
Personality Traits As Predictors of Career Adaptability of ...
Upload
uni-freiburgCategory
view
5download
0
Not fo
r Dist
ribut
ion
9 Close-to-nature forest management in Europe
Compatible with managing forests as complex adaptive forest ecosystems?
Jürgen Bauhus, Klaus J. Puettmann and Christian Kühne
What is close-to-nature forest management?
Close-to-nature forest management (CTNFM) is a production system that devel-
oped from concepts of multifunctional forest management. The main historical
and philosophical roots of close-to-nature forest management include Möller
(1922), who first described the ‘Dauerwald’ (perpetual forest) approach as an
alternative to the conventional even-aged forest management focused on conifer
monocultures. Earlier, Karl Gayer (1886) had already promoted mixed uneven-
aged forests as an ecologically more stable alternative to even-aged monocultures.
Another important influence in the development of this system came from Leibun-
dgut in Switzerland, who emphasized that silvicultural practices should not be
forced into specific themes but instead should accentuate the natural development
of forests (Schütz, 2001). This agreed well with Möller’s suggestions to employ all
suitable forest structures in forest management, and not to carry out the Dauer-
wald approach in a rigid systematic way (Möller, 1922).
While the Dauerwald and later the close-to-nature movement still show that
their origin and applications were strongly influenced by practitioners who had to
address specific management situations and goals, Möller (1922) developed a con-
ceptual basis for this approach early on. His writings represent an early attempt
to describe what is now known as the ecosystem concept (Huss, 1990; Thomasius,
1996). It is important to note that Möller developed his ideas before ecosystem
ecology became of major interest to the science community. This explains what
now appears as an incomplete understanding of ecosystems in his writings. Philo-
sophically, Möller’s view of ecosystems as an organism suggests close linkages with
Clements’ view of ecosystems as meta-organism dynamics (Clements, 1936) and
with the Gaia hypothesis, which views the whole globe as a super-organism; both
of which imply top-down control mechanisms (Levin, 1998). This view expressed
itself in the idea of bringing together productivity, diversity and especially continu-
ity and stability of forest conditions. Möller and his followers believed that simul-
taneously emphasizing these attributes would provide forests with the capacity
for self-regeneration and self-regulation. While this appears to be philosophically
SW_545_Ch 9.indd 187SW_545_Ch 9.indd 187 10/22/2012 3:11:45 PM10/22/2012 3:11:45 PM

188 Jürgen Bauhus
Not fo
r Dist
ribut
ion
aligned with the concept of self-organization of complex adaptive systems, several
philosophical distinctions are already evident. For example, complexity science
suggests that self-organization originates from a bottom-up approach, rather than
from top-down control. Several other distinctions, such as the emphasis of close-
to-nature forest management on continuity and stability, rather than on system
dynamics as highlighted in complexity science, are discussed in detail below. In
addressing the title question, we will specifically focus on the characteristics of
complex adaptive systems (Chapters 1 and 2), including openness, heterogeneity
and diversity, hierarchy, uncertainty, self-organization and adaptation.
The management settings of the main proponents of CTNFM lead to an
emphasis on stability, productivity, diversity and continuity of forest conditions,
which resulted in attempts to integrate multiple forest management goals at small
spatial scales, ideally within individual forest stands. This is in contrast to a segre-
gated forest management approach, such as the TRIAD concept, which comprises
separate areas in the landscape for wood production and biodiversity conservation
functions, in addition to areas of less intensive, multiple use forestry (Seymour and
Hunter, 1992). The attempt to simultaneously integrate multiple forest functions
at smaller scales, typically forest stands, has led to an emphasis on mixed-spe-
cies and uneven-aged forests across the landscape as a way to provide for wood
production, recreation, biodiversity, aesthetic values, and so on. More recently, a
stronger consideration of conservation goals has prompted the integration of per-
manent or semi-permanent structural elements of old forests at the stand level (e.g.
Gustafsson et al., 2012). Whereas the emphasis on managing forest at small spatial
scales and on maintaining mixed-species and uneven-aged forests everywhere may
appear to lead to homogenous forest landscapes, the situation in Central Europe
today is quite variable, due in part to the diversity in land ownership and manage-
ment histories.
Pro Silva, a non-governmental organization, serves as an umbrella for national
non-governmental organizations promoting close-to-nature forest management in
Europe. Pro Silva is now 23 years old, but its ideas and several national organi-
zations are several decades older. For a long time, the close-to-nature approach
was only practiced by a minority of mostly private landowners. Their main goals
of providing a stable supply of high value wood from small stands holdings had a
strong influence on the development of CTNFM practices. The large windstorms
in the early 1990s, in combination with public pressures concerned with biodi-
versity issues and budget limitations, raised interest in alternative management
regimes, specifically in CNTFM. Other ownerships with a wider set of manage-
ment settings and goals subsequently endorsed close-to-nature, or a derivative
thereof, as their official management approach (Hockenjos, 1999).
Members of Pro Silva agree to management that is guided by well-accepted
principles; see, for example, Pro Silva (1999). In addition to these private hold-
ings, many public forest owners, including city and state forests, have based their
management on principles of CTFNM (e.g. Lower Saxony Ministry for Nutrition,
Agriculture, Consumer Protection and Regional Development, 2007). Today,
CTNFM is a prominent paradigm in central Europe, shared by many forest
SW_545_Ch 9.indd 188SW_545_Ch 9.indd 188 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 189
Not fo
r Dist
ribut
ion
owners as well as governmental and environmental organizations (e.g. NABU,
2008). Its principles form the basis for certification standards of the Forest Steward-
ship Council (FSC) as well as the Programme for the Endorsement of Forest Certi-
fication Schemes (PEFC) for Central Europe. In addition, a common assumption
by many forest practitioners, forest administrations and policy makers in central
Europe is that CTNFM is the most suitable approach to cope with future climate
change (e.g. Reif et al., 2010).
Due to its origin and wide-spread application, CTNFM cannot be regarded as
an approach with a single commonly agreed definition and a well-defined, estab-
lished scientific basis (e.g. Pommerening and Murphy, 2004). Instead, it is defined
by a set of general principles that are derived from the common goal of managing
for high value wood production. The widespread application of these principles
accommodates local conditions and challenges, as different managers and agen-
cies will emphasize different principles in different situations. Here, we provide
a short description of the main principles and rationales of CTNFM, discuss the
strength and limitations of these, and analyze how these principles relate to the
concept of managing complex adaptive systems (see Table 1).
The principles and rationales of close-to-nature forest management
Forest development types and site adapted species composition
Description. The species composition of forest stands is commonly planned on the
basis of so-called forest development types. Based on the spatially explicit analysis
of site conditions, the desired forest functions (e.g. high value wood production, soil
conservation, water protection, recreation) or conservation status, one or multiple
forest development types are selected as target stand conditions, which are assumed
to provide the desired future forest functions (e.g. Niedersächsische Landesforsten,
2004). Forest development types serve as guides for future silvicultural activities
to direct the development of forest stand condition towards desired states, typi-
cally defined as fully stocked, multi-age, mixed species stands (Larsen and Nielsen,
2007). Forest development types are comprised of site adapted tree species with
physiological niches compatible with current or predicted future site conditions
(based on climate change predictions), high growth and value potential and little
or acceptable production risks (Frischbier et al., 2010). Depending on landowner
objectives, stand composition may comprise exclusively native species based on the
potentially natural vegetation. Alternatively, it may include a variable amount of
non-native or exotic species to promote the production function, or – increasingly
important – to promote the resistance of forests to future stresses and disturbances.
For example, forest development types for sub-montane sites in central Europe at
around 500 m elevation with approximately 800 mm precipitation and acidic soils
could include a) a highly natural, native European Beech (Fagus sylvatica) forest, with
few or no admixed species, b) a mixed sessile oak (Quercus petraea)-beech forest, or c)
SW_545_Ch 9.indd 189SW_545_Ch 9.indd 189 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM
190 Jürgen Bauhus
Not fo
r Dist
ribut
ion
a mixed beech-conifer forest, where conifer species could include larch, Douglas fir,
Norway spruce or Silver fir. The tree species choice may be based on observations
of species performance in adjacent stands and empirical knowledge about compat-
ibility of species in mixtures in terms of competition for light and other resources.
The selected forest development types and target stand conditions are designed to
fully use the site potential without deteriorating site conditions, such as through lit-
ter of low decomposability which may acidify the surface soils. Ideally these species
should have the potential to regenerate naturally under the range of acceptable
stand conditions to facilitate the use of natural processes for future stand manage-
ment and maintenance of ecosystem functions (Röhrig et al., 2006).
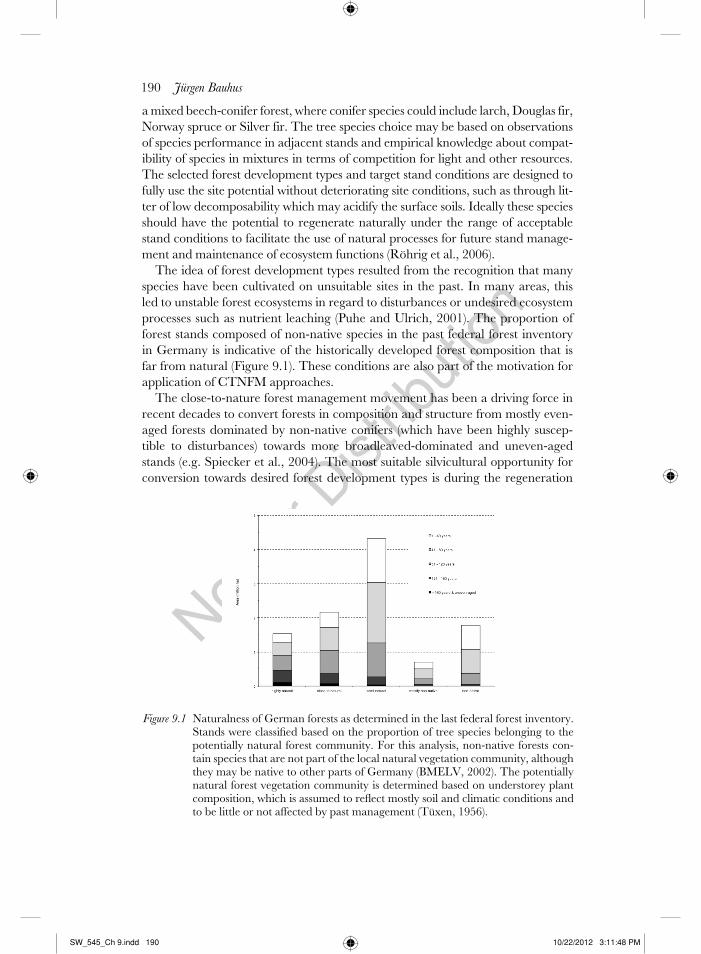
The idea of forest development types resulted from the recognition that many
species have been cultivated on unsuitable sites in the past. In many areas, this
led to unstable forest ecosystems in regard to disturbances or undesired ecosystem
processes such as nutrient leaching (Puhe and Ulrich, 2001). The proportion of
forest stands composed of non-native species in the past federal forest inventory
in Germany is indicative of the historically developed forest composition that is
far from natural (Figure 9.1). These conditions are also part of the motivation for
application of CTNFM approaches.
The close-to-nature forest management movement has been a driving force in
recent decades to convert forests in composition and structure from mostly even-
aged forests dominated by non-native conifers (which have been highly suscep-
tible to disturbances) towards more broadleaved-dominated and uneven-aged
stands (e.g. Spiecker et al., 2004). The most suitable silvicultural opportunity for
conversion towards desired forest development types is during the regeneration
Figure 9.1 Naturalness of German forests as determined in the last federal forest inventory. Stands were classifi ed based on the proportion of tree species belonging to the potentially natural forest community. For this analysis, non-native forests con-tain species that are not part of the local natural vegetation community, although they may be native to other parts of Germany (BMELV, 2002). The potentially natural forest vegetation community is determined based on understorey plant composition, which is assumed to refl ect mostly soil and climatic conditions and to be little or not affected by past management (Tüxen, 1956).
SW_545_Ch 9.indd 190SW_545_Ch 9.indd 190 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 191
Not fo
r Dist
ribut
ion
phase. Conversion practices often utilize advanced natural regeneration, if desir-
able species are present, or advance planting, to introduce desirable species. How-
ever, CTNFM also gained wide-scale recognition as an opportunity to respond
to unplanned natural disturbances. After several large-scale storms in the past
three decades, capacity and resources were insufficient for artificial regeneration
of many disturbed sites. Following a CTNFM approach, spontaneous successional
development of forests was permitted in many situations, even though this led
to species compositions in the natural regeneration that did not conform to the
intended forest development types (e.g. Schmidt-Schütz and Huss, 1998).
Critique of the principle and relationship to complex systems theory. The use of
site-adapted (rather than native) species is also discussed as an important strategy
to allow forests to adapt to climate change. However in this discussion, forest man-
agers focus more on adaptation to past and current site conditions and disturbance
regimes than on predictions of what will happen in the future (see also Figure 1.2 in
Chapter 1). This strategy is opposed by many environmental organizations, which
promote the exclusive use of native species (Reif et al., 2010). Their main argu-
ment is based on the importance of co-evolution; the idea being that native spe-
cies should have developed tighter ecological interactions with other forest organ-
isms than introduced species (Southwood, 1961). Such tight ecological coupling is
believed to favour resistance and adaptability to unexpected changes or extreme
events. However, history is telling a different story. There are no documented cases
of species being lost (or similar changes) that can be attributed to an introduced
tree species per se. In some cases, native species appear to benefit from their interac-
tions with introduced species (Goßner and Ammer, 2006) and introduced species,
such as Douglas-fir, are expected to better cope with increased drought events in
the future. On the other hand, there is ample evidence of native species struggling
to deal with the introduction of pest species or pathogens such as Ulmus sp. being
affected by Dutch Elm disease, Castanea sp. through chestnut blight, or currently
Fraxinus excelsior through the fungus Chalara fraxinea. Introduced tree species are,
however, not immune to introduced pests, such as Pinus strobus in Europe being
affected by white pine blister rust (McDonald and Hoff, 2001; Kirisits, 2007). Up
to now, there does not seem to be any scientific evidence that sole reliance on
native species will offer advantages with regard to potential adaptations to climate
change or to the supply of ecosystem services, compared with exotic species. Any
species may have a greater capacity to resist or recover quickly from unexpected
or extreme events, if these have been part of their historical natural disturbance
regime. Clearly, the perceived great advantage of native species is that they har-
bour more native biodiversity than exotics, and hence provide for more ecological
interactions within the ecosystem, but that may not always be the case (Goßner
and Ammer, 2006; Quine and Humphrey, 2010). Clearly, the decision about the
use of exotic tree species in the context of CTNFM will be a compromise between
landowner constraints and objectives, and political considerations.
The principle of using site-adapted species is primarily focused on local site
conditions and thus on the spatial scale of individual stands. It does not explicitly
consider landscape conditions. At the same time, in the long-term planning efforts
SW_545_Ch 9.indd 191SW_545_Ch 9.indd 191 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

192 Jürgen Bauhus
Not fo
r Dist
ribut
ion
of large forest owners such as state forests, this approach is guided by targets for
regional proportions of different tree species or forest types (e.g. Niedersächsische
Landesforsten, 2004). In practice, such regional level planning does account only
to a small extent for the goals and practices of other landowners, and thus cannot
provide specific goals regarding species composition and diversity of actual or for-
est development types at landscape scales.
The focus on local site conditions matches the emphasis on bottom-up regulation
of ecosystems as highlighted in complexity science. But, as described above, there
is no emphasis in CTNFM on cross-scale interactions, an important principle of
complex systems; for example, there is no tight linkage between local and landscape
or regional targets of species composition. The notion of forest development types
is nicely linked to the concept of self-organization, if the development types are
viewed in the broadest context, allowing for maximum flexibility. An example for
this can be seen in the reliance on spontaneous natural regeneration and succession
on disturbed sites following wind throw. However, reflecting the management set-
tings and goals of the landowners practicing CTNFM, the emphasis on target stand
structures associated with development types is not necessarily aligned with the
notion of adaptability and self-organization, which are important characteristics
of complex adaptive systems. Fully stocked uneven-aged and mixed-species stands
may have many advantages, but focusing on maintaining these conditions continu-
ously throughout the landscape suggests that landowners’ approach to achieving
their management goals is based (at least partially) on assumptions underlying a
static view of ecosystems. This contrasts with the dynamic view of complexity sci-
ence, often expressed as ‘stability is the exception, rather than the rule’.
Avoidance of clearfelling
Description. The avoidance of clearfelling and the focus on continuity of ecosystem
processes throughout forest developmental stages highlights the linkage of close-
to-nature forest management to other silvicultural approaches, especially continu-
ous cover forestry (Mason et al., 1999). Möller’s concept of forests as ‘organisms’
– that all elements are top-down controlled, interconnected and together provide
for a continuity of ecosystem processes – stood in strong contrast to clearfelling
practices common at the time, with a subsequent simplification in forest structure
and dominant functions and processes (e.g. Schütz et al., 2012). He linked the poor
growth of pine monocultures to the practice of clearfelling, which presumably led
to reductions in soil organic matter content in the sandy soils of north-eastern
Germany (Möller, 1922). Subsequently, this largely empirical knowledge has been
substantiated by many other studies that have demonstrated the potentially strong
impact of clearfelling on reducing ecosystem nutrient pools and fluxes (e.g. Swank
and Crossley, 1988), although the magnitude of this effect can be highly variable.
In addition, clearfelling is avoided under CTNFM for a number of other reasons,
such as the objective of shifting the focus from optimizing stand values to optimiz-
ing tree values. Given a range of tree sizes in even the most homogenous stand,
clearfelling results in individual trees being harvested prematurely (trees at the
SW_545_Ch 9.indd 192SW_545_Ch 9.indd 192 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 193
Not fo
r Dist
ribut
ion
smaller end of size distributions) or post maturity (trees at the larger end of size dis-
tributions), that is, before or after trees have reached their optimal dimension and
value, respectively. Also, one aim of CTNFM is to produce and maintain valuable
timber in harvestable dimensions continuously in every stand. In addition, the
avoidance of clearfelling is associated with a reliance on natural regeneration and
associated savings in planting costs. The retained overstorey following harvesting
is presumed to have a positive influence not only on survival but also on the quality
development of advance regeneration (form and branch size). While this is gener-
ally observed for conifer advance growth, it is questionable whether such canopy
shading has a similarly beneficial influence on quality development in hardwood
regeneration (Leonhardt and Wagner, 2006). Thus, while, technically, large open-
ings can be allowed under special conditions, the obvious bias against them (e.g.
evident in expressions, such as continuous cover forestry) has meant that they are
practically never implemented.
Partially as a result of the CTNFM movement, many German states have imple-
mented forest laws that prohibit clearfelling. Varying by state, clearfellings are
defined as units larger than contiguous areas of 0.3 to 2.0 ha, where stocking vol-
umes are reduced to less than 40% of a fully stocked stand (Klose and Orf, 1998).
Clearing sites at the scale of clearfells is only permitted as salvage operation and to
‘clean up’ following disturbances, or where it serves the conservation of habitat or
species. These new laws encouraged landowners to limit harvesting operations to
partial harvests, including small gaps, strips and shelterwood cuttings.
Critique of the principle and relationship with complex systems
theory. Concerns have been voiced that cutting small-scale gaps as dominant
harvest operations to initiate regeneration will eventually reduce the proportion of
light demanding tree species, such as oaks (c.f. von Lüpke, 2008). These species are
of special interest for nature conservation, future economic interests and for their
potential to adapt to future climatic conditions (e.g. von Lüpke, 2004).
From the perspective of complexity science, the avoidance of clearfelling, which
is a strong and central principle of CTNFM, shows an emphasis on memory and
legacies. The retention of forest structures during forest harvesting has evolved as
a central concept of ecologically sustainable forest management (e.g. Gustafsson
et al., 2012). It helps ensure the maintenance of structural heterogeneity and eco-
logical interactions on site throughout the regeneration phase of forests, such as
in the form of mycorrhizal networks (Chapter 7). However, limiting regeneration
options to favour a selective set of mostly shade tolerant species has the potential
to decrease species diversity in the long term. Such trends in central European
forests have been attributed to the continuous maintenance of mid- to late-suc-
cessional conditions and a lack of diversity of anthropogenic disturbances (e.g.
Wohlgemuth et al., 2002, see also Swanson et al., 2010 for a similar discussion
in North America). It has been suggested that a greater variety of management
practices is likely to provide for greater biodiversity, especially if they are designed
to provide niches for a wider variety of species (Bunnell and Huggard, 1999;
Wohlgemuth et al., 2002). In the context of complexity science, these management
SW_545_Ch 9.indd 193SW_545_Ch 9.indd 193 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

194 Jürgen Bauhus
Not fo
r Dist
ribut
ion
limitations imposed by CTNFM appear to conflict with the notion of maintaining
or increasing the ability of forest ecosystems to adapt to changes in future condi-
tions, because species that depend on regular disturbance (wind-dispersed species,
long-lived seed bank species, resprouters) may be lost. An important question in
this context is whether avoidance of clearfelling in CTNFM actually influences the
extent, intensity and severity of natural disturbances and the ability of forests to
respond. If CTNFM does not reduce the occurrence and severity of natural dis-
turbance events, then the combination of partial harvests and natural disturbances
may still provide ample opportunities for a diversity of self-organization processes.
However, these will depend on how disturbed sites are treated silviculturally; for
example, CTNFM approaches encourage maintenance of early successional spe-
cies, when target species are planted or established as part of accelerating stand
development towards target structures.
Focus on ecosystem stability
Description. The popularity of close-to-nature forest management in Germany
increased considerably during the public debate about forest decline related to
acid rain and other atmospheric pollution in the 1980s (Schmidt, 2009) and fol-
lowing the large storm events of the early 1990s (Hockenjos, 1999). The large
research efforts triggered by the forest decline improved the understanding of for-
est ecosystems considerably. Specifically, it expanded the concept of ecosystem
stability to include development of plant and faunal communities and processes,
such as carbon and nutrient cycling (Puhe and Ulrich, 2001). Further, research
demonstrated that highly cultivated stand types created under traditional even-
aged management (i.e. large-scale coniferous monocultures) were very productive
in terms of forest biomass growth (Spiecker et al., 1996), but were also particu-
larly vulnerable to storms (Schütz et al., 2006) and predicted climatic extremes
(e.g. Kölling et al., 2009), or even destabilized the systems through nutrient
leaching and accelerated soil acidification (Puhe and Ulrich, 2001).
Ecological stability in the context of CTNFM relates mostly to factors that
are currently or predicted to become the most important disturbances in Cen-
tral Europe, including wind storms (Schelhaas et al., 2003), insect outbreaks, in
particular bark beetles feeding in coniferous trees (Jönsson et al., 2011) and snow
damage (Nykänen et al., 1997). These disturbances typically interact with each
other; for example, bark beetles often follow wind or snow damage in Norway
spruce or Scots pine forests (Bakke, 1983). The development and maintenance
of ecologically stable forests focuses, therefore, on the physical stability of trees
and stands and on the reduction in the susceptibility to insect outbreaks. As is
described in Chapter 1, ecological stability is closely linked to resistance, resilience
and adaptability. The former refers to the ability of the ecosystem to withstand
the forces of disturbances and to maintain structure, composition and processes,
that is, to minimize disturbance severity. Silvicultural approaches as elements
of CTNFM aiming to increase resistance typically employ the use of compara-
tively ‘storm-resistant’ tree species. Also, tending schemes are specifically aimed
SW_545_Ch 9.indd 194SW_545_Ch 9.indd 194 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 195
Not fo
r Dist
ribut
ion
at developing forest stands with a high collective (stand) and individual tree stabil-
ity (Klädtke and Abetz, 2010). For example, mixed-species forests that include
deep-rooted species are more wind-firm than monocultures of shallow-rooted spe-
cies (Knoke et al., 2008). However, this increased stability may be only detectable
when assessing whole stands and is not necessarily found for shallow-rooted trees
within mixed stands (von Lüpke and Spellmann, 1999). Collective stand stability
is further pursued through spacing and thinning practices aimed at reducing the
height at which certain target diameters are being reached. Tree height has been
identified as the most important single predictor of wind damage in forests (e.g.
Albrecht et al., 2012). Hence, reducing the length of production cycles is viewed
by forest practitioners as an important element of silvicultural strategies to avoid
or minimize storm damage as well as other risks (Reif et al., 2010). Consequently,
managed forest stands are likely to become increasingly young at the time of final
harvest. The production cycle is further reduced through recent trends showing
improved growth of forests in Central Europe (Spiecker et al., 1996). However,
reduced production cycles and consequently lower stocking volumes contrast with
other goals of CTNFM and nature conservation NGOs (Reif et al., 2010), which
emphasize the values of fully stocked and older stands. Therefore, reduction in
production cycle length is contested amongst followers of CTNFM.
The preferred methods in CTNFM to increase ecological stability with regard
to insect pest outbreaks focus on development of mixed-species stands, increasing
diversity and abundance of antagonists to pest species, and reducing the rate of
spread of pests (e.g. Jactel and Brockerhoff, 2007; Dulaurent et al., 2012), as well
as on modifications in stand structures that create less favourable microclimatic or
soil conditions for pest species (e.g. Heiermann and Schütz, 2008).
An interesting question is whether forests managed under CTNFM principles
have been less affected by disturbances. Large-scale disturbances have certainly
been a major factor shaping forests in Central Europe (Scheelhas et al., 2003).
For example, in the German State of Baden-Württemberg, the average amount
of unplanned harvesting (largely from storms and insect outbreaks) in state forests
typically constitutes more than one third of the total harvested volume. However,
for the same region, single tree selection forests (Plenterwald) have been affected
by disturbances to a much smaller degree (Lenk and Kenk, 2007). The volume
of unplanned harvests after disturbances for these forests was 21% for the period
1953–2001, compared with 36% for all state forests. Of course, these figures
have to be viewed with caution. Single tree selection forests could easily be biased
towards sites that are less susceptible to disturbances and hence have permitted
this type of forest management in the past.
In the context of forest management, the notion of adaptability is typically dis-
cussed in terms of re-establishment of forest conditions and associated provision
of ecosystem services. For CTNFM this is often assessed as the (re-)development
of forests towards a desired tree species composition and size distribution. In this
regard, CTNFM places particular value on the continual presence of advance
regeneration, which normally survives the most common disturbances in central
Europe, such as wind throw and insects outbreaks.
SW_545_Ch 9.indd 195SW_545_Ch 9.indd 195 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

196 Jürgen Bauhus
Not fo
r Dist
ribut
ion
Critique of the principle and relationship with complex systems
theory. Despite the prominence of the central principle of close-to-nature for-
est management to develop or maintain forest stands that ‘are stable in the face
of biotic and abiotic disturbances’ (Davies et al., 2008), limitations of viewing
ecosystem as stable are well recognized within the CTNFM community (e.g.
Schütz et al., 2012). As viewed within CTNFM, this principle implies a heavy
reliance on resistance, with less emphasis on resilience, adaptability and realign-
ment (Stephens et al., 2010). In contrast, complexity science emphasizes the
importance of the latter two principles, as stability is considered the exception
rather than the rule in forest ecosystems. In this context, fully stocked uneven-
aged mixed stands can be viewed as a stable domain that is desirable and ideally
maintained on all sites. However, research in old-growth forests suggested that
it is only a temporary phase and only one component of a natural forest devel-
opment cycle (e.g. Leibundgut, 1978). Nevertheless, through repeated manage-
ment inputs and in absence of major disturbances, CTNFM efforts have been
quite successful in maintaining ecosystems in this phase for relatively long peri-
ods. Thus, much of the management activity in CTNFM is aimed at minimizing
variability in stand conditions at larger spatial scales, to ensure a continuity of
ecosystem services and economic gains at small spatial scales. In contrast, com-
plexity science suggests that there is much value in variability and changes which
are likely to favour what Holling and Gunderson (2002) called ‘creativity’ in for-
est dynamics. Especially extreme events provide ecosystems with opportunities
to adapt to new conditions (Holling and Gunderson, 2002). A discussion of the
panarchy cycle (for a detailed description, see Holling and Gunderson, 2002)
highlights this point. In this context, CTNFM can be viewed as an attempt to
prevent larger-scale release and reorganization periods through repeated inter-
vention at small scales. The avoidance of clearcutting and the associated amount
of residual trees after harvest operations can be viewed as an emphasis on lega-
cies and memory in a smaller nested cycle. If successful, CTNFM continuously
eliminates the ‘back loop’ (release and reorganization) phases, through mini-
mizing impacts of wind storms and insect outbreaks. However, in the panar-
chy cycle, the back loop is also considered important for maximizing ‘creative’
changes that are likely to increase the ecological resilience of forests to possible
future increase of such disturbances. As such, Holling and Gunderson (2002)
suggest that these processes are crucial for ecosystem’s ability to adapt to future
perturbations. Consequently, complexity science places a strong emphasis on
these phases, when external forces or variability can more easily influence eco-
systems, leading to novel combinations of components, including more extreme
events (Drever et al., 2006).
The brief discussion of the panarchy cycle highlights the issue about risks
associated with reductions in disturbance frequencies and intensities through
CTNFM (see above). However, whether CTNFM has been successful at reduc-
ing the occurrence and severity of disturbances, perhaps below the background
level of natural disturbances typical for a particular region, is extremely difficult
to answer.
SW_545_Ch 9.indd 196SW_545_Ch 9.indd 196 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 197
Not fo
r Dist
ribut
ion
Reliance on natural processes
Description. Close-to-nature forest management aims to minimize management
inputs and relies as much as possible on natural processes. This does not mean that
CTNFM approaches do not apply intensive management and active silvicultural
interventions. The reliance on natural processes is mainly focused on stages in
which silvicultural activities are not producing revenues, such as regeneration and
early stand development phases. For example, wherever possible, CTNFM relies
on natural regeneration, ideally of site-adapted species. Large-scale artificial regen-
eration through planting or sowing is generally avoided. Exceptions include the
conversion of stands dominated by species that are not site-adapted or to reforest
sites following large-scale disturbances. The last German federal forest inventory
revealed that, across all ownerships and management approaches, approximately
80% of forest stands in sapling stages or younger (up to 4 m tall trees) originated
from natural regeneration (BMELV, 2002). With the exception of forest domi-
nated by Douglas-fir and larch, natural regeneration was the most important form
of regeneration for all species, in particular in broadleaved forests.
Advantages of natural regeneration include primarily reduced establishment
costs and avoidance of problems due to distorted root systems, which may result
from planting of bare-root or container stock, in particular of species with tap
root systems (Nörr and Baumer, 2002). A further advantage of high densities and
multiple species in naturally regenerating stands is that natural selection processes
select for species and individuals best adapted to micro-site conditions (selection
filter of site conditions, e.g. drought years), which can be viewed as promoting
self-organisation of ecosystems. Also, natural regeneration established over long
periods from many parent trees ensures high genetic variation within seedling
populations (Finkeldey and Ziehe, 2004).
Under close-to-nature forest management, natural regeneration is mostly
achieved through reproduction methods that avoid large openings (Figure 9.2),
which favours a limited number of shade-tolerant species and discriminates
against light-demanding tree species (Wagner et al., 2011). The resulting extended
regeneration periods, which also conform with the aim to increase or maintain
resilience through the presence of advance growth (see above), may last up to four
decades in beech shelterwood systems or become a permanent feature in all-aged
Plenterforest. Extended establishment periods beneath the canopy of mature trees
protect seedling and saplings from frost and thus may reduce damages that lead to
reductions in tree quality, such as forking as result of bud damage (e.g. Kerr and
Boswell, 2001). The long regeneration periods, which are characterized by initially
fairly dark conditions in the understory, promote very shade-tolerant species such
as European beech (Fagus sylvatica) and Silver fir (Abies alba), which gain a growth
advantage over less shade-tolerant but (in full light) potentially more fast growing
species (e.g. Kühne and Bartsch, 2004).
As more landowners switched to natural regeneration of all tree species, it became
more and more obvious that herbivory by ungulates, mainly roe deer (Capreo-
lus capreolus) and red deer (Cervus elaphus), is a major factor limiting regeneration
SW_545_Ch 9.indd 197SW_545_Ch 9.indd 197 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

198 Jürgen Bauhus
Not fo
r Dist
ribut
ionsuccess (Ammer et al., 2010). The requirement that population densities of browsing
wildlife species should permit natural regeneration of forest tree species is stated in
most German state forest laws. However, this issue did not receive as much attention
when practices were focused on monocultures, often of species not very palatable to
deer and when investments in regeneration could be justified, such as browse pro-
tection. Thus, CTNFM revived the long-standing discussion about increasing hunt-
ing pressures to reduce populations of herbivores. Recently, this topic gained even
further attention, as the current browsing situation in German forests is considered
by many to be a strong impediment to forests adapting to climate change (Ammer
et al., 2010). Under current conditions, regeneration layers are often missing species
deemed more suitable to future climates (e.g. oaks, fir), as these are particularly sus-
ceptible to browsing (Gill and Beardall, 2001).
After regeneration establishment, natural processes (such as self-thinning and
self-pruning) may also have to be complemented by active stand tending to ensure
the maintenance of species diversity and removal of undesired species and trees
(e.g. wolf trees). The reliance on self-thinning and self-pruning is most common in
young hardwood stands (Röhrig et al., 2006). Self-thinning involves a strong selec-
tion process, and tree numbers can decline by orders of magnitude before the first
silvicultural thinning entry occurs. For example, numbers of seedlings in naturally
Figure 9.2 Long regeneration periods beneath the shelter of mature trees that are harvested based on target diameters promote advance regeneration of shade-tolerant spe-cies, such as Silver fi r and Norway spruce.
SW_545_Ch 9.indd 198SW_545_Ch 9.indd 198 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 199
Not fo
r Dist
ribut
ion
regenerated hardwood stands may decline from 100 000–200 000 germinants per
hectare to fewer than 1000 trees per hectare at 40 to 60 years, when the first crop
tree selection and associated thinning typically takes place (Pretzsch, 2009). Dur-
ing this process, asymmetric competition for light leads to size-dependent mortal-
ity of smaller, slower growing trees. Assuming that tree growth is related to genetic
fitness, self-thinning leads to selection for fitter plants and is expressed in higher
proportions of heterozygous individuals (e.g. Baker-Brosh, 1996). Hence, the reli-
ance of CTNFM on natural regeneration, often resulting in high seedling densities
and on self-thinning offers a large potential for natural selection to act and thus
promote adaptability of tree populations.
In contrast to hardwood stands, self-thinning in very dense, young conifer stands
is not commonly desirable. Here, high initial stand densities may lead to restricted
development of root systems, with long-lasting effects that may not even be rem-
edied by thinning, as has been shown for Norway spruce (Nielsen, 1990). Impaired
root development results in physically less stable trees, which is particularly critical
for shallow-rooted species, such as Norway spruce (Peltola et al., 2000). In addi-
tion, high densities in sapling and poles sized conifer stands may lead to smaller
branches, but not to improvement of stem quality through self-pruning (e.g. Hein
et al., 2008). Consequently, silvicultural approaches in conifer-dominated stands
do not typically rely on self-thinning, even in close-to-nature forest management,
but focus on promotion of tree and stand stability through spacing treatments
early in stand development.
Critique of the principle and relationship with complex systems
theory. The reliance on natural regeneration in the form of shade tolerant
advance growth restricts establishment and growth of many shade intolerant spe-
cies. In addition, exposure to root competition and shading through mature trees
may lead to restricted root development, with possible problems for the recovery
of structural roots in some species (Kühne et al., 2011). Consistent widespread
downplaying of the role of a large subset of species that potentially could be impor-
tant in determining the ecosystem’s response to disturbances or environmental
changes runs counter to the emphasis in complexity science on maintaining or
increasing the adaptive capacity of systems (Chapters 1 and 2). One may assume
that the likely recurrence of large-scale natural disturbances may provide ample
opportunities to promote these light demanding tree species and lead to land-
scapes with the full suite of species compositions (e.g. Schmidt-Schütz and Huss,
1998). However, this will only occur if seed-trees and propagules of all species have
been maintained within the landscape.
The strong reliance on natural processes is linked to the concept of self-organi-
zation (Camazine et al., 2001), an important characteristic of complex adaptive
systems. In the case of CNTFM, however, self-organizing principles are utilized
in a limited context. Clear boundaries defined by management objectives are evi-
dent, for example, from the different utilization of self-thinning in deciduous and
in coniferous stands. The need for deviation from reliance on natural processes
and self-organization, and thus the implementation of management practices such
SW_545_Ch 9.indd 199SW_545_Ch 9.indd 199 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

200 Jürgen Bauhus
Not fo
r Dist
ribut
ion
as planting, thinning or pruning, will be decided by an assessment of whether or
not the stands are developing towards desired target structures and compositions.
This highlights how the focus on desired target structures and compositions can
subsume other aspects important for complex adaptive systems. How decisions to
plant, thin or prune affect the ability of ecosystems to respond to a variety of distur-
bances or environmental changes receives less attention in this context.
The importance of the emphasis on natural processes in terms of the public
acceptance of CTNFM cannot be underestimated (e.g. Edwards et al., 2012) and
is an indication of forests as open systems that cannot be separated from their
social context (Olsson et al., 2004). The general public acceptance of CTNFM
is at least partially due to a general positive perception of ‘nature’. In addition,
the emphasis on natural processes implies less reliance on social factors to ensure
adaptability of forest ecosystems. This is expressed in less costly remedies and
fewer external inputs, such as herbicides or fertilizer. In addition, this approach
requires less tolerance from the public in terms of accepting unsightly forest condi-
tions, such as recently clearfelled areas or perfectly aligned trees in monoculture
plantations. CTNFM also has high acceptance and is promoted by environmental
organizations (e.g. NABU, 2008).
Focus on the development of individual trees
Description. This principle is expressed in silvicultural practices that focus on
individual trees, specifically to benefit future crop trees. For example, harvesting
decisions are made on a tree-by-tree basis to ensure that trees are cut as close
to target diameters as possible. This ensures maximum value increment. Target
diameters are typically a function of size premiums, penalties for large diameters,
size-dependent growth rates and production risks (such as potential pathogens or
pests that increase with tree size and/or age) (e.g. Zell et al., 2004). Thus, target
diameters represent a tree’s economic optimum or economic maturity, when the
value increment culminates. It is important to note that the relatively long produc-
tion cycles and large dimensions are partially a result of using low interest rates in
economic calculations by many landowners in central Europe (often zero or less
than 2% or 3 %).
Through focusing silvicultural practices on individual trees, forest stands become
less important as management units. Instead, silvicultural interventions may be
focused on tending blocks that comprise multiple adjacent stands of variable ages,
compositions and structures. Using tending blocks also saves costs by concentrat-
ing management activities in one part of forest management districts, such as har-
vesting, log grading, road maintenance, regeneration, monitoring and tending
treatments. Also, operations are concentrated, leaving other parts of the forests
undisturbed. Tending efforts to optimize diameter and quality development of
individual trees often include frequent thinning operations during selected phases
of tree and stand development (e.g. Röhrig et al., 2006). Thus, typical CNTFM
operations may use tending blocks that comprise one fifth of management districts,
resulting in five-yearly return intervals for tending and harvesting operations. The
SW_545_Ch 9.indd 200SW_545_Ch 9.indd 200 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 201
Not fo
r Dist
ribut
ion
frequent entries also highlight the importance of a good road infrastructure, espe-
cially in steeper terrain, which appears crucial for implementation of CTNFM.
Critique of the principle and relationship with complex systems
theory. The strong focus of CTNFM on the economic maturity of individual
trees has become a point of criticism. As has been pointed out, structural elements
important for wildlife habitat, such as habitat trees and standing or downed dead
wood, may be missing from forests managed under CTNFM, just as in forests
managed under the traditional even-aged monoculture paradigm (Bauhus et al.,
2009). Recent efforts push for increased integration of such structural elements
also in forests managed under CTNFM (Gustafsson et al., 2012, and see below).
However, the choice in scope and scale of these structural elements often appears
to be driven by specific wildlife habitat or biodiversity conservation concerns or
conditions found in nearby older forests. Whether these elements actually increase
the ability of ecosystems to respond to unexpected changes receives less attention.
For example, the size of retention patches may be driven by aspects of stability,
work safety, and so on, rather than ensuring that ecosystem processes are main-
tained at multiple spatial scales, which has been suggested as an important charac-
teristic of complex adaptive systems (Puettmann, 2011).
As mentioned above, in the absence of major disturbances, the avoidance of
clearfelling and associated use of single-tree or group cuttings leads to long regen-
eration periods. While this may allow many trees to participate in the regenera-
tion process (see above), it has also led to concerns that selection felling could lead
to a reduction in genetic variation and fitness, if the most vigorous trees with the
fastest diameter, which are harvested first, had no or only limited opportunities to
regenerate (e.g. Ziehe and Hattemer, 2002). However, the current understanding
of relationships between cutting systems, structural diversity of forests and genetic
diversity of tree species is not well developed. The general concern is that pref-
erential early removal of heterozygote individuals would affect allele frequencies
of uncommon alleles. These alleles, which are asymmetrically distributed, would
be discriminated against during target diameter harvesting and hence become
less frequent in the remaining stands, leading to lower frequencies of heterozy-
gote individuals in the following tree generation (Finkeldey and Ziehe, 2004). For
example, an inventory based on isozyme gene loci in Abies alba from Switzerland
and southern Germany showed slightly reduced levels of genetic variation in for-
ests managed under the CTNFM paradigm when compared with conventional
age-class forests (Konnert and Hussendörfer, 2001).
The focus on economic optimization of individual tree values highlights a shift
in scale of silvicultural interventions in close-to-nature forest management from
stands to smaller spatial scales, that is, single tree or small groups of trees (Schütz et
al., 2012). This is also reflected in various management practices, such as replacing
stand level rotation ages, which optimized the economic value of the whole stand,
with production cycles or target diameter harvesting. While the stand concept has
received much criticism (e.g. Puettmann et al., 2009), CTNFM has practically
replaced the focus on one spatial scale (stands) with that on another (trees). While
SW_545_Ch 9.indd 201SW_545_Ch 9.indd 201 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

202 Jürgen Bauhus
Not fo
r Dist
ribut
ion
smaller spatial scales inherently lend themselves better to bottom-up manage-
ment, complexity science strongly emphasizes the need for multi-scale approaches
(Chapters 1 and 2). Especially, concepts such as nonlinear feedback loops and
cross-scale linkages are hard to reconcile with the narrow focus on a single small
spatial scale. Further research is necessary to investigate whether optimizing value
and function at individual tree or small group scales will also result in optimal
accomplishment of stand and ownership or landscape level goals and especially
what the implications are for the adaptive capacity of ecosystems.
Mixed and uneven-aged, structurally diverse forests
Description. As mentioned above, recognition of the ecological value of mixed
species forests was one of the early motivations and inspirations for close-to-nature
forest management approaches. Since then, many studies in central Europe have
demonstrated that mixed species stands often have higher productivities than mono-
cultures and that they often offer greater economic and ecological stability (e.g.
Knoke et al., 2008; Pretzsch et al., 2010). Thus, this CTNFM principle and associ-
ated empirical knowledge and experimental evidence now have been integrated
in ecological research efforts investigating relationships between biodiversity and
ecosystem functioning. These efforts focus on two main hypotheses: a) ecosystem
productivity increases with the level of biodiversity and b) biodiversity enhances
ecosystem stability, which is interpreted as meaning that an increase in biodiver-
sity decreases the temporal variability of the provision of ecosystem services under
changing environmental conditions (Hooper et al., 2005; Loreau et al., 2002). The
latter is also called the ecological insurance concept and is based on the premise that
more diverse communities are better able to cope with new conditions when subject
to unpredictable stress or disturbance (Yachi and Loreau, 1999). For all these rea-
sons, the maintenance and further development of mixed-species forests is viewed
as one of the most important silvicultural strategies to facilitate forest ecosystems’
ability to adapt to climate change (Reif et al., 2010; Puettmann, 2011).
The general principles underlying productivity and stability of specific mixtures
in forestry are well researched, at least in two-species mixtures. However, great
uncertainty exists about the precise level and direction of interactions for specific
species combinations and site conditions (c.f. Pretzsch et al., 2010). For that rea-
son, silvicultural practices based on CTNFM (and other silvicultural approaches)
do not simply follow strategies to maximize tree species diversity, but rather aim
for specific mixtures of ‘compatible’ species, that is, selection and proportions of
species and their spatial distribution that maximize provision of ecosystem serv-
ices while minimizing tending efforts (Thomasius, 1996). For example, oaks and
other valuable hardwoods are cultivated with shade-tolerant, broadleaved trainer
species. Alternatively, conifers species (such as Norway spruce or Scots pine) are
mixed with shade tolerant broadleaves species to improve various aspects of bio-
diversity, nutrient cycling and productivity. Newer alternatives, such as cluster
plantings, provide for a long term mix of planted target and naturally regenerated
species (Figure 9.3).
SW_545_Ch 9.indd 202SW_545_Ch 9.indd 202 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

Close-to-nature forest management in Europe 203
Not fo
r Dist
ribut
ionForests managed according to CTNFM principles typically have a high struc-
tural diversity with regards to different tree size classes and canopy layers at small
spatial scales. This is a direct consequence of the approaches outlined above,
including target diameter cutting, promotion of advance regeneration and spe-
cies mixtures. Assuming that biotic and abiotic disturbance agents affect trees of
specific size classes differently, diversity in tree sizes may offer a similar ‘insurance’
as species diversity (Brang et al., 2012). For example, large, thick barked trees are
likely to be more resistant to fires but they are also more susceptible to wind throw
than small trees of the same species. Other elements of structural diversity, such
as a diversity of differently sized dead wood, may provide more niches for a wider
variety of species (McElhinny et al., 2005). Also, a diversity of microsites in diverse
structured forests, such as microsites receiving different amount of sunlight, may
offer more regeneration opportunities than would otherwise be available (e.g. Sze-
wczyk and Szwagrzyk, 1996). Hence, in structurally diverse forests, it is more likely
that trees surviving specific disturbances may provide more legacy elements that
will facilitate self-organization than in less structurally diverse forests.
Figure 9.3 Planting of widely-spaced clusters of trees, such as oaks, to permit spontaneous regeneration of diverse tree communities between the clusters promotes the self-organization potential of forests when compared with conventional row planting.
SW_545_Ch 9.indd 203SW_545_Ch 9.indd 203 10/22/2012 3:11:48 PM10/22/2012 3:11:48 PM

204 Jürgen Bauhus
Not fo
r Dist
ribut
ion
Critique of the principle and relationship with complex systems the-
ory. Increasing tree species’ diversity and stand structural attributes accords well
with the theory of complex systems of maintaining diverse and heterogeneous
elements to increase self-organization and adaptability of ecosystems. However,
much of the above considerations regarding mixing of species have been focused
on functional trait groups, that is, species traits that directly impact ecosystems
functions, such as timber production (Chapter 1). Less attention has been paid
to the mixing of species to maximize diversity of response type traits, meaning
traits that determine how species and ecosystems respond to perturbations, such
as different regeneration modes (with notable exceptions: Chapter 1; von Lüpke,
2009). This may be viewed as another indication of the relative importance of
economic factors under CTNFM and the underlying assumption of stable eco-
systems. In addition, the management of mixed-species stands has focused on
particular desired species combinations, deliberately excluding undesirable tree
species from the dynamic development of stands, and thus restricting the self-
organization of the system.
Another issue is the increasing pressure by conservation groups to restrict spe-
cies composition in CTNFM largely to native species (NABU, 2008). For example,
the enrichment of stands with non-native species, such as Douglas-fir (Pseudotsuga
menziesii) with a presumably high adaptive capacity towards future climatic condi-
tions, is discouraged (Reif et al., 2010). With the relatively low native tree species
diversity in central European forest, the desired adaptability to future conditions
may require a fresh look at introduced species. Clearly, assessing non-native tree
species as options to increase adaptive capacity of ecosystems will lead to chal-
lenges of the value of ‘naturalness’ under CTNFM (Reif et al, 2010).
Is close-to-nature forest management in central Europe compatible with managing forests as complex adaptive systems?
The question of whether close-to-nature forest management results in ecosystems
that are able to self-organize and adapt to changing conditions while providing the
desired ecosystem goods and services is at the heart of the previous discussion. It
is important to note that for a long time CTNFM was promoted by practitioners
without the benefit of a solid conceptual framework, such as is now available in
ecosystem ecology and complexity science. Clearly, many CTNFM principles and
practices, such as the emphasis on tree species diversity and structural heterogene-
ity at small spatial scales, can be viewed as an early attempt to apply principles that
are also embedded in complexity science (Chapters 1 and 2). Other aspects, such
as the emphasis on a limited set of spatial, temporal and hierarchical scales, the
focus on tree species diversity that emphasizes compatibility of growth pattern and
pays less attention to increasing the response type diversity (Chapter 1), as well as
limitations in terms of accepting the full suite of natural processes (such as larger-
scale disturbances), are not as closely aligned with characteristics and elements of
complex adaptive systems (Table 9.1).
SW_545_Ch 9.indd 204SW_545_Ch 9.indd 204 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

Close-to-nature forest management in Europe 205
Not fo
r Dist
ribut
ion
Differences in the view of hierarchical scales, an important concept in complex-
ity science, appear to show up in multiple dimensions of CTNFM. In complex-
ity science, the variety of scale – and especially cross-scale linkages and feedback
loops – are considered crucial for ecosystem adaptability; any silvicultural prac-
tice should be assessed on a variety of spatial and temporal scales using multiple
dimension, including ecological and social criteria. In contrast, most management
Table 9.1 Overview of the main principles of close-to-nature forest management and how they relate to characteristics of complex adaptive systems.
Principle Critique Relationship to complexity science
Site adapted Promotes ecological stability and Focus on local site conditions relatesspecies naturalness. Focus on native to bottom-up regulation and composition species may restrict adaptability self-organization of ecosystems. to future conditions/events. However, cross-scale interactions to the landscape level are largely ignored.Avoidance of Maintains structure and ecological Principle is in agreement withclearfelling processes of undisturbed forests emphasis on memory and legacies. (e.g. nutrient cycling). Facilitates If occurrence and severity of natural natural regeneration and focuses disturbance events are reduced, on development of individual trees. opportunities for self-organization Lack of disturbance may limit processes that require such large light-demanding tree species and disturbances are limited. reduce biodiversity. Focus on Reduced disturbances may Principle focuses on resistance andstability improve economic yields and less on resilience and adaptability provision of ecosystem services at of ecosystems. small spatial scales. Disturbance- dependent species may decline. Reliance on Reduces management inputs and Principle is linked to the concept ofnatural costs. Promotes natural selection self-organization, but clearprocesses processes. Improves public boundaries for stand development acceptance of forest management. are set by targeted stand structure and composition.Focus on Optimizing economic development Whereas smaller spatial scales lenddevelopment of individual trees. Reduced themselves better to bottom-upof individual regeneration opportunities for management, complexity sciencetrees shade-intolerant species and strongly emphasizes the need for reduced genetic diversity through multi-scale approaches. removal of fittest trees. Mixed and Higher productivities and greater Increasing tree species diversityuneven-aged, economic and ecological stability promotes diversity and heterogeneitystructurally than monocultures. that favours adaptive capacity ofdiverse forests ecosystems. Focus on functional types (e.g. compatibility of growth patterns), with less attention paid to response type traits, i.e. response to perturbations, limits adaptive capacity.
SW_545_Ch 9.indd 205SW_545_Ch 9.indd 205 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

206 Jürgen Bauhus
Not fo
r Dist
ribut
ion
activities under CTNFM are narrowly focused on trees and do not give the same
attention to larger spatial scales, such as a landscape or region.
For many management and conservation aspects, as well as elements of propa-
gation of pests and fire, these large-scale properties (such as distributions of tree
species and forest types, forest ages, areas under protection or conservation ease-
ments and viability of populations or species) are of particular importance. More
recently, such landscape considerations have also entered forest planning of large
public forest owners (Braunisch and Suchant, 2007) leading to substantial modi-
fications of close-to-nature forest management approaches; for example, the sub-
stantial reduction in stocking volumes in high elevations of the Black Forest to
increase habitat suitability for capercaillie.
A second aspect, which appears in the various sections above, is the accept-
ance of the inherent unpredictability of complex adaptive systems. On the one
hand, CTNFM embraces uncertainty and self-organization as it allows for natu-
ral processes to play out, such as natural regeneration and self-thinning. On the
other hand, CTNFM places firm boundaries on how to deal with uncertainty and
for the use and development of natural processes in managed forests, based on a
limited set of target stand structures and compositions, as evident in the emphasis
on forest development types and avoidance of larger-scale disturbances, such as
clearfelling.
Overall, the above discussion indicates that CTNFM already addresses many of
the characteristics of complex adaptive systems and has a great potential to benefit
from embracing complexity science as a conceptual framework. This framework
would allow researchers and foresters to assess how forest management practices
can be modified to improve ecosystems’ adaptive capacity and self-organization.
In the meantime, general claims about ecosystem adaptability should be viewed
with caution, such as claims that CTNFM is a suitable management option to pro-
vide for adaptation of forest to climate change (e.g. Schütz et al., 2012; see Brang
et al., 2012). These claims typically use traditional forest management as refer-
ence, and simply highlight the higher species and structural diversity at smaller
spatial scales without taking into consideration the highly uncertain future and the
ability of such an approach to be able to adapt to unknown future ecological and
socio-economic conditions. The apparent tradeoffs between adaptation to cur-
rent conditions and adaptability to future conditions (Puettmann, 2011; Chapter
1) suggest that a closer examination of CTNFM in terms of adaptability may be
beneficial in this context. Consequently, an analysis of ecosystem adaptability of
forests managed under CTNFM should be based on broad assessments that go
beyond specific goals and intentions of landowners and include landscape aspects,
such as natural disturbance patterns and the full suite of societal demands and
expectations.
Lastly, close-to-nature forest management poses numerous challenges as many
ecological interactions and processes and their responses to silvicultural treatments
and disturbances (such as climate change) are not well understood (Puettmann,
2011). The outcomes of any specific treatments or treatment intensities cannot
be predicted within tight boundaries. Hence, CTNFM with its relatively short
SW_545_Ch 9.indd 206SW_545_Ch 9.indd 206 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

Close-to-nature forest management in Europe 207
Not fo
r Dist
ribut
ion
history must be understood in the context of an adaptive management approach
that requires close observation and monitoring of outcomes and adjustment of the
approach as needed. Clearly, setting wider boundaries for future development of
forest ecosystems would probably provide more opportunities for diversification of
forests over the European landscapes. Less predictable development of forests and
landscapes, however, also require the willingness to adjust silvicultural concepts on
a continuous basis, and complexity science appears to provide a framework that
will facilitate this process.
Conclusion
Close-to-nature forest management (CTNFM) is widely held as an approach
that optimizes multiple forest functions at small spatial scales. Here we reviewed
the main principles of this approach, discussed their merits and limitations and
analysed to what extent they compared to principles expressed in complexity sci-
ence. The main principles of close-to-nature forest management reviewed here
comprise the use of site adapted tree species, development of mixed and uneven-
aged structurally diverse forests, avoidance of clearfelling, focus on stand stabil-
ity, reliance on natural processes and focusing on development of individual
trees. Many principles and practices of close-to-nature forest management, such
as the emphasis on diversity and structural heterogeneity, agree with principles
that are also embedded in complexity science. However, other aspects, such as
the emphasis on a limited set of spatial, temporal and hierarchical scales and
tight boundaries set for target structures and compositions, pay less attention
to increasing the response type diversity, and hence adaptability of forest eco-
systems are not as closely aligned with characteristics and elements of complex
adaptive systems. Although CTNFM embraces uncertainty and self-organiza-
tion as it allows for natural processes such as natural regeneration and self-thin-
ning, there is limited acceptance of the inherent unpredictability of complex
adaptive systems as evident in the emphasis on forest development types and
avoidance of larger-scale disturbances. We discuss such discrepancies in the
context of the different origins and goals of the two approaches. Our analysis
indicates that CTNFM could benefit from embracing complexity science as a
conceptual framework to assess the influence of its management practices and to
develop or adopt complementary approaches to promote the adaptive capacity
of forest ecosystems.
Acknowledgements
We are very grateful for the helpful and constructive suggestions for improve-
ment that we received for earlier versions of this chapter from Christian Messier,
Christian Ammer, Timo Kuuluvainen and Lluis Coll. Thanks to Ursula Eggert
for formatting the text and references. And we wish to thank the many colleagues
with whom we had the pleasure of discussing our views on close-to-nature forest
management and complex adaptive systems over the years.
SW_545_Ch 9.indd 207SW_545_Ch 9.indd 207 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

208 Jürgen Bauhus
Not fo
r Dist
ribut
ion
Literature Cited
Albrecht, A., Hanewinkel, M., Bauhus, J. and Kohnle, U. (2012) ‘How does Silviculture
affect Storm Damage in Forests of South-western Germany? Results from Empirical
Modeling based on Long-term Observations’, European Journal of Forest Research, 131, pp.
229–247.
Ammer, C., Vor, T., Knoke, T. and Wagner, S. (2010) Der Wald-Wild-Konflikt. Analyse und
Lösungsansätze vor dem Hintergrund Rechtlicher, Ökologischer und Ökonomischer Zusammenhänge,
Göttinger Forstwissenschaften 5, Universitätsverlag Göttingen, 175 pp.
Baker-Brosh, K.F. (1996) The Genetic Consequences of Self-Thinning in Two Populations of Loblolly
Pine (Pinus taeda L.), PhD thesis, The University of North Carolina, Chapel Hill, NC.
Bakke, A. (1983) ‘Host Tree and Bark Beetle Interaction during a Mass Outbreak of Ips
typographus in Norway’, Zeitschrift für Angewandte Entomologie, vol 96, pp. 118–125.
Bauhus, J, Puettmann, K. and Messier, C. (2009) ‘Silviculture for Old-growth Attributes’,
Forest Ecology and Management, vol 258, pp. 525–537.
BMELV (2002) Bundeswaldinventur 2. Bundesministerium für Ernährung, Landwirtschaft
und Verbraucherschutz, www.bundeswaldinventur.de.
Brang, P., Larsen, J.B., Spathelf, P., Bauhus, J., Boncina, A., Chauvin, C., Drössler, L.,
García-Güemes, C., Heiri, C., Kerr, G., Lexer, M., Mason, W.L., Mohren, F., Müh-
lethaler, U., Nocentini, S. and Svoboda, M. (2012) ‘Opportunities and Constraints of
Close-to-nature Silviculture as Adaptation Strategy to Climate Change’, European Journal
of Forest Research (in review).
Braunisch, V. and Suchant, R. (2007) ‘A Model for Evaluating the ‘Habitat Potential’ of a
Landscape for Capercaillie Tetrao urogallus: a Tool for Conservation Planning’, Wildlife
Biology, vol 13, pp. 21–33.
Bunnell, F.L. and Huggard, D.J. (1999) ‘Biodiversity across Spatial and Temporal Scales:
Problems and Opportunities’, Forest Ecology and Management, vol 115, pp. 113–126.
Camazine, S., Deneuborg, J.L., Franks, N.R., Sneyd, J., Theraulaz, G. and Bonabeau, E.
(2001) Self-Organization in Biological Systems, Princeton University Press: Princeton, NJ, 538
pp.
Clements, F.E. (1936) ‘Nature and Structure of the Climax’, Journal of Ecology, vol 24, pp.
252–284.
Davies, O., Haufe, J. and Pommerening, A. (2008) Silvicultural Principles of Continuous Cover
Forestry: A Guide to Best Practice, The Forestry Commission Wales, Bangor University,
School of the Environment and Natural Resources, Tyfiant Coed Project Report,
http://tyfcoed.bangor.ac.uk/BPG_final.pdf.
Drever, C.R., Peterson, G., Messier, C., Bergeron, Y. and Flannigan, M. (2006) ‘Can For-
est Management based on Natural Disturbances Maintain Ecological Resilience?’, Cana-
dian Journal of Forest Research, vol 36, pp. 2285–2299.
Dulaurent, A.M., Porté, A.J., van Halder, I., Vétillard, F., Menassieu, P. and Jactel, H.
(2012) ‘Hide and Seek in Forests: Colonization by the Pine Processionary Moth is
impeded by the Presence of Non-host Trees’, Agricultural and Forest Entomology, vol 14, pp.
19–27.
Edwards, D.M., Jay M., Jensen F.S., Lucas, B., Marzano M., Montagné, C., Peace A.
and Weiss, G. (2012) ‘Public Preferences across Europe for Different Forest Stand
Types as Sites for Recreation’. Ecology and Society, vol 17, no 1, article 27, http://dx.doi.
org/10.5751/ES-04520-170127.
Frischbier, N., Profft, I. and Arenhövel, W. (2010) ‘Thuringian Tree Species Recommen-
dations for Adaptation to Climate Change (in German)’, Forst und Holz 65, pp. 28–35.
SW_545_Ch 9.indd 208SW_545_Ch 9.indd 208 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

Close-to-nature forest management in Europe 209
Not fo
r Dist
ribut
ion
Finkeldey, R. and Ziehe, M. (2004) ‘Genetic Implications of Silvicultural Regimes’, Forest
Ecology and Management, vol 197, pp. 231–244.
ForstBW (ed) (2010) Alt- und Totholzkonzept Baden-Württemberg, Stuttgart, Germany, 37 pp,
www.fva-bw.de/publikationen/sonstiges/aut_konzept.pdf.
Gayer, K. (1886) Der Gemischte Wald – Seine Begründung und Pflege, Insbesondere Durch Horst- und
Gruppenwirtschaft, Paul Parey Verlag: Berlin.
Gill, R.M.A. and Beardall, V. (2001) ‘The Impact of Deer on Woodlands: the Effects of
Browsing and Seed Dispersal on Vegetation Structure and Composition’, Forestry, vol
74, pp. 209–218.
Goßner, M. and Ammer, U. (2006) ‘The Effects of Douglas-fir on Tree-specific Arthropod
Communities in Mixed Species Stands with European Beech and Norway Spruce’, Euro-
pean Journal of Forest Research, vol 125, pp. 221–235.
Gustafsson, L., Baker, S.C., Bauhus, J., Beese, W.J., Brodie, A., Kouki, J., Lindenmayer,
D.B., Lõhmus, A., Martínez Pastur, G., Messier C., Neyland, M., Palik, B., Sverdrup-
Thygeson, A., Volney, W.J.A., Wayne, A. and Franklin, J.F. (2012) ‘Retention Forestry to
Maintain Multifunctional Forests: a World Perspective’, Bioscience, vol 62, pp. 633–645.
Heiermann, J. and Schütz, S. (2008) ‘The Effect of the Tree Species Ratio of European
Beech (Fagus sylvatica L.) and Norway Spruce (Picea abies (L.) Karst.) on Polyphagous and
Monophagous Pest Species – Lymantria monacha L. and Calliteara pudibunda L. (Lepidoptera:
Lymantriidae) as an Example’, Forest Ecology and Management, vol 255, pp. 1161–1166.
Hein, S., Weiskittel, A.R. and Kohnle, U. (2008) ‘Effect of Wide Spacing on Tree Growth,
Branch and Sapwood Properties of Young Douglas-fir (Pseudotsuga menziesii [Mirb.]
Franco) in South-western Germany’, European Journal of Forest Research, vol 127, pp.
481–493.
Hockenjos, W. (1999) ‘Vom Forst zum Wald. Entwicklungstendenzen im naturnahen
Waldbau (From a production-oriented forest to a multipurpose forest)’, New Trends in
Near-Natural Silviculture, Schweizerische Zeitschrift fur Forstwesen, vol 150, no 12, pp. 484–488.
Holling, C.S. and Gunderson, L. (2002) ‘Resilience and Adaptive Cycles’, in L. Gunderson
and C.S. Holling (eds.) Panarchy: Understanding Transformations in Human and Natural Systems,
Island Press: Washington DC, pp. 25–62.
Hooper, D.U., Chapin, F.S.I., Ewel, J.J., Hector, A., Inchausti, P., Lavorel, S., Lawton,
J.H., Lodge, D., Loreau, M., Naeem, S., Schmid, B., Setälä, H., Symstad, A.J., Vander-
meer, J. and Wardle, D.A. (2005) ‘Effects of Biodiversity on Ecosystem Functioning: a
Consensus of Current Knowledge and Needs for Future Research’, Ecological Monographs,
vol 75, pp. 3–36.
Huss, J. (1990) ‘Die Entwicklung des Dauerwaldgedankens bis zum Dritten Reich’, Forst und
Holz, vol 45, pp. 163–176.
Jactel, H. and Brockerhoff, E.G. (2007) ‘Tree Diversity Reduces Herbivory by Forest
Insects’, Ecology Letters, vol 10, pp. 835–848.
Jönsson, A.M., Harding, S., Krokene, P., Lange, H., Lindelöw, A., Økland, B., Ravn,
H.P. and Schroeder, L.M. (2011) ‘Modelling the Potential Impact of Global Warming
on Ips typographus voltinism and Reproductive Diapause’, Climate Change, vol 109, pp.
791–825.
Kerr, G. and Boswell, R.C. (2001) ‘The Influence of Spring Frosts, Ash Bud Moth (Prays
fraxinella) and Site Factors on Forking of Young Ash (Fraxinus excelsior) in Southern Brit-
ain’, Forestry, vol 74, pp. 29–40.
Kirisits, T. (2007) ‘Der Stroben-Blasenrost’, Forstzeitung, vol 118, pp. 15.
Klädtke, J. and Abetz, P. (2010) Durchforstungshilfe 2010. Merkblätter der Forstlichen Versuchs- und
Forschungsanstalt Baden-Württemberg, Heft 53: Freiburg, Germany, 11 pp.
SW_545_Ch 9.indd 209SW_545_Ch 9.indd 209 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

210 Jürgen Bauhus
Not fo
r Dist
ribut
ion
Klose, F. and Orf, S. (1998) Forstrecht: Kommentar zum Waldrecht des Bundes und der Länder,
Aschendorff Münster, Germany, 746 pp.
Knoke, T., Ammer, C., Stimm, B. and Mosandl, R. (2008) ‘Admixing Broadleaved to
Coniferous Tree Species: a Review on Yield, Ecological Stability and Economics’, Euro-
pean Journal of Forest Research, vol 127, pp. 89–101.
Kölling, C., Knoke, T., Schall, P. and Ammer, C. (2009) ‘Überlegungen zum Risiko des
Fichtenanbaus in Deutschland vor dem Hintergrund des Klimawandels (Cultivation of
Norway Spruce (Picea abies (L.) Karst.) in Germany: Considerations on Risk against the
Background of Climate Change)’, Forstarchiv, vol 80, pp. 42–54.
Konnert, M. and Hussendörfer, E. (2001) ‘Genetic Variation of Silver Fir (Abies alba) in
Unevenaged Forests (‘Plenter’ Forest) in Comparison with Even-aged Forests (Altersklas-
senwald)’, in G. Müller-Starck and R. Schubert (eds.), Genetic Response of Forest Systems to
Changing Environmental Conditions, Kluwer academic publishers: Dordrecht, The Nether-
lands, pp. 307–320.
Kühne, C. and Bartsch, N. (2004) ‘Zur Naturverjüngung von Fichten-Buchen- Misch-
beständen im Solling’, Forst und Holz, vol 58, pp. 3–7.
Kühne, C., Bauhus J., Hörnig T. and Oh, S. (2011) ‘Effects of Canopy Closure, Crowd-
ing and Plant Size on Root System Development in Douglas-fir Saplings (in German)’,
Forstarchiv, vol 82, pp. 184–194.
Larsen, J.B. and Nielsen, A.B. (2007) ‘Nature-based Forest Management—Where are we
Going? Elaborating Forest Development Types in and with Practice’, Forest Ecology and
Management, vol 238, pp. 107–117.
Leibundgut, H. (1978) ‘Über die Dynamik europäischer Urwälder’, Allgemeine Forst und Jag-
dzeitung, vol 33, pp. 686–690.
Lenk, E. and Kenk, G. (2007) ‘Sortenproduktion und Risiken Schwarzwälder Plenter-
wälder’, AFZ/Der Wald, vol 23, pp. 136–139.
Leonhardt, B. and Wagner, S. (2006) ‘Qualitative Entwicklung von Buchen-Voranbauten
unter Fichtenschirm’, Forst und Holz, vol 61, pp. 454–457.
Levin, S.A. (1998) ‘Ecosystems and the Biosphere as Complex Adaptive Systems’, Ecosys-
tems, vol 1, pp. 431–436.
Loreau, M., Naeem, S. and Inchausti, P. (2002) Biodiversity and Ecosystem Functioning, Oxford
University Press: New York.
McDonald, G.I. and Hoff, R.J. (2001) ‘Blister Rust: an Introduced Plague’, in D.F. Tom-
back, S.F. Arno and R.E. Keane (eds.), Whitebark Pine Communities: Ecology and Restoration,
Island Press: Washington DC, pp. 193–220.
McElhinny, C., Gibbons, P., Brack, C. and Bauhus, J. (2005) ‘Forest and Woodland Stand
Structural Complexity: Its Definition and Measurement’, Forest Ecology and Management,
vol 218, pp. 1–24.
Mason, B., Kerr, G. and Simpson, J. (1999) What Is Continuous Cover Forestry? Forestry Com-
mission Information Note, October 1999, 8 pp.
Möller, A. (1922) Der Dauerwaldgedanke – Sein Sinn und Seine Bedeutung, Springer Verlag: Berlin,
136 pp.
NABU (2008) Waldwirtschaft 2020. Perspektiven und Anforderungen aus Sicht des Naturschutzes,
Strategiepapier, Berlin, www.nabu.de/imperia/md/content/nabude/wald/4.pdf.
Niedersächsische Landesforsten (2004) ‘Langfristige Ökologische Waldentwicklung – Rich-
tlinie zur Baumartenwahl. Aus dem Walde – Schriftenreihe Waldentwicklung in Nie-
dersachsen. No.54. Niedersächsisches Ministerium für ländlichen Raum, Ernährung,
Landwirtschaft und Verbraucherschutz, Hannover, Germany, 150 pp.
Niedersächsisches Ministerium für Ernährung, Landwirtschaft, Verbraucherschutz und
SW_545_Ch 9.indd 210SW_545_Ch 9.indd 210 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

Close-to-nature forest management in Europe 211
Not fo
r Dist
ribut
ion
Landesentwicklung (2007) Langfristige, ökologische Waldentwicklung in den Niedersächsischen
Landesforsten (LÖWE-Erlass), 21 pp.
Nielsen, C.C.N. (1990) Einflüsse von Pflanzenabstand und Stammzahlhaltung auf Wurzelform, Wur-
zelbiomasse, Verankerung sowie auf Biomassenverteilung im Hinblick auf die Sturmfestigkeit der Fichte,
Schriften aus der Forstlichen Fakultät der Universität Göttingen und der Niedersäch-
sischen Forstlichen Versuchsanstalt 100, Frankfurt am Main, Sauerländer, Germany.
Nörr, R. and Baumer, M. (2002) Pflanzung – ein Risiko für die Bestandesstabilität? Die Bedeutung
Wurzelschonender Pflanzung und ihre Umsetzung im Forstbetrieb, Berichte aus der Bayrischen
Landesanstalt für Wald und Forstwirtschaft 37, LWF, Freising, Germany, 58 pp.
Nykänen, M.L., Peltola, H., Quine, C.P., Kellomäki, S. and Broadgate, M. (1997) ‘Factors
Affecting Snow Damage of Trees with Particular Reference to European Conditions’,
Silva Fennica, vol 31, pp. 193–213.
Olsson, P., Folke, C. and Berkes, F. (2004) ‘Adaptive Comanagement for Building Resil-
ience in Social-ecological Systems’, Environmental Management, vol 34, pp. 75–90.
Peltola, H., Kellomäki, S., Hassinen, A. and Granander, M. (2000) ‘Mechanical Stability
of Scots Pine, Norway Spruce and Birch: an Analysis of Tree-pulling Experiments in
Finland’, Forest Ecology and Management, vol 135, pp. 143–153.
Pommerening, A. and Murphy, S.T. (2004) ‘A Review of the History, Definitions and
Methods of Continuous Cover Forestry with Special Attention to Afforestation and
Restocking’, Forestry, vol 77, pp. 27–44.
Puettmann, K.J., Coates, K.D. and Messier, C. (2009) A Critique of Silviculture: Managing for
Complexity, Island Press: Washington, DC.
Pretzsch, H. (2009) Forest Dynamics, Growth and Yield. From Measurement to Model, Springer:
Berlin and Heidelberg, 664 pp.
Pretzsch, H., Spellmann, H., Nagel, J., Kohnle, U., Zingg, A., Dong, P.H., Block, J. and
Dieler, J. (2010) ‘Comparison between the Productivity of Pure and Mixed Stands of
Norway Spruce and European Beech along an Ecological Gradient’, Annals of Forest Sci-
ence, vol 67, article 712.
Pro Silva (1999) Pro Silva Europe Policy http://prosilvaeurope.org/?q=about#Founding.
Puettmann, K.J. (2011) ‘Silvicultural Challenges and Options in the Context of Global
Change – ‘Simple’ Fixes and Opportunities for New Management Approaches’, Journal
of Forestry, vol 109, pp. 321–331.
Puettmann, K.J., Coates, K.D. and Messier, C. (2009) A Critique of Silviculture: Managing for
Complexity, Island Press: Washington DC.
Puhe, J. and Ulrich, B. (2001) Global Climate Change and Human Impact on Forest Ecosystems:
Postglacial Development, Present Situation and Future Trends in Central Europe, Ecological Studies,
No. 143, Springer: Berlin and Heidelberg.
Quine, C.P. and Humphrey, J.W. (2010) ‘Plantations of Exotic Tree Species in Britain:
Irrelevant for Biodiversity or Novel Habitat for Native Species?’, Biodiversity and Conserva-
tion, vol 19, pp. 1503–1512.
Reif, A., Brucker, U., Kratzer, R., Schmiedinger, A. and Bauhus, J. (2010) ‘Waldbe-
wirtschaftung in Zeiten des Klimawandels - Synergien und Konfliktpotenziale zwischen
Forstwirtschaft und Naturschutz’, Naturschutz und Landschaftsplanung, vol 42, pp. 261–266.
Röhrig, E., Bartsch, N. and von Lüpke, B. (2006) Waldbau auf ökologischer Grundlage. Eugen
Ulmer: Stuttgart 479 pp.
Schelhaas, M.J., Nabuurs, G.J. and Schuck, A. (2003) ‘Natural Disturbances in the European
Forests in the 19th and 20th Centuries’, Global Change Biology, vol 9, pp. 1620–1633.
Schmidt, U.E. (2009) ‘Continuous Cover Forests – a Success? A Historical Analysis’, Sch-
weizerische Zeitschrift Forstwesen, vol 160, pp. 144–151.
SW_545_Ch 9.indd 211SW_545_Ch 9.indd 211 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

212 Jürgen Bauhus
Not fo
r Dist
ribut
ion
Schmidt-Schütz, A. and Huss J. (1998) ‘Wiederbewaldung von Fichten-Sturmwurfflächen
auf vernässenden Standorten mit Hilfe von Pioniergehölzen’, in A. Fischer (ed.), Die Ent-
wicklung der Waldbiozönose nach Sturmwurf, Wiley-VCH Verlag GmbH, Ecomed, Lands-
berg, pp. 188–211.
Schütz, J.P. (2001) Der Plenterwald und weitere Formen strukturierter und gemischter Wälder, Parey:
Berlin 207 pp.
Schütz, J.P., Götz, M., Schmid, W. and Mandallaz, D. (2006) ‘Vulnerability of Spruce
(Picea abies) and Beech (Fagus sylvatica) Forest Stands to Storms and Consequences for
Silviculture’, European Journal of Forest Research , vol 125, pp. 291–302.
Schütz, J.P., Pukkula, T., Donoso, P. and von Gadow, K. (2012) ‘Historical Emer-
gence and Current Application of CCF’, in T.T. Pukkala and K. von Gadow
(eds.), Continuous Cover Forestry, Managing Forest Ecosystems, vol 23, pp. 1–28,
doi:10.1007/978–94–007–2202–6.
Seymour, R.S. and Hunter, M.L. Jr. (1992) New Forestry in Eastern Spruce-Fir Forests: Principles
and Applications to Maine, Orono, Maine, Maine Agricultural Experiment Station. Misc.
Publ. 716..
Spiecker, H., Mielikäinen, K., Köhl, M. and Skovsgaard, J.P. (1996) Growth Trends in
European Forests—Studies from 12 Countries, European Forest Institute Research Report 5,
Springer Verlag: Heidelberg 372 pp.
Spiecker, H., Hansen, J., Klimo, E., Skovsgaard, J.P., Sterba, H. and von Teuffel, K. (2004)
Norway Spruce Conversion – Options and Consequences, European Forest Institute Research
Reports 18. S. Brill, Leiden, The Netherlands, 269 pp.
Stephens, S.L, Millar, C.I. and Collins, B.M. (2010) ‘Operational Approaches to Managing
Forests of the Future in Mediterranean Regions within a Context of Changing Climates’,
Environmental Research Letters, vol 5, pp. 1–9.
Southwood, T.R.E. (1961) ‘The Number of Species of Insect Associated with Various
Trees’, Journal of Animal Ecology, vol 30, pp. 1–8.
Swank, T.W. and Crossley, D.A. (1988) Forest Hydrology and Ecology at Coweeta, Springer: New
York, 469 pp.
Swanson, M.E., Franklin, J.F., Beschta, R.L., Crisafulli, C.M, DellaSala, D.A., Hutto, R.L.,
Lindenmayer, D.B. and Swanson, F.J. (2010) ‘The Forgotten Stage of Forest Succession:
Early Successional Ecosystems on Forest Sites’, Frontiers in Ecology and the Environment, vol
10, pp. 1–10.
Szewczyk, J. and Szwagrzyk, J. (1996) ‘Tree Regeneration on Rotten Wood and on Soil in
Old-growth Stand’, Vegetatio, vol 122, pp. 37–46.
Thomasius, H. (1996) Geschichte, Theorie und Praxis des Dauerwaldes, Stassfurt, Landesforstver-
ein Sachsen-Anhalt, e. V., Straßfurt, Germany, 64 pp.
Tüxen, R. (1956) ‘Die heutige potentielle natürliche Vegetation als Gegenstand der Veg-
etationskartierung’, Angewandte Pflanzensoziologie, vol 13, pp. 5–42.
von Lüpke, B. and Spellmann, H. (1999) ‘Aspects of Stability, Growth and Natural Regen-
eration in Mixed Norway Spruce-European Beech Stands as a Basis of Silvicultural Deci-
sions’, in A.F.M. Olsthoorn, H.H. Bartelink, J.J. Gardiner, H. Pretzsch, H.J. Hekhuis
and A. Franc (eds.), Management of Mixed-species Forest: Silviculture and Economics, IBN-DLO
Scientific Contributions 15, Wageningen, The Netherlands, pp. 245–267.
von Lüpke, B. (2004) ‘Mixed Species Forests as a Means for Risk Reduction and Close to
Nature Silviculture: an Area of Conflict (in German)’, Forstarchiv, vol 57, pp. 43–50.
von Lüpke, B. (2008) ‘Influence of Various Cutting Types on Natural Regeneration of a
Sessile Oak-beech Mixed Stand (in German)’, Forstarchiv, vol 79, pp. 4–15.
von Lüpke, B. (2009) ‘Considerations about Tree Species Choice and Regeneration
SW_545_Ch 9.indd 212SW_545_Ch 9.indd 212 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM

Close-to-nature forest management in Europe 213
Not fo
r Dist
ribut
ion
Methods under Advancing Climate Change in Germany (in German)’, Forstarchiv, vol
80, pp. 67–75.
Wagner, S., Fischer, H. and Huth, F. (2011) ‘Canopy Effects on Vegetation Caused by Har-
vesting and Regeneration Treatments’, European Journal Forest Research, vol 130, pp. 17–40.
Wohlgemuth, T., Bürgi, M., Scheidegger, C. and Schütz, M. (2002) ‘Dominance Reduc-
tion of Species through Disturbance – a Proposed Management Principle for Central
European Forests’, Forest Ecology and Management, vol 166, pp. 1–15.
Yachi S. and Loreau, M. 1999. ‘Biodiversity and Ecosystem Productivity in a Fluctuating
Environment: the Insurance Hypothesis’, Proceedings of the National Academy of Sciences of the
United States of America, vol 96, pp. 57–64.
Zell, J., Hanewinkel, M. and Seeling, U. (2004) ‘Financial Optimisation of Target Diameter
Harvest of European Beech (Fagus sylvatica ) Considering the Risk of Decrease of timber
Quality due to Red Heartwood’, Forest Policy and Economics, vol 6, pp. 579–593.
Ziehe, M. and Hattemer, H.H. (2002) ‘Target-diameter Felling and Consequences for
Genetic Structures in a Beech Stand (Fagus sylvatica L.)’, in K. von Gadow, J. Nagel and
Saborowski, J. (eds.), Continuous Cover Forestry: Assessment, Analysis, Scenarios. Managing Forest
Ecosystems 4, Kluwer: Dordrecht, The Netherlands, pp. 91–105
SW_545_Ch 9.indd 213SW_545_Ch 9.indd 213 10/22/2012 3:11:49 PM10/22/2012 3:11:49 PM
Copyright © 2022 FDOKUMEN




















