguidelines.
Tissue Disaggregation and Cell IsolationCommon carotid arteries were washed in PBS containing 10 U/mLheparin, cut into �1-mm pieces, and incubated in RPMI containing
Received September 13, 2001; revision accepted September 19, 2001.From Genetic Therapy, Inc (C.H., G.L., H.D.L.), Gaithersburg, Md; Atlanta Cardiovascular Research Institute (C.M., K.R., H.D.L.), Norcross, Ga; and
Novartis Pharmaceuticals (M.F.P.), East Hanover, NJ.Correspondence to Hector De Leon, MD, PhD, Atlanta Cardiovascular Research Institute, 3155 Northwoods Place, Norcross, GA 30071. E-mail
[email protected]© 2001 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org
1948 by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from
collagenase (2 mg/mL, type IV, Sigma) and elastase (0.4 mg/mLpancreatic, Sigma) for 1 hour at 37°C. Twenty percent FBS wasadded to the resulting cell suspension, and the cells were passedthrough a 100-�m cell strainer (Becton Dickinson). Microscopicmonitoring through the digestion period revealed a rapid andextensive disaggregation of arterial tissue and the presence ofspherical cells. The number of cells obtained from whole commoncarotid artery digestion ranged from 2�105 to 6�105.
Flow CytometryAfter disaggregation, cell suspensions were pelleted and washedtwice with PBS containing 3% FBS and divided into aliquots(0.4�105 to 1�105 cells per tube). Selected samples were stained for15 minutes at 4°C with a phycoerythrin-labeled Ab against rat CD45(clone OX-1, 1/100, BD Pharmingen) or a matched isotype control(phycoerythrin-labeled IgG1, Sigma). Cells were then fixed with 4%formaldehyde, permeabilized with 0.1% saponin, and stained forBrdU incorporation by use of an FITC-labeled anti-BrdU Ab (BrdUFlow Kit, BD Pharmingen). Selected samples were then stained withan anti–�-smooth muscle actin (anti–�-SMA) Ab (1/100, CedarlaneLaboratories) or an isotype control (IgG2a, Sigma). The unconju-gated primary �-SMA Ab cell samples and the correspondingisotype controls were incubated with phycoerythrin-labeled goatanti-mouse IgG (1/80, Sigma). Cell samples were stained for DNAcontent by using 7-aminoactinomycin D (7-AAD, 20 �L per sam-ple). For analysis of leukocyte subpopulations, cells were double-stained with phycoerythrin-labeled anti-rat CD45 and FITC-labeledanti-rat CD3 (clone G4.18), granulocyte (clone HIS48), or mononu-clear phagocyte (ED-1 antigen, clone 1C7) Abs (1/100, BD Pharm-ingen). Samples were also double-stained with the appropriateisotype controls for flow cytometry analysis gating. The unconju-gated primary mononuclear phagocyte Ab cell samples and corre-sponding isotype controls were incubated with FITC-labeled goatanti-mouse IgG (1/40, Sigma). Samples were then resuspended in750 �L of PBS and analyzed by flow cytometry by using a Becton
Dickinson FACSCalibur flow cytometer with a 15-mW argon-ionlaser operating at 488 nm.
ImmunohistochemistryImmunohistochemical localization of leukocytes with the use ofmonoclonal anti-CD45 and mononuclear phagocyte Abs (cloneOX-1 and 1C7, 1/100 dilution, BD Pharmingen) was performed onfrozen sections of injured and contralateral uninjured carotid arteriesharvested 1 day after balloon injury. The primary Abs were appliedat the indicated dilutions, and the slides were incubated with abiotinylated secondary Ab and stained by use of the ABC-AP system(Vector Laboratories).14
Statistical AnalysisThe results are expressed as mean�SEM. The data were evaluatedby 1-way ANOVA. When the overall F test of the ANOVA analysiswas significant, a multiple comparison Dunnett test was applied. AStudent t test was used in 2-mean comparisons.
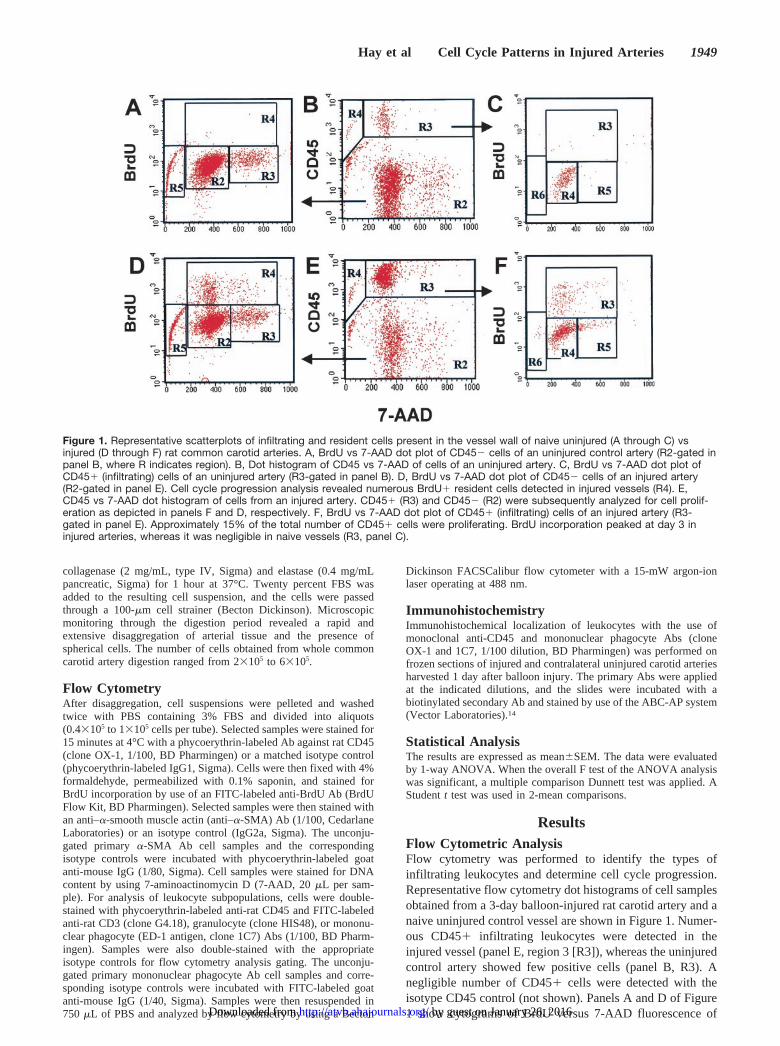
ResultsFlow Cytometric AnalysisFlow cytometry was performed to identify the types ofinfiltrating leukocytes and determine cell cycle progression.Representative flow cytometry dot histograms of cell samplesobtained from a 3-day balloon-injured rat carotid artery and anaive uninjured control vessel are shown in Figure 1. Numer-ous CD45� infiltrating leukocytes were detected in theinjured vessel (panel E, region 3 [R3]), whereas the uninjuredcontrol artery showed few positive cells (panel B, R3). Anegligible number of CD45� cells were detected with theisotype CD45 control (not shown). Panels A and D of Figure1 show cytograms of BrdU versus 7-AAD fluorescence of
Figure 1. Representative scatterplots of infiltrating and resident cells present in the vessel wall of naive uninjured (A through C) vsinjured (D through F) rat common carotid arteries. A, BrdU vs 7-AAD dot plot of CD45� cells of an uninjured control artery (R2-gated inpanel B, where R indicates region). B, Dot histogram of CD45 vs 7-AAD of cells of an uninjured artery. C, BrdU vs 7-AAD dot plot ofCD45� (infiltrating) cells of an uninjured artery (R3-gated in panel B). D, BrdU vs 7-AAD dot plot of CD45� cells of an injured artery(R2-gated in panel E). Cell cycle progression analysis revealed numerous BrdU� resident cells detected in injured vessels (R4). E,CD45 vs 7-AAD dot histogram of cells from an injured artery. CD45� (R3) and CD45� (R2) were subsequently analyzed for cell prolif-eration as depicted in panels F and D, respectively. F, BrdU vs 7-AAD dot plot of CD45� (infiltrating) cells of an injured artery (R3-gated in panel E). Approximately 15% of the total number of CD45� cells were proliferating. BrdU incorporation peaked at day 3 ininjured arteries, whereas it was negligible in naive vessels (R3, panel C).
Hay et al Cell Cycle Patterns in Injured Arteries 1949
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from

CD45� cells from uninjured and injured arteries, respec-tively. Gated region 4 (R4) in panels A and D representCD45� cells in S phase, ie, the fraction of cells with ameasured DNA content between that of the G0/G1 phase(diploid or 2C) and G2/M phase (tetraploid or 4C). Gated R3sections in panels C and F show the fraction of inflammatorycells (CD45�), which were proliferating (BrdU�) in unin-jured and injured arteries, respectively. Compared with theuninjured vessel (panel C, R3), the injured artery (panel F)showed a higher number of proliferating CD45� cells (R3).The same analysis performed with an isotype control Abshowed few scattered background events in the same R3region (not shown). Therefore, using flow cytometry analysis,we demonstrated a large increase in CD45� cells in theinjured artery and increased proliferation of the CD45� andCD45� populations.
Time Course of Recruitment of InflammatoryCells to the Injury SiteWe observed a 11.9-fold increase in cells that stained posi-tively for the pan leukocyte Ab CD45 at day 1 after injury(Figure 2). Indeed, 26.1�4.5% of the total number of cellspresent in injured arteries at day 1 was CD45�. However, 3days after arterial damage, the percentage of infiltrating cellsdropped to 10.8�1.2%, representing an almost 5-fold in-crease over control values (2.2�0.3%). At 7 and 15 days,CD45� cells accounted for 17.4�1.1% and 11.2�0.4%,representing 7.8- and 4.6-fold increases, respectively, overvalues in naive uninjured arteries (Figure 2).
Subpopulations of Infiltrating LeukocytesTo further characterize the circulating cell types recruited tothe injury site, we carried out simultaneous labeling withCD45 and specific Abs against subpopulations of T cells(CD3), monocyte/macrophages (ED1), and granulocytes(HIS148 monoclonal Ab). Figure 3 depicts 2-parameter dothistograms of a representative experiment with cells from1-day injured arteries and their contralateral controls. TheCD45� vascular infiltrate (black dots) in injured arteries wasprimarily due to the recruitment of granulocytes (49.1%,
panel E) and monocytes (42.2%, panel F). The proportion ofinfiltrating T cells (2.7%, panel D) was small compared withthe proportion of granulocytes and monocytes. The largestincrease in cell recruitment was observed with granulocytes,which showed a 32-fold increase (from 38.8�12.3 to1255.0�131.4; Table 1). T cells demonstrated a 8.7-foldincrease (from 34.3�11.4 to 298.3�22.6; Table 1). Whereasthere was almost an equivalent number of monocytes andgranulocytes in the injured tissue, the monocytic enhance-ment was only 6-fold because of the presence of ED-1� cellsin the contralateral uninjured vessels (from 275.5�52.2 to1628.0�515.0; Table 1). A slightly higher percent of CD45�cells was detected in contralateral arteries of balloon-injuredrats compared with naive vessels from non–balloon-injuredanimals (4.1�0.2% versus 2.2�0.3%, respectively; P�0.05).Almost 40% of CD45� cells in contralateral vessels wereED-1� (Figure 3, panel C, red dots).
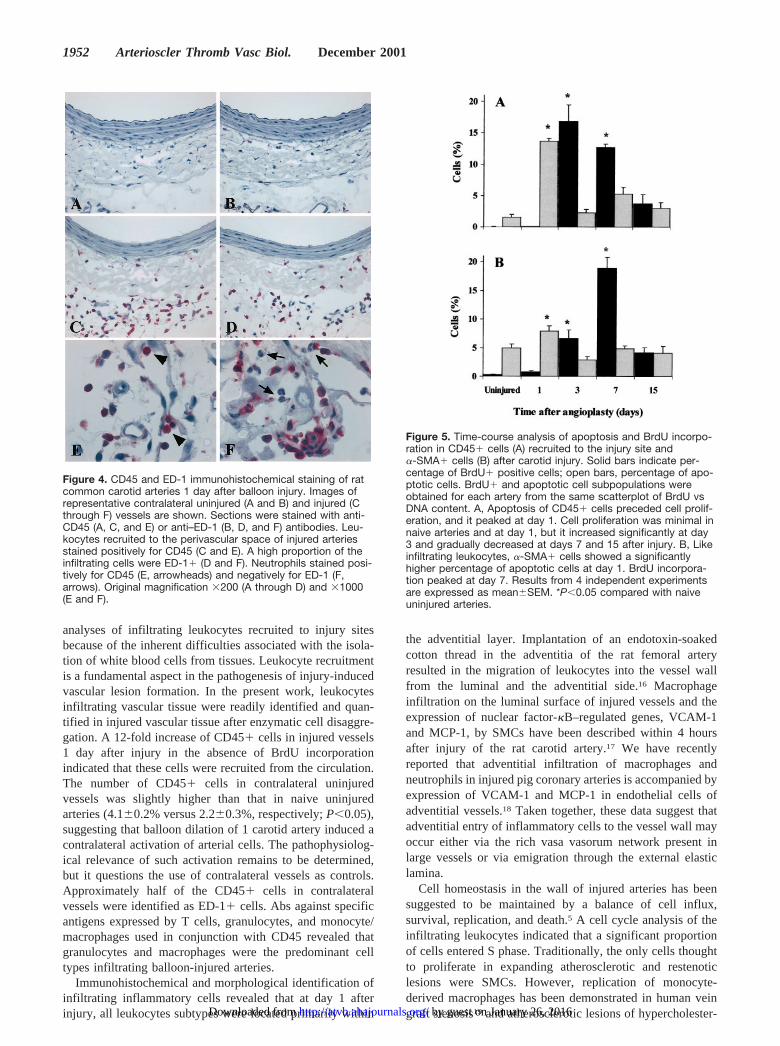
Immunohistochemical Localization ofInfiltrating CellsTo aid in determining the localization of infiltrating CD45�cells within the arterial wall, immunohistochemistry wasperformed on histological sections from an additional groupof animals euthanized at day 1 after injury, which is the peaktime for infiltration of CD45� cells as revealed by flowcytometry. Uninjured contralateral arteries demonstrated fewscattered CD45� and ED-1� cells in the adventitia andperivascular region (Figure 4A and 4B). One day after injury,however, a large number of CD45� cells were observedwithin the adventitia of the vessel (Figure 4C and 4E). Asimilar localization pattern was found for monocytes/macro-phages (Figure 4D and 4F) and granulocytes (identified bytheir nuclear morphology). In CD45-stained and ED-1–stained frozen sections, granulocytes were stained positivelyfor CD45 (Figure 4E) and negatively for ED-1 (Figure 4F).
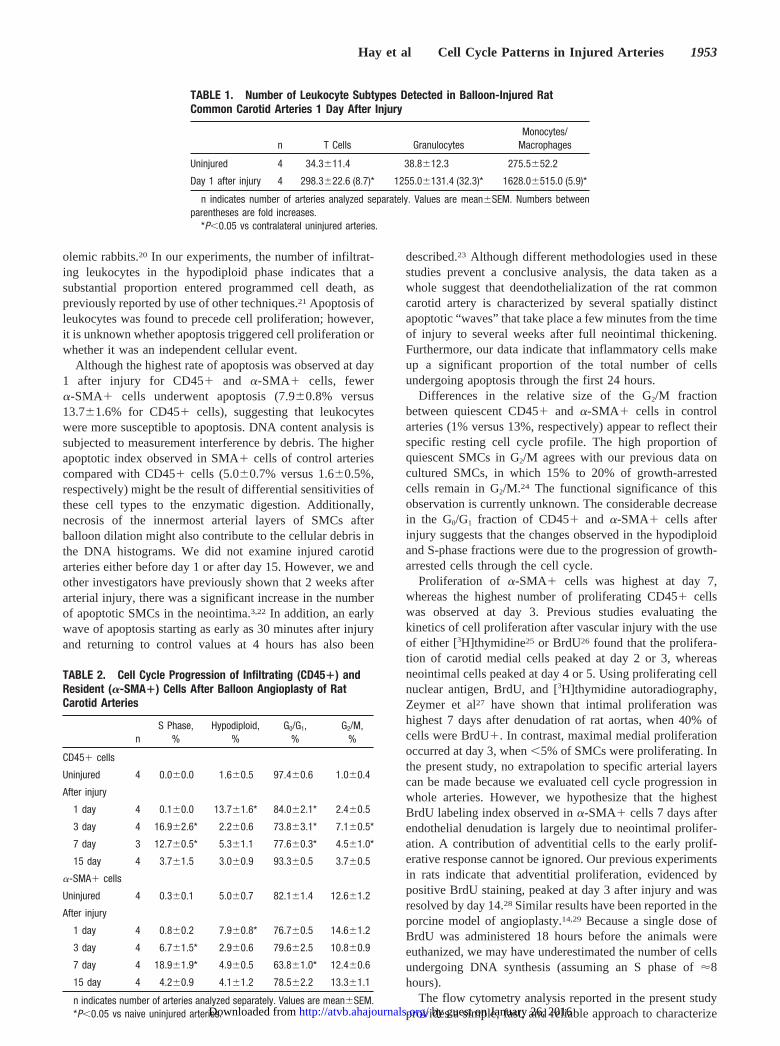
Cell Cycle Progression of Infiltrating CD45� CellsCell distribution in G0/G1, G2/M, S, and sub-G1 phases wasestimated by flow cytometric determinations of DNA ploidyafter DNA counterstaining with 7-AAD. Sub-G1 cells (hyp-odiploid) were identified as apoptotic cells. The number ofBrdU� cells present in S phase was used to compare theproliferating index in injured vessels to that of naive unin-jured arteries. Figure 5A illustrates that 13.7�1.6% of thetotal number of CD45� cells were apoptotic on day 1 afterinjury. This was an almost 9-fold increase over uninjuredarteries and represented the highest percentage of apoptoticcells observed at any time point examined. The percentage ofapoptotic cells at days 3 (2.2�0.6%), 7 (5.3�1.1%), and 15(3.0�0.9%) after injury was not significantly different fromcontrol values (P�0.05, Figure 5A). Cell proliferation wasnegligible in CD45� cells present in the vessel wall ofuninjured and injured (1-day) carotid arteries (Figure 5A).However, 3 days after injury, cell proliferation was signifi-cantly increased in injured vessels, with 16.9�2.6% ofCD45� cells BrdU positive (P�0.05, Figure 5A). At day 7,proliferation of infiltrating leukocytes remained elevatedcompared with control uninjured vessels (P�0.05, Figure5A). By day 15, BrdU�/CD45� cells had decreased to3.7�1.5%.
Figure 2. Time course infiltration of CD45� to arterial tissueafter balloon injury of the rat common carotid artery. An almost12-fold increase of CD45� cells was detected at day 1. By day3, this increase was reduced by more than half, but the numberof CD45� cells was significantly higher than control values atdays 3, 7, and 15. Results from 4 independent experiments areexpressed as mean�SEM. *P�0.05 compared with naive unin-jured arteries (Uninj).
1950 Arterioscler Thromb Vasc Biol. December 2001
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from

The small number of CD45� cells present in uninjuredvessels was quiescent. No cells in S phase were detected innaive uninjured vessels, whereas �97% of cells were inG0/G1 phase, and 1% was in G2/M phase (Table 2). By day 1after injury, the number of CD45� cells in the hypodiploidphase was increased, whereas the number of cells in the G0/G1
compartment was significantly decreased (P�0.05, Table 2).At day 3, a further significant reduction in the G0/G1 phasereflected the transition of numerous cells to S phase and G2/Mphase (Table 2). Seven days after injury, a cell cycle patternsimilar to that on day 3, with a significant reduction in G0/G1
and increases in S and G2/M phases, was observed (P�0.05,Table 2). By day 15, the cell cycle progression profile ofCD45� cells had returned to control values.
Determination of the Percentage of �-SMA� Cellsin the Apoptotic and Proliferative CompartmentTo further explore the proliferative status of cells expressing�-SMA (SMCs, SMC-derived neointimal cells, and adventi-tial myofibroblasts), samples were labeled simultaneouslywith fluorescent monoclonal anti–�-SMA and anti-BrdU Absafter permeabilization and staining with 7-AAD. CellularDNA content and BrdU incorporation were then analyzed by2-parameter flow cytometry. Figure 5B shows that 5.0�0.7%of �-SMA� cells in naive uninjured arteries were apoptotic.The number of apoptotic cells increased to 7.9�0.8% at day
1 (P�0.05, Figure 5B) but returned to levels similar to thoseof control vessels at days 3, 7, and 15. As observed withinfiltrating CD45� cells, replication of �-SMA� cells wasnegligible in uninjured arteries (0.3�0.1%) and arteries 1 dayafter injury (0.8�0.2%, Figure 5B). Three days after injury,6.7�1.5% of �-SMA� cells were BrdU positive. The highestrate of proliferation in �-SMA� cells was observed at day 7(18.9�1.9%, Figure 5B), whereas the highest rate of prolif-eration of CD45� cells was observed at day 3 (Figure 5A). Inaddition, a higher percentage of �-SMA� cells in the vesselwall of uninjured arteries was observed in G2/M phasecompared with CD45� cells (12.6�1.2 versus 1.0�0.4,Table 2). Few �-SMA� cells in S phase were detected inuninjured arteries and 1 day after balloon injury. The mostsignificant change in cell cycle progression of �-SMA� cellswas observed at day 7, with a decrease in the G0/G1
population and a concomitant increase in the S-phase fraction(P�0.05, Table 2). In contrast to CD45� cells, the percent-age of �-SMA� cells in G2/M remained constant at all timepoints examined (Table 2).
DiscussionThe phenotypic characterization of peripheral blood leuko-cytes has become a routine assay because of the ease, speed,and reproducibility of flow cytometric analysis of single cellsuspensions.15 However, there is a paucity of quantitative
Figure 3. Two-parameter representative histograms of CD45 vs side scatter of leukocytes from an injured (D through F) rat commoncarotid artery and its contralateral uninjured (A through C) vessel stained with 3 separate antibodies to detect T cells (CD3), granulo-cytes (HIS48 clone), and monocytes/macrophages (ED-1). Each leukocyte subpopulation cluster was defined in a 2D plot, assigned thered color, and plotted in that color on the CD45 dot plot (black). In the contralateral vessel, T cells (8.4%), granulocytes (16.7%), andmonocytes (38.8%) accounted for 64% of CD45� cells, whereas in the injured vessel, the same cell types combined represented�90% of infiltrating cells (2.7%, 49.1%, and 42.2%, respectively). Fold increases in relation to the contralateral vessel are printed inparentheses.
Hay et al Cell Cycle Patterns in Injured Arteries 1951
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from
analyses of infiltrating leukocytes recruited to injury sitesbecause of the inherent difficulties associated with the isola-tion of white blood cells from tissues. Leukocyte recruitmentis a fundamental aspect in the pathogenesis of injury-inducedvascular lesion formation. In the present work, leukocytesinfiltrating vascular tissue were readily identified and quan-tified in injured vascular tissue after enzymatic cell disaggre-gation. A 12-fold increase of CD45� cells in injured vessels1 day after injury in the absence of BrdU incorporationindicated that these cells were recruited from the circulation.The number of CD45� cells in contralateral uninjuredvessels was slightly higher than that in naive uninjuredarteries (4.1�0.2% versus 2.2�0.3%, respectively; P�0.05),suggesting that balloon dilation of 1 carotid artery induced acontralateral activation of arterial cells. The pathophysiolog-ical relevance of such activation remains to be determined,but it questions the use of contralateral vessels as controls.Approximately half of the CD45� cells in contralateralvessels were identified as ED-1� cells. Abs against specificantigens expressed by T cells, granulocytes, and monocyte/macrophages used in conjunction with CD45 revealed thatgranulocytes and macrophages were the predominant celltypes infiltrating balloon-injured arteries.
Immunohistochemical and morphological identification ofinfiltrating inflammatory cells revealed that at day 1 afterinjury, all leukocytes subtypes were located primarily within
the adventitial layer. Implantation of an endotoxin-soakedcotton thread in the adventitia of the rat femoral arteryresulted in the migration of leukocytes into the vessel wallfrom the luminal and the adventitial side.16 Macrophageinfiltration on the luminal surface of injured vessels and theexpression of nuclear factor-�B–regulated genes, VCAM-1and MCP-1, by SMCs have been described within 4 hoursafter injury of the rat carotid artery.17 We have recentlyreported that adventitial infiltration of macrophages andneutrophils in injured pig coronary arteries is accompanied byexpression of VCAM-1 and MCP-1 in endothelial cells ofadventitial vessels.18 Taken together, these data suggest thatadventitial entry of inflammatory cells to the vessel wall mayoccur either via the rich vasa vasorum network present inlarge vessels or via emigration through the external elasticlamina.
Cell homeostasis in the wall of injured arteries has beensuggested to be maintained by a balance of cell influx,survival, replication, and death.5 A cell cycle analysis of theinfiltrating leukocytes indicated that a significant proportionof cells entered S phase. Traditionally, the only cells thoughtto proliferate in expanding atherosclerotic and restenoticlesions were SMCs. However, replication of monocyte-derived macrophages has been demonstrated in human veingraft stenosis19 and atherosclerotic lesions of hypercholester-
Figure 4. CD45 and ED-1 immunohistochemical staining of ratcommon carotid arteries 1 day after balloon injury. Images ofrepresentative contralateral uninjured (A and B) and injured (Cthrough F) vessels are shown. Sections were stained with anti-CD45 (A, C, and E) or anti–ED-1 (B, D, and F) antibodies. Leu-kocytes recruited to the perivascular space of injured arteriesstained positively for CD45 (C and E). A high proportion of theinfiltrating cells were ED-1� (D and F). Neutrophils stained posi-tively for CD45 (E, arrowheads) and negatively for ED-1 (F,arrows). Original magnification �200 (A through D) and �1000(E and F).
Figure 5. Time-course analysis of apoptosis and BrdU incorpo-ration in CD45� cells (A) recruited to the injury site and�-SMA� cells (B) after carotid injury. Solid bars indicate per-centage of BrdU� positive cells; open bars, percentage of apo-ptotic cells. BrdU� and apoptotic cell subpopulations wereobtained for each artery from the same scatterplot of BrdU vsDNA content. A, Apoptosis of CD45� cells preceded cell prolif-eration, and it peaked at day 1. Cell proliferation was minimal innaive arteries and at day 1, but it increased significantly at day3 and gradually decreased at days 7 and 15 after injury. B, Likeinfiltrating leukocytes, �-SMA� cells showed a significantlyhigher percentage of apoptotic cells at day 1. BrdU incorpora-tion peaked at day 7. Results from 4 independent experimentsare expressed as mean�SEM. *P�0.05 compared with naiveuninjured arteries.
1952 Arterioscler Thromb Vasc Biol. December 2001
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from
olemic rabbits.20 In our experiments, the number of infiltrat-ing leukocytes in the hypodiploid phase indicates that asubstantial proportion entered programmed cell death, aspreviously reported by use of other techniques.21 Apoptosis ofleukocytes was found to precede cell proliferation; however,it is unknown whether apoptosis triggered cell proliferation orwhether it was an independent cellular event.
Although the highest rate of apoptosis was observed at day1 after injury for CD45� and �-SMA� cells, fewer�-SMA� cells underwent apoptosis (7.9�0.8% versus13.7�1.6% for CD45� cells), suggesting that leukocyteswere more susceptible to apoptosis. DNA content analysis issubjected to measurement interference by debris. The higherapoptotic index observed in SMA� cells of control arteriescompared with CD45� cells (5.0�0.7% versus 1.6�0.5%,respectively) might be the result of differential sensitivities ofthese cell types to the enzymatic digestion. Additionally,necrosis of the innermost arterial layers of SMCs afterballoon dilation might also contribute to the cellular debris inthe DNA histograms. We did not examine injured carotidarteries either before day 1 or after day 15. However, we andother investigators have previously shown that 2 weeks afterarterial injury, there was a significant increase in the numberof apoptotic SMCs in the neointima.3,22 In addition, an earlywave of apoptosis starting as early as 30 minutes after injuryand returning to control values at 4 hours has also been
described.23 Although different methodologies used in thesestudies prevent a conclusive analysis, the data taken as awhole suggest that deendothelialization of the rat commoncarotid artery is characterized by several spatially distinctapoptotic “waves” that take place a few minutes from the timeof injury to several weeks after full neointimal thickening.Furthermore, our data indicate that inflammatory cells makeup a significant proportion of the total number of cellsundergoing apoptosis through the first 24 hours.
Differences in the relative size of the G2/M fractionbetween quiescent CD45� and �-SMA� cells in controlarteries (1% versus 13%, respectively) appear to reflect theirspecific resting cell cycle profile. The high proportion ofquiescent SMCs in G2/M agrees with our previous data oncultured SMCs, in which 15% to 20% of growth-arrestedcells remain in G2/M.24 The functional significance of thisobservation is currently unknown. The considerable decreasein the G0/G1 fraction of CD45� and �-SMA� cells afterinjury suggests that the changes observed in the hypodiploidand S-phase fractions were due to the progression of growth-arrested cells through the cell cycle.
Proliferation of �-SMA� cells was highest at day 7,whereas the highest number of proliferating CD45� cellswas observed at day 3. Previous studies evaluating thekinetics of cell proliferation after vascular injury with the useof either [3H]thymidine25 or BrdU26 found that the prolifera-tion of carotid medial cells peaked at day 2 or 3, whereasneointimal cells peaked at day 4 or 5. Using proliferating cellnuclear antigen, BrdU, and [3H]thymidine autoradiography,Zeymer et al27 have shown that intimal proliferation washighest 7 days after denudation of rat aortas, when 40% ofcells were BrdU�. In contrast, maximal medial proliferationoccurred at day 3, when �5% of SMCs were proliferating. Inthe present study, no extrapolation to specific arterial layerscan be made because we evaluated cell cycle progression inwhole arteries. However, we hypothesize that the highestBrdU labeling index observed in �-SMA� cells 7 days afterendothelial denudation is largely due to neointimal prolifer-ation. A contribution of adventitial cells to the early prolif-erative response cannot be ignored. Our previous experimentsin rats indicate that adventitial proliferation, evidenced bypositive BrdU staining, peaked at day 3 after injury and wasresolved by day 14.28 Similar results have been reported in theporcine model of angioplasty.14,29 Because a single dose ofBrdU was administered 18 hours before the animals wereeuthanized, we may have underestimated the number of cellsundergoing DNA synthesis (assuming an S phase of �8hours).
The flow cytometry analysis reported in the present studyprovides a simple, fast, and reliable approach to characterize
TABLE 1. Number of Leukocyte Subtypes Detected in Balloon-Injured RatCommon Carotid Arteries 1 Day After Injury
n T Cells GranulocytesMonocytes/
Macrophages
Uninjured 4 34.3�11.4 38.8�12.3 275.5�52.2
Day 1 after injury 4 298.3�22.6 (8.7)* 1255.0�131.4 (32.3)* 1628.0�515.0 (5.9)*
n indicates number of arteries analyzed separately. Values are mean�SEM. Numbers betweenparentheses are fold increases.
*P�0.05 vs contralateral uninjured arteries.
TABLE 2. Cell Cycle Progression of Infiltrating (CD45�) andResident (�-SMA�) Cells After Balloon Angioplasty of RatCarotid Arteries
nS Phase,
%Hypodiploid,
%G0/G1,
%G2/M,
%
CD45� cells
Uninjured 4 0.0�0.0 1.6�0.5 97.4�0.6 1.0�0.4
After injury
1 day 4 0.1�0.0 13.7�1.6* 84.0�2.1* 2.4�0.5
3 day 4 16.9�2.6* 2.2�0.6 73.8�3.1* 7.1�0.5*
7 day 3 12.7�0.5* 5.3�1.1 77.6�0.3* 4.5�1.0*
15 day 4 3.7�1.5 3.0�0.9 93.3�0.5 3.7�0.5
�-SMA� cells
Uninjured 4 0.3�0.1 5.0�0.7 82.1�1.4 12.6�1.2
After injury
1 day 4 0.8�0.2 7.9�0.8* 76.7�0.5 14.6�1.2
3 day 4 6.7�1.5* 2.9�0.6 79.6�2.5 10.8�0.9
7 day 4 18.9�1.9* 4.9�0.5 63.8�1.0* 12.4�0.6
15 day 4 4.2�0.9 4.1�1.2 78.5�2.2 13.3�1.1
n indicates number of arteries analyzed separately. Values are mean�SEM.*P�0.05 vs naive uninjured arteries.
Hay et al Cell Cycle Patterns in Injured Arteries 1953
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from

the dynamic infiltration of leukocyte subtypes as well as theircell cycle progression in injured vascular tissues. The currentavailability of monoclonal Abs against a wide array ofcytokines and chemokines would also allow for the study ofthe cell-specific expression of these factors on the multipleresident and infiltrating cell types involved in vascularinflammation under various pathological conditions.
AcknowledgmentThis work was partially supported by a grant from the RichFoundation.
References1. Majesky MW, Giachelli CM, Reidy MA, Schwartz SM. Rat carotid
neointimal smooth muscle cells reexpress a developmentally regulatedmRNA phenotype during repair of arterial injury. Circ Res. 1992;71:759–768.
2. Schwartz SM, Majesky MW, Murry CE. The intima: development andmonoclonal responses to injury. Atherosclerosis. 1995;118(suppl):S125–S140.
3. Han DK, Haudenschild CC, Hong MK, Tinkle BT, Leon MB, Liau G.Evidence for apoptosis in human atherogenesis and in a rat vascularinjury model. Am J Pathol. 1995;147:267–277.
4. Jackson CL, Schwartz SM. Pharmacology of smooth muscle cell repli-cation. Hypertension. 1992;20:713–736.
5. Ross R. Atherosclerosis is an inflammatory disease. Am Heart J. 1999;138:S419–S420.
6. Mukaida N, Harada A, Matsushima K. Interleukin-8 (IL-8) and monocytechemotactic and activating factor (MCAF/MCP-1), chemokines essen-tially involved in inflammatory and immune reactions. Cytokine GrowthFactor Rev. 1998;9:9–23.
7. Printseva OY, Peclo MM, Gown AM. Various cell types in humanatherosclerotic lesions express ICAM-1: further immunocytochemicaland immunochemical studies employing monoclonal antibody 10F3.Am J Pathol. 1992;140:889–896.
8. Bevilacqua MP. Endothelial-leukocyte adhesion molecules. Annu RevImmunol. 1993;11:767–804.
9. Yla-Herttuala S, Lipton BA, Rosenfeld ME, Sarkioja T, Yoshimura T,Leonard EJ, Witztum JL, Steinberg D. Expression of monocyte chemoat-tractant protein 1 in macrophage-rich areas of human and rabbit athero-sclerotic lesions. Proc Natl Acad Sci U S A. 1991;88:5252–5256.
10. Wysocki SJ, Zheng MH, Smith A, Lamawansa MD, Iacopetta BJ, RobertsonTA, Papadimitriou JM, House AK, Norman PE. Monocyte chemoattractantprotein-1 gene expression in injured pig artery coincides with earlyappearance of infiltrating monocyte/macrophages. J Cell Biochem. 1996;62:303–313.
11. Wang X, Li X, Yue TL, Ohlstein EH. Expression of monocyte chemo-tactic protein-3 mRNA in rat vascular smooth muscle cells and in carotidartery after balloon angioplasty. Biochim Biophys Acta. 2000;1500:41–48.
12. Furukawa Y, Matsumori A, Ohashi N, Shioi T, Ono K, Harada A,Matsushima K, Sasayama S. Anti-monocyte chemoattractant protein-1/
monocyte chemotactic and activating factor antibody inhibits neointimalhyperplasia in injured rat carotid arteries. Circ Res. 1999;84:306–314.
13. Clowes AW, Reidy MA, Clowes MM. Mechanisms of stenosis afterarterial injury. Lab Invest. 1983;49:208–215.
14. Scott NA, Cipolla GD, Ross CE, Dunn B, Martin FH, Simonet L, WilcoxJN. Identification of a potential role for the adventitia in vascular lesionformation after balloon overstretch injury of porcine coronary arteries.Circulation. 1996;93:2178–2187.
15. Shapiro, HM. Practical Flow Cytometry. New York, NY: Wiley-Liss Inc;2000.
16. Prescott MF, McBride CK, Court M. Development of intimal lesions afterleukocyte migration into the vascular wall. Am J Pathol. 1989;135:835–846.
17. Landry DB, Couper LL, Bryant SR, Lindner V. Activation of theNF-kappa B and I kappa B system in smooth muscle cells after rat arterialinjury: induction of vascular cell adhesion molecule-1 and monocytechemoattractant protein-1. Am J Pathol. 1997;151:1085–1095.
18. Okamoto E, Bennet T, De Leon H, Vinten-Johansen J, Goodman RB,Scott NA, Wilcox JN. Perivascular inflammation after balloon angio-plasty of porcine coronary arteries. Circulation. 2001;104:2228–2235.
19. Westerband A, Mills JL, Marek JM, Heimark RL, Hunter GC, WilliamsSK. Immunocytochemical determination of cell type and proliferationrate in human vein graft stenoses. J Vasc Surg. 1997;25:64–73.
20. Rosenfeld ME, Ross R. Macrophage and smooth muscle cell proliferationin atherosclerotic lesions of WHHL and comparably hypercholesterol-emic fat-fed rabbits. Arteriosclerosis. 1990;10:680–687.
21. Niebauer J, Schwarzacher SP, Hayase M, Wang B, Kernoff RS, CookeJP, Yeung AC. Local L-arginine delivery after balloon angioplastyreduces monocyte binding and induces apoptosis. Circulation. 1999;100:1830–1835.
22. Bochaton-Piallat ML, Gabbiani F, Redard M, Desmouliere A, GabbianiG. Apoptosis participates in cellularity regulation during rat aortic intimalthickening. Am J Pathol. 1995;146:1059–1064.
23. Perlman H, Maillard L, Krasinski K, Walsh K. Evidence for the rapidonset of apoptosis in medial smooth muscle cells after balloon injury.Circulation. 1997;95:981–987.
24. Liau G, Janat MF, Wirth PJ. Regulation of alpha-smooth muscle actin andother polypeptides in proliferating and density-arrested vascular smoothmuscle cells. J Cell Physiol. 1990;142:236–246.
25. Clowes AW, Reidy MA, Clowes MM. Kinetics of cellular proliferationafter arterial injury, I: smooth muscle growth in the absence of endothe-lium. Lab Invest. 1983;49:327–333.
26. London SM, Mayberg MR. Kinetics of bromodeoxyuridine uptake bysmooth muscle cells after arterial injury. J Vasc Res. 1994;31:247–255.
27. Zeymer U, Fishbein MC, Forrester JS, Cercek B. Proliferating cellnuclear antigen immunohistochemistry in rat aorta after balloon denu-dation: comparison with thymidine and bromodeoxyuridine labeling.Am J Pathol. 1992;141:685–690.
28. De Leon H, Ollerenshaw JD, Griendling KK, Wilcox JN. Adventitialcells do not contribute to the neointimal mass after balloon angioplasty ofthe rat common carotid artery. Circulation. 2001;104:1591–1593.
29. Shi Y, O’Brien JE, Fard A, Mannion JD, Wang D, Zalewski A. Adven-titial myofibroblasts contribute to neointimal formation in injured porcinecoronary arteries. Circulation. 1996;94:1655–1664.
1954 Arterioscler Thromb Vasc Biol. December 2001
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from

LeonCarl Hay, Connie Micko, Margaret Forney Prescott, Gene Liau, Keith Robinson and Hector De
After Balloon Injury of the Rat Carotid ArteryDifferential Cell Cycle Progression Patterns of Infiltrating Leukocytes and Resident Cells
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2001 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/hq1201.1002562001;21:1948-1954Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/21/12/1948World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. AnswerPermissions and Rights Question andunder Services. Further information about this process is available in the
permission is being requested is located, click Request Permissions in the middle column of the Web page whichCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on January 26, 2016http://atvb.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















