Controlled surface modification of tissue culture polystyrene for selective cell binding using...
10
Controlled surface modification of tissue culture polystyrene for selective cell binding using resilin-inspired polypeptides This article has been downloaded from IOPscience. Please scroll down to see the full text article. 2013 Biofabrication 5 035005 (http://iopscience.iop.org/1758-5090/5/3/035005) Download details: IP Address: 140.253.154.103 The article was downloaded on 17/06/2013 at 00:42 Please note that terms and conditions apply. View the table of contents for this issue, or go to the journal homepage for more Home Search Collections Journals About Contact us My IOPscience
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Controlled surface modification of tissue culture polystyrene for selective cell binding using...

Controlled surface modification of tissue culture polystyrene for selective cell binding using
resilin-inspired polypeptides
This article has been downloaded from IOPscience. Please scroll down to see the full text article.
2013 Biofabrication 5 035005
(http://iopscience.iop.org/1758-5090/5/3/035005)
Download details:
IP Address: 140.253.154.103
The article was downloaded on 17/06/2013 at 00:42
Please note that terms and conditions apply.
View the table of contents for this issue, or go to the journal homepage for more
Home Search Collections Journals About Contact us My IOPscience

IOP PUBLISHING BIOFABRICATION
Biofabrication 5 (2013) 035005 (9pp) doi:10.1088/1758-5082/5/3/035005
Controlled surface modification of tissueculture polystyrene for selective cellbinding using resilin-inspiredpolypeptidesAditya V Vashi1, John A M Ramshaw1,3, Veronica Glattauer1,Christopher M Elvin2, Russell E Lyons2 and Jerome A Werkmeister1
1 CSIRO Materials Science and Engineering, Private Bag 10, Clayton South, VIC 3169, Australia2 CSIRO Animal, Food and Health Sciences, Queensland Bioscience Precinct, 306 Carmody Rd,St Lucia, QLD 4067, Australia
E-mail: [email protected]
Received 17 January 2013Accepted for publication 20 May 2013Published 10 June 2013Online at stacks.iop.org/BF/5/035005
AbstractModified tissue culture polystyrene (TCP) surfaces have been fabricated by attachment ofrecombinant polypeptides based on Drosophila melanogaster resilin and the Anophelesgambiae resilin-like protein. The D. melanogaster polypeptide (Rec-1) was from the first exonof resilin and consisted of 17 very similar repeats of a 15 residue sequence. The A. gambiaepolypeptide consisted of 16 repeats of an 11 residue consensus sequence (An16). Polypeptideswere attached to the TCP surface through tyrosine-based photo-crosslinking using blue light incombination with (RuII(bpy)3)Cl2 and sodium persulfate. TCP that has been manufactured bymild oxidation has surface phenolic groups that are believed to participate in this crosslinkingprocess. X-ray photoelectron spectroscopy and contact angle analyses were used todemonstrate polypeptide binding. At higher coating concentrations of Rec-1 and An16, thesurface was passivated and fibroblasts no longer attached and spread. At coatingconcentrations of 1 mg ml−1 for Rec-1 and 0.1 mg ml−1 for An16, where the surface was fullypassivated against fibroblast attachment, addition of a cell attachment peptide,cyclo(Arg-Gly-Asp-D-Tyr-Lys) during coating and photo-crosslinking at >0.1 mg ml−1, led tothe restoration of fibroblast binding that was dependent on the integrin αV chain.
(Some figures may appear in colour only in the online journal)
1. Introduction
Resilin is an elastomeric protein that is found in mostinsects, typically in specialized regions of the cuticle, andhas several unique properties including high elasticity andhigh resilience [1, 2]. In vivo, the protein assemblies arestabilized by the formation of di-tyrosine crosslinks betweenthe numerous tyrosine residues [3, 4]. With the completionof the Drosophila melanogaster genome, a gene (CG15920)
3 Author to whom any correspondence should be addressed.
was tentatively identified for pro-resilin [5]. Subsequently,recombinant proteins for the first exon [6] and for the entiregene [7] have been produced. After crosslinking, either byperoxidase [7], which may reflect the in vivo process [8] or amore efficient Ru-based photochemical process [6, 9], theseproducts had very high elasticity and resilience, consistentwith native resilin [6, 7].
The amino acid sequence of the N-terminal, first exondomain of D. melanogaster resilin contains a major sectionconsisting of 17 highly similar repeating amino acid sequences[5, 6]. These are each typically 15 amino acids long,
1758-5082/13/035005+09$33.00 1 © 2013 IOP Publishing Ltd Printed in the UK & the USA

Biofabrication 5 (2013) 035005 A V Vashi et al
and include a constant Tyr-Gly-Ala-Pro (YGAP) sequence[6]. Subsequently, genes and recombinant proteins thatcontain repeating sequences that include a YGAP elementhave also been isolated from a number of other species,including the mosquito, Anopheles gambiae [10], the cat flea,Ctenocephalides felis and the buffalo fly, Haematobia irritansexigua [11]. Despite similarity to resilin, the A. gambiaeproteins may not have the same in vivo functional properties[12]. Novel, multimeric constructs have also been madeusing a recursive strategy based on the consensus sequencesderived from the repetitive sequence observed in resilin andin these other proteins. These include constructs based on theconsensus sequence AQTPSSYGAP, derived from A. gambiae[10]. Resilin-inspired polypeptides, containing multiple copiesof consensus repeat sequences, have been expressed as solubleproducts in Escherichia coli with structures ranging from 4 to64 repeats [10].
In the present study, we have examined the recombinantproduct of the first exon of the D. melanogaster resilin,Rec-1 [6], and the construct with 16 repeats of the 11residue A. gambiae consensus sequence (An16). The An16polypeptide could be produced as a soluble and stable proteinat greater than 400 mg l−1 using the Studier method for auto-induction of protein expression [10, 13]. The recombinantRec-1 and An16 have previously been shown to form rubber-like biomaterials when cast in glass molds [6, 10] using arapid, Ru-based photochemical crosslinking method [6, 9].This crosslinking method leads to the formation of dityrosinecrosslinks with high specificity between, and possibly within,the protein chains [6, 9]. However, in initial attempts to use24-well tissue culture plates as molds to make disks of cross-linked Rec-1, it was found, unexpectedly, that the disks couldnot be released from the plate. The hypothesis was that theprotein had reacted and covalently attached to the phenolicgroups that are present on tissue culture polystyrene (TCP).
TCP is based on polystyrene that has been modified bycontrolled oxidation to enhance cell attachment. A variety ofmethods have been used, including treating with H2SO4 [14],ultraviolet/ozone [15], gamma [16], electron beam irradiation[17] and plasma oxidation [18]. Little information on thepropriety processes that manufacturers use is available, butit seems likely that corona discharge or plasma treatments arenormally used [14, 19], which would induce a mild oxidationof the surface. With these mild conditions, hydroxyl additionoccurs, leading to formation of phenol entities as an importantreaction product [20]. With more vigorous approaches, suchas UV irradiation or gamma irradiation, the predominantreactions involve scission of the phenyl C–C ring bonds andof the backbone, leading to carboxylate formation [20].
In this study, we have used Rec-1 and An16 tofabricate thin coatings on tissue culture ware, through photo-crosslinking onto the surface. It was a surprising finding thatunder certain conditions these treated surfaces were no longeradherent to fibroblasts. This was examined further, and itwas found that the protein coating concentrations could betitrated to provide uniform surfaces on TCP that were notadherent to fibroblasts. However, at the lower concentrationsexamined some cell attachment was found, indicating that
neither of the proteins nor the process used for crosslinkingwas inherently cytotoxic. It was postulated that selected,passivated surfaces could possibly be selectively modified. Byincluding in the coating process with Rec-1 or An-16, a cellattachment peptide, cyclo(Arg-Gly-Asp-D-Tyr-Lys), that alsocontains a Tyr residue that is available for photo-crosslinking,it was found that fibroblast binding could be restored withcells attaching and spreading, on the otherwise non-adherentsurfaces. This approach could, therefore, provide a simplemethod for fabricating passivated TCP plates or for assessmentof peptides, singly or in combinations, for their cell interactioncapacities.
2. Materials and methods
2.1. Construct design and expression
A recombinant construct derived from the majority of thefirst exon of the Drosophila melanogaster CG15920 gene,encoding the elastomeric domain, from nucleotide 48 275 to49 183 [6], termed Rec-1 resilin, as previously described, wasused for recombinant protein expression [6]. A 946-base-pairfragment (including codons for an N-terminal His6 affinitytag) was inserted into a T7 promoter bacterial expressionvector pETMCSI [21]. Recombinant Rec-1 protein productionwas induced with IPTG in E. coli BL21 StarTM(DE3)/pLysS(Invitrogen). Cells were collected and stored frozen at −80 ◦Cprior to extraction of the recombinant protein.
As previously described [10], an 11 residue consensusamino acid sequence, AQTPSSQYGAP, based on therepetitive domains of A. gambiae (African malaria mosquito)resilin-like gene (GenBank accession no. BX619161) wasused to make a multimeric construct by modification of arecursive method for generating periodic polypeptides [22].The construct containing 16 repeats of this repeat sequence,termed An16, was also inserted into a T7 promoter bacterialexpression vector pETMCSI. Recombinant An16 productionalso used auto-induction of protein expression [13] in E. coliBL21 StarTM(DE3)/pLysS. Cells were collected and storedfrozen at −80 ◦C prior to extraction of the recombinant protein
2.2. Extraction and purification of soluble proteins
Frozen cell pellets, for An16 and Rec-1, were thawedand resuspended in lysis buffer (50 mM Tris HCl, pH7.2, 10 mM benzamidine HCl, 10 mM EDTA, 1% TritonX-100, 10 mM β-mercaptoethanol), and homogenized usingan UltraTurrax T25 (IKA), followed by an UltrasonicsTM
(Melbourne, Australia) A180 sonicator 10 mm ultrasonicprobe. Polyethyleneimine (PEI) solution was added to soluble,clarified protein supernatant to 0.5% final concentration andclear supernatant was collected by centrifugation. Ammoniumsulfate was added to a final concentration of 20% saturation.Precipitated protein was collected by centrifugation anddialyzed at 4 ◦C against phosphate-buffered saline (PBS).Dialyzed protein was heated at 80 ◦C for 10 min anddenatured proteins removed by centrifugation. Protein identityand purity were assessed by SDS-PAGE analysis [23]and by mass spectroscopy using a Voyager DESTR mass
2

Biofabrication 5 (2013) 035005 A V Vashi et al
spectrometer [10]. Protein concentrations were estimatedspectrophotometrically [10].
2.3. Coating of tissue culture polystyrene
Stock solutions of Rec-1 and An16 at 10 mg ml−1 were madein sterile PBS, and all further steps were also performed ina biohazard cabinet. Appropriate dilutions of 10 mg ml−1,1 mg ml−1, 0.1 mg ml−1 and 10 μg ml−1 were made in PBSand 100 μl per well was used to coat 96 well TCP plates(Nunc NucleonTM, Denmark). (TCP plates and their reactivitymay vary between manufacturers.) TCP plates were incubatedovernight at 4 ◦C and excess solution was removed from thewells using suction. Protein was crosslinked through additionof 100 μl per well of sterile filtered 2 mM (RuII(bpy)3)Cl2 and10 mM sodium persulfate (SPS) in PBS and then crosslinkedby irradiation for 20 s per well at a distance of 15–20 mm witha LED dental curing lamp (430–480 nm, peak wavelength455 nm ± 10 nm, 1200 mW cm−2 at source, 3M ESPETM S10LED curing Light). The peak wavelength of this light source,at 455 nm, is close to the preferred irradiation wavelength of452 nm [9]. Wells were then washed with PBS including 5 mMEDTA once, then washed twice with PBS.
For studies on coatings that contain the adhesion peptide,cyclo(Arg-Gly-Asp-D-Tyr-Lys) (cycloRGDyK) (PCI-3662-PI: Peptides International Inc., Louisville KY), the sameprotocol was used except that the coating solutions were10 mg ml−1 for Rec-1 and 100 μg ml−1 for An-16, with theadhesion peptide added at 1 mg ml−1 and at 0.1 mg ml−1 toboth coating solutions. As controls for the covalent surfacemodifications on TCP, identical solutions were added but notcross-linked by omission of either the 2 mM (RuII(bpy)3)Cl2 or10 mM sodium persulfate (SPS). In addition TCP plates werealso irradiated with the curing light.
2.4. Surface examination by x-ray photoelectron spectroscopy
To demonstrate the presence of protein on the TCPsurface after coating, x-ray photoelectron spectroscopy (XPS)analyses were performed using an AXIS HSi spectrometer(Kratos Analytical Ltd, Manchester, UK) equipped with amonochromated Al Kα x-ray source at a power of 144 W(12 mA, 12 kV). A hemispherical analyzer was used, operatingin the fixed analyzer transmission mode with a standardaperture (1.0 × 0.5 mm). Charging of the samples duringirradiation was compensated for by the internal flood gun,coupled with a magnetic immersion lens. The pressure inthe main vacuum chamber during analysis was typically5 × 10−6 Pa. Survey spectra were acquired at a pass energyof 320 eV to identify the elements present in the untreatedTCP and the coated TCP and spectra for individual peakswere recorded for further analysis and quantification (80 eVpass energy). Atomic concentrations were calculated from theintegral peak intensities of O 1 s, N 1 s and C 1 s spectra, usinga linear background and applying sensitivity factors suppliedby the instrument manufacturer [24].
2.5. Surface properties by contact angle measurements
The surface properties of the Rec-1 and An16 coated materialswere analyzed by sessile contact angle measurement withMilliQ water using a KSV Cam200 (Helsinki, Finland)instrument, based on a previously described method [25].
2.6. Cell culture
Cell adhesion was assessed using the common, highly adherentcell line, L929 fibroblasts that were added at 104 cells/wellfor 96-well TCP plates. Cells were grown for 24 h in MEMcontaining GlutaMax (Life Technologies), plus 10% v/vfoetal bovine serum, 1% NEAA (Gibco) and 1% Antibiotic-Antimycotic (Gibco-Invitrogen) at 37 ◦C in 5% CO2. Theextent of cell adhesion and spreading was photographedusing a Nikon TE2000-U microscope. The viability ofattached and spread cells was assessed using a Live/Dead-Viability/Cytotoxicity Kit assay (Molecular Probes) at 20 haccording to the manufacturer’s instructions, using a NikonTE2000-U fluorescent microscope. Live cells were stainedgreen by calcein AM while dead cells stained red with ethidiumhomodimer-1. Cell numbers (green or red) in representativeimages were counted using ImageJ software. Results ofpercentage viability are expressed as Means ± StandardError. A two-tailed Student’s t-test was used to evaluate thestatistical significance between comparisons of cell viabilityon unmodified TCP and on modified TCP where cell bindinghad been restored through addition of cycloRGDyK peptide.The specificity of cell binding to the RGD epitope on thecycloRGDyK modified surfaces was assessed using a ratanti-mouse monoclonal antibody, 12-0512 (eBioscience) thatis specific for the αV integrin chain that is involved withbinding to the L929 fibroblasts [26]. TCP was modified usingRec-1 at 10 mg ml−1 and An-16 at 0.1 mg ml−1, togetherwith cycloRGDyK peptide at 1 mg ml−1, as above. L929fibroblasts, at 104 cells/well for 96-well TCP plates, werepre-incubated with anti mouse CD51 αV chain antibody (12-0512, eBioscience) at 10 μg ml−1 for 30 min at 4 ◦C [27]. Cellattachment was then examined after 2 h and 20 h incubations.
3. Results
3.1. Protein attachment to tissue culture polystyrene plates
Protein attachment to the TCP plates, after coating andcrosslinking, using 10 mg ml−1 concentrations of purifiedRec-1 and An-16 (figure 1), and followed by standard washing,was validated by XPS and contact angle measurements. TheXPS data, normalized to C = 1.00 (table 1), showed thatcoating with both Rec-1 and with An16 increased the Ncontent of the surface, as well as elevating the O content,consistent with protein/polypeptide binding. The amount ofprotein binding was greater than for absorbed protein withoutcrosslinking. This was also the case for samples that had beenwashed with daily changes for ten days (table 1), showing thepersistence of the covalent crosslinking of the polypeptides tothe TCP surfaces.
The sessile contact angle measurements showed that thecontact angle for un-coated TCP was 62◦ ± 1◦ (S.D.), while
3

Biofabrication 5 (2013) 035005 A V Vashi et al
PeptidePeptide
Figure 1. A schematic representation of the photo-crosslinking modification process. (A) Tissue culture polystyrene (TCP), with activegroups arising from the oxidative modifications during manufacture, (B) polypeptide, such as Rec-1 or An16, absorbed onto the TCP,(C) new crosslinks, formed by photo-crosslinking, that covalently bind the polypeptide to the TCP, and (D) photo-crosslinking ofpolypeptide and Tyr-containing peptide, leading to covalent binding of the polypeptide and the peptide to the TCP.
Table 1. Elemental analysis by XPS for C, O and N concentrations(normalized to C = 1.00) on TCP and TCP coated with10 mg ml−1 Rec-1 and An16 and stabilized throughphoto-crosslinking.
Sample C O N
TCP 1.00 0.12 0.01TCP coated with 10 mg ml−1 Rec-1 1.00 0.39 0.23TCP coated with 10 mg ml−1 Rec-1 1.00 0.34 0.21plus 10 day washTCP with absorbed 10 mg ml−1 Rec 1 1.00 0.19 0.10and single washTCP coated with 10 mg ml−1 An16 1.00 0.36 0.21TCP coated with 10 mg ml−1 An16 1.00 0.37 0.21plus 10 day washTCP coated with 10 mg ml−1 An16 1.00 0.37 0.21and single wash
that for the Rec-1 coating was 27◦ ± 2◦ (S.D.), and that forAn-16 was 17◦ ± 2◦ (S.D.).
3.2. Cell culture on coated surfaces
Surfaces that had been modified with both Rec-1 and An-16were assessed for cell attachment using L-929 fibroblasts.With the Rec-1 coated surfaces (figure 2), those coated at10 mg ml−1 showed no fibroblast binding with the cellspresent in a rounded phenotype. However, at lower coatingconcentrations, from 1 mg ml−1 and below for Rec-1, thefibroblasts showed an increasing tendency to attach andshowed a spreading phenotype, such that at 10 μg ml−1 coatingconcentration, all cells were attached (figure 2).
A similar inhibition of cell attachment was seen forAn16 coated plates (figure 3). In this case, coatings from10 mg ml−1 down to 0.1 mg ml−1 all led to inhibition offibroblast attachment, as shown by the rounded cell phenotypeof all the cells. At the lower concentration of 10 μg ml−1, somecell attachment was observed, as shown by a few spreadingcells, although most cells still remained round and unbound(figure 3). Thus, the coating by An16 was more effective for agiven concentration of polypeptide than Rec-1.
Also, a range of control plates were examined, none ofwhich generated non-adherent surfaces (figure 4). Thus TCP
treated with either blue light or the reagents alone, 2 mM(RuII(bpy)3)Cl2 and 10 mM SPS in PBS, remained adherentto cells. Also TCP treated with Rec-1 or An16 at elevatedconcentrations (figure 4) but without the photo-crosslinking,followed by the standard EDTA/PBS then PBS washed alsoremained adherent to cells (figure 4).
Cell binding to otherwise non-adherent Rec-1 and An16coated surfaces could be restored by co-crosslinking of theRec-1 or An-16 with a cell binding peptide (figure 1). Inthe present case, the cycloRGDyK peptide was chosen asthis cyclic peptide is known to show a high degree ofspecificity for cell binding via the αVβ3 integrin [28] andit contains a Tyr residue that can participate in the photo-crosslinking process. In the present study the Rec-1 and An-16 concentrations were chosen on the basis of the previousresults (figures 3 and 4) and used at a dilution where therewas clearly no cell adhesion. For Rec-1 this was takenas a 10 mg ml−1 concentration, while for An16 it wastaken as a 100 μg ml−1 concentration. During coating, thecycloRGDyK peptide was added to the Rec-1 and An16coating solutions at 1 and at 0.1 mg ml−1, respectively. Afterphoto-cross-linking the surfaces were thoroughly washed. Atboth concentrations, for both Rec-1 and An16 coatings, theaddition of the cycloRGDyK during the crosslinking processled to recovery of fibroblast binding (figure 5), with themajority of cells showing a flattened, attached phenotype, withvery few rounded cells present. This was the case for both of thecycloRGDyK peptide concentrations in this study (figure 5),although lower concentrations may also be effective.
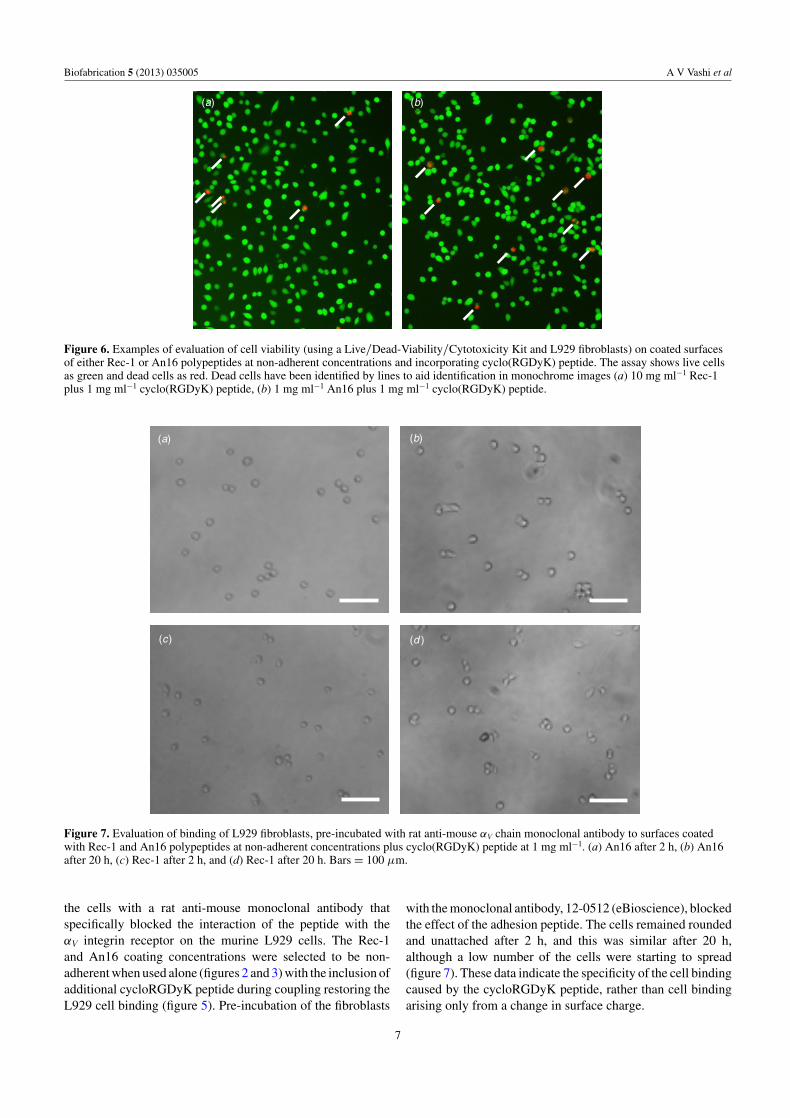
The observed effects on restoration of cell binding weredue to the protein and peptide coating. A lack of cytotoxicityfor the adherent, spreading cells was seen through useof Live/Dead-Viability/Cytotoxicity assay (figure 6). Thepercentage cell viability was 97.8 ± 0.6 for Rec-1/RGDsurfaces, while for An16/RGD surfaces, percentage cellviability was 96.4 ± 0.6. There were no significant differences(p > 0.05) when compared with themselves or when comparedto control, unmodified TCP surfaces, where a percentage cellviability of 98.8 ± 0.2 was observed.
The specificity of the cycloRGDyK peptide interactionwith the murine L929 fibroblasts was shown by pre-incubating
4

Biofabrication 5 (2013) 035005 A V Vashi et al
(a) (b)
(c) (d )
Figure 2. Binding of L929 fibroblasts to different coating concentrations Rec-1 polypeptide. (a) 10 mg ml−1 Rec-1, (b) 1 mg ml−1 Rec-1,(c) 0.1 mg ml−1 Rec-1, and (d) 10 μg ml−1 Rec-1. Bars = 100 μm.
(a) (b)
(c ) (d )
Figure 3. Binding of L929 fibroblasts to different coating concentrations An16 polypeptide. (a) 10 mg ml−1 An16, (b) 1 mg ml−1 An16,(c) 0.1 mg ml−1 An16, and (d) 10 μg ml−1 An16. Bars = 100 μm.
5

Biofabrication 5 (2013) 035005 A V Vashi et al
(a) (b)
(c) (d )
Figure 4. Binding of L929 fibroblasts to control TCP plates treated with, (a) blue light only, with no polypeptide, (b) 2 mM (RuII(bpy)3)Cl2 and 10 mM SPS in PBS only, with no polypeptide, (c) Rec-1 (300 mg ml−1) only followed by washing, and (d) An16 (230 mg ml−1)only followed by washing. Bars = 100 μm.
(a) (b)
(c) (d )
Figure 5. Binding of L929 fibroblasts to different coated surfaces of either Rec-1 or An16 polypeptides at non-adherent concentrations andincorporating cyclo(RGDyK) peptide. (a) 10 mg ml−1 Rec-1 plus 1 mg ml−1 cyclo(RGDyK) peptide, (b) 10 mg ml−1 Rec-1 plus0.1 mg ml−1 cyclo(RGDyK) peptide, (c) 0.1 mg ml−1 An16 plus 1 mg ml−1 cyclo(RGDyK) peptide, and (d) 0.1 mg ml−1 An16 plus0.1 mg ml−1 cyclo(RGDyK) peptide. Bars = 100 μm.
6
Biofabrication 5 (2013) 035005 A V Vashi et al
(a) (b)
Figure 6. Examples of evaluation of cell viability (using a Live/Dead-Viability/Cytotoxicity Kit and L929 fibroblasts) on coated surfacesof either Rec-1 or An16 polypeptides at non-adherent concentrations and incorporating cyclo(RGDyK) peptide. The assay shows live cellsas green and dead cells as red. Dead cells have been identified by lines to aid identification in monochrome images (a) 10 mg ml−1 Rec-1plus 1 mg ml−1 cyclo(RGDyK) peptide, (b) 1 mg ml−1 An16 plus 1 mg ml−1 cyclo(RGDyK) peptide.
(a) (b)
(c) (d )
Figure 7. Evaluation of binding of L929 fibroblasts, pre-incubated with rat anti-mouse αV chain monoclonal antibody to surfaces coatedwith Rec-1 and An16 polypeptides at non-adherent concentrations plus cyclo(RGDyK) peptide at 1 mg ml−1. (a) An16 after 2 h, (b) An16after 20 h, (c) Rec-1 after 2 h, and (d) Rec-1 after 20 h. Bars = 100 μm.
the cells with a rat anti-mouse monoclonal antibody thatspecifically blocked the interaction of the peptide with theαV integrin receptor on the murine L929 cells. The Rec-1and An16 coating concentrations were selected to be non-adherent when used alone (figures 2 and 3) with the inclusion ofadditional cycloRGDyK peptide during coupling restoring theL929 cell binding (figure 5). Pre-incubation of the fibroblasts
with the monoclonal antibody, 12-0512 (eBioscience), blockedthe effect of the adhesion peptide. The cells remained roundedand unattached after 2 h, and this was similar after 20 h,although a low number of the cells were starting to spread(figure 7). These data indicate the specificity of the cell bindingcaused by the cycloRGDyK peptide, rather than cell bindingarising only from a change in surface charge.
7

Biofabrication 5 (2013) 035005 A V Vashi et al
4. Discussion
The present study has demonstrated a simple, rapid andefficient method for controlling the surface binding propertiesfor cells of commonly used TCP culture ware, including flasks,plates or individual wells. Using the current approach, thesurface of TCP is readily modified so that it becomes non-adherent to fibroblasts. The method can be used to introducesimultaneously a peptide for selective attachment of cells.L929 cells were used as these are one of the commonestcell lines used in cell culture and are known to be stronglyadhesive to TCP. The attachment of the polypeptides, Rec-1and An16, with photo-crosslinking, was shown by both XPSand contact angle studies. Changes in surface compositiondemonstrated by XPS were consistent with the presence ofthe polypeptide backbone and N or O containing side chainsof the polypeptides attached to the surface. It seems highlylikely, given the stability of the polypeptide layer to extensivewashing, as shown by XPS (table 1), that the protein layeris crosslinked to the TCP as well as itself to give a stronglybonded layer through formation of dityrosine crosslinks.
The contact angle data showed that the two coatingshad substantially changed the observed contact angles.Commercially, TCP is treated to increase the hydrophilicityof the polystyrene substrate to improve cell attachment, withvariations existing between manufacturers. Good cell bindingis often observed for various cells at contact angles between50◦ and 70◦ [29, 30], but at higher angles, particularly above90◦, and at lower angles, particularly below 30◦, cell bindingis poor [29].
The present polypeptide coatings substantially reducedthe observed contact angles, to less than 30◦, giving conditionswhere fibroblasts no longer adhered. In the present case,we have used polypeptides based on resilin and resilin-like sequences, which contain Tyr residues from the YGAPrepeating motif that is reactive in photo-crosslinking. It iswell known that the hydrophobic or hydrophilic nature ofthe surface is one of many parameters that will impact oncell adhesion as mentioned above [29, 30]. In this case theintrinsic nature of the resilin or resilin-like sequences aresufficient to alter the cell adhesion properties of the surface,but the uniqueness of the photo-crosslinking approach isthat it is quick, cheap and simple to use; the presence ofdefined tyrosines allow these large polypeptides to be readilyintroduced and crosslinked to a non-specific surface eitheralone or in combination with more selective and specific cellbinding ligands, to generate a controlled and highly definedsurface. Other sequences containing a Tyr could also bedesigned that may be equally or more effective in preventingcell adhesion, but which could include sites for subsequentpeptide modification that do not rely on photo-crosslinking.The greater effectiveness of An16 compared to Rec-1 inproviding passivation probably reflected the differences inamino acid sequences, rather than molecular weight. TheRec-1 sequences contained almost 10% charged residues (1Glu, 2 Lys, 13 Asp and 14 Arg in 310 residues), whereas theAn16 is devoid of this type of residue.
The fabrication method used to form this layer is simpleand easy to perform as it is a solution chemistry approach
and does not need reactors or high energy light sources. Forexample, surface modification or passivation where UV or gasplasma methods have been used requires the availability ofmore complex apparatus [31]. In the present case, a dentallamp was used, but a quartz-halogen lamp is also suitable forthe irradiation during crosslinking [32].
The present method is suitable for single well passivationof TCP plates, and could potentially be used for generatingpatterned surfaces. For example, during photo-crosslinkinga light pattern could be used to give cross-linking only atspecific locations, followed by quenching of un-reacted SPSand washing of unbound protein, although the resolution thatcould be achieved would need further study and optimization.The method is also readily applied to modification of tissueculture flasks as these are transparent to the blue light that isused, and so a direct line of sight is not required. The coatingcan also be modified to introduce a specific function onto anotherwise non-adherent control surface. In the present case,this was achieved by using a peptide that contained a D-Tyrresidue that could participate in the crosslinking process. Inthe present example, the peptide cyclo(Arg-Gly-Asp-D-Tyr-Lys) was chosen as this cyclic peptide is known to show cellbinding, especially to newly growing endothelial cells [33],but also at a lesser extent to fibroblasts [26, 28], both via theαVβ3 integrin. However, other functional peptides, or ligandsincorporating a phenolic moiety, could be equally applicableand allow studies on a wide range of different cell types andfunctions.
If other peptides are to be examined they could besynthesized to contain a Tyr residue so that they couldbe incorporated during the photo-crosslinking. Alternatively,previously synthesized or commercially available peptidescould be modified to introduce a phenolic function, eitherthrough 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide-based coupling of tyramine to the carboxy-terminal of thepeptide, or by Bolton–Hunter reagent (N-succinimidyl-3-(4-hydroxyphenyl) propionate) to modify the amino-terminal ofa peptide. Peptides that contain an existing Tyr as part ofthe function, such as the laminin derived peptide Tyr-Ile-Gly-Ser-Arg [34] can be made using orthogonal protection duringpeptide synthesis [27] with final deprotection of functional Tyrresidues after peptide coupling to the plate.
5. Conclusions
Modification of TCP surfaces by coating with solutions ofRec-1 or An16 polypeptides, followed by photo-crosslinkingusing (RuII(bpy)3)Cl2 and SPS with brief, blue light irradiation,provides a simple and easy way of passivating the mildlyoxidized surface of TCP so that normally adherent fibroblastsno longer bind. The surface can be made selectively adherentby attachment of suitable peptides that are synthesized ormodified to contain a Tyr or other phenolic function and soparticipate in the crosslinking process. This coating methodcould, therefore, provide a simple method for assessmentof many peptides, singly or in combinations, for their cellinteraction capacities, or to provide passivated control surfaceswhen required for other studies.
8

Biofabrication 5 (2013) 035005 A V Vashi et al
Acknowledgments
We wish to thank Dr Thomas Gengenbach and Dr Chris Eastonfor assistance with XPS determinations.
References
[1] Jensen M and Weis-Fogh T 1962 Biology and physics oflocust flight: V. Strength and elasticity of locust cuticle Phil.Trans. R. Soc. London B 245 137–69
[2] Weis-Fogh T 1960 A rubber-like protein in insect cuticleJ. Exp. Biol. 37 889–907
[3] Andersen S O 1964 The cross-links in resilin identified asdityrosine and trityrosine Biochem. Biophys. Acta 93 213–5
[4] Andersen S O 1966 Covalent cross-links in a structuralprotein, resilin Acta Physiol. Scand. Suppl. 263 1–81
[5] Ardell D H and Andersen S O 2001 Tentative identification ofa resilin gene in Drosophila melanogaster Insect Biochem.Mol. Biol. 31 965–70
[6] Elvin C M, Carr A G, Huson M G, Maxwell J M,Pearson R D, Vuocolo T, Liyou N E, Wong D C C,Merritt D J and Dixon N E 2005 Synthesis and properties ofcrosslinked recombinant pro-resilin Nature 437 999–1002
[7] Qin G, Lapidot S, Numata K, Hu X, Meirovitch S, Dekel M,Podoler I, Shoseyov O and Kaplan D L 2009 Expression,cross-linking, and characterization of recombinant chitinbinding resilin Biomacromolecules 10 3227–34
[8] Coles G S 1966 Studies on resilin biosynthesis J. InsectPhysiol. 12 679–91
[9] Fancy D A and Kodadek T 1999 Chemistry for the analysis ofprotein-protein interactions: rapid and efficientcross-linking triggered by long wavelength light Proc. NatlAcad. Sci. USA 96 6020–4
[10] Lyons R E, Lesieur E, Kim M, Wong D C C, Huson M G,Nairn K M, Brownlee A G, Pearson R D and Elvin C M2007 Design and facile production of recombinantresilin-like polypeptides: gene construction and a rapidprotein purification method Protein Eng. Des. Sel. 20 25–32
[11] Lyons R E et al 2011 Molecular and functionalcharacterisation of resilin across three insect orders InsectBiochem. Mol. Biol. 41 881–90
[12] Andersen S O 2010 Studies on resilin-like gene products ininsects Insect Biochem. Mol. Biol. 40 541–51
[13] Studier F W 2005 Protein production by auto-induction inhigh-density shaking cultures Protein Exp. Purif. 41 207–34
[14] Curtis A S, Forrester J V, McInnes C and Lawrie F 1983Adhesion of cells to polystyrene surfaces J. Cell Biol.97 1500–6
[15] Teare D O H, Emmison N, Ton-That C and Bradley R H 2000Cellular attachment to ultraviolet ozone modifiedpolystyrene surfaces Langmuir 16 2818–24
[16] Onyiriuka E C, Hersh L S and Hertl W 1990 Surfacemodification of polystyrene by gamma-radiation Appl.Spectrosc. 44 808–11
[17] Onyiriuka E C 1994 Electron beam surface modification ofpolystyrene used for cell cultures J. Adhes. Sci. Technol.8 1–9
[18] Wells R K, Badyal J P S, Drummond I W, Robinson K Sand Street F J 1993 A comparison of plasma-oxidized andphoto-oxidized polystyrene surfaces Polymer 34 3611–3
[19] Onyiriuka E C, Hersch L S and Hertl W 1991 Solubilization ofcorona discharge- and plasma-treated polystyrene J. ColloidInterface Sci. 144 98–102
[20] Onyiriuka E C 1993 The effects of high-energy radiation onthe surface chemistry of polystyrene: a mechanistic studyJ. Appl. Polym. Sci. 47 2187–94
[21] Neylon C, Brown S E, Kralicek A V, Miles C S, Love C Aand Dixon N E 2000 Interaction of the Escherichia colireplication terminator protein (Tus) with DNA: a modelderived from DNA-binding studies of mutant proteins bysurface plasmon resonance Biochemistry 39 11989–99
[22] Elmorjani K, Thievin M, Michon T, Popineau Y, Hallet J Nand Gueguen J 1997 Synthetic genes specifying periodicpolymers modelled on the repetitive domain of wheatgliadins: conception and expression Biochem. Biophys. Res.Commun. 239 240–6
[23] Laemmli U K 1970 Cleavage of structural proteins duringassembly of head of bacteriophage-T4 Nature227 680–5
[24] Hamilton-Brown P, Gengenbach T, Griesser H Jand Meagher L 2009 End terminal, poly(ethylene oxide)graft layers: surface forces and protein adsorption Langmuir25 9149–56
[25] Truong Y B, Glattauer V, Lang G, Hands K, Kyratzis I L,Werkmeister J A and Ramshaw J A M 2010 A comparisonof the effects of fibre alignment of smooth and texturedfibres in electrospun membranes on fibroblast cell adhesionBiomed. Mater. 5 025005
[26] Massia S P and Hubbell J A 1991 An RGD spacing of 440 nmis sufficient for integrin alpha V beta 3-mediated fibroblastspreading and 140 nm for focal contact and stress fiberformation J. Cell Biol. 114 1089–100
[27] Taleski D, Butler S J, Stone M J and Payne R J 2011 Divergentand site-selective solid-phase synthesis of sulfopeptidesChem. Asian J. 6 1316–20
[28] van Hagen P M, Breeman W A P, Bernard H F, Schaar M,Mooij C M, Srinivasan A, Schmidt M A, Krenning E Pand de Jong M 2000 Evaluation of a radiolabelled cyclicDTPA-RGD analogue for tumour imaging and radionuclidetherapy Int. J. Cancer 90 186–98
[29] Tamada Y and Ikada Y 1993 Cell adhesion to plasma treatedpolymer surfaces Polymer 34 2208–12
[30] Arima Y and Iwata H 2007 Effect of wettability and surfacefunctional groups on protein adsorption and cell adhesionusing well-defined mixed self-assembled monolayersBiomaterials 28 3074–82
[31] Kingshott P, Thissen H and Griesser H J 2002 Effects ofcloud-point grafting, chain length, and density of PEGlayers on competitive adsorption of ocular proteinsBiomaterials 23 2043–56
[32] Elvin C M et al 2009 The development of photochemicallycrosslinked native fibrinogen as a rapidly formed andmechanically strong surgical tissue sealant Biomaterials30 2059–65
[33] Brooks P C, Clark R A and Cheresh D A 1994 Requirement ofvascular integrin alpha v beta 3 for angiogenesis Science264 569–71
[34] Graf J, Ogle R C, Robey F A, Sasaki M, Martin G R,Yamada Y and Kleinman H K 1987 A pentapeptide fromthe laminin B1 chain mediates cell adhesion and binds the67,000 laminin receptor Biochemistry 26 6896–900
9