
Insights from letter position dyslexia on morphological decomposition in reading

Neuropsychologia 50 (2012) 2044–2055
Contents lists available at SciVerse ScienceDirect
Neuropsychologia
0028-39
http://d
n Corr
E-m
journal homepage: www.elsevier.com/locate/neuropsychologia
Deficit in the preattentive processing of syllabic duration and VOTin children with dyslexia
Julie Chobert a,n, Clement Franc-ois b, Michel Habib a,c, Mireille Besson a,d
a Laboratoire de Neurosciences Cognitives, CNRS – Aix-Marseille Universite, Franceb Institut de Neurosciences des Syst�emes, INSERM – Aix-Marseille Universite, Francec Departement de Neurologie Pediatrique, CHU Timone, Marseille, Franced Cuban Neuroscience Center, La Havane, Cuba
a r t i c l e i n f o
Article history:
Received 2 May 2011
Received in revised form
23 March 2012
Accepted 4 May 2012Available online 15 May 2012
Keywords:
Dyslexia
Children
Mismatch Negativity (MMN)
Frequency
Duration
Voice Onset Time (VOT)
32/$ - see front matter & 2012 Elsevier Ltd. A
x.doi.org/10.1016/j.neuropsychologia.2012.05
esponding author.
ail address: [email protected] (J. Cho
a b s t r a c t
The aim of this experiment was to examine the preattentive processing of syllables in 9–11-year-old
children with dyslexia and matched controls using the Mismatch Negativity (MMN), an auditory Event-
Related brain potential (ERP) related to preattentive discrimination. Children were presented with a
sequence of syllables that included standards (the syllable ‘‘Ba’’) and deviants in vowel frequency,
vowel duration and Voice Onset Time (VOT) that were either close to or far from the standard (Small
and Large deviants). No between-group differences were found for frequency deviants. However, whilst
normal-reading children showed larger MMNs to Large than to Small deviants in vowel duration and
VOT, no such deviance size effect was found in children with dyslexia. These results are taken to
indicate that the preattentive processing of vowel duration and VOT is impaired in children with
dyslexia, with no impairment in the processing of vowel frequency deviants. By revealing processing
deficits of both duration and VOT deviants, these results suggest a strong link between acoustical and
phonological processing.
& 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Developmental dyslexia is a specific disability in acquiringnormal reading skills despite conventional instruction and socio-cultural opportunities, normal intelligence and motivation, andthe absence of sensory deficits (Collective expertise INSERM,2007; Demonet, Taylor, & Chaix, 2004; Lyon, Shaywitz, &Shaywitz, 2003; Vellutino, Fletcher, Snowling, & Scanlon, 2004).However, the nature and origin of dyslexia still remain unclear.Visual and auditory deficits have been reported in children withdyslexia (e.g., Lallier, Donnadieu, Berger, & Valdois, 2010; Ramus,2003), with a phonological deficit often found to be the mostcommon (for reviews see, Goswami, 2011; Liberman &Shankweiler, 1985; Mody, 2003; Snowling, 1998). Although thereis still a debate as to whether this phonological deficit is causallylinked to a more general deficit in acoustic processing, there issome agreement that the acoustic content of speech, especially itstemporal features, are processed abnormally by dyslexic children(Bishop, 2007; Goswami, 2011).
Functional brain imaging has prompted great interest into thistopic and electrophysiological methods are widely used to
ll rights reserved.
.004
bert).
investigate the auditory correlates of dyslexia, especially exploit-ing two characteristics: (1) their excellent temporal resolutionand (2) the possibility they offer to investigate preattentiveprocesses (i.e., to test for the presence of deficits not onlyinterpretable by the subjects’ own attentional and/or motiva-tional state).
In the present experiment, we investigated the preattentiveprocessing of speech sounds in dyslexic children using theMismatch Negativity (MMN). The MMN is a change-specificcomponent of auditory Event-Related brain Potentials (ERPs)elicited by infrequently occurring discriminable deviant soundswithin a repetitive sequence of standard stimuli and is taken as ameasure of preattentive, implicit auditory processing (Naatanen,Gaillard, & Mantysalo, 1978; for review, see Naatanen,Paavilainen, Rinne, & Alho, 2007). The MMN, obtained by sub-tracting the ERPs to the standard from the ERPs to the deviantstimuli, typically peaks between 100–300 ms from change onsetand exhibits maximal amplitude over frontal and temporalregions of the scalp. Moreover, when the magnitude of thedeviance increases, the amplitude of the MMN also increasesand its latency shortens (Novitski, Tervaniemi, Huotilainen, &Naatanen, 2004; Sams, Paavilainen, Alho, & Naatanen, 1985).Finally, even if MMN amplitude can be modulated by attention(Woldorff, Hackley, & Hillyard, 1991), the MMN is also obtainedwhen the subject’s attention is oriented away from the auditory

J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2045
stimuli (for a review, see Naatanen et al., 2007), which is animportant advantage when studying dyslexic children who oftenalso show attention deficits. As a consequence, the MMN hasoften been used to compare dyslexics and normal-readers (forreviews see Bishop, 2007; Goswami, 2011; Kujala & Naatanen,2001; Schulte-Korne & Bruder, 2010).
Whereas early studies mainly aimed at investigating proces-sing of the temporal features of speech or non-speech stimuli,abnormalities for a larger variety of acoustic properties, includingpitch, duration, intensity and pattern perception, have been morerecently reported, though to date these have yielded poorlyconsistent results (Bishop, 2007; Schulte-Korne & Bruder, 2010).Here, we aimed to determine which basic attributes of speechperception, both at the acoustic and phonological levels, areimpaired in dyslexics. To this end, we examined the preattentiveprocessing of speech Consonant Vowel (CV) syllables in childrenwith dyslexia and in normal reading children by using a multi-feature MMN paradigm in which standard sounds alternate in thesame MMN sequence with several types of deviants, each differ-ing from the standard in one specific feature (Naatanen,Pakarinen, Rinne, & Takegata, 2004). We used the syllable ‘‘Ba’’as standard and alterations of vowel frequency, vowel durationand Voice Onset Time (VOT) as deviants. Moreover, deviants ineach condition were either close to or far from the standard(Small and Large deviants).
Lovio et al. (2009) have used a new variant of the multi-featureparadigm that was especially targeted for the study of speech-sound discrimination in 6-year-old children. They used CV sylla-bles as standard stimuli and created deviants in vowel, consonant,vowel-frequency (F0), vowel-duration and syllable-intensity.Importantly, they compared the MMNs recorded in this multi-feature paradigm with the MMNs in the traditional oddballparadigm (i.e., only one deviant per sound sequence) and showedthat the amplitude and latency of the effects were similar in bothcases. Lovio, Naatanen, and Kujala (2010) used the same design tocompare 6-year-old children at risk for dyslexia and normal-reading children. They found significantly smaller MMN ampli-tudes in children at risk of dyslexia compared to normal-readersfor consonant deviants that differed from the standard on theplace of articulation (that allows one to distinguish /ba/ and /da/syllables, for example) and for vowel, vowel-duration, consonantand intensity deviants but no between-group differences forvowel-frequency deviants.
The preattentive processing of frequency deviants has beeninvestigated in several studies using pure tones. Typically, theMMN to large deviants does not differ between dyslexic andcontrols, whether adults (Schulte-Korne, Deimel, Bartling, &Remschmidt, 2001; Standard: 2200 Hz; deviant: 2640 Hz) or8–12-year-old children (Sharma et al. 2006; Standard: 1000 Hz;deviants; 1100 Hz and 1500 Hz). By contrast, the MMN to smallfrequency deviants is smaller in adults with dyslexia compared tonormal-readers (Baldeweg, Richardson, Watkins, Foale, &Gruzelier 1999; Kujala, Lovio, Lepisto, Laasonen, & Naatanen,2006a; Lachmann, Berti, Kujala & Schroger, 2005; Renvall &Hari, 2003) but no between-group differences have been reportedin children (Schulte-Korne, Deimel, Bartling, & Remschmidt, 1998,Standard: 1000 Hz, deviant: 1050 Hz; Corbera, Escera, & Artigas,2006, Standard: 500 Hz, deviant: 550 Hz).
To our knowledge, no study has yet examined the preattentiveprocessing of the frequency of speech sounds in children withdyslexia. The only related study was the above-mentioned experi-ment by Lovio et al. (2010) who found no MMN amplitude orlatency differences between children at risk of dyslexia andnormal-reading children when using standard syllables with afrequency of 101 Hz and 8 Hz deviants (8% change). Thus, whilethe evidence in the literature is still sparse, it seems that children
with dyslexia are not impaired in the preattentive processing ofpitch variations in syllables (Lovio et al., 2010). Such results standin contrast with findings from our own laboratory in an attentiveERP design that varied the pitch level of sentence final words(Santos, Joly-Pottuz, Moreno, Habib, & Besson, 2007). Dyslexicswere clearly impaired at actively detecting large frequencyvariations and the P300 component was reduced in amplitudein dyslexics compared to normal-readers matched on reading-age. Thus, it was of interest to determine whether frequencydeviants would elicit MMNs of similar amplitude in dyslexics andin normal-readers, as in Lovio et al. (2010), or whether theywould elicit smaller MMN in dyslexics than in normal-readers.
Studies of the preattentive processing of duration using tonesand syllables have produced mixed findings. While some resultsshow similar MMN amplitude to duration deviants for dyslexicsand normal-readers (Baldeweg et al., 1999; Huttunen, Halonen,Kaartinen, & Lyytinen, 2007; Kujala et al., 2006a, 2006b), otherresults reveal smaller MMN to duration deviants in dyslexics(Corbera et al., 2006; Huttunen-Scott, Kaartinen, Tolvanen, &Lyytinen, 2008). For instance, using tones rather than speechsounds, Corbera et al. (2006) and Huttunen-Scott et al. (2008)showed that standard tones lasting for 100 ms with deviants of33 ms and 30 ms in duration elicited decreased MMNs in dys-lexics compared to controls, whereas Baldeweg et al. (1999) didnot report any between-group differences when standard toneslasted for 200 ms with deviants of 160 ms, 120 ms, 80 ms and40 ms in duration. In line with the hypothesis that dyslexics areimpaired in processing short stimuli and rapid transitions(Frumkin & Rapin, 1980; Tallal & Piercy, 1974; see Tallal (2001)for a review), these contradictory findings suggest that shortdurations are more problematic than long durations for childrenwith dyslexia.
Using speech sounds, Kujala et al. (2006b) compared the ability fordyslexic and normal-reading children to passively and activelydiscriminate vowel duration in CV syllables (e.g., /taatata/, /tataata/or /tatataa/). The MMN showed no between-group differences butwhen children had to report at which position in the sequence thelong vowel occurred (1st, 2nd or 3rd position), N2b amplitude wassmaller in dyslexics than in normal-readers. Thus, dyslexics wereable to detect trials with prolonged vowels but they failed toaccurately discriminate where, among the three possible locations,the longest vowels occurred. These results were taken as evidencethat dyslexia exerts detrimental effects when children were activelyprocessing vowel duration but not when they were passively listeningto the same sounds. However, compared to normal-readers,Shankarnarayan & Maruthy (2007) found increased MMN latencyto vowel duration deviants (standard¼250 ms; deviants¼175 ms) inIndian dyslexic children. More recently, Lovio et al. (2010) reporteddecreased MMN amplitude (standard¼170 ms, deviant¼100 ms) inFinnish children at risk for dyslexia.
In the present experiment, standard syllables lasted for 278 ms(with 208 ms vowel duration) and we used both Large and Smalldeviants that were 80 ms and 50 ms shorter (38% and 24% change,respectively) than the Standard. Based on Lovio et al. (2010), wepredicted that the MMN to duration deviants should be smaller inamplitude for children with dyslexia than for normal-readers. Ofmain interest was to determine whether the deviance size effect(i.e., difference between Large and Small deviants) would besimilar or different in children with dyslexia and controls.
VOT is an important parameter that allows perceiving stopconsonant as voiced (e.g., /b/) or voiceless (e.g., /p/). The VOT isdefined as the interval between the noise burst produced atconsonant release and the onset of the waveform periodicityassociated with vocal cord vibration (Lisker & Abramson, 1967). InFrench, consonants with VOT values shorter than 0 ms tend to becategorized as voiced (negative VOT around �100 ms; /b/) and

1 This experiment is part of a longitudinal study aimed at testing the influence
of musical training on several aspects of speech processing in dyslexics and in
normal-readers. As a consequence and for ethical reasons, all children from the
class did participate in the project. This explains why, out of the 24 dyslexics
children, 12 could be matched with controls on reading-age but only 7 children
could be matched on chronological-age.
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–20552046
consonants with VOT values longer than 0 ms tend to be categorizedas voiceless (positive VOT around þ30 ms; /p/): the phonemicboundary is around 0 ms (Serniclaes, 1987). The perception of VOTin French has been examined both in normal-reading adults and ininfants (e.g., Medina, Hoonhorst, Bogliotti, & Serniclaes, 2010) withresults typically showing smaller MMNs for within-category changesthan for across-category changes, despite an equal size of deviation(Serniclaes, Sprenger-Charolles, Carre, & Demonet, 2001).
In contrast, results of several experiments have shown loweracross-category discrimination together with higher within-cate-gory discrimination in French dyslexics when compared to bothchronological-age and reading-age matched controls (Bogliotti,Serniclaes, Messaoud-Galusi, & Sprenger-Charolles, 2008). Forinstance, Serniclaes, Heghe, Mousty, Carre, & Sprenger-Charolles(2004) used a discrimination task with pairs of syllables from/ba–pa/ and /ga–ka/ continua. They showed that dyslexics exhibithigher sensitivity to phonetic features between different membersof the same phonemic category than chronological-age matchedcontrols. This increased within-category discrimination was taken toreflect higher allophonic sensitivity rather than an overall highersensitivity to acoustic features (Serniclaes et al., 2004). Based onthese findings it has been argued that the developmental deficitfound in dyslexia may result from the persistence of more categoriesthan are necessary for perceiving the phonemes of one’s nativelanguage, thereby generating weaker across-category perceptionand higher within-category discrimination (Dufor, Serniclaes,Sprenger-Charolles, & Demonet, 2009; Serniclaes et al., 2004).
To our knowledge, only two studies have directly comparedMMNs to VOT deviants in dyslexics and normal-readers. First,Csepe (2003) presented different sequences that included deviantsin VOT (/ba/ vs. /pa/, VOT difference¼80 ms) or place of articula-tion (/ba/ vs. /da) as well as pitch deviants in a pure tone sequence,to compare 7–14-year-old Hungarian dyslexic and normal-readingchildren. Results showed no differences between 7–14-year-olddyslexics and age-matched normal-readers in either MMN ampli-tude or latency to pitch changes in pure tone sequences. Bycontrast, MMNs to VOT and to place of articulation deviants wereboth smaller and developed later in children with dyslexia than innormal-readers. Based on these results, the author argued thatdeficits in these specific aspects of speech sound processing inchildren with dyslexia were linked to their phonological deficits.Second, Shankarnarayan and Maruthy (2007) used deviants in VOT(/tSa/ vs. /dZa/), place of articulation (/da/ vs. /d8a/), manner (/tSa/vs. /sa/) as well as deviants in vowel duration of the /da/ syllable(250 ms vs. 175 ms) to compare 7–12-year-old Indian dyslexic andnormal-reading children. Moreover, they used a non-linguisticsequence (standard¼1000 Hz, 250 ms) with deviants in frequency(1100 Hz, 250 ms) and duration (1000 Hz, 175 ms). Dyslexicsdisplayed longer MMN latency than controls for all linguisticcontrasts and for vowel duration deviants, as well as for thefrequency deviants in pure tones. According to the authors, theseresults indicate that both speech and non-speech processing areaffected in children with dyslexia, thereby arguing for a general,non-linguistic acoustic processing deficit. In sum, results of thetwo studies that investigated the preattentive processing of VOTlead to different conclusions, in terms of either phonologicaldeficits or general acoustic processing deficits.
To investigate this issue further, we used VOT deviants issuedfrom a /ba-pa/ continuum, with Small deviants located at middleposition (‘‘Ba�40ms’’) and Large deviants at the extreme position(‘‘Ba0ms’’), very close to ‘‘Pa’’. We hypothesized that both normal-readers and dyslexics should perceive Large deviants as across-category changes (‘‘Pa’’) and should consequently develop largeMMNs (Dehaene-Lambertz, 1997; Dehaene-Lambertz & Baillet,1998). However, if dyslexic children are less sensitive to across-category changes than controls (Dufor et al., 2009; Serniclaes
et al., 2004), the MMN to Large deviants should be smaller inamplitude and longer in latency for children with dyslexia thanfor normal-reading children. Moreover, if children with dyslexiaare more sensitive to within-category changes, they may developlarger MMNs to Small deviants than normal-reading children(Serniclaes et al., 2004; Dufor et al., 2009). In other words, thedifference between Large and Small deviants (i.e., the deviancesize effect) should be smaller in dyslexics than in normal-readers.
2. Methods
2.1. Participants
A total of 48 children with 24 normal-reading children (NR; mean age¼8.2
years; s.d.¼0.4 year, 16 boys, 19 right-handers) and 24 children with dyslexia
(Dys; mean age¼10.2 years; s.d.¼0.9 year, 16 boys, 16 right-handers) attending
the 3rd grade in 2 elementary schools (in Marseille and in Aix-en-Provence,
southern France) participated in the experiment.1 The 48 children (24 Dys and 24
NR) were divided in 4 sub-groups. Two groups were matched on Chronological
(C) age ([F(1,12)¼3.10; p¼ .10]; 7 NR-C: mean age¼8.9 years, s.d.¼0.2, 6 boys,
6 right-handers and 7 Dys-C: mean age¼9.3 years, s.d.¼0.6, 6 boys, 5 right-
handers) to ensure that potential between-group differences were not due to
maturational effects. Two other groups were matched on their reading (R) age as
assessed with the standardized ‘‘Alouette’’ reading test (Lefavrais, 1967;
[F(1,22)¼1.10; p¼0.31]; 12 NR-R: mean reading age¼7.2 years, s.d.¼0.3, 8 boys,
10 right-handers; and 12 Dys-R: mean reading age¼7.0 years, s.d.¼0.7, 10 boys,
9 right-handers) in order to examine the effect of dyslexia when the reading level
of Dys-R is similar to NR-R.
All children were native speakers of French and had normal or corrected-to-
normal vision, normal audition and no known neurological deficits as determined
from a detailed questionnaire completed by parents prior to the experiment. This
questionnaire also allowed determining that all children had similar socioeconomic
status (the French middle to low class), as revealed by the profession of the parents
and that all children were involved in extra-curricular activities (e.g., mainly sports).
Children were tested twice, individually and in a quiet classroom of the school. In one
session, children were tested using standard neuropsychological tests and in the
other session, their EEG was recorded while they performed various in-house tests.
The entire procedure was explained to the children to ensure that they agreed to
participate in the experiment. This study was conducted in accordance with local
norms and guidelines for the protection of human subjects. The inspector of schools,
as well as the school director and the teachers approved of this research project, and
parents signed an informed consent sheet prior to the experiment. Children were
given presents at the end of the sessions to thank them for their participation.
2.2. Procedure
During the first session, lasting an average of 1.5 h, each child was tested on a
large battery of standard neuropsychological and speech tests. Dyslexic children
had already been diagnosed by a neurologist and a speech therapist according to
the criteria defined in the Diagnostic and Statistical Manual of Mental Disorders IV
(American Psychiatric Association, 1994). Nevertheless, these further evaluations
allowed us to compare the level of performance of dyslexics and controls when
tested in the same conditions. Standardized tests included the Alouette’s reading
test (Lefavrais, 1967), reading strategy and phonological awareness tests (ODEDYS,
Jacquier-Roux, Valdois, & Zorman, 2005), verbal and non-verbal IQ (WISC IV,
Wechsler, 2003; Raven matrices, Raven, Corporation, & Lewis, 1962) and attention
tests (NEPSY, Korkman, Kirk, & Kemp, 1998).
During the second session, again lasting for an average of 1.5 h, each child
participated in an electrophysiological recording session. Children sat in a comfor-
table chair at 1 m distance from a computer screen. In the passive listening
experiment, EEG was recorded while the child watched a silent subtitled movie
displayed on the computer screen. Children were asked to watch the movie
without paying attention to the sounds that were presented through headphones.
Frequency, duration and VOT deviants, each with two levels of deviance size (Small
and Large deviance from the standard) were randomly presented within the
auditory sequence with a Sound Onset Asynchrony (SOA) of 600 ms synchronized
with vowel onset. A total of 1200 stimuli were used with 432 deviants (72 for each
of the 6 deviant types; 6% probability). All stimuli were presented within a single

J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2047
block that lasted for 12.2 min. At the end of the experiment, children were asked
questions to ensure they had paid attention to the movie. Finally, children were
asked to listen to twenty VOT stimuli (standard syllables, Large and Small VOT
deviants), pseudo-randomly presented through headphones, and to say aloud after
each stimulus which syllable they heard (/ba/ or /pa/). This syllable identification
test was aimed at specifying how dyslexic and control children perceived the Large
and Small VOT deviants.
2.3. Stimuli
Stimuli were syllables with Consonant–Vowel structure. The standard stimu-
lus ‘‘Ba’’ had a fundamental frequency (F0) of 103 Hz, a vowel duration of 208 ms
for a total syllabic duration of 278 ms and a VOT of �70 ms.
For frequency deviants, vowel duration and VOT were the same as for the
Standard but the F0 of the vowel was increased using the software Praat (Boersma
& Weenink, 2001). For Large deviants, the F0 was increased to 154 Hz (i.e., 51 Hz
higher than Standard, 49% increase) and for Small deviants to 117 Hz (i.e., 14 Hz
higher than Standard, 13% increase).
For duration deviants, F0 and VOT were the same as for the Standard but
vowel duration was shortened using Adobe audition software. For Large deviants,
vowel duration was 128 ms (i.e., 80 ms shorter than the Standard, 38% decrease;
Total duration Large deviant¼198 ms) and 158 ms for the Small deviant (i.e.,
50 ms shorter than the Standard, 24% decrease; Total duration Small
deviant¼228 ms).
For VOT deviants, F0 and vowel duration were the same as for the Standard but
VOT changed. Small and Large deviants were selected on a ‘‘Ba-Pa’’ continuum that
comprised 9 sounds. The Large deviant was ‘‘Ba0 ms’’ (VOT¼0 ms; i.e., 70 ms shorter
than the Standard, 100% decrease) and the Small deviant was ‘‘Ba�40 ms’’
(VOT¼�40 ms; i.e., 30 ms shorter than the Standard, 42% decrease).
2.4. ERPs recording and processing
The electroencephalogram (EEG) was continuously recorded at a sampling
rate of 512 Hz and the band-pass was of 0-102.4 Hz using a Biosemi amplifier
system (Amsterdam, BioSemi Active 2) from 32 active Ag-Cl electrodes mounted
on a child-sized elastic cap (Biosemi Pintype) at standard positions of the
International 10/20 System (Jasper, 1958): Fz, Cz, Pz, Oz, Fp1, Fp2, AF3, AF4, F7,
F8, F3, F4, Fc5, Fc6, Fc1, Fc2, T7, T8, C3, C4, Cp5, Cp6, Cp1, Cp2, P3, P4, PO3, PO4, P7,
P8, O1, O2. Data were re-referenced off-line to the algebraic average of the left and
right mastoids (or to the nose recordings) and filtered with a bandpass of 1–30 Hz
(12 dB/oct). To detect horizontal eye movements and blinks, the electro-oculo-
gram (EOG) was recorded from Flat-type active electrodes placed 1 cm to the left
and right of the external canthi, and from an electrode beneath the right eye.
Three additional electrodes were placed on the left and right mastoids and on the
nose. EEG data were analyzed using the Brain Vision Analyser software (Version
01/04/2002; Brain Products, Gmbh). Recordings were segmented into 700 ms
epochs (from �100 ms until 600 ms post-stimulus onset). Epochs with electric
activity exceeding baseline activity by 775 mV were considered as artifacts and
were automatically rejected from further processing (around 20%, with Frequency
small deviants: 22%, Frequency large deviants: 21%, Duration small deviants: 22%,
Duration large deviants: 21%, VOT small deviants: 22%, VOT large deviants: 17%).
2.5. Data analysis
Repeated measures Analyses of Variance (ANOVAs) were used to analyze data
from the various neuropsychological and speech tests. They included Group
(dyslexics vs. normal-readers) as a between-subjects factor.
For the final syllable identification test, the percentages of ‘‘Ba’’ identification
were computed for each stimulus and for each child. ANOVAs included Group
(dyslexics vs. normal-readers) as a between-subjects factor and VOT stimuli (Bastd
vs. Ba�40 ms vs. Ba0 ms) as a within-subject factor.
Mastoid off-line referenced ERP data were averaged for each child and for each
experimental condition. Difference waveforms were obtained by subtracting the
ERPs elicited by the Standard from those elicited by each deviant. The MMN was
also computed by using the nose reference to ensure the typical MMN inversion
between Fz/Cz and the mastoid electrodes (for review, see Naatanen et al., 2007).
However, because mastoid-referenced averages typically show a better signal-to-
noise ratio than the nose-referenced averages (e.g., Kujala, Tervaniemi, & Schroger,
2007), the former were used to quantify MMN amplitude.
To this aim, the MMN was first identified at Fz as the most negative peak in the
grand-average difference waveform in each condition and a 50 ms window was
chosen, centered on the peak of the MMN to each deviant. Mean amplitudes were
then measured within this latency window for each participant and for each
deviant. Maximum MMN amplitude developed between 270 and 370 ms for both
frequency and duration deviants and between 50 ms and 150 ms for VOT deviants.
Moreover, P3a was identified at Cz between 200 ms and 400 ms post-deviant onset
and N1 was identified in the ERPs to Standard, Large deviants and Small deviants at
Fz between 50 ms and 150 ms post-deviant onset. In all cases, mean amplitude
were computed for each electrode and for each subject in a 50 ms window centered
at the peak.
Five-way repeated-measures ANOVAs were first conducted on MMN and P3a
and N1 amplitude that included Group (dyslexics and normal-readers) as a
between-subjects factor, and Dimension (Frequency, Duration and VOT), Deviance
size (Small and Large), Laterality (Left: F3, C3, P3; Midlines: Fz, Cz, Pz; and Right:
F4, C4, P4) and Anterior–Posterior locus (Frontal, Central and Parietal) as within-
subject factors. If the Group by Dimension interaction was significant, four-way
ANOVAs were conducted for each Dimension separately (by including Group as a
between-subject factor and Deviance size, Laterality and Anterior–posterior locus
as within-subject factors). Greenhouse–Geisser corrections were applied when
appropriate and Tukey post-hoc tests were conducted to determine the source of
significant interactions.
3. Results
3.1. Neuropsychological and speech assessments
Results of the neuropsychological and speech assessments arereported in Tables 1, 2 and 3.
–
All children (24 Dys and 24 NR; see Table 1)The level of performance in the standardized Alouette’s read-ing test was significantly lower (po .001) for children withdyslexia, even if they were 2 years older than normal-readers(chronological age, po .001). Children with dyslexia were alsoimpaired in regular word-reading (po .001), as well as inpseudo-word- and irregular word-reading (po .001, in bothcases). No significant differences were found between the twogroups in phoneme suppression and fusion or in peudowordsrepetition. Results in the digit span test revealed a deficit inphonological memory (Direct digit span; po .001) and inworking memory (Reverse digit span; po .001) as has oftenbeen reported in phonological dyslexia (Ackerman & Dykman,1993; Helland & Asbjornsen, 2004).Other tests of the WISC IV battery (e.g., similarities andsymbols), of the PM47 and tests of the NEPSY battery did notreveal significant between-group differences, thereby showingthat children with dyslexia had verbal and non-verbal IQwithin the normal range and no specific attentional deficitscompared to normal-readers.–
Children matched on Chronological age (7 Dys-C and 7 NR-C;see Table 2)When the two groups of children were matched on chronolo-gical age (p¼ .10) and in line with the definition of dyslexiaDys-C showed lower scores than NR-C in the standardizedAlouette’s reading test (po .04) as well as in all words’ readingtests (regular words, po .001; irregular words, po .003 andpseudowords, po .001). Moreover, Dys-C performed signifi-cantly worse than NR-C in the phoneme fusion (po .01), inpseudoword repetition (po .03) as well as in the digit spantest (both direct, po .03, and reverse, po .02). By contrast, thelevel of performance of Dys-C was not significantly differentfrom NR-C in the several tests that assessed IQ and attention,and only marginally significant in naming speed errors(po .14) and phoneme suppression (po .09).–
Children matched on reading age (12 Dys-R and 12 NR-R;Table 3)When the two groups of children were matched on theirreading level (standardized Alouette reading test, p¼ .31),chronological age was significantly different (po .001) withDys-R being 2½ years older, on average, than NR-R. This agedifference can account for the higher scores of Dys-R than NR-R at some WISC-IV tests (similarities and symbols, bothpo .05) and PM47 test (po .005). Similarly, the age differencecan also explain the lack of between-group differences on digitspan (po1) and on phonological awareness tests (po1).
Table 2Neuropsychological and speech assessments in 12 children with dyslexia and 12 normal-reading children matched on reading-age (df¼(1, 22)). Significant effects
and interactions are highlighted in gray.
Table 1Neuropsychological and speech assessments in 24 children with dyslexia and 24 normal-reading children (df¼(1, 46)). Significant effects and interactions are
highlighted in gray.
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–20552048
However, Dys-R still showed lower scores than NR-R in regularword- and pseudoword-reading (both po .02), as well as invisual attention (po .02) and orientation tests (po .06) withno deficit in the other attention tests (p 4 .20).
3.2. Identification test with VOT deviants
For technical reasons, this test could only be conducted in asub-group of 12 children with dyslexia and 12 normal-readingchildren. The main effect of Group was marginally significant
(p¼ .07) with more ‘‘Ba’’ identifications in dyslexics (83%) than innormal-readers (71%). Moreover, the main effect of stimuli wassignificant [F(2,44)¼50.9; po .001]: the standard ‘‘Ba-70 ms’’ (97%)and the Small deviant ‘‘Ba�40 ms’’ (89%) were more often identi-fied as ‘‘Ba’’ than the Large deviant ‘‘Ba0 ms’’ (45%) withoutsignificant differences between dyslexics and normal-readers.However, ‘‘Ba0 ms’’ syllables (Large deviants) were more oftencategorized as ‘‘Ba’’ by children with dyslexia (60%) than bynormal-readers (30%; Tukey test: po .01; significant Group byStimulus interaction: [F(2,80)¼7.53; po .001]). By contrast, nobetween-group differences were found for the Small deviants
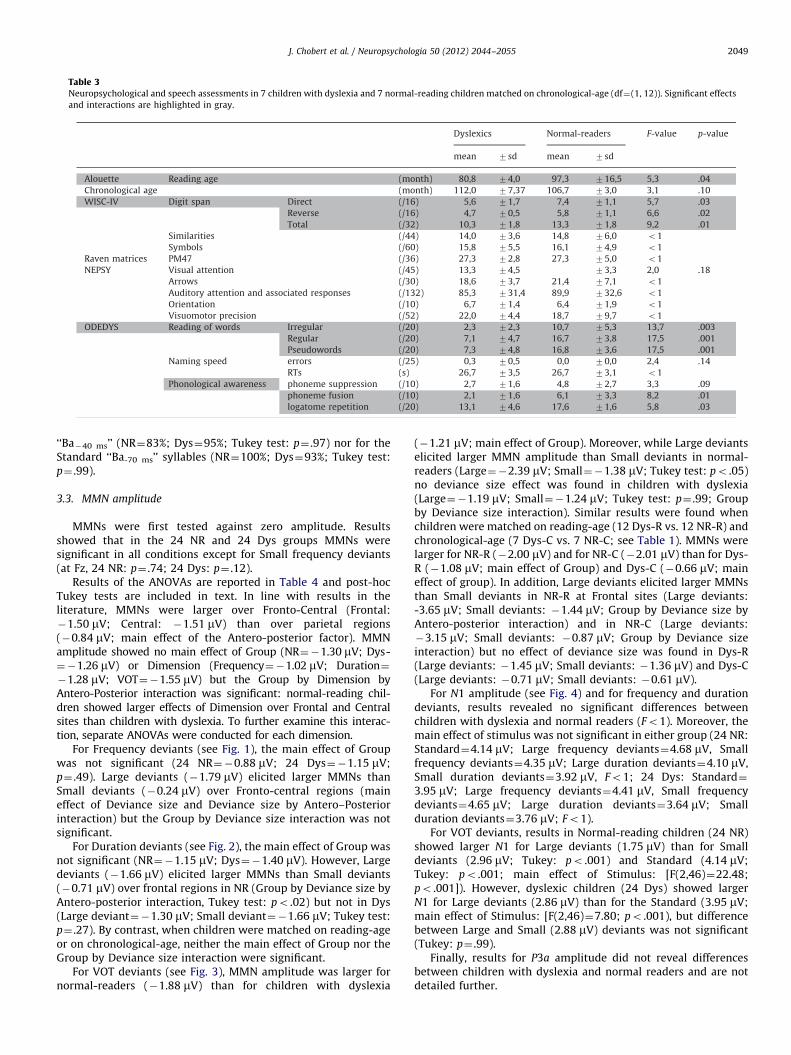
Table 3Neuropsychological and speech assessments in 7 children with dyslexia and 7 normal-reading children matched on chronological-age (df¼(1, 12)). Significant effects
and interactions are highlighted in gray.
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2049
‘‘Ba�40 ms’’ (NR¼83%; Dys¼95%; Tukey test: p¼ .97) nor for theStandard ‘‘Ba-70 ms’’ syllables (NR¼100%; Dys¼93%; Tukey test:p¼ .99).
3.3. MMN amplitude
MMNs were first tested against zero amplitude. Resultsshowed that in the 24 NR and 24 Dys groups MMNs weresignificant in all conditions except for Small frequency deviants(at Fz, 24 NR: p¼ .74; 24 Dys: p¼ .12).
Results of the ANOVAs are reported in Table 4 and post-hocTukey tests are included in text. In line with results in theliterature, MMNs were larger over Fronto-Central (Frontal:�1.50 mV; Central: �1.51 mV) than over parietal regions(�0.84 mV; main effect of the Antero-posterior factor). MMNamplitude showed no main effect of Group (NR¼�1.30 mV; Dys-¼�1.26 mV) or Dimension (Frequency¼�1.02 mV; Duration¼�1.28 mV; VOT¼�1.55 mV) but the Group by Dimension byAntero-Posterior interaction was significant: normal-reading chil-dren showed larger effects of Dimension over Frontal and Centralsites than children with dyslexia. To further examine this interac-tion, separate ANOVAs were conducted for each dimension.
For Frequency deviants (see Fig. 1), the main effect of Groupwas not significant (24 NR¼�0.88 mV; 24 Dys¼�1.15 mV;p¼ .49). Large deviants (�1.79 mV) elicited larger MMNs thanSmall deviants (�0.24 mV) over Fronto-central regions (maineffect of Deviance size and Deviance size by Antero–Posteriorinteraction) but the Group by Deviance size interaction was notsignificant.
For Duration deviants (see Fig. 2), the main effect of Group wasnot significant (NR¼�1.15 mV; Dys¼�1.40 mV). However, Largedeviants (�1.66 mV) elicited larger MMNs than Small deviants(�0.71 mV) over frontal regions in NR (Group by Deviance size byAntero-posterior interaction, Tukey test: po .02) but not in Dys(Large deviant¼�1.30 mV; Small deviant¼�1.66 mV; Tukey test:p¼ .27). By contrast, when children were matched on reading-ageor on chronological-age, neither the main effect of Group nor theGroup by Deviance size interaction were significant.
For VOT deviants (see Fig. 3), MMN amplitude was larger fornormal-readers (�1.88 mV) than for children with dyslexia
(�1.21 mV; main effect of Group). Moreover, while Large deviantselicited larger MMN amplitude than Small deviants in normal-readers (Large¼�2.39 mV; Small¼�1.38 mV; Tukey test: po .05)no deviance size effect was found in children with dyslexia(Large¼�1.19 mV; Small¼�1.24 mV; Tukey test: p¼ .99; Groupby Deviance size interaction). Similar results were found whenchildren were matched on reading-age (12 Dys-R vs. 12 NR-R) andchronological-age (7 Dys-C vs. 7 NR-C; see Table 1). MMNs werelarger for NR-R (�2.00 mV) and for NR-C (�2.01 mV) than for Dys-R (�1.08 mV; main effect of Group) and Dys-C (�0.66 mV; maineffect of group). In addition, Large deviants elicited larger MMNsthan Small deviants in NR-R at Frontal sites (Large deviants:-3.65 mV; Small deviants: �1.44 mV; Group by Deviance size byAntero-posterior interaction) and in NR-C (Large deviants:�3.15 mV; Small deviants: �0.87 mV; Group by Deviance sizeinteraction) but no effect of deviance size was found in Dys-R(Large deviants: �1.45 mV; Small deviants: �1.36 mV) and Dys-C(Large deviants: �0.71 mV; Small deviants: �0.61 mV).
For N1 amplitude (see Fig. 4) and for frequency and durationdeviants, results revealed no significant differences betweenchildren with dyslexia and normal readers (Fo1). Moreover, themain effect of stimulus was not significant in either group (24 NR:Standard¼4.14 mV; Large frequency deviants¼4.68 mV, Smallfrequency deviants¼4.35 mV; Large duration deviants¼4.10 mV,Small duration deviants¼3.92 mV, Fo1; 24 Dys: Standard¼3.95 mV; Large frequency deviants¼4.41 mV, Small frequencydeviants¼4.65 mV; Large duration deviants¼3.64 mV; Smallduration deviants¼3.76 mV; Fo1).
For VOT deviants, results in Normal-reading children (24 NR)showed larger N1 for Large deviants (1.75 mV) than for Smalldeviants (2.96 mV; Tukey: po .001) and Standard (4.14 mV;Tukey: po .001; main effect of Stimulus: [F(2,46)¼22.48;po .001]). However, dyslexic children (24 Dys) showed largerN1 for Large deviants (2.86 mV) than for the Standard (3.95 mV;main effect of Stimulus: [F(2,46)¼7.80; po .001), but differencebetween Large and Small (2.88 mV) deviants was not significant(Tukey: p¼ .99).
Finally, results for P3a amplitude did not reveal differencesbetween children with dyslexia and normal readers and are notdetailed further.
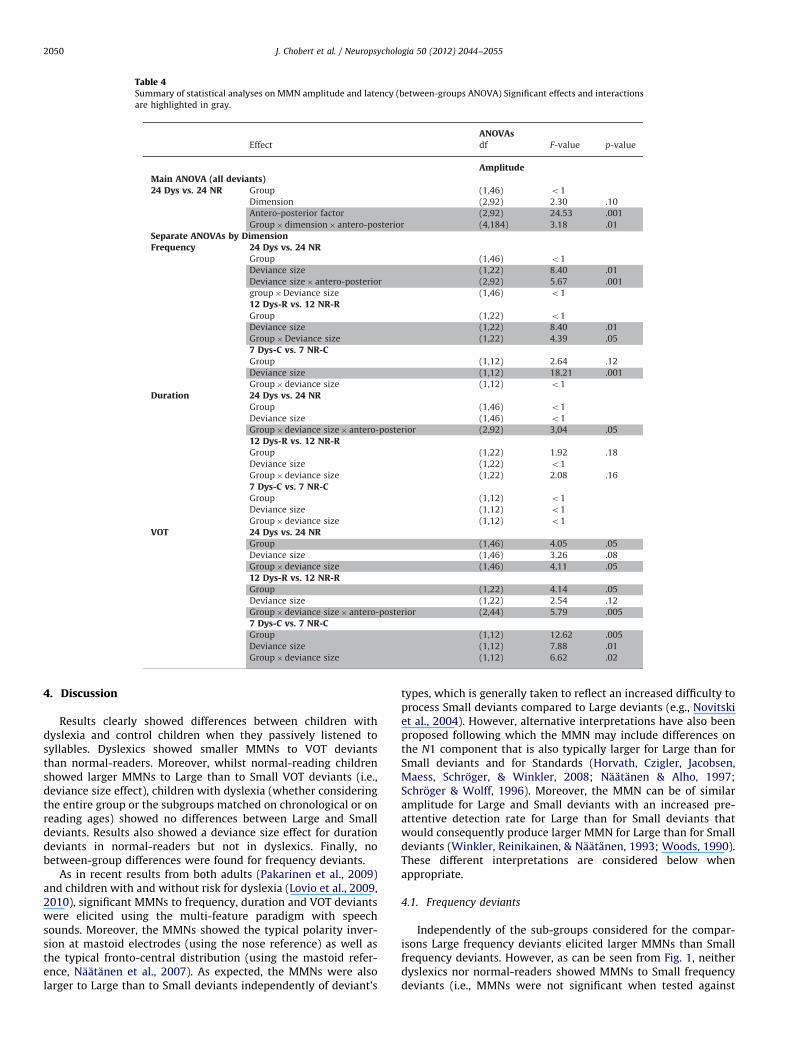
Table 4Summary of statistical analyses on MMN amplitude and latency (between-groups ANOVA) Significant effects and interactions
are highlighted in gray.
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–20552050
4. Discussion
Results clearly showed differences between children withdyslexia and control children when they passively listened tosyllables. Dyslexics showed smaller MMNs to VOT deviantsthan normal-readers. Moreover, whilst normal-reading childrenshowed larger MMNs to Large than to Small VOT deviants (i.e.,deviance size effect), children with dyslexia (whether consideringthe entire group or the subgroups matched on chronological or onreading ages) showed no differences between Large and Smalldeviants. Results also showed a deviance size effect for durationdeviants in normal-readers but not in dyslexics. Finally, nobetween-group differences were found for frequency deviants.
As in recent results from both adults (Pakarinen et al., 2009)and children with and without risk for dyslexia (Lovio et al., 2009,2010), significant MMNs to frequency, duration and VOT deviantswere elicited using the multi-feature paradigm with speechsounds. Moreover, the MMNs showed the typical polarity inver-sion at mastoid electrodes (using the nose reference) as well asthe typical fronto-central distribution (using the mastoid refer-ence, Naatanen et al., 2007). As expected, the MMNs were alsolarger to Large than to Small deviants independently of deviant’s
types, which is generally taken to reflect an increased difficulty toprocess Small deviants compared to Large deviants (e.g., Novitskiet al., 2004). However, alternative interpretations have also beenproposed following which the MMN may include differences onthe N1 component that is also typically larger for Large than forSmall deviants and for Standards (Horvath, Czigler, Jacobsen,Maess, Schroger, & Winkler, 2008; Naatanen & Alho, 1997;Schroger & Wolff, 1996). Moreover, the MMN can be of similaramplitude for Large and Small deviants with an increased pre-attentive detection rate for Large than for Small deviants thatwould consequently produce larger MMN for Large than for Smalldeviants (Winkler, Reinikainen, & Naatanen, 1993; Woods, 1990).These different interpretations are considered below whenappropriate.
4.1. Frequency deviants
Independently of the sub-groups considered for the compar-isons Large frequency deviants elicited larger MMNs than Smallfrequency deviants. However, as can be seen from Fig. 1, neitherdyslexics nor normal-readers showed MMNs to Small frequencydeviants (i.e., MMNs were not significant when tested against

Fig. 1. Difference waves (Deviant–Standard) illustrating the MMNs to Large (solid line) and to Small frequency deviants (dashed line) for dyslexics (left) and normal-
readers (right). At the Top: MMNs for 24 dyslexics and 24 normal-readers. In the middle: MMNs for 12 dyslexics and 12 normal-readers matched on reading-age. Bottom:
MMNs for 7 dyslexics and 7 normal-readers matched on chronological-age.
Fig. 2. Difference waves (Deviant–Standard) illustrating the MMNs to Large (solid line) and to Small duration deviants (dashed line) for dyslexics (left) and normal-readers
(right). At the Top: MMNs for 24 dyslexics and 24 normal-readers. In the middle: MMNs for 12 dyslexics and 12 normal-readers matched on reading-age. Bottom: MMNs
for 7 dyslexics and 7 normal-readers matched on chronological-age.
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2051
zero amplitude) probably because they were too close from theStandard to be preattentively discriminated. Most importantly,and in line with previous results in children at risk for dyslexia(Lovio et al., 2010), the preattentive processing of pitch variationsin syllables was not different between children with dyslexia andnormal-readers. Similarly, perceptual processes (as measured
through the N1 component) were not significantly differentbetween the two groups of children (see Fig. 4).
In sum, and in contrast with previous results by Santos et al.(2007) showing impaired attentive pitch processing of finalsentence words in dyslexic children, the present results did notreveal deficits in the preattentive processing of vowel-frequency

Fig. 3. Difference waves (Deviant–Standard) illustrating the MMNs to Large (solid line) and to Small VOT deviants (dashed line) for dyslexics (left) and normal-readers
(right). At the Top: MMNs for 24 dyslexics and 24 normal-readers. In the middle: MMNs for 12 dyslexics and 12 normal-readers matched on reading-age. Bottom: MMNs
for 7 dyslexics and 7 normal-readers matched on chronological-age.
Fig. 4. Event-Related Potentials (N1 and P2) at Fz electrode to Standard (black),
Large deviant (light gray) and Small deviant (dark gray) in frequency (top),
duration (middle) and VOT (bottom) for 24 children with dyslexia (left) and 24
normal-reading children (right).
J. Chobert et al. / Neuropsychologia 50 (2012) 2044–20552052
in children with dyslexia. This interesting dissociation betweenthe preattentive and attentive level of pitch processing in dyslexicchildren will need to be directly tested in further experiments.
4.2. Duration deviants
For normal readers, Large duration deviants elicited largerMMN amplitude than Small deviants (see Fig. 2). While this effectwas present in all normal-readers, it was only significant for theentire group (24) but did not reach significance in the subgroupsof 12 NR-R and the 7 NR-C probably because of lower signal-to-noise ratio. By contrast, none of the 3 groups of dyslexics showeda deviance size effect. As observed for the 24 Dys, the 12 Dys-Ralso showed a tendency for a reverse effect (larger MMNs to Smallthan to Large deviants, Fig. 2). However, careful examination ofthe individual traces revealed that this tendency was accountedfor by only three dyslexic children, thereby explaining why theeffect did not reach significance. Thus, in line with previousreports by Shankarnarayan and Maruthy (2007) and Lovio et al.(2010), and based on the statistically reliable comparisonbetween the 24 NR and the 24 Dys, the present results show thatonly normal-readers seemed to process the acoustical differencebetween Large and Small duration deviants, thereby revealing apreattentive processing deficit of syllable duration in childrenwith dyslexia. As mentioned above, differences in the amplitudeof the N1 between the Standard, Small and Large deviants canalso contribute to the MMN (Horvath et al., 2008; Naatanen &Alho, 1997; Schroger & Wolff, 1996). However, this confoundingis unlikely here since all stimuli were identical until 198 ms, thatis well after the occurrence of the N1 component. Accordingly,results revealed no deviance effect on N1 amplitude in eithergroup of children (see Fig. 4).
4.3. VOT deviants
When comparing the 24 Dys to the 24 NR children, VOTdeviants elicited larger MMN amplitude in normal-readers thanin children with dyslexia. This result is in line with previousresults of Csepe (2003) and Shankarnarayan and Maruthy (2007)and is taken to reflect impaired preattentive processing of VOT inchildren with dyslexia. Moreover, normal-readers showed larger

J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2053
MMN to Large deviants than to Small deviants (significantdeviance size effect). Importantly, this finding parallels the resultsreported in the syllable identification task showing that normal-readers identified Large deviants ‘‘Ba0ms’’ more often as a ‘‘Pa’’than as a ‘‘Ba’’ (i.e., as across category deviants) and Smalldeviants ‘‘Ba�40 ms’’ more often as ‘‘Ba’’ (i.e., as within-categorydeviants). Thus, in normal readers, syllables perceived as across-category deviants elicited larger MMNs than syllables perceivedas within-category deviants (Dehaene-Lambertz, 1997; Dehaene-Lambertz & Baillet, 1998). By contrast, and in line with ourhypothesis, the deviance size effect was not significant in childrenwith dyslexia. In this group, the MMNs to Large and Smalldeviants were not significantly different whether consideringthe entire group of dyslexics (24 Dys), the 12 Dys-R (matchedwith 12 NR-R on reading-age) or the 7 Dys-C (matched with 7NR-C on chronological-age). Insofar as the deviance-size effect wasnot significant when dyslexics and controls were matched onchronological-age, this finding does not reflect a maturationeffect. Rather, these results support the interpretation that chil-dren with dyslexia are impaired in the processing of VOTindependently of their chronological age or their reading level.Moreover, as for normal readers, it is also important to point outthat the dyslexics’ results on MMN amplitude clearly parallelresults in the syllable identification task. Children with dyslexiaidentified Small deviants ‘‘Ba�40ms’’ more often as ‘‘Ba’’ than as‘‘Pa’’ (i.e., as within category deviants), as did normal readers. Thisfinding is in line with the electrophysiological data showing thatthe MMNs to Small deviants were not different between the twogroups of children. However, in contrast to normal readers,dyslexics also identified Large deviants ‘‘Ba0ms’’ more often as‘‘Ba’’ than as ‘‘Pa’’ (i.e., again as within category deviants). Thus, indyslexics, both Large and Small deviants were more likely to beidentified as within category deviants and both elicited MMN ofsimilar amplitude, thereby explaining why the deviance sizeeffect was not significant in this group.
However, and as noted above, alternative interpretations needto be considered. In contrast to duration deviants, VOT deviantsdiffered from standards right at stimulus onset. Consequently,they were possibly associated with larger N1 components thanthe standard stimuli and the observed effect may also include anN1 effect (Horvath et al., 2008; Naatanen & Alho, 1997; Schroger& Wolff, 1996). Results of N1 amplitude analyses revealed that inNormal-reading children (24 NL) the N1 was larger for Largedeviants than for Small deviants and for Standard. By contrast, thedifference between Large and Small deviants was not significantin children with dyslexia (24 Dys; see Fig. 4). Therefore, results onN1 amplitude paralleled the results found for the MMN of asignificant deviance size effect for VOT deviants in normal readersbut not in Dyslexics. This suggests that differences in bothperceptual processing (as reflected by the N1) and in the mis-match detection processes (as reflected by the MMN) may alsocontribute to explain the differences between children withdyslexia and normal readers.
Finally, it should be noted that the rate of pre-attentivedetection was possibly higher for Large than for Small VOTdeviants each being associated with MMN of similar amplitude.In this case, the differences between Large and Small VOTdeviants in normal readers does not reflect genuine differencesin the processing of within and between category deviants but thehigher pre-attentive detection rate of across-category deviants.Following this interpretation, the finding of MMNs of similaramplitude for Large and Small VOT deviants in the dyslexic group,and for Small VOT deviants in normal readers, would reflect lowerpre-attentive detection rates for across-category deviants indyslexics. Importantly, however, whether the deviance sizeMMN effect depends upon the rate of pre-attentive detection of
VOT deviants or from Larger MMN to Large than to Smalldeviants, the most important point is that it was significant innormal readers but not in children with dyslexia.
In sum, the findings of MMNs of similar amplitude in dyslexicsand controls for Small VOT deviants but of MMNs of differentamplitude in the two groups for Large VOT deviants is in line withthe interpretation proposed by (Serniclaes, Sprenger-Charolles,Carre, and Demonet, 2001; Serniclaes et al., 2004) followingwhich children with dyslexia are more sensitive to within-category than to across-category deviants. However, our resultsdid not provide evidence that dyslexics were more sensitive thannormal-readers to within-category deviants. Moreover, the pre-sent results also highlight a clear relationship between theprocessing of VOT at the attentive level (as in discrimination oridentification tasks) and at the preattentive (as reflected by theMMN) and perceptual levels (as reflected by the N1). Finally, theyare in line with the interpretation proposed by Csepe (2003) thatimpaired preattentive processing of VOT in children with dyslexiacontributes to the phonological deficits often found in thislanguage disorder.
This interpretation is also in line with the model proposed byHickok & Poeppel (2007) following which speech perceptionrelies on the mapping of acoustic information and phonologicalrepresentations. Analysis of the acoustic (spectro-temporal) prop-erties of speech sounds in the superior temporal gyrus is thentransformed into phonological codes in the superior temporalsulcus. Thus, the deficit in processing syllabic duration, revealedby the present results in children with dyslexia, may impact onthe processing of voicing, a phonological parameter important forspeech perception. More generally, these findings fairly wellintegrate within the general hypothesis of an extended temporaldeficit in dyslexics (Habib, 2000; Goswami, 2011).
5. Conclusion
By providing evidence of two-level impairments in the proces-sing of syllables, our results lend support to the proposition thatbasic acoustic deficits contribute to poor reading abilities inphonological dyslexia (e.g., Goswami, 2011). If such deficits arecausally and developmentally linked to the build-in of phonolo-gical representations, it becomes highly plausible that the pho-nological roots of reading cannot be used as efficiently by childrenwith dyslexia than by normal readers. More specifically, integrat-ing these various aspects of basic speech perception into more orless complex patterns of temporally organized clusters thatmakes up natural speech would be necessary to reach properphonological achievement, and thereby efficient letter-to-soundmapping, which is specifically compromised in dyslexia.
Beyond these theoretical considerations, our results also havepractical implications: since intensive training of one or morespecific areas of deficit in dyslexia have been repeatedly shown toyield significant improvement (e.g., Kujala et al. 2001; Shaywitzet al. 2004), specific remediation techniques can be designed toeach individual pattern of deficits. For example, music trainingcould be one way of improving one or more of the aspects of basicspeech processing which have been examined in this paper. Forinstance, recent results by Chobert, Marie, Franc-ois, Schon andBesson (2011) have shown that musical expertise in 9-year-oldnormal-readers improves the processing of both duration andVOT variations. Thus, by improving the perception of acoustic andphonological cues in speech sounds, musical training mayincrease the perception of changes in VOT in children withdyslexia, thereby helping them to develop more robust phonolo-gical representations, and possibly reading abilities. Obviously,this area of research is at the very beginning but it is our hope

J. Chobert et al. / Neuropsychologia 50 (2012) 2044–20552054
that it will provide important results for diagnosis, treatmentevaluation, as well as for prevention purposes.
Acknowledgments
This research was supported by a grant from the ANR-Neuro(#024-01) to Mireille Besson. At the time the study was con-ducted, Julie Chobert and Clement Franc-ois were PhD studentssupported by the ANR-Neuro (#024-01). Julie Chobert is now apost-doctoral researcher supported by a grant from the Fondationde France (#00015167) to Mireille Besson. We thank Noel Nguyenand Daniele Schon for helping us prepare the stimuli, JitpanutMakchoay and Usanee Sotthiwat for helping us run part of theexperiment, Carine Verse and Amandine Dettori-Campus for run-ning the neuropsychological tests and Nia Cason for her attentivereading of the manuscript. We also thank all the children whoparticipated in this study and their parents as well as the directorsof the schools in which this research was conducted.
References
Ackerman, P. T., & Dykman, R. A. (1993). Phonological processes, confrontationalnaming, and immediate memory in dyslexia. Journal of Learning Disabilities,26(9), 597–609.
American Psychiatric Association (1994). Diagnostic and statistical manual ofmental disorders. Washington, DC: American Psychiatric Association.
Baldeweg, T., Richardson, A., Watkins, S., Foale, C., & Gruzelier, J. (1999). Impairedauditory frequency discrimination in dyslexia detected with mismatch evokedpotentials. Annals of Neurology, 45(4), 495–503.
Bishop, D. V. M. (2007). Using mismatch negativity to study central auditoryprocessing in developmental language and literacy impairments: where arewe, and where should we be going? Psychological Bulletin, 133(4), 651.
Boersma, P., & Weenink, D. (2001). Praat (version 4.0)[Computer software]. Avail-able: http.
Bogliotti, C., Serniclaes, W., Messaoud-Galusi, S., & Sprenger-Charolles, L. (2008).Discrimination of speech sounds by children with dyslexia: comparisons withchronological age and reading level controls. Journal of Experimental ChildPsychology, 101(2), 137–155.
Chobert, J., Marie, C., Franc-ois, C., Schon, D., & Besson, M. (2011). Enhanced Passiveand Active Processing of Syllables in Musician Children. Journal of CognitiveNeuroscience, 23(12), 3874–3887.
Collective expertise INSERM, C. N. D. R. S. D. I. (2007). Dyslexie, dysorthographie,dyscalculie: Bilan des donnees scientifiques. INSERM.
Corbera, S., Escera, C., & Artigas, J. (2006). Impaired duration mismatch negativityin developmental dyslexia. NeuroReport, 17(10), 1051–1055.
Csepe, V. (2003). Dyslexia: Different Brain, Different Behavior. Springer.Dehaene-Lambertz, G. (1997). Electrophysiological correlates of categorical pho-
neme perception in adults. NeuroReport, 8(4), 919.Dehaene-Lambertz, G., & Baillet, S. (1998). A phonological representation in the
infant brain. Neuroreport, 9(8), 1885.Demonet, J. F., Taylor, M. J., & Chaix, Y. (2004). Developmental dyslexia. The Lancet,
363(9419), 1451–1460.Dufor, O., Serniclaes, W., Sprenger-Charolles, L., & Demonet, J. F. (2009). Left
premotor cortex and allophonic speech perception in dyslexia: a PET study.Neuroimage, 46(1), 241–248.
Frumkin, B., & Rapin, I. (1980). Perception of vowels and consonant-vowels ofvarying duration in language impaired children. Neuropsychologia, 18(4–5),443–454.
Goswami, U. (2011). A temporal sampling framework for developmental dyslexia.Trends in cognitive sciences, 15(1), 3–10.
Habib, M. (2000). The neurological basis of developmental dyslexia. Brain, 123(12),2373.
Helland, T., & Asbjornsen, A. (2004). Digit span in dyslexia: variations according tolanguage comprehension and mathematics skills. Journal of Clinical andExperimental Neuropsychology, 26(1), 31–42.
Hickok, G., & Poeppel, D. (2007). The cortical organization of speech processing.Nature Reviews Neuroscience, 8(5), 393–402.
Horvath, J., Czigler, I., Jacobsen, T., Maess, B., Schroger, E., & Winkler, I. (2008).MMN or no MMN: No magnitude deviance effect on the MMN amplitude.Psychophysiology, 45, 60–69.
Huttunen, T., Halonen, A., Kaartinen, J., & Lyytinen, H. (2007). Does mismatchnegativity show differences in reading-disabled children compared to normalchildren and children with attention deficit?. Developmental neuropsychology,31(3), 453–470.
Huttunen-Scott, T., Kaartinen, J., Tolvanen, A., & Lyytinen, H. (2008). Mismatchnegativity (MMN) elicited by duration deviations in children with readingdisorder, attention deficit or both. International Journal of Psychophysiology,69(1), 69–77.
Jacquier-Roux, M., Valdois, S., & Zorman, M. O. (2005). Outil de Depistage desDyslexies. Grenoble: Cogni-Sciences.
Korkman, M., Kirk, U., & Kemp, S. (1998). NEPSY: A Developmental Neuropsycholo-gical Assessment. San Antonio, TX: Psychological Corporation.
Kujala, T., Lovio, R., Lepisto, T., Laasonen, M., & Naatanen, R. (2006a). Evaluation ofmulti-attribute auditory discrimination in dyslexia with the mismatch nega-tivity. Clinical Neurophysiology, 117(4), 885–893.
Kujala, T., Halmetoja, J., Naatanen, R., Alku, P., Lyytinen, H., & Sussman, E. (2006b).Speech-and sound-segmentation in dyslexia: evidence for a multiple-levelcortical impairment. European Journal of Neuroscience, 24(8), 2420–2427.
Kujala, T., Karma, K., Ceponiene, R., Belitz, S., Turkkila, P., Tervaniemi, M., et al.(2001). Plastic neural changes and reading improvement caused by audio-visual training in reading-impaired children. Proceedings of the NationalAcademy of Sciences of the United States of America, 98(18), 10509.
Kujala, T., & Naatanen, R. (2001). The mismatch negativity in evaluating centralauditory dysfunction in dyslexia. Neuroscience & Biobehavioral Reviews, 25(6),535–543.
Kujala, T., Tervaniemi, M., & Schroger, E. (2007). The mismatch negativity incognitive and clinical neuroscience: theoretical and methodological consid-erations. Biological Psychology, 74(1), 1–19.
Lachmann, T., Berti, S., Kujala, T., & Schroger, E. (2005). Diagnostic subgroups ofdevelopmental dyslexia have different deficits in neural processing of tonesand phonemes. International Journal of Psychophysiology, 56(2), 105–120.
Lallier, M., Donnadieu, S., Berger, C., & Valdois, S. (2010). A case study ofdevelopmental phonological dyslexia: is the attentional deficit in the percep-tion of rapid stimuli sequences amodal? Cortex, 46(2), 231–241, http://dx.doi.org/10.1016/j.cortex.2009.03.014.
Lefavrais, P. (1967). L’alouette. Paris: ECPA.Liberman, I. Y., & Shankweiler, D. (1985). Phonology and the problems of learning
to read and write. Remedial and Special Education, 6(6), 8.Lisker, L., & Abramson, A. S. (1967). Some effects of context on voice onset time in
English stops. Language and Speech, 10(1), 1–28.Lovio, R., Naatanen, R., & Kujala, T. (2010). Abnormal pattern of cortical speech
feature discrimination in 6-year-old children at risk for dyslexia. BrainResearch, 1335, 53–62.
Lovio, R., Pakarinen, S., Huotilainen, M., Alku, P., Silvennoinen, S., Naatanen, R.,et al. (2009). Auditory discrimination profiles of speech sound changes in6-year-old children as determined with the multi-feature MMN paradigm.Clinical Neurophysiology, 120(5), 916–921.
Lyon, G. R., Shaywitz, S. E., & Shaywitz, B. A. (2003). A definition of dyslexia. Annalsof Dyslexia, 53(1), 1–14.
Medina, V., Hoonhorst, I., Bogliotti, C., & Serniclaes, W. (2010). Development ofvoicing perception in French: Comparing adults, adolescents, and children.Journal of Phonetics, 38(4), 493–503.
Mody, M. (2003). Phonological basis in reading disability: a review and analysis ofthe evidence. Reading and Writing, 16(1), 21–39.
Naatanen, R., & Alho, K. (1997). Mismatch Negativity–the measure for centralsound representation accuracy. Audiology and Neuro-otology, 2, 341–353.
Naatanen, R., Gaillard, A. W. K., & Mantysalo, S. (1978). Early selective-attentioneffect on evoked potential reinterpreted. Acta Psychologica, 42(4), 313–329.
Naatanen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The Mismatch Negativity(MMN) in basic research of central auditory processing: a review. ClinicalNeurophysiology, 118(12), 2544–2590.
Naatanen, R., Pakarinen, S., Rinne, T., & Takegata, R. (2004). The mismatchnegativity (MMN): towards the optimal paradigm. Clinical Neurophysiology,115(1), 140–144.
Novitski, N., Tervaniemi, M., Huotilainen, M., & Naatanen, R. (2004). Frequencydiscrimination at different frequency levels as indexed by electrophysiologicaland behavioral measures. Cognitive Brain Research, 20(1), 26–36.
Pakarinen, S., Lovio, R., Huotilainen, M., Alku, P., Naatanen, R., & Kujala, T. (2009).Fast multi-feature paradigm for recording several mismatch negativities(MMNs) to phonetic and acoustic changes in speech sounds. Biologicalpsychology, 82(3), 219–226.
Ramus, F. (2003). Developmental dyslexia: specific phonological deficit or generalsensorimotor dysfunction?. Current Opinion in Neurobiology, 13(2), 212–218.
Raven, J. C., Corporation, P., & Lewis, H. K. (1962). Coloured Progressive Matrices:Sets A, AB, B. London: HK Lewis.
Renvall, H., & Hari, R. (2003). Diminished auditory mismatch fields in dyslexicadults. Annals of Neurology, 53(5), 551–557.
Sams, M., Paavilainen, P., Alho, K., & Naatanen, R. (1985). Auditory frequencydiscrimination and event-related potentials. Electroencephalography and Clin-ical Neurophysiology/Evoked Potentials Section, 62(6), 437–448.
Santos, A., Joly-Pottuz, B., Moreno, S., Habib, M., & Besson, M. (2007). Behaviouraland event-related potentials evidence for pitch discrimination deficits indyslexic children: Improvement after intensive phonic intervention. Neurop-sychologia, 45(5), 1080–1090.
Schulte-Korne, G., & Bruder, J. (2010). Clinical neurophysiology of visual andauditory processing in dyslexia: a review. Clinical Neurophysiology: OfficialJournal of the International Federation of Clinical Neurophysiology, 121(11),1794–1809.
Schulte-Korne, G., Deimel, W., Bartling, J., & Remschmidt, H. (1998). Auditoryprocessing and dyslexia: evidence for a specific speech processing deficit.Neuroreport, 9(2), 337–340.
Schulte-Korne, G., Deimel, W., Bartling, J., & Remschmidt, H. (2001). Speechperception deficit in dyslexic adults as measured by mismatch negativity(MMN). International Journal of Psychophysiology, 40(1), 77–87.

J. Chobert et al. / Neuropsychologia 50 (2012) 2044–2055 2055
Serniclaes, W. (1987). Etude experimentale de la perception du trait de voisement des
occlusives du franc-ais. Unpublished Doctoral Thesis. Universite Libre deBruxelles. Telechargeable �a /http://www.vjf.cnrs.fr/umr8606S.
Serniclaes, W., Heghe, S. V., Mousty, P., Carre, R., & Sprenger-Charolles, L. (2004).Allophonic mode of speech perception in dyslexia. Journal of Experimental Child
Psychology, 87(4), 336–361.Serniclaes, W., Sprenger-Charolles, L., Carre, R., & Demonet, J. F. (2001). Perceptual
discrimination of speech sounds in developmental dyslexia. Journal of Speech,
Language, and Hearing Research, 44(2), 384.Shankarnarayan, V. C., & Maruthy, S. (2007). Mismatch Negativity in children with
dyslexia speaking Indian languages. Behavioral and Brain Functions, 3(1), 36.Sharma, M., Purdy, S., Newall, P., Wheldall, K., Beaman, R., & Dillon, H. (2006).
Electrophysiological and behavioral evidence of auditory processing deficits inchildren with reading disorder. Clinical Neurophysiology, 117(5), 1130–1144.
Shaywitz, B. A., Shaywitz, S. E., Blachman, B. A., Pugh, K. R., Fulbright, R. K.,Skudlarski, P., et al. (2004). Development of left occipitotemporal systems for
skilled reading in children after a phonologically-based intervention. Biological
Psychiatry, 55(9), 926–933.Schroger, E., & Wolff, C. (1996). Mismatch response to changes in sound location.
NeuroReport, 7, 3005–3008.
Snowling, M. (1998). Dyslexia as a Phonological Deficit: evidence and implications.Child and Adolescent Mental Health, 3(1), 4–11.
Tallal, P., & Piercy, M. (1974). Developmental aphasia: rate of auditory processingand selective impairment of consonant perception. Neuropsychologia, 12(1),83–93.
Tallal, P. (2001). Experimental studies of language learning impairments: Fromresearch to remediation. Speech and Language Impairments in Children: Causes,Characteristics, Intervention and Outcome. Psychology Press.
Vellutino, F. R., Fletcher, J. M., Snowling, M. J., & Scanlon, D. M. (2004). Specificreading disability (dyslexia): What have we learned in the past four decades?.Journal of Child Psychology and Psychiatry, 45(1), 2–40.
Wechsler, D. (2003). Wechsler Intelligence Scale for Children (Fourth edition). SanAntonio, TX: The Psychological Corporation (WISC-IV).
Winkler, I., Reinikainen, K., & Naatanen, R. (1993). Event related brain potentialsreflect traces of the echoic memory in humans. Perception & Psychophysics, 53,443–449.
Woldorff, M. G., Hackley, S. A., & Hillyard, S. A. (1991). The effects of channel-selective attention on the Mismatch Negativity wave elicited by deviant tones.Psychophysiology, 28(1), 30–42.
Woods, D. L. (1990). Selective auditory attention: Complex processes and complexERP generators. Behavioral and Brain Sciences, 13, 260–261.
Copyright © 2022 FDOKUMEN




















