
CLIC6, a member of the intracellular chloride channel family, interacts with dopamine D 2-like...
11
Molecular Brain Research 117 (2003) 47–57 www.elsevier.com / locate / molbrainres Research report CLIC6, a member of the intracellular chloride channel family, interacts with dopamine D -like receptors 2 a, a a b a * Nathalie Griffon , Freddy Jeanneteau , Fanny Prieur , Jorge Diaz , Pierre Sokoloff a ´ ´ ´ Unite de Neurobiologie et Pharmacologie Moleculaire, INSERM U-573, Centre Paul Broca,2ter Rue d’ Alesia, 75014 Paris, France b ´ ´ Laboratoire de Physiologie, Universite Rene Descartes,4 Avenue de l’ Observatoire, 75006 Paris, France Accepted 25 June 2003 Abstract To identify proteins interacting with the C-terminal cytoplasmic tail of the dopamine D receptor (D R), we used the two-hybrid 3 3 system to screen a rat brain cDNA library. We isolated three partial cDNAs encoding, respectively, for the MUPP1 multi-PDZ protein, for the N-terminal region of radixin, for GIPC and for a 160-amino acid open reading frame sharing high homology with the human CLIC6, also identified as parchorin in rabbit. In the two-hybrid system, CLIC6 was also able to interact with the D R and D R. The interaction 2 4 between D R and CLIC6 was confirmed by the use of a GST-D R C-terminus fusion protein and COS cell extracts transiently expressing 3 3 epitope-tagged CLIC6. In adult rat brain, CLIC6 mRNA expression was restricted to the choroid plexus, the striatal proliferative subventricular zone and the cerebellum where it is co-expressed with the D R in the Purkinje cells of the lobules IX and X. CLIC6 3 mRNA was also detected in the pituitary in the posterior lobe and in cells co-expressing the D R at the border between the intermediate 2 and anterior lobes. In transfected HEK293 cells, D R and CLIC6 co-localized at the plasma-membrane. No effect of CLIC6 transfection 3 was observed on either intracellular chloride concentration or D R / D R-mediated response. In two-hybrid system, CLIC6 also interacted 3 2 with MUPP1 and radixin but not GIPC, suggesting it could take part in a complex with D -like receptors, not only by direct interaction 2 with their C-termini, but also through interactions with scaffolding proteins. 2003 Elsevier B.V. All rights reserved. Theme: Neurotransmitters, modulators, transporters, and receptors Topic: Catecholamine receptors Keywords: Dopamine receptor; Two-hybrid system; Pituitary; Chloride channel; Parchorin; Scaffolding protein 1. Introduction coupling to G /G proteins, regulates ion channel activities i o and stimulates mitogenesis through activation of mitogen- The pleiotropic actions of dopamine, as well as of drugs activated protein kinases (MAPKs) (for a review, see Refs. used in the treatment of Parkinson’s disease and schizo- [2,8] ). However, a distinctive pattern of intracellular phrenia are mediated through interaction with five dopa- cascades seems to be activated by the D R. Indeed, 3 mine receptor subtypes, usually classified as D -like (D , mitogenesis induced by DR stimulation is markedly 1 1 3 D ) and D -like (D , D , D ) receptors [1,2]. Among enhanced when the cAMP pathway is activated, suggesting 5 2 2 3 4 them, the D R [3] is expressed in discrete adult rat brain that the transduction of the D R can involve both opposite 3 3 regions including the ventral striatum, mammillary bodies, and synergistic interactions with cAMP [9]. Furthermore, archicerebellum and the striatal proliferative subventricular D R-mediated MAPK phosphorylation involves the activa- 3 zone [4–7]. In heterologous expression systems, like the tion of phosphatidylinositol 3-kinase and of an atypical D R and D R and in agreement with their structural PKC isoform [10]. 2 4 homology, the D R inhibits the formation of cAMP by The structural and functional organization of receptors 3 within the cells requires sequence-specific interactions between proteins and signalling molecules. For several G *Corresponding author. Tel.: 133-1-4078-9256; fax: 133-1-4580- protein-coupled receptors, these interactions involve their 7293. E-mail address: [email protected] (N. Griffon). C-termini and PDZ (PSD-95 /discs large /ZO-1) domain- ] ] ] 0169-328X / 03 / $ – see front matter 2003 Elsevier B.V. All rights reserved. doi:10.1016 / S0169-328X(03)00283-3
Transcript of CLIC6, a member of the intracellular chloride channel family, interacts with dopamine D 2-like...

Molecular Brain Research 117 (2003) 47–57www.elsevier.com/ locate/molbrainres
Research report
C LIC6, a member of the intracellular chloride channel family,interacts with dopamine D -like receptors2a , a a b a*Nathalie Griffon , Freddy Jeanneteau , Fanny Prieur , Jorge Diaz , Pierre Sokoloff
a ´ ´ ´Unite de Neurobiologie et Pharmacologie Moleculaire, INSERM U-573, Centre Paul Broca, 2ter Rue d’ Alesia, 75014Paris, Franceb ´ ´Laboratoire de Physiologie, Universite Rene Descartes, 4 Avenue de l’ Observatoire, 75006 Paris, France
Accepted 25 June 2003
Abstract
To identify proteins interacting with the C-terminal cytoplasmic tail of the dopamine D receptor (D R), we used the two-hybrid3 3
system to screen a rat brain cDNA library. We isolated three partial cDNAs encoding, respectively, for the MUPP1 multi-PDZ protein, forthe N-terminal region of radixin, for GIPC and for a 160-amino acid open reading frame sharing high homology with the human CLIC6,also identified as parchorin in rabbit. In the two-hybrid system, CLIC6 was also able to interact with the D R and D R. The interaction2 4
between D R and CLIC6 was confirmed by the use of a GST-D R C-terminus fusion protein and COS cell extracts transiently expressing3 3
epitope-tagged CLIC6. In adult rat brain, CLIC6 mRNA expression was restricted to the choroid plexus, the striatal proliferativesubventricular zone and the cerebellum where it is co-expressed with the D R in the Purkinje cells of the lobules IX and X. CLIC63
mRNA was also detected in the pituitary in the posterior lobe and in cells co-expressing the D R at the border between the intermediate2
and anterior lobes. In transfected HEK293 cells, D R and CLIC6 co-localized at the plasma-membrane. No effect of CLIC6 transfection3
was observed on either intracellular chloride concentration or D R/D R-mediated response. In two-hybrid system, CLIC6 also interacted3 2
with MUPP1 and radixin but not GIPC, suggesting it could take part in a complex with D -like receptors, not only by direct interaction2
with their C-termini, but also through interactions with scaffolding proteins. 2003 Elsevier B.V. All rights reserved.
Theme: Neurotransmitters, modulators, transporters, and receptors
Topic: Catecholamine receptors
Keywords: Dopamine receptor; Two-hybrid system; Pituitary; Chloride channel; Parchorin; Scaffolding protein
1 . Introduction coupling to G /G proteins, regulates ion channel activitiesi o
and stimulates mitogenesis through activation of mitogen-The pleiotropic actions of dopamine, as well as of drugs activated protein kinases (MAPKs) (for a review, see Refs.
used in the treatment of Parkinson’s disease and schizo-[2,8]). However, a distinctive pattern of intracellularphrenia are mediated through interaction with five dopa- cascades seems to be activated by the D R. Indeed,3
mine receptor subtypes, usually classified as D -like (D , mitogenesis induced by D R stimulation is markedly1 1 3
D ) and D -like (D , D , D ) receptors[1,2]. Among enhanced when the cAMP pathway is activated, suggesting5 2 2 3 4
them, the D R[3] is expressed in discrete adult rat brain that the transduction of the D R can involve both opposite3 3
regions including the ventral striatum, mammillary bodies, and synergistic interactions with cAMP[9]. Furthermore,archicerebellum and the striatal proliferative subventricular D R-mediated MAPK phosphorylation involves the activa-3
zone [4–7]. In heterologous expression systems, like the tion of phosphatidylinositol 3-kinase and of an atypicalD R and D R and in agreement with their structural PKC isoform[10].2 4
homology, the D R inhibits the formation of cAMP by The structural and functional organization of receptors3
within the cells requires sequence-specific interactionsbetween proteins and signalling molecules. For several G*Corresponding author. Tel.:133-1-4078-9256; fax:133-1-4580-protein-coupled receptors, these interactions involve their7293.
E-mail address: [email protected](N. Griffon). C-termini and PDZ (PSD-95/discs large/ZO-1) domain-] ] ]
0169-328X/03/$ – see front matter 2003 Elsevier B.V. All rights reserved.doi:10.1016/S0169-328X(03)00283-3

48 N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57
T able 1containing proteins. In the case of the D -like receptors,2Isolation of binding partners to the D R by two-hybrid system3association of the D R and D R with filamin A provides a2 3
Protein Interaction with D R-CTmechanism whereby they may be functionally linked to 3
downstream signalling components via the actin cyto- CLIC6 (CT5) 1
skeleton[11], a role similarly reported for spinophilin[12] MUPP1 (PDZ 6–13) 1
Radixin (AA 106–379) 1and the 4.1N protein[13], two other actin-binding proteins.GIPC 1To search for partners involved in D R subcellular3 CT3 1
localization and/or transduction, we have sought forYeast two-hybrid screen of a rat brain cDNA library with the D R3proteins that interact with the C-terminal tail of the D R,3C-terminus (CT) as bait identified five interacting proteins: CLIC6, aby conducting a yeast two-hybrid screen of a rat brainpotential intracellular chloride channel, MUPP1, a multi-PDZ domain
cDNA library. In the present study, we describe the protein, radixin, a protein of the ERM family, GIPC, the GAIP-interactingisolation and characterization of a member of the chloride protein, C-terminus and CT3, an unidentified protein. To confirm the
specificity of the interaction, EGY48 yeast cells were co-transformed withintracellular channel (CLIC) family, the rat CLIC6, whichthese plasmids and the bait and selected for growth on leucine-deprivedspecifically interacts with D -like receptors. The family2medium andb-galactosidase colouration.consists of six 6 members, termed CLIC1 to CLIC6, the
bovine CLIC5B or p64 protein being the prototypicalmember. These proteins have been implicated in chloride in frame with the B42 activation domain in pEG202 andion transport within various subcellular compartments. tested for interaction with the CT5 clone by specificThese proteins have highly conserved carboxy-terminal growth on selective medium as well asb-galactosidaseends, containing a putative hydrophobic membrane-span- activity. A cDNA encoding for the CLIC6 coiled-coil-likening domain, whereas their N-termini differ markedly, domain (amino acids 480–519) was generated by PCRbeing very short for the CLIC1 to CLIC4 and longer for with the CT5 clone as template and subcloned in frameCLIC5B and CLIC6[14–20].The identification of CLIC’s with either the B42 activation domain in pEG202 or thebinding partners may help define the function of these Lex-A binding domain in pJG4-5. CT5 was digested byproteins, which is presently unknown. EcoRI/XhoI from pJG4-5 and the insert was subcloned in
frame with the B42 activation domain in pEG202. MUPP1(PDZ domains 6–13), radixin (amino acids 106–363) and
2 . Materials and methods the full-length GIPC subcloned in the pJG4-5 were isolatedfrom the original two-hybrid screen.
2 .1. Yeast two-hybrid screening2 .3. Full-length cDNA cloning
Residues 435–446 of the D R were generated by3
polymerase chain reaction (PCR), inserted in the pEG202 The 59 part of the CLIC6 mRNA was determined by6bait vector in frame with the B42 activation domain and screening a rat brain cDNA library (2310 phages) in
32used with the DupLex-A Two-Hybrid system kit (OriGene lZAPII (Stratagene) with a P-labelled PCR fragment6Technologies). A rat brain cDNA library (4.5310 in- obtained from the CT5 insert as template. Five positive
dependent recombinant clones) in the pJG4-5 downstream clones were analyzed. None of them contained the full-Lex-A binding domain (OriGene Technologies) was length cDNA, so the CLIC6 coding sequence was con-screened using the supplied yeast strain EGY48. Among structed by ligation with anEcoRI–XmnI-digested 5950 blue colonies picked from the original galactose-in- fragment from a partiallZAPII clone and anXmnI–XhoIduced selection plates (-Ura, -Trp, -Leu, -His), one con- digested 39 fragment from the CT5 two-hybrid clone andtained a plasmid, that we named CT5, which allowed subcloned into the pcDNA3.1/HIS C vector (Invitrogen,specific and reproducible growth on selective medium, as San Diego, CA, USA) in frame with a polyhistidine metal-well as b-galactosidase induction upon isolation and binding tag and an Xpress� epitope for mammalianretransformation with the bait vector. This clone contained expression (Xpress/His-CLIC6).an in-frame insertion of 160 amino acids. Other plasmidscorresponded to full length or partial sequences encoded2 .4. Cell culture and transfectionsfor radixin (1300 bp), MUPP1 (5500 bp), GIPC (950 and550 bp) and two other encoded for an unidentified protein COS-7 and Chinese hamster ovary (CHO) cells were(700 and 900 bp) (Table 1). grown in a humidified atmosphere of 5% CO , 95% air in2
Dulbecco’s modified Eagle’s medium (DMEM; Life Tech-2 .2. Yeast two-hybrid binary assays nologies) supplemented with 10% fetal calf serum. CHO-
D R and CHO-D R cells[3] were grown in DMEM2 3
C-Termini of rat D R (amino acids 433–444), D R supplemented with 10% dialyzed fetal calf serum. HEK2 4
(amino acids 372–385) and D R C-terminus mutants were 293 cells were grown inaMEM supplemented with 10%3
constructed by oligonucleotides hybridization, subcloned fetal calf serum.

N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57 49
COS-7 cells were transfected with pcDNA3.1/HIS C or hybridized as described[7]. Slices were apposed tobmaxXpress/His-CLIC6 plasmid using electroporation. After 48 Hyperfilms (Amersham, Freiburg, Germany) or dipped in ah, cells were collected and resuspended in 50 mM potas- photographic emulsion (LM1; Amersham Pharmaciasium phosphate buffer containing 150 mM NaCl (PBS) Biotech). Dipped sections were counterstained withand a cocktail of protease inhibitors (Sigma, St. Louis, Mayer’s hemalun solution and examined with a photo-MO, USA) and centrifuged at 45,000g for 30 min. The microscope Axiophot (Zeiss).supernatants were used directly for affinity adsorption onglutathione-Sepharose (Amersham Pharmacia Biotech). 2 .7. Intracellular chloride measurements
CHO, CHO-D R or CHO-D R cells were transfected2 3
with Lipofectine� (Life Technologies) with Xpress/His- Intracellular chloride concentration was monitored byCLIC6. After selection with Geneticin (4 mg/ml, Life the chloride-sensitive fluorescent dye,n-(ethoxycar-Technologies) for 15 days, individual clones were isolated bonylmethyl)-6-methoxyquinolinium (MQAE), as de-by ring cloning and tested by western blotting with Anti- scribed[21]. Briefly, cells were incubated with 10 mMXpress� antibody for CLIC6 expression. MQAE in loading buffer [DMEM, 0.1% bovine serum
albumin (BSA), 10 mM 4-(2-hydroxyethyl)-1-piperazine-ethanesulphonic acid (HEPES), pH 7.5] for 45 min at
2 .5. Bacterial glutathione S-transferase (GST) fusion37 8C. Cells were washed with and resuspended in loading
protein expression and binding studiesbuffer. For measurement, cells were diluted in DMEM orin chloride-free medium (101 mM sodium gluconate, 5
Residues 435–446 of the D R were fused to GST in the3 mM potassium gluconate, 2 mM MgSO , 2 mM calcium4pGEX-2TK vector (Amersham Pharmacia Biotech) andacetate, 10 mM HEPES, pH 7.5, 10 mM glucose). Fluores-
expressed in theEscherichia coli BL21 strain after induc-cence was excited at 350 nm and detected at 460 nm. The
tion with 0.5 mM isopropylb-D-thiogalactopyranoside fortime course of MQAE was monitored using a fluorescence
3 h. Cells were sonicated and incubated in PBS with 1%spectrophotometer F2000 (Hitachi). Dopamine agonist
n-octyl b-D-glucoside for 2 h and then centrifuged atquinpirole (Ely-Lilly, Indianapolis, IN, USA), antagonist
45,000 g for 30 min. Supernatants were incubated withhaloperidol (Janssen Pharmaceuticals, Bersee, Belgium) or
250 ml of a 50% suspension of glutathione-SepharoseNaCl (25 mM) effect on intracellular chloride concen-
(Amersham Pharmacia Biotech) beads for 30 min. and thentration were tested. Reduction of intracellular (efflux)
washed three times with 10 ml PBS. The immobilisedchloride concentration is expected as an increase in
fusion proteins where then incubated with soluble extractsfluorescence.
from Xpress/His-CLIC6 transfected COS-7 cells at 48Cfor 1 h and washed again three times with 10 ml PBS.
2 .8. Measurement of mitogenesisPellets were resuspended in sodium dodecyl sulphate(SDS) sample buffer and separated by 8% SDS–poly-
Cells were plated in 96-well plates at a density of 5,000acrylamide gel electrophoresis (PAGE) followed by elec-
cells /well. After a 24-h culture, cells were washed twicetrophoretic transfer onto a nitro-cellulose membrane. The
with DMEM and incubated for 16 h with 100 nMmembrane was blocked and reacted with the Anti-Xpress� 3quinpirole (Eli-Lilly). Then, [ H]thymidine (1mCi/well;antibody (Invitrogen).
Amersham Pharmacia Biotech) was added for 2 h and cellswere harvested by vacuum filtration through Whatman
2 .6. In situ hybridization GF/C fibre filters using an automated cell harvester(Brandel-Beckman) and rinsed 15 times with water.
Probes for rat D R and D R mRNAs corresponded to Radioactivity retained on filters was counted by liquid2 3
respective sequences of the third intracellular loop of the scintigraphy.receptor and are described elsewhere[3,4]. Probes 1 and 2(Fig. 1A) for CLIC6 mRNA corresponded, respectively, to 2 .9. ImmunocytochemistryN- and C-terminal fragments (nucleotides 449–874 and1567–1831) obtained by digestion and subcloned into A c-myc-tag was introduced into the N-terminus of the
33pGEM-4Z. P-Labelled riboprobes were synthesized with rat D R cDNA after the first methionine codon by per-3
the Riboprobe Gemini System (Promega), treated by forming PCR with a Not I-flanked myc-tag-containingRNAse–DNAse free (Roche), and recovered from Chroma sense primer and a Xba I-flanked antisense primer en-Spin-30 columns (Clontech, Cambridge, UK). Digox- compassing the stop codon and subcloned in the pRC-igenin-labelled (Roche) D R probe was synthezised with CMV (Invitrogen) (myc-D R). HEK 293 cells grown on2 3
the Ampliscribe� transcription kit (Epicentre Tech- fibronectin-coated cover slips were transfected with thenologies). Cryostat sections (10mm) of adult male Wistar Xpress/His-CLIC6, the myc-D R or both with the3
rat brain and pituitary and of embryo at the 18th gestation- Superfect� reagent (Qiagen). Forty-eight hours after trans-al day were mounted on slides. Slides were treated and fection, cells were washed with PBS and fixed with 2%

50 N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57
Fig. 1. Domain structure of the rat CLIC 6 and sequence comparison with other members of the CLIC family. (A) Schematic representation of the CT5two-hybrid clone and of the identified domains on CLIC6. TM: Predicted transmembrane domain, CC: coiled-coil like domain, probes 1 and 2 representthose used in situ hybridization. (B) Sequence alignment of members of the CLIC family. The underlined and overlined sequences represent the predictedtransmembrane domain and the putative coiled-coil domain, respectively. A 17 amino acid repeat is indicated by bold letters.
paraformaldehyde in PBS at room temperature for 30 min. (Sigma) and the rabbit polyclonal anti-myc antibodyAfter blocking in Tris-buffered saline (TBS) containing (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were5% donkey serum, 0.1% gelatine, 0.05% Tween 20 and used as primary antibodies at 1:3,000 and 1:400 dilutions,0.4% BSA, the mouse monoclonal anti-His antibody respectively, in the blocking buffer diluted five times.

N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57 51
After washing, fluorescin isothiocyanate (FITC)- and CY3- interaction whereas deletion of the LKI motif, mutationconjugated secondary antibodies (Interchim) were em- K442A and F440A weakened the association. Deletion ofployed to differentially visualise the primary antibodies. the EFR motif, proximal to the seventh transmembraneConfocal pictures were taken using the Leica confocal domain abolished the interaction and replacement of thesesystem TSC SPII. amino acids by neutral residues which restore bait length
did not allow one to recover the interaction (Fig. 2B).Furthermore, interaction of the D R with the C-terminal3
3 . Results region of CLIC6 was not mediated by the coiled-coil-likedomain of CLIC6 (Fig. 2C). In binary two-hybrid assays,
3 .1. Isolation of dopamine D receptor binding partners we also tested the ability of CLIC6 to interact with itself3
by a two-hybrid screen and showed that the coiled-coil-like domain dimerizedwith the C-terminal region of CLIC6 (CT5) but not with
To identify proteins that interact with the D R, we itself (Fig. 2C).3
screened a yeast two-hybrid cDNA library constructedfrom adult rat brain using the cytoplasmic C-terminal tail 3 .4. CLIC6 and D R C-terminus interacts in vitro3
6region as ‘bait’ (amino acids 435–446). From 4.5310clones screened, 50 colonies were isolated from selection We expressed an epitope-tagged Xpress/His-CLIC6 inplates, representing five different cDNAs (Table 1). These COS-7 cells. The recombinant CLIC6, predominantlycDNAs encoded for the multi-PDZ-domain protein cytosolic, migrated as two bands with apparent size of 135MUPP1 (PDZ domains 6–13)[22], the N-terminal part of and 140 kDa in SDS–PAGE, whereas a 65 kDa molecularradixin, a scaffolding protein mediated membrane–protein mass is predicted from the sequence (Fig. 3, lane 2) whichassociation (for a review, see Ref.[23]) and for GIPC could be explain by the dimerization observed in the(GAIP interacting protein C-terminus)[24], respectively. two-hybrid system (Fig. 2C). The two bands observedTwo other cDNAs encoded an unidentified protein, with- could be due to distinct post-translational modifications,out homology with any known domain, that we termed since the CLIC6 sequence contains consensus motifs forCT3. Finally, we identified a clone that we named CT5 and protein kinase C phosphorylation (five sites), casein kinasecharacterized further, encoding for a 160-amino acid open II phosphorylation (14 sites) andN-myristoylation (sixreading frame, homologous to the CLIC family members sites) but no predicted site for proteine kinase A phos-(Fig. 1A). phorylation.
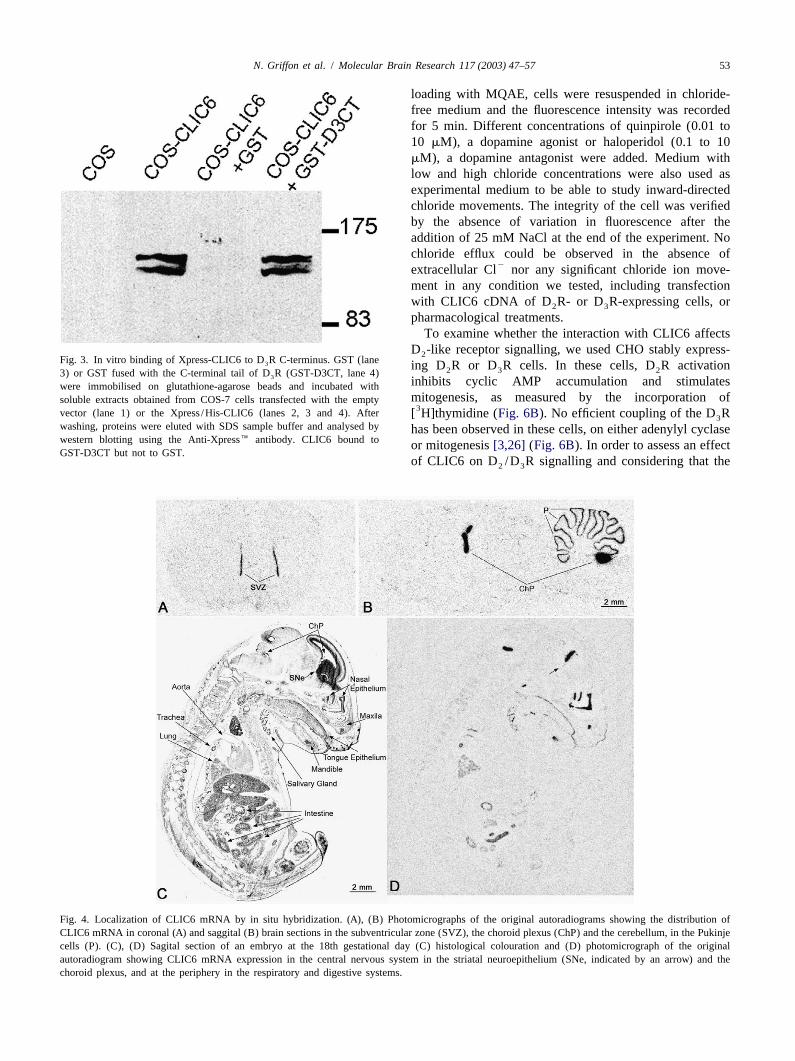
To biochemically confirm the interaction of CLIC6 with3 .2. Cloning of the rat CLIC6 the D R C-terminal tail by a biochemical approach, we3
used Xpress/His-CLIC6 transfected COS-7 cells cytosolicTo obtain the full-length cDNA, we screened a rat brain extracts to perform a pull-down assay using either GST or
cDNA library using the CT5 coding sequence as a probe the GST fusion protein with the D R C-terminal tail3
and obtained one positive clone encoding a cDNA of 2.2 (GST-D3CT) immobilized on glutathione-Sepharosekbp partially encompassing the 39-coding region of CT5. beads. Xpress/His-CLIC6 bound to GST-D3CT, but not toThe full-length cDNA encoded for a 612-amino acid GST (Fig. 3). This confirms the yeast two-hybrid resultsprotein, homologous to the human CLIC6 sequence (acces- and shows that, in vitro, the C-terminal tail of the D R3
sion number AY191249). The rat CLIC6 contains a binds to CLIC6.predicted hydrophobic membrane-spanning domain (aminoacids 402–417) and a putative coiled-coil domain (amino 3 .5. Expression of CLIC6 mRNA in rat brain and co-acids 490–512) also found in CLIC3 and CLIC5 (predic- expression with D -like receptors2
tion of coiled coil regions in proteins by the Lupas’s33method at http: / /www.ch.embnet.org/software/COIL- Northern blot hybridization with a P-labelled cDNA
S form.html) (Fig. 1A and B). probe located at the 39 coding region of CLIC6 on rat]
mRNAs revealed the predominant expression of a 3 kb3 .3. CLIC6 bound to all dopamine D -like receptors transcript in the stomach, pituitary and brain (not shown).2
33In situ hybridization using P-labelled cRNA probesThe C-terminal is highly conserved among the dopamine designed at either the 39 or 59 coding region of the CLIC6
D -like receptors. To examine whether CLIC6 interacts (Fig. 1A) revealed a high expression in the choroid plexus2
with other dopamine D -like receptor, we also tested the (Fig. 4B) and a restricted pattern of expression in the brain2
D R and D R C-terminal tail domains in binary two- (Fig. 4A and B). CLIC6 mRNA was expressed in the2 4
hybrid assays (Fig. 2A). Both receptors were found to bind Purkinje cells of the cerebellum, where it was co-expressedto CLIC6, suggesting a consensus motif for binding among with D R mRNA in lobules IX and X (all the Purkinje3
conserved amino acids. Mutations in the D R C-terminal cells of these lobules expressed the D R, see Ref.[6])3 3
tail showed that the last cysteine was not involved in the (Figs. 4B and 5A and B). CLIC6 mRNA was also
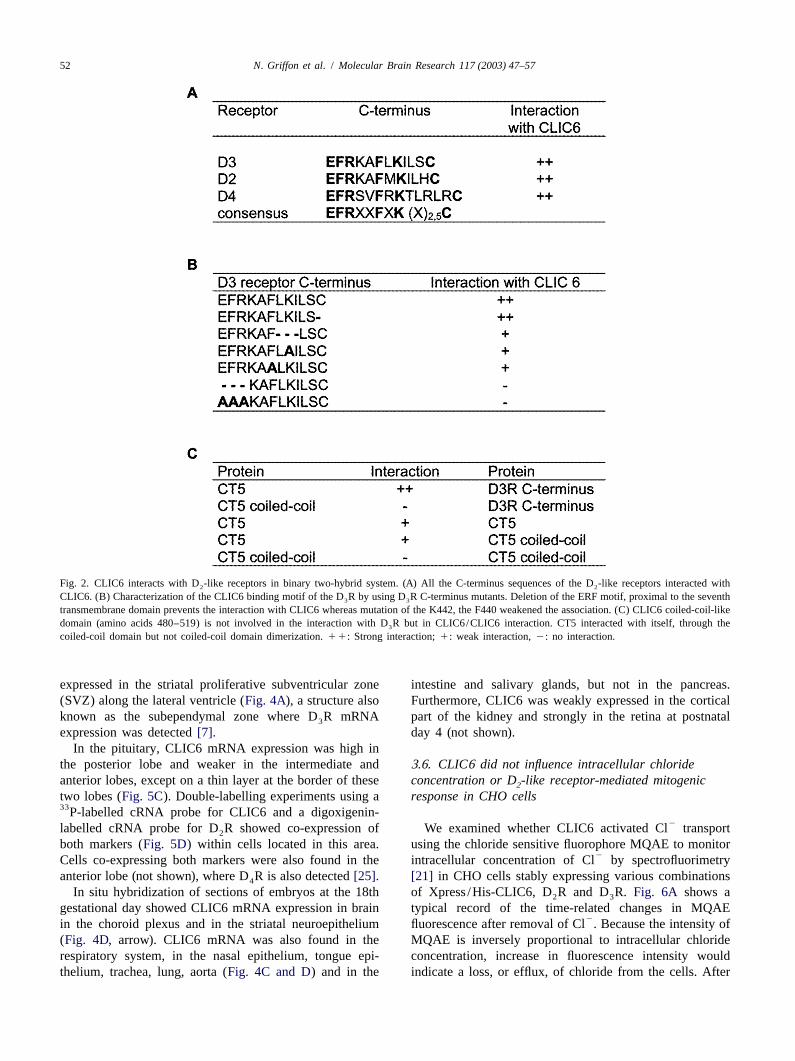
52 N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57
Fig. 2. CLIC6 interacts with D -like receptors in binary two-hybrid system. (A) All the C-terminus sequences of the D -like receptors interacted with2 2
CLIC6. (B) Characterization of the CLIC6 binding motif of the D R by using D R C-terminus mutants. Deletion of the ERF motif, proximal to the seventh3 3
transmembrane domain prevents the interaction with CLIC6 whereas mutation of the K442, the F440 weakened the association. (C) CLIC6 coiled-coil-likedomain (amino acids 480–519) is not involved in the interaction with D R but in CLIC6/CLIC6 interaction. CT5 interacted with itself, through the3
coiled-coil domain but not coiled-coil domain dimerization.11: Strong interaction;1: weak interaction,2: no interaction.
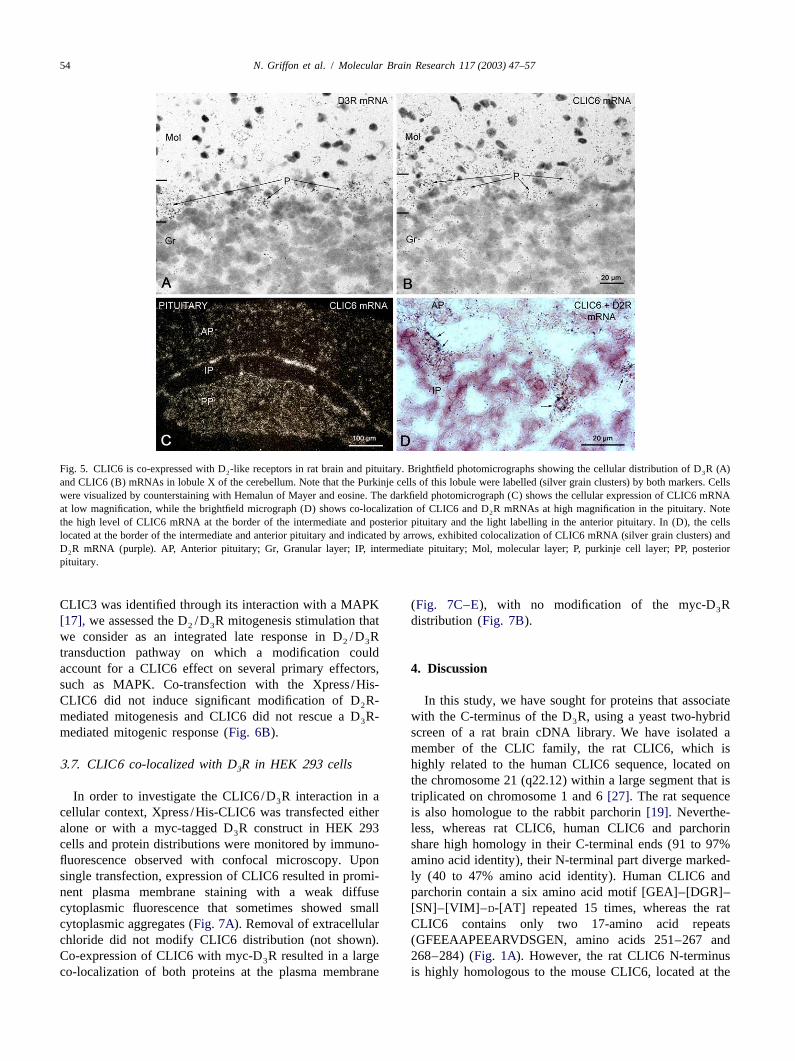
expressed in the striatal proliferative subventricular zone intestine and salivary glands, but not in the pancreas.(SVZ) along the lateral ventricle (Fig. 4A), a structure also Furthermore, CLIC6 was weakly expressed in the corticalknown as the subependymal zone where D R mRNA part of the kidney and strongly in the retina at postnatal3
expression was detected[7]. day 4 (not shown).In the pituitary, CLIC6 mRNA expression was high in
the posterior lobe and weaker in the intermediate and 3 .6. CLIC6 did not influence intracellular chlorideanterior lobes, except on a thin layer at the border of theseconcentration or D -like receptor-mediated mitogenic2
two lobes (Fig. 5C). Double-labelling experiments using a response in CHO cells33P-labelled cRNA probe for CLIC6 and a digoxigenin-
2labelled cRNA probe for D R showed co-expression of We examined whether CLIC6 activated Cl transport2
both markers (Fig. 5D) within cells located in this area. using the chloride sensitive fluorophore MQAE to monitor2Cells co-expressing both markers were also found in the intracellular concentration of Cl by spectrofluorimetry
anterior lobe (not shown), where D R is also detected[25]. [21] in CHO cells stably expressing various combinations4
In situ hybridization of sections of embryos at the 18th of Xpress/His-CLIC6, D R and D R.Fig. 6A shows a2 3
gestational day showed CLIC6 mRNA expression in brain typical record of the time-related changes in MQAE2in the choroid plexus and in the striatal neuroepithelium fluorescence after removal of Cl . Because the intensity of
(Fig. 4D, arrow). CLIC6 mRNA was also found in the MQAE is inversely proportional to intracellular chloriderespiratory system, in the nasal epithelium, tongue epi- concentration, increase in fluorescence intensity wouldthelium, trachea, lung, aorta (Fig. 4C and D) and in the indicate a loss, or efflux, of chloride from the cells. After
N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57 53
loading with MQAE, cells were resuspended in chloride-free medium and the fluorescence intensity was recordedfor 5 min. Different concentrations of quinpirole (0.01 to10 mM), a dopamine agonist or haloperidol (0.1 to 10mM), a dopamine antagonist were added. Medium withlow and high chloride concentrations were also used asexperimental medium to be able to study inward-directedchloride movements. The integrity of the cell was verifiedby the absence of variation in fluorescence after theaddition of 25 mM NaCl at the end of the experiment. Nochloride efflux could be observed in the absence of
2extracellular Cl nor any significant chloride ion move-ment in any condition we tested, including transfectionwith CLIC6 cDNA of D R- or D R-expressing cells, or2 3
pharmacological treatments.To examine whether the interaction with CLIC6 affects
D -like receptor signalling, we used CHO stably express-2Fig. 3. In vitro binding of Xpress-CLIC6 to D R C-terminus. GST (lane3 ing D R or D R cells. In these cells, D R activation2 3 23) or GST fused with the C-terminal tail of D R (GST-D3CT, lane 4)3 inhibits cyclic AMP accumulation and stimulateswere immobilised on glutathione-agarose beads and incubated with
mitogenesis, as measured by the incorporation ofsoluble extracts obtained from COS-7 cells transfected with the empty3vector (lane 1) or the Xpress/His-CLIC6 (lanes 2, 3 and 4). After [ H]thymidine (Fig. 6B). No efficient coupling of the D R3
washing, proteins were eluted with SDS sample buffer and analysed by has been observed in these cells, on either adenylyl cyclasewestern blotting using the Anti-Xpress� antibody. CLIC6 bound to or mitogenesis[3,26] (Fig. 6B). In order to assess an effectGST-D3CT but not to GST.
of CLIC6 on D /D R signalling and considering that the2 3
Fig. 4. Localization of CLIC6 mRNA by in situ hybridization. (A), (B) Photomicrographs of the original autoradiograms showing the distribution ofCLIC6 mRNA in coronal (A) and saggital (B) brain sections in the subventricular zone (SVZ), the choroid plexus (ChP) and the cerebellum, in the Pukinjecells (P). (C), (D) Sagital section of an embryo at the 18th gestational day (C) histological colouration and (D) photomicrograph of the originalautoradiogram showing CLIC6 mRNA expression in the central nervous system in the striatal neuroepithelium (SNe, indicated by an arrow) and thechoroid plexus, and at the periphery in the respiratory and digestive systems.
54 N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57
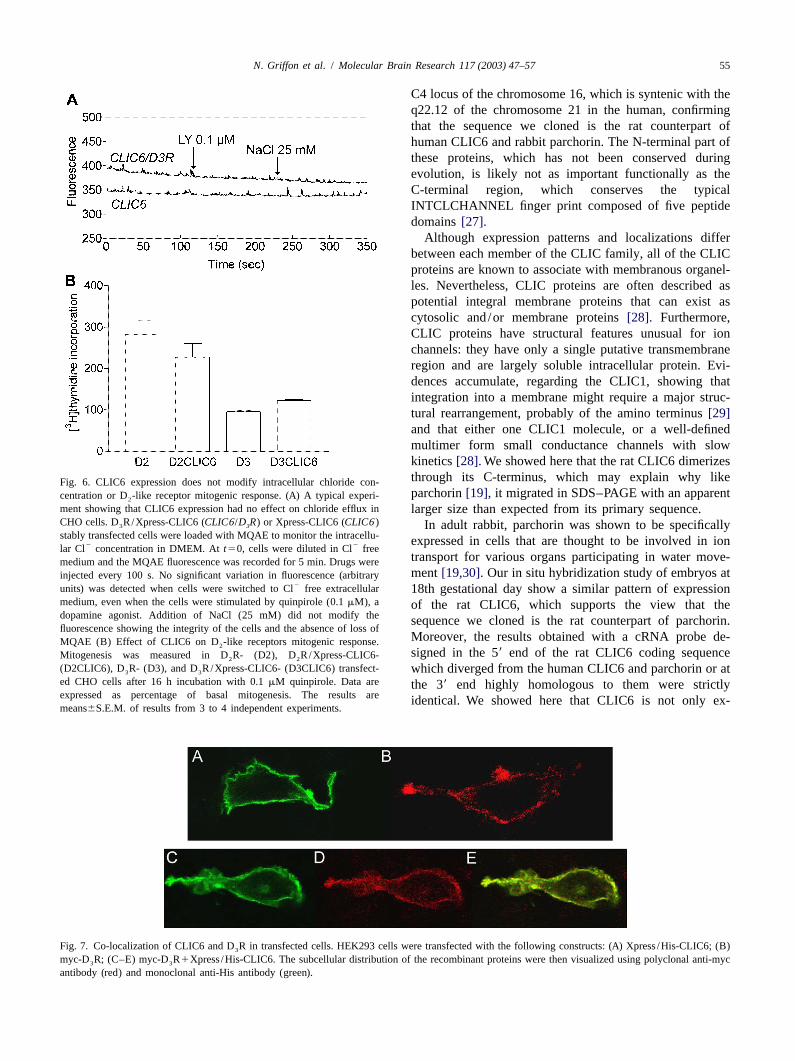
Fig. 5. CLIC6 is co-expressed with D -like receptors in rat brain and pituitary. Brightfield photomicrographs showing the cellular distribution of D R(A)2 3
and CLIC6 (B) mRNAs in lobule X of the cerebellum. Note that the Purkinje cells of this lobule were labelled (silver grain clusters) by both markers. Cellswere visualized by counterstaining with Hemalun of Mayer and eosine. The darkfield photomicrograph (C) shows the cellular expression of CLIC6 mRNAat low magnification, while the brightfield micrograph (D) shows co-localization of CLIC6 and D R mRNAs at high magnification in the pituitary. Note2
the high level of CLIC6 mRNA at the border of the intermediate and posterior pituitary and the light labelling in the anterior pituitary. In (D), the cellslocated at the border of the intermediate and anterior pituitary and indicated by arrows, exhibited colocalization of CLIC6 mRNA (silver grain clusters) andD R mRNA (purple). AP, Anterior pituitary; Gr, Granular layer; IP, intermediate pituitary; Mol, molecular layer; P, purkinje cell layer; PP, posterior2
pituitary.
CLIC3 was identified through its interaction with a MAPK (Fig. 7C–E), with no modification of the myc-D R3
[17], we assessed the D /D R mitogenesis stimulation that distribution (Fig. 7B).2 3
we consider as an integrated late response in D /D R2 3
transduction pathway on which a modification couldaccount for a CLIC6 effect on several primary effectors, 4 . Discussionsuch as MAPK. Co-transfection with the Xpress/His-CLIC6 did not induce significant modification of D R- In this study, we have sought for proteins that associate2
mediated mitogenesis and CLIC6 did not rescue a D R- with the C-terminus of the D R, using a yeast two-hybrid3 3
mediated mitogenic response (Fig. 6B). screen of a rat brain cDNA library. We have isolated amember of the CLIC family, the rat CLIC6, which is
3 .7. CLIC6 co-localized with D R in HEK 293 cells highly related to the human CLIC6 sequence, located on3
the chromosome 21 (q22.12) within a large segment that isIn order to investigate the CLIC6/D R interaction in a triplicated on chromosome 1 and 6[27]. The rat sequence3
cellular context, Xpress/His-CLIC6 was transfected either is also homologue to the rabbit parchorin[19]. Neverthe-alone or with a myc-tagged D R construct in HEK 293 less, whereas rat CLIC6, human CLIC6 and parchorin3
cells and protein distributions were monitored by immuno- share high homology in their C-terminal ends (91 to 97%fluorescence observed with confocal microscopy. Upon amino acid identity), their N-terminal part diverge marked-single transfection, expression of CLIC6 resulted in promi- ly (40 to 47% amino acid identity). Human CLIC6 andnent plasma membrane staining with a weak diffuse parchorin contain a six amino acid motif [GEA]–[DGR]–cytoplasmic fluorescence that sometimes showed small [SN]–[VIM]–D-[AT] repeated 15 times, whereas the ratcytoplasmic aggregates (Fig. 7A). Removal of extracellular CLIC6 contains only two 17-amino acid repeatschloride did not modify CLIC6 distribution (not shown). (GFEEAAPEEARVDSGEN, amino acids 251–267 andCo-expression of CLIC6 with myc-D R resulted in a large 268–284) (Fig. 1A). However, the rat CLIC6 N-terminus3
co-localization of both proteins at the plasma membrane is highly homologous to the mouse CLIC6, located at the
N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57 55
C4 locus of the chromosome 16, which is syntenic with theq22.12 of the chromosome 21 in the human, confirmingthat the sequence we cloned is the rat counterpart ofhuman CLIC6 and rabbit parchorin. The N-terminal part ofthese proteins, which has not been conserved duringevolution, is likely not as important functionally as theC-terminal region, which conserves the typicalINTCLCHANNEL finger print composed of five peptidedomains[27].
Although expression patterns and localizations differbetween each member of the CLIC family, all of the CLICproteins are known to associate with membranous organel-les. Nevertheless, CLIC proteins are often described aspotential integral membrane proteins that can exist ascytosolic and/or membrane proteins[28]. Furthermore,CLIC proteins have structural features unusual for ionchannels: they have only a single putative transmembraneregion and are largely soluble intracellular protein. Evi-dences accumulate, regarding the CLIC1, showing thatintegration into a membrane might require a major struc-tural rearrangement, probably of the amino terminus[29]and that either one CLIC1 molecule, or a well-definedmultimer form small conductance channels with slowkinetics[28]. We showed here that the rat CLIC6 dimerizesthrough its C-terminus, which may explain why likeFig. 6. CLIC6 expression does not modify intracellular chloride con-parchorin[19], it migrated in SDS–PAGE with an apparentcentration or D -like receptor mitogenic response. (A) A typical experi-2
ment showing that CLIC6 expression had no effect on chloride efflux in larger size than expected from its primary sequence.CHO cells. D R/Xpress-CLIC6 (CLIC6/D R) or Xpress-CLIC6 (CLIC6)3 3 In adult rabbit, parchorin was shown to be specificallystably transfected cells were loaded with MQAE to monitor the intracellu- expressed in cells that are thought to be involved in ion2 2lar Cl concentration in DMEM. Att50, cells were diluted in Cl free
transport for various organs participating in water move-medium and the MQAE fluorescence was recorded for 5 min. Drugs werement[19,30].Our in situ hybridization study of embryos atinjected every 100 s. No significant variation in fluorescence (arbitrary
2units) was detected when cells were switched to Cl free extracellular 18th gestational day show a similar pattern of expressionmedium, even when the cells were stimulated by quinpirole (0.1mM), a of the rat CLIC6, which supports the view that thedopamine agonist. Addition of NaCl (25 mM) did not modify the sequence we cloned is the rat counterpart of parchorin.fluorescence showing the integrity of the cells and the absence of loss of
Moreover, the results obtained with a cRNA probe de-MQAE (B) Effect of CLIC6 on D -like receptors mitogenic response.2
signed in the 59 end of the rat CLIC6 coding sequenceMitogenesis was measured in D R- (D2), D R/Xpress-CLIC6-2 2
(D2CLIC6), D R- (D3), and D R/Xpress-CLIC6- (D3CLIC6) transfect- which diverged from the human CLIC6 and parchorin or at3 3
ed CHO cells after 16 h incubation with 0.1mM quinpirole. Data are the 39 end highly homologous to them were strictlyexpressed as percentage of basal mitogenesis. The results areidentical. We showed here that CLIC6 is not only ex-means6S.E.M. of results from 3 to 4 independent experiments.
Fig. 7. Co-localization of CLIC6 and D R in transfected cells. HEK293 cells were transfected with the following constructs: (A) Xpress/His-CLIC6; (B)3
myc-D R; (C–E) myc-D R1Xpress/His-CLIC6. The subcellular distribution of the recombinant proteins were then visualized using polyclonal anti-myc3 3
antibody (red) and monoclonal anti-His antibody (green).

56 N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57
pressed in water-secreting cells, but also in endocrine cells, part in a complex with the D -like receptors with scaffold-2
such as in the pituitary where it co-exists with the D R ing proteins, such as the multi-PDZ protein, MUPP1 and2
which is known to be present in both melanotroph and radixin, which bind to D2-like receptor C-terminus (Tablelactotroph cells[31]. In these cells, the D R mediates a 1). It is interesting to note that like CLIC6, MUPP1 is2
tonic inhibitory control of hypothalamic dopamine ona- expressed in the choroid plexus where it interacts with themelanocyte-stimulating hormone and prolactin secretion, G-protein coupled receptor 5-HT2C[41]. This G proteinrespectively. This suggests that CLIC6 might also be coupled receptor presents a YRRAF motif in its C-terminalinvolved in hormone secretion, a physiological process cytoplasmic tail, close to the E–F–R–X–[AV]–F–X–Kknown to be regulated by chloride through GTP-binding consensus sequence that could be determined on dopamineproteins[32–34].Furthermore, it seems that CLIC6 is also receptor by mutagenesis for interaction with CLIC6 (Fig.expressed in astrocytes in particular in the neurohypo- 3). Swiss-PROT scan identified only 92 proteins bearingphysis consisting mainly in the axon terminals of endo- the consensus motif [FY]–R–X–[AV]–F, among whom 23crine neurons and in specialized astrocytes, named are GPCR (http: / /us.expasy.org/cgi-bin /scanprosite). Forpituicytes, which may participate in the modulation of comparison, cAMP- and cGMP-dependent protein kinasesecretion of neurohypophyseal hormones[35,36]. phosphorylation consensus site [RK](2)–x–[ST] is found
In brain, we also showed that CLIC6 expression is not in thousands of proteins.restricted to water-secreting cells since CLIC6 mRNA was In conclusion, we showed that the rat CLIC6 is afound not only in the choroid plexus, but also in neurons, submembranous protein which can form a multimericsuch as the Purkinje cells of the cerebellum where it is complex with GPCR, like the dopamine D -like receptors2
co-expressed with the D R. The SVZ, where CLIC6 and and with scaffolding proteins. These associations com-3
D R mRNA co-exist, corresponds to a remnant of the bined with the CLIC6 distribution in water- and hormone-3
neuroepithelium in which both D R[7] and CLIC6 secreting cells suggest that this CLIC member could be3
mRNAs occur during the gestational development, where involved in the regulation of secretion, presumably throughcells continue to proliferate, suggesting a role of the the regulation of chloride ion transport.CLIC6 in cell proliferation. An interaction between CLIC3and ERK7, a member of the mitogen-activated proteinkinase family, suggests that CLIC family members may A cknowledgementsplay a role in the regulation of cell growth[17]. TheD -like receptors all inhibit adenylyl cyclase and stimulate2 The authors thank O. El Far and S. Kins for advice onmitogenesis, through ERK1 and ERK2 activation
the two-hybrid system and V. Mignon for technical assis-[10,26,37,38].We used stably transfected CHO cell lines to
tance.assess the effect of CLIC6 on the D R or D R mitogenic2 3
response. We did not observed any effect of CLIC6 on3either basal or quinpirole-stimulated [ H]thymidine in-
R eferencescorporation. Similarly, we did not observe any effect ofCLIC6 on chloride efflux, whereas such an effect has been
[1] P . Sokoloff, J.C. Schwartz, The novel dopamine receptors half ashown with parchorin in LLC-PK1 cells, however in adecade later, Trends Pharmacol. Sci. 16 (1995) 270–275.different intracellular chloride measurement system[19].
[2] C . Missale, S.R. Nash, S.W. Robinson, M. Jaber, M.G. Caron,Those negative results may be explained by an absence ofDopamine receptors: from structure to function, Physiol. Rev. 78
the adequate proteins for CLIC6 to be functional in the (1998) 189–225.recipient cell we used. Additional experiments using other [3] P . Sokoloff, B. Giros, M.P. Martres, M.L. Bouthenet, J.C. Schwartz,cell recipients would be necessary to determine whether Molecular cloning and characterization of a novel dopamine re-
ceptor (D ) as a target for neuroleptics, Nature 347 (1990) 146–151.CLIC6 is involved in D -like transduction and the po- 32[4] M .L. Bouthenet, E. Souil, M.P. Martres, P. Sokoloff, B. Giros, J.C.tential functional consequences on cell proliferation in the
Schwartz, Localization of dopamine D receptor mRNA in the rat3subventricular zone or in hormone secretion in the pitui- brain using in situ hybridization histochemistry: comparison with D2tary. receptor mRNA, Brain Res. 564 (1991) 203–219.
´Emerging evidence has suggested the existence, beside[5] D . Levesque, J. Diaz, C. Pilon, M.P. Martres, B. Giros, E. Souil, D.Schott, J.L. Morgat, J.C. Schwartz, P. Sokoloff, Identification,the classical G protein-mediated receptor signalling, of acharacterization and localization of the dopamine D receptor in rat3multiprotein signalling complex and signalling compart- 3brain using 7-[ H]-hydroxy-N,N-di-n-propyl-2-aminotetralin, Proc.
mentalization, through anchoring proteins. Several CLIC Natl. Acad. Sci. USA 89 (1992) 8155–8159.family members have been shown to bind to AKAP350, a ´[6] J . Diaz, D. Levesque, C.H. Lammers, N. Griffon, M.P. Martres, J.C.protein which is able to scaffold a number of protein Schwartz, P. Sokoloff, Phenotypical characterization of neurons
expressing the dopamine D receptor, Neuroscience 65 (1995)kinases and phosphatases, through their carboxyl termini 3
731–745.[39] whereas CLIC4 binds directly to brain dynamin I in a[7] J . Diaz, S. Ridray,V. Mignon, N. Griffon, J.C. Schwartz, P. Sokoloff,
complex containing actin, tubulin and 14-3-3 proteins[40] Selective expression of dopamine D receptor mRNA in prolifer-3and CLIC5 interacts with both ezrin and F-actin cyto- ative zones during embryonic development of the rat brain, J.skeleton of placental microvilli[18]. CLIC6 could take Neurosci. 17 (1997) 4282–4292.

N. Griffon et al. / Molecular Brain Research 117 (2003) 47–57 57
[8] D . Vallone, R. Picetti, E. Borrelli, Structure and function of D receptor in a transfected NG 108-15 neuroblastoma-glioma3
dopamine receptors, Neurosci. Biobehav. Rev. 24 (2000) 125–132. hybrid cell line, Eur. J. Pharmacol. [Mol. Pharmacol. Sect.] 268[9] N . Griffon, C. Pilon, F. Sautel, J.C. Schwartz, P. Sokoloff, Two (1994) 129–139.
intracellular pathways for the dopamine D receptor: opposite and [27] P . Strippoli, P. D’addabbo, L. Lenzi, S. Giannone, S. Canaider, R.3
synergistic interactions with cyclic AMP, J. Neurochem. 68 (1997) Casadei, L. Vitale, P. Carinci, M. Zannotti, Segmental parology in1–9. the human genome: a large-scale triplication on 1p, 6p and 21q,
[10] D . Cussac, A. Newman-Tancredi, V. Pasteau, M.J. Millan, Human Mamm. Genome 13 (2002) 456–462.dopamine D receptors mediate mitogen-activated protein kinase [28] K . Warton, R. Tonini, W.D. Fairlie, J.M. Matthews, S.M. Valenzuela,3
activation via phosphatidylinositol 3-kinase and an atypical protein M.R. Qiu, W.M. Wu, S. Pankhurst, A.R. Bauskin, S.J. Harrop, T.J.kinase C-dependent mechanism, Mol. Pharmacol. 56 (1999) 1025– Campbell, P.M. Curmi, S.N. Breit, M. Mazzanti, Recombinant1030. CLIC1 (NCC27) assembles in lipid bilayers via a pH-dependent
[11] R . Lin, K. Karpa, N. Kabbani, P. Goldman-Rakic, R. Levenson, two-state process to form chloride ion channels with identicalDopamine D2 and D3 receptors are linked to the actin cytoskeleton characterisitcs to those observed in chinese hamster ovary cellsvia interaction with filamin A, Proc. Natl. Acad. Sci. USA 98 (2001) expressing CLIC1, J. Biol. Chem. 277 (2002) 26003–26011.5258–5263. [29] S .J. Harrop, M.Z. DeMaere, W.D. Fairlie, T. Resztva, S.M. Val-
[12] F .D. Smith, G.S. Oxford, S.L. Milgram, Association of the D2 enzuela, M. Mazzanti, R. Tonini, M.R. Qiu, L. Jankova, K. Warton,dopamine receptor third cytoplasmic loop with spinophilin, a protein A.R. Bauskin, W.M. Wu, S. Pankhurst, T.J. Campbell, S.N. Breit,phosphatase interacting protein, J. Biol. Chem. 247 (1999) 19894– P.M. Curmi, Crystal structure of a soluble form of the intracellular19900. chloride ion channel CLIC1 (NCC27) at 1.4-A resolution, J. Biol.
[13] A .V. Binda, N. Kabbani, R. Lin, R. Levenson, D2 and D3 dopamine Chem. 276 (2001) 44993–45000.receptor cell surface localization mediated by interaction with [30] Y . Mizukawa, T. Nishizawa, T. Nagao, K. Kitamura, T. Urushidani,protein 4.1N, Mol. Pharmacol. 62 (2002) 507–513. Cellular distribution of parchorin, a chloride intracellular channel-
[14] D . Landry, S. Sullivan, M. Nicolaides, C. Redhead, A. Edelman, M. related protein, in various tissues, Am. J. Physiol. Cell Physiol. 282Field, Q. Al-Awqati, J. Edwards, Molecular cloning and characteri- (2002) C786–795.zation of p64, a chloride channel protein from kidney microsomes, J. [31] J .H. Meador-Woodruff, A. Mansour, J.R. Bunzow, H.H.M. Van Tol,Biol. Chem. 268 (1993) 14948–14955. S.J. Watson, O. Civelli, Distribution of D dopamine receptor2
[15] S . Howell, R.R. Duncan, R.H. Ashley, Identification and characteri- mRNA in rat brain, Proc. Natl. Acad. Sci. 86 (1989) 7625–7628.sation of a homologue of p64 in rat tissues, FEBS Lett. 390 (1996) [32] L . Garcia, B. Couderc, M.F. Odessa, L. Dufy-Barbe, P. Sartor,207–210. Effects of Cl(2) substitution on electrophysiological properties,
[16] S .M. Valenzuela, D.K. Martin, S.B. Por, J.M. Robbins, K. Warton, Ca(21) influx and prolactin secretion of rat lactotropes in vitro,M.R. Bootcov, P.R. Schofield, T.J. Campbell, S.N. Breit, Molecular Neuroendocrinology 70 (1999) 332–342.cloning and expression of a chloride ion channel of cell nuclei, J. [33] P . Sharma, K.E. Hagler, N.O. Dybdal, B.M. Chronwall, Salt-loadingBiol. Chem. 272 (1997) 12575–12582. induces decreased POMC mRNA levels, increased alpha-MSH
[17] Z . Qian, D. Okuhara, M.K. Abe, M.R. Rosner, Molecular cloning immunoreactivity, and sustained elevated fos expression in ratand characterization of a mitogen-activated protein kinase-associated pituitary intermediate lobe melanotropes, Ann. N. Y. Acad. Sci. 814intracellular chloride channel, J. Biol. Chem. 274 (1999) 1621– (1997) 295–299.1627. [34] M . Rupnik, R. Zorec, Intracellular Cl2 modulates Ca21-induced
[18] M . Berryman, A. Bretscher, Identification of a novel member of the exocytosis from rat melanotrophs through GTP-binding proteins,chloride intracellular channel gene family (CLIC5) that associates Pflugers Arch. 431 (1995) 76–83.with the actin cytoskeleton of placental microvilli, Mol. Biol. Cell [35] L . Garcia-Segura, J.A. Chowen, F. Naftolin, Endocrine glia: roles of11 (2000) 1509–1521. glial cells in the brain actions of steroid and thyroid hormones and
[19] T . Nishizawa, T. Nagao, T. Iwatsubo, J.G. Fort, T. Urushidani, in the regulation of hormone secretion, Front. Neuroendocrinol. 17Molecular cloning and characterization of a novel chloride intracel- (1996) 180–211.lular channel-related protein, Parchorin, expressed in water-secreting [36] C .J. Boersma, F.W. Van Leeuwen, Neuron–glia interactions in thecells, J. Biol. Chem. 275 (2000) 11164–11173. release of oxytocin and vasopressin from the rat neural lobe: the role
[20] N .S. Heiss, A. Poustka, Genomic structure of a novel chloride of opioids, other neuropeptides and their receptors, Neuroscience 62channel gene, CLIC2, in Xq28, Genomics 45 (1997) 224–228. (1994) 1003–1020.
[21] A .C. Engblom, K.E. Akerman, Determination of the chloride [37] M .E. Lajiness, C.L. Chio, R.M. Huff, D dopamine receptor2
concentration in rat brain synaptoneurosomes using a chloride- stimulation of mitogenesis in transfected Chinese hamster ovarysensitive fluorescent indicator, Biochim. Biophys. Acta 1153 (1993) cells: relationship to dopamine stimulation of tyrosine phosphoryla-262–266. tions, J. Pharmacol. Exp. Ther. 267 (1993) 1573–1581.
¨[22] C . Ullmer, K. Schmuk, A. Figge, H. Lubbert, Cloning and charac- [38] Y . Luo, G.C. Kokkonen, X. Wang, K.A. Neve, G.S. Roth, D2terization of MUPP1, a novel PDZ domain protein, FEBS Lett. 424 dopamine receptors stimulate mitogenesis through pertussis toxin-(1998) 63–68. sensitive G proteins and Ras-involved ERK and SAP/JNK pathways
[23] A . Bretscher, K. Edwards, R.G. Fehon, ERM proteins and merlin: in rat C6-D2L glioma cells, J. Neurochem. 71 (1998) 980–990.integrators at the cell cortex, Nat. Rev. Mol. Cell Biol. 3 (2002) [39] R .A. Shanks, B.T. Steadman, P.H. Schmidt, J.R. Goldenring,586–599. AKAP350 at the Golgi apparatus. I. Identification of a distinct Golgi
[24] F . Jeanneteau, F. Prieur, J. Diaz, P. Sokoloff, N. Griffon, Protective apparatus targeting motif in AKAP350, J. Biol. Chem. 277 (2002)effect of GIPC upon the dopamine D3 receptor degradation, in: 40967–40972.Dopamine 2002, IUPHAR Congress, Portland, OR, 2002, p. 68, [40] W . Suginta, N. Karoulias, A. Aitken, H. Ashley, Chloride intracellu-P1.18. lar channel protein CLIC4 (p64H1) binds directly to brain dynamin
[25] H .H.M. Van Tol, C.M.H.C. Wu Guan, K. O’Hara, J.R. Bunzow, O. I in a complex containing actin, tubulin and 14-3-3 isoforms,Civelli, J. Kennedy, P. Seeman, H.B. Niznik, V. Janovic, Multiple Biochem. J. 359 (2001) 55–64.dopamine D receptor variants in the human population, Nature 358 [41] C . Becamel, A. Figge, S. Poliak, A. Dumuis, E. Peles, J. Bockaert,4
(1992) 149–152. H. Lubbert, C. Ullmer, Interaction of serotonin 5-hydroxytryptamine´[26] C . Pilon, D. Levesque,V. Dimitriadou, N. Griffon, M.P. Martres, J.C. type 2C receptors with PDZ10 of the multi-PDZ domain protein
Schwartz, P. Sokoloff, Functional coupling of the human dopamine MUPP1, J. Biol. Chem. 276 (2001) 12974–12982.








![Non-amine dopamine transporter probe [3H]tropoxene distributes to dopamine-rich regions of monkey brain](https://static.fdokumen.com/doc/165x107/63224d2f050768990e0fcb6c/non-amine-dopamine-transporter-probe-3htropoxene-distributes-to-dopamine-rich.jpg)











