
Cysteine peptidases in the tomato trypanosomatid Phytomonas serpens: Influence of growth conditions,...
11
This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript of Cysteine peptidases in the tomato trypanosomatid Phytomonas serpens: Influence of growth conditions,...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Cysteine peptidases in the tomato trypanosomatid Phytomonas serpens: Influenceof growth conditions, similarities with cruzipain and secretion to theextracellular environment
Camila G.R. Elias a, Fernanda M. Pereira a, Felipe A. Dias b, Thiago L.A. Silva b, Angela H.C.S. Lopes b,Claudia M. d’Avila-Levy c, Marta H. Branquinha d, André L.S. Santos a,*
a Laboratório de Estudos Integrados em Bioquímica Microbiana, Departamento de Microbiologia Geral, Instituto de Microbiologia Prof. Paulo de Góes (IMPPG), Centro de Ciências daSaúde (CCS), Bloco I-subsolo, Universidade Federal do Rio de Janeiro (UFRJ), Av. Carlos Chagas Filho, 373, Cidade Universitária, Rio de Janeiro, RJ 21941-902, Brazilb Laboratório de Bioquímica de Microrganismos, Departamento de Microbiologia Geral, IMPPG, CCS, UFRJ, Brazilc Laboratório de Biologia Molecular e Doenças Endêmicas, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, RJ, Brazild Laboratório de Bioquímica de Proteases, Departamento de Microbiologia Geral, IMPPG, CCS, UFRJ, Brazil
a r t i c l e i n f o
Article history:Received 30 June 2008Received in revised form 13 August 2008Accepted 22 August 2008Available online 31 August 2008
Keywords:Cysteine peptidaseCruzipainGrowth conditionsPhytomonas serpensPlant trypanosomatidSecretion
a b s t r a c t
We have characterized the cysteine peptidase production by Phytomonas serpens, a tomato trypanoso-matid. The parasites were cultivated in four distinct media, since growth conditions could modulatethe synthesis of bioactive molecules. The proteolytic profile has not changed qualitatively regardlessthe media, showing two peptidases of 38 and 40 kDa; however, few quantitative changes were observedincluding a drastic reduction (around 70%) on the 40 and 38 kDa peptidase activities when parasites weregrown in yeast extract and liver infusion trypticase medium, respectively, in comparison with parasitescultured in Warren medium. The time-span of growth did not significantly alter the protein and pepti-dase expression. The proteolytic activities were blocked by classical cysteine peptidase inhibitors (E-64, leupeptin, and cystatin), being more active at pH 5.0 and showing complete dependence to reducingagents (dithiothreitol and L-cysteine) for full activity. The cysteine peptidases were able to hydrolyze sev-eral proteinaceous substrates, including salivary gland proteins from Oncopeltus fasciatus, suggestingbroad substrate utilization. By means of agglutination, fluorescence microscopy, flow cytometry andWestern blotting analyses we showed that both cysteine peptidases produced by P. serpens share com-mon epitopes with cruzipain, the major cysteine peptidase of Trypanosoma cruzi. Moreover, our data sug-gest that the 40 kDa cysteine peptidase was located at the P. serpens cell surface, attached to membranedomains via a glycosylphosphatidylinositol anchor. The 40 kDa peptidase was also detected in the cell-free culture supernatant, in an active form, which suggests secretion of this peptidase to the extracellularenvironment.
� 2008 Elsevier Inc. All rights reserved.
1. Introduction
Peptidases represent a very important group of proteins withgreat medical and biotechnological applications capable in cleav-ing peptide bonds in protein and peptides (Barrett et al., 2001).Cysteine peptidases are widely distributed in nature and can becompartmentalized in vesicles of the endo-lysosomal system andmay conceivably play a role in protein catabolism and/or promotethe processing of selective polypeptide precursors (Sajid andMcKerrow, 2002; Vermelho et al., 2007). Cysteine peptidases playa key role in the penetration of parasites into host cells, participatein the parasite nutrition at the expense of the host, and are in-volved in the escape mechanisms of the parasite from the host’s
immune system. In accordance, cysteine peptidases appear to berelevant to several aspects of the Trypanosoma, Leishmania andPlasmodium life cycles and are implicated in parasite–host relation-ships. In fact, cysteine peptidase inhibitors block Trypanosoma,Leishmania and Plasmodium replication and differentiation, provid-ing an alternative to the traditional therapy against parasites(Cazzulo et al., 2001; Rosenthal et al., 2002; Sajid and McKerrow,2002; Mottram et al., 2004; Vermelho et al., 2007). Similarly, cys-teine peptidase inhibitors block some essential biological processesin Phytomonas serpens, including cellular growth and interactionwith the invertebrate vector (Santos et al., 2006a).
The discovery of protists belonging to the Trypanosomatidaefamily parasitizing plants occurred in 1909, when Alexander Lafontfound single flagellated parasites in Euphorbia pilulifera (Lafont,1909). In the 1970s, the unearthing of trypanosomatids causinginjuries to plants and the economic losses associated to those inju-
0014-4894/$ - see front matter � 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.exppara.2008.08.011
* Corresponding author.E-mail address: [email protected] (A.L.S. Santos).
Experimental Parasitology 120 (2008) 343–352
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/ locate/yexpr

Author's personal copy
ries changed the scenario and specialists started to draw theirattention to the subject (Parthasarathy, 1976; Dollet et al., 1977;Dollet and Wallace, 1987). Currently it is known that these para-sites are able to parasitize a diverse range of plants (cassava, coco-nut, oil palm, coffee, Alpinia purpurata, Euphorbes and Asclepiades)in Latin America, the Caribbean, Africa, Europe and India causingserious diseases, such as hartrot and marchitez, which in manycases lead the cultivate to death (Dollet, 1984; Camargo, 1999).P. serpens is an important plant flagellate capable of causing dam-age in tomatoes (Gibbs, 1957, 1984). Phytomonads are responsiblefor significant economical losses in agriculture, but they also repre-sent a biological model, since these trypanosomatids present met-abolic routes similar to those of other trypanosomatids pathogenicto humans and animals. Indeed, P. serpens parasites share antigenswith pathogenic species, such as T. cruzi and Leishmania spp (re-viewed by Santos et al. (2007)).
The present study aims the identification and partial charac-terization of proteolytic enzymes from P. serpens promastigotecells. The influence of the medium composition and the time-span of growth on the peptidase expression were also reported.In addition, we have assessed, by antibody cross-reactivity, therelationship of the P. serpens proteolytic enzymes to cruzipain,the major peptidase synthesized by T. cruzi that is a well-de-scribed virulence factor for this parasite. We also demonstratedthat cruzipain-like molecules were able to cleave several protein-aceous substrates, including salivary gland polypeptides fromOncopeltus fasciatus, an insect phytophagous. Lastly, we showedthat the 40 kDa peptidase was extracellularly released by P. ser-pens cells.
2. Materials and methods
2.1. Chemicals
Media constituents, reagents used in electrophoresis, buffercomponents, nitrocellulose membrane and reagents for chemilu-minescence detection (ECL kit) were obtained from Amersham LifeScience (Little Chalfont, England). Sulfosuccinimidyl-6-(biotina-mide) hexanoate (Sulfo-NHS-LC-biotin) and avidin–peroxidasewere products of Pierce Chemical Co. (Rockford, IL, USA). Lowand high molecular mass standards and fetal bovine serum (FBS)were acquired from Gibco BRL (Grand Island, NY, USA). Bacillusthuringiensis phospholipase C (PLC), trypsin, dithiothreitol (DTT),L-cysteine, all protein substrates (gelatin, hemoglobin, casein, bo-vine serum albumin [BSA], non-immune immunoglobulin G andmucin), all proteolytic inhibitors (cystatin, leupeptin, trans-epox-ysuccinyl L-leucylamido-(4-guanidino) butane [E-64], phenylm-ethylsulphonyl fluoride (PMSF), N-p-tosyl-L-lysine chloromethylketone [TLCK], antipain, pepstatin A, ethylenediamine tetraaceticacid [EDTA], ethylene glycol-bis (b-aminoethyl) ether [EGTA] and1,10-phenanthroline) and the secondary antibodies were pur-chased from Sigma Chemical Co. (St Louis, MO, USA). All other re-agents were analytical grade.
2.2. Parasites and cultivation
Phytomonas serpens (isolate 9T; CT-IOC-189), isolated from to-mato (Lycopersicon esculentum), was provided by the Coleção deTripanossomatídeos, Instituto Oswaldo Cruz, Rio de Janeiro, Brazil.The plant flagellate was grown and maintained by weekly transfersin 50-ml Erlenmeyer flasks containing 20 ml of four different cul-ture media: yeast extract—YE (YE, 5 g/l; NaCl, 5 g/l), brain heartinfusion—BHI (BHI, 3.5 g/l; casein, 10 g/l; peptone, 12 g/l; YE, 2 g/l, dextrose, 2 g/l; NaCl, 2 g/l), liver infusion trypticase—LIT (KCl,0.4 g/l; NaCl, 4 g/l; Na2HPO4, 8 g/l; glucose, 2 g/l; tryptone, 5 g/l; li-ver infusion, 5 g/l; YE, 15 g/l) and Warren (BHI, 37 g/l; folic acid,
10 lg/l; hemin, 1 mg/l). All culture media were supplemented with10% FBS. The flasks were inoculated with a 48 h culture containing1 � 106 cells. The parasites were grown at 26 �C up to 96 h. The fla-gellate Trypanosoma cruzi (Dm28c strain) is part of our laboratorycollection, and epimastigote forms were cultured in LIT mediumsupplemented with 10% FBS, for 72 h at 26 �C. Cellular growthwas estimated by counting the parasites in a Neubauerchamber. The cellular viability was monitored through parasitemotility.
2.3. Insects
Adult insects (Oncopeltus fasciatus, Hemiptera: Lygaeidae) werekindly provided by Dr Patrícia Azambuja Penna (Instituto OswaldoCruz, Rio de Janeiro, Brazil). The insects were kept at room temper-ature in plastic pitchers and fed with peeled and toasted sunflowerseeds and distilled water (Romeiro et al., 2003). Salivary glandswere carefully dissected, rinsed three times in cold phosphate-buf-fered saline (PBS; 150 mM NaCl, 20 mM phosphate buffer, pH 7.2)at 4 �C and immediately used for the experiments.
2.4. Parasite extracts
The parasites (1 � 108 cells) were harvested by centrifugationfor 5 min at 1500g at 4 �C, and washed three times with coldPBS. The cells were resuspended in 100 ll of PBS and lysed bythe addition of 1% SDS. The cells were broken in a vortex by alter-nating 1 min shaking and 2 min cooling intervals, followed by acentrifugation at 5000g for 15 min at 4 �C. The supernatants ob-tained after centrifugation corresponded to the whole parasite cel-lular extracts (Santos et al., 2006a). Alternatively, parasites(3 � 108 cells) were extracted with 500 ll of 2% precondensed Tri-ton X-114 in TBS (150 mM NaCl; 10 mM Tris, pH 7.4) as describedby Bordier (1981), to obtain the membrane-rich fractions.
2.5. Salivary gland extract
To obtain the soluble salivary gland extract, 20 insect glandswere resuspended in 500 ll of PBS, supplemented with a proteo-lytic inhibitor cocktail (100 lM E-64, 10 lM 1,10-phenanthroline,10 lM pepstatin A and 1 mM PMSF), homogenized with a Teflon-coated microtissue grinder and vigorously vortexed (three timesfor 30 s) in the presence of 1% SDS, followed by centrifugation at5000g for 10 min at 4 �C. The supernatant obtained after centrifu-gation corresponded to the insect salivary gland extract and con-tained approximately 1 lg protein per ll. Then, the extract washeated for 5 min at 100 �C, alternating 10 min cooling intervals(three times), to abolish the endogenous proteolytic activity (d’Avi-la-Levy et al., 2006a, 2006b).
2.6. Culture supernatant extract
The spent culture medium from 1 � 1010 cells, obtained after P.serpens growth in Warren medium for 48 h, was centrifuged,passed over a 0.22 lm filtration membrane unit (Millipore), andthen concentrated 100-fold in a 10,000 molecular mass cut-offAmicon microconcentrator (AMICON, Beverly, MA, USA). The samevolume of fresh Warren medium was also concentrated and usedas a control to check for possible proteolytic activity. In order toascertain that lysis of promastigotes did not occur during growthunder these conditions and that released peptidases were not anartefact resulting from uncontrolled cell lysis, the presence of lac-tate dehydrogenase activity, an intracellular cytoplasmatic en-zyme, was measured in the cell-free culture supernatant aspreviously described (Elias et al., 2006), presenting negativeresults.
344 C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352

Author's personal copy
2.7. Protein analysis
Protein concentration was determined by the method describedby Lowry et al. (1951), using BSA as standard. Samples containing50 lg of protein were resuspended in SDS–PAGE sample buffer(125 mM Tris, pH 6.8, 4% SDS, 20% glycerol, 0.002% bromophenolblue) and mixed with 5% b-mercaptoethanol, followed by heatingat 100 �C for 5 min. Proteins were analyzed in 12% SDS–PAGE. Elec-trophoresis was carried out at 4 �C, at 120 V, and the gels werestained with 0.2% Coomassie brilliant blue R-250 in methanol–ace-tic acid–water (50:10:40) and destained in the same solvent.Molecular mass of sample polypeptides was calculated by compar-ison with the mobility of molecular mass standards.
2.8. Substrate-SDS–PAGE
Peptidases were assayed and characterized by electrophoresison 10% SDS–PAGE with 0.1% co-polymerized gelatin as substrate(Santos et al., 2005). The gels were loaded with 50 lg of proteinper slot. After electrophoresis, at a constant voltage of 120 V at4 �C, SDS was removed by incubation with 10 volumes of 1% TritonX-100 for 1 h at room temperature under constant agitation. Then,the gels were incubated in 50 mM sodium phosphate buffer sup-plemented with 2 mM DTT, pH 5.0, for 48 h, to promote the prote-olysis. The gels were stained for 2 h with 0.2% Coomassie brilliantblue R-250 in methanol–acetic acid–water (50:10:40) and de-stained overnight in a solution containing methanol–acetic acid–water (5:10:85), to intensify the digestion halos (Santos et al.,2005). The molecular mass of the peptidases was calculated bycomparison with the mobility of low molecular mass standards.The gels were dried, scanned and the densitometric analysis wasperformed with the use of the Kodak Digital Science EDAS 120 soft-ware (Elias et al., 2006).
2.9. Effect of pH and reducing agents on the proteolytic activities
The effect of pH was determined by incubating the gel strips un-der standard assay conditions described above at 37 �C for 48 h inthe following buffers: 10 mM sodium citrate (pH 3.0), 50 mM so-dium phosphate (pH 5.0 and 7.0) and 20 mM glycine–NaOH (pH9.0). All buffers were supplemented with 2 mM DTT. Alternatively,the requirement of reducing agent was determined by incubatingthe gel strips in 50 mM sodium phosphate buffer, pH 5.0, for48 h at 37 �C, in the absence (control) or in the presence of DTT,L-cysteine or b-mercaptoethanol at 2 mM (Elias et al., 2006).
2.10. Effect of different proteinaceous substrates on the proteolyticactivities
The ability of the peptidases to degrade different proteinaceoussubstrates was also evaluated by 10% SDS–PAGE co-polymerizedwith 0.1% casein, hemoglobin, IgG, BSA, mucin or 100 lg of salivarygland extract from O. fasciatus. Electrophoresis was performed asdescribed before and gel strips were incubated in 50 mM sodiumphosphate buffer, pH 5.0, supplemented with 2 mM DTT for 48 hat 37 �C (Elias et al., 2006).
2.11. Degradation of salivary gland surface proteins by live P. serpenscells
In this set of experiments, initially the salivary glands from O. fas-ciatus were dissected, washed twice in PBS and then biotinylated for30 min at 4 �C through the incubation with 0.1 mg/ml Sulfo-NHS-LC-biotin (Santos et al., 2001; Dias et al., 2007). Afterwards, glandswere washed five times in PBS to remove non-reacted biotin. In par-allel, live P. serpens promastigotes (1 � 108 cells) were incubated for
30 min at 26 �C in the presence or absence of 10 lM E-64. As previ-ously reported, the parasites maintained their viability after E-64treatment as judged by their unaltered morphology, mobility, try-pan blue dye exclusion (in which more than 99% of the parasiteswere viable) (Santos et al., 2006a). Finally, the untreated and prote-olytic inhibitor-treated parasites were incubated for 4 h at 26 �Cwith the biotinylated salivary glands (three glands for each system).After incubation, the glands were added to SDS–PAGE sample bufferand mixed with 10% b-mercaptoethanol, followed by heating at100 �C for 5 min. Thereafter, the polypeptides were separated in12% SDS–PAGE and electrophoretically transferred at 100 V/300 mA for 1 h at 4 �C to a nitrocellulose membrane. The membranewas blocked with 2% BSA in TBS containing 0.05% Tween 20 (TBS/Tween) for 1 h at room temperature. Then, membranes were washedthree times (10 min each) with the blocking solution and incubatedwith avidin–peroxidase at 1:10,000 dilution in TBS/Tween for 1 h.The reactive polypeptides were detected by chemiluminescenceafter reaction with ECL reagents.
2.12. Quantitative assay for detection of secreted proteolytic activity
Extracellular proteolytic activity was also measured spectro-photometrically using the substrate gelatin, according to the meth-od described by Buroker-Kilgore and Wang (1993). Briefly,concentrated supernatant was incubated for 30 min at 37 �C inthe presence or absence of the following proteolytic inhibitors:1 mM 1,10-phenanthroline, 1 mM EDTA, 1 mM EGTA, 1 lM pepsta-tin A, 1 mM PMSF, 1 mM TLCK, 1 lM antipain, 1 lM cystatin, 1 lMleupeptin and 1 lM E-64. For assaying the remaining proteolyticactivity, gelatin (0.5 mg/ml), 2 mM DTT, 50 mM sodium phosphate(pH 5.0) and 20 ll of the supernatant were added to a microcentri-fuge tube (350 ll) and incubated for 2 h at 37 �C. After this incuba-tion, three aliquots (100 ll each) of the reaction mixture weretransferred to wells on a microtiter plate containing 50 ll of waterand 100 ll of a Coomassie solution (0.025% Coomassie brilliantblue G-250, 11.75% ethanol, and 21.25% phosphoric acid). After10 min to allow dye binding, the plate was read on a ThermomaxMolecular Device microplate reader (sensitivity: 0.0001) at anabsorbance of 595 nm. One unit of proteolytic activity was definedas the amount of enzyme that caused an increase of 0.001 in theabsorbance unit, under standard assay conditions. The results areexpressed as relative percentage of activity with inhibitors sub-tracted from the activity without inhibitors.
2.13. Agglutination test
The agglutination assay was performed by incubating 100 ll ofa suspension containing 1 � 108 parasites/ml with different dilu-tions (1:100, 1:250, 1:500, 1:1000 and 1:2500) of rabbit anti-cruzi-pain polyclonal antibody raised against T. cruzi (kindly provided byDr Juan-Jose Cazzulo, Instituto de Investigaciones Biotecnologicas,Universidad Nacional de General San Martin, Buenos Aires, Argen-tina). The agglutination was evaluated after 1 h at room tempera-ture by comparison with untreated parasites observed in a ZeissAxiovert light inverted microscope.
2.14. Phospholipase C and trypsin treatments
Parasites (1 � 107 cells) were fixed in 0.4% paraformaldehydediluted in PBS for 30 min at room temperature, followed by exten-sive washing in the same buffer. These fixed cells maintained theirmorphological integrity, as verified by optical microscopic obser-vation. Cells (5 � 106) were incubated with 0.2 U/l of PLC in a finalreaction mixture (400 ll) containing 100 mM Tris–acetate, pH 7.4,supplemented with 0.1% Triton X-100 for 4 h at 37 �C (Santos et al.,2002). Alternatively, the parasites were incubated with 1 mg/ml of
C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352 345
Author's personal copy
trypsin for 2 h at 37 �C. Control cells were subjected to the sameexperimental conditions, except for the presence of PLC or trypsin.
2.15. Flow cytometry and fluorescence microscopy
Parasites (5 � 106 cells) previously treated or not with PLC ortrypsin were incubated for 1 h with a 1:250 dilution of anti-cruzi-pain antibody, and then incubated for an additional hour with a1:250 dilution of fluorescein isothiocyanate (FITC)-labeled goatanti-rabbit IgG. These cells were washed three times in PBS and ob-served in a Zeiss epifluorescence microscope (Axioplan). For flowcytometry analysis, these cells were examined in an EPICS ELITEflow cytometer (Coulter Electronics, Hialeah, Fla.) equipped witha 15 mW argon laser emitting at 488 nm. Untreated cells and thosetreated only with the secondary antibody were used as controls.Each experimental population was then mapped by using a two-parameter histogram of forward-angle light scatter versus sidescatter. The mapped population (n = 10,000) was then analyzedfor log green fluorescence by using a single parameter histogram(Santos et al., 2006a).
2.16. Assay for cruzipain-like polypeptides
Parasite extracts (cell-associated and concentrated culturesupernatant), equivalent to 50 lg of proteins, were separated in12% SDS–PAGE and electrophoretically transferred to a nitrocellu-lose membrane, which was blocked and washed as describedabove. Then, membranes were incubated with the anti-cruzipainantibody at 1:250 dilutions for 2 h. The secondary antibody usedwas peroxidase-conjugated goat anti-rabbit IgG at 1:2500. Immu-noblots were exposed to X-ray film after reaction with ECL re-agents for chemiluminescence (Santos et al. 2006a).
2.17. Statistical analysis
The experiments were performed in triplicate, in three indepen-dent experimental sets. The data were analyzed statistically bymeans of Student’s t-test using EPI-INFO 6.04 (Database and Statis-tics Program for Public Health) computer software. P values of 0.05or less were considered statistically significant.
3. Results
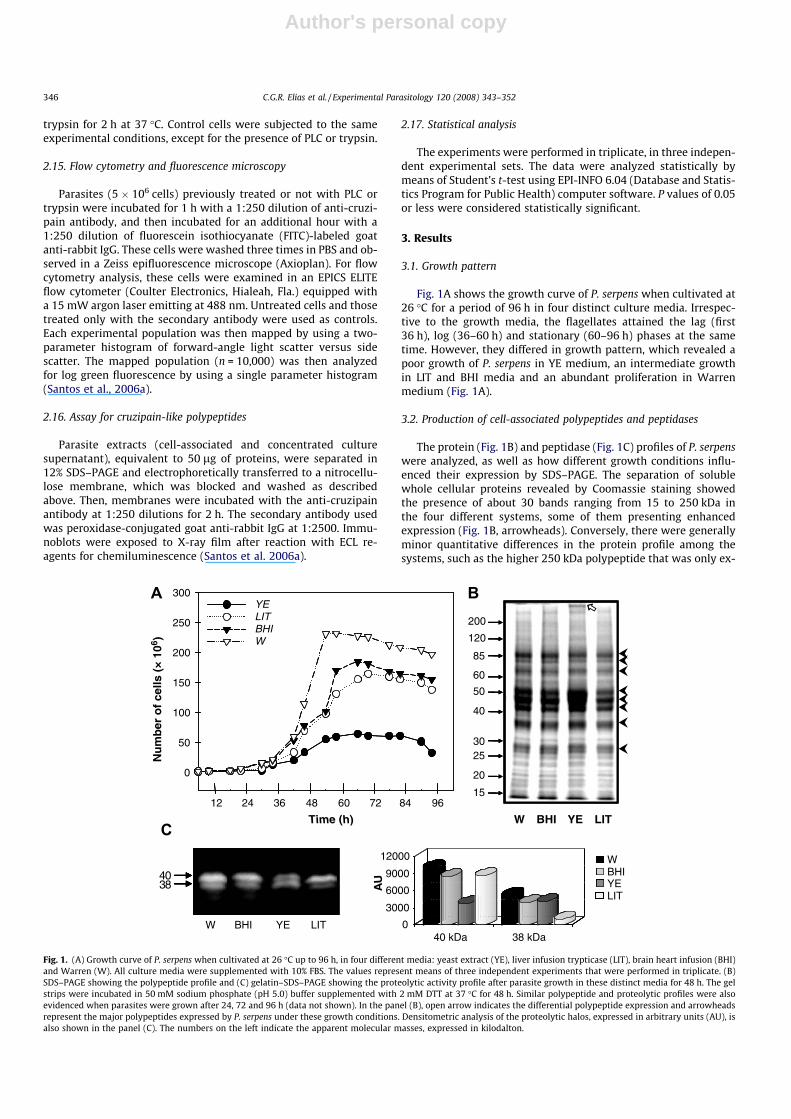
3.1. Growth pattern
Fig. 1A shows the growth curve of P. serpens when cultivated at26 �C for a period of 96 h in four distinct culture media. Irrespec-tive to the growth media, the flagellates attained the lag (first36 h), log (36–60 h) and stationary (60–96 h) phases at the sametime. However, they differed in growth pattern, which revealed apoor growth of P. serpens in YE medium, an intermediate growthin LIT and BHI media and an abundant proliferation in Warrenmedium (Fig. 1A).
3.2. Production of cell-associated polypeptides and peptidases
The protein (Fig. 1B) and peptidase (Fig. 1C) profiles of P. serpenswere analyzed, as well as how different growth conditions influ-enced their expression by SDS–PAGE. The separation of solublewhole cellular proteins revealed by Coomassie staining showedthe presence of about 30 bands ranging from 15 to 250 kDa inthe four different systems, some of them presenting enhancedexpression (Fig. 1B, arrowheads). Conversely, there were generallyminor quantitative differences in the protein profile among thesystems, such as the higher 250 kDa polypeptide that was only ex-
Time (h)
200
120
85
60
50
40
3025
20
15
W BHI YE LIT
0
50
100
150
200
250
300YELITBHIW
482412 60
4038
W BHI YE LIT 040 kDa
3000
6000
9000
12000
38 kDa
WBHIYELIT
AU
36 72 84 96
Nu
mb
er o
f ce
lls (
× 1
06 )
Fig. 1. (A) Growth curve of P. serpens when cultivated at 26 �C up to 96 h, in four different media: yeast extract (YE), liver infusion trypticase (LIT), brain heart infusion (BHI)and Warren (W). All culture media were supplemented with 10% FBS. The values represent means of three independent experiments that were performed in triplicate. (B)SDS–PAGE showing the polypeptide profile and (C) gelatin–SDS–PAGE showing the proteolytic activity profile after parasite growth in these distinct media for 48 h. The gelstrips were incubated in 50 mM sodium phosphate (pH 5.0) buffer supplemented with 2 mM DTT at 37 �C for 48 h. Similar polypeptide and proteolytic profiles were alsoevidenced when parasites were grown after 24, 72 and 96 h (data not shown). In the panel (B), open arrow indicates the differential polypeptide expression and arrowheadsrepresent the major polypeptides expressed by P. serpens under these growth conditions. Densitometric analysis of the proteolytic halos, expressed in arbitrary units (AU), isalso shown in the panel (C). The numbers on the left indicate the apparent molecular masses, expressed in kilodalton.
346 C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352
Author's personal copy
pressed in parasites cultured in yeast extract medium (Fig. 1B,open arrow in lane YE).
As well known, trypanosomatids produce mainly metallo andcysteine peptidases (Branquinha et al., 1996; Santos et al., 2005).In addition, acidic pH added with a reducing agent is currentlyused to evidence these peptidase classes. With these consider-ations in mind, we evaluated the cell-associated peptidase ex-pressed by P. serpens using these experimental conditions. Ourresults showed that the proteolytic profiles were also similar inthe four distinct media, being composed of two major peptidasesof 38 and 40 kDa (Fig. 1C); however, few quantitative changes wereobserved including a drastic reduction (around 70%) on the 40 and38 kDa peptidase activities when parasites were grown in YE andLIT medium, respectively, in comparison with parasites culturedin Warren medium (Fig. 1C, graphic). Interestingly, the analysis
of protein and peptidase expression was not altered during the96 h of in vitro growth (data not shown). Taking it in consideration,parasites grown in Warren medium for 48 h (mid-exponentialphase) were chosen to carry out further analyses.
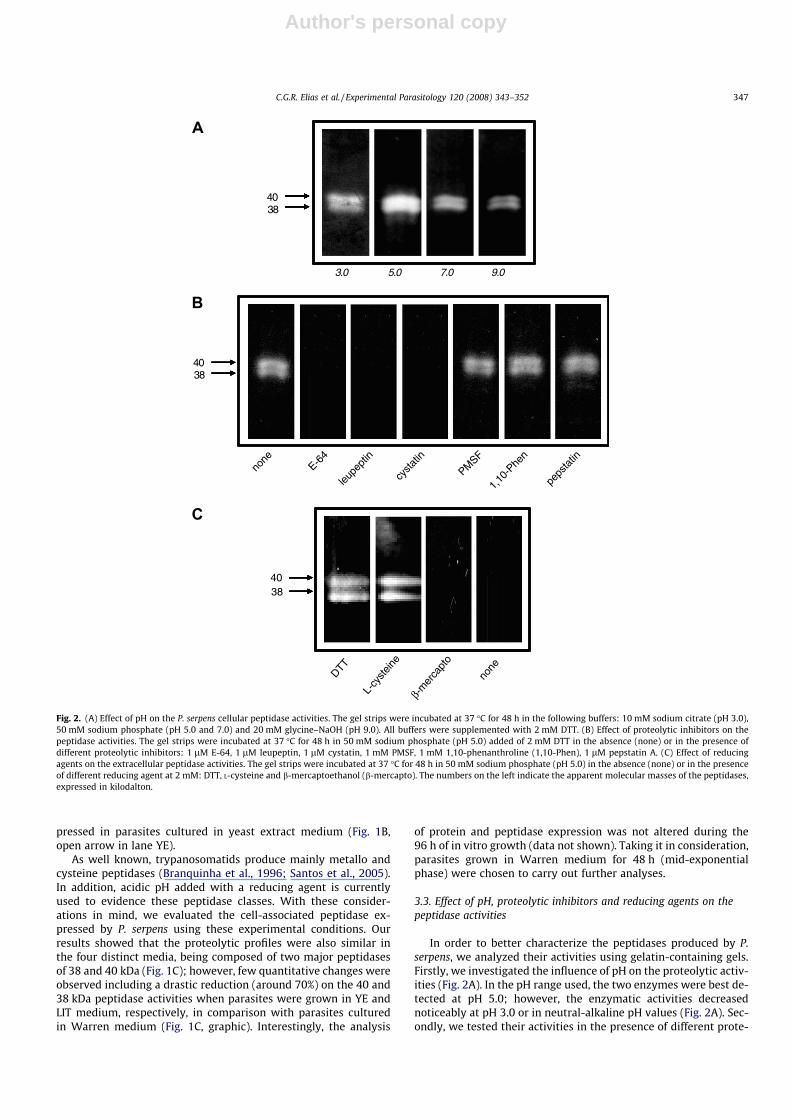
3.3. Effect of pH, proteolytic inhibitors and reducing agents on thepeptidase activities
In order to better characterize the peptidases produced by P.serpens, we analyzed their activities using gelatin-containing gels.Firstly, we investigated the influence of pH on the proteolytic activ-ities (Fig. 2A). In the pH range used, the two enzymes were best de-tected at pH 5.0; however, the enzymatic activities decreasednoticeably at pH 3.0 or in neutral-alkaline pH values (Fig. 2A). Sec-ondly, we tested their activities in the presence of different prote-
40
3.0 5.0 7.0 9.0
3840
none
E-64
leupe
ptin
cysta
tin
PMSF
1,10
-Phe
n
peps
tatin
38
DTT
L-cy
steine
β-m
erca
pto
none
3840
Fig. 2. (A) Effect of pH on the P. serpens cellular peptidase activities. The gel strips were incubated at 37 �C for 48 h in the following buffers: 10 mM sodium citrate (pH 3.0),50 mM sodium phosphate (pH 5.0 and 7.0) and 20 mM glycine–NaOH (pH 9.0). All buffers were supplemented with 2 mM DTT. (B) Effect of proteolytic inhibitors on thepeptidase activities. The gel strips were incubated at 37 �C for 48 h in 50 mM sodium phosphate (pH 5.0) added of 2 mM DTT in the absence (none) or in the presence ofdifferent proteolytic inhibitors: 1 lM E-64, 1 lM leupeptin, 1 lM cystatin, 1 mM PMSF, 1 mM 1,10-phenanthroline (1,10-Phen), 1 lM pepstatin A. (C) Effect of reducingagents on the extracellular peptidase activities. The gel strips were incubated at 37 �C for 48 h in 50 mM sodium phosphate (pH 5.0) in the absence (none) or in the presenceof different reducing agent at 2 mM: DTT, L-cysteine and b-mercaptoethanol (b-mercapto). The numbers on the left indicate the apparent molecular masses of the peptidases,expressed in kilodalton.
C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352 347
Author's personal copy
olytic inhibitors (Fig. 2B). The two proteolytic enzymes synthesizedby P. serpens exhibited total sensibility to E-64, leupeptin and cyst-atin, three well-known cysteine peptidase inhibitors. Conversely,PMSF (a serine peptidase inhibitor), 1,10-phenanthroline (a metal-lopeptidase inhibitor) and pepstatin A (an aspartyl peptidaseinhibitor) did not promote any significant alterations in the behav-ior of the 38 and 40 kDa proteolytic activities (Fig. 2B). Since cys-teine peptidases require a reducing agent for their full activity,we checked the influence of these compounds in the P. serpens en-zymes (Fig. 2C). The activities of cysteine peptidases were abso-lutely dependent on the presence of a reducing agent, sincesupplementation of the digesting buffer with DTT or L-cysteinecompletely restored the proteolytic activities when compared withnon-supplemented buffer (Fig. 2C). b-mercaptoethanol did not re-store the peptidase activities (Fig. 2C).
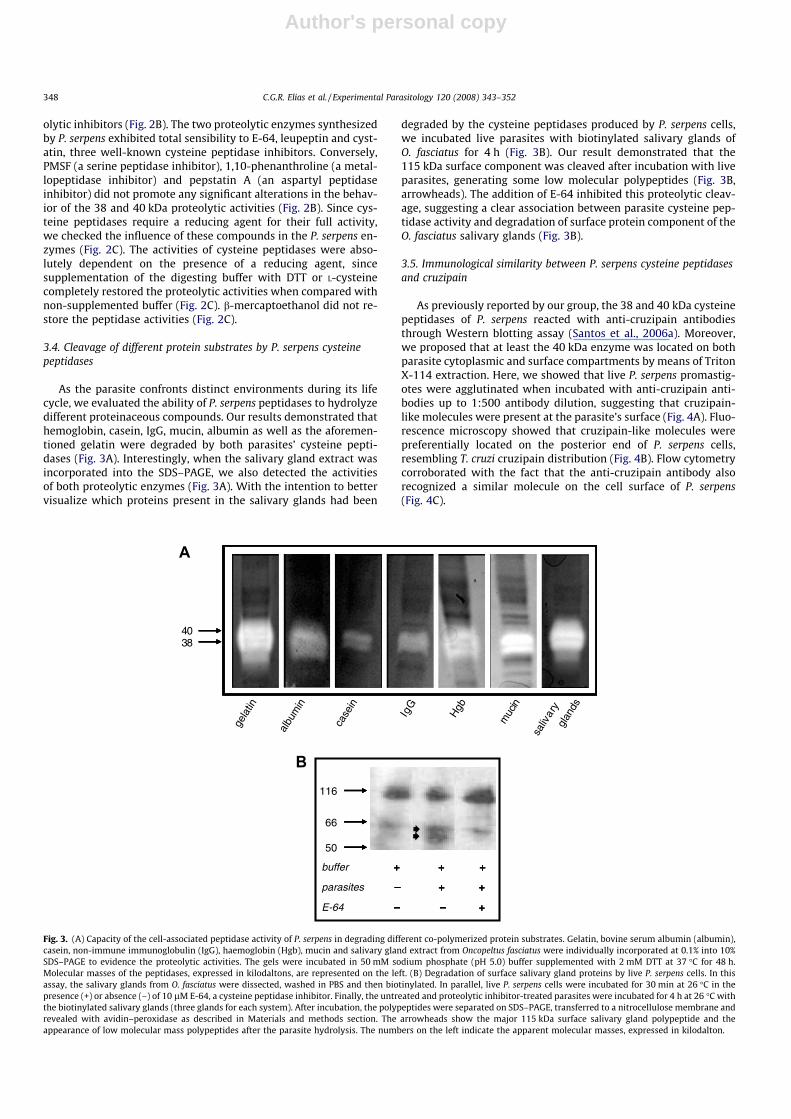
3.4. Cleavage of different protein substrates by P. serpens cysteinepeptidases
As the parasite confronts distinct environments during its lifecycle, we evaluated the ability of P. serpens peptidases to hydrolyzedifferent proteinaceous compounds. Our results demonstrated thathemoglobin, casein, IgG, mucin, albumin as well as the aforemen-tioned gelatin were degraded by both parasites’ cysteine pepti-dases (Fig. 3A). Interestingly, when the salivary gland extract wasincorporated into the SDS–PAGE, we also detected the activitiesof both proteolytic enzymes (Fig. 3A). With the intention to bettervisualize which proteins present in the salivary glands had been
degraded by the cysteine peptidases produced by P. serpens cells,we incubated live parasites with biotinylated salivary glands ofO. fasciatus for 4 h (Fig. 3B). Our result demonstrated that the115 kDa surface component was cleaved after incubation with liveparasites, generating some low molecular polypeptides (Fig. 3B,arrowheads). The addition of E-64 inhibited this proteolytic cleav-age, suggesting a clear association between parasite cysteine pep-tidase activity and degradation of surface protein component of theO. fasciatus salivary glands (Fig. 3B).
3.5. Immunological similarity between P. serpens cysteine peptidasesand cruzipain
As previously reported by our group, the 38 and 40 kDa cysteinepeptidases of P. serpens reacted with anti-cruzipain antibodiesthrough Western blotting assay (Santos et al., 2006a). Moreover,we proposed that at least the 40 kDa enzyme was located on bothparasite cytoplasmic and surface compartments by means of TritonX-114 extraction. Here, we showed that live P. serpens promastig-otes were agglutinated when incubated with anti-cruzipain anti-bodies up to 1:500 antibody dilution, suggesting that cruzipain-like molecules were present at the parasite’s surface (Fig. 4A). Fluo-rescence microscopy showed that cruzipain-like molecules werepreferentially located on the posterior end of P. serpens cells,resembling T. cruzi cruzipain distribution (Fig. 4B). Flow cytometrycorroborated with the fact that the anti-cruzipain antibody alsorecognized a similar molecule on the cell surface of P. serpens(Fig. 4C).
gela
tin
4038
albu
min
case
in
IgG
Hgb
muc
in
saliv
ary
glan
ds
+ + +
– + +
– – +
+ + +
– + +
– – +
+ + +
– + +
– – +
116
66
50
buffer
parasites
E-64
+ + +
– + +
– – +
Fig. 3. (A) Capacity of the cell-associated peptidase activity of P. serpens in degrading different co-polymerized protein substrates. Gelatin, bovine serum albumin (albumin),casein, non-immune immunoglobulin (IgG), haemoglobin (Hgb), mucin and salivary gland extract from Oncopeltus fasciatus were individually incorporated at 0.1% into 10%SDS–PAGE to evidence the proteolytic activities. The gels were incubated in 50 mM sodium phosphate (pH 5.0) buffer supplemented with 2 mM DTT at 37 �C for 48 h.Molecular masses of the peptidases, expressed in kilodaltons, are represented on the left. (B) Degradation of surface salivary gland proteins by live P. serpens cells. In thisassay, the salivary glands from O. fasciatus were dissected, washed in PBS and then biotinylated. In parallel, live P. serpens cells were incubated for 30 min at 26 �C in thepresence (+) or absence (–) of 10 lM E-64, a cysteine peptidase inhibitor. Finally, the untreated and proteolytic inhibitor-treated parasites were incubated for 4 h at 26 �C withthe biotinylated salivary glands (three glands for each system). After incubation, the polypeptides were separated on SDS–PAGE, transferred to a nitrocellulose membrane andrevealed with avidin–peroxidase as described in Materials and methods section. The arrowheads show the major 115 kDa surface salivary gland polypeptide and theappearance of low molecular mass polypeptides after the parasite hydrolysis. The numbers on the left indicate the apparent molecular masses, expressed in kilodalton.
348 C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352

Author's personal copy
3.6. Attachment mode of the cruzipain-like molecules to the parasitesurface
The cell surface of trypanosomatids is coated by a glycocalyxthat comprises a high number of glycosylphosphatidylinositol(GPI)-anchored molecules. Here, we looked for a GPI-anchor inthe cruzipain-like molecules expressed at the surface of P. serpenspromastigote cells. Evidence for the occurrence of GPI-anchoredproteins has been obtained by loss of surface components afterin vitro treatment of intact parasites with PLC (Cardoso de Almeidaand Heise, 1993; Santos et al., 2002; d’Avila-Levy et al., 2006b). Inthis sense, fixed parasites were treated or not with PLC, followedby the incubation with anti-cruzipain antibody and IgG–FITC.Then, the cruzipain-like molecule amount was assessed by flowcytometry. PLC-treated P. serpens showed a reduction on the bind-ing of anti-cruzipain antibody when compared to the non-treatedflagellates (Fig. 4C), suggesting that these molecules are expressedon the cell surface through a GPI anchor. As expected, parasitestreated with trypsin, which is a proteolytic enzyme able to cleavearginine–lysine linkages, presented a powerful reduction on thesurface amount of cuzipain-like molecules (Fig. 4C).
3.7. Secretion of cruzipain-like molecules by P. serpens
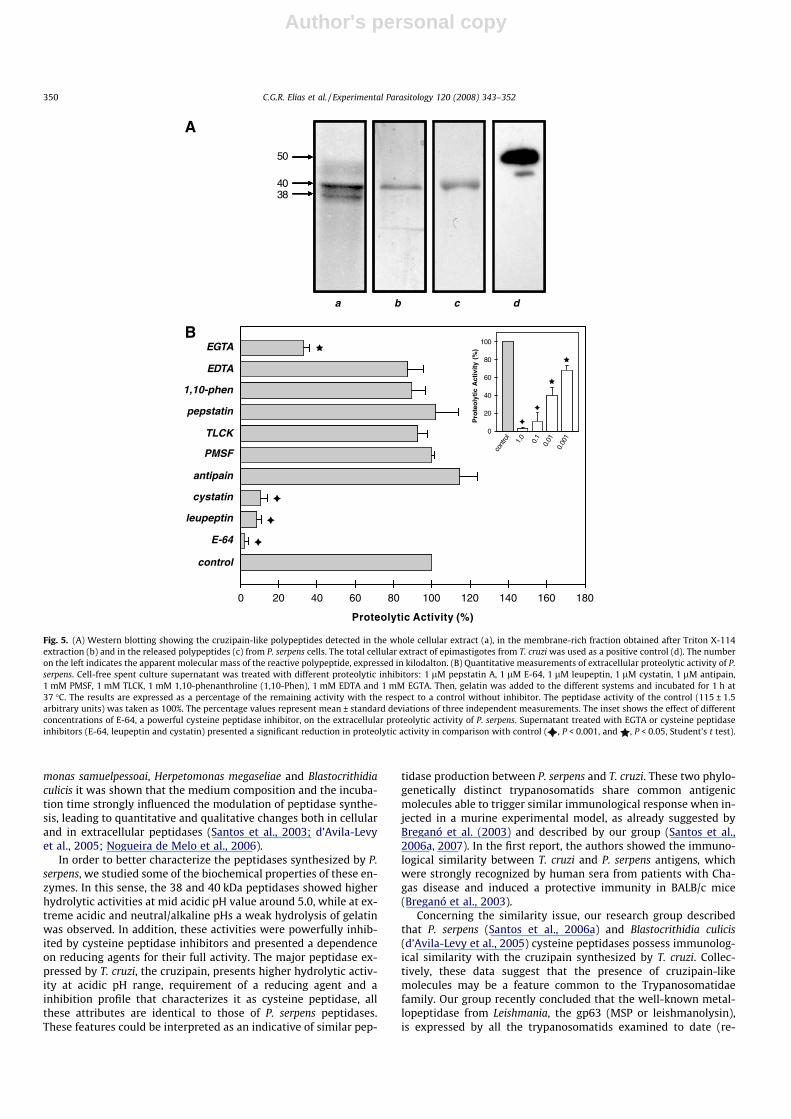
P. serpens promastigote cells, when immunocytochemically la-beled with anti-cruzipain antibodies associated with colloidal goldand subsequently analyzed by transmission electron microscopy,showed gold particles in all the extension of the membrane, coat-ing the cell body and the flagellum, as well as in the cytoplasmicregion (Santos et al., 2006a). Interestingly, gold particles were alsodetected in the flagellar pocket, some of them free or bound tosmall membrane vesicles, suggesting that the cruzipain-like mole-cules could be released and/or shed from the parasite surface (San-tos et al., 2006a). To ascertain the presence of cruzipain-likemolecules in the extracellular environment, we performed a wes-tern blotting assay using the concentrated cell-free culture super-natant obtained after P. serpens growth in vitro (Fig. 5A). Ourresults showed a reactive polypeptide band of 40 kDa that was pre-sented in the total cellular extract of P. serpens, in the membrane-
rich fraction as well as in the culture supernatant extract (Fig. 5A).In this set of experiments, we used the cellular extract from T. cruziepimastigote forms as a positive control (Fig. 5A).
In addition, we evidenced the proteolytic activity in the culturesupernatant of P. serpens by a quantitative measurement (Fig. 5B).To determine the nature of the P. serpens peptidase that was cleav-ing the soluble substrate gelatin, we incubated the supernatant inthe presence of inhibitors of the four major peptidase classes. Theaddition of the cysteine peptidase inhibitors (E-64, leupeptin andcystatin) powerfully inhibited the proteolytic cleavage (Fig. 5B).On the other hand, serine (PMSF and TLCK), metallo (1,10-phenan-throline and EDTA) and aspartic (pepstatin A) peptidase inhibitorsdid not significantly diminish the proteolytic activity (Fig. 5B), withthe exception to EGTA. E-64, a powerful cysteine peptidase inhib-itor, restrained the extracellular proteolytic activity in a concentra-tion-dependent manner (Fig. 5B, inset).
4. Discussion
Trypanosomatids orchestrate extensive changes in cellularorganization and metabolism to obtain the nutrients they need.Our understanding of the molecular exchanges in P. serpens is, asyet, rudimentary. However, some adaptive forms, especially inthe insects, use amino acids present in their host for energy pro-duction. Proteolytic enzymes might participate in acquiring aminoacids for the trypanosomatid cells, by efficiently hydrolyzing pro-tein substrates. However, the production of peptidases by trypan-osomatids is closely related to the main conditions determiningculture growth and development: first and foremost, the composi-tion of the nutritive medium (Nogueira de Melo et al., 2002, 2006;d’Avila-Levy et al., 2005). In the present study we showed that,irrespective to the culture medium used and the growth phaseanalyzed, P. serpens produced the same cellular peptidase profile,composed of two distinct activities of 38 and 40 kDa. However,some quantitative differences on both peptidases expression werenoticed as judged by densitometric analysis of the proteolyticbands after SDS–PAGE analysis. Similarly, Crithidia guilhermeigrown in four distinct media did not alter its proteolytic enzymeproduction (Nogueira de Melo et al., 2002). Conversely, in Herpeto-
Fig. 4. (A) Binding of anti-cruzipain antibodies to the cell surface of P. serpens cells. The live parasites were incubated in the presence of the rabbit pre-immune serum (a) orrabbit anti-cruzipain serum (b) at 1:125 dilution for 1 h. Then, cells were fixed and experimental systems were analyzed under phase contrast images. The arrow indicates theagglutination of parasites by the anti-cruzipain antibody. (B) Phase contrast (a) and fluorescence microscopy (b) images of parasites sequentially incubated with anti-cruzipain and FITC-secondary antibody. Note an intense fluorescence signal on the posterior end of the parasite cellular body. The bars represent 10 lm. (C) Flow cytometryanalyses showing the distribution of cruzipain-like molecules on the parasite cell surface. In this set of experiments, parasites were initially non-treated or treated with 1 mg/ml trypsin or 0.2 U/ll PLC and then incubated in the presence of anti-cruzipain antibody (1:250 dilution) as described in the material and methods, and then analyzed by flowcytometry. Grey curve represents the autofluorescence of P. serpens promastigote cells without any enzymatic or antibody treatment.
C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352 349
Author's personal copy
monas samuelpessoai, Herpetomonas megaseliae and Blastocrithidiaculicis it was shown that the medium composition and the incuba-tion time strongly influenced the modulation of peptidase synthe-sis, leading to quantitative and qualitative changes both in cellularand in extracellular peptidases (Santos et al., 2003; d’Avila-Levyet al., 2005; Nogueira de Melo et al., 2006).
In order to better characterize the peptidases synthesized by P.serpens, we studied some of the biochemical properties of these en-zymes. In this sense, the 38 and 40 kDa peptidases showed higherhydrolytic activities at mid acidic pH value around 5.0, while at ex-treme acidic and neutral/alkaline pHs a weak hydrolysis of gelatinwas observed. In addition, these activities were powerfully inhib-ited by cysteine peptidase inhibitors and presented a dependenceon reducing agents for their full activity. The major peptidase ex-pressed by T. cruzi, the cruzipain, presents higher hydrolytic activ-ity at acidic pH range, requirement of a reducing agent and ainhibition profile that characterizes it as cysteine peptidase, allthese attributes are identical to those of P. serpens peptidases.These features could be interpreted as an indicative of similar pep-
tidase production between P. serpens and T. cruzi. These two phylo-genetically distinct trypanosomatids share common antigenicmolecules able to trigger similar immunological response when in-jected in a murine experimental model, as already suggested byBreganó et al. (2003) and described by our group (Santos et al.,2006a, 2007). In the first report, the authors showed the immuno-logical similarity between T. cruzi and P. serpens antigens, whichwere strongly recognized by human sera from patients with Cha-gas disease and induced a protective immunity in BALB/c mice(Breganó et al., 2003).
Concerning the similarity issue, our research group describedthat P. serpens (Santos et al., 2006a) and Blastocrithidia culicis(d’Avila-Levy et al., 2005) cysteine peptidases possess immunolog-ical similarity with the cruzipain synthesized by T. cruzi. Collec-tively, these data suggest that the presence of cruzipain-likemolecules may be a feature common to the Trypanosomatidaefamily. Our group recently concluded that the well-known metal-lopeptidase from Leishmania, the gp63 (MSP or leishmanolysin),is expressed by all the trypanosomatids examined to date (re-
0
20
40
60
80
100
0
20
40
60
80
100
a b c d
4038
50
0
20
40
60
80
100
0 20 40 60 80 100 120 140 160 180
control
E-64
leupeptin
cystatin
antipain
PMSF
TLCK
pepstatin
1,10-phen
EDTA
EGTA
0
20
40
60
80
100
cont
rol
1.0
0.1
0.01
0.00
1
Proteolytic Activity (%)
Pro
teo
lyti
c A
ctiv
ity
(%)
Fig. 5. (A) Western blotting showing the cruzipain-like polypeptides detected in the whole cellular extract (a), in the membrane-rich fraction obtained after Triton X-114extraction (b) and in the released polypeptides (c) from P. serpens cells. The total cellular extract of epimastigotes from T. cruzi was used as a positive control (d). The numberon the left indicates the apparent molecular mass of the reactive polypeptide, expressed in kilodalton. (B) Quantitative measurements of extracellular proteolytic activity of P.serpens. Cell-free spent culture supernatant was treated with different proteolytic inhibitors: 1 lM pepstatin A, 1 lM E-64, 1 lM leupeptin, 1 lM cystatin, 1 lM antipain,1 mM PMSF, 1 mM TLCK, 1 mM 1,10-phenanthroline (1,10-Phen), 1 mM EDTA and 1 mM EGTA. Then, gelatin was added to the different systems and incubated for 1 h at37 �C. The results are expressed as a percentage of the remaining activity with the respect to a control without inhibitor. The peptidase activity of the control (115 ± 1.5arbitrary units) was taken as 100%. The percentage values represent mean ± standard deviations of three independent measurements. The inset shows the effect of differentconcentrations of E-64, a powerful cysteine peptidase inhibitor, on the extracellular proteolytic activity of P. serpens. Supernatant treated with EGTA or cysteine peptidaseinhibitors (E-64, leupeptin and cystatin) presented a significant reduction in proteolytic activity in comparison with control ( , P < 0.001, and , P < 0.05, Student’s t test).
350 C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352

Author's personal copy
viewed by Santos et al., 2006b), including T. cruzi (Cuevas et al.,2003). So far, B. culicis and P. serpens, that are the only trypanoso-matids screened for the presence of cruzipain-like molecules up tonow, showed to possess this molecule. Further efforts should be di-rected to screen distinct trypanosomatids in order to evaluate thedistribution of cruzipain-like molecules in this family. We mayspeculate that cruzipain is subjected to the selective pressures ofsimilar environments (invertebrate host) and it is expected to bestructurally and functionally conserved in distincttrypanosomatids.
The use of proteolytic enzymes to degrade the host barriersseems to be a common strategy used by microorganisms to colo-nize host tissues, cause disease and overcome host defenses. In thiscontext, the cysteine peptidases of P. serpens were able to hydro-lyze a broad spectrum of protein components, including: gelatin,albumin, casein, immunoglobulin, hemoglobin, mucin and O. fasci-atus salivary gland polypeptide extract. The broad hydrolytic abil-ity of P. serpens proteolytic enzymes might help the parasites inobtaining amino acid for its nutrition. Moreover, plants commonlypresent several protection mechanisms against phytopathogenicmicroorganisms, such as production of peptides with antimicrobialaction (e.g. lectin-like molecules) or expression of peptidase inhib-itors (Fritig et al., 1998; Garcia-Olmedo et al., 1998; Malek and Die-trich, 1999). In this sense, plant microorganisms have to escapethese mechanisms in order to act as parasites. The production ofpeptidases is one of the most important apparatus that parasitesdispose for escaping of plant protection; these enzymes are ableto cleave antimicrobial peptides and degrade protein componentsof cell wall (reviewed by Mosolov et al. (2001)). Alternatively, theseenzymes may also help phytomonads to bypass the insect defensemechanisms such as the prophenoloxidase (proPO) system, whichis dependable on activation by insect peptidases (Azambuja et al.,1999), which in turn can be degraded by P. serpens peptidases.Thereby, the broad spectrum of action of P. serpens peptidasesmay improve its ability to break the plant and/or insect barriersof defense. Moreover, this could also mean a degradation of differ-ent substrates in vivo by the same enzyme, which is true for somepeptidases such as cruzipain that is able to degrade gelatin, immu-noglobulin, hemoglobin and casein (Del Nery et al., 1997).
As previously reported by our group, the cellular cysteine pep-tidases of P. serpens share common epitopes with the major cys-teine peptidase from T. cruzi (Santos et al., 2006a). At least the40 kDa peptidase was detected in the intracellular compartmentsas well as in cell surface. These results were corroborated hereinby flow cytometry analysis and by agglutination assay usinganti-cruzipain antibodies. In addition, the surface cruzipain-likemolecules of P. serpens were cleaved by the PLC enzyme, suggest-ing a possible interaction with membrane domains via a GPI an-chor. These features concerning the cysteine peptidaselocalization and membrane interaction through GPI also testifythe similarity between P. serpens and T. cruzi. Aside from the factthat the P. serpens peptidases were recognized by anti-cruzipainantibodies, the cruzipain itself presents equal characteristicsregarding the localization issue: it can be found on the parasitesurface (Souto-Padrón et al., 1990) and it was suggested by Fresnoet al. (1994) a possible connection to the plasma membrane by aGPI anchor.
Few studies have been done in order to identify and characterizereleased/secreted peptidases in trypanosomatids. The secretion ofmolecules to the extracellular environment may be an importantadaptive mechanism during the life cycle of the parasitic protozoan.Metallopeptidases are the main proteolytic class secreted by Leish-mania (Yao et al., 2007) and by all monoxenic trypanosomatids stud-ied so far (reviewed by Santos et al., 2006b), with the exception of C.deanei and C. desouzai, which also releases cysteine peptidases(d’Avila-Levy et al., 2001). Intriguingly, in the present report, we de-
tected a cysteine peptidase activity in the culture supernatant of P.serpens using an in-solution assay with gelatin. This extracellularpeptidase activity has also strongly sensitivity to three distinct cys-teine peptidases inhibitors (E-64, leupeptin and cystatin). Moreover,by means of western blotting we identified in the supernatant ex-tract a 40 kDa polypeptide that reacted with the anti-cruzipain anti-body. This polypeptide was also evidenced in the total cellularextract as well as in the membrane-rich fraction of P. serpens. Theseresults corroborated our previous reports in which cruzipain-likemolecules were immunolocalized in the parasite flagellar pocket,suggesting the release of this molecule to the extracellular environ-ment (Santos et al., 2006a). Inquisitively, it has been reported thatthe trypomastigote forms of T. cruzi are able to secrete cysteine pep-tidases, which includes cruzipain, into the extracellular medium(Yokoyama-Yasunaka et al., 1994).
The cysteine peptidases expressed by P. serpens and the cruzi-pain itself share a great number of common features. Therefore,it is feasible to suggest some functions to the enzymes expressedand secreted by P. serpens based on the well-described functionsof cruzipain. In this context, the cruzipain-like peptidases in P. ser-pens could be helpful for the interactions between the parasite andits invertebrate host in a direct or indirect manner. Our group hasalready suggested the importance of these molecules for directprocesses of adhesion between the parasites and the salivaryglands explanted from O. fasciatus, a phytophagous insect experi-mental model (Santos et al., 2006a, 2007). Herein, we demon-strated that cysteine peptidase activity produced by live P.serpens promastigote cells was capable of cleaving at least one sur-face polypeptide of 115 kDa located in the surface of salivaryglands of O. fasciatus. Therefore, we suggest a possible indirectfunction for the surface and/or secreted molecules on these inter-action processes: the enzymes could be cleaving important surfaceprotein receptors, exposing essential epitopes for parasite–hostcell connections.
Acknowledgments
This work was supported by grants from the Brazilian Agencies:Fundação Universitária José Bonifácio (FUJB), Financiadora de Estu-dos e Projetos (FINEP), Conselho Nacional de DesenvolvimentoCientífico e Tecnológico (CNPq), Conselho de Ensino para Gradua-dos e Pesquisa (CEPG/UFRJ), and Fundação de Amparo à Pesquisado Estado do Rio de Janeiro (FAPERJ).
References
Azambuja, P., Feder, D., Mello, C., Gomes, S., Garcia, E., 1999. Immunity in Rhodniusprolixus: trypanosomatid-vector interactions. Memórias do Instituto OswaldoCruz 94, 219–222.
Barrett, A.J., Rawlings, N.D., O’brien, E.A., 2001. The MEROPS database as a proteaseinformation system. Journal of Structural Biology 134, 95–102.
Bordier, C., 1981. Phase separation of integral membrane proteins in Triton X-114solutions. Journal of Biology Chemistry 256, 1604–1607.
Branquinha, M.H., Vermelho, A.B., Goldenberg, S., Bonaldo, M.C., 1996. Ubiquity ofcysteine- and metalloproteinase activities in a wide range of trypanosomatids.Journal of Eukaryotic Microbiology 43, 131–135.
Breganó, J.W., Picão, R.C., Graça, V.K., Menolli, R.A., Jankevicius, S.I., Filho, P.P.,Jankevicius, J.V., 2003. Phytomonas serpens, a tomato parasite, shares antigenswith Trypanosoma cruzi that are recognized by human sera and induceprotective immunity in mice. FEMS Immunology and Medical Microbiology39, 257–264.
Buroker-Kilgore, M., Wang, K.K.W., 1993. A Coomassie brilliant blue G-250-basedcolorimetric assay for measuring activity of calpain and other proteases.Analytical Biochemistry 208, 387–392.
Camargo, E.P., 1999. Phytomonas and other trypanosomatid parasites of plants andfruit. Advanced Parasitology 42, 29–112.
Cardoso de Almeida, M.L., Heise, N., 1993. Proteins anchored viaglycosylphosphatidylinositol and solubilizing phospholipases in Trypanosomacruzi. Biological Research 26, 285–312.
Cazzulo, J.J., Stoka, V., Turk, V., 2001. The major cysteine proteinase of Trypanosomacruzi: a valid target for chemotherapy of Chagas disease. CurrentPharmaceutical Design 7, 1143–1156.
C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352 351

Author's personal copy
Cuevas, I.C., Cazzulo, J.J., Sanchez, D.O., 2003. Gp63 homologues in Trypanosomacruzi: surface antigens with metalloprotease activity and a possible role in hostcell infection. Infection and Immunity 71, 5739–5749.
d’Avila-Levy, C.M., Melo, A.C., Vermelho, A.B., Branquinha, M.H., 2001. Differentialexpression of proteolytic enzymes in endosymbiont-harboring Crithidia species.FEMS Microbiology Letters 202, 73–77.
d’Avila-Levy, C.M., Araújo, F.M., Vermelho, A.B., Soares, R.M.A., Santos, A.L.S.,Branquinha, M.H., 2005. Proteolytic expression in Blastocrithidia culicis:influence of the endosymbiont and similarities with virulence factors ofpathogenic trypanosomatids. Parasitology 130, 413–420.
d’Avila-Levy, C.M., Dias, F.A., Nogueira de Melo, A.C., Martins, J.L., Lopes, A.H.C.S.,Santos, A.L.S., Vermelho, A.B., Branquinha, M.H., 2006a. Insights into the role ofgp63-like proteins in lower trypanosomatids. FEMS Microbiology Letters 254,149–156.
d’Avila-Levy, C.M., Santos, L.O., Marinho, F.A., Dias, F.A., Lopes, A.H.C.S., Santos,A.L.S., Branquinha, M.H., 2006b. Gp63-like molecules in Phytomonas serpens:possible role in the insect interaction. Current Microbiology 52, 439–444.
Del Nery, E., Juliano, M.A., Meldan, M., Svendsen, I., Scharfstein, J., Walmsley, A.,Juliano, L., 1997. Characterization of the substrate specificty of the majorcysteine protease (cruzipain) from Trypanosoma cruzi using a portion-mixingcombinatorial library and fluorogenic peptides. Biochemistry Journal 323, 427–433.
Dias, F.A., Santos, A.L.S., Ferreira-Pereira, A., Romeiro, A., Zimmermann, L.T.,Nascimento, M.T.C., Atella, G.C., Saraiva, E.M., Linden, R., Lopes, A.H.C.S., 2007.A novel ligand blot assay using biotinylated live cells. Journal of BiomolecularScreening 12, 1006–1010.
Dollet, M., 1984. Plant diseases caused by flagellate protozoa (Phytomonas). AnnualReview of Phytopathology 22, 115–132.
Dollet, M., Wallace, F.G., 1987. Compte rendu du premier Phytomonas workshop,Cayenne. Oleagineux 42, 461–468.
Dollet, M., Giannotti, J., Ollagnier, M., 1977. Observation de protozoaires flagellesdans les tubes cribces de palmiers a huile malades. Les Comptes Rendus del0Académie des Sciences 284, 643–645.
Elias, C.G.R., Pereira, F.M., Silva, B.A., Alviano, C.S., Soares, R.M.A., Santos, A.L.S., 2006.Leishmanolysin (gp63 metallopeptidase)-like activity extracellularly releasedby Herpetomonas samuelpessoai. Parasitology 130, 79–88.
Fresno, M., Hernandez-Munain, C., de-Diego, J., Rivas, L., Scharsftein, J., Bonay, P.,1994. Trypanosoma cruzi: identification of a membrane cysteine proteinaselinked through a GPI anchor. Brazilian Journal of Medical and BiologicalResearch 27, 431–437.
Fritig, B., Heitz, T., Legrand, M., 1998. Antimicrobial proteins in induced plantdefense. Current Opinion in Immunology 10, 16–22.
Garcia-Olmedo, F., Molina, A., Alamillo, J.M., Rodriguez-Palanzuela, P., 1998. Plantdefense peptides. Biopolymers 47, 479–491.
Gibbs, A.J., 1957. Leptomonas serpens n. sp., parasitic in the digestive tract and thesalivary glands of Nezara viridula (Pentatomidae) and in the sap of Solanumlycopersicun (tomato) and other plants. Parasitology 47, 297–303.
Lafont, A., 1909. Sur la présence d’un parasite de la classe des flagellés dans le latexde l’Euphorbia pilulifera. Comptes Rendus des Séances et Mémoires de laSociété de Biologie 66, 1011–1013.
Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J., 1951. Protein measurementwith the Folin phenol reagent. The Journal of Biological Chemistry 193, 265–275.
Malek, K., Dietrich, R.A., 1999. Defense on multiple fronts: how do plants cope withdiverse endemies? Trends in Plant Science 4, 215–219.
Mosolov, V.V., Grigoreva, L.I., Valueva, T.A., 2001. Involvement of proteolyticenzymes and their inhibitors in plant protection. Applied of BiochemistryMicrobiology 37, 115–123.
Mottram, J.C., Coombs, G.H., Alexander, G., 2004. Cysteine peptidases as virulencefactors of Leishmania. Current Opinion in Microbiology 7, 375–381.
Nogueira de Melo, A.C., d’Avila-Levy, C.M., Branquinha, M.H., Vermelho, A.B., 2002.Crithidia guilhermei: gelatin and haemoglobin-degrading extracellularmetalloproteinases. Experimental Parasitology 102, 150–156.
Nogueira de Melo, A.C., d’Avila-Levy, C.M., Dias, F.A., Armada, J.L.A., Silva, H.D.,Lopes, A.H.C.S., Santos, A.L.S., Branquinha, M.H., Vermelho, A.B., 2006.Peptidases and gp63-like proteins in Herpetomonas megaseliae: possibleinvolvement in the adhesion to the invertebrate host. International Journalfor Parasitology 36, 415–422.
Parthasarathy, M.V., 1976. Trypanosomatid flagellates in the phloem of diseasedcoconut palms. Science 192, 1346–1348.
Romeiro, A., Leal, L.H.M., De Souza, W., Attias, M., 2003. Interaction of Leptomonaswallacei with the intestinal tract of its natural host Oncopeltus fasciatus(Hemiptera: Lygaeidae). Journal of Invertebrate Pathology 82, 41–49.
Rosenthal, P.J., Sijwali, P.S., Singh, A., Shenai, B.R., 2002. Cysteine proteases ofmalaria parasites: targets for chemotherapy. Current Pharmacological Design 8,1659–1672.
Sajid, M., McKerrow, J.H., 2002. Cysteine proteases of parasitic organisms. Molecularand Biochemical Parasitology 120, 1–21.
Santos, A.L.S., Batista, L.M., Abreu, C.M., Alviano, C.S., Angluster, J., Soares, R.M.A.,2001. Developmentally regulated protein expression mediated bydimethylsulfoxide in Herpetomonas samuelpessoai. Current Microbiology 42,111–116.
Santos, A.L.S., Alviano, C.S., Soares, R.M.A., 2002. Detection of sialoglycomolecules infive plant trypanosomatids and in an insect phytophagous isolate. FEMSMicrobiology Letters 214, 19–23.
Santos, A.L.S., Rodrigues, M.L., Alviano, C.S., Angluster, J., Soares, R.M.A., 2003.Herpetomonas samuelpessoai: dimethylsulfoxide-induced differentiation isinfluenced by proteinase expression. Current Microbiology 46, 11–17.
Santos, A.L.S., Alviano, C.S., Soares, R.M.A., 2005. Use of proteolytic enzymes asan additional tool for trypanosomatid identification. Parasitology 130, 79–88.
Santos, A.L.S., d’Avila-Levy, C.M., Dias, F.A., Ribeiro, R.O., Pereira, F.M., Elias, C.G.R.,Souto-Padrón, T., Lopes, A.H.C.S., Alviano, C.S., Branquinha, M.H., Soares, R.M.A.,2006a. Phytomonas serpens: cysteine peptidase inhibitors interfere with growth,ultrastructure and host adhesion. International Journal for Parasitology 36, 47–56.
Santos, A.L.S., Branquinha, M.H., d’Avila-Levy, C.M., 2006b. The ubiquitous gp63-likemetalloprotease from lower trypanosomatids: in the search for a function.Anais da Academia Brasileira de Ciências 78, 387–714.
Santos, A.L.S., d’Avila-Levy, C.M., Elias, C.G.R., Vermelho, A.B., Branquinha, M.H.,2007. Phytomonas serpens: immunological similarities with the humantrypanosomatid pathogens. Microbes and Infection 9, 915–921.
Souto-Padrón, T., Campetella, O.E., Cazzulo, J.J., De Souza, W., 1990. Cysteineproteinase in Trypanosoma cruzi: immunocytochemical localization andinvolvement in parasite-host cell interaction. The Journal of Cell Science 96,485–490.
Vermelho, A.B., Giovanni-de-Simone, S., d’Avila-Levy, C.M., Santos, A.L.S., Nogueirade Melo, A.C., Silva-Junior, F.P., Bom, E.P., Branquinha, M.H., 2007.Trypanosomatidae peptidases: a target for drugs development. CurrentEnzyme Inhibition 3, 19–48.
Yao, C., Donelson, J.E., Wilson, M.E., 2007. Internal and surface-localized MSP ofLeishmania and their differential release from promastigotes. Eukaryotic Cell 6,1905–1912.
Yokoyama-Yasunaka, J.K., Pral, E.M., Oliveira-Junior, O.C., Alfieri, S.C., Stolf, A.M.,1994. Trypanosoma cruzi: identification of proteinases in shed components oftrypomastigote forms. Acta Tropica 57, 307–315.
352 C.G.R. Elias et al. / Experimental Parasitology 120 (2008) 343–352






