
Two new species of Erysiphe sect. Uncinula (Erysiphales): Erysiphe fernandoae and E. michikoae

Cortinarius sect. Armillati in northern Europe
Tuula Niskanen1
Department of Biosciences, Plant Biology, P.O. Box 65,FI-00014 University of Helsinki, Finland
Ilkka KytovuoriBotanical Museum, P.O. Box 7, FI-00014 University ofHelsinki, Finland
Kare LiimatainenDepartment of Biosciences, Plant Biology, P.O. Box 65,FI-00014 University of Helsinki, Finland
Abstract: Cortinarius sect. Armillati (subgenus Tela-monia) was studied extensively based on morphologyand molecular data. A total of about 1000 specimens,mostly from Fennoscandia, were revised. The nomen-clature of the species was confirmed by sequencingthe type material. Phylogenetic relationships wereinferred by analyses of ITS, and the results werecompared with the morphological and ecologicaldata. Based on macro- and micromorphologicalcharacters, as well as molecular data, section Armillaticontains only the medium to large species withslightly hygrophanous pileus and 6 reddish or insome species yellowish brown to rose brown universalveils. The other red-brown-veiled species, previouslyincluded in Armillati, seem to belong to at least sevendifferent sections or clades: sect. Anthracini, sect.Boulderenses, sect. Brunneotincti p.p., sect. Cinnabar-ini, sect. Fulvescentes, /Fuscoperonatus, and /Praesti-giosus. Our study recognized six Armillati speciesfrom northern Europe: C. armillatus, C. luteo-ornatus,C. paragaudis, and three species described as new, C.pinigaudis, C. roseoarmillatus, and C. suboenochelis.The former three also occur in North America. Twoadditional species, C. subarmillatus (Japan) and C.quercoarmillatus (Costa Rica), are known outside thearea. Based on the phylogenetic analysis, the speciesassociated with deciduous trees, C. armillatus, C.quercoarmillatus, and C. roseoarmillatus, all withdextrinoid, thick-walled spores, formed a separategroup from the mainly conifer-associated species, C.luteo-ornatus, C. paragaudis, C. pinigaudis and C.suboenochelis, all with fairly thin to moderately thick-walled, indextrinoid to moderately dextrinoid spores.Descriptions of the northern European species areprovided, the distribution is mapped and theirtaxonomy, ecology, distribution, and relationships
are discussed. A total of 64 new sequences of 12species are reported including 17 sequences fromtype material. Our study also suggests that ITSsequences are not always sufficiently variable forspecies-rank recognition (barcoding) in Cortinarius.
Key words: Agaricales, barcoding, ITS, molecularsystematics, phylogeny, taxonomy
INTRODUCTION
Cortinarius is the largest genus of the Agaricales witha worldwide distribution. All the species are mycor-rhizal and associated with different shrubs and treesof for example Pinaceae, Fagales and Salicaceae. Thegenus is divided into several subgenera, and sectionArmillati traditionally is placed in subgenus Telamo-nia, which is the most species rich and taxonomicallydifficult (e.g. Bidaud et al. 1995; Brandrud et al. 1989,1992; Moser 1965; Melot 1990).
The name Armillati first was introduced by Kuhner& Romagnesi (1953), who included only threespecies, C. armillatus (Fr. : Fr.) Fr., C. veregregiusRob. Henry and C. fuscoperonatus Kuhner. Thesection was validly described by Moser in 1965. Heplaced almost all known red-veiled telamonioidspecies in it and further divided it into groupsArmillatus, Bulliardii and Miniatopus. Høiland(1983) restricted the section to include only speciescontaining anthraquinoid pigments. He described anew subsection, Anthracini, in addition to Moser’sgroups. Melot (1990) restricted section Armillati evenfurther to include only species with slightly hygro-phanous pilei, brown to red (rarely yellow) veils orbasidiospores reddening in KOH. The hygrophanousspecies were placed into sections Anthracini, Cinna-barini, and Fulvescentes. Bidaud et al. (1995) consid-ered sect. Armillati, more widely dividing it into fourseries, Armillatus, Craticius, Paragaudis, and Praesti-giosus. We detailed these classification systems (SUP-
PLEMENTARY TABLE I). The red-veiled Telamoniaspecies also were studied by Soop (1990, 1997).Outside Europe two Armillati species have beendescribed, C. subarmillatus Hongo from Japan(Hongo 1965) and C. quercoarmillatus Ammirati,Halling & Garnica from Costa Rica (Ammirati et al.2007).
The taxonomy of species in sect. Armillati hasrelied on morphology, pigment chemistry and ecol-ogy, resulting in the diverse classifications describedabove. Recently the use of molecular data has aidedtaxonomic work. The infrageneric classification of
Submitted 4 Nov 2010; accepted for publication 16 Feb 2011.1 Corresponding author. E-mail: [email protected]
Mycologia, 103(5), 2011, pp. 1080–1101. DOI: 10.3852/10-350# 2011 by The Mycological Society of America, Lawrence, KS 66044-8897
1080

Cortinarius has been studied based on rDNA ITS1and ITS2 and/or adjacent LSU region, for exampleby Garnica et al. (2005) and Peintner et al. (2004).They supported the placement of sect. Armillati insubgenus Telamonia and suggested that C. armillatusis not closely related to Cinnabarini, Anthracini orFulvescentes.
The most widely used gene region for studies atspecies rank is the internal transcribed spacer region(ITS), which has proven to be useful in Cortinarius insubgenus Telamonia by Kytovuori et al. (2005),Lindstrom et al. (2008), Niskanen et al. (2009) andSuarez-Santiago et al. (2009), in subgenus Phlegmaciumby Garnica et al. (2009), Frøslev et al. (2007) andOrtega et al. (2008) and in subgenus Myxacium byPeintner (2008). In most studies the majority of speciesdiffer with respect to the degree of intra- andinterspecific variation, and most polymorphic speciestypically do not have closely related sister taxa; theamount of intraspecific variation reported usually wasfewer than six base pairs (bp) while the interspecificvariation was more than six bp. Frøslev et al. (2007)and Niskanen et al. (2009) reported some exceptionswhere morphologically distinguishable species wereseparated only by 3–5 nucleotides. FurthermoreFrøslev et al. (2007), Garnica et al. (2005) and Peintner(2008) found that ITS sequences did not differentiatea few morphological species. Nearly all the aforemen-tioned studies found morphologically indistinguish-able subgroups inside morphologically delimitedspecies. Unlike most studies comparing taxa at speciesrank, Ortega et al. (2008) and Suarez-Santiago et al.(2009) also compared infraspecific varieties. In theirstudy species differed by at least 10 diagnostic positionsand varieties by 2–9 diagnostic positions.
In all recent Cortinarius studies, regardless of thedegree of genetic variation within or among clades,the amount of morphological difference determinesthe taxonomic rank of the clades obtained fromphylogenetic analysis based on ITS sequences. There-fore no consensus of the limits between the intra- andinterspecific variation in ITS regions exists. Despitethese problems ITS sequences have been proposed asspecies-identifier sequences (barcoding) in Cortinar-ius (Frøslev et al. 2007, Ortega et al. 2008).
Our aim was to study the taxonomy, ecology anddistribution of the species of section Armillati innorthern Europe as well as study the limitation ofsect. Armillati based on morphology and moleculardata. We also studied all relevant type specimens.
MATERIALS AND METHODS
Material.—The herbarium material of Armillati speciesfrom Denmark (C), Norway (O, BG, TRH, TROM), Sweden
(S, GB, LD, UME, UPS) and Finland (H, JOE, KUO, TUR,OULU) was studied as well as specimens gathered by theauthors, a total of about 1000 specimens. We also examinedthe type specimens, mentioned in the beginning ofMolecular analysis. Herbarium acronyms follow Thiers(continuously updated) and the vegetation zones of Ahtiet al. (1968) and Knudsen and Vesterholt (2008). Forbiogeographical provinces see Knudsen and Vesterholt(2008). Collectors are abbreviated by TN, IK, KL and PK,in collections made by us and Pirjo Kytovuori.
Molecular analyses.—Several collections (n 5 46) fromdifferent geographical areas (specimens marked with ‘‘D’’ inSUPPLEMENTARY TABLES II–VII) and type material of thesetaxa were sequenced: Cortinarius armillatus var. luteo-ornatus MM1962/0027 (IB), C. armillatus var. miniatusGK432464 (G), C. armillatus var. subcroceofulvus GK432465(G), C. bresadolianus GK432466 (G), C. paragaudis CFP564(S), C. paragaudis ssp. oenochelis CFP546 (S), C. salmoneo-tomentosus var. pseudoparagaudis GK432479 (G), C. sub-armillatus var. picetorum GK432481 (G), C. vaccinochelisPC90323 (PC), C. badiovinaceus MM1950/0061 (IB), C.caput-medusae CFP1300 (S), C. fillioni GK432471 (G), C.subtestaceus AHS15458 (UPS) and C. veregregius PC0090322(PC). A collection of C. suberi IK95-349 (H) also wassequenced. Altogether we generated 64 sequences. We didnot succeed in sequencing the type material of C.subarmillatus. The sequenced specimens belonging tosection Armillati are listed under each species.
Total DNA was extracted from a few milligrams of driedmaterial (a piece of lamella) with the NucleoSpin Plant kit(Macherey-Nagel). Primers ITS 1F and ITS 4 (Gardes andBruns 1993, White et al. 1990) were used to amplify ITSregions. Primer combinations ITS 1F/ITS 2 and ITS 3/ITS4 were used on problematic material. The same primer pairswere used in direct sequencing. PCR amplification andsequencing followed Niskanen et al. (2009). Sequenceswere assembled and edited with Sequencher 4.1 (GeneCodes, Ann Arbor, Michigan).
Intragenomic polymorphisms were observed as mixedpeaks in chromatographic data.
Base polymorphisms are marked with ambiguous IUBcodes and length polymorphisms with N (informationprovided on request). Frøslev et al. (2007), Niskanen et al.(2009) and Suarez-Santiago et al. (2009) reported theintragenomic variation in Cortinarius. The sequences werecompared with the material in public databases (GenBank:http://www.ncbi.nlm.nih.gov/; UNITE: http://unite.ut.ee/) with BLAST queries.
A sequence of every species was compared with all othersequences of the Armillati species by using BLAST toestimate their genetic distances. These sequences and thesequences of the closest ones then were aligned with theMuscle program (Edgar 2004) on the European Bioinfor-matics Institute server (http://www.ebi.ac.uk/Tools/muscle/index.html). These differences were counted fromthe alignments: (i) the observed number of variable sitesshowing how many sites contain infraspecific and/orinfragenomic polymorphisms and (ii) the differencesbetween the closest species as minimum evolutionary events
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1081

including indels, transitions and transversions, and forexample multiple base indels are treated as one change.Only differences shared by all the specimens of the samespecies were counted. The results are presented in point‘‘ITS regions’’ under every species.
For the preliminary analysis (POY [Gladstein andWheeler 2001, Wheeler 1996], not shown) we chose 200of our own unpublished and published ITS sequencescovering all the major clades of subgenus Telamonia s. s.Using a BLAST query of the public databases we also choseall the sequences similar to our Armillati material, andpreviously published sequences of the type material of C.boulderensis, C. hinnuleoarmillatus and C. quercoarmillatusand a sequence of C. anthracinus, C. bulliardii, C.cinnabarinus, C. colus, C. praestigiosus and C. raphanoides.The analysis was run with Cortinarius norrlandicus (subge-nus Phlegmacium) as outgroup. Section Armillati, aspresented in this paper, formed a monophyletic group,and for the final analysis we chose sect. Armillati and 33other Telamonia species to represent the various clades ofsubgenus Telamonia, and which previously been classified insect. Armillati, and/or appear related to Armillati by treetopology. Based on preliminary analyses, three species fromsubgenus Phlegmacium were chosen as outgroup species. Analignment of 110 sequences for phylogenetic analyses wasproduced with Muscle under default settings and followedby manual adjustments in BioEdit (www.mbio.ncsu.edu/BioEdit/bioedit.html). The alignment comprised 722 nu-cleotides (including gaps) and is available at TreeBASEunder S11133 (http://www.treebase.org/treebase-web/home.html).
Bayesian inference (BI) was performed with Mr.Bayes 3.1.1(Huelsenbeck and Ronquist 2003). The entire dataset wasanalyzed with the GTR model including a gamma shapeparameter and estimating the proportion of invariable sites.Two independent runs with four chains in each were performedfor 1 000 000 generations with sampling every 100th generation.All trees sampled before stationarity were discarded with a 25%
safety margin (burn-in of 2500 trees [250 000 generations]).Sampled trees from both runs were combined in a 50% majorityrule consensus phylogram with posterior probabilities (PP). Theanalysis was run with computer clusters of the CSC, IT Centrefor Science, Espoo, Finland.
Morphological studies.—Morphological descriptions arebased on material collected by the authors, includingspecimens in all stages of development. Macroscopiccharacteristics were observed from fresh basidiomata. Someof the collections also were photographed fresh. Codes werenot used to describe colors, however color photographs ofthe new species, C. pinigaudis, C. roseoarmillatus, and C.suboenochelis, are provided along with photographs of C.armillatus, C. luteo-ornatus, and C. paragaudis for compar-ison. The macrochemical reaction of the universal veil wastested with 40% KOH on fresh material, but it was not testedsystematically.
We observed microscopic characteristics from driedmaterial mounted in Melzer’s reagent (MLZ) and com-pared them with observations made on dried materialmounted in 5% KOH. Measurements were made in MLZ
with an ocular micrometer using 1003 oil-immersion lens.We measured 20 basidiospores from one fruit body fromeach collection (specimens marked with an ‘‘S’’ inSUPPLEMENTARY TABLES II–VII), from the veil or top of thestipe. In C. luteo-ornatus and C. subarmillatus the differencein basidiospore size between the lamellae and veil wasgreater than usual for Cortinarius; so 20 basidiospores fromlamellae were measured for comparison. The length andwidth were measured from each spore, and their length/width ratios (Q value) were calculated. Extreme measure-ments, clearly differing from the others, were excludedfrom the final values. The spore size is indicated as min.–average of all the collections–max. The lamellar trama andbasidia also were examined and the pileipellis structure wasstudied from both radial freehand sections and scalps fromthe pileus center or midway between the center and margin.The measurements of the elements of pileipellis were madefrom scalp preparations.
RESULTS
Molecular analyses.—All six Armillati species aremorphologically distinct. Four of them have a clearbarcoding gap, a smaller intraspecific pairwise dis-tance than interspecific, in the ITS sequences. C.paragaudis and C. pinigaudis differ by only one basepair, while the other four species differ from theclosest species by more than 13 evolutionary events.
Four species were genetically polymorphic, andmore than 30% of samples from those species hadintragenomic polymorphisms. Polymorphisms werenot observed in two species, C. roseoarmillatus and C.suboenochelis. (See ITS regions for each species.)
The 50% majority rule phylogram resulting fromthe BI analysis is provided (FIG. 1) with posteriorprobabilities indicated above the branches. Theingroup representing sect. Armillati was supportedwith 100 PP, and within sect. Armillati all speciesexcept C. paragaudis complex also were supportedwith 100 PP; Cortinarius pinigaudis was supportedonly by a 51 PP value and C. paragaudis did not forma monophyletic group. Monophyletic groups with ahigh PP value were formed inside species C.armillatus, C. luteo-ornatus, and C. paragaudis-com-plex. Within sect. Armillati three species groups werewell or somewhat supported, C. armillatus/C. quer-coarmillatus/C. roseoarmillatus clade (80 PP), C.suboenochelis/C. luteo-ornatus (95 PP) and C. sub-oenochelis/C. luteo-ornatus/C. paragaudis/C. pinigau-dis clade (96 PP). We present species in the order ofthis classification in TAXONOMY.
These analyses supported the Telamonia sensu latosections Anthracini (96), Brunnei (100 PP), Fulves-centes (100 PP), Firmiores (100 PP), Hinnulei (99 PP)and Malachi (100 PP) and somewhat supported cladeBovini (76 PP) and sect. Brunneotincti p.p. (84 PP).
1082 MYCOLOGIA
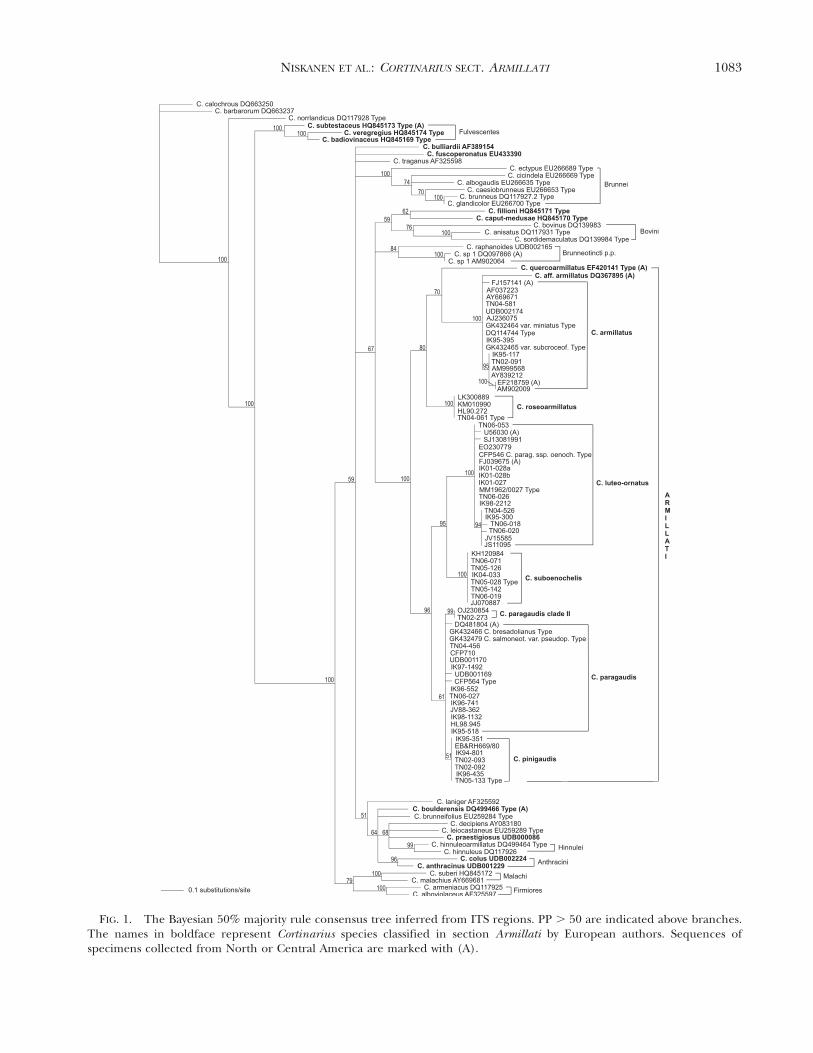
FIG. 1. The Bayesian 50% majority rule consensus tree inferred from ITS regions. PP . 50 are indicated above branches.The names in boldface represent Cortinarius species classified in section Armillati by European authors. Sequences ofspecimens collected from North or Central America are marked with (A).
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1083

A comparison of all GenBank and UNITE sequenc-es with sequences generated in this study revealed 20belonging to sect. Armillati representing three of thesix species from northern Europe (see point sequenc-es retrieved from the public databases under eachspecies and SUPPLEMENTARY TABLE VIII). Of these 65%
were deposited in public databases under the wrongname or without a species designated, leaving only35% correctly identified. Possibly one new Armillatispecies from British Columbia, Canada, (DQ367895,C. aff. armillatus) was found among these sequences.
TAXONOMY
Cortinarius sect. Armillati Kuhner & Romagn. exM.M. Moser, Schweiz. Z. Pilzk. 43(8):114 (1965)em. Niskanen, Kytov. & Liimat.Type species. Cortinarius armillatus (Fr. : Fr.) Fr.Medium to large, fruit body usually 6 fleshy. Pileus
6 innately fibrillose and/or with small scales, onlysomewhat hygrophanous and often with hygropha-nous streaks, yellowish red-brown to ochraceousbrown to grayish brown. Universal veil pinkish tovinaceous red or in some species yellowish brown orbrownish pink, at least when young. Stipe clavate tocylindrical, white fibrillose when young, later grayishbrown. Context marbled brown to gray brown whenfresh and moist, becoming whitish brown with loss ofmoisture. Lamellae odor indistinct or faintly rapha-noid. Exsiccata with brown, grayish brown to darkbrown pileus and grayish to brownish stipe, 6 reduniversal veil often visible on the stipe, sometimes thewhole fruiting body almost blackish, at least in C.armillatus, C. roseoarmillatus, and C. luteo-ornatus.Spores subglobose to amygdaloid to ellipsoid, some 6
reddening in KOH, but not in every specimen of thesame species, best seen with 403 lens. Lamellar tramahyphae not encrusted or finely scabrous to spot-likeencrusted, best seen in the exsiccata. Epicutis usuallymedium thick; narrow uppermost hyphae withyellowish brown to olivaceous yellowish brown con-tents; hypoderm weakly developed. Anthraquinoidpigments are found at least from C. armillatus and C.paragaudis (Bendiksen and Bendiksen 1993, Høiland1983), and the universal veil of at least C. armillatus,C.luteo-ornatus, C. paragaudis and C. pinigaudis turnspurplish with 40% KOH, but we have not systemati-cally tested this characteristic. Species are associatedwith coniferous and/or deciduous trees.
Macroscopic characteristics are best seen in youngto middle-aged fresh fruit bodies. With age the pileusloses its fine structure and becomes fibrillose or scalyand the universal veil can become darker or vinaceousred. Although macroscopical characteristics exist, thespecies are most reliably distinguished by the spore
size, shape and dextrinoidity. The spores howeverseemingly develop fairly late, so older fruit bodiesalways should be included in the exsiccata. The sporesof young fruit bodies are often somewhat smaller thanthose from older ones.
Species with a 6 reddish or yellowish brownuniversal veil in sections Brunneotincti M.M. Moser,Fulvescentes Melot and Boulderenses Niskanen, Liimat.& Kytov. can resemble Armillati species, but Brunneo-tincti species usually have olivaceous tints in thefruiting body and obovoid to ellipsoid spores. If thespores are broadly ellipsoid to almost subglobose thenlarge, extracellular olivaceous spots deep in thepileipellis exist. Fulvescentes and Boulderenses specieshave hygrophanous pileus, and the species of sect.Fulvescentes are smaller and slenderer with yellowishbrown flesh. Cortinarius spilomeus also has a brownishred to vinaceous universal veil, but the veil usuallyforms small spots instead of bands on the stipe; thepileus is pale grayish brown, the lamellae and top ofthe stipe often have a violet tint when young, and thespores are pale, indextrinoid and subglobose. Corti-narius hinnuleoarmillatus has an abundant red-orange universal veil but differs from the Armillatispecies by the hygrophanous pileus, ochraceouscontext, strongly ornamented, obovoidly ellipsoidspores, and shared habitat with Quercus. The univer-sal veil of Armillati species sometimes is indistinct,and the exsiccata can be blackish and easily confusedwith species of section Brunnei (see C. luteo-ornatusand C. paragaudis). The key to the species of sectionArmillati in Fennoscandia is presented in Niskanenand Kytovuori (2008) and to the colored-veiledspecies in Niskanen et al. (2008b).
Cortinarius armillatus (Fr. : Fr.) Fr., Epicr. Syst. Mycol.295 (1838). FIGS. 2A, 3A, 4–7Basionym. Agaricus armillatus Fr., Observ. Mycol.
2:61 (1818): sanctioned in Fr., Syst. Mycol. 1:214(1821). Type: SWEDEN. ANGERMANLAND: Sabra,Furuhultsan 2 km N Innerbran, in wet coniferousforest with blueberries (Picea, Betula), 23 Aug 1987,Lindstrom et al. CFP584 (S, NEOTYPE, neotypus hicdesignatus). GenBank DQ114744.
Cortinarius armillatus var. miniatus Reumaux,Atlas des Cortinaires 7:227 (1995). Type: FRANCE.ARDENNES: La Croix aux Bois, under Betula, 200 m,2 Oct 1985, P. Reumaux 3127 (G, HOLOTYPE).GenBank HQ845115.
Cortinarius armillatus var. subcroceofulvus Bidaud,Moenne-Locc. & Reumaux, Atlas des Cortinaires7:228 (1995). Type: FRANCE. ARDENNES: Foretde Belval, under Carpinus betulus, 200 m, 20 Sep1992, P. Reumaux 3135 (G, HOLOTYPE). GenBank
1084 MYCOLOGIA

HQ845116. Cortinarius haematochelis sensu Bidaud,Moenne-Locc. & Reumaux, Atlas des Cortinaires 7:fiche 265 (1995).
Illustrations. Bidaud et al. (1995: pl. 146, 147),Brandrud et al. (1992: pl. B09), Consiglio et al. (2004:pl. B17), Fries (1867–1884: pl. 158).
Pileus. 5–11 cm, convex with an incurved marginwhen young, later plano-convex with a low, broadumbo; surface often has distinct, small red scalesespecially at the center, finely innately fibrillosetoward the edge, often with orange-red patches orcoating of veil in the margin, yellowish red-brown toyellow-brown, often somewhat paler toward the edge;only somewhat hygrophanous, with fine, dark hygro-
phanous streaks. Lamellae: medium-spaced (45–65reaching the stipe), emarginate, medium thick tothick, light brown when young, dark brown with age;edge somewhat lighter, rather uneven. Stipe: 7–17(–20) 3 0.8–1.5 cm (top), cylindrical, clavate at base(23.0 cm) rarely wholly cylindric, white fibrillosewhen young, later grayish brown. Basal mycelium:white to pale pinkish. Universal veil on stipe: vividorange-red, usually forming an annular band on themiddle part and distinct belts or zones lower down.Context: marbled with gray brown to brown whenfresh and moist, becoming whitish brown with loss ofmoisture, darker toward the base of the stipe. Odor: inlamellae slightly raphanoid. Exsiccata: pileus red-
FIG. 2. Photos of the North European Armillati species. A. Cortinarius armillatus Liimatainen & Niskanen 04-581. B. C.roseoarmillatus, Kytovuori (Niskanen 04-061) (type, H). C. C. suboenochelis, Kytovuori (Niskanen 05-028) (type, H). D. C. luteo-ornatus Liimatainen & Niskanen 04-526. E. C. paragaudis Liimatainen & Niskanen 04-456. F. C. pinigaudis, Liimatainen &Niskanen 05-133 (type, H). Photographs by K. Liimatainen.
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1085
brown to dark brown to sometimes blackish brown;stipe grayish to grayish brown; red universal veil stillvisible on the stipe; basal mycelium white or pinkish.Chemical reactions: KOH on universal veil on stipepurple.
Spores. 9.7–10.8–11.8 3 5.9–6.3–7.0 mm, Q 5 (1.53–)1.58–1.71–1.79(–1.87), av. 5 10.2–11.2 3 6.1–6.5 mm,Qav. 5 1.65–1.78 (320 spores, 16 collections, FIGS. 3A,4), weakly sigmoid to narrowly amygdaloid to narrowlyellipsoid, sometimes with a low suprahilar depressionslightly curved apex to the abaxial side of the spore,making the spores unique; thick-walled, fairly finely tomoderately, densely verrucose, most strongly at theapex; warts somewhat anastomosing, strongly dextri-noid, at least some spores becoming brownish red inKOH, but this character was not observed in all
specimens. Spores on the lamellae usually less dex-trinoid and slightly narrower. Lamellar trama hyphae:in the overall view (pale) olivaceous yellowish toolivaceous brownish in MLZ and KOH, smooth to
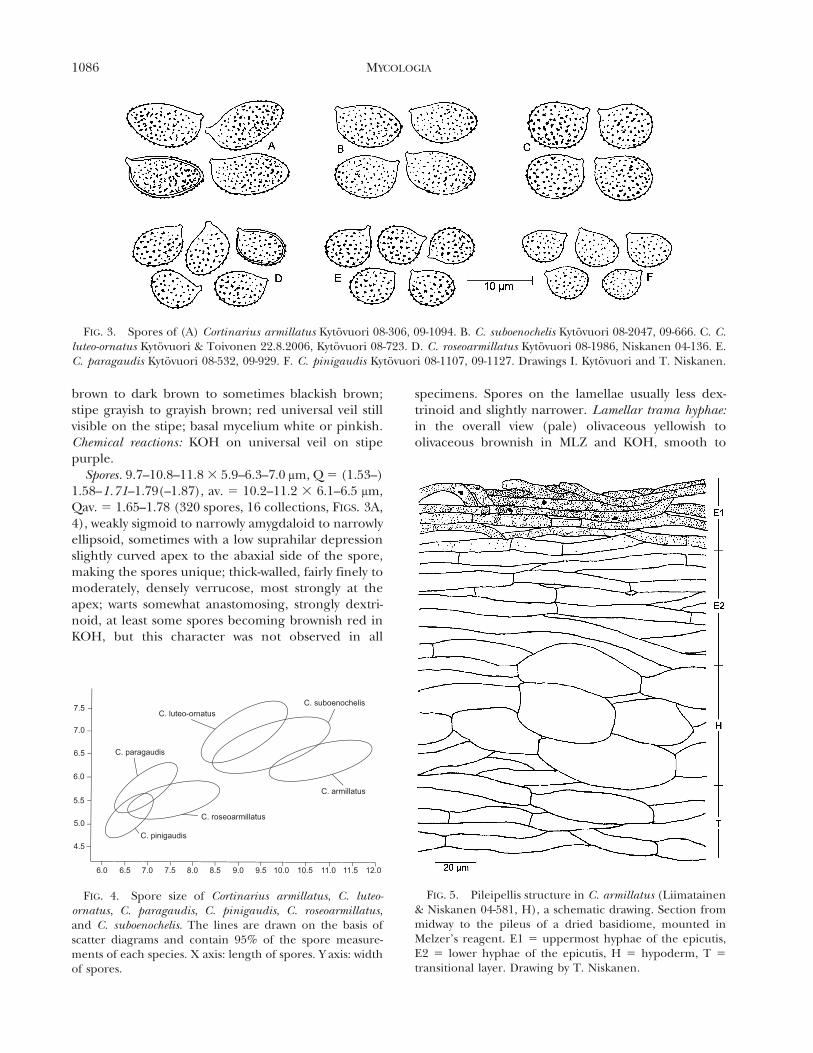
FIG. 3. Spores of (A) Cortinarius armillatus Kytovuori 08-306, 09-1094. B. C. suboenochelis Kytovuori 08-2047, 09-666. C. C.luteo-ornatus Kytovuori & Toivonen 22.8.2006, Kytovuori 08-723. D. C. roseoarmillatus Kytovuori 08-1986, Niskanen 04-136. E.C. paragaudis Kytovuori 08-532, 09-929. F. C. pinigaudis Kytovuori 08-1107, 09-1127. Drawings I. Kytovuori and T. Niskanen.
FIG. 4. Spore size of Cortinarius armillatus, C. luteo-ornatus, C. paragaudis, C. pinigaudis, C. roseoarmillatus,and C. suboenochelis. The lines are drawn on the basis ofscatter diagrams and contain 95% of the spore measure-ments of each species. X axis: length of spores. Y axis: widthof spores.
FIG. 5. Pileipellis structure in C. armillatus (Liimatainen& Niskanen 04-581, H), a schematic drawing. Section frommidway to the pileus of a dried basidiome, mounted inMelzer’s reagent. E1 5 uppermost hyphae of the epicutis,E2 5 lower hyphae of the epicutis, H 5 hypoderm, T 5
transitional layer. Drawing by T. Niskanen.
1086 MYCOLOGIA

distinctly finely scabrous, with small to large, olivaceousbrown spot-like incrustations. Basidia: four-spored, 30–35–41 3 8–8.5–10 mm, av. 33–40 3 7.6–9.4 mm (100basidia, 10 collections), with pale yellowish to oliva-ceous brownish contents in MLZ and KOH. Lamellaredge fertile with basidia and narrowly clavate cells.Pileipellis (FIG. 5): Epicutis medium thick; uppermosthyphae 4–10 mm wide, yellowish brown to olivaceousyellowish brown in MLZ and KOH, smooth (to veryfinely scabrous); lower hyphae 6–12 mm wide, almosthyaline, smooth to finely scabrous. Hypoderm weaklydeveloped with elongated elements about 40–75(–90)3 17–25 mm, almost hyaline, fairly thin-walled, smoothto finely encrusted. Hyphae in the transition to tramasmooth to finely encrusted in KOH, in MLZ withscanty, small, pale olivaceous brownish, spot-likeincrustations. Clamp connections: present.
ITS regions (including 5.8S region). 514–515 baseslong (based on a total of seven sequences, includingsequences from three type specimens). Sequencesfrom the type material of C. armillatus var. miniatusand C. armillatus var. subcroceofulvus were identical tothe neotype sequence. The other four sequences have
one indel and two intragenomic length polymor-phisms, but we found no morphological or ecologicalcharacteristics to support the groups. Cortinariusarmillatus differs from all other Armillati species bymore than 20 evolutionary events. Sequences retrievedfrom the public databases (a total of 10): Six of themwere from Nordic countries (AJ236075, UDB002174,AM902009, AM999568, AY839212, AY839206), onefrom British Columbia, Canada, (EF218759) andno locality was mentioned for two (AY669671,AF037223). The intraspecific variation in the se-quences is much higher than in our own sequences(0–7 evolutionary events), despite most also beingfrom Nordic countries. Some differences are probablyreading errors, such as AY839206 from Sweden thatdiffers by seven evolutionary events. Most of thedifferences in this sequence are indels at the end ofthe ITS 2 region, and they are double bases instead ofone. A common phenomenon is that at the end of thesequence signal quality is poor and peaks start to gettails that sometimes are read as separate bases.Therefore chromatograms should be examined toconfirm the differences. One sequence from NorthAmerica (DQ367895) however differs so much (14evolutionary events) that it could be a separate speciesfrom C. armillatus, but further studies are needed.One peculiar thing in the sequences retrieved fromthe public databases is their source. One of them isfrom house dust and one from bryophytes, which hasto be a consequence of C. armillatus’s abundantfruiting.
Ecology and distribution. Cortinarius armillatusgrows in deciduous and mixed forests with Betula,
FIG. 6. Fruiting periods of Cortinarius armillatus,C. luteo-ornatus, C. paragaudis, C. pinigaudis, C. roseoarmil-latus, and C. suboenochelis in the examined material. X axis:the season 1.VII–26.XI divided in pentads. Y axis: number ofspecimens collected in each pentad.
FIG. 7. Distribution of Cortinarius armillatus in northernEurope according to the examined material.
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1087

possibly also with Carpinus, on damp to fairly dry,usually oligotrophic soil. It also is common inconiferous forests under solitary birches and inmanmade habitats such as yards, parks, cemeteriesand roadsides. It grows in loose groups and often isabundant. It is one of the most common andabundant Cortinarius species in northern Europe.Fruit bodies occur late Jul–mid-Oct, but the peak ofthe fruiting season is often from mid- to late Aug(FIG. 6). Cortinarius armillatus is common in hemi-boreal to subarctic zones and rare to occasional in thetemperate zone (FIG. 7). The species is known inEurope and North America.
Differential diagnosis. Cortinarius armillatus is theeasiest species to identify in the group. Typical for thespecies are the vivid orange-red universal veil, whichusually forms distinct belts or zones on the stipe, fairlylarge, thick-walled, strongly dextrinoid spores andhabitats with Betula. The pileus also is typical withsmall red scales, especially at the center. All the otherArmillati species have less pronounced, pinkish tovinaceous red, rarely yellowish brown to pink brownuniversal veils and smaller and/or less dextrinoidand/or thin-walled spores. C. luteo-ornatus and C.pinigaudis sometimes can have almost C. armillatus-like bands of the universal veil on the stipe.
The description of C. armillatus by Fries (1821)‘‘pileo squamuloso testaceo-rubro … stipite bulboso,velo contexto rubro’’ and the fruiting bodies illustrat-ed in Fries (1867–1884: pl. 158) fit our species well.The collection Lindstrom et al. CFP584 (Brandrud etal. 1992: pl. B09) represents a typical C. armillatus, andwe propose it as a neotype for the species.
Two varieties of C. armillatus were described inBidaud et al. (1995): Cortinarius armillatus var.miniatus Reumaux and C. armillatus var. subcroceo-fulvus Bidaud, Moenne-Locc. & Reumaux. Cortinar-ius armillatus var. miniatus is a brilliant variety ofCortinarius armillatus with a minium red pileus anduniversal veil. The type material is collected underBetula. The exsiccatum is similar to those of C.armillatus and the spores (9.8–10.2–10.6[–11.2] 3
[5.6–]5.9–6.1–6.4 mm, Q 5 ]1.56–]1.60–1.67–1.75 [20spores]) fit our material well. Cortinarius armillatusvar. subcroceofulvus is, according to the authors, aslender variety of Cortinarius armillatus, which growsunder Carpinus betulus instead of Betula. Theexsiccatum and the spores are C. armillatus-like([10.1–]10.3–10.7–11.2[–11.7] 3 [5.9–]6.1–6.3–6.4[–6.9] mm, Q 5 [1.60–]1.63–1.69–1.78 [20 spores]).In both types the ITS regions are identical to those ofthe neotype of C. armillatus. Based on the morpho-logical, ecological and molecular data, we concludethat Cortinarius armillatus var. miniatus and C.armillatus var. subcroceofulvus essentially do not differ
from C. armillatus and should be considered syno-nyms of the latter. Cortinarius haematochelis sensuBidaud, Moenne-Locc. & Reumaux (1995) most likelyrepresents C. armillatus. The figure is C. armillatus-like and the spores are almost similar in size. Formore about C. haematochelis see C. paragaudis.
Specimens examined (a total of 565 collections, forthe complete list of specimens see SUPPLEMENTARY TABLE
II). SWEDEN. ANGERMANLAND: Sabra, Furuhultsan2 km N Innerbran, 23 Aug 1987, H. Lindstrom et al.CFP584 (S, NEOTYPE), GenBank DQ114744. FINLAND.POHJOIS-HAME: Uurainen, Kutula, 5 Aug 1995, I.K.95-117 (H, TUR), GenBank HQ845111. KOILISMAA:Kuusamo, Oulanka, Puukkorinne, 26 Aug 2002, K.L. &T.N. 02-091 (H), GenBank HQ845112. PERA-POHJAN-MAA: Rovaniemi, Pisavaara, 31 Aug 2004, K.L. & T.N. 04-581 (H), GenBank HQ845113. INARIN LAPPI: Inari, N sideof Lutto River, 14 Aug 1995, I.K. 95-395 (H, TUR),GenBank HQ845114. FRANCE. ARDENNES: Foret deBelval, 20 Sep 1992, P. Reumaux 3135 (G, HOLOTYPE ofC. armillatus var. subcroceofulvus), GenBank HQ845116. LaCroix aux Bois, 2 Oct 1985, P. Reumaux 3127 (G,HOLOTYPE of C. armillatus var. miniatus), GenBankHQ845115.
Cortinarius roseoarmillatus Niskanen, Kytov. & Lii-mat., sp. nov. FIGS. 2B, 3D, 4, 6, 8
MycoBank MB519556Cortinarius roseoarmillatus Niskanen, Kytov. & Lii-
mat., Funga Nordica: 727 (2008) (invalid).Pileus. 3.5–7.5 cm, convexus, dein planoconvexus, roseo-
brunneus, cum venis hygrophanis. Lamellae subconfertae,pallide brunneae, dein obscure brunneae. Stipes 6–9 3 0.6–1.3 cm, clavatus (22.7 cm), albide fibrillosus, dein pallido
FIG. 8. Distribution of Cortinarius roseoarmillatus innorthern Europe according to the examined material.
1088 MYCOLOGIA

griseobrunneus, velo roseo, cingulato. Caro brunnea,variegata. Odor nullus. Sporae 6.6–8.6 3 5.0–6.1 mm,amygdaliformes vel ovatae vel obovatoellipsoidae, subvaldeverrucosae. In silvis mediocriter humidis, cum Betulis.
Typus. FINLAND. POHJOIS-HAME: Virrat com-mune, Salmela, under Betula, also near Pinussylvestris and Picea abies, 16 Aug 2004, coll. Kytovuori(Niskanen 04-061) (H, HOLOTYPE; NY, ISOTYPE).GenBank HQ845117.
Etymology. roseoarmillatus (Latin), roseus 5 rose-colored, armillatus 5 with a ring. The epithet refersto the vivid rose-red universal veil bands and affinityto C. armillatus.
Pileus. 3.5–7.5 cm, convex with an incurved marginwhen young, plano-convex with a low, broad umbowhen fairly old, small-scaled at the center, finelyfibrillose toward the edge, with some rose-red coatingof veil in the margin, rose brown to pinkish brown (inold fruit bodies color best seen at the margin of thepileus), only somewhat hygrophanous, with finebranching hygrophanous streaks. Lamellae: medium-spaced (50–65 reaching the stipe), emarginate,medium thick to thick, light brown when young, darkbrown with age; edge somewhat lighter, uneven. Stipe:6–9 3 0.6–1.3 cm (top), cylindrical, at the base clavate(22.7 cm) rarely wholly cylindrical, white fibrillose,later more grayish brown. Basal mycelium: white.Universal veil on stipe: vivid rose-red, forming somebelts or zones usually on the lower half of the stipe.Context: marbled with brown when fresh and moist,becoming whitish brown with moisture loss, darkertoward the base of the stipe. Odor: in lamellaeindistinct. Exsiccata: pileus fairly dark gray brown tobrown to blackish brown, rosered tints usually at leastin the margin; stipe grayish white, in old fruit bodiesdark; rose to vinaceous red universal veil still visibleon the stipe; basal mycelium white; sometimes thewhole exsiccatum almost blackish. Chemical reactions:not tested.
Spores. 6.6–7.7–8.6 3 5.0–5.5–6.1 mm, Q 5 1.22–1.39–1.56, av. 5 6.9–8.2 3 5.3–5.7 mm, Qav. 5 1.31–1.46 (320 spores, 16 collections, FIGS. 3D, 4), amyg-daloid to ovoid to obovoid-ellipsoid with a somewhattapering apex, and possibly a low suprahilar depres-sion (sometimes almost subglobose), thick-walled,separately, sharply verrucose, more strongly at theapex, strongly dextrinoid, dark, some spores becomeslightly blackish red in KOH. Spores on the lamellaeusually less dextrinoid and slightly larger, especiallyyoung spores may have dark umber contents. Lamellartrama hyphae: in the overall view olivaceous yellow toolivaceous brownish in MLZ and KOH, smooth todistinctly finely scabrous, often strongly guttulate,with olivaceous brownish spot-like incrustations.Basidia: four-spored, 29–35–41 3 7.5–8.1–9.0 mm,
av. 31–38 3 7.9–8.5 mm (110 basidia, 11 collections),with olivaceous yellow to olivaceous brown, sometimesgranulose contents in MLZ and KOH. Both the sporesand structures of the lamellae darken strongly withage. Lamellar edge mostly fertile with basidia and fairlywide, clavate marginal cells. Pileipellis: Epicutis medi-um thick; uppermost hyphae 4–10 mm wide, yellowishbrown to olivaceous yellowish brown in MLZ andKOH, smooth to finely scabrous; lower hyphae scanty,5–11 mm wide, almost hyaline, mostly distinctly finelyencrusted. Hypoderm weakly developed with elongat-ed elements about 38–55 3 15–25 mm, finelyencrusted, almost hyaline in KOH, olivaceous toreddish brown in MLZ. Hyphae in the transition totrama smooth to finely encrusted in KOH, weakly tostrongly spot-like encrusted in MLZ. Clamp connec-tions: present.
ITS regions (including 5.8S region). 519 bases long(based on four sequences, including the sequencefrom the type specimen). All sequences are identical.The difference compared to other Armillati-species ismore than 20 evolutionary events. Sequences retrievedfrom the public databases: No sequences of this speciesexist in public databases.
Ecology and distribution. Cortinarius roseoarmillatusgrows often in groups, sometimes solitary in mesic,mixed forests and in wooded gardens, presumablywith Betula. It often occurs on somewhat richer soil,especially in the northern parts of the distributionarea. The species might be somewhat continental,and so far it is known only from northern hemiborealand/or boreal zones of Sweden and Finland andconsidered rare (FIG. 8). It has been found mid-Aug–late Sep (FIG. 6).
Differential diagnosis. Cortinarius roseoarmillatusmacroscopically resembles C. paragaudis and C.pinigaudis, but in good condition it has rose-brownto pinkish brown pileus and rose-red universal veil. Itis distinguished most easily from the latter and otherspecies of the section however by its fairly small,amygdaloid to ovoid to obovoid-ellipsoid, thick-walledand strongly dextrinoid spores. Both the spores andlamellae structures typically darken strongly with age.
Comments. The epithet roseoarmillatus first wasintroduced by Niskanen and Kytovuori (2008), andthe species is now validly described here.
Specimens examined (a total of 19 collections, for thecomplete list of specimens see SUPPLEMENTARY TABLE III).SWEDEN. MEDELPAD: Stode, Vigge, Sep 1990, H.Lindstrom 90.272 (UPS), GenBank HQ845118. FINLAND.SATAKUNTA: Vammala, Perkojarvi, 30 Aug 1989, L.Kosonen (TUR), GenBank HQ845119. ETELA-SAVO: Kan-gasniemi, Luusniemi, 1 Sep 1990, K. Malmstrom 14 (H),GenBank HQ845120. POHJOIS-HAME: Virrat, Hauhuu, 16Aug 2004, leg. I.K. (T.N. 04-061) (H, HOLOTYPE, NY,ISOTYPE), GenBank HQ845117.
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1089

Cortinarius suboenochelis Kytov., Liimat. & Niskanensp. nov. FIGS. 2C, 3B, 4, 6, 9
MycoBank MB519557Cortinarius suboenochelis Kytov., Liimat. & Niskanen,
Funga Nordica: 727 (2008) (invalid)Cortinarius subarmillatus var. picetorum Moenne-
Locc., Atlas des Cortinaires 7:231 (1995).Type. FRANCE. HTE-SAVOIE: Merdassier, in Vac-
cinium-Picea community (no Betula), 1700 m, 2 Oct1989, P. Moenne-Loccoz 1239 (G part of holotype).GenBank HQ845129.
Pileus 3.5–10 cm, convexus, dein planoconvexus, areno-sobrunneus, dein brunneus, cum venis hygrophanis. La-mellae subconfertae, pallide caesiobrunneae, dein obscurebrunneae. Stipes 8–14 3 0.7–1.4 cm, cylindraceus velclavatus (22.5 cm), albide fibrillosus, apice caesiogriseo,velo roseo vel brunneoroseo, cingulato. Caro caesiobrun-nea, variegata. Odor nullus. Sporae 8.6–11.1 3 5.9–7.3 mm,amygdaliformes vel ellipsoidae, paulo verrucosae. In silvisconiferis mediocriter humidis, cum Picea abiete.
Typus. FINLAND. VARSINAIS-SUOMI: Vihti com-mune, Salmenkartano, mesic Picea forest with someBetula, Populus and Pinus, 30 Aug 2005, coll.Kytovuori (Niskanen 05-028) (H, HOLOTYPE; NY,ISOTYPE). GenBank HQ845121.
Illustrations. Bidaud et al. (1995: pl. 145 as C.subarmillatus var. picetorum).
Etymology. suboenochelis (Latin and Greek), sub 5
near. The epithet refers to the affinity with C.paragaudis ssp. oenochelis (5 C. luteo-ornatus).
Pileus. 3.5–10 cm, convex with incurved marginwhen young, soon plano-convex and sometimes with alow, small umbo; usually small especially at the centerwhen mature, finely fibrillose toward the edge, with
some veil remnants at margin, sand-brown, pale brown,to pale grayish brown when young, later more brown;young pilei sometimes with bluish tints; only somewhathygrophanous, with fine, dark hygrophanous streaks.Lamellae: medium-spaced to distant (40–48 reachingthe stipe), emarginate, thick, broad, bluish brownwhen young, dark yellowish brown with age, edgesomewhat lighter, uneven. Stipe: 8–14 3 0.7–1.4 cm(top), cylindrical, at least slightly clavate at the base(22.5 cm), whitish fibrillose, later more grayish brown,upper part with bluish tints vanishing with age. Basalmycelium: white to pale pinkish. Universal veil on stipe:vivid rose-pink to brownish pink, often forming someincomplete zones. Context: marbled with brown whenfresh and moist, becoming whitish brown with loss ofmoisture, bluish at the top of the stipe, darker towardthe base. Odor: in lamellae indistinct. Exsiccata: pileusgrayish brown to dark brown; stipe grayish to brownish;pinkish rose to brownish rose universal veil still visibleon the stipe; basal mycelium white to pinkish. Chemicalreactions: not tested.
Spores. 8.6–9.6–11.1 3 5.9–6.6–7.3 mm, Q 5 1.30–1.45–1.61, av. 5 8.9–10.5 3 6.2–7.0 mm, Qav. 5 1.33–1.55 (540 spores, 23 collections, FIGS. 3B, 4), (broad-ly) amygdaloid to ovoid-ellipsoid to ellipsoid, thin-walled, with more or less tapering apex, finely to fairlyfinely verrucose, more strongly at the apex, some-times warts slightly anastomosing, fairly faintly tomoderately dextrinoid, few spores becoming slightlybrownish red in KOH, but not all the specimensexhibited these characteristics. Spores on the lamel-lae (av. 5 9.1–10.3 3 6.1–6.9 mm, Qav. 5 1.37–1.60(440 spores, 22 collections) are somewhat narrowerthan those on the veil. The size and shape of thespores are variable both within and between thecollections. Lamellar trama hyphae: in the overall viewpale yellowish to pale olivaceous in MLZ and KOH,smooth to finely, densely scabrous sometimes with fewolivaceous spot-like incrustations. Basidia: four-spored, 33–40–48 3 8–9.6–11 mm, av. 37–46 3 9.0–10.1 mm (120 basidia, 12 collections), with olivaceousyellowish to olivaceous brownish, often somewhatgranulose contents in MLZ and KOH. Lamellar edgefertile to partly sterile with basidia and few toabundant narrowly clavate cells. Pileipellis: Epicutismedium thick; uppermost hyphae 4–10 mm wide, dirtyyellow to olivaceous brownish in MLZ and KOH,smooth to finely scabrous; lower hyphae 7–12(–15)mm wide, almost hyaline, smooth to finely encrusted.Hypoderm weakly developed, with elongated ele-ments about 45–65(–75) 3 17–25(–30) mm, thin-walled, hyaline, smooth or very finely encrusted.Hyphae in the transition to trama smooth to finelyencrusted in KOH, with spot-like incrustations inMLZ. Clamp connections: present.
FIG. 9. Distribution of Cortinarius suboenchelis in north-ern Europe according to the examined material.
1090 MYCOLOGIA

ITS regions (including 5.8S region): 522 bases long(based on nine sequences, including sequences fromtwo type specimens, although only ITS 1 region fromC. subarmillatus var. picetorum was sequenced success-fully). All sequences are identical without anypolymorphisms. The difference compared to C.luteo-ornatus is at least 13 evolutionary events and toC. paragaudis-complex at least 16. Sequences retrievedfrom the public databases: No sequences of this speciesexist in the public databases.
Ecology and distribution. Cortinarius suboenochelisoccurs in mesic to dryish coniferous forests mostly withPicea, but perhaps also with Pinus, often on somewhatricher soil. It usually grows in groups, but also solitary.It is fairly common in the boreal zone; in thehemiboreal zone it seems to be occasional or rare. Itis known in Norway, Sweden, Finland and montaneareas of France (FIG. 9). Cortinarius suboenochelis fruitsearly Aug–late Sep with its optimum in early Sep, acouple of weeks later than C. luteo-ornatus (FIG. 6).
Differential diagnosis. Cortinarius suboenochelis isrecognized by the medium to stout, fleshy fruit body,small-scaled pileus, vivid rose-pink to brownish pinkuniversal veil, and fairly large, slightly amygdaloid,moderately to fairly strongly dextrinoid spores,although the size of the spores varies. Bluish tintsare typical on the top of the stipe, which no otherEuropean Armillati species has, but vanishes easilywith age. The sister species, C. luteo-ornatus, is similarto C. suboenochelis, but has a more abundant palepinkish to vinaceous red, sometimes yellowish brownuniversal veil and often more reddish brown pileus. Inaddition spores of C. luteo-ornatus are broadly ovoidto subglobose, sometimes broadly ellipsoid withbroad, rounded apex, and slightly less dextrinoidand a bit more strongly and sharply verrucose than inC. suboenochelis. The difference in the force of theverrucosity, height and width of warts, between theapex and middle is smaller than in the latter. Thebirch-associated C. armillatus has vivid orange-reduniversal veil and more amygdaloid, thick-walled,strongly dextrinoid spores. C. paragaudis and C.pinigaudis have distinctly innately fibrillose pileus andsmaller spores (, 8 mm long).
Based on molecular and morphological data, C.suboenochelis is a distinct species. The epithetsuboenochelis was introduced in Niskanen and Kyto-vuori (2008) and the species is validly described here.The species was described first as a variety of C.subarmillatus, a species that grows with deciduoustrees in Japan and has bluish tints in the fruit body(Hongo 1965). We studied the type material of C.subarmillatus, which includes two whole and one halffruitbodies. The universal veil was orange-red tobrickred, and the spores were 9.5–11 3 6.8–7.5 mm,
ovoid, thick-walled, fairly finely, evenly verrucose andweakly dextrinoid, although something in the ex-siccatum seemed to hinder the dextrinoid reaction.The molecular studies did not succeed, despite theseemingly good condition of the type material. Basedon the morphology and ecology however we conclud-ed that C. subarmillatus must be a separate speciesfrom C. suboenochelis. In the original description of C.subarmillatus var. picetorum (Moenne-Loccoz in Bi-daud et al. 1995) the spores are described as 10–13.5(–14) 3 7.0–8.0(–8.5) mm, but based on ourmeasurements of the type material the spores weresmaller 10–10.5–11.2(–11.5) 3 6.3–6.8–7.3 mm, Q 5
1.48–1.54–1.64 (20 spores). This variety also lacksbluish tints, but this most likely is due to thecondition of the fruit bodies (the illustration showstwo mature fruiting bodies with cracked, dry pilei).The ITS1 sequence was identical to C. suboenochelis(ITS2 did not succeed).
Specimens examined (a total of 85 collections, for thecomplete list of specimens see SUPPLEMENTARY TABLE IV).NORWAY. OPPLAND: Nordle Land, Røste, 12 Sep 1984, K.Høiland (O), GenBank HQ845122. TROMS: Bardu, Seter-moen, 5 Aug 2006, K.L. & T.N. 06-019 (H), GenBankHQ845123. Storfjord, by the road from E8 Kilpisjarvi toSkibotn, 25 Aug 2006, K.L. & T.N. 06-071 (H), GenBankHQ845124. FINLAND. VARSINAIS-SUOMI: Vihti, Salmen-kartano, 30 Aug 2004, P.K. & I.K. 04-033 (H), GenBankHQ845125, loc. cit., 30 Aug 2005, leg. I.K. (T.N. 05-028) (H,HOLOTYPE), GenBank HQ845121. POHJOIS-KARJALA:Ilomantsi, Patrikka, 7 Aug 1987, J.-P. Jappinen (JOE),GenBank HQ845126. KAINUU: Suomussalmi, Huurun-vaaran lehto, 16 Sep 2005, T.N. et al. 05-126 (H), GenBankHQ845127. KOILLISMAA: Kuusamo, Oulanka, Ampuma-vaara, 17 Sep 2005, T.N. et al. 05-142 (H), GenBankHQ845128. FRANCE. HTE-SAVOIE: Merdassier, 2 Oct1989, P. Moenne-Loccoz 1239 (G, HOLOTYPE of C.subarmillatus var. picetorum), GenBank HQ845129.
Cortinarius luteo-ornatus (M.M. Moser) Bidaud,Moenne-Locc. & Reumaux, Atlas des Cortinaires7:229 (1995). FIGS. 2D, 3C, 4, 6, 10Basionym. Cortinarius armillatus var. luteo-ornatus
M.M. Moser, Schweiz. Z. Pilzk. 43:117 (1965). Type:AUSTRIA. TIROL: Pitztal (Tiefental), Betula, Pinuscembra and Picea excelsa, 13 Sep 1962, Moser 62/27(IB, HOLOTYPE). GenBank HQ845130.
Cortinarius paragaudis ssp. oenochelis H. Lindstr.,Cortin. Fl. Photogr. 2:33 (1992). Type: SWEDEN.JAMTLAND: Hallesjo, in coniferous forest on richground (Picea, Pinus), 8 Aug 1987, Lindstrom et al.CFP546 (S, HOLOTYPE). GenBank HQ845145.
Cortinarius oenochelis (H. Lindstr.) Bidaud, Moenne-Locc. & Reumaux, Atlas des Cortinaires 7: 230 (1995).(Nom. inval., Art. 33.3, full and direct reference tobasionym omitted: year [1992] not given).
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1091

Cortinarius paragaudis sensu Moser, Schweiz. Z.Pilzk. 43(8):118 (1965), and Kleine Krypt.-fl. 2 b/2, 4.Aufl.: 395 (1978).
Illustrations. Bidaud et al. (1995: pl. 155 as C.oenochelis), Brandrud et al. (1992: pl. B10 as C.paragaudis ssp. oenochelis).
Pileus. 3.5–10 cm, convex with an incurved marginwhen young, soon plano-convex and sometimes with alow, broad umbo, sometimes small at the center,finely fibrillose toward the edge, with some pinkishred veil remnants at margin, reddish brown toochraceous brown, only somewhat hygrophanous,with fine, branching hygrophanous streaks. Lamellae:medium-spaced (45–60 reaching the stipe), emargin-ate, medium thick to thick, light brown when young,darkening with age, edge somewhat lighter, uneven.Stipe: 6–17 3 0.8–1.5 cm (top), cylindrical, at leastslightly clavate at the base (22.5 cm), firm, whitefibrillose, later more grayish brown. Basal mycelium:white to pale pinkish. Universal veil on stipe: usuallypale to more vivid pinkish when young, sometimesyellowish brown, turning often vinaceous red to brick-red with age, rarely whitish, often fairly abundant,forming incomplete and complete zones. Context:marbled with brown when fresh and moist, becomingwhitish brown with moisture loss, darker toward thebase of the stipe. Odor: in lamellae indistinct.Exsiccata: pileus grayish brown to dark brown, rarelyblackish brown; stipe grayish white to brownish;pinkish to red universal veil still visible on the stipe;basal mycelium white to pinkish. Chemical reactions:KOH on universal veil on stipe purple.
Spores. 8.2–9.1–10.0 3 6.3–7.0–7.7 mm, Q 5 1.18–1.30–1.43, av. 5 8.4–9.7 3 6.4–7.4 mm, Qav. 5 1.26–1.38 (640 spores, 28 collections; FIGS. 3D, 4), broadlyovoid to subglobose, sometimes broadly ellipsoid,with a broad, rounded apex, thin-walled, finely tomoderately verrucose, more strongly at the apex,sometimes warts slightly anastomozing, faintly tomoderately dextrinoid, few spores becoming slightlybrownish red in KOH, but all specimens did notdisplay these characteristics. Spores on the lamellae(av. 5 8.4–9.9 3 6.3–7.3 mm, Qav. 5 1.32–1.40 (300spores, 15 collections) are slightly narrower thanthose on the veil. Lamellar trama hyphae: in theoverall view pale olivaceous yellowish to olivaceousbrownish in MLZ and KOH, smooth to finely, denselyscabrous, sometimes weakly granulose-guttulate, withsome olivaceous spot-like incrustations. Basidia: four-spored, 33–39–45 3 8.5–9.5–11 mm, av. 35–42 3 8.7–10.0 mm (120 basidia, 12 collections), with olivaceousyellow to olivaceous brown contents in MLZ andKOH. Lamellar edge: fertile with basidia and clavatecells. Pileipellis: Epicutis medium thick; uppermosthyphae 4–10 mm wide, yellowish brown to olivaceousyellowish brown in MLZ and KOH, smooth to finelyscabrous; lower hyphae 5–11 mm wide, almost hyaline,smooth to finely scabrous. Hypoderm weakly devel-oped with elongated elements about 40–60 3 15–25 mm, almost hyaline, finely encrusted. Hyphae inthe transition to trama smooth to finely encrusted inKOH, with olivaceous to sepia brownish, small tofairly large, spot-like incrustations. Clamp connections:present.
ITS regions (including 5.8S region): Total 522–524bases long (based on 16 sequences, includingsequences from two type specimens). Sequences fromthe type material of C. paragaudis ssp. oenochelis andC. armillatus var. luteo-ornatus differ from the othersequences in the one length intragenomic polymor-phism site. C. luteo-ornatus generally has nineintraspesific variation sites in the sequences, eight ofthose include base or length intragenomic polymor-phisms; therefore intermediate individuals also occur.The amount of variation is high for a species. Thespecimens also can be divided into two subgroupsbased on one base site. This is similar to the C.paragaudis complex, except we found no morpho-logical or ecological characteristics support thegroups. However a comparatively high variation ofspore sizes was found among collections of C. luteo-ornatus. Our finding is similar to the one reported bySuarez-Santiago et al. (2009) about C. gallurae, whereno correlation was found between molecular data andspore-size variation. The difference compared to C.suboenochelis is at least 13 and to C. paragaudis morethan 20 evolutionary events. Sequences retrieved from
FIG. 10. Distribution of Cortinarius luteo-ornatus innorthern Europe according to the examined material.
1092 MYCOLOGIA

the public databases (six sequences): All are fromBritish Columbia, Canada, and all (FJ157138,FJ039675, FJ039674, FJ157119, FJ717545) exceptU56030 were identical to our data. One sequence(FJ157119) was deposited under C. cacaocolor, a namethat belongs to a species in section Brunnei or Uracei.
Ecology and distribution. Cortinarius luteo-ornatusgrows in mesic to dry coniferous forests with Picea,possibly with Pinus, and in subalpine Betula forests,often on somewhat richer soil. It seems to prefer welldeveloped mesic Picea forests with a thick moss cover,often growing in groups, rarely solitary. Occursoccasionally in subarctic zone and commonly inboreal zone. It is known only in hemiboreal zone innorthern regions (FIG. 10), unlike reported in Niska-nen et Kytovuori (2008). C. luteo-ornatus is known inEurope and North America. It fruits mid-Jul–late Sepwith its optimum in mid-Aug, approximately a weekearlier than C. armillatus and a couple of weeksearlier than C. suboenochelis (FIG. 6).
Differential diagnosis. In the field C. luteo-ornatus ismost easily identified when it is robust and has anabundant pinkish to vinaceous red universal veil, andwhen old almost plane, red-brown pileus. A habitatwith conifers also aides identification, except in thesubarctic zone where it associates with Betula. Thebest characteristic however is the large, broadly ovoidto subglobose to broadly ellipsoid, slightly to moder-ately dextrinoid spores. The sister species, C.suboenochelis, that often grows in similar Picea-dominated habitats usually has bluish tints on/inthe top of the stipe, small, sand-brown to pale grayishbrown pileus at least when mature, and rose pinkishto brownish pink universal veil that forms only someincomplete girdles on the stipe. The spores areamygdaloid to ovoid-ellipsoid to ellipsoid, slightlymore dextrinoid and slightly less strongly verrucosethan in C. luteo-ornatus. Plus the difference in theforce of the verrucosity, height and width of wartsbetween the apex and the middle center is stronger.Cortinarius armillatus has small red scales on thepileus, amygdaloid to slightly ellipsoid, stronglydextrinoid spores and habitat with Betula. Cortinariusparagaudis and C. pinigaudis have a distinctly innatelyfibrillose pileus and smaller spores (, 8 mm long).Sometimes the universal veil of C. luteo-ornatus can bewhitish (Niskanen and Kytovuori 3 Aug 2001); thenthe exsiccatum might be difficult to distinguish fromspecies in sect. Brunnei.
Cortinarius luteo-ornatus originally was described byMoser (1965) as a variety of C. armillatus withyellowish to brownish yellow universal veil, less vividcolors, and shorter, slightly broader spores. Lind-strom (1992) considered the variety to be similar to C.paragaudis ssp. oenochelis H. Lindstr., except for the
brownish yellow veil and therefore did not synony-mise it with his ssp. oenochelis. Although C. luteo-ornatus usually has 6 pinkish universal veil that oftenturns vinaceous red to brickred with age, we havefound fruit bodies with brownish yellow veil (Niska-nen and Liimatainen 06-053). We studied both typesand additional material from Fennoscandia. Thesequence data and morphology show that C. luteo-ornatus is separate from C. armillatus and C.paragaudis and identical to Cortinarius paragaudisssp. oenochelis H. Lindstr.
Cortinarius paragaudis Fr. sensu Moser (1965 and1978) most likely also represents C. luteo-ornatus.Previous treatments of the latter also might include C.suboenochelis because the species was not known; forexample the collection Kytovuori et al. 7 Sep 2001,identified as C. ‘‘oenochelis’’ in Kytovuori et al. (2002),is in fact C. suboenochelis, and in the comments on C.luteo-ornatus (as C. paragaudis) Soop (2004) men-tions that occasionally the whole fruit body has apurple tint, typical for C. suboenochelis but lackingfrom C. luteo-ornatus.
Specimens examined (a total of 172 collections, for thecomplete list of specimens see SUPPLEMENTARY TABLE V).NORWAY. BUSKERUD: Gol, skogen ost For Svartevann, 5Oct 1957, J. Stordal 11095 (O), GenBank HQ845131.TROMS: Bardu, Setermoen, 5 Aug 2006, K.L. & T.N. 06-018 (H), GenBank HQ845132, loc. cit., K.L. & T.N. 06-020(H), GenBank HQ845133. Malselv, about 5 km S from thecenter of Skjold near Road 87, 8 Aug 2006, K.L. & T.N. 06-026 (H), GenBank HQ845134 , loc. cit., 10 Aug 2006, K.L.& T.N. 06-053 (H), GenBank HQ845135. Storfjord,Lullesletta, 3 Aug 2001, H. Tuovila & I.K. 01-027 (H),GenBank HQ845136, loc. cit., H. Tuovila & I.K. 01-028a(H), GenBank HQ845137, loc. cit., H. Tuovila & I.K. 01-028b (H), GenBank HQ845138. SWEDEN. JAMTLAND:Hallesjo, 1 km S Ansjo, 8 Aug 1987, H. Lindstrom et al. CFP546 (S, HOLOTYPE of C. paragaudis ssp. oenochelis),GenBank HQ845145. Ostersund, OSK-omradet, 13 Aug1991, S. Jacobsson (GB), GenBank HQ845139. FINLAND.VARSINAIS-SUOMI: Suomusjarvi, Lahnajarvi, 21 Sep 1998,I.K. 98-2212 (H), GenBank HQ845140. ETELA-HAME:Vilppula, Elamanmaki, 16 Sep 1999, J. Vauras 15585(TUR), GenBank HQ845141. PERA-POHJANMAA: Tornio,Arpela, Korkiamaa, 30 Aug 2004, K.L. & T.N. 04-526 (H),GenBank HQ845142. Simo, Alaniemi, 23 Jul 1979, E.Ohenoja (OULU), GenBank HQ845143. SOMPION LAPPI:Sodankyla, Tankajoki, 13 Aug 1995, I.K. 95-300 (H),GenBank HQ845144. AUSTRIA. TIROL: Pitztal (Tiefental),13 Sep 1962, Moser 62/27 (IB, HOLOTYPE of C. armillatusvar. luteo-ornatus), GenBank HQ845130.
Cortinarius paragaudis Fr., Epicr. Syst. Mycol.: 295(1838). FIGS. 2E, 3E, 4, 6, 11Type. NORWAY. SØR-TRØNDELAG: Røros, in wet
forest with Pinus and Betula on rich ground, 16 Aug1987, Lindstrom et al. CFP564 (S, NEOTYPE, desig-
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1093

nated in Cortin. Fl. Photogr. II [Swedish version]:22,1992). GenBank HQ845146.
Cortinarius bresadolianus Moenne-Locc. & Reu-maux, Atlas des Cortinaires 7:228 (1995)—Type:FRANCE. LOIRET: Montargis, under Picea abies,150 m, 25 Oct 1983, J. Poirier 930 (G, HOLOTYPE).GenBank HQ845159.
Cortinarius vaccinochelis Chevassut & Rob. Henry,Documents Mycologiques 8:52 (1978)—Type: Hen-ry 70676 (PC, HOLOTYPE). GenBank HQ845161.
Cortinarius haematochelis sensu Moser, Schweiz. Z.Pilzk. 43(8):119 (1965), Kleine Krypt.-fl. 2 b/2, 4.Aufl.: 395 (1978).
? Cortinarius salmoneotomentosus var. pseudoparagau-dis Bidaud, Moenne-Locc. & Reumaux, Atlas desCortinaires 7:230 (1995).Type. FRANCE. ARDENNES: Foret de Belval, on
argillaceo-calcareous soil under Quercus, 200 m, 9 Oct1992, P. & F. Reumaux 3120 (G, HOLOTYPE).GenBank HQ845160.
Illustrations. Bidaud et al. (1995: pl. 151 as C.bresadolianus), Brandrud et al. (1992: pl. B32).
Pileus. 3–6(–7) cm, convex with an incurved marginwhen young, plano-convex with a small umbo whenold, distinctly innately fibrillose, often with vinaceousred veil patches in the margin, grayish brown toochraceous brown, only somewhat hygrophanous,with hygrophanous streaks. Lamellae: medium-spaced(45–55 reaching the stipe), emarginate, mediumthick to thick, light brown when young, dark brownwith age, edge somewhat lighter, uneven. Stipe: 6–153 0.6–1.2 cm (top), cylindrical, at the base clavate
(22.5 cm) rarely wholly cylindrical, usually slender,white fibrillose when young, later grayish brown.Basal mycelium: white or pale pinkish. Universal veilon stipe: vinaceous red, sometimes yellowish brown inyoung fruit bodies, usually forming distinct belts orzones or several spots. Context: marbled with graybrown when fresh and moist, becoming whitish brownwith moisture loss, darker toward the base of thestipe. Odor: in lamellae slightly raphanoid. Exsiccata:pileus brown to dark gray brown, rarely blackishbrown; stipe grayish to grayish brown; vinaceous reduniversal veil still visible on the stipe; basal myceliumoften pinkish or with pinkish tints, sometimescompletely white. Chemical reactions: KOH on univer-sal veil on stipe purple.
Spores. 6.3–7.0–7.7 3 5.2–5.8–6.3 mm, Q 5 1.14–1.19–1.27, av. 5 6.4–7.5 3 5.5–6.3 mm, Qav. 5 1.16–1.24 (340 spores, 17 collections, FIGS. 3E, 4), broadlyovoid to subglobose, wall medium thick, moderately,fairly coarsely verrucose, somewhat more strongly atthe apex, almost indextrinoid to fairly slightlydextrinoid, somewhat dark, some spores becomingslightly brownish red in KOH, not observed in all thespecimens. Spores on the lamellae usually narrowerand more ovoid than those on the veil. Lamellartrama hyphae: in the overall view pale olivaceousyellowish to olivaceous brownish in MLZ and KOH,almost smooth to distinctly finely, densely scabrous,sometimes guttulate, with some olivaceous spot-likeincrustations. Basidia: four-spored, 26–31–36 3 7.0–7.8–8.5 mm, av. 28–33 3 7.2–8.4 mm (110 basidia, 11collections), with dirty olivaceous to olivaceous browncontents in MLZ and KOH. Pileipellis: Epicutismedium thick; uppermost hyphae 4–9 mm wide,olivaceous yellowish brown in MLZ and KOH, smoothto finely scabrous or with small to large spots; lowerhyphae 5–11 mm wide, almost hyaline to yellowishbrown, finely to more strongly encrusted. Hypodermweakly developed with elongated elements about 40–60 3 15–23 mm, almost hyaline to pale yellowishbrownish, fairly thin-walled, finely encrusted. Hyphaein the transition to trama smooth to finely encrustedin KOH, often with fairly abundant spot-like incrus-tations in MLZ. Clamp connections: present.
ITS regions (including 5.8S region): 518–524 baseslong (based on 14 sequences, including sequencesfrom four type specimens, although only ITS 1 regionof the type material of C. vaccinochelis was obtained).The C. paragaudis neotype differs from all other typesequences with one base site and C. bresadolianus withone length intragenomic polymorphism site. C.paragaudis generally has seven infraspecific variationsites, all except one with intragenomic polymor-phisms. Compared to C. pinigaudis only one differ-ence exists: In one base site C. pinigaudis has C and
FIG. 11. Distribution of Cortinarius paragaudis innorthern Europe according to the examined material.
1094 MYCOLOGIA

C. paragaudis has T. Two sequences (C. paragaudisclade II, FIG. 1), which formed a monophyletic groupwith high PP value, were identical and had three basechanges and one indel compared to others. Thedifference with C. suboenochelis is at least 16 evolu-tionary events and with C. luteo-ornatus more than 20.Sequences retrieved from the public databases (threesequences): Two of the sequences were from Sweden(UDB001169, UDB001170) and one from BritishColumbia, Canada, (DQ481804). Based on the sitein ITS1 region, which distinguishes C. paragaudisfrom C. pinigaudis, the retreived public databasesequences were determined as C. paragaudis, despitethe presence of additional variation. In additionUDB001169 seemed to be C. paragaudis, regardlessof its Pinus association.
Ecology and distribution. Cortinarius paragaudisoccurs in a wide range of habitats from submesic todamp Picea forests, dryish coniferous forests (Picea,Pinus), subalpine Betula forests and presumably alsotemperate Quercus forests. The most characteristichabitat is submesic to damp Picea forest. In oceanicareas it also occurs in dryish Pinus forests mixed withBetula. It is common in boreal zone, common tooccasional in hemiboreal zone, occasional in subarc-tic zone and rare in temperate zone (FIG. 11). It isknown from Europe and North America. Fruit bodiesoccur late Jul–early Oct, but the peak of the fruitingseason is late Aug–early Sep (FIG. 6).
Differential diagnosis. Cortinarius paragaudis variesgreatly in appearance, it can be slender or stout andthe vinaceous red universal veil can be distinct oralmost indistinct. Typical for the species however arethe distinctly innately fibrillose pileus and broadlyovoid to subglobose, non-dextrinoid to slightlydextrinoid and somewhat dark spores. Cortinariuspinigaudis, C. roseoarmillatus, and slender C. luteo-ornatus are similar to C. paragaudis, but C. pinigaudishas smaller (6.1–6.6–7.0 3 4.7–5.2–5.7 mm), roundedtriangular ovoid to ventrally flattened subglobosespores with weakly thinner and lighter wall, andslightly weaker and sharper verrucosity. In addition C.pinigaudis mostly occurs in dry to dryish habitats withPinus. Cortinarius roseoarmillatus has rose-red tints inthe pileus and universal veil, and amygdaloid to ovoidto obovoid-ellipsoid, thick-walled, and strongly dex-trinoid spores, while C. luteo-ornatus has distinctlylarger spores (8.2–9.1–10.0 3 6.3–7.0–7.7 mm) andtypically soon plano-convex, less fibrillose pileus.
Cortinarius fillioni Bidaud, Moenne-Locc. & Reu-maux and C. raphanoides (Pers.) Fr. also can resembleC. paragaudis because they have finely innatelyfibrillose pileus and colorful universal veil, which isusually yellowish brown to olivaceous brown. Further-more the former two have broadly ellipsoid to almost
subglobose, moderately to strongly dextrinoid sporesand large olivaceous spots deep in the pileipellis.Cortinarius suberi Soop with an innately fibrillosepileus and commonly observed pinkish tint in theuniversal veil in an exsiccatum can resemble C.paragaudis. C. suberi however has a whitish universalveil when fresh and ellipsoid to somewhat amygdaloidspores (7.5–9.0 3 5.0–5.5 mm).
The small-spored Brunnei species, C. ectypus J.Favre and C. albogaudis Kytov., Niskanen & Liimat.,also with innately fibrillose pileus, sometimes can beconfused with C. paragaudis; for example in Bran-drud et al. (1992) the photographed collection of C.ectypus (pl. B28, CFP710) includes four C. paragaudisfruiting bodies: two fruit bodies on the right (the 2ndand 3rd), the lower right pileus of the three pilei andone slender half fruit body in the middle, butexsiccata are blackish and the universal veil isindistinct or brownish white. Spores are about thesame size as in C. paragaudis, but those of C. ectypusare finely verrucose (5.5–7.0 3 4.5–5.5 mm) and thoseof C. albogaudis obovoid-subglobose and echinate-verrucose (6.0–7.0 3 5.0–5.5 mm). The vinaceous red-veiled C. badiovinaceus M.M. Moser has a hygropha-nous pileus, yellow brown context, and yellow,somewhat encrusted lamellar trama hyphae.
The nomenclature of C. paragaudis is extensivelydiscussed by Lindstrom (1992), who also proposed aneotype for the species. The chosen neotype repre-sents a small-spored, conifer-associated species. C.paragaudis is a small species resembling C. brunneus,similar to C. haematochelis, and grows in damp forests(‘‘Locis humidis pinetorum … inter Sphagna’’), ac-cording to Fries (1838). We think the species chosenfor the neotype fits Fries’ description well and theselection is well founded. Furthermore the othercommon red-veiled species, C. luteo-ornatus, has alargely boreal distribution and at the time ofdescription Fries worked in Uppsala so he most likelyencountered this species, a fact already brought up byLindstrom (1992).
At least two species from France, C. bresadolianus(spores from the type 6.6–7.0–7.1 3 5.5–5.7–5.9 mm,Q 5 1.18–1.22–1.25(–1.27)) and C. vaccinochelis(spores from the type [6.1–]6.5–6.8–7.1[–7.4] 3
[5.3]5.5–5.7–6.2 mm, Q 5 [1.11–]1.14–1.20–1.25[–1.32]), based on our studies, are synonymous withC. paragaudis, which most likely is due to the variableappearance, commonness as well as the wide distri-bution of the species. The color of the exsiccata,spores, ecology, and ITS regions fit well with C.paragaudis. Cortinarius salmoneotomentosus Rob. Hen-ry (1977) also might belong in this group, but withoutavailable type material this could not be determinedsolely by the Latin description. The recorded spore is
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1095

6.5 3 5.2–7.9 3 5.8–6.8 mm, somewhat larger than inour material, and the habitat is montaneous conifer-ous and deciduous forest ‘‘In silvis abiegnis velnemorosis montanis’’. A variety C. salmoneotomentosusvar. pseudoparagaudis was described by Bidaud et al.(1995). The exsiccatum of the holotype was otherwiseC. paragaudis-like, but the base of the stipe wasorange-red. Furthermore the spores were small (6.7–7.1–7.4 3 5.2–5.5–5.7 mm, Q 5 1.23–1.29–1.34 [20spores]) but more ovoid and/or somewhat moreelongated than in C. paragaudis, and the holotypewas collected under Quercus. The ITS regions weresimilar to C. paragaudis. We also studied onecollection (JV88-362, GenBank HQ845147) fromDenmark from a temperate deciduous forest col-lected under Quercus and Populus. Based on mor-phology and DNA data, it represented a typical C.paragaudis. Therefore we concluded C. salmoneoto-mentosus var. pseudoparagaudis is most likely asynonym for C. paragaudis, but more material fromQuercus forests should be studied.
Cortinarius haematochelis (Bull.) Fr. sensu Moser(1965, 1978) most likely represents our species aswell. It has small spores, 5.5–7(–8) 3 5–6(–6.5) mm,and shares a habitat with coniferous and deciduoustrees (Moser 1978). The name haematochelis has beeninterpreted in various ways. Fries (1838) considered itas a rare species in Fagus forests in the vicinity of C.craticius and with a C. brunneus-like habitus, andBidaud et al. (1995) used the name for a C. armillatus-like species. The species epithet, haematochelis, wasvalidly published by Bulliard in 1791, but the reportincluded only a Latin name and plate(t. 527). The plate clearly represents a species ofsection Armillati and includes illustrations of fourfruiting bodies in all developmental stages. The statureof fruiting bodies is C. armillatus-like, but the reddishuniversal veil is scanty and forms only one band on thestipe. From the illustration it is impossible to say forcertain which species it represents. Bulliard includedanother plate (1793, t. 596), which likely represents C.armillatus. In 1812 he reported a description with areference to both plates (Bulliard and Ventenat 1812)but included no new information to aid interpretation.Bulliard studied fungi in the Paris region wheredeciduous forests are dominant (Lindstrom 1992).Based on our current knowledge at least two Armillatispecies could come into question, C. armillatus and arepresentative of the C. paragaudis group (C. haema-tochelis ss. Moser p.p., C. salmoneotomentosus p.p., C.salmoneotomentosus var. pseudoparagaudis, see above).Because the name cannot be interpreted unambigu-ously and it has been used in different ways by differentauthors it should be excluded, as suggested byLindstrom (1992).
In our phylogenetic analysis two specimens, Schul-mann 23 Aug 1954 (H) and Niskanen et al. 02-273 (H),formed a monophyletic group with a high PP value (C.paragaudis clade II, FIG. 1), and they differed from theother C. paragaudis specimens by four evolutionaryevents. C. paragaudis clade II could represent aseparate species, based on the ITS data. No notes onmacromorphology exist, but C. paragaudis clade IIspores were somewhat larger, more dextrinoid and lessovoid than generally found in C. paragaudis (6.7–7.8 3
5.9–6.4(–6.7) mm, Q 5 1.15–1.26, av. 5 7.3–7.4 3
6.1 mm, Qav. 5 1.17–1.20 (40 spores, two collections).The spores of the collection Liimatainen & Niskanen06-057 also were larger (7.0–7.5–7.9 3 5.9–6.3–6.8 mm,Q 5 1.15–1.19–1.25) than generally in C. paragaudis;however, although based on ITS regions, it was closer toC. paragaudis than the other two large-spored collec-tions (the quality of the sequence was poor due to thetwo intragenomic length polymorphisms and was notincluded in the phylogenetic analysis). Of interest theuniversal veil of Liimatainen & Niskanen 06-057 wasvivid pink reddish unlike typically in C. paragaudis.Theese non-sequenced specimens also had somewhatlarger spores than generally in C. paragaudis ingeneral: P. & I. Kytovuori 09-667 (H), 09-870 (H) andKytovuori 08-1254 (H). Further studies are needed toclarify the taxonomy of the group.
Specimens examined (a total of 148 collections, for thecomplete list of specimens see SUPPLEMENTARY TABLE VI).DENMARK. WESTERN JYLLAND: Strasø Plantage E ofUlfborg, 17 Aug 1983, A. Nielsen & J. Vesterholt 88-362 (C),GenBank HQ845147. NORWAY. SØR-TRØNDELAG:Røros, 16 Aug 1987, Lindstrom et al. CFP564 (S, NEO-TYPE), GenBank HQ845146. TROMS: Malselv, about 5 kmS from the center of Skjold near Road 87, 10 Aug 2006, K.L.& T.N. 06-027 (H), GenBank HQ845148. SWEDEN.JAMTLAND: Undersaker, Edsasdalen, 14 Aug 1988, H.Lindstrom et al. CFP710 (S, as C. ectypus), GenBankHQ845149. FINLAND. VARSINAIS-SUOMI: Karkkila, Tuor-ila, 23 Aug 1954, O. v. Schulmann (H), GenBankHQ845157. ETELA-HAME: Virrat, Monoskyla, 22 Aug1996, I.K. 96-552 (H), GenBank HQ845150. POHJOIS-KARJALA: Vartsila, Kaustajarvi, 31 Aug 1996, I.K. 96-741(H), GenBank HQ845151. KAINUU: Puolanka, Vayryla, 15Sep 1997, IK 97-1492 (H), GenBank HQ845152. KOILIS-MAA: Kuusamo, Liikasenvaara. 29 Aug 2002, K.L. & T.N.02-273 (H), GenBank HQ845158. PERA-POHJANMAA:Rovaniemi, Louevaara, 29 Aug 2004, K.L. & T.N. 04-456(H), GenBank HQ845153. KITTILAN LAPPI: Kolari,Lappea, 18 Aug 1998, I.K. 98-1132 (H), GenBankHQ845154. INARIN LAPPI: Utsjoki, Kevo, 16 Aug 1995,I.K. 95-518 (H), GenBank HQ845155. FRANCE. Henry70676 (PC, HOLOTYPE of C. vaccinochelis), GenBHQ845161. JURA: Arbois, 2 Oct 1998, H. Lindstrom98.945 (UPS), GenBank HQ845156. LOIRET: Montargis,25 Oct 1983, J. Poirier 930 (G, HOLOTYPE of C.bresadolianus), GenBank HQ845159.
1096 MYCOLOGIA

Cortinarius pinigaudis Niskanen, Kytov. & Liimat., sp.nov. FIGS. 2F, 3F, 4, 6, 12
MycoBank MB519558Cortinarius pinigaudis Niskanen, Kytov. & Liimat.,
Funga Nordica: 728 (2008) (invalid)Pileus 3–8 cm, convexus, dein planoconvexus, ochraceo-
brunneus vel griseobrunneus, cum venis hygrophanis.Lamellae subconfertae, pallide brunneae, dein obscurebrunneae. Stipes 7–11(–15) 3 0.8–1.5 cm, clavatus(23.0 cm), griseoalbide fibrillosus, velo saepe primobrunneoochraceo, dein vinaceo, cingulato. Caro brunnea,variegata. Odor nullus vel obsolete raphanoideus. Sporae6.1–7.0 3 4.7–5.7 mm, ovatae vel subglobosae, pauloverrucosae. In silvis coniferis aridis et semiaridis, cum Pinosylvestri.
Typus. FINLAND. KOILLISMAA: Kuusamo com-mune, Oulanka National Park, dry Pinus heath foreston sandy soil, 17 Sep 2005, Liimatainen & Niskanen05-133 (H, HOLOTYPE; NY, ISOTYPE). GenBankHQ845162.
Etymology. pinigaudis (Latin), Pinus 5 pine,gaudium 5 joy, rejoicing, pleasure. The epithet refersto the affinity with C. paragaudis and habitat withpine.
Pileus. 3–8 cm, convex with an incurved marginwhen young, plano-convex with a low, broad umbowhen old, moderately innately fibrillose, sometimeswith yellowish to reddish veil patches in the margin,grayish brown to yellowish brown, only somewhathygrophanous, with hygrophanous streaks. Lamellae:medium-spaced (45–65 reaching the stipe), emargin-ate, medium thick to thick, light brown when young,dark brown with age, with somewhat lighter, unevenedge. Stipe: 7–11(–15) 3 0.8–1.5 cm (top), clavate
(23.0 cm), rarely cylindrical, firm, grayish whitefibrillose when young, later grayish brown. Basalmycelium: white to pale pinkish. Universal veil on stipe:sparse, yellowish brown when young, later morevinaceous red, usually forming only one or a fewzones on the lower half of the stipe. Context: marbledwith gray-brown when fresh and moist, becomingwhitish brown with loss of moisture, darker toward thebase of the stipe. Odor: in lamellae slightly raphanoid.Exsiccata: pileus brown to dark gray brown to blackishbrown, stipe grayish to brownish, red universal veilhardly visible on the stipe, basal mycelium white orsometimes pinkish. Chemical reactions: universal veilon the stipe purple with KOH.
Spores. 6.1–6.6–7.0 3 4.7–5.2–5.7 mm, Q 5 1.21–1.28–1.36, av. 5 6.3–6.8 3 5.0–5.4 mm, Qav. 5 1.25–1.35 (280 spores, 14 collections, FIGS. 3F, 4), roundedtriangular ovoid to ventrally flattened subglobose; wallmedium thick, fairly finely to moderately, sharplyverrucose, somewhat more strongly at the apex,indextrinoid to faintly dextrinoid but slightly dark;few spores becoming slightly brownish red in KOH,not observed in all specimens. Spores on the lamellaeusually narrower and more ovoid than those on theveil. Lamellar trama hyphae: in the overall view paleolivaceous yellowish to pale olivaceous brownish inMLZ and KOH, smooth to finely, densely scabrous,sometimes guttulate, with few olivaceous brownishspot-like incrustations. Basidia: four-spored, 25–30–36 3 6.5–7.2–8.5 mm, with dirty olivaceous-browncontents in MLZ and KOH. Lamellar edge fertile withbasidia and narrowly clavate cells, sometimes palebluish or turquoise. Pileipellis: Epicutis medium thick;uppermost hyphae 3–8 mm wide, (olivaceous) yellow-ish brown in MLZ and KOH, smooth to finelyscabrous; lower hyphae 5–11 mm wide, fairly scanty,almost hyaline, finely scabrous. Hypoderm weaklydeveloped with elongated elements about 40–65 3
15–25 mm, almost hyaline, fairly thin-walled, finelyencrusted. Hyphae in the transition to trama smoothto finely encrusted in KOH, smooth to finely spot-likeencrusted in MLZ. Clamp connections: present.
ITS regions (including 5.8S region): 518–520 baseslong (based on seven sequences, including thesequence from the type specimen). The sequenceshave two intragenomic polymorphism sites. Only onenucleotide change exists between C. paragaudis andC. pinigaudis. Sequences from C. pinigaudis share a Cin this position, while C. paragaudis collections sharea T. The difference compared to C. suboenochelis is 16evolutionary events and to C. luteo-ornatus more than20. Sequences retrieved from the public databases: Noneexist.
Ecology and distribution. Cortinarius pinigaudisoccurs in dry to dryish, oligotrophic coniferous forests
FIG. 12. Distribution of Cortinarius pinigaudis in north-ern Europe according to the examined material.
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1097

with Pinus. Most characteristic to dry pine heaths onsandy soil. It also is found but much less commonly inrocky pine forests and submesic spruce forests mixedwith pine. It grows in loose groups, rarely solitary. It iscommon in the boreal zone but becomes lesscommon toward the south, and in the hemiborealzone it is recorded only from the northern parts(FIG. 12). It might be somewhat continental. C.pinigaudis is so far known from Norway, Swedenand Finland. It fruits early Aug–early Oct with itsoptimum in late Aug and early Sep (FIG. 6).
Differential diagnosis. Typical for Cortinarius pini-gaudis is the innately fibrillose pileus, often clavateand stout stipe, when young yellowish brown, latermore vinaceous red universal veil usually formingonly some zones on the lower half of the stipe, androunded triangular ovoid to ventrally flattened sub-globose spores. The sister species, C. paragaudis, ismost easily distinguished from C. pinigaudis by themore roundish, stronger verrucose and somewhatlarger spores with a slightly thicker wall (6.3–7.0–7.73 5.2–5.8–6.3 mm, FIG. 3), often with a somewhatdifferent ecology and more abundant universal veil.Cortinarius pinigaudis grows in dry to dryish Pinusdominated forests, whereas C. paragaudis has a widerecological range from submesic Picea-dominatedforests to subalpine Betula forests. In oceanic areasthe latter also occurs in dryish Pinus forests mixedwith Betula. The ITS sequences of the two species donot differ much, only one site in the ITS 1 region isdifferent. Nevertheless C. pinigaudis formed a mono-phyletic group in our phylogenetic analysis, althoughtwo distinct groups were not formed. Based on thecorrelation of morphological and ecological differ-ences however, we conclude that C. paragaudis and C.pinigaudis represent two different species. Theepithet pinigaudis was introduced in Niskanen andKytovuori (2008) and is now validly described here.
All other Armillati species have larger spores thanC. pinigaudis. In addition C. roseoarmillatus hasstrongly dextrinoid spores and rose-red to pinkishtints in the pileus and universal veil, and C. luteo-ornatus and C. suboenochelis have less clavate stipe. InPinus-dominated forests C. suberi Soop with innatelyfibrillose pileus can resemble C. pinigaudis. Thesometimes observed faint pinkish tint in the universalveil in its exsiccata also can lead to confusion withArmillati species. C. suberi however has a whitishuniversal veil when fresh and ellipsoid to somewhatamygdaloid spores (7.5–9.0 3 5–5.5 mm).
Specimens examined (a total of 109 collections, for thecomplete list of specimens see SUPPLEMENTARY TABLE VII).NORWAY. HEDMARK: Alvdal, Sollia, 4 Sep 1980, E.Bendiksen & R. Halvorsen EB & RH 669/80 (O), GenBankHQ845163. FINLAND. ETELA-KARJALA: Anjalankoski,
Kaipiainen, 19 Sep 1994, I.K. 94-801(H), GenBankHQ845165. ETELA-HAME: Leivonmaki, Selanpohja, 10Aug 1996, I.K. 96-435(H), GenBank HQ845164. KOILLIS-MAA: Kuusamo, Oulanka Biological Station, 26 Aug 2002,K.L. & T.N. 02-092, 02-093 (H), GenBank HQ845166,HQ845167, loc. cit., 17 Sep 2005, T.N. et al. 05-133 (H,HOLOTYPE, NY, ISOTYPE), GenBank HQ845162. INARINLAPPI: Inari, Lutto, 14 Aug 1995, I.K. 95-351(H), GenBankHQ845168.
KEY TO THE SPECIES OF SECT. ARMILLATI IN
NORTHERN EUROPE
1. Spores on average . 8.5 mm long . . . . . . . . . . . . . 21.9 Spores on average , 8.5 mmlong . . . . . . . . . . . . . . 4
2. Spores strongly dextrinoid, thick-walled; univer-sal veil vivid orange red; with Betula, possiblyalso with Carpinus . . . . . . . . . . . 1. C. armillatus
2.9 Spores weakly to moderately dextrinoid, thin-walled; universal veil usually 6 pinkish tovinaceous red, sometimes yellowish brown; withconifers . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3
3. Spores broadly ovoid to subglobose, sometimesbroadly ellipsoid, with broad, rounded apex, Q-value on average , 1.35; fruit body without bluishtints . . . . . . . . . . . . . . . . . . . . . . 3. C. luteo-ornatus
3.9 Spores (broadly) amygdaloid to ovoid-ellipsoid toellipsoid with a more or less tapering apex, Q-valueon average . 1.35; fruit body often with bluish tintson the top of the stipe and/or lamellae, especiallywhen young . . . . . . . . . . . . . . . . 4. C. suboenochelis
4. Spores amygdaloid to ovoid to obovoid-ellip-soid, thick-walled, strongly dextrinoid; universalveil rose-red . . . . . . . . . . . 2. C. roseoarmillatus
4.9 Spores ovoid to subglobose, fairly thin-walled,indextrinoid to weakly dextrinoid; universal veilvinaceous red or yellowish brown . . . . . . . . . . 5
5. Spores on average $ 5.4 mm wide; in mesic to dryishconiferous forests and in subalpine Betula for-ests . . . . . . . . . . . . . . . . . . . . . . . . 5. C. paragaudis
5.9 Spores on average # 5.4 mm wide; in dry Pinusforests and in submesic mixed forests with Pinus. . . . . . . . . . . . . . . . . . . . . . . . . . . . 6. C. pinigaudis
DISCUSSION
Armillati.—Section Armillati includes at least sixspecies in northern Europe. Two deciduous forestspecies, C. subarmillatus (Japan) and C. quercoarmil-latus (Costa Rica), also are known. All northernEuropean species are medium-sized to large, thepileus is 6 innately fibrillose and/or with small scales,only somewhat hygrophanous, and most often withhygrophanous streaks. The universal veil is pinkish tovinaceous red or in some species yellowish brown orrose-brown at least when young, the context ismarbled brown to gray-brown, and the mycelium is
1098 MYCOLOGIA

whitish to pale pinkish. Brandrud et al. (1989) andHøiland (1983) stated that spores turn reddish inKOH. We also observed this, but usually only somespores became reddish and it did not occur in allspecimens. The reddening was strongest in the twospecies with thick-walled spores, C. armillatus and C.roseoarmillatus.
The species of section Armillati have some distin-guishing macromorphological characteristics but,especially in C. paragaudis, intraspecific variationcan make identification in the field difficult. All thespecies however have typical basidiospore size, shapeand dextrinoidity based on which they can be reliablyidentified. The macromorphological characteristicsare best seen in young to middle-aged fruiting bodies,but basidiospores seem to develop fairly late, so onlymature fruiting bodies should be used for sporestudies.
The species grow in association with coniferousand/or deciduous trees. Locally they are associatedwith certain tree species, but for at least some of themhost association can vary depending on locality. In theboreal zone Cortinarius paragaudis and C. luteo-ornatus associate with conifers but in the subalpinezone with Betula spp. Betula-associated C. armillatusalso may grow with Carpinus spp. in temperate zones.Most of the species are considered common, al-though more than 1000 collections have beenexamined to date; C. roseoarmillatus is known onlyfrom Sweden and Finland. Cortinarius luteo-ornatusand C. pinigaudis seem to have boreal distributions,and C. luteo-ornatus also occurs in montane areas ofcentral Europe and the subalpine zone of northernEurope. The majority of Armillati species typicallystart fruiting early, often in late Jul or early Aug, theearliest species being C. armillatus, C. luteo-ornatus,and C. paragaudis, which have been recorded to fruitin mid–late July. Although peak fruiting occurs in thebeginning of the Cortinarius season, most species alsoare found in the autumn.
Only three sect. Armillati species were representedin public sequence databases, indicating poor cover-age of the group. In addition 65% of the sequenceswere deposited under a wrong name or as Cortinariussp. A similar situation was observed for sequences ofsect. Brunnei by Niskanen et al. (2009). The datahowever revealed that at least C. armillatus, C. luteo-ornatus, and C. paragaudis occur in British Columbia,Canada.
Although sect. Armillati is one of the most studiedgroups of subgenus Telamonia, two of the six specieswere unknown (33%). In section Brunnei theproportion of unknown species was 50% (Niskanenet al. 2009). Furthermore, based on morphologicaland molecular data, several new Telamonia species
representing various sections have been described(Kytovuori et al. 2005, Lindstrom et al. 2008,Niskanen et al. 2008a, Niskanen et al. 2009),reflecting the extent of the lack of information ongenus Cortinarius in Europe.
Section and species-taxonomy.—Our phylogenetic anal-ysis shows that section Armillati is paraphyletic astraditionally classified. The classification by Brandrudet al. (1992) is most congruent with ours includingonly one unrelated species, C. fuscoperonatus. As awhole sect. Armillati is phylogenetically well delimitedand the species share several morphological charac-teristics. The other red-brown-veiled species previous-ly included in sect. Armillati seem to belong to at leastseven different sections or clades: sect. Fulvescentes,sect. Boulderenses, sect. Anthracini, sect. Cinnabarini,/Praestigiosus, sect. Brunneotincti p.p., and /Fuscoper-onatus. Of these the species of the first five groups arehygrophanous and the species of sect. Brunneotinctioften have olivaceous tints in the fruiting body andobovoid to ellipsoid spores. If the spores are broadlyellipsoid to almost subglobose then large, extracellu-lar olivaceous spots deep in the pileipellis exist.Cortinarius fuscoperonatus, a rare species in Nordiccountries, is an isolated species with no close relativesand differs from the species of sect. Armillati byhaving a grayish brown universal veil, large (10.5–12.03 6.5–7.5 mm), strongly verrucose spores and moredistinctly duplex pileipellis.
The relationships of the species in section Armillatiwere well deciphered with ITS data, and theclassification also is supported by morphological andecological data. Two main groups were formed, C.quercoarmillatus/C. armillatus/C. roseoarmillatus andC. luteo-ornatus/C. suboenochelis/C. paragaudis/C.pinigaudis. The former three species are associatedwith deciduous trees and have thick-walled, stronglydextrinoid spores. The latter four associate mainlywith conifers and have thin- to moderately thick-walled, non-dextrinoid to moderately dextrinoidspores. Furthermore all species having brownish tintsin the universal veil belong in the latter group. Theinternal classification in the latter group is compara-ble to morphological distintinction. Based on mor-phology, C. paragaudis/C. pinigaudis forms a sistergroup with C. luteo-ornatus/C. suboenochelis, whichwas regarded as a subspecies by Lindstrom (1992).The former have small spores and distinctly innatelyfibrillose pileus, and the latter have larger spores andfinely fibrillose to small pileus.
Four of the six species we recognized fulfill thecriteria of a morphogenetic species (Niskanen et al.2009). They have distinguishing morphological char-acters, and pairwise infraspecific differences in the
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1099

ITS regions are smaller than interspecific ones,correlating well with the results of Frøslev et al.(2007). The two remaining species, C. paragaudis andC. pinigaudis, have morphological and ecologicaldifferences but differ only by one base pair in the ITS1 region. Although genetic studies (e.g. Kytovuori etal. 2005, Lindstrom et al. 2008, Suarez-Santiago et al.2009) on genus Cortinarius have treated this amountof variation as infraspecific, we are aware that somemorphological species are not distinguishable basedon ITS sequences (Frøslev et al. 2007, Garnica et al.2005, Peintner 2008). It is important to realize thatidentical ITS sequences are not necessarily in conflictwith the idea of distinct taxa. We lack sufficient datato confirm that ITS regions can separate allCortinarius species. Using more variable regionsmight solve the problem. For instance Hebelomashowed that IGS1 has more informative characteris-tics than ITS regions (Aanen et al. 2000). Further-more species delimitation based on IGS1 was morecongruent with results gained by using the biologicalspecies concept.
Cortinarius roseoarmillatus and C. suboenochelis hadno genetic variation in their ITS regions. Frøslev et al.(2007), Garnica et al. (2009) and Niskanen et al.(2009) have reported low genetic variation in someCortinarius species, even from a wide geographicalrange. An interesting species pair was C. paragaudis/C. pinigaudis, which resembles the species pair C.brunneus/C. glandicolor in sect. Brunnei (Niskanen etal. 2009). The other species (C. glandicolor, C.paragaudis) are genetically and morphologicallyvariable and grow in a wide range of habitats, whilethe sister species are less variable and ecologicallymore restricted.
Our data do not support the broader morpholog-ical species concept used in Brandrud et al. (1989,1992). C. paragaudis ssp. oenochelis appeared to begenetically and morphologically well delimited andtherefore we consider it a distinct species. Similarresults were gained on sect. Brunnei by Niskanen et al.(2009). We also do not agree with the narrow speciesconcept of Bidaud et al. (1995), and our resultscorrelate well with those of Frøslev et al. (2007) insect. Calochroi. Several names, such as C. luteo-ornatusand C. oenochelis, and C. paragaudis, C. bresadolianusand C. vaccinochelis, were synonyms. The two varietiesof C. armillatus were not morphologically distinguish-able and had ITS sequences identical to othersequenced C. armillatus specimens, although theITS region is probably too conserved to be useful forvariety-rank taxonomy in Cortinarius.
ITS regions have been proposed as a barcodingsequence for Cortinarius (Frøslev et al. 2007, Ortegaet al. 2008). The studies above demonstrate however
that a clear difference between the amount ofintraspecific and interspecific variation in ITS se-quences does not exist for every species. The totalnumber of Cortinarius species not having a clearbarcoding gap is unclear and most likely varies fromgroup to group. ITS sequences however are clearlynot sufficiently variable for species-rank recognition(barcoding) of all Cortinarius species.
ACKNOWLEDGMENTS
We thank Heino Vanska for revision of the Latin descrip-tions, Teuvo Ahti for advice on nomenclatural issues,Shannon Kuismanen for the linguistic revision of themanuscript and the reviewers for valuable comments. Weare grateful for the help of the curators of these herbaria:BG, C, G, GB, H, IB, JOE, KUO, LD, O, OULU, PC, S, TRH,TROM, TUR, UME and UPS. We also thank collectionlenders Katriina and Egil Bendiksen, Tea von Bonsdorff-Salminen, Lasse Kosonen, Hakan Lindstrom, Esteri Ohe-noja, Juhani Ruotsalainen, Jukka Vauras and Jan Vesterholt.This work was supported by Emil Aaltonen Foundation, theMinistry of Environment, Finland (YM38/5512/2009) andSocietas pro Fauna et Flora Fennica.
LITERATURE CITED
Aanen DK, Kuyper TW, Mes THM, Hoekstra RF. 2000. Theevolution of reproductive isolation in the ectomycor-rhizal Hebeloma crustuliniforme aggregate (Basidiomy-cetes) in northwestern Europe: a phylogenetic ap-proach. Evolution 54:1192–1206.
Ahti T, Hamet-Ahti L, Jalas J. 1968. Vegetation zones andtheir sections in northwestern Europe. Ann. BotFennici 5:169–211.
Ammirati J, Garnica S, Halling RE, Mata M, Mueller GM,Carranza J. 2007. New Cortinarius species associatedwith Quercus and Comarostaphylis in Costa Rica. Can JBot 85:794–812, doi:10.1139/B07-067
Bendiksen E, Bendiksen K. 1993. A new red-veiled species ofCortinarius subgenus Telamonia, C. rubrovioleipes sp.nov. Windahlia 20:29–34.
Bidaud A, Moenne-Loccoz P, Reumaux P. 1995. Atlas desCortinaires. Pars VII. Editions Federation mycologiqueDauphine-Savoie, La Roche-sur-Foron. 24 pl.
Brandrud TE, Lindstrom H, Marklund H, Melot J, MuskosS. 1989. Cortinarius Flora Photographica I (Swedishversion). Matfors: Cortinarius HB. 60 pl.
———, ———, ———, ———, ———. 1992. CortinariusFlora Photographica II (Swedish version). Matfors:Cortinarius HB. 60 pl.
Bulliard JB. 1791. Herbier de la France 11. Paris. 48 pl.———. 1793. Herbier de la France 13. Paris. 24 pl.———, Ventenat EP. 1812. Histoire des champignons de la
France II(2). Paris: Leblanc. 160 p.Consiglio G, Antonini D, Antonini M. 2004. Il genere
Cortinarius in Italia II. Luglio: Associazione MicologicaBresadola. 50 pl.
Edgar RC. 2004. Muscle: multiple sequence alignment with
1100 MYCOLOGIA

high accuracy and high throughput. Nucleic Acids Res32:1792–1797, doi:10.1093/nar/gkh340
Fries EM. 1821. Systema mycologicum I. Lundae, 520 p.———. 1838. Epicrisis systematis mycologici. Upsaliae.
610 p.———. 1877–1884. Icones selectæ Hymenomycetum non-
dum delineatorum I, II. Upsaliæ & Holmiæ. 100 pl.Frøslev TG, Jeppesen TS, Læssoe T, Kjøller R. 2007.
Molecular phylogenetics and delimitation of speciesin Cortinarius section Calochroi (Basidiomycota, Agar-icales) in Europe. Mol Phyl Evol 44:217–227,doi:10.1016/j.ympev.2006.11.013
Gardes M, Bruns TD. 1993. ITS primers with enhancedspecifity for basidiomycetes. Application to the identi-fication of mycorrhizae and rusts. Mol Ecol 2:113–118,doi:10.1111/j.1365-294X.1993.tb00005.x
Garnica S, Weiß M, Oertel B, Ammirati J, Oberwinkler F.2009. Phylogenetic relationships in Cortinarius, sectionCalochroi, inferred from nuclear DNA sequences. BMCEvol Biol 9:1, (http://www.biomedcentral.com/1471-2148/9/1), doi:10.1186/1471-2148-9-1
———, ———, ———, Oberwinkler F. 2005. A frameworkfor a phylogenetic classification in the genus Cortinar-ius (Basidiomycota, Agaricales) derived from morpho-logical and molecular data. Can J Bot 83:1457–1477,doi:10.1139/b05-107
Gladstein D, Wheeler W. 2001. POY (phylogeny reconstruc-tion via direct optimization of DNA data). 2.7. NewYork: Dept. Invertebrates, American Museum of Natu-ral History.
Henry R. 1977. Nouveau regard sur les cortinaires. Bull SocMycol Fr 93:313–371.
Høiland K. 1983. Cortinarius subgenus Dermocybe. OperaBotanica 71:1–113.
Hongo T. 1965. Notes on Japanese larger fungi 17. J Jpn Bot40:23–30.
Huelsenbeck JP, Ronquist F. 2003. MrBayes 3: Bayesianphylogenetic inference under mixed models. Bioinfor-matics 19:1572–1574, doi:10.1093/bioinformatics/btg180
Knudsen H, Vesterholt J. 2008. Funga Nordica. Agaricoid,Boletoid and Cyphelloid genera. Copenhagen: Nords-vamp. 965 p.
Kuhner R, Romagnesi H. 1953. Flore analytique deschampignons superieurs. Paris: Masson etc. 554 p.
Kytovuori I, Niskanen T, Liimatainen K, Lindstrom H. 2005.Cortinarius sordidemaculatus and two new relatedspecies, C. anisatus and C. neofurvolaesus, in Fenno-scandia (Basidiomycota, Agaricales). Karstenia 45:33–49.
———, ———, Ohenoja E. 2002. Cortinarius species foundin Kainuu during the XIXth Journees europeennes duCortinaire in 2001. J JEC Band 5:1–10.
Lindstrom H. 1992. Cortinarius paragaudis Fr. In: BrandrudTE, Lindstrom H, Marklund H, Melot J, Muskos S, eds.Cortinarius Flora Photographica II (in Swedish).Matfors: Cortinarius HB. p 22–24.
———, Bendiksen E, Bendiksen K, Larsson E. 2008.Studies of the Cortinarius saniosus (Fr.: Fr.) Fr.complex and a new closely related species, C.
aureovelatus (Basidiomycota, Agaricales). Sommerfeltia31:139–159.
Melot J. 1990. Une classification du genre Cortinarius(Pers.) S.F. Gray. Doc Mycol 80:43–59.
Moser M. 1965. Studien zu Cortinarius Fr. subgen.Telamonia sect. Armillati. Schweiz Z Pilzk 43:113–124.
———. 1978. Die Rohrlinge und Blatterpilze. 4th ed. In:Gams H, ed. Kleine Kryptogamenflora, 2b/2. Stuttgart,Germany: Gustav Fischer Verlag. 532 p.
Niskanen T, Kytovuori I. 2008. Key A: subgenus Telamoniasect. Armillati M.M. Moser. In: Knudsen H, Vesterholt J,eds. Funga Nordica. Agaricoid, Boletoid and Cyphel-loid genera. Copenhagen, Denmark: Nordsvamp.p 726–728.
———, Liimatainen K, Kytovuori I. 2008a. Two newCortinarius (Basidiomycota, Agaricales) species fromnorth Europe: C. brunneifolius and C. leiocastaneus.Mycol Prog 7:239–247, doi:10.1007/s11557-008-0565-1
———, ———, ———, Lindstrom H. 2008b. Subgen.Telamonia (Fr.) Trog. In: Knudsen H, Vesterholt J,eds. Funga Nordica. Agaricoid, Boletoid and Cyphelloidgenera. Copenhagen, Denmark: Nordsvamp. p 721–726.
———, ———, ———. 2009. Cortinarius sect. Brunnei(Basidiomycota, Agaricales) in north Europe. MycolRes 113:182–206, doi:10.1016/j.mycres.2008.10.006
Ortega A, Suarez-Santiago VN, Reyes JD. 2008. Morpholog-ical and ITS identification of Cortinarius species(section Calochroi) collected in Mediterranean Quercuswoodlands. Fungal Divers 29:73–88.
Peintner U. 2008. Cortinarius alpinus as an example formorphological and phylogenetic species concepts inectomycorrhizal fungi. Sommerfeltia 31:161–178.
———, Moncalvo J-M, Vilgalys R. 2004. Toward a betterunderstanding of the infrageneric relationships inCortinarius (Agaricales, Basidiomycota). Mycologia 96:1042–1058, doi:10.2307/3762088
Soop K. 1990. The group Cortinarius paragaudis Fries inSweden. Agarica 10/11:98–107.
———. 1997. Spindlingar med rod sloja. Jordstjarnan 18:26–35.
———. 2004. Cortinarius in Sweden, 9th rev ed. Mora:Editions Scientrix. 24 pl.
Suarez-Santiago VN, Ortega A, Peintner U, Lopez-Flores I.2009. Study on Cortinarius subgenus Telamonia sectionHydrocybe in Europe, with especial emphasis onMediterranean taxa. Mycol Res 113:1070–1090,doi:10.1016/j.mycres.2009.07.006
Thiers B. Continuously updated. Index Herbariorum: aglobal directory of public herbaria and associated staff.New York Botanical Gardens Virtual Herbarium.http://sweetgum.nybg.org/ih/
Wheeler WC. 1996. Optimization alignment. The end ofmultiple sequence alignment in phylogenetics? Cladis-tics 12:1–9, doi:10.1111/j.1096-0031.1996.tb00189.x
White TJ, Bruns T, Lee S, Taylor J. 1990. Amplification anddirect sequencing of fungal ribosomal RNA genes forphylogenetics. In: Michael AJ, Gelfand DH, Sninsky JJ,White TJ, eds. PCR protocols: a guide to the methodsand applications. New York: Academic Press. p 315–322.
NISKANEN ET AL.: CORTINARIUS SECT. ARMILLATI 1101
Copyright © 2022 FDOKUMEN



















