
Coccolith Sr/Ca ratios in the eastern Mediterranean: Production versus export processes
11
Coccolith Sr/Ca ratios in the eastern Mediterranean: Production versus export processes L. Auliaherliaty a, ⁎, H.M. Stoll b,c , P. Ziveri a,d , E. Malinverno e , M. Triantaphyllou f , S. Stravrakakis g , V. Lykousis g a Section Marine Biogeology, Faculty of Earth and Life Sciences, VU University Amsterdam, De Boelelaan 1085, 1081 HV Amsterdam, The Netherlands b Department of Geoscience, Williams College, Williamstown, MA 01267, USA c Department of Geology, University of Oviedo, Arias de Velasco, s/n 30005, Oviedo, Asturias, Spain d ICTA, Universitat Autònoma de Barcelona (UAB), Edifici Cn-Campus UAB, 08193 Bellaterra, Spain e Department of Geological Sciences and Geotechnologies, University of Milano-Bicocca, Milano, Italy f Department of Historical Geology–Paleontology, Faculty of Geology, University of Athens, Panepistimiopolis 15784, Athens, Greece g Hellenic Centre for Marine Research, Institute of Oceanography, 19013 Anavyssos, Greece abstract article info Article history: Received 6 March 2009 Received in revised form 18 September 2009 Accepted 1 October 2009 Keywords: Mediterranean Sea Coccolith Sr/Ca ratio Sediment traps Productivity Export processes Samples collected by two sediment traps located southwest of Crete in the eastern Mediterranean (EMED) [48A (1953 m) and 48B (950 m)] from June 2005 to May 2006 were used to study fluxes of organic carbon, carbonate and coccolithophores in combination with the variations of Sr/Ca ratios in different individually picked coccolith species. Considering the complexity of the EMED, we validate the use of Sr/Ca ratios as productivity proxy and unravel the varied processes which may influence it. We examined the relationship between the seasonal peaks in export fluxes and the Sr/Ca ratio in coccoliths of three upper photic zone coccolithophores species collected in the traps, Calcidiscus leptoporus, Helicosphaera carteri and Emiliania huxleyi. We aimed at testing whether high export fluxes are correlated with high Sr/Ca ratios, suggestive of higher nutrient-stimulated production, or Sr/Ca ratios are unchanged during high export periods, suggestive of increased export efficiency or scavenging. Periods of enhanced trap fluxes in March and June result from surface water blooms recognized in satellite imagery. An additional peak flux was found in January, but this peak represents re-suspended or recycled material in the water column. The amplitude of seasonal variations in the Sr/Ca ratios of the three investigated species is small in both traps. In the shallow trap, a decrease in the Sr/Ca ratio of C. leptoporus occurred synchronously with minimal fluxes. The other two species were not measured for this period. In the deep trap, no such decrease in Sr/Ca was observed during minimal fluxes, in either C. leptoporus or H. carteri, probably due to a long residence of coccoliths in the water column, recycling and low export efficiency. Absolute Sr/Ca ratios for all species are lower than in other more productive environments like the Bay of Bengal, Arabian Sea, or Sargasso Sea. We conclude that Sr/Ca ratios in coccoliths of surface sediments in the EMED reflect mainly spring–summer bloom conditions averaged over hundreds to thousands of years. In addition, the origin of varying calcite thickness in H. carteri was investigated. The similarity of average Sr/ Ca ratios in differently-calcified specimens confirms that coccolith thickness variations in this species result from primary biomineralization processes and not from variable overgrowth by (low Sr) abiogenic calcite in the water column or the sediments. © 2009 Elsevier B.V. All rights reserved. 1. Introduction The production of coccolithophores, like that of many other phytoplankton groups, is seasonally variable and stimulated by optimum nutrient concentrations and light. However, different coccolithophore species are adapted to different light conditions, so that also deep species exist which can live at low light levels. Coccolithophores have an excellent continuous fossil record and are commonly used for paleo-environmental reconstructions. Recently the Sr/Ca ratio of coccolith calcite has been suggested as a proxy of algal growth rate (Stoll and Schrag, 2000; Stoll et al., 2007a,b). For example, in the high productivity region of the Arabian Sea, (Stoll et al., 2007a) showed that high coccolith Sr/Ca ratios were well correlated to high coccolithophore fluxes in sediment traps, demon- strating that in this setting the export processes are strictly linked to surface production. This positive correlation has been shown also in the Sargasso Sea during the fall bloom period (Stoll et al., 2007a). Marine Micropaleontology 73 (2009) 196–206 ⁎ Corresponding author. Tel.: +31 20 5987412; fax: +31 20 5989941. E-mail address: [email protected] (L. Auliaherliaty). 0377-8398/$ – see front matter © 2009 Elsevier B.V. All rights reserved. doi:10.1016/j.marmicro.2009.10.001 Contents lists available at ScienceDirect Marine Micropaleontology journal homepage: www.elsevier.com/locate/marmicro
Transcript of Coccolith Sr/Ca ratios in the eastern Mediterranean: Production versus export processes

Marine Micropaleontology 73 (2009) 196ndash206
Contents lists available at ScienceDirect
Marine Micropaleontology
j ourna l homepage wwwe lsev ie rcom locate marmicro
Coccolith SrCa ratios in the eastern Mediterranean Production versusexport processes
L Auliaherliaty a HM Stoll bc P Ziveri ad E Malinverno e M Triantaphyllou fS Stravrakakis g V Lykousis g
a Section Marine Biogeology Faculty of Earth and Life Sciences VU University Amsterdam De Boelelaan 1085 1081 HV Amsterdam The Netherlandsb Department of Geoscience Williams College Williamstown MA 01267 USAc Department of Geology University of Oviedo Arias de Velasco sn 30005 Oviedo Asturias Spaind ICTA Universitat Autogravenoma de Barcelona (UAB) Edifici Cn-Campus UAB 08193 Bellaterra Spaine Department of Geological Sciences and Geotechnologies University of Milano-Bicocca Milano Italyf Department of Historical GeologyndashPaleontology Faculty of Geology University of Athens Panepistimiopolis 15784 Athens Greeceg Hellenic Centre for Marine Research Institute of Oceanography 19013 Anavyssos Greece
Corresponding author Tel +31 20 5987412 fax +E-mail address lay500falwvunl (L Auliaherliaty)
0377-8398$ ndash see front matter copy 2009 Elsevier BV Adoi101016jmarmicro200910001
a b s t r a c t
a r t i c l e i n f o
Article historyReceived 6 March 2009Received in revised form 18 September 2009Accepted 1 October 2009
KeywordsMediterranean SeaCoccolithSrCa ratioSediment trapsProductivityExport processes
Samples collected by two sediment traps located southwest of Crete in the eastern Mediterranean (EMED)[48A (1953 m) and 48B (950 m)] from June 2005 to May 2006 were used to study fluxes of organic carboncarbonate and coccolithophores in combination with the variations of SrCa ratios in different individuallypicked coccolith species Considering the complexity of the EMED we validate the use of SrCa ratios asproductivity proxy and unravel the varied processes which may influence it We examined the relationshipbetween the seasonal peaks in export fluxes and the SrCa ratio in coccoliths of three upper photic zonecoccolithophores species collected in the traps Calcidiscus leptoporus Helicosphaera carteri and Emilianiahuxleyi We aimed at testing whether high export fluxes are correlated with high SrCa ratios suggestive ofhigher nutrient-stimulated production or SrCa ratios are unchanged during high export periods suggestiveof increased export efficiency or scavenging Periods of enhanced trap fluxes in March and June result fromsurface water blooms recognized in satellite imagery An additional peak flux was found in January but thispeak represents re-suspended or recycled material in the water columnThe amplitude of seasonal variations in the SrCa ratios of the three investigated species is small in bothtraps In the shallow trap a decrease in the SrCa ratio of C leptoporus occurred synchronously with minimalfluxes The other two species were not measured for this period In the deep trap no such decrease in SrCawas observed during minimal fluxes in either C leptoporus or H carteri probably due to a long residence ofcoccoliths in the water column recycling and low export efficiency Absolute SrCa ratios for all species arelower than in other more productive environments like the Bay of Bengal Arabian Sea or Sargasso Sea Weconclude that SrCa ratios in coccoliths of surface sediments in the EMED reflect mainly springndashsummerbloom conditions averaged over hundreds to thousands of yearsIn addition the origin of varying calcite thickness in H carteri was investigated The similarity of average SrCa ratios in differently-calcified specimens confirms that coccolith thickness variations in this species resultfrom primary biomineralization processes and not from variable overgrowth by (low Sr) abiogenic calcite inthe water column or the sediments
31 20 5989941
ll rights reserved
copy 2009 Elsevier BV All rights reserved
1 Introduction
The production of coccolithophores like that of many otherphytoplankton groups is seasonally variable and stimulated byoptimum nutrient concentrations and light However differentcoccolithophore species are adapted to different light conditions sothat also deep species exist which can live at low light levels
Coccolithophores have an excellent continuous fossil record and arecommonly used for paleo-environmental reconstructions Recentlythe SrCa ratio of coccolith calcite has been suggested as a proxy ofalgal growth rate (Stoll and Schrag 2000 Stoll et al 2007ab) Forexample in the high productivity region of the Arabian Sea (Stoll etal 2007a) showed that high coccolith SrCa ratios were wellcorrelated to high coccolithophore fluxes in sediment traps demon-strating that in this setting the export processes are strictly linked tosurface production This positive correlation has been shown also inthe Sargasso Sea during the fall bloom period (Stoll et al 2007a)
197L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Coccoliths (micrometric calcite plates produced by coccolitho-phore algae) are major contributors of the carbonate flux andsedimentation to the deep sea (Milliman 1993 Steinmetz 1994Chiu and Broecker 2008) Theymay act as mineral ballast which playsa role in the export of particulate organic matter to the deep ocean(Ziveri et al 2007) In the North Atlantic a positive correlationbetween daily coccolith carbonate mass fluxes of relatively heavyspecies ie Calcidiscus leptoporus and Helicosphaera carteri and dailyorganic carbon fluxes suggests their effectiveness in this exportprocess (Ziveri et al 2007)
In this study we focus on the eastern Mediterranean (EMED) andaim at assessing the relationship between seasonal surface productionas obtained through satellite data particle fluxes in the sediment trapsand SrCa ratios in coccoliths In the sediment trap flux data areavailable for both loose coccoliths and intact coccospheres (inter-connected coccoliths forming the cell cover) This is particularly usefulbecause in the living coccolithophores coccoliths are well connectedto cover the cell but after death the coccospheres are rapidlydisaggregated into individual coccoliths Therefore the presence ofintact coccospheres in sinking particle fluxes collected by sedimenttraps represents freshly delivered production from the photic zone(Honjo 1976 Honjo 1982 Ziveri et al 1995 Ziveri and Thunnel2000 Broerse et al 2000) During sinking the proportion ofcoccoliths over intact coccospheres increases as a consequence ofnatural coccosphere disruption grazing activity lateral transport andresuspension
By comparing coccolith SrCa ratios and coccosphere fluxes insediment trap samples we also aim at establishing whether thesurface production signals are transferred synchronously to the seafloor or are obscured by export processes during transport to the deepsea in this part of the EMED We also ascertain the link between SrCaratios of coccoliths in sediment traps and those in surface sedimentsThis will further enhance our knowledge on transport processesMoreover we compare the SrCa ratio results from the sediment trapsin EMED with those of other locations to evaluate whether the SrCaratio signal in EMED which is related to relatively small blooms iscomparable to SrCa signals in other parts of the ocean where largeblooms occur Therefore the validation of SrCa ratios as a productivityproxy can be established regarding the complexity of differentdisturbing processes ie oligotrophic condition low export andsecondary calcification Based on the fact that secondary calcifica-tion has been found in coccoliths of Emiliania huxleyi and Reticulo-fenestra spp from sediments of the EMED (Ziveri et al 2000 Crudeliet al 2004) and in the Red Sea (Winter 1982) we test here for
Fig 1 (a) Surface sediment samples distribution and hydrological circulations in the EMEDData View packages (Schlitzer 2008) The wind system and hydrological circulations are adIonian gyre 2ab The AtlanticndashIonian stream 2c The Atlantic Tunisian Current 2dMid-Med Jgyre 7 Cilician and Asia Minor current
coccoliths of H carteri whether secondary calcite precipitation isalso occurring in the water column since these types of calcite arereadily distinguished geochemically
2 Study area
The Mediterranean climate is characterized by mild wet wintersand warm dry summers The seasonal variability of the surface watermass properties and its large scale circulation are influenced by theexternal forcing particularly by wind stress Westerly winds prevailduring winter while stronger northerly winds prevail during latesummer over the EMED (Pinardi and Masetti 2000) as illustrated inFig 1
The physical circulation of the EMED leads to a ldquonutrient desertrdquosituation in which nutrient-rich deep waters are exported from thebasin and replaced by nutrient-poor Atlantic surface waters (Bethoux1979 Sarmiento et al 1988) The evaporative density increase in thebasin causes the sinking of surface water in the eastern part of thebasin The Levantine Intermediate Water (LIW) is formed by wintercooling of surface waters that sink to depths of 150ndash600 m(Ovchinnikov 1984 Malanotte-Rizzoli and Hecht 1988) Thisintermediate water spills throughout the basin westwards and duringthe cold winter period merges with the dense surface water along thenorthern margin of Mediterranean resulting in the formation of deepwater Western Mediterranean Deep Water (WMDW) is formed inthe Gulf of Lions Eastern Mediterranean Deep Water (EMDW) isformed in the Adriatic and Aegean Seas (POEM group 1992 Millot1999 Pinardi and Masetti 2000) Moreover this downwelling andsubsequent export of nutrient-rich intermediate water through theGibraltar Strait removes nutrients from the surface water andprevents their renewal in the photic zone producing the oligotrophicconditions of the modern Mediterranean (Pinet 1992 Krom et al2005)
The EMED including our studied region (South Cretan MarginCretan Sea and adjacent area) is characterized by phosphorus limitation(Krom et al 1991 Pinet 1992 Carbo et al 2005) This unusual Plimitation of the EMEDmaybecausedbyunequal supply ofNandP fromexternal sources (Herut et al 1999) by adsorption of phosphate byinorganic particles (Krom et al 1991) or by N2 fixation (Sachs andRepeta 1999) as also observed in the Central Pacific (Karl et al 1997)
Fig 1 shows seasonal eddies which occur in winter the cyclonicWestern Cretan Gyre and a minor anticyclonic Iera-Petra Gyre(Souvermezoglou et al 1999 Pinardi and Masetti 2000) A combina-tion between these seasonal eddies and the outflow from the Aegean
(b) sediment trap location with detailed bathymetric map is created through the Oceanapted from Pinardi and Masetti (2000) Detailed circulations are as follows 1 Westernet 3 Anticyclone in the Gulf of Syrte 4Western Cretan gyre 5 Iera-Petra gyre 6 Rhodes
Table 1Sediment trap site position water and trap depths sample selection for this study andrelated time interval as adapted from Malinverno et al (2009)
No Site Latitude Longitude Waterdepth(m)
Trapdepth(m)
Sub-samples Period ofsampling
1 48B 351455degN 237387degE 1967 995 48IB1a 1ndash15 Jun2005
2 48IB2 16ndash30Jun 2005
3 48IB4 16ndash31 Jul2005
4 48IB6 16ndash31Aug 2005
5 48IIB1 16 Octndash15 Nov2005
6 48IIB4b 1ndash15 Jan2005
7 48IIB9 16ndash31Mar 2006
8 48A 351455degN 237387degE 1967 1953 48IA3 1ndash15 Jul2005
9 48IA6 16ndash31Aug 2005
10 48IIA1 16 Octndash15 Nov2005
11 48IIA4 1ndash15 Jan2006
12 48IIA9 16ndash31Mar 2006
13 48IIA11c 15ndash30Apr 2006
48B is the shallow trap and 48A is the deep trapa Sample was also picked for individual H carterib Sample was only picked for H carteri populationc Sample was only picked for E huxleyi population
Table 2Surface sediment sample positions and water depth used for SrCa ratio analysis of Cleptoporus and H carteri
No Sample code Cruise Year Coordinates Waterdepth(m)
Longitude Latitude
1 M51-3 562 RV Meteor 2001 191910degE 327743degN 13902 M51-3 575 RV Meteor 2001 317862degE 345272degN 23373 M51-3 594 RV Meteor 2001 262185degE 379025degN 10054 M51-3 604 RV Meteor 2001 277883degE 408393degN 5525 Bio Deep 01 MCAT01a RV Urania 1997 213992degE 353012degN 32816 Bio Deep 03 AB12MCa RV Urania 1997 215103degE 352310degN 3310
a Sample was also picked for H carteri population
198 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Sea the so called ldquoTransition Mediterranean Waterrdquo (TMW) which isdefined as a ldquonutrient (phosphate) richndashoxygen poorrdquo intermediatewater mass (a layer between the LIW and Eastern Mediterranean DeepWater) can stimulate the productivity in the region in spring and fall(Souvermezoglou et al 1999)
In the EMED the main sources of fertilization originate from runoff from the surrounding land masses river inflow (mainly the Nile)by upwelling of the TMW and a combination between Saharan dustinput and precipitation (wet dust deposition) [Guerzoni et al 1999]which may be driven by two seasonal wind systems (Krom et al1991 2005 Carbo et al 2005 Eker-Develi et al 2006 Triantha-phyllou et al 2004 Ziveri et al 2000)
The maximum coccolithophore production is suggested to takeplace in late winter for the EMED (Kleijne 1991 Knappertsbusch1993) springndashsummer for the Cretan Sea and the Straits of Cretan Arc(Gotsis-Skretas et al 1999 Trianthaphyllou et al 2004) and in lateautumn and early spring for the coastal waters of Andros Island andthe Middle Aegean Sea (Dimiza et al 2008)
The high coccolithophore production in autumn as mentionedabove is incoherent with the results of Malinverno et al (2009) whichindicate the highest coccosphere flux occurring in spring and earlysummer Therefore by employing SrCa ratio of individual coccolithspecies we may unravel the timing of the highest coccosphereproduction and also elucidate whether the highest coccosphere fluxin spring is a real production signal or a remnant of greater exportefficiency due to aggregation promoted by diatoms and otherphytoplankton which dominated the winterndashspring blooms
3 Material and methods
Samples used in this studywere collected from two sediment trapsat one mooring station located off southwest Crete (35deg08732primeN23deg44322primeE) with a bottom depth of 1967 m (Fig 1) The depth ofdeployment of the two traps were at 995 m (48B) and 1953 m (48A)Sediment traps are PPS33 Technicap with a total of 12 collecting cupsand a collection area of 0125 m2 The 48B and 48A traps wereemployed from June 2005 until May 2006 with a sampling timeresolution of 15 to 16 days Detailed sampling processes samplepreservation sample preparation and coccolithophores speciescounting are explained by Stavrakakis et al (2000) and Malinvernoet al (2009) The biogeochemical and coccosphere export flux fromthese traps are from Malinverno et al (2009) Detailed sampleinformation used in this study is shown in Tables 1 and 2
SrCa measurements were carried out on individually pickedcoccoliths in order to obtain species-specific SrCa ratio time seriesand elucidate such a bias on SrCa ratio variation related to productivitywhichmay be given by different coccolith species (Stoll et al 2007a) Atotal of 10 samples (5 samples each from 48A and 48B) have beenselected for elemental chemistry using the new developed technique(Stoll et al 2007cd) of individually picked coccoliths We focused onthe surface dwellers C leptoporus andH carteri and the cosmopolitan Ehuxleyi (Fig 2) In order to assess whether the SrCa ratio signals fromthe EMED traps are similar to those from the seafloor sediments sixsurface sediment samples recovered from the EMEDwere used for SrCaratio analysis in the population of individually picked coccolithspecimens
Individual coccolith specimens were picked from a dry smear slideby a tungsten needle which is operated by a micromanipulator (waterrobot) under 400times magnification on an inverted polarizationmicroscope We picked 15ndash20 specimens for both ldquolargerrdquo speciesand 25ndash30 specimens for E huxleyi We isolated the specimens andthen placed them on thewell of a Tacky Dot slide The slidewas coatedby Epoxy (Buehler Epoxide) that was poured into an aluminium ringmounted on the slide Application of ethanol on the Tacky Dotremoved the epoxy mold from the slide The epoxy mold was thenpolished on an ethanol-wetted felt pad to remove the Tacky Dot
polymer coating Afterwards the mold was mounted on a CamecaIMS3f Secondary Ion Mass Spectrometer for SrCa ratio analysis Weshot a primary ion beam on a population of the individual coccolithsto obtain an average value of that particular species Data werecollected in three consecutive analyses of five-cycle scans of magneticfields covering 24 Mg 40 Ca and 88 Sr with a typical counting error of1 The standard deviation of collected data is 005 In addition wealso conducted profiles through single individual coccoliths of Hcarteri from the sediment traps which exhibited differing coccoliththickness and weight in order to observe the presence of secondarycalcification Detailed procedure of this analysis can be found in Stollet al (2007cd)
The time series (2004 until 2006) of surface water environmentparameters derived from SeaWiFS and MODIS-Aqua satellite datasets(httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) areshown in Fig 3 The surface water parameters are chlorophyll-aconcentration sea surface temperature (SST) and aerosol opticalthickness as indicator of aeolian dust The area coverage is 22degndash26degE
Fig 2 Scanning electron microscope pictures of the target species which were used for SrCa ratio analysis (a) C leptoporus (b) H carteri (c) E huxleyi
199L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
and 34degndash36degN The seasonal variation of surface production rangesfrom ~008 to ~022 mg mminus3 The maximum value of chlorophyll-aconcentration is recorded mostly in late winter until early spring andthe minimum value occurs in summer
In order to verify the species identification and the preservationstate of individual coccoliths Scanning Electron Microscope (SEM)analysis on each mounted stub was done at the SEM Laboratory VUUniversity Amsterdam The SrCa sample preparation and picking ofsingle coccoliths specimens were done at the Geology Department ofthe University of Oviedo Spain The Secondary IonMass Spectrometeranalysis of SrCa was conducted at the Northeast National IonMicroprobe Facility at Woods Hole Oceanographic Institution(WHOI) USA
4 Results
41 Shallow sediment trap 48B (995 m)
In the shallower sediment trap (48B) the total mass carbonateorganic carbon coccosphere and coccolith fluxes exhibit a strongseasonal pattern of particle fluxes (Fig 4ab Malinverno et al 2009) Afirst maximum in total mass carbonate organic carbon coccosphereand coccolith fluxes occurred between June and August 2005 Aminimum in fluxes occurred in autumn between September and earlyDecember During winter (December 2005ndashearly February 2006) totalmassflux including carbonate organic carbon and coccoliths increasedagain although the pronounced increase in coccosphere flux laggedbehind by several months and increased in March 2006 E huxleyi
Fig 3 Time series (December 2004ndashJune 2006) of chlorophyll-a concentration (OBPG SeaProducts for SST 11 microm day) and aerosol optical thickness (SeaWiFS Monthly Global 9-km
dominated both coccosphere and coccolith assemblages during thewhole investigated period andwasmost abundant during the intervalsof high coccosphere export fluxes in JunendashJuly 2005 andMarch 2006 Bycontrast during the high mass flux period in December 2005ndashearlyFebruary 2006 when coccosphere fluxes were not elevated thecoccosphere fluxes displayed a high abundance of deep-dwellingspecies in particular Algirosphaera robusta (Malinverno et al 2009)The fluxes of H carteri were most abundant during the periods of hightotal coccosphere flux Instead C leptoporus fluxes were always below1 of the total coccolithophore assemblage with slight increases duringthe total coccolith and coccosphere high flux intervals
The SrCa ratios of C leptoporus generally follow the seasonal trendin coccosphereexport in the shallower trap (Fig 5ab) HighC leptoporusSrCa ratios (299 mmolmol) accompanied the first peak in cocco-sphere and coccolith fluxes of this species in JunendashAugust Lower Cleptoporus SrCa (265 mmolmol) occurred in early autumn duringminima in fluxes In March 2006 an increase in coccosphere exportcorresponds toan increase in SrCa ratio (293 mmolmol)During JunendashJuly 2005 the peak of coccosphere and coccolith fluxes coincided withincreased SST and low values of the aerosol optical index andchlorophyll-a concentration (Fig 5h) In contrast the March 2006coccosphere peak lags by one month the peak values of chlorophyll-aconcentration and coincides with low SST
42 Deep sediment trap 48A (1953 m)
In the deeper sediment trap (48A) the total mass carbonate andorganic carbon show a slightly different seasonal pattern in fluxes
WiFS Monthly Global 9-km Products) SST (OBPG Modis-Aqua Monthly Global 9-kmProducts)
Fig 4 Total mass flux carbonate organic carbon coccosphere and coccolith fluxes from 48B (a b) and 48A (c d) according toMalinverno et al (2009) Periods A B C are indicated ingrey bars (see description in 51) Note timespans with no sample point and unconnected symbols indicate that no data are available
200 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
than the shallow trap although the pattern in coccosphere fluxes ismore similar (Fig 4cd Malinverno et al 2009) As in the shallowtrap there was a maximum in total mass carbonate organic carboncoccosphere and coccolith fluxes between June and July 2005however there was no spring 2006 peak in mass flux as observed inthe shallow trap Coccosphere fluxes in March 2006 in the deeper trapare significantly higher than those of winter 2005 From December2005 until February 2006 all fluxes show low values
H carteri SrCa ratios are not clearly related to total coccosphereflux patterns or coccosphere fluxes of H carteri (Fig 5cf) Minimumvalues occur in the early summer and early autumn (313 mmolmol)and maximum value (34 mmolmol) in January 2006 The peak in Hcarteri SrCa in January 2006 coincideswith a peak in total mass flux inthe shallow trap where the coccospheres of H carteri and Helico-sphaera spp also showed peak fluxes and increased relativeabundance
For the same trap periods there are differences in SrCa ratios in Cleptoporus in the deep and shallow sediment traps (Fig 5ce) Therange of SrCa ratios in the deeper trap (283 to 306 mmolmol) is
smaller than in the shallower trap (265 to 299 mmolmol) and noanalysis was performed for mid-October The early summer 2005value in the deeper trap is lower than the corresponding one in theshallow trap
The seasonal trend in E huxleyi SrCa ratios is similar to that seenin C leptoporus in the shallow trap E huxleyi SrCa ratio exhibitedmaximum values (195 to 201 mmolmol) during the early summer(June ndash July 2005) and spring (March 2006) periods of high fluxes of Ehuxleyi coccospheres and lower SrCa ratios (184 mmolmol) duringthe period of low fluxes (Fig 5dg) Overall E huxleyi SrCa ratios arelower than those of C leptoporus and H carteri
43 SrCa ratios along a profile of H carteri
We analysed SrCa ratios on individual coccoliths of H carteri(sample 48IB1 Table 1) having similar length but different carbonatemass (ldquothicknessrdquo or degree of calcification) The SrCa ratios of allspecimens (Fig 6) are within the analytical uncertainty for analysis ofa single coccolith (2 standard errors of the mean) No significant
Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

197L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Coccoliths (micrometric calcite plates produced by coccolitho-phore algae) are major contributors of the carbonate flux andsedimentation to the deep sea (Milliman 1993 Steinmetz 1994Chiu and Broecker 2008) Theymay act as mineral ballast which playsa role in the export of particulate organic matter to the deep ocean(Ziveri et al 2007) In the North Atlantic a positive correlationbetween daily coccolith carbonate mass fluxes of relatively heavyspecies ie Calcidiscus leptoporus and Helicosphaera carteri and dailyorganic carbon fluxes suggests their effectiveness in this exportprocess (Ziveri et al 2007)
In this study we focus on the eastern Mediterranean (EMED) andaim at assessing the relationship between seasonal surface productionas obtained through satellite data particle fluxes in the sediment trapsand SrCa ratios in coccoliths In the sediment trap flux data areavailable for both loose coccoliths and intact coccospheres (inter-connected coccoliths forming the cell cover) This is particularly usefulbecause in the living coccolithophores coccoliths are well connectedto cover the cell but after death the coccospheres are rapidlydisaggregated into individual coccoliths Therefore the presence ofintact coccospheres in sinking particle fluxes collected by sedimenttraps represents freshly delivered production from the photic zone(Honjo 1976 Honjo 1982 Ziveri et al 1995 Ziveri and Thunnel2000 Broerse et al 2000) During sinking the proportion ofcoccoliths over intact coccospheres increases as a consequence ofnatural coccosphere disruption grazing activity lateral transport andresuspension
By comparing coccolith SrCa ratios and coccosphere fluxes insediment trap samples we also aim at establishing whether thesurface production signals are transferred synchronously to the seafloor or are obscured by export processes during transport to the deepsea in this part of the EMED We also ascertain the link between SrCaratios of coccoliths in sediment traps and those in surface sedimentsThis will further enhance our knowledge on transport processesMoreover we compare the SrCa ratio results from the sediment trapsin EMED with those of other locations to evaluate whether the SrCaratio signal in EMED which is related to relatively small blooms iscomparable to SrCa signals in other parts of the ocean where largeblooms occur Therefore the validation of SrCa ratios as a productivityproxy can be established regarding the complexity of differentdisturbing processes ie oligotrophic condition low export andsecondary calcification Based on the fact that secondary calcifica-tion has been found in coccoliths of Emiliania huxleyi and Reticulo-fenestra spp from sediments of the EMED (Ziveri et al 2000 Crudeliet al 2004) and in the Red Sea (Winter 1982) we test here for
Fig 1 (a) Surface sediment samples distribution and hydrological circulations in the EMEDData View packages (Schlitzer 2008) The wind system and hydrological circulations are adIonian gyre 2ab The AtlanticndashIonian stream 2c The Atlantic Tunisian Current 2dMid-Med Jgyre 7 Cilician and Asia Minor current
coccoliths of H carteri whether secondary calcite precipitation isalso occurring in the water column since these types of calcite arereadily distinguished geochemically
2 Study area
The Mediterranean climate is characterized by mild wet wintersand warm dry summers The seasonal variability of the surface watermass properties and its large scale circulation are influenced by theexternal forcing particularly by wind stress Westerly winds prevailduring winter while stronger northerly winds prevail during latesummer over the EMED (Pinardi and Masetti 2000) as illustrated inFig 1
The physical circulation of the EMED leads to a ldquonutrient desertrdquosituation in which nutrient-rich deep waters are exported from thebasin and replaced by nutrient-poor Atlantic surface waters (Bethoux1979 Sarmiento et al 1988) The evaporative density increase in thebasin causes the sinking of surface water in the eastern part of thebasin The Levantine Intermediate Water (LIW) is formed by wintercooling of surface waters that sink to depths of 150ndash600 m(Ovchinnikov 1984 Malanotte-Rizzoli and Hecht 1988) Thisintermediate water spills throughout the basin westwards and duringthe cold winter period merges with the dense surface water along thenorthern margin of Mediterranean resulting in the formation of deepwater Western Mediterranean Deep Water (WMDW) is formed inthe Gulf of Lions Eastern Mediterranean Deep Water (EMDW) isformed in the Adriatic and Aegean Seas (POEM group 1992 Millot1999 Pinardi and Masetti 2000) Moreover this downwelling andsubsequent export of nutrient-rich intermediate water through theGibraltar Strait removes nutrients from the surface water andprevents their renewal in the photic zone producing the oligotrophicconditions of the modern Mediterranean (Pinet 1992 Krom et al2005)
The EMED including our studied region (South Cretan MarginCretan Sea and adjacent area) is characterized by phosphorus limitation(Krom et al 1991 Pinet 1992 Carbo et al 2005) This unusual Plimitation of the EMEDmaybecausedbyunequal supply ofNandP fromexternal sources (Herut et al 1999) by adsorption of phosphate byinorganic particles (Krom et al 1991) or by N2 fixation (Sachs andRepeta 1999) as also observed in the Central Pacific (Karl et al 1997)
Fig 1 shows seasonal eddies which occur in winter the cyclonicWestern Cretan Gyre and a minor anticyclonic Iera-Petra Gyre(Souvermezoglou et al 1999 Pinardi and Masetti 2000) A combina-tion between these seasonal eddies and the outflow from the Aegean
(b) sediment trap location with detailed bathymetric map is created through the Oceanapted from Pinardi and Masetti (2000) Detailed circulations are as follows 1 Westernet 3 Anticyclone in the Gulf of Syrte 4Western Cretan gyre 5 Iera-Petra gyre 6 Rhodes
Table 1Sediment trap site position water and trap depths sample selection for this study andrelated time interval as adapted from Malinverno et al (2009)
No Site Latitude Longitude Waterdepth(m)
Trapdepth(m)
Sub-samples Period ofsampling
1 48B 351455degN 237387degE 1967 995 48IB1a 1ndash15 Jun2005
2 48IB2 16ndash30Jun 2005
3 48IB4 16ndash31 Jul2005
4 48IB6 16ndash31Aug 2005
5 48IIB1 16 Octndash15 Nov2005
6 48IIB4b 1ndash15 Jan2005
7 48IIB9 16ndash31Mar 2006
8 48A 351455degN 237387degE 1967 1953 48IA3 1ndash15 Jul2005
9 48IA6 16ndash31Aug 2005
10 48IIA1 16 Octndash15 Nov2005
11 48IIA4 1ndash15 Jan2006
12 48IIA9 16ndash31Mar 2006
13 48IIA11c 15ndash30Apr 2006
48B is the shallow trap and 48A is the deep trapa Sample was also picked for individual H carterib Sample was only picked for H carteri populationc Sample was only picked for E huxleyi population
Table 2Surface sediment sample positions and water depth used for SrCa ratio analysis of Cleptoporus and H carteri
No Sample code Cruise Year Coordinates Waterdepth(m)
Longitude Latitude
1 M51-3 562 RV Meteor 2001 191910degE 327743degN 13902 M51-3 575 RV Meteor 2001 317862degE 345272degN 23373 M51-3 594 RV Meteor 2001 262185degE 379025degN 10054 M51-3 604 RV Meteor 2001 277883degE 408393degN 5525 Bio Deep 01 MCAT01a RV Urania 1997 213992degE 353012degN 32816 Bio Deep 03 AB12MCa RV Urania 1997 215103degE 352310degN 3310
a Sample was also picked for H carteri population
198 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Sea the so called ldquoTransition Mediterranean Waterrdquo (TMW) which isdefined as a ldquonutrient (phosphate) richndashoxygen poorrdquo intermediatewater mass (a layer between the LIW and Eastern Mediterranean DeepWater) can stimulate the productivity in the region in spring and fall(Souvermezoglou et al 1999)
In the EMED the main sources of fertilization originate from runoff from the surrounding land masses river inflow (mainly the Nile)by upwelling of the TMW and a combination between Saharan dustinput and precipitation (wet dust deposition) [Guerzoni et al 1999]which may be driven by two seasonal wind systems (Krom et al1991 2005 Carbo et al 2005 Eker-Develi et al 2006 Triantha-phyllou et al 2004 Ziveri et al 2000)
The maximum coccolithophore production is suggested to takeplace in late winter for the EMED (Kleijne 1991 Knappertsbusch1993) springndashsummer for the Cretan Sea and the Straits of Cretan Arc(Gotsis-Skretas et al 1999 Trianthaphyllou et al 2004) and in lateautumn and early spring for the coastal waters of Andros Island andthe Middle Aegean Sea (Dimiza et al 2008)
The high coccolithophore production in autumn as mentionedabove is incoherent with the results of Malinverno et al (2009) whichindicate the highest coccosphere flux occurring in spring and earlysummer Therefore by employing SrCa ratio of individual coccolithspecies we may unravel the timing of the highest coccosphereproduction and also elucidate whether the highest coccosphere fluxin spring is a real production signal or a remnant of greater exportefficiency due to aggregation promoted by diatoms and otherphytoplankton which dominated the winterndashspring blooms
3 Material and methods
Samples used in this studywere collected from two sediment trapsat one mooring station located off southwest Crete (35deg08732primeN23deg44322primeE) with a bottom depth of 1967 m (Fig 1) The depth ofdeployment of the two traps were at 995 m (48B) and 1953 m (48A)Sediment traps are PPS33 Technicap with a total of 12 collecting cupsand a collection area of 0125 m2 The 48B and 48A traps wereemployed from June 2005 until May 2006 with a sampling timeresolution of 15 to 16 days Detailed sampling processes samplepreservation sample preparation and coccolithophores speciescounting are explained by Stavrakakis et al (2000) and Malinvernoet al (2009) The biogeochemical and coccosphere export flux fromthese traps are from Malinverno et al (2009) Detailed sampleinformation used in this study is shown in Tables 1 and 2
SrCa measurements were carried out on individually pickedcoccoliths in order to obtain species-specific SrCa ratio time seriesand elucidate such a bias on SrCa ratio variation related to productivitywhichmay be given by different coccolith species (Stoll et al 2007a) Atotal of 10 samples (5 samples each from 48A and 48B) have beenselected for elemental chemistry using the new developed technique(Stoll et al 2007cd) of individually picked coccoliths We focused onthe surface dwellers C leptoporus andH carteri and the cosmopolitan Ehuxleyi (Fig 2) In order to assess whether the SrCa ratio signals fromthe EMED traps are similar to those from the seafloor sediments sixsurface sediment samples recovered from the EMEDwere used for SrCaratio analysis in the population of individually picked coccolithspecimens
Individual coccolith specimens were picked from a dry smear slideby a tungsten needle which is operated by a micromanipulator (waterrobot) under 400times magnification on an inverted polarizationmicroscope We picked 15ndash20 specimens for both ldquolargerrdquo speciesand 25ndash30 specimens for E huxleyi We isolated the specimens andthen placed them on thewell of a Tacky Dot slide The slidewas coatedby Epoxy (Buehler Epoxide) that was poured into an aluminium ringmounted on the slide Application of ethanol on the Tacky Dotremoved the epoxy mold from the slide The epoxy mold was thenpolished on an ethanol-wetted felt pad to remove the Tacky Dot
polymer coating Afterwards the mold was mounted on a CamecaIMS3f Secondary Ion Mass Spectrometer for SrCa ratio analysis Weshot a primary ion beam on a population of the individual coccolithsto obtain an average value of that particular species Data werecollected in three consecutive analyses of five-cycle scans of magneticfields covering 24 Mg 40 Ca and 88 Sr with a typical counting error of1 The standard deviation of collected data is 005 In addition wealso conducted profiles through single individual coccoliths of Hcarteri from the sediment traps which exhibited differing coccoliththickness and weight in order to observe the presence of secondarycalcification Detailed procedure of this analysis can be found in Stollet al (2007cd)
The time series (2004 until 2006) of surface water environmentparameters derived from SeaWiFS and MODIS-Aqua satellite datasets(httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) areshown in Fig 3 The surface water parameters are chlorophyll-aconcentration sea surface temperature (SST) and aerosol opticalthickness as indicator of aeolian dust The area coverage is 22degndash26degE
Fig 2 Scanning electron microscope pictures of the target species which were used for SrCa ratio analysis (a) C leptoporus (b) H carteri (c) E huxleyi
199L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
and 34degndash36degN The seasonal variation of surface production rangesfrom ~008 to ~022 mg mminus3 The maximum value of chlorophyll-aconcentration is recorded mostly in late winter until early spring andthe minimum value occurs in summer
In order to verify the species identification and the preservationstate of individual coccoliths Scanning Electron Microscope (SEM)analysis on each mounted stub was done at the SEM Laboratory VUUniversity Amsterdam The SrCa sample preparation and picking ofsingle coccoliths specimens were done at the Geology Department ofthe University of Oviedo Spain The Secondary IonMass Spectrometeranalysis of SrCa was conducted at the Northeast National IonMicroprobe Facility at Woods Hole Oceanographic Institution(WHOI) USA
4 Results
41 Shallow sediment trap 48B (995 m)
In the shallower sediment trap (48B) the total mass carbonateorganic carbon coccosphere and coccolith fluxes exhibit a strongseasonal pattern of particle fluxes (Fig 4ab Malinverno et al 2009) Afirst maximum in total mass carbonate organic carbon coccosphereand coccolith fluxes occurred between June and August 2005 Aminimum in fluxes occurred in autumn between September and earlyDecember During winter (December 2005ndashearly February 2006) totalmassflux including carbonate organic carbon and coccoliths increasedagain although the pronounced increase in coccosphere flux laggedbehind by several months and increased in March 2006 E huxleyi
Fig 3 Time series (December 2004ndashJune 2006) of chlorophyll-a concentration (OBPG SeaProducts for SST 11 microm day) and aerosol optical thickness (SeaWiFS Monthly Global 9-km
dominated both coccosphere and coccolith assemblages during thewhole investigated period andwasmost abundant during the intervalsof high coccosphere export fluxes in JunendashJuly 2005 andMarch 2006 Bycontrast during the high mass flux period in December 2005ndashearlyFebruary 2006 when coccosphere fluxes were not elevated thecoccosphere fluxes displayed a high abundance of deep-dwellingspecies in particular Algirosphaera robusta (Malinverno et al 2009)The fluxes of H carteri were most abundant during the periods of hightotal coccosphere flux Instead C leptoporus fluxes were always below1 of the total coccolithophore assemblage with slight increases duringthe total coccolith and coccosphere high flux intervals
The SrCa ratios of C leptoporus generally follow the seasonal trendin coccosphereexport in the shallower trap (Fig 5ab) HighC leptoporusSrCa ratios (299 mmolmol) accompanied the first peak in cocco-sphere and coccolith fluxes of this species in JunendashAugust Lower Cleptoporus SrCa (265 mmolmol) occurred in early autumn duringminima in fluxes In March 2006 an increase in coccosphere exportcorresponds toan increase in SrCa ratio (293 mmolmol)During JunendashJuly 2005 the peak of coccosphere and coccolith fluxes coincided withincreased SST and low values of the aerosol optical index andchlorophyll-a concentration (Fig 5h) In contrast the March 2006coccosphere peak lags by one month the peak values of chlorophyll-aconcentration and coincides with low SST
42 Deep sediment trap 48A (1953 m)
In the deeper sediment trap (48A) the total mass carbonate andorganic carbon show a slightly different seasonal pattern in fluxes
WiFS Monthly Global 9-km Products) SST (OBPG Modis-Aqua Monthly Global 9-kmProducts)
Fig 4 Total mass flux carbonate organic carbon coccosphere and coccolith fluxes from 48B (a b) and 48A (c d) according toMalinverno et al (2009) Periods A B C are indicated ingrey bars (see description in 51) Note timespans with no sample point and unconnected symbols indicate that no data are available
200 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
than the shallow trap although the pattern in coccosphere fluxes ismore similar (Fig 4cd Malinverno et al 2009) As in the shallowtrap there was a maximum in total mass carbonate organic carboncoccosphere and coccolith fluxes between June and July 2005however there was no spring 2006 peak in mass flux as observed inthe shallow trap Coccosphere fluxes in March 2006 in the deeper trapare significantly higher than those of winter 2005 From December2005 until February 2006 all fluxes show low values
H carteri SrCa ratios are not clearly related to total coccosphereflux patterns or coccosphere fluxes of H carteri (Fig 5cf) Minimumvalues occur in the early summer and early autumn (313 mmolmol)and maximum value (34 mmolmol) in January 2006 The peak in Hcarteri SrCa in January 2006 coincideswith a peak in total mass flux inthe shallow trap where the coccospheres of H carteri and Helico-sphaera spp also showed peak fluxes and increased relativeabundance
For the same trap periods there are differences in SrCa ratios in Cleptoporus in the deep and shallow sediment traps (Fig 5ce) Therange of SrCa ratios in the deeper trap (283 to 306 mmolmol) is
smaller than in the shallower trap (265 to 299 mmolmol) and noanalysis was performed for mid-October The early summer 2005value in the deeper trap is lower than the corresponding one in theshallow trap
The seasonal trend in E huxleyi SrCa ratios is similar to that seenin C leptoporus in the shallow trap E huxleyi SrCa ratio exhibitedmaximum values (195 to 201 mmolmol) during the early summer(June ndash July 2005) and spring (March 2006) periods of high fluxes of Ehuxleyi coccospheres and lower SrCa ratios (184 mmolmol) duringthe period of low fluxes (Fig 5dg) Overall E huxleyi SrCa ratios arelower than those of C leptoporus and H carteri
43 SrCa ratios along a profile of H carteri
We analysed SrCa ratios on individual coccoliths of H carteri(sample 48IB1 Table 1) having similar length but different carbonatemass (ldquothicknessrdquo or degree of calcification) The SrCa ratios of allspecimens (Fig 6) are within the analytical uncertainty for analysis ofa single coccolith (2 standard errors of the mean) No significant
Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675
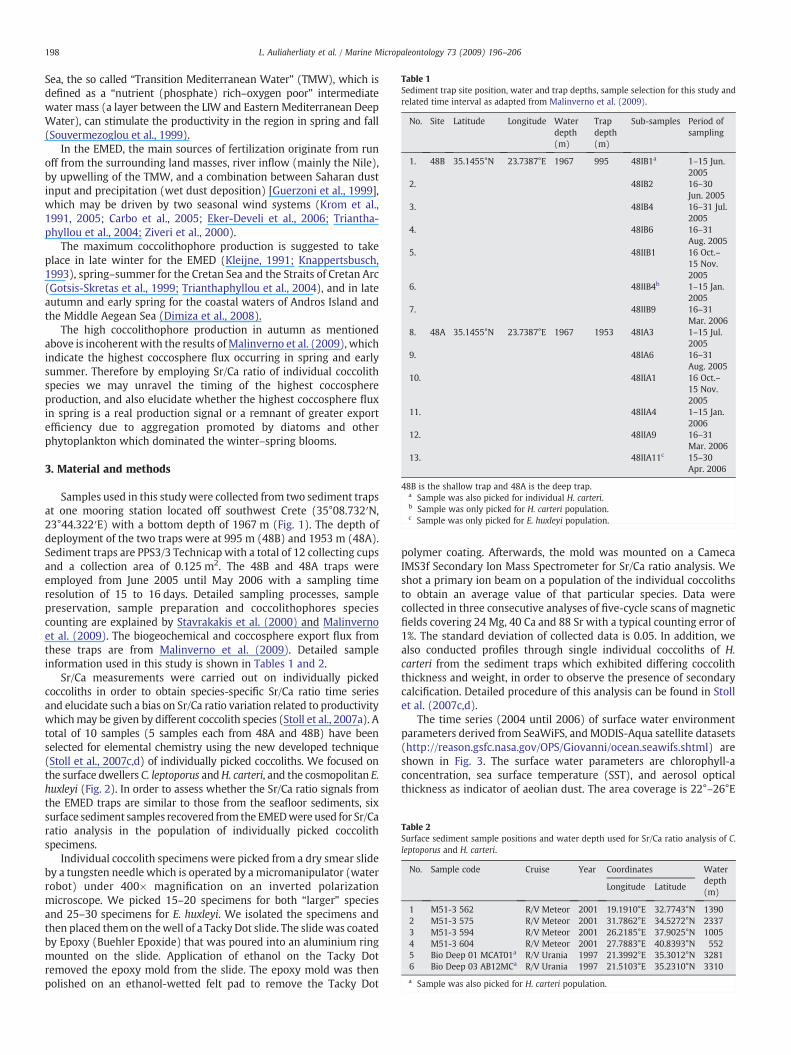
Table 1Sediment trap site position water and trap depths sample selection for this study andrelated time interval as adapted from Malinverno et al (2009)
No Site Latitude Longitude Waterdepth(m)
Trapdepth(m)
Sub-samples Period ofsampling
1 48B 351455degN 237387degE 1967 995 48IB1a 1ndash15 Jun2005
2 48IB2 16ndash30Jun 2005
3 48IB4 16ndash31 Jul2005
4 48IB6 16ndash31Aug 2005
5 48IIB1 16 Octndash15 Nov2005
6 48IIB4b 1ndash15 Jan2005
7 48IIB9 16ndash31Mar 2006
8 48A 351455degN 237387degE 1967 1953 48IA3 1ndash15 Jul2005
9 48IA6 16ndash31Aug 2005
10 48IIA1 16 Octndash15 Nov2005
11 48IIA4 1ndash15 Jan2006
12 48IIA9 16ndash31Mar 2006
13 48IIA11c 15ndash30Apr 2006
48B is the shallow trap and 48A is the deep trapa Sample was also picked for individual H carterib Sample was only picked for H carteri populationc Sample was only picked for E huxleyi population
Table 2Surface sediment sample positions and water depth used for SrCa ratio analysis of Cleptoporus and H carteri
No Sample code Cruise Year Coordinates Waterdepth(m)
Longitude Latitude
1 M51-3 562 RV Meteor 2001 191910degE 327743degN 13902 M51-3 575 RV Meteor 2001 317862degE 345272degN 23373 M51-3 594 RV Meteor 2001 262185degE 379025degN 10054 M51-3 604 RV Meteor 2001 277883degE 408393degN 5525 Bio Deep 01 MCAT01a RV Urania 1997 213992degE 353012degN 32816 Bio Deep 03 AB12MCa RV Urania 1997 215103degE 352310degN 3310
a Sample was also picked for H carteri population
198 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Sea the so called ldquoTransition Mediterranean Waterrdquo (TMW) which isdefined as a ldquonutrient (phosphate) richndashoxygen poorrdquo intermediatewater mass (a layer between the LIW and Eastern Mediterranean DeepWater) can stimulate the productivity in the region in spring and fall(Souvermezoglou et al 1999)
In the EMED the main sources of fertilization originate from runoff from the surrounding land masses river inflow (mainly the Nile)by upwelling of the TMW and a combination between Saharan dustinput and precipitation (wet dust deposition) [Guerzoni et al 1999]which may be driven by two seasonal wind systems (Krom et al1991 2005 Carbo et al 2005 Eker-Develi et al 2006 Triantha-phyllou et al 2004 Ziveri et al 2000)
The maximum coccolithophore production is suggested to takeplace in late winter for the EMED (Kleijne 1991 Knappertsbusch1993) springndashsummer for the Cretan Sea and the Straits of Cretan Arc(Gotsis-Skretas et al 1999 Trianthaphyllou et al 2004) and in lateautumn and early spring for the coastal waters of Andros Island andthe Middle Aegean Sea (Dimiza et al 2008)
The high coccolithophore production in autumn as mentionedabove is incoherent with the results of Malinverno et al (2009) whichindicate the highest coccosphere flux occurring in spring and earlysummer Therefore by employing SrCa ratio of individual coccolithspecies we may unravel the timing of the highest coccosphereproduction and also elucidate whether the highest coccosphere fluxin spring is a real production signal or a remnant of greater exportefficiency due to aggregation promoted by diatoms and otherphytoplankton which dominated the winterndashspring blooms
3 Material and methods
Samples used in this studywere collected from two sediment trapsat one mooring station located off southwest Crete (35deg08732primeN23deg44322primeE) with a bottom depth of 1967 m (Fig 1) The depth ofdeployment of the two traps were at 995 m (48B) and 1953 m (48A)Sediment traps are PPS33 Technicap with a total of 12 collecting cupsand a collection area of 0125 m2 The 48B and 48A traps wereemployed from June 2005 until May 2006 with a sampling timeresolution of 15 to 16 days Detailed sampling processes samplepreservation sample preparation and coccolithophores speciescounting are explained by Stavrakakis et al (2000) and Malinvernoet al (2009) The biogeochemical and coccosphere export flux fromthese traps are from Malinverno et al (2009) Detailed sampleinformation used in this study is shown in Tables 1 and 2
SrCa measurements were carried out on individually pickedcoccoliths in order to obtain species-specific SrCa ratio time seriesand elucidate such a bias on SrCa ratio variation related to productivitywhichmay be given by different coccolith species (Stoll et al 2007a) Atotal of 10 samples (5 samples each from 48A and 48B) have beenselected for elemental chemistry using the new developed technique(Stoll et al 2007cd) of individually picked coccoliths We focused onthe surface dwellers C leptoporus andH carteri and the cosmopolitan Ehuxleyi (Fig 2) In order to assess whether the SrCa ratio signals fromthe EMED traps are similar to those from the seafloor sediments sixsurface sediment samples recovered from the EMEDwere used for SrCaratio analysis in the population of individually picked coccolithspecimens
Individual coccolith specimens were picked from a dry smear slideby a tungsten needle which is operated by a micromanipulator (waterrobot) under 400times magnification on an inverted polarizationmicroscope We picked 15ndash20 specimens for both ldquolargerrdquo speciesand 25ndash30 specimens for E huxleyi We isolated the specimens andthen placed them on thewell of a Tacky Dot slide The slidewas coatedby Epoxy (Buehler Epoxide) that was poured into an aluminium ringmounted on the slide Application of ethanol on the Tacky Dotremoved the epoxy mold from the slide The epoxy mold was thenpolished on an ethanol-wetted felt pad to remove the Tacky Dot
polymer coating Afterwards the mold was mounted on a CamecaIMS3f Secondary Ion Mass Spectrometer for SrCa ratio analysis Weshot a primary ion beam on a population of the individual coccolithsto obtain an average value of that particular species Data werecollected in three consecutive analyses of five-cycle scans of magneticfields covering 24 Mg 40 Ca and 88 Sr with a typical counting error of1 The standard deviation of collected data is 005 In addition wealso conducted profiles through single individual coccoliths of Hcarteri from the sediment traps which exhibited differing coccoliththickness and weight in order to observe the presence of secondarycalcification Detailed procedure of this analysis can be found in Stollet al (2007cd)
The time series (2004 until 2006) of surface water environmentparameters derived from SeaWiFS and MODIS-Aqua satellite datasets(httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) areshown in Fig 3 The surface water parameters are chlorophyll-aconcentration sea surface temperature (SST) and aerosol opticalthickness as indicator of aeolian dust The area coverage is 22degndash26degE
Fig 2 Scanning electron microscope pictures of the target species which were used for SrCa ratio analysis (a) C leptoporus (b) H carteri (c) E huxleyi
199L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
and 34degndash36degN The seasonal variation of surface production rangesfrom ~008 to ~022 mg mminus3 The maximum value of chlorophyll-aconcentration is recorded mostly in late winter until early spring andthe minimum value occurs in summer
In order to verify the species identification and the preservationstate of individual coccoliths Scanning Electron Microscope (SEM)analysis on each mounted stub was done at the SEM Laboratory VUUniversity Amsterdam The SrCa sample preparation and picking ofsingle coccoliths specimens were done at the Geology Department ofthe University of Oviedo Spain The Secondary IonMass Spectrometeranalysis of SrCa was conducted at the Northeast National IonMicroprobe Facility at Woods Hole Oceanographic Institution(WHOI) USA
4 Results
41 Shallow sediment trap 48B (995 m)
In the shallower sediment trap (48B) the total mass carbonateorganic carbon coccosphere and coccolith fluxes exhibit a strongseasonal pattern of particle fluxes (Fig 4ab Malinverno et al 2009) Afirst maximum in total mass carbonate organic carbon coccosphereand coccolith fluxes occurred between June and August 2005 Aminimum in fluxes occurred in autumn between September and earlyDecember During winter (December 2005ndashearly February 2006) totalmassflux including carbonate organic carbon and coccoliths increasedagain although the pronounced increase in coccosphere flux laggedbehind by several months and increased in March 2006 E huxleyi
Fig 3 Time series (December 2004ndashJune 2006) of chlorophyll-a concentration (OBPG SeaProducts for SST 11 microm day) and aerosol optical thickness (SeaWiFS Monthly Global 9-km
dominated both coccosphere and coccolith assemblages during thewhole investigated period andwasmost abundant during the intervalsof high coccosphere export fluxes in JunendashJuly 2005 andMarch 2006 Bycontrast during the high mass flux period in December 2005ndashearlyFebruary 2006 when coccosphere fluxes were not elevated thecoccosphere fluxes displayed a high abundance of deep-dwellingspecies in particular Algirosphaera robusta (Malinverno et al 2009)The fluxes of H carteri were most abundant during the periods of hightotal coccosphere flux Instead C leptoporus fluxes were always below1 of the total coccolithophore assemblage with slight increases duringthe total coccolith and coccosphere high flux intervals
The SrCa ratios of C leptoporus generally follow the seasonal trendin coccosphereexport in the shallower trap (Fig 5ab) HighC leptoporusSrCa ratios (299 mmolmol) accompanied the first peak in cocco-sphere and coccolith fluxes of this species in JunendashAugust Lower Cleptoporus SrCa (265 mmolmol) occurred in early autumn duringminima in fluxes In March 2006 an increase in coccosphere exportcorresponds toan increase in SrCa ratio (293 mmolmol)During JunendashJuly 2005 the peak of coccosphere and coccolith fluxes coincided withincreased SST and low values of the aerosol optical index andchlorophyll-a concentration (Fig 5h) In contrast the March 2006coccosphere peak lags by one month the peak values of chlorophyll-aconcentration and coincides with low SST
42 Deep sediment trap 48A (1953 m)
In the deeper sediment trap (48A) the total mass carbonate andorganic carbon show a slightly different seasonal pattern in fluxes
WiFS Monthly Global 9-km Products) SST (OBPG Modis-Aqua Monthly Global 9-kmProducts)
Fig 4 Total mass flux carbonate organic carbon coccosphere and coccolith fluxes from 48B (a b) and 48A (c d) according toMalinverno et al (2009) Periods A B C are indicated ingrey bars (see description in 51) Note timespans with no sample point and unconnected symbols indicate that no data are available
200 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
than the shallow trap although the pattern in coccosphere fluxes ismore similar (Fig 4cd Malinverno et al 2009) As in the shallowtrap there was a maximum in total mass carbonate organic carboncoccosphere and coccolith fluxes between June and July 2005however there was no spring 2006 peak in mass flux as observed inthe shallow trap Coccosphere fluxes in March 2006 in the deeper trapare significantly higher than those of winter 2005 From December2005 until February 2006 all fluxes show low values
H carteri SrCa ratios are not clearly related to total coccosphereflux patterns or coccosphere fluxes of H carteri (Fig 5cf) Minimumvalues occur in the early summer and early autumn (313 mmolmol)and maximum value (34 mmolmol) in January 2006 The peak in Hcarteri SrCa in January 2006 coincideswith a peak in total mass flux inthe shallow trap where the coccospheres of H carteri and Helico-sphaera spp also showed peak fluxes and increased relativeabundance
For the same trap periods there are differences in SrCa ratios in Cleptoporus in the deep and shallow sediment traps (Fig 5ce) Therange of SrCa ratios in the deeper trap (283 to 306 mmolmol) is
smaller than in the shallower trap (265 to 299 mmolmol) and noanalysis was performed for mid-October The early summer 2005value in the deeper trap is lower than the corresponding one in theshallow trap
The seasonal trend in E huxleyi SrCa ratios is similar to that seenin C leptoporus in the shallow trap E huxleyi SrCa ratio exhibitedmaximum values (195 to 201 mmolmol) during the early summer(June ndash July 2005) and spring (March 2006) periods of high fluxes of Ehuxleyi coccospheres and lower SrCa ratios (184 mmolmol) duringthe period of low fluxes (Fig 5dg) Overall E huxleyi SrCa ratios arelower than those of C leptoporus and H carteri
43 SrCa ratios along a profile of H carteri
We analysed SrCa ratios on individual coccoliths of H carteri(sample 48IB1 Table 1) having similar length but different carbonatemass (ldquothicknessrdquo or degree of calcification) The SrCa ratios of allspecimens (Fig 6) are within the analytical uncertainty for analysis ofa single coccolith (2 standard errors of the mean) No significant
Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

Fig 2 Scanning electron microscope pictures of the target species which were used for SrCa ratio analysis (a) C leptoporus (b) H carteri (c) E huxleyi
199L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
and 34degndash36degN The seasonal variation of surface production rangesfrom ~008 to ~022 mg mminus3 The maximum value of chlorophyll-aconcentration is recorded mostly in late winter until early spring andthe minimum value occurs in summer
In order to verify the species identification and the preservationstate of individual coccoliths Scanning Electron Microscope (SEM)analysis on each mounted stub was done at the SEM Laboratory VUUniversity Amsterdam The SrCa sample preparation and picking ofsingle coccoliths specimens were done at the Geology Department ofthe University of Oviedo Spain The Secondary IonMass Spectrometeranalysis of SrCa was conducted at the Northeast National IonMicroprobe Facility at Woods Hole Oceanographic Institution(WHOI) USA
4 Results
41 Shallow sediment trap 48B (995 m)
In the shallower sediment trap (48B) the total mass carbonateorganic carbon coccosphere and coccolith fluxes exhibit a strongseasonal pattern of particle fluxes (Fig 4ab Malinverno et al 2009) Afirst maximum in total mass carbonate organic carbon coccosphereand coccolith fluxes occurred between June and August 2005 Aminimum in fluxes occurred in autumn between September and earlyDecember During winter (December 2005ndashearly February 2006) totalmassflux including carbonate organic carbon and coccoliths increasedagain although the pronounced increase in coccosphere flux laggedbehind by several months and increased in March 2006 E huxleyi
Fig 3 Time series (December 2004ndashJune 2006) of chlorophyll-a concentration (OBPG SeaProducts for SST 11 microm day) and aerosol optical thickness (SeaWiFS Monthly Global 9-km
dominated both coccosphere and coccolith assemblages during thewhole investigated period andwasmost abundant during the intervalsof high coccosphere export fluxes in JunendashJuly 2005 andMarch 2006 Bycontrast during the high mass flux period in December 2005ndashearlyFebruary 2006 when coccosphere fluxes were not elevated thecoccosphere fluxes displayed a high abundance of deep-dwellingspecies in particular Algirosphaera robusta (Malinverno et al 2009)The fluxes of H carteri were most abundant during the periods of hightotal coccosphere flux Instead C leptoporus fluxes were always below1 of the total coccolithophore assemblage with slight increases duringthe total coccolith and coccosphere high flux intervals
The SrCa ratios of C leptoporus generally follow the seasonal trendin coccosphereexport in the shallower trap (Fig 5ab) HighC leptoporusSrCa ratios (299 mmolmol) accompanied the first peak in cocco-sphere and coccolith fluxes of this species in JunendashAugust Lower Cleptoporus SrCa (265 mmolmol) occurred in early autumn duringminima in fluxes In March 2006 an increase in coccosphere exportcorresponds toan increase in SrCa ratio (293 mmolmol)During JunendashJuly 2005 the peak of coccosphere and coccolith fluxes coincided withincreased SST and low values of the aerosol optical index andchlorophyll-a concentration (Fig 5h) In contrast the March 2006coccosphere peak lags by one month the peak values of chlorophyll-aconcentration and coincides with low SST
42 Deep sediment trap 48A (1953 m)
In the deeper sediment trap (48A) the total mass carbonate andorganic carbon show a slightly different seasonal pattern in fluxes
WiFS Monthly Global 9-km Products) SST (OBPG Modis-Aqua Monthly Global 9-kmProducts)
Fig 4 Total mass flux carbonate organic carbon coccosphere and coccolith fluxes from 48B (a b) and 48A (c d) according toMalinverno et al (2009) Periods A B C are indicated ingrey bars (see description in 51) Note timespans with no sample point and unconnected symbols indicate that no data are available
200 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
than the shallow trap although the pattern in coccosphere fluxes ismore similar (Fig 4cd Malinverno et al 2009) As in the shallowtrap there was a maximum in total mass carbonate organic carboncoccosphere and coccolith fluxes between June and July 2005however there was no spring 2006 peak in mass flux as observed inthe shallow trap Coccosphere fluxes in March 2006 in the deeper trapare significantly higher than those of winter 2005 From December2005 until February 2006 all fluxes show low values
H carteri SrCa ratios are not clearly related to total coccosphereflux patterns or coccosphere fluxes of H carteri (Fig 5cf) Minimumvalues occur in the early summer and early autumn (313 mmolmol)and maximum value (34 mmolmol) in January 2006 The peak in Hcarteri SrCa in January 2006 coincideswith a peak in total mass flux inthe shallow trap where the coccospheres of H carteri and Helico-sphaera spp also showed peak fluxes and increased relativeabundance
For the same trap periods there are differences in SrCa ratios in Cleptoporus in the deep and shallow sediment traps (Fig 5ce) Therange of SrCa ratios in the deeper trap (283 to 306 mmolmol) is
smaller than in the shallower trap (265 to 299 mmolmol) and noanalysis was performed for mid-October The early summer 2005value in the deeper trap is lower than the corresponding one in theshallow trap
The seasonal trend in E huxleyi SrCa ratios is similar to that seenin C leptoporus in the shallow trap E huxleyi SrCa ratio exhibitedmaximum values (195 to 201 mmolmol) during the early summer(June ndash July 2005) and spring (March 2006) periods of high fluxes of Ehuxleyi coccospheres and lower SrCa ratios (184 mmolmol) duringthe period of low fluxes (Fig 5dg) Overall E huxleyi SrCa ratios arelower than those of C leptoporus and H carteri
43 SrCa ratios along a profile of H carteri
We analysed SrCa ratios on individual coccoliths of H carteri(sample 48IB1 Table 1) having similar length but different carbonatemass (ldquothicknessrdquo or degree of calcification) The SrCa ratios of allspecimens (Fig 6) are within the analytical uncertainty for analysis ofa single coccolith (2 standard errors of the mean) No significant
Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

Fig 4 Total mass flux carbonate organic carbon coccosphere and coccolith fluxes from 48B (a b) and 48A (c d) according toMalinverno et al (2009) Periods A B C are indicated ingrey bars (see description in 51) Note timespans with no sample point and unconnected symbols indicate that no data are available
200 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
than the shallow trap although the pattern in coccosphere fluxes ismore similar (Fig 4cd Malinverno et al 2009) As in the shallowtrap there was a maximum in total mass carbonate organic carboncoccosphere and coccolith fluxes between June and July 2005however there was no spring 2006 peak in mass flux as observed inthe shallow trap Coccosphere fluxes in March 2006 in the deeper trapare significantly higher than those of winter 2005 From December2005 until February 2006 all fluxes show low values
H carteri SrCa ratios are not clearly related to total coccosphereflux patterns or coccosphere fluxes of H carteri (Fig 5cf) Minimumvalues occur in the early summer and early autumn (313 mmolmol)and maximum value (34 mmolmol) in January 2006 The peak in Hcarteri SrCa in January 2006 coincideswith a peak in total mass flux inthe shallow trap where the coccospheres of H carteri and Helico-sphaera spp also showed peak fluxes and increased relativeabundance
For the same trap periods there are differences in SrCa ratios in Cleptoporus in the deep and shallow sediment traps (Fig 5ce) Therange of SrCa ratios in the deeper trap (283 to 306 mmolmol) is
smaller than in the shallower trap (265 to 299 mmolmol) and noanalysis was performed for mid-October The early summer 2005value in the deeper trap is lower than the corresponding one in theshallow trap
The seasonal trend in E huxleyi SrCa ratios is similar to that seenin C leptoporus in the shallow trap E huxleyi SrCa ratio exhibitedmaximum values (195 to 201 mmolmol) during the early summer(June ndash July 2005) and spring (March 2006) periods of high fluxes of Ehuxleyi coccospheres and lower SrCa ratios (184 mmolmol) duringthe period of low fluxes (Fig 5dg) Overall E huxleyi SrCa ratios arelower than those of C leptoporus and H carteri
43 SrCa ratios along a profile of H carteri
We analysed SrCa ratios on individual coccoliths of H carteri(sample 48IB1 Table 1) having similar length but different carbonatemass (ldquothicknessrdquo or degree of calcification) The SrCa ratios of allspecimens (Fig 6) are within the analytical uncertainty for analysis ofa single coccolith (2 standard errors of the mean) No significant
Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

Fig 5 (a) C leptoporus SrCa ratios from the shallow trap (48B) (b) Total coccosphere (Malinverno et al 2009) and coccolith fluxes of C leptoporus from the shallow trap 48B (c) Cleptoporus and H carteri SrCa ratios from the deep trap (48A) (d) E huxleyi SrCa ratio from the deep trap (48A) (e f g) Total coccosphere (Malinverno et al 2009) and coccolithfluxes of C leptoporus H carteri and E huxleyi from the deep trap 48A (h) Surface water parameters derived from SeaWiFS and MODIS-Aqua satellite datasets Periods A B C areshown in grey bars Note timespans with no sample point and unconnected symbols indicate that no data are available
201L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
changes of SrCa ratios are recorded The SrCa of the thin specimen is~256 mmolmol The normal specimen shows the value of~265 mmolmol and the value for the thicker one is ~258 mmolmol
44 Surface sediments of EMED
C leptoporus SrCa ratios of EMED surface sediment samples rangefrom 215 to 279 mmolmol H carteri SrCa ratios show slightlyhigher values ranging from 303 to 318 mmolmol The SrCa ratios ofall surface sediments for both species (Fig 7) are lower than thevalues we obtained from sediment traps
5 Discussion
51 Surface production and dynamic of export processes offshore thesouth-western margin of Crete
According to the occurrences of coccosphere flux maxima andminima in the sediment traps (Fig 4) we discuss the data in threeseparate intervals from June to July 2005 as period A from Decemberto early February 2006 as period B and March 2006 as period C Wepropose that during periods A and C high production was directlyrecorded in the traps as evidenced by high coccosphere fluxes whilstduring period B productionwas low and a pulse in fluxes in December
Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

Fig 6 (a) SrCa ratios of H carteri with different thickness versus peak of Ca40 intensity
202 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
2005 mainly represented re-suspended material as evidenced by theabsence of a high coccosphere flux
For C leptoporus SrCa ratios we focus on the higher amplitudeand more complete signal in the shallower 48B trap (Fig 5) Thedeeper 48A trap (Fig 6) may have more attenuated signals because oflonger residence times mixing and scavenging of materials of varyingproduction times in the water column (Honjo 1976 Honjo 1982)Furthermore the proximity of the deeper 48A trap to the seafloor(14m) provides potential for resuspension from the bottom sedi-ments The canyon system where this mooring station is located mayalso contribute to lateral transport from the coastal area Moreover adecrease in abundance of about ~3 of holococcolithophores withincreasing sediment trap depth is detected from ~1000 to ~2000 mwater depth (Malinverno et al 2009) and may be indicative of watercolumn selective dissolution of more dissolution-susceptible speciesduring the transit from the shallow to the deeper trap
511 Early summer bloom (JunendashJuly 2005) Period AIn early summer 2005 maxima of coccosphere coccolith organic
carbon and daily fluxes of carbonate were recorded (Fig 4) Thesemaxima show correlation with relatively heavy coccoliths ie Cleptoporus and H carteri (Young and Ziveri 2000) and organic carbondaily fluxes This correlation may indicate a role of these coccoliths asmineral ballast as suggested for the North Atlantic (Ziveri et al 2007)
In period A coccosphere fluxes and higher SrCa ratios in E huxleyiindicate an interval of high surface production and export processes(Fig 5dg) E huxleyi is the dominant coccolithophore species at thattime period (Malinverno et al 2009) SrCa maxima in C leptoporuspersisted several months after the relatively low coccosphere andslightly high coccolith fluxes (Fig 5abe) The same mechanism wasalso observed in response to a fall production period triggered by aneddy in the Sargasso Sea in 1996 (Stoll et al 2007a) Similarly weconsider that there is a long ldquotailrdquo to the distribution of settling ratesso that material from the production period still arrived at thesediment traps long after the actual bloom ended For the late springndash
Fig 7 Distribution of the SrCa ratios values of C leptoporus (
early summer 2005 period there is no corresponding chlorophyllpeak detected by satellite SeaWIFS chlorophyll-a (httpreasongsfcnasagovOPSGiovannioceanseawifsshtml) peaked in February2005 but was low by June 2005 Instead E huxleyi and C leptoporusSrCa values indicative of high production are recorded in mid-June2005 in our traps Given that coccolithophores are the mainphytoplankton group contributing to the biogenic fluxes to the trapsas well as in other Mediterranean settings we should consider thesurface chlorophyll-a signal as indicative of coccolithophore produc-tion This would imply a 4 month delay between the surfaceproduction period and the export flux into 995m water depthHowever we should also consider that satellite-sensed chlorophyll islimited to the very surface of the ocean while phytoplanktonproduction may develop at greater depth at our sediment trap ahigh flux of deep-dwelling coccolithophore species is recorded in latespring indicating a flourishing deep community (Malinverno et al2009) Therefore long sinking times are not required to account forthe apparent delay between surface chlorophyll-a and flux peaks as itis shown for March 2006 which will be discussed later
Late springndashearly summer peaks in export production could betriggered by other mechanisms in addition to deep water mixing Forexample the peak of export production in late spring 2001 fromanother nearby sediment trap located in the Cretan Sea was strictlyrelated to fertilization of the TMW and possibly due to Saharan dustinput at the sea surface (Trianthaphyllou et al 2004) However oursatellite dataset shows there is no correlation between high cocco-sphere flux and the aerosol optical index as indicator of dust inputHigh coccosphere flux coincided with low aerosol optical index andincreased SST instead This corroborates Saharan dust ineffectivenessin fertilization during in situ experiment after dust storm (Krom et al2005) and in the paleo record (Incarbona et al 2008) in contrast withthe results from microcosm experiments (Herut et al 2005)Combination of these findings may suggest that the role of Saharandust in fertilization applies only in exceptional conditions
Eddies and gyres may influence the dynamics of surface produc-tion and export processes since cold core cyclonic eddies wouldenhance the efficiency of nutrient diffusion from deeper in the watercolumn (Gotsis-Skretas et al 1999 Moutin and Raimbault 2002)However the Western Cretan and Iera-Petra gyres are typicallyestablished later between late summer and early winter when north-easterly winds interact with the relief of Crete Island and attainmaximum wind speed (Malanotte-Rizzoli et al 1997 Pinardi andMasetti 2000 Robinson et al 2001 Hamad et al 2006)
Therefore we may summarize that the E huxleyi bloom andincreased SrCa ratios in period Bmay originate as an in situ signal dueto the ability of this species to inhabit also the deeper photic zone thusescaping satellite detection In addition the SrCa ratios of Cleptoporus are related to the coccolith signal which may remain in
a) and H carteri (b) from surface sediments in the EMED
203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

203L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
the water column after transport from a nearby area whichexperienced such a bloom
512 Lower production autumn and early winter (December 2005ndashearly February 2006) Period B
The late summer fall and early winter 2005were characterized bylower surface productivity and a scavenging event Berman et al(2003) mentioned that in the EMED SSTs differ ~12 degC betweensummer (~28 degC) and winter (~16 degC) thus an increase in SST and awater column re-stratification may cause a shallow surface mixedlayer and consequently a low nutrient supply to the upper photiczone This is in agreement with an increase of deep water speciesduring this interval (Malinverno et al 2009)
The minimum in C leptoporus SrCa ratios in the shallow trapsoccurs in mid-October coincident with low coccosphere and coccolithfluxes and high SST Likewise theminimum in E huxleyi SrCa ratios isreached during this time period We interpret the fallndashearly winterperiod as one of low surface coccolithophore production especiallyfor these species which are associated with moderate to highturbulence andor upwelling
The response of the SrCa ratios in H carteri is more complex as itis opposite to that of C leptoporus and E huxleyi with peak SrCaratios during the time interval of low overall production Therelatively low Helicosphaera coccosphere flux in the samples withhigh H carteri SrCa ratios might suggest a slightly different behaviourin this species yet from this limited dataset we are uncertain whetherthere is anti-correlation in term of production between C leptoporusand H carteri in the photic zone
Despite the fact that in the EMED we observe a general correlationbetween coccolith SrCa ratio and the export of C leptoporus and Ehuxleyi the amplitude of this change is smaller (~03 mmolmol) thanthose observed over seasonal productivity cycles in sediment traps inthe Arabian Sea and Bermuda (~10 mmolmol) (Stoll et al 2007a)This may reflect the different food web in the EMED which featuresmuch lower primary production aswell as export efficiency and hencemuch greater mixing of production from different seasons Stoll et al(2007a) demonstrated that the SrCa ratios of larger species from asediment trap recovered from the Arabian Sea are high duringsouthwest and northwest monsoon induced upwelling and alsocorrespond to high export In the Sargasso Sea during the fall bloomtriggered by mode water eddy higher SrCa ratios correspond tohigher export However the summer cyclonic eddies in the SargassoSea trigger higher C leptoporus standing stocks and higher SrCa ratiosbut do not cause a higher export (decoupling of export fromproduction and SrCa)
513 High productivity~early spring bloom (March 2006) Period CC leptoporus and E huxleyi SrCa ratios in sediment traps
significantly correlate to the coccosphere flux demonstrating thelink with surface production and export Increase of SrCa ratios in Cleptoporus and E huxleyi in period C reflects a late winterearly springbloom The peak of surface production is recorded by satellite inFebruary 2006 through chlorophyll-a data Therefore a possible onemonth delay occurred between production and sedimentationMalinverno et al (2009) suggested that maximum surface productionin February coincides with intervals of maximum water columnmixing which supplied maximum nutrients to the upper photic zonedetectable by satellite-sensed chlorophyll-a measurement The lowoverall fluxes in this period suggest that the water column may nothave been completely mixed even though SST was low and the mixedlayer deepened (Malinverno et al 2009) Spring blooms emerge to bea common feature in the EMED Knappertsbusch (1993) quantifiedthe coccolithophore abundance in the water column and observedthat during late winter (FebruaryndashMarch 1988) the coccolithophoreconcentration was 5 to 10 times higher than the average in summerand E huxleyi dominated the community In addition Dimiza et al
(2008) reported highest coccolithophore total cell densities duringearly spring in the Aegean Sea The sediment trap deployed fromJanuary 2001 to February 2002 in the Cretan Sea (Trianthaphyllou etal 2004) showedmaximum coccolith flux at the 500 m trap from lateMarch to late June 2001 (~9times108mminus2 dminus1) and in the deeper 700 mtrap in late April 2001 (~39times105mminus2 dminus1) However such a highseasonal-interannual variability of particle sinking rate (exportprocess to the deeper depth) occurs in this sub-basin as reported byMalinverno et al (2003a) Ziveri et al (2000) and Rutten et al (2000)The surface sediments from the EMED are dominated by E huxleyi (50to 60 of the fossil assemblage) implying that this species likelyproduces regular blooms in surface waters during late winter orspring in the region (Knappertsbusch 1993 Ziveri et al 2000Malinverno et al 2003b Trianthaphyllou et al 2004 Dimiza et al2008 Turgoklu 2008) and that their production is efficientlyexported to the bottom
52 Primary calcite precipitation in H carteri
According to previous studies in the surface and Quaternary coresediments from the EMED calcite overgrowth of E huxleyi and Reti-culofenestra have been documented (Ziveri et al 2000 Crudeli et al2004) The overgrowth has been interpreted as secondary calciteprecipitation possibly related to carbonate diagenesis which occurredwithin the sediment as it was found in the Red Sea (Winter 1982) and(or) at the sediment-water interface in the EMED as proposed byCrudeli et al (2004) However clear mechanistic understanding ofthis process remains unsolved Moreover heavily calcified E huxleyispecimens have been also reported in the photic zone of the EMEDespecially in the Aegean Sea during winter (Dimiza et al 2008Triantaphyllou et al in press) However this type of overcalcificationis morphologically different from the former and is shown to bephysiologically controlled (Triantaphyllou et al in press)
Our results on H carteri suggest that no secondary calcification hasoccurred on the selected specimens Abiogenic calcite is expected tohave SrCa ratios of 026 mmolmol based on equilibrium partitioning(Tesoriero and Pankow 1996) If abiogenic secondary overgrowthwas a significant part of the coccolith calcite the SrCa ratio of theabiogenic component (which is one order of magnitude lower) woulddecrease the mean SrCa ratio of the overgrown thickened coccolithsrelative to normal or thin ones The measured average SrCa ratios arenot significantly different among thick normal and thin specimensThis confirms that variations in coccolith weight are primary featuresof coccolith biomineralization in this genus as it has been found earlierin E huxleyi (Winter 1985 Samtleben and Bickert 1990 Ziveri et al1995 Beaufort and Heussner 2001 Engel et al 2005) at differentlocations Triantaphyllou et al (in press) shows that the calcificationof E huxleyi coccoliths in the Aegean Sea is seasonally affected withmore calcified specimens occurring in the winter season SrCa ratiosfor all individuals is slightly lower than the mean values of populationof H carteri extracted from surface sediments (~28 mmolmol) andsediment traps (~32 mmolmol) however it is still within the rangeof primary H carteri production in the area
53 EMED as an oligotrophic region
The EMED is a ldquonutrient desertrdquo regionwhere phosphorous limitationis occurring (Krom et al 1991 Pinet 1992 Carbo et al 2005) due toseveral suggested processes ie unequal N and P supply from externalsources adsorptionof phosphate by inorganic particles N2fixation (Herutet al 1999 Karl et al 1997 Sachs and Repeta 1999)
The rangeof SrCa values for each species is shown in Fig 8 The SrCaratios of C leptoporus ratios show larger variation among sites thanthose of H carteri and E huxleyi C leptoporus SrCa ratios in EMEDsediment traps during both high production events and mixing periodare slightly higher than in surface sediment samples The reason for this
Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

Fig 8 Comparison of C leptoporus H carteri and E huxleyi SrCa ratios from ion probe measurements of sediment trap (ST) and core top (CT) samples recovered from differentenvironmental settings The bars give the range of values from the individual SrCa ratio measurements The SrCa ratios of coccolithophores from the sediment traps from theSargasso Sea Arabian Sea and Bay of Bengal are from Stoll et al (2007ad) The numbers in X axis are related with the locations as follow 1=EMED 48B (ST) high productivity earlyspringndashsummer bloom 2=EMED 48A (ST) high productivity early springndashsummer and winter blooms 3=EMED 48A (ST) low productivity and mixing 4=EMED coretop (CT)5=Sargasso Sea (ST) high productivity fall bloom 6=Arabian Sea (ST) high productivity fall bloom 7=Arabian Sea (ST) low productivity intermonsoon 8=Bay of Bengal NBBTS(ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 9=Bay of Bengal NBBTS (ST) low productivity 10=EMED 48A (ST) high productivityearly springndashsummer bloom 11=EMED coretop (CT) 12=Arabian Sea (ST) high productivity SW monsoon-upwelling and NE monsoon mixing 13=Arabian Sea (ST) lowproductivity intermonsoon 14=Bay of Bengal NBTTS (ST) high productivity spring bloom and late summer to early fall eddy pumping-upwelling 15=Bay of Bengal NBTTS (ST)low productivity 16=EMED individual specimen 481B1 (ST) high productivity early summer bloom 17=EMED 48A (ST) high productivity early springndashsummer bloom18=EMED 48A (ST) low productivity and mixing 19=Sargasso Sea (ST) high productivity fall bloom
204 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
could be that recorded values ofC leptoporus SrCa ratios from sedimenttrapdonot resemble values of a real lowproductioneventdue tomixingand scavenging or that the sediment trap period is not typical of theaverage conditions over the time spanof hundreds of years integrated insurface sediments In addition SrCa ratios of H carteri and E huxleyishow constant values between sediment trap and surface sedimentsamples in the EMED We also take into consideration that the dataderived from these sediment traps are possibly from an annually highproductive period in comparisonwith other sediment trap studies in theEMED
Average SrCa ratios of EMED sediment traps are lower than thoseof all other locations during their high production period studied todate (Bay of Bengal Sargasso Sea and Arabian Sea) by a significantmargin of ~04 mmolmol lower for C leptoporus ~03 mmolmollower for H carteri and ~08 mmolmol for E huxleyi
6 Conclusions
The spring bloom period (March 2006 - period C) in the EMED ischaracterized by evidence of both high production and high exportHigh rates of production in surface waters are indicated by highchlorophyll contents and high SrCa in individually picked coccolithsin the sediment traps and strong export is indicated by highcoccosphere and coccolith fluxes to sediment traps In addition thepotential effectiveness of coccoliths as mineral ballast is shownmainly in periods A (JunendashJuly 2005) and C (March 2006) wherecorrelation between coccospheres and organic carbon fluxes is wellestablished For the period A and period B (December 2005ndashearlyFebruary 2006) we suggest a combination of wind speed gyre andeddies wind-generated circulation SSTs differences water columnmixing and sea bottom topography as factors controlling seasonal
productivity in the EMED sub-basin SrCa maxima in E huxleyi and Cleptoporus generally follow maxima in coccosphere export andinferred production maxima (high export processes) More attenuat-ed signals are recorded in the deeper 48A sediment trap (low exportprocesses and scavenging event)
H carteri and C leptoporus SrCa ratios in the EMED are higher thanthose of E huxleyi found in sediment trap samples an offset consistentwith sediment traps studied elsewhere SrCa ratios in C leptoporus inEMED range from 26 to 31mmolmol which is lower than that foundin sediment traps from more productive regions like the Arabian Seaand Sargasso Sea These results corroborate that coccolith SrCa ratiois reflecting the oligotrophy of the EMED relative to other moreproductive environments and can be a tool to develop quantitativeestimations of past primary production
SrCa ratios of H carteri specimens with different thickness do notcorrespondwith abiogenic calcite precipitation because the average SrCa ratio values obtained from the different specimens are stillwithin therangeof primary calcificationvalues (~25 to ~27 mmolmol)while thelaboratory based abiogenic calcite SrCa ratio shows values of~01 mmolmol Moreover SrCa ratios of E huxleyi (population) sug-gest that secondary carbonate calcification does not occur in the EMEDwater column However regarding previous discovery of overgrownE huxleyi and Reticulofenestra spp in surface sediments careful at-tention should be paid to the selection of pristine samples for SrCa ratioanalysis of population and individually- picked coccoliths
Acknowledgements
We acknowledge Dick Kroon and Simon R Troelstra for sugges-tions and comments on this article We thank Alessandro Incarbonaand Sebastian Meier for their constructive comments and discussion
205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

205L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
to improve this manuscript We also thank Saskia Kars for SEManalysis The bathymetric map is created through the Ocean DataView packages httpodvawide (Schlitzer 2008) The satellite dataused in this study were acquired using the GES-DISC InteractiveOnline Visualization ANd aNalysis Infrastructure (Giovanni) as part ofthe NASAs Goddard Earth Sciences (GES) Data and InformationServices Center (DISC) This research is part of theMERF Project whichwas made possible thanks to the support from the European ScienceFoundation (ESF) under the EUROCORES Programme EuroCLIMATEthrough contract No ERAS-CT-2003-980490 of the European Com-mission DG Research Fp6 The MERF project (04 ECLIM Fp35) isfunded by the Netherlands Organisatie Voor WetenschappelijkOnderzoek (NWO Division for the Earth and Life Sciences85501086) Funding for ion probe analyses was provided by NSFResearch OCE-0424474 and EAR-0628336 to HM Stoll and afellowship to H Stoll and P Ziveri from the Spanish Ministry ofEducation cofunded by the European Social Fund Funding forcoccolithophores flux quantification analysis has been provided bythe 04EP59 ENTER Project of the European Union and the GeneralSecretariat for Research and Technology Greek Ministry of Develop-ment The sediment trap samples for this study were provided byHERMES project (European Commission FP6)
References
Beaufort L Heussner S 2001 Seasonal dynamics of calcareous nannoplankton on aWest European continental margin the Bay of Biscay Marine Micropaleontology43 27ndash55
Berman T Paldor N Brenner S 2003 Annual SST cycle in the Eastern MediterraneanRed Sea and Gulf of Elat Geophysical Research Letters 30 (5) 1261 doi1010292002GL015860
Bethoux J-P 1979 Budgets of the Mediterranean Sea Their dependence on the localclimate and on the characteristics of the Atlantic waters Oceanologica Acta 2157ndash163
Broerse ATC Ziveri P van Hinte JE Honjo S 2000 Coccolithophore exportproduction species composition and coccolithndashCaCO3 fluxes in the NE Atlantic(34 N 21 W and 48 N 21 W) Deep-Sea Research II 47 1877ndash1905
Carbo P Krom MD HomokyWB Benning LG Herut B 2005 Impact of atmosphericdeposition on N and P geochemistry in the southern Levantine basin Deep-SeaResearch II 52 3041ndash3053
Chiu T-Z Broecker WS 2008 Toward better paleocarbonate ion reconstructions Newinsights regarding the CaCO3 size index Paleoceanography 23 PA2216 doi1010292008PA001599
Crudeli D Young JR Erba E de Lange GJ Henriksen K Kinkel H Slomp CP Ziveri P2004 Abnormal carbonate diagenesis in Holocenendashlate Pleistocene sapropel-associated sediments from the Eastern Mediterranean evidence from Emilianiahuxleyi coccolith morphology Marine Micropaleontology 52 217ndash240
Dimiza MD Triantaphyllou MV Dermitzakis MD 2008 Seasonality and ecology ofliving coccolithophores in E Mediterranean coastal environments (Andros IslandMiddle Aegean Sea) Micropaleontology 54 (2) 159ndash175
Eker-Develi E Kideys AE Tugrul S 2006 Role of Saharan dust on phytoplanktondynamics in the northeastern Mediterranean Marine Ecology Progress Series 31461ndash75
Engel A Zondervan I Aerts K Beaufort L Benthien A Chou L Delille B Gattuso JP Harlay J Heermann C Hoffmann L Jacquet S Nejstgaard J Pizay MDRochelle-Newall E Schneider U Terbrueggen A Riebesell U 2005 Testing thedirect effect of CO2 concentration on a bloom of the coccolithophorid Emilianiahuxleyi in mesocosm experiments Limnology and Oceanography 50 (2) 493ndash507
Gotsis-Skretas O Pagou K Moraitou-Apostolopoulou M Ignatiades L 1999 Seasonalhorizontal and vertical variability in primary production and standing stocks ofphytoplankton and zooplankton in the Cretan Sea and the Straits of the Cretan Arc(March 1994ndashJanuary 1995) Progress in Oceanography 44 625ndash649
Guerzoni S Chester R Dulac F Herut B Loye-Pilot MD Measures C Migon CMolinaroli EMoulin C Rossini P Saydam C Soudine A Ziveri P 1999 The role ofatmospheric deposition in the biogeochemistry of theMediterranean Sea Progress inOceanography 44 147ndash190
Hamad N Millot C Taupier-Letage I 2006 The surface circulation in the easternbasin of the Mediterranean Sea Scientia Marina 70 (3) 457ndash503
Herut B Krom MD Pan G Mortimer R 1999 Atmospheric input of nitrogen andphosphorus to the SE Mediterranean sources fluxes and possible impactLimnology and Oceanography 44 (7) 1683ndash1692
Herut B Zohary T Krom MD Mantoura RFC Pitta P Psarra S RassaoulzadeganF Tanaka T Thingstad TF 2005 Response of East Mediterranean surface waterto Saharan dust on-board microcosm experiment and field observations Deep-SeaResearch II 52 3024ndash3040
Honjo S 1976 Coccoliths production transportation sedimentation Marine Micropa-leontology 1 65ndash79
Honjo S 1982 Seasonality and interaction of biogenic and lithogenic particulate flux atthe Panama Basin Science 218 883ndash884
Incarbona A Di Stefano E Sprovieri R Bonomo S Censi P Dinaregraves-Turell J SpotoS 2008 Vertical structure variability of the water column and paleoproductivityreconstruction in the central-western Mediterranean during the late PleistoceneMarine Micropaleontology 69 26ndash41
Karl D Letellier R Tupas L Dare J Christian J Hebel D 1997 The role of nitrogenfixationin biogeochemical cycling in the subtropical North Pacific Ocean Nature 398 57ndash61
Kleijne A 1991 Holococcolithophorids from the Indian Ocean Red Sea MediterraneanSea and North Atlantic Ocean Marine Micropaleontology 17 1ndash76
Knappertsbusch M 1993 Geographic distribution of living and Holocene coccolitho-phores in the Mediterranean Sea Marine Micropaleontology 21 219ndash247
Krom MD Kress N Brenner S Gordon LI 1991 Phosphorus limitation of primaryproductivity in the eastern Mediterranean Sea Limnology and Oceanography 36(3) 424ndash432
Krom MD Thingstad TF Brenner S Carbo P Drakopoulos P Fileman TW FlatenGAF Groom S Herut B Kitidis V Kress N Law CS Liddicoat MI MantouraRFC Pasternal A Pitta P Polychronaki T Psarra S Rassoulzadegan F SkjodalEF Spyres G Tanaka T Tselepides A Wassmann P Wexels Riser CWoodward EMS Zodiatis G Zohary T 2005 Summary and overview of theCYCLOPS P addition Langrangian experiment in the eastern Mediterranean Deep-Sea Research II 52 3090ndash3108
Malanotte-Rizzoli P Hecht A 1988 Large-scale properties of the Eastern Mediter-ranean a review Oceanologica Acta 11 323ndash335
Malanotte-Rizzoli P Manca BB Ribera dAlcala M Theocharis A Bergamasso ABregant D Budillon G Civitarese G Georgopoulus D Michelato A Sansone EScarazzato P Souvermezoglou E 1997 A synthesis of the Ionian Sea hydrogra-phy circulation and water mass pathways during POEMmdashPhase I Progress inOceanography 39 153ndash204
Malinverno E Corselli C Ziveri P de Lange GHuumlbnerA 2003a Coccolithophorid exportproduction and fluxes in the Ionian Basin eastern Mediterranean In TriantaphyllouMV (Ed) Coccolithophores 2003 Gaia Heraklion Crete Greece 64 pp
Malinverno E Ziveri P Corselli C 2003b Coccolithophorid distribution in the IonianSea and its relationship to eastern Mediterranean circulation during late fall toearly winter 1997 Journal of Geophysical Research 108 (C9) 8115 doi1010292002JC001346
Malinverno E Triantaphyllou MV Stavrakakis S Ziveri P Lykousis V 2009 Seasonaland spatial variability of coccolithophore export production at the South-Westernmargin of Crete (Eastern Mediterranean) Marine Micropaleontology 71 131ndash147
Milliman JD 1993 Production and accumulation of calcium carbonate in the oceanbudget of a nonsteady state Global Biogeochemical Cycles 7 (4) 927ndash957
Millot C 1999 Circulation in the western Mediterranean Sea Journal of MarineSystems 20 423ndash442
Moutin T Raimbault P 2002 Primary production carbon export and nutrientsavailability in western and eastern Mediterranean Sea in early summer 1996(MINOS cruise) Journal of Marine System 33ndash34 273ndash288
Ovchinnikov IM 1984 The formation of intermediate water in the MediterraneanOceanology 24 168ndash173
Pinardi N Masetti E 2000 Variability of the large scale general circulation of theMediterranean Sea from observations and modeling a review PalaeogeographyPalaeoclimatology Palaeoecology 158 153ndash173
Pinet RP 1992 Oceanography an introduction to the planet oceanus West PublishingCompany USA 571 pp
Poem group (Physical Oceanography of the Eastern Mediterranean) 1992 Generalcirculation of the Eastern Mediterranean Earth Science Reviews 32 285ndash309
Robinson AR Leslie WG Theocharis A Lascaratos A 2001 Mediterranean SeaCirculation Enciclopedia of Ocean Sciences Academia Press pp 1689ndash1706doi101006rwos20010376
Rutten A de Lange GJ Ziveri P Thomson J van Santvoort PJM Colley S CorselliC 2000 Recent terrestrial and carbonate fluxes in the pelagic eastern Mediter-ranean a comparison between sediment trap and surface sediment Palaeogeo-graphy Palaeoclimatology Palaeoecology 158 197ndash213
Sachs JP Repeta DJ 1999 Oligotrophy and nitrogen fixation during easternMediterranean sapropel events Science 286 2485ndash2488
Samtleben C Bickert T 1990 Coccoliths in sediment traps from the Norwegian SeaMarine Micropaleontology 16 39ndash64
Sarmiento J Herbert T Toggweiler JR 1988 Mediterranean nutrients balance andepisodes of anoxia Global Biogeochemical Cycles 2 (4) 427ndash444
Schlitzer R 2008 Ocean Data View httpodvawideSouvermezoglou Egrave Krasakopoulou E Pavlidou A 1999 Temporal variability in
oxygen and nutrients concentrations in the southern Aegean Sea and the straits ofthe Cretan Arc Progress in Oceanography 44 573ndash600
Stavrakakis S Chronis G Tselepides A Heussner S Monaco A Abassis A 2000Downward fluxes of settling particles in the deep Cretan Sea (NE Mediterranean)Progress in Oceanography 46 217ndash240
Steinmetz JC 1994 Sedimentation of coccolithophores In Winter A Sissier WG(Eds) Coccolithophores Cambridge University Press Cambridge pp 179ndash197
Stoll HM Schrag DP 2000 Coccolith SrCa as a new indicator of coccolithophoridcalcification and growth rate Geochemistry Geophysics Geosystems 1 (5) 1006doi1010291999GC000015
Stoll HM Ziveri P Shimizu N Conte M Theroux S 2007a Relationship betweencoccolith SrCa ratios and coccolithophore production and export in the ArabianSea and Sargasso Sea Deep-Sea Research II 54 581ndash600
Stoll HM Shimizu N Archer D Ziveri P 2007b Coccolithophore productivityresponse to greenhouse event of the PaleocenendashEocene Thermal Maximum Earthand Planetary Science Letters 258 192ndash206
206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675

206 L Auliaherliaty et al Marine Micropaleontology 73 (2009) 196ndash206
Stoll HM Shimizu N Arevalos A Matell N Banasiak A Zeren S 2007c Insights oncoccolith chemistry from a new ion probe method for analysis of individuallypicked coccoliths Geochemistry Geophysics Geosystems 8 (6) Q06020doi1010292006GC001546
Stoll HM Arevalos A Burke A Ziveri P Mortyn G Shimizu N Unger D 2007dSeasonal cycles in biogenic production and export in Northern Bay of Bengalsediment traps Deep-Sea Research II 54 558ndash580
Tesoriero AJ Pankow JF 1996 Solid solution partitioning of Sr+2 Ba+2 and Cd+2 tocalcite Geochimica et Cosmochimica Acta 60 l053ndashl1063
Triantaphyllou MV Dimiza M Krasakopoulou E Malinverno E Lianou VSouvermezoglou E in press Seasonal variation in Emiliania huxleyi coccolithmorphology and calcification in the Aegean Sea (Eastern Mediterranean) Geobiosdoi101016jgeobios200909002
Trianthaphyllou MV Ziveri P Tselepides A 2004 Coccolithophore exportproduction and response to seasonal surface water variability in the oligothrophicCretan Sea (NE Mediterranean) Micropaleontology 50 (1) 127ndash144
Turgoklu M 2008 Synchronous blooms of the coccolithophore Emiliania huxleyi andthree dinoflagellates in the Dardanelles (Turkish Straits System) Journal of theMarine Biological Association (United Kingdom) 88 (3) 433ndash441
Winter A 1982 Post-depositional shape modification in Red Sea coccoliths Micropale-ontology 28 (3) 319ndash323
Winter A 1985 Distribution of living coccolithophores in the California current systemSouthern California borderland Marine Micropaleontology 9 385ndash393
Young J Ziveri P 2000 Calculation of coccolith volume and its use in calibration ofcarbonate flux estimates Deep-Sea Research II 47 1679ndash1700
Ziveri P Thunnel RC 2000 Coccolithophore export production in Guaymas BasinGulf of California Response to climate forcing In Ganssen GM Wefer G (Eds)Deep-Sea Research Special Part II Topical studies in Oceanography Particle fluxesand their preservation in deep sea sediments vol 47 pp 2073ndash2100
Ziveri P Thunell RC Rio D 1995 Export production of coccolithophores in anupwelling region results from San Pedro Basin Southern California BorderlandsMarine Micropaleontology 24 335ndash358
Ziveri P Rutten A de Lange GJ Thomson J Corselli C 2000 Present day coccolithfluxes recorded in central eastern Mediterranean sediment traps and surfacesediments Palaeogeography Palaeoclimatology Palaeoecology 158 175ndash195
Ziveri P de Bernardi B Baumann K-H Stoll HM Mortyn G 2007 Sinking ofcoccolith carbonate and potential contribution to organic carbon ballasting in thedeep ocean Deep-Sea Research II 54 659ndash675




















