
Components of task-set reconfiguration: Differential effects of ‘ switch-to’ and ‘...
17
Research Report Components of task-set reconfiguration: Differential effects of ‘switch-to’ and ‘switch-away’ cues Rebecca Nicholson a,b, ⁎ , Frini Karayanidis a,b,c , Anna Davies, Patricia T. Michie a,b,c a Functional Neuroimaging Lab, School of Psychology, University of Newcastle, Australia b Neuroscience Institute of Schizophrenia and Allied Disorders (NISAD), Australia c Hunter Medical Research Institute, Newcastle, Australia ARTICLE INFO ABSTRACT Article history: Accepted 26 August 2006 Available online 29 September 2006 Preparation for a switch in task was manipulated using two types of switch cues: ‘switch- away’ from the previous task-set and ‘switch-to’ a different task-set. Increasing cue–stimulus interval resulted in a reduction in reaction time switch cost for switch-to trials only. Cue- locked difference waveforms for both switch-to and switch-away trials showed a large, broad differential positivity, relative to repeat waveforms. However, the later part of the differential positivity was significantly reduced on switch-away trials. A differential positivity then emerged after stimulus onset for switch-away trials only. This suggests that, with a long cue–stimulus interval, the new task-set was implemented before stimulus onset for switch-to trials, whereas on switch-away trials this process was delayed until after stimulus onset leading to increased switch cost. These results demonstrate dissociable effects of switching away from the current task-set and switching to the upcoming task-set and support the interpretation that the differential positivity observed for switch-to trials reflects processes associated with anticipatory task-set reconfiguration. © 2006 Elsevier B.V. All rights reserved. Keywords: ERP Task-switching Task-set reconfiguration Attentional control 1. Introduction As contextual requirements change, previously relevant information becomes irrelevant and cognitive processes need to be reconfigured to deal with changing contextual demands, suppressing previously relevant task-sets and implementing the currently active task-set (i.e., the procedures required for task completion, Monsell, 1996). Task-switching paradigms have been used to investigate this process of task-set reconfiguration (for review see Monsell, 2003; Logan, 2003). In a typical task-switching experiment, participants alternate between two simple tasks in a fixed sequence or using task cues. Reaction time (RT) is longer on trials that require a switch, as compared to a repeat, in task. This switch cost has been attributed to passive processes associated with the relative level of activation of relevant and irrelevant task-sets and stimulus-response interference (Allport et al., 1994; Allport and Wylie, 2000), as well as the engagement of active task-set reconfiguration processes on switch trials (Rogers and Monsell, 1995). 1.1. Processes underlying RT switch cost It has been consistently found that increasing the time available to prepare in anticipation of a switch in task reduces RT switch cost (e.g., Goschke, 2000; Meiran, 1996; Rogers and Monsell, 1995). This finding has been interpreted as evidence for a cognitive control process of task-set reconfiguration that BRAIN RESEARCH 1121 (2006) 160 – 176 ⁎ Corresponding author. School of Psychology, University of Newcastle, Callaghan, NSW 2308, Australia. Fax: +61 2 49216980. E-mail address: [email protected] (R. Nicholson). 0006-8993/$ – see front matter © 2006 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2006.08.101 available at www.sciencedirect.com www.elsevier.com/locate/brainres
-
Upload
newcastle-au -
Category
Documents
-
view
0 -
download
0
Transcript of Components of task-set reconfiguration: Differential effects of ‘ switch-to’ and ‘...

B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res
Research Report
Components of task-set reconfiguration: Differential effects of‘switch-to’ and ‘switch-away’ cues
Rebecca Nicholsona,b,⁎, Frini Karayanidisa,b,c, Anna Davies, Patricia T. Michiea,b,c
aFunctional Neuroimaging Lab, School of Psychology, University of Newcastle, AustraliabNeuroscience Institute of Schizophrenia and Allied Disorders (NISAD), AustraliacHunter Medical Research Institute, Newcastle, Australia
A R T I C L E I N F O
⁎ Corresponding author. School of PsychologyE-mail address: Rebecca.Nicholson@newc
0006-8993/$ – see front matter © 2006 Elsevidoi:10.1016/j.brainres.2006.08.101
A B S T R A C T
Article history:Accepted 26 August 2006Available online 29 September 2006
Preparation for a switch in task was manipulated using two types of switch cues: ‘switch-away’ from the previous task-set and ‘switch-to’ a different task-set. Increasing cue–stimulusinterval resulted in a reduction in reaction time switch cost for switch-to trials only. Cue-locked difference waveforms for both switch-to and switch-away trials showed a large, broaddifferential positivity, relative to repeat waveforms. However, the later part of thedifferential positivity was significantly reduced on switch-away trials. A differentialpositivity then emerged after stimulus onset for switch-away trials only. This suggeststhat, with a long cue–stimulus interval, the new task-set was implemented before stimulusonset for switch-to trials, whereas on switch-away trials this process was delayed until afterstimulus onset leading to increased switch cost. These results demonstrate dissociableeffects of switching away from the current task-set and switching to the upcoming task-setand support the interpretation that the differential positivity observed for switch-to trialsreflects processes associated with anticipatory task-set reconfiguration.
© 2006 Elsevier B.V. All rights reserved.
Keywords:ERPTask-switchingTask-set reconfigurationAttentional control
1. Introduction
As contextual requirements change, previously relevantinformation becomes irrelevant and cognitive processes needto be reconfigured to deal with changing contextual demands,suppressing previously relevant task-sets and implementingthe currently active task-set (i.e., the procedures required fortask completion, Monsell, 1996). Task-switching paradigmshave been used to investigate this process of task-setreconfiguration (for review see Monsell, 2003; Logan, 2003). Ina typical task-switching experiment, participants alternatebetween two simple tasks in a fixed sequence or using taskcues. Reaction time (RT) is longer on trials that require a switch,as compared to a repeat, in task. This switch cost has been
, University of Newcastleastle.edu.au (R. Nicholson
er B.V. All rights reserved
attributed to passive processes associated with the relativelevel of activation of relevant and irrelevant task-sets andstimulus-response interference (Allport et al., 1994;Allport andWylie, 2000), as well as the engagement of active task-setreconfigurationprocesseson switch trials (Rogers andMonsell,1995).
1.1. Processes underlying RT switch cost
It has been consistently found that increasing the timeavailable to prepare in anticipation of a switch in task reducesRT switch cost (e.g., Goschke, 2000; Meiran, 1996; Rogers andMonsell, 1995). This finding has been interpreted as evidencefor a cognitive control process of task-set reconfiguration that
, Callaghan, NSW 2308, Australia. Fax: +61 2 49216980.).
.

161B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
can be activated prior to the onset of an imperative stimulus,leading to a reduction in the RT switch cost (Rogers andMonsell). According to De Jong (2000), task-set reconfigurationis an all-or-none process that can be activated before or afterstimulus onset depending on task and subject parameters (seeSection 1.2 below).Whilemore recent studies have questionedthe notion of an ‘all-or-none’ task-set reconfiguration process(e.g., Brown et al., 2006; Lien et al., 2005), failures to engage inanticipatory task-set reconfiguration on some proportion oftrials are now recognized to at least partially account for theresidual switch cost that remains even at long preparationintervals (Rogers and Monsell).
In contrast to task-set reconfiguration accounts of RTswitch cost, Allport and colleagues (Allport et al., 1994; Allportand Wylie, 2000; Wylie and Allport, 2000; Waszak et al., 2003)argue that the switch cost results from differences in task-setinertia or performance readiness between switch and repeattrials. Performance readiness is affected by factors such aspositive and negative priming from previous task practice,cued activation of relevant/irrelevant stimulus, response and/or task-set attributes and inhibition of incongruent attributes.Meiran and colleagues (Meiran, 1996, 2000; Meiran et al., 2000;see also Goschke, 2000) offered a conciliatory position showingevidence for both active task-set reconfiguration and passivetask-set inertia effects on RT switch cost. Independentmanipulation of the opportunity for anticipatory task-setreconfiguration in the cue–stimulus interval (CSI) and thetime available for passive dissipation of activation across theoverall response-stimulus interval (RSI) both resulted insignificant reductions in RT switch cost (Meiran, 1996; Meiranet al., 2000; Nicholson et al., 2005).
1.2. Components of task-set reconfiguration
Although there is considerable evidence regarding the exis-tence of a process of anticipatory task-set reconfiguration (butsee Koch, 2005; Ruthruff et al., 2001; Sohn and Carlson, 2000),relatively little is known about what this process actuallyentails. For task-set reconfiguration to be activated prior tostimulus onset, it is necessary for participants to haveforeknowledge that the next trial will require a switch intask, as is the case with alternating runs paradigms that use apredictable task sequence (e.g., task AABB) and with taskcueing paradigms that validly cue the relevant task prior tostimulus onset. Foreknowledge is only useful if the length ofthe preparation interval (e.g., the RSI in alternating runsparadigms or the CSI in cued task paradigms) is sufficient toallow activation of anticipatory task-set reconfiguration(Rogers and Monsell, 1995) and the participant activelyengages this process prior to stimulus onset (De Jong, 2000).If these conditions are not met, task-set reconfiguration isinitiated and/or completed after stimulus onset, resulting inlonger switch trial RT and hence increased RT switch cost.
Anticipatory task-set reconfiguration can also be affectedby parameters that promote or impede active engagement inthese processes. Goschke (2000) found that interruptinganticipatory task-set reconfiguration by requiring participantsto recite irrelevant verbal material during a long RSI (1200 ms)resulted in RT switch cost equivalent to the no preparation orshort RSI (150 ms) condition. In contrast, when participants
verbalized the task to be completed on the next trial, therewasa significant reduction in RT switch cost compared to either ofthe above conditions and equivalent to that obtained with along RSI when no verbalization is required. This suggests thatretrieval and application of the new task-set from long termintoworkingmemory are important parts of anticipatory task-set reconfiguration (Mayr and Kliegl, 2000).
Changing switch trial salience by reducing the proportionof switch relative to repeat trials has also been shown toincrease the engagement of active task-set reconfigurationprocesses (Monsell and Mizon, 2006). Using cues that mani-pulate subjective expectancy of equiprobable switch andrepeat trials, Dreisbach et al. (2002) found equivalent effectsof subjective expectancy on both switch and repeat trials,suggesting that task preparation may occur in anticipation ofboth switch and repeat trials. Furthermore, task preparationdid not interact with task foreknowledge (see also Sohn andCarlson, 2000), suggesting that these manipulations affectindependent processes and supporting a preparation readi-ness account of switch cost. Alternatively, Monsell and Mizon(2006) argue that these effects may be at least partly attributedto cue complexity as complex cues may inadvertentlyintroduce an additional task, resulting in task-set reconfigu-ration for both switch and repeat trials.
Inhibitory processes may also affect task-set reconfigura-tion. Mayr and Keele (2000) found that switching back to arecently abandoned task-set resulted in larger RT switch costas compared to switching to a third task—a phenomenon theyrefer to as backward inhibition. Specifically, RT was larger onthe third trial of a CBC task sequence as compared to an ABCsequence (see also Arbuthnott and Frank, 2000; Koch et al.,2004). This suggests that the previously relevant, but nowirrelevant, task-set (task C) was inhibited in the former tasksequence, thereby resulting in longer RT when this task-setwas reactivated within a short period. Using a go/no-goparadigm, Schuch and Koch (2003) found that backwardinhibition did not occur following no-go trials that requiredtask-set preparation but no response execution, suggestingthat selection or execution of the response triggers theinhibitory processes. Mayr and Keele (2000) found that theamount of backward inhibition was not reduced withincreasing CSI, suggesting that the inhibition process is notaffected by the amount of time available for anticipatorytask-set reconfiguration. In contrast, backward inhibitionwas reduced at longer response–cue intervals (RCI) thatfacilitated greater passive dissipation of task-set interference(see also Koch et al., 2004). Furthermore, Gade, 2005 showedthat the opportunity for passive dissipation of interferenceacross the interval between trial n and trial n−2 had agreater effect than the interval between n and n−1 (e.g., intask sequence ABA, the amount of backward inhibition wasaffected more by the interval between the two task A trialsthan between task B and task A trials). Dreisbach et al. (2002)reported that ‘semi-specific’ cues (i.e., cues that signal animpending switch trial without identifying which specifictask to prepare) resulted in larger RT switch cost than‘specific’ cues that indicated which task to switch to. Unlikespecific cues, semi-specific cues did not produce subjectiveexpectancy effects, suggesting that knowledge that the taskwould change without specification of which task would be

1 The term D-Pos is used as a convenience label to refer to thedifferential switch minus repeat positivity without necessarilyimplying that the label represents a specific or new ERPcomponent that is not evident in the original waveforms.
162 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
performed did not result in any differential response benefit.Hubner et al. (2003) showed that backward inhibition effectswere comparable for an uncued switch in task and a cuedswitch in task with semi-specific cues.
Taken together, these findings suggest that task-set recon-figuration may involve not only activation of the currentlyactive task-set but also inhibition of the previously active task-set. Furthermore, it appears as if the inhibition process cannotbe initiated independently of activation of the currently activetask-set. Mayr and Keele (2000) argue that inhibition mayreflect a low-level control process, such as lateral inhibition ofcompeting action schemas. Dreisbach et al. (2002) suggest thatactive engagement of the cued task-set triggers automaticinhibition of the previously relevant task-set when theresponse is executed. That is, they argue that inhibition ofthe irrelevant task-set is not under endogenous control andcannot occur independently of activation of the relevant task-set (see also Hubner et al., 2003).
Overall, there are still considerable gaps in our knowledgeregarding the number and type of cognitive processes thatunderlie anticipatory task-set reconfiguration. Although thereis evidence of a role for an inhibitory process, its timing anddegree of dependence on other activation processes remainunclear. Furthermore, the precise set of operations thatconstitute activation of the relevant task-set in anticipationof a switch in task remains to be defined.
1.3. Task-switching and event-related brain potentials(ERPs)
Event-related brain potentials (ERPs) in task-switchingexperiments can help elucidate the cognitive processesthat lead up to behavioral differences between switch andrepeat trials in general, as well as those processes morespecifically involved in task-set reconfiguration. Most ERPtask-switching studies have focused on differences be-tween ERPs to switch and repeat stimuli occurring afterstimulus onset (e.g., Barcelo et al., 2000, 2002; Gehring etal., 2003; Hsieh and Yu, 2003; Hsieh and Liu, 2005; Poulsenet al., 2005; Swainson et al., 2003). ERP differences betweenswitch and repeat trials occurring after stimulus onset mayreflect processing differences occurring as a result ofdifferential levels of proactive interference for switchcompared to repeat stimuli, differential level of activationof the relevant task-set at stimulus onset, as well asdifferential stimulus-response interference elicited by thestimulus itself. Therefore, although anticipatory task-setreconfiguration may indirectly affect stimulus-locked ERPs(e.g., Barcelo et al., 2000), it is difficult to isolate this effectfrom that of other passive interference or stimulus-elicitedprocesses.
It is possible to isolate ERP effects associated withanticipatory task-set reconfiguration by examining either theinterval preceding stimulus onset in an alternating runsparadigm or the interval following cue onset in a cued task-switching paradigm. Using Rogers and Monsell's (1995)predictable task-switching paradigm, Karayanidis et al.(2003) identified an increased positivity for switch as com-pared to repeat trials in the interval between the response tothe previous trial and the onset of the next trial (i.e., during the
RSI). This differential positivity for switch as compared torepeat waveforms (labeled D-Pos1) was largest parietally andpeaked around 400 ms after the previous response. At shortpreparation intervals (150 or 300 ms), D-Pos began prior to butdid not peak until after stimulus onset, whereas with longpreparation intervals (600 and 1200 ms), D-Pos peaked andreturned to baseline before the stimulus was presented. Asimilar switch-related positivity has been reported in otherstudies. Wylie et al. (2003) found that trials preceding apredictable switch trial were associated with a larger latepositivity over posterior scalp electrodes compared to trialspreceding a repeat trial. Other studies found that ERP wave-forms time-locked to cue presentation show a larger positivityfor switch compared to repeat cues over parietal sites around500 ms after cue onset (Miniussi et al., 2005; Rushworth et al.,2002, 2005). Using a cued task-switching paradigm, Nicholsonet al. (2005) found that the morphology of the parietal switchpositivity was modified by manipulation of the CSI (150 to600 ms) but not the RSI (750 to 1200 ms), indicating that it isaffected by opportunity to engage in active task-set reconfig-uration but not by the passive passage of time between theprevious and the current trial.
In summary, anticipatory task-set reconfiguration is asso-ciated with increased positivity in anticipation of switchrelative to repeat trials, particularly over parietal electrodes.At longer preparation intervals, this differential activity forswitch trials can be completed prior to stimulus presentationand is associated with reduced RT switch cost. With shortpreparation intervals, which provide little or no opportunityfor anticipatory task-set reconfiguration, these processesoccur after stimulus onset and are associated with larger RTswitch cost.
1.4. The current study
The current study investigates one component process oftask-set reconfiguration, the activation of the currentlyrelevant task-set. Previous ERP studies have identified adifferential positivity for switch compared to repeat trials inthe interval preceding stimulus presentation that is believedto reflect anticipatory task-set reconfiguration processes(Karayanidis et al., 2003; Nicholson et al., 2005). The currentstudy examined whether this anticipatory positivity is specif-ically associated with activation of the relevant task-set or isaffected by other processes that may occur during preparationfor a switch in task.
A cued-trials task-switching paradigm was used. Partici-pants randomly alternated between three tasks defined forthe same stimulus set. Three types of cues were used tosignal the requirements of the next trial. The first cue typesignaled task repetition, the second cue type signaled a taskswitch and specified which task to switch to (switch-to orspecific cues), whereas the third cue type signaled a taskswitch but did not specify which of the two alternate taskswould be active (switch-away or semi-specific cues). With the

163B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
latter cues, participants knew that they would not berepeating the same task as on the previous trial but hadno information about which task-set would be relevant andhad to await stimulus onset before the new task-set could beactivated. Therefore, the use of switch-to and switch-awaycues was designed to isolate processes associated withactivation of the relevant task-set.
Switch-to trials were identical to the switch trials defined inprevious ERP and behavioral studies. Therefore, switch-totrials were expected to produce a large RT switch cost thatreduced with increasing CSI and with increasing RSI, as wellas a large switch-related positivity time-locked to cue onsetand peaking before stimulus onset in the long CSI condition(Karayanidis et al., 2003; Nicholson et al., 2005). Semi-specificor switch-away cues index that the current task-set will not beactive upon stimulus onset, but do not specify which of theremaining two task-sets will be active. Since for switch-awaycues, activation of the relevant task-set could not becompleted until after stimulus onset regardless of prepara-tion interval, it was expected that switch-away cues wouldhave larger RT switch cost than switch-to cues and thatswitch-away RT cost would not reduce with increasing CSI.However, as passive dissipation of task-set interferenceshould not be affected by the information provided by thetwo switch cue types, increasing RSI for constant CSI wasexpected to reduce RT switch cost equally for switch-to andswitch-away trials.
ERP waveforms for switch-to and switch-away trials werecompared only at the long CSI (1000 ms) that affords theopportunity for activation of anticipatory task-set reconfigu-ration and provides temporal dissociation between cue andstimulus related processes. If the cue-locked differentialswitch positivity that peaks before stimulus onset with longCSIs for switch-to trials indexes activation of the relevanttask-set in anticipation of a cued switch in task-set, thensince this process cannot be activated until after stimulusonset for switch-away trials, there should be no evidence of acue-locked differential switch positivity for switch-away trials.Instead, for switch-away trials, this cue-locked differentialswitch positivity should emerge after stimulus onset just as itdoes when there is no advance cueing or when the CSI is veryshort (Nicholson et al., 2005). If a differential switch positivityis still evident within the CSI for switch-away trials, thenclearly it cannot indicate activation of the relevant task-set,
Table 1 – Mean reaction time (RT) and percentage error rate by
RSI-1200:CSI-200 RSI-
Repeat Switch-to Switch-away Repeat Sw
Mean RT (ms)Letter task 773 (14) 949 (9) 1008 (12) 703 (13) 84Digit task 776 (14) 1021 (14) 1072 (14) 721 (13) 90Average 775 (12) 986 (9) 1040 (10) 713 (11) 87
Error %Letter task 2.1 (0.6) 3.9 (0.4) 4.1 (0.5) 1.4 (0.4) 3Digit task 2.8 (0.4) 5.2 (0.8) 4.6 (0.4) 2.5 (0.5) 3Average 2.5 (0.4) 4.6 (0.5) 4.4 (0.2) 1.9 (0.4) 3
Standard error in parentheses.
but presumably some other component of task-set reconfig-uration that can be triggered by both switch-to and switch-away cues.
2. Results
2.1. Behavioral data
Mean RT and error rate for each timing condition, trial typeand task are shown in Table 1. The main effects of condition,trial type and task (F(2,70)=36.2, p<0.001; F(1,48)=229.3,p<0.001; F(1,35)=22.2, p<0.001, respectively) and the condi-tion×trial type and trial type×task interactions (F(4,140)=6.9,p<0.001; F(2,58)=8.5, p<0.005) were significant. Overall, RTwas longer for the digit task compared to letter task, and thiseffect was larger for both types of switch trials compared torepeat trials (Table 1). RT was also larger for the short CSI(RSI-1200:CSI-200) compared to the two long CSI conditions.The reduction in RT with increasing CSI was significant forall trial types, indicating a general preparation effect. Thiseffect did not differ between repeat and switch-away trials(F<1) but was significantly larger for switch-to as compared torepeat trials (F(2,70)=9.29, p<0.001).
These effects were further examined by calculating RTswitch cost for switch-to and switch-away trials (Fig. 1).Switch cost was larger for the digit than for the letter task(F(1,35)=11.3, p<.005), however task did not interact withswitch type or condition. As shown in Fig. 1, RT switch costwas larger for switch-away than for switch-to trials across allthree conditions (F(1,35)=139.8, p<0.001). Paired compari-sons between conditions showed that, for switch-to cues, RTswitch cost did not differ significantly between short(1200 ms) and long (1600 ms) RSI (162 ms and 159 ms,respectively), but it was significantly greater in the short(200) compared to the long (1000) CSI (210 ms and 162 ms,respectively, F(1,35)=13.9, p<0.005). For switch-away cues,RT switch cost did not differ significantly between short andlong RSI (269 ms and 252 ms, respectively) nor betweenshort and long CSI (265 ms and 269 ms, respectively).
The overall error rate was very low, ranging between 1.3and 4.6% (Table 1). Transformed error scores showed asignificant main effect of condition (F(2,70)=6.5, p<.005)reflecting slightly higher error rates in the short CSI (3.8%)
condition, trial type and task (average over task also shown)
1200:CSI-1000 RSI-1600:CSI-1000
itch-to Switch-away Repeat Switch-to Switch-away
3 (10) 955 (12) 701 (13) 840 (13) 942 (11)5 (16) 1007 (14) 740 (13) 918 (15) 1004 (15)5 (10) 982 (9) 721 (11) 880 (11) 973 (9)
.4 (0.3) 2.9 (0.3) 1.3 (0.5) 4.3 (0.6) 3.5 (0.4)
.5 (0.6) 4.1 (0.4) 1.9 (0.4) 3.6 (0.4) 4.4 (0.6)
.4 (0.3) 3.5 (0.3) 1.6 (0.3) 3.9 (0.3) 4.0 (0.4)

Fig. 1 – Switch-away switch cost (switch-awayminus repeat)and switch-to switch cost (switch-to minus repeat) for eachcondition averaged across task. Standard error bars areshown. Top: Mean reaction time (RT) switch cost. Bottom: %error switch cost.
2 Difference waveforms for the four midline sites (averaged overthe two long CSI conditions) are shown in Fig. 4B (left).
164 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
compared to the long CSI conditions (3.0–3.2%). Both switchtrial types produced more errors (3.9% for switch-away and4% for switch-to) than repeat trials (2%, F(2,68) =23.5,p<0.001). There were no further significant effects in theerror data.
2.2. ERP data
Behavioral data indicated that, although there was anoverall effect of task on RT, task did not interact with RTswitch cost type or condition. Therefore, in order tomaximize signal to noise ratio, all ERP data were averagedover task (letter/digit).
2.2.1. Cue-locked effects of switching to a taskCue-locked waveforms averaged separately for repeat andswitch-to trials for each timing condition at midline sites areshown in Fig. 2A (top). Long CSI conditions (middle and right)show a broad centroparietally maximal positivity beginningapproximately 200 ms after cue onset at most midline sites.This positivity returned to baseline by 450 ms at frontal andcentral sites, but persisted to approximately 700ms at parietal
and occipital sites. Differences between repeat and switch-tocue-locked ERPs emerged as early as 100 ms occipitally andspread beyond 800 ms parietally. The variations in ERPmorphology between these two long CSI conditions are likelyto reflect differential resolution of post-response processes.Specifically, although CSI was constant at 1000 ms for bothconditions, the response–cue interval was only 200ms for RSI-1200 but 600 ms for RSI-1600ms. In the former condition, bothswitch and repeat trials may have included an overlap of post-response processes that had resolved in the longer RSIcondition.
Cue-locked waveforms for the short CSI condition (left)differed more notably from the two long CSI conditions(middle, right) due to the partial temporal overlap of cue-locked and stimulus-locked ERPs. In this short CSI condition,frontal and central sites showed a series of positive andnegative deflections that do not differentiate between repeatand switch-to trials as clearly as the long CSI conditions andare likely to reflect cue and stimulus processing. Central andparietal electrodes do, however, show a broad positivedeflection extending from 150 ms to approximately1000 ms after cue onset reflecting the temporal overlap ofa post-cue positivity, similar to that seen in the long C–Sconditions, and a late positive component (LPC) to stimulusonset.
2.2.1.1. Cue-locked switch-to difference waveforms. Fig. 2A(bottom) depicts switch-to cue-locked difference waveformsfor the three timing conditions at Pz2 where the effectswere maximal. Positive deviations from baseline indicateareas where the switch-to waveform was more positiverelative to the repeat waveform (see Table 2). Switch-todifference waveforms showed a broad positive deflectionsimilar to that previously reported by Karayanidis et al.(2003) and Nicholson et al. (2005). This differential positivity(D-Pos) emerged 150–190 ms after cue onset for all threeconditions. In both long CSI conditions, D-Pos spreadbeyond 800 ms covering the entire interval until stimulusonset, whereas in the short CSI condition, it resolvedaround 500 ms post-cue.
2.2.2. Stimulus-locked effects of switching to a taskERP waveforms time-locked to stimulus onset for repeat andswitch-to trials are shown at midline sites in Fig. 2B (top).Stimulus-locked waveforms tended to show a pattern ofearly occipital obligatory ERPs (P1, N1 and P2) followed by abroad LPC that spread beyond 700 ms and was maximalparietally. Frontally, a large P2-type component emergedaround 200 ms but there was little evidence of furtheractivity. These effects are most clearly evident in the longCSI conditions that had no temporal overlap with the cue-locked period. In comparison, the short CSI condition (RSI-1200:CSI-200) had a sharp shifting pre-stimulus baseline,especially at frontal to parietal sites, reflecting the partialtemporal overlap between cue-locked and stimulus-lockedERPs. Overall, the stimulus-locked ERP morphology was verysimilar for repeat and switch-to waveforms; however,

Fig. 2 – Top: (A) Cue-locked and (B) stimulus-locked ERP waveforms for switch-to and repeat trials compared at four midline sites for each timing condition. Negative isplotted up. In the cue-locked waveforms, the solid vertical line indicates the onset of the cue and the dotted vertical line indicates the onset of the stimulus. In thestimulus-locked waveforms, the solid vertical line indicates the onset of the stimulus. Bottom: (A) Cue-locked and B) stimulus-locked ERP switch-to difference waveforms(switch-to waveform minus repeat waveform) at Pz. Black bars indicate regions of significant deviation between the switch-to and repeat waveform. See Table 2 forexact values at the four midline sites.
165BR
AIN
RESEA
RC
H1121
(2006)
160–176
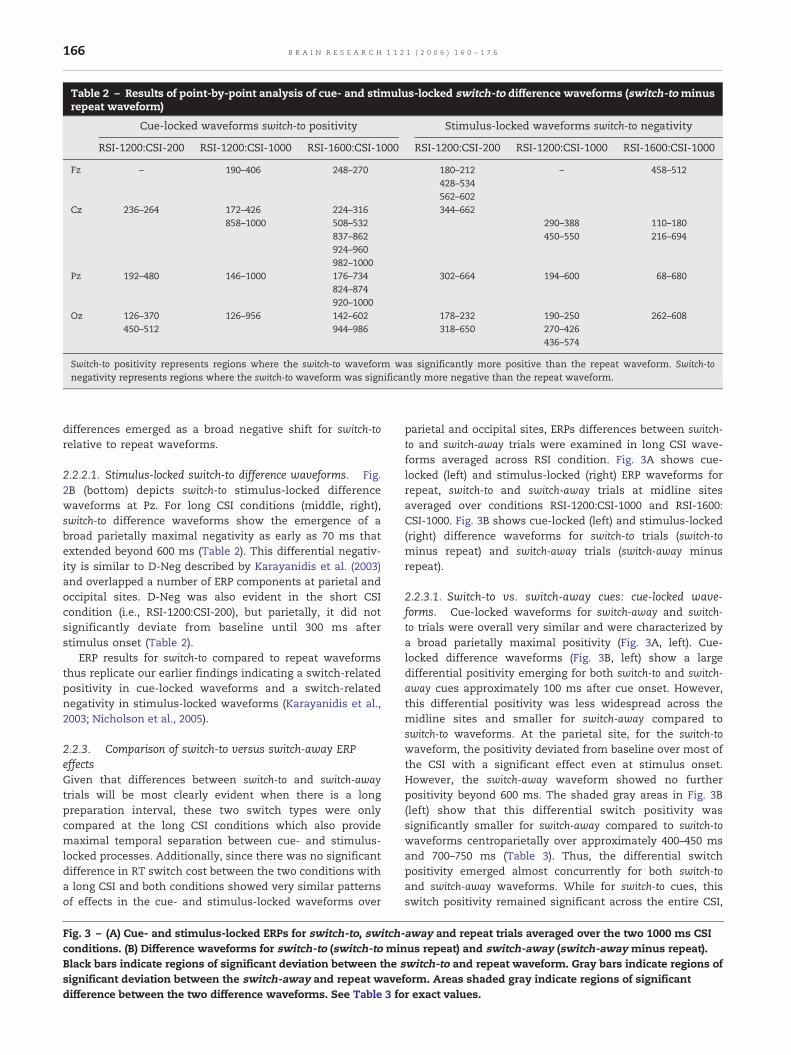
Table 2 – Results of point-by-point analysis of cue- and stimulus-locked switch-to difference waveforms (switch-to minusrepeat waveform)
Cue-locked waveforms switch-to positivity Stimulus-locked waveforms switch-to negativity
RSI-1200:CSI-200 RSI-1200:CSI-1000 RSI-1600:CSI-1000 RSI-1200:CSI-200 RSI-1200:CSI-1000 RSI-1600:CSI-1000
Fz – 190–406 248–270 180–212 – 458–512428–534562–602
Cz 236–264 172–426 224–316 344–662858–1000 508–532 290–388 110–180
837–862 450–550 216–694924–960982–1000
Pz 192–480 146–1000 176–734 302–664 194–600 68–680824–874920–1000
Oz 126–370 126–956 142–602 178–232 190–250 262–608450–512 944–986 318–650 270–426
436–574
Switch-to positivity represents regions where the switch-to waveform was significantly more positive than the repeat waveform. Switch-tonegativity represents regions where the switch-to waveform was significantly more negative than the repeat waveform.
166 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
differences emerged as a broad negative shift for switch-torelative to repeat waveforms.
2.2.2.1. Stimulus-locked switch-to difference waveforms. Fig.2B (bottom) depicts switch-to stimulus-locked differencewaveforms at Pz. For long CSI conditions (middle, right),switch-to difference waveforms show the emergence of abroad parietally maximal negativity as early as 70 ms thatextended beyond 600 ms (Table 2). This differential negativ-ity is similar to D-Neg described by Karayanidis et al. (2003)and overlapped a number of ERP components at parietal andoccipital sites. D-Neg was also evident in the short CSIcondition (i.e., RSI-1200:CSI-200), but parietally, it did notsignificantly deviate from baseline until 300 ms afterstimulus onset (Table 2).
ERP results for switch-to compared to repeat waveformsthus replicate our earlier findings indicating a switch-relatedpositivity in cue-locked waveforms and a switch-relatednegativity in stimulus-locked waveforms (Karayanidis et al.,2003; Nicholson et al., 2005).
2.2.3. Comparison of switch-to versus switch-away ERPeffectsGiven that differences between switch-to and switch-awaytrials will be most clearly evident when there is a longpreparation interval, these two switch types were onlycompared at the long CSI conditions which also providemaximal temporal separation between cue- and stimulus-locked processes. Additionally, since there was no significantdifference in RT switch cost between the two conditions witha long CSI and both conditions showed very similar patternsof effects in the cue- and stimulus-locked waveforms over
Fig. 3 – (A) Cue- and stimulus-locked ERPs for switch-to, switch-conditions. (B) Difference waveforms for switch-to (switch-to minBlack bars indicate regions of significant deviation between the ssignificant deviation between the switch-away and repeat wavedifference between the two difference waveforms. See Table 3 fo
parietal and occipital sites, ERPs differences between switch-to and switch-away trials were examined in long CSI wave-forms averaged across RSI condition. Fig. 3A shows cue-locked (left) and stimulus-locked (right) ERP waveforms forrepeat, switch-to and switch-away trials at midline sitesaveraged over conditions RSI-1200:CSI-1000 and RSI-1600:CSI-1000. Fig. 3B shows cue-locked (left) and stimulus-locked(right) difference waveforms for switch-to trials (switch-tominus repeat) and switch-away trials (switch-away minusrepeat).
2.2.3.1. Switch-to vs. switch-away cues: cue-locked wave-forms. Cue-locked waveforms for switch-away and switch-to trials were overall very similar and were characterized bya broad parietally maximal positivity (Fig. 3A, left). Cue-locked difference waveforms (Fig. 3B, left) show a largedifferential positivity emerging for both switch-to and switch-away cues approximately 100 ms after cue onset. However,this differential positivity was less widespread across themidline sites and smaller for switch-away compared toswitch-to waveforms. At the parietal site, for the switch-towaveform, the positivity deviated from baseline over most ofthe CSI with a significant effect even at stimulus onset.However, the switch-away waveform showed no furtherpositivity beyond 600 ms. The shaded gray areas in Fig. 3B(left) show that this differential switch positivity wassignificantly smaller for switch-away compared to switch-towaveforms centroparietally over approximately 400–450 msand 700–750 ms (Table 3). Thus, the differential switchpositivity emerged almost concurrently for both switch-toand switch-away waveforms. While for switch-to cues, thisswitch positivity remained significant across the entire CSI,
away and repeat trials averaged over the two 1000 ms CSIus repeat) and switch-away (switch-away minus repeat).witch-to and repeat waveform. Gray bars indicate regions ofform. Areas shaded gray indicate regions of significantr exact values.

167B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6

Table 3 – Results of point-by-point analysis of cue- and stimulus-locked difference waveforms averaged over the twoconditions with a 1000 ms CSI
Cue-locked difference waveforms Stimulus-locked difference waveforms
Switch-awaypositivity
Switch-topositivity
To/Awaydifference
Switch-awaypositivity
Switch-awaynegativity
Switch-tonegativity
To/Awaydifference
Fz 202–264 226–388 – 172–332 392–556 – 222–332432–500
Cz 150–278 194–420 392–424 172–276 370–654 232–640 162–336324–374576–602
Pz 140–400 162–766 394–442 164–254 374–556 188–634 156–354470–594 810–1000 690–742
Oz 110–408 134–666 – 160–264 414–488 266–548 160–278470–510 684–764
844–884938–978
Switch-to and switch-away positivity represents regions where the switch-to waveform and the switch-away waveform were significantly morepositive than the repeat waveform, respectively. Switch-to and switch-away negativity represents regions where the switch-to waveform and theswitch-away waveform were significantly more negative than the repeat waveform, respectively. The To/Away difference values representregions where there was a significant difference between the two difference waveforms.
168 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
for switch-away cues, it reduced in amplitude around 400 msand returned to baseline before stimulus onset.
2.2.3.2. Switch-to vs. switch-away cues: stimulus-locked wave-forms. The overall morphology of stimulus-locked ERPwaveforms was similar for repeat, switch-to and switch-awaytrials (Fig. 3A, right). However, switch-away and switch-towaveforms differed in their pattern of deviation from therepeat waveform. As shown earlier, switch-to trials showed anearly and prolonged centroparietally maximal negative shiftcompared to repeat waveforms. In contrast, switch-awaywaveforms show an early positive deviation emerging around150 ms after stimulus onset. This can be most clearly seen inthe difference waveforms (Fig. 3B, right). Switch-away trialsshowed a large, sharp positive deviation from repeat trialsapproximately 150–300 ms after stimulus onset (Table 3). Thispositivity was spread across all midline sites, overlapping P2anteriorly and P2 and early P3 posteriorly. This early positivityin the switch-away difference waveform was quickly suc-ceeded by a differential switch negativity that differed fromthe switch-to negativity in that it began much later (360 ms)and spread more broadly across the scalp. However, like theswitch-to negativity, the negativity for switch-away trials alsoreturned to baseline around 700 ms (Table 3). Direct point-by-point comparison of switch-to and switch-away differencewaveforms resulted in significant differences over approxi-mately 160–350 ms spanning across all midline sites (Table 3),but no further differences across the analysis epoch. Thisconfirmed that switch-away trials showed a significant earlydifferential positivity which was replaced by a differentialswitch negativity that began later than but resolved in linewith the switch-to negativity.
2.2.4. Summary of ERP findingsSwitch-to cues showed the expected pattern of a parietallymaximal broad differential cue-locked positivity emergingaround 150 ms and extending across the entire CSI and acentroparietal differential negativity emerging shortly after
stimulus onset and extending beyond 600 ms. In cue-lockedwaveforms, a differential positivity also emerged around150 ms for switch-away cues, but had a smaller amplitudeand shorter duration than for switch-to cues. In stimulus-locked waveforms, switch-away trials showed an early differ-ential positivity relative to repeat trials extending over 150–300 ms that was followed by a differential negativity thatemerged much later and was more widespread than forswitch-to trials.
3. Discussion
The present study used behavioral and ERP measures todissociate components of anticipatory task-set reconfigura-tion. Specifically, it was investigated whether the differentialpositivity observed during preparation for a switch in task(Karayanidis et al., 2003; Nicholson et al., 2005) can be shownto be associated with activation of the currently relevant task-set. The point at which the new task-set could be activatedwas manipulated by varying the amount of informationprovided by the task cue. Cues signaled either that the presenttask would be repeated (task repeat cue), that the next trialwould require a switch in task and specified which task was tobe performed (switch-to cue), or that the next trial wouldrequire a switch in task but did not specify which task was tobe performed (switch-away cue).
3.1. Behavioral effects of switching to versus switchingaway from a task-set
The switch-to cues used here were identical to the switch cuesused in most previous task-switching studies in that theyspecified both the need to shift away from the current task-setand identified which task-set would be relevant on theupcoming trial. RT on switch-to trials was much slower ascompared to repeat trials and RT switch cost ranged from159 to211 ms, which is larger than that found in our earlier studies

169B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
(Nicholson et al., 2005). A number of factors may havecontributed to this inflated switch cost. Firstly, this studyincluded only bivalent incongruent stimuli that have beenshown to produce slower RT and greater switch cost (Rogers andMonsell, 1995; Woodward et al., 2003). Secondly, the taskinvolved a larger switch trial probability (two-thirds of all trials)compared to our previous studies (Nicholson et al., 2005).Monsell andMizon (2006) suggest that increasing theproportionof switch trials reduces participants' tendency to engage inanticipatory task-set reconfiguration, thus potentially leading toincreased mean RT switch cost (De Jong, 2000). Thirdly, thisstudy involved switching between three instead of two task-sets. This may have resulted in larger memory requirementsand more stimulus-response mapping interference.
As found in many previous studies (Meiran, 1996; Nichol-son et al., 2005; Rogers and Monsell, 1995), increasing thepreparation interval resulted in a reduction in switch-to RT cost(i.e., 48 ms decline as CSI increased from 200 to 1000 ms).However, contrary to previous studies (Meiran et al., 2000;Nicholson et al., 2005), increasing opportunity for passivedissipation of task-set interference (i.e., increasing RSI from1200 to 1600 ms) had no further effect on switch-to RT cost.This may be attributable to the specific RSI values used in thecurrent study. Although Nicholson et al. found that switchcost significantly declined as the RSI increased from 750 to1200 ms, Meiran et al. found no reduction in RT switch cost asthe RSI increased from 1000 to 3000 ms. At the shortest RSIvalue used in the current study (1200 ms), passive dissipationof task-set interferencemay have already plateaued. Thus, thefurther increase to 1600 ms provided no additional benefit.
A large ‘residual’ switch cost (Rogers and Monsell, 1995)remained on switch-to trials, even with a preparation intervalof 1 s (see also Nicholson et al., 2005). This ‘residual’ switchcost may reflect failures to engage in anticipatory task-setreconfiguration on some proportion of switch-to trials, therebyresulting in an overall increase in mean switch-to RT (De Jong,2000). Upon stimulus onset, the spatial location of thestimulus provided a redundant cue as to which task wasrelevant. Therefore, although participants were encouraged touse switch-to cues to prepare in anticipation of stimulus onset,if they failed to do this on some trials, they could still respondsuccessfully by initiating task-set reconfiguration after stim-ulus onset. Nicholson et al. (2006) found that eliminatingredundant task cues and removing the cue prior to stimulusonset encouraged anticipatory task-set reconfiguration andreduced variability in strategy implementation both betweenparticipants and between trials for the same participant.
Overall, RT switch cost was significantly larger for switch-away (∼260 ms) compared to switch-to (∼180 ms) trials. Incontrast to switch-to trials, increasing the preparation intervalfrom 200 to 1000 ms did not have any differential effect onswitch-away as compared to repeat trial RT. This finding iscompatible with the earlier findings by Dreisbach et al. (2002)and Hubner et al. (2003) that information about an impendingswitch trial without specific knowledge about which task toswitch to does not provide any differential behavioraladvantage and supports the suggestion by Dreisbach et al.that irrelevant task-set inhibition cannot occur independentlyof relevant task-set activation. Interestingly, RT cost waslarger for switch-away than switch-to cues even at the short
(200 ms) preparation interval. Nicholson et al. (2005) foundthat RT switch cost was smaller for 150 ms CSI compared to ano cue condition. These findings suggest that active task-setreconfiguration can at least be initiated prior to stimulus onseteven with a very short CSI.
3.2. Electrophysiological effects of switching to versusswitching away from a task-set
Cue-locked ERP waveforms for switch-to trials showed asignificant differential positivity that began approximately100 ms after cue onset, peaked around 350–400 ms and waslargest parietally. At long preparation intervals, this differen-tial switch-related positivity extended across most of the CSIbut resolved prior to stimulus onset. At the short preparationinterval (200 ms), the differential positivity peaked afterstimulus onset and had a shorter duration. For all conditions,switch-to trials showed a large stimulus-locked differentialnegativity. This emerged earlier for long CSI conditions(around 150 ms) than for the short CSI condition (around300 ms) but peaked around 500 ms in all conditions. Thesefindings are consistent with those of previous studies(Karayanidis et al., 2003; Miniussi et al., 2005; Nicholson etal., 2005; Rushworth et al., 2002, 2005) showing differentialprocessing in anticipation of a switch trial and differentialprocessing of switch and repeat stimuli, thereby supportingthe contribution of both active task-set reconfiguration andstimulus-elicited interference processes on RT switch cost.
ERP differences between switch-to and switch-away trialswere examined at the long preparation interval that providedopportunity for anticipatory task-set reconfiguration process-es to be initiated prior to stimulus onset and temporallyseparated cue- and stimulus-locked processes. Switch-awaydifference waveforms showed a cue-locked differential posi-tivity emerging approximately 150 ms after cue onset similarto that for switch-to difference waveforms (Fig. 3B, left).However, while for switch-to trials, this positivity remainedsignificant centroparietally across the entire CSI, for switch-away trials, it reduced in amplitude over 350–450 ms andreturned to baseline by 600 ms. Specific point-by-pointcomparisons between switch-to and switch-away waveformsshowed that the later portion of the positivity was signifi-cantly smaller centroparietally over 400–450 ms and parietallyover 650–750 ms after cue onset for switch-away trials. So, adifferential switch-related positivity emerged within the CSIfor both switch-to and switch-away waveforms but had asmaller amplitude and shorter duration in the latter cuetype. Additionally, switch-away difference waveforms showedthe emergence of differential positivity after stimulus onset(Fig. 3B, right). This post-stimulus differential positivity forswitch-away trials was broadly distributed emerging around150 ms and extending frontally beyond 300 ms. It wassucceeded by a centroparietal differential switch negativitythat peaked around 500 ms after stimulus onset, similar tothat obtained for switch-to trials.
3.3. Implications for models of task-switching
Previous work (e.g., Nicholson et al., 2005) has suggested thatthe differential positivity to specific switch cues (i.e., cues that

3 Although our switch-away cues and Barceló's 3D shift feedbackcues both indicate that the previously active task-set is no longerrelevant, they convey different information about how toproceed. In the WCST, participants select one of the alternativetask-sets, implement it upon stimulus onset and wait for the nextfeedback cue to determine whether it was correct. In the presenttask, the relevant task-set is defined upon stimulus onset and, asdiscussed below, there was no evidence that participants eitherprepared both task-sets or prepared one task-set based onsubjective sequence expectancies. Despite these task differences,in both studies, the late parietal positivity to shift cues behavedquite consistently; Barceló et al. found that P3b amplitudereduced with increasing certainty about which task-set wouldbe active upon stimulus onset (e.g., 3D shift vs. 2D shift feedbackcues), and the present data showed a larger late differentialswitch positivity for switch-to than for switch-away cues.4 We thank Dr. Leuthold for this suggestion.
170 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
both signal an impending switch trial and specify which taskwill be active on the next trial) reflects task-set reconfigura-tion. The finding that, at long preparation intervals, thispositivity is restricted within the CSI suggests that at leastsome aspects of task-set reconfiguration can be completed inanticipation of stimulus onset (see current switch-to data, butalso Karayanidis et al., 2003; Nicholson et al., 2005; Rushworthet al., 2002, 2005). However, the fact that a residual RT switchcost remains at even the longest preparation intervalscombined with the evidence for differential post-stimulusprocessing of switch and repeat trials suggests that thisanticipatory component of task-set reconfiguration cannotfully account for RT switch cost. Stimulus and/or responseinterference, as well as passive dissipation of the previouslyactivated task-set may account for the post-stimulus differ-ential switch negativity and contribute to the RT switch cost.Additionally, at short preparation intervals and in no cueconditions, the switch-related positivity emerges or peaksafter stimulus onset (see current switch-to data for short CSI,but also Karayanidis et al., 2003; Nicholson et al., 2005),suggesting that task-set reconfiguration is a crucial processthat is completed either before or after stimulus onset,depending on task parameters.
Within this framework, the current findings for switch-away trials suggest that the process of task-set reconfigura-tion consists of multiple subcomponents that can betemporally dissociated. One process appears to be reflectedin the emergence of a cue-locked differential positivity forboth switch-to and switch-away trials. This finding indicatesthat forewarning of an impending switch trial initiatesdifferential processing between switch and repeat trials,even in the absence of information about which specific taskwill be active on the subsequent trial. This process cannotrelate to activation of the task-set that will be relevant forthe next stimulus because this information is not yetavailable for switch-away trials. A second process is reflectedin the post-stimulus differential positivity for switch-awaycompared to repeat trials. This is believed to reflect theactive engagement of the relevant task-set, which for switch-away trials is only possible after the stimulus has beenpresented and its position has indicated which task toperform.
Therefore, it is suggested that the late component of thecue-locked differential positivity for switch-to trials and thestimulus-locked differential positivity for switch-away trialsboth reflect activation of the relevant task-set. This inter-pretation is supported by the finding that switch-away trialshave a larger RT switch cost than switch-to trials, even at theshort 200 ms CSI (i.e., even with a CSI of only 200 ms,participants were able to begin preparation for the new task-set following cue presentation on switch-to trials, resulting inreduced RT switch cost compared to switch-away trials). It isalso supported by earlier findings that, at short preparationintervals (e.g., see short CSI condition in Fig. 2A) and in nocue conditions (Nicholson et al., 2005), the differential switchpositivity emerges and/or peaks after stimulus onset.
The functional significance of the early cue-locked positi-vity for both switch-to and switch-away trials is more obscureand a number of possible interpretations will be examined.Given that the only common information provided by switch-
to and switch-away cues is that the following trial will not be arepeat trial, it is possible that the common early differentialpositivity may reflect suppression of the previously active butnow irrelevant task-set (e.g., having just completed a lettertask, the letter task can be safely inhibited). As switch-awaytrials are associated with larger RT switch cost than switch-totrials and show no reduction in RT switch cost with increasingCSI, it would appear that, if such a suppression process exists,it has no effect on speed of responding. This, however, is notnecessarily true. It is possible that any behavioral effect of thissuppression or inhibition process would have reached aplateau by 200ms andwould only be evident when comparingperformance on trials with a completely non-informative cuevs. switch-away cues. This interpretation appears largelycompatible with Barceló et al.'s (2002) frontal P3a componentthat is elicited to feedback cues indicating a shift in task-setwithin the Wisconsin Card Sorting Task (WCST) and that canbe differentiated from a later parietal component that Barcelóet al. suggest is a P3B, which reflects retrieval of task-set intoworking memory. Like our switch-away cues, Barceló's 3D shiftfeedback cues indicate that the previous task-set is no longerrelevant but do not specify which task-set will be relevant onthe subsequent trial.3 However, this interpretation still contra-dicts the conclusion drawn on the basis of recent behavioraldata that inhibition is not an independent process, but a by-product of activation of the relevant task-set (e.g., Dreisbach etal., 2002; Hubner et al., 2003) or is triggered upon responseselection (Schuch and Koch, 2003). Clearly, there are differ-ences in the type and timing of information provided by ERPand behavioral data and further research is necessary toreconcile these discrepancies.
An alternative possibility is that the early differentialpositivity for both switch-away and switch-to relative to repeatcues reflects differing amounts of information provided by thecue.4 Specifically, repeat cues may be said to provide no newinformation as the same task-set will be implemented again,switch-away cues provide information about an impendingtask change, whereas switch-to cues also indicate which task-set needs to be implemented. Given the sensitivity of the P300component to stimulus information value (e.g., Rugg andColes, 1995), it is possible that the early portion of thedifferential positivity for switch-to and switch-away cuesrepresents modulation of P300 by cue information value.Within this framework, given that the early differential

171B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
positivity did not differ in amplitude between switch-to andswitch-away cues, it would then appear to reflect generalprocesses associated with preparing for a switch in task-set,whereas the prolonged differential positivity for switch-to cueswould reflect processes more specifically associated withactivation of the new task-set. Further work is needed todefine what is encompassed by the general preparationprocesses that are evident for both switch-to and switch-awaycues and to explain why this general preparation does notappear to contribute to behavioral performance (see earlierdiscussion).
Another possibility is that switch-away cues elicit theactivation of both possible task-sets and their maintenancein working memory. This could account for the overall largerRT cost on switch-away compared to switch-to trials and for thelack of a switch cost reduction with increasing CSI for switch-away trials as activation and maintenance of two task-setstake longer to be completed. However, if this were the case,then switch-away trials would be expected to show a larger andmore prolonged differential positivity in the CSI than switch-totrials, reflecting greater activation of task-set reconfigurationprocesses. This was clearly not the case in the current data.Furthermore, this interpretation cannot also account for theadditional post-stimulus positivity that occurred exclusivelyfor switch-away trials.
A related possibility is that, on switch-away trials, partici-pants activated one of the two possible task-sets basing theirchoice on subjective expectancies derived using local proba-bilities (e.g., if the digit task has not been presented recently, aparticipant may activate that task-set based on their assump-tion that there is an increased probability that it will bepresented on the next trial). In this case, on roughly half of thetrials they would have successfully reconfigured the correcttask-set, whereas on the other half they will have toreconfigure again after stimulus onset. This explanation canaccount for the overall larger RT switch cost for switch-awaythan switch-to trials, for the emergence of an early differentialpositivity for both types of switch trials and for the brief post-stimulus differential positivity on switch-away trials only.However, it is hard to reconcile with the reduced amplitudeand duration of the cue-locked differential positivity forswitch-away compared to switch-to trials as well as with theabsence of any reduction in RT switch cost with increasing CSIfor switch-away trials.
The differential early positivity for switch-to and switch-away trials could, alternatively, reflect differential processingof the cue on repeat compared to switch trials. That is, it maynot represent a process that is common to the two types ofswitch trials, but a process that occurs for repeat trials only.This differential positivity may then represent processes thatcontribute to a repetition benefit (Dreisbach et al., 2002; Loganand Bundesen, 2003; Mayr and Kliegl, 2003). This appears to beunlikely in the current data. The outline of the circle and thesix wedges were continuously displayed, and the cue involvedhighlighting two of the six wedges that were associated withparticular tasks (Fig. 4A). The cue itself was thus identical onall trials, only its spatial position across the circle differed.Additionally, Nicholson et al. (2006) show that, in wellpracticed participants, alternating between explicitly mappedand easy to process cues produces no behavioral effects of a
switch in cue, while ERP effects of a cue switch are restricted toearly endogenous components. On the other hand, based onthe argument by Dreisbach et al. (2002) that active preparationoccurs for the least expected trial type and the fact that,overall, there were fewer repeat than switch trials in thecurrent paradigm, it could be argued that task-set reconfigu-ration occurred here for repeat rather than switch trials.However, in that instance, one would expect that the switch-related differential positivity would occur for repeat trials andthat no switch cost would be observed, which was clearly notthe case.
3.4. Summary
The electrophysiological data are compatible with the con-struct of task-set reconfiguration as an active control processthat is necessary for successful switching between tasks.These findings suggest that the process of task-set reconfig-uration consists of multiple subcomponents, however it is theactivation of the currently relevant task-set prior to stimulusonset that facilitates the reduction in RT switch cost previ-ously observedwith increasing preparation interval.With longpreparation intervals and complete information about anupcoming switch trial, task-set reconfiguration can be com-pleted before stimulus onset. However, if the preparationinterval is insufficient or the information provided is inade-quate, then all or part of this process will be completed afterstimulus onset.
4. Experimental procedures
4.1. Participants
Thirty-six undergraduate students (mean age 23 years±6.8years, range 18 to 40 years; 29 female) participated for coursecredit in an introductory psychology course. Participants hadno prior exposure to task-switching paradigms and providedwritten informed consent. The study was approved by theNewcastle University Human Research Ethics Committee.
4.2. Stimuli and tasks
For each run of trials, a circle (230 pixels diameter) divided into6 equal wedges was continuously displayed in the middle of acomputer monitor at approximately 90 cm viewing distance.On each trial, a stimulus was displayed in the center of one ofthe six wedges (Fig. 4A). Pairs of adjacent wedges weregrouped by thicker lines extending slightly beyond theperimeter of the circle, thus demarcating three sections.Each of the three sections was assigned to one of threetasks. The assignment of task to section was counterbalancedacross participants.
In the letter task, participants respondedwhether the letterpresented was a vowel (A, E, I, U) or a consonant (G, K, M, R). Inthe digit task, participants responded whether the digit pre-sented was odd (3, 5, 7, 9) or even (2, 4, 6, 8). In the color task,participants responded whether the color of the stimulus washot (red, pink, orange, burgundy) or cold (ocean blue, emeraldgreen, sky blue, turquoise). Participants used their left and

172 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
right index fingers to respond (Fig. 4B), and the hand assignedto each response was counterbalanced across participants.
Stimuli consisted of a pair of characters in Times NewRoman font (60 by 60 pixels). Characters could be a letter, adigit or a non-alphanumeric character (#, ?, *, %) that was not
Fig. 4 – (A) Circle fixation grid in which participants are instructedthe stimulus is presented in section 1, task B (is the digit odd or evthe color hot or cold?) when the stimulus is presented in sectionhand is mapped to one response option for each of the three tassequence with either repeat task cue (indicates the same task wthat the task will switch on the next trial and showswhich task wtask will switch on the next trial but does not show which task wthe outline of the circle sections. (D) The three conditions used inactive and passive components to task-switching.
mapped to a response (neutral). Stimuli were presented eitherin gray (neutral) or in one of eight colors listed above. Eachstimulus therefore consisted of three dimensions: character 1,character 2 and color. One dimension was selected from thecurrently relevant task-set (i.e., a digit for the digit task). The
to perform task A (is the letter a vowel or a consonant?) whenen?) when the stimulus is displayed in section 2, and task C (is3. (B) Stimulus-response mappings for the three tasks: eachks. (C) Circle grid showing unpredictable task alternationill be repeated on the next trial), switch-to task cue (indicatesill be switched to) or switch-away task cue (indicates that theill be switched to). The cue was a highlight displayed aroundthe present study to differentiate between the contribution of

173B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
second dimension was selected from one of the two alterna-tive task-sets (i.e., a letter or a color). This second dimensionwas always incongruently mapped to the first dimension,whereas the third dimension was always neutral. For exam-ple, assume that the digit taskwas relevant on the current trialand that the number ‘3’ was presented, thus requiring a lefthand response. The second character could be either a letter ora neutral stimulus. If it was a letter, then it would have to be aconsonant, so as to be mapped to an incongruent response(i.e., right hand). In this case, the third stimulus dimension(color) would be neutral (i.e., gray). For example, the stimulus‘3G’ would be presented in gray and the correct responsewould be left. Alternatively, if the second character wasneutral (e.g., #), then the stimulus would be presented in oneof the cold colors, so that again it would be mapped to anincongruent response (right hand). For example, the stimulus‘3#’would be presented in green. The combination of stimulusdimensions and the position of task-relevant and task-irrelevant characters (e.g., 4U, U4) were varied pseudoran-domly across trials. Stimuli were selected pseudorandomlyfrom their character set with the only restriction that the samestimulus could not appear on two successive trials.
Each trial began with a cue that highlighted the bordersurrounding two of the six wedges (Fig. 4C). Three differenttrial types were defined by the location of the cue and werepseudorandomly selected with equal probability. The sametrial type was never repeated more than 3 times. On repeattrials (1/3 of trials), the cue highlighted the section of the circleassigned to the same task as on the previous trial. Fig. 4C(repeat sequence) shows that, having completed a letter tasktrial, the next cue signals that the following trial will also be aletter task trial. On half of the repeat trials, the stimulusappeared in the same wedge as on the previous trial, while onthe other half it appeared in the adjacent wedge within thattask section.5 The other two types of cues indicated that thenext trial would involve a switch in task, but provideddifferent degrees of information regarding what the nexttask would be. Switch-to cues (1/3 of trials) highlighted asection assigned to one of the other two tasks, thereby validlycueing which task-set would be active on the upcoming trial.Fig. 4C (switch-to sequence) shows that, having performed aletter task trial, the next cue indicates that the following trialwill involve a switch-to the digit task. Switch-away cues (1/3 oftrials) transgressed task sections and highlighted wedgesbelonging to the two tasks that were irrelevant on the previoustrial, thereby indicating that the current task would not berepeated, but not indicating which task-set would be active onthe upcoming trial. As shown in Fig. 4C (switch-awaysequence), the cue highlighted two segments, one assignedto the digit task and one assigned to the color task. Therefore,
5 As noted by an anonymous reviewer, stimulus locationrepetition is thus possible for task repetitions, whereas it cannotoccur for task-switch trials, possibly providing some extrapositive priming on task repeat trials. Unfortunately, this wasunavoidable as the alternative (i.e., changing the location onevery repeat trial) would have produced an even greaterconfound. That is, participants would have known the exactlocation of the upcoming stimulus on repeat trials only, possiblyfacilitating faster RT compared to switch trials where thestimulus could appear in one of two alternative locations.
having completed a letter trial, the highlight indicated that theupcoming trial would require a switch-away from the lettertask-set. The active task-set (digit or color) is then determinedby the position of the stimulus itself and thus the new task-setcannot be activated until after stimulus onset. The cue and thestimulus remained on the screen until a response was gene-rated or 5000 ms had elapsed.
Three timing conditions were used to vary the CSI and RSI(Fig. 4D). RSI was manipulated across two levels: short(1200 ms) and long (1600 ms). At the shorter RSI, CSI variedacross two levels: short (200ms) and long (1000ms). Conditionlabels represent the duration of the RSI and CSI (e.g., RSI-1200:CSI-200=RSI of 1200 ms and CSI of 200 ms).
4.3. Procedure
All participants attended 2 sessions scheduled 2 to 14 daysapart. The first session provided substantial task training andpractice. Training began with two runs (48 trials/run) on eachtask alone. This was followed by two runs (72 trials/run) ofrepeat and switch-to trials and two runs of repeat and switch-away trials. Session 1 finished with 2 runs (228 trials/run) ofcombined repeat, switch-to and switch-away trials. Throughouttraining, stimulus-response mapping (Fig. 4B) and task loca-tion (Fig. 4A) were displayed continuously at the bottom of thecomputer monitor and the CSI was a minimum of 600 ms.
The second session included further practice followed bythe behavioral and ERP testing session. Second day practiceincluded one run (48 trials/run) on each task alone, two runs(72 trials/run) with repeat and switch-to, and repeat and switch-away trials, respectively, and two runs (228 trials/run) ofcombined repeat, switch-to and switch-away trials (RSI-1200:CSI-1000 and RSI-1200:CSI-200, respectively). Participants thuscompleted a total of 1776 practice trials across the 2 trainingand practice sessions.
The behavioral and ERP testing session consisted of the 3timing conditions presented in blocks of 3 runs (228 trials/run).Each condition was presented in a separate block, and theorder of block presentation was counterbalanced acrossparticipants using a Latin square design. The first 12 trials ofevery run were consider ‘warm-up’ trials and were discardedfrom analysis. For all participants, stimulus-response map-ping and assignment of tasks to specific sections within thecircle remained constant across training and testing sessions.Participants were instructed to respond as quickly as possible,without making too many errors. At the start of each block,participants were informed of the specific RSI and CSI to beused and were encouraged to use the CSI to prepare for thenext trial based on the information provided by the cue.Incorrect responses resulted in auditory feedback, and theonset of the next cue was delayed by 1000 ms. A short breakwas provided at the end of each run, and behavioral feedback(overall average RT and percentage of trials correct) wasdisplayed. Participants were encouraged to monitor andimprove their performance.
4.4. Data analysis
The main analysis of both behavioral and ERP measures wasconducted on the data from digit and letter task trials alone.

174 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
Data from color task trials were excluded from the mainanalyses for a number of reasons. Firstly, this increasedcomparability with our earlier work that had included onlythe letter and digit tasks (Karayanidis et al., 2003; Nicholsonet al., 2005). Secondly, selective attention to color attributeshas been shown to result in differences in behavioral andERP measures of attention as compared to attention to otherattributes such as location, orientation, size and pattern (e.g.,Karayanidis and Michie, 1997; Michie et al., 1999), suggestingpossible differences in attentional processing of color andother attributes. Finally, initial inspection of the dataindicated that the pattern of behavioral findings wasdifferent for the color task than for the other two tasks.Mean RT was considerably faster for the color task (771 ms)than the other two tasks (883 ms; F(1,35)=51.58, p<0.001)and showed significantly less switch cost (switch-to cost: 122vs. 177 ms; F(1,35)= 18.36, p<0.001; switch-away cost: 220 vs.262 ms F(1,35)= 7.5, p<0.01). Further inspection of ERPwaveforms (Fig. 5) showed that the repeat trial waveformfor the color task was noticeably different to those for letterand digit tasks, resulting in spurious task differences in theswitch-related difference waveforms. These differences areinteresting insofar as they suggest a difference in task-switching processes dependent on task attributes and/ortask difficulty that need to be further explored in their ownright. However, as these effects are not central to the aims ofthis paper, color task trials were excluded from behavioraland ERP data analysis.
Trials associated with an incorrect response, trialsimmediately following an incorrect response and trialson which the response occurred outside a window of 200to 2000 ms after stimulus onset were excluded fromanalysis. Where necessary, critical values were adjustedusing the Greenhouse–Geisser correction to avoid violatingthe assumption of sphericity (Vasey and Thayer, 1987).Standard error values were calculated using the methodfor within-subjects designs described by Loftus and Mas-son (1994).
Fig. 5 – Cue-locked ERPs for repeat, switch-to and switch-away trfor each task.
4.4.1. Behavioral data analysisMean RT and error rates were computed for repeat, switch-toand switch-away trials separately for each condition and task.Error and RT switch cost for switch-to and switch-away trialswas calculated by subtracting the mean value for repeattrials from the mean value on switch-to and switch-awaytrials, respectively (i.e., switch-to cost= switch-to–repeat;switch-away cost=switch-away–repeat).
RT andarc sine transformedproportion error datawere firstanalyzed using a 3 condition (RSI-1200:CSI-200, RSI-1200:CSI-1000, RSI-1600:CSI-1000) by 3 trial type (repeat, switch-to,switch-away) by 2 task (letter, digit) repeated-measuresANOVA. Given significant main effects of trial type and inter-actions between trial type and other factors, RT and errorswitch cost measures were then examined in a 3 condition(RSI-1200:CSI-200, RSI-1200:CSI-1000, RSI-1600:CSI-1000) by 2switch cost type (switch-to switch cost, switch-away switch cost)by 2 task (letter, digit) repeated-measures ANOVA. The effectsof condition on switch cost type were examined in two sets ofplanned comparisons that were run separately for switch-toand switch-away costs (averaged across task with alphaadjusted using the Bonferonni correction). The effect ofincreasing CSI was examined by comparing RSI-1200:CSI-200and RSI-1200:CSI-1000. The effect of increasing RSI wasexamined by comparing RSI-1200:CSI-1000 and RSI-1600:CSI-1000.
4.4.2. EEG recording and data analysisElectroencephalogram (EEG) was recorded continuously from12 scalp electrodes according to the 10/20 system (Fz, Cz, Pz,Oz, F3, C3, P3, T5, F4, C4, P4, T6) using an electrode cap (Electro-cap International) and linked mastoids reference. Verticalelectro-oculogram (VEOG) was recorded bipolarly from elec-trodes attached to the supra-orbital and infra-orbital ridges ofthe left eye. Horizontal electro-oculogram (HEOG) wasrecorded bipolarly from electrodes placed on the outer canthiof each eye. EEG and EOG were continuously sampled at500 Hz/channel using NeuroScan Inc. and amplified (×5000 for
ials averaged over the two 1000 ms CSI conditions separately

175B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
EOG and frontal channels; ×20,000 for other EEG channels)using a Grass Neurodata system (Model 12) with a bandpass of0.01–30 Hz (−6 dB down).
Vertical eye movement artifact was corrected (Semlitsch etal., 1986), and the continuous EEG files were manually inspec-ted and sections with muscle artifact or channel saturationwere excluded from analysis (rejection criteria of outside −100to 100 μV). Cue- and stimulus-locked averages were created byextracting 1400 ms epochs around the onset of the cue orstimulus, respectively, with a 200mspre-onset interval. Due tolarge pre-baseline shifts in some conditions, baseline correc-tion was set −50 to 50 ms around the onset of the cue orstimulus (see Karayanidis et al., 2003).
For all three conditions, ERP waveforms were averagedacross task (letter/digit) in order to increase signal to noiseratio. Cue-locked and stimulus-locked epochs were averagedseparately for each condition and trial type. Nine cue- andninestimulus-locked (3 timing conditions×3 trial types) ERPaverage waveforms were created for each participant at eachelectrode site. Across all conditions and trial types, theretended to be slowpotential drift from the fronto-temporal sitesthat extended beyond the range of the epoch. This wasmostlylikely attributable to drift from the HEOG channel, which wasconsistent for all three trial types, as participants wererequired to make horizontal eye movements around the circlegrid. All average files were thus linearly detrended across theentire interval in order to remove this eye drift artifact.
As with the behavioral switch cost data, switch-to andswitch-away difference waveforms were calculated by sub-tracting the ERP repeat waveform from the ERP switch-towaveform and the ERP switch-away waveform, respectively.Six cue-locked and six stimulus-locked difference waveformswere thus created for each participant at each electrode site.Difference waveforms were analyzed using point-by-point t-tests over 50 to 1000 ms to identify points of significantdeviation of each waveform from baseline and points ofsignificant deviation between the two waveforms (i.e., switch-to vs. switch-away difference waveforms). This analysis wasconducted at the midline sites (Fz, Cz, Pz and Oz). The Guthrieand Buchwald (1991) procedure was used to control for Type 1error at α=0.01 using an autocorrelation coefficient of 0.9. Onlyeffects significant by these criteria are reported.
Acknowledgments
This work was supported by a postgraduate scholarship fromthe Neuroscience Institute of Schizophrenia and AlliedDisorders (NISAD) and the University of Newcastle to RebeccaNicholson. Thanks to Dr. Leuthold and an anonymousreviewer for insightful suggestions on earlier versions of themanuscript.
R E F E R E N C E S
Allport, D.A., Wylie, G., 2000. Task-switching, stimulus responsebindings, and negative priming. In: Monsell, S., Driver, J. (Eds.),Attention and Performance XVIII. MIT Press, Cambridge, MA,pp. 35–70.
Allport, A., Styles, E.A., Hsieh, S., 1994. Shifting intentional set:exploring the dynamic control of tasks. In: Umilta, C.,Moscovitvh, M. (Eds.), Attention and Performance XV. MITPress, Cambridge, MA, pp. 421–452.
Arbuthnott, K., Frank, J., 2000. Executive control in set switching:residual switch cost and task-set inhibition. Can. J. Exp.Psychol. 54 (1), 33–41.
Barcelo, F., Munoz-Cespedes, J.M., Pozo, M.A., Rubia, F.J., 2000.Attentional set shifting modulates the target P3b response inthe Wisconsin Card Sorting Test. Neuropsychologia 38 (10),1342–1355.
Barcelo, F.C.A., Perianez, J.A., Knight, R.T., 2002. Think differently:a brain orienting response to task novelty. NeuroReport 13 (15),1887–1892.
Brown, S., Lehmann, C., Poboka, D., 2006. A critical test of thefailure-to-engage theory of task switching. Psychon. Bull. Rev.13, 152–159.
De Jong, R., 2000. An intention-activation account of residualswitch costs. In: Monsell, S., Driver, J. (Eds.), Attention andPerformance XVIII. MIT Press, Cambridge, MA, pp. 357–376.
Dreisbach, G., Haider, H., Kluwe, R.H., 2002. Preparatory processesin the task-switching paradigm: evidence from the use ofprobability cues. J. Exper. Psychol., Learn., Mem., Cogn. 28 (3),468–483.
Gade, M., Koch, I., 2005. Linking inhibition to activation in thecontrol of task sequences. Psychon. Bull. Rev. 12 (3), 530–534.
Gehring, W.J., Bryck, R.L., Jonides, J., Albin, R.L., Badre, D., 2003.The mind's eye, looking inward? In search of executivecontrol in internal attention shifting. Psychophysiology 40 (4),572–585.
Goschke, T., 2000. Intentional reconfiguration and involuntarypersistence in task set switching. In: Monsell, S., Driver, J. (Eds.),Attention and Performance XVIII. MIT Press, Cambridge, MA,pp. 331–355.
Guthrie, D., Buchwald, J.S., 1991. Significance testing of differencepotentials. Psychophysiology 28, 240–244.
Hsieh, S., Liu, L.-C., 2005. The nature of switch cost: task setconfiguration or carry-over effect? Cogn. Brain Res. 22 (2),165–175.
Hsieh, S., Yu, Y.-T., 2003. Switching between simple response-sets:inferences from the lateralized readiness potential. Cogn. BrainRes. 17 (2), 228–237.
Hubner, M., Dreisbach, G., Haider, H., Kluwe, R.H., 2003. Backwardinhibition as a means of sequential task-set control: evidencefor reduction of task competition. J. Exper. Psychol., Learn.,Mem., Cogn. 29 (2), 289–297.
Karayanidis, F., Michie, P.T., 1997. Evidence of visual processingnegativity with attention to orientation and color in centralspace. Electroencephalogr. Clin. Neurophysiol. 103 (2),282–297.
Karayanidis, F., Coltheart, M., Michie, P.T., Murphy, K., 2003.Electrophysiological correlates of anticipatory andpost-stimulus components of task-switchingPsychophysiology 40 (3), 329–348.
Koch, I., 2005. Sequential task predictability in task switching.Psychon. Bull. Rev. 12 (1), 107–112.
Koch, I., Gade, M., Philipp, A.M., 2004. Inhibition of response modein task switching. Exp. Psychol. 51 (1), 52–58.
Lien, M.C., Ruthruff, E., Remington, R.W., Johnston, J.C., 2005. Onthe limits of advance preparation for a task switch: do peopleprepare all the task some of the time or some of the task allthe time? J. Exp. Psychol. Hum. Percept. Perform. 31 (2),299–315.
Loftus, G.R., Masson, M.E., 1994. Using confidence intervals inwithin-subject designs. Psychon. Bull. Rev. 1, 476–490.
Logan, G.D., 2003. Executive control of thought and action: insearch of the wild homunculus. Curr. Dir. Psychol. Sci. 12 (2),45–48.
Logan, G.D., Bundesen, C., 2003. Clever homunculus: is there an

176 B R A I N R E S E A R C H 1 1 2 1 ( 2 0 0 6 ) 1 6 0 – 1 7 6
endogenous act of control in the explicit task-cuing procedure?J. Exp. Psychol. Hum. Percept. Perform. 29 (3), 575–599.
Mayr, U., Keele, S.W., 2000. Changing internal constraints onaction: the role of backward inhibition. J. Exp. Psychol. Gen. 129(1), 4–26.
Mayr, U., Kliegl, R., 2000. Task-set switching and long-termmemory retrieval. J. Exper. Psychol., Learn., Mem., Cogn. 26 (5),1124–1140.
Mayr, U., Kliegl, R., 2003. Differential effects of cue changes andtask changes on task-set selection costs. J. Exper. Psychol.,Learn., Mem., Cogn. 29 (3), 362–372.
Meiran, N., 1996. Reconfiguration of processing mode prior to taskperformance. J. Exper. Psychol., Learn., Mem., Cogn. 22,1423–1442.
Meiran, N., 2000. Reconfiguration of stimulus task sets andresponse task sets during task switching. In: Monsell, S., Driver,J. (Eds.), Attention and Performance XVIII. MIT Press,Cambridge, MA, pp. 377–399.
Meiran, N., Chorev, Z., Sapir, A., 2000. Component processes intask switching. Cogn. Psychol. 41, 211–253.
Michie, P.T., Karayanidis, F., Smith, G.L., Barrett, N.A., Large, M.M.,O'Sullivan, B.T., Kavanagh, D.J., 1999. An exploration ofvarieties of visual attention: ERP findings. Cogn. Brain Res. 7 (4),419–450.
Miniussi, C., Marzi, C.A., Nobre, A.C., 2005. Modulation of brainactivity by selective task sets observed using event-relatedpotentials. Neuropsychologia 43, 1514–1528.
Monsell, S., 1996. Control of mental process. In: Bruce, V. (Ed.),Unsolved Mysteries of the Mind: Tutorial Essays in Cognition.Erlbaum, Taylor, and Francis, Hove, England, pp. 93–148.
Monsell, S., 2003. Task switching. Trends Cogn. Sci. 7 (3),134–140.
Monsell, S., Mizon, G.A., 2006. Can the task-cueing paradigmmeasure an “endogenous” task-set reconfiguration process?J. Exp. Psychol. Hum. Percept. Perform. 32 (3), 493–516.
Nicholson, R., Karayanidis, F., Poboka, D., Heathcote, A., Michie, P.,2005. Electrophysiological correlates of anticipatorytask-switching processes. Psychophysiology 42, 540–554.
Nicholson, R., Karayanidis, F., Bumak, E., Poboka, D., Michie, P.,2006. ERPs dissociate the effects of switching task-sets andtask-cues. Brain Res. 1095, 107–123.
Poulsen, C., Luu, P., Davey, C., Tucker, D.M., 2005. Dynamics of tasksets: evidence from dense-array event-related potentials.Cogn. Brain Res. 24, 133–154.
Rogers, R.D., Monsell, S., 1995. Costs of a predictable switchbetween simple cognitive tasks. J. Exp. Psychol. 124 (2),207–231.
Rugg, M.D., Coles, M.G.H., 1995. Electrophysiology of Mind:Event-related Brain Potentials and Cognition. Oxford Univ.Press, New York.
Rushworth, M.F.S., Passingham, R.E., Nobre, A.C., 2002.Components of switching intentional set. J. Cogn. Neurosci. 14(8), 1139–1150.
Rushworth, M.F.S., Passingham, R.E., Nobre, A.C., 2005.Components of attentional set-switching. Exp. Psychol. 52 (2),83–98.
Ruthruff, E., Remington, R.W., Johnston, J.C., 2001. Switchingbetween simple cognitive tasks: the interaction of top–downand bottom–up factors. J. Exp. Psychol. Hum. Percept. Perform.27 (6), 1404–1419.
Schuch, S., Koch, I., 2003. The role of response selection forinhibition of task sets in task shifting. J. Exp. Psychol. Hum.Percept. Perform. 29 (1), 92–105.
Semlitsch, H.V., Anderer, P., Schuster, P., Presslich, O., 1986. Asolution for reliable and valid reduction of ocular artifacts.Psychophysiology 23, 695–703.
Sohn, M.H., Carlson, R.A., 2000. Effects of repetition andforeknowledge in task-set reconfiguration. J. Exper. Psychol.,Learn., Mem., Cogn. 26 (6), 1445–1460.
Swainson, R., Cunnington, R., Jackson, G., Rorden, C., Peters, A.,Morris, P., Jackson, S.R., 2003. Cognitive control mechanismsrevealed by ERP and fMRI: evidence from repeatedtask-switching. J. Cogn. Neurosci. 15 (6), 785–799.
Vasey, M., Thayer, J., 1987. The continuing problem of falsepositives in repeated measures ANOVA inpsychophysiology: A multivariate solution.Psychophysiology 24 (4), 479–486.
Waszak, F., Hommel, B., Allport, A., 2003. Task-switching and longterm priming: role of episodic stimulus-task bindings intask-shift costs. Cogn. Psychol. 46 (4), 361–413.
Woodward, T.S., Meier, B., Tipper, C., Graf, P., 2003. Bivalency iscostly: bivalent stimuli elicit cautious responding. Exp. Psychol.50 (4), 233–238.
Wylie, G., Allport, A., 2000. Task switching and the measurementof ‘switch costs’. Psychol. Res. 63, 212–233.
Wylie, G.R., Javitt, D.C., Foxe, J.J., 2003. Task switching: ahigh-density electrical mapping study. NeuroImage 20 (4),2322–2342.




















