
DARPP-32 expression in rat brain after electroconvulsive stimulation
7
Research Report DARPP-32 expression in rat brain after electroconvulsive stimulation Daniela V.F. Rosa a,b , Renan P. Souza a,b , Bruno R. Souza a,b , Bernardo S. Motta a,b , Fernando Caetano a,b , Luciano K. Jornada c , Gustavo Feier c , Marcus V. Gomez b , João Quevedo c , Marco A. Romano-Silva a,b, ⁎ a Grupo de Pesquisa em Neuropsiquiatria Clínica e Molecular, Universidade Federal de Minas Gerais, Av Antonio Carlos 6627, Belo Horizonte 31270-901 Minas Gerais, Brazil b Departamento de Farmacologia, ICB, Universidade Federal de Minas Gerais, Av Antonio Carlos 6627, Belo Horizonte 31270-901 Minas Gerais, Brazil c Laboratorio de Neurociências, Programa de Pós-Graduação em Ciências da Saúde, Universidade do Extremo Sul Catarinense, 88806-000 Criciúma, SC, Brazil ARTICLE INFO ABSTRACT Article history: Accepted 17 August 2007 Available online 25 August 2007 Although electroconvulsive therapy (ECT) has been used as a treatment for mental disorder since 1930s, little progress has been made in the mechanisms underlying its therapeutic or adverse effects. The aim of this work was to analyze the expression of DARPP-32 (a protein with a central role in dopaminergic signaling) in striatum, cortex, hippocampus and cerebellum of Wistar rats subjected to acute or chronic electroconvulsive stimulation (ECS). Rats were submitted to a single stimulation (acute) or to a series of eight stimulations, applied one every 48 h (chronic). Animals were killed for collection of tissue samples at time zero, 0.5, 3, 12, 24 and 48 h after stimulation in the acute model and at the same time intervals after the last stimulation in the chronic model. Our results indicated that acute ECS produces smaller changes in the expression of DARPP-32 but, interestingly, chronic ECS increased transient expression of DARPP-32 in several time frames, in striatum and hippocampus, after the last stimulation. Results on the expression of proteins involved in signaling pathways are relevant for neuropsychiatric disorders and treatment, in particular ECT, and can contribute to shed light on the mechanisms related to therapeutic and adverse effects. © 2007 Elsevier B.V. All rights reserved. Keywords: Electroconvulsive therapy DARPP-32 Electroconvulsive stimulation Depression Schizophrenia BRAIN RESEARCH 1179 (2007) 35 – 41 ⁎ Corresponding author. Departamento de Farmacologia – ICB, Universidade Federal de Minas Gerais, Av Antonio Carlos, 6627, 31270-901 Belo Horizonte-MG, Brazil. Fax: +5531 3499 2983. E-mail address: [email protected] (M.A. Romano-Silva). Abbreviations: BDNF, brain derived neurotrophic factor; CRE, cyclic AMP responsive element; CREB, cyclic AMP response element binding protein; DA, dopamine; DARPP-32, dopamine and cAMP regulated phosphoprotein of Mr 32 kDa; D1R, dopamine receptor type 1; D2R, dopamine receptor type 2; ECS, electroconvulsive stimulation; ECT, electroconvulsive therapy; PI3K, phosphatidylinositide 3-kinase; PKA, cAMP-dependent protein kinase; PP1, protein phosphatase-1; Ser, serine; Thr, threonine; TrkB, tyrosine kinase receptor B; VEGF, vascular endothelial growth factor; 5-HTT, serotonin receptor; 5-HT 1A , serotonin receptor type 1 A ; 5-HT 2A , serotonin receptor type 2 A 0006-8993/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2007.08.043 available at www.sciencedirect.com www.elsevier.com/locate/brainres
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of DARPP-32 expression in rat brain after electroconvulsive stimulation

B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ l oca te /b ra in res
Research Report
DARPP-32 expression in rat brain afterelectroconvulsive stimulation
Daniela V.F. Rosaa,b, Renan P. Souzaa,b, Bruno R. Souzaa,b, Bernardo S. Mottaa,b,Fernando Caetanoa,b, Luciano K. Jornadac, Gustavo Feierc, Marcus V. Gomezb,João Quevedoc, Marco A. Romano-Silvaa,b,⁎aGrupo de Pesquisa em Neuropsiquiatria Clínica e Molecular, Universidade Federal de Minas Gerais, Av Antonio Carlos 6627,Belo Horizonte 31270-901 Minas Gerais, BrazilbDepartamento de Farmacologia, ICB, Universidade Federal de Minas Gerais, Av Antonio Carlos 6627,Belo Horizonte 31270-901 Minas Gerais, BrazilcLaboratorio de Neurociências, Programa de Pós-Graduação em Ciências da Saúde, Universidade do Extremo Sul Catarinense,88806-000 Criciúma, SC, Brazil
A R T I C L E I N F O
⁎ Corresponding author. Departamento de FaBelo Horizonte-MG, Brazil. Fax: +5531 3499 2
E-mail address: [email protected] (MAbbreviations: BDNF, brain derived neuro
binding protein; DA, dopamine; DARPP-32, dD2R, dopamine receptor type 2; ECS, electrocPKA, cAMP-dependent protein kinase; PP1, pvascular endothelial growth factor; 5-HTT, s
0006-8993/$ – see front matter © 2007 Elsevidoi:10.1016/j.brainres.2007.08.043
A B S T R A C T
Article history:Accepted 17 August 2007Available online 25 August 2007
Although electroconvulsive therapy (ECT) has been used as a treatment for mental disordersince 1930s, little progress has been made in the mechanisms underlying its therapeutic oradverse effects. The aim of this work was to analyze the expression of DARPP-32 (a proteinwith a central role in dopaminergic signaling) in striatum, cortex, hippocampus andcerebellum of Wistar rats subjected to acute or chronic electroconvulsive stimulation (ECS).Rats were submitted to a single stimulation (acute) or to a series of eight stimulations,applied one every 48 h (chronic). Animals were killed for collection of tissue samples at timezero, 0.5, 3, 12, 24 and 48 h after stimulation in the acutemodel and at the same time intervalsafter the last stimulation in the chronicmodel. Our results indicated that acute ECS producessmaller changes in the expression of DARPP-32 but, interestingly, chronic ECS increasedtransient expression ofDARPP-32 in several time frames, in striatumandhippocampus, afterthe last stimulation. Results on the expression of proteins involved in signaling pathways arerelevant for neuropsychiatric disorders and treatment, in particular ECT, and can contributeto shed light on the mechanisms related to therapeutic and adverse effects.
© 2007 Elsevier B.V. All rights reserved.
Keywords:Electroconvulsive therapyDARPP-32Electroconvulsive stimulationDepressionSchizophrenia
rmacologia – ICB, Universidade Federal de Minas Gerais, Av Antonio Carlos, 6627, 31270-901983..A. Romano-Silva).trophic factor; CRE, cyclic AMP responsive element; CREB, cyclic AMP response elementopamine and cAMP regulated phosphoprotein of Mr 32 kDa; D1R, dopamine receptor type 1;onvulsive stimulation; ECT, electroconvulsive therapy; PI3K, phosphatidylinositide 3-kinase;rotein phosphatase-1; Ser, serine; Thr, threonine; TrkB, tyrosine kinase receptor B; VEGF,
erotonin receptor; 5-HT1A, serotonin receptor type 1A; 5-HT2A, serotonin receptor type 2A
er B.V. All rights reserved.

36 B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
1. Introduction
Electroconvulsive therapy (ECT) has been used as a treatmentfor mental disorders since 1930s. It is defined as a medicalprocedure inwhich a brief electrical stimulus is used to inducea cerebral seizure under controlled conditions (Rosen et al.,2003). Studies about ECT have progressed rapidly over the last20 years, providing new insights into themechanismof action,improving both its acute and long-term efficacy and decreas-ing cognitive problems associatedwith this treatment. Despitea large number of hypothesis on the mechanism of action ofECT have been proposed, it remains not well established. Themain indications for ECT include depression, mania, catatoniaand schizophrenia (Abrams, 1998). Particularly in the treat-ment of severe major depression, evidences for the effective-ness of ECT are clear and convincing (The UK ECT ReviewGroup, 2003; Barichello et al., 2004).
Evidence for dopaminergic systemdysregulation in depres-sion is supported by a variety of reports, ranging from studiesof dopamine (DA) and DA-metabolite levels, to neuroimaging,histopathological and neuroendocrine studies. Specifically, anumber of reports suggest not only that depression may belinked to abnormally low dopamine levels, but also that theseverity of depression is inversely correlated to central nervoussystem DA metabolite levels (Papakostas, 2006).
DARPP-32 (dopamine and cAMP regulated phosphoproteinof Mr 32 kDa) is a cytosolic protein that is selectively enrichedinmediumspiny neurons in the neostriatum.WhenDARPP-32is phosphorylated by cAMP-dependent protein kinase (PKA)on Thr34, it is converted into a potent inhibitor of proteinphosphatase-1 (PP-1). DARPP-32 Thr34 phosphorylation leadsto an increase in the state of phosphorylation of downstreamPP1 substrates, including several neurotransmitter receptorsand voltage-gated ion channels (Greengard et al., 1999). Inaddition to Thr34, DARPP-32 is phosphorylated at Thr75 bycyclin-dependent kinase 5 (Cdk-5). DARPP-32 phosphorylatedat Thr75 inhibits PKA activity and thereby reduces the efficacyof DA signaling (Bibb et al., 1999). Svenningsson et al. (2006)showed that acute administration of fluoxetine in miceproduced changes in phosphorylation of DARPP-32 in pre-frontal cortex, hippocampus and striatum, while chronictreatment, in addition to phosphorylation changes, increasedlevels of DARPP-32 mRNA and protein. It has also beenrecently shown that the expression of DARPP-32 was down-regulated in post-mortem brains of patients with schizophre-nia (Albert et al., 2002).
Electroconvulsive stimulation (ECS) was shown to havemany effects in experimental animals and those findingscontributed toward a better understanding of the therapeuticand side effects of ECT (Newman et al., 1998). To ourknowledge, no studies have examined the effect of acute orchronic ECS onDARPP-32 expression. To demonstrate changesin the protein expression profile after acute or chronic ECS, weanalyzed DARPP-32 levels in the striatum, cortex, hippocam-pus and cerebellum of Wistar rats. Our results showed thatacute ECS induced small changes in the expression of DARPP-32 but, interestingly, chronic ECS induced a sustained increaseof the expression of DARPP-32 for several time frames after thelast stimulation.
2. Results
2.1. Effect of ECS on DARPP-32 expression in striatum
DARPP-32 expression in the striatum of rats submitted toacute and chronic ECS was examined. Acute ECS did not showchanges in expression levels of DARPP-32 during the timeanalyzed (Fig. 1A). DARPP-32 expression increased afterchronic ECS at 3, 12 and 24 h when compared to sham andtime zero groups (pb0.05). After 48 h, DARPP-32 expressionreturned to basal (Fig. 1B).
2.2. Effect of ECS on DARPP-32 expression in cortex
Twenty-four hours after acute ECS, DARPP-32 expression wasincreased compared to the sham group (pb0.05) (Fig. 2A). Nodifference was observed in DARPP-32 levels after chronic ECS(Fig. 2B).
2.3. Effect of ECS on DARPP-32 expression in hippocampus
DARPP-32 expression was not altered in the hippocampusafter acute stimulation (Fig. 3A). The greatest increase ofDARPP-32 levels (1888±180%), after chronic stimulation, wasat time zero (pb0.05) when compared to sham group. Whencompared to time zero, differences were decreased in 30 min(338±18%), 3 h (860±112%), 12 h (676±101%), 24 h (559±2%)and 48 h (1339±29%) (Fig. 3B).
2.4. Effect of ECS on DARPP-32 expression in cerebellum
Acute or chronic ECS did not show alteration in DARPP-32expression in cerebellum (Figs. 4A and B).
3. Discussion
ECS affects several brain regions, particularly hippocampus,frontal cortex, neostriatum, entorhinal cortex, temporal-parietal cortex and several monoaminergic nuclei that projectinto these areas (Fochtmann, 1994). It is known that sine waveECT can lead to memory deficits and attention/executivefunctions deterioration (Fujita et al., 2006). DARPP-32 seems tobe related to these processes (Heyser et al., 2000).
Acute ECS increased DARPP-32 levels in the cerebral cortex24 h after stimulation. In all other regions examined (striatum,hippocampus and cerebellum), no differences were observed.Chronic ECS induced more significant changes in DARPP-32expression in striatum and especially in the hippocampus. Ithas been proposed that the great benefits of ECT are derivedfrom chronic treatment and repeated sessions in the clinicalpractice (Thienhaus et al., 1990).
Guitary and Nestler (1992) demonstrated that regulation ofDARPP-32 immunoreactivity was induced by chronic admin-istration of lithium; however, it was not observed in severalother examined brain regions. Moreover, chronic administra-tion of the antidepressant imipramine or tranylcypromineproduced a similar increase in DARPP-32 levels in frontal cor-tex. Increased levels of DARPP-32 could reflect a common
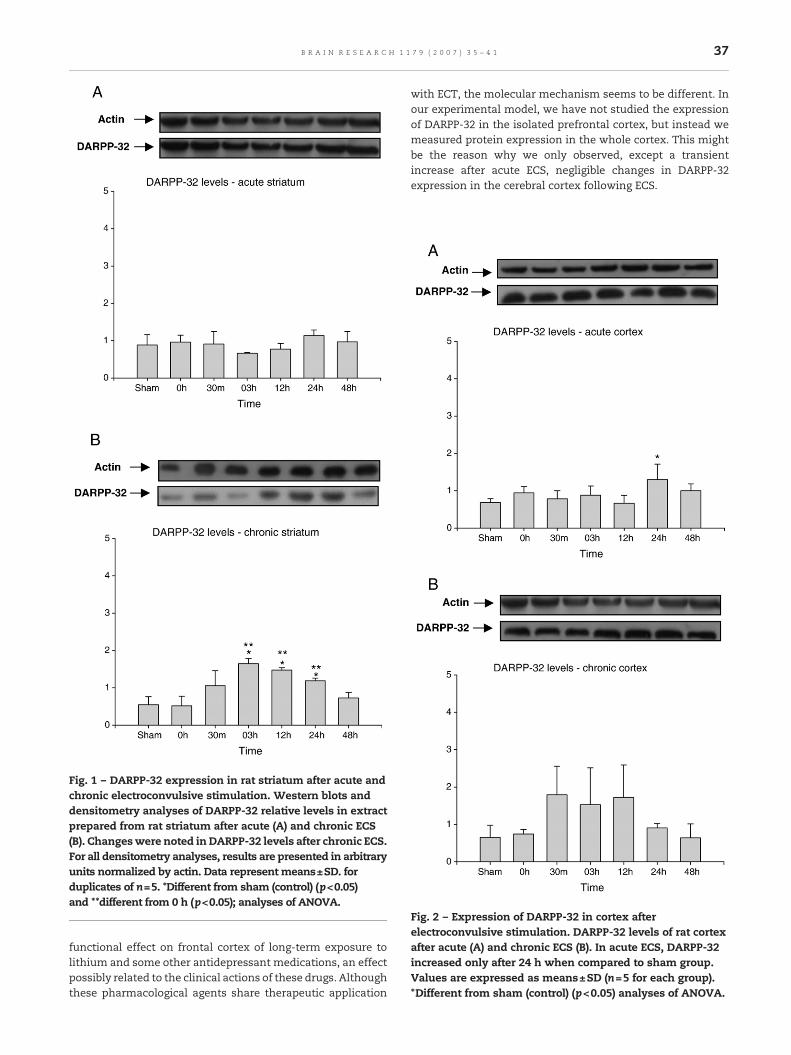
Fig. 2 – Expression of DARPP-32 in cortex afterelectroconvulsive stimulation. DARPP-32 levels of rat cortexafter acute (A) and chronic ECS (B). In acute ECS, DARPP-32increased only after 24 h when compared to sham group.Values are expressed as means±SD (n=5 for each group).*Different from sham (control) (p<0.05) analyses of ANOVA.
Fig. 1 – DARPP-32 expression in rat striatum after acute andchronic electroconvulsive stimulation. Western blots anddensitometry analyses of DARPP-32 relative levels in extractprepared from rat striatum after acute (A) and chronic ECS(B). Changeswere noted in DARPP-32 levels after chronic ECS.For all densitometry analyses, results are presented in arbitraryunits normalized by actin. Data represent means±SD. forduplicates of n=5. *Different from sham (control) (p<0.05)and **different from 0 h (p<0.05); analyses of ANOVA.
37B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
functional effect on frontal cortex of long-term exposure tolithium and some other antidepressant medications, an effectpossibly related to the clinical actions of these drugs. Althoughthese pharmacological agents share therapeutic application
with ECT, the molecular mechanism seems to be different. Inour experimental model, we have not studied the expressionof DARPP-32 in the isolated prefrontal cortex, but instead wemeasured protein expression in the whole cortex. This mightbe the reason why we only observed, except a transientincrease after acute ECS, negligible changes in DARPP-32expression in the cerebral cortex following ECS.
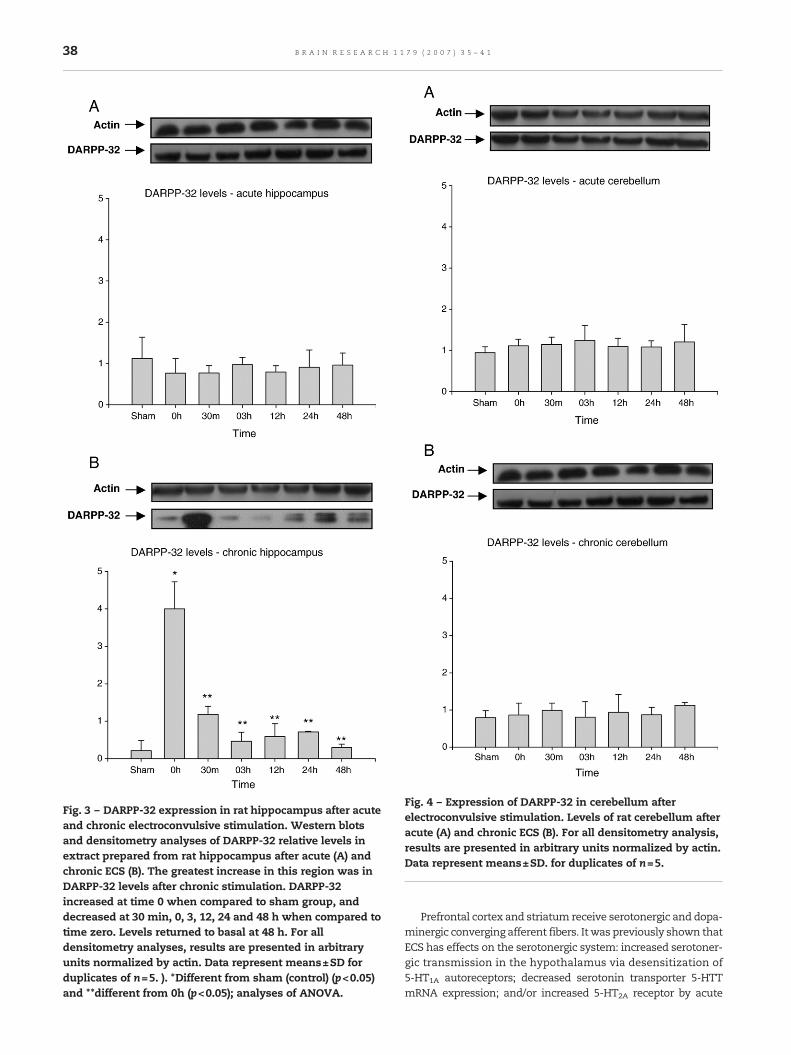
Fig. 4 – Expression of DARPP-32 in cerebellum afterelectroconvulsive stimulation. Levels of rat cerebellum afteracute (A) and chronic ECS (B). For all densitometry analysis,results are presented in arbitrary units normalized by actin.Data represent means±SD. for duplicates of n=5.
Fig. 3 – DARPP-32 expression in rat hippocampus after acuteand chronic electroconvulsive stimulation. Western blotsand densitometry analyses of DARPP-32 relative levels inextract prepared from rat hippocampus after acute (A) andchronic ECS (B). The greatest increase in this region was inDARPP-32 levels after chronic stimulation. DARPP-32increased at time 0 when compared to sham group, anddecreased at 30 min, 0, 3, 12, 24 and 48 h when compared totime zero. Levels returned to basal at 48 h. For alldensitometry analyses, results are presented in arbitraryunits normalized by actin. Data represent means±SD forduplicates of n=5. ). *Different from sham (control) (p<0.05)and **different from 0h (p<0.05); analyses of ANOVA.
38 B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
Prefrontal cortex and striatum receive serotonergic and dopa-minergic converging afferent fibers. Itwas previously shown thatECS has effects on the serotonergic system: increased serotoner-gic transmission in the hypothalamus via desensitization of5-HT1A autoreceptors; decreased serotonin transporter 5-HTTmRNA expression; and/or increased 5-HT2A receptor by acute

39B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
and/or chronic ECS protocols. These data could partially explainthe efficiency of ECT on medication-resistant depression (Dre-mencov et al., 2002; Gur et al., 2002; Shen et al., 2001). In addition,regulation of DARPP-32 phosphorylation induced by serotoninwasalsopreviously shown (Svenningssonet al., 2002),whichwasmediated primarily via activation of 5-HT4 and 5-HT6, regulatingDARPP-32 phosphorylation at Thr34 and Thr75, whereas regula-tion at Ser137 was mediated primarily by 5-HT2 receptors. Thesethree pathways appeared to inhibit PP1 through synergisticmechanisms.
Yoshida et al. (1998, 1997) investigated the effect ofrepeated ECS on extracellular concentrations of DA in thefrontal cortex of rats, using in vivomicrodialysis, and conclud-ed that the ECS effect was greater in the striatum than in thefrontal cortex. Our results showed transient increase ofDARPP-32 expression in striatum after chronic ECS. It hasbeen reported that dopaminergic pathways are able toenhance DARPP-32 expression and Nomikos et al. (1991)showed interstitial striatal DA elevation up to 1310% ofbaseline when ECS was administered. In cultured embryonicmouse striatal cells, dopamine was shown to regulate theexpression of BDNF mRNA and protein through activation ofD1 receptor (Kuppers and Beyer, 2001). BDNF belongs to theneurotrophin family and interacts with the high-affinitytyrosine kinase receptor B (TrkB). Previous studies haveshown that both acute and chronic ECS enhance the ex-pression of BDNF and TrkB in rat brain (Nibuya et al., 1995;Zetterstrom et al., 1998; Conti et al., 2007; Ploski et al., 2006).Furthermore, functional cAMP-responsive element (CRE) sitewas very recently identified in the promoter III of humanBDNF, which participates in dopamine modulation of BDNFexpression induced via D1R (Nibuya et al., 1996; Fang et al.,2003) and it has also been shown that CREB and its phosphor-ylated state were increased after ECS (Yagasaki et al., 2006).
Chronic ECS induces significant alterations in hippocam-pus. Zis et al. (1992) showed an increase in homovanilic acid, aDA metabolite, in hippocampus. Interestingly, seizure activityinduced by the convulsant agent flurothyl did not influencehippocampusdialysateDAconcentrations, suggesting that theECS-induced DA release was related to the passage of currentand not to the seizure activity (Zis et al., 1991). Convergingevidence point to hippocampal neurogenesis as an importantfactor in the pathophysiology of depression and as target ofantidepressants (Dranovsky and Hen, 2006). ECS has beenshown to have robust effects in rat hippocampus trophicfactors such as the induction of BDNF/TrkB/MAP kinasepathway, VEGF and other regulators of neurogenesis andangiogenesis (Newton et al., 2003, 2006; Altar et al., 2004). Thus,the ECS-dependent induction of DARPP-32 could be mediatedby activation of PI3K via BDNF acting through its tyrosinekinase receptor (TrkB) (Stroppolo et al., 2001). Such regulationof DARPP-32 expression was also found in other normal andneoplastic tissues (El-Rifai et al., 2002; Belkhiri et al., 2005;Garcia-Jimenez et al., 2005; Hansen et al., 2006; Souza et al.,2006a,b), linking the expression and phosphorylation ofDARPP-32 to cell survival and proliferation.
We observed marked changes in DARPP-32 expression inthe hippocampus after chronic ECS, with a high increase justafter the last session of ECS. Although the expression levelsare kept significantly higher when compared to sham shock,
the difference between 0 and 30 min was striking. We haveruled out possible artifacts since the data were obtained fromfive different animals with small variability among them, asshown by the standard deviation. Although there is nosupporting data in the literature regarding synthesis andturnover of DARPP-32, we could speculate that application ofthe last session on a animal that was already primed by theanterior sessions would produce a peak due to the presence oflarger amounts of mRNA; however, the turnover machinerywould quickly reduce DARPP-32 to levels that are still higherthan those observed in sham shock animals. We areperforming experiments involving parallel mRNA and proteinquantification in order to further clarify that observation.
Although cerebellum has been classically identified as aregion involved in motor coordination there is increasingevidence that it plays an important role in higher cognitivefunctions such as memory, learning and attention (Leineret al., 1989). Our data showed that ECS was not able to producechanges in DARPP-32 protein levels in cerebellum.
In conclusion, our results showed that ECS was able tochange transient expression of DARPP-32, especially in thehippocampus, which has a strategic role in signaling path-ways relevant to the pathophysiology and treatment of sever-al psychiatric disorders, where the use of ECT is evidence-based. The induction of DARPP-32 affects several differentdownstream pathways such as those involving receptors, ionchannels and transcription factors (Greengard, 2001). Furtherexperiments are needed to shed more light on the role ofDARPP-32 and other mechanisms linked to signaling path-ways related to the biological effects of neuropsychiatrictreatments such as ECT.
4. Experimental procedures
4.1. Animals and study design
Adult male Wistar rats (250–300 g) were caged in groups offive with food and water ad libitum and were maintained on a12 h-light-dark cycle (lights on at 7 AM), at a temperature of23±1 °C. Experiments were performed between 2 and 5 PM.The rats were divided into two groups: acute and chronictreatment. In the acute treatment, animals received a singleECS. In the chronic treatment group, animals received eightECS every other day. In both groups, they were killed bydecapitation at different time points after the last ECS, i.e.,time 0, 0.5, 3, 12, 24 and 48 h after stimulation (n=5 animalsper group). The hippocampus, striatum, total cortex andcerebellum were immediately dissected out after decapita-tion and stored at −80 °C for posterior analysis. In vivostudies were performed in accordance with the NationalInstitutes of Health guidelines and with the approval ofthe Ethics Committee from Universidade do Extremo SulCatarinense.
4.2. Electroconvulsive stimulation
ECSwas applied via bilateral ear clip electrodes (Barichello et al.,2004). The stimulus parameters were 150 V, 60 Hz, sine wave,during 2 s. Each stimulation elicited tonic–clonic seizures. The

40 B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
shamgroups (acute andchronic)werehandled identically to theECS-treated rats except no current was applied. This protocolclosely resembles ECT in clinical practice, either in number ofsessions or in stimulation intensity.
4.3. Preparation of tissue sample
Frozen tissue samples of different brain areas were soni-cated in lysis buffer (20 mM Tris, pH 8.0, 137 mM NaCl, 0.5 mMsodium orthovanadate, 2 mM okadaic acid, 10% glycerol, 1%Nonidet P40, 2% protease inhibitor) for 90 s (1 s on, 9 s off) andwere kept on ice throughout the procedure until centri-fugation at 13,000×g for 20 min at 4 °C. Supernatants weretransferred to plastic tubes and protein was quantified(Bradford, 1976).
4.4. Immunoblotting
Equal amounts of protein (100 μg) were separated by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE15%) and transferred to nitrocellulosemembranes (Towbin et al.,1979). The membranes were sequentially immunoblotted usingantibodies raised against DARPP-32 (H-62, 1:500 dilution, SantaCruz Biotechnology) and actin (1:2000 dilution, Chemicon Inter-national,) for 2 h.
Antibody binding was revealed by incubation with a goatanti-rabbit horseradish peroxidase-linked IgG (1:20000 dilu-tion, Molecular Probes,) or a goat anti-mouse horseradishperoxidase-linked IgG (1:7000 dilution, Molecular Probes) for 1h, and the ECL immunoblotting detection system (AmershamBiosciences). Chemiluminescense was detected by autoradi-ography using Kodak autoradiography film and apparentbands of DARPP-32 and actin were quantified by densitometryand analyzed using Scion Image version Beta 4.0.2 (ScionImage software Frederick, MO, USA).
Samples from the sham group and groups exposed to ECSwere run on the same immunoblots and then analyzed together.For each experiment, values obtained for DARPP-32 were cor-rected using actin values as a reference. Normalized data frommultiple experiments were averaged and statistical analysis wascarried out as described in the figure legends.
4.5. Statistical analysis
Data were analyzed by analysis of variance (ANOVA). Values areexpressed asmean±SD (n=5 for each group). The sham (control)group was handled identical to the ECS-treated rats except thatno current was applied. Differences were considered significantwhen pb0.05.
Acknowledgments
This research was supported by grants from CNPq, FAPESC,Instituto Cérebro e Mente and UNESC to J. Quevedo; MVG andMAR-S are CNPq research fellows and RPS, BSM and FSC areholders of CNPq studentships andDVFR and BRS are holders ofCAPES studentships. Financial support from CNPq Universalgrant proc. #471837/2004-0, Programa Institutos do Milênio/CNPq/FINEP and FAPEMIG # CBB-453/04.
R E F E R E N C E S
Abrams, R., 1998. Electroconvulsive Therapy. Oxford UniversityPress, Oxford.
Altar, C.A., Laeng, P., Jurata, L.W., Brockman, J.A., Lemire, A.,Bullard, J., Bukhman, Y.V., Young, T.A., Charles, V., Palfreyman,M.G., 2004. Electroconvulsive seizures regulate gene expressionof distinct neurotrophic signaling pathways. J. Neurosci. 17(24 (11)), 2667–2677.
Albert, K.A., Hemmings Jr., H.C., Adamo, A.I., Potkin, S.G.,Akbarian, S., Sandman, C.A., Cotman, C.W., Bunney, W.E.,Greengard, P., 2002. Evidence for decreased DARPP-32 in theprefrontal cortex of patients with schizophrenia. Arch. Gen.Psychiatry 59, 705–712.
Barichello, T., Bonatto, F., Agostinho, F.R., Reinke, A., Moreira, J.C.,Dal-Pizzol, F., Izquierdo, I., Quevedo, J., 2004. Structure-relatedoxidative damage in rat brain after acute and chronicelectroshock. Neurochem. Res. 29, 1749–1753.
Belkhiri, A., Zaika, A., Pidkovka, N., Knuutila, S., Moskaluk, C.,El-Rifai, W., 2005. Darpp-32: a novel antiapoptotic gene inupper gastrointestinal carcinomas. Cancer Res. 65, 6583–6592.
Bibb, J.A., Snyder, G.L., Nishi, A., Yan, Z., Meijer, L., Fienberg, A.A.,Tsai, L.H., Kwon, Y.T., Girault, J.A., Czernik, A.J., Huganir, R.L.,Hemmings Jr., H.C., Nairn, A.C., Greengard, P., 1999.Phosphorylation of DARPP-32 by Cdk5 modulates dopaminesignalling in neurons. Nature 402, 669–671.
Bradford, M.M., 1976. A rapid and sensitive method for thequantitation of microgram quantities of protein utilizing theprinciple of protein-dye binding. Anal. Biochem. 72, 248–254.
Conti, B., Maier, R., Barr, A.M., Morale, M.C., Lu, X., Sanna, P.P.,Bilbe, G., Hoyer, D., Bartfai, T., 2007. Region-specifictranscriptional changes following the three antidepressanttreatments electro convulsive therapy, sleep deprivation andfluoxetine. Mol. Psychiatry 12, 167–189.
Dranovsky, A., Hen, R., 2006. Hippocampal neurogenesis:regulation by stress and antidepressants. Biol. Psychiatry 59,1136–1143.
Dremencov, E., Gur, E., Lerer, B., Newman, M.E., 2002.Effects of chronic antidepressants and electroconvulsive shockon serotonergic neurotransmission in the rat hypothalamus.Prog. Neuro-Psychopharmacol. Biol. Psychiatry 26,1029–1034.
El-Rifai, W., Smith Jr., M.F., Li, G., Beckler, A., Carl, V.S.,Montgomery, E., Knuutila, S., Moskaluk, C.A., Frierson, H.F.,Powell, S.M., 2002. Gastric cancers overexpress DARPP-32 and anovel isoform, t-DARPP. Cancer Res. 62, 4061–4064.
Fang, H., Chartier, J., Sodja, C., Desbois, A., Ribecco-Lutkiewicz, M.,Walker, P.R., Sikorska, M., 2003. Transcriptional activation ofthe human brain-derived neurotrophic factor gene promoter IIIby dopamine signaling in NT2/N neurons. J. Biol. Chem. 278,26401–26409.
Fochtmann, L.J., 1994. Animal studies of electroconvulsivetherapy: foundations for future research. Psychopharmacol.Bull. 30, 321–444.
Fujita, A., Nakaaki, S., Segawa, K., Azuma, H., Sato, K., Arahata, K.,Otsuki, K., Hori, M., Mochida, Y., Uchida, M., Yamada, T.,Nakamura, C., Akechi, T., Furukawa, T.A., 2006. Memory,attention, and executive functions before and after sine andpulse wave electroconvulsive therapies for treatment-resistantmajor depression. J. ECT 22 (2), 107–112.
Garcia-Jimenez, C., Zaballos, M.A., Santisteban, P., 2005. DARPP-32(dopamine and 3',5'-cyclic adenosine monophosphate-regulated neuronal phosphoprotein) is essential for themaintenance of thyroid differentiation. Mol. Endocrinol. 19,3060–3072.
Greengard, P., 2001. The neurobiology of dopamine signaling.Biosci. rep. 21 (3), 247–269.
Greengard, P., Allen, P.B., Nairn, A.C., 1999. Beyond the dopamine

41B R A I N R E S E A R C H 1 1 7 9 ( 2 0 0 7 ) 3 5 – 4 1
receptor: the DARPP-32/Protein phosphatase-1 cascadeNeuron 23, 435–447.
Guitary, X., Nestler, E.J., 1992. Chronic administration of lithium orother antidepressants increases levels of DARPP-32 in ratfrontal cortex. J. Neurochem. 59, 1164–1167.
Gur, E., Dremencov, E., Garcia, F., van de Kar, L.D., Lerer, B.,Newman, M.E., 2002. Functional effects of chronicelectroconvulsive shock on serotonergic 5-HT(1A) and 5-HT(1B)receptor activity in rat hippocampus and hypothalamusBrain Res. 952, 52–60.
Hansen, C., Greengard, P., Nairn, A.C., Andersson, T., Vogel, W.F.,2006. Phosphorylation of DARPP-32 regulates breast cancer cellmigration downstream of the receptor tyrosine kinase DDR1.Exp. Cell Res. 312, 4011–4018.
Heyser, C.J., Fienberg, A.A., Greengard, P., Gold, L.H., 2000. DARPP-32knockout mice exhibit impaired reversal learning in adiscriminated operant task. Brain Res. 867 (1–2), 122–130.
Kuppers, E., Beyer, C, 2001. Dopamine regulates brain-derivedneurotrophic factor (BDNF) expression in cultured embryonicmouse striatal cells. Neuroreport 12, 1175–1179.
Leiner, H.C., Leiner, A.L., Dow, R.S., 1989. Reappraising thecerebellum: what does the hindbrain contribute to theforebrain? Behav. Neurosci. 103 (5), 998–1008.
Newman, M.E., Gur, B., Shapira Lerer, B., 1998. Neurochemicalmechanism of action of ECS: evidence from in vivo studies.J. ECT 14, 151–171.
Newton, S.S., Collier, E.F., Hunsberger, J., Adams, D., Terwilliger, R.,Selvanayagam, E., Duman, R.S., 2003. Gene profile ofelectroconvulsive seizures: induction of neurotrophic andangiogenic factors. J. Neurosci. 26 (23 (34)), 10841–10851.
Newton, S.S., Girgent, M.J., Collier, E.F., Duman, R.S., 2006.Electroconvulsive seizure increases adult hippocampalangiogenesis in rats. Eur. J. Neurosci. 24 (3), 819–828.
Nibuya, M., Morinobu, S., Duman, R.S., 1995. Regulation of BDNF andtrkB mRNA in rat brain by chronic electroconvulsive seizure andantidepressant drug treatments. J. Neurosci. 15, 7539–7547.
Nibuya, M., Nestler, E.J., Duman, R.S., 1996. Chronic antidepressantadministration increases the expression of cAMP responseelement binding protein (CREB) in rat hippocampus.J. Neurosci. 16, 2365–2372.
Nomikos, G.G., Zia, A.P., Damsma, G., Fibiger, H.C., 1991.Electroconvulsive shock produces large increases in interstitialconcentrations of dopamine in the rat striatum: an in vivomicrodialysis study. Neuropsychopharmacology 4, 65–69.
Papakostas, G.I., 2006. Dopaminergic-based pharmacotherapiesfor depression. Eur. Neuropsychopharmacol. 16, 391–402.
Ploski, J.E., Newton, S.S., Duman, R.S., 2006. Electroconvulsiveseizure-induced gene expression profile of the hippocampusdentate gyrus granule cell layer. J. Neurochem. 99, 1122–1132.
Rosen, Y., Resinik, I., Sluvis, A., Kaplan, D., Mester, R., 2003. Thesignificance of the nitric oxide in electro-convulsive therapy: aproposed neurophysiological mechanism.Med. Hypotheses 60,424–429.
Shen, H., Numachi, Y., Yoshida, S., Toda, S., Awata, S., Matsuoka,H., Sato, M., 2001. Electroconvulsive shock regulates serotonintransporter mRNA expression in rat raphe nucleus. PsychiatryClin. Neurosci. 55, 75–77.
Souza, B.R., Souza, R.P., Rosa, D.V.F., Guimarães, M.M., Correa, H.,Romano-Silva, M.A., 2006a. Dopaminergic intracellular signalintegrating proteins: relevance to schizophrenia. DialoguesClin. Neurosci. 7 (4), 95–100.
Souza, R.P., Rosa, D.V., Souza, B.R., Romano-Silva, M.A., 2006b.Darpp32. AfCS-Nature Molecule Pages. doi:10.1038/mp.a000752.01.
Stroppolo, A., Guinea, B., Tian, C., Sommer, J., Ehrlich, M.E., 2001.Role of phosphatidylinositide 3-kinase in brain-derivedneurotrophic factor-induced DARPP-32 expression in mediumsize spiny neurons in vitro. J. Neurochem. 79, 1027–1032.
Svenningsson, P., Tzavara, E.T., Liu, F., Fienberg, A.A., Nomikos, G.G.,Greengard, P., 2002. DARPP-32 mediates serotonergicneurotransmission in the forebrain. Proc. Natl. Acad. Sci. U. S. A.99, 3188–3193.
Svenningsson, P., Chergui, K., Rachleff, I., Flajolet, M., Zhang, X.,Yaacoubi, M., Vaugeois, J.M., Nomikos, G., Greengard, P., 2006.Alterations in 5-HT1B receptor function by p11 indepression-like states. Science 311, 77–80.
The UK ECT Review Group, 2003. Efficacy and safety ofelectroconvulsive therapy in depressive disorders: asystematic review and meta-analysis. The Lancet 361,799–808.
Thienhaus, O.J., Margletta, S., Bennet, J.A., 1990. A study of theclinical efficacy of maintenance ECT. J. Clin. Psychiatry 51,141–144.
Towbin, H., Staehelin, T., Gordon, J., 1979. Electrophoretic transferof proteins from polyacrylamide gels to nitrocellulose sheets:procedure and some applications. Proc. Natl. Acad. Sci. U. S. A.76, 4350–4354.
Yagasaki, Y., Numakawa, T., Kumamaru, E., Hayashi, T., Su, T.,Kunugi, H., 2006. Chronic antidepressants potentiate viasigma-1 receptors the brain-derived neurotrophicfactor-induced signaling for glutamate release. J. Biol. Chem.281, 12941–12949.
Yoshida, K., Higushi, H., Kamata, M., Yoshimoto, M., Shimizu, T.,Hishikawa, Y., 1997. Dopamine releasing response in ratstriatum to single and repeated electroconvulsive shocktreatment. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 21,707–715.
Yoshida, K., Higushi, H., Kamata, M., Yoshimoto, M., Shimizu, T.,Hishikawa, Y., 1998. Single and repeated electroconvulsiveshocks activate dopaminergic and 5-hydroytryptaminergicneurotransmission in the frontal cortex of rats. Prog.Neuro-Psychopharmacol. Biol. Psychiatry 22, 435–444.
Zetterstrom, T.S., Pei, Q., Grahame-Smith, D.G., 1998. Repeatedelectroconvulsive shock extends the duration of enhancedgene expression for BDNF in rat brain compared with a singleadministration. Brain Res. Mol. Brain Res. 57, 106–110.
Zis, A.P, Nomikos, G.G., Damsma, G., Fibiger, H.C., 1991. In vivoneurochemical effects of electroconvulsive shock studied bymicrodialysis in the rat striatum. Psychopharmacology 103,343–350.
Zis, A.P., Nomikos, G.G., Brown, E.E., Damsma, G., Fibiger, H.C.,1992. Neurochemical effects of electrically and chemicallyinduced seizures: an in vivo microdialysis study in the rathippocampus. Neuropsychopharmacology 7, 189–195.




















