
Combining histology, stable isotope analysis and ZooMS collagen fingerprinting to investigate the...
12
Combining histology, stable isotope analysis and ZooMS collagen fingerprinting to investigate the taphonomic history and dietary behaviour of extinct giant tortoises from the Mare aux Songes deposit on Mauritius L.G. van der Sluis a, ⁎, H.I. Hollund b , M. Buckley c , P.G.B. De Louw d , K.F. Rijsdijk e , H. Kars a a VU University Amsterdam, Institute for Geo- and Bioarchaeology, De Boelelaan 1085, 1081 HV Amsterdam, the Netherlands b Archaeological Museum, University of Stavanger, Peder Klows gate 30 A 4010 Stavanger, Norway c Manchester Institute of Biotechnology, Faculty of Life Sciences, 131 Princess Street, Manchester M1 7DN, UK d Deltares, Department of Soil and Groundwater, P.O. Box 85467, 3508 AL Utrecht, the Netherlands e Computational GeoEcology Group, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, P.O. Box 94248, 1090 GE Amsterdam, the Netherlands abstract article info Article history: Received 15 February 2014 Received in revised form 29 May 2014 Accepted 4 June 2014 Available online xxxx Keywords: Taphonomy Diagenesis Stable isotopes ZooMS Cylindraspis Taphonomic research of bones can provide additional insight into a site's formation and development, the burial environment and ongoing post-mortem processes. A total of 30 tortoise (Cylindraspis) femur bone samples from the Mare aux Songes site (Mauritius) were studied histologically, assessing parameters such as presence and type of microbial alteration, inclusions, staining/infiltrations, the degree of microcracking and birefringence. The absence of microbial attack in the 4200 year old Mare aux Songes bones suggests the animals rapidly entered the soil whole-bodied and were sealed anoxically, although they suffered from biological and chemical degrada- tion (i.e. pyrite formation/oxidation, mineral dissolution and staining) related to changes in the site's hydrology. Additionally, carbon and nitrogen stable isotopes were analysed to obtain information on the animals' feeding be- haviour. The results show narrowly distributed δ 13 C ratios, indicating a terrestrial C 3 plant-based diet, combined with a wide range in δ 15 N ratios. This is most likely related to the tortoises' drought-adaptive ability to change their metabolic processes, which can affect the δ 15 N ratios. Furthermore, ZooMS collagen fingerprinting analysis successfully identified two tortoise species (C. triserrata and C. inepta) in the bone assemblage, which, when com- bined with stable isotope data, revealed significantly different δ 15 N ratios between the two tortoise species. As climatic changes around this period resulted in increased aridity in the Mascarene Islands, this could explain the extremely elevated δ 15 N ratio in our dataset. The endemic fauna was able to endure the climatic changes 4200 years ago, although human arrival in the 17th century changed the original habitat to such an extent that it resulted in the extinction of several species. Fortunately we are still able to study these extinct tortoises due to the beneficial conditions of their burial environment, resulting in excellent bone preservation. © 2014 Elsevier B.V. All rights reserved. 1. Introduction The Mare aux Songes (MaS) site, initially discovered by Higginson and Clark in 1865 on Mauritius (Clark, 1866; Hume and Prys-Jones, 2005), has been subject to several excavations (Cheke, 1987; Staub, 1996; Hume, 2005). New excavations were performed in 2006 and 2007 after the rediscovery of an undisturbed fossil layer, which contained more than 20 native vertebrate species (Rijsdijk et al., 2009). Fossil assemblages are rare on volcanic islands due to their usually acidic soils, which are unfavourable for bone preservation. Additionally, the presence of bones from the extinct dodo and two giant tortoise species augments the uniqueness of this multitaxic assemblage. Rijsdijk et al. (2009, 2011) reconstructed that climatic changes around 4200 years ago resulted in a dry period in the Mascarene Islands, probably affecting vertebrate faunal populations in this region. This paper will report the results obtained from histological, stable isotope analysis and ZooMS collagen fingerprinting performed on 30 giant tortoise samples from the Mare aux Songes site to reconstruct the taphonomic history of the bones and to provide insight into the feeding behaviour of these extinct animals. To understand what happened at this site in more detail, it is essential to study taphonomic processes and development of the burial environment. Taphonomic research comprises:”… the study of processes of preservation and how they affect information in the fossil record” Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx ⁎ Corresponding author at: School of Geography, Archaeology and Palaeoecology, Queen's University Belfast, BT7 1 NN Northern Ireland, UK. Tel.: +44 78 478 02897. E-mail addresses: [email protected] (L.G. van der Sluis), [email protected] (H.I. Hollund), [email protected] (M. Buckley), [email protected] (P.G.B. De Louw), [email protected] (K.F. Rijsdijk), [email protected] (H. Kars). PALAEO-06900; No of Pages 12 http://dx.doi.org/10.1016/j.palaeo.2014.06.003 0031-0182/© 2014 Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo Please cite this article as: van der Sluis, L.G., et al., Combining histology, stable isotope analysis and ZooMS collagen fingerprinting to investigate the taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeoecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Combining histology, stable isotope analysis and ZooMS collagen fingerprinting to investigate the...

Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
PALAEO-06900; No of Pages 12
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
Combining histology, stable isotope analysis and ZooMS collagenfingerprinting to investigate the taphonomic history and dietarybehaviour of extinct giant tortoises from the Mare aux Songes depositon Mauritius
L.G. van der Sluis a,⁎, H.I. Hollund b, M. Buckley c, P.G.B. De Louw d, K.F. Rijsdijk e, H. Kars a
a VU University Amsterdam, Institute for Geo- and Bioarchaeology, De Boelelaan 1085, 1081 HV Amsterdam, the Netherlandsb Archaeological Museum, University of Stavanger, Peder Klows gate 30 A 4010 Stavanger, Norwayc Manchester Institute of Biotechnology, Faculty of Life Sciences, 131 Princess Street, Manchester M1 7DN, UKd Deltares, Department of Soil and Groundwater, P.O. Box 85467, 3508 AL Utrecht, the Netherlandse Computational GeoEcology Group, Institute for Biodiversity and Ecosystem Dynamics, University of Amsterdam, P.O. Box 94248, 1090 GE Amsterdam, the Netherlands
⁎ Corresponding author at: School of Geography, ArQueen's University Belfast, BT7 1 NN Northern Ireland, UK
E-mail addresses: [email protected] (L.G. van(H.I. Hollund), [email protected] (M. Buckley)(P.G.B. De Louw), [email protected] (K.F. Rijsdijk), h.kars
http://dx.doi.org/10.1016/j.palaeo.2014.06.0030031-0182/© 2014 Elsevier B.V. All rights reserved.
Please cite this article as: van der Sluis, L.G.,the taphonomic history and dietary..., Palaeo
a b s t r a c t
a r t i c l e i n f oArticle history:Received 15 February 2014Received in revised form 29 May 2014Accepted 4 June 2014Available online xxxx
Keywords:TaphonomyDiagenesisStable isotopesZooMSCylindraspis
Taphonomic research of bones can provide additional insight into a site's formation and development, the burialenvironment and ongoing post-mortem processes. A total of 30 tortoise (Cylindraspis) femur bone samples fromtheMare aux Songes site (Mauritius)were studied histologically, assessing parameters such as presence and typeof microbial alteration, inclusions, staining/infiltrations, the degree of microcracking and birefringence. Theabsence of microbial attack in the 4200 year old Mare aux Songes bones suggests the animals rapidly enteredthe soil whole-bodied and were sealed anoxically, although they suffered from biological and chemical degrada-tion (i.e. pyrite formation/oxidation, mineral dissolution and staining) related to changes in the site's hydrology.Additionally, carbon andnitrogen stable isotopeswere analysed to obtain information on the animals' feeding be-haviour. The results show narrowly distributed δ13C ratios, indicating a terrestrial C3 plant-based diet, combinedwith a wide range in δ15N ratios. This is most likely related to the tortoises' drought-adaptive ability to changetheir metabolic processes, which can affect the δ15N ratios. Furthermore, ZooMS collagen fingerprinting analysissuccessfully identified two tortoise species (C. triserrata and C. inepta) in the bone assemblage, which, when com-bined with stable isotope data, revealed significantly different δ15N ratios between the two tortoise species. Asclimatic changes around this period resulted in increased aridity in the Mascarene Islands, this could explainthe extremely elevated δ15N ratio in our dataset. The endemic fauna was able to endure the climatic changes4200 years ago, although human arrival in the 17th century changed the original habitat to such an extent thatit resulted in the extinction of several species. Fortunately we are still able to study these extinct tortoises dueto the beneficial conditions of their burial environment, resulting in excellent bone preservation.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The Mare aux Songes (MaS) site, initially discovered by Higginsonand Clark in 1865 on Mauritius (Clark, 1866; Hume and Prys-Jones,2005), has been subject to several excavations (Cheke, 1987; Staub,1996; Hume, 2005). New excavations were performed in 2006 and2007 after the rediscovery of an undisturbed fossil layer, whichcontained more than 20 native vertebrate species (Rijsdijk et al.,2009). Fossil assemblages are rare on volcanic islands due to their
chaeology and Palaeoecology,. Tel.: +44 78 478 02897.der Sluis), [email protected], [email protected]@vu.nl (H. Kars).
et al., Combining histology, stgeogr. Palaeoclimatol. Palaeo
usually acidic soils, which are unfavourable for bone preservation.Additionally, the presence of bones from the extinct dodo and twogiant tortoise species augments the uniqueness of this multitaxicassemblage. Rijsdijk et al. (2009, 2011) reconstructed that climaticchanges around 4200 years ago resulted in a dry period in theMascarene Islands, probably affecting vertebrate faunal populations inthis region. This paper will report the results obtained from histological,stable isotope analysis and ZooMS collagen fingerprinting performed on30 giant tortoise samples from the Mare aux Songes site to reconstructthe taphonomic history of the bones and to provide insight into thefeeding behaviour of these extinct animals.
To understand what happened at this site in more detail, it isessential to study taphonomic processes and development of the burialenvironment. Taphonomic research comprises:”… the study of processesof preservation and how they affect information in the fossil record”
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

2 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
(Behrensmeyer and Kidwell, 1985). Prior to performing [biomolecularor chemical] analyses, it is important to understand to what extent thesamples and information recorded inside them has been affected bypost-mortem processes in the burial environment. Previous researchinto taphonomy of the bone assemblage (Meijer et al., 2012) reportedan overrepresentation of tortoise hind limb elements, which suggestedmiring, whereby the heavy animals got trappedwith their hind limbs inthe swampy area, while their cranial and front limbs were exposed toscavengers and weathering. Meijer et al.'s macroscopic examinationalso revealed overall good preservation of the bones with minor degreeof bioerosion or mechanical degradation.
A microscopic, or histological, assessment of the bones may provideadditional evidence on taphonomic processes and the dynamics of theburial environment. Histology has proved to be a valuable tool in study-ing taphonomic processes (Jans et al., 2004; Turner-Walker and Jans,2008; Hollund et al., 2012), which can reveal traces (occasionally evena sequence) of changes in the burial environment. For example, the de-gree and pattern of bacterial alteration observed on a histological scalemay suggest whether or not the corpse was defleshed or intact whenit entered the burial environment, thus revealing information on theearly post-mortem stages (Schoeninger et al., 1989; Jans et al., 2004;Hollund et al., 2012). As bone can undergo numerous post-mortemchanges, before, during and after burial, the sequence of these changescan be referred to as the taphonomic history of bones (Turner-Walkerand Jans, 2008). Changes in site hydrology or pH can have significant in-fluence on bone preservation state and stability. Histological analyses canthus revealwhich bones suffered fromdegradation and are less suited foranalyses. The MaS site constitutes a unique example of a large faunalbone depositionwithout human interference, thus providing an excellentopportunity for improving our understanding of bone diagenetic andtaphonomic processes which could be relevant for similar buried sites.
Whereas histological investigations may elucidate what happenedto the tortoises after death, stable isotope analysis can provide informa-tion on the animals' feeding behaviour during life. Mauritius used to beinhabited by two giant tortoise species which were characterised bydifferent carapaces (shells). As the two tortoise species are extinct, study-ing their diet may reveal parts of their lives that would otherwise remainunknown. For example, it has been suggested that carapace morphologywas related to the type of vegetation present in their habitat (Arnold,1979; Burleigh and Arnold, 1986), which in turn might have affectedthe tortoises' choice of food as well. Carbon stable isotope values can re-veal the type of consumed vegetation, i.e. C3, C4/CAM and marine plants(DeNiro and Epstein, 1978), while nitrogen stable isotopes are used to de-termine the trophic level of an animal (Schoeller, 1999). However, somedifficulties exist in the interpretation of the nitrogen stable isotope ratios,which can be influenced by factors other than diet (Hedges and Reynard,2007). For example, salt spray in coastal areas can result in elevated δ15Nvalues (Virginia and Delwiche, 1982; Heaton, 1987), while aridity can in-fluence the animal's metabolism to retain as much water as possible,which can result in adjustments of thenitrogen stable isotope distribution(Heaton, 1986, 1987; Sealy et al., 1987; Ambrose, 1991; Koch, 1998). Thisis likely to occur in tortoises due to their ability to modify their nitrogenexcretion for water retention (Styring et al., 2010).
As the bone assemblage most likely consisted of two giant tortoisespecies, it would be worthwhile to investigate any inter-species dietarypreferences. However, asmost boneswere incomplete, it was not possi-ble to separate species based on measurements. To potentially over-come this, Zooarchaeology by Mass Spectrometry (ZooMS) analysiswas performed to examine whether or not it would be possible to dis-tinguish the two species based on collagen fingerprinting.
Specifically, this study aims to:
• Use bone histological analysis to provide information on site forma-tion processes
• Assess the state of bone preservation and identify the type of degrada-tion processes
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
• Assess palaeodietary behaviour of extinct giant tortoises• Explore possibilities of using ZooMS analysis to separate two closelyrelated tortoise species
1.1. Description of the site
Located in the southwest Indian Ocean roughly 800 km east ofMadagascar (Fig. 1A),Mauritius is part of theMascarene archipelago to-gether with Réunion and Rodrigues. Due to the island's volcanic origin,lava tunnels created in coastal areas collapsed with time, resulting inhydrologically inter-connected basins (Fig. 1B), which became filledwith carbonate sands originating from the reef surrounding the island(Rijsdijk et al., 2009). As the basins were hydrologically still connectedto the ocean, rising sea levels supported the formation of a small freshwater lens [see below], which supposedly attracted numerous terrestri-al animals. Circa 4200 B.P. the lake became rapidly filled with animalcorpses and considerable amounts of plantmaterial. Radiocarbon datingsuggests the entire fossil assemblage was accumulated over just twocenturies (Rijsdijk et al., 2009). However, the presence of laminatedorganic sediments indicative of low energetic deposition rejects thepossibility of a catastrophic event (e.g. tsunamis). Rijsdijk et al. (2009)suggest the animals died of water shortage caused by an extremearidification event that affected Mauritius, resulting from climaticchanges in the Indian Ocean (Marchant and Hooghiemstra, 2004; DeBoer et al., 2014). Inmore recent years, the sitewas infilledwith doleriteblocks in an attempt to control malaria (Rijsdijk et al., 2009).
1.2. Hydrological setting of the site
Situated in a depression at mean ocean level, the MaS site is con-stantly fed by a flow of groundwater originating from higher grounds.Groundwater levels in MaS show a tidal fluctuation of about 10 cmindicating a strong subsurface connection to the ocean. Present ground-water levels correlate with mean ocean levels (0 m mean sea levels(M.S.L.)) and water levels are unlikely to drop below ocean level.Groundwater flowing through coral sands located in the subsoil dis-solves CaCO3, resulting in alkaline conditions conducive to bone preser-vation (Nielsen-Marsh et al., 2007). The present chemical compositionof MaS's groundwater confirms these alkaline conditions (averageCa2+ = 60 mg/l; average HCO3
− = 280 mg/l; pH = 7.1–7.8). Inacidic volcanic settings as on Mauritius, alkaline conditions arerare and occur exclusively in the coastal belt (Willaime, 1984).Groundwater in MaS also contains elevated concentrations of iron(average Fe2+ = 17 mg/l), sulphate (average SO4
2− = 11 mg/l)and chloride (average Cl− =50 mg/l). Solute enrichment of ground-water exfiltrating in MaS results from the mineralisation of the or-ganic peat layer, dissolution of the coral sands and solutes like SO4
and Cl originate from ocean water. Salinity measurements publishedby Rijsdijk et al. (2009) show an increasing groundwater salinity gra-dient towards the ocean, indicating the presence of a nearby saltwa-ter wedge.
1.3. Extinct giant tortoises
Giant tortoises used to inhabit many islands in the Indian Oceanwith extinct species such as Aldabrachelys inhabiting Madagascar,Aldabra, the Seychelles and surrounding islands, and Cylindraspisinhabiting the Mascarene Islands (Arnold, 1979). The latter were hostto five different species of giant tortoises, one on Réunion (C. indica),two on Rodrigues (C. peltastes and C. vosmaeri) and two on Mauritius(C. inepta and C. triserrata). Although currently extinct, written sourcessuggest that Mascarene tortoise populations were quite large aroundthe year 1500 AD (Austin and Arnold, 2001).
It is still unclear where the founder species of the Mascarenetortoises originated. Phylogenetic research revealed that Mauritiuswas the first of the Mascarene Islands colonised by the giant tortoises,
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

Fig. 1. (A) Location of MaS site on Mauritius. (B) The MaS site with three deeper basins and different trenches (Rijsdijk et al., 2009).
Table 1Sample information of the bone material.
Sample Genus Bone element Bone number Trench
MaS-tr0-01 Cylindraspis Femur dex AG-717 0MaS-tr0-02 Cylindraspis Femur dex AG-718 0
3L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
after which they floated to Réunion and Rodrigues (Austin and Arnold,2001). A weak relationship between the Mascarene tortoises andtortoises from Western Africa, Madagascar and the Seychelles was ob-served, while Mauritian lizards and geckos were originating from bothMadagascar and surrounding islands, and Australasia. However, due tothe enormous distance between Australasia and Mauritius, a westernorigin for the tortoises appears more likely (Austin and Arnold, 2001).
The two Mauritian species are morphologically distinct in mostbones, although their carapaces differ the most. C. inepta carried adome-shaped carapace, while C. triserrata was characterised by asaddleback-shaped carapace (Fig. 2). It has been suggested that thesedifferences in carapace are related to the degree of openness of the veg-etation (Arnold, 1979; Burleigh and Arnold, 1986) and perhaps climaticchanges; a drier period could increase the vegetation openness, benefi-cial for themobility of saddleback-shaped tortoises,while dome-shapedtortoises would thrive in wetter periods with more closed vegetation.Remarkably, the radiocarbon-dated fossil record shows that both spe-cies were coexisting and flourishing on the same island. Assumingthat C. inepta and C. triserrata were sharing their habitat, the difference
Fig. 2. Dome- and saddelback-shaped carapaces (Arnold, 1979, 134).
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
in carapacemorphology could result from different dietary preferences.A case study (Burleigh and Arnold, 1986) fromMadagascar using stableisotope analysis on two giant tortoise species lacking the differences incarapace morphology revealed that tortoises focused on different foodsources (C3 and C4/CAM plants) to minimise interspecies competition.Something similar may have been the case for the Mauritian tortoisespecies.
2. Material
Frommaterial excavated in 2006, 30 tortoise femur bones were sam-pled, originating from four trenches in basin I and basin III (Table 1).
MaS-tr0-03 Cylindraspis Femur dex AG-699 0MaS-tr0-04 Cylindraspis Femur dex AG-731 0MaS-tr0-05 Cylindraspis Femur dex AG-732 0MaS-tr0-06 Cylindraspis Femur dex AG-760 0MaS-tr0-07 Cylindraspis Femur dex AG-668 0MaS-tr0-08 Cylindraspis Femur dex AG-670 0MaS-tr1-01 Cylindraspis Femur dex mas 2006 626 1MaS-tr1-02 Cylindraspis Femur dex mas 2006 625 1MaS-tr1-03 Cylindraspis Femur dex MAS 2006 661 1MaS-tr1-04 Cylindraspis Femur dex MAS 2006 645 1MaS-tr2-01 Cylindraspis Femur dex mas 2006 913 2MaS-tr2-02 Cylindraspis Femur dex mas 2006 915 2MaS-tr2-03 Cylindraspis Femur sin AG-251 2MaS-tr2-04 Cylindraspis Femur dex mas 2006 1392 2MaS-tr2-05 Cylindraspis Femur dex AG-568 2MaS-tr2-06 Cylindraspis Femur dex AG-601 2MaS-tr2-07 Cylindraspis Femur dex AG-240 2MaS-tr2-08 Cylindraspis Femur dex? mas 2006 889 2MaS-tr2-09 Cylindraspis Femur sin? AG-632 2MaS-tr2-10 Cylindraspis Femur sin? AG-631 2MaS-tr2-11 Cylindraspis Femur sin mas 2006 1276 2MaS-tr2-12 Cylindraspis Femur sin mas 2006 1020 2MaS-tr2-13 Cylindraspis Femur sin mas 2006 1284 2MaS-tr2-14 Cylindraspis Femur sin mas 2006 031 2MaS-tr2-15 Cylindraspis Femur dex AG-219 2MaS-tr2-16 Cylindraspis Femur sin AG-250 2MaS-tr2-17 Cylindraspis Femur dex MAS 2006 458 2MaS-tr3-01 Cylindraspis Femur dex mas 2006 1376 3
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

4 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
Trench 0 and trench 1 are located close to the edge of basin I, while trench2 and 3were positioned closer to the centre of basins I and III respectively(Fig. 1B). Eight bones originated from trench 0, four bones from trench 1,17 bones from trench 2 and one bone from trench 3. No depths wererecorded during excavation of the bones. The fossil layer (layer ‘D’) waslocated directly underneath the dolerite blocks, circa 10 cm underneaththe surface level (Rijsdijk et al., 2009). No bones with pathologicalfeatures or other surficial alterations were sampled. Right femurs weresampled when possible to ensure different individuals were analysed.U-shaped pieces of cortex of circa 1 cm by 1 cm were taken from eachbone. As complete bone specimens from the MaS site could not leavethe island, sampling was performed on site. Sampling was done indoorsand precautions were taken to reduce inter-bone contamination consid-ering future ancient DNA analyses.
3. Methodology
3.1. Histology
In order to reduce inter-bone contamination, gloves and dremeldrill equipment were cleaned with a wire brush and ethanol, whiledremel cutting wheels were replaced between sampling specimens.To study the bone histology, thin sections weremade by cutting boneslices of approximately 3–8 mm, which were embedded in AralditeDBF and hardener H956. From these embedded samples, thin sliceswere cut and ground using a diamond grinding wheel on a Jacobson618 grinding machine to circa 70 μm and afterwards covered withCanada balsam and a glass cover slip. Thin sections were studiedusing a Leica Dialux 20 microscope with magnifications of 40, 100,160, 250 and 400.
Scanning electron microscopy (SEM) was performed on one sample(MaS-tr0-07). This sample was selected to determine the orange grains'constituents, which were observed in the thin section. A thick sectionembedded in epoxy resin was polished and coated with carbon. Theanalysis was performed at the Dutch Cultural Heritage Agency usingthe SEM-EDS (Energy Dispersive Spectroscopy) JEOL JSM 5910 LVwith SiliconDrift Detector (SDD) fromThermoScientific andNoran Sys-tem Seven (NSS) software, producing images in backscattered electron(BSE) mode.
Diagenetic features visible in histological thin sections have been de-scribed extensively by several authors (Hackett, 1981; Garland, 1987;Jans, 2005). For the current study, five diagenetic features were exam-ined: (1) the presence and type of microbial alteration, (2) inclusions,(3) infiltrations (staining), (4) extent of microcracking and (5) extentof birefringence. Microbial attack (or bioerosion) can consist of tunnelscaused by bacteria (Hackett, 1981; Bell, 1990; Bell et al., 1996),fungi (Hackett, 1981; Piepenbrink, 1986; Garland, 1987; Martill, 1989;Child, 1995) or algae (Arnaud et al., 1978; Davis, 1997). The overall ex-tent of bioerosion is summarised in the Oxford Histological Index (OHI)(Hedges et al., 1995), ranging from 5 (unaltered) to 0 (no original mi-crostructure present). Inclusions consist of exogenous material locatedwithin the voids of the bone, such as framboidal pyrite grains, fungalfruiting bodies, soil and plant material (Jans, 2005). Soluble inclusionscan infiltrate the bone matrix and can cause staining of the bone;brown to dark red stains are often caused by humics, orange stains bymetal ions (Fe2+), while pink stains can result from fungal action(Piepenbrink, 1986; Jans et al., 2002). A cracking index was obtained,following Jans et al. (2002): the number of cracked versus intact osteonswas assessed in five microscopic fields with magnification 160 and re-ported as a percentage. As large cracks may be post-excavation arte-facts, these were excluded from the cracking index (Jans et al., 2002).The degree of birefringence, which is related to the orientation of theapatite crystals and/or the intactness of collagen fibrils (Schoeningeret al., 1989; Jans, 2005), was assessed and reported as 1 (perfect), 0.5(reduced) or 0 (absent) (Jans, 2005). Generalised destruction, referringto the loss of original microstructure due to mineral dissolution, was
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
also assessed. The bone appears ‘blank’ and lacks characteristic featuressuch as osteocyte lacunae, canaliculi and lamellar bone structure(Garland, 1993). The General Histological Index (GHI) (Hollund et al.,2012), ranging from 5 to 0, assesses the proportion of microstructuredestroyed by staining and mineral dissolution as well as bioerosion.
3.2. Stable isotope analysis
Collagen extraction was based on the Longin (1971) method, whichis similar to the protocol described by Brock et al. (2010). The bone sur-face was mechanically cleaned, after which the bone was crushed intosmaller pieces. An acid solution of 0.6 M hydrochloric acid (HCl) (aq.)was added to 200 mg bone to dissolve the mineral component in thebone at 4 ºC for 24 h. The acidwas renewed several times to ensure com-plete mineral dissolution. The dissolved mineral was discarded and theacid insoluble fraction was rinsed three times with Milli-Q water for pHneutralisation. An acid solution of pH 3 HCl (aq.) was added to the boneto dissolve the collagen at 80 ºC for 24h. The dissolved collagenwas sep-arated from the undissolved particles using 60–90 μm polyethyleneEzee-filters™ (Elkay Laboratory Product, UK). After freezing andlyophilisation, 1–1.05 mg of the collagen was sampled for the carbonand nitrogen stable isotope measurements, which were performedusing an Elemental Analyzer (NC2500, ThermoQuest Italia, Rodana,Italy) connected to a continuous-flow IRMS (Isotope Ratio Mass Spec-trometer) (Delta Plus, ThermoQuest Finnigan, Bremen, Germany) atthe department of Systems Ecology, faculty Earth- and Life Sciences,VU University Amsterdam, the Netherlands. The machine has a preci-sion of 0.15‰ for carbon and nitrogen stable isotope measurements.To ensure high quality results, collagen yields should be higher than1% (van Klinken, 1999), atomic C/N ratios should be between 2.9 and3.6 (DeNiro, 1985) and mass percentage of collagen should consist for15.3–47% of carbon and for 5.5–17.3% of nitrogen (Ambrose, 1990).
3.3. ZooMS analysis
ZooMS collagenfingerprinting analyseswere carried out on 29of the30 samples of bone powder leftover from stable isotope analysis follow-ingmodifiedmethods of Buckley et al. (2009) inwhich acid-soluble col-lagen was also utilised due to the limited amount of bone powderavailable. Approximately 5 mg bone powder was demineralised with0.5mL 0.6 M HCl (Fisher Scientific, UK) for 18 h at 4 °C and then centri-fuged for 5 min at 14,000 × g. The supernatant, containing acid-solublecollagen, was applied to a 10 kDa molecular weight cut-off ultrafilter(Vivaspin, UK) and spun at 14,000 × g for 90 min. The acid-insolublepellet was then incubated with ammonium bicarbonate buffer(pH 7.4; ABC; Sigma-Aldrich, UK) at 75 °C for 3 h prior to centrifugationfor 1 min. The ultrafilter flow-through for each sample was thendiscarded and 0.5 mL ABC was added and centrifuged for another90 min. The supernatant from the acid-insoluble fraction incubated inABC was then added to the ultrafilter and centrifuged as before. Finally,the filtrate was resuspended with 0.2 mL ABC and digested with 0.4 μgtrypsin (Promega, UK) for 18 h at 37 °C.
Following digestion, the protein fragments (peptides) were thenacidified to 0.1% trifluoroacetic acid (TFA; Signa-Aldrich, UK) andthen further purified and fractionated using C18 pipette tips(OMIX) into a 0–10% acetonitrile (ACN) fraction and a 10–50% ACNfraction following Buckley et al. (2009). These fractions were drieddown and resuspended with 10 μL 0.1% TFA and then 1 μL wasco-crystallised with 1 μL α-cyano-4-hydroxycinnamic acid (Sigma-Aldrich, UK) solution (10 mg/mL in 50% ACN) onto a Bruker stainlesssteel matrix-assisted laser desorption-ionization (MALDI) targetplate. MALDI analysis was carried out using a Bruker Ultraflex II, inwhich 2000 laser acquisitions were accumulated over a m/z range of700–3700. Spectra were inspected manually using Bruker DaltonicsflexAnalysis 3.3 software.
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003
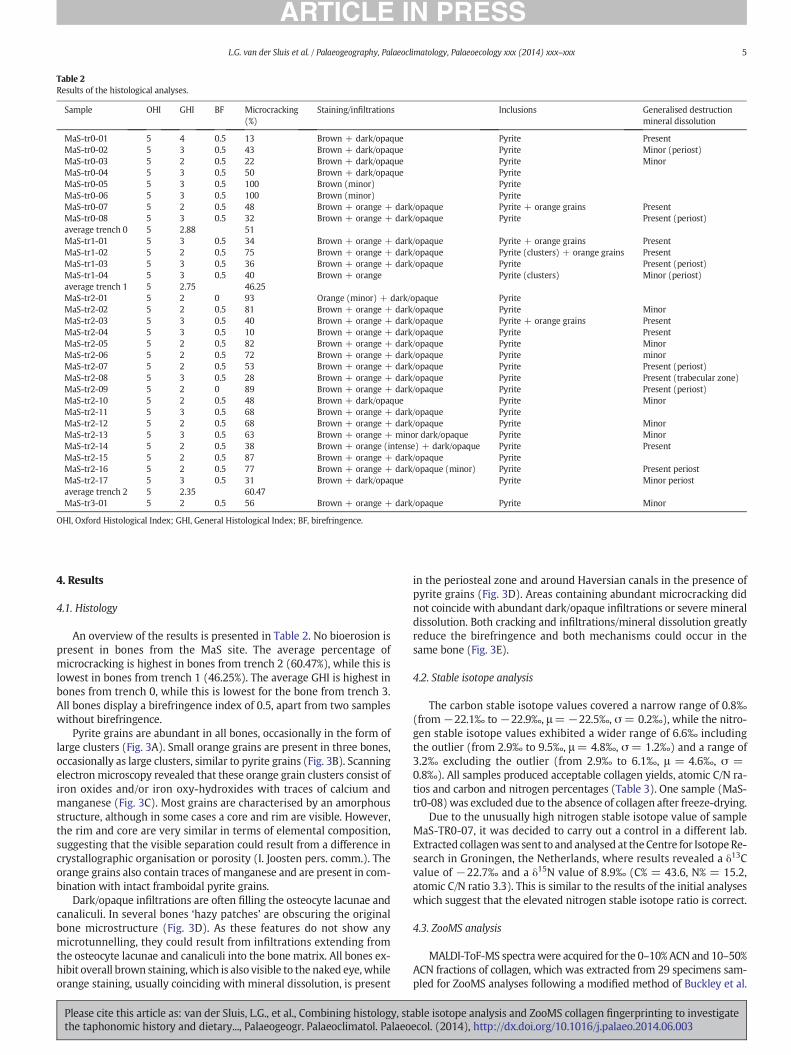
Table 2Results of the histological analyses.
Sample OHI GHI BF Microcracking(%)
Staining/infiltrations Inclusions Generalised destructionmineral dissolution
MaS-tr0-01 5 4 0.5 13 Brown + dark/opaque Pyrite PresentMaS-tr0-02 5 3 0.5 43 Brown + dark/opaque Pyrite Minor (periost)MaS-tr0-03 5 2 0.5 22 Brown + dark/opaque Pyrite MinorMaS-tr0-04 5 3 0.5 50 Brown + dark/opaque PyriteMaS-tr0-05 5 3 0.5 100 Brown (minor) PyriteMaS-tr0-06 5 3 0.5 100 Brown (minor) PyriteMaS-tr0-07 5 2 0.5 48 Brown + orange + dark/opaque Pyrite + orange grains PresentMaS-tr0-08 5 3 0.5 32 Brown + orange + dark/opaque Pyrite Present (periost)average trench 0 5 2.88 51MaS-tr1-01 5 3 0.5 34 Brown + orange + dark/opaque Pyrite + orange grains PresentMaS-tr1-02 5 2 0.5 75 Brown + orange + dark/opaque Pyrite (clusters) + orange grains PresentMaS-tr1-03 5 3 0.5 36 Brown + orange + dark/opaque Pyrite Present (periost)MaS-tr1-04 5 3 0.5 40 Brown + orange Pyrite (clusters) Minor (periost)average trench 1 5 2.75 46.25MaS-tr2-01 5 2 0 93 Orange (minor) + dark/opaque PyriteMaS-tr2-02 5 2 0.5 81 Brown + orange + dark/opaque Pyrite MinorMaS-tr2-03 5 3 0.5 40 Brown + orange + dark/opaque Pyrite + orange grains PresentMaS-tr2-04 5 3 0.5 10 Brown + orange + dark/opaque Pyrite PresentMaS-tr2-05 5 2 0.5 82 Brown + orange + dark/opaque Pyrite MinorMaS-tr2-06 5 2 0.5 72 Brown + orange + dark/opaque Pyrite minorMaS-tr2-07 5 2 0.5 53 Brown + orange + dark/opaque Pyrite Present (periost)MaS-tr2-08 5 3 0.5 28 Brown + orange + dark/opaque Pyrite Present (trabecular zone)MaS-tr2-09 5 2 0 89 Brown + orange + dark/opaque Pyrite Present (periost)MaS-tr2-10 5 2 0.5 48 Brown + dark/opaque Pyrite MinorMaS-tr2-11 5 3 0.5 68 Brown + orange + dark/opaque PyriteMaS-tr2-12 5 2 0.5 68 Brown + orange + dark/opaque Pyrite MinorMaS-tr2-13 5 3 0.5 63 Brown + orange + minor dark/opaque Pyrite MinorMaS-tr2-14 5 2 0.5 38 Brown + orange (intense) + dark/opaque Pyrite PresentMaS-tr2-15 5 2 0.5 87 Brown + orange + dark/opaque PyriteMaS-tr2-16 5 2 0.5 77 Brown + orange + dark/opaque (minor) Pyrite Present periostMaS-tr2-17 5 3 0.5 31 Brown + dark/opaque Pyrite Minor periostaverage trench 2 5 2.35 60.47MaS-tr3-01 5 2 0.5 56 Brown + orange + dark/opaque Pyrite Minor
OHI, Oxford Histological Index; GHI, General Histological Index; BF, birefringence.
5L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
4. Results
4.1. Histology
An overview of the results is presented in Table 2. No bioerosion ispresent in bones from the MaS site. The average percentage ofmicrocracking is highest in bones from trench 2 (60.47%), while this islowest in bones from trench 1 (46.25%). The average GHI is highest inbones from trench 0, while this is lowest for the bone from trench 3.All bones display a birefringence index of 0.5, apart from two sampleswithout birefringence.
Pyrite grains are abundant in all bones, occasionally in the form oflarge clusters (Fig. 3A). Small orange grains are present in three bones,occasionally as large clusters, similar to pyrite grains (Fig. 3B). Scanningelectron microscopy revealed that these orange grain clusters consist ofiron oxides and/or iron oxy-hydroxides with traces of calcium andmanganese (Fig. 3C). Most grains are characterised by an amorphousstructure, although in some cases a core and rim are visible. However,the rim and core are very similar in terms of elemental composition,suggesting that the visible separation could result from a difference incrystallographic organisation or porosity (I. Joosten pers. comm.). Theorange grains also contain traces of manganese and are present in com-bination with intact framboidal pyrite grains.
Dark/opaque infiltrations are often filling the osteocyte lacunae andcanaliculi. In several bones ‘hazy patches’ are obscuring the originalbone microstructure (Fig. 3D). As these features do not show anymicrotunnelling, they could result from infiltrations extending fromthe osteocyte lacunae and canaliculi into the bone matrix. All bones ex-hibit overall brown staining,which is also visible to the naked eye,whileorange staining, usually coinciding with mineral dissolution, is present
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
in the periosteal zone and around Haversian canals in the presence ofpyrite grains (Fig. 3D). Areas containing abundant microcracking didnot coincide with abundant dark/opaque infiltrations or severe mineraldissolution. Both cracking and infiltrations/mineral dissolution greatlyreduce the birefringence and both mechanisms could occur in thesame bone (Fig. 3E).
4.2. Stable isotope analysis
The carbon stable isotope values covered a narrow range of 0.8‰(from −22.1‰ to −22.9‰, μ = −22.5‰, σ = 0.2‰), while the nitro-gen stable isotope values exhibited a wider range of 6.6‰ includingthe outlier (from 2.9‰ to 9.5‰, μ = 4.8‰, σ = 1.2‰) and a range of3.2‰ excluding the outlier (from 2.9‰ to 6.1‰, μ = 4.6‰, σ =0.8‰). All samples produced acceptable collagen yields, atomic C/N ra-tios and carbon and nitrogen percentages (Table 3). One sample (MaS-tr0-08) was excluded due to the absence of collagen after freeze-drying.
Due to the unusually high nitrogen stable isotope value of sampleMaS-TR0-07, it was decided to carry out a control in a different lab.Extracted collagenwas sent to and analysed at theCentre for IsotopeRe-search in Groningen, the Netherlands, where results revealed a δ13Cvalue of −22.7‰ and a δ15N value of 8.9‰ (C% = 43.6, N% = 15.2,atomic C/N ratio 3.3). This is similar to the results of the initial analyseswhich suggest that the elevated nitrogen stable isotope ratio is correct.
4.3. ZooMS analysis
MALDI-ToF-MS spectrawere acquired for the 0–10%ACN and 10–50%ACN fractions of collagen, which was extracted from 29 specimens sam-pled for ZooMS analyses following a modified method of Buckley et al.
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

Fig. 3. (A) MaS-tr1-04 pyrite cluster in the trabecular bone (×40). (B) MaS-tr2-03 orange grain cluster (×250). (C) Elemental spot analysis MaS-tr0-07 on framboidal pyrite (1) and acluster of orange grains (2). Accelerating voltage: 15.0 kV Magnification: 3000. (D) MaS-tr1-01 mineral dissolution and pyrite grains in the Haversian canals (×100). Some ‘hazy patches’are visible in the bonematrix (arrow). (E)MaS-tr1-02microcracking is visible in themidsection of the bone,whilemineral dissolution and orange staining prevail in the periosteal zone (×40).
6 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
(2009). All peaks (representative of collagen tryptic peptides) observedin the 10–50% ACN fractions were also observed in the 0–10% ACN frac-tions, indicating that the fractionation developed as amethod to separatesomemammals, such as sheep from goats (Buckley et al., 2010) ormam-moths from mastodons (Buckley et al., 2011), may not be necessary forthe analysis of non-mammalian remains. As expected, most of the‘giant tortoise’ collagen fingerprints yielded very similar peakm/z values(Fig. 4). However, a small number of samples displayed clear differencesbetween them. The most notable of these differences being the presenceof a series of peaks (labelled C in Fig. 4 inset; likely from a single peptidewith +16 hydroxylation variants) at m/z 2827, 2843 and 2859 inmost samples (n = 21) being replaced by a series of peaks at m/z2839, 2855 and 2871 in some (n = 8). Another series of discrimina-tory peaks was also observed atm/z 2783 and 2799 (n=21) andm/z2825 and 2841 (labelled B) as well as at m/z 1261 (n = 21) and m/z1209 (n = 8) (labelled A).
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
Of the 30 bones, only 8 had intact proximal ends, 5 were missingminor parts, while the remainder had broken ends. Two types ofmorphology, observed in bones with intact femoral proximal ends,corresponded to the two groups that emerged from the ZooMS results.One type displayed two protrusions, producing an s-shaped proximalend, whereas the other type had one protrusion, resulting in a relativelystraight proximal end (Fig. 5). Thesemorphological differences are sim-ilar to those described byGünther (1877), who assigned femurswith ans-shaped proximal end to tortoises with a saddleback-shaped carapaceand femurs with a straight proximal end to tortoises with a dome-shaped carapace (Günther, 1877; J.P. Hume pers. comm.). The fewbones with intact femoral proximal ends allowed us to connect femurmorphology to carapace morphology and ZooMS spectra. As a result,the ZooMS spectra could be used to further separate between C. ineptaand C. triserrata species in morphologically non-diagnostic bone frag-ments as well. The smaller sample set in this dataset (n = 8) can be
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

Table 3Results of the carbon and nitrogen stable isotope and ZooMS analyses.
Samples δ15N (‰) δ13C (‰) %N %C C/N ratio Collagen yield (%) Biomarkers:1209 (A), 2825/2841 (B/B+16),2839/2855/2871 (C/C+16/C+32)
Biomarkers:1261 (A), 2783/2799 (B/B+16),2827/2843/2859 (C/C+16/C+32)
Species
MaS-tr0-01 4.1 −22.2 16.0 44.0 3.2 12.6 x C. triserrataMaS-tr0-02 4.5 −22.8 15.8 43.4 3.2 11.9 x C. triserrataMaS-tr0-03 4.2 −22.4 15.8 44.0 3.3 13.7 x C. triserrataMaS-tr0-04 3.1 −22.2 16.0 44.5 3.2 13.5 x C. triserrataMaS-tr0-05 2.9 −22.3 16.0 44.6 3.3 13.9 x C. triserrataMaS-tr0-06 3.8 −22.3 15.1 41.6 3.2 12.8 x C. triserrataMaS-tr0-07 9.5 −22.4 15.6 43.1 3.2 6.8 x C. triserrataMaS-tr0-07a 8.9 −22.7 15.2 43.6 3.3MaS-tr0-08b – – – – – – x C. triserrataMaS-tr1-01 5.0 −22.8 15.2 42.1 3.2 11.9 x C. triserrataMaS-tr1-02 4.0 −22.4 15.4 43.1 3.3 10.1 x C. triserrataMaS-tr1-03 5.2 −22.6 15.4 42.9 3.3 10.8 x C. ineptaMaS-tr1-04 4.2 −22.6 15.2 42.5 3.3 13.1 x C. triserrataMaS-tr2-01 5.0 −22.5 15.5 44.0 3.3 12.8 x C. ineptaMaS-tr2-02 5.8 −22.3 15.2 43.3 3.3 13.3 x C. ineptaMaS-tr2-03 4.0 −22.8 15.3 41.9 3.2 12.6 x C. triserrataMaS-tr2-04 5.5 −22.5 15.2 43.3 3.3 7.6 x C. ineptaMaS-tr2-05 4.2 −22.7 15.4 43.3 3.3 12.8 x C. triserrataMaS-tr2-06 4.3 −22.7 15.9 45.0 3.3 15.1 x C. triserrataMaS-tr2-07 6.1 −22.9 15.0 44.1 3.4 4.9 x C. ineptaMaS-tr2-08c 4.2 −22.5 15.2 43.2 3.3 9.8 -MaS-tr2-09 4.8 −22.3 15.0 43.1 3.4 14.7 x C. triserrataMaS-tr2-10 5.2 −22.3 15.7 44.0 3.3 12.0 x C. triserrataMaS-tr2-11 5.1 −22.8 15.4 43.8 3.3 13.3 x C. triserrataMaS-tr2-12 3.8 −22.9 15.6 44.3 3.3 14.0 x C. triserrataMaS-tr2-13 5.8 −22.6 15.5 44.6 3.4 14.9 x C. ineptaMaS-tr2-14 5.5 −22.4 15.7 44.9 3.3 14.8 x C. ineptaMaS-tr2-15 4.0 −22.1 15.7 44.5 3.3 14.4 x C. triserrataMaS-tr2-16 4.5 −22.3 15.2 42.3 3.2 10.3 x C. triserrataMaS-tr2-17 4.1 −22.2 15.3 43.1 3.3 7.4 x C. ineptaMaS-tr3-01 5.5 −22.6 16.1 45.0 3.3 16.0 x C. triserrata
a Control analysis of sample MaS-tr0-07 carried out at the Centre for Isotope Research in Groningen.b Collagen extraction of sample MaS-tr0-08 failed.c Not enough bone powder was available from sample MaS-Tr2-08 for ZooMS analysis.
Fig. 4.MALDI-ToF-MS spectra of the 0–10% ACN fraction from collagen extracted from samplesMAS-TR2-04 andMAS-TR1-01 highlighting the two distinct fingerprints obtained through-out the sample set. *Cylindraspis inepta (top) is characterised by dominant markers at m/z 1209 (A), 2825/2841 (B/B+16) and 2839/2855/2871 (C/C+16/C+32), while Cylindraspistriserrata produced distinctive markers at m/z 1261 (A), 2783/2799 (B/B+16) and 2827/2843/2859 (C/C+16/C+32); in the absence of sequencing, the likely homology betweensome of these markers (particularly B and C) is supported by the similar mass shifts of +16 observed, likely representing homologous peptides with additional hydroxylations, a post-translational modification that commonly occurs in collagen.
7L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stable isotope analysis and ZooMS collagen fingerprinting to investigatethe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeoecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

Fig. 5. Two types of morphology observed in femoral proximal ends. Femurs on the leftdisplay an s-shaped proximal end (C. triserrata), while the femurs on the right arecharacterised by a relatively straight proximal end (C. inepta).
8 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
identified as the dome-shaped species, C. inepta, while the larger group(n=21) represents the saddleback-shaped species, C. triserrata (Fig. 5)(Table 3).
5. Discussion
5.1. Taphonomy and bone preservation
All MaS bones revealed similar characteristics, although some weremore pronounced in certain trenches. Bones from trench 0 revealedthe least amount of orange staining and mineral dissolution comparedto bones from other trenches, while pyrite clusters were only presentin bones from trench 1. All bones contained framboidal pyrite grains,the presence of which is closely connected to the metabolism ofsulphate reducing bacteria. Sulphate reducing bacteria oxidise simple or-ganic compounds from organic matter in anaerobic circumstances, whileproducing dissolved sulphides (Berner et al., 1985; Turner-Walker,1998). These sulphides can subsequently react with iron (soluble orpresent inminerals) to precipitate in a range of different sulphide phasesand eventually as pyrite (Berner et al., 1985). Elevated concentrations ofFe2+ in the site's groundwater are providing the iron source, whereasdecomposing organic matter covering or occupying the spaces insidebone can act as a rich sulphur source for pyrite formation in non-marine environments (Canfield and Raiswell, 1991; Briggs et al., 1996;Pfretzschner, 2001; Leduc, 2012), which could have been the case forbones at the MaS site. Rapid burial in an anoxic environment ensuredlarge amounts of highly reactive organic matter were available for thesulphate reducing bacteria (Berner et al., 1985), resulting in abundantframboidal pyrites.
However, exposure to atmospheric oxygen at high relative humidi-ties can initiate pyrite oxidation, leading to the release of acid in the di-rect bone environment and mobilisation of iron (Canfield and Raiswell,1991). This small scale acid production can subsequently demineralisethe surrounding bone, while themobile iron ions stain the bone orange(Turner-Walker, 1999; Jans, 2005; Hollund et al., 2012). Ferric iron can
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
also precipitate as iron oxide hydroxides (Nicholson et al., 1988), whichexplains the presence of orange grains in some bones. Similar grainsfound in a fossil elephant bone from West Runton (Turner-Walker,1999) and in bones from Castricum (Hollund et al., 2012) have beeninterpreted as oxidised pyrite due to their low sulphur and high ironcontent. However, a similar sphere found in an experimental bonestudy in Taiwan was interpreted as a precursor of pyrite due to theabsence of framboidal pyrites and the lack of time for oxidation totake place (Turner-Walker, 2012). The presence of framboidal pyrites,combined with orange staining in the MaS bones, is, however,supporting the initial explanation. Pyrite oxidation could be responsiblefor the localised mineral dissolution and loss of microstructure at peri-osteal zones and someHaversian canals in theMaS bones. This suggeststhe pyrite grains were in contact with oxygen at some point. This prob-ably relates to hydrological changes as theMaS site's basins are still con-nected to the ocean and influenced by tidal and sea level changes(Rijsdijk et al., 2009). Staining caused by infiltrated metal ions orhumic acids has a bactericidal effect, thus inhibiting bioerosion and im-proving the histological preservation (Jans, 2005; Aufderheide, 2011).An initially anoxic environment combined with infiltrations is thenmost likely responsible for the absence of bioerosional features in theMaS bones.
The non-biological decay observed is probably linked to an activehydrology in the upper layers of the site. Microcracking in bones fromtemperate environments (i.e. high latitudes) can result from seasonalfreezing-thawing cycles (Jans, 2005), of which the equivalent in tropicalregions may consist of seasonal wetting-drying cycles. All MaS bonesprobably experienced alternating oxic and anoxic periods resultingfrom hydrological changes, during which the bones at the top layercould have experienced more intense stress, also caused by pyriteoxidation and precipitation of iron oxides. Thus, bones displayingmicrocracking were possibly located somewhere in the top section ofthe MaS site, while bones characterised by more intense staining andgreater numbers of pyrite clusters are likely to originate from lowerlayers.
The site's unique location in the landscape combined with afavourable hydrology provided exceptional conditions for bone preser-vation. The MaS site is characterised by a benign soil, in which the earlytaphonomic factors determined the bones' state of preservation(Nielsen-Marsh et al., 2007). We can assume the tortoises entered thesoil archive as complete bodies due to the absence of gnawing andweathering marks, while rapid burial sealed the bones anoxically,resulting in the absence of microbial attack and abundant pyrite forma-tion. Thus, the MaS bones are well equilibrated with their burial envi-ronment and could remain this way for many years, assuming noradical changes occur in the hydrology or pH of the site. The lack ofmicrobial alteration is reassuring, although the destructive effect ofoxidising pyrite grains at high relative humidities and oxic conditionsmust be kept inmind. Continued cycles of wetting and dryingwill resultin greater damages, in terms of increased extent of microcracking, min-eral dissolution and infiltrations.
5.2. Giant tortoise ecology and dietary behaviour
Comparative stable isotopic data is limited (Fig. 6), as few studies ontortoise ecology were based on both carbon and nitrogen stableisotopes. Burleigh and Arnold (1986) analysed carbon stable isotopesof several giant tortoise species. Two of their samples were probablyoriginating from the Mare aux Songes. Bone collagen from the fe-murs yielded δ13C values of −23.8‰ and −25.6‰ for C. inepta andC. triserrata respectively. Bone collagen derived from the carapaceof two tortoises from Rodrigues, the second nearest island, displayedδ13C values of −22.5‰ and −22.9‰. These values correspond wellwith our carbon stable isotope results, which ranged from −22.1‰to −22.9‰. Giant tortoise species from Madagascar, Rodrigues,Aldabra and Assumption Island produced carbon stable isotope ratios
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

9L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
ranging from−25.8‰ to−13.3‰. Apparently, tortoise δ13C values fromMauritius and Rodrigues reveal a terrestrial C3 diet, while tortoises ofmorewestern origin reveal elevated carbon stable isotope ratios, indicat-ing C4 and/or CAM plant consumption (Burleigh and Arnold, 1986).
Investigation of floral remains from the MaS site revealed this areawas covered with marsh, wet forest and dry forest (Rijsdijk et al.,2009). It is unclear if these tropical plant species had a C3 or C4 photo-synthetic pathway, although seeds from the Sideroxylon grandiflorumand an unidentified piece of wood revealed δ13C values of −28.1‰and −27.7‰ respectively, suggesting a C3 pathway. Additionally,carbon and nitrogen stable isotope analysis performed on two dodoand three tortoise bones revealed a herbivorous terrestrial C3 diet(Rijsdijk et al., 2009), similar to our results (Fig. 7), which means C3
plants were present 4200 years ago on Mauritius. A factor that can in-fluence δ13C ratios is density of forest vegetation, due to the so-calledcanopy effect (van der Merwe and Medina, 1989, 1991; Vogel et al.,1990). In densely vegetated areas, plants growing underneath the can-opy can use CO2 that has already been fractionated, resulting in lowerδ13C values compared to plants (or leaves) in the forest canopy or plantsin more open environments (van Klinken et al., 2000). The tortoisespresent very uniform δ13C values, although the slight variation of 0.8‰could be induced by the canopy effect.
Tortoise bones (n=4) from two sites near Cape Town, South Africa,revealed δ13C values ranging from−21.0‰ to−22.2‰ and δ15N valuesconsisting of 2.9‰, 2.5‰, 8.8‰ and 10.3‰ (Corr et al., 2005) (Fig. 6).Two of these elevated nitrogen stable isotope values are similar tothe outlier in this dataset (9.52‰). Although tortoises are known tooccasionally scavenge on bone (Hume, 2005), it seems unlikely that oc-casional bone scavenging results in a nitrogen stable isotope enrich-ment of 6‰. However, other factors than protein consumption areknown to increase the δ15N values. Several authors found a relationshipbetween precipitation, temperature and elevated δ15N values in soils,plants and terrestrial animals (Heaton, 1986; Heaton, 1987; Sealyet al., 1987; Ambrose, 1991; Gröcke et al., 1997; Handley et al., 1999;Amundson et al., 2003; Aranibar et al., 2004; Hollund et al., 2010;Ugan and Coltrain, 2012). Furthermore, drought tolerant species areable to influence their amino acid δ15N values by changing theirmetabolic processes (Styring et al., 2010). Tortoises are able to ex-crete nitrogen as both uric acid and urea, which can help them retainwater in periods of drought. The extreme drought conditions pre-vailing in Mauritius 4200 years ago (De Boer et al., 2014) may havebeen involved inmodifying themetabolic processes leading to nitrogenstable isotope enrichment. Thiswas observed by Styring et al. (2010) bystudying stable isotope ratios in amino acids (i.e. compound specific
Fig. 6. Carbon and nitrogen stable isotope data from the literature: Madagascar (n=10), Rodri(Burleigh and Arnold, 1986), dodo (n = 2) and Cylindraspis (n = 3) (Rijsdijk et al., 2009), and
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
analysis) and currently appears to be the most likely explanation forthe extremely elevated nitrogen stable isotope ratio in this dataset.
The ZooMS analyses allowed successful species identification of all ofthe analysed bones. This is a significant result. Although collagen finger-prints have recently been shown to be species-specific in a few mam-malian groups (e.g., camels (Rybczynski et al., 2013) and some whales(e.g., Balaenoptera, see Buckley et al., 2014)), this represents thefirst in-stance of species-specificity in non-mammalian vertebrates. This alsoallows investigation of variation in stable isotope values and dietarybehaviour for the two tortoise species. Apart from the C. triserrataindividual outlier, the extent of variation in δ13C and δ15N values issimilar for the two species. In fact, the average for the carbon stableisotope values is the same. The two species largely overlap in terms ofnitrogen stable isotope ratios, although some C. triserrata individualsdisplay lower δ15N values and the average δ15N value is 0.9‰ lowerthan C. inepta. This is not a great difference, but a two tailed T-testshowed that the two groups differ significantly in their nitrogen stableisotope values (including the outlier p = 0.034, α = 0.05, two tailedT-test with unequal variances, excluding the outlier p = 0.0001,α = 0.05, two tailed T-test with unequal variances). Variation innitrogen stable isotope values may be caused by consumption ofdifferent plant parts, as δ15N values are known to vary anatomically inplants, e.g. between leaves, stems, seeds and roots (Yoneyama et al.,1997; Codron et al., 2005; Pechenkina et al., 2005; Pardo et al., 2013).However, Codron et al. (2005) also report that δ15N values can vary con-siderably, up to 4‰, betweenmicrohabitats due to environmental influ-ences. Due to their different carapace morphologies, tortoises perhapsdid not share their direct habitats, but could have been more dispersedbetween such microhabitats, consisting of adjacent areas alternatingwith densely covered and open vegetation. Alternatively, the dome-shaped tortoise (C. inepta) possibly consumedmore plants near ground,while the saddleback-shaped tortoise (C. triserrata) possibly preferredhigher growing plants on bushes and trees (Arnold, 1979). This wouldallow the two species to share their habitat without interfering witheach other's dietary preference. If carapace morphology is related tothe degree of openness of the vegetation (Arnold, 1979; Burleighand Arnold, 1986), this may also be a factor influencing the observedinter-species differences.
The high δ15N value of the outlier is most likely resulting from met-abolic drought adjustments, which could similarly have affected othertortoises, though to a much lesser extent, as their δ15N ratios are bestexplained in terms of tortoise species. This dichotomy in the dataset issimilar to what Corr et al. (2005) observed in their tortoise specimens.Still, interpreting δ15N values in these extinct tortoises remains
gues (n=2), Mare aux Songes C. inepta (n=2), C. triserrata (n=2) and Aldabra (n=5)South Africa (n = 4) (Corr et al., 2005).
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003
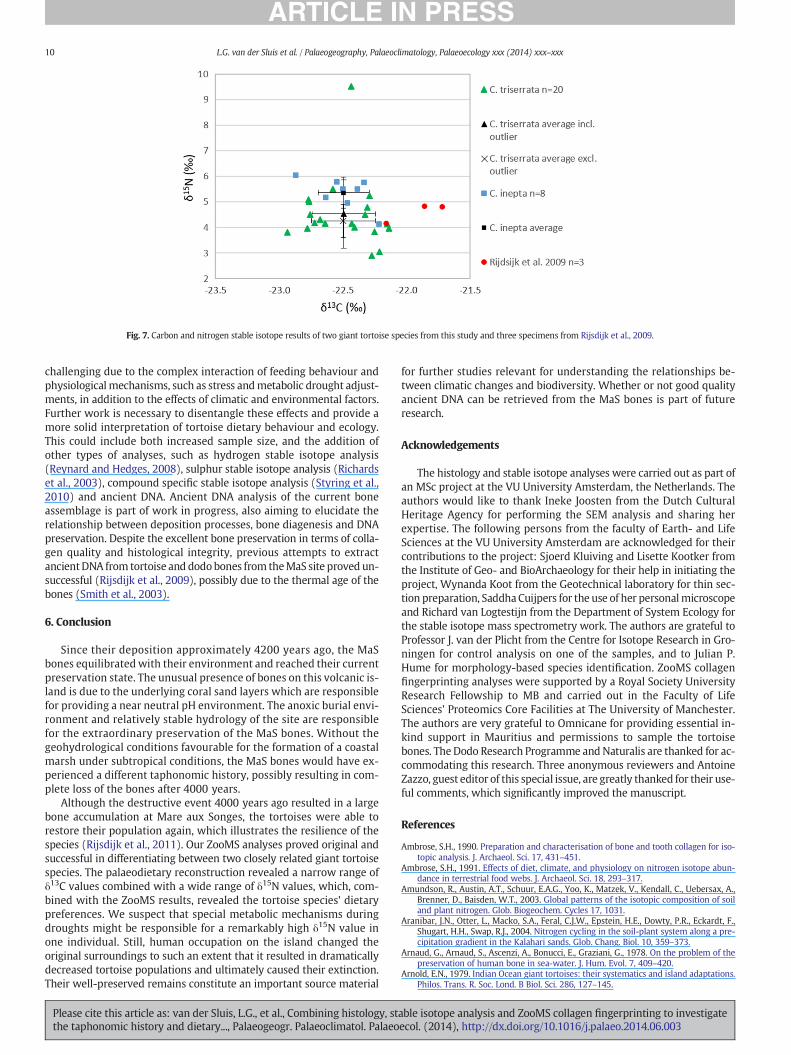
Fig. 7. Carbon and nitrogen stable isotope results of two giant tortoise species from this study and three specimens from Rijsdijk et al., 2009.
10 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
challenging due to the complex interaction of feeding behaviour andphysiologicalmechanisms, such as stress andmetabolic drought adjust-ments, in addition to the effects of climatic and environmental factors.Further work is necessary to disentangle these effects and provide amore solid interpretation of tortoise dietary behaviour and ecology.This could include both increased sample size, and the addition ofother types of analyses, such as hydrogen stable isotope analysis(Reynard and Hedges, 2008), sulphur stable isotope analysis (Richardset al., 2003), compound specific stable isotope analysis (Styring et al.,2010) and ancient DNA. Ancient DNA analysis of the current boneassemblage is part of work in progress, also aiming to elucidate therelationship between deposition processes, bone diagenesis and DNApreservation. Despite the excellent bone preservation in terms of colla-gen quality and histological integrity, previous attempts to extractancientDNA from tortoise and dodobones from theMaS site proved un-successful (Rijsdijk et al., 2009), possibly due to the thermal age of thebones (Smith et al., 2003).
6. Conclusion
Since their deposition approximately 4200 years ago, the MaSbones equilibratedwith their environment and reached their currentpreservation state. The unusual presence of bones on this volcanic is-land is due to the underlying coral sand layers which are responsiblefor providing a near neutral pH environment. The anoxic burial envi-ronment and relatively stable hydrology of the site are responsiblefor the extraordinary preservation of the MaS bones. Without thegeohydrological conditions favourable for the formation of a coastalmarsh under subtropical conditions, the MaS bones would have ex-perienced a different taphonomic history, possibly resulting in com-plete loss of the bones after 4000 years.
Although the destructive event 4000 years ago resulted in a largebone accumulation at Mare aux Songes, the tortoises were able torestore their population again, which illustrates the resilience of thespecies (Rijsdijk et al., 2011). Our ZooMS analyses proved original andsuccessful in differentiating between two closely related giant tortoisespecies. The palaeodietary reconstruction revealed a narrow range ofδ13C values combined with a wide range of δ15N values, which, com-bined with the ZooMS results, revealed the tortoise species' dietarypreferences. We suspect that special metabolic mechanisms duringdroughts might be responsible for a remarkably high δ15N value inone individual. Still, human occupation on the island changed theoriginal surroundings to such an extent that it resulted in dramaticallydecreased tortoise populations and ultimately caused their extinction.Their well-preserved remains constitute an important source material
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
for further studies relevant for understanding the relationships be-tween climatic changes and biodiversity. Whether or not good qualityancient DNA can be retrieved from the MaS bones is part of futureresearch.
Acknowledgements
The histology and stable isotope analyses were carried out as part ofan MSc project at the VU University Amsterdam, the Netherlands. Theauthors would like to thank Ineke Joosten from the Dutch CulturalHeritage Agency for performing the SEM analysis and sharing herexpertise. The following persons from the faculty of Earth- and LifeSciences at the VU University Amsterdam are acknowledged for theircontributions to the project: Sjoerd Kluiving and Lisette Kootker fromthe Institute of Geo- and BioArchaeology for their help in initiating theproject, Wynanda Koot from the Geotechnical laboratory for thin sec-tion preparation, SaddhaCuijpers for the use of her personalmicroscopeand Richard van Logtestijn from the Department of System Ecology forthe stable isotope mass spectrometry work. The authors are grateful toProfessor J. van der Plicht from the Centre for Isotope Research in Gro-ningen for control analysis on one of the samples, and to Julian P.Hume for morphology-based species identification. ZooMS collagenfingerprinting analyses were supported by a Royal Society UniversityResearch Fellowship to MB and carried out in the Faculty of LifeSciences' Proteomics Core Facilities at The University of Manchester.The authors are very grateful to Omnicane for providing essential in-kind support in Mauritius and permissions to sample the tortoisebones. TheDodo Research Programme andNaturalis are thanked for ac-commodating this research. Three anonymous reviewers and AntoineZazzo, guest editor of this special issue, are greatly thanked for their use-ful comments, which significantly improved the manuscript.
References
Ambrose, S.H., 1990. Preparation and characterisation of bone and tooth collagen for iso-topic analysis. J. Archaeol. Sci. 17, 431–451.
Ambrose, S.H., 1991. Effects of diet, climate, and physiology on nitrogen isotope abun-dance in terrestrial food webs. J. Archaeol. Sci. 18, 293–317.
Amundson, R., Austin, A.T., Schuur, E.A.G., Yoo, K., Matzek, V., Kendall, C., Uebersax, A.,Brenner, D., Baisden, W.T., 2003. Global patterns of the isotopic composition of soiland plant nitrogen. Glob. Biogeochem. Cycles 17, 1031.
Aranibar, J.N., Otter, L., Macko, S.A., Feral, C.J.W., Epstein, H.E., Dowty, P.R., Eckardt, F.,Shugart, H.H., Swap, R.J., 2004. Nitrogen cycling in the soil-plant system along a pre-cipitation gradient in the Kalahari sands. Glob. Chang. Biol. 10, 359–373.
Arnaud, G., Arnaud, S., Ascenzi, A., Bonucci, E., Graziani, G., 1978. On the problem of thepreservation of human bone in sea-water. J. Hum. Evol. 7, 409–420.
Arnold, E.N., 1979. Indian Ocean giant tortoises: their systematics and island adaptations.Philos. Trans. R. Soc. Lond. B Biol. Sci. 286, 127–145.
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

11L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
Aufderheide, A.C., 2011. Soft tissue taphonomy: a paleopathology perspective. Int. J.Paleopathol. 1, 75–80.
Austin, J.J., Arnold, E.N., 2001. Ancient mitochondrial DNA and morphology elucidate anextinct island radiation of Indian Ocean giant tortoises (Cylindraspis). R. Soc.London B Ser. Biol. Sci. 268, 2515–2523.
Behrensmeyer, A.K., Kidwell, S.M., 1985. Taphonomy's contributions to paleobiology.Paleobiology 11, 105–119.
Bell, L.S., 1990. Palaeopathology and diagenesis: an SEM evaluation of structural changesusing backscattered electron imaging. J. Archaeol. Sci. 17, 85–102.
Bell, L.S., Skinner, M.F., Jones, S.J., 1996. The speed of post mortem change to the humanskeleton and its taphonomic significance. Forensic Sci. Int. 82, 129–140.
Berner, R.A., De Leeuw, J.W., Spiro, B., Murchison, D.G., Eglinton, G., 1985. Sulphate reduc-tion, organic matter decomposition and pyrite formation [and discussion]. Philos.Trans. R. Soc. London Ser. A Math. Phys. Sci. 315, 25–38.
Briggs, D.E.G., Raiswell, R., Bottrel, S.H., Hatfield, D., Bartels, C., 1996. Controls on thepyritization of exceptionally preserved fossils: an analysis of the Lower DevonianHunsbrück Slate of Germany. Am. J. Sci. 296, 633–663.
Brock, F., Higham, T., Ditchfield, P., Bronk Ramsey, C., 2010. Current pre-treatmentmethods for AMS radiocarbon dating at the Oxford radiocarbon accelerator unit(ORAU). Radiocarbon 52, 103–112.
Buckley, M., Collins, M., Thomas-Oates, J., Wilson, J.C., 2009. Species identificationby analysis of bone collagen using matrix-assisted laser description/ionisationtime-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 23,3843–3854.
Buckley, M., Whitcher Kansa, S., Howard, S., Campbell, S., Thomas-Oates, J., Collins, M.,2010. Distinguishing between archaeological sheep and goat bones using a singlecollagen peptide. J. Archaeol. Sci. 37, 13–20.
Buckley, M., Larkin, N., Collins, M., 2011. Mammoth and Mastodon collagen sequences;survival and utility. Geochim. Cosmochim. Acta 75, 2007–2016.
Buckley, M., Fraser, S., Herman, Jeremy.S., Melton, N.D., Mulville, J., Pálsdóttir, A.H., 2014.Species identification of archaeological marine mammals using collagen fingerprint-ing. J. Archaeol. Sci. 41, 631–641.
Burleigh, R., Arnold, E.N., 1986. Age and dietary differences of recently extinct IndianOcean tortoises (Geochelone s. lat.) revealed by carbon isotope analysis. Proc. R.Soc. Lond. B Ser. Biol. Sci. 227, 137–144.
Canfield, D.E., Raiswell, R., 1991. Pyrite formation and fossil preservation. In: Allison, P.A.,Briggs, D.E. (Eds.), Taphonomy. Plenum Press, New York, pp. 337–387.
Cheke, A., 1987. An ecological history of the Mascarene Islands, with special refer-ence to extinctions and introductions of land vertebrates. In: Diamond, A.W.(Ed.), Studies of Mascarene Island Birds. Cambridge University Press, Cam-bridge, pp. 5–89.
Child, A.M., 1995. Towards an understanding of the microbial decomposition of archaeo-logical bone in the burial environment. J. Archaeol. Sci. 22, 165–174.
Clark, G., 1866. Account of the late discovery of Dodo remains in the island of Mauritius.Ibis 2, 141–146.
Codron, J., Codron, D., Lee-Thorp, J.A., Sponheimer, M., Bond, W.J., de Ruiter, D., Grant, R.,2005. Taxonomic, anatomical, and spatio-temporal variations in the stable carbonand nitrogen isotopic compositions of plants from an African savanna. J. Archaeol.Sci. 32, 1757–1772.
Corr, L.T., Sealy, J.C., Horton, M.C., Evershed, R.P., 2005. A novel marine dietary indicatorutilising compound-specific bone collagen amino acid δ13C values of ancient humans.J. Archaeol. Sci. 32, 321–330.
Davis, P.G., 1997. The bioerosion of bird bones. Int. J. Osteoarchaeol. 7, 388–401.De Boer, E.J., Tjallingii, R., Vélez, M.I., Rijsdijk, K.F., Vlug, A., Reichart, G.J., Prendergast, A.L.,
De Louw, P.G.B., Florens, F.B., Baider, C., Hooghiemstra, H., 2014. Climate variability inthe SW Indian Ocean from an 8000-yr long multi-proxy record in the Mauritian low-lands shows a middle to late Holocene shift from negative IOD-state to ENSO-state.Quat. Sci. Rev. 86, 175–189.
DeNiro, M.J., 1985. Postmortem preservation and alteration of in vivo bonecollagen isotope ratios in relation to palaeodietary reconstruction. Nature 317,806–809.
DeNiro, M.J., Epstein, S., 1978. Influence of diet on the distribution of carbon isotopes inanimals. Geochim. Cosmochim. Acta 42, 495–506.
Garland, A.N., 1987. A histological study of archaeological bone decomposition. In:Boddington, A., Garland, A.N., Janaway, R.C. (Eds.), Death, Decay and reconstruction.Manchester University Press, Manchester, pp. 109–126.
Garland, A.N., 1993. An introduction to the histology of exhumed mineralized tissue. In:Grupe, G., Garland, A.N. (Eds.), Histology of Ancient Human Bone: Methods and Diag-nosis. Springer-Verlag, New York, pp. 1–16.
Gröcke, D.R., Bocherens, H., Mariotti, A., 1997. Annual rainfall and nitrogen-isotope corre-lation in macropod collagen: application as a palaeoprecipitation indicator. EarthPlanet. Sci. Lett. 153, 279–285.
Günther, A., 1877. The gigantic land-tortoises (living and extinct) in the collection of theBritish Museum. British Museum, London (iv + 96 p., pls. I-LIV.).
Hackett, C.J., 1981. Microscopical focal destruction (tunnels) in exhumed human bones.Med. Sci. Law 21, 243–265.
Handley, L.L., Austin, A.T., Robinson, D., Scrimgeour, C.M., Raven, J.A., Heaton, T.H.E.,Schmidt, S., Stewart, G.R., 1999. The 15 N natural abundance (d15N) of ecosystemsamples reflects measures of water availability. Aust. J. Plant Physiol. 26,185–199.
Heaton, T.H.E., 1986. Climatic influence of the isotopic composition of bone nitrogen.Nature 322, 822–823.
Heaton, T.H.E., 1987. The 15 N/14N ratios of plants in South Africa and Namibia: relation-ship to climate and coastal/saline environments. Oecologia 74, 236–246.
Hedges, R.E.M., Reynard, L.M., 2007. Nitrogen isotopes and the trophic level of humans inarchaeology. J. Archaeol. Sci. 34, 1240–1251.
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
Hedges, R.E.M., Millard, A.R., Pike, A.W.G., 1995. Measurements and relationships ofdiagenetic alteration of bone from three archaeological sites. J. Archaeol. Sci. 22,201–209.
Hollund, H.I., Higham, T., Belinskij, A., Korenevskij, S., 2010. Investigation ofpalaeodiet in the North Caucasus (South Russia) Bronze Age using stable iso-tope analysis and AMS dating of human and animal bones. J. Archaeol. Sci. 37,2971–2983.
Hollund, H.I., Jans, M.M.E., Collins, M.J., Kars, H., Joosten, I., Kars, S.M., 2012. What hap-pened here? Bone histology as a tool in decoding the postmortem histories of archae-ological bone from Castricum, The Netherlands. Int. J. Osteoarchaeol. 22, 537–548.http://dx.doi.org/10.1002/oa.1273.
Hume, J.P., 2005. Contrasting taphofacies in ocean island settings: the fossil record ofMascarene vertebrates. Proceedings of the International Symposium “Insular Verte-brate Evolution: the Paleontological Approach”. Mon Soc d'Hist Nat Bal 12, 129–144.
Hume, J.P., Prys-Jones, R.P., 2005. New discoveries from old sources with reference to theoriginal bird andmammal fauna of theMascarene Islands, Indian Ocean. Zoöl. Meded.Leiden 97, 85–95.
Jans, M.M.E., 2005. Histological Characterisation of Diagenetic Alteration of ArchaeologicalBone. (PhD Thesis) Institute for Geo- and BioArchaeology, Vrije Universiteit Amsterdam.
Jans, M.M.E., Kars, H., Nielsen-Marsh, C.M., Smith, C.I., Nord, A.G., Arthur, P., Earl, N., 2002.In situ preservation of archaeological bone. A histological study within a multidisci-plinary approach. Archaeometry 44, 343–352.
Jans, M.M.E., Nielsen-Marsh, C.M., Smith, C.I., Collins, M.J., Kars, H., 2004. Characterisationof microbial attack on archaeological bone. J. Archaeol. Sci. 31, 87–95.
Koch, P.L., 1998. Isotopic reconstruction of past continental environments. Annu. Rev.Earth Planet. Sci. 26, 573–613.
Leduc, T., 2012. Diagenesis of the fossil bones of Iguanodon bernissartensis from the Iguan-odon sinkhole. In: Godefroit, P. (Ed.), Bernissart Dinosaurs and early Cretaceous ter-restrial ecosystems. Indiana University Press, Indiana, pp. 113–136.
Longin, R., 1971. New method of collagen extraction for radiocarbon dating. Nature 230,241–242.
Marchant, R., Hooghiemstra, H., 2004. Rapid environmental change in African and SouthAmerican tropics around 4000 years before present: a review. Earth Sci. Rev. 66,217–260.
Martill, D.M., 1989. Fungal borings in neoselacian teeth from the Lower Oxford Clay ofPeterborough. Mercian Geol. 12, 1–4.
Meijer, H.J.M., Gill, A., de Louw, P.G.B., van den Hoek Ostende, L.W., Hume, J.P., Rijsdijk, K.F., 2012. Dodo remains from an in situ context from Mare aux Songes, Mauritius.Naturwissenschaft 99, 177–184.
Nicholson, R.V., Gillham, R.W., Reardon, E.J., 1988. Pyrite oxidation in carbonate-buffered solution: 1. Experimental kinetics. Geochim. Cosmochim. Acta 52,1077–1085.
Nielsen-Marsh, C.M., Smith, C.I., Nord, A., Kars, H., Collins, M.J., 2007. Bone diagen-esis in the European Holocene II: taphonomic and environmental consider-ations. J. Archaeol. Sci. 34, 1523–1531.
Pardo, L.H., Semaoune, P., Schaberg, P.G., Eagar, C., Sebilo, M., 2013. Patterns in δ15N inroots, stems and leaves of sugar maple and American beech seedlinds, saplings, andmature trees. Biogeochemistry 112, 275–291.
Pechenkina, E.A., Ambrose, S.H., Xiaolin, M., Benfer Jr., R.A., 2005. Reconstructing northernChinese Neolithic subsistence practices by isotopic analysis. J. Archaeol. Sci. 32,1176–1189.
Pfretzschner, H.-U., 2001. Pyrite in fossil bone. Neues Jb. Geol. Paläontol. Abh. 220, 1–23.Piepenbrink, H., 1986. Two examples of biogenous dead bone decomposition and their
consequences for taphonomic interpretation. J. Archaeol. Sci. 13, 417–430.Reynard, L.M., Hedges, R.E.M., 2008. Stable hydrogen isotopes of bone collagen
in palaeodietary and palaeoenvironmental reconstruction. J. Archaeol. Sci. 35,1934–1942.
Richards, M.P., Fuller, B.T., Sponheimer, M., Robinson, T., Ayliffe, L., 2003. Sulphur isotopein palaeodietary studies: a review and results from a controlled feeding experiment.Int. J. Osteoarchaeol. 13, 37–45.
Rijsdijk, K.F., Hume, J.P., Bunnik, F., Florens, F.B.V., Baider, C., Shapiro, B., van der Plicht, J.,Janoo, A., Griffiths, O., van den Hoek Ostende, L.W., Cremer, H., Vernimmen, T., deLouw, P.G.B., Bholah, A., Saumtally, A., Porch, N., Haile, J., Buckley, M., Collins, M.,Gittenberger, E., 2009. Mid-Holocene vertebrate bone concentration-Lagerstätte onoceanic island Mauritius provides a window into the ecosystem of the dodo (Raphuscucullatus). Quat. Sci. Rev. 28, 14–24.
Rijsdijk, K.F., Zinke, J., de Louw, P.G.B., Hume, J.P., van der Plicht, H.J., Hooghiemstra,H., Meijer, H.J.M., Vonhof, H.B., Porch, N., Florens, F.B.V., Baider, C., van Geel, B.,Brinkkemper, J., Vernimmen, T., Janoo, A., 2011. Mid-Holocene (4200 kyr BP)mass mortalities in Mauritius (Mascarenes): insular vertebrates resilient to cli-matic extremes but vulnerable to human impact. The Holocene 21, 1179–1194.
Rybczynski, N., Gosse, J.C., Harington, C.R., Wogelius, R.A., Hidy, A.J., Buckley, M., 2013.Mid-Pliocene warm-period deposits in the High Arctic yield insight into camel evolu-tion. Nat. Commun. 4, 1550. http://dx.doi.org/10.1038/ncomms2516.
Schoeller, D.A., 1999. Isotope fractionation: why aren't we what we eat? J. Archaeol. Sci.26, 667–673.
Schoeninger, M.J., Moore, K.M., Murray, M.L., Kingston, J.D., 1989. Detection of bone pres-ervation in archaeological and fossil samples. Appl. Geochem. 4, 281–292.
Sealy, J., van der Merwe, N., Lee-Thorpe, J., Lanham, J., 1987. Nitrogen isotope ecology insouthern Africa: implications for environmental and dietary tracing. Geochim.Cosmochim. Acta 51, 2707–2717.
Smith, C.I., Chamberlain, A.T., Riley, M.S., Stringer, C., Collins, M.J., 2003. The thermal his-tory of human fossils and the likelihood of successful DNA amplification. J. Hum. Evol.45, 203–217.
Staub, F., 1996. Dodos and Solitaires, myth and reality. Proc. R. Soc. Arts Sci. Mauritius 6,89–122.
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003

12 L.G. van der Sluis et al. / Palaeogeography, Palaeoclimatology, Palaeoecology xxx (2014) xxx–xxx
Styring, A.K., Sealy, J.C., Evershed, R.P., 2010. Resolving the bulk δ15N values of ancienthuman and animal bone collagen via compound-specific nitrogen isotope analysisof constituent amino acids. Geochim. Cosmochim. Acta 74, 241–251.
Turner-Walker, G., 1998. The West Runton fossil elephant: a pre-conservation evalu-ation of its condition, chemistry and burial environment. The Conservator 22,26–35.
Turner-Walker, G., 1999. Pyrite and bone diagenesis in terrestrial sediments: evidencefrom the West Runton freshwater bed. Bull. Geol. Soc. Norfolk 48, 3–26.
Turner-Walker, G., 2012. Early bioerosion in skeletal tissues: persistence through deeptime. N. Jb. Geol. Paläont. 265, 165–183.
Turner-Walker, G., Jans, M.M.E., 2008. Reconstructing taphonomic histories using histo-logical analysis. Palaeogeogr. Palaeoclimatol. Palaeoecol. 266, 227–235.
Ugan, A., Coltrain, J., 2012. Variation in collagen stable nitrogen values in black-tailed jack-rabbits (Lepus californicus) in relation to small-scale differences in climate, soil andtopography. J. Archaeol. Sci. 38, 1417–1429.
van der Merwe, N.J., Medina, E., 1989. Photosynthesis and 13C/12C ratios in Amazonianrain forests. Geochim. Cosmochim. Acta 53, 1091–1094.
van der Merwe, N.J., Medina, E., 1991. The canopy effect, carbon isotope ratios andfoodwebs in Amazonia. J. Archaeol. Sci. 18, 249–259.
Please cite this article as: van der Sluis, L.G., et al., Combining histology, stthe taphonomic history and dietary..., Palaeogeogr. Palaeoclimatol. Palaeo
van Klinken, G.J., 1999. Bone collagen quality indicators for palaeodietary and radiocarbonmeasurements. J. Archaeol. Sci. 26, 687–695.
van Klinken, G.J., Richards, M.P., Hedges, R.E.M., 2000. An overview of causes for stableisotopic variations in past European human populations: environmental, ecophysio-logical, and cultural effects. In: Ambrose, S., Katzenberg, M.A. (Eds.), BiogeochemicalApproaches to Palaeodietary Analysis. Kluwer Academic/Plenum Publishers NewYork.
Virginia, R.A., Delwiche, C.C., 1982. Natural 15 N abundance of presumed N2-fixing andnon-N2-fixing plants from selected ecosystems. Oecologia 54, 317–325.
Vogel, J.C., Eglinton, B., Auret, J.M., 1990. Isotopic fingerprints in elephant bone and ivory.Nature 346, 747–749.
Willaime, P., 1984. Carte Pedologique de l´Ile Maurice 1/50.000. Office de la RechercheScientifique et Technique Outre/Mer, France. Mauritius Sugar Industry ResearchInstitute.
Yoneyama, T., Handley, L.L., Scrimgeour, C.M., Fisher, D.B., Raven, J.A., 1997. Variations ofthe natural abundances of nitrogen and carbon isotopes in Triticum aestivum, withspecial reference to phloem and xylem exudates. New Phytol. 137, 205–213.
able isotope analysis and ZooMS collagen fingerprinting to investigateecol. (2014), http://dx.doi.org/10.1016/j.palaeo.2014.06.003




















