
Comparison between disturbed and undisturbed areas of the Patagonian scallop (Zygochlamys...
10
Comparison between disturbed and undisturbed areas of the Patagonian scallop (Zygochlamys patagonica) fishing ground “Reclutas” in the Argentine Sea Laura Schejter a,b, ⁎, Claudia S. Bremec a,b , Daniel Hernández b a CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentina b INIDEP, Instituto Nacional de Investigación y Desarrollo Pesquero, Paseo Victoria Ocampo 1, B7602HSA, Mar del Plata, Argentina ABSTRACT ARTICLE INFO Article history: Received 13 November 2007 Received in revised form 10 April 2008 Accepted 29 April 2008 Available online 17 June 2008 Keywords: Zygochlamys patagonica Scallop Fichery Benthic Assemblages Fishing Disturbance Argentina In this study we compare the composition, structure and biomass of the species assemblage in the Patagonian scallop management unit “Reclutas” (39°00′–39°30′S and 55°45′–56°05′W), in areas subjected to fishing effort (FG) and in the exclusion area (REX), between 1998 and 2002. Data obtained during 1995 and previous to the beginning of the scallop fishery are also considered as baseline condition. No differences in species richness or species composition were detected between areas in the study period. Patagonian scallop biomass was always higher at REX, but by-catch biomass was similar in both areas. The kind of analysis made during this study only allowed us to detect differences in the Patagonian scallop biomass between areas during the study period, while by-catch biomass, as a whole, apparently presented no differences between disturbed and undisturbed areas. Nonetheless, ABC curves in FG in 1998 matched with an undisturbed area, but from 2001 they showed a pattern coincident with a moderately disturbed area. In REX, the pattern observed in 1998 was not quite clear, but in 2002, this unfished area matched with and undisturbed area, as expected for an exclusion area. In addition, multidimensional analysis suggested a differentiation between FG and REX after 1998. Based on analysis of species (dis)similarities we could attribute this differentiation to an increase in predators biomass (Volutidae) and the detritivore ofiuroid Ophiactis asperula, a decrease in the fragile species (Sterechinus agassizii) and the target species of the fishery, Zygochlamys patagonica, and the founding of Chaetopterus variopedatus and Idanthyrsus armatus tubes in the fishing ground. © 2008 Elsevier B.V. All rights reserved. 1. Introduction Fishing results in the removal of a huge biomass of both target and non-target species. Trawling and dredging over the marine bottom produce an important effect on the ecosystem, damaging not only the substrate, but also the invertebrates associated with the bottom. These modifications could cause destruction of the benthic habitat and long and short-term changes in the composition and structure of the associated fauna (National Research Council, 2002). Undisturbed areas are usually characterized by high species richness and an abundance of erect and sessile epifauna, which also creates a complex of microhabitats available for other species. On the other hand, disturbed areas, subjected to moderate or severe fishing, suffer from reduced structural diversity and heterogeneity of the habitat and a dominance of thick-shelled bivalves, echinoderms and scavengers (Kaiser and Spencer, 1994; Lindley et al., 1995; National Research Council, 2002; Turner et al., 1999; Talman et al., 2004). Most of the studies of fishing impact on the benthic communities compare areas subjected to different levels of fishing effort, or they consider closure areas as undisturbed areas (Eleftheriou and Robertson, 1992; Lindeboom and de Groot, 1998; Lindegarth et al., 2000; McConnaughey et al., 2000; Dinmore et al., 2003). The Patagonian scallop, Zygochlamys patagonica (King and Broderip, 1832) is the target species of a new fishery in the Argentine Sea along the 100 m isobath, and it is distributed in the Magellanic Biogeographic Province (Lasta and Bremec, 1998). The management unit “Reclutas” is located between 39°00′–39°30′S and 55°45′–56°05′W. An exclusion area (39°20′–39°30′S and 55°52′–56°00′W, not allowed for commercial fishing of scallops) was established inside this management unit in 1996 (Resolution N°150, 19/3/1996, Secretaría de Agricultura, Pesca y Alimentación, Argentina). The aim of this study was to compare the biomass, composition and structure of the assemblage in the Patagonian scallop management unit “Reclutas”, in areas subjected to fishing effort (FG) and in the exclusion area (REX), in the period comprised between 1998 and 2002, in order to identify short-term faunistic changes. We discuss our results considering the baseline condition, the data obtained during 1995, previous to the beginning of the scallop fishery, reported by Bremec and Lasta (2002). 2. Materials and methods A total of 94 samples was collected at “Reclutas” (39°00′–39°30′S and 55°45′–56°05′W), 71 of which were taken in the fishing ground Journal of Sea Research 60 (2008) 193–200 ⁎ Corresponding author. CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentina. E-mail address: [email protected] (L. Schejter). 1385-1101/$ – see front matter © 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.seares.2008.04.007 Contents lists available at ScienceDirect Journal of Sea Research journal homepage: www.elsevier.com/locate/seares
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Comparison between disturbed and undisturbed areas of the Patagonian scallop (Zygochlamys...

Journal of Sea Research 60 (2008) 193–200
Contents lists available at ScienceDirect
Journal of Sea Research
j ourna l homepage: www.e lsev ie r.com/ locate /seares
Comparison between disturbed and undisturbed areas of the Patagonian scallop(Zygochlamys patagonica) fishing ground “Reclutas” in the Argentine Sea
Laura Schejter a,b,⁎, Claudia S. Bremec a,b, Daniel Hernández b
a CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentinab INIDEP, Instituto Nacional de Investigación y Desarrollo Pesquero, Paseo Victoria Ocampo 1, B7602HSA, Mar del Plata, Argentina
⁎ Corresponding author. CONICET, Consejo Nacional dTécnicas, Buenos Aires, Argentina.
E-mail address: [email protected] (L. Schejter).
1385-1101/$ – see front matter © 2008 Elsevier B.V. Alldoi:10.1016/j.seares.2008.04.007
A B S T R A C T
A R T I C L E I N F OArticle history:
In this study we compare Received 13 November 2007Received in revised form 10 April 2008Accepted 29 April 2008Available online 17 June 2008Keywords:Zygochlamys patagonicaScallop FicheryBenthic AssemblagesFishing DisturbanceArgentina
the composition, structure and biomass of the species assemblage in thePatagonian scallop management unit “Reclutas” (39°00′–39°30′S and 55°45′–56°05′W), in areas subjected tofishing effort (FG) and in the exclusion area (REX), between 1998 and 2002. Data obtained during 1995 andprevious to the beginning of the scallop fishery are also considered as baseline condition. No differences inspecies richness or species composition were detected between areas in the study period. Patagonian scallopbiomass was always higher at REX, but by-catch biomass was similar in both areas. The kind of analysis madeduring this study only allowed us to detect differences in the Patagonian scallop biomass between areasduring the study period, while by-catch biomass, as a whole, apparently presented no differences betweendisturbed and undisturbed areas. Nonetheless, ABC curves in FG in 1998 matched with an undisturbed area,but from 2001 they showed a pattern coincident with a moderately disturbed area. In REX, the patternobserved in 1998 was not quite clear, but in 2002, this unfished area matched with and undisturbed area, asexpected for an exclusion area. In addition, multidimensional analysis suggested a differentiation between FGand REX after 1998. Based on analysis of species (dis)similarities we could attribute this differentiation to anincrease in predators biomass (Volutidae) and the detritivore ofiuroid Ophiactis asperula, a decrease in thefragile species (Sterechinus agassizii) and the target species of the fishery, Zygochlamys patagonica, and thefounding of Chaetopterus variopedatus and Idanthyrsus armatus tubes in the fishing ground.
© 2008 Elsevier B.V. All rights reserved.
1. Introduction
Fishing results in the removal of a huge biomass of both target andnon-target species. Trawling and dredging over the marine bottomproduce an important effect on the ecosystem, damaging not only thesubstrate, but also the invertebrates associated with the bottom. Thesemodifications could cause destruction of the benthic habitat and longand short-term changes in the composition and structure of theassociated fauna (National Research Council, 2002). Undisturbed areasare usually characterized by high species richness and an abundance oferect and sessile epifauna,which also creates a complexofmicrohabitatsavailable for other species. On the other hand, disturbed areas, subjectedto moderate or severe fishing, suffer from reduced structural diversityand heterogeneity of the habitat and a dominance of thick-shelledbivalves, echinoderms and scavengers (Kaiser and Spencer,1994; Lindleyet al., 1995; National Research Council, 2002; Turner et al., 1999; Talmanet al., 2004). Most of the studies of fishing impact on the benthiccommunities compare areas subjected to different levels offishing effort,or they consider closure areas as undisturbed areas (Eleftheriou and
e Investigaciones Científicas y
rights reserved.
Robertson,1992; Lindeboom and de Groot,1998; Lindegarth et al., 2000;McConnaughey et al., 2000; Dinmore et al., 2003).
The Patagonian scallop, Zygochlamys patagonica (King and Broderip,1832) is the target species of a newfishery in theArgentine Sea along the100 m isobath, and it is distributed in the Magellanic BiogeographicProvince (Lasta and Bremec, 1998). The management unit “Reclutas” islocated between 39°00′–39°30′S and 55°45′–56°05′W. An exclusionarea (39°20′–39°30′S and 55°52′–56°00′W, not allowed for commercialfishing of scallops)was established inside thismanagement unit in 1996(Resolution N°150, 19/3/1996, Secretaría de Agricultura, Pesca yAlimentación, Argentina). The aim of this study was to compare thebiomass, composition and structure of the assemblage in the Patagonianscallop management unit “Reclutas”, in areas subjected to fishing effort(FG) and in the exclusion area (REX), in the period comprised between1998 and 2002, in order to identify short-term faunistic changes. Wediscuss our results considering the baseline condition, the data obtainedduring1995, previous to the beginningof the scallopfishery, reportedbyBremec and Lasta (2002).
2. Materials and methods
A total of 94 samples was collected at “Reclutas” (39°00′–39°30′Sand 55°45′–56°05′W), 71 of which were taken in the fishing ground

Table 1Number of analyzed samples in the fishing ground (FG) and in the exclusion area (REX)of the Patagonian scallop management unit “Reclutas” during the study period
Year/area FG REX
1998 35 82001 24 62002 12 9
194 L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
(FG) and 23 in the exclusion area (REX) of the same management unitduring assessment research cruises (R/V Capitán Cánepa, INIDEP) in1998, 2001 and 2002 (Table 1, Fig. 1). Samples were collected using a2.5 m wide dredge of 100 mm mesh. This gear is not selective forPatagonian scallops; it permits the collection of the associatedepibenthos that conforms the by-catch of the fishery (Lasta andBremec, 1998; Bremec et al., 2000). This dredge performs efficientlywhen used in the assessment of Patagonian scallops from softbottoms, reaching 43%, as demonstrated by Valero (unpublished).This value highly exceeds the fraction caught in coastal rockyenvironments (18%, Iribarne et al., 1991) and the fraction caught byotter trawls in fishing grounds (21–31%, Lasta and Iribarne, 1997).
The sampling procedure was designed for stock assessment of Z.patagonica in the fishing grounds and not particularly for every singleby-catch species; it only considers the proportion (% in wet weight) ofinvertebrates by-catch. The sampling was performed every yearfollowing the same design of stations, which are spaced regularly at5 nm intervals. In general, given the geo-statistical methods used forthe resource evaluation, the total number of sampling sites in thedifferent areas or beds is in accordance with their area in km2 (Lasta etal., 2001). Consequently, our samples represent the only availablesource of data of associated fauna to make comparisons among areasacross time, and they add ecological information about the benthiccommunity in a fisheries context. Identification of organisms,abundance and biomass data were obtained on board (1998 and
Fig. 1. Localization of the Zygochlamys patagonica management unit “Reclutas” in the Argrepresent sample sites in the exclusion area (REX).
2001) and at the laboratory (2002). Data indicate abundance andbiomass of organisms per 100 m2. Epibiont invertebrates on scallopsand other organisms were excluded from this analysis.
The Mann–Whitney Test (Zar, 1984) was employed to testdifferences of taxa richness and scallop biomass and by-catch biomassbetween areas (FG-REX) during the study period (1998–2002). Multi-variate analysis using PRIMER software was performed (Clarke, 1993;Clarke and Gorley, 2001), as recommended for these kinds of studies(Lindeboom and de Groot, 1998). Hierarchical and multidimensionalanalysis (Bray–Curtis similarity index, fourth root transformation) andSIMPER test were developed to establish similarities and dissimilaritiesbetween stations and species assemblages. Abundance/biomass com-parative method (ABC curves) (Warwick, 1986; Warwick and Clarke,1994) was used to assign disturbance levels in the different studiedareas and years. This method permits the assessment of the pollution/disturbance status of a marinemacrobenthic community (undisturbed,moderately disturbed, heavily disturbed) following a conceptual modeldescribed inWarwick (1986).More details and examples of thismethodcould also be found in Warwick and Clarke (1994).
3. Results
Fifty six taxa were registered at “Reclutas” (FG+REX), most of thempresent in both areas (Table 2). Number of taxa per station in thefishing area (FG) varied between 6 and 25 while in the exclusion area(REX), it varied between 11 and 24. No differences in species richnesswere detected between areas in the study period (Mann–Whitneytest, pN0.05 in all cases).
Four invertebrate taxa (Austrocidaris canaliculata, Cosmasterias lurida,Ctenodiscus australis and Porifera) were closely associated with thePatagonian scallop in every year and in both areas, representingapproximately 40% (ormore) in biomass contribution (SIMPER analysis)to the community assemblage. The addition of eight other common taxa(Ophiactis asperula, Ophiacanta vivipara, Libidoclaea granaria, Actinostolacrassicornis, Fusitriton magellanicus, Calyptraster sp., Flabellum sp. and
entine Sea. Circles represent sampled sites of the fishing ground (FG) while triangles
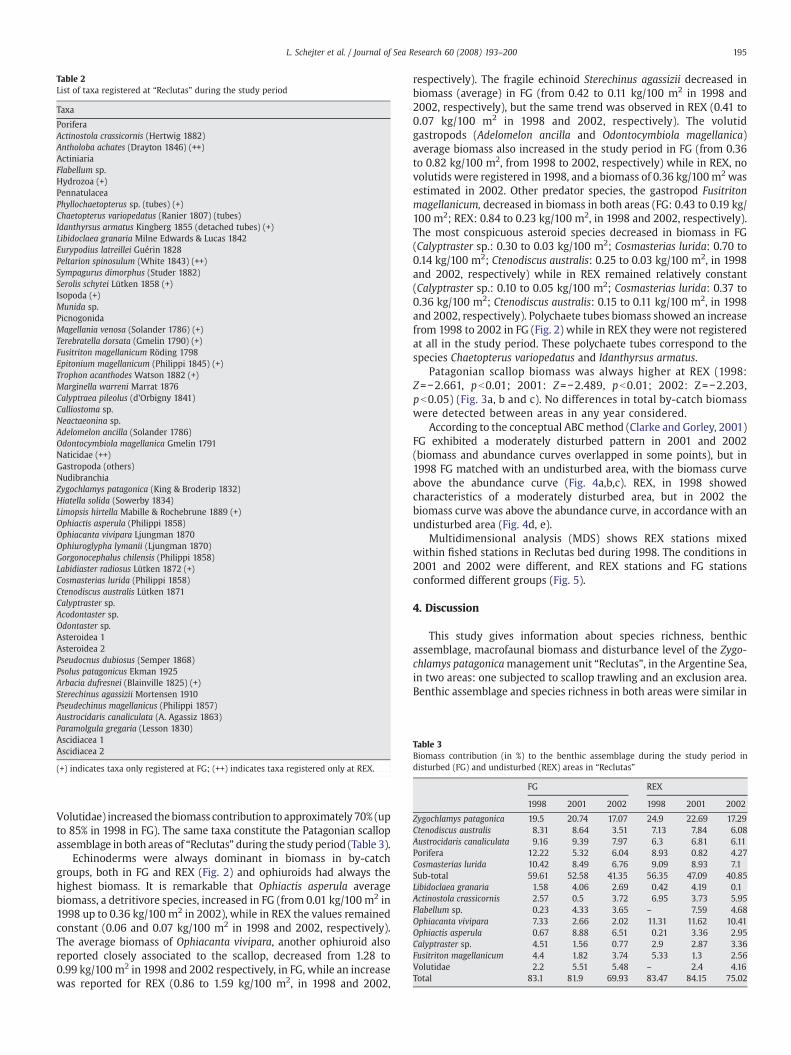
Table 2List of taxa registered at “Reclutas” during the study period
Taxa
PoriferaActinostola crassicornis (Hertwig 1882)Antholoba achates (Drayton 1846) (++)ActiniariaFlabellum sp.Hydrozoa (+)PennatulaceaPhyllochaetopterus sp. (tubes) (+)Chaetopterus variopedatus (Ranier 1807) (tubes)Idanthyrsus armatus Kingberg 1855 (detached tubes) (+)Libidoclaea granaria Milne Edwards & Lucas 1842Eurypodius latreillei Guérin 1828Peltarion spinosulum (White 1843) (++)Sympagurus dimorphus (Studer 1882)Serolis schytei Lütken 1858 (+)Isopoda (+)Munida sp.PicnogonidaMagellania venosa (Solander 1786) (+)Terebratella dorsata (Gmelin 1790) (+)Fusitriton magellanicum Röding 1798Epitonium magellanicum (Philippi 1845) (+)Trophon acanthodes Watson 1882 (+)Marginella warreni Marrat 1876Calyptraea pileolus (d'Orbigny 1841)Calliostoma sp.Neactaeonina sp.Adelomelon ancilla (Solander 1786)Odontocymbiola magellanica Gmelin 1791Naticidae (++)Gastropoda (others)NudibranchiaZygochlamys patagonica (King & Broderip 1832)Hiatella solida (Sowerby 1834)Limopsis hirtella Mabille & Rochebrune 1889 (+)Ophiactis asperula (Philippi 1858)Ophiacanta vivipara Ljungman 1870Ophiuroglypha lymanii (Ljungman 1870)Gorgonocephalus chilensis (Philippi 1858)Labidiaster radiosus Lütken 1872 (+)Cosmasterias lurida (Philippi 1858)Ctenodiscus australis Lütken 1871Calyptraster sp.Acodontaster sp.Odontaster sp.Asteroidea 1Asteroidea 2Pseudocnus dubiosus (Semper 1868)Psolus patagonicus Ekman 1925Arbacia dufresnei (Blainville 1825) (+)Sterechinus agassizii Mortensen 1910Pseudechinus magellanicus (Philippi 1857)Austrocidaris canaliculata (A. Agassiz 1863)Paramolgula gregaria (Lesson 1830)Ascidiacea 1Ascidiacea 2
(+) indicates taxa only registered at FG; (++) indicates taxa registered only at REX.
Table 3Biomass contribution (in %) to the benthic assemblage during the study period indisturbed (FG) and undisturbed (REX) areas in “Reclutas”
FG REX
1998 2001 2002 1998 2001 2002
Zygochlamys patagonica 19.5 20.74 17.07 24.9 22.69 17.29Ctenodiscus australis 8.31 8.64 3.51 7.13 7.84 6.08Austrocidaris canaliculata 9.16 9.39 7.97 6.3 6.81 6.11Porifera 12.22 5.32 6.04 8.93 0.82 4.27Cosmasterias lurida 10.42 8.49 6.76 9.09 8.93 7.1Sub-total 59.61 52.58 41.35 56.35 47.09 40.85Libidoclaea granaria 1.58 4.06 2.69 0.42 4.19 0.1Actinostola crassicornis 2.57 0.5 3.72 6.95 3.73 5.95Flabellum sp. 0.23 4.33 3.65 – 7.59 4.68Ophiacanta vivipara 7.33 2.66 2.02 11.31 11.62 10.41Ophiactis asperula 0.67 8.88 6.51 0.21 3.36 2.95Calyptraster sp. 4.51 1.56 0.77 2.9 2.87 3.36Fusitriton magellanicum 4.4 1.82 3.74 5.33 1.3 2.56Volutidae 2.2 5.51 5.48 – 2.4 4.16Total 83.1 81.9 69.93 83.47 84.15 75.02
195L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
Volutidae) increased thebiomass contribution to approximately 70% (upto 85% in 1998 in FG). The same taxa constitute the Patagonian scallopassemblage in both areas of “Reclutas” during the study period (Table 3).
Echinoderms were always dominant in biomass in by-catchgroups, both in FG and REX (Fig. 2) and ophiuroids had always thehighest biomass. It is remarkable that Ophiactis asperula averagebiomass, a detritivore species, increased in FG (from 0.01 kg/100 m2 in1998 up to 0.36 kg/100 m2 in 2002), while in REX the values remainedconstant (0.06 and 0.07 kg/100 m2 in 1998 and 2002, respectively).The average biomass of Ophiacanta vivipara, another ophiuroid alsoreported closely associated to the scallop, decreased from 1.28 to0.99 kg/100m2 in 1998 and 2002 respectively, in FG, while an increasewas reported for REX (0.86 to 1.59 kg/100 m2, in 1998 and 2002,
respectively). The fragile echinoid Sterechinus agassizii decreased inbiomass (average) in FG (from 0.42 to 0.11 kg/100 m2 in 1998 and2002, respectively), but the same trend was observed in REX (0.41 to0.07 kg/100 m2 in 1998 and 2002, respectively). The volutidgastropods (Adelomelon ancilla and Odontocymbiola magellanica)average biomass also increased in the study period in FG (from 0.36to 0.82 kg/100 m2, from 1998 to 2002, respectively) while in REX, novolutids were registered in 1998, and a biomass of 0.36 kg/100m2 wasestimated in 2002. Other predator species, the gastropod Fusitritonmagellanicum, decreased in biomass in both areas (FG: 0.43 to 0.19 kg/100 m2; REX: 0.84 to 0.23 kg/100 m2, in 1998 and 2002, respectively).The most conspicuous asteroid species decreased in biomass in FG(Calyptraster sp.: 0.30 to 0.03 kg/100 m2; Cosmasterias lurida: 0.70 to0.14 kg/100 m2; Ctenodiscus australis: 0.25 to 0.03 kg/100 m2, in 1998and 2002, respectively) while in REX remained relatively constant(Calyptraster sp.: 0.10 to 0.05 kg/100 m2; Cosmasterias lurida: 0.37 to0.36 kg/100 m2; Ctenodiscus australis: 0.15 to 0.11 kg/100 m2, in 1998and 2002, respectively). Polychaete tubes biomass showed an increasefrom 1998 to 2002 in FG (Fig. 2) while in REX they were not registeredat all in the study period. These polychaete tubes correspond to thespecies Chaetopterus variopedatus and Idanthyrsus armatus.
Patagonian scallop biomass was always higher at REX (1998:Z=−2.661, pb0.01; 2001: Z=−2.489, pb0.01; 2002: Z=−2.203,pb0.05) (Fig. 3a, b and c). No differences in total by-catch biomasswere detected between areas in any year considered.
According to the conceptual ABCmethod (Clarke and Gorley, 2001)FG exhibited a moderately disturbed pattern in 2001 and 2002(biomass and abundance curves overlapped in some points), but in1998 FG matched with an undisturbed area, with the biomass curveabove the abundance curve (Fig. 4a,b,c). REX, in 1998 showedcharacteristics of a moderately disturbed area, but in 2002 thebiomass curve was above the abundance curve, in accordance with anundisturbed area (Fig. 4d, e).
Multidimensional analysis (MDS) shows REX stations mixedwithin fished stations in Reclutas bed during 1998. The conditions in2001 and 2002 were different, and REX stations and FG stationsconformed different groups (Fig. 5).
4. Discussion
This study gives information about species richness, benthicassemblage, macrofaunal biomass and disturbance level of the Zygo-chlamys patagonicamanagement unit “Reclutas”, in the Argentine Sea,in two areas: one subjected to scallop trawling and an exclusion area.Benthic assemblage and species richness in both areas were similar in

Fig. 2. Mean biomass of by-catch groups (in kg/100 m2) registered for the study periodat the fishing grounds (FG) and at the exclusion area (REX) in “Reclutas”.
196 L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
the study period. Moreover, the mentioned species are also commonto other Patagonian scallop management units in the Argentine Sea,and were also present in 1995 (pre-fished condition) (Bremec andLasta, 2002; Bremec et al., 2003). Differences in richness during thestudy period were mainly due to differences in sampling effort (thatwas higher in the last sampled year) and to the expertise acquired inthe identification of the taxa recorded. In addition, fewer sampleswere taken from REX than FG, so rare or occasional species that wererecorded in FG were less likely to be observed in REX. Biodiversity andrichness values of the benthic communities subjected to fishing effortare very variable (Thrush and Dayton, 2002). Brocken and Kenching-ton (1999) found no differences in the richness and composition ofbenthic communities between fished and unfished areas. Otherstudies (Veale et al., 2000; McConnaughey et al., 2000) have detecteda loss in biodiversity and richness and a decrease in the biomass of
colonial and fragile organisms in trawled areas. However, not all thebenthic communities have shown this trend (Collie et al., 1997;National Research Council, 2002). Additionally, there are evidences ofprofound and highly statistically-significant change in the mega-benthic community in scallop fishing grounds in the Bay of Fundy(Canada) where no loss of species and no change in average number oftaxa per station were reported but dominance of many speciesdrastically changed (Kenchington et al., 2007).
In Lunenberg County, Canada, Placopecten magellanicus recruitswere more abundant in an exclusion area than in fished areas(Brocken and Kenchington, 1999). In Argentina, since the beginning ofthe Patagonian scallop fishery, important recruitment events wereregistered in 1994, 1995 and 1999 (Valero, unpublished). The lastsuccessful and massive recruitment in all commercial managementunits of the Argentine Seawere registered in 2000 and 2001 (M. Lasta,pers. com). In 2002 only a low frequency of recruits was detected infew samples of “Reclutas” (in both, FG and REX) (Campodónico andLasta, 2002). Considering this data, no differences in recruitabundance were found between FG and REX.
Fluctuations in biomass and abundance of by-catch groups havebeen documented in other regions. Reduction of sessile and fragilespecies were reported by Eleftheriou and Robertson (1992), NationalResearch Council (2002) and Wassenberg et al. (2002), whileincrements in scavenger, opportunistic and predator species werementioned by Kaiser and Spencer (1996), Ramsay et al. (1998), Vealeet al. (2000). Kaiser and Spencer (1996) and Demestre et al. (2000)pointed out that the arrival of scavengers and predators following atrawling event occurred in the first hours (8–10 h in average,depending on the species). After 24–48 h, this rapid increase inpredators and scavengers as a consequence of damaged or deadorganisms diminished to normal levels due to the fast consumption ofavailable food. In our study, no significant changes in total by-catchbiomass were observed among areas in the studied years. Notwith-standing, some species were reported with variations in biomass, bothbetween areas and years. Biomass variations of scavengers, predators,fragile and sessile species in our study could be partially attributed tofishing activities. For example, the increment in biomass of theophiuroid Ophiactis asperula and the predator volutid gastropods, thedecrease of the fragile echinoid species Sterechinus agassizzi and theincrease in polychaete tubes biomass in FG could be a consequence offishing disturbance. Chaetopterus variopedatus is an infaunal, filter-feeder tubicolous polychaete that lives permanently inside its tube(Avant, 2002). The tubes registered in the samples were sometimesempty or broken. These tubes could be removed from the bottomwiththe nets and could be transported because of the fishery fleet activitiesor the bottom currents. On the other hand, I. armatus builds sandytubes that are usually registered as epibionts on Zygochlamyspatagonica valves (Schejter and Bremec, 2007). The detection andthe increase of these isolated tubes in FG suggest their detachmentfrom the valves, perhaps attributable to trawling activities. Sessiletaxa, like Porifera and Cnidaria, decreased in biomass from 1995 to1998 (Bremec et al., 2000), but our present data only show irregularfluctuations. Moreover, life cycles and population dynamics of themajority of the invertebrate species living in the benthic communitydominated by the Patagonian scallop and herein considered areunknown (Bremec and Echeverría, 2005), and hence, it is not possibleto differentiate between natural and anthropogenic variations.
The exclusion area (REX) had a higher biomass of scallops than thefishing area in the study period. However, total biomass and by-catchbiomass were similar in both areas between 1998 and 2002. Bremec(unpubl. data) estimated an average value of 850 scallops per 100 m2
(up to 3962 ind. 100 m2) that correspond to 4.5 kg whole scallops per100 m2 (range: 0.6–15.2 kg per 100 m2) in 1995. Although theexclusion area was established in a patch with a high density ofscallops, other small patches with the same high densities of scallopswere present in all the bed, suggesting that our results are not

Fig. 3. Zygochlamys patagonica and by-catch biomass in 1998 (a), 2001 (b) and 2002 (c) in the undisturbed area (REX) and the fishing ground (FG) of “Reclutas”. ⁎ indicates significancedifferences between areas (Mann–Whitney test, pN0.05).
197L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
reflecting a “site effect”. The average scallop biomass in FGwas 5.79 kgper 100m2 in 1998, 5.44 kg per 100m2 in 2001 and 4.67 kg per 100m2
in 2002. On the contrary, the average scallop biomass in REX was14.12 kg per 100 m2 in 1998, 14.54 kg per 100 m2 in 2001 and 7.86 kgper 100 m2 in 2003. Based on general results and considering also thepre-fished condition in 1995, it is probable that REX was preserved bythe commercial fleet during the study period, allowing the growth ofthe scallop recruits registered in 1995. The high values of biomassregistered in this area coincide with the peaks registered in 1995,while in FG average scallop biomass values were always lower. Thebiomass registered in REX in 2002 was lower than the biomass
registered for the same area in previous years and this could be relatedto natural mortality of scallops, but sporadic fishery events cannot bediscounted. Scallop biomass was still higher in REX than in FG during2002.
Bremec et al. (2000) concluded that at least eight samples arenecessary for the majority of the species to be represented. Therefore,precaution is needed for interpretation of data from 2001 in theexclusion area. The statistical analysis donewas limited due to the lownumber of samples collected at REX and because samples were notindependent between years. For that reason comparisons were madefor average biomass values from both areas but only within each year.

Fig. 4. ABC curves estimated for the fishing area (FG) and the exclusion area (REX) of “Reclutas” during the study period. An UNDISTURBED condition (biomass curve above abundancecurve) could be observed in 1998 in the FG and in 2002 in REX (the 1998 condition in REX is not clear). The other years in FG presented aMODERATELY DISTURBED condition, showingthe abundance and the biomass curves overlapped in some points.
198 L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
Under stable unpolluted/undisturbed conditions, where thebenthic community is approaching equilibrium, the biomass willbecame increasingly dominated by one or few large species (Warwick,1986). ABC curves in FG in 1998 matched with an undisturbed area,but they showed a pattern coincident with a moderately disturbedarea during 2001 and 2002, with almost no difference between theabundance and biomass curves, which were very close and crossingeach other one or more times. This fact is probably related to thefishing effort in this management unit. Between 1996 (when fisherystarted) and 1998, only ~600 commercial hauls were done in the studyarea, which could be considered as a very low fishing effort. However,Reclutas bed was heavily fished between 1998 and 2001, before oursampling period, reaching ~4500 commercial hauls (the equivalent tomore than 30% of the total catch recorded for the total PatagonianScallop Fishery in the Argentine Sea). Again, before our samplingperiod in 2002, ~800 commercial hauls were performed, which couldbe considered at least as a moderate fishing effort (Data Source:Patagonian Scallop Project, INIDEP). This fishing effort explains whythe multidimensional scaling diagram of 1998 showed the REX
stations and FG stations in one grouping, while in 2001 and 2002,they constituted separate groups. Although there was not loss ofspecies, no difference in species richness was reported and associatedspecies to the Patagonian scallop were the same in both areas duringthe whole study period. We can attribute differentiation of FG andREX after 1998 to the moderately disturbed condition reported, basedin the biomass changes of many species: increase in FG of the detri-tivore ophiuroid Ophiactis asperula and the predator volutid gastro-pods, the founding and increase of polychaete tubes (these last ones,absent in REX), and a decrease in the fragile echinoid Sterechinusagassizii and in the Patagonian scallop. These biomass changes agreewith the general pattern already mentioned (increment of scavenger,predator and opportunistic species and a decrease of fragile and sessilespecies) for other disturbed areas (i.e. Eleftheriou and Robertson,1992;National Research Council, 2002; Wassenberg et al., 2002; Kaiser andSpencer, 1996; Ramsay et al., 1998; Veale et al., 2000). However, in thiscommunity, the decrease of other predators (starfishes and the gas-tropod Fusitriton magellanicum) cannot be explained by these generaltrends.

Fig. 5.MDS ordination plots of the 3 study years. Circles correspond to FG stations whileblack triangles correspond to REX stations.
199L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
The analysis here performed allowed us to detect differences inPatagonian scallop biomass between areas and to establish amoderate disturbance level of the community in the fishing ground.Our results represent a case of study with information about pre-fishery condition and annual sampling since 1995; they provide areference tool to monitor the evolution of the benthic communityassociated with Patagonian scallops in areas subjected to anthropo-genic disturbance in the Argentine Sea.
Acknowledgements
This study was part of a PhD Thesis to Laura Schejter. We wouldlike to thank very much Gaby and Guilla (INIDEP Librarians) forhelping with the bibliography and Dr. O. Iribarne for the criticalrevision and suggestions on the preliminary versions of this study. Wewould also like to thank very much the anonymous reviewers whocontribute to the improvement of this article. Finally, we would like tothank Corina Bremec for the revision of English text. This study waspartially supported by the Fund. Antorchas No. 13900-13, SECyT PICT01-15080, CONICET PIP 5009. This is INIDEP Contribution 1522.
References
Avant, P., 2002. Chaetopterus variopedatus. Parchment Worm. Marine Life InformationNetwork: Biology and Sensitivity Key Information Sub-programme [on-line].Marine Biological Association of the United Kingdom, Plymouth. bhttp://www.marlin.ac.uk/species/Chaetopterusvariopedatus.htmN.
Bremec, C., Brey, T., Lasta, M., Valero, J., Lucifora, L., 2000. Zygochlamys patagonica bedson the Argentinian shelf: Part I: energy flow through the scallop bed community.Arch. Fish. Mar. Res. 48, 295–303.
Bremec, C.S., Lasta, M.L., 2002. Epibenthic assemblage associated with scallop(Zygochlamys patagonica) beds in the Argentinian shelf. Bull. Mar. Sci. 70, 89–105.
Bremec, C., Marecos, A., Schejter, L., Lasta, M., 2003. Guía técnica para la identificaciónde invertebrados epibentónicos asociados a los bancos de vieira patagónica(Zygochlamys patagonica) en el Mar Argentino. Publ. Esp. INIDEP, pp. 28.
Bremec, C., Echeverría, A., 2005. Zygochlamys patagonica fishery in the Argentine Sea.Invertebrates by-catch: a preliminary review on life spans of taxa. INIDEP Tech. Rep.70, pp. 23.
Brocken, F., Kenchington, E., 1999. A comparison of scallop (Placopecten magellanicus)population and community characteristics between fished and unfished areas inLunenburg country, N.S., Canada. Can. Tech. Rep. Fish. Aq. Sci. 2258, 93.
Campodónico, S., Lasta,M., 2002. InformedeCampañaCC-10-2002. EvaluacióndeBiomasade Vieira Patagónica. Unidad Norte de Manejo: Bancos Reclutas y MDQ, área “a”.Complementariamente: Estación Fija Banco Reclutas. Tech. Rep., INIDEP, pp. 50.
Clarke, K.R., 1993. Non-parametric multivariate analysis of changes in communitystructure. Austr. Ecol. 18, 117–143.
Clarke, K.R., Gorley, R.N., 2001. PRIMER v5: user manual/Tutorial. PRIMER-E:, Plymouth,United Kingdom, p. 91.
Collie, J.S., Escanero, G.A., Valentine, P.C., 1997. Effects of bottom fishing on the benthicmegafauna of Geaorges Bank. In: Lindeboom, H.J., de Groot, S.J. (Eds.), Impact II. Theeffects of different types of fisheries on the North Sea and Irish Sea benthicecosystems. Nioz-rapport 1998-1, RIVO-DLO Report C003/98.
Demestre, M., Sánchez, P., Kaiser, M.J., 2000. The behavioural response of benthicscavengers to otter trawling disturbance in the Mediterranean. In: Kaiser, M.J., deGroot , S.J. (Eds.), Effects of fishing on non-target species and habitats. BlackwellScience, Oxford, pp. 121–129.
Dinmore, T.A., Duplisea, D.E., Rackham, B.D., Maxwell, D.L., Jennings, S., 2003. Impact oflarge-scale area closure on patterns of fishing disturbance and consequences forbenthic communities. ICES J. Mar. Sci. 60, 371–380.
Eleftheriou, A., Robertson, M.R., 1992. The effects of experimental scallop dredging on thefauna and physical environment of a shallow sandy community. J. Sea Res. 30, 289–299.
Iribarne, O.O., Lasta, M.L., Vacas, H.C., Parma, A.M., Pascual, M.S., 1991. Assessment ofabundance, gear efficiency and disturbance in a scallop dredge fishery: results of adepletion experiment. In: Shumway, S.E., Sandifer , P.A. (Eds.), Scallop biology andculture. The world aquaculture society, selected papers from the 7th Int. Pect.Workshop, pp. 244–248.
Kaiser, M.J., Spencer, B.E., 1994. Fish scavenging behavior in recently trawled areas. Mar.Ecol., Prog. Ser. 112, 41–49.
Kaiser, M.J., Spencer, B.E., 1996. Behavioral responses of scavengers to beam trawldisturbance In: Greenstreet, S.P.R., Tasker , M.L. (Eds.), Aquatic predators and theirprey. Liverpool University Press, Liverpool, pp. 169–218. Chapter 16.
Kenchington, E.L., Kenchigton, J.T., Henry, L.A., Fuller, S., Gonzalez, P., 2007. Multi-decadal changes in the megabenthos of the Bay of Fundy: the effects of fishing. J.Sea Res. 58, 220–240.
Lasta, M.L., Iribarne, O.O., 1997. Southwester Atlantic scallop (Zygochlamys patagonica)fishery: assessment of gear efficiency through a depletion experiment. J. ShellfishRes. 16, 59–62.
Lasta, M.L., Bremec, C.S., 1998. Zygochlamys patagonica in the Argentine Sea: a newscallop fishery. J. Shellfish Res. 17, 103–111.
Lasta, M., Hernández, D.R., Bogazzi, E., Burgos, G.E., Valero, J.L., Lucifora, L., 2001. Uso detécnicas geoestadísticas en la estimación de la abundancia de vieira patagónica(Zygochlamys patagonica). Rev. Invest. Desarro. Pesq. 14, 95–108.
Lindeboom, H.J., de Groot, S.J., 1998. Impact-II: the effects of different types of fisherieson the North Sea and Irish Sea benthic ecosystems. Nioz-rapport 1998-1, RIVO-DLOReport C003/98, p. 404.
Lindegarth, M., Valentinsson, D., Hansson, M., Ulmestrand, M., 2000. Effects of trawlingdisturbances on temporal and spatial structure on benthic soft-sedimentassemblages in Gullmarsfjorden, Sweden. ICES J. Mar. Sci. 57, 1369–1376.
Lindley, J.A., Gamble, J.C., Hunt, H.G., 1995. A change in the zooplancton of the centralNorth Sea (55° to 58°N): a possible consequence of changes in the benthos. Mar.Ecol., Prog. Ser. 119, 299–303.
McConnaughey, R.A., Mier, K.L., Dew, C.B., 2000. An examination of chronic fishing effectson soft bottom benthos of the eastern Bering Sea. ICES J. Mar. Sci. 57, 1377–1388.
National Research Council, 2002. Effects of trawling and dredging on seafloor habitat.Committee on Ecosystem Effects of Fishing: Phase 1– Effects of Bottom Trawling onSeafloor Habitats, Ocean Studies Board, Division on Earth and Life Studies. NationalAcademy, Washington DC, p. 126.
Ramsay, K., Kaiser, M.J., Hughe, R.N., 1998. Responses of benthic scavengers to fishingdisturbance by towed gears in different habitats. J. Exp. Mar. Biol. Ecol. 224, 73–89.
Schejter, L., Bremec, C., 2007. Benthic richness in the Argentine continental shelf: therole of Zygochlamys patagonica (Mollusca: Bivalvia: Pectinidae) as settlementsubstrate. J. Mar. Biol. Assoc. U. K. 87, 917–925.
Talman, S.G., Norkko, A., Thrush, S.F., Hewitt, J.E., 2004. Habitat structure and thesurvival of juvenile scallops Pecten novazelandiae: comparing predation in habitatswith varying complexity. Mar. Ecol., Prog. Ser. 269, 197–207.
Thrush, S.F., Dayton, P.K., 2002. Disturbance to marine benthic habitats by trawling anddredging: implications for marine biodiversity. Ann. Rev. Ecolog. Syst. 33, 449–473.

200 L. Schejter et al. / Journal of Sea Research 60 (2008) 193–200
Turner, S.J., Thrush, S.F., Hewitt, J.E., Cummings, V.J., Funnel, G., 1999. Fishing impactsand the degradation or loss of habitat structure. Fish Manag. Ecol. 6, 401–420.
Valero, J., Unpublished 2002. Analysis of temporal and spatial variation in growth andnatural mortality estimation with an integrated dynamic model in the Patagonianscallop (Zygochalmys patagonica). Thesis of Master Science (School of Aquatic andFishery Sciences, University of Washington, Seattle, USA), unpublished.
Veale, L.O., Hill, A.S., Hawkins, S.J., Brand, A.R., 2000. Effects of long term physicaldisturbance by commercial scallop fishing on subtidal epifaunal assemblages andhabitats. Mar. Biol. 137, 325–337.
Warwick, R.M., 1986. A new method for detecting pollution effects on marinemacrobenthic communities. Mar. Biol. 92, 557–562.
Warwick, R.M., Clarke, K.R., 1994. Relearning the ABC: taxonomic changes andabundance/biomass relationships in disturbed benthic communities. Mar. Biol.118, 736–744.
Wassenberg, T.J., Dews, G., Cook, S.D., 2002. The impact of fish trawls on megabenthos(sponges) on the north-west shelf of Australia. Fish. Res. 58, 141–151.
Zar, J.H., 1984. Biostatistical Analysis. Prentice-Hall, New York. 622 pp.

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Erratum
Erratum to “Comparison between disturbed and undisturbed areas of the Patagonianscallop (Zygochlamys patagonica) fishing ground “Reclutas” in the Argentine Sea”[J. Sea Research 60/3 (2008) 193]
Laura Schejter a,b,⁎, Claudia S. Bremec a,b, Daniel Hernández b
a CONICET, Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentinab INIDEP, Instituto Nacional de Investigación y Desarrollo Pesquero, Paseo Victoria Ocampo 1, B7602HSA, Mar del Plata, Argentina
The published article mentioned in M&M section a mesh size of 100 mm, but this is not correct, because the mesh size of the dredge is 10 mm.
Journal of Sea Research 61 (2009) 275
DOI of original article: 10.1016/j.seares.2008.04.007.⁎ Corresponding author. CONICET, Consejo Nacional de Investigaciones Científicas y
E-mail address: [email protected] (L. Schejter).
1385-1101/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.seares.2009.01.006
Contents lists available at ScienceDirect
Journal of Sea Research
j ourna l homepage: www.e lsev ie r.com/ locate /seares










