
Cortical activity related to accuracy of letter recognition
13
Cortical Activity Related to Accuracy of Letter Recognition 1 A. S. Garrett,* D. L. Flowers,² J. R.Absher,² F. H. Fahey,² H. D. Gage,² J. W. Keyes,² L. J. Porrino,² and F. B. Wood² *University of California at Davis, Davis, California 95616; and ²Wake Forest University School of Medicine, Winston-Salem, North Carolina 27157-1043 Received April 6, 1999 Previous imaging and neurophysiological studies have suggested that the posterior inferior temporal region participates in tasks requiring the recognition of objects, including faces, words, and letters; however, the relationship between accuracy of recognition and activity in that region has not been systematically investigated. In this study, positron emission tomogra- phy was used to estimate glucose metabolism in 60 normal adults performing a computer-generated letter- recognition task. Both a region of interest and a voxel- based method of analysis, with subject state and trait variables statistically controlled, found task accuracy to be: (1) negatively related to metabolism in the left ventrolateral inferior temporal occipital cortex (Brod- mann’s area 37, or ventrolateral BA 37) and (2) posi- tively related to metabolism in a region of the right ventrolateral frontal cortex (Brodmann’s areas 47 and 11, or right BA 47/11). Left ventrolateral BA 37 was significantly related both to hits and to false alarms, whereas the right BA 47/11 finding was related only to false alarms. The results were taken as supporting an automaticity mechanism for left ventrolateral BA 37, whereby task accuracy was associated with automatic letter recognition and in turn to reduced metabolism in this extrastriate area. The right BA 47/11 finding was interpreted as reflecting a separate component of task accuracy, associated with selectivity of attention broadly and with inhibition of erroneous responding in particular. The findings are interpreted as support- ing the need for control of variance due to subject and task variables, not only in correlational but also in subtraction designs. r 2000 Academic Press INTRODUCTION The processing of certain visual functions of the perception of discrete objects is widely recognized to be mediated by a ventral visual processing system that originates in striate and extrastriate visual cortex itself and extends through the lower lateral and inferior occipital–temporal cortical region. Also known as the ‘‘object’’ or ‘‘what is it’’ pathway (Ungerleider and Mish- kin, 1982), it is distinguished from a dorsal ‘‘location’’ or ‘‘where is it’’ pathway coursing upward into the parietal region. Successive regions in the ventral pathway are usually considered to involve progressively elaborated aspects of visual processing (Mishkin et al., 1983; Gaffan et al., 1986; Maunsell and Newsome, 1987; Felleman and Van Essen, 1991; Young, 1992), and within this putative hierarchy the posterior inferior temporal portion (Brodmann’s area 37) is selectively involved in object recognition, as first demonstrated in monkeys (Schwartz et al., 1983; Desimone et al., 1984, 1991; Tanaka et al., 1991; Perrett et al., 1987). Studies in human subjects also show a posterior inferior temporal role in object recognition and object naming (Ungerleider and Haxby, 1994; Price et al., 1996; Ungerleider et al., 1998; Moore and Price, 1999). Functional magnetic resonance imaging (fMRI) and positron emission tomography studies (PET) of re- gional cerebral blood flow (rCBF) show activation of BA 37 by tasks involving face (Haxby et al., 1991; Sergent et al., 1992), visual patterns (Roland and Gulyas, 1995; Schacter et al., 1995; Kawashima et al., 1998), and objects viewed from different perspectives (Kosslyn et al., 1994). From a clinical perspective, intraoperative electrophysiological recordings in the human extrastri- ate cortex of epileptic patients carried out by Allison et al. (1994) have shown that cells in the bilateral inferior temporo-occipital cortex, in the region of BA 37, re- spond selectively to the presentation of faces, letter strings, and numbers. Also, patients with damage to the occipito-temporal region often are unable to recog- nize familiar faces (Damasio et al., 1982), words (Binder and Mohr, 1992), or symbolic representations of words (Soma et al., 1989). Studies of nonpatient volunteers have demonstrated that the left BA 37 role in object recognition also includes language-relevant objects such as words, pho- netically regular nonwords, and letter strings. For 1 This work was supported by National Institute of Child Health and Development PHS Grant P01 HD 21887. NeuroImage 11, 111–123 (2000) doi:10.1006/nimg.1999.0528, available online at http://www.idealibrary.com on 111 1053-8119/00 $35.00 Copyright r 2000 by Academic Press All rights of reproduction in any form reserved.
Transcript of Cortical activity related to accuracy of letter recognition

hrotaipnrbvtvmtv1swfawliiabiits
pm
a
NeuroImage 11, 111–123 (2000)doi:10.1006/nimg.1999.0528, available online at http://www.idealibrary.com on
Cortical Activity Related to Accuracy of Letter Recognition1
A. S. Garrett,* D. L. Flowers,† J. R. Absher,† F. H. Fahey,† H. D. Gage,† J. W. Keyes,†L. J. Porrino,† and F. B. Wood†
*University of California at Davis, Davis, California 95616; and †Wake Forest University School of Medicine,Winston-Salem, North Carolina 27157-1043
Received April 6, 1999
oao‘k‘ruaGFwtim1
in1Fpg3eSoaeaatsstna(
ti
Previous imaging and neurophysiological studiesave suggested that the posterior inferior temporalegion participates in tasks requiring the recognitionf objects, including faces, words, and letters; however,he relationship between accuracy of recognition andctivity in that region has not been systematicallynvestigated. In this study, positron emission tomogra-hy was used to estimate glucose metabolism in 60ormal adults performing a computer-generated letter-ecognition task. Both a region of interest and a voxel-ased method of analysis, with subject state and traitariables statistically controlled, found task accuracyo be: (1) negatively related to metabolism in the leftentrolateral inferior temporal occipital cortex (Brod-ann’s area 37, or ventrolateral BA 37) and (2) posi-
ively related to metabolism in a region of the rightentrolateral frontal cortex (Brodmann’s areas 47 and1, or right BA 47/11). Left ventrolateral BA 37 wasignificantly related both to hits and to false alarms,hereas the right BA 47/11 finding was related only to
alse alarms. The results were taken as supporting anutomaticity mechanism for left ventrolateral BA 37,hereby task accuracy was associated with automatic
etter recognition and in turn to reduced metabolismn this extrastriate area. The right BA 47/11 finding wasnterpreted as reflecting a separate component of taskccuracy, associated with selectivity of attentionroadly and with inhibition of erroneous respondingn particular. The findings are interpreted as support-ng the need for control of variance due to subject andask variables, not only in correlational but also inubtraction designs. r 2000 Academic Press
INTRODUCTION
The processing of certain visual functions of theerception of discrete objects is widely recognized to beediated by a ventral visual processing system that
1 This work was supported by National Institute of Child Health
nnd Development PHS Grant P01 HD 21887.111
riginates in striate and extrastriate visual cortex itselfnd extends through the lower lateral and inferiorccipital–temporal cortical region. Also known as the
‘object’’ or ‘‘what is it’’ pathway (Ungerleider and Mish-in, 1982), it is distinguished from a dorsal ‘‘location’’ or
‘where is it’’ pathway coursing upward into the parietalegion. Successive regions in the ventral pathway aresually considered to involve progressively elaboratedspects of visual processing (Mishkin et al., 1983;affan et al., 1986; Maunsell and Newsome, 1987;elleman and Van Essen, 1991; Young, 1992), andithin this putative hierarchy the posterior inferior
emporal portion (Brodmann’s area 37) is selectivelynvolved in object recognition, as first demonstrated in
onkeys (Schwartz et al., 1983; Desimone et al., 1984,991; Tanaka et al., 1991; Perrett et al., 1987).Studies in human subjects also show a posterior
nferior temporal role in object recognition and objectaming (Ungerleider and Haxby, 1994; Price et al.,996; Ungerleider et al., 1998; Moore and Price, 1999).unctional magnetic resonance imaging (fMRI) andositron emission tomography studies (PET) of re-ional cerebral blood flow (rCBF) show activation of BA7 by tasks involving face (Haxby et al., 1991; Sergentt al., 1992), visual patterns (Roland and Gulyas, 1995;chacter et al., 1995; Kawashima et al., 1998), andbjects viewed from different perspectives (Kosslyn etl., 1994). From a clinical perspective, intraoperativelectrophysiological recordings in the human extrastri-te cortex of epileptic patients carried out by Allison etl. (1994) have shown that cells in the bilateral inferioremporo-occipital cortex, in the region of BA 37, re-pond selectively to the presentation of faces, lettertrings, and numbers. Also, patients with damage tohe occipito-temporal region often are unable to recog-ize familiar faces (Damasio et al., 1982), words (Bindernd Mohr, 1992), or symbolic representations of wordsSoma et al., 1989).
Studies of nonpatient volunteers have demonstratedhat the left BA 37 role in object recognition alsoncludes language-relevant objects such as words, pho-
etically regular nonwords, and letter strings. For1053-8119/00 $35.00Copyright r 2000 by Academic Press
All rights of reproduction in any form reserved.

egw1oao1e
oasiradttiWiidrAga
usrfoohacomaHrspctm
afa
tBHapri
nmsutsiAsrntnsdttRstJt
ipap
AEAPPTNWWWWW
W
112 GARRETT ET AL.
xample, fMRI has shown left BA 37 activation byeneration of rhymes and semantic categories (Shay-itz et al., 1995) and viewing letter strings (Puce et al.,996), and PET studies of rCBF have shown activationf portions of left BA 37 by visual presentation of wordsnd nonwords (Petersen et al., 1989, 1990) or by viewingr reading words (Bookheimer et al., 1995; Buchel et al.,998, Moore and Price, 1999). Activation in the ventrolat-ral portion of BA37 is common to most of these studies.While the above studies indicate a role for BA 37 in
bject recognition, it is unclear how that role relates toccuracy of task performance. If the activation is dueolely to the stimuli involved, without regard to successn recognizing them, then accuracy of performance inecognizing those stimuli should have no relation toctivation. On the other hand, if activation is depen-ent on actual object recognition itself, as indexed byask accuracy, then the activation should be propor-ional to task accuracy. However, accurate performances often less effortful than inaccurate performance (see
ood, 1990, for a review). Consequently, if BA37 activations to any substantial extent proportional to the effortnvolved, and if poor performance indexes greater effort orifficulty, then the activation might be greater with inaccu-ate performance than with more accurate performance.lso the roles of demographic variables, such as age andender, or extraneous task-related variables, such as statenxiety, remain to be clarified.The present study addresses the above questions by
sing a correlational approach that tests the relation-hip of ventrolateral BA 37 activity, during letterecognition, to task performance accuracy, with controlor variance in age, gender, and state anxiety. Of course,ther regions may be expected to be involved in variousf these relationships, but we offer no specific a prioriypotheses about these except to test several appropri-te candidate regions of interest. This multivariateorrelational approach requires larger sample sizes inrder to achieve adequate degrees of freedom (approxi-ately 10 subjects for each variable including global
ctivation, according to the conservative criterion ofarris, 1975), so the present study invests scanning
esources across a large sample of N 5 60, using aingle activation condition. This between-subjects ap-roach, as in a classic study of individual differences,ontrasts with the classic subtraction rationale in whichhe scanning resources are invested within subjects acrossultiple activation conditions and baselines.
METHODS AND MATERIALS
Subjects
Sixty healthy adults (50% male, 18% non-Caucasianll of whom were African American) were recruitedrom the surrounding communities by advertisement
nd word of mouth. The subjects ranged in age from 20 Wo 66 years (mean 40.6, standard deviation 12.3). Theriggs and Nebes (1975) modification of the Annettandedness Inventory classified 51 of the 60 subjectss strongly preferring the right hand, 3 as stronglyreferring the left hand, and 6 as not strongly prefer-ing either hand. Table 1 provides further descriptivenformation of the sample.
Subjects were included if they had no history ofeurological disease, head injury, diabetes or otheretabolic disease, heart disease, drug or alcohol abuse,
eizures, liver disease, or glaucoma or current or recentse of psychoactive or metabolically relevant medica-ions. Subjects were also excluded if their urine drugcreen on the day of scanning showed any evidence ofllegal or centrally acting (e.g., antihistimine) drug use.ll subjects included in the sample had the absence ofpike discharges or observable slow wave activity onesting EEG, had normal MRI of the brain, and hadormal fasting blood glucose levels. By the criteria ofhe Schedule for Affective Disorders and Schizophre-ia—Lifetime Version (Endicott and Spitzer, 1978),ubjects were excluded for histories of bipolar affectiveisorder or schizophrenia or for current unipolar affec-ive disorder. Subjects were selected without referenceo reading ability or Wechsler Adult Intelligence Test—evised subtests (Wechsler, 1981), and their scores oningle-word reading (Letter–Word Identification Sub-est of the Woodcock Johnson—Revised; Woodcock andohnson, 1989) and selected Wechsler subtests wereypical of normal adults, as shown in Table 1.
All subjects were scanned in the morning and werenstructed to abstain from nicotine and caffeine for 24 hrior to the study and from food or drink except waterfter midnight on the day of the study. Subjects wereaid for their participation, the study was approved by
TABLE 1
Demographic and Task Performance Characteristics
Mean Minimum MaximumStandarddeviation
ge 40.6 20 66 12.3ducation (years) 16.2 11 21 2.2nxiety (STAI-S) 30.0 20 64 8.1ercentage hits 96.5 79 100 4.4ercentage false alarms 5.8 0 26 6.0ask d-prime 3.9 1.98 5.58 1.0umber of trials 554 332 762 90.9AIS-R vocabulary 11.5 5 18 2.6AIS-R block design 10.1 5 15 2.4AIS-R digit–symbol 10.2 5 15 2.4AIS-R digit span 9.3 6 17 2.4J-R Word ID 49.3 39.7 51.0 3.6
Note. STAI-S, State–Trait Anxiety Inventory of Spielberger;AIS-R, Wechsler Adult Intelligence Scale, Revised; WJ-R Word ID,
oodcock Johnson—Revised Letter–Word Identification Task.
ts
wpPgtclSobmrlioaphts
lh_mtncwEfio
ptaptmst
ftotsA
atwtpascsFpbstfs1
Mgwat
Sttfic
g(ti1iuMaM
tsictn
113CORTICAL ACTIVITY RELATED TO TASK ACCURACY
he medical school’s Institutional Review Board, and allubjects gave their written consent.
Procedure
Consistent positioning in the PET and MRI scannersas facilitated by individually molded thermosettinglastic masks along with fiducial markers on the mask.rior to FDG uptake, subjects practiced a computer-enerated letter-recognition task (described below) un-il confident of their performance of the task and until ariterion score was achieved (greater than 75% hits andess than 25% false alarms during the trial period).ubjects wore glasses if needed, and none complainedf problems seeing the stimuli. State anxiety (Spiel-erger et al., 1983) was measured just prior to the 10Ci FDG bolus injection, which was delivered into the
ight arm antecubital vein, through an intravenous (iv)ine previously placed but immediately removed afternjection. Blood samples were withdrawn every 15 sver the first 2 min and at minutes 3, 4, 6, 8, 13, 18, 28,nd 38 and following scanning, through a previouslylaced indwelling iv in the left arm, which had beeneated to 110°F. The subject performed the computerask for 35 min, voided, and returned to the PETcanner for acquisition of the emission scan.
Letter Task during FDG Uptake
Stimuli for the letter-recognition task included 12etters (6 uppercase and 6 lowercase; for example, K, V,, and c) and 12 unfamiliar characters (for example, \,0 , ¥, and <) presented in either black or white inside a
agenta box at the center of a computer monitor. Thearget was any of the 12 letters regardless of color,ontarget characters were any of the 12 unfamiliarharacters. Probability of target versus nontarget itemsas 50%. The stimuli subtended a 0.7° visual angle.ach stimulus was flashed for 50 ms. A black dotxation point was displayed continuously in the centerf the screen within the magenta box.The subject controlled the pace of the game by
ressing and holding down the mouse button to start arial and lifting the right finger from the mouse buttons quickly as possible when a target stimulus wasresented. Subjects again depressed the mouse buttono start the next trial. If no finger lift response wasade, and the button remained down, then the next
timulus was presented at a random 1.5 to 2.0 s afterhe prior stimulus.
The computer provided auditory and visual feedbackor both correct and incorrect responses. The differen-ial auditory feedback was a short, higher pitched toner ‘‘beep’’ for correct and a moderately lower pitchedone or ‘‘boop’’ for incorrect. Tones were of equal inten-ity. Visually, several feedbacks were given, as follows.
correct response caused two dots, one below and one ebove the fixation point, to move by a small incrementoward the fixation point. When five correct responsesere accumulated, then the dots ended up close outside
he magenta box in middle field and a message wasrinted on the screen informing the subject of theward of bonus points for that group of correct re-ponses. The dots were then reset. Incorrect responsesaused a printed statement to appear notifying theubject of the mistake, whether miss or false alarm.inally, after every 17 trials, a rest message wasrinted that informed the subject of a brief (3 s) restefore resuming the task. A critical reaction time waset at 900 ms (i.e., target relevant responses after thisime were recorded as misses). Percentages of hits andalse alarms were used to calculate d-prime, the mea-ure of signal detection accuracy (Green and Swets,966).
MRI Scanning
A 1.5-T GE Signa MRI scanner (GE Medical Systems,ilwaukee, WI) was used to acquire axial 3D spoiled
radient-echo T1-weighted images (2.5-mm thicknessith no interslice gap, TR 5 45 ms, TE 5 5 ms, flipngle 5 45°, NEX 5 1). This anatomic image was usedo localize regions of interest (ROIs) on the PET image.
PET Scanning
All scans in this study were performed using aiemens/CTI 951/31 ECAT scanner which has a resolu-ion of approximately 6.0 mm in all axes. The attenua-ion-corrected emission data were reconstructed withltered back-projection using a Hann filter with a 0.4ycle/pixel cutoff.Quantitative images of glucose metabolic rate were
enerated by applying the method of Phelps et al.1979), and the reconstructed PET images were regis-ered by trained investigators to their respective MRImages using the ‘‘Register’’ program (Neelin et al.,993), validated by Woods (1996). The MRI was placedn stereotaxic space (Talairach and Tournoux, 1988)sing an algorithm developed by Louis Collins ofontreal Neurological Institute (Collins et al., 1994)
nd the PET data were resampled to correspond to theRI image.
PET Regions of Interest Analysis
Gemini, a locally developed software package, simul-aneously displays the MRI and its coregistered PETtudy, allowing any point to be simultaneously viewedn three orthogonal planes (transverse, sagittal, andoronal). It also allows the placement of a spherical ROIo be guided by viewing all three planes simulta-eously.MRI segmentation to isolate the functionally rel-
vant (Kadekaro et al., 1985; Juliano et al., 1981, 1983)

gp0(ncgsctomtt
adiswrgr
rliTl
C
B
A
I
O
D
T
A
Tvi
114 GARRETT ET AL.
ray matter signal intensity boundaries was accom-lished by placing small spherical ROIs, of diameter.67 cm, bilaterally in the genu of the corpus callosumrepresenting white matter), the head of the caudateucleus (representing gray matter), and the anteriorerebral ventricles (representing CSF). Relative homo-eneity of tissue type was achieved by accepting onlypheres whose distribution of pixel values had a coeffi-ient of variation (standard deviation as percentage ofhe mean) not exceeding 5% for gray and white matterr 12% for cerebrospinal fluid. The average of theedian pixel values in the two spheres for each tissue
ype was then taken as the criterion pixel intensity forhat tissue. Midpoints between the gray and the white
TAB
Regions
Region of interest(ROI)
Size of spherical Rto locate
alcarine fissure 2.0 cm. Locate the calcarine fissure ontion the sphere entirely within the ocfissure. On coronal view, move the ce1 cm from midline.
rodmann area 37 2.5 cm. Find the most lateral sagittal pporal sulcus and the preoccipital inciT-intersects the occipitotemporal sulintersection by paging through the asphere at this intersection, moving itthe cerebrum.
ngular gyrus 2.0 cm. On saggital views, locate the sumedially to its termination or that ofverify on the axial view. Page down tSTS terminus, and center the sphereessary to place sphere wholly in the c
nferior frontal 2.5 cm. Observe the coronal view tangedisplays the superior, middle (when(IFS). The IFS is easily located as a nverified from the sagittal view. Centemoving it medially until sphere is wh
rbital frontal 2.5 cm. Beginning from a low axial viewbone is visualized around the eyes. Tboth orbits and locate the most medinext gyrus laterally will be the orbitasphere in the gyrus so that the edgesrounding the gyrus. Adjust it verticaoften includes other short sulci.
orsal caudate 2.0 cm. Select the axial slice showing thCenter the sphere on this plane anddoes not infringe on the putamen. It
halamus 3.0 cm. Using all views, position the spmidline and excluding all adjacent bmay include portions of the ventricle
nterior cingulate 2.5 cm. Center the sphere midway betwintersection of the axial and coronaland anterior surfaces of the callosum
Note. Anatomical description for positioning on high-resolution MRalairach and Tournoux (1988); x, sagittal plane (negative indicateertical plane through the anterior commissure); z, axial plane (neg
n mm.nd between the gray and the CSF tissue criteria thenefined the thresholds for segmenting the MRI pixelsnto tissue types. Gemini then allowed into the analy-es only the PET voxels whose MRI intensities wereithin the gray matter thresholds. The interscorer
eliability (A.S.G. and D.L.F.) for both upper and lowerray matter thresholds, calculated as described for 15andomly selected MRIs, was 0.99.Three-dimensional ROI spheres, chosen a priori to
epresent candidate areas of general interest, wereocated on the MRI image, according to rules presentedn Table 2, which also shows their average location inalairach space (for a more complete description of ROI
ocation rules, see Wood and Flowers, 1999). Separate
2
nterest
and instructione ROI
Average Talairachcoordinates
(x, y, z)
sagittal view nearest midline. Posi-ital pole, its center in the calcariner laterally in the calcarine to a point
Left: 210, 282, 8Right: 11, 282, 8
e passing through the inferior tem-a (occipital notch), which usuallyon the ventral surface. Locate that
l planes in this region. Center thertically to ensure it is wholly within
Left: 246, 268, 211Right: 48, 263, 212
ior temporal sulcus (STS). Pagesuperior (angular) branch andaxial plane that is 1 cm below the
the STS. Page down farther if nec-brum.
Left: 246, 258, 41Right: 47, 253, 42
to the genu of the corpus callosum; itsent), and inferior frontal sulcirly horizontal feature and can bee sphere in the IFS on this plane,
y within the cerebrum.
Left: 240, 35, 12Right: 44, 35, 12
age upward until the orbital frontale observe a coronal plane throughentral gyrus, the gyrus rectus. Theontal gyrus (OFG). Center thethe sphere include both sulci sur-to avoid ocular muscle. The ROI
Left: 230, 40, 29Right: 32, 39, 29
ead of the caudate at its widest.ust from all views, so the spherey extend into the lateral ventricle.
Left: 210, 15, 4Right: 11, 15, 4
e within the thalamus, tangent tol ganglia structures. The sphere
Left: 211, 216, 8Right: 11, 216, 8
n the hemispheres, on the line ofnes tangent to the midline dorsal
(Mid:) 0, 33, 37
izes, and average locations of their centers in the stereotaxic space offt hemisphere); y, coronal plane (negative indicates posterior to a
ve indicates inferior to a horizontal plane through the AC-PC line);
LE
of I
OIth
thecipnte
lansurcusxiave
perits
o aninerentpreear tholl, p
heral vl frof
lly
e hadjmaherasas.ee
pla.
I, ss leati
omamode
twcbi1wd
R0(
gbda1eatT
P
115CORTICAL ACTIVITY RELATED TO TASK ACCURACY
bservers (A.S.G. vs D.L.F.) had especially close agree-ent (between 0 and 3 mm difference in any plane) for
ll regions except the angular gyri (between 4.5 and 6.0m absolute difference in the x and y planes) and the
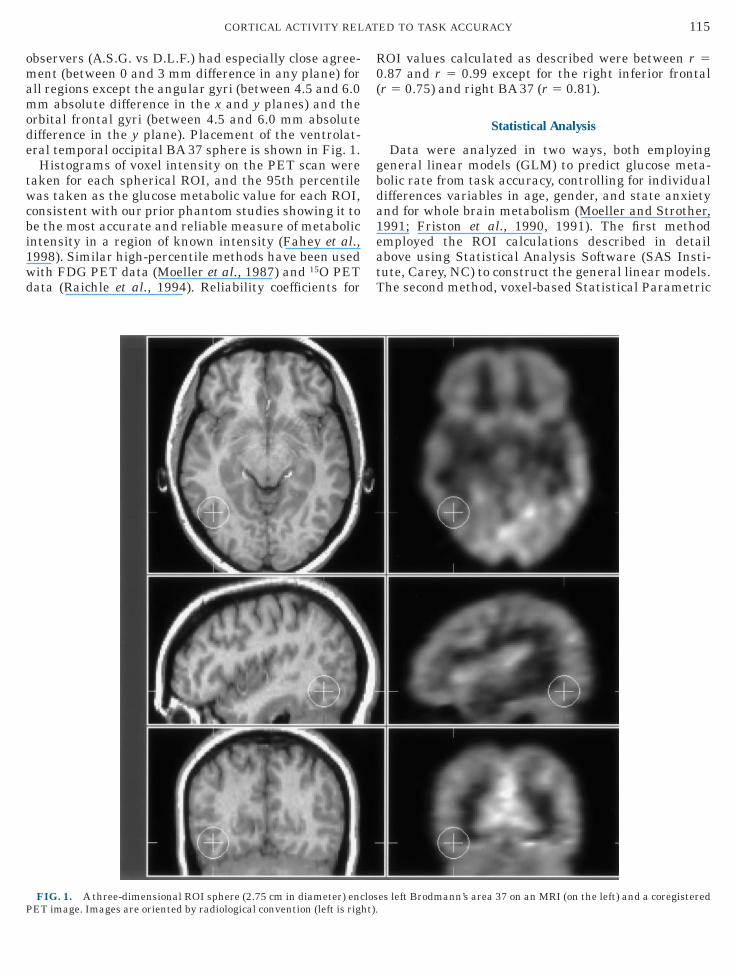
rbital frontal gyri (between 4.5 and 6.0 mm absoluteifference in the y plane). Placement of the ventrolat-ral temporal occipital BA 37 sphere is shown in Fig. 1.Histograms of voxel intensity on the PET scan were
aken for each spherical ROI, and the 95th percentileas taken as the glucose metabolic value for each ROI,
onsistent with our prior phantom studies showing it toe the most accurate and reliable measure of metabolicntensity in a region of known intensity (Fahey et al.,998). Similar high-percentile methods have been usedith FDG PET data (Moeller et al., 1987) and 15O PETata (Raichle et al., 1994). Reliability coefficients for
FIG. 1. A three-dimensional ROI sphere (2.75 cm in diameter) enc
ET image. Images are oriented by radiological convention (left is right).OI values calculated as described were between r 5.87 and r 5 0.99 except for the right inferior frontalr 5 0.75) and right BA 37 (r 5 0.81).
Statistical Analysis
Data were analyzed in two ways, both employingeneral linear models (GLM) to predict glucose meta-olic rate from task accuracy, controlling for individualifferences variables in age, gender, and state anxietynd for whole brain metabolism (Moeller and Strother,991; Friston et al., 1990, 1991). The first methodmployed the ROI calculations described in detailbove using Statistical Analysis Software (SAS Insti-ute, Carey, NC) to construct the general linear models.he second method, voxel-based Statistical Parametric
es left Brodmann’s area 37 on an MRI (on the left) and a coregistered
los
M(aR(ltns
st8tccs
s
favG
tshEnenw
oRsnpaasetAsfifsb
ttudwdvt
A%%S
P
At
sD
LRLRLRLRLRLRLRA
G
116 GARRETT ET AL.
apping (SPM), was carried out using SPM96 softwareFriston et al., 1991, 1994, 1995; Worsley et al., 1992),fter conversion to ANALYZE format (Mayo Clinic,ochester, Minnesota). Conversion to standard space
Talairach and Tournoux, 1988) was done using aocally constructed template that is the average of 40 ofhe 60 normal glucose metabolized PET brains. Theormalized PET images were then conventionallymoothed using a 13-mm (FWHM) Gaussian kernel.In the SPM multisubject, single-condition design,
ubject and covariate effects were estimated accordingo the general linear model at every voxel exceeding0% of the mean global gray matter threshold. A heighthreshold of P 5 0.001, uncorrected, was set to identifylusters. Inasmuch as the ROI analyses had alreadyonstrained the total volume to prespecified areas, amall volume correction was employed (Worsley, 1996).
RESULTS
General Linear Models of Regional Metabolism
Pearson correlations between task variables andubject variables are listed in Table 3.The formal GLM predicted metabolism for each ROI
rom d-prime (task accuracy) age, gender, state anxiety,nd whole brain average metabolism. Table 4 lists the Falues and significance levels for each variable in eachLM.A significant F value indicates unique contribution of
hat variable to the regional metabolic-dependent mea-ure after all other sources of variance in the modelave been taken into account (Type III sums of squares).ven after statistical control, d-prime contributed sig-ificantly—and inversely—to the variance in left lat-ral BA 37 metabolism (P , 0.0005). D-prime alsoegatively predicted left angular gyrus metabolismhile it positively predicted left thalamus and right
TABLE 3
Pearson Product-Moment Correlations between TaskPerformance Variables and Individual Difference Variables
Task variable Age GenderAnxiety(STAI-S) Handedness
ccuracy (d-prime) 20.60*** 20.04 20.23 20.08Hits 20.57*** 0.01 20.19 20.12False alarms 0.52*** 0.05 0.33* 20.12
timulus rate(items per minute) 0.12 0.27* 20.14 0.08
rinted messages 0.45** 0.23 0.21 0.03
Note. Gender is coded as male, 1; female, 2. STAI-S, State–Traitnxiety Inventory of Spielberger. ‘‘2’’ indicates an inverse correla-
ion. Nonsignificant correlations with task variables are omitted.* P , 0.05.
** P , 0.005.
s*** P , 0.0001.rbital frontal metabolism (all at P , 0.05). No otherOI was significantly related to task accuracy. As apecial test for possible confounding effects of handed-ess, rate of stimulus presentation, and number ofrinted error messages delivered to the subjects, welso produced subsequent GLMs for these four regions,dding each of the potential confounding variableseparately. These confirmed that the addition of hand-dness or stimulus presentation rate had little effect onhe relationship of d-prime to regional metabolism.dding the number of printed error messages as atatistical covariate control did not change the area 37nding, but did abolish, i.e., explain, the right orbital
rontal (BA 47/11) relationship to d-prime. Table 5ummarizes these effects on the d-prime affect in theasic general linear model.The calculation of glucose metabolic rate is exponen-
ially weighted from the moment of injection and isherefore dominated by the earlier minutes of theptake period. We addressed this issue by examiningata from the 37 subjects in the experiment on whome had individual trial data. These subjects did notiffer from the total 60 in any demographic or taskariables or in the regional metabolic findings. Withinhe 37 subjects, d-prime for the first 40 or 80 trials was
TABLE 4
F Values (Type III Sums of Squares) for Linear Regres-ion Models of ROI Metabolism Predicted by Individualifferences
ROI
Individual difference variables includedin each general linear model
Globalmetabolism Age Gender
Stateanxiety
Taskaccuracy(d-prime)
calcarine 250.44***calcarine 290.09***BA 37 484.88*** 6.15* (2)14.61***BA 37 568.63*** 12.83**inf frontal 450.89***inf frontal 631.04***orbital 212.51***orbital 272.56*** 4.80*thalamus 334.55*** 4.34*thalamus 386.84*** 9.78**angular G 325.68*** (2)6.34* (2)5.26*angular G 322.76***caudate 243.51***caudate 473.60***nt cing 309.06*** (2)6.13*
Note. ‘‘2’’ indicates an inverse relation. L, left; R, right; Inf, inferior;, gyrus; Ant cing, anterior cingulate.P . 0.05 not shown.
* P , 0.05.** P , 0.005.
*** P , 0.0005.
ignificantly related (r 5 0.64, P , 0.0001 and r 5 0.77,
Ptawafii
ctha
m
sst
a(l(tttad
2tfiia1Ni
Slc(4sstFdtrs
(roahirf(ar
iAHGDt
BBLL
(
fRa
%%A
117CORTICAL ACTIVITY RELATED TO TASK ACCURACY
, 0.0001, respectively) to d-prime over the entireask period. More importantly, all relations to taskccuracy were the same if the d-prime accuracy scoreas calculated only from the first 40 or 80 trials (onverage, the first 2.5 or 5 min). There was a tendencyor d-prime to improve over trials, but measures of thismprovement showed no relation to the metabolic find-ngs.
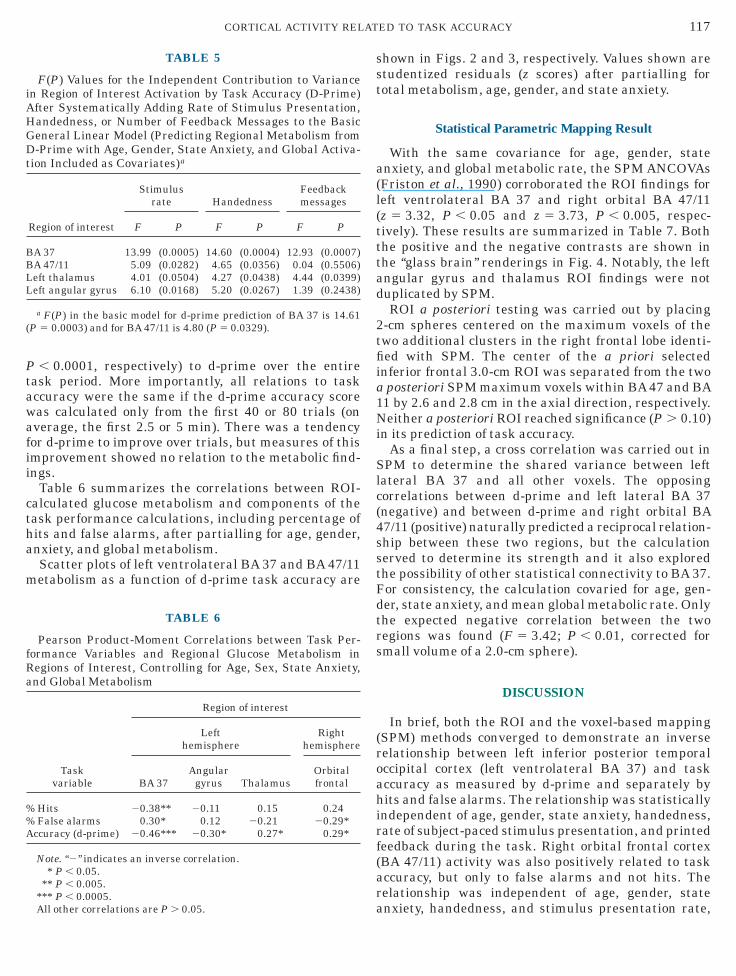
Table 6 summarizes the correlations between ROI-alculated glucose metabolism and components of theask performance calculations, including percentage ofits and false alarms, after partialling for age, gender,nxiety, and global metabolism.Scatter plots of left ventrolateral BA 37 and BA 47/11etabolism as a function of d-prime task accuracy are
TABLE 5
F(P) Values for the Independent Contribution to Variancen Region of Interest Activation by Task Accuracy (D-Prime)fter Systematically Adding Rate of Stimulus Presentation,andedness, or Number of Feedback Messages to the Basiceneral Linear Model (Predicting Regional Metabolism from-Prime with Age, Gender, State Anxiety, and Global Activa-
ion Included as Covariates)a
Region of interest
Stimulusrate Handedness
Feedbackmessages
F P F P F P
A 37 13.99 (0.0005) 14.60 (0.0004) 12.93 (0.0007)A 47/11 5.09 (0.0282) 4.65 (0.0356) 0.04 (0.5506)eft thalamus 4.01 (0.0504) 4.27 (0.0438) 4.44 (0.0399)eft angular gyrus 6.10 (0.0168) 5.20 (0.0267) 1.39 (0.2438)
a F(P) in the basic model for d-prime prediction of BA 37 is 14.61P 5 0.0003) and for BA 47/11 is 4.80 (P 5 0.0329).
TABLE 6
Pearson Product-Moment Correlations between Task Per-ormance Variables and Regional Glucose Metabolism inegions of Interest, Controlling for Age, Sex, State Anxiety,nd Global Metabolism
Taskvariable
Region of interest
Lefthemisphere
Righthemisphere
BA 37Angular
gyrus ThalamusOrbitalfrontal
Hits 20.38** 20.11 0.15 0.24False alarms 0.30* 0.12 20.21 20.29*
ccuracy (d-prime) 20.46*** 20.30* 0.27* 0.29*
Note. ‘‘2’’ indicates an inverse correlation.* P , 0.05.
** P , 0.005.*** P , 0.0005.
a
hown in Figs. 2 and 3, respectively. Values shown aretudentized residuals (z scores) after partialling forotal metabolism, age, gender, and state anxiety.
Statistical Parametric Mapping Result
With the same covariance for age, gender, statenxiety, and global metabolic rate, the SPM ANCOVAsFriston et al., 1990) corroborated the ROI findings foreft ventrolateral BA 37 and right orbital BA 47/11z 5 3.32, P , 0.05 and z 5 3.73, P , 0.005, respec-ively). These results are summarized in Table 7. Bothhe positive and the negative contrasts are shown inhe ‘‘glass brain’’ renderings in Fig. 4. Notably, the leftngular gyrus and thalamus ROI findings were notuplicated by SPM.ROI a posteriori testing was carried out by placing
-cm spheres centered on the maximum voxels of thewo additional clusters in the right frontal lobe identi-ed with SPM. The center of the a priori selected
nferior frontal 3.0-cm ROI was separated from the twoposteriori SPM maximum voxels within BA 47 and BA1 by 2.6 and 2.8 cm in the axial direction, respectively.either a posteriori ROI reached significance (P . 0.10)
n its prediction of task accuracy.As a final step, a cross correlation was carried out in
PM to determine the shared variance between leftateral BA 37 and all other voxels. The opposingorrelations between d-prime and left lateral BA 37negative) and between d-prime and right orbital BA7/11 (positive) naturally predicted a reciprocal relation-hip between these two regions, but the calculationerved to determine its strength and it also exploredhe possibility of other statistical connectivity to BA 37.or consistency, the calculation covaried for age, gen-er, state anxiety, and mean global metabolic rate. Onlyhe expected negative correlation between the twoegions was found (F 5 3.42; P , 0.01, corrected formall volume of a 2.0-cm sphere).
DISCUSSION
In brief, both the ROI and the voxel-based mappingSPM) methods converged to demonstrate an inverseelationship between left inferior posterior temporalccipital cortex (left ventrolateral BA 37) and taskccuracy as measured by d-prime and separately byits and false alarms. The relationship was statistically
ndependent of age, gender, state anxiety, handedness,ate of subject-paced stimulus presentation, and printedeedback during the task. Right orbital frontal cortexBA 47/11) activity was also positively related to taskccuracy, but only to false alarms and not hits. Theelationship was independent of age, gender, state
nxiety, handedness, and stimulus presentation rate, All other correlations are P . 0.05.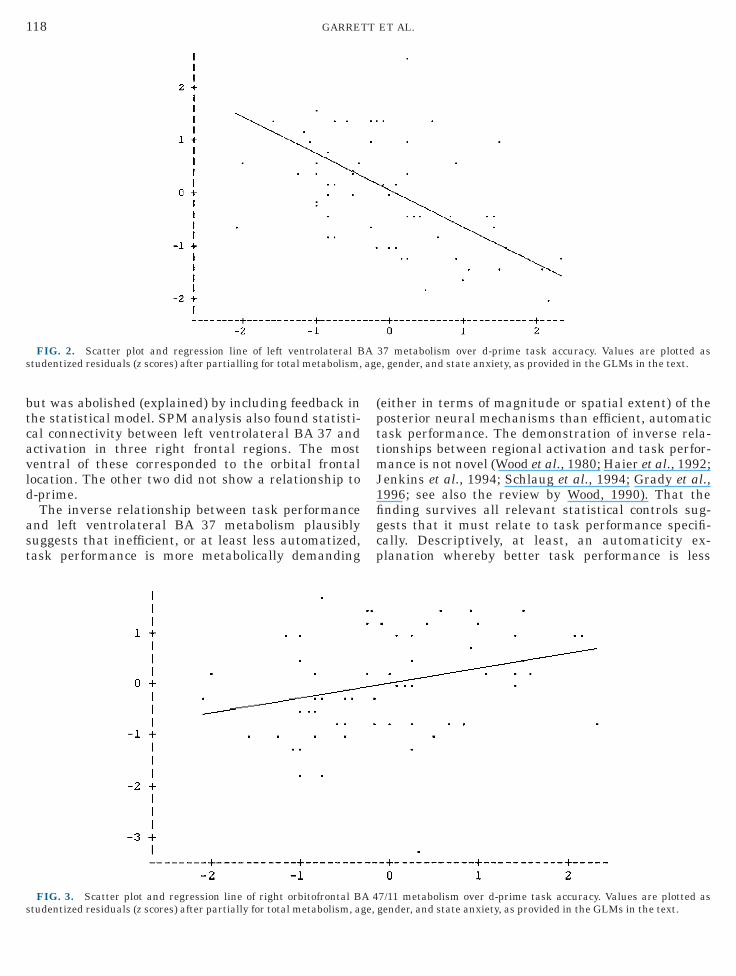
btcavld
ast
(pttmJ1figcp
s ag
s
118 GARRETT ET AL.
ut was abolished (explained) by including feedback inhe statistical model. SPM analysis also found statisti-al connectivity between left ventrolateral BA 37 andctivation in three right frontal regions. The mostentral of these corresponded to the orbital frontalocation. The other two did not show a relationship to-prime.The inverse relationship between task performance
nd left ventrolateral BA 37 metabolism plausiblyuggests that inefficient, or at least less automatized,ask performance is more metabolically demanding
FIG. 2. Scatter plot and regression line of left ventrolateral Btudentized residuals (z scores) after partialling for total metabolism,
FIG. 3. Scatter plot and regression line of right orbitofrontal B
tudentized residuals (z scores) after partially for total metabolism, age,either in terms of magnitude or spatial extent) of theosterior neural mechanisms than efficient, automaticask performance. The demonstration of inverse rela-ionships between regional activation and task perfor-ance is not novel (Wood et al., 1980; Haier et al., 1992;
enkins et al., 1994; Schlaug et al., 1994; Grady et al.,996; see also the review by Wood, 1990). That thending survives all relevant statistical controls sug-ests that it must relate to task performance specifi-ally. Descriptively, at least, an automaticity ex-lanation whereby better task performance is less
37 metabolism over d-prime task accuracy. Values are plotted ase, gender, and state anxiety, as provided in the GLMs in the text.
7/11 metabolism over d-prime task accuracy. Values are plotted as
A
A 4
gender, and state anxiety, as provided in the GLMs in the text.
et
ei(acsaopB
svosan1sr(tsmbabMe1itp
rlpampna
VtC
f
Ac
nms
119CORTICAL ACTIVITY RELATED TO TASK ACCURACY
ffortful and less metabolically demanding, is difficulto avoid.
Of course, ventrolateral BA 37 is not the only ventralxtrastriate region to be implicated in lexical process-ng and discrimination. For example, Petersen et al.1990) showed both lateral and medial extrastriatectivation by words and nonwords compared to fixationontrol, but the effect disappeared in the Howard et al.tudy (1992) when the control task was unpronounce-ble letter strings or false fonts. In contrast, activationf the ventrolateral posterior temporo-occipital visualrocessing stream, near or including our ventrolateralA 37 ROI, does survive a number of different compari-
TABLE 7
Stereotaxic Location and Confidence Levels for Maximumoxels Within Clusters Identified by SPM in Predicted Loca-ions (P , 0.001 for Height, Uncorrected, with Small Volumeorrection)
z scoreor height
Coordinates:x, y, z
Brodmann’sarea Cortical area
3.73 40, 46, 214 47/11** Right orbital frontal
Negative contrast
3.32 246, 258, 212 37* Left temporal occipital
Note. Gray matter threshold at 0.80 of mean global metabolic rate.NCOVA normalization for mean global metabolism; confounding
ovariates age, gender, and state anxiety.* P , 0.05.
** P , 0.005.
FIG. 4. Statistical parametric maps (thresholded at P , 0.001,ormal adults during a letter-identification task with age, gender, aetabolism. The orthogonal set on the left shows higher glucose met
hows lower glucose metabolic rate relative to better performance.
on conditions, as in simple viewing of real words vsiewing of face or texture controls (Puce et al., 1996) orvert discriminations such as real words vs lettertrings (Buchel et al., 1998). Activation in this area alsoppears to involve tasks in which silent or overtaming of visual objects is required (Bookheimer et al.,995; Price et al., 1996; Moore and Price, 1999). To beure, objects other than lexical ones also activate thisegion—e.g., pattern matching vs pattern scanningKawashima et al., 1998). Consistent with the above,hen, the ventrolateral BA 37 activity we observe in ourtudy is plausibly relevant to discrimination task perfor-ance, more so than medial extrastriate activity would
e. Furthermore, the fact that ventrolateral BA 37ctivation is inverse to task performance is explainabley the naming data reviewed above (Price et al., 1996;oore and Price, 1999): if subjects adopted a strategy of
ven subvocal naming of the stimuli (Bookheimer et al.,995), and if that strategy is more often adopted byndividuals having more difficulty with the task, thenhe excess ventrolateral BA 37 activity with poorerformance is understandable.The role of the right orbital cortex in this letter
ecognition task was not explicitly predicted. Neverthe-ess, right frontal lateral and orbital activity has beenositively related to percentage of correct responses onface recognition task (Grady et al., 1996), to perfor-ance on a Stroop task (Bench et al., 1993), to task
erformance feedback (Elliot et al., 1997), and to theumber of successive correct responses while shiftingttention between color and shape (Nagahama et al.,
corrected) predicting normalized glucose metabolic rate in N 5 60state anxiety covaried. Voxel values are normalized by global meanlic rate with better performance (d-prime), and the set on the right
unndabo

1tr
wlto1Osl(catwwpt(arwtps
riRd(TrrciArbi(imiNso
wcpffs
riictbicgrAmcrnTcpei
etrpoitebpismbt1iiitbaAdtiw
srsNin
120 GARRETT ET AL.
998). Of special note is the fact that the first two ofhese studies showed reciprocal decreases in otheregions, including extrastriate visual cortex.Reciprocities of activation (some regions increasinghile others decrease) are elsewhere familiar in the
iterature. For example, reciprocal changes in activa-ion with diminishing difficulty over trials of practicen motor tasks have been demonstrated (Grafton et al.,992, 1994; Schlaug et al., 1994; Jenkins et al., 1994).ther studies involving identification of object or word
timuli have suggested an inverse response betweeneft posterior cortical and frontal areas. Howard et al.1992) found a reduction in right inferior prefrontalortex activation (and in other right hemisphere areas)nd increased activation of the left posterior middleemporal gyrus during a single-word reading taskhen the control task was viewing false font stringshile repeating a single word (thus controlling forrocessing of visual patterns, articulation, and audi-ory feedback of subject’s own voice). Buckner et al.1996) also reported opposing anterior versus posteriorctivation, but in the reverse direction with increasedight frontal and decreased bilateral parietal activity,hen subjects performed an auditory semantic memory
ask wherein they recalled practiced word–word oricture–word pairs when the control task was theimple repetition of words.Another example of reciprocity—demonstrating neu-
al system responses to both increases and decreasesn difficulty—occurs in a cleverly designed study byaichle et al. (1994), who examined cortical activationuring verb generation to visually presented nounscontrasted with simply reading a string of nouns).hey found increased activation in left posterior tempo-al cortices (including BA 37) and other regions (ante-ior cingulate, left prefrontal, and right cerebellum) butoncomitant decreased activation in bilateral Sylviannsular cortex and the left medial extrastriate cortex.fter 15 min of practice, however, activity in theseegions reversed while task performance, as measuredy reaction time, decreased simultaneously. Introduc-ng a novel list of nouns returned most of these regionsand reaction time) to the ‘‘naive’’ state, leading thesenvestigators to conclude that separate circuits serve
ore effortful and automatic response choice selectionn a word generation task when visual stimuli are used.otably, however, the method of averaging scans across
ubjects did not allow for a direct analysis of the effectf individual task accuracy on focal neuronal activity.A relationship similar to that of the present studyas reported by Grady et al. (1996) who found that,
ompared to a sensorimotor control task (viewing noiseatterns), the task of matching progressively degradedaces was related to progressive increases in rightrontal BA 9/46 and concomitant decreases in medial
triate and bilateral fusiform (BA 19/37) regions. Accu- tate judgments during this self-paced task correspond-ngly declined, as might be expected. It is tempting tonfer a relationship between task accuracy and specificortical changes; however, unexpectedly—and consis-ent with the present findings—a direct correlationetween task performance and cortical blood flow dur-ng the high degradation condition revealed positiveorrelations between accuracy and a right frontal re-ion (BA 45) and negative correlations between accu-acy and left prefrontal cortex and striate cortex.lthough the present study does not systematicallyanipulate quality of the stimuli, targets are made
hallenging to detect by their 50-ms duration and byestricting response time, whereas the Grady study didot constrain stimulus duration or response time.hus, both studies point to the interpretative error thatould be made if the relationship between strength oferformance and regional activity is not specificallyxamined. Of course, forces driving these similar find-ngs may not be the same.
Even though working memory was not explictlylicited by the present task, if a memory strategy wereo be adopted by the subject—such as maintaining aepresentation of ‘‘letters’’ or ‘‘nonletter characters’’—erformance might be enhanced. Consider the findingsf McIntosh et al. (1996) that as retention delay wasncreased on a delayed match-to-sample face percep-ion task there was a decrease in striate and ventralxtrastriate area activity as measured by cerebrallood flow, interpreted as reflecting transient visualerceptual processes, while right prefrontal activityncreased, interpreted as a greater reliance in the moreustained task of holding the object’s icon in workingemory. Right frontal BA 47/11 activation has also
een reported in response to delayed match-to-sampleasks involving face matching (Haxby et al., 1994,995). Thus, if the subjects in the present study differedn their reliance on a strategy that minimized the effortn earlier sensory processing and maximized the effortn maintaining a working memory representation ofhe target stimuli, and if that strategy engenderedetter performance, then the reciprocity of left extrastri-te and right orbital activation could be explained.gainst this explanation, however, is the fact that theegree of a subject’s improvement in performance overrials—which would be expected to reflect an increas-ngly available working memory of the target stimuli—as not related to localized brain activity in our study.Perhaps a more satisfying interpretation of the pre-
ent results relates to task demands, which in this caseequire subjects to withhold responses to particulartimuli as well as to emit responses to target stimuli.otably, in the present study orbital metabolism is
nversely related to the percentage of false alarms butot significantly related to percentage of hits. In con-
rast, BA 37 is related to both hits and false alarms. The
olicvwbvt
thtf
saabdtb
rmaHgrdbpef
avvlaraotaabscaa
mmcc
vvplss
iat
A
B
B
B
B
B
B
C
C
D
D
D
E
E
E
F
F
121CORTICAL ACTIVITY RELATED TO TASK ACCURACY
rbital frontal component, therefore, may be particu-arly related to the allocation of resources to inhibitmpulsive responses, whereas the ventrolateral BA 37omponent may be more directly related to basic targets nontarget discriminations. That would also explainhy statistical control for the number of printed feed-ack messages, which were more highly correlated withariance in errors than in hits, would abolish or explainhe right orbital frontal finding.
Consistent with our a priori decision to interpret onlyhose sites corroborated both by ROI and by SPM, weave declined to interpret the angular gyrus andhalamic findings, which should then be the object ofurther investigation.
Our technique of validating ROI methods by SPM isimilar, albeit directionally reversed, to that of Koepp etl. (1997). They propose that, because of SPM’s moder-te liability to partial volume effects, the conservativeest approach would be to use SPM to generate candi-ate maps and then test them by an ROI method. Inhis case, we had a priori candidate regions, to confirmy SPM.Finally, the role of stimulus familiarity in object
ecognition is not addressed by this study. Hits andisses reflect responses to letters, while false alarms
nd correct rejections are responses to characters.owever, since left ventrolateral BA 37 metabolism is
reater for any correct decision, whether hits or correctejections, then familiarity is not confounded with theecision process itself. However, this explanation muste tested further. The length of the glucose uptakeeriod (35 min) may allow the subject to gain enoughxperience with the foil characters to consider themamiliar; however, no posttest of this was administered.
The present study reduces unexplained variance byccounting for age, gender, and state anxiety. This isariance that would otherwise be treated as errorariance, so some inconsistencies across studies in theiterature may be explained by these sources of vari-nce. On the other hand, despite these controls, thereemains a significant proportion of unexplained vari-nce in localized activation, in the present study andther studies. Future strategies for exploring this ques-ion should include both a correlational approach thatddresses other demographic or state variables andlso a subtraction approach that uses appropriateaseline controls. Both strategies remain important,ince the subtraction strategy would still be subject toonfounds due to such factors as accuracy and statenxiety, which could vary not only between subjects butlso within subjects.The present study, therefore, suggests the followingethodological considerations. Individual task perfor-ance measures should be considered in interpreting
ortical activation. Subject samples need to be of suffi-
ient size to allow for statistical control of subjectariables. Interactions among task variables, subjectariables, and localized brain activity are not onlyossible but should be expected. Finally, inverse corre-ations and reciprocities of localized brain activityhould not be overlooked, and data analytic strategieshould routinely examine for them.
ACKNOWLEDGMENTS
The authors thank Cathy Eades for her invaluable help in develop-ng and refining the Gemini software used to measure the PET datand also Dennis Vickland, Natalie Price, and Beth Harkness for theirechnical assistance.
REFERENCES
llison, T., McCarthy, G., Nobre, A., Puce, A., and Belger, A. 1994.Human extrastriate visual cortex and the perception of faces,words, numbers, and colors. Cereb. Cortex 4:544–554.ench, C. J., Frith, C. D., Grasby, P. M., Frackowiak, R. S., and Dolan,R. J. 1993. Investigations of the functional anatomy of attentionusing the Stroop test. Neuropsychologia 31:907–922.inder, J. R., and Mohr, J. P. 1992. The topography of callosal readingpathways. Brain 115:1807–1826.
ookheimer, S. Y., Zeffiro, T. A., Blaxton, T., Gaillard, W., andTheodore, W. 1995. Regional cerebral blood flow during objectnaming and word reading. Hum. Brain Mapp. 3:93–106.riggs, G. G., and Nebes, R. D. 1975. Patterns of hand preference in astudent population. Cortex 11:230–238.
uckner, R. L., Raichle, M. E., Meizin, F. M., and Petersen, S. E. 1996.Functional anatomic studies of memory retrieval for auditorywords and visual pictures. J. Neurosci. 16:6219–6235.uchel, C., Price, C., and Kriston, K. 1998. A multimodal languageregion in the ventral visual pathway. Nature 394:274–277.ollins, D. L., Neelin, P., Peters, T. M., and Evans, A. C. 1994.Automatic 3D intersubject registration of MR volumetric data instandardized Talairach space. J. Comput. Assisted Tomogr. 18:192–205.ourtney, S. M., Ungerleider, L. G., Keil, K., and Haxby, J. V. 1996.Object and spatial visual working memory activate separate neuralsystems in human cortex. Cereb. Cortex 6:39–49.amasio, A. R., Damasio, H., and Van Hoesen, G. W. 1982. Prosopag-nosia: Anatomic basis and behavioral mechanisms. Neurology32:331–341.esimone, R. 1991. Face-selective cells in the temporal cortex ofmonkeys. Neuroscience 3:1–8.esimone, R., Albright, T. D., Gross, C. G., and Bruce, C. 1984.Stimulus selective properties of inferior temporal neurons in themacaque. J. Neurosci. 4:2051–2062.lliot, R., and Dolan, R. J. 1998. Activation of different anteriorcingulate foci in association with hypothesis testing and responseselection. NeuroImage 8:17–29.lliot, R., Frith, C. D., and Dolan, R. J. 1997. Differential neuralresponse to positive and negative feedback in planning and guess-ing tasks. Neuropsychologia 35:1395–404.ndicott, J., and Spitzer, R. L. 1978. A diagnostic interview: TheSchedule for Affective Disorders and Schizophrenia. Arch. Gen.Psychiatry 35:837–844.
ahey, F. H., Wood, F. W., Flowers, D. L., Eades, C. G., Gage, H. D.,and Harkness, B. A. 1998. Evaluation of brain activation in FDGPET studies. J. Comput. Assisted Tomogr. 22:953–961.
elleman, D. J., and Van Essen, D. C. 1991. Distributed hierarchical
processing in the primate cerebral cortex. Cereb. Cortex 1:1–47.
F
F
F
F
G
G
G
G
G
H
H
H
H
H
H
J
J
J
K
K
K
K
M
M
M
M
M
M
N
N
P
P
P
P
P
P
R
122 GARRETT ET AL.
riston, K. J., Frith, C. D., Liddle, P. F., Dolan, R. J., Lammertsma,A. A., and Frackowiak, R. S. K. 1990. The relationship betweenglobal and local changes in PET scans. J. Cereb. Blood Flow Metab.10:458–66.
riston, K. J., Frith, C. D., Liddle, P. F., and Frackowiak, R. S. J.1991. Comparing functional (PET) images: The assessment ofsignificant change. J. Cereb. Blood Flow Metab. 11:690–699.
riston, K. J., Worsley, K. J., Frackowiak, R. S. J., Mazziotta, J. C.,and Evans, A. C. 1994. Assessing the significance of focal activa-tions using their spatial extent. Hum. Brain Mapp. 1:214–220.
riston, K. J., Holmes, A. P., Worsley, K. J., Poline, J. B., Frith, C. D.,and Frackowiak, R. S. J. 1995. Statistical parametric maps infunctional imaging: A general approach. Hum. Brain Mapp. 2:189–210.affan, D., Harrison, S., and Gaffan, E. A. 1986. Visual identificationfollowing infero-temporal ablation in the monkey. J. Exp. Psychol.38:5–30.rady, C. L., Horwitz, B., Pietrini, P., Mentis, M. J., Ungerleider,L. G., Rapoport, S. I., and Haxby, J. V. 1996. Effect of task difficultyon cerebral blood flow during perceptual matching of faces. Hum.Brain Mapp. 4:227–239.rafton, S. T., Mazziotta, J. C., Presty, S., Friston, K. J., Frackowiak,R. S. J., and Phelps, M. E. 1992. Functional anatomy of humanprocedural learning determined with regional cerebral blood flowand PET. J. Neurosci. 12:2542–2548.rafton, S. T., Woods, R. P., and Tyszka, M. 1994. Functional imagingof procedural motor learning: Relating cerebral blood flow withindividual subject performance. Hum. Brain Mapp. 1:221–234.reen, D. M., and Swets, J. A. 1966. Signal Detection Theory andPsychophysics. Wiley, New York.aier, R. J., Siegel, B. V., Jr., MacLachlan, A., Soderling, E., Lotten-berg, S., and Buchsbaum, M. S. 1992. Regional glucose metabolicchanges after learning a complex visuospatial/motor task: A posi-tron emission tomographic study. Brain Res. 570:134–143.arris, R. J. 1975. A Primer of Multivariate Statistics. AcademicPress, New York.axby, J. V., Grady, C. L., Horwitz, B., Ungerleider, L. G., Mishkin,M., Carson, R. E., Herscovitch, P., Schapiro, M. B., and Rapoport,S. I. 1991. Dissociation of object and spatial visual processingpathways in human extrastriate cortex. Proc. Natl. Acad. Sci. USA88:1621–1625.axby, J. V., Horwitz, B., Ungerleider, L. G., Maisog, J. M., Pietrini,P., and Grady, C. L. 1994. The functional organization of humanextrastriate cortex: A PET–rCBF study of selective attention tofaces and locations. J. Neurosci. 14:6336–6353.axby, J. V., Ungerleider, L. G., Horwitz, B., Rapoport, S. I., andGrady, C. L. 1995. Hemispheric differences in neural systems forface working memory: A PET–rCBF study. Hum. Brain Mapp.3:68–82.oward, D., Patterson, K., Wise, R., Brown, D., Friston, K., Weiller,C., and Frackowiak, R. 1992. The cortical localization of thelexicons. Positron emission tomography evidence. Brain 115:1769–1782.
enkins, I. H., Brooks, D. J., Nixon, P. D., Frackowiak, R. S. J., andPassingham, R. E. 1994. Motor sequence learning: A study withpositron emission tomography. J. Neurosci. 14:3775–3790.
uliano, S. L., Hand, P. J., and Whitsel, B. L. 1981. Patterns ofincreased metabolic activity in somatosensory cortex of the monkeyMacaca fascicularis, subjected to controlled cutaneous stimulation:A 2-deoxyglucose study. J. Neurophysiol. 46:1260–1284.
uliano, S. L., Whitsel, B. L., and Hand, P. J. 1983. Patterns ofmetabolic activity in cytoarchitectural area SII and surroundingcortical fields of the monkey. J. Neurophysiol. 50:961–980.
adekaro, M., Grane, A. M., and Sokoloff, L. 1985. Differential effects Rof electrical stimulation of sciatic nerve on metabolic activity inspinal cord and dorsal root ganglion in the rat. Proc. Natl. Acad.Sci. USA 82:2337–6013.awashima, R., Satoh, K., Goto, R., Inoue, D., Itoh, M., and Fukuda,H. 1998. The role of the left inferior temporal cortex for visualpattern discrimination—A PET study. NeuroReport 9:1581–1586.oepp, M. J., Labbe, D., Richardson, M. P., Brooks, B. J., Paesschen,W., Cunningham, B. J., and Duncan, J. S. 1987. Regional hippocam-pal [11C] flumazenil PET in temporal lobe epilepsy with unilateraland bilateral hippocampal sclerosis. Brain 120:1865–1876.osslyn, S. M., Alpert, N. M., Thompson, W. L., Chabris, C. F., Rauch,S. L., and Anderson, A. K. 1994. Identifying objects seen fromdifferent viewpoints. A PET investigation. Brain 117:1055–1071.aunsell, J. H. R., and Newsome, W. T. 1987. Visual processing inmonkey extrastriate cortex. Annu. Rev. Neurosci. 10:363–401.cIntosh, A. R., Grady, C. L., Haxby, J. V., Ungerleider, L. G.,Rapoport, S. I., and Horwitz, B. 1996. Changes in limbic andprefrontal functional interactions in a working memory task forfaces. Cereb. Cortex 6:571–584.ishkin, M., Ungerleider, L. G., and Macko, K. A. 1983. Object visionand spatial vision: Two cortical pathways. Trends Neurosci. 6:414–17.oeller, J. R., Strother, S. C., Sidtis, J. J., and Rottenberg, D. A. 1987.Scaled subprofile model: A statistical approach to the analysis offunctional patterns in positron emission tomographic data. J.Cereb. Blood Flow Metab. 7:649–658.oeller, J. R., and Strother, S. C. 1991. A regional covarianceapproach to the analysis of functional patterns in positron emissiontomographic data. J. Cereb. Blood Flow Metab. 11:A121–A135.oore, C. J., and Price, C. J. 1998. Three distinct ventral occipitotem-poral regions for reading and object naming. NeuroImage 10:181–192.agahama, Y., Sadato, N., Yamauchi, H., Katsumi, Y., Hayashi, T.,Fukuyama, H., Kimura, J., Shibasaki, H., and Yonekura, Y. 1998.Neural activity during attention shifts between object features.NeuroReport 9:2633–2638.eelin, P., Crossman, J., Hawkes, D. J., Ma, Y., and Evans, A. C. 1993.Validation of an MRI/PET landmark registration method using 3Dsimulated PET images and point simulations. Comput. Med. Imag.Graphics 17:351–356.
errett, D. I., Mistlin, A. J., and Chitty, A. J. 1987. Visual neuronesresponsive to faces. Trends Neurosci. 10:358–364.
etersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., and Raichle,M. E. 1989. Positron emission tomographic studies of the process-ing of single words. J. Cognit. Neurosci. 1:153–170.
etersen, S. E., Fox, P. T., Snyder, A. Z., and Raichle, M. E. 1990.Activation of extrastriate and frontal cortical areas by visual wordsand word-like stimuli. Science 249:1041–1044.
helps, M. E., Huang, S. C., Hoffman, E. J., Selin, C., Sokoloff, L., andKuhl, D. E. 1979. Tomographic measurement of local cerebralglucose metabolic rate in humans with [18F]2-fluoro-2-deoxy-2-D-glucose: Validation of method. Ann. Neurol. 6:371–388.
rice, C. J., Moore, C. J., Humphreys, G. W., Frackowiak, R. S. J., andFriston, K. J. 1996. The neural regions sustaining object recogni-tion and naming. Proc. R. Soc. London 262:1501–1507.
uce, A., Allison, T., Asgari, M., Gore, J. C., and McCarthy, G. 1996.Differential sensitivity of human visual cortex to faces, letter-strings, and textures: A functional magnetic resonance imagingstudy. J. Neurosci. 16:5205–5215.
aichle, M. E., Fiez, J. A., Videen, T. O., MacLeod, A.-M. K., Pardo,J. V., Fox, P. T., and Petersen, S. E. 1994. Practice-related changesin human brain functional anatomy during nonmotor learning.Cereb. Cortex 4:8–26.
oland, P. E., and Gulyas, B. 1995. Visual memory, visual imagery,
S
S
S
S
S
S
S
T
T
U
U
U
W
W
W
W
W
W
W
W
Y
123CORTICAL ACTIVITY RELATED TO TASK ACCURACY
and visual recognition of large field patterns by the human brain:Functional anatomy by positron emission tomography. Cereb.Cortex 1:79–93.
chacter, D. L., Reiman, E., Uecker, A., Polster, M. R., Yun, L. S., andCooper, L. A. 1995. Brain regions associated with retrieval ofstructurally coherent visual information. Nature 376:587–590.
chlaug, G., Knorr, U., and Seitz, R. J. 1994. Inter-subject variabilityof cerebral activations in acquiring a motor skill: A study withpositron emission tomography. Exp. Brain Res. 98:523–534.
chwartz, E. L., Desimone, R., Albright, T. D., and Gross, C. G. 1983.Shape recognition and inferior temporal neurons. Proc. Natl. Acad.Sci. USA 80:5776–5778.
ergent, J., Ohta, S., and MacDonald, B. 1992. Functional neuro-anatomy of face and object processing. A positron emission tomogra-phy study. Brain 115:15–36.
haywitz, B. A., Pugh, K. R., Constable, R. T., Shaywitz, S. E.,Bronen, R. A., Fulbright, R. K., Shankweiler, D. P., Katz, L.,Fletcher, J. M., Skudlarski, P., and Gore, J. C. 1995. Localization ofsemantic processing using functional magnetic resonance imaging.Hum. Brain Mapp. 2:149–158.
oma, Y., Sugishita, M., Kitamura, K., Maruyama, S., and Imanaga,H. 1989. Lexical agraphia in the Japanese language. Brain 112:1549–1561.
pielberger, C. D., Gorsuch, R. L., Luchene, R., Bagg, P. R., andJacobs, G. A. 1983. Manual for the State–Trait Anxiety Inventory(STAI)—Form Y. Consulting Psychology Press, Palo Alto, CA.
alairach, J., and Tournoux, P. 1988. Coplanar Stereotaxic Atlas ofthe Human Brain. Thieme, New York.
anaka, K., Saito, H., Fukuda, Y., and Moriya, M. 1991. Coding visualimages of objects in the infero-temporal cortex of the macaquemonkey. J. Neurophysiol. 66:179–189.ngerleider, L. G., Courtney, S. M., and Haxby, J. V. 1998. A neuralsystem for human visual working memory. Proc. Natl. Acad. Sci.
USA 95:883–890.ngerleider, L. G., and Haxby, J. V. 1994. ‘What’ and ‘where’ in thehuman brain. Curr. Opin. Neurobiol. 4:157–165.ngerleider, L. G., and Mishkin, M. 1982. Two cortical visualsystems. In Analysis of Visual Behavior (D. J. Ingle, M. A. Goodale,and R. J. W. Mansfield, Eds.), pp. 549–586. MIT Press, Cambridge,MA.echsler, D. 1981. Wechsler Adult Intelligence Scale—Revised. Psy-chological Corp., New York.ood, F. 1990. Functional neuroimaging in neurobehavioral re-search. In Neuromethods, Vol. 17, Neuropsychology (A. A. Boulton,G. B. Gaker, and M. Hiscock, Eds.). Humana Press, Clifton, NJ.ood, F., Armentrout, R., Toole, J., McHenry, L., and Stump, D. 1980.Regional cerebral blood flow during rest and memory activation ina patient with global amnesia. Brain Lang. 9:124–136.ood, F. B., and Flowers, D. L. 1999. Functional neuroanatomy ofdyslexic subtypes: A survey of 43 candidate regions with a factoranalytic validation across 100 cases. In Reading and AttentionDisorders: Neurobiological Correlates (D. D. Duane, Ed.), pp.131–161. Yorkton Press, Parkton.oodcock, R. W., and Johnson, M. B. 1989. Woodcock–JohnsonPsychoeducational Battery—Revised. DLM Teaching Resources,Allen, TX.oods, R. P. 1996. Correlation of brain structure and function. InBrain Mapping: The Methods (A. W. Toga and J. C. Mazziotta,Eds.), pp. 313–341. Academic Press, San Diego.orsley, K. J., Evans, A. C., Marrett, S., and Neelin, P. 1992. Athree-dimensional statistical analysis for rCBF activation studiesin human brain. J. Cereb. Blood Flow Metab. 12:900–918.orsley, K. J., Marrett, S., Neelin, P., Vandal, A. C., Friston, K. J., andEvans, A. C. 1996. A unified statistical approach for determiningsignificant signals in images of cerebral activation. Hum. BrainMapp. 4:58–73.
oung, M. P. 1992. Objective analysis of the topological organization
of the primate cortical visual system. Nature 358:152–154.











![[Posterior cortical atrophy]](https://static.fdokumen.com/doc/165x107/6331b9d14e01430403005392/posterior-cortical-atrophy.jpg)







