
CONFORMING MARINE DECAPOD CRUSTACEANS
20
J. exp. Biol. (1978), T*. 107-126 It y7 With 8 figures Printed in Great Britain PATTERNS OF WATER AND SOLUTE REGULATION IN THE MUSCLE FIBRES OF OSMO- CONFORMING MARINE DECAPOD CRUSTACEANS BY ROBERT W. FREEL Department of Biology, University of California, Los Angeles, California 90024 (Received 26 May 1977) SUMMARY 1. The patterns of myoplasmic water and solute regulation were examined in several species of marine decapod crustaceans acclimated to various salinities. Three osmoconformers (Callianassa cahformemsis, Cancer anten- narius, Emerita analoga) and one weak osmoregulator {Pachygrapsus crassipes) were studied. 2. Although haemolymph water and solute activities varied with adap- tational salinity in these species, regulation of cell water content was apparent in all but 25 % s.w. acclimated Pachygrapsus. Cell water regulation was cor- related with the regulation of ninhydrin-positive solutes. An estimate of the extent of the regulation was achieved by comparing the observed cell hydration changes to that predicted for an ideal osmometer and to that of a perfect volume regulator. It was concluded that the extent of cell volume regulation was variable and incomplete, particularly in dilute salinities. 3. Net losses of myoplasmic sodium, chloride, and ninhydrin-positive nitrogen were observed in animals acclimated to dilute sea water, while net gains of these solutes were noted in crabs acclimated to concentrated sea water. In contrast, the amounts of cell potassium and calcium plus mag- nesium were relatively constant fractions of the cell dry weight at any salinity. 4. A simple mechanism, br.sed upon Donnan considerations, is sufficient to account for the constancy of myoplasmic potassium and the variability of cell chloride with changing extracellular osmolarity. INTRODUCTION A variety of marine decapod crustaceans survive substantial changes in the osmotic pressure of their environment (Kinne, 1971). Such tolerance is not restricted to those with moderately developed capacities of extracellular osmoregulation, but is also observed in strictly osmoconforming species. In both osmoconforming and weakly osmoregulating crustaceans the haemolymph osmotic pressure and ionic composition will vary with external salinity. Studies of the consequences of such variable haemo- lymph osmotic composition have, for the most part, centered about the phenomenon of isosmotic intracellular volume regulation (Schoffeniels & Gilles, 1970). Since the cell volume regulation process is not necessarily complete (Lange, 1970), alterations in
-
Upload
khangminh22 -
Category
Documents
-
view
5 -
download
0
Transcript of CONFORMING MARINE DECAPOD CRUSTACEANS

J. exp. Biol. (1978), T*. 107-126 I ty7With 8 figures
Printed in Great Britain
PATTERNS OF WATER AND SOLUTEREGULATION IN THE MUSCLE FIBRES OF OSMO-CONFORMING MARINE DECAPOD CRUSTACEANS
BY ROBERT W. FREEL
Department of Biology, University of California,Los Angeles, California 90024
(Received 26 May 1977)
SUMMARY
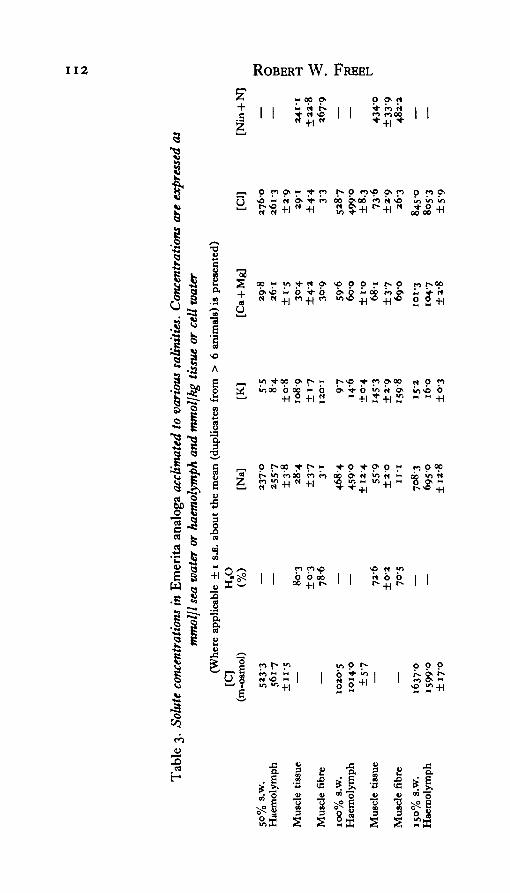
1. The patterns of myoplasmic water and solute regulation were examinedin several species of marine decapod crustaceans acclimated to varioussalinities. Three osmoconformers (Callianassa cahformemsis, Cancer anten-narius, Emerita analoga) and one weak osmoregulator {Pachygrapsus crassipes)were studied.
2. Although haemolymph water and solute activities varied with adap-tational salinity in these species, regulation of cell water content was apparentin all but 25 % s.w. acclimated Pachygrapsus. Cell water regulation was cor-related with the regulation of ninhydrin-positive solutes. An estimate of theextent of the regulation was achieved by comparing the observed cellhydration changes to that predicted for an ideal osmometer and to that of aperfect volume regulator. It was concluded that the extent of cell volumeregulation was variable and incomplete, particularly in dilute salinities.
3. Net losses of myoplasmic sodium, chloride, and ninhydrin-positivenitrogen were observed in animals acclimated to dilute sea water, while netgains of these solutes were noted in crabs acclimated to concentrated seawater. In contrast, the amounts of cell potassium and calcium plus mag-nesium were relatively constant fractions of the cell dry weight at any salinity.
4. A simple mechanism, br.sed upon Donnan considerations, is sufficientto account for the constancy of myoplasmic potassium and the variability ofcell chloride with changing extracellular osmolarity.
INTRODUCTION
A variety of marine decapod crustaceans survive substantial changes in the osmoticpressure of their environment (Kinne, 1971). Such tolerance is not restricted to thosewith moderately developed capacities of extracellular osmoregulation, but is alsoobserved in strictly osmoconforming species. In both osmoconforming and weaklyosmoregulating crustaceans the haemolymph osmotic pressure and ionic compositionwill vary with external salinity. Studies of the consequences of such variable haemo-lymph osmotic composition have, for the most part, centered about the phenomenon ofisosmotic intracellular volume regulation (Schoffeniels & Gilles, 1970). Since the cellvolume regulation process is not necessarily complete (Lange, 1970), alterations in

108 ROBERT W. FREEL
the concentrations of intracellular solutes not involved in the regulation process mayalso be observed.
With few exceptions (Shaw, 1955, 1958; Hays, Lang & Gainer, 1968), little atten-tion has been directed to the description and ultimate understanding of the effectsof- alterations in the ratios (haemolymph/cellular) of inorganic solutes in osmo-conforming or weakly osmoregulating marine crustaceans. The potential physio-logical consequences of such water and solute adjustments are manifold, rangingfrom the disruption of normal membrane electrophysiological properties (Pichon &Treherne, 1976) to alterations in the activities of intracellular biochemical pathways(Hochachka & Somero, 1973).
The present study was initiated to provide a comparative description of the patternsand magnitudes of intracellular water and solute regulation in decapod crustaceansinhabiting marine environments of varying degrees of osmotic stability. The ultimateaim of such studies is to provide a unified theoretical account for the observed patternsof intracellular water and solute adjustments in the tissues of these and other osmo-conforming marine organisms. The results of this study also serve as a foundation foranother report dealing with some passive membrane electrophysiological properties(Freel, 1977a) and for future studies on the active membrane properties of muscletissue from these crustaceans.
MATERIALS AND METHODSExperimental animals
Ghost shrimp, Callianassa californiensis Dana, were collected from the mud flatsat Morro Bay, California, and purchased from bait dealers in San Diego, California.The subtidal rock crab, Cancer antennarius Stimpson, the semi-terrestrial shore crab,Pachygrapsus crassipes Randall, and the sandy intertidal mole crab, Emerita analogaRathbun, were collected at various localities in Los Angeles County, California. Crabswere maintained in a recirculating sea water system containing natural sea water(100% s.w. 2: 1000 m-osmol) at 18 °C. Mature, intermoult specimens of eithergender were used in the following analyses.
Experimental animals were challenged with stepwise changes in salinity (ca. 100m-osmol per day) and subsequently maintained at the final acclimation salinity for aminimum of 5 days. The final lower acclimation salinity, selected on the basis ofpreliminary tolerance studies, varied for each species. These final dilutions werechosen such that the survivorship and activity of the experimental group was com-parable to control animals in natural sea water. An upper limit of approximately1500 m-osmol was arbitrarily chosen as the other experimental extreme. Reducedsalinities were made by tap water dilution of natural sea water, and 150% s.w. wasprepared by the addition of a synthetic marine salt mixture (Instant Ocean).
Analytical methods
The following analyses were carried out on the carpopodite flexor and extensormuscles dissected from the meropodites of the walking legs of the two brachyurancrabs (Cancer and Pachygrapsus). In the two anomurans (Callianassa and Emerita)the abdominal flexor and extensor musculature was used. Excised tissue samples,

Water and solute regulation in muscle fibres 109
weighing 25-100 mg, were vigorously rinsed in an isotonic sucrose solution for 5 sto remove adherent haemolymph, then blotted on filter paper prior to wet weightdetermination. After overnight drying at 105 °C, the inorganic solutes were extractedfrom the dried tissues in o-1 N - H N O 3 in sealed tubes for 24 h at 60 °C. This pro-cedure proved to be the most efficient and economical of the several commonlyemployed extraction techniques.
Sodium and potassium were determined with a Beckman DU flame spectrophoto-meter. Standard solutions were prepared to contain the appropriate levels of interfer-ing ions. Total calcium plus magnesium (Ca + Mg) was determined by microtitrationwith EDTA in buffered alkaline solutions using Eriochrome Black T as an indicator(Walser, i960). The osmolarities of the haemolymph and sea water samples weremeasured with a Mechrolab vapour pressure osmometer.
Free amino acids were determined as ninhydrin-positive nitrogen (Nin + N) onethanolic extracts of fresh muscle (25-100 mg) following the method of Clark (1973).Glycine was used as a standard. Since proline, taurine, and other low molecularweight nitrogenous solutes are not detected by ninhydrin, the Nin + N levels reportedhere are minimal estimates of the free amino acid pools of these crustacean tissues.
The extracellular space (ECS) of the muscle tissue was determined by injectingthe crabs with 25-50 /A of artificial haemolymph containing 14C-inulin (ICN Radio-chemicals). After equilibration of the probe (about 4 h), haemolymph samples weretaken and dispensed into scintillation vials. Muscle tissue of the same size and typeas used for the chemical analyses was excised and vigorously rinsed for 5 sec in label-free saline before blotting and weighing. Both muscle and haemolymph sampleswere prepared for liquid scintillation counting by digesting in 0-5 ml of a tissuesolubilizer (ICN Tissue Solubilizer) for several days at room temperature. Ten mlof a scintillation cocktail (Freel, Medler & Clark, 1973) were added to the digestedsamples. The samples were later counted in a Beckman liquid scintillation counter. Allcount rates were quench corrected.
RESULTS
Haemolymph solutes and osmotic pressure
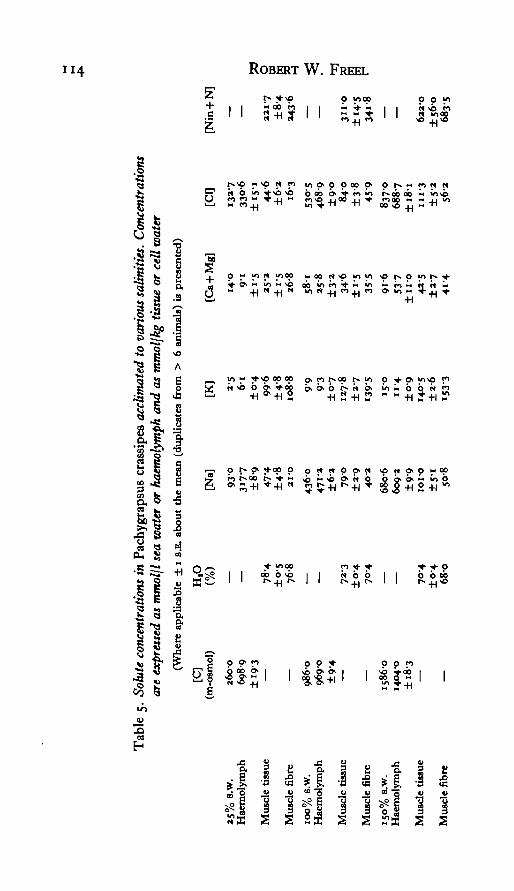
Both anomurans, Callianassa and Emerita, are essentially complete osmotic andionic conformers over the imposed salinity ranges (Tables 2, 3). Cancer is also anosmoconformer (Table 4), but it strongly regulates haemolymph Ca + Mg levels,as does the other brachyuran, Pachygrapsus (Table 5). In addition, Pachygrapsusis a weak osmoregulator in both concentrated and dilute sea water. The osmoregu-latory capacity of this species is the result of the moderate regulation of blood sodiumand chloride. It is evident, therefore, that haemolymph water activity varies con-siderably with adaptational salinity in each of the species considered here.
Muscle extracellular space
The 14C-inulin spaces for the different species and osmotic conditions are presentedin Table 1. The size of the extracellular space of Callianassa abdominal musculatureis independent of haemolymph osmolarity. The effect of salinity on the ECS of
muscle from Cancer, Pachygrapsus, and Emerita was not determined. Therefore, in

n o ROBERT W. FREEL
Table i. Extracellular space (ECS) estimates of the muscle tissue from various decapodsat different acclimation salinities. X±i s.E. (number of animals measured in triplicate)
Species
CaUianattaCallianatsaCalUantutaPachygrapsutEmeritaCancer
Sea water(%)
35IOO
1 5 0IOO
IOO
IOO
ECS(gm H,O/gm tissue H,O)
o-iai 10-015 (5)0-125±0-009 (7)013510-015(3)009010-005(9)0-10010-0x4(3)003910004(8)
the calculations of intracellular water and solute levels which follow the 100%s.w. ECS estimates were used for all salinity conditions in these three species.
The interspecific variation in the ECS measurements of muscle from 100% s.w.acclimated crabs is a clear demonstration of the morphological differences of themuscle masses examined. Callianassa and Emerita abdominal musculature consists oflarge, compact associations of many, relatively small fibres. The extensor musclesof the walking legs of Packygrapsus are composed of loosely associated bundles offewer, but larger muscle fibres. The bulk of Cancer carpopodite muscles are, incontrast, essentially single fibres which are readily separated in the tissue rinsingprocess. As shown in Table 1, even these single fibres have a measurable extracellularspace.
Total intracellular water
Muscle cell water contents were determined by subtracting the contribution ofextracellular water from the muscle tissue water values in Tables 2-5. It is apparentthat the changes in haemolymph water activity, incurred during salinity adaptation,produce changes in the muscle fibre water contents of these species (Tables 2-5).The magnitude of the observed change in cell water content is best expressed as therelative change in cell hydration (i?H,o) *& follows:
_ (gm ff2O/gm DW),
where the control (100% s.w.) and experimental values (subscripts 1 and 2, respec-tively) are given in grams of cell water per gram of dry cell weight (DW). In Fig. 1 thecalculated values of i?Hlo for the various species have been plotted as a function ofthe observed relative change in haemolymph osmotic pressure (CJCj). The data arepresented on log-log co-ordinates to emphasize the linear behaviour expected of aperfect osmometer (solid line in Fig. 1) in which i?H|o = CJCt. An i?H,o = I"°°implies strict regulation of both cell water and solids. A more realistic situation isdescribed by the hatched curve in Fig. 1. This curve describes the behaviour of aperfect volume regulator in which the cell volume is maintained by the accumulationor release of a solution isosmotic to Ca. In this model the weight of solids lost orgained in the regulation process is considered to be replaced by an equivalent amountof water. The /?H,o value for such a perfect volume regulator is dependent uponboth the initial cell water content (here taken to be 2-33 gm H20/gm DW) and theaverage molecular weight of the solutes involved in the regulation process. In Fig.

Tab
le 2
. Sol
ute
conc
entr
atio
ns in
Cal
liana
ssa
calif
orni
ensi
s ac
clim
ated
to
vari
ous
salin
ities
. C
once
ntra
tions
are
exp
ress
edas
mm
olfl
sea
wat
er o
r ha
emol
ymph
an
d m
mol
jkg
tissu
e or
cel
l w
ater
(Whe
re a
pp
licab
le ±
I S
.E. a
bou
t th
e m
ean
(d
up
licat
es f
rom
> 6
an
imal
s) U
pre
sen
ted
)
35%
s-w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
100%
s.w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
150%
s.w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
[C]
(m-o
smol
)
338-
53
61
3±
12
6
995-
610
17-0
±5-8
1660
-116
27-0
±22-
8
H,O 82
6±0
-380
7
72-8
±o-
670
-1
69
1±
0-3
65
9
[Na]
141-
114
3 3
±5-
82
48
±0-
58-
547
2-2
46
48
±3
774
6±
55
189
7135
6891
±9-4
128-
1±7
-440
-6
[K]
39
47
±0
275
-8±o
-68
56
107
115
±O-2
1274
±2'O
1440
15
0I5
-I±
0-3
167-
0±
60
1907
[Ca
+ M
g]
19-2
192
±0
9249
±i-
5257
592
586
±i-
3397
±39
37O
83
4920
45
7±
2-8
38-5
[Cl]
1705
160-
8±
47
24
5±
0-9 57
5041
506-
0±
6-i
863
±4-4
263
81
27
751-
9±
96
160-
7±
8-5
68
4
[Nin
+ N
]
1580
±11-2
1797 -
£3469
§
±15-3
3
-3965
„
51
29
±35-6
593
1
^
Tab
le 3
. S
olu
te c
once
ntr
atio
ns
in E
mer
ita
anal
oga
accl
imat
ed t
o va
riou
s sa
lin
itie
s. C
once
ntr
atio
ns
are
expr
esse
d as
mm
ol\l
sea
wat
er o
r h
aem
olym
ph a
nd
mm
oljk
g ti
ssu
e or
cel
l w
ater
(Whe
re a
pplic
able
± i
S.B
. abo
ut t
he
mea
n (d
uplic
ates
fro
m >
6 a
nim
als)
is p
rese
nted
)[C
] H
,O(m
-osm
ol)
(%)
[Na]
[K
] [C
a +
Mg]
[C
l][N
in +
N]
50%
s.
w.
Hae
mol
ymph
Mus
cle
tissu
e
Mus
cle
fibr
e10
0%
s.w
.H
aem
olym
ph
Mus
cle
tissu
e
Mus
cle
fibr
e15
0%
s.w
.H
aem
olym
ph
5233
5617
±n-
5— —
1020
-510
14-0
±5
7— —
1637
015
990
±1
70
— — 80
3±
03
78-6
— — 73-6
±0
-37O
-5
—
2370
2557
±3
82
84
±3
73
1
4684
459O
±12
-45
59
±3
0II
-I
70
83
69
50
±13
-8
55
8-4
±o-
810
89±
17
I2O
-I
97
14
6
±0-
414
53±
39
15
98
15
316
-0±0
-3
39-8
36-1
±1-5
30-4
±4-
23
09
59-6
6o-o
±t-
o6
81
±3
769
-0
101-
310
47±3
-8
376-
036
1-3
±3-9
29
1
±4-
43
353
8749
90±
8.3
73-6
±3-9
36
3
845-
080
5-3
±5
9
— — 341-
1±2
2-8
3679
434O
±33
'948
33— —

Ta
ble
4.
Solu
te co
ncen
tra
tio
ns
in C
an
cer
an
ten
nari
us
accli
mate
d t
o v
ari
ous
sali
nit
ies.
C
oncentr
ati
ons
are
exp
ress
ed
as
mrn
olj
l se
a w
ate
r or
haem
oly
mph and a
s m
molj
kg ti
ssue o
r cell
wate
r
(Wh
ere
app
licab
le ±
i s
.E. a
bout
th
e m
ean
(d
up
lica
tes
from
>
6 a
nim
als)
is
pre
sen
ted
)
6o%
s.w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
I O
O%
S.W
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
150%
s.w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
[C]
(m-o
smol
)
6oo-
o61
3-1
±8
2— — 99
55
997-
8±
7-0
— —
1514
-015
020
±3-
5— —
H,O
(%)
— — 76
5±
04
75-8
— — 72
5±
03
71
7
— — 70
9
±0-
570
-1
[Na]
28
22
27
69
+ I
3-5
20
5
±2-
3IO
-I
47
00
46
96
±5
63
50
±1-
21
74
68
50
67
84
±8
755
'4±
2-8
30
1
[K]
6-o
77
±0
311
1-3
±3
4"
5-5
IO-I
12
5
±0
114
85±
3-6
1540
15
01
52
±0-
316
88±3
»17
5-0
[Ca
+ M
g]
38
02
44
±0
72
58
±2
82
59
6o-o
36
1±
i-3
32-9
±3
03
28
93 4
56-5
±2
-63
77
±2
73
69
[Cl]
30
50
291-
0±
38
25
2
±i-
91
41
5<H
45
02
5±
70
48
2
±1
729
-8
74
50
743-
6±
59
88
5±
2-6
61-o
[Nin
+ N
]
— — 274-
7±
IO
-I2
85
8
— 44
57
±2
8-4
46
38
— — 6350
±25
-16607
a c £. c 1 a «<> s"
Tab
le 5
. So
lute
con
cent
ratio
ns in
Pac
hygr
apsu
s cr
assi
pes
accl
imat
ed t
o va
riou
s sa
liniti
es.
Con
cent
ratio
nsar
e ex
pres
sed
as m
mol
jl se
a w
ater
or
haem
olym
ph a
nd
as m
mol
lkg
tissu
e or
cel
l w
ater
(Wh
ere
app
lica
ble
±
1 s
.E.
abou
t th
e m
ean
(d
up
lica
tes
from
>
6 an
imal
s) i
s p
rese
nte
d)
25%
8.W
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
100%
s.w
.H
aem
olym
ph
Mu
scle
tis
sue
Mu
scle
fib
re
ISO
% 8
.W.
Hae
mol
ymp
h
Mu
scle
tis
sue
Mu
scle
fib
re
[C]
(m-o
smol
)
260-
069
89±
19
3— — 08
6096
90±
94
— —15
860
1404
-0±1
8-3
— —
H,O
(%)
— — 78-4
±0-5
76
8
— — 7*
3±0
-47
04
— 70-4
±0-4
68
0
[Na]
93
°31
77±8
-947
-4±4
-82I
-O
4360
471-
a±
62
79-0
±a-9
40-2
680-
660
9-2
±9
9IO
I-O
±5-
i5
08
[K]
2-5
6-i
±0
49
96
±4-
810
8-8
99
93
±0
712
7-8
±2-
713
951
50
u-4
±0-
914
05±
2-6
15
33
[Ca
+ M
g]
14
09
1
±1-5
25-2
±i-
52
68
58
1»5
-8±3
-*3
46
±i-
53
55
91-6
53-7
± II
-O42
-5±
27
4i 4
[cq
1327
330-
6±
15
-144
-6±
62
16
3
53O
-54
68
9±
90
84-0
±3
845
9
837O
68
87
±1
81
m-3
±5
-35
62
[Nin
+ T
— — 221-
7±
84
2436
— — 311-
0±
14
534
1-8
— 622-
0±
56
068
35

Water and solute regulation in muscle fibres
2-8
2-4
2-0>
1-8
1-6
1-4
1-2
10
ORu o
0-6
O-
1
/
/Complete volume
regulation
-
X+ \
•0
ocA
+
i i
Callianassa
Cancer
Pachygrapsus
Emerita
\ ^
i i i i\i
- 85
- 80
75 |c
s70 2
cd
- 65 3
- 60
0-2 0-4 0-6 0-8
eye,1 0 1-2 1-4 1-6 1-8 2 0
Fig. i. The relative change in cell hydration as a function of the relative change in extracellularosmolarity. The corresponding cell water contents are presented on the right ordinate. Thelines describing perfect volume regulation (hatched curve) and an ideal osmometer (diagonalline) were generated assuming an initial water content of 70%. The closed and numberedcircles in this and the following figures represent: 1, Carcitau maenat (Shaw, 195s, 1958); a,Potamon nHoticut (Shaw, 1959); 3, IJbima emargmata (Gilles, 1970); 4, CalUnectes tapidus(Gerard & Gilles, 1972); 5, Nereis succmea (Freel et a!. 1973); 6, Bttfo vtridis (Gordon, 1965).
the limits on the hatched curve were generated by considering solutes having mole-cular weights ranging between 50 and 100 grams per mole. The utility of this approachlies in the fact that reasonable limits may be applied to the osmotic behaviour of agiven cell type. These then permit a more realistic assessment of the magnitude of cellvolume regulation within and between species subjected to various osmotic con-ditions.
Two major conclusions may be drawn from the results presented in Fig. 1. First,all of the crustaceans examined in the present study (with the exception of 25 % s.w.acclimated Pachygrapsus) regulate the water content of their muscle to varyingdegrees, particularly when exposed to hyperosmotic conditions. Secondly, the volumeregulation process is not necessarily complete, but even when it is, significant changesin cell water content would be observed.
Intracellular solute concentrations
The muscle tissue solute concentrations and apparent intracellular solute con-centrations are presented in Tables 2-5 for the different species and osmotic con-ditions. The apparent intracellular solute concentrations were computed from thefollowing relation:
[ (ECS)( 2 )1-(ECS) '
where [C ]̂ = apparent intracellular solute concentration, mmol/kg cell H2O; [Ct] =tissue solute concentration, mmol/kg tissue H2O; [C/(] = haemolymph solute concen-
n6
20
1-8
1-6
1-4
1-2
10
0-8
0-6
0-4
0-2
0
-
-
-
•
-
•
0
3«1«
" # 5
) 0-2 0-4
c A*4
»
0-6 0-8 1-0 1-2
eye,Fig. a
ROBERT
0 »6
1-4 1-6 1-8 20
w2-2
20
1-8
1-6
1-4
1-2
10
0-8
0-6
0-4
0-2n
FREEL--------
3
•
D 0-2 0-4 0-6 0-8
0
•
A
1 1 1 1 1 1 1
10 1-2 1-4 1-6 1-8 2-0 2-2
eye,Fig. 3
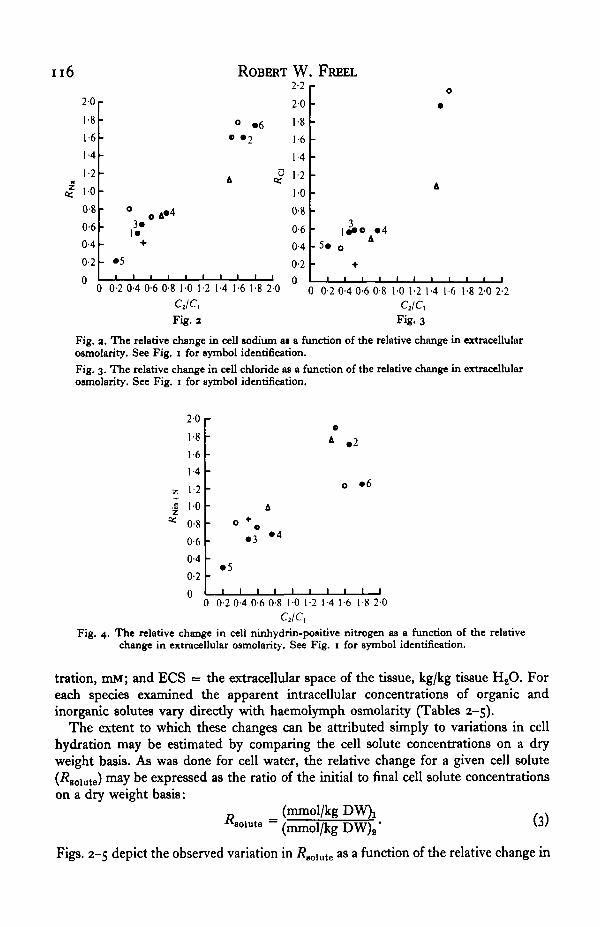
Fig. a. The relative change in cell sodium as a function of the relative change in extracellularosmolarity. See Fig. i for symbol identification.Fig. 3. The relative change in cell chloride as a function of the relative change in extracellularownolarity. See Fig. 1 for symbol identification.
zDC
20
8
6
4
•2
0
0-8
0-6
0-4
0-2
0
A . 2
• 6
»3 • 4
• 5
0 0-2 0-4 0-6 0-8 10 1-2 1-4 1-6 1-8 20C2/C,
Fig. 4. The relative change in cell ninhydrin-positive nitrogen as a function of the relativechange in extracellular osmolarity. See Fig. 1 for symbol identification.
tration, DIM; and ECS = the extracellular space of the tissue, kg/kg tissue H2O. Foreach species examined the apparent intracellular concentrations of organic andinorganic solutes vary directly with haemolymph osmolarity (Tables 2-5).
The extent to which these changes can be attributed simply to variations in cellhydration may be estimated by comparing the cell solute concentrations on a dryweight basis. As was done for cell water, the relative change for a given cell solute(^solute) n^y be expressed as the ratio of the initial to final cell solute concentrationson a dry weight basis:
(mmol/kg80lute ~ (mmol/kg DW)2"
(3)
Figs. 2-5 depict the observed variation in a s a function of the relative change in
Water and solute regulation in muscle fibres 117
1-6
1-4
1-2
10
08
0-6ftdJ *+
1-6
1-4
1-2
10
0-8
0-6
0-4
-(a)
+. 5 . o . 3 A
1* • «4•
-
i i i i i
3 0-2 0-4 0-6 0-8 10
o
• •+
1 1 1 1 1
A
I i
1-2 1-4
i i
• 2
*° .6
i i i
1-6 1-8 20
D
0
1 1 1
0 0 - 2 0 - 4 0 - 6 0 8 1 - 0 1 -2 1 - 4 1 -6 1-8 2 0CJC,
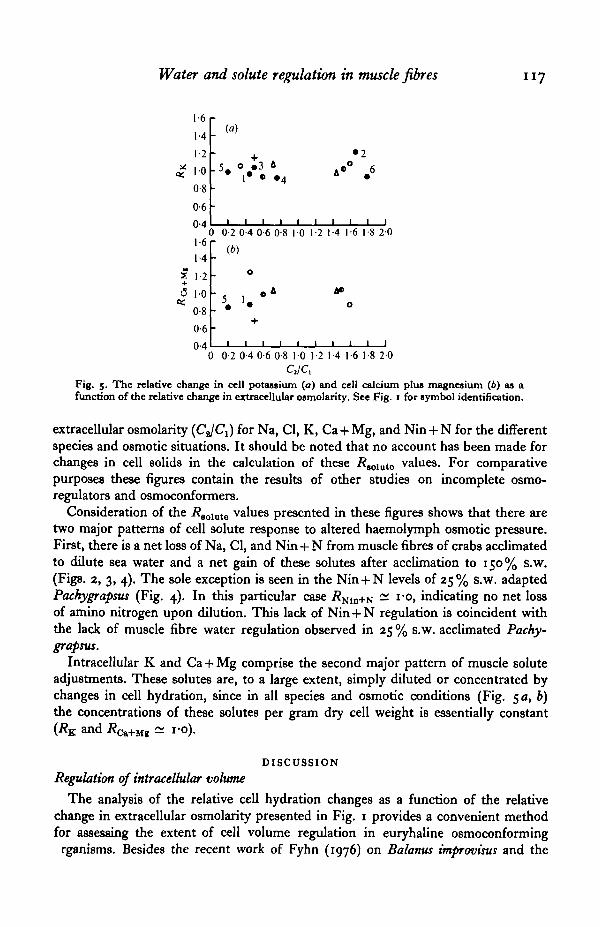
Fig. 5. The relative change in cell potassium (a) and cell calcium plus magnesium (6) as afunction of the relative change in extracellular osmolarity. See Fig. 1 for symbol identification.
extracellular osmolarity (CJCj) for Na, Cl, K, Ca + Mg, and Nin + N for the differentspecies and osmotic situations. It should be noted that no account has been made forchanges in cell solids in the calculation of these ^goiute values. For comparativepurposes these figures contain the results of other studies on incomplete osmo-regulators and osmoconformers.
Consideration of the /?aolnte values presented in these figures shows that there aretwo major patterns of cell solute response to altered haemolymph osmotic pressure.First, there is a net loss of Na, Cl, and Nin + N from muscle fibres of crabs acclimatedto dilute sea water and a net gain of these solutes after acclimation to 150% s.w.(Figs. 2, 3, 4). The sole exception is seen in the Nin + N levels of 25% s.w. adaptedPachygrapsus (Fig. 4). In this particular case Rmn+s — i-o, indicating no net lossof amino nitrogen upon dilution. This lack of Nin + N regulation is coincident withthe lack of muscle fibre water regulation observed in 25 % s.w. acclimated Pachy-grapsus.
Intracellular K and Ca + Mg comprise the second major pattern of muscle soluteadjustments. These solutes are, to a large extent, simply diluted or concentrated bychanges in cell hydration, since in all species and osmotic conditions (Fig. 50, b)the concentrations of these solutes per gram dry cell weight is essentially constant(RK and R^+iu ^ i-o).
DISCUSSIONRegulation of intracellular volume
The analysis of the relative cell hydration changes as a function of the relativechange in extracellular osmolarity presented in Fig. 1 provides a convenient methodfor assessing the extent of cell volume regulation in euryhaline osmoconforming
rganisms. Besides the recent work of Fyhn (1976) on Balanus improvisus and the

n 8 ROBERT W. FREEL
earlier studies of Lange & Mostad (1967) on euryhaline bivalves, no attempt has beenmade to quantify the capacity of intracellular volume regulation. The analysis ofLange & Mostad (1967) is more precise than that presented here in that the bounds ofthe regulation process are experimentally defined. Their method is, however, limitedto the few situations where tissue densities have been determined (Lange & Mostad,1967; Lange, 1970; Staaland, 1970). The major advantage of the present approachlies in its applicability to a rather large body of literature regarding cell or tissuewater contents of different organisms exposed to various osmotic conditions. Themajor disadvantage to this approach is in the estimation of the cell dry weight changesaccompanying the volume regulation process. The maximum capacity presented inFig. i was based upon the assumption that the average molecular weight of the solutesemployed during regulation ranged between 50 and 100 grams per mole. This is areasonable approximation since previous studies have documented the quantitativesignificance of the common, low molecular weight nitrogenous solutes in the regula-tory process (Schoffeniels & Gilles, 1970).
It is generally assumed in any analysis of cell volume regulation that all intracellularwater is osmotically active. However, in crustacean muscle fibres it has been repeatedlydemonstrated that only about 70% of the total intracellular water is osmoticallyactive (Hays et ah 1968; Hinke, 1970; Freel, 19776). If the osmotically active fractionrepresents a constant percentage of the total cell water, independent of extracellularosmolarity or cell hydration, then complications arising from the heterogeneousnature of cell water are resolved. This appears to be the case for isolated nerve fromEriocheir smensis (Gilles, 1973) and in Callianassa muscle (Freel, 19776). In the lattercrustacean, single fibre preparations from both 35% s.w. and 150% s.w. acclimatedspecimens have osmotically active volumes of about 65%.
From the foregoing discussion it is concluded that the relative change in cellhydration (#H,O) i8 a useful and reasonable approximation of the magnitude of cellvolume regulation. Based on such an analysis the results presented in Fig. 1 indicatethat the cell volume regulation process is not necessarily complete. This feature hasalso been observed in the isolated muscle fibres of Callinectes sapidus (Lang & Gainer,1969) and in the muscle of several bivalves (Lange, 1970). It may also be concludedfrom Fig. 1 that the magnitude of the regulatory response to hyposmotic stress is notnecessarily similar to that observed for an equivalent hyperosmotic stress. Thisimplies that a unified mechanism for cell volume control in hypo- and hyperosmoticsituations may not exist for these organisms. Distinct mechanisms are furtherindicated by the differences in the time courses for volume control of isolated tissueschallenged with hypo- or hyperosmotic salines (Lang & Gainer, 1969; Gilles, 1973).In the general scheme proposed by Schoffeniels (Schoffeniels & Gilles, 1970) theaccumulation and release of intracellular free amino acids is considered to be governedby the activity of key enzymes involved in amino metabolism. It is further proposedthat the activities of these enzymes are in turn modulated by the levels of intracellularions. In the case of Pachygrapsus this type of unified mechanism is not indicatedsince this crab cell volume regulates in concentrated salines, but not in dilute seawater, even though the changes in the intracellular inorganic ions are similar to thoseof the cell volume regulating species.
It is also interesting to point-out that the ability to regulate cell volume is noi

Water and solute regulation in muscle fibres 119
necessarily restricted to those crustaceans inhabiting osmotically unstable environ-ments, but is also found among the inhabitants of more strictly marine environmentssuch as Cancer and Emerita. Like the tolerances of the latter species to graduallyimposed salinity stresses, the cell volume regulating abilities of these typically open-coast crustaceans is interesting. It is difficult to imagine natural situations whereenvironmental osmolarities would ever reach the extremes imposed here experi-mentally. Ltbirna emarginata is another subtidal and reportedly stenohaline marinedecapod which has been shown to cell volume regulate in sea water concentrations aslow as 500 m-osmol (Gilles, 1970).
The above observations are consistent with the notion that laboratory salinitytolerances are not necessarily related to the salinity ranges encountered or tolerated bymarine organisms in their natural environment (Kinne, 1971). Thus, it would appearthat distinctions between ecological and physiological potential must be made whendiscussing the biological significance of cell volume regulation. The observationspresented here, together with the paucity of evidence documenting non-regulation,suggests that the physiological potential for cell volume regulation is more funda-mental to marine organisms than is generally appreciated.
Patterns of intracellular electrolyte regulation
Besides the regulation of cellular water contents by net amino acid exchanges,other features of intracellular solute regulation are apparent. The general patternwhich has emerged from the present study is that there is a net exchange of cellularNa and Cl during adaptation to new osmotic environments. In sharp contrast,cellular K and, to a large extent, cellular Ca + Mg are relatively constant per gramdry cell weight. These results are in accord with observations made on a variety ofother organisms for which the relevant data are available (Figs. 2-5).
Such consistent patterns of intracellular electrolyte regulation possibly reflect thepassive re-establishment of equilibria which had been disrupted by changes in theactivities of plasma water and solutes during salinity adaptation. If the transmem-brane ionic gradients of K and Cl are maintained by a Donnan equilibrium, then amodel may be proposed here to account for several of the observed patterns of inor-ganic solute regulation.
What is first required is a physical explanation for the experimental observationthat potassium, the major diffusible cation within the myoplasma, is a relatively con-stant fraction of the cell dry weight (RK ~ i-o). The fact that intracellular K concen-trations (mmol/kg cell water) lie within a relatively narrow range has been recognizedfor many years (Schlieper, 1958; Steinbach, 1962; Burton, 1968, 1973) and variousproposals have been presented to account for this phenomenon. For example, Stein-bach (1962) considered the ubiquity of the 150 IBM value for [K<] in terms of theevolution of cell systems, arguing that this particular cation is the most benevolent toprotoplasmic machinery. Such hypotheses provide little insight to the physicalnature of K accumulation in cells. Burton (1973) has provided a most interestingaccount of the significance of ion concentrations in animal body fluids and tissues.The starting point for his discussion is the classic analysis of Boyle & Con way (1941).However, the Burton model also considers the possibility that potassium is not inpassive equilibrium across the membrane, but rather that it is actively accumulated

120 ROBERT W. FREEL
in exchange for sodium. As a result of this Na/K exchange the ionic gradients areconsidered to be dependent upon the critical energy barrier to Na excretion from thecell as well as on the permeability characteristics of the cell membrane and theconcentration and valence of the non-diffusible solutes within the myoplasm.
The following analysis of the patterns of cellular ion adjustments observed in thedecapod crustaceans studied here is also based upon the model of Boyle & Conway(1941). In contrast to Burton's representation, the distribution of potassium is con-sidered to be entirely passive. Therefore, the major factors governing the ionicgradients across resting cell membranes are the concentration and charge of non-diffusible solutes within the myoplasm. Aside from the fact that this simplified schemeis sufficient to account for the experimental observations presented here and else-where (Freel, 1977 a) for the behaviour of K and Cl, the quantitative significance of aNa/K exchange pump in tissues such as crustacean muscle, where calcium carriesthe inward active current (Reuter, 1975), may be questionable. Furthermore, basedupon the experimental evidence presented here, it appears that the absolute amountof potassium per gram dry cell weight is more constant than is its concentrationper se. Indeed, the variations in cell K concentration can be quite large (86-191mmol/kg cell water in Callianassa, Table 2), but the bulk of such changes are broughtabout chiefly by water movements.
The present model considers the myoplasm of a marine osmoconforming organismto be in double Donnan equilibrium with the extracellular milieu. A specific andsimplified situation for an ideal muscle fibre in equilibrium with a solution resemblingsea water is depicted in Fig. 6. The definitions of the symbols used in this figure andin the equations which follow are given below.
[A~n] = concentration of intracellular, non-diffusible anion with a valence of n(mmol/kg cell HaO). For the present analysis n = 1.
[K,] .= total intracellular K concentration (mmol/kg cell HaO).[K,]A = intracellular K concentration acting as counter-ion to [A~n] (mmol/kg
cell H20).= intracellular K concentration acting as counter-ion to [Clj (mmol/kg
cell H20).= intracellular Cl concentration (mmol/kg cell H80).
[N] = concentration of intracellular non-diffusible, neutral solutes (mmol/kgcell H20).
V = volume of intracellular water (kg).[Ko], [Na0], and [Cl0] = extracellular ion concentrations (mM).[Q] and [Co] = molar sum of intra- and extracellular solutes, respectively.
For such an ideal system the following equilibrium relations are applicable (Boyle& Conway, 1941):
Osmotic equilibrium
[CJ = [Ai + [K,] + [Cy + [N] = [Ko] + [Na J + [Cl0] = [Co]. (4)
Macroelectroneutrality
(56)

Water and solute regulation in muscle fibres
Intracellular Extracellular
1 2 1
Non-diffusible
[A"](116-7)
[N](700)
ICI7](33-3)
^-~ Diffusible - -
t K i ] A
(116-7)
P^c(33-3)
(10)
- — —
[ci-](10)
[ci-](490)
*~—
K
Na
Non-diffusible
[NaJ](490)
Fig. 6. A simplified model of an ideal cell in osmotic, electrical, and Donnan equilibriumwith a sea water composed of only sodium, potassium, and chloride. The notation is ex-plained in the text. The numbers in parentheses refer to the concentrations (mM or mmol/kgcell water) of the various components of this hypothetical system. The water content ofthis cell is 3 gm HtO/gm dry weight (75%). Note that when extracellular osmolarity isaltered N becomes diffusible and represents the solute source for volume regulation.
Donnan equilibrium[KJ = [KJ [C1J. (6)
For the following it will also be convenient to describe the extracellular product of[KJ [Cl0] in terms of [Co]. For a perfect osmoconformer [Co] = [Ko] + [Na0] +[Cl0] and [KJ = [ClJ-[NaJ. Thus [Cl0] = | [CJ. Also, [Ko] = io-» [CJ fornormal sea water ([KJ = 10 mM; [CJ = 1 M). Thus the [KJ [C1J product maybe given as:
[ K J [ C 1 J = 5 X I O - 3 [ C J ' . (7)
How are the concentrations of potassium and chloride expected to change withadaptational salinity in an ideal osmoconformer? Given the equilibrium conditions(equations (4), (5 a), (56) and (6)) the variations in [Cy and [KJ may be calculatedstrictly in terms of [A~] and [CJ. By definition we have the following relationships:
where[KJci = [OJ and [KJA = [A-].
Therefore,[ K j [ c y = ( [A- ]+[cy) [cy .
From this relation, together with equations (6) and (7), we have the following:
[Cli]» + [A-] [Cy- S x io-»[CJ« = o.
Solving for [C1J we obtain the internal chloride concentration as a function of theextracellular osmotic concentration:
[Cy = (8)
122 ROBERT W. FREEL
1-8
1-6
1-4
1-2
1 0
0-6
0-4
0-2
0
Constant hydration
Constant volume
0 0-2 0 4 0-6 0-8 1 0 1-2 1-4
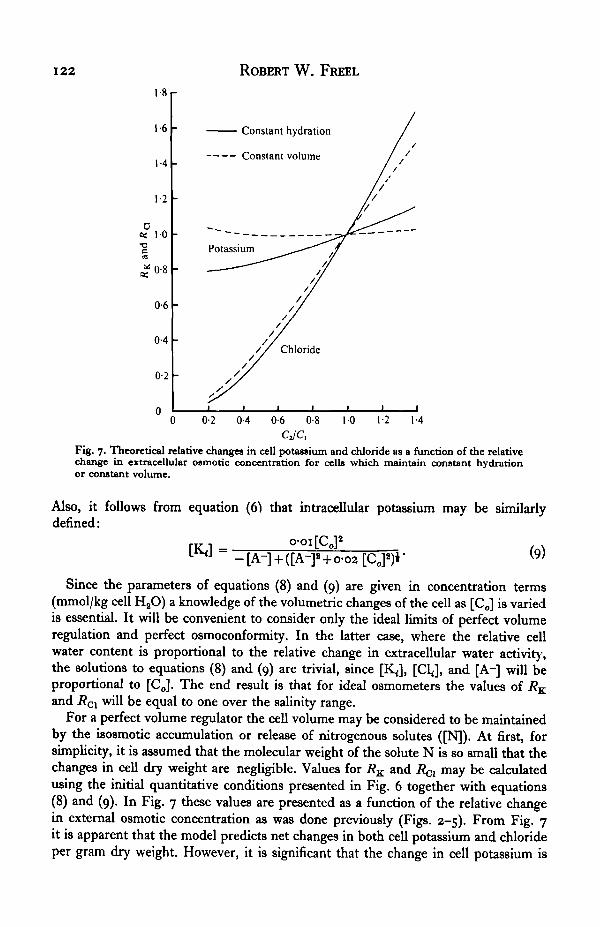
CJC,Fig. 7. Theoretical relative changes in cell potassium and chloride as a function of the relativechange in extracellular osmotic concentration for cells which maintain constant hydrationor constant volume.
Also, it follows from equation (6^ that intracellular potassium may be similarlydefined:
„ rr MM- (9)
Since the parameters of equations (8) and (9) are given in concentration terms(mmol/kg cell H2O) a knowledge of the volumetric changes of the cell as [Co] is variedis essential. It will be convenient to consider only the ideal limits of perfect volumeregulation and perfect osmoconformity. In the latter case, where the relative cellwater content is proportional to the relative change in extracellular water activity,the solutions to equations (8) and (9) are trivial, since [KJ, [Cl<], and [A~] will beproportional to [CJ. The end result is that for ideal osmometers the values of RK
and R01 will be equal to one over the salinity range.For a perfect volume regulator the cell volume may be considered to be maintained
by the isosmotic accumulation or release of nitrogenous solutes ([N]). At first, forsimplicity, it is assumed that the molecular weight of the solute N is so small that thechanges in cell dry weight are negligible. Values for RK and 7?^ may be calculatedusing the initial quantitative conditions presented in Fig. 6 together with equations(8) and (9). In Fig. 7 these values are presented as a function of the relative changein external osmotic concentration as was done previously (Figs. 2-5). From Fig. 7it is apparent that the model predicts net changes in both cell potassium and chlorideper gram dry weight. However, it is significant that the change in cell potassium is

Water and solute regulation in muscle fibres 123
considerably less than that of cell chloride. Any factor which increases the cell watercontent during adaptation to dilute sea water will tend to bring the RK and RO1 valuescloser to unity. It has already been noted that in a more realistic situation the celldry weight will change with adaptational salinity, since the molecular weight of N can-not be ignored. The dashed lines in Fig. 7 describe the behaviour of RK and R^ for aperfect volume regulator in which the dry weight is changing as a result of theaccumulation or release of a solute having a molecular weight of 100 grams/mole.In addition, it is assumed that water replaces or is displaced by such changes in celldry weight. Clearly, this more reasonable model for perfect volume regulation pre-dicts that cell potassium per gram dry cell weight will remain relatively constant,while chloride will vary with external osmotic concentration. This is precisely whatwas observed here experimentally (Figs. 3, 5 a).
In this analysis of ion gradient adjustments in osmoconforming marine organismsthe assumption has been made that the non-diffusible anion fraction (A~) and the freeamino acid pool (N) are not related. In striated muscle it can be expected that cellularprotein, together with the various phosphate fractions, comprise the bulk of the non-diffusible anion fraction, since the majority of the amino acids (glycine, alanine, andproline) should be neutral under normal cellular pH conditions.
The situation for the nervous tissues of marine osmoconformers is, however,likely to be quite different. Thus, in CalUnectes sapidus nerve, 94 % of the total freeamino acid composition is supplied by aspartate and glutamate (Gerard & Gilles,1972). These negatively charged solutes represent the bulk of the anion fractionrequired to balance the intracellular potassium concentration. If these amino acidswere also utilized in the volume regulation process, then the potassium concentrationper gram dry cell weight would be expected to be more variable than that observed formuscle. Unfortunately, it is difficult to state at the present time whether or not thisis the case, since no detailed studies have been directed towards this end. The chemicalanalyses of CalUnectes nerves at 50% and 100% s.w. supplied by Gerard & Gilles(1972) are difficult to interpret in this respect, since the magnitude of the relativewater exchange for these tissues was close to that expected for an osmometer andalso because the total solute change in these fibres was negligible, indicating littlecapacity for cell volume control. Certainly, cellular solute adjustments in the nervoustissues of osmoconforming marine organisms deserves further experimental attentionand would serve as a reasonable test of the model presented here.
As a final comment on the utility of the present model it is worth emphasizingthat in their original development Boyle & Conway (1941) were able to demonstratethat in the frog sartorius muscle the molar sum of all the non-diffusible solutes in themuscle fibre (7/V) was approximately equal to the molar sum of the non-diffusibleanion fraction (e/V). In terms of the present discussion of the adaptation of marineosmoconformers to dilute situations the significance of this observation (17/V = e/V)may be generalized as follows. Combining the definitions utilized here with those ofBoyle & Conway (1941) we have:
7/V=[C0]-(K,/V)-(Cl,/V) (10)and
(11)
124 ROBERT W. FREEL
3500 r
3000
^ 2500
af 2000oE£ 1500
QZ 1000
500
Constant volume
- Constant hydration
0 200 400 600[Co
800 1000 1200 1400
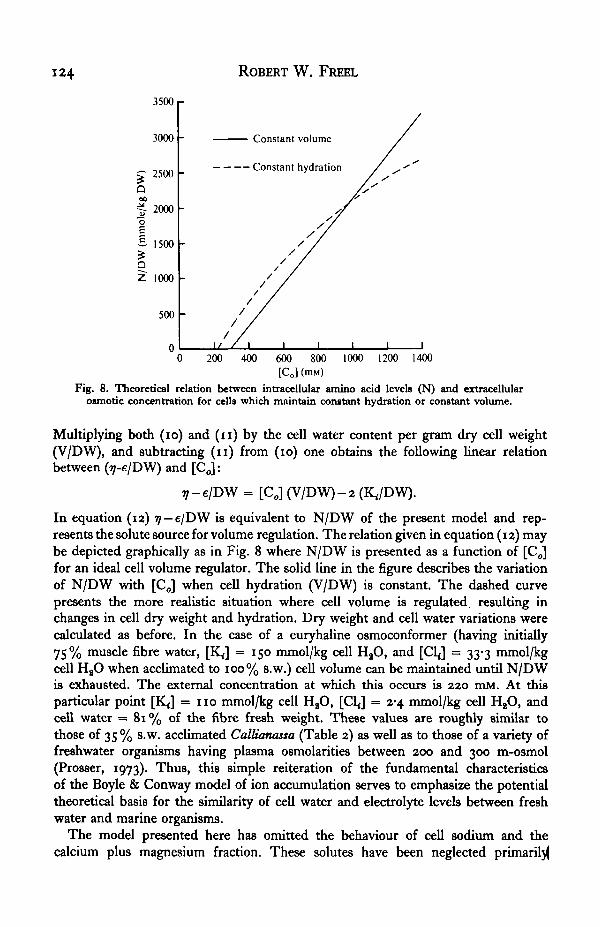
Fig. 8. Theoretical relation between intracellular amino acid levels (N) and extracellularosmotic concentration for cells which maintain constant hydration or constant volume.
Multiplying both (io) and (n ) by the cell water content per gram dry cell weight(V/DW), and subtracting (n ) from (io) one obtains the following linear relationbetween (ij-e/DW) and [C<J:
7-e/DW = [CJ (V/DW)-2 (K./DW).
In equation (12) rj — e/DW is equivalent to N/DW of the present model and rep-resents the solute source for volume regulation. The relation given in equation (12) maybe depicted graphically as in Fig. 8 where N/DW is presented as a function of [Co]for an ideal cell volume regulator. The solid line in the figure describes the variationof N/DW with [Co] when cell hydration (V/DW) is constant. The dashed curvepresents the more realistic situation where cell volume is regulated, resulting inchanges in cell dry weight and hydration. Dry weight and cell water variations werecalculated as before. In the case of a euryhaline osmoconformer (having initially75% muscle fibre water, [K ]̂ = 150 mmol/kg cell HaO, and [Cl<] = 33-3 mmol/kgcell H2O when acclimated to 100% s.w.) cell volume can be maintained until N/DWis exhausted. The external concentration at which this occurs is 220 mM. At thisparticular point [KJ = 110 mmol/kg cell HaO, [C1J = 2-4 mmol/kg cell HaO, andcell water = 81% of the fibre fresh weight. These values are roughly similar tothose of 35 % s.w. acclimated Callianassa (Table 2) as well as to those of a variety offreshwater organisms having plasma osmolarities between 200 and 300 m-osmol(Prosser, 1973). Thus, this simple reiteration of the fundamental characteristicsof the Boyle & Conway model of ion accumulation serves to emphasize the potentialtheoretical basis for the similarity of cell water and electrolyte levels between freshwater and marine organisms.
The model presented here has omitted the behaviour of cell sodium and thecalcium plus magnesium fraction. These solutes have been neglected primarily

Water and solute regulation in muscle fibres 125
because of the uncertainty of their physical status within the myoplasm and themechanism of their equilibrium disposition across the membrane. Equations similarto (8) and (9) cannot be developed for [Na ]̂ without introducing additional variables.It should also be pointed out that predicting [Na<] changes from relations such asthe Goldman-Hodgkin-Katz equation (Hodgkin & Katz, 1949) is useless, since[Na^] can take on a wide range of values without altering the [K ]̂ or [CLJ values whenthe permeability of the fibre to Na is low.
A most appropriate test of the Donnan model discussed here requires that theproducts of the concentrations (or more accurately, activities) of the diffusible ionsin the myoplasm equals their product on the outside (equation (6)). Previous attemptsto relate the concentration gradients of K and Cl across crustacean muscle fibremembranes, in terms of the Donnan principle, have met with varying success (seeFreel, 1977 a). The major difficulty arises from the use of apparent intracellularconcentrations without regard for the heterogeneous nature of cell water and solutesin these tissues (Hays et al. 1968; Hinke & Gayton, 1971; Freel, 19776). A quantita-tive appraisal of the potassium and chloride concentration gradients observed in thepresent study is likewise inadequate. Therefore, definitive statements concerningthe role of the Donnan principle in the transmembrane distributions of potassiumand chloride can only be achieved by considering the thermodynamic activity gradientsof these ions.
In another report (Freel, 1977 a) the transmembrane activity gradients for potassiumand chloride are presented for several of the marine decapods studied here. Theresults of this study support the hypothesis that these ions are distributed in accor-dance with the Donnan principle and that during the adaptation to new osmoticenvironments adjustments in the internal product of [K ]̂ [CLJ are met chiefly bynet changes in the intracellular chloride level.
This paper represents part of a Ph.D. dissertation submitted to the Department ofBiology, University of California at Los Angeles. I would like to thank Drs M. S.Gordon, G. N. Somero, S. Hagiwara, M. E. Clark, S. S. Hillman and Messrs C.Loretz and R. Putnam for their interest and helpful discussions throughout thedevelopment of this report. Part of this work was supported by the University ofCalifornia Patent Foundation and by the UCLA Laboratory of Fisheries and MarineBiology.
REFERENCES
BOYLE, P. J. & CONWAY, E. J. (1941). Potassium accumulation in muscle and associated changes.y. Pkyriol., Lond. 101, 1-63.
BURTON, R. F. (1968). Cell potassium and the significance of osmolarity in vertebrates. Comp. Biochem.Pkytiol. xj, nbz-mi.
BURTON, R. F. (1973). The significance of ionic concentrations in the internal media of animals. Biol.Rev. 48, 195-331.
CLARK, M. E. (1973). Amino acids in osmoregulation. In Experiments in Physiology and Biochemistry,vol. 6 (ed. G. A. Kerkut), pp. 81-114. London: Academic Press.
FREEL, R. W. (10770). Transmembrane activity gradients for K and Cl in the muscle fibres of osmo-conforming marine crustaceans. J. exp. Biol. 73, 127-140.
FREEL, R. W. (19776). Cellular hydration and solvent volumes of crustacean muscle fibres. In pre-paration.
FREEL, R. W. MEDLER, S. G. & CLARK, M. E. (1073). Solute adjustments in the coelomic fluid andmuscle fibres of a euryhaline polychaete, Neanthes sucdnea, adapted to various salinities. Biol. Bull.144, 280-303.
5 EXB 27

126 ROBERT W. FREEL
FYHN, H. J. (1976). Holeuryhalinity and its mechanism in a cirriped crustacean, Balanus improving.Comp. Biochem. Phytiol. 53A, 19-30.
GERARD, J. F. & GILLES, R. (197a). The free amino acid pool in Callinectet sapidut (Rathbun) tissuesand its role in the osmotic intracellular regulation. J. exp. mar. Biol. Ecol. 10, 125-136.
GILLES, R. (1970). Osmoregulation in the stenohaline crab Libma emarginata Leach. Archs. Int.Phytiol. Biochim. 78, 91-99.
GILLES, R. (1973). Osmotic behaviour of isolated axons of a euryhaline and a stenohaline crustacean.Experience 39, 1354-1355-
GORDON, M. S. (1965). Intracellular osmoregulation in skeletal muscle during salinity adaptation intwo species of toads. Biol. Bull. 138, 318-329.
HAYS, E. A., LANG, M. A. & GAINER, H. (1968). A re-examination of the Donnan distribution as amechanism for membrane potentials and potassium and chloride ion distribution in crab musclefibres. Comp. Biochem. Phytiol. 36, 761-792.
HINKE, J. A. M. (1970). Solvent water for electrolytes in the muscle fibres of the giant barnacle. J. gen.Phytiol. 56, 521-541.
HINKE, J. A. M. & GAYTON, D. C. (1971). Transmembrane K and Cl activity gradients for the musclefibre of the giant barnacle. Can. J. Phytiol. Pharmacol. 49, 312-322.
HOCHACHKA, P. W. & SOMERO, G. N. (1973). Strategies of Biochemical Adaptation. 358 pp. Philadelphia:Saunders.
HODGKIN, A. L. & KATZ, B. (1949). The effect of sodium on the electrical activity of the giant azon ofthe squid. J. Phytiol., Land. 108, 37-77.
KINNE, O. (1971). Salinity-invertebrates. In Marine Ecology, vol. 1 (ed. O. Kinne), pp. 821-995. NewYork: Wiley Interscience.
LANG, M. A. & GAINER, H. (1967). Volume control by muscle fibres of the blue crab. Volume readjust-ment in hypotonic salines. J. gen. Phytiol. 53, 323-340.
LANGE, R. (1970). Isosmotic intracellular regulation and euryhalinity in marine bivalves. J. exp. mar.Biol. Ecol. 5, 170-179.
LANGE, R. & MOSTAD, A. (1967). Cell volume regulation in osmotically adjusting marine animals.J. exp. mar. Biol. Ecol. x, 209-319.
PICHON, Y. & TREHERNB, J. E. (1976). The effects of osmotic stress on the electrical properties of theaxons of a marine oxmoconformer (Maia tquinado. Brachyura: Crustacea). J. exp. Biol. 65, 553-563.
PROSSER, C. L. (1973). Comparative AnimalPhytiology, 3rd edn., 966 pp. Philadelphia: W. B. Saunders.REUTER, H. (1975). Divalent cations as charge carriers in excitable membranes. In Calcium Movement in
Excitable Ceils, pp. 57-97. Oxford: Pergamon Press.SCHLIEPER, C. (1958). Sur l'adaptation des invertebres marins a l'eau de mer diluee. Vie tt Milieu 9,
139-152.SCHOFFKNIELS, E. & GILLES, R. (1970). Osmoregulation in aquatic arthropods. In Chemical Zoology,
vol. 5 (ed. M. Florkin and B. T. Scheer), pp. 255-286. New York: Academic Press.SHAW, J. (1955). Ionic regulation in the muscle fibres of Cardnus maenat. II. The effect of reduced
blood concentration. J. exp. Biol. 3a, 644-680.SHAW, J. (1958). Further studies on ionic regulation in the muscle fibres of Cardnut maenat. J. exp.
Biol. 35, 902-919.SHAW, J. (1959). Solute and water balance in the muscle fibres of the fresh water crab, Potamon mloticvs
(M. Edw.). j . exp. Biol. 36, 145-156.STAALAND, H. (1970). Volume regulation in the common whelk, Buccinum undulatum L. Comp. Biochem.
Physiol. 34, 355-365.STEINBACH, H. B. (1962). The prevalence of K. Perspect. Biol. Med. 5, 338-355-WALSER, M. (i960). Determination of free magnesium ions in body fluids. Improved methods for free,
calcium ions, total calcium, and total magnesium. Analyt. Chem. 33, 711-717.




















