
Defining Octopus vulgaris populations: A comparative study of the morphology and chromatophore...
10
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopus vulgaris populations: A comparative study of the morphology and chromatophore pattern of paralarvae from Northeastern and Southwestern Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009 ARTICLE IN PRESS G Model FISH-3000; No. of Pages 10 Fisheries Research xxx (2010) xxx–xxx Contents lists available at ScienceDirect Fisheries Research journal homepage: www.elsevier.com/locate/fishres Defining Octopus vulgaris populations: A comparative study of the morphology and chromatophore pattern of paralarvae from Northeastern and Southwestern Atlantic Érica A.G. Vidal a,∗ , Lidia Fuentes b , Lisiane B. da Silva c a Centro de Estudos do Mar, Universidade Federal do Paraná - UFPR, Cx. Postal 50.002, Pontal do Paraná, PR 83255-000, Brazil b Centro Oceanográfico de Vigo, Instituto Espa˜ nol de Oceanografía, Cabo Estay, Canido, Apartado 1552, 36280 Vigo, Spain c Curso de Pós-Graduac ¸ ão em Aquicultura, Fundac ¸ ão Universidade Federal do Rio Grande, Cx. P. 474, Rio Grande, RS 96.201-900, Brazil article info Article history: Received 7 June 2010 Received in revised form 18 August 2010 Accepted 19 August 2010 Keywords: Chromatophore Cryptic Octopus vulgaris Paralarvae Populations abstract The chromatophore pattern of cephalopods provides a rapid and relatively easy means to identify species. Therefore, this paper describes and compares the morphology and chromatophore pattern of Octopus vul- garis paralarvae from different geographic regions, specifically, the notheastern (Galicia, Spain) and the southwestern (Southern Brazil), Atlantic, and ultimately, contributes to knowledge for future discrimi- nation of populations of O. vulgaris from both regions. The eggs and paralarvae of both localities were incubated in the laboratory and reared at 21 and 22 ◦ C from hatching to 17 or 45 days of age. Important differences were observed on the chromatophores pattern of paralarvae from both regions. In the ventral mantle, between 23 and 36 chromatophores were found in the Brazilian paralarvae, whereas between 16 and 20 were found in the Spanish paralarvae. However, Brazilian paralarvae showed considerably less extrategumental chromatophores over the viscera (6–9) in the dorsal mantle than the Spanish paralar- vae (6–20). Differences were also detected in the funnel, 4–6 chromatophores (four different patterns) versus 4 (single pattern), respectively. These results showed substantial differences between the chro- matophore patterns of paralarvae from both regions, which could provide evidence for the existence of distinct populations or even cryptic O. vulgaris-like species along the Southern Brazilian coast. © 2010 Elsevier B.V. All rights reserved. 1. Introduction The common octopus, Octopus vulgaris Cuvier, 1797 is one of world’s most important commercial marine resources and is sub- ject to active fisheries. The considerable interannual fluctuations in catches are largely influenced by environmental factors (Otero et al., 2008, 2009). Due to its biological characteristics, substan- tial market and high value, the species constitutes a potential new candidate for aquaculture (Vaz-Pires et al., 2004; Iglesias et al., 2007). This species is also one of the best-studied cephalopods that has long been considered cosmopolitan in temperate and tropi- cal seas with unknown distributional limits (Roper et al., 1984). More recently, the wide distribution has become quite controver- sial, with some studies suggesting that O. vulgaris may be restricted to the Mediterranean and the Eastern Atlantic (Mangold, 1997; Mangold and Hochberg, 1991), and others imply that O. vulgaris ∗ Corresponding author. Tel.: +55 41 3511 8600; fax: +55 41 3511 8635. E-mail address: [email protected] (E.A.G. Vidal). is a complex of cryptic species among O. vulgaris-like octopods (Norman, 2003). Genetic studies on the DNA sequences of the Octopus from the Southern Hemisphere using the mitochondrial cytochrome oxidase III gene pointed towards a wide distributional range for O. vulgaris, including specimens collected from Venezuela, Isla Margarita and Central (Rio de Janeiro) and Southern Brazil (Warnke et al., 2004). Teske et al. (2007) found that the species was also present on both coasts of South Africa (Atlantic and Indian), but two specimens from Durban (Indian Ocean) were genetically so different that they could represent an undescribed species. To address this problem, those authors suggested that further sampling was needed in regions for which no genetic data were yet available, such as east Africa, India and southeast Asia. Furthermore, later molecular studies have shown that octopus specimens from Saint Paul and Amsterdam Islands (Southern Indian Ocean), are O. vulgaris sensu Cuvier, 1797 or sensu stricto, strengthening the evidence that this species has a wide geographic distribution, which until present, comprises the Mediterranean Sea, the Eastern Atlantic (from Southern England to southwestern Africa), the Azores, the Canary Islands, Cape Verde, St Helena, the Tristan da Cunha Island, the southeast coast of South Africa in the Indian Ocean, the oceanic islands of the central South- 0165-7836/$ – see front matter © 2010 Elsevier B.V. All rights reserved. doi:10.1016/j.fishres.2010.08.009
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Defining Octopus vulgaris populations: A comparative study of the morphology and chromatophore...

G
F
DmN
Éa
b
c
a
ARRA
KCCOPP
1
wjietc2hcMstM
0d
ARTICLE IN PRESSModel
ISH-3000; No. of Pages 10
Fisheries Research xxx (2010) xxx–xxx
Contents lists available at ScienceDirect
Fisheries Research
journa l homepage: www.e lsev ier .com/ locate / f i shres
efining Octopus vulgaris populations: A comparative study of theorphology and chromatophore pattern of paralarvae fromortheastern and Southwestern Atlantic
rica A.G. Vidala,∗, Lidia Fuentesb, Lisiane B. da Silvac
Centro de Estudos do Mar, Universidade Federal do Paraná - UFPR, Cx. Postal 50.002, Pontal do Paraná, PR 83255-000, BrazilCentro Oceanográfico de Vigo, Instituto Espanol de Oceanografía, Cabo Estay, Canido, Apartado 1552, 36280 Vigo, SpainCurso de Pós-Graduacão em Aquicultura, Fundacão Universidade Federal do Rio Grande, Cx. P. 474, Rio Grande, RS 96.201-900, Brazil
r t i c l e i n f o
rticle history:eceived 7 June 2010eceived in revised form 18 August 2010ccepted 19 August 2010
eywords:hromatophoreryptic
a b s t r a c t
The chromatophore pattern of cephalopods provides a rapid and relatively easy means to identify species.Therefore, this paper describes and compares the morphology and chromatophore pattern of Octopus vul-garis paralarvae from different geographic regions, specifically, the notheastern (Galicia, Spain) and thesouthwestern (Southern Brazil), Atlantic, and ultimately, contributes to knowledge for future discrimi-nation of populations of O. vulgaris from both regions. The eggs and paralarvae of both localities wereincubated in the laboratory and reared at 21 and 22 ◦C from hatching to 17 or 45 days of age. Importantdifferences were observed on the chromatophores pattern of paralarvae from both regions. In the ventral
ctopus vulgarisaralarvaeopulations
mantle, between 23 and 36 chromatophores were found in the Brazilian paralarvae, whereas between16 and 20 were found in the Spanish paralarvae. However, Brazilian paralarvae showed considerably lessextrategumental chromatophores over the viscera (6–9) in the dorsal mantle than the Spanish paralar-vae (6–20). Differences were also detected in the funnel, 4–6 chromatophores (four different patterns)versus 4 (single pattern), respectively. These results showed substantial differences between the chro-matophore patterns of paralarvae from both regions, which could provide evidence for the existence of
en cr
distinct populations or ev. Introduction
The common octopus, Octopus vulgaris Cuvier, 1797 is one oforld’s most important commercial marine resources and is sub-
ect to active fisheries. The considerable interannual fluctuationsn catches are largely influenced by environmental factors (Oterot al., 2008, 2009). Due to its biological characteristics, substan-ial market and high value, the species constitutes a potential newandidate for aquaculture (Vaz-Pires et al., 2004; Iglesias et al.,007). This species is also one of the best-studied cephalopods thatas long been considered cosmopolitan in temperate and tropi-
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
al seas with unknown distributional limits (Roper et al., 1984).ore recently, the wide distribution has become quite controver-
ial, with some studies suggesting that O. vulgaris may be restrictedo the Mediterranean and the Eastern Atlantic (Mangold, 1997;
angold and Hochberg, 1991), and others imply that O. vulgaris
∗ Corresponding author. Tel.: +55 41 3511 8600; fax: +55 41 3511 8635.E-mail address: [email protected] (E.A.G. Vidal).
165-7836/$ – see front matter © 2010 Elsevier B.V. All rights reserved.oi:10.1016/j.fishres.2010.08.009
yptic O. vulgaris-like species along the Southern Brazilian coast.© 2010 Elsevier B.V. All rights reserved.
is a complex of cryptic species among O. vulgaris-like octopods(Norman, 2003).
Genetic studies on the DNA sequences of the Octopus from theSouthern Hemisphere using the mitochondrial cytochrome oxidaseIII gene pointed towards a wide distributional range for O. vulgaris,including specimens collected from Venezuela, Isla Margarita andCentral (Rio de Janeiro) and Southern Brazil (Warnke et al., 2004).Teske et al. (2007) found that the species was also present on bothcoasts of South Africa (Atlantic and Indian), but two specimens fromDurban (Indian Ocean) were genetically so different that they couldrepresent an undescribed species. To address this problem, thoseauthors suggested that further sampling was needed in regionsfor which no genetic data were yet available, such as east Africa,India and southeast Asia. Furthermore, later molecular studies haveshown that octopus specimens from Saint Paul and AmsterdamIslands (Southern Indian Ocean), are O. vulgaris sensu Cuvier, 1797or sensu stricto, strengthening the evidence that this species has a
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
wide geographic distribution, which until present, comprises theMediterranean Sea, the Eastern Atlantic (from Southern England tosouthwestern Africa), the Azores, the Canary Islands, Cape Verde,St Helena, the Tristan da Cunha Island, the southeast coast of SouthAfrica in the Indian Ocean, the oceanic islands of the central South-

ING
F
2 es Rese
eTsic
tsBlS(tvMvnwmfinlO
scmtrmIsii1
dalctcimoeaa(
aenamae
cefspfs
ARTICLEModel
ISH-3000; No. of Pages 10
E.A.G. Vidal et al. / Fisheri
rn Indian Ocean, and the northwestern Pacific, that is, the waters ofaiwan and Japan (Guerra et al., 2010). Those authors also empha-ized the need to clarify the geographical range of O. vulgaris andts phylogenetic relationship to other species of the O. vulgaris-likeomplex.
In relation to the Brazilian coast, no correlation linking genetico geographical distances could be established between the almostympatric O. vulgaris-like specimens from Southern and Northernrazil (Söller et al., 2000). Additionally, the genotype of O. vulgaris-
ike specimens collected in northeastern Brazil (Recife, Pernanbucotate) differed from that of Southern Brazil and the MediterraneanWarnke, 1999). It is of note that a relatively small genetic dis-ance was found between Mediterranean O. vulgaris and the O.ulgaris-like individuals from Southern Brazil (Söller et al., 2000).ore recently, a new species, Octopus insularis, belonging to the O.
ulgaris-like complex was recognized as a separate species fromortheastern Brazil, confirming that the specimens from Recifeere in fact genetically similar to O. insularis (Leite et al., 2008). Thisedium-sized species appears to be the most common octopus
ound in shallow waters along the mainland shelf and in the oceanicslands of northeastern Brazil. This new discovery underlines theeed to undertake comprehensive and integrated morphological,
ife history and molecular studies to review the systematics of thectopodinae.
Information on the morphology and distribution of youngtages of cephalopods could be of potential importance for dis-rimination of populations and identifying species. Although theorphological differentiation of young stages is often difficult,
he chromatophore patterns are conservative and consideredeliable taxonomic features, as they provide a rapid and easyethod to identify paralarval cephalopods (Young et al., 1989).
n fact, the number and distribution of chromatophores on thekin of paralarvae are species-specific and, when combined withridophore pattern, body dimensions and arm length formulae,s a reliable tool used in species recognition (Hochberg et al.,992).
In Octopodidae, founder chromatophores are the first to appearuring late embryogenesis in a fixed sequence. They are the largestnd the deepest in the body of hatchlings and are still visible inate planktonic stages and post-settlement juveniles. As a result,hromatophores appearing later in development are smaller thanhose already established and the initial spatial arrangement ofhromatophores influences all subsequent arrangements, resultingn spatio-temporal patterns (Packard, 1985). In addition, extrategu-
ental chromatophores appear in the connective tissue at the basef the arms, covering the dorsal and ventral surfaces of the brain andyes and overlying the visceral mass. Tegumental chromatophoresre located in the epithelium covering the surface of the mantle andre often smaller and more conspicuous for species identificationPackard, 1985).
O. vulgaris paralarvae have been described by various authorsnd a review of the available information is found in Hochbergt al. (1992). Nonetheless, the critical chromatophore pattern hasever been fully described until now. Laboratory studies provideunique opportunity to observe the continuous development oforphological features from hatchlings to juveniles obtained from
dults of known locality and taxonomic status and under controllednvironmental conditions.
The aim of this study was to describe the morphology and thehromatophore pattern of O. vulgaris paralarvae from the north-astern (Galicia, Spain) and southwestern (Southern Brazil) Atlantic
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
rom hatching to the age of 45 and 17 days, respectively. In doingo, we attempt to contribute with taxonomic features of O. vulgarisaralarvae from both regions that could broaden knowledge foruture discrimination of populations or cryptic speciation of thispecies.
PRESSarch xxx (2010) xxx–xxx
2. Materials and methods
2.1. Broodstook information
The broodstock consisted of octopus males and females (ratio1:2) captured at sea in the northeastern (Vigo – Galicia, Spain)(42◦15′04′ ′N, 8◦53′30′ ′W) and the southwestern (Bombinhas –Santa Catarina, Brazil) (27◦07′54′ ′S, 48◦31′40′ ′W) Atlantic andmaintained in captivity. They were fed a diet consisting of 80%crustaceans and 20% fish. PVC pipes of 40 cm in length and 20 cmin diameter were placed into the broodstock tanks as shelters forspawning females. When a female started to spawn it was isolatedin a smaller tank until hatching of paralarvae.
2.2. Paralarval rearing
Paralarvae from Spain were reared in August 2005 in one 1000 Ltank, with an initial density of 0.5 individuals L−1 at a mean tem-perature of 22.74 ± 0.50 ◦C, salinity between 32 and 34 and exposedto 24 h light. They were fed with 5–6 days old Artemia sp. enrichedwith microalgae (Isochrysis galbana and Nannochloropsis sp.) andsporadically with crab zoeae (Maja brachydactyla).
Paralarvae from Brazil were obtained from two broodingfemales and reared between February and March 2004 in six 90 Ltanks, at an initial density of 0.4 individuals L−1, a mean temper-ature of 21 ± 2 ◦C, salinity between 32 and 34 and photoperiodof 10 h light:14 h dark. They were fed with Artemia sp. naupliienriched with SUPER SELCO (INVE®), copepods (Acartia tonsa,Temora turbinata, Oncaea spp. and Paracalanus sp.), crab zoeae(Callinectes sapidus, Cyrtograpsus angulatus) and juvenile mysids(Metamysidopsis elongata atlantica).
2.3. Description of the morphology and the chromatophorepattern of paralarvae
Descriptions of paralarvae and their chromatophore patternwere based on samples of 5–10 paralarvae each taken from therearing tanks on 0, 1, 6, 10, 13, 15, 17 and 20 days after hatching,approximately 60 paralarvae from each region. Additionally, few(2–3) paralarvae of 27, 36 and 45 days old from Spain were alsodescribed. Live paralarvae were sampled from the rearing tankswith a beaker and anaesthetized with magnesium chloride (75 mgMgCl in 1 L of distilled water; 1:2 diluted with seawater) beforeimages were taken.
In Spain, images from each paralarvae were taken in both dor-sal and ventral views and occasionally in the lateral view andrecorded through a Leica MZ 8 dissecting microscope at 25, 50,64, 80 and100× magnification with the ocular replaced by a LeicaICA digital camera. This procedure allowed the observation of onlycontracted chromatophores; no preserved material was used. InBrazil, images of paralarvae were taken in dorsal, ventral andlateral views and recorded through an Olympus SZX-9 dissect-ing microscope at 10, 20 and 50× magnification with the ocularreplaced by a SONY DXC- 197 camera connected to a computer.Immediately after images were taken, Brazilian paralarvae werepreserved in buffered 4–6% seawater formalin for observation ofsemi-extended cromatophores in the following weeks. Problemscaused by abrasion of the tips of the arms due to friction againstthe tank walls prevented complete observation of the number ofsuckers and the chromatophore pattern of the arms of Brazilianparalarvae.
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
Measurements of several body dimensions such as the man-tle length (ML), the mantle width (MW), the head width (HW)and the funnel length (FuL) were obtained by image analysisperformed using NIH-Image public domain software (1.61 ver.) fol-lowing Roper and Voss (1983). The body dimensions measured

IN PRESSG
F
es Research xxx (2010) xxx–xxx 3
wdrMM%wr1
3
3
mmaavtwmrdtaamrIdfm
3A
T1aWhoAeCtHattnMnvpTweoccb
uck
ers
per
arm
and
chro
mat
oph
ore
pat
tern
ofO
.vul
gari
sp
aral
arva
efr
omth
en
orth
east
ern
Atl
anti
c(S
pai
n).
Age
(day
s)Su
cker
sp
erar
mC
hro
mat
oph
ores
dor
salv
iew
Tota
lC
hro
mat
oph
ores
ven
tral
view
Tota
l
Arm
sH
ead
Eyes
Man
tle
edge
Man
tle
Man
tle
Tota
lA
rmH
ead
Eyes
Man
tle
Fun
nel
03
2–4
(1ro
w)
2+
2+
2+
21
2-3
R8-
9(6
-7V
S)La
rge
10-1
221
-25
2–3
(1ro
w)
2+
21
R16
–19B
2+
227
–31
13
3–4
(1ro
w)
2+
2+
2+
21
2–3
R8–
10(6
–7V
S)10
–13
22–2
62–
3(1
row
)2
+2
1R
16–1
92
+2
27–3
16
3–4
3–4
(1ro
w)
2+
2+
2+
21
2–3
R8–
10(6
–7V
S)10
–13
22–2
63–
4(1
row
)2
+2
1R
16–1
92
+2
28–3
210
3–6
3–4
(1ro
w)
2+
2+
2+
21
2–3
R8–
10(6
–7V
S)10
–13
22–2
63–
4(1
row
)2
+2
1R
16–2
02
+2
28–3
315
4–8
3–9
(1+
2ro
ws)
2+
2+
2+
21
2–4
8–19
(6–1
7V
S)10
–23
22–4
13–
6(1
+2
row
s)2
+2
1R
16–2
02
+2
28–3
517
5–9
5–9
(1+
2ro
ws)
2+
2+
2+
1R+
2+
21
2–4
13–2
2(1
0–20
VS)
15–2
532
–47
4–7(
1+
2ro
ws)
2+
2+
21
R16
–20
2+
231
–38
205–
105–
13(1
+2
row
s)2+
2+
2+
2+
1R+
2+
21
2–4
13–2
6(1
1–22
VS)
15–3
034
–57
4–11
(1+
2ro
ws)
2+
2+
21
R16
–20
2+
231
–42
277–
147–
13(1
+2
row
s)2
+2
+2
+2
+8
3∼2
5–34
(in
crea
sin
g)∼5
1–66
7–11
(1+
2ro
ws)
2+
2+
21
18–2
42
+2
∼36–
46
3610
–20
10–1
5(1
+2
row
s)2
+2
+2
+2
+10
(in
crea
sin
g)3
∼32–
40(i
ncr
easi
ng)
∼63–
7611
–14
(1+
2ro
ws)
2+
2+
21
∼22–
24(i
ncr
easi
ng)
2+
2∼4
4–49
45∼2
7∼2
0(1
+2
row
s)2
+2
+2
+2
+20
(in
crea
sin
g)3
∼46
(in
crea
sin
g)∼9
7∼1
7(1
+2
row
s)2
+2
+2
1∼3
3(i
ncr
easi
ng)
2+
2∼6
1
th;
B:
blac
k;R
:re
d;
VS:
visc
eral
.Hea
dm
ean
sh
ead
wit
hou
tey
es.R
igh
tan
dle
ftey
ear
eal
way
sid
enti
cal.
Som
etim
esre
dco
lou
rw
asre
late
dto
inci
pie
nt
chro
mat
oph
ores
.
ARTICLEModel
ISH-3000; No. of Pages 10
E.A.G. Vidal et al. / Fisheri
ere compared between regions at each sampling age using Stu-ent’s t-tests (Zar, 1996). Growth rates of paralarvae from eachegion were calculated using the standard exponential function:L = ML0 ebA, where ML is mantle length in mm, MLo is the initialL, e is the natural logarithm, b is the instantaneous growth rate inbody ML d−1 and, A is age in days post-hatching. Mantle lengthsere log-transformed and plotted against age and differences in
egression slopes between localities were tested by ANCOVA (Zar,996).
. Results
.1. General considerations to the description of paralarvae
Paralarvae from both localities were compared based on theirean ML. The chromatophore pattern and the definition of chro-atophore fields were based on the terminology of Hochberg et
l. (1992). Thus, descriptions of the ventral view have taken intoccount arms, head, funnel and mantle, and those on the dorsaliew, besides arms, head and eye, also a new field was defined ashe mantle edge to include those chromatophores around this area,hich were sometimes difficult to assign to either the head or theantle. The pattern was described from the anterior to the poste-
ior parts of the body. For example, in the funnel, the pattern usuallyescribed is 2 + 2, that means there are two chromatophores closeo the orifice and other two close to the base of the funnel. Rightnd left eye are always identical. When variability was observedmong individuals, the most frequent number and pattern of chro-atophores was listed. The sucker counts per arm were based on
anges from the minimum to the maximum number observed.n general, the colour of the chromatophores did not changeuring development so the colour pattern was only describedor newly-hatched paralarvae. Measurements indicated are
eans ± SD.
.2. Description of O. vulgaris paralarvae from northeasterntlantic (Galicia, Spain)
2.18 ± 0.04 mm ML (newly hatched and 1-day old) (Fig. 1a,ables 1 and 2). Mantle: Globose with an average width of.46 ± 0.03 mm, representing 67.0% of ML. Head: Squarish, aver-ge width of 1.15 ± 0.05 mm and representing 52.7% of ML. Funnel:ell developed and occupies almost the entire length of the
ead, reaching the base of the fourth pair of arms, average lengthf 0.96 ± 0.01 mm. Eyes: Conspicuous and slightly ventral. Arms:pproximately of same length with attenuated tips and three suck-rs each. The length of the arm I right represents 47.7% of the ML.hromatophores: Arms: In the 1st and 2nd pairs from 2 to 4; inhe 3rd and 4th pairs from 2 to 3, all arranged in a single row.ead: Dorsally eight, two between the eyes midhead, other twot the basis of the eyes, and four covered by the mantle edge. Ven-rally four, two behind the funnel and other two near the base ofhe arms. Eyes: One posterodorsally and one on midventral. Fun-el: Four, two near the orifice and two adjacent to the mantle edge.antle: Dorsally from 10 to 12, 2 to 3 on the mantle edge. Under-
eath the mantle and above the digestive gland there are 6–7 largeisceral chromatophores distributed in oval form, and two on theosterior cap. Ventrally, 16–19 randomly distributed. Colouration:he colour depends on the expansion level of the elastic sacculushere the granules of pigments are contained. However, incipi-
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopus vulgaris populations: A comparative study of the morphology andchromatophore pattern of paralarvae from Northeastern and Southwestern Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
nt chromatophores are reddish and then become darker, brownr black. This was clearly observed on the arm tips. Contractedhromatophore colours: Mantle: Dorsally, slightly red; visceralhromatophores are black or quite dark. Ventrally, black. Head: Inoth dorsal and ventral view they are black. Funnel: Black. Eyes: Ta
ble
1M
ean
ML,
age,
s
Mea
nM
L±
SD(m
m)
2.18
±0.
042.
18±
0.05
2.47
±0.
093.
18±
0.16
3.61
±0.
183.
76±
0.18
4.07
±0.
144.
43±
0.30
5.31
±0.
29
6.60
±0.
00
ML:
man
tle
len
g

Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopus vulgaris populations: A comparative study of the morphology andchromatophore pattern of paralarvae from Northeastern and Southwestern Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
ARTICLE IN PRESSG Model
FISH-3000; No. of Pages 10
4 E.A.G. Vidal et al. / Fisheries Research xxx (2010) xxx–xxx
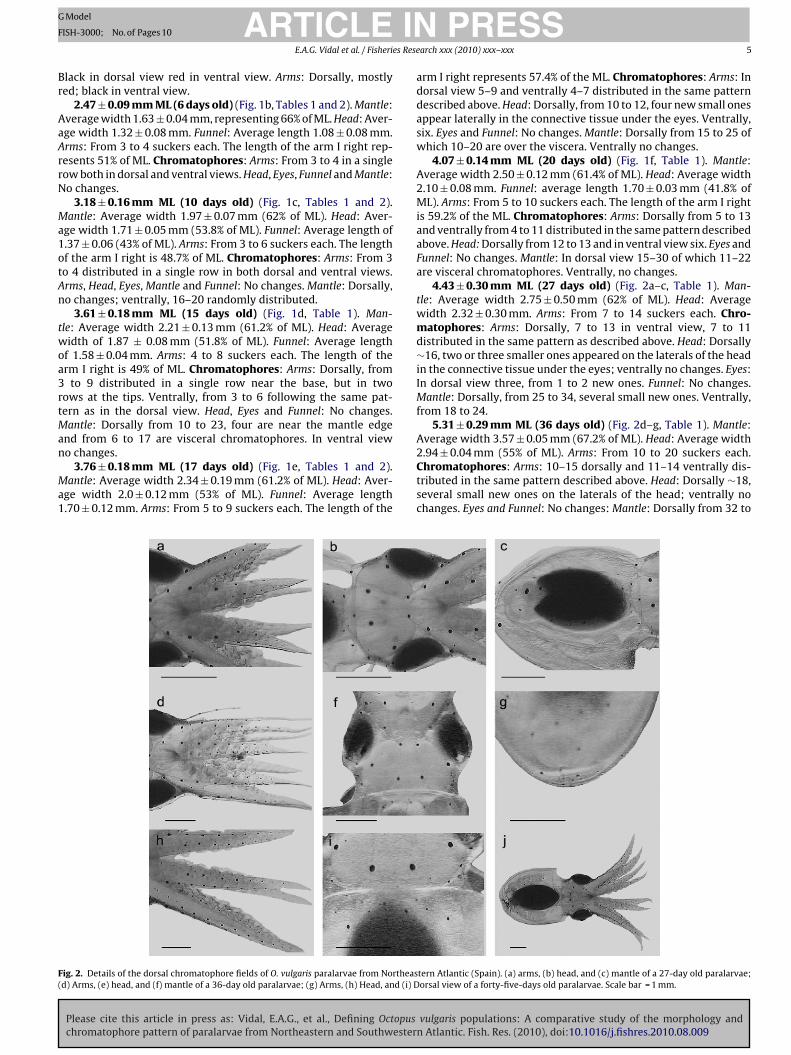
Fig. 1. Chromatophore development, arrangement and pattering in paralarvae of Octopus vulgaris from the northeastern Atlantic (Spain). (a) Newly-hatched to 1-day old;(b) 6-day old; (c) 10-day old; (d) 15-day old; (e) 17-day old and (f) 20-day old. Scale bar = 0.5 mm.
Table 2Measurements of O. vulgaris paralarvae from the northeastern and southwestern Atlantic maintained in the laboratory for 17 days after hatching. Values are means ± SD.
Age (days) Northeastern Atlantic (Spain) Southwestern Atlantic (Southern Brazil)
ML MW HW FuL ML MW HW FuL
0 2.18 ± 0.04 1.46 ± 0.03 1.15 ± 0.05 0.96 ± 0.01 2.21 ± 0.09 1.28 ± 0.05 0.96 ± 0.04 0.92 ± 0.036 2.47 ± 0.09 1.63 ± 0.04 1.32 ± 0.08 1.08 ± 0.08 2.40 ± 0.08 1.47 ± 0.11 1.10 ± 0.12 0.97 ± 0.0510 3.18 ± 0.16 1.97 ± 0.07 1.71 ± 0.05 1.37 ± 0.06 2.80 ± 0.18 1.81 ± 0.13 1.62 ± 0.16 1.18 ± 0.0917 3.76 ± 0.18 2.34 ± 0.19 2.00 ± 0.12 1.70 ± 0.12 3.38 ± 0.34 2.11 ± 0.15 1.87 ± 0.19 1.54 ± 0.13
ML: mantle length; MW: mantle width; HW: Head width; FuL: funnel length. No statistical difference was found (P > 0.05. t-test) when body dimensions of same age paralarvaefrom northeastern and southwestern Atlantic were compared.
ING
F
es Rese
Br
AaArrN
Ma1otAn
twoa3rtMan
Ma1
F(
ARTICLEModel
ISH-3000; No. of Pages 10
E.A.G. Vidal et al. / Fisheri
lack in dorsal view red in ventral view. Arms: Dorsally, mostlyed; black in ventral view.
2.47 ± 0.09 mm ML (6 days old) (Fig. 1b, Tables 1 and 2). Mantle:verage width 1.63 ± 0.04 mm, representing 66% of ML. Head: Aver-ge width 1.32 ± 0.08 mm. Funnel: Average length 1.08 ± 0.08 mm.rms: From 3 to 4 suckers each. The length of the arm I right rep-esents 51% of ML. Chromatophores: Arms: From 3 to 4 in a singleow both in dorsal and ventral views. Head, Eyes, Funnel and Mantle:o changes.
3.18 ± 0.16 mm ML (10 days old) (Fig. 1c, Tables 1 and 2).antle: Average width 1.97 ± 0.07 mm (62% of ML). Head: Aver-
ge width 1.71 ± 0.05 mm (53.8% of ML). Funnel: Average length of.37 ± 0.06 (43% of ML). Arms: From 3 to 6 suckers each. The lengthf the arm I right is 48.7% of ML. Chromatophores: Arms: From 3o 4 distributed in a single row in both dorsal and ventral views.rms, Head, Eyes, Mantle and Funnel: No changes. Mantle: Dorsally,o changes; ventrally, 16–20 randomly distributed.
3.61 ± 0.18 mm ML (15 days old) (Fig. 1d, Table 1). Man-le: Average width 2.21 ± 0.13 mm (61.2% of ML). Head: Averageidth of 1.87 ± 0.08 mm (51.8% of ML). Funnel: Average length
f 1.58 ± 0.04 mm. Arms: 4 to 8 suckers each. The length of therm I right is 49% of ML. Chromatophores: Arms: Dorsally, from
to 9 distributed in a single row near the base, but in twoows at the tips. Ventrally, from 3 to 6 following the same pat-ern as in the dorsal view. Head, Eyes and Funnel: No changes.antle: Dorsally from 10 to 23, four are near the mantle edge
nd from 6 to 17 are visceral chromatophores. In ventral view
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
o changes.3.76 ± 0.18 mm ML (17 days old) (Fig. 1e, Tables 1 and 2).
antle: Average width 2.34 ± 0.19 mm (61.2% of ML). Head: Aver-ge width 2.0 ± 0.12 mm (53% of ML). Funnel: Average length.70 ± 0.12 mm. Arms: From 5 to 9 suckers each. The length of the
ig. 2. Details of the dorsal chromatophore fields of O. vulgaris paralarvae from Northeasd) Arms, (e) head, and (f) mantle of a 36-day old paralarvae; (g) Arms, (h) Head, and (i) D
PRESSarch xxx (2010) xxx–xxx 5
arm I right represents 57.4% of the ML. Chromatophores: Arms: Indorsal view 5–9 and ventrally 4–7 distributed in the same patterndescribed above. Head: Dorsally, from 10 to 12, four new small onesappear laterally in the connective tissue under the eyes. Ventrally,six. Eyes and Funnel: No changes. Mantle: Dorsally from 15 to 25 ofwhich 10–20 are over the viscera. Ventrally no changes.
4.07 ± 0.14 mm ML (20 days old) (Fig. 1f, Table 1). Mantle:Average width 2.50 ± 0.12 mm (61.4% of ML). Head: Average width2.10 ± 0.08 mm. Funnel: average length 1.70 ± 0.03 mm (41.8% ofML). Arms: From 5 to 10 suckers each. The length of the arm I rightis 59.2% of the ML. Chromatophores: Arms: Dorsally from 5 to 13and ventrally from 4 to 11 distributed in the same pattern describedabove. Head: Dorsally from 12 to 13 and in ventral view six. Eyes andFunnel: No changes. Mantle: In dorsal view 15–30 of which 11–22are visceral chromatophores. Ventrally, no changes.
4.43 ± 0.30 mm ML (27 days old) (Fig. 2a–c, Table 1). Man-tle: Average width 2.75 ± 0.50 mm (62% of ML). Head: Averagewidth 2.32 ± 0.30 mm. Arms: From 7 to 14 suckers each. Chro-matophores: Arms: Dorsally, 7 to 13 in ventral view, 7 to 11distributed in the same pattern as described above. Head: Dorsally∼16, two or three smaller ones appeared on the laterals of the headin the connective tissue under the eyes; ventrally no changes. Eyes:In dorsal view three, from 1 to 2 new ones. Funnel: No changes.Mantle: Dorsally, from 25 to 34, several small new ones. Ventrally,from 18 to 24.
5.31 ± 0.29 mm ML (36 days old) (Fig. 2d–g, Table 1). Mantle:Average width 3.57 ± 0.05 mm (67.2% of ML). Head: Average width
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
2.94 ± 0.04 mm (55% of ML). Arms: From 10 to 20 suckers each.Chromatophores: Arms: 10–15 dorsally and 11–14 ventrally dis-tributed in the same pattern described above. Head: Dorsally ∼18,several small new ones on the laterals of the head; ventrally nochanges. Eyes and Funnel: No changes: Mantle: Dorsally from 32 to
tern Atlantic (Spain). (a) arms, (b) head, and (c) mantle of a 27-day old paralarvae;orsal view of a forty-five-days old paralarvae. Scale bar = 1 mm.
ARTICLE IN PRESSG Model
FISH-3000; No. of Pages 10
6 E.A.G. Vidal et al. / Fisheries Research xxx (2010) xxx–xxx
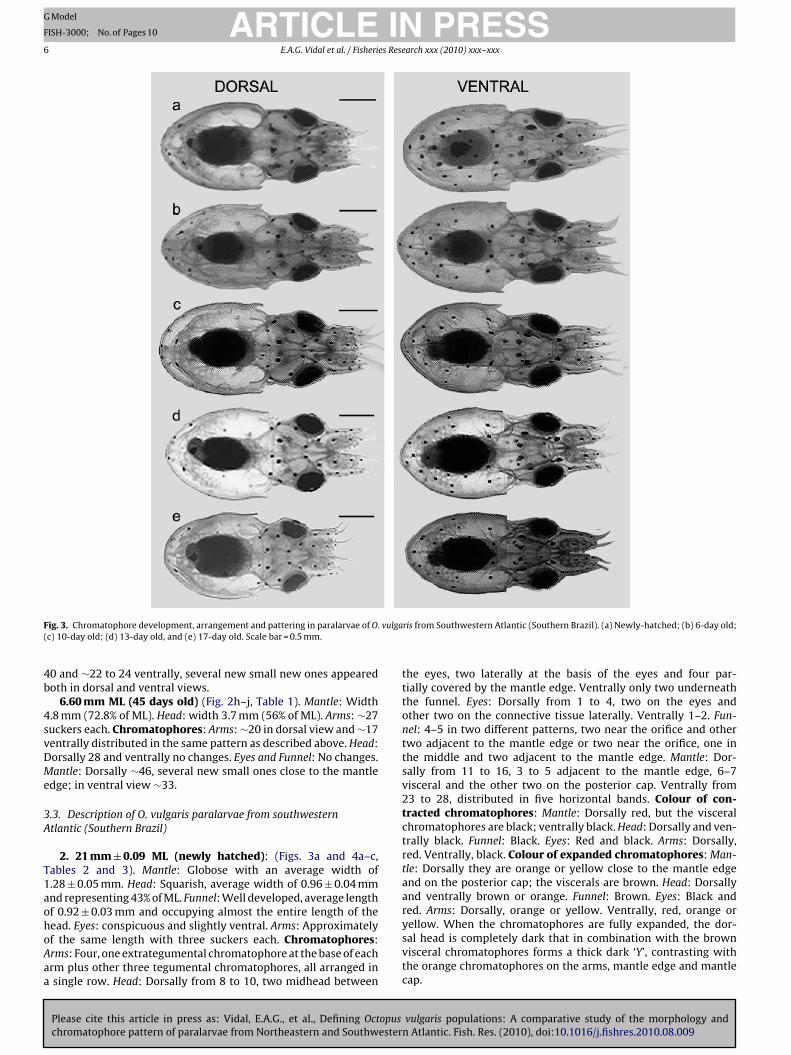
F . vulga(
4b
4svDMe
3A
T1aohoAaa
ig. 3. Chromatophore development, arrangement and pattering in paralarvae of Oc) 10-day old; (d) 13-day old, and (e) 17-day old. Scale bar = 0.5 mm.
0 and ∼22 to 24 ventrally, several new small new ones appearedoth in dorsal and ventral views.
6.60 mm ML (45 days old) (Fig. 2h–j, Table 1). Mantle: Width.8 mm (72.8% of ML). Head: width 3.7 mm (56% of ML). Arms: ∼27uckers each. Chromatophores: Arms: ∼20 in dorsal view and ∼17entrally distributed in the same pattern as described above. Head:orsally 28 and ventrally no changes. Eyes and Funnel: No changes.antle: Dorsally ∼46, several new small ones close to the mantle
dge; in ventral view ∼33.
.3. Description of O. vulgaris paralarvae from southwesterntlantic (Southern Brazil)
2. 21 mm ± 0.09 ML (newly hatched): (Figs. 3a and 4a–c,ables 2 and 3). Mantle: Globose with an average width of.28 ± 0.05 mm. Head: Squarish, average width of 0.96 ± 0.04 mmnd representing 43% of ML. Funnel: Well developed, average lengthf 0.92 ± 0.03 mm and occupying almost the entire length of the
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
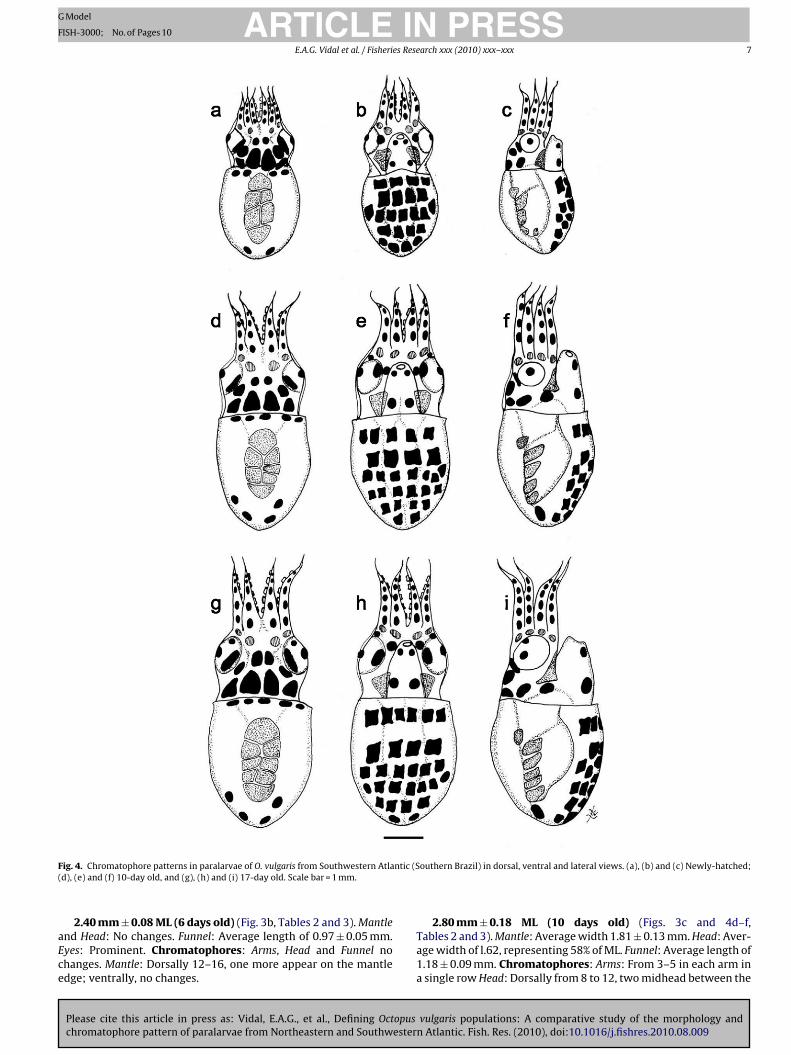
ead. Eyes: conspicuous and slightly ventral. Arms: Approximatelyf the same length with three suckers each. Chromatophores:rms: Four, one extrategumental chromatophore at the base of eachrm plus other three tegumental chromatophores, all arranged insingle row. Head: Dorsally from 8 to 10, two midhead between
ris from Southwestern Atlantic (Southern Brazil). (a) Newly-hatched; (b) 6-day old;
the eyes, two laterally at the basis of the eyes and four par-tially covered by the mantle edge. Ventrally only two underneaththe funnel. Eyes: Dorsally from 1 to 4, two on the eyes andother two on the connective tissue laterally. Ventrally 1–2. Fun-nel: 4–5 in two different patterns, two near the orifice and othertwo adjacent to the mantle edge or two near the orifice, one inthe middle and two adjacent to the mantle edge. Mantle: Dor-sally from 11 to 16, 3 to 5 adjacent to the mantle edge, 6–7visceral and the other two on the posterior cap. Ventrally from23 to 28, distributed in five horizontal bands. Colour of con-tracted chromatophores: Mantle: Dorsally red, but the visceralchromatophores are black; ventrally black. Head: Dorsally and ven-trally black. Funnel: Black. Eyes: Red and black. Arms: Dorsally,red. Ventrally, black. Colour of expanded chromatophores: Man-tle: Dorsally they are orange or yellow close to the mantle edgeand on the posterior cap; the viscerals are brown. Head: Dorsallyand ventrally brown or orange. Funnel: Brown. Eyes: Black andred. Arms: Dorsally, orange or yellow. Ventrally, red, orange or
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
yellow. When the chromatophores are fully expanded, the dor-sal head is completely dark that in combination with the brownvisceral chromatophores forms a thick dark ‘Y’, contrasting withthe orange chromatophores on the arms, mantle edge and mantlecap.
ARTICLE IN PRESSG Model
FISH-3000; No. of Pages 10
E.A.G. Vidal et al. / Fisheries Research xxx (2010) xxx–xxx 7
F ntic (S(
aEce
ig. 4. Chromatophore patterns in paralarvae of O. vulgaris from Southwestern Atlad), (e) and (f) 10-day old, and (g), (h) and (i) 17-day old. Scale bar = 1 mm.
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
2.40 mm ± 0.08 ML (6 days old) (Fig. 3b, Tables 2 and 3). Mantlend Head: No changes. Funnel: Average length of 0.97 ± 0.05 mm.yes: Prominent. Chromatophores: Arms, Head and Funnel nohanges. Mantle: Dorsally 12–16, one more appear on the mantledge; ventrally, no changes.
outhern Brazil) in dorsal, ventral and lateral views. (a), (b) and (c) Newly-hatched;
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
2.80 mm ± 0.18 ML (10 days old) (Figs. 3c and 4d–f,Tables 2 and 3). Mantle: Average width 1.81 ± 0.13 mm. Head: Aver-age width of l.62, representing 58% of ML. Funnel: Average length of1.18 ± 0.09 mm. Chromatophores: Arms: From 3–5 in each arm ina single row Head: Dorsally from 8 to 12, two midhead between the
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
ARTICLE ING Model
FISH-3000; No. of Pages 10
8 E.A.G. Vidal et al. / Fisheries Rese
Tab
le3
Mea
nM
L,ag
ean
dch
rom
atop
hor
ep
atte
rnof
O.v
ulga
ris
par
alar
vae
from
the
sou
thw
este
rnA
tlan
tic
(Sou
ther
nB
razi
l).C
hro
mat
oph
ores
inth
ear
ms
are
arra
nge
din
asi
ngl
ero
w.
Mea
nM
L±
SD(m
m)
Age
(day
s)C
hro
mat
oph
ores
dor
salv
iew
Tota
lC
hro
mat
oph
ores
ven
tral
view
Tota
l
Arm
sH
ead
Eyes
Man
tle
edge
Man
tle
Man
tle
tota
lA
rms
Hea
dEy
esM
antl
eFu
nn
el
2.21
±0.
090
3–4
(1ro
w)
2+
2+
4;2
+3
+4;
2+
2+
2+
4;2
+2
+3
+4;
2+
4+
2+
41–
43–
58–
11(6
–8V
S)11
–16
23–3
63–
4(1
row
)2
1–2
23–2
82
+2
or2
+1
+2
33–4
1
2.40
±0.
086
3–4
(1ro
w)
2+
2+
4;2
+3
+4;
3+
2+
4;2
+4
+2
+4
1–4
4–5
8–11
(6–8
VS)
12–1
624
–36
3–4
(1ro
w)
21–
223
–28
2+
2or
2+
1+
233
–41
2.80
±0.
1810
3–5
(1ro
w)
2+
2+
4;2
+2
+3
+4;
2+
4+
2+
42–
44–
58–
12(6
–9V
S)14
–17
27–3
73–
4(1
row
)6
1–2
28–3
22
+2;
2+
1+
2;3
+2;
3+
1+
238
–46
2.95
±0.
2113
3–5
(1ro
w)
2+
2+
4;2
+2
+3
+4;
2+
4+
2+
42–
44–
59–
12(8
–9V
S)15
–17
28–3
73–
4(1
row
)6
1–2
29–3
42
+2;
2+
1+
2;3
+2;
3+
1+
238
–50
3.38
±0.
3417
4–7
(1ro
w)
2+
2+
4;2
+2
+4
+2;
2+
4+
2+
43–
44–
59–
12(8
–9V
S)17
–20
31–4
03–
4(1
row
)6
1–2
29–3
62
+2;
2+
1+
2;3
+2;
3+
1+
239
–52
ML:
man
tle
len
gth
;V
S:vi
scer
al.H
ead
mea
ns
hea
dw
ith
out
eyes
.Rig
ht
and
left
eye
are
alw
ays
iden
tica
l.
PRESSarch xxx (2010) xxx–xxx
eyes, two laterally at the base of the eyes and four close to the man-tle edge. Ventrally six, two small ones above the eyes, two midheadclose to the funnel orifice and two partially hidden by the funnel.Eyes: No changes. Funnel: 4 to 6, 2 to 3 adjacent to the orifice andtwo close to the base of the funnel or 2 to 3 adjacent to the orifice,one at midcentral and two close to the base of the funnel. Mantle:Dorsally 14 to 17, one new visceral appears and there are two newones on the posterior region. Ventrally 28–32, five near the mantleedge.
2.95 ± 0.21 mm ML (13 days old) (Fig. 3d, Table 3). Mantle andHead: No major changes. Chromatophores: Arms, Head, Eyes andFunnel: No changes. Mantle: Dorsally from 15 to 17. Ventrally from29 to 34.
3.38 ± 0.34 mm ML (17 days old) (Fig. 3e and 4g–i,Tables 2 and 3). Mantle: Average width 2.11 ± 0.15 mm; Head: aver-age width of 1.87 ± 0.19 mm and Funnel Length 1.54 ± 0.13 mm.Chromatophores: Arms: 6–7 in a single row. Head: no changes.Eyes: From 3 to 4, one more appears dorsally. Funnel: Same patternsas above. Mantle: Dorsally, from 17 to 20. Ventrally from 29–36,six near the mantle edge.
3.4. Main morphological and chromatophore pattern differencesof O. vulgaris paralarvae from the northeastern and southwesternAtlantic up to 3.7 mm ML and 17 days after hatching
When several body dimensions of same-age paralarvae werecompared between regions, no significant differences were found(P > 0.05) (Table 2). Additionally, growth rates of Spanish paralarvae(ML = 2.11e0.034A; R2 = 0.95) were not significantly different fromthose of Brazilian paralarvae (ML = 2.15e0.027A; R2 = 0.97) (P > 0.05).
In relation to the chromatophore pattern, several differenceswere observed, as follows: Head: Dorsally, the number of chro-matophores was similar (8–13). However, the distribution patternwas quite different, Spanish paralarvae showed only three patternsagainst five of Brazilian paralarvae (Table 4). Ventrally, paralarvaefrom Spain had from 2 to 6 chromatophores and those from Brazilfrom 2 to 4. Eyes: In Spanish paralarvae only one both in dorsaland ventral views, but in Brazilian paralarvae from 1 to 4 dorsallyand from 1 to 2 ventrally. Mantle: In paralarvae from Spain, 10 to25 dorsally of which 6–20 are viscerals, and ventrally, 16 to 20. InBrazilian paralarvae, 11–20 dorsally of which 6–9 are viscerals and23–36 ventrally. Funnel: Only four chromatophores distributed in asingle pattern in Spanish paralarvae against from 4 to 6 distributedin four different patterns in Brazilian paralarvae.
4. Discussion
This study showed that there are marked differences in the chro-matophore pattern of O. vulgaris paralarvae from the northeasternand southwestern Atlantic, through an outline of the changes in thechromatophore fields during early development. Newly-hatchedparalarvae from Southern Brazil have a total number of chro-matophores ranging from 56 to 77 against 48 to 56 in those fromSpain. Brazilian paralarvae had more chromatophores on the ven-tral mantle, on the eyes and on the funnel than Spanish paralarvaeand both the number and pattern observed on the dorsal headand funnel were quite different. On the other hand, Spanish par-alarvae have more chromatophores on the dorsal mantle than theBrazilian paralarvae. However, no significant difference was foundbetween regions for mean ML, MW, HW, FuL and growth rate when
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
same age paralarvae were compared (Table 2). This is of note,considering the large intraspecific variability in size, weight andyolk content of cephalopods at hatching (Villanueva, 2000; Vidalet al., 2002, 2006). Perhaps, the similarities between the incuba-tion and rearing temperatures in the two regions had a major

ARTICLE IN PRESSG Model
FISH-3000; No. of Pages 10
E.A.G. Vidal et al. / Fisheries Research xxx (2010) xxx–xxx 9
Table 4Main differences in the chromatophore pattern of O. vulgaris paralarvae from the northeastern and southwestern Atlantic up to 3.76 mm ML and 17 days after hatching.
Chromatophorepattern
Northeastern Atlantic (Spain) Southwestern Atlantic (Southern Brazil)
Dorsal Ventral Dorsal Ventral
Head 8–13 (2 + 2 + 2 + 2; 2 + 2 + 2 + 2 + 2 + 2 or(2 + 2 + 2 + 2 + 1 + 2 + 2)
2–6 8–12 (2 + 2 + 4; 2 + 2 + 2 + 4; 2 + 2 + 3 + 4;2 + 3 + 4; or 2 + 4 + 2 + 4)
2–4
Eyes 1 1 1–4 1–2–11
H
etsasprdppt1alMfi
tuiaotp
vcmcvltsoivcapdte(isttwtwtst
Funnel – 4 (2 + 2)Mantle 10–25 (6–20 VS) 16–20
ead means head without eyes. Right and left eye are always identical.
ffect on growth rate. Nevertheless, variability was observed inhe number of chromatophores per field and in the number ofuckers per arm of paralarvae. Individual variability in the numbernd location of chromatophores has been observed before in latetage embryos (Fioroni, 1965), and also in the number of suckerser arm for hatchlings of O. vulgaris and Enteroctopus dofleini (foreview see Villanueva and Norman, 2008). However, our resultsemonstrated that this variability tend to increase with size. Otherossible sources of the variability observed in the chromatophoreatterns of paralarvae could be attributed to the different diet, pho-operiod or even related to the thermal latitude (Atkinson and Sibly,997), since animals hatching at higher latitudes tend to be larger,nd thus could have a greater number of chromatophores. Nonethe-ess, paralarvae from both regions hatched out at almost the same
L and Spanish paralarvae had lesser chromatophores in someelds (ventral mantle and funnel) than the Brazillian paralarvae.
Regardless of the variability observed the results also revealedhat some chromatophore fields display a reliable and almostnchangeable number of chromatophores during the size/age
nterval evaluated, i.e. the dorsal and ventral view of eyes, the headnd the dorsal mantle under the viscera. Therefore, the occurrencef dissimilarities in the chromatophore pattern of paralarvae onhese conservative fields should not be attributed to phenotypiclasticity.
One of the most characteristic and consistent patterns in O.ulgaris early paralarvae from both localities is the “few largehromatophores” that includes from 6 to 8 large visceral chro-atophores located on the dorsal surface of the viscera. This
hromatophore field is easily recognized in hatchlings and paralar-ae up to 15 days of age (3.6 mm ML) from both regions, forming aarge dark oval spot on the dorsal mantle and thus is a conspicuousaxonomic character. Afterwards, other smaller chromatophorestart to be found in the middle of the larger ones and at the agef 20 days (4.0 mm ML) there are from 11 to 22 chromatophoresn this area. The pattern observed in the ventral head was alsoery conservative in early paralarvae. In contrast, the number ofhromatophores increased continually on the dorsal mantle, headnd arms, particularly in older octopus (>20 days old). O. vulgarisaralarvae hatched out with a few large chromatophores on theorsal view and a greater number of smaller chromatophores onhe ventral view. This enables them to display a translucent silhou-tte (contracted chromatophores) or a dark and colourful silhouettebrown and orange/yellow chromatophores expanded), character-zed by a dark head and a transparent mantle with an oval darkpot over the viscera on the dorsal view and a all dark pattern inhe ventral view. In spite of this, the colour changes during its plank-onic life on the dorsal view are far more pronounced, culminatingith a wide array of chromatophores and a complex colour pat-
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
ern as grounding for the benthic life. Our results are in agreementith those of Packard (1985), who observed a rapid increase in
he number of tegumental and visceral chromatophores in the dor-al mantle of O. vulgaris during the transition from the planktonico the benthonic life style that happens between 40 and 52 days
4–6 (2 + 2; 2 + 1 + 2; 3 + 2; 3 + 1 + 2)–20 (6–9 VS) 23–36
after hatching at a mean temperature of 21–22 ◦C (Villanueva andNorman, 2008). During this transition, a major change in morphol-ogy, physiology and behaviour takes place during a relatively shortperiod (Villanueva et al., 1996; Villanueva and Norman, 2008) anda more complex chromatophore pattern is required on settlementfor camouflage, hunting and survival in the benthos.
The chromatophore pattern of O. vulgaris hatchlings(1.5–2.0 mm ML), 2.4 mm ML (∼10 days old) and 6 mm MLparalarvae from the Western Mediterranean was described byHochberg et al. (1992). The chromatophore pattern of both thedorsal and ventral head of the paralarvae from the Mediterraneanwas very similar to the pattern described here for Brazilian paralar-vae, although hatchlings from Brazil (2.21 mm ML) had a similarnumber of chromatophores to a 6.0 mm ML paralarvae from theMediterranean. Additionally, the pattern observed by Hochberg etal. (1992) on the ventral mantle of hatchlings was comparable tothat observed in this study for hatchlings from Spain. However,it was not possible to draw further comparisons with our resultsdue to differences in ML. Mainly contracted chromatophoreswere described in the present study, but this does not seemsto affect patterns at all, i.e. the number of chromatophores isconservative, both in contracted and in extended form. Extendedchromatophores have shown a more reliable definition of colourpatterns in Brazilian paralarvae (Fig. 4), but in every case, thecolour of chromatophores depends on the degree of expansion ofthe elastic sacculus in which the granules of pigment are found.
The underlying causes of the differences observed on the chro-matophore pattern of O. vulgaris paralarvae from Brazil and Spainshould be investigated further, since the chromatophore patternhas long been recognized as one of the most useful tools for iden-tification of paralarval cephalopods to species level (Young et al.,1989; Hochberg et al., 1992). It is possible that the differencesobserved in the chromatophore pattern of paralarvae from twogeographically separated regions are indicating the existence ofdistinct populations of a supposed cosmopolitan species, or even,a cryptic O. vulgaris-like species along the Southern Brazilian coast.Hence, detailed genetic studies are required to establish the degreeof taxonomic divergence between Octopus specimens from Spainand Southern Brazil.
Recently, a wide variety of molecular studies with phylogeneticinference and morphological observations have been carried out forOctopus and the O. vulgaris complex to increase the understandingof biogeography and phylogenetic relationships. One example is O.insularis, a recently described species belonging to the O. vulgariscomplex off the northwestern coast of Brazil (Leite et al., 2008). Alsoworking on this subject, Guerra et al. (2010), extended the distribu-tion of O. vulgaris into the Indian Ocean (Amsterdam and Saint PaulIslands, halfway between South Africa and Australia). Their study
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
reinforces the idea that O. vulgaris is not a complex of species but asingle species, which has a wide geographic distribution. Therefore,further studies are necessary to clarify the distribution and phylo-genetic status of Octopus species off the Brazilian coast. It would beof considerable importance to carry out a multidisciplinary study

ING
F
1 es Rese
ca
A
mTiot(
R
A
F
G
H
I
L
M
M
N
ARTICLEModel
ISH-3000; No. of Pages 10
0 E.A.G. Vidal et al. / Fisheri
ombining genetic studies and morphological observations of par-larvae, juveniles and adults along the Atlantic.
cknowledgements
We thank two anonymous reviewers for valued input to theanuscript. L. Fuentes was supported by the PETRI (Proyectos de
ransferencia de Resultados de la Investigación) Program of Span-sh Research and L.B. Silva was supported by a Brazilian Ministryf Education (CAPES) scholarship. E.A.G. Vidal would like to thankhe PROFIX Program from the Brazilian National Research CouncilCNPq-Proc. no. 540542/01-6).
eferences
tkinson, D., Sibly, R.M., 1997. Why are organisms usually bigger in colder environ-ments? Making sense of a life history puzzle. Trends. Ecol. Evol. 12, 235–239.
ioroni, P., 1965. Die embryonale Musterentwicklung bei einigen Mediterranen Tin-tenfischarten. Vie et Milieu 16, 655–756.
uerra, A., Roura, A., González, A.F., Pascual, S., Cherel, I., Pérez-Losada, M., 2010.Morphological and genetic evidences that octopus from Amsterdam and SaintPaul Islands waters (southern Indian Ocean) is Octopus vulgaris. ICES J. Mar. Sci.67, 000–10.
ochberg, F.G., Nixon, M., Toll, R.B., 1992. Orden Octopoda Leach 1818. In: Sweeney,M.J., Roper, C.F.E., Mangold, K.M., Clarke, M.R., Boletzky, S.V. (Eds.), Larval” andJuvenile Cephalopods: A Manual for their Identification. Smithsonian InstitutionPress. D.C., number 513. Smithsonian Contrib. Zool, Washington, pp. 213–279.
glesias, J., Sánchez, F.J., Bersano, J.G.F., Carrasco, F.J., Dhont, J., Linares, F., Okumura,S., Meeren, T., van der, Munoz, J.L., Roo, J., Vidal, E.A.G., Villanueva, R., 2007.Rearing of Octopus vulgaris paralarvae: present status, bottlenecks and trends.Aquaculture 266, 1–15.
eite, T.S., Haimovici, M., Molina, W., Warnke, K., 2008. Morphological and geneticdescription of Octopus insularis, a new cryptic species in the Octopus vulgariscomplex (Cephalopoda: Octopodidae) from the tropical southwestern Atlantic.J. Moluscan Stud. 74, 63–74.
angold, K., 1997. Octopus vulgaris: review of the biology. In: Lang, M.A., Hochberg,
Please cite this article in press as: Vidal, E.A.G., et al., Defining Octopuschromatophore pattern of paralarvae from Northeastern and Southwester
F.G. (Eds.), Proceedings of the Workshop on “The Fishery and Market Potentialof Octopus in California”. Smithsonian Institution Press, Washington, pp. 85–90.
angold, K., Hochberg, F.G., 1991. Defining the genus Octopus: redescription ofOctopus vulgaris. Bull. Mar. Sci. 49, 665.
orman, M., 2003. Cephalopods of the World: A World Guide. ConchBooks, Haken-heim, Germany.
PRESSarch xxx (2010) xxx–xxx
Otero, J., Salgado, X.A., González, Á.F., Groom, S.B., Miranda, A., Cabanas, J.M., Guerra,Á., 2008. Bottom–up and top–down controls of common octopus (Octopus vul-garis) in a wind-driven upwelling area (Galicia NE Atlantic). Mar. Ecol. Prog. Ser.362, 181–192.
Otero, J., Salgado, X.A., González, Á.F., Gilcoto, M., Guerra, Á., 2009. High-frequencycoastal upwelling events influence Octopus vulgaris larval dynamics on the NWIberian shelf. Mar. Ecol. Prog. Ser. 386, 123–132.
Packard, A., 1985. Sizes and distribution of chromatophores during post-embryonicdevelopment in Cephalopods. Vie Milieu 35 (3/4), 285–298.
Roper, C.F.E., Voss, G., 1983. Guidelines for taxonomic descriptions of cephalopodspecies. Mem. Nat. Mus. Victoria 44, 49–63.
Roper, C.F.E., Sweeny, M.J., Nauen, C.E., 1984. FAO species catalogue Vol. 3Cephalopods of the world. An annotated and illustrated catalogue of speciesof interest to fisheries. FAO, Rome Fisheries. Synopsis 125, 3, pp. 1–277.
Söller, R., Warnke, K., Saint Paul, U., Blohm, D., 2000. Sequence divergence of mito-chondrial DNA indicates cryptic biodiversity in Octopus vulgaris and supportsthe taxonomic distinctiveness of Octopus mimus (Cephalopoda: Octopodidae).Mar. Biol. 136, 29–35.
Teske, P.R., Oosthuizen, A., Papadopoulos, I., Barker, N.P., 2007. Phylogeographicstructure of Octopus vulgaris in South Africa revisited: identification of a secondlineage near Durban harbour. Mar. Biol. 151, 2119–2122.
Vaz-Pires, P., Seixas, P., Barbosa, A., 2004. Aquaculture potential of the commonoctopus (Octopus vulgaris Cuvier, 1797): a review. Aquaculture 238, 221–238.
Vidal, E.A.G., DiMarco, F.P., Wormuth, J.H., Lee, P.G., 2002. Influence of temperatureand food availability on survival, growth and yolk utilization in hatchling squid.Bull. Mar. Sci. 71, 915–931.
Vidal, E.A.G., DiMarco, P., Lee, P.G., 2006. Effects of starvation and recovery on thesurvival, growth and RNA:DNA ratio in loliginid squid paralarvae. Aquaculture260, 94–105.
Villanueva, R., 2000. Effect of temperature on statolith growth of the European squidLoligo vulgaris during early life. Mar. Biol. 136, 449–460.
Villanueva, R., Norman, M.D., 2008. Biology of the planktonic stages of benthic octo-puses. Oceanography Marine Biol.: Annu. Rev. 46, 105–202.
Villanueva, R., Nozais, C., Boletzky, S.v., 1996. Swimming behaviour and food search-ing in planktonic Octopus vulgaris Cuvier from hatching to settlement. J. Exp. Mar.Biol. Ecol. 208, 169–184.
Warnke, K., 1999. Diversität des Artenkomplexes Octopus cf. vulgaris Cuvier, 1797in Beziehung zu seiner Verbreitung an der Ost- und Westküste Lateinamerikas.Ph.D Thesis, University of Bremen, Shaker Verlag, Aachen.
Warnke, K., Söller, R., Blohm, D., Saint-Paul, U., 2004. A new look at geographic
vulgaris populations: A comparative study of the morphology andn Atlantic. Fish. Res. (2010), doi:10.1016/j.fishres.2010.08.009
and phylogenetic relationships within the species group surrounding Octopusvulgaris (Mollusca Cephalopod): indications of very wide distribution from mito-chondrial DNA sequences. J. Zool. Syst. Evol. Res. 42, 306–312.
Young, R.E., Harman, R.F., Hochberg, F.G., 1989. Octopodid paralarvae from Hawaiianwaters. Veliger 32, 152–165.
Zar, J.H., 1996. Biostatistical Analysis, 3rd ed. Prentice-Hall, New Jersey.




















