
Đa dạng sinh học cùa vi khuẩn kỵ khỉ ôxy hóa Fe(II), khử - VNUF
93
ĐẠI HỌC QUỐC GIA HÀ NỘI BÁO CÁO KẾT QUẢ THỰC HIỆN ĐỀ TÀI KHCN ĐẶC BIỆT CẮP ĐHQG Tên đề tài: Đa dạng sinh học cùa vi khuẩn kỵ khỉ ôxy hóa Fe(II), khử nitrate trong một số môi trường sình thái và khả năng ứng dụng của chủng Mã sổ: QG. 07. 23 Chủ trì đề tài: TS. Đinh Thúy Hằng Cơ quan: Viện Vi sinh vật & Công nghệ Sinh học - ĐHQGHN ' HỌC Quoc GIA HA NỌI tNJG TÁM THỎNG TIN THƯ VIỆN o ự ẹ i u Hà Nội, tháng ỉ năm 20ỉ 0
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Đa dạng sinh học cùa vi khuẩn kỵ khỉ ôxy hóa Fe(II), khử - VNUF

ĐẠI HỌC QUỐC GIA HÀ NỘI
BÁO CÁO KẾT QUẢ THỰC HIỆN
ĐỀ TÀI KHCN ĐẶC BIỆT CẮP ĐHQG
Tên đề tài: Đa dạng sinh học cùa vi khuẩn kỵ khỉ ôxy hóa Fe(II), khử nitrate
trong một số môi trường sình thái và khả năng ứng dụng của chủng
Mã sổ: QG. 07. 23
Chủ trì đề tài: TS. Đinh Thúy Hằng
Cơ quan: Viện Vi sinh vật & Công nghệ Sinh học - ĐHQGHN
' H Ọ C Quoc GIA HA NỌI tNJG TÁM THỎNG TIN THƯ VIỆN
o ự ẹ i u
Hà Nội, tháng ỉ năm 20ỉ 0

BÁO CÁO KÉT QUẢ THỰC H Ệ N ĐỀ TÀI KHCN ĐẶC BIỆT CÁP DHQG
BÁO CÁO TÓM TẮT1. Tên đề tài: Đa dạng sinh học cùa vỉ khuẩn kỵ khí ôxy hoả Fe(II), khử nitrate trongmột số môi
trường sinh thái và khả nâng ứng dụng của chúng.2. Mã số: QG.07.233. Thời gian thực biện: 2 năm (2007 - 2009)4. Cấp quản lý: Đại học Quốc gia Hà nội5. Chủ trì đề tài: TS. Đinh Thuý Hằng
Viện Vi sinh vật và Công nghệ sinh học - ĐHQGHNĐiện thoại: 37547694; Fax: 37547407; Email: [email protected]
6 . Cán bộ tham gia: CN. Nguyễn Thị TuyềnThS. Nguyễn Minh Giảng
7. Mục tỉêu và nội dung nghiên cứuMục tiêu:- Nghiên cứu vả so sánh đa dạng sinh học của vi khuẩn ôxy hoá Fe2+/khử NO3- tại một số môi
trường sinh thái khác nhau.- Phân lập các nhóm vi khuẩn đại diện, chiếm đa số trong mỗi loại môi trường sinh thái và
nghiên cứu các đặc điểm phân loại, sinh lý, sinh hoá của chúng.- Nghiên cứu khả năng ứng dụng các chủng vi khuẩn phân lập được vào việc kết hợp loại bỏ
Fe2+, Mn2+, NO3” trong môi trường ô nhiễm.Nội dung nghiền cửu:- Mau trầm tích ở đáy hồ, mẫu đất ở ruộng lúa ngập nước và mẫu trầm tích ven biển sẽ được
lấy theo yêu cầu đảm bảo kỵ khí (tránh tiếp xúc với ôxy), đưa về phòng thí nghiệm và giữ ở 4°c cho đến khi tiến hành phân tích vi sinh vật.
- Số lượng vi khuẩn ôxy hoá Fe(II)/khử NO3- sẽ được xác định theo phương pháp MPN (Most
Propable Number) trong môi trường dịch thể chứa Fe(II) làm nguồn điện tử và NO3- là chẩt nhận điện tử trong điều kiện kỵ khí hoàn toàn. Thành phần khoáng của môi trường đàm bào tương ứng với môi trường sinh thái tại địa điểm lấy mẫu.
- Đa dạng về thành phần loài ttong các mẫu khác nhau sẽ được xác định thông qua phương pháp sinh học phân tử, tách DNA tổng số trục tiếp từ các lô thí nghiệm ở nồng độ pha loãng khác nhau trong dãy MPN và phân tích tính đa dạng cùa một đoạn gen 16S rARN cùa tập đoàn vi sinh vật thu được trong mỗi !ô bằng phưomg pháp PCR/DGGE.
- Các chùng vi khuẩn ôxy hoả Fe(ÍI)/khừ NCV đại diện, chiếm đa số tại mỗi địa điểm lấy mẫusẽ được phàn lập từ ống pha loãng cao nhất có vi sinh vật phát triển cùa dăy MPN bằngphương pháp ống thạch bán lòng.
- Các chùng vi khuẩn phân lập sẽ được nghiên cứu về đặc điểm sinh lý sinh hoá và phân loại dựa trên trình tự 16S rDNA. Ket quả cùa nghiên cứu này sẽ làm cơ sở để tuyển chọn các

chùng cỏ hoạt tính sinh học (khả năng chuyển hóa Fe(II) và NO3- ) cao và nghiên cứu khả
năng ứng dụng của chúng vào việc kết hợp loại bỏ Fe(II), Mn(II), NO3- trong môi trường ô nhiễm.
8. Các kết quả đạt được
- Ba dạng môi trường khác nhau gồm có ao nước ngọt, ruộng lúa ngập nước và trầm tích ven biển được chọn để tiến hành nghiên cứu sự đa dạng của vi khuẩn ôxy hoá Fe(II) khử NO3- . Đây là những dạng môi trường trong đó vi khuẩn tham gia chu trình chuyển hoá sắt và nitơ đóng vai trò quan trọng. Mầu bùn đáy (dưới 10 cm bề mặt bùn) tại các môi trường sinh thái kể trên được thu thập và sử dụng để phân tích nhóm vi khuẩn kỵ khí này.
Sổ lượng vi khuẩn ôxy hoá Fe(II), khử NO3- đã được xác định bằng phương pháp MPN trong môi trường dịch thể nước ngọt (đối với mẫu ao và mẫu ruộng lúa) hoậc nước lợ (đối với mẫu trầm tích ven biển) chứa Fe(II) làm chất cho điện tử, NO3 ' làm chất nhận điện tử và acetate/C0 2 làm nguồn cacbon. Sự sinh trưởng của vi khuẩn trong các ống MPN được nhận biết thông qua thay đổi màu sắc của môi trường nuôi cấy sang màu vàng nâu do Fe(II) bị ôxy hoá thành Fe(III).
- DNA tổng số từ các ống MPN đại diện có vi sinh vật phát triển được tách chiết và sử dụng làm khuôn trong phân ứng PCR khuyếch đại đoạn gen I6 S rDNA có độ dài 550 bp. Phân tích điện đi biến tính đoạn gen trên cho thấy tính đa dạng khá cao của vi khuẩn trong các ống MPN. Một số băng điện di đại diện được cắt và thôi gel để đọc trình tự, qua đó đã xác định được các nhóm vi khuẩn chiếm ưu thế trong các môi trường nghiên cứu, cụ thể là Anaeromyxobacter, Pseudomonas và Paracoccus.
- Đã tiến hành nghiên cứu tính đa dạng của vi sinh vật sinh trưởng trong điều kiện oxy hoả Fe(II) khử NO3- trong các dãy MPN thông qua phương pháp lai in situ sử dụng các đầu dò
huỳnh quang (FISH) đặc hiệu cho ba nhóm vi khuẩn a-, 0- và y-Proteobacterỉa.
12 chùng vi khuẩn kỵ khí được phân lập và tinh sạch (4 chùng đối với mỗi dạng môi trường sinh thái). Tính đa dạng cùa các chủng này về mật di truyền được xác định thông qua phân tích ARDRA đoạn gen mã hoá cho 16S rRNA đài 1500 bp sừ dụng hai enzyme Mspỉ và Haeììl. Các chùng đại diện cho các nhóm ARDRA được giải trình tự để xác định vị trí phân loại của cá nhóm vi khuẩn mà chúng đại diện.
- Bằng phương pháp xác định lượng Fe(ll) và NO3" trong môi trường nuôi cấy theo then gian, chùng vi khuẩn IN 12 đã được chứng minh có khả năng chuyển hoá hai hợp chất này khá cao, như vậy chủng này có tiềm năng sử dụng trong lĩnh vực môi trường. Bên cạnh đó khả năng sử dụng Mn(II) thay cho Fe(II) cũng đâ được nghiên cứu, tuy nhiên chùng IN 12 không thể hiện khả năng ửng dụng để loại bò Mn(IĨ) trong môi trường.

- Các kết quả nghiên cứu đã được công bố trong 02 bài báo khoa học, 01 báo cáo Poster tham gia hội nghị về sinh thái sinh vật và 02 trình tự gen đăng ký trong GenBank.
- Đã tham gia đào tạo 01 học viên cao học
9. Tình hình sử dụng kỉnh phí- Tổng kinh phí của đề tài được duyệt: 60.000.000 VNĐ- Tổng kinh phí của đề tài đã chi: 60.027.400, bao gồm các mục:
+ Chi phí thuê mướn: 24.000.000+ Chi phí nghiệp vụ chuyên môn: 29.522.400+ Văn phòng phẩm: 505.000+ Chi khác (lệ phí của đơn vị DT): 6.000.000

SUMMARY
1. Project title: Diversity o f anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria insome ecological environments and their potential use.
2. Code: QG.07.233. Duration: 2007 - 20094. Organizer: Vietnam National University Hanoi (VNUH)5. Project leader: Dr. Dinh Thuy Hang
Institute of Microbiology and Biotechnology, VNUH6 . Key participants: BSc. Nguyen Thi Tuyen
MSc. Nguyen Minh Giang7. Objectives and study contentsObjectives
- Comparatively study biodiversity of Fe(II) oxidizing - nitrate reducing bacteria in some ecological environments in Vietnam.
- Isolate representative strains that dominate in each environment and study their physiological, and phylogenetic characteristics.
- Study the application potential of the isolates in elumination of Fe(II), Mn(II) and NO3- in contaminated water sources.
Study contents
- Collect anaerobic mud samples from representative environments in Vietnam and keep at 4°c as the sources for microbiological studies.
- Determine the number of Fe(II) oxidizing-nitrate reducing bacteria in these environments by using MPN method carried out in anoxic liquid media containing Fe(II) as electron donor, N 03- as electron acceptor and trace acetate as corbo source. Mineral content of the media would respect to the natural conditions where the samples have been collected.
Species composition of the microbial communities in these environments would be analyzed via PCR-DGGE method applied for 16S rDNA of the samples of the MPN series.
- Isolate representative strains from the MPN tube of the highest dilution by performing dilution series in semi-liquid agar tubes.
Study physiology and phylogeny of the isolated strains, afterward select strains for the study
of application potential in elimination of Fe(II), Mn(II) and NO3- in contaminated waters.
8 . The obtained results
- Samples from three ecological environments, namely mud sediment freshwater pond, flooded paddy soil and sediment from estuarine environment were collected for studying Fe(II) oxidizing, nitrate reducing bacteria. These environments are known for significant

involvement o f bacteria in Fe and nitrogen cycles. Samples at 10 cm depth below the sediment surface were used for this study.
Number of Fe(II) oxidizing, nitrate reducing bacteria was enumerated via MPN method earned out in anoxic media with freshwater mineral content (for the samples from freshwater pond and paddy soil) or brackish water mineral content (for the sample from estuarine environment). These media contained Fe(II) as electron donor, NO3- as electron acceptor and trace acetate as carbon source. Growth of bacteria in the MPN series was recognized by colour changing in the media, from white (Fell precipitation) to dark brown (Felll precipitation).
Total DNA from representative MPN samples was purified and used in the PCR experiments to amplify 550 bp fragments of the V3 region of the 16S rDNA and used for analyzing diversity of bacteria in the communities with DGGE method. Most prominent DGGE bands were excised, reamlified and sequenced to determine the phylogenetic affiliation of the respective baterial groups.
Diversity of the bacterial communities in the samples was also analyzed by using in situ hybridization (FISH) with fluorescent probes specific for the a-, P- and y-Proteobacteria.
12 bacterial strains were isolated and purified (4 strains for each studied environment). Genetic diversity of these isolates was studied via ARDRA analyzes of Ỉ6 S rDNA with two endonucleases Mspl and HaelU. 16S rDNA of the strains represented for the RFLP groups were sequenced and comparatively aligned with the database to identify the phylogenetic affiliation of the ARDRA groups.
Quantitative analyzes of the amount of Fe(II) and N o r in growth media of the isolated strains allowed to select strain IN 12 as a candidate for application for simultaneous elimination of Fe(II) and NO3 ' in contaminated waters. Besides that, the ability of strain IN12 in oxidation of Mn(II) with nitrate was also studied, however it was revealed that tis strain did not growth with Mn(II) as electron donor for nitrate reduction.

MỤC LỤC
MỞ ĐẦU 1
CHƯƠNG I. TỐNG QUAN TÀI LIỆU1.1. Vi khuẩn tham gia chu trình Fe 21.2. Vi khuẩn oxy boá Fe(II) ở pH trung tính 3
1.2.1. Vi khuẩn hiếu khí oxy hoá Fe(II) 31.2.2. Vi khuẩn quang hợp kỵ khí oxy hoá Fe(II) 31.2.3. Vi khuẩn khử nitrate oxy hoá Fe(II) 4
1.3. Tiềm năng ứng dụng của vi khuẩn oxy hoá Fe(II), khử nitrate 41.3.1. Ảnh hưởng của nhiễm nitrogen trong các nguồn nước 41.3.2. Ảnh hưởng của nhiễm sắt trong các nguồn nước 51.3.3. Khả năng ứng dụng của vi khuẩn oxy hoá Fe(II), khử nitrate 6
CHƯƠNG II. NGUYÊN VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN c ử u 72.1. Nguyên vật liệu 7
2.1.1. Đối tượng nghiên cứu 72.1.2. Hoá chẩt 72.1.3. Thiết bị và dụng cụ 7
2.2. Phuxmg pháp nghiên cứu 72.2.1. Xác định số lượng vi khuẩn oxy hoá Fe(II) khử nitrate 72.2.2. Phân lập vi khuẩn oxy hoá Fe(II), khử nitrate 8
2.2.3. Tách DNA tổng sổ từ mẫu quần thể và từ chùng vi sinh vật 92.2.4. Phân tích đa dạng vi sinh vật bằng phương pháp ARDRA 92.2.5. Phưcmg pháp điện di biến tính DGGE 92.2.6. Giải trình tự gen 16S rDNA và dựng cây phân loại 102.2.7. Phương pháp FISH (Fluorescence In Situ Hybridization) 112.2.8. Định lượng Fe(II), Mn(II) và nitrate 12
CHƯƠNG III. KẾT QUẢ VÀ THẢO LUẬN 153.1. Xác định sổ lượng vi khuẩn oxy hóa Fe(II), khử nitrate tại ] 5các môi trường Dghiên cứu
3.2. Phân tích cấu trúc quần thể bằng điện di biến tính (DGGE) 163.3. Mức độ oxy hóa Fe(II), khử nitrate của vi sinh vật trong các mẫu 173.4. Phản lập vi khuẩn oxy hóa Fe(ll) khử nitrate từ các mẫu nghiên cứu 17và đánh giá tính đa dạng của các chủng phân lập.3.5. Phân tích đa dạng vi khuẩn trong các môi trường nghiên cứu 21bằng phương pháp FISH3.6. Nghiên cứu đặc điểm sinh lý, phân loại và hoạt tính sinh học 22

của các chủng vi khuẩn đại diện
KẾT LUẬN
KIẾN NGHỊ
TÀI LIỆU THAM KHẢO
PHỤ LỤCPhụ lục 1. Các trình tự genPhụ lục 2. Tham gia đào tạo khoa họcPhụ lục 3. Các công trình khoa học đã công

DANH MỤC CÁC CHỬVIÉT TẮTALF968 Fluorescence probe against a -ProteobacteriaARDRA Amplified ribosomal DNA restriction analysesBET42a Fluorescence probe against fi-ProteobacteriaBSA Bovin serum albuminCl Chloroform - isoamylalcoholCTAB Cetyl trimethyl ammonium bromideDAP1 4’,6-diamino-2-phenylindoleDNA Acid deoxyribonucleicdNTP Deoxyribonucleotide triphosphateddNTP Dideoxyribonucleotide triphosphateDGGE Denaturing gradient gel electrophoresisEDTA Ethylene-diamine-tetraacetic acidFISH Fluorescence in situ hybridizationGAM42a Fluorescence probe against y-ProteobacteriaMPN Most probable numberMQ Milli Q waterPBS Phosphate buffered salinePCI Phenol-chloroform-isoamylalcoholPCR Polymerase chain reactionOD Optical densityrDNA Ribosomal DNARNA Ribonucleic acidSDS Sodium dodecy] sulfateTAE Tris/Acetate/EDTATBE Tris/Borate/EDTAu v Ultraviolet

MỞ ĐẦU
Trong tự nhiên sát là một trong những nguyên tố cỏ mặt với hàm lượng đáng kể, sau carbon, nitơ, phospho và lưu huỳnh (Ehlich, 1996). Là một nguyên tố kim ioại, sất tồn tại ở các dạng ion với điện thế oxy hoá khử khác nhau, gồm Fe(II) và Fe(III). Trong hai dạng kể trên chi có Fe(II) tồn tại ở dạng hoà tan trong nước. Tuy nhiên, do có tính khử cao, Fe(II) nhanh chóng bị oxy hoá thành Fe(III) trong phản ứng hoá học với oxygen không khí. Do vậy ion Fe(II) thường chỉ được tìm thấy trong các điều kiện môi trường không có oxygen, ví dụ như ở đáy các thuỷ vực, các tầng nước ngầm hay môi trường có pH thấp (Konhauser, 1997; Nealson, Saffarini, 1994).
Trong khi oxy hoá Fe(II) bằng oxygen theo con đường sinh học chỉ diễn ra ờ môi trường có pH thấp thì oxy hoá Fe(II) bằng nitrate là quá trình diễn ra ở điều kiện pH trung tính (Ehrlich, 1996). Trong tự nhiên, quá trình oxy hoá sắt (II) với chất nhận điện từ là nitrate chủ yếu diễn ra ở ranh giới hiếu khí (có oxygen) và kỵ khí (không có oxygen) trong lớp trầm tích ở đáy các thuỷ vực. Oxy hoá Fe(II) kết hợp với khử nitrate có thể đóng vai trò quan trọng trong môi trường ô nhiễm với nồng độ Fe(II) cao (do thiếu oxygen) và NƠ3- cao (đo chất hữu cơ bị phân huỳ tạo thành) (Weber et aL, 2006b). Các loài vi khuẩn với khả năng tiến hành phàn ứng oxy hoá khử này có thể cùng một lúc thực hiện được hai nhiệm vụ, thứ nhất là chuyển Fe(II) hoà tan trong nước về dạng Fe(III) kết tủa, và hai là loại bỏ NO3-, chuyền thành dạng N2 không độc hại.
Nhiều công trình nghiên cứu trên thế giới cho thấy sự có mặt khá phổ biển của nhóm vi khuẩn oxy hoá Fe(II) khử nitrate với mật độ khá cao (106 tể bào/g trầm tích) trong các điều kiện môi trường khác nhau, bao gồm cả nước ngọt, nước lợ và nước mặn và tại nhiều vị trí địa iý khác nhau trên thế giới (Straub, Buchholz-Cleven, 1998). Tuy nhiên ờ Việt nam cho đến này chưa có công trình nghiên cứu nào đề cập đến quá trình sinh học dùng Fe(II) để khử NO3” cũng như các loài vi khuẩn đảm nhiệm quá trình này. Do vậy, đề tài ' Đa dạng sinh học của vi khuẩn kỵ kh í oxy hoả Fe(IĨ), khử nitrate trong một sổ môi trường sinh thái và khả năng ứng dụng của chúng ’’ được đặt ra với mục tiêu nghiên cứu tính đa dạng của vi khuẩn khử nitrate sử dụng Fe(II) là nguồn điện tử duy nhất trong một sổ dạng môi trường sinh thái đặc trung ở Việt Nam, đồng thời tìm hiểu khả năng ứng dụng của chúng trong việc xử lý các nguồn nước nhiễm ion sắt kim loại và nitrogen.
1

CHƯƠNG I. TỔNG QUAN TÀI LIỆU
1.1. Vi khuẩn tham gia chu trình Fe
Sắt là một trong những kim loại phổ biến nhất và là nguyên tố có mặt trên trái đất với hàm lượng lớn thứ tư, sau carbon, oxygen và nitrogen. Thông thường sắt tồn tại ờ đạng Fe2Ơ3 ít tan trong nước và có màu vàng nâu. Trong môi trường pH trung tính, dạng hòa tan trong nước Fe(II) chi tồn tại ở điều kiện không có oxygen, ví dụ như ở đáy các thủy vực, nơi oxygen hòa tan trong nước đã bị các vi sinh vật hiếu khí sử đụng đề phân hủy các hợp chất hữu cơ. Quá trình oxy hóa - khử giữa Fe(II) và Fe(III) có vai trò thiểt yếu trong chu trình sinh địa hóa môi trưởng và là một trong những quá trình chuyển hoá vật chất quan ưọng có mặt từ rất sớm trên ưái đất. Fe(II) là thành phần của các dạng khoáng phổ biến như siderite (khoáng chất có chứa FeCƠ3), vivianite (Fe3(PC>4)2 .8 H2 0 ) hoặc pyrite (Fe$2), trong môi trường kỵ khí có tính acid yểu đến môi trường trung tính (Straub et ai, 2001). Oxy hóa sắt diễn ra theo cả hai con đường hoá học và sinh học, trong đó các quá trình sinh học cỏ vai trò đặc biệt quan trọng tại nhiều dạng môi trường sinh thái khác nhau (hình 1.1). Với thế oxy hóa khử của cặp Fe(II)/Fe(III) là +770 mV đối với môi trường acid và +200 mV dối với môi truờng trung tính, Fe(II) có thể được sử dụng như chất cho điện tử để cung cấp đương lượng khử cho các quá trình đồng hóa carbon thành sinh khối nhờ các vi khuẩn oxy hóa Fe(II) trong cả điều kiện kỵ khí và hiếu khí, còn Fe(III) có thể được sử dụng như là chất nhận điện tử cuối cùng trong điều kiện kỵ khí để oxy hoá các hợp chất hữu cơ (Weber et a i, 2006a).
Hình 1.1. Chu trình chuyển hoá sắt trong tự nhiên (Ehrlich, Newman, 2008).1 - Vi sinh vật trong môi trường acid; 2 - Vi sính vật kỵ khỉ ở môi tnxờng trung tính (khừ nitrate, quang hợp kỵ khi)'
3 - Quá trình hóa học trong môi trưcmg trung tinh với nồng độ oxygen cao; 4 - Quá trinh hóa học; 5 - Quá trình sinh
học; 6 - H2S từ vi sinh vật; 7 - + 0 2, quá trình sinh học hoặc hóa học; 8 - -“-CO}2", quá trinh hóa học; 9 - chuyền H+
quá trình sinh học hoặc hỏa học; Í0 - Ọuá trình sinh học hoặc hóa học.
2.3
2

Vi khuẩn oxy hóa Fe(II) bao gồm nhiều nhóm có các đặc điểm sinh lý khác nhau, ví dụ như tự dưỡng và dị dưỡng, quang dưỡng và hóa dưỡng, ưa acid và trung tính, hiếu khí và ky khí. Mặc dù vai trò của quá trình chuyển hóa sắt nhờ vi khuẩn trong môi trường là rất lớn nhưng những hiểu biết hiện nay về sinh lý cũng như sinh hóa của nhóm ví sinh vật này còn nhiều giới hạn. Hầu hết các nghiên cứu và công bổ về quá trinh oxy hóa Fe(II) đều tập trung vào các vi khuẩn hiếu khí ưa acid như Thiobacciỉlus ferrooxidans (Temple, Colmer, 1951; Ehrlich, 1996), phát triển trong môi trường có pH 1-2, nơi có Fe(II) và Fe(III) ờ dạng ion hòa tan. Tuy nhiên, ở pH trung tính cơ chất và sản phẩm của quá trình chuyển hóa sắt lại ít hòa tan, điều này gây khó khăn cho việc nghiên cửu sinh ]ý của các vi sinh vật tham gia (Straub et a i, 2001). Mặc dù vậy, vi khuẩn tham gia chuyển hoá sắt ở điều kiện môi trường trung tính, bao gồm oxy hoá Fe(II) và khử Fe(III) lại rất đa dạng và đóng vai trò quan trọng hơn trong chu trình vật chất trong tự nhiên.
1.2. Vi khuẩn oxy boá Fe(II) ở pH trung tính
1.2.1. Vi khuẩn hiếu khí oxy hóa Fe(II)
Mặc dù quá trình oxy hóa Fe(II) bằng oxygen không khí tại pH trung tính xảy rất nhanh, nhiều nghiên cứu cho thấy oxy hoá Fe(II) bằng còn đường sinh học hoàn toàn có thể cạnh tranh với quá trình hoá học. Vi khuẩn hiếu khí oxy hoá Fe(II) ở pH trung tính được mô tả đầu tiên là các đại điện thuộc ba chi Gaỉỉionelỉa, Leptothrix và Marinobacier (Kuetzing, 1919; Ehrenberg, 1919). Ngoài ra, theo các nghiên cứu gần đây, một số vi khuẩn oxy hoá Fe(II) hiếu khí mới tìm thấy được xếp vào các phân lớp a-, p- và Ỵ- Proteobacterìa (Edwards et a l, 2003; Emerson, Weiss, 2004; Dhillon et a i, 2005; Rentz et a i, 2007). Nhu vậy cùng với sự phát triển của các phương pháp nghiên cứu vi sinh vật, bức tranh đa dạng của nhóm vi sinh vật này ngày càng trở nên phong phú hơn.
1.2.2. Vi khuẩn quang họp ky khí oxy hỏa Fe(II)
Trong những khu vực có ánh sáng, Fe(III) có the cũng được tạo thành thông qua hoạt tính oxy hóa Fe(II) của vi khuẩn quang hợp có khà năng sử dụng Fe(II) như một nguồn điện tử để tạo các đương lượng khử cho quá trình đồng hóa carbon vô cơ (Weber et aỉ., 2006a). Vi khuẩn quang hợp kỵ khí là vi khuẩn oxy hỏa Fe(II) kỵ khí được biết đến đầu tiên (Widdel et aỉ., 1993). Hiện nay nhóm vi khuẩn này đã được mô tà với nhiều đại diện thuộc các chi Chỉorobium, Rhodobacier, Thiodictyon, Rhodovuỉum, Rhodomicrobium (Ehrenreich, Widdel, 1994; Heising, Schink, 1998; Heising et aì., 1999; Straub et a l, 1999; Kappler, Newman, 2004).
Tuy nhiên quá trình oxy hóa Fe(II) bàng quang hợp trong môi trường tự nhiên bị giới hạn ở độ sâu 200 um trong đất và trầm tích do hạn chế ánh sáng (Ciani et a l, 2005). Hơn nữa, những nghiên cứu gần đây đã chỉ ra rầng vi khuẩn quang hợp oxy hóa Fe(II) không có khả năng sử dụng Fe(II) ờ dạng khoảng mà chi có thể oxy hóa Fe(II) ở dạng hòa tan (Kappler, Newman. 2004), do đó, chúng không đỏng vai trò đáng kể trong chu trình chuyển hoá sắt trên cạn.
3

1.2.3. Vi khuẩn khử nitrate, oxy hóa Fe(II)
Vi khuẩn oxy hoá Fe(II) kỵ khí đóng vai trò quan trọng trong chu trinh chuyển hoá sắt, đặc biệt trong các lóp trầm tích nơi vi sinh vật bị hạn chế về oxygen hoà tan (Lack eí al., 2002; Weber et al., 2001; Senn, Hemond, 2002; Straub et al., 2001; Weber et a i, 2006c). Với hiệu điện thế oxy hóa khử Fe(III)/Fe(II) tại pH 7 vào khoảng +200 mV, Fe(ll) có thể ừờ thành nguồn điện tử cho quá ưình hô hấp kỵ khí, khử nitrate thành N2 do một số nhóm vi khuẩn đảm nhiệm (Straub et a i, 1996; Benz et a i, 1998; Weber et a i, 2006c). Quá trình oxy hóa Fe(II), khử nitrate được tóm tát theo phương trình phản ứng như sau:
10 Fe2+ + 2 N 03“ + 24 H20 = 10 Fe(OH)3 ị + N2 T + 9 H2 1
Trong tự nhiên, quá trình oxy hóa Fe(II) với chất nhận điện tử là nitrate chù yểu diễn ra ở ranh giới hiếu khí (có oxygen) và kỵ khí (không có oxygen) trong lớp trầm tích ờ đáy các thủy vực. Vi khuẩn dùng ion Fe(II) làm nguồn cho điện tử để khử nitrate được phân lập đầu tiên từ các lớp trầm tích ao, hồ nước ngọt tại Bremen (Đức) năm 1996 (Straub et a i, 1996). Một số công trình nghiên cứu tiếp sau cho thấy sự có mặt phổ biển của nhóm vi khuẩn này với mật độ khá cao ( 1 0 6 tế bào/g trầm tích) trong các điều kiện môi trường khác nhau, bao gồm cà nước ngọt, nước lợ và nước mặn và tại nhiều vị trí địa lý khác nhau trên thế giới (Straub et a l, 1998). Các loài vi khuẩn phổ biến nhất trong nhóm nảy được biết đến hiện nay lả các loài thuộc chi Chromobacteríum và Klebsiella (Benz et aỉ.y 1998; Senko eí a i , 2005; Weber etaỉ., 2006b).
Vi khuẩn oxy hóa Fe(II), khử nitrate phần lớn sinh trường dị dưỡng, phụ thuộc vào chất hữu cơ (vi dụ như acetate) để cung cấp nguồn carbon cho việc tổng hợp thành phần tế bào (Straub et al., 1996; Benz et a l, Ỉ998). Bằng phương pháp MPN, người ta đã xác định được vi khuẩn oxy hóa Fe(II) khử nitrate chiếm 0,8% tổng số vi khuẩn khử nitrate nói chung, trong đó nhóm sinh trưởng dị dưỡng thường gặp hơn nhóm sinh trưởng tự dưỡng (Straub et a l, 1998). Vi khuẩn oxy hóa Fe(II), khử nitrate tự dưỡng mới chỉ được biết điến với hai đại diện là Ferroglobus pỉacidus, một vi khuẩn cổ ưa nhiệt (Hafenbradl et a l, 1996), và một vi khuẩn ưa
ấm thuộc phân lớp p-Proteobacteria (Weber et a l, 2006b). Đối với lịch sừ tiến hoá, các sinh vật sinh trường tự dưỡng vô cơ như nhóm vi khuẩn oxy hoả Fe(II) khử nitrate tự dưỡng đóng một vai trò quan trọng, góp phần tạo nguồn carbon hữu cơ ban đẩu để duy trì sự sống.
1.3. Tiềm năng ứng dụng vi khuẩn oxy hóa Fe(II), khử nitrate
1.3.1. Ảnh hưởng của nhiễm nitrogen trong các nguồn nướcNước thải với nồng độ ammonium hay nitrate cao xả ra ngoài môi truờng có thề gây ra
nhiều vấn đề nghiêm trọng liên quan đến môi trường sinh thái và sức khỏe cộng đồng.
4

Ảnh hưởng tới môi trường sinh thái
- Ảnh hưởng độc hại: Nitrogen ammonia ở dạng ammonia rất độc hại đối vói các loài thủy sinh, đặc biệt là cá. Ở pH trang tính, 99% N-ammonia tồn tại ở dạng NRị+, trong khi đó khi pH ở khoảng trên 9, dạng NH3 tăng đảng kể. Do vậy mức độ độc hại của N- ammonia rất nghiêm trọng khi pH trong môi trường tăng, thường là hậu quả sau khi có lượng nước thải với độ kiềm cao đổ ra ngoài môi trường.
- Giảm oxygen hòa tan trong các thủy vực: ammonia thường dẫn đến việc tăng nhu cầu oxygen cho các quá trình sinh hóa ở các thủy vực. Theo tính toán, 1 mg ammonia thải ra ngoài môi trường đòi hỏi lượng oxygen hòa tan tăng thêm 4,6 mg, chủ yếu do hoạt động của nhóm vi khuẩn tham gia vào quá trình nitrate hóa. Hậu quả của việc tăng nhu cầu về oxygen này là làm thiếu oxygen cho các loài thủy sinh khác cùng sống trong môi trường.
- Gây hiện tượng phủ dưỡng trong các ao hồ: việc tăng hàm lượng nitrogen trong các ao hồ kéo theo sinh trưởng rầm rộ cùa tảo và các thực vật thủy sinh, dẫn đến tâng nhu cầu oxygen về đêm và làm ảnh hưởng đến hoạt động sống của các động vật thủy sinh.
- Làm giảm hiệu quả của chỉorỉn: ammonium kết hợp với chlorin tạo hợp chất chloramin có tác dụng kháng khuẩn kém hom so với chlorin ở dạng tự do.
- Ần mòn: ammonium ở nồng độ trên 1 mg/L có thể gây ăn mòn cho các đường ống dẫn nước và các công trình kim loại dưới nước.
Ảnh hưởng tới sức khỏe cộng đồng
- Gây bệnh thiếu máu: nitrate là nguyên nhân gây bệnh thiếu máu (Methemoglobinemia) ở người do bị khử thành nitrite trong hệ đường ruột, sau đó liên kết với hemoglobin dưới dạng methemoglobin, không cỏ khả năng vận chuyển oxygen. Ở người khỏe mạnh, hàm lượng methemogỉobin trong máu được giữ ổn định ở mức 1 - 2% nhờ enzyme methemoglobin reductase. Trè em dễ bị ảnh hường cùa bệnh này hơn so với người trưởng thành vì có độ pH trong dạ dày cao hom, tạo điều kiện thuận lợi cho vi khuẩn khử nitrate sinh trưởng. Vitamin c được biết đến với khả năng bảo vệ và giữ nồng độ methemoglobin ở mức thấp.
- Gây bệnh ung thư: nitrite được tạo ra từ nitrate còn có thể kết hợp với các amine thứ cấp để tạo thành nitrosamine, là tác nhân gây đột biến gen và một số bệnh ung thư như ung thư dạ dày, ung thư bàng quang (Tricker, Preussmann, 1991; Lundberg, 2004). Theo quy định quốc tế, giới hạn nitrate trong nước uổng là 10 mg/L.
1.3.2. Ảnh hirởng của nhiễm sắt trong các nguồn nước
Sắt là nguyên tố phổ biến trong tự nhiên, là một thành phần quan trọng trong tồng hợp hemoglobin (chất vận chuyển oxygen cho các tế bào trong cơ thề) và myoglobin (chất dự trữ

oxygen cho cơ thể). Ngoài ra sắt còn tham gia vào thành phần một số enzyme oxy hoá khử như cataỉase, peroxydase vả các cytochrome (những chất xúc tác sinh học quan trọng trong cơ thể). Sắt đóng vai trò quan ưọng trong việc sản xuất ra năng lượng oxy hoá, vận chuyển oxygen, hô hấp của ty lạp thể và bất hoạt các gốc oxy hóa cỏ hại. Tuy nhiên quá tải sắt trong cơ thể cũng gây ứ đọng sắt tại các mô như tim, gan, tuyến nội tiết..., dẫn đến rối loạn trầm trọng chức năng các cơ quan này (Trần Thị Kiều My và Nguyễn Hà Thanh, 2006).
Cơ thể hấp thu sắt ở dạng Fe(II). Pepsin tách sắt khỏi các hợp chất hữu cơ và chuyển thành dạng gắn với các acid amine hoặc đường. Acid chlohydric khử Fe(III) thành Fe)II) để hấp thu. Vì vậy nên lượng sắt dư thừa trong nước uống cho dù ở dạng Fe(II) hay Fe(III) đều gây nguy hiểm cho con người.
Tác hại của thừa sắt trong cơ thể hiện nay vẫn còn đang gây nhiều tranh cãi. Người ta cho rằng bộ não là mục tiêu chính của sự thừa sắt, sự tích lũy sắt trong mô não là một trong những nguyên nhân của các bệnh về thần kinh như Parkinson và Alzheimer, sắt còn được coi như là một chất gây ung thư rất mạnh, thường gặp nhất là ung thư gan. sắt du thừa tích luỹ trong tim và các động mạch có thể gây nên các bệnh về tim hoặc phá vỡ động mạch, sắt thừa lắng đọng trong lá lách sẽ phá vỡ sự bài tiết insulin và gây ra bệnh đái tháo đường nghiên trọng.
1.3.3. Khả năng ứng dụng của vi khuẩn oxy hoá Fe(II), khử nitrate
Với khả năng hô hấp kỵ khí bằng nitrate, vi khuẩn oxy hóa Fe(II) khử nitrate có thể tham gia vào quả trình loại bỏ nitrogen trong các nguồn nước thài hay nước sinh hoạt có nồng độ nitrate cao (Straub et ai., 1996). Quá trình này diễn ra khi có mặt các hợp chất hữu cơ làm nguồn điện tử thích hợp để khử nitrate, ví dụ như lactate, acetate hay methanol, là những sàn phẩm của quá trình phân giải chất hữu cơ cao phân tử. Bằng các phương pháp sinh học phân tử người ta đã xác định được tỷ lệ khá cao của vi khuẩn oxy hóa Fe(II) khử nitrate trong các hệ thống xử lý nước thải nhiễm nitrogen (Bitton, 1999). Bên cạnh đó, khả năng sinh trường vô cơ sử dụng Fe(II) làm nguồn điện tử để khử nitrate là một ưu thế của nhóm vi khuẩn này so với các loài khử nitrate thông thường. Do tác động của vi khuẩn oxy hóa Fe(II) khử nitrate, cùng một lúc ion Fe(II) vả nitrate dư trong nguồn nước có thể được loại bỏ. Khả năng ứng dụng cùa nhóm vi khuẩn này đặc biệt có ý nghĩa đối vói việc xừ lý các nguồn nước ngầm dùng cho sinh hoạt bị nhiễm sắt và nitrogen ngấm xuống từ tầng nước mặt. Ngoài ra, trong một số nghiên cứu gần đây, vai trò cùa vi khuẩn oxy hóa Fe(II) khử nitrate còn được tìm thấy trong việc cố định các ion arsen thông qua khả năng tạo Fe(III) là sàn phẩm cùa quá trình trao đổi chất (Hohmann et a i, 2009).
6

CHƯƠNG II - NGUYÊN VẬT LIỆU VÀ PHƯƠNG PHÁP NGHIÊN c ứ u
2.1. Nguyên vật liệu
2.1.1. Dối tượng nghiên cứuMầu bủn vả trầm tích được thu thập ờ ba môi trường sinh thái đại diện khác nhau: mầu
bùn đáy ao nước ngọt và chân ruộng ngập nước được thu ở Từ Liêm, Hà Nội, mẫu trầm tích nước lợ được thu ở ven biển Vân Đồn, Quảng Ninh. Mâu chân ruộng và ữầm tích ven biển được thu thập ừong các ống thủy tính nút xoáy cho phép khoan sâu tới 60 cm dưới bề mặt trầm tích (hình2.1). Mẫu đáy ao được thu thập trong binh thuỷ tinh, sau đó bổ sung nước vào toàn bộ thề tích để giảm thiểu sự ảnh hường cùa oxygen trong không khí. Mầu được bảo quản ở nhiệt độ 4°c cho đến khi tiến bành các thí nghỉệm tiếp theo.
Ỉ Hình 2.1. Mẫu bùn và trầm tích thu thập ngoài
môi trường tự nhiên, được đảm bảo kỵ khí nghiêm ngặt.
2.1.2. Hóa chấtCác hóa chất được sừ đụng trong nghiên cứu vi sinh vật đều là những hóa chất có tiêu
chuẩn chất lượng cao của các hãng cung cấp lớn nhu Merck (Đức), Sigma (Mỹ). Hoá chất và một số kit dùng cho phân tích sinh học phân tử do các hãng Bioneer (Hàn Quốc), Fermentas (Đức), Qiagen (Mỹ), ABI (Mỹ), BioRad (Mỹ) cung cấp.
2.1.3. Thiết bị, dụng cụ sử dụng trong nghiên cửuNghiên cứu được thực hiện tại Viện Vi sinh vật và Công nghệ sinh học - ĐHQGHN, sử
dụng các thiết bị chuyên môn dùng ừong vi sinh vật học, sinh học phân tử và hoá phân tích đạt tiêu chuẩn quốc tế.
2.2. Phirong pháp nghiên cứu
2.2.1. Xác định số lượng vi khuẩn oxy hoá Fe(II), khử nitrateSố lượng vi khuẩn oxy hoá Fe(II) khử nitrate được xác định thông qua phương pháp MPN
(American Public Health Association, 1969) trong môi trường dịch thể kỵ khí hoàn toàn có thành phần khoáng tương úmg với nước ngọt (dùng cho mẫu bùn đáy ao và chân ruộng ngập nước) hoặc nước lợ (dùng cho mẫu trầm tích ven biển) (bàng 2 .1 ).
7

Băng 2.1. Môi trường kboáng kỵ khí nirớc ngọt và nước lợ cho vi khuẩn oxy hoá Fe(II) khử nitrate (Ratering, 1999).
Thàob phần Nước ngọt Nước lợNước cất 1 lít 1 lítNaCl 1 g 13,7 gMgCl2 .6H20 0,4 g 5,9 gCaCl2 .2H20 0,15 g 0,81 gKC1 0,5 g 0,58 gKBr - 0,05 gMgS04 .7H20 0,25 g 0,25 gNH4CI 0,25 g Hỗn hợp NH4 CI (0,25 g) vàKH2PO4 0 , 2 g KH2PO4 (0,25 g) được chuẩn
bị trước trong 50 ml nước cấtKhử trùng ờ 121 °c trong 25 phút, lấy ra ờ 80 °c, sục khí N2 trong 5 phút, làm nguội trongnước máy, sau đó thêm các dung dịch sau (đã khử trùng riêng bằng nhiệt hoặc màng lọc):Hỗn hợp Vitamin 1 ml 1 mlHỗn hợp vi lượng 1 ml 1 mlVitamin BI (Thiamin) 1 ml 1 mlVitamin B12 0 , 1 ml 0 , 1 mlVitamin B2 1 ml 1 mlNa-Acetate 1 M I ml 1 mlNaNƠ3 I M (chất nhận điện tử) 5 ml (5 mM) 5 ml (5 mM)FeS04 1 M (chất cho điện tử) 10 ml (10 mM) 1 0 ml (10 mM)NaHC03 1 M 30 ml 30 mlChuẩn pH ở 7-7,2, sau đó chia môi trường sang các bình serum và ống nghiệm rồi sục khí N2)đậy bình và ống nghiệm bầng nút cao su có kẹp nhôm hay nút vặn có lỗ hở.
Mẩu được bổ sung vào ống MPN đầu tiên với tỷ lệ 10% thể tích. Thí nghiệm được ủ tại 28 °c trong 8 tuần, sự phát triển của vi khuẩn trong ống MPN được ghi nhận thông qua đổi màu môi trường từ trắng đục (màu của Fell) sang vàng nâu (màu cùa Felll).
2.2.2. Phân lập vỉ khuẩn kỵ khíoxy hoả Fe(IỈ) khử nitrateỐng MPN ờ nồng độ pha loãng cao nhất có vi khuẩn oxy hóa Fe(II), khử nitrate phát triển
được sử đụng để làm nguồn phân lập các khuẩn lạc đơn. Việc phân lập được tiến hành theo phương pháp pha loãng trên dãy ổng thạch bán lỏng ( 1 %) với môi trường có thành phần tương tự như môi trường dùng trong phương pháp MPN (Widdel, Bak, 1992). Ống thạch bán lòng sau khi bổ sung nguồn vi sinh vật (10%) từ ống MPN (nguồn phân lập) được sục khí N2 và ủ ở tư thế đảo
8

ngược đầu tại 28°c trong bóng tối. Khuẩn lạc đom phát triển trong các ổng pha loãng được tách bằng pipet Pasteur và chuyển sang môi trường dịch thể thích hợp (nước ngọt hay nước lợ).
2.2.3. Tách DNA tồng sổ từ mẫu quần thể và chủng vi sinh vậtDNA tổng số của quẩn thể vi sinh vật từ các ống MPN được tách chiết theo phương pháp
do Zhou và cộng sự (1996) mô tà với cải biến về nồng độ đệm phosphate là 120 mM thay cho 100 mM và nồng độ proteinase K là 14 mg/ml thay cho 10 mg/ml. DNA genome của các chủng đom được tách theo phương pháp cùa Marmur (1961). Sản phẩm DNA sau khi tách chiết được
hoà tan trong nước và bảo quản tại - 2 0 °c.
2.2.4. Phân tích đa dạng vi sinh vật bằng phương pháp AKDRAARDRA (Amplified Ribosomal DNA Restriction Analyses) đuợc sử dụng để nghiên cứu
mức độ đa dạng về di truyền của vi sinh vật dựa trên phân tích kết quả xử lý gen 16S rDNA bằng enzyme giới hạn (Ravenschlag et a i, 1999). Gen 16S rDNA của các chủng đơn được khuếch đại trong phản ứng PCR sử dụng cặp mồi 27 F (AGA GTT TGA TCC TGG CTC AG) và 1492R (GGT TAC CTT GTT ACG ACT T), sau đó được xử lý bàng hai enzyme giới hạn Mspỉ và HaeIII (Fermentas) (bảng 2.2). Sản phẩm DNA sau khi đã xử lý enzyme được phân tách bằng điện đi trên gel agarose 2% tại 100 V trong thời gian 30 phút. Các chủng vi khuẩn sẽ được xếp nhỏm đựa trên sự tương đồng về phổ các băng điện di.
Bảng 22. Phản ứng cắt gen Ỉ6 S rDNA bằng các enzyme giói hạn Mspl và Haeìll
Nước cắt vô trùng 18 ụl
1 Ox đệm R (hoặc đệm Tango) 2 p.1
Trộn đều và ly tâm nhanh, sau đó ủ hỗn hợp phản ứng tại 37 °c trong 3 giờ
2.2.5. Phương pháp điện di biến tính DGGE
Khuếch đại đoạn gen 16S rDNA: Vùng V3-V5 của gen I6 S rDNA với độ dài 550 bp (hình 2.2) được khuếch đại trong phản ứng PCR sử dụng cặp mồi GM5F (CCT ACG GGA GGC AGC AG) và 907R (CCG TCA ATT CCT TTR AGT TT) (Muyzer et a i, 1993). “Touch-down” PCR được sừ dụng để tăng độ đặc hiệu và giảm sự hình thành các sàn phẩm phụ trong phản ứng khuyeechs đại. Đẻ tạo tính ổn định cho việc phân tách các trình tự DNA trên gel điện di biến tính, kẹp GC khoảng 40 bp (CGC CCG CCG CGC GCG GCG GCC GGG GCG GGG GCA CGG GGG G) được gắn vào đầu 5’ cùa mồi GM5F.
Sản phầm PCR
Enzyme Hae III (hoặc Msp I)10^1 (0,1 -0,5 p.g ADN)1 |il ( 1 0 u/|il)
9

6U 5f
£<*># Tuel Ec1f *“ . / A. ___^ mi
16S rDNA 0VI V2 V3 , V4 j V5
i. ♦+ EcS1BrLacĩ WIAB2 WBAC7 £*1406
HÕA2 ♦itw
Hình 2.2. Vùng V3-V5 và vị trí các mồi trên gen 16S rDNA ở Lactobacillusplantarum
Thành phần phản ứng và chu kỳ nhiệt của PCR khuyếch đại đoạn V3-V5 ờ vi khuẩn như sau:
Bảng 2.3. Thành phần phản ứng và chu kỳ nhiệt của PCR khuyếch đại vùng V3-V5 của gen 16S rĐNA
Thành phần phản ứng Thể tích (jil) Chu kỳ nhiệt (touch-down PCR)h 2 0 m q 27 Taq polymerase được đưa vào phàn ứng sau
IOx Taq buffer 5 bước biến tính đầu tiên ờ 94 °c trong 1 phút
BSA 5 khi nhiệt độ đạt 80 °c. Nhiệt độ gắn mồi
MgCl2(20 mM) 4 được đặt cao hon 1 0 °c so với nhiệt độ lý
dNTP (2,5 mM mỗi loại) 4 thuyết (tức là 65 °C). Sau mỗi chu kỳ nhiệt
Mồi GM5F-GC (50 pmol/nl) 1 độ gắn mồi giảm đi 0,5 °c cho tới khi đạt
Mồi 907R (50 pmol/fi.l) 1 tới 55 °c, ờ nhiệt độ này phản ứng tiến hành
ADN khuôn 2 thêm 15 chu kỳ. Thời gian kéo dài chuỗi
Taq polymerase 1cùa môi là 3 phút.
Tổng thể tích phản ứng 50
DGGE: Điện di được tiến hành trên gel polyacrylamide 6 % với dải biến tính urea/formamid từ 30% đến 60%. Quá trình điện di được thực hiện bằng bộ điện di Dcode™ System (BioRad) ở nhiệt độ 60 °c tại 200 V trong 3,5 h. Sau khi điện di, gel polyacrylamide được nhuộm trong dung dịch ethidium bromide (5 mg/ml) trong 30 phút, sau đó rửa nước và chụp ảnh duới tia u v trên máy GelDoc (BioRad). Băng điện di được cắt và thôi trong nước qua đêm tại 4 °c. Dịch thôi DNA được dùng lảm khuôn để thục hiện phản ứng PCR như chu trình nhiệt của phản ứng PCR cho DGGE với cặp mồi GM5F (không có kẹp GC) và 907R. Sàn phẩm PCR tiếp theo được tinh sạch và giải trình tự.
2.2.6. Giải trình tự gen 16S rDNA và dựng cây phân loại
Gen 16S rDNA cùa các chủng đơn (1500 bp) hoặc sản phẩm PCR từ các bãng điện di biến tính (550 bp) sau khi tinh sạch bằng kít (Bioneer - Hàn Quốc) được tiến hành phản ửng đọc trình tự với ABI Prism BigDye Terminator cycle sequencing kit và đọc trình tự trên máy tự động
10

3110 Avant Appied Biosystems. Trinh tự gen sau đó được phân tích so sánh với trình tự 16S rDNA của các loài có liên quan hiện đã công bố trên Database DDBJ/EMBL/GenBank sử dụng phần mềm BLAST Search. Cây phân loại được dựng theo phương pháp neighbour-joining (Saítou, Nei, 1987), trong đó định dạng cây được tiến hành dựa trên 1000 phép so sánh đa chiều (Felsenstein, 1985).
2.2.7. Phương pháp FISH (Fluorescence In Situ Hybridization) (Amann et al., 1992)
Nguyên lý: FISH là phương pháp lai sử dụng các đầu dò thích hợp bắt cặp với rRNA nhàm xác định phả hệ cùa vi sinh vật trong quần thể một cách trực tiếp, không qua bước phân lập và nuôi
Chuẩn bị bóa chất:Formaldehyde 37% PBS I Ox5 M NaCl I M Tris-HCl0,5 M EDTA SDSỈ0%Formamide DAPI (0,02 mg/ml)Đầu dò: là các đoạn oligonucleotide đài 18-20 nucleotide gán chất huỳnh quang sulphoindocyonide (CY3, hấp phụ bước sóng 552 nm). Các đầu dò đã sử dụng trong nghiên cứu này được liệt kê trong bảng dưới đây (nồng độ 2 ng/^1).
Bảng 2.4. Các đầu dò sử dụng trong nghiên cứu
Tên đầu dò Đối tượng bắt cặp Trình tự (5’-> 3’) Formamide (%)
ALF968 a -Proteobacteria GGTAAGGTTCTGCGCGTT 2 0
BET42a p -Proteobacíeria GCCTTCCCACTTCGTTT 35
GAM42a y-Proteobacteria AAACGATGTGGGAAGGC 35
Bảng 2.5. Chuẩn bị đệm lai và đệm rửa
Dung dịch gổcThê tích
2 ml đệm lai 50 mi đệm rửa5 M NaCl 360 9 ml 900 mM1 M Tris/HCl 40 ụ\ 1 ml 20 mM
Form amide % phụ thuộc từng đâu dò0,5 M EDTA 0,5 ml 5 mMNước cât vô trùng dân tới 2 ml dân tới 50 miSDS 10% (bô sung cuối cùng để tránh kết tủa)
2ịil 50 ịiị 0 ,0 1 %
Các bước tiến hành: Cố đinh mẫu:

- Hoà 0,5 g mẫu đất/bùn ưong 1,5 ml PBS lx- Bổ sung formaldehyde vào mẫu tới nồng độ 2 - 4% và cố định ở 4°c trong thời gian ít nhất ỉà 30 phút nhưng không quá 24 h.- Ly tâm, loại phần dịch và rửa lại cặn tế bào bàng PBS IX- Ly tâm, loại PBS lx và bổ sung 1,5 ml hồn hợp dung dịch etanol/PBS (1:1)
- Bảo quản tế bào đã cố định tại -20°c.Lai với đầu dò:
- Lấy 15 - 20 mẫu hoà vớí 2 ml nước cất vô trùng vả đưa lên màng polycarbonate (0 , 2 ịim).
- Để khô mẫu trong không khí và bảo quản trong hộp kín tại -20°c cho đến khi sử dụng.- Chia màng thành các phần nhỏ để tiến hành lai (một màng có đường kính 25 mm có thể chia làm 6 - 8 phần sử dụng cho các đầu dò khác nhau). Dùng bút chì đánh dấu mẫu (bằng số).- Chuẩn bị 2 ml đệm lai trong ổng eppendorf (bảng 2.5)- Chuẩn bị dung dịch đầu dò: trộn 2 fil đầu dò (2 ng/|il) với 18 jnl đệm lai trong ống eppendorf0,5 ml, giữ trong đá, tránh ánh sảng.- Đặt một miếng giấy thẩm vào ống Falcone 50 ml, đổ phần đệm lai còn lại vào ống.- Đặt mảnh màng polycarbonate lên lam kính, mặt có mẫu lên trên. Các mẫu lai với cùng một đẩu dò có thể đặt cùng trẽn một lam kính.- Nhỏ hỗn hợp đầu dò lên các mẫu và đặt toàn bộ lam kính vào ổng falcon có miếng giấy lọc thấm đệm lai đã chuẩn bị trước (ở tư thế nằm ngang) và lai ờ 46°c trong thời gian ít nhất là 90 phút (nhưng không quá 3 h).- Chuẩn bị 50 ml đệm rửa trong ống falcon 50 ml- Chuyển nhanh mẫu sau khi lai vào đệm rửa đã được làm nóng trước đó trong bể ổn nhiệt tại 48 °c trong 15 phút. Gạn bớt đệm và đổ mẫu ra đĩa Petri. Dùng kẹp gắp mẫu, nhúng qua nước cất vô trùng trong vài giây và đặt lên giấy thấm cho khô (mật có mẫu ở trên).
Nhuôm đối chửng và quan sát:- Đe nhuộm đối chứng, nhỏ 50 nl dung dịch DAPI lên mồi mẫu, giữ 3 phút trong tối, sau đó rửa nhanh trong nước cất và rửa trong cồn 80% trong vài giây rồi để khô trong không khí (đặt trong hộp tối). Mầu có thể được bảo quản trong -20°c trong vài ngày mà tín hiệu huỳnh quang không bị thay đổi.- Phủ một lớp Citifluor/Vecta shield (4:1 v/v) và quan sát dưới kính hiển vi huỳnh quang.
2.2.9. Định lượng Fe(II), Mn(II) và nitrate
Định Iưựng Fe(II) bằng thuổc thử phenanthroiin {DIN 38406 E1-], 1983)Nguyên lý: O- phenanthrolin phản ứng với Fe(II) tạo phức có màu tím đỏ trong khoảng pH 3 -9, đo được ở bước sóng 510 run. Nồng độ Fe(II) cho phép là 0,01 - 5 mg/L. Ket quà của phép đo có thể bị ảnh hưởng bời ion Mn và Cu.
12

Chuẩn bị:Dung dịch HC1 25% (rửa dụng cụ): dụng cụ thủy tinh thí nghiệm phải được rửa qua dung dịch HC125% sau đó tráng bằng nước cất trước khi sử dụng.Dung dịch H2SO4 loãng (hạ pH): H2 SO4 đặc : nước = 1 : 4 Dung dịch ammonium acetate 5,2 M (đệm):
CH3COONH4 40 gCH3COOH 50 miNước cất đủ 100 ml
Dung dịch phenanthrolin 21 mM (tạo phức):C12H9C1N2.H20 0,5 gNước cất đủ 100 ml
(Bảo quản trong lọ tối, hạn sử dụng trong 1 tuần)Dung dịch chuẩn:
FeS04 .7H20 2,78 mgHC1 25% 6,25 mlNước cất đù 100 ml
Tiến hành:- Sau khi thu mẫu, ngay lập tức hạ pH tới 1 bàng dung dịch H2 SO4 loãng (1% thể tích)- Thêm vào 50 ml mẫu 5 ml dung dịch ammonium acetate, trộn đều bàng vortex. Hỗn hợp phải có pH nàm trong khoảng 3,4 - 5,5 (tối ưu là 4,5).- Thêm 2 ml dung dịch phenanthrolin, trộn đều.- Thêm nước cho tới 100 ml, trộn đều. Giữ ở nhiệt độ phòng trong 15 phút.- Đo OD tại bước sóng 510 nm.- Đường cong chuẩn được tiến hành với nồng độ Fe(C) trong khoảng từ 5 - 50 ỊiM.
Định Iưọng nitrate bằng tbuổc thử acid disulfofemic (Lê Đức, 2004)
Nguyên lý: ĩon NƠ3~ tác dụng với acid disulfofemic tạo thành nitrodisulfofemic, phàn ứng vớiammonia (NH3) đặc tạo thảnh phức có màu vàng, xác định được ở bước sóng 410 nm. Nồng độNO3- cho phép là 0,5 - 5 mg/L.Chuẩn bị:
Dung dịch acid disulfofemic:Phenol tinh khiết không màu 25g H2SO4 đặc (tinh khiết) 150 ml H2 SO4 bốc khói (oleum) 75 mlTrộn đều, đun sôi cách thủy trong bình cầu gắn ống sinh hàn hồi lun trong 2 giờ.
Dung dịch chuẩn:KN03(đãsấykhôờ 105 °C) 0,1631g Nước cất 500 mỉ
13

CHClj lmlNước cất dẫn đủ 1000 ml
Tiến bành:- Mầu nước (chúa không quá 5 mg/L NO3) lấy vảo ống nghiệm, trung hòa đến pH 7.- Chuyển mẫu sang chén sứ, cô cạn bằng đun cách thủy.- Thêm 2 ml dung dịch acid disulfofemic, hòa tan phần cặn bằng đũa thủy tinh.- Thêm 20 ml nước cất, 6 ml NH3 đặc hoặc 6 ml KOH 12N.- Thêm nước cất cho đủ 50 ml (trong bình định mức).- Đo OD tại bước sóng 410 tun.- Đường chuẩn được xây dựng trên các nồng độ dung dịch chuẩn trong khoảng từ 0-10 mM.
Định lượng Mn(n) bằng thuốc thử formaldoxime (Brewer and Spencer, 1971)Nguyên lý: Mangan phản ứng màu với formaldoxime (formaldehyde oxime) trong môi trường kiềm tạo thành phức có màu cam đò đậm, xác định ờ bước sóng 450 nm. Để không hình thành các kết tủa hydroxiđ, pH tối ưu cùa phương pháp này lả 8 , 8 - 8,9.Chuẩn bị:
Dung dịch formaldoxime:20g Hydroxygenlamine hydrochloride hòa tan trong 450 ml nước cất, thêm 10 ml dung dịch formaldehyde 37% và thêm nước đến 500m].
Dung dịch ammonia (NH3)Hỗn hợp formalđoxime:ammonia theo ti lệ 5:2 (thể tích)Dung dịch chuẩn (100 )ag mangan/ml):
Hòa tan 0,2876 g permanganate kali (KMn04) trong 100 ml nước cất, thêm 3 ml dung dịch H2 SO4 đầm đặc, dẫn acid sunfurous (SO2 trong nước) cho đán khi dung dịch mất màu. Đun sôi dung dịch để loại bỏ SO2 thừa sau đó iàm lạnh. Bổ sung nuớc cho đù 1 lít.
Tiến hành:- Đưa 35 mi mẫu về pH = 8 , 8 - 8,9- Thêm 3 ml hỗn hợp Formaldoxime/Ammonia vào lắc xoay tròn, đều.- Sau 2 - 30 phút đo quang phổ hấp thụ ờ bước sóng 450 nm.- Đường chuẩn được xây dựng trên các nồng độ dung địch chuẩn trong khoảng từ 0 - 10 mM.
14

CHƯƠNG IU. KẾT QUẢ VÀ THẢO LUẬN
3.1. Xấc định số lưọng vi ldraẩn oxy hỏa Fe(]I), khử NO3' tại các mỏi trưừng nghiên cứu
Ba môi trường đại diện được lựa chọn để tiến hành nghiên cứu gồm có bủn đáy ao nước ngọt, chân ruộng lúa ngâp nước và trầm tích nước lợ ven biển. Đây là những môi trường có hoạt động vi sinh vật trong chu trình chuyển hoá sắt và nitrogen cao (Ratering, Schnell, 2000). số
lượng vi khuẩn oxy hóa Fe(II), khử NO3" trong các mẫu thu thập được xác định thông qua pbưcmg pháp MPN sử dụng môi trường địch thể chứa FeSC>4 làm chất cho điện tử và NaNƠ3 làm chất nhận điện tử cuối cùng. Thành phần khoáng trong môi tnrờng tương ứng với điều kiện nước ngọt (đối với mẫu bùn đáy ao và chân ruộng ngập nước) hoặc nước lợ (đối với mẫu bùn ven biển). Sự phát triển của vi sinh vật sinh trưởng nhờ oxy hoá Fe(n) trong các ống MPN được nhận biết thông qua biến đổi màu sắc của môi trưởng từ ừấng xanh (màu của Fe(II)) sang màu vàng nâu (màu của Fe(ni)) (hình 3.la).
Hình 3.1. Xác định số lượng vi khuẩn oxy hoá Fe(II) khử N O 3 - trong các môi trưởng sinh thái khác nhau, (a) - Nhận biết sự có mặt của vĩ khuẩn trong các ống MPN thông qua biến đổi màu sắc của môi trường từ trắng xanh (Fell) sang vàng nâu (Felll). (b) - Sổ lượng vi khuẩn oxy hóa Fe(n), khử N (V xác định thÔDg qua phương pháp MPN.
Kết quả cùa thí nghiệm MPN (hình 3.1b) cho thấy số lượng vi khuẩn oxy hóa Fe(II), khử NO3 cao nhất trong mẫu bùn chân ruộng ngập nước (9,3 X 103 tế bào/g bùn), cao hcm hẳn so với mẫu trầm tích nước lợ ven biển (4,3 X 103 tế bào/g trầm tích) và mẫu bùn đáy ao nước ngọt (1,5 X
1 0 3 tế bào/g bùn). Mật độ và mối tương quan giữa số lượng vi khuẩn oxy hoá Fe(II) khử NCV với các điều kiện môi trường tại mỗi vùng sinh thái như trên cũng đã được tìm thấy ữong một số nghiên cứu trước đây (Weber et al., 2006b,c; Ratering, Schnell, 2000; Hauck et al., 2001; Ratering, 1999; Straub et a i, 1996).
15

3.2. Phân tích cấu trúc quần thể bằng điện di biến tính DGGE
Để Xác định các nhóm vi khuẩn oxy hoá Fe(II)/khử NO3- chiếm ưu thế tại các môi trường nghiên cứu, chúng tôi tiến hành phân tích cấu trúc quần thể trong các ống MPN ở độ pha loãng 10" 3 (là nồng độ gần tới hạn cùa dãy MPN đối với cả 3 mẫu) bẳng phương pháp PCR-DGGE đoạn gen 16S rDNA (hình 3.2). Các băng điện di đuợc cắt từ gel và sử dụng lảm khuôn cho phản ứng PCR tiếp sau để xác định trình tự và so sánh với các trình tự 16S rDNA đã công bố trong ngân hàng đữ liệu GenBank.
R lì
•+• Pseudomonas sp.Paracoccus SD. -►
Anaeromvxobơcter SV.+-
Hình 3.2. Phổ điện di biến tính (DGGE) phân tích đoạn gen Ỉ 6 S rDNA của quần thể vi sinh vật trong các ống MPN của các mẫu nghiên cứu. A - Bùn ao nước ngọt; R- Bùn chân ruộng ngập nước; B - Bùn trầm tích ven biển.
Cỏ thể thấy rằng các loài Anaeromyxobacter có mặt trong cả 3 dạng môi trường nghiên cứu. Đây là nhóm vi khuẩn nằm trong phân lớp h-Proteobacteria, hiện mới chi có một loài duy nhất được công bố là A. dehaỉogenans cùng với một sổ đại diện chưa định đanh đến loài. Các chủng Anaeromyxobacter đã công bố đều sinh trường kỵ khí khử Fe(III), chưa có chùng nào 4
được nghiên cứu về khả năng sinh trưởng khừ NO3”, sử dụng Fe(II) làm chất cho điện tử (Treude et ai., 2003; Straub, Buchholz-Cleven, 1998; Straub et al., 1996). Trong môi trường nuôi cấy sử dụng ờ đây (cũng như trong điều kiện tự nhiên), Fe(III) đồng thời tồn tại với Fe(II) đo kết quả chuyển hoá Fe(II) bằng con đường hoá học (phàn ứng với lượng nhò oxygen trong môi trường) và con đường sinh học (đo các vi sinh vật oxy hoá Fe(II) khử NC>3_). Do vậy, sự có mặt cùa các loài sinh trưởng kỵ khí khử Fe(III) như Anaeromyxobacter trong điều kiện môi trường nghiên cứu trên (cũng như trong tự nhiên) là mắt xích khép kín chu trình chuyển hoá sẳt tại đây.
Hai nhóm vi khuẩn Paracoccus và Pseudomonas tương ứng chiếm ưu thế trong các mẫu bùn ao nước ngọt và chân ruộng ngập nước. Đây là hai chi thuộc lófp Proíeobacteria, có nhiều đại
diện sinh trưởng kỵ khí khử NOj", bao gồm cả các loài có khả nãng sử dụng Fe(II) làm chất cho điện tử (Weber et aỉ., 2006b,c; Ratering, Schnell, 2000). Như vậy có thề kết luận ràng trong môi trường nước ngọt tại Việt Nam (đại diện là ao nước ngọt và ruộng ngập nước), Paracoccus và
16

Pseudomonas là các nhóm vi khuẩn đóng vai trò chính trong việc oxy hoá Fe(II) bằng NO3 . Tuy nhiên, trong môi trường nước lợ (và nước biển) bàng phương pháp điện di biến tính hiện chưa xác định được nhỏm chiếm ưu thế. Nguyên nhân có thể do sự kém cạnh tranh của vi khuẩn khử NO3- nói chung so với các nhóm kỵ khí khác, đặc biệt là khử sulfate trong môi trường này (Schlafleigh, 2000).
3 J . Mức độ oxy hoá Fe(II) và khử NƠ3~ của vi sinh vật trong các mẫu nghiên cứuNồng độ Fe(II) và NO3” ưong môi trường nuôi cấy được xác định theo thời gian để đánh
giá khả năng sinh trưởng của vi sinh vật trong các mẫu làm giàu (hình 3.3).
£3$
Thời gian (ngày)1 2 3 4
Thời gian (ngày)
Hình 3.3. Hoạt tính oxy boá Fe(II) (A) và khử N O 3 (B) của các quần thể vi sinh vật tại ba môi trường nghiên cứu sau khi làm giàu bằng phưong pháp MPN. Kýhiệu: o đối chứng không có vi sinh vật; ■ mẫu bùn đáy ao nước ngọt; •mẫu bùn chân ruộng ngập nước; A mẫu trầm tích nước lợ ven biển.
Như vậy các quần thể vi sinh vật được làm giàu thể hiện khả năng sinh trường với Fe(II) và NO3- khá cao. Ở cả 3 mẫu, lưọng nitrate trong môi trường bị khử gần hoàn toàn sau 4 ngày (hình 3.3B), tuy nhiên lượng Fe(II) không giảm tương ứng (hình 3.3A). Hiện tượng này cỏ thể lý giải bằng sự có mặt của nhóm vi khuẩn Anaeromyxobacter trong cà 3 mẫu phân tích, làm chuyển hoá Fe(III) ngược lại thành Fe(II).
3.4. Phân lập vi khuẩn oxy hoá Fe(II), khử NO3 - tử các mẫu nghiên cứu và đánh giá tính đa dạng của các chủng phân lập
Vi khuẩn oxy hóa Fe(II), khử NO3" được phân lập theo phương pháp pha loâng trong dãy ống thạch bán lòng (1%) (Widdel, Bak, 1992) (hình 3.4a). Các khuẩn lạc đom được phân lập dựa trên sự khác nhau về hình dạng và kích thước khuẩn lạc. Mồi khuẩn lạc được tách riêng và nuôi
cấy kỵ khí trong môi trường dịch thể chứa Fe(II) (10 mM) và NƠ3“ (5 mM) trong binh serum có nút cao su và kẹp nhôm (hình 3.4b). 12 khuẩn lạc đơn được phân lập từ các ống MPN ở độ pha
V** hỌC tìUOC GIA HM NỤI ‘VỤNG TÂM THÒNG TIN THỰ VIỆN
O ĩ / e i i

loãng cao nhếỉ có vi sinh vật phát triển (bảng 3.1) là đại diện cho vi khuẩn oxy hoá Fe(n), khừ
NO3" chiếm đa số tại ba môi trưởng nghiên cứu.
Hình 3.4. Phân lập vi khuẩn oxy hoá Fe(II) khử NOj đại diện tại các môi trườg nghiên cứu. (a) Phân lập chủng đơn thông qua phương pháp ống thạch kỵ khí bán lỏng; (b) Nuôi cấy chủng đơn vi khuẩn oxy hóa Fe(n), khử NO3” trong môi trường dịch thể ở điều kiện ky khí hoàn toàn.
Bảng 3.1. Vi khuẩn oxy hóa Fe(II), khửNC>3 phân lập được từ các môi tnròug nghiên cứu
Nguồn phân lập ChÙDg vi khuấn Đặc điểm hình thái khuẩn lạc
Chân ruộng lúa ngập nước (Tù Liêm, Hà Nội)
INI Hình tròn, bề mật nhẵn, kích thước 1 -2 mm
EN2 Hình tròn, bề mặt không nhăn, kích thước 0,25-0,5 mmIN 3 Hình tròn, bể mặt xù xi, kích thước 0,25-0,5 mmIN4 Hình tròn, bề mặt xù xi, kích thước 1 -2 mm
Trầm tích nước lợ tại biển (Vân Đồn, Quảng Ninh)
IN5 Hình tròn, bề mặt nhẵn, kích thuớc 0,25 mmIN6 Hình bầu dực, be mặt nhẵn, kích thước 0,25-0,5 mmIN 7 Hình tròn, bề mặt xù xi, kích thước 0,8 mm
EN8Hình tròn tia, mật đọ tể bao ở ngoài ít hơn phía ừong, kích thuớc 1-1,5 mm
Đáy ao nước ngọt (Từ Liêm, Hà Nội)
IN9 Hình tròn, bề mặt xù xi, kích thước 0,5-1 mm
IN10 Hình tròn không đều, kích thước 0,5-1 mmINI 1 Hình tròn, bề mặt nhẵn, kích thước 0,2-0,3 nun
INI 2 Hình đĩa lồi hai mặt, kích thước 2-2,5 mm
Tính đa dạng cùa 12 chủng vi khuẩn đã phân ỉập ờ trên được phân tích bầng phương pháp ARDRA sử đụng hai enzyme giới hạn Haelỉỉ và Mspì (hình 3.5). Kết quả thu được cho thấy các chủng này có thể xếp vào 5 nhóm đi truyền khác nhau (bảng 3.2).
18

Hue III \Isp\
Hình 3.5. Phổ điện di gen 16S rDNA của 12 chủng vi khuẩn oxy hóa Fe(II), khử NC>3~ sau khi xử lý bằng các enzyme giới hạn HaeIII và Mspl. INI - IN 12: Kỷ hiệu cùa 12 chùng đơn phân lập được từ các mẫu nghiên cứu; M: Marker 1 kb (Bioneer, Hàn quốc)
Từ kết quả phân nhóm ở bảng 3.2 kết hợp với nguồn gốc phân lập 12 chủng này (bảng3.1), có thể nhận thấy tính đa dạng di truyền của 12 chủng vi khuẩn oxy hóa Fe(II), khử NCV phụ thuộc vào địa điểm thu mẫu ban đầu. Mầu trầm tích nước lợ ven biến thể hiện mức độ đa dạng di truyền cao nhất với 4 chủng phân lập từ mẫu này (IN5, IN6 , IN7, ĨN8 ) thuộc vào 4 nhóm ARDRA khác nhau (nhóm 2, nhóm 3, nhóm 4 và nhỏm 5). Tiếp đến ỉà mẫu bùn đáy ao nước ngọt với 4 chủng (IN9, IN 10, INI 1, IN 12) thuộc vào 3 nhóm ARJDRA khác nhau (nhỏm 1, nhổm2 và nhóm 4). Thể hiện tính đa dạng di truyền thấp nhất !à mẫu bùn chân ruộng ngập nước với 4 chủng (INI, IN2, IN3, IN4) thuộc vào 2 nhóm ARDRA khác nhau (nhóm 1 và nhóm 2).
Bàng 3.2. Tính đa dạng về di truyền của 12 chủng vi khuẩn oxy hóa Fe(II) khử nitrate đã phân lập (INI - IN 12) dựa trên phân tích ARDRA
NhỏmARDRA
Chủng vi khuẩnCác đoạn DNA tạo ra sau khi xử lý 16S rDNA băng
các enzyme giói hạn (bp)H aelll Mspl
1 INI, IN2, IN4, INI 1 200, 300 300, 500
2 IN3, IN8 200, 300 500
3 IN5, IN9 200, 300 500, 8004 IN6 , IN 10, IN12 200, 300, 500 300, 500
5 IN7 200, 900 500
Ba chủng IN2, ĨN7 và IN 12 đại điện cho 3 nguồn phân lập và đại diện cho các nhóm ARDRA chính (bảng 3.2, tên chủng in đậm) được lựa chọn để tiến hành phân tích trình tự đầy đù của gen 16S rDNA và định danh khoa học. Chùng ĨN2 đại diện cho nhóm ARDRA-1 gồm 4 chùng từ hai dạng môi trường nước ngọt, chiếm trên 30% trong tổng số các chủng phân lập được. Tương tự, chùng INI2 đại diện cho nhóm ARDRA-4 gồm 3 chủng từ môi trường nước lợ và ao nước ngọt, chiếm 25% trong tổng số các chùng phân lập được. Chủng IN7 làm thành một nhóm
19

riêng biệt (ARDRA-5), chi tìm thấy ở môi trường nước lợ ven biển. Kết quả so sánh trình tự 16S rDNA của các chùng nảy với ngân hàng dữ liệu GenBank cho thấy chủng IN2 có độ tương đồng cao nhất với Anaeromyxobacter dehalogenans (99%), chùng IN7 với Pseudomonas stutzeri (98%) và chủng IN 12 với Paracoccus ferooxydant (97%) (hình 3.6).
.0.02.
100
100
100
-B acillus sub íiỉis (FJ544386)
f
rLttc
671----- -•
E
Anaeromyxobacíer sp. FAcl2 (AJ504438) Anaeromyxo bacler dehalogenans 2CP1 (NR027547)
•Aỉtaeromyxobacler clone (AB293367)IN2
Paracoccus marinus (AB185959)'aracoccusferooxydant (AY954687)
cIN12100 \-Paracoccvs sp (AM989037)
P seudom onaspuứda (FJ217182) Pseudom onas stuizeri (EU518705) TN7
• Ô-Proteobacteria
a-Proteobacteria
100
ĩ I— IPseudom onas alcaìigenes XB5 (GQ150490)
Pseudom onas nitroreducens PS2 (FJ588866)
y-Proteobacteria
Hình 3.6. Cây phân loại thể hiện mối liên quan giữa các chủng IN2, IN7, IN12 và các loài gần gũi dựa trên trình tự gen 16S rDNA. Cây được dựng theo phương pháp neighbor-joining, đơn vị = 0,02 Knuc trong trình tự nucleotide. Các số hiển thị ờ các vị trí phân nhánh là kết quả phân tích bootstrap đối với 1 0 0 0 phép so sánh (chì có các giá trị trên 50% được trình bày trên hình). B. subtiỉis là loài vi khuẩn được chọn làm outgroup. Các ký hiệu hiển thị trong ngoặc kép iả mã của trình tự gen 16S rDNA tại các ngân hàng gen.
Như vậy nhóm ARDRA-4 vói chủng IN 12 làm đại diện là các loài Paracoccus và được tìm thấy trong hai dạng môi trường nước lợ ven biển và ao nước ngọt. Paracoccus là chi vi khuẩn nằm trong phân lớp a -Proteobacteria, gồm các loài sinh trường kỵ khí tuỳ tiện, hô hấp bàng nitrate hoặc oxygen (Schapleigh, 2000). Kết hợp với kết quả phân tích PCR-DGGE ở trên có thể thấy rằng Paracoccus và Pseudomonas là hai nhóm chính oxy hoá Fe(II) khử NO3” chiếm ưu thế trong ba dạng môi trường được nghiên cứu, trong đó Paracoccus có vai trò quan trọng hom ờ ao nước ngọt và Pseudomonas ở ruộng ngập nước. Đại diện cùa cả hai nhóm này được tìm thấy ờ môi trường nước lợ ven biển, có thể là đo ảnh hường của nguồn nước ngọt từ đất liền.
Dựa trẽn kết quả so sánh trình tự 16S rDNA ở trên, các chủng ĨN2, IN7 và IN 12 được định danh lần lượt 1 à Anaeromyxobacter sp. ĨN2, Pseudomonas sp. ÍN7 và Paracoccus sp. ĨN12. Trong ba chủng được nghiên cửu chi tiết ở trên chỉ có chùng IN7 là đại diện được phân lập từ môi trường nước lợ ven biển. Bầng phương pháp PCR-DGGE trình bày ờ trên {xem phần 3.2), nhóm vi sinh vật chiếm ưu thế trong môi trường nước lợ chưa xác định được. Tuy nhiên việc phân tích các chủng đại diện phân lập được tử cà ba môi trường nghiên cứu cho phép kết luận cả

Paracoccus (đại diện là chủng IN6 ) và Pseudomonas (đại diện là chủng IN7)cũng có mặt trong dạng môi trưởng náy.
3.5. Phân tích đa dạng vỉ khuẩn trong các mối trvờng nghiên cứu bằng phưong pháp FISHFISH (Fluorescence In Situ Hybridization) là phương pháp trực tiếp xác định mối liên
quan phả hệ của vi kHiiẩn trong môi trường sống của chúng thông qua các đầu dò đánh dấu huỳnh quang có thể bắt cặp với RNA ribosome. Theo kết quả so sánh trình tự gen 16S rDNA, các chủng đại diện cho các nhóm ARDRA c hính được xếp vào các phân lớp a-, p- và y-Proteobacteria (hình 3.6), vì vậy trong thí nghiệm FISH chúng tôi sử dụng ba đầu dò ALF968, BET42a và GAM42a tương ứng bắt cặp đặc hiệu với vi khuẩn thuộc ba phân lớp trên (hỉnh 3.7).
Hình 3.7. Hinh ảnh hiển vi của tế bào vi khuẩn ưong các mẫu nghiên cứu bắt cặp với các
Tỷ lệ vi khuẩn mỗi nhóm được tính theo tỷ lệ số tế bào bắt cặp với đầu dò tương ứng trên tổng số tế bào được nhuộm DAPI. Phần không bắt cặp với một trong ba đầu dò được sử dụng ở đây được gọi là phần không xác định (KXĐ). Thí nghiệm lai với các đầu dò chi cho kết quả đối với mẫu từ hai môi trường nước ngọt là ruộng ngập nước và bùn đáy ao, mẫu trầm tích ven biển không cho kết quả dương tính đối với cả 3 đầu dò. Kết quả thống kê số tế bào bắt cặp với các đầu dò trên tổng số tế bào nhuộm DAPI được trình bày trên hình 3.8.
Hình 3.8. Kết quả phân tích đa dạng vi sinh vật trong các mẫu bằng phương pháp FISH
Kết quà thu được cho thấy số vi sinh vật không bắt cặp với một trong ba đầu dò sử dựng ờ thí nghiệm FISH chiếm một phần khá lớn trên tổng số tế bào nhuộm DAPI trong các mẫu ruộng
đầu dò huỳnh quang.
Ruộng ngập nước Bùn đáy ao
ALF96820%
ALF968 _ 30%
GAM42a20%
BET42a20%
21

ngập nước (60%) và bùn đáy ao (50%). Trong phần tế bào có bắt cặp với các đầu dò, nhóm a- Proteobacteria chiếm 20% và 30% (trên tổng số tế bào nhuộm DAPI) tương ứng trong các mẫu ruộng ngập nước và ao. Theo kết quả nghiên cứu bằng PCR-DGGE gen I6 S rDNA (phần 3.2), vi khuẩn thuộc phân lớp a -Proteobacteria với đại diện là chi Paracoccus đâ được tìm thấy chiếm đa số trong môi trường ruộng ngập nuớc, nhưng lại không được tìm thấy ờ môi trường bùn đáy ao. Tiểp theo, đầu dò BET42a (p-Proteobacteria) cho kết quả dương tính với 20% tổng số tế bào nhuộm DAPI trong mẫu bùn đáy ao nhưng không có tín hiệu với mẫu ruộng ngập nước. Điều này
cũng phù hợp với kết quả phân tích PCR-DGGE (phần 3.2), vi khuẩn thuộc phân lớp y- Proteobacíeria với đại diện là chi Pseudomonas được tìm thấy chiếm ưu thế trong môi trường ruộng ngập nước, nhưng không tìm thấy trong mẫu bùn đáy ao. Cuối cùng, đầu đò GAM42a (y- Proteobacterià) cho kết quả duơng tính với 20% tổng sổ tế bào nhuộm DAPI trong mẫu ruộng ngập nước nhưng lại không cỏ tín hiệu đối với mẫu bùn đáy ao, tuy nhiên đại diện cùa phân lớp p-Proteobacteria lại không được xác định trong phân tích PCR-DGGE ở trên. Như vậy việc bổ sung thông tin từ thí nghiệm phân tích FISH đã góp phần làm rõ nét hơn bức tranh đa dạng của vi khuẩn oxy hóa Fe(II)/khù nitrate ở các môi trường nghiên cứu.
Đối với mẫu trầm tích nước ]ợ ven biển, cả hai phương pháp phân tích đa đạng không phụ thuộc bước phân lập là PRC-DGGE và FISH đều không đem lại thông tin về tính đa dạng của vi sinh vật oxy hóa Fe(II)/khử nitrate tại đây. Tuy các đại diện thuộc chi Paracoccus (a-
Proteobacteria) và Pseudomonas (y-Proteobacteria) đều đã được tìm thấy trong môi trường này (phân tích ARDRA, phần 3.4), nhưng vai trò ưu thế cùa chúng trong môi trường không được xác định. Sự có mặt của các chủng Paracoccus và Pseudomonas trong môi trường trầm tích ven biển nhiều khả năng do ảnh hường của nguồn nước ngọt từ đất liền. Nhóm vi sinh vật thực sự chiếm ưu thế trong môi trường này có thể vẫn nằm ngoài giới hạn đặc hiệu của các đầu dò đã sử dụng trong nghiên cứu F1SH cũng như cặp mồi sử dụng trong PCR-DGGE.
3.6. Nghiên cứu đặc điểm sinh lý, phân hoại và hoạt tính sinh bọc của các chủng vỉ khuẩn đại diện
Hai chủng vi khuẩn IN2 và IN12 đại diện cho hai nhóm vi khuẩn chiếm ưu thế trong các môi trường nghiên cứu là Anaeromyxobacter và Paracoccus (phần 3.4). Hơn thế, trong quá trình thí nghiệm có thể nhận thấy rầng đây là hai chùng thể hiện khả năng sinh trường cao nhất trong số các chủng đã phân lập được. Để tìm hiểu khả năng ứng dụng của nhóm vi khuẩn oxy hoá Fe(II) khử nitrate, chúng tôi tiến hành nghiên cứu các đặc điểm sinh lý cùa hai chủng này.
Kết quả nghiên cứu cho thấy hai chùng IN2, IN 12 rất khác nhau về đặc điểm sinh lý (bảng 3.3). Trong khi chủng IN2 là vi khuẩn kỵ khí bắt buộc thỉ chủng IN 12 là vi khuẩn kỵ khí
tùy tiện, có khả năng hô hấp bằng oxygen (sinh trưởng hiếu khí). Ngoài NO3" được sử dụng làm chất nhận điện tử cuối cùng, chùng IN2 còn có khả năng sử dụng Fe(III) làm chất nhận điện từ còn chùng IN 12 không cỏ khả năng này, thể hiện trong kết quà thí nghiệm xác định sự biến đồi
22

của hàm lượng protein tổng số theo thời gian ưong dịch nuôi vi khuẩn với Fe(III) làm chất nhận điện tò cuối cùng (hỉnh 3.9). Cả hai chủng đều không sinh trường bằng khử SO42".
Thời gian (ngày)
Hình 3.9. Khả năng sinh trưởng bằng khử Fe(III) cùa hai chủng IN2 (■) và IN 12 ( • ) thông qua xác đinh hàm lượng protein tổng số trong dịch tế bào biến đổi theo thời gian.
Băng 3.3. Đặc điểm sinh lý của hai chủng vi khuẩn IN2 và IN12
Đặc điểm sinh lý IN2 IN12
Hô hấp bằng oxygen (sinh trưởng hiếu khí) - + +
0 % - + + +
1% + + + + + +Sinh trưởng phụ thuộc độ mặn cùa 2 % + + + + + +môi trường (% NaCl) 3% + + + + + +
4% + + + + +
5% + 4- + +
Lactate + + + + + +
Chất cho điên tử (để khử nitrate)Acetate + + + ■+ H—b
Ethanol + + + + + +
Benzoate + + + +
Chất nhận điện tử (để oxy hoá Na- lactate)
n o 3~
Fe(III)
SO42"
+ + +
+ + +
+ + +
Chú thích: + + +: sinh trưởng mạnh; + +: sinh trưởng trung bình; +: sinh trường yếu; không sinh trưởng.
Đối với các nguồn điện từ khác nhau phục vụ cho quá trình khử nitrate, ngoài Fe(II) đã được sử đụng để phân lập và nuôi cấy từ ban đầu, lactate, acetate, benzoate và ethanol đều được
23

oxy hoá bởi hai chủng IN2 và IN12. Trong trường hợp benzoate, một acid hữu cơ chứa vòng thơm và thường khó bị oxy hoá hơn các hợp chất còn lại, chủng INI 2 thể hiện khả năng oxy hoá kém hơn hẳn so với chủng IN2.
Độ mận lả một Ưong các yếu tố môi trường quan trọng ảnh hưởng đến sinh lý cùa vi sinh vật trong tự nhiên, do vậy ở đây chúng tôi tiến hành nghiên cứu ảnh hường của nồng độ muối trong môi trường đối với khả năng sinh trưởng của hai chủng vi khuẩn quan tâm. Kết quả nghiên cứu cho thấy chủng IN2 có nhu cầu về muối để sinh trưởng, chủng INI2 thi không. Trong khi chủng INI2 có thể sinh trưởng trong môi trường không cần bổ sung NaCl thì chủng IN2 chi phát triển khi NaCl có mặt ở nồng độ 1% trở lên. Tuy nhiên, chủng IN2 lại có khả năng chịu muối cao hơn IN12, chủng này vẫn phát triển tốt ở nồng độ muối 5% trong khi đó tốc độ sinh trường của IN 12 bị ảnh hưởng đáng kể ở các nồng độ muối cao hom 3%. Điều này cũng phù hợp với đặc điểm phân bố của hai chùng vi khuẩn trong tự nhiên, cụ thể là chủng IN2 đại diện cho nhóm vi khuẩn được tìm thấy ở nhiều dạng môi trường khác nhau, bao gồm cả nước ngọt và nước lợ, trong khi đó chùng IN 12 đại diện cho nhóm chi tim thấy ở bùn đáy thuỷ vực nước ngọt.
M
4 & 9 •r
ۥ
0.02
Hỉnh 3.10.
9301000
1000
SaciiầíS subttito (FJ544 386)
AnMTomyivbacter clone (AB2933Ổ7)
Anotromyxìbacter dehabgtnans (EP067314) N2K Mr
UArumromyìobactor sp. FAcl2 (AJ504438) La dehabgenans 2CP1 (NR027547)
^Anaervmymbacter clone (DQ39499Ố)
Bacúkiĩ subttỈB (FJ544386) -Paracoccus arninopMus (D32239)
■Paracoccus aminovorans (D3224Q) 'aracoccus sp (AM989037)
■Paracoccus marịms (AB185959)_'Pơracoocus denUrựkans (AI288159)
iÓÕÕÍParacoccus jimoxydant (AY954687)
Hình thái tể bào và vị trí phân loại của hai chủng IN2 và ĨN12. Hình thái tế bào được quan sát trên kính hiển vi quang học, độ phóng đại 1 0 0 0 lần (đơn vị = 5
Jim). Cây phân loại được dựng theo phương pháp neighbor-joining, đom vị = 0 02 Knuc trong trình tự nucleotide. Các số hiển thị ở các vị tri phân nhánh là kết quả phân tích bootstrap đối với 1 0 0 0 phép so sánh (chi có các giá trị trên 500 được trình bày trên hình). B. subtilis là loài vi khuẩn được chọn làm outgroup.
Chủng IN2 gồm nhừng tế bào dạng trực khuẩn, kích thước 1x4-5 um, chuyển động tích cực (hình 3.10). So sánh trinh tự gần đủ gen mã hóa cho 16S rDNA với ngân hàng dữ liệu
24

GenBank cho phép xếp chùng IN2 vào chi Anaeromyxobacter (!oài gần gũi nhất là A. dehaỉogenans, 99% tương đồng). Như vậy chùng IN2 được bổ sung vào danh sách chi Anaeromyxobacter với tên khoa học là Anaeromyxobacter sp. IN2 và mã trình tự gen 16S rDNA trong GenBank là FJ939131 (hình 3.10). Anaeromyxobacter là một chi được biết đến với các đại diện sinh trưởng kỵ khí bắt buộc bàng oxy hoá các hợp chất hữu cơ để khử Fe(III) (Treude et aỉ., 2003). Trong nghiên cứu này, lần đầu tiên vi khuẩn Anaeromyxobacter mà đại diện là chùng ĨN2 thể hiện khả năng sinh trưcmg bằng oxy hoá Fe(II)/khử nitrate, tuy nhiên hình thức sinh trưởng này không vượt trội so với sinh trường kỵ khí khử Fe(III),
Chủng IN12 gồm những tế bào hình oval có kích thước 1,5x1.5-2 um, chuyển động chậm. So sánh trình tự gần đủ của gen 16S rDNA với ngân hàng dữ liệu GenBank cho phép xếp chúng IN 12 vào chi Paracoccus (loài gần gũi nhất là p. aminovorans, 98% tương đồng). Paracoccus là chi vi khuẩn nằm trong phân lớp a-Proteobacíeria, gồm nhiều loài có khả năng hô hấp bằng oxygen (sinh trưởng hiếu khí), đồng thời hô hấp bằng khử nitrate (Shapleigh, 2000). Nhiều đại diện của chi Paracccus được tìm thấy trong các điều kiện tối ưu cho khử nitrate, như đáy các thuỷ vực (Shapleigh, 2000) hay trong các hệ thống xử lý nước thải loại bỏ nitrogen, ví dụ như P. denitrificans, p. ferooxygendans (Bitton, 1999). Chùng IN 12 được định danh khoa học là Paracoccus sp. IN 12 và có mã trinh tự gen 16S rDNA trong GenBank là GU084390 (hình 3.10).
Mặc đù cùng được phân lập thông qua bước làm giàu bàng dãy MPN với chất cho và nhận điện tử tương ứng là Fe(II) và nitrate, hai chủng IN2 và IN 12 lại đại diện cho hai nhóm vi khuẩn khác nhau tham gia chu trình sắt ở môi trường trung tính, cụ thể là oxy hoá Fe(II) thành Fe(IlI) bằng nitrate và khử Fe(III) thành Fe(II) bàng các hợp chất hữu cơ. Tuy cũng có khả năng sinh trường cao trong môi trường có Fe(II) và nitrate nhung chủng IN2 sẽ khỏ có khả năng úng dụng vào mục đích loại bỏ Fe(II) và nitrate trong các nguồn nước ô nhiễm vì bị ảnh hưởng bởi khả năng sừ dụng Fe(III) làm chất nhận điện từ cuối cùng để tạo năng lượng. Khác với IN2, chủng IN12 là một vi khuẩn chi sinh trưởng nhờ oxy hoá Fe(ll) bằng nitrate, không có khả năng khử Fe(III). Hơn thế nữa, chủng vi khuẩn này có khả năng sinh trường trong nhiều đều kiện môi trường khác nhau như có mặt hay không có mặt oxygen, thích nghi với nhiều loại nguồn điện tử khác nhau (như Fe(II), các acid hữu cơ, rượu) và sinh trường tốt trong môi trường có độ mặn dao động trong dải 0-3% (đặc trưng cho cả môi trường nước ngọt và nước lợ). Với những ưu điểm kể trên, chùng IN 12 hứa hẹn có tiềm năng ứng dụng vào việc loại bò nitrogen và Fe(II) trong các nguồn nước ô nhiễm.
25

3.5 3.5
- 3 _2 E
■ 2.5 a-QJ Uh
- 2
- 1.50 1 2 3 4 5
Thời gian (ngày)
Hình 3.11. Loại bò Fe(II) (•) và N 03- (■) trong môi truờng do chủng vi khuẩn IN 12.
Hoạt tính oxy hóa Fe(II) và khử nitrate của chủng IN 12 được xác định ờ điều kiện kỵ khí với Fe(II) là chất cho điện từ và nitrate làm chất nhận điện tử cuối cùng. Kết quả xác định nồng độ các chất cho và nhận điện từ trong môi trường biến đổi theo thời gian cho thấy sự giảm nồng độ Fe(II) và nitrate diễn ra song song với tốc độ đáng kể (hình 3.11).
Bên cạnh đó khả năng sử dụng Mn(II) làm chất cho điện tử để khử nitrate cũng được kiểm tra, tuy nhiên chủng IN 12 không thể hiện sinh trưởng rõ rệt trong điều kiện nuôi cấy này. Theo một số nghiên cứu đã công bố, hoạt tính oxy hoá Mn(II), khử nitrate bằng con đường sinh học được xác định trong một số quần thể vi sinh vật ở đáy các thuỷ vực, nhưng cho đến nay chưa có chủng vi sính vật nào được phân lập trong phòng thí nghiệm với hoạt tính kể trên (Vandenebeele eí a l, 1995; Gounot, 1994).
26

KẾT LUẬN
- Số lượng vi khuẩn oxy hoá Fe(II) khử NOị” trong các mẫu bùn thu thập ở thủy vực nước ngọt, chân ruộng ngập nước và vùng nước lợ ven biển được xác định nằm trong khoảng 103 - 104 TB/g, trong đó mẫu bùn ờ chân ruộng ngập nước có số lượng tế bào vi khuẩn
này cao hơn cả (9,3 X 103 TB/g).
- Phân tích thành phân loài bằng phương pháp PCR-DGGE đối với gen 16S rDNA cho thấy Anaeromyxobacter là một trong những nhóm chiếm ưu thế có mặt cả ở ba dạng môi trường sinh thái trên, đóng vai trò khép kín chu trình chuyển hoá sẩt trong các môi trường nghiên cứu thông qua khả năng sinh trưởng kỵ khí khử Fe(III). Hai nhóm Paracoccus và Pseudomonas được tìm thấy tương ứng ở mẫu bùn ao và ruộng, là hai nhóm vi khuẩn chính thực hiện quả trình oxy hoá Fe(II) khử NƠ3- trong hai dạng môi trường sinh thái kể trên. Ngoài ra, các chủng vi khuẩn thuộc chi Paracoccus (ÍN6 ) và Pseudomonas (IN7) còn được tìm thấy ở môi trường nước lợ ven biển, chứng tỏ các nhóm vi khuẩn này cũng đóng vai trò nhất định trong môi trường nước lợ.
- Vi khuẩn oxy hoá Fe(II) khử NCV tại các môi trường nghiên cứu có tính đa dạng khá cao. Mười hai chủng đom phân lập từ các mẫu MPN được xếp vào 5 nhóm ARDRA khác nhau trong phân tích sử dụng hai enzyme giới hạn HaeIII và Mspỉ. Ba chủng đại diện là IN2, ĨN7 và IN 12 có ưình tự gen 16S rDNA có độ tương đồng cao với các loài vi khuẩn Anaeromyxobacíer dehaỉogenans (99%), Pseudomonas stutzeri (97%) và Paracoccus ferrooxydant (98%).
- Hai chùng vi khuẩn IN2 và IN 12 đại điện cho hai nhóm tham gia chu trình chuyển hoá sắt tại một số môi trường sinh thái khác nhau ờ Việt nam, trong đó chủng IN2 tham gia khừ Fe(III) thành Fe(II) bằng các hợp chất hữu cơ và chùng IN 12 tham gia oxy hoá Fe(IÍ) thành Fe(III) bằng nitrate. Vai ưò sinh thái của hai chủng này được khẳng định thông qua các đặc điểm sinh lý của chúng, cụ thể là khả năng sinh trưởng với các chất cho và nhận điện tử khác nhau và mức độ sinh trưởng tại các nồng độ muối khác nhau trong môi trường.
- So sánh trinh tự gần đủ cùa gen 16S rDNA với các loài đã công bố trên GenBank cho phép định danh hai chủng vi khuẩn này là Anaeromyxobacter sp. IN2 (mã trình tự đăng ký trên GenBank là FJ939131) và Paracoccus sp. IN 12 (mã trinh tự đăng ký trên GenBank là GU084390).
- Với hoạt tính cao trong việc oxy hoá Fe(II) và khử nitrate, chủng IN 12 có tiềm năng úng đụng thực tế trong việc xử ]ý các nguồn nước nhiễm nitrogen và ion kim loại Fe(II).
27

KIẾN NGHỊ- Nghiên cứu tìm giá thể phù hợp để tạo chế phẩm chứa vi khuẩn oxy hóa Fe(II) khử nitrate.- Nghiên cứu thử nghiệm áp dụng vi khuẩn oxy hóa Fe(II) khử nitrate để xử lý nước ngầm
nhiễm sắt và nitơ.
TÀI LIỆU THAM KHẢO
Tiếng Việt
1. Lê Đửc (2004) Một sổ phương pháp phân tích môi trường. NXB ĐHQGHN.
2. Trần Thị Kiều My, Nguyễn Hà Thanh (2006) Chuyền hóa sắt trong cơ thể và quá tải sắt ở một số bệnh máu. Hội nghị Khoa học ngành Huyết học - Truyền máu Quốc gia
Tiếng Anh
3. Amann RI, Stromley J, Devereux R. Key R, Stahl DA (1992). Molecular and microscopic identification of sulfate-reducing bacteria in multispecies biofilms. Appl. Environ. Microbiol. 58: 614-623.
4. American public health association (1996) Standard methods for the examination of water and waste water including bottom sediments and sludge. 604-609.
5. Benz M, Brune A, Schink B (1998) Anaerobic and aerobic oxidation of ferrous iron at neutral pH by chemoheterotrophic nitrate-reducing bacteria. Arch Microbiol 169: 159-165.
6 . Bitton G. Wastewater Microbiology. Willey-Liss, Inc. Toronto, Canada, 1999.
7. Brewer PG, Spencer DW (1971) Colorimetric determination of manganese in anoxic waters. Limnology and Oceanography. 16: 107-110
8 . Ciani A, Goss KU, Schwarzenbach RP (2005) Light penetration in soil and particulate minerals. Europ. J. Soil Sci. 56: 561-574.
9. Dhillon A, Edwards K, Webb E, Roger D, Sogin M (2005) Marinobacter aquaeolei gene expression studies for clues to neutrophilic iron oxydation. NAỈ 2005 biannual meeting o f the NASA Astrology Institute.
10. DIN 38406-EỈ-1 (1983) German standard methods for the examination of water, waste water and sludge; cation (group E); determination of ừon (El).
11. Edwards KJ, Rogers DR, Wirsen CO, McCollom TM (2003) Isolation and characterization of novel psychrophilic, neutrophilic, Fe-oxidizing, chemolithoautotrophic a- and p- Proteobacteria from the deep sea. Appl. Environ. Microbiol. 69: 2906-2913.
12. Ehrlich H.L (1996) Geomicrobiology, 3rd Ed, revised and expanded, Marcel Dekker Inc New York.
28

13. Ehrenberg (1919) Microbiology o f the iron - depositing bacteria, “Gallionella ferruginea". Ellis D, Methuen & Co. Ltd.
14. Ehrenreich A, Widdel F (1994) Anaerobic oxidation of ferrous iron purple bacteria, a new type of phototrophic metabolism. Appl Environ. Microbiol. 60 (12) 4517-4526.
15. Ehrlich HL, Newman DK (2008) Geomicrobiology, 5th Edition, CRC Press.
16. Emerson D, Weiss JV (2004) Bacterial iron oxidation in circumneutral freshwater habitats: findings from the field and the laboratory. Geomicrobiol. J. 21:405 - 414.
17. Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39: 783-791.
18. Gounot AM (1994) Microbial oxidation and reduction of manganese: consequences in groundwater and applications. FEMS Microbiol. Rev. 14: 339.
19. Hafenbradl D, Keller M, Dirmeier R, Rachel R, Rossagel p, Burggraf s, Huber H, Stetter KO (1996) Ferroglobus placidus gen. nov., sp. nov., a novel hyperthermophilic archaeum that oxidizes Fe2+ at neutral pH under anoxic conditions. Arch. Microbiol. 166: 308-314.
20. Hauck s, Benz M, Brune A, Schink B (2001) Ferrous iron oxidation by denitrifying bacteria in profiindal sediments of a deep lake (Lake Constance). FEMS Microbiol Ecol 37: 127-134.
21. Heising s, Schink B (1998) Phototrophic oxidation of ferrous iron by a Rhodomicrobium vannielii strain. Microbiol. 144: 2263-2269.
22. Heising s, Richter L, Ludwig w , Schink B (1999) Chlorobium ferrooxidans sp. nov., a phototrophic green sulfur bacterium that oxidizes ferrous iron in coculture with a “Geospirillum" sp. strain. Arch. Microbiol. 172:116-124.
23. Hohmann c , Winkler E, Morin G, Kappler A (2009) Anaerobic Fe(II)-oxygendizing bacteria showAs resistance and immobilize As during Fe(III) mineral precipitation. Environ. Sci. Technol.
24. Kappler A, Newman DK (2004) Formation of Fe(III)-minerals by Fe(II)-oxidizing photoautotrophic bacteria. Geochim Cosmochim Acta. 68:1217-1226.
25. Konhauser KO (Ỉ997) Bacterial iron biomineralisation in nature. FEMS Microbiol. Rev. 20' 315.
26. Kuetzing (1919) Microbiology o f the iron - depositing bacteria, “Leptothrix ochracea Ellis D, Methuen & Co. Ltd.
27. Lack JG, Chaudhuri SK, Kelly SD, Kemner KM, O'Connor SM, Coates JD (2002) Immobilization of radionuclides and heavy metals through anaerobic bio-oxidation of Fe(II) Appl. Environ. Microbiol. 6 8 : 2704-2710.
29

28. Lundberg JO, Weitzberg E, Cole JA, Benjamin N (2004) Nitrate, bacteria and human health.
Nature'. 593-602
29. Lovley D.R. (1991) Dissimilatory Fe(III) and Mn(IV) reduction. Microbiol. Rev. 55: 259.
30. Lovley D.R. (1997) Microbial Fe(III) reduction in subsurface environments. FEMS Microbiol. Rev. 20: 305.
31. Marmur Ỉ (1961) A procedure for the isolation of deoxygenribonucleic acid from microorganisms. JM ol Biol 3: 208-218.
32. Muyzer G, De Waal EC, Utterlinden AG (1993) Profiling of complex microbial population by denaturing gradient gel electrophoresis analysis of polymerase chain reaction amplified genes coding for 16S rARN. Appl Environ Microbiol 59: 695-700.
33. Nealson K.H, Saffarini D. (1994) Iron and manganese in anaerobic respiration: environmental significance, physiology and regulation. Annu. Rev. Microbiol. 48: 311.
34. Ratering s (1999) Iron cycle in Italian rice field soil: localization of the redox processes and charactarization of the involved microorganisms. PhD Dissertation. Department of Microbiology, University of Marburg (Germany).
35. Ratering s, Schnell s (2000) Nitrate-dependent iron(II) oxidation in paddy soil. Environ Microbiol 3: 100-109.
36. Ravenschlag K, Sahm K, Pemthaler J, Amann R (1999) High bacterial diversity in permanently cold marine sediments. Appl. Environ. Microbiol. 65: 3982.
37. Renz JA, Kraiya c , Luther GW, Emerson D (2007) Control of ferrous iron oxydation within circumneutral microbial marts by cellular activity and autocatalysis. Environ. Sci. Technol. 41: 6084-6089.
38. Saitou N, Nei M (1987) The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol Evol 4: 406-425.
39. Shapleigh JP (2000) The Denitrifying Prokaryotes. In Dworkin M, Falkow s, Rosenberg E Schleifer KH, Stackerbrandt E, eds. The prokaryotes: an evolving electronic resource for the microbiological community. Springer-Verlag, New York.
40. Senko JM, Dewers TA, Krumholz LR (2005) Effect of oxidation rate and Fe(II) state on microbial nitrate-dependent Fe(III) mineral formation. ,4/?/?/. Environ. Microbiol 71: 7172- 7177.
41. Senn DB, Hemond HF (2002) Nitrate controls on iron and arsenic in an urban lake. Science 296: 2373-2376.
42. Straub KL, Benz M, Schink B, Widdel F (1996) Anaerobic, nitrate-dependent microbial oxidation of ferrous iron. Appl Environ Microbiol 62: 1458-1460.
30

43. Straub KL, Buchholz-Cleven BEE (1998) Enumeration and detection of anaerobic ferrous ừon-oxidizing, nitrate-reducing bacteria from diverse European sediments. Appl Environ
Microbiol 64:4846-4856.
44. Straub KL, Rainey FA, Widdel F (1999) Rhodovulum iodosum sp. nov. and Rhodovulumrobiginosum sp. nov., two new marine phototrophic ferrous-iron-oxidizing purple bacteria. Ị
Int. J. Syst. Bacteriol. 49: 729-735. 1
45. Straub KL, Benz M, Schink B (2001) Iron metabolism in anoxic environments at near '
neutral pH. FEMS Microbiol Ecol. 34: 181-186.
46. Temple KL, Colmer AR (1951) The autotrophic oxidation of iron by a new bacterium: Thiobacilỉus ferrooxidans. J. Bacteriol. 62: 605-611.
47. Treude N, Rosencrantz D, Liesack w, Schnell s (2003) Strain FAcl2, a dissimilatory iron- reducing member of the Anaeromyxobacter subgroup of Myxococcales. FEMS Microbiol E col44 : 261-269.
48. Tricker AR, Preussmann R (1991) Carcinogenic N-nitrosamines in the diet: occurrence, formation, mechanisms and carcinogenic potential. Mut. Res. 259: 277-289.
49. Vandenebeele J, De Beer D, Germonpre R, Van De Sande R, Verstraete w (1995)Influence of nitrate on manganese removing microbial consortia from sand filters. Wat. Res.29: 579.
50. Weber KA, Picardal FW, Roden EE (2001) Microbially catalyzed nitrate-dependent ■oxidation of biogenic solid-phase Fe(II) compounds. Environ. Sci. Technol. 35: 1644-1650.
51. Weber KA, Achenbach LA, Coates JD (2006a) Microorganisms pumping iron: anaerobic microbial iron oxidation and reduction. Nature 4: 752-764.
52. Weber KA, Pollock J, Cole KA, O’Connor SM, Achenbach LA, Coates JD (2006b) Anaerobic nitrate-dependent iron(II) bio-oxidation by a novel Lithoautotrophic Betaproteobacterium, strain 2002. Appl Environ Microbiol. 72: 686-694.
53. Weber KA, Urrutia MM, Churchill PF, Kukkadapu RK, Roden EE (2006c) Anaerobic redox cycling of iron by freshwater sediment microorganisms. Environ Microbiol 8 : 100- 113.
54. Widdel F, Schneil s, Heising s, Ehrenreich A, Assmus A, Schink B (1993) Ferrous iron oxygendation by anoxygengenic phototrophic bacteria. Nature 362: 834.
55. Widdel F, Bak F (1992) Gram-negative mesophilic sulfate-reducing bacteria. In Balows A Trilper HG, Dworkin M, Harder w, Schleifer KH eds. The Prokaryotes, 2nd ed. Springer Berlin Heidelberg New York: 3352-3378.
56. Zhou J, Bruns MA, Tiedje JM (1996) DNA recovery from soils of diverse composition Appl Environ Microbiol, 62: 316-322.
31

PHỤ LỤC
IN2 16S rDNA 1429 bp, Ánaeromyxobacíer sp.ta ca ca tg c a g tc g a g c g a g a a a g cc c g c a a g g g tg a g ta a a g c g g cg c a c g g g tg cg ta a c a c g tg g g ta a tc tg ccc ta g a g tc c g g a a ta a c tc g c c g a a a g g c g tg c ta a tg c c g g a tg a g a c c a c g g g a g cc tcg g c tc c tg c g g g a a a a gg tg g c c tc tg ta c a c a a g c ta tc g c tc ta g g a tg a g c c c g c g g c c c a tc a g c tc g ttg g c g g g g ta a tg g c c c a c c a ag g ca a cg a cg g g ta g c tg g tc tg a g a g g a c g a tca g c c a c a ctg g a a c tg a g a c a c g g tcc a g a c tc c ta cg g g a g g ca g c a g tg g g g a a tc ttg c g c a a tg g g c g a a a g c c tg a c g c a g c a a c g c c g c g tg tg tg a tg a a g g tc ttc g g a tc g taa a g c a c tg tc g cg a g g g a cg a a ta a g g g a cg g g cg a a ca g tccg tttcg a tg a cg g ta cc tcg a g a g g a a g ca ccg g cta a c tc tg tg c c a g c a g c c g c g g ta a ta c a g a g g g tg c a a g c g ttg ttc g g a a tta ttg g g c g ta a a g c g c g tg ta g gc g g c c ta g c a a g tc g g a tg tg a a a g c c c tc g g c tta a c c g a g g a a g tg c g ttc g a a a c ta c tg g g c ttg a g ta c c g g ag a g g g tg g cg g a a ttcc c g g tg ta g a g g tg a a a ttc g ta g a ta tc g g g a g g a a c a c c a g tg g c g a a g g c g g cc a c c tgg a c g g a ta c tg a c g c tg a g a c g c g a a a g c g tg g g ta g c a a a c a g g a tta g a ta c cc tg g ta g tc ca c g c tg ta a a c g atg a g c g c ta g g tg ttg c g g g tg ttg a c c c c tg c a g tg c c g c a g c ta a c g c a tta a g c g c tc c g c c tg g g a a g ta c g g cc g c a a g g c ta a a a c tc a a a g g a a ttg a c g g g g g c c cg c a c a a g c g g tg g a g c a tg tg g ttta a ttc g a c g c a a cg c g ca g a a c c tta c c tg g tc ttg a c a tc c tc g g a a tc c c tc a g a g a tg a g g g g g tg c c c g c a a g g g a a c c g a g a g a c a g g tgc tg c a tg g c tg tc g tc a g c tc g tg tc g tg a g a tg t tg g g t ta a g tc c c g c a a c g a g c g c a a c c c c tg c c g tta g t tg cc a tc a ttc a g ttg g g c a c tc ta a c g g g a c tg c c g g c g tc a a g c c g g a g g a a g g tg g g g a tg a c g tc a a g tc c tc a tg gc c tt ta tg a c c a g g g c ta c a c a c g tg c ta c a a tg g c c g g ta c a g a g g g tc g c c a a g tc g c g a g a c g g a g c ta a tc c c ag a a a a c c g g tc tc a g ttc g g a ttg g a g tc tg c a a c tc g a c tc c a tg a a g tc g g a a tc g c ta g ta a tc g c g g a tc a g c ac g c c g c g g tg a a ta c g ttc c c g g g c c ttg ta c a c a c c g c c c g tc a c a c c a tg g g a g tc a g ttg c tc c a g a a g tg g c c gcgccaacccgcaagggaggcaggtc
IN12 16S rDNA 1319 bp, Paracoccus sp.TAACGCGTGGGAATGTGCCCTTCTCTACGGAATAGCCCTGGGAAACTGGGAGTAATACCGTATACGCCCTATTGGGGAAAGATTTATCGGAGAAGGATCAGCCCGCGTTGGATTAGGTAGTTGGTGGGGTAATGGCCTACCAAGCCGACGATCCATAGCTGGTTTGAGAGGATGATCAGCCACACTGGGACTGAGACACGGCCCAGACTCCTACGGGAGGCAGCAGTGGGGAATCTTAGACNATGNGGGCAACCCTGATCTAGCCATGCCGCGTGAGTGATGAAGGCCTTAGGGTTGTAAAGCTCTTTCAGCTGGGAAGATAATGACGGTACCAGCAGAAGAAGCCCCGGCTAACTCCGTGCCAGCAGCCGCGGTAATACGGAGGGGGCTAGCGTTGTTCGGAATTACTGGGCGTAAAGCGCACGTAGGCGGACCGGAAAGTCAGAGGTGAAATCCCAGGGCTCAACCTTGGAACTGCCTTTGAAACTATCGGTCTGGAGTTCGAGAGAGGTGAGTGGAATTCCGAGTGTAGAGGTGAAATTCGTAGATATTCGGAGGAACACCAGTGGCGAAGGCGGCTCACTGGCTCGATACTGACGCTGAGGTGCGAAAGCGTGGGGAGCAAACAGGATTAGATACCCTGGTAGTCCACGCCGTAAACGATGAATGCCAGTCGTCGGGCAGCATGCTGTTCGGTGACACACCTAACGGATTAAGCATTCCGCCTGGGGAGTACGGTCGCAAGATTAAAACTCAAAGGAATTGACGGGGGCCCGCACAAGCGGTGGAGCATGTGGTTTAATTCGAAGCAACGCGCAGAACCTTACCAACCCTTGACATCGCAGGACAGCCCGAGAGATCGGGTCTTCTCGTAAGAGACCTGTGGACAGGTGCTGCATGGCTGTCGTCAGCTCGTGTCGTGAGATGTTCGGTTAAGTCCGGCAACGAGCGCAACCCACGTCTTTAGTTGCCAGCATTCAGTTGGGCACTCTAAAGAAACTGCCGATGATAAGTCGGAGGAAGGTGTGGATGACGTCAAGTCCTCATGGCCCTTACGGGTTGGGCTACACACGTGCTACAATGGTGGTGACAGTGGGTTAATCCCCAAAAGCCATCTCAGTTCGGATTGGGGTCTGCAACTCGACCCCATGAAGTTGGAATCGCTAGTAATCGCGGAACAGCATGCCGCGGTGAATACGTTCCCGGGCCTTGTACACACCGCCCGTCACACCATGGGAGTTGGGTCTACCCGACGGCCGTGCGCCAACCTCGCAAGAGGAGGCAGCGGACCACGGTAGGCTCAGCGACTGGGGTGAAGT
IN7 I6S rDNA 576 bp, Pseudomonas sp.
TGCAGTCGAGCGGATGAGTGGAGCTTGCTCCATGATAAGCGGCGGACGGGTGAGTAATGCCTAGGAATCTGCCTGGTAGTGGGGGACAACGTTTCGAAAGGAACGCTAATACCGCATACGTCCTACGGGAGAAAGT
Phụ lục 1. Các trình tự gen

GGGGGATCTTCGGACCTCACGCTATCAGATGAGCCTAGGTCGGATTAGCTAGTTGGTGAGGTAAAGGCTCACCAAGGCGACGATCCGTAACTGGTCTGAGAGGATGATCAGTCACACTGGAACTGAGACACGGTCCAGACTCCTACGGGAGGCAGCAGTGGGGAATATTGGACAATGGGCGAAAGCCTGATCCAGCCATGCCGCGTGTGTGAAGAAGGTCTTCGGATTGTAAAGCACTTTAAGTTGGGAGGAAGGGCAGTAAGTTAATACCTTGCTGTTTTGACGTTACCAACAGAATAAGCACCGGCTAACTTCGTGCCAGCAGCCGCGGTAATACGAAGGGTGCAAGCGTTAATCGGAATTACTGGGCGTAAAGCGCGCGTAGGTGGTTCGTTAAGTTGGATGTGAAAGCCCCGGGCTCAACCTGGGAACTGCATCCA

Phụ lục 2. Tham gia đào tạo sau đại họcHọc viên cao học:
Nguyễn Thị TuyềnKhoá cao học 2007 — 2009, Bộ môn Vi sinh, ĐHKHTN, ĐHQGHN Tên đề tài: Vi khuẩn sinh tnrởng nhờ ôxy hoá Fe(II), khử nitrat ở Việt nam: tỉnh đa
dạng và khả năng ứng dụng.Thời gian bảo vệ luận vãn: ngày 11 tháng 12 năm 2009

ĐẠI HỌC QUỐC GIA HÀ NỘI
TRƯỜNG ĐẠI HỌC KHOA HỌC Tự NHIÊN
Nguyễn Thị Tuyền
VI KHUẨN OXY HÓA Fe(H) VÀ KHỬ NITRATE Ở VIỆT NAM:
TÍNH ĐA DẠNG VÀ TlỂM NĂNG ỨNG DỤNG
Chuyên ngành: Vi sinh vật học
Mã số: 60 42 40
LUẬN VĂN THẠC s ĩ KHOA HỌC
NGƯỜI HƯỚNG DÂN KHOA HỌC: TS. ĐINH THÚY HẰNG

Phụ lục 3. Các công trình khoa học đã công bốSTT Tên công trình công bố Tác giả Noi đăng
1 Đa dạng vi khuẩn oxy hoá Fe(II), khử NO3 tại một số môi trường sinh thái ở Việt Nam
Nguyễn Thị Tuyển, Đinh Thuý Hằng
Tạp chí Công nghệ Sinh học, VAST
2 Nghiên cứu vi khuẩn ưu thế tham gia chu trinh Fe trong các điều kiện môi trưởng khác nhau tại Việt Nam
Nguyễn Thị Tuyển, Nguyễn Minh Giảng, Đinh Thuý Hằng
Tạp chí Khoa học Công nghệ, ĐHQGHN
3 Nghiên cứu đa dạng vi khuẩn oxy hoá Fe(II), khử N 03' tại một số môi trường sinh thái ở Việt Nam và khả năng ứng dụng.
Nguyền Thị Tuyền, Đinh Thuý Hẳng
Poster tham gia Hội nghị toàn quốc về đa dạng và sinh thái sinh vật, Hà Nội 10/2009
4 Trình tự gen 16S rDNA cùa chùng vi khuẩn Anaeromyxobacter sp. IN2
Nguyên Thị Tuyên, Đinh Thuý Hằng
FJ939131, GenBank
5 Trinh tự gen 16S rDNA của chủng vi khuẩn Paracoccus sp. ĨN1 2
Nguyễn Thị Tuyền, Đinh Thuý Hằng
GU084390, GenBank

Luân vãn th ạ c e ĩkhoâ học * 9
LỜ I CẢM Ơ N
v ề tài luận văn được thực hiện với sự hỗ trợ kinh phí từ để tà i P ặc biệt cấp
Đại học Quốc gia Hà Nội, mã số (3(3.07.23.
Để có th ể hoàn thành luận văn này, trước tiên, tôi muốn bày tổ lỏng biết ơn
õâu ôắc tới Tiến õTĐinh Thúy Hằng, Trưởng phòng 5inh thái */i 5inh vật, Viện Vi ỗítih
vật và Công nghệ 5inh học, Đạ\ học Quốc gia Hà Nội dã định hướng nghiên cứu, trực
tiếp hướng dẫn và chỉ bảo tận tình cho tôi trong suốt thời gian nghiến cứu.
Tôi cũng mong muốn dược 0Ởi lời cảm ơn chân thành nhât tới Ban lãnh ảạo vằ
các cán bộ Viện v\ sinh vật và Công nghệ Sinh học, Đại học Quôc gia Hà Nội ăẵ tạo
diểu kiện thuận lợi vẩ trang thiất bị và cơ sở v |t chất cho tôi Hoàn thành nghiên cứu
này.
Qua đây, 'Dổi cũng muốn được bày tỏ ỉòng biết ơn chân thành tới cấc thầy cô
giáo, cán bộ Khoa Sinh học, Trường Pại Học Khoa học Tự nhiên, Đại học Quốc Gia Hà
Nội dã giúp đỡ và trang bị những kiến thức hừu ích cho tôi trong suốt thời gian học
tập tại trường.
Cuối cùng, tôi xin bày tỏ lòng biết ơn õâu sắc tới ỳ a đmh, bạn bè thân thiết,
nhữn người đã ỉuỗn cổ vũ, độnq viên tôi vượt CỊU3 mọi khố khăn trong quá trình học
tập và nghiên cứu.
//à /Vọ/, ngày # tháng ÀA năm S009
Học viên
Nguyễn Thị Tuyển
Chuyên ngành Vi õinh v ậ t học

Phụ lục 3. Các công trình khoa bọc đã công bốSTT Tên công trình công bổ Tác giả Nơi đăng
1 Đa dạng vi khuẩn oxy hoá Fe(ỈI), khử
NC>3~ tại một số môi trường sinh thái ở
Việt Nam
Nguyễn Thị Tuyển, Đinh Thuý Hằng
Tạp chí Công nghệ Sinh học, VAST
2 Nghiên cứu vi khuẩn ưu thể tham gia chu trình Fe trong các điều kiện môi trường khác nhau tại Việt Nam
Nguyễn Thị Tuyền, Nguyễn Minh Giảng, Đinh Thuý Hằng
Tạp chí Khoa học Công nghệ, ĐHQGHN
3 Nghiên cứu đa dạng vi khuẩn oxy hoá
Fe(II), khử NO3" tại một sổ môi trường sinh thái ở Việt Nam và khả năng ứng dụng.
Nguyền Thị Tuyền, Đinh Thuý Hằng
Poster tham gia Hội nghị toàn quốc về đa dạng và sinh thái sinh vật, Hà Nội 10/2009
4 Trình tự gen I6 S rDNA của chùng vi khuẩn Anaeromyxobacter sp. IN2
Nguyễn Thị Tuyền, Đinh Thuý Hằng
FJ93913I, GenBank
5 Trình tự gen 16S rDNA của chủng vi khuẩn Paracoccus sp. ĨN12
Nguyên Thị Tuyên, Đinh Thuý Hằng
GU084390, GenBank

VIỆN KHOA HỌC VÀ CÔNG NGHỆ VIỆT NAM• • » «
VIETNAM ACADEMY OF SCIENCE AND TECHNOLOGY
ISSN 1811-4989
TẠP CHÍ
CÔNG NGHỆ S inh h ọ cJOURNAL OF BIOTECHNOLOGY

Tọp chí Cõng nghệ Sinh học 7(3): 371-379,2009
NGHIÊN CỨU ĐA DẠNG VI KHUẨN OXY HÓA Fe(n), KHỬ N 03" TẠI MỘT SÓ MÔI TRƯỜNG SINH THÁI Ở VIỆT NAM
N g n y ễn T h ị T u y ền 1, Đ ỉn h T h ú y H ăn g1
1 Trường Đại học Khoa học Tự nhiên, Hà Nội2 Viện Vi sừth vật vò công nghệ sinh học, Đợi học Quôc gừi Hà Nội
TÓMTẲT
Vi khuẩn oxy hỏa-Fe(II), khủ NOj_ trong các mẫu bùn thu thập ở thủy vực nước Qgọt, chân ruộng ngập nước và vủng nước lọ ven biển được nghiên cứu về số lượng và tính đa đạng về thành phần loài. Kết quả thí nghiệm MPN (Most Probable Number) cho thẩy trong ba dạng môi trưởng Qghiên cứu, mẫu bùn ờ chân ruộng ngập nước có sá lượng tể bào vi khuán này cao hơn cả (9,3 ■ 1Ọ3 TB/g), gấp 5 Ịần mẫu bùn thủy vực nước ngọt vả 2 Lần so với mẫu bùn ven biển. Dựa trên những khác biệt về hình thái khuẩn lạc, 12 chủng đơn được phân Lập từ các mẫu MPN khác nhau. Theo kết quả phân tích ARDRA sử đụng hai enzyme hạn chế H aéĩĩỊ và M spl, các chủng náy được xếp vào 5 nhóm di truyền khác nhau, chúng tỏ tỉnh đa dạng khá cao của vi khuân oxy hóa Fe(n), khử NO}~ tại các môi trường nghiên cửu. Phân tích thành phẩn loài trong quần thể ờ độ pha loãng tới hạn cùa dãy MPN bằng phương pháp PCR-DGGE đối với 16S iĐNA cho thây Anaeromyxobacter sp. có mặt ò cả ba dạng môi trường sinh thái trên. Kết hợp với nghiên cửu các chùng đơn phân lập được từ các dạng môi trưởng nghiên cứu trên cho thấy, nhóm Paracoccus và Pseudomonas là hai nhỏm vi khuẩn chinh thực hiện quá trinh oxy hóa Fe(U), khử NO}’, lương úng đóng vai trò quan trọng bung môi trườnẹ 30 nước Dgọt và chân ruộng ngập nước. Giải trình tự 16S rDNA và 50 sánh kêt quả với Qgân hàng dử liệu đôi với ba chủng đại diện là IN2, IN7 và IN 12 cho phép xếp các chùng này tương ứng vào các chi vi khuân Anaeromyxobacter, Pseudomonas và Paracoccus. Ba chi vi khuẩn kể trên là các nhóm chính chiếm ưu thế trong chu trinh chuyển hỏa sắt tại ba dạng môi tniờng sinh thái được nghiên cửu ờ đây.
Từ kkổa: Anaeromyxobacter, ARDRA, DGGE, khứ NOị~, Paracoccus, Pseudomonas, vi khuẩn oxy hóa Fe(Il), ỉó S rD N A
MỞĐẰU
sát lả một ừong nhũng kim loại phổ biến nhất trên trái đất. Thông thường sắt tồn tại ở dạng oxide Fe(UI) ỉt tan trong oước và có màu vàng nâu. Trong môi trường pH bung tính, dạng hòa tan trong nước (Fe(n)) chi tồn tại ở điêu kiện không có oxy, ví dụ như ở đáy các thủy vực, nơi oxy hòa tan trong nước đã bị cảc vị sinh vật hiếu kỉú sủ dụng để phân hủy các hợp chât hữu cơ. Với hiệu điện the oxy hóa khừ Feỉ+/Feĩ+ tại pH 7 vào khoảng +200 mV, ion Fe(n) có thể trở thảnh nguồn điện từ cho các quá trinh hỏ hấp kỵ khí, điển hình là khử N03“ thành N2 do một số nhóm vi khuẩn đảm nhiệm (Benz et ai, 1998). Quá trình oxy hóa Fe(n) bằng NOr diễn ra như sau:
10 Fe2+ + 2 NOf + 24 H20 — 10 Fe(OH) 3 ị +N2 T + 9H2 T
Trong tự nhiên, quá trinh oxy hóa Fe(II) với chất nhận điện tử là NƠ3~ chủ yếu diễn ra ở ranh giới hiếu khí (có oxy) và kỵ khí (không có oxy) tronẹ lớp tràm tích ờ đáy các thủy vực. Oxy hóa Fe(II) ket hợp với
khử N(V có thể đóng vai trò quan trọng trong môi tnrcmg ô nhiễm với Dồng độ Fe(n) cao (do thiếu oxy) và N(V cao (do chất hữu cơ bị phân hủy tạo thành) (Weber et ai., 2006). Các loài v i khuẩn v ó ị khả năng tiến hành phản ứng oxy hóa khử nảy có thể cùng một lúc thực hiện được hai nhiệm vụ, thứ nhất là chuyển Fe(H) hỏa tan trong nước về dạng Fe(in) kết tủa, và hai là loại bò NO}~, chuyển thành dạng N2.
Vi khuẩn dùng ion Fe(n) làm nguồn cho điện từ để khử NỌr được phân lập đầu tiên từ các lớp trầm tích ao, hồ nước ngọt tại Bremen, Đức năm 1996 (Straub et ai, Ị996). Một số công trinh nghiên cứu tiếp saụ cho thấy sự có mặt khá phổ biển của nhóm vi khuân này với mật độ khá cao ( 1 0 6 tế bào/g trầm tích) trong các điêu kiện môi trường khác nhau bao gôm cả nuớc ngọt, nước lợ và nước mặn và tại nhiều vị trí địa lý khác nhau trên thế giới (Straub Buchholz, 1998). Các loài vị khuẩn phổ biển nhất trong nhóm này được biết đến hiện nay là các loài
, thuộc chi C h ro m o b a c ter iu m và K le b s ie lla (Benz et ai., 1998; Weber et al, 2006). Các nghiên cứu trên thế giới mới chi đưa ra k ết quả khảo sát về nhóm VI
371

Nguyễn Th| Tuyên & Đinh Thuý Hầog
khuẩn nảy ở châu Áu vói điểu kiện sinh thái hoàn toàn khác biệt vớ i nước ta. Trong nghiên cứu này, chủng tôi tiến hành tìm hiểu tính đa dạng cùa vi khuẩn khứ N 0 3_ sử dụng Fe(IĨ) là nguồn điện từ tại một số môi trường sinh thái đại điện ở V iệt Nam .
VẶT LỆU VÀ PHƯƠNG PHÁP NGHIÊN c ứ u
Phtnnig pbỉp tha mẫu
Mầu được thu thập ở ba m ôi trường sinh thái đại diện khác nhau: mẫu bùn đảy ao nưởc Dgọt và chăn ruộng ngập nước được thu ờ ngoại thành Hà N ội, mẫu trâm tích nước lợ được thu ở ven biên Vân Đôn- Quáng Ninh. Mầu được thu thập trong các ống thúy rinh nút xoáy với toàn bộ thê tích binh đê giậm thiêu sự ảnh hưởng cùa oxy không khi tới quân thê vi sinh vật trong mẫu. Mâu được bảo quàn tại nhiệt độ 4 ° c cho đến khi tiến hành thí nghiệm.
X ic định sổ lưựng vỉ khuẩn oxy hóa F e(II), k h ử NO]" b ằng phương pháp M PN
Số lượng vi khuấn oxy hóa Fe(II), khử N C V được xác định theo phương pháp M PN (M ost Propable Number), American Public Health Association, 1969) với thế tích 10% bùn đáy hoặc tram tích trong môi trường khoáng dịch thể chúa NạNO) (5 mM), FeSƠ 4 (10 mM) làm chất nhận và chất cho điện tử. Môi trường dịch thể kỵ khi hoàn toàn có thành phần khoáng tương ứng với nước ngọt (dùng cho mau tù ao til và chân ruộng ngập nước) hoặc nước [ợ (dùng cho mẫu lấy từ Vân Đ ồn - Quảng N inh) được chuấn bị theo phương pháp do Widdel và Bak (1992) mô tá. Dảy pha loàng được thực hiện đên độ pha loăng 10‘\ mẫu được ũ trong tủ ấm lại 2 8 ° c trong 8 tuần.
Phân lập vi khuẩn osy hóa Fe(II), k bử NỌ T băng phương pháp ấng thạch bán lỏng
Óng MPN ở nồng độ pha loảng cao nhất có vi khuân oxy hóa Fe{II), khử NOj' phát triền được sử dụng làm nguồn phân lập cảc khuẩn lạc đơn. V iệc phán lập được tiến hành theo phương pháp pha loãng trên dày õng thạch bán lóng (1%) (W iddel, Bak 1992) với mòi trướng có thành phần tương tụ như mỏi trướng dùng trong phương pháp MPN. Óng thạch bán lóng sau khi bổ sung nguon vi sinh vật (]0°o) từ ông MPN (nguồn phản lặp) được sục khí N; và Ú ứ tư the dáo ngược đầu tại 28°c trong bóng tôi. Khuân lạc đơn phát triển trong các ống pha loãng
được tách bằng pippet Pasteur và chuyên sang raôi trường dịch thể thích hợp (nư ớc n gọt hay nước lợ).
X ác định h àm lư ợ n g F e(II) v ì N Ọ f tron g môi trư ờ n g nuôi cấ y
Mầu vi sinh vật oxy hóa Fe(II), khử NCV đưọc nuôi cẩy trong môi trường ky khỉ dạng dịch thể chúa Fe(II) (10 m M ) và N O j~ (5 m M ) trong binh serum đậy kin băng nút cao su. M ầu dịch nuôi cấy {1 mỉ) được thu sau m ỗi 2 4 h để xác định n ông độ Fe(II) vả N03‘. Nồng độ Fe(II) được xác định bằng phương pháp so màu ở bước són g S10 nm , sử dụng thuốc thừ phenanthrolin) (DIN 38406-E1-1, 1983). Nồng độ N 0 3‘ được xác định bàng phương pháp so màu ở bước són g 410 nm, sử dụng thuôc thử disulfofemic acid (Lê Đ ức, 2004).
T ách D N A tổng số
D N A tổng sổ của quần thể v i sinh vật từ các ổng M PN được tách ch iết theo phương pháp đo Zhou và đồng tác giả (1 9 9 6 ) m ô tà v ó i cải biến về nồng độ đệm phosphate và nồng độ proteinase KL DNA genom e của các cbủng đơn được tách theo phương pháp của Marmur (1961). DNA sau khi tách chiết được hòa tan trong nước và bảo quản tại - 2 0 ° c cho các thí nghiệm tiếp theo.
P hân tích đa d ạn g củ a các ch ủ n g đơn bằng pbirữQg pháp A R D R A
A R D R A (A m plified R ibosom al DNA Restriction A nalyses) là phương pháp thường được sù dụng để nghiên cứu m ức độ đa dạng về di truyền cùa vi sinh vặt dựa trẽn phân tích kết quả xừ lý 16S rDNA bằng enzym e hạn chế. 16S rD N A cùa các chùng đan sau khi đuợc khuếch đại trong phản ứng PCR Sừ dụng cặp m ồi 27F (A G A G TT TGA TCC TGG CTC A G ) và 1492R (G G T T A C CTT GTT ACG AC T T) được xử lý bàng hai en zym e hạn chế Ađspl và H a e ỉĩ ỉ (Ferm entas). Sản phẩm D N A sau khi đã xử lý enzyme được phân tách bàng điện di trên gel agarose 2% tại 100 V trong thời gian 30 phút. Các chùng vi khuân sẽ được xêp nhóm dựa trên sự tiromg đông về phổ các băng đ iện di.
Phân tk h cấu trú c quần th ể b ằ n g phuromg pháp điện di b iến tính (D G G E )
Đoạn 16S rD N A với độ dài 550 bp được khuếch
đại ĩ 0 n g Phàn_ửng PCR sừ d ụ n 8 c*p m°' GM5F(CCT ACÕ GGA GGC AGC AG) và 907R (CCG TCA ATT CCT TTR AG T TT) (M uyzer e t a i ,
372

Tạp chỉ Công nghệ Sinh học 7(3): 371-379,2009
1993). Đ ể tạo tính ổn định ch o v iệc phân tách các trinh tự D N A trên g e l đ iện di biên tính, kẹp GC gôm 4 0 bp được gắn vào đầu 5 ’ của m ồi G M 5F. Đ iện di được tiến hành trên gel polyacrylam ide 6% vớ i dải biến tính urea/form am id từ 3 0 đến 60%. Quá trinh điện di đuợc thực hiện bằng bộ đ iện di D cod e™ System (B ioR ad) ở nhiệt đ ộ 6 0 ° c tại 2 0 0 V trong 3,5 h. Sau khi đ iệu di, g e l polyacrylam ide được nhuộm trong dung dịch etjiidium brom ide (5 m g/m l) trong 30 phút, sau đó rửa nước và chụp ảnh duới tia u v trên m áy G elD oc (B ioR ad). B ăng điện di được căt và thôi D N A ữ on g nước qua đêm tại 4 ° c , sau đỏ dùng làm khuôn để thực hiện PCR với cặp m ồi GM 5F, 907R. Sản phẩm PCR tiếp theo được tinh sạch vả giải trình tự.
Giải trình tự 16S rDNAĐ oạn 16S rD N A của các chùng đơn hoặc sàn
phẩm PCR từ các băng điện đi biến tính được tiến hành phản ứng đọc trình tự vớ i A B l Prism B igD ye Terminator cycle sequencing kit và đọc trình tự trên m áy tự động 3100 A vant A ppied B iosystem s. Trinh tự gen sau đó được phân tích so sánh với trình tự 16S rD N A cùa các loài có liên quan hiện đã công bổ trên Database D D BJ/EM BL/G enBank sừ đụng phần m ềm B L A ST Search. Cây phân loại được dựng theo phương pháp neighbour-joining (Saitou, N ei, 1987), trong đó định dạng cây được tiên hành dựa trên 1000 phép so sảnh đa ch iều (Felsenstein, 1985).
KẾT Q U Ả V À TH ẢO L U Ậ N
X ác đ ịnh sổ lư ợ n g vi khuẩn oxy hóa F e(II), khử N 0 3~ tại các m ôi trư ờ n g n gh iên cứu
Sổ lượng vi khuẩn o x y hóa Fe(II), khử N O f trong các mẫu bùn đáy thu thập tại 3 m ôi trường sinh thái khác nhau được xác đinh thõng qua phưcmg pháp MPN sừ dụng môi trường dịch thê chứa FeS04
làm chất cho điện tử vả N aN O j ỉàra chất nhận điện tử cuối cùng. Thành phẩn khoáng trong m ôi trường tương ứng với điêu kiện nước ngọt (đối với mẫu bùn đáy ao và chân ruộng ngập nước) hoặc nước lợ (đối với mẫu bùn ven biền). Sự phát triển cùa vi sinh vật sinh tmờng nhờ oxy hóa Fe(U) trong các ống MPN được nhận biết thông qua biến đổi màu sắc của môi trường từ trắng xanh (màu cùa Fe(II)) sang màu vàng nâu (màu cùa Fe(lII)) (Hình la).
Kết quà cùa thí nghiệm MPN (Hình lb) cho thấy sô lượng vi khuằn oxy hóa Fe(II), khử NOj" cao nhất trong mẫu bùn chân ruộng ngập nước (9,3 X 1 o 3 tế
bào/g bùn), cao hơn hẳn so vớ i m ẫu trầm tích nước lợ ven biển (4,3 X 103 tể b ào /ẹ trầm tích) và mẫu bùn đáỵ ao nước ngọt (1 ,5 * 103 tê bào/g bùn). M ật độ và m oi tuơng quan giữa số lượng v i khuân o x y hóa Fe(n), khử NO-f với các điều kiện môi trường tại m ỗi vùng sinh thái như trên cũng đã được tìm thấy trong m ột số nghiên cửu trước đầy. Ratering (1999 ) khi nghiên cứu chu trình chuyên hỏa săt trong đật trồng lúa tại Italy đã phát hiện nồng độ các ion sắt ữ on g m ôi trường này rất cao vả các loài tham g ia chu trinh chuyển hóa sắt đóng vai ưò quan trọng trong chu trình chuyển h óa vật chất tại đây. B ên cạnh đó, nhiều nghiên cứu khác cũng ch o thây số lượng vi khuần oxy hóa Fe(U ), khử NO ị~ tại nhiều vùng khác nhau trên thế g iớ i dao đ ộn g trong khoảng từ 1 • 103 đến 5 • 10* tế bào/g mẫu khô (W eber e i a l., 2006; Ratering, Schnell, 2000; Hauck e t a l., 2001 ; Straub et ai, 1996).
P h ân tích cấu trức q u ần th ể b ằ n g đ iện d ỉ b iến tính D G G E
Đ ể xảc định các nhóm v i khuẩn o x y hỏa F e(II), khử N03" chiếm ưu thể tại các môi trường nghiên cứu, chúng tôi tiến hành phân tích cấu trúc quần thể trong cảc ống M PN ờ độ pha loãng 10_J (là n ồn g độ gần tới hạn cùa dãy M P N đối vớ i cà 3 m ẫu) bàng phương pháp P C R -D G G E đoạn 16S rD N A (H ình 2). Các băng đ iện di được cắt từ ge l và sử dụng làm khuôn cho phàn ứng PC R tiêp sau để xác đ ịnh trình tự và so sánh vớ i các trinh tự 16S rD N A đã côn g bố ừ o n g ngân hàng dữ liệu G enBank.
C ó thê thây răng, các loài A n a e ro m y x o b a c te r có mặt trong cả 3 dạng m ôi trường nghiên cứu. Đ ây là nhỏm vi khuẩn nam trong phàn lớp Ỗ -P roteobacteria , hiện mới chi có một loài duy nhất được công bổ là A. deh a lo g ẹn a n s cùng với m ột số đại diện chưa định danh đên loài. Các chủng A n a e ro m y x o b a c te r đã công bố đều sinh trường kỵ khi khừ Fe(III), chưa có chủng nào được nghiên cứu về khả năng sinh trường khử N O f , sử dụng Fe(II) làm chất cho đ iện tư (Treude e t a l., 2003; Straub, B u ch holz-C leven , 1998 Straub e t a l., 1996). Trong m ôi trường nuôi cấy sử dụng ớ đây (cũ nẹ như trong đ iều kiện tự nhiên), Fe(III) đ ồng thời ton tại vớ i Fe(II) d o kết quà chuyển hóa Fe(II) băng con đường hóa học (phản ứng với lượng nhó oxy trong môi tniờng) và con đưcmg sinh học (do các vi sinh vật oxy hóa Fe(II), khừ NO3'). D o vậy, sự cỏ mặt của các loài sinh trường k ỵ khí khừ Fe(III) như Anaeromỵxobacter trong điều kiện m ôi trường nghiên cứu trên (cùng như trong tự
373
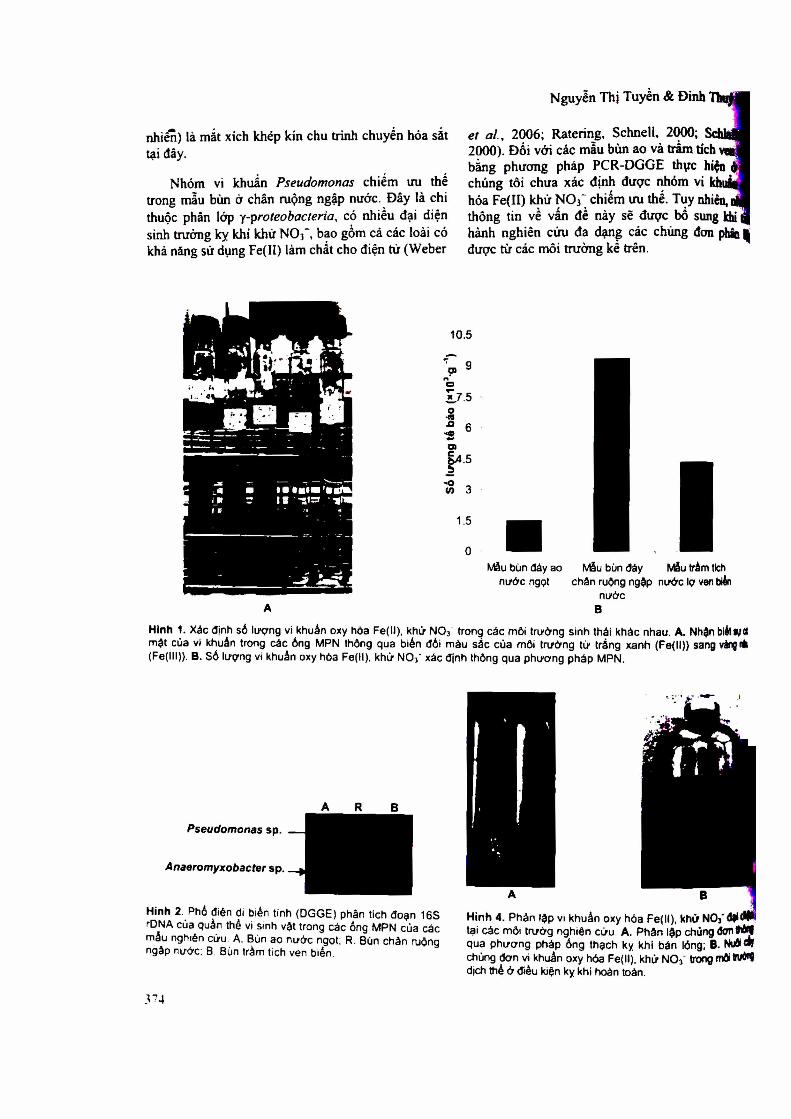
Nguyễn Thị Tuyển & Đinh
nhiên) là mẩt xích khép kín chu trình chuyến hóa sắt tại đây.
Nhóm vi khuẩn Pseudomonas chiếm ưu thế ơong mẫu bùn ớ chân ruộng ngập nước. Đây là chi th u ộ c p h ân lớp Ỵ-pro teobacteria , cỏ n h iề u đại diện sinh trưởng ky khi khứ NƠ3', bạo gồm cả các loài có khá năng sú dụng Fe(II) làm chât cho điện từ (Weber
et ai, 2006; Ratering, Schnell, 2000; Schtad 2000). Đối vởi các mẫu bùn ao và trâm tích vtafl bằng phương pháp PCR-DG G E thực hiện Ỉ l chúng tôi chưa xác định được nhóm vi khuỈH hóa Fe(II) khử N03~ chiếm uru thê. Tụy nhiên, « 1 thông tin về vẩn đề này sẽ được bổ sung khỉ t] hành nghiên cứu đa dạng các chủng đcm phầQ| được từ các môi trường kể trên.
Hlnh 1. Xác định số lượng vi khuần oxy hóa Fe(ll), khử N03' trong các môi trường sinh thái khác nhau. A. Nhận biỉtsựot mặt của vl khuản trong các ống MPN thông qua biến đồi màu sác của môi trường từ trẳng xanh (Fe(ll)) sang vàngiéi (Fe(lll)). B. Số lượng vi khuản oxy hóa Fe(ll), khừ N03' xác định thông qua phương phảp MPN.
A R B
Pseudomonas sp.
Anaeromyxobacter sp.
Hinh 2. Phổ điện di biến tinh (DGGE) phân tích đoạn 16S rDNA của quần thề vi sinh vật trong các ống MPN của các mảu nghién cửu A. Bùn ao nưởc ngọt; R. Bún chần ruộng ngâp nước: B Bún trầm tich ven biẻn.
JHình 4. Phản lặp vi khuần oxy hóa Fe(ll), khử NOj‘ tại các môi trưóg nghiên cứu A. Phân lập chửng (ton qua phương pháp ống thạch kỵ khi bán lỏng; B. Nuởor chủng đơn vi khuần oxy hóa Fe{ll), khử NO3' trong mfli Iwfr’i d|Ch thẻ ở điều kiện kỵ khi hoàn toàn.
374
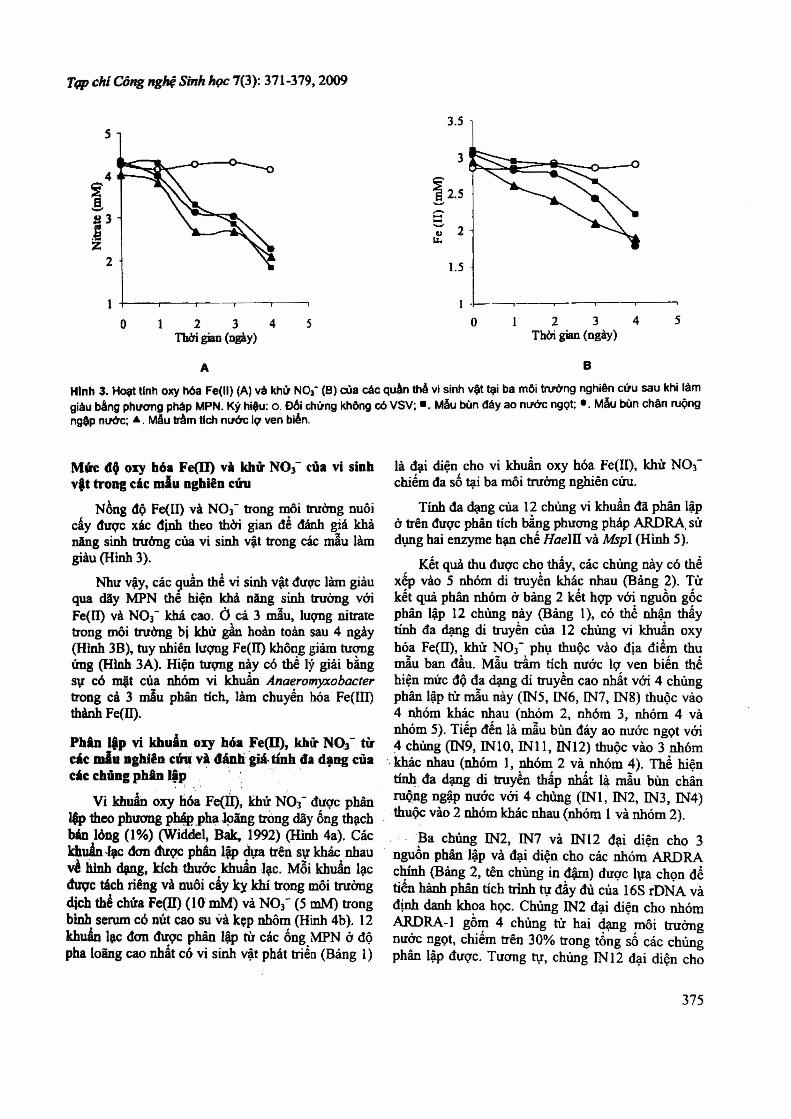
Tạp chí Công nghệ Smh học 7(3): 371-379,2009
Thời gãn (ngày)2 3
Thời gian (ngày)
B
Hlnh 3. Hoạt tinh oxy hốa Fe(ll) (A) vả khử NCV (B) của các quần thề vi sinh vật tại ba môi trường nghiên cứu sau khi làm giàu bẳng phương pháp MPN. Ký hiệu: o. Đối chứng không cỏ VSV; ■. Mẫu bùn đáy ao nước ngọt; Mẫu bùn chân mộng ngập nước; A , Mău trầm tlch nước lợ ven biển.
Mức độ oxy hóa Fe(n) và khử NO) của vi sinh vật trong các mSu nghiên cứu
Nồng độ Fe(II) và NO}' ừong môi trường nuôi cấy được xác định theo thời gian để đánh giá khà năng sinh tmởng của vi sinh vật trong các mẫu làm giàu (Hình 3).
Như vậy, các guần thể vi sinh vật được làm giàu qua dãy MPN the hiện khả năng sinh trường với Fe(H) và NOị' khá cao. Ở cà 3 mẫu, luợng nitrate trong môi truờng bị khử gần hoàD toàn sau 4 ngày (Hình 3B), tuy nhiên lượng Fe(n) không giảm tương ứng (Hinh 3A). Hiện tượng này cỏ thể lý giải bằng sự có mặt của nhóm v i khuẩn A n a ero m yxo b a c ter trong cả 3 mẫu phân tích, làm chuyển hóa Fe(in) thảnh Fe(Et).
P h ân lập v ỉ k h u ầ n o x y h óa F e ( n ) , k h ử N 0 3~ từ cic m ẫu n g h iên cứu và đ á n b g iá tín h đa d ạ n g củ a cic chủng phân lập
Vi khuẩn oxy hỏa Fe(ÌI), khử N03" được phân lệp theo phương pháp pha lọãng trong dãy ống thạch bán lỏng (1%) (Widdel, Bak, 1992) (Hình 4a) Các khuẩn 'lạc đơn được phân lập dựa trên sự khác nhau về hỉnh dạng, kích thước khuẳn lạc. Mỗi khuẩn lạc đutợc tách riêng và nuôi cấy kỵ khí trong môi trường dịch thể chửa Fe(II) (10 mM) và NO3" (5 mM) ừong bỉnh serum có nút cao su và kẹp nhôra (Hinh 4b). 12 khuẩn lạc đom được phân lập từ các ổng MPN ở độ pha loãng cao nhât có vi sinh vặt phát triển (Bàng 1)
là đại diện cho vi khuẩn oxy hóa Fe(II), khử NOj" chiếm đa số tại ba m ôi tnròmg nghiên cứu.
Tính đa dạng của 12 chủng vi khuẩn đã phân lập ở trên được phân tích bẳng phương pháp ARDRA, sử dụng hai enzym e bạn ch ế H aeìữ . và M sp l {H ình 5).
Kết quả thu được cho thấy, các chủng này có thể xếp vào 5 nhóm di truyền khác nhau (Bảng 2). Từ kết quà phân nhóm ờ bàng 2 kết hợp với nguồn gốc phân lập 12 chủng này (Bàng 1), có thể nhận thấy tính đa dạng di truyền của 1 2 chùng vi khuẩn oxy hóa Fe(H), khử NO3’ phụ thuộc vào địa điểm thu mẫu ban đầu. Mầu trầm tích nước lợ ven biến thể hiện mức độ đa dạng đi truyền cao nhất với 4 chủng phần lập từ mẫu này (IN5, IN6 , IN7, EN8 ) thuộc vào 4 nhóm khác nhau (nhóm 2, nhóm 3, nhóm 4 và nhóm 5). Tiếp đến là mẫu bùn đáy ao nước ngọt với 4 chủng (IN9, IN10, IN11, IN12) thuộc vào 3 nhóm khác nhau (nhóm 1, nhóm 2 và nhóm 4). T hể hiện tính đa dạng di truyền thấp nhất lậ mẫu bùn chân ruộng ngập nước với 4 chủng (INI, IN2, IN3, IN4) thuộc vào 2 nhóm khác nhau (nhóra 1 và nhóm 2 ).
Ba chủng IN2, IN7 và IN12 đại diện cho 3 nguôn phân lập và đại diện cho các nhóm ARDRA chính (Bàng 2, tên chủng in đậm) được lựa chọn để tiến hành phân tích trinh tự đầy đủ của 16S rDNA và định danh khọa học. Chủng EN2 đại diện cho nhóm ARDRA-1 gôm 4 chủng từ hai dạng môi truờng nirớc ngọt, chiếm trên 30% trong tổng số các chủng phán lập được. Tương tự, chủng IN 12 đại diện cho

Nguyễn Thị Tuyên & Đinh Tbý
nhỏm A R D R A -4 gồm 3 chủng từ m ôi trường nước lợ và ao nước ngọt, ch iếm 25% trong tổng số các chủng phân lập được. Chủng EN7 làm thánh m ột nbóm riêng biẹt (ARDRA-5^, chỉ tìm thấy ờ m ôi trường nước lợ veo biên. K êt quà so sánh trình tự 16S rD N A của các chủng này v ó i ngân hàng dữ liệu GenBank cho thấy, chùng IN 2 có độ tương đồng cao nhẩt với A n a ero m ỵxo b a c ter d eh a ỉo g en a n s (99% ), chủng IN7 với P seu d o m o n a s s tu tzer i (98% ) và chủng IN 12 với P aracoccus fe ro o x y d a n t (97% ) (Hình 6).
N hư vậy, nhóm A R D R A -4 vớ i chùng EN 12 làm đại diện là các loài P a ra co ccu s và được tim thây trong hai dạng m ôi trường nước lợ ven biển và ao nước ngọt. P aracoccus là chi v i khuẩn nầm trong
phân lớp a-proteobacterria, gồm các loài sinh! kỵ khí tùy tiện , bô hấp b ằn g nitrate boệcl (Schap leigh , 2 0 0 0 ). K ết hợp vớ i kết q u i PC R-D G G E ở trên c ó thể thấy răng, P a ra P seu d o m o n a s là b ỉ i nhỏm c hinh o x y hóa L A [Ậ N 0 3" ch iếm ưu thế trong ba dạng m ôi buòogí nghiên cứu, trong đ ó P a ra co c cu s có vai trò trọng h on ở ao nước ngọt và Pseudomonas ờ IL, Dgập nước. Đ ại d iện của cả hai nhóm nảy đuợcl thấy ở m ôi trường nước lợ ven b iển, có thể ỉà dai hướng của nguồn nước ngọt từ đấỉ liền.
D ựa trên kết quả so sánh trình tự 16S rONA trên, các chủng IN 2, IN7 và EN12 được đ ịnhdM lần lượt là A n a ero m y xo b a c te r sp. EN2, P seudom ^Ế sp. IN7 và P a ra c o c c u s sp. IN 12.
Béng 1. VI khuản oxy hóa Fe(ll), khử NO3' phân lập được từ các môi trường nghiên cứu.
Nguồn phin lfp Chủng vi khuần Đặc điẻni hình thái khuân lạc
Chân ruộng lúa ngập nước (ngoại thánh Há Nội)
IN1IN2
Hlnti tròn, bè mặt nhẵn, kich thước 1 -2 mm
Hình ừủn, bề mặt khdng nhến. Kich thước 0,25 - 0,5 mm
IN3 Hỉnh tròn, bề mặt xù xì, kích thước 0,25 - 0,5 mmIN4 Hinh tròn, bề mặt xù xỉ, kích thước 1 - 211)171
Trầm tlch nước lợ tại biển (Vãn Đòn, Quảng Ninh)
IN5IN6IN7
Hỉnh tròn, bề mặt nhân, kích thưởc 0,25 rrtm
Hlnh bầu đục, bề mặt nhẵn, klch thước 0,25 - 0,5 mm
Hlnh tròn, bè mặt *0 xi, klch thước 0,8 mmIN8 Hình tròn tía. mật độ té bảo ở ngoài ít han phla trong, klch thước 1 ■
1,5 mmĐây ao nirớc ngọt (ngoại thành Há Nội)
IN9IN10
Hlnh tròn, bề mặt xù xl, ktch thước 0,5 -1 mm
Hlnh trỏn không đèu, klch thước 0,5 -1 mmIN11 Hinh tròn, bè mặt nhẵn, kfcti thước 0.2 - 0,3 mmIN12 Hinh đìa lồi hai mặt, kich thước 2 - 2,5 mm
12 Wl n oxy Fe(">- NOì sau khi xử lý bằng cảc * K C fm WHàn Ouòc^ ~ °9 đơn phâ" lập k các mẫu nghién cưu; M; Master 1 tò(Ejẽynfl«
376

Tạp chi C ồ n g nghệ S in h học 7(3): 371-379, 2009
BAng 2. Tinh đa dạng vè <& truyèn của 12 chủng vi khuẩn oxy hỏa Fe(ll), khử NOT đâ phân lập (IN1 - IN12) dựa trén phân tích ARDRA.
NhòmARDRA
Chùng vi khuẩn Các đoạn DNAtạo ra sau khi xử lý 16S rONA bằng các enzyme hạn ché (bp)
H a m MsfA
1 IN1.IN2, IN4. IN11 200.300 300,500
2 IN3, IN8 200,300 500
3 IN5, IN9 200,300 500, 800
4 INS, IN10, IN12 200, 300, 500 300,500
5 BI7 200, 900 500
m .
100
100
M 2
-B acillu s subtU is (FJ544386)
n-ịUl/MI-IN2
Anaeromyxobacter sp FAcl2 (AJ504438)A n a ero m yxo b a c ter d eha ìogenans 2CP1 (N R 027547) Y Ô-Proteobacteria
-A naerom yxobacter clone (A B 293367)
621------- P aracoccus m arim ts (A B 185959)I------Pa°a ra co ccu sfa ro o xyà a n í (A Y 954687)
rIN 1 2
100 \-P aracoccus sp. (A M 9890J7)
k- a-Proteobacteria
P seu d o m o n a sp u tid a (FJ217182)
P seudom onas stu izeri (E U 518705)I N 7
P seudom onas a ỉcahgenes X B 5 (G Q 1504 90) ■Pseudomonas m tro rsd u cen s P S2 (FJ588866)
>, T'-Proteobacteria
Hỉnh 6. Cây phân loại mề hiện mối Nén quan giữa các chủng IN2, IN7, IN12 và các loảí gằn gũi dựa trẽn trình tự 16S rONA. Cây được dựng theo phương pháp neighbor-joining, đom vị = 0,02 K^c trong trinh tự nucleotide. Các số hiền thị ờ các vi tri phân nhánh lả Kết quỏ phân tích bootstrap đối với 1000 phép so sánh (chì có các giá tri trên 50% được binh bày trên hlnh). B. subtilis là loài vi khuẩn được chọn lám outgroup.
K ÉT L U Ậ N
Số lượng vi khuẩn oxy hóa Fe(H), khử N03
toong các mui bùn thu thập ở thùy vực nước ngọt, chân ruộng ngập nước và vùng nước lợ ven biển được xác định Dằm trong khoảng 103 - 104 T B /g, trong đó mẫu bùn ờ chân ruộng ngập nưóc cỏ số lượng tể bào vi khuẩo này cao hơn cả (9 , 3 • 1 0 3
rB/g). Vi khuẩn oxy hóa Fe(ĨI), khử N03~ tại các nôi trường nghiên cứu có tính đa dạng khá cao. tíười hai chùng đơn phân tập từ các mẫu MPN được tếp vào 5 nhóm ARDRA khác nhau ữong phân tích ử dụng hai enzyme giới hạn HaeĩU và Mspl Phân ích thảnh phàn loài bằng phương pháp PCR-DGGE
đ ôi vớ i 16S rD N A cho thấy A n a e ro m y x o b a c te r là m ột trong những nhóm ch iếm ưu thế có m ặt cả ở ba dạng m ôi tnrcmg sinJb thái trên, đóng vai trò khép k ín chu trinh chuyên hóa sắt trong các m ôi trường nghiên cứu thông qua khả năng sinh trưởng k ỵ khí khử Fe(IH). K ết hợp với kết quà nghiên cứu các chủng đom phân lập được từ các dạng m ôi trường nghien cưu trên cho thây P a rữ co c cu s và P se u d o m o n a s là hai nhóm v i khuẩn chính thực h iện quá trình o x y hỏa Fe<n), khử N O f , tương ứ n g đ ón g vai trò quan trọng trong m ôi trường ao nước ngọt va chân ruộng ngập nước. Ba chùng đại d iện la IN 2 IN7 và IN 12 có trình tự 16S rD N A có độ tương đ ồng cao v ó i các loài v i khuẩn A n a e ro m y x o b a c te r
377

Nguyên Thi Iuyén & Đ i n f
d eh a ỉo g en a n s (99% ), Pseudomonas s tu tze r i (97% ) và Paracoccus ferrooxydant (98%), do vậy được đặt tên lần lượt lả Anaeromyxobacter sp. IN2, Pseudomonas sp. IN7 và Paracoccus sp. IN 12. Hai chùng IN2 và EN7 là các chủng vi khuẩa có tiềm năng ứng dụng trong việc xử lý các nguồn ũ ước ngầm nhiễm sắt và nitrate.
L ờ i cảm o n : Nghiên cứ u n à y đ ư ợ c th ự c hiện n h ờ s ự hỗ trợ kinh phi từ đề tài QG.07.26. Tác già xin trán trọng cám ơn Viện Vi sinh vật và Cóng nghệ sinh học, Đại học Quốc gia Hà Nội đã tạo điêu kiện trong quả trinh thực hiện để tài.
TÀI LIỆU THAM KHẢO
American public health association (1969) Standard methods for the examination of water and waste water including bottom sediments and sludge. 604-609.
Benz M, Bnine A, Schiok B (1998) Anaerobic and aerobic oxidation of ferrous iron at neutral pH by chemoheterotrophic nitrate-reducing bacteria. Arch Microbiol 169: 159-165.
DIN 38406-E1-1 (1983) German standard methods for the examination of water, waste water and sludge; cation (group E); determination of iron (El).
Felsenstein J (1985) Confidence limits on phytogenies: an approach using the bootstrap. Evolution 39: 783-791.
Hauck s, Benz M, Brune A, Schink B (2001) Ferrous ừon oxidation by denitrifying bacleria in profundal sediments of a deep lake (Lake Constance). FEMS Microbiol Ecoi 37; 127-134.
Lê Đức (2004) Một Jố phựcmg pháp phán tích môi trường. Nhà xuât bàn Đại học Quõc gia Hà Nội.
Marmur J (i 961) A procedure for the isolation of deoxyribonucleic acid from microorganisms. J Mol BioI 3 208-218.
Muyzer G, De Waal EC, Utterlindea AG (1993) Profiling of complex microbial population by denaturing gradient
ge! electrophoresis analysis of polymerase amplified genes coding for 16S rARN.Microbiol 59: 695-700.
Ratering s (1999) Iron cycle in Italian localization of the redox processes and chi the involved microorganisms. PhD Department of Microbiology, University of (Germany).
Ratering s, Schnell s (2000) Nitrate-depeodeatl oxidation in paddy soil. Environ Microbiol Ỉ: I094J
Saitou N, Nei M (1987) The neighbor-joining new method for reconstructing phylogenetic tm BiolEvo! 4:406-425.
Shapleigh JP (2000) The Denitrifying Dworkin M, Falkow s, Rosenberg E, Stackerbrandt E, eds. The prokaryotes: t electronic resource for the microbiological Springer-Verlag, New York.
Straub KL, Benz M, Scbink B, Widdel F (1996) Ah nitrate-dependent microbial oxidation of fenoui nt Environ Microbiol 62: 1458-1460.
Straub KL, Buchholz-Cleven BEE (1998) Em and detection of anaerobic ferrous iron-oxiđiàDti reducing bacteria from diverse European sedanofc Environ Microbiol 64: 4846-4856.
Treude N, Rosencrantz D, Liesack w, ScbndlSI Strain FAcl2, a dissimilatory iron-reducing meata Anaeromyxobacier subgroup of Myxococcda. I Microbiol Ecol 44: 261 -269.
Weber KA, Umitia MM, Churchill PF, Kukbàp Roden EE (2006) Anaerobic redox cycling flf ãí freshwater sediment microorganisms. Environ lit* 8: 100-113.
Widdel F, Bak F (1992) Gram-negative mesoph&i reducing bacteria. In BaJows A, TrQper HG, Dual Harder w, Schleifer KH eds. The Prokaryota, Í Springer, Berlin Heidelberg New York: 3 3 52-3371
Zhou J, Bruns MA, Tiedje JM (1996) DNA reantf. soils of diverse composition. Appl Environ hôơdi 316-322.
378

Tạp chí Công nghệ Sinh học 7(3): 371-379, 2009
STUDY ON DIVERSITY OF ANAEROBIC Fe(II)-OXIDIZING, N O 3 -REDUCING BACTERIA IN SOME ECOLOGICAL ENVIRONMENTS IN VIETNAM
Nguyen Thỉ Toyen1, Dính Thuy Hang2, *'H a n o i U n iversity o f Science, V N U 2Institute o f Microbiology and Biotechnology, VNJJ
SU M M A R Y
Fe(H)-oxydizing, NOj'-reducmg bacteria in sediment samples collected from freshwater aquifer, flooded paddy soil and estuarine area were quantified and analysed on species composition. The MPN results showed that among the three studied environments, flooded paddy soil had the highest number o f these bacteria (9.3 • 103g_l), five and two times higher than that of freshwater aquifer and estuarine sediment, respectively. Based on distinguish colony characteristics, 12 strains of Fe{n)-oxidizing, NOj-- reduing bacteria were isolated from MPN series and were set into five genetically different groups as proposed by ARDRA analyses with two restricition enzymes HaeUl and M spl, showing relatively high diversity of this bacterial group in the studied environments. Analysing species composition in the communities developed in MPN tubes at the highest dilution via PCR-DGGE of 16S rDNA revealed that Anaeromyxobacter species presented in all three above environments. In combination with the culture- dependent studies, it was revealed that Paracoccus and Pseudomonas were dominant groups playing important roles in freshwater aquifer and flooded paddy soil, respectively. Sequencing of 16S rDNA from three representative strains IN2, IN7 and EN 12 suggested that they are respectively belonged to Anaeromyxobacter, Pseudomonas and Paracoccus genera.
Keywords: Fe(II)-oxidizing, NO3~-reducing bacteria, DGGE, ARDRA, Anaeromyxobacter, Paracoccus, Pseudomonas, ỈÓSrDNA
'A u th o r fo r co rresp o n d en ce: Tel: 8 4 -4 -3 7 5 4 7 6 9 4 ; E-mail: d th a n e ia ivn u p.rhi vn
379

chí Khoa học ĐHQGHN, Khoa học Tự nhiên và Công nghệ X X (2009) 0-0
Nghiên cứu vi khuẩn ưu thế tham gia chu trình Fe trong các điều kiện môi trường khác nhau tại Việt Nam
Nguyễn Thị Tuyền1, Nguyễn Minh Giảng2, Đinh Thúy Hằng2-*
'Khoa Sinh học. Đại học Khoa học Tự nhiên, Đại học Quốc gia Hù Nội.334 Nguyễn Trãi, Hà Nội2 Viện Vi s in h vậ t và Công n g h ệ Sinh học, Đ ạ i họ c Q u ố c g ia H à N ộ i, ì 4 4 Xuân Thúy, Hà N ộ i
Nhận ngày tháng năm
Tóm tắt. Hai chúng vi khuấn IN2 và IN 12 được phân lập từ bùn đáy nước ngọt, đại diện cho hainhóm vi khuẩn chính tham gia chu trinh chuyển hoá sảt tại đây, bao gôm khứ Fe(IIl) băng các họp chất hữu cơ và oxy hóa Fe(II) bàng nitrate. Hai chủng vi khuấn nói trẽn có các đặc điêm sinh lý hoàn toàn khác biệt nhau. Chùng IN2 được phân lập từ mẫu bùn chân ruộng ngập nước là trực khuẩn, sinh trướng ky khí bảt buộc bằng Fe(III) hoặc nitrate, thích hợp với môi trường có nông độmuối cao hom 1%. Chừng IN I2 được phàn lập từ mầu bùn đáy ao nước ngọt là vi khuắn kỵ khí tùytiện, có tế bào hình oval và thích hợp với môi trướng có nồng độ muối từ 0 - 3%. Chúng IN 12 thề hiện hoại tính khứ nitrate, oxy hóa Fe(II) cao, do đó có tiềm năng ứng dụng thực tế trong việc xừ lý các nguồn nước nhiềm nitơ và ion kim loại Fe(li). Giải trinh tự 16S rADN và so sánh kết quà với ngân hàng dữ liệu đối với hai chúng IN2 và IN 12 cho phép định đanh các chúng này tương ừng tà A naerom yxobacter sp. 1N2 (mã trình tự FJ939131) và P aracoccus sp. IN 12 (mã trinh tự GU084390).
Từ khóa: A naerom yxobacler , Paracoccus, Vi khuân khứ F e(lll). Vi khuẩn khứ nitrate, oxv hóa Fe(II), 16S rADN.
M ỡ đ ầ u
Trong tự nhiên sắt lá một trong những uyên tổ có mặt với hàm lượng đáng kể. sau ‘bon, nitơ, phospho và lưu huỳnh []]. Là >t nguyên tố kim loại, sắt tồn tại ở các dạng I với điện thế oxy hoá khử khác nhau, gồm [II) và Fe(Ill). Trong hai dạng ké trên chi có ;ỉl) tồn tại ỏ' dạng hoà tan trong nước, tuv lên. do có tính ,khừ cao. Fe{ll) nhanh chóng
ic giá liên hệ. ĐÌ': t84 4 37547694 •mail: dlliiiimaAnu.cdu.ui
bị oxy hoá thành Fe(IIl) trong phản ứng hoá học với oxy không khí. Do vậy ion Fe(II) thường chi được tìm thấy trong các điều kiện môi trường không có oxy, ví dụ như ở đáy các thuỳ vực hay các tầng nước ngầm hay môi trường có pH thấp [2,3]-
Các vi sinh vật tham gia chu trình sắt gồm c ó ( i ) vi khuân OXV lio á Fe(11) bằng oxy như
Thiobaúllus ferrooxydam [I], bằng nitrate như một số loài Chromobacterium, Klebsiella [4 ] hay bang quang hợp như Chìorobium, Rhodobucter [5] và (ii) vi khuẩn khử Fe(IỊ|)

N.T. Tuyên và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tụ Nhiên và Công nghệ XX (2009) 0-0
í các loài Geobacter, Sewanella, aeromyxobacter [6 , 7, 8 ]. Trong khi oxy hoá II) bằng oxy theo con đường sinh học chi n ra ở môi trường có pH thấp thì oxy hoá (II) thành Fe(lU) bàng nitrate và khừ Fe(III) tnh Fe(U) bàng một số hợp chất hữu co là i quá trình diễn ra ở điều kiện pH trung tính, ép kín chu trình chuyển hoá sẳt tại đây [ 1]. liều công trình nghiên cứu cho thấy sự có ít khá phả biến của hai nhóm vi khuẩn này ở iều điều kiện môi trường khác nhau, bao m cà nước ngọt, nước lợ và nước mặn và tại iều vị trí địa lý khác nhau trên thế giới [7,
Trong bài báo này chủng tòi trình bày .ưng kết quả nghiên cứu thực hiện đối với hai ủng vi khuẩn đại diện cho nhóm vi khuẩn ,y hoá Fe(ll) bang nitrate và khứ Fe(lll), rợc phân lập từ môi triròng thiếu oxy trong in đáy nước ngọt tại Việt nam.
Nguyên liệu và phương pháp
/ . N g u ồ n vi s in h vậ t
Các chúng vi khuẩn đưcrc phân lập từ thi >hiệm nuôi tich lũy trẽn mẫu bùn tự nhiên Dng môi trượng kỵ khí sir dụng Fe(ll) và Oi" làm chất cho và nhận điện tứ tương ửng ]. Mau bùn tự nhiên được thu thập ò' đáy ao rớc ngọt và chân ruộng ngập nước ờ ngoại ành Hà Nội.
I
?. N g h iê n c ứ u c á c đ ặ c đ iè tỉì s in h lý
Đặc điếm sinh trướng cùa các chủng vi uẩn được tien liành nghiên cứu trên môi rờiig kỵ khi dịcli thể. sử dụng các chất cho và ận điện tử khác nhau (bao gồm cả oxy) và nồng độ muối khác nhau. Sự sinh trương
ỉ vi khuẩn tại các điều kiện nuôi cấy khác
nhau được đánh giá thông qua thay đổi về độ đục của môi trường so với đôi chứng không co vi khuẩn.
2.3. Nghiên cứu hình thái
Hinh thái cùa các chủng vi khuẩn đurợc xác định đối với tế bào ờ pha sinh trường lũy tiến. Dịch tế bào được ly tâm 10.000 vòng/phút trong 5 phút, sau đó sinh khối được hòa trong 200 Ịi! nước muối 0,9% và dùng làm tiêu bản trên lam kính phù agar. Ảnh tế bào được chụp duới kinh hiển vi đối pha, độ phòng đại 1 0 0 0
lẩn.
2.4 . T á c h A D N tổ n g số , n h â n g e n ỉ 6 S r A D N và x á c đ ịn h v ị t r í p h â n lo ạ i
ADN tổng số từ các chùng vi khuẩn được tách chiết theo phương pháp cùa Marmur (196 ]). [1 0 ] với một số thay đổi để tối ưu hóa. Gen mà hóa cho I6 S rARN (1500 bp) được khuếch đại trong phàn ứng PCR sử dụng cập mồi 27F (AGA GTT TGA TCC TGG CTC AG) và I492R (GGT TAC CTT GTT ACG ACT T). Sản phẩm PCR được tinh sạch và giải trinh tự với ABI Prism BigDye Terminator cycle sequencing kit và đọc trình tự trên máy tự động 3110 Avant Appied Biosystems. Trình tự gen sau đó được phân tích so sánh với trình tự 16S rADN cùa các loài có liên quan hiện đã công bố trên Database DDBJ/EMBL/GenBank sử dụng phần mềm BLAST Search. Cây phân loại được dựng theo phương pháp neighbour- joining [ 1 1 ], trong đó định dạng cây được tiến hành dựa trên 1 0 0 0 phép so sánh đa chiều [ 1 2 ].
2.5 . Đ á n h g iã h o ạ t t in h s in h h ọ c c u a c á c c h u n g 17 khuân
Cluing vi khuẩn khứ nitrate được nuôi cấy trên mòi trường k>; khí dạng dịch thể chứa

N.T. Tuyềh và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tự Nhiên và Công nghệ tập (2009) sõ trang 3
trate (5 mM) íàm chất nhận điện tử và Fe(II) tặc Mn (II) (10 mM mồi loại) ỉàm chất cho ện tử trong bình serum đậy kín bằng nút cao . Mầu dịch nuôi cấy (5 ml) được thu thập sau ỗi 24 h và xác định nồng độ các nguồn cho t nhận điện từ trong môi trường. Nồng độ trate được xác định bằng phương pháp so àu sử dụng thuốc thử axit desulfofemic [13]. ồng độ Fe(ll) được xác định bàng phương láp so màu ỡ bước sóng 510 nm, sữ dụng uốc thử phenanthrolin [14], Nồng độ Mn (II) rợc xác định theo phương pháp 'imaidoxime [15].
Vi khuẩn kliử Fe(Iỉl) được nuôi cấy trong ôi trường kỵ khí dịch thể, chứa Fe(IIi) ở Ịng phức hợp với citrat (tan trong nước) làm lất nhận điện từ (15 mM) và Na-acetate (10 M) làm chất cho điện từ. Sinh trường cùa vi luẩn được nhận biết thông qua sự mất màu ia mòi trường (màu xanh lá cây đậm do e(lll)-citrate tạo ra).
. Kết quả và thào luận
/ . P h â n lậ p v i k h u â n
Hai chủng 1N2 và INI2 đưọc phân lập từ í nghiệm làrtí giàu bùn đáy thủy vực nước ;ọt và chân ruộng ngập nước qua dãy MPN r dụng môi trường kỵ khi dịch thế chứa Fe(l[) 1 NO,- làm chít cho và nhận điện tử tirơng Ig. Nglìiêtr cíhỊ đa dạng vê đặc điểm di lyền gen I6 S pADN bằng phtrong pháp 3GE và ARDRA đôi với nhóm vi khuân oxy á Fe(ll) khữ NOi" tại hai môi trường sinh íi kề trên ở Việt nam cho thấy chùng IN2 đại ịn cho nhóm vi khuân luôn có mặt ờ cà 2
■>i trưòng đã nghiên cứu. trong khi đó chùng 1 2 chi tim thây ớ bùn đáy thuy vực nước ?t [16]. Dựa trên tinh đại diện đổi với hai 5m vi khuấn chính tham liia chu trình lyên hoá sắt tại các môi trưòng nghiên cửu,
hai chủng này được lựa chọn đê tìm hiêu sâu về các đặc tính sinh học cũng như khả năng ứng đụng thực tế.
3.J. Nghiên cửu các đặc điếm sinh lý của hai chúng vi khuân 1N2 và INI2
Báng ]. Đặc điểm sinh lý cúa hai chúng vi khuẩn IN2 va IN 12
Đặc điểm sinh lý 1N2 IN12
Hô hấp bằng oxy (sinh trường hiếu khí)
- + +
0% - + + +
Sinh trưởng 1% + + + + + +phụ thuộc độ mặn của môi
2% + + + + + +
trường 3% + + + + + +
(% NaCl) 4% + + + + +
5% + + + +
Lactate + + + + + +Chất cho điên từ
Acetate + + + + + +
(khử nitrate) Ethanol + + + + "4" +
Benzoate + + + +
Chấi nhận
2 o v»1
+ + + + 4* +
điện tứ (oxy hoá lactate)
Fe( III) - + + +
so42~ — -
Kết quả nghiên cứu cho thấy hai chùng IN2, IN 12 rất khác nhau về đặc điểm sinh ly (bảng I). Trong khi chùng IN2 là vi khuẩn kỵ khi bắt buộc thi chúng IN 12 là vi khuẩn ky khi tùy tiện, có khả năng hô hấp bàng oxy (sinh trường hiếu khi). Ngoài N (V được sừ dụng làm chât nhận điện tử cuối cùng, chúng IN2
còn có khá năng sử dụng Fe(III) làm chất nhận

N.T. Tiạ/ỈH và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tự Nhiên và Công nghệ XX (2009) 0-0
ện từ còn chủng INI2 không có khà năng iy. Cả hai chùng đều không sinh trường bằnglử so/".
Đối với các nguồn điện tử khác nhau phụccho quá trình khử nitrate, ngoài Fe(ll) đã
irợc sừ dụng để phân lập và nuôi cấy từ ban ầu, lactate, acetate, benzoate và ethanol đều trợc oxy hoá bời hai chùng IN2 và [NI2. rong trường hợp benzoate, một axit hữu cơ nứa vòng thơm và thường khó bị oxy hoá hơn íc hợp chất còn lại, chủng IN 12 thể hiện khà ẫng oxy hoá kém hom hẳn so với chủng 1N2 .
Độ mặn là một trong các yếu tố môi trường uart trọng ảnh hưởng đến sinh lý của vi sinh ật trong tự nhiên, do vậy ờ đây chủng tỏi tiến ành nghiên cứu ảnh hưởng cùa nồng độ muối ong môi trường đối với khả năng sinh trưòng ủa hai chùng vi khuẩn quan tâm. Kết quả ghiẽn cứu cho thấy chimg IN2 có nhu cẩu về tuổi để sinh trưởng, chùng INI2 thì không, 'rong khi chùng IN 12 có thế sinh trường trong lôi trường không cần bồ sung NaCI thì chùng N2 chi phát triển khi NaCI có mặt ờ nồng độ % trờ lên. Tuy nhiên, chủng IN2 lại cỏ khả lăng chịu muối cao hon IN 12, chùng này vẫn 'hát triển tốt ở nồng độ muối 5% trong khi đỏ 3C độ sinh trưởng của IN 12 bị ảnh hường áng kề ở các nong độ muối cao hom 3%. Điều ày cùng phù hợp với đặc điểm phân bố của ai chúng vi kịiuần trong tự nhiên, cụ thể ]à húng 1N2 đạhdi,ện cho nhóm vi khuẩn được m thấy ờ í>h(ềủ /Ịạng môi trường khác nhau, ỈO gôm cả nước ngọt và nƯỚC lợ. trong khi đó
lùng IN 12 đại diện cho nhóm chi tim thấy ỏ1
in đáy thuý vực nưóc ngọt [16].
2. Nghiên cứu hình thái tế bào và g iai trình ‘ gen Ỉ6S rADN cua hai chưng vi khuân ỈN2 r ỈN ỉ 2
Chúng IN2 gồm nhũng tế bào dạng trực uẩn, kích thước 1x4-5 |im. chuyến động tích
cực (hình 1); chủng rN 12 gồm những tế bào hinh oval có kích thước 1 ,5 x 1 .5-2 fim, chuyên động chậm (hình 1 ).
Giải trình tự gần đủ gen mã hóa cho I6 S rARN (1300 bp) của hai chủng ĨN2 và IN 12 và so sánh với ngân hàng dừ liệu GenBank cho phép chúng tôi xếp chủng ỈN2 vảo chi Anaeromỵxobacler (ioài gân gũi nhât là A. dehalogenans, 99% tương đồng) và chùng INI2 vào chi Paracoccus (loài gần gũi nhất là p. aminovorans, 98% tương đồng).
Anaeromyxobacter là một chi thuộc phản lớp Ồ-Proteobacteria, được biết đếnvới các đại diện sinh trưởng kỵ khí bắt buộc bằng oxy hoá các hợp chất hữu cơ để khừ Fe(IlI) [8 ]. Trong ngân hảng dữ liệu về gen 16S rADN cùa chi này hiện mới có loài A. dehaỉogenans và một số các chùng chưa được định danh đến tên chi. Chùng IN2 đưọc bổ sung vào danh sách chi Anaeromyxubacter với tên khoa học là Anaeromyxobacter sp. IN2 và mã trình tụ gen ]6 S rADN trong GenBank là FJ939I31 (hình 1C). Trong nghiên cứu này, lần đẩu tiên vi khuân Anaeromyxobơcter mà đại diện [à chùng IN2 thê hiện khả năng sinh trưởng bang oxy hoá Fe([l), khử nitrate, tuy nhiên hình thức sinh trường này không vượt trội so với sinh trưởng kỵ khí khử Fe(!il).
Paracoccus là chi vi khuẩn nằm trong phân lớp a-Proteobacteria, gồm nhiều ioài có khả năng hô hấp bằng oxy (sinh trường hiếu khí), đông thời hô hấp bàng khử nitrate [17]. Nhiều đại diện của chi Paracccus được tim thấy trong các điều kiện tối ưu cho khừ nitrate, như đáy các thuý vực [ 17] hay trong các hệ thống xử lý nước thải loại bó ni tơ, ví dụ như p. denitrificam, p. ferooxydcms [18]. Chúng INI2 đưọc đinh danh khoa học là P u r a c o c c u s sp IN 12 và có mã trinh tự gen 16S rADN trong GenBank là GU084390 (hình 1D).

N.T. Tựyá và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tự Nhiên và Công nghệ tập (2009) sô'trang
0 02
mế ễ v hP * ■
f %• .
; V • ;Ị * / é ,
ệ ^ *• * Paracoccus sp. IN12
?3Ũ
] 000
1000
-Bãciỉíis subtiìis (FJ544 3SÕ!
- AnơenimvĩữbiKler clone (AB293367)
i Anjenmyrnhjcter dehnii-gi-.'U'S ! EF067314)
1N2
0.02
-Anaeromyxobacter sp FAcl2 (AJ504438)
A dehabgenans 2CP1 (HR027547)
—AnaeromyT&bacter clone (L‘Q394ỹ9ó)
-Bacúkỉs subtiíis (FJ5443S6)
588
—Paracoccus aminophiíus (D32239)r-Parucoccus aminovonans (D3224Ũ)
■Paracoccus sp (AM939Ũ37)1Ũ0õCị]V)12-Panacữccus marinus (AB185959)IParacoccus dem tnficans (/J288159)
lũũõlParacoccus fcrooxydant (AY954687)
Hinh I. Hình thái tế bào và vị tri phân loại cùa hai chủng IN2 và IN 12. Hình thái tể bào được quan sát trẽn kính hiến vi quang học, độ phóng đại 1000 lẩn (đơn vị = 5 Ị.im). Cây phân loại được dựng theo phương pháp
neighbor-joining, đơn vị = 0,02 Kmii: trong trinh tự nucleotide. Các số hiên thị ớ các vị trí phân nhánh là kết quả phân tích bootstrap đôi với 1000 phép so sánh (chi có các giá trị trẽn 500 được trình bày trên hình). B. subtilis là
loài vi khuẩn được chọn làm outgroup.
3.3. N g h iên ciiru h o ạ t tin h s in h h ọ c v à k h á n ă n g ứ n g d ụ n g cu a c h u n g vi k h u â n I N 12
Trong nghiên cứu về đa dạng vi khuẩn khứ nitrate, oxy hoá Fe(ll) ờ Việt Nam, P a ra c o c c u s sp. vói đại diện là chủng IN 12 chiếm ưu thé trong điều kiện nước ngọt [16]. Chủng vi khuân này thẻ hiện khá năng sinh trưởng trong nhiều đều kiện môi trường khác nhau như có mặt hay không có mặt oxy, thích nghi với nhiéu loại nguồn điện tứ khác nhau (như Fe(II), các axit hữu co, rưọu) và sinh trường tốt trong môi trường có độ mặn dao động trong dải 0-3% (đặc trưng cho cả mỏi trường nước ngọt và nước lợ). Đây cũng là lý
do để chúng tôi tiến hành tìm hiểu khá năng ứng dụng của chùng vi khuấn này trong việc loại bò nitơ và Fe(II) trong các nguồn nước thài õ nhiễm,
Hoạt tính ôxy hóa Fe(ll) và khử nitrate của chủng IN 12 được xác định ở điều kiện kỵ khí với Fe(II) là chât cho điện từ và nitrate làm chất nhận điện từ cuối cùng. Kết quà xác định nòng độ các chát cho và nhận điện tử trong môi trượng biến đồi theo thời gian cho thấy sự giảm nônẹ độ Fe(][) và nitrate diễn ra song song với tôc độ đáng kể (hình 2).

6 NT. Tuyên và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tự Nhiên và Cõng nghệ XX (2009) 0-0
0 1 2 3 4 5
Thời gian (ngây)
Hlnh 2. Loại bò Fe(ll) (•) vả NO," (■) trong môi trường do tác động cùa chủng vi khuẩn IN ] 2.
Bên cạnh đó khà năng sử dụng Mn(II) làm chất cho diện từ để khử nitrate cũng được kiểm tra, tuy nhiên chùng IN 12 không thề hiện sinh trường rỏ rệt trong điều kiện nuôi cấy này. Theo một số nghiên cứu đã công bố, hoạt tính oxy hoá M 11(11). khử nitrate bàng con đường sinh học được xác định trong một số quẩn thể vi sinh vật ỏ đáy các thuỳ vực, nhưng cho đến nay chưa có chủng vi sinh vật nào đưọc phán lập trong phòng thí nghiêm với hoạt tính kể tren [! 9, 20].
4. Kết luận
Hai chủng vi khuẩn 1N2 và IN 12 đại diện cho hai nhóm tham gia chu trình chuyển hoá sắt tại một số môi trường sinh thái khác nhau ở Việt nam, trong đó chủng IN2 tham gia khử Fe(ill) thành Fe(ll) bẩng các họp chất hữu cơ và chùng 1N12 tham gia oxy hoá Fe(ll) thành Fe(lll) bằng nitrate. Vai trò sinh thái cua hai chúng này được khãng định thông qua các đặc điềm sinh lý của chúng, cụ thể lả khá nãng sinh trưởng với các chất cho và nhặn điện tử khác
nhau và mức độ sinh trường tại các nồng độ muối khác nhau trong môi trường.
So sánh trinh tự gần đù của gen 16S rADN với các loài đã công bố trên GenBank cho
B phép định danh hai chùng vi khuân này là— Anaeromyxobacter sp. IN2 (mã trình tự đăng£ ký trên GenBank là FJ939 ] 31) và Paracoccus
sp. IN 12 (mã trình tự đăng ký trên GenBank là GU084390).
Với hoạt tính cao trong việc oxy hoá Fe(II) và khử nitrate, chùng IN 12 có tiềm năng ứng dụng thực tế trong việc xử lý các nguồn nước nhiễm nitơ và ion kim loại Fe(II).
Lòi cảm ơn
Nghiên cứu này được thực hiện nhờ sự hỗ trợ kinh phí từ đề tài ỌG.07.23. Tác già xin chân thành cảm ơn Viện Vi sinh vật và Công nghệ sinh học, ĐHQGHN đã tạo điều kiện trong quá trình thực hiện đề tải.
Tài liệu tham khảo
[1] H.L. Ehrlich, Geomicrobiology. 3'J Ed. revised and expanded, Marcel Dekker Inc., New York, 1996.
[2] K.H. Nealson and D. Saffarini, Iron and manganese in anaerobic respiration: environmental significance, physiology and regulation, Annu. Rev Microbiol. 48 (1994) 311.
[3] K..O. Konhauser, Bacterial iron biomineralisation in nature. FEMS Microbiol. Rev 20(1997)315.
[4Ị K.L. Straub, M. Benz. B. Schink, F. Widdel Anaerobic. nitrate-dependent microbial oxidation of ferrous iron. Appl Environ. Microbiol. 62(1996) 145
[5] F. Widdel, s. Schnell, s.. Heising A Ehrenreich, A. Assmus, B. Schink. Ferrous iron

N.T. Tuyên và nnk. / Tạp chi Khoa học ĐHQGHN, Khoa học Tự Nhiên và Công nghệ tập (2009) só trang 1
oxydationby anoxygenic phototrophic bacteria, Nature 362 (1993) 834.
[6] D.R. Lovley, Dissimilatory Fe(Ill) and Mn(IV) reduction, Microbiol. Rev. 55 (1991) 259.
[7] D.R. Lovley, Microbial Fe(lll) reduction in subsurface environments, FEMS Microbiol Rev. 20(1997)305.
[8] N. Treude. D. Rosencrantz, w. Liesack. s. Schnell. Strain FAc12, a dissimilatory iron- reducing member of the Anaeromyxobac/er subgroup of Myxococcales, FEMS Microbiol. Ecol. 44 (2003)261.
[9] K.L. Straub, B.E.E Buchholz-Cleven, Enumeration and detection of anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria from diverse European sediments. Appl. Environ Microbiol 64 ( 1998) 4846.
[10] J. Marmur J, A procedure for the isolation of deoxyribonucleic acid from microorganisms. J Mol Biol 3 (1961)208.
[11] N Saitou. M. Nei. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Mol Biol EraI 4 (1987) 406.
[12] J. Felsenstein. Confidence limits on phyloeenies: an approach using the bootstrap. Evolution 39 (1985) 783.
(1.1] Lê Đức, MỘI so phương pháp phân lich mõi trường, NXB ĐHỌGHN, 2004.
[14] DIN 38406-E1 - ], German standard methods for the examination of water, waste water and sludge; cation (group E); determination of iron (El )~ 1983.
[15] P.G. Brewer, D.w. Spencer, American Sociaty of Limnology and Oceanography, Colorimetric determination of manganese in anoxic water, 16 (1971) 107.
[16] Nguyễn Thị Tuyển. Đinh Thúy Hằng, Đa dạng vi khuân oxy hoá Fe(ll), khứ NOì tại một số môi trường sinh thái ớ Việt Nam, Tạp chí Cóng nghệ sinh học, 2009 (sẩp đăng).
[17] J.p. Shapleigh, The Denitrifying Prokaryotes. In M. Dworkin, s. Falkow, E. Rosenberg, K.H. Schleifer, E. Stackerbrandt, eds. The prokaryotes', an evolving electronic resource for the microbiological community, Springer- Verlag, New York, 2000.
[18] G. Bitton, Wastewater Microbiology, Wiley- Liss, Inc. Toronto, Canada. 1999.
ỊI9] A.M. Gounot, Microbial oxidation and reduction of manganese: Consequences in groundwater and applications, l-'UMS Microbiology Review 14 (1994) 339.
[20] J. Vandenebeele, D. De Beer, R. Germonpre. R. Van De Sande, w. Verstraete. Influence of nitrate on manganese removing microbial consortia from sand filters. Hal Rex. lo t 29(2) (1995) 579.
Study, bacteria o f the iron cycle that dominate in different t 6: environments in Vietnam
Nguyen Thi Tuyen1, Nguyen Minh Giang2, Dinh Thuy Hang2
1 Department o f Biology,College ofSciences, Vietnam National University Hanoi, Ì34 Nguven Trai. Hanoi ■ Institute o f Microbiology andBiatechnnlogv, Vietnam National University Hanoi. 144 Xuan thuv. Hanoi
A b str a c t. T w o bacterial strains IN2 and IN 12 w ere isolated from fresh water sediments and
represented for two major groups of the iron cycle in these environments, including the reduction of Fe(lll) with organic compounds and oxydation of Fe(II) with nitrate. These two strains possessed different p h y sio lo g ica l ch aracteristics. Strain IN2 w as isolated from flood ed rice paddv so il, con sisted

8 NT. Tuyên và nnk. / Tạp chí Khoa học ĐHQGHN, Khoa học Tự Nhiên và Công nghệ XX (2009) 0-0
of rod-shaped cells, grown strictly anaerobicaly with Fe(IH) or nitrate and best growth was observed at the concentration of NaCI in the medium above 1%. Strain IN 12 was isolated from sediment of a fresh water aquifer, contained oval cells that were facultatively anaerobic, grown best at the concentration of NaCI in the medium in the range of 0 - 3%. Strain [N12 showed a high activity of Fe(ll) oxidation with nitrate, therefore would have application potential in treatment of waters contaminated with nitrogen and metal ion Fe(II). Sequencing of 16S rDNA and comparative alignment with database allowed to designate these strains with the names Anaeromyxobucler sp. ]N2 (sequence number FJ939I31) and Paracoccus sp. IN 12 (sequence number GU084390).
Key words: Anaeromyxobacter, Paracoccus, Fe(lll) reducing bacteria, nitrate reducing bacteria, Fe(ll) oxydation, I6 S rDNA.

VIỆN KHOA HỌC VÀ CÔNG NGHỆ VIỆT NAM VIỆN SINH THÁI VÀ TÀI NGUYÊN SINH VẬT
Ban tổ chức Hội nghị Khoa học Toàn quốc về Sình thái và Tài nguyên sinh vật lần thứ 3
........................... 0 O 0 ................. ....
Ban tồ chức Hội nghị Khoa học toàn quốc về Sinh thái và Tài nguyên sinh vật ỉần thứ 3 đã nhận được bài poster:
'Đa dạng vi khuẩn oxy hóa Fe(II), khử nitrate tại một số môi trường sinh thái ở Việt Nam”
Của các tác giả:Nguyễn Thị Tuyền
Đại học Khoa học Tự nhiên, Đại học Quôc gia Hà NộiĐỉnh Thúy Hằng
Viện Vi sinh vật và Công nghệ Sinh học, Đại học Quôc gia Hà Nội
Bài poster của các tác giả đã được biên tập và dự kiến sẽ trưng bầy tại Hội nghị.
Xin chân thành cảm ơn sự cộng tác của các tác giả
G IẤY N H Ậ N BÀI

Đa dạng vi khuẩn oxy hóa Fe(ll), khử nitrate tại một số môi trường
sinh thái ở Việt Nam
N g u yễn T h i T u y è n V B in h T h u y h t ịn g 1
'Đ»1 hoc Khoa hoe Tự nínén. Đai hoc Quốc gia Hâ NỘI 1 VHn Vi jmh vậl vi C6ng nghệ Sinh hoc. Đai hpc Quốc gia Hả NỘI
Mờ đẩuTrong lự nhiện quâ lónh 0* y hôa Fe (ll) châi rvhôn diện lừ kđ n tra ie chũ yéu
d iễ n r a ở r a n h Q1Ớ1 h ié u k h i (Củ Oxy) v ú ky k h i (K hôog c ó 0 *y ) ( /o n g ựjrọ t r á m (ic t i ở
đ é y c i c Ih ũ y v ư c O x y t>òd F e (H ) Kéi h o p vứ< K hữ n i ir a te c ó t h í S ô n g v a i Irớ q u a n
t r ọ n g iro n g mộ< t r ư ơ n g 6 n h iê m vón n ô n g đ ộ F e (ll> c a o (ớ o tfi<éu Qxy) v à o ô n g d ỏ
m lr a i* c a o (ỜQ crtAl h ữ u c o tx p h â n H úy t a o th a n ĩi) íV V eber e i a l 2 006}
C i c I03J VI k rtu á n VỬ1 k h á n â n g t ié n h â n h p h á n ưr»fl 0 »¥ h o a h h ũ n ã y c o th é C ung
môí luc Ihưc mén đirợc ftai Íìíiiệm vụ. (0 crtuyẻo Fe<ll> rtòa lan troog nvrO-c vè
d a n g Fe<lll> k é l l ũ a . v á {a) loạ i b ố m ir a le c rtu y Ể n Ih ầ n h d a n g N-
Q u â tr in h Dxy f iổ a F e ( l l ) b a n g n i l r a t e d i ỉ n ra nhu- l a u
to Fe;‘ ♦ 2 NO,- * 24 H?0 - >OFe(OHj, 1 ♦ Nj I *9h;ỊT ro n g n g r tiê n c ứ u n á y c t tu n g tỏ i t i ê n n à n h liro h i i u l in h d a <J«rig c ù a VI k h u i n k h ứ
nrtrale sừ đụny Fe(ll) là nguồn diện li/lífc môi sổ mỗi Irưởng Sính Ihâi dai diện ờ V iệ t N a m
Sơ đồ thí nghiệmc iy m lu
(tìu r OềỊ M Bun ỠArt rLi r>9 ngập nưcrc Trim l< * V«A m/Oc lơ)
Ũỉk ra n pM o m t u i MPN(F*tll)iTíO» * A cru i« i 1
ũ (1n w u h o n*r*noc C=I 1 Phâr l*fi oKmg đ» <Ho 1 . PCR DGG£16SfO*tA 1 1 1MI
ũa ũ
1 APQRAì&SiONA 1 C « M r g ttv ^ 9 * 1binh tự
rtAngưng Ị1 ũ
Kết quả và Thảo luậnsố lượng té bào trong các mẫu ngmèn cửu
Níiin b<ếĩ w co mai CUJ »1 tmuAn o>r to a FtlM j Iroogf.*c 6*B thống Qua b>ếr\ dố»m tu i i c cua mò' iroof'S lư 1'ềTQ »*nh (Fe>ii Mf>g vang r»Ju iF f i li i
Số lưOi*^ «1 «»y K * 1 4H>1| hfHín 4i»le u c Oiríi Ihồng Qua P*^IOtìg ph*p MPN Sò n/ong 1* bâo 10: - 10* n ong đo ►i » /0 0 5 (♦ bao I'ong n»i(* lự rưtng ■» n ự « >0 '
Nghiên ciru các nhóm vi khuản chièm ưu thé bẳng PCR-DGGE
Phố«*» 01 w*« tinh (DOGỉ) ( M n K h 4o#ni(3
rONA eúa quin Ihề ví tinh vặt Irong cAc ống
MPW cua c*c mếu r^ iể n c w
A - §0 fHrti rtọot. R - 8un chén i u V ị n^tỉ
<VỨC. 0 - Ệun dèm lích v«n b4 o
Ar>*r/vrr\ỵ, o b a tie ' (.vProí«o©acífd*j co
•nil Irorg ci } múi (lựong rvạniío (Ui
Ps»ưđtyn»viís P'0/Í0ti»cle''»>cnié»n u iné (rong m iu bu" o c^ản ruồng n g jp rx/
Đánh giá tinh đa dạng cùa các chùng đơn đâ phân lập bàng ARDRA
0 ^ c**n rụoog T rim K lỉnuO c Son d*> »0lu* *ỊÌữ n ir tc w ^ (a^f\ ryỊM
P lú f l Itc h AROflA 8 *n 16S rũNA C M l ỉ cn u n g «1 fchuln o » y HOI Flrtilj. trtt/
rvlratc «ỈU t h tir ly blng CMC eruyme J01 fun «1 Mtpl 'N1 •• M I? Kì
n>«v «ya 12 pí\an <40 đưo< 'ự e*e mÌM ngr.itn c v v M Mame»
<• »2 ngh<r- ÍV I/M đ ư o t » tp v»» 5 r* n rr' di fiv»rè" ►*»ac nnav '•"»! 4 i -ÍVVỊ o Vư-fệfì e#lu
thvxic *»0 đ • 'f>v n-k- OV' đ*v ni<fi >.v> 'V/OC lơ -,e< 1» ln*i da đv>a fi. trtjfinc*0 ''*'4' '-<c ,ra ti Dun díy 40 <a i*!Ập \a mk bu''efií'> ‘v-yig ''flit <■ &» CN y'J 'N ? ữ i ĩ V* d ế -ư ito cr-c 3 rtguò" llp < a ữm *140 cKoc*r. ohorr. ARDRA{hir#i
oe 1«/» 1.IV5'' 4 Ỉ p ^ á '' 1<n t '-n r tu đ i r -ivC j t 9** 16S '0*1* «* ứ*^i <5»V1 r<K

K há n à n g ứ n g d ụ n g của c h ủ n g Paracoccus s p IN12
A B
::::n u
I ’ • mPmữceu* *p I»1J
1 iTom *■« |Kf tyi
Hojrt tính oiy hò* khir nHr«4> ỊA) V* hkrrh lh*i 1« Mo cú* chu»9 «12
Kết luậnÝ Ỉ i lượng Ự1 Utuin 0«T F*<llị khư n4rM» Ironfl c t í rn iu bur> IÍH1 IKập ứ m úf w c 'KfOc ngợ c tiín ruòng nọ4fr nưrx V* «v*fl «wơc <0 v*n b itn đươc a < <íkV> nế*« Vong khoảng 10’ »0* (rong dó
m iu txr> à thjkl >uộr«g n0 l p nuO>c co tổ l i M o »1 M 'uifl A tr c*0 rvyn CA (9 3 • lỡ ’ TD'fll
*V tU w in 01) h o t F•<r)> u> j n * |t« !•> cềc <XÒI IrưOng ng ti^ n c ư u co linh đa d«ng Kh* cỞO 12 chung to n p N n 43 tư CẬC m lu MPN đươc >(p «40 5 nhóm ARDSA »ư^*c níMư Iroog p h in (>C«1 tư đung n il
'rxrtT* 9«1 nạn M#*m V* Ut4*tPnftnlttft fftftrn pftftn loa> bịng c+n/png p n tp PCH OOGE ứò\ «01 0*n 16 S íONA cho m iy
4#wwom|rjo6*ct« It rré< bong nNfrnj r tw * OìArri ưu lh£ ca rritt d ò h t mối Kư eng 4rf»fi iru i
rtn, đỏng VN lí* M*p *k> cfxi lilrft tííụyln hc4 »4t lror>fl cầe mử« irưonọ nghiện cưu (nôoo Qua *h» nấng K t i Irưứrtg k | tư* I fỳ f Mill] Ké* hop vOi k tt ÍỊ\JÌ ngNt^n c ư u o íc thu n g đ o n pf»jn lip đươc lưE*c d*>9 m6* fiwO«9 r^i4n Cw tfi0 »M|P Paiicoccưt v» Pitưito^ửAii 4 hat nhom w fchuin chim to re M « qut líirth 0>r ho* f »{ll| kfx> n t ia l i lương ưng dong v» Iro Quan dong Iroog <nửi (<uorvg ao IW*C ngự «• c*4o luòng ng4p mrức
■>6*<fKx>s 4 » a ^ n t è l k ỉ IN? v i IN1?C0 I/»V> tư ọ tn l6 S rO N A ce a i <ưor\g dốnfl c*o v9ì eae loai VI *n»utw nynữ4K*t ơ#u»ỈOQtnttrt (W%J P isuO em ertts tỉu i ỉở '1 |97*> VI P»<Ko<tut
60 vẳy đưọc đ * lén h/ơl to A n* ũw rr\yiữ t>»cft *fi \ n ĩ P * iu 9o m o o tt H> >N7 v» P trtcóccu tap M U
♦Cn wni lÉ«*M*g*<Unjềrtajii*fiinến<}ưnQ<lyngnon9 v>*e «v ly c*c rtguổn nv/ừe riỊỊèm «W\4m »é< «* nrlr«t
X Ỉ N C H Ấ N T H À N H C Ả M Ơ N
N jihu'n lU u n.it tlirữi. it<ựk h iê n "t»t> >ư h ỗ «|Ọ Im li I>hi lự .tè 1 . 1 1 n ' *«• I.IV
Ịí«i 'UI ..... rOiijiiiin.nl V4» Vi »11.1. 'ÔI V., I .1.. I^j.i Milll l»Ov IUM .UN AiI9i>»]ié>i Lifrii <!«<•> Im iti iliUv h ién 'l è «.n
2

ĐẠI HỌC QUỐC GIA HÀ NỘI
Đ Ề C Ư Ơ N G
Đ È T À I K H C N T R Ọ N G Đ IẺ M / Đ Ặ C B IỆ T / C Ấ P Đ H Q G H N
Tên đề tài; Đa dạng sinh học của vi khuẩn kỵ khỉ ôxy hỏa sắt (II), khử nitrat
trong một số môi trường sinh thái và khả năng ứng dụng của chúng
Mã số: QG. 07. 26
Chủ trì đề tài: TS. Đinh Thúy Hằng
Cơ quan: Trung tâm Công nghệ Sinh học - Đại học Quốc gia Hà nội
HÀ NỘI, 2007

ĐẠI HỌC QUỐC GIA HÀ NỘIĐỀ CƯƠNG
ĐỀ TÀI KHCN TRỌNG ĐIỂM/ĐẶC BIỆT/CÁP ĐHQGHN1. Tên để tàiTiếng Việt: Đ a dạng sinh học của vi khuẩn kỵ khỉ ôxy hoá sẳt (II), khử nitrat trongmột sổ
môi trường sinh thái và khả năng ứng dụng của chúng.
Tiếng Anh: D iversity o f anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria in some
ecological environments and their potential use.
2. Thòi gian thực hiện: 24 tháng (Bắt đầu từ tháng 5/2007 đến tháng 5/2009)------------------------ (■-----------------------------------------------------------------------------------------------------------------------------
3. Đe tài thuộc lĩnh vực ưu tiên: Công nghệ sinh học môi trường
4. Đe tài có trùng vói một đe tài đã hoặc đang tỉen hành không? Nêu có, nêu lý do
Để tài cỏ liên quan với đề tài Nghiên cứu cơ bản đăng ký thực hiện từ tháng 1 năm 2006 đến tháng 12 năm 2007. Với tiêu đề: “Nghiên cửu một sổ nhỏm vi sinh vật tham gia chu trình
chuyển hoá nitơ và phospho trong môi trường ô nhiễm và khả năng ứng dụng của chúng trong xử lý ô nhiễm môi trường", đề tài nghiên cứu cơ bản đăng ký thực hiện với Bộ Khoa học và Công nghệ tập trung nghiên cứu các nhóm vi khuẩn đóng vai trò chủ đạo trong quá trình chuyển hoá nitơ và phospho như ôxy hoá ammonium thành NCV, ôxy hoá N 0 2~ thành NOj-, khử N 03" thành N2 , tích luỹ phosphat dưới dạng polyphosphat trong tế bào. Trong các qui trình xừ lý nước thải giàu chất hữu cơ ammonium được tạo ra nhiều do các hợp chất hữu cơ, đặc biệt là protein bị phân huỷ. Khâu cuổi cùng trong việc loại bỏ các hợp chất nitơ trong nước thải hữu cơ do vi khuẩn khử nitrat đảm nhiệm. Đây là các loài vi khuẩn sổng kỵ khỉ hoàn toàn, sử dụng chất hữu cơ trong nguồn nước ô nhiễm để khử nitrat thành khí nitơ và tích luỹ năng lượng.
Đề tài Đặc biệt đăng kỷ với Đại học Quốc gia trinh bày trong bản đề cương này tập trung nghiên cửu một nhóm vi khuẩn đặc biệt có khả năng sinh trường tự dưỡng, sử dụng iông sắt II làm nguồn điện tử để khử nitrat. Các loài vi khuẩn thuộc nhóm này đóng vai ừò quan trọng trong tự nhiên, chúng không chi tham gia vào chu trình nitơ mà còn có ảnh hường lớn đến chu trình chuyển hoá kim loại. Nghiên cứu này được đề ra với hai mục tiêu. Mộĩ là đặt ra nhiệm vụ nghiên cứu một nhỏm vi sinh vật mới ờ Việt nam, và hai là tìm hiểu khả năng ứng đụng của chúng vào việc loại bỏ kim loại nặng trong nước thải công nghiệp.

5. Chủ trì để tài (Kèm theo Lý lịch khoa học theo biểu mẫu 02/KHCN/ĐHQGHN)
- Họ và tên: Đinh Thuý H ằng Nữ
- Năm sinh: ỉ 970
- Chuyên môn đào tạo: Vi sinh vật
- Học hàm, học vị: Tiến s ĩ
- Chức vụ: Nghiên cứu viên
- Đơn vị công tác: Trụng tăm Câng nghệ Sinh học, ĐHQG Hà nội- Đ ịa chỉ liên hệ: E2, 144 Xuân thuỷ - cầ u giấy, Hà nội
- Số điện thoại: 9Ỉ10402; Fax: 7547407; Email: [email protected]
Tóm tắt hoạt động nghiên cứu của chủ trì đề tài (Các chương trình, đề tài nghiên cứu
khoa học đã tham gia, các công trình đã công bố liên quan tới phương hưởng của đề tài)Thời gian Tên để tài/ công trình Tư cách tham gia Cấp quản lỷ/noi công bố
1/2006-12/2006
Nghiên cứu qui trình sản xuất chế phẩm vi sinh dùng trong nuôi trồng thuỷ sản. '
Chủ tri Cấp Đại học Quốc gia
6/2006-6/2008
Nghiên cứu các nhóm vi khuẩn tham gia chu trình nitơ và phospho và khả năng ứng dụng của chúng trong xử lý ô nhiễm môi trường.
Chủ trì Bộ Khoa học và Công nghệ
11/2005-nay
Quĩ gen Vi sinh vật Thành viên tham gia, phụ trách nhóm Prokaryotes
Đê tài độc lập, Bộ khoa học và Công nghệ
Tóm tắt hoạt động đào tạo sau đại học của chủ trì đề tài trong 5 năm trở lại đây
Thỏi gian Tên nghiên cửu sinh Tên hoc viên cao hoc • •
6 . Cơ quan phối hựp và cộng tác viên chính của đề tài (ghi rõ các đơn vị và các cánhân đã được mời và nhặr\ ỉệil mờị ịham gia để tài, mỗi cả nhân tham gia Đe tài phải có bản Lý lịch khoa học biểu, mâu 02/KHCN/ĐHQGHN, và ý kiến xác nhận đồng ý tham gia đồng thực hiện ti&ifiiho òếr.
TT Cơ quan phổi^yp* ...' 1 * ' l ? ’ 4 • ' C i-k - i i , ' u b y t A ,
Cộng tác viênHọ và tên Chuyên ngành
1 Trung tâm CNSH - -ÍV1 Nguyên Minh Giảng Công nghệ môi trường

2 Trung tâm CNSH Nguyễn Thị Vân Công nghệ sinh học3 Trung tâm CNSH Nguyên Thị Kim Quy Vi sinh vật học
7. Thuyết minh sự cần thiết hình thành dự án
- Tổng quan các công trình nghiên cửu trong và ngoài nước liên quan tới vấn đề
nghiên cứu của đề tài (trích dẫn những tài liệu mới nhẩt trong và ngoài nước)Sắt là một trong những kim loại phổ biến nhất ưên trái đất. Thông thường sắt tồn tại ở dạng ôxit sát (III) ít tan trong nước và có màu vàng. Trong môi trường pH trung tính, dạng hoà tan trong nước (Fe2+) chỉ tồn tại ở điều kiện không có ôxy, ví dụ như ở đáy các thuỷ vực, nơi ôxy hoà tan trong nước đã bị các vi sinh vật hiếu khí sử dụng để phân huỷ cốc hợp chất hữu cơ. Với hiệu điện thế ôxy hoá khử Fe3+/Fe2+ tại pH 7 vào khoảng +200 mV, iông sắt (II) cỏ thể trở thành nguồn điện tử cho các quá trình hô hẩp kỵ khí, điển hình là khử nitrat thành N 2 do một số nhóm vi khuẩn đảm nhiệm (Benz và cộng sự, 1998). Phản ứng ôxy hoá khử do vi khuẩn thực hiện như sau:
10 Fe2+ + 2 NCV + 24 H2 0 -► 1 0 Fe(OH)3ị + N2 t+ 9 H2ÍTrong tự nhiên, quá trình ôxy hoá sất (II) với chất nhận điện tử là nitrat chủ yếu
diễn ra ở ranh gịới iiiếu khí (có ôxy) và kỵ khí (không có ôxy) trong lớp trầm tích ở đáy các thuỷ vực. ôxy hoá sắt (II) kết hợp với khử nitrat có thể đóng vai trò quan trọng trong môi trường ô nhiễm với nồng độ Fe2+ cao (do thiếu ôxy) và NO3 ' cao (đo chất hữu cơ bị phân huỷ tạo thành) (Weber và cộng sự, 2006). Các loài vi khuẩn với khả năng tiến hành phản ứng ôxy hoả khử này có thể cùng một lúc thực hiện được hai nhiệm vụ, thứ nhất là chuyển Fe2+ hoà tan trong nước về dạng Fe3+ kết tủa, và hai là loại bỏ NO3 , chuyển thành
dạng N2 không độc hại.Vi khuẩn dùng iông sắt (II) làm nguồn cho điện tử để khử nitrat được phân lập đầu
tiên tò các lớp trầm tích ao, hồ nước ngọt tại Bremen, CHLB Đức năm 1996 (Straub và cộng sự 1996). Một sổ công trình nghiên cứu tiếp sau cho thây sự có mật khá phổ biên của nhóm vi khuẩn này với mật độ khá cao ( 1 0 6 tế bào/g trầm tích) trong các điêu kiện môi trường khác nhau, bao gồm cả nước ngọt, nước lợ và nước mặn và tại nhiều vị trí địa lý khác nhau trên thế giới (Straub và cộng sự, 1998). Các loài vi khuẩn phổ biến nhất trong nhóm này được biết đến hiện nay là các loài thuộc chi Chromobacterium vàKlebsiella (Benz và cộng sự, 1998; Weber và cộng sự, 2006).
Tuy nhiên ở Việt nam cho đến này chưa có công trình nghiên cứu nào đê cập đênquả trình sinh học dùng Fe2+ để khử NO3' cũng như các loài vi khuẩn đảm nhiệm quá trình này Do vậy đề tài được đặt ra với mục tiêu nghiên cứu tính đa dạng của vi khuân khử nitrat sử dụng Fe2+ là nguồn điện tử duy nhất cũng như tìm hiểu khả năng ứng đụng

của Cftung trong việc phát triên chế phẩm sinh học để xử lý các nguồn thải đồng thời nhiễm kim loại nặng và chất hữu cơ.- Lý do chọn đề tài
Tỉnh th ờ i s ự của đề tà i
Phân huỷ sinh học là một trong những biện pháp giải quyết ô nhiễm hữu hiệu đã được kiểm chứng tại nhiều nơi trên thế giới. Yếu tổ quan trọng hảng đầu quyết định thành công của biện pháp nàỵ thuộc vê các vi sinh vật có khả năng phân huỷ đặc biệt. Để cỏ thể cho ra đời các sản phẩm sinh học làm sạch môi trường, trước tiên các nhà khoa học phải hiểu rõ chưc nâng va cơ chc hoạt động của nhiêu nhóm vi sinh vật khác nhau, cũng như sự phân bô của chúng trong môi trường ô nhiễm và không ô nhiễm. Mặc dù đỏng vai trò không nhỏ trong chu trình chuyển hoá vật chất ngoài tự nhiên nhưng các loài vi khuẩn có khả năng đùng Fe2+ để khử nitrat còn chưa được đề cập đến trong các nghiên cửu về vi sinh vật, sinh-địa-hoá hay môi trường ờ Việt nam. Bên cạnh đó, các nghiên cứu đã công bố trên thế giới mới chỉ đưa ra kết quả khảo sát về nhóm vi khuẩn này ở châu Âu với điều kiện sinh thái hoàn toàn khác biệt với nuởc ta. Do vậy, đề tài được đặt ra không những đề cập đến một vấn đề còn chưa được nghiên cửu ờ Việt nam mà còn nhàm đem lại những hiểu biết thêm về vi khuẩn ôxy hoá Fe2+ để khừ nitrat cho khoa học nói chung.Tinh cấp th iết đáp ứng nhu cầu ph át triển kinh tế - xã hội
Mục tiêu thứ hai của đề tài là tìm hiểu khả năng ứng dụng của các loài vi khuẩn ôxy hoá Fe2+ để khử nitrat phân lập được từ các môi trường khác nhau ở Việt nam. Thông qua phản ứng ôxy hoá Fe2+ thành Fe3+ và khử N0 3 ~ thành N2, các loài vi khuẩn này có khả nãng loại bỏ NO3- và Fe2+ cỏ trong môi trường ô nhiễm. Hơn thế nữa, các loài vi khuẩn phân lập được sẽ dùng để nghiên cứu thêm về khả nãng loại bỏ một số kim loại khác theo cơ chế tương tự như đổi với sắt, ví dụ như chuyển Mn2+ tan trong nước thành Mn4+ kết tùa.
8 . Địa bàn tiến hành nghiên cứu (xã, huyện, tỉnh, vùng)Hiểu biết thực tễ của tác giả về địa bàn nghiên cứu:Mẩu để nghiên cứu đa dạng và phân lập vi khuẩn ôxy hoả Fe2+/khử N 03” sẽ được lấy ờ các môi trường nước ngọt và nước lợ. Mầu môi trưởng nước ngọt là trầm tích ở đáy các hồ trong địa bàn Hà nội hoặc chân ruộng lúa ngập nước ờ vùng ngoại thành quanh Hà nội; mẫu môi trường nước lợ là mẫu trầm tích ven biển tại vịnh Hạ long.Tinh đại diện của địa bàn nghiên cửu:Địa điểm lấy mẫu Hà nội và Hạ long được chọn làm đại diện cho môi trường nước ngọt và môi trường nước lợ. Bên cạnh đó, các chi sổ về môi trường của hai địa điểm được chọn này đã công bố trong các nghiên cứu trong nước cho phép lưa chọn lấy mẫu một cách chính xác

theo tiêu chí mà để tài đã đặt ra.
9. Mục tiêu của đề tài
- Nghiên cửu vả so sánh đa dạng sinh học của vi khuẩn ôxy hoá Fe2+/khử N 03' tại một sổ môi trưởng sinh thái khác nhau.
Phan lạp cac nhom VI khuan đại diện, chiêm đa sô trong mỗi loại môi trường sinh thái và nghiên cửu các đặc điểm phân loại, sinh lý, sinh hoá của chúng.
- Nghiên cửu khả năng ứng đụng các chủng vi khuẩn phân lập được vào việc kết hợp loại bỏ Fe2+, Mn2+, NO3" trong môi trường ô nhiễm.
10. Tóm tắt nội dung nghiên cứu của đề tài
- Mầu trầm tích ở đáy hồ, mẫu đất ờ ruộng lúa ngập nước và mẫu trầm tích ven biển sẽ được lấy theo yêu cầu đảm bảo kỵ khí (tránh tiếp xúc với ôxy), đưa về phòng thí nghiệm và giữ ở 4°c cho đến khi tiến hành phân tích vi sinh vật.
- Số lượng vi khuẩn ôxy hoá Fe2+/khử NO3 ' sẽ dược xác định theo phương pháp MPN(Most Propable Number) trong môi trường địch thể chứa Fe2+ làm nguồn điện tử và NO3 - là chất nhận điện tử trong điều kiện kỵ khí hoàn toàn. Thành phẩn khoáng cùa môi trường dảm bào tương ứng với môi trường sinh thái tại địa điểm lấy mẫu.
- Đa dạng về thành phần loài trong các mẫu khác nhau sẽ được xác định thông qua phương pháp sinh học phân tử, tách ADN tổng sổ trực tiếp từ các lô thí nghiệm ở nồng độ pha loãne khác nhau trong dãy MPN và phân tích tính đa dạng cùa một đoạn gen 16S rARN của tập đoàn vi sinh vật thu được trong mỗi lô bàng phương pháp PCR/DGGE.
- Các chủng vi khuẩn ôxy hoá Fe2 7khử N 03~ dại diện, chiếm đa số tại mỗi địa điểm lẩy mẫu sẽ được phân lập từ ống pha loãng cao nhất có vi sinh vật phát triển của dãy MPN bằng phương pháp ống thạch bán lỏng.
- Các chủng vilchuẩn phân lập sẽ được nghiên cứu về đặc điểm sinh lý sinh hoá và phân loại dựa trên trình tự 16S rADN. Kết quả của nghiên cứu này sẽ làm cơ sở để tuyển chọn các chủng có hoạt tính sinh học (khả năng chuyển hóa Fe2+ và N 03“ ) cao và nghiên cứu khả năng ứng dụng của chúng vào việc kết hợp loại bỏ Fe2+, Mn2+, N 03“ trong môi
trường ô nhiễm.
11. Các chuyên đề nghiên cứu dự kiến của đề tài (ỉẽn và nội dung cùa từng chuyên đề)1. Nghiên cứu sự đa dạng của vi khuẩn ôxy hoá Fe2+/ khử N 03" trong một sổ môi trường
sinh thải khác nhau.2. Phân lập phân loại và nghiên cứu đặc điểm sinh lý sinh hoá của các chủng vi khuẩn ôxy
hoá Fe2+/ khử NO3" từ một sổ môi trường sinh thái khác nhau ở Việt nam._____________

trong lỉnh vực môi
mvc tiêu nghiên cứu
3. Tim hiểu khả năng ứng dụng của các chủng vi khuẩrTphanlạịrđ— trưởng. Ọc
12. Câu trúc dự kiến báo cáo kêt quả của đê tài (chi t iế th ^ á lả ^ k ^ —--------1. Tổng quan tà i liệu: về vi khuẩn ôxy hoá Fe2+/ khử NCV và đưa ra^ mục)
2. Vật liệu và phư ơng ph áp dùng trong nghiên cửu:
- Kỹ thuật lấy mẫu kỵ khí và bảo quản mẫu.Kỹ thuật lây mâu kỵ khí và bảo quản mẫu.
Các phương pháp vi sinh vật: phân lập và nuôi cấy vi khuẩn kỵ khí định lượn • kh ẩ băng phượng pháp MPN, nghiên cửu các đặc tính sinh lý cùa v s v (điều kiện s h trườntAi tm ntiA /"*<■* oViát tkí/ili °
mã hóa 16S rARN.
- Các phương pháp phân tích hoá học: phân tích một số yếu tố sinh địa hóa trong môi trường, đo nồng độ Fe2 +/Fe3+ và NO3- trong các thí nghiệm với vi sinh vật
3. K ết quả và thảo luận:
- Xác định số lượng vi khuẩn ôxy hoá Fe2+/khừ N 03~ thông qua phương pháp MPN- So sánh đa dạng của nhỏm vi khẩn này tại các địa điểm lấy mẫu khác nhau thông qua
phương pháp PCR/DGGE đoạn gen 16S rARN.- Phân lập các chủng vi khuẩn đại điện.- Phân loại các chủng vi khuẩn phân lập được.- Đặc điểm sinh lý, sinh hoá của các chủng vi khuẩn phân lập được.- Khả năng ứng đụng cùa các chủng vi khuẩn phân lập được.4. Kết luận, s. Kiến nghị6. Tài liệu tham khảo.
13. Tính đa ngành và liên quan của đê tàiVới nội dung đề ra như trên, đề tài sẽ sử dụng chủ yếu các kiến thức về vi sinh vật học, sinh học phân tử vả công nghệ môi trường.14. Phương pháp luận và phương pháp khoa học sử dụng trong đe tàiCác vấn đề đặt ra trong khuôn khổ của đê tài được giải quyêt thông qua các phương pháp vi sinh vật học truyền thống kết hợp với các phương pháp sinh học phân tử tiên tiến15. Khả năng sử dụng cơ sở vật chat, trang thiet bị (tên các phòng thí nghiệm sẽ được sử

Đề tài sẽ được thực hiện trong điều kiện trang thiết bị và cơ sờ vật chất hiện cỏ của Trung tâm Cồng nghệ sinh học, Đại học Quốc gia Hà nội, đặc biệt là các trang thiết bị của phòng Bảo tàng giống chuẩn Vi sinh vật.16. Khà năng hợp tác quoc te- Đề tải cô sự trợ giúp cổ vấn khoa học từ viện Max Planck Vi sinh vật biển Bremen CHLB Đức và Viện Kỹ thuật & kiểm tra chất lượng Quốc gia (NITE) Nhật bản.17. Các hoạt động nghiên cứu của đe tài
- Nghiên cứu lý thuyết: tổng quan tài liệu về vi khuẩn ôxy hoá Fe2+/khử N 03'- Điêu tra khảo sát: xác định sô lượng và đa dạng vê thành phần loài của vi khuẩn ôxy hoá Fe2+/khử N 0 3“ trong các môi trường sinh thái khác nhau.- Viết báo cáo khoa học: đưa ra bức tranh tổng quát về sự phân bổ của vi khuẩn ôxy hoá Fe2+/khử NO3 ” trong tự nhiên và vai trò của chúng trong chu trình chuyển hoá vật chất- Nghiên cứu khả năng ứng đụng các chủng vi khuẩn phân lập được từ đề tài vào việc loại bỏ Fe2+, NO3 - và Mn2+ trong môi trường ô nhiễm
18. Kết quả dự kiến 18.ĩ K êt quả khoa học- Có khả năng bổ sung các loài vi khuẩn mới cho khoa học ở Việt nam và thế giới- Dự kiến sẽ công bổ kểt quả nghiên cứu thu được từ đề tài trong 2 bài báo khoa học18.2 Kết quả ứng dụng- Các kết quả điều tra và đánh giá thu được từ đề tài sẽ làm cơ sở khoa học cho việc phát triển sinh phẩm làm sạch ô nhiễm môi trường.- Các chủng vi khuẩn thu được ừong đề tài nhiều khả năng là nguồn nguyên liệu để phát
triển sinh phẩm.18.3 Kết quả đào tạo
- Số cử nhân được đào tạo trong khuôn khổ của đề tài: 0 1
- Số thạc sĩ được đào tạo trong khuôn khổ của đề tài: 0 1
18.4 Kết quả về tăng cường tiềm lực cho đơn vị- Thông qua đề tài các cán bộ tham gia sẽ có cơ hội củng cô kỹ năng làm việc với nhỏm vi sinh vật kỵ khí tiếp cận với các phương pháp sinh học phân tử tiên tiến dùng trong nghiên
cứu đa dạng, sinh thái và phân loại vi sinh vật.
19 Nội dung và tiến độ thực hiện cùa đề tài (các công việc cân triêrì khai, thời hạn thực
hiện và sản phẩm đạt được)
dụng trong để tài)

TT Hoạt động nghiên cứu Thời gian thực hiện Sản phẩm khoa họcTừ thảng Đến tháng
1 Thu thập và viét tổng quan tài liệu 1/2007 3/2007 Tông quan tài liệu
2 Xây dựng để cương nghiên cứu chi tiết 2/207 3/2007 Đê cương KHChuyên đê 1: Nghiên cứu sự đa dạng của vi khuẩn ôxy hoá Fe2+/ khử NO f trong một số môi trường sinh thái khác nhau.Chuyên để 2: Phân lập, phân loại và nghiên cửu đặc điểm sinh lý sinh hoả của các chủng vi khuẩn ôxy hoá Fe2+/ khử NOj" từ một số môi trường sinh thái khác nhau ở Việt nam.Chuyên đề 3: Tìm hiểu khả năng ứng dụng của các chủng vi khuẩn 'phân lập được trong lĩnh vực môi trường.
3 Lây mâu, đặt thí nghiệm, xử lý kêt quả 3/2007 9/2008Chuyên đê 1: Nghiên cứu sự đa dạng của
vi khuẩn ôxy hoá Fe2+/ khử N 03~ trong một sổ môi trường sinh thái khác nhau.
4/2007 10/2007 Báo cáo chuyên đề, viết 1 bài báo khoa học
Chuyên đề 2: Phân lập, phân loại và nghiên cứtr-đặc điểm sinh lý sinh hoá của các chủng vi khuẩn ôxy hoá Fe2+/ khử N 0 3' từ một sổ môi trường sinh thái khác
nhau ờ Việt nam.
10/2007 4/2008 Báo cáo chuyên đề, viết 1 bài báo khoa học
C h u y ên đề 3: Tìm hiểu khả năng ứng dụng của các chủng vi khuân phân lập được trong lĩnh vực môi trường.
4/2008 8/2008
Xừ ]ý kêt quả 8/2008 9/2008
4 Viết báo cáo các chuyên đê 8/2008 9/2008 Hoàn chinh các báo cáo chuyên đề
Số chuyên để (như mục 2 )Hội thảo giữa kỳ

5 Bổ sung số liệu/thử nghiệm/ứng dụng 9/2008 11/2008Tống kết số liệu 11/2008 12/2008
6 Viêt báo cáo tông hợp 11/2008 12/2008Hội thảo lân cuôiHoàn thiện báo cáo
7 Nộp sản phẩm 11/2008 12/20088 Nghiệm thu để tài 1/2009
20. Phân bo kỉnh phí (Tưỳ theo đặc điêm chuyên môn của từng đề tài, các mục/tiểu mục trong bảng sẽ có những thay đổi cho phù hợp)
TT Nội dung Kinh phíNăm thử 1 Năm thử 2
1 Điểu tra khảo sát, thí nghiệm, thu thập số liệu 12 000 000 12 000 000Thuê khoán công lao động trong nước 1 2 0 0 0 0 0 0 1 2 0 0 0 0 0 0
2 Thuê, mua săm trang thiêt bị, nguyên vật liệu 14 500 000 14 500 000Mua vật tư, hoá chât dùng cho chuyên môn 14 500 000 14 500 000
3 Chi khác 3 500 000 3 500 000Mua văn phòng phâm 500 000In ân, photocopy 500 000
Quản lý phí 3 000 000 3 000 000
4 Tông kinh phí 30 000 000 30 000 000
21. Tài liệu tham khảo để viết đề cương- Tài liệu tiếng Việt
1 . Báo cáo khoa học. Hội nghị công nghệ sinh học toàn quốc, Hà nội 1999. Nhà xuât
bản Khoa học và Kỹ thuật.2. Bộ khoa học công nghệ và môi trường. Tuyển tập các báo cáo khoa học tại hội
nghị môi trường toàn quốc, năm 1998. Nhà xuất bản Khoa học và Kỹ thuật.
- Tài liệu tiếng A nh1 KL Straub M. Benz, B. Schink, F. Widdel. Anaerobic, nitrate-dependent
m icrobial oxidation o f ferrous iron. Applied and Environmental Microbiology
(1996). 62: 1458-1460.2 M Benz A. Brune, B. Schink. Anaerobic and aerobic oxidation of ferrous iron at
neutral pH by chemoheterotrophic nitrate-reducing bacteria. Archives o f
Microbiology (1998). 169: 159-165.

3. K.L. Straub and B.E.E. Buchholz-Cleven. Enumeration and detection of anaerobic ferrous iron-oxidizing, nitrate-reducing bacteria from diverse European sediments. Applied and Environmental M icrobiology (1998) 64: 4846 - 4856.
4. J.M. Senko, T.A. Dewers and L.R. Krumholz. Effect of oxidation rate and Fe(II) state on microbial nitrate-dependent Fe(III) mineral formation. Applied
Environmental Microbiololy (2005), 71: 7172-7177.5. K. A. Weber, J. Pollock, K.A. Cole, S.M. O’Connor, L.A. Achenbach and J.D.
Coates. Anaerobic nitrate-dependent iron (II) bio-oxidation by a novel lithoautotrophic betaproteobacterium, strain 2002. Applied and Environmental Microbiology (2006), 72: 686-694.
6. K.A. Weber, M.M. Urrutia, P.F. Churchill, R.K. Kukkadapu and E. E. Roden. Anaerobic redox cycling of iron by freshwater sediment Microorganisms. Environmental Microbiology (2006), 8: ỈOO-Ỉ13.
7. K. Zengler, 2000. Anaerobic, nitrate-dependent utilization of hydrocarbons by marine members of the alpha and, gamma subclass of Proteobacterỉa. Luận vãn Tiến sỹ, Đại học Bremen, CHLB Đức.
Ngày 26 thảng 01 năm 2007 CHỦ TRÌ ĐỀ TÀI
Đinh Thuý Hằng
Ngày 'ỉ tháng năm 2007 THÙ TRƯỞNG ĐƠN VỊ
Ngày OS tháng ó~ năm 2007 PHÊ DUYỆT CỦA ĐHQGHN
TL.GIÁM ĐÓC ĐẠI HỌC QUỐC GIA HÀ NỘI

-----------L U -----------
Số-stfâl /KHCN
CỘNG HÒA XẢ HỘI CHỦ NGHĨA VIỆT n a m 'Độc lập - Tự do - Hạnh phúc
Hà Nội, ngày ũẫ. tháng 5 ” nâm 2007
QUYẾT ĐỊNH CỬA GIÁM Đ ốc ĐẠI HỌC QUỐC GIA HÀ NỘI Về vỉệc phê duyệt danh mục đé tài đăc biệt cấp ĐHQGHN năm 2007
củá Trung tầm Công nghệ Sinh học
GIÁM ĐỐC ĐẠI HỌC QUỐC GIA HÀ NỘICăn cứ Nghị định số 07/2001/NĐ-CP ngày 1 tháng 2 năm 2001 của Chính phù về Đại học
Quốc gia;Căn cứ Quyết định số 14/2001/QĐ-TTg ngày 12 tháng 2 năm 2001 của Thủ tướng Chính phủ
về việc tổ chức lại Đại học Quốc gia Hà Nội;Căn cứ vào Quy chế Tổ chức và Hoạt động của Đại học Quốc gia được ban hành theo Quyết
định số 16/2001/QĐ-TTg ngày 12 tháng 2 năm 2001 của Thủ tướng Chính phủ;Căn cứ Hướng đẫn số 973/KHCN ngày 19/3/2007 của Giám đốc ĐHQGHN về “Quản lý
các đề tài khoa học và công nghệ ở ĐHQGHN";Theo đề nghị của Hội đồng ngành/liên ngành và Ông Trường Ban Khoa học - Công nghệ,
ĐHQGHN.
Điều ỉ : Phê duyệt danh sách đề tài đặc biêt cấp ĐHQGHN nẫm 2007 của Trung tâm Công nghệ Sinh học (Phụ lục kèm theo).
Điều 2: Kinh phí của các đề tài đặc biệt nói trên sẽ được chuyển vào tài khoản của Trung tâm Công nghệ Sinh học.
Điều 3: Giám đốc Trung tâm Công nghệ Sinh học có trách nhiệm:
- Lập hội đổng thẩm định để cương đối với các đề tài có mức kinh phí 100 triệu đồng.- Ký hợp đổng với chủ nhiệm đề tài sau khi có đề cương chi tiết đã được ĐHQGHN phê duyệt.- Cấp kinh phí cho chủ nhiệm đề tài theo Quyết định này.- Theo dõi và quản lý việc triển khai thực hiện đề tài; Đánh giá tiến độ và chất lượng triển khai đề tài sau 1 năm thực hiện.- Báo cáo lãnh đạo ĐHQGHN và đề xuất kiến nghị cho phép đề tài tiếp tục hoặc đình chỉ việc nghiên cứu tiếp tục năm 2008.Điểu 4: Ông Chánh Vãn phòng, ông Trưởng Ban Khoa học - Công nghệ, Giám đốc Trung tâm Công nghệ Sinh học và chủ nhiệm đề tài có trách nhiệm thi hành quyết định này.
QUYẾT ĐỊNH
Nơi nhận:- Như Điều 4.- Lưu VP, KHCN

ĐẠI HỌC QUỐC GIA HÀ NỘI CỘNG HÒA XÃ HỘI CHỦ NGHĨA VIỆT NAM* * * Độc lập - Tự do — Hanh phúc
Đơn vị: Trung tâm Công nghệ Sinh học = = = = = = = = =Số: /HĐ-KHCN
HỢP ĐỒNG THỰC HIỆN ĐẺ TÀI KHCN CỦA ĐHQGHN(DÀNH CHO CÁC ĐÊ TÀI ĐẶC BIỆT, CẤP ĐHQGHN, NCCB, CÁP c ơ SỞ)
• Căn cư quyet đinh so 1562/KHCN ngày 02 tháng 05 năiĩỉ 2007 củâ Giám đốc Đậi học Quốc gia Hà nội về việc thành lập đề tài và bổ nhiệm chủ nhiệm đề tài NCKH cấp ĐHQGHN.
- Căn cư vào bản dê cương nghiên cứu của đê tài mã số QG.07.26 và số kinh phí đã được phê duyệt.
Chúng tôi gồm:Bên giao (Bên A, đơn vị chủ ữì): Trung tâm Công nghệ Sinh học - ĐHQGHNĐại diện: TS. Dương Văn HợpChức vụ: Giám đốc
Bên nhận (Bên B, chủ trì đề tài): TS. Đinh Thúy HằngĐom vị công tác: Trung tâm Công nghệ Sinh học - ĐHQGHNĐiện thoại: +4 911 04 02Là chủ nhiệm để tài NCKH của ĐHQGHN, mã số QG.07.26
Hai bên thỏa thuận như sau:
Điều I: Bên B chịu ưách nhiệm tổ chức triển khai các nội dung nghiên cứu ừong năm 2007 - 2009 cụ thể dưới đây:
- Nghiên cứu sự đa dạng cùa vi khuẩn ồxy hỏa Fe2+/khừ N O 3' ữong một số môi trường sinh thái khác nhau ở Việt nam.
- Phân lập, phân loại và nghiên cứu đặc điểm sinh lý sinh hóa của các chùng vi khuẩn ôxy
hóa Fe2+/khử N o r đại diện.- Tìm hiểu khả năng ứng dụng của các chủng vi khuẩn phân lập được trong lĩnh vực môi
trường.
Điều 2: Bên B nộp cho bên A các sản phẩm khoa học theo nội dung và tiến độ thực hiện cùa đề tài theo 2 đợt: đợt 1 trước ngày 01/06/2008 và đựt 2 trước ngày 01/06/2009.
Đợt 1: Báo cáo tiến độĐợt 2: Báo cáo nghiệm thu vả các sản phẩm cùa đề tài (theo đề cương)
Điều 3: Kinh phí cùa đề tài:Tồng kinh phí: 60 000 000 (Sáu mươi triệu đông).

— l±J —
CỘNG HÒA XÃ HỘI CHỦ NGHĨA VIỆT n a m 'Độc lập - Tự do - Hanh phúc
S Ố ^ a / K H C NHà Nội, ngấyŨẴ. tháng 5* năm 2007
QUYẾT ĐỊNH CỦA GIÁM Đốc ĐẠI HỌC QUỐC GIA HÀ NỘI vể việc phê duyệt danh mục để tài đặc biệt cấp ĐHQGHN năm 2007
củá Trung tám Công nghệ Sinh học
GIẮM ĐỐC ĐẠI HỌC QUỐC GIA HÀ NỘICăn cứ Nghị định số 07/2001/NĐ-CP ngày 1 tháng 2 năm 2001 của Chính phù về Đại học
Quốc gia;Căn cứ Quyết định số 14/2001/QĐ-TTg ngày 12 tháng 2 năm 2001 của Thủ tướng Chính phủ
vé việc tổ chức lại Đại học Quốc gia Hà Nội;Cản cứ vào Quy chế Tổ chức và Hoạt động của Đại học Quốc gia được ban hành theo Quyết
định số 16/2001/QĐ-TTg ngày 12 tháng 2 năm 2001 của Thủ tướng Chinh phủ;Căn cứ Hướng dẫn số 973/KHCN ngày 19/3/2007 của Giám đốc ĐHQGHN về “Quản lý
các để tài khoa học và công nghệ ở ĐHQGHN” ;Theo để nghị của Hội đổng ngành/liên ngành và ồng Trường Ban Khoa học - Công nghệ,
ĐHQGHN.
Điều 1: Phê duyệt danh sách đề tài đặc biệt cấp ĐHQGHN năm 2007 cùa Trung tâm Công nghệ Sinh học (Phụ lục kèm theo).
Điều 2: Kinh phí của các đề tài đặc biệt nói trên sẽ được chuyển vào tài khoản của Trung tâm Công nghộ Sinh học.
Điểu 3: Giám đốc Trung tâm Cồng nghệ Sinh học có trách nhiộm:
- Lập hội đồng thẩm định đề cương đối với các đề tài có mức kinh phí 100 triệu đổng.- Ký hợp đồng với chủ nhiệm đề tài sau khi có đề cương chi tiết đã được ĐHQGHN phê duyệt.- Cấp kinh phí cho chủ nhiệm đề tài theo Quyết định này.- Theo dõi và quản lý viêc triển khai thực hiên đề tài; Đánh giá tiến độ và chất lượng triển khai đề tài sau 1 năm thực hiện.- Báo cáo ỉãnh đạo ĐHQGHN và đề xuất kiến nghị cho phép đề tài tiếp tục hoặc đình chi việc nghiên cứu tiếp tục năm 2008.Điêu 4 : Ông Chánh Vãn phòng, ông Trưởng Ban Khoa học - Công nghệ, Giám đốc Trung tâm Công nghê Sinh học và chủ nhiộm đề tài có trách nhiệm thi hành quyết định này.
QUYẾT ĐỊNH
Nơi nhận:- Như Điểu 4.- Lưu VP, KHCN

Đại học Quốc gia Hà Xôi
DANH SÁCH ĐỀ TÀI ĐẶC BIỆT ĐUỢC PHÊ DƯYỆT NĂM 2007 (Kèm theo quyết định số 4S6& /KHCN ngày QÍL tháng 5"năm 2007)
Đơn vị kinh phí: triệu đồnTT Mã sôTTẻn đề tài/Chủ trì Tổng
kinh phíKinhp
2007
1. QG.07.23Đa dạng sinh học của vi khuẩn kỵ khí oxy hoá sắt 2, khử nitrat trong các môi tnrờng sinh thái khác nhau và khả năng ứng dụng của chúng TS. Đinh Thúy Hằng
60 30
2 . QG.07.24Nghiên cứu đặc điểm sinh học và thăm dò khả nãng phân huỷ độc tố của một số chủng Microcystis phân lập ở hổ Hoàn Kiếm, Hà Nội TS. Nguyễn Thị Hoài Hà
100 50
3. Tổng cộng 160 80

ĐẠI HỌC QƯÓC GIA HÀ NỘI CỘNG HÒA XÃ HỘI CHỦ NGHĨA VIỆT NAM-------- M A - — — Độc lập — Tự do - Hạnh phúc
Đon vị: Trung tâm Công nghệ Sinh học = = = = = = = = =Sổ: /HĐ-KHCN
HỢP ĐỎNG TH ựC HIỆN ĐÈ TÀI KHCN CỦA ĐHQGHN (DÀNH CHO CÁC ĐỀ TÀI ĐẶC BIỆT, CẤP ĐHQGHN, NCCB, CẮP c ơ sở)
- Căn cứ quyết định số 1562/KHCN ngày 02 tháng 05 năm 2007 của Giám đốc Đại học Quốc gia Hả nội về việc thành lập đề tài và bổ nhiệm chù nhiệm đề tài NCKH cấp ĐHQGHN.
- Căn cứ vào bàn đề cương nghiên cứu của đề tài mẵ số QG.07.26 và số kinh phí đã được phê duyệt.
Chúng tôi gồm:Bên giao (Bên A, đơn vị chủ ừì): Trung tâm Công nghệ Sinh học - ĐHQGHNĐại diện: TS. Dương Văn HợpChức vụ: Giám đốc
Bên nhận (Bên B, chủ tri đề tài): TS. Đinh Thúy HằngĐơn vị công tác: Trung tâm Công nghệ Sinh học - ĐHQGHNĐiện thoại: +4 911 04 02Là chủ nhiệm đề tài NCKH của ĐHQGHN, mã sổ QG.07.26
Hai bên thỏa thuận như sau:
Điều ỉ : Bên B chịu trách nhiệm tổ chức triển khai các nội dung nghiên cứu trong năm 2007 - 2009 cụ thể dưới đây:
Nghiên cửu sự đa dạng của vi khuẩn ôxy hóa Fe2+/khử NO3- trong một số môi trường sinh thái khác nhau ờ Việt nam.
- Phân lập, phân loại và nghiên cứu đặc điểm sinh lý sinh hóa cùa các chùng vi khuẩn ôxy
hóa Fe /khừ NOj đại diện.- Tỉm hiểu khả năng ứng dụng của các chủng vi khuẩn phân lập được trong lĩnh vực môi
trường.
Điều 2: Bên B nộp cho bên A các sản phẩm khoa học theo nội dung và tiến độ thực hiện của đề tài theo 2 đợt: đợt 1 trước ngày 01/06/2008 và đợt 2 trước ngày 01/06/2009.
Đợt 1: Báo cáo tiến độĐợt 2: Báo cáo nghiệm thu và các sản phẩm của đề tài (theo đề cương)
Điều 3: Kinh phí cùa đề tài:Tổng kinh phí: 60 000 000 (Sáu mươi triệu đồng).

- Kinh phí năm 2007 - 200«: 30 000 000 (Ba mươi triệu đồng).- Kinh phí năm 2008 - 2009:30 000 000 (Ba mươi triệu đồng).
Kinh phí nảy bao gồm những khoản đổng góp nghĩa vụ theo qui định hiện hành của Nhà nước. Sổ kinh phí này chủ trì đề tài sỗ nhận tại Phòng Tài vụ của đơn vị.
Điều 4: Bên B có trách nhiệm chi tiêu kinii phí được cấp theo đúng mục đích, đúng ché độ tài chính hiện hành và quyết toán vói phồng tài vụ, chậm nhất trước n g ày .......................................
Điều 5ỉ Hai bên cam kết thực hiện đúng các điều khoản đã được ghi trong hợp đồng. Trong quá trình thưc hiện hợp đồng, hai bên phải thông báo cho nhau những vấn đề nảy sinh vả cùng nhau bàn bạc giải quyết.
Điều 6: Hợp đồng này làm thành 4 bản, Bên A giữ 2 bàn, Bên B giữ ỉ bàn, 1 bản gửi đến Phòng Tài vụ cơ quan.
Hà nội, ngày 14 tháng 5 năm 2007
ĐẠI DIỆN BÊN B
TS. Dương Văn Hợp

ĐẠI HỌCQUOC GIA HA NỤ1 Vỉện VI sinh vật và Công Bghệ *áni» học
CỘNG HOÀ XẢ HỘI CHỦ NGHĨA VIỆT NAMĐộc lập - Tự do - Hạnh phúc
PHIẾU ĐỀ NGHỊ
THAY ĐỔI TRONG QƯÁ TRÌNH THựC HIỆN ĐỀ TÀI KHCN CỦA ĐHQGHN
1. Tên đề tài: Đa dạng sinh học cùa vi khuẩn kỵ khí ôxy hoả sẳt (II), khử nitrat trong một sổ môi trường sinh thái và khả năng ứng dụng cùa chủng
2. Mă số đề tài: QG.07.26
3. Chủ tri đề tài: TS. Đinh Thúy Hằng
4. Cơ quan chủ trì: Viện Vi sinh vật và Công nghệ sinh học, Đại học Quốc gia Hà nội
5. Những thay đổi về tiến độ, thời gian nghiên cứu:Xỉn lui lai thời gian nghiệm thu đề tài 4 tháng (từ tháng 6/2009 sang tháng 10/2009) do chủ trì đề tài nghỉ gián đoạn 4 tháng (từ 9/2008 đến hểt tháng 12/2008) theo chế độ của nhà nurớc dành cho Đà Mẹ và ừẻ sơ sinh.
ố. Những thay đổi khác:- Xin rút không đăng ký hưởng dẫn sinh viên làm khoá luận tốt nghiệp do đối tượng nghiên cửu là vi khuẩn kỵ khí đòi hỏi kỹ năng thí nghiệm cao và thời gian dài, khó phù hợp với loại hình đào tạo này (đã đề xuất trong báo cáo tiến độ giữa kỳ tháng 1 năm 2008).- Hoạt động khoa học thay thể: gửi bài tham gia hội nghị khoa học quốc gia về sinh thái và tài nguyên sinh vật.
Ngày thảng năm 2009 Ngày ì/ tháng ) nám 2009 Ngày 18 tháng 3 năm 2009Cơ quan chủ quản duyệt Cơ quan chủ tri Chủ nhiệm đe tài
ỈSTSít

CỘNG HOÀ XÃ HỘI CHỦ NGHĨA VIỆT NAMĐộc lập - Tự do - Hạnh phúc
BIÊN BẢN HỌPHỘI ĐỒNG KHOA HỌC NGHIỆM THU CAP c ơ SỞ ĐẾ TÀI ĐẶC BIỆT CAP ĐHQGHN MÃ số: QG.07.23
1.Tin đé tài:
Đa dạng sinh học của vi khuẩn kị khí oxy hóa sắt n, khử nitrate trong 1 số môi trường sinh
thái và khả năng ứng dụng của chúng
2. Chủ trì: T$, &ĩ'ỵUL
3. Quyết định thành lập H ộ i đồng Khoa học nghiệm thu đề tài số 149 /QĐ-CNSH ngày
23 tháng 10 năm 2009
4. Ngày họp H ội đồng: 28/1012009
5. Danh sách các thành viên của H ội đồng:
1. TS. Dương Văn Hợp2. TS. Lê Thu Hiền
3. TS. Nguyễn Huỳnh Minh Quyên4. TS. Đào Thị Lương
5. TS. Nguyễn Quỳnh Uyển6 . TS. Nghiêm Ngọc Minh7. TS. Nguyễn Thị Hoài Hà
Chủ tịch Hội đồng Phản biện 1 Phản biện 2
Thư ký ủy viên ù y viên ửy viên
a. Có mạt; Ỷ người
b. Vắng mặt 0 ngưòi
6. người trình bày báo cáo:
Họ và tên - 7 S t W i TLi. >.'ij
7. Kết quả đánh giá:
Sô phiêu phát ra:
Số phiếu hợp lệ: 7-
Kết quả: -ffí
í kt 'V/

8. Tháo luận của Hội đồng
- 7 $ . Ì^Cị\u.yCrỉ th iy iĩ ỉì kin\t} C\ I U | 'ì) dec ✓ A ù / ; M\a7viị Ìàtì
Hu ì d (>\*Ị á c \ t \h <\ < y3 fc ít' J
- C ^ v u 'K t / i H-D >vìiA (Or/j ' Ì ^ i Í i / ị I r u ~ r t K ' ừ \ .
~ *7~ s . -0 » » \./j ^ /u ì t / ) /vny i'l n t? Ă ĩ
N K a o X í ' f G<Uy H ũ ỉ doxjj
— Ị) I \ A o b t £* ; i , Till T A l t f C o
-V* i ) ỉ f r \ . ị t o Y O u ị Í i i O I C Ì k -c s \ c v a h < r ) C í K O
+ 'Ỵĩửr Ị ỢítCi n <4 à y W\n ỵj f i n•+ í \ ĩ c I f ^w<.K(- i í " i r u Ậ} C i i f í ^ u o n Ậ i \ f i . n h 1 c 1 r a i . 1 \ Ĩ > I ít / i
J , ơ > J'WC \Ắ vi '1 O . U u A m
4 ‘V T Ai , , , ..• l \ U v f z . 7- i t A c £ A 0 ^ + Í V | ũ Oi-.-) (■•Ki ỷi > f ir>*
* 1 1 ụ ' ■' j '. i ■• *• ft )V*- ■•> f ‘Ợ '. ' > •

V c . c L ^ .ís ^ A n (VC,,
C í l c < v í v ^ i ; _ \ / ị £ CỴ(_J ự a ( I I I ) á <\ ể à J c . { M J y ^ o i y
l<^ n ù ó c 7 e ù c l * f c f i i a o ' n K o i ) V ' / C /Uiy 7
)<^L . H orT u -H''-A*Wt sCi.t2x.-f 9 A t >1 ^ 1' * ->J '.'ii fV *? .'Vi
PJuih it*iTíJ i T S' A/'juiyio ^ i ‘*y»i/> /Iíim6
+■ Co VỊ IV (virf* if/. , ^ j n (. w \ "KvvvC A «• i) r ( I.4 / ^
■4 ICiTf ^ I f ic-v i\ + k r \ iL l ; -I* /- <r. > r,t<- u ù ' r l IIIV. * y ' ' ỹ ^ ' '
' r ' i \ II ',1 iU t ' 1 ■' ' , IM r ì . ; ( )»w t o r / H/, < ;,,, ,u ; yl / i f .
* ỹlvã.) ' t m ^ h i ‘ Vj V -uf c i u , k y ctưC (c Ọ J./1 /J ci U1 i-\ I~x rí nt C ( ũ | l v o i : D i t ,(1 ^ Ki'ii.^j- ivjCift]' CHA hthipj** 1 n U f j ; x J ir> J j :tr
fc L . ( V >.' r I • I \ I\ IV 11 ( f \ c M il I J , k I/ > '• • ' ■ > ' • • ■ ,
;■ u . jU . ' ( i „ K .jU ^ '.M -, u " <‘f ^ H ^ ( r1 >
) c iV Í j I I I Í > J v -. *. t. ; 1 I / \ I *■ > j ' < ' )
• N í í í ì I A . ’ f X , .-J r" , ^ : " J ( w W O l ( ' - ■■•
K L ■ỉnt .; . k c - ; - 1 -"M \ f\ t v J )
■+v ‘ j 1 1 Uv ^ i I \ I A 1 » > f
7 c iV < j u ^ (/■
• 1 ■ V ■ .. V
tHI I »j'«'
• ^ t \ p X i jl 1 A x' V c . 'i c
1 <; . rv:0 ltl l < «\ 1 ^ ~1 1 . r- 1 / u 1 ^

V ! t ' C ị • Ị
ÌẴịi
' 7 $ ^ !(J í C A |ù l / | ■
• T t ' T qf uo. l i Ể* c „ / , y ^ • 'r
l T . . . . . . > . /.
■ỉ. - ' • t ĩ o a - ị r<t.i ^Vĩ r ri it tu /Ị f t< 4 :-ì
cVv^ q f u a . l i Ể * c „ / , y H \.ctrị f <~ì'ỳ ^ i i> i '7
u i.ữL V IC Ic J a i’ II v'À »ii ^ •• '■ ‘■1 ^
« I&/vtiV'jj t-ì JCÍm b A J it io c cJ-ct- i1. c ^ i '1 y h s t i x A »vr c. l
' Ộ h k ỉ ) Tì\.ii[j H ru WỊ TArt / ò r i ì t fÃ-M Jvc/ r<íí'i Al-D ■ 'f . w

Kết luận của Chả tịch Hội đổng: . .p j * 7 n ~ p . - r ' í : o 4 ; »\
2 T V<5yívì f í X ■ i i u ' c H u Ị r h p -fi }'l c* ■•/
- D l ' f r J o U - ^ i « 1 ^ 1 -Ij CÌ<Ả H-ỡ ^ 'v ío r iM A
THƯKÝ HỘI ĐỔNG
(Họ, tên và chữ ký)
Y
' f t t ' w ĩ ì i ì ừ í m1
CHỦ TỊCH HỘI ĐỒNG
(Họ, tên và chữ ký)

PHIÉU ĐĂNG KÝ
KÉT QUẢ NGHIÊN c ứ u ĐẺ TÀI KHCN
Tên đề tài:
Mã sổ: Cơ quan quẳn lý đề tài: Địa chi:Điện thoại:Ctf quan chủ trì đề tài: Địa chi:Điện thoại:Tổng chi phí thực chi: Trong đó:
- Từ ngân sách NN:- Nguồn khác:
Thời gian nghiên cú u:
- Thời gian bất đầu:- Thời gian kết thúc:
Tên các cán bộ phối họ p
- Chù trì đề tải:- Cán bộ tham gia:
Đa dạng sinh học của vi khuân kỵ khí oxy hỏa Fe(]l). khừ nitrate trong một sô môi trường sinh thái vả khà năng ừng dụng cùa chúng. QG.07.23Đại học Quốc gia Hả Nội144 đường Xuân Thủy, cầu Giấy - Hà Nội37 54 86 64
Viện Vi sinh vật vá Công nghệ sinh học. ĐHQGHN Nhà E2. 144 Xuân Thúy, cầu Giấv - Hà Nội 37 54 74 07 60.027.400
100%
không có.2 năm
6/2007 10/2009
nghiên cứu:
TS. Đinh Thúy Hằng
CN Nguyễn T hị Tuyền ThS. Nguyền Minh Giáng
Sô đăng ký đê tài Sò chứng nhận dãrití ký KQNC Bào mậiA. Phô biến rộng rãi
Ngày Ngày
Tóm tắt ý nghĩa, kết quả nghiên cứu:- Ba dạng môi trường khác nhau gồm có ao nước ngọt, ruộng lúa ngập nước và irầm tích ven biến
được chọn đê tiến hành nghiên cứu sự đa dạng cua vi khuân òx> hoá Fe(ll) khứ NO','. Đáy là những dạng môi trường trong đó vi khuân tham gia chu trinh chuyên hoá sắt và nitơ đóng vai trò quan trọng. Mầu bùn đáy (dưới 10 cm bề mặt bùn) tại các mòi trường sinh thái kẻ trên được thu thập và sử dụng đê phân tích nhóm vi khuân kv khí này .
- Số lượng vi khuẩn ôxy hoá Fe(ll). khư NO;, dD dược xác định bằng phương pháp MPN trong môi trườn ti dịch thê nước ngọt {đối vói mẫu ao và màu ruộng lúa) hoặc nước lự (đòi với máu
trầm tích ven biển) chứa Fe(ll) làm chấl cho diện tư. NO; làm chát nhặn diện tư và aceiaie/CO:
làm nguồn cacbon. Sự sinh trường cua vi khuân trong các ống MPN dược nhận biêt thòng qua thay đồi màu sắc cua môi Irirờnu nuôi cắ\ santi máu xàng nâu do 1 c(ll) bị ò.\\ hoá thánh f:e(lll).

DNA tổng số tử các ống MPN đại diện có vi sinh vật phát triển được tách chiết và sử dụng làm khuôn trong phản ứng PCR khuyếch đại đoạn gen I6 S rDNA có độ dài 550 bp. Phàn tích điện di biến tính đoạn gen trên cho thấy tính đa dạng khá cao cùa vi khuấn trong các ống MPN. Một số băng điện di đại diện được cắt và thôi gel để đọc trình tự, qua đó đã xác định đưọc các nhóm vi khuẩn chiếm ưu thế trong các môi trường nghiên cứu, cụ thê là Anaeromvxobacter Pseudomonas và Paracoccus.
- Đã tiến hành nghiên cứu tính đa dạng của vi sinh vật sinh trường trong điều kiện oxy hoá Fe((l)
khừ NO} trong các dãy MPN thông qua phương pháp tai in situ sứ dụng các đầu dò huỳnh quang (FISH) đặc hiệu cho ba nhóm vi khuân CX-. P- và ỵ-Proieobacteria.
12 chủng vi khuân kỵ khí được phân lập và tinh sạch (4 chúng đối vói mỗi dạng môi trường sinh thái). Tính đa dạng của các chúng này về mặt di truyền được xác định thông qua phân tích ARDRA đoạn gen mã hoá cho 16S rRNA dài 1500 bp sừ dụng hai enzvme M.spị và Haeìỉỉ. Các chùng đại diện cho các nhóm ARDRA được giài trình tự đè xác định vị trí phân loại cùa cá nhóm vi khuẩn mà chúng đại diện.
- Bằng phương pháp xác định lượng Fe(II) và NO}' trong môi trường nuòi cấy theo thời gian, chúng vi khuân IN 12 đã được chúng minh cỏ khá năng chuyên hoá hai hợp chất này khá cao, như vậy chủng này có tiềm năng sử dụng trong lình vực môi trường. Bên cạnh đó khá năng sừ dụngMn(I!) thay cho Fe(ll)cũngđã được nghiẻn cứu, tuv nhiên chung IN 12 không thê hiện kha nãng ứng dụng để loại bó Mn(IỈ) trong môi trưòng.
- Các kết quà nghiên cứu đã được công bố trong 02 bài báo khoa học. 0 1 báo cáo Poster iham gia hội n gh ị v ề s in h thái s in h vặt và 02 trinh tự g en dăntỉ ký tron g G e n B a n k .
- Đã tham gia đào tạo 01 học viên cao họcKiến nghị về qui mô và đối tuọng áp dụng kết quá nghiên cứu:- Nghiên cứu tìm giả thể phù hợp đè tạo chế phẩm chứa vi khuẩn oxlì Fe(II) khư nitrate.- Nghiên cứu thừ nghiệm áp dụng vi khuân oxy hóa Fe(II) khư nitrate dề xử lý nước ngầm nhiễm
sắt và ni tơ.Chức vụ Chù nhiệm đê tài Thú trường cơ
quan chù trì đe tàiChủ tịch hội đôiiii
đánh uiá chính thức ĩ i lii-'f/Mj
Thu trưởng cơ quan
Họ và tên Đinh Thúy Hăng Dươníì Vãn Họp Dưong Vãn Hợp p
Học vị TSKý tên
Đóng dấu
ý h ị V SINH H O ầ y ậ ìì V '•*.-»‘'71 i \ V --
' - - I ,
Pítli N ỷuỷto 3 S t n 4




















