
Chemically-modified polysaccharide extract derived from Leucaena leucocephala alters Raw 264.7...
8
Preliminary report Chemically-modified polysaccharide extract derived from Leucaena leucocephala alters Raw 264.7 murine macrophage functions Amira M. Gamal-Eldeen a, ⁎ , Hassan Amer b , Wafaa A. Helmy b , Roba M. Talaat c , Halla Ragab a a Cancer Biology Laboratory, Nobel Project, Department of Biochemistry, National Research Centre, Dokki, Cairo, Egypt b Department of Natural and Microbial Products Chemistry, National Research Center, Dokki, Cairo, Egypt c Department of Molecular Biology, Genetic Engineering and Biotechnology Research Institute, Minofyia University, Egypt Received 8 October 2006; received in revised form 5 February 2007; accepted 8 February 2007 Abstract In this study, a chemical modification of the polysaccharides extract (E) derived from Leucaena leucocephala seeds was performed to prepare C-glycosidic 2-propanol derivative (PE), and its sulphated derivative (SPE). This study aimed to characterize immunomodulatory activities of the original extract and its derivatives by exploring their effects on Raw macrophage 264.7 functions and their antioxidant activity. Our results indicated that PE was an effective radical scavenger to hydroxyl, peroxyl, and superoxide anion radicals, and SPE was a peroxyl radical scavenger. PE and SPE were found to influence the macrophage functions. Both of PE and SPE enhanced the macrophage proliferation and phagocytosis of FITC-zymosan; PE inhibited nitric oxide (NO) generation and tumor necrosis factor-α (TNF-α) secretion in lipopolysaccharide (LPS)-stimulated Raw macrophage 264.7. In contrast, SPE over-induced NO generation and TNF-α secretion. Moreover, PE strongly inhibited the binding affinity of FITC-LPS to Raw 264.7, as indicated by flow cytometry analysis. These findings revealed that PE may act as a potent anti- inflammatory agent; however SPE may act as an inducer of macrophage functions against pathogens. © 2007 Elsevier B.V. All rights reserved. Keywords: Leucaena leucocephala; Macrophage; Nitric oxide; Tumor necrosis factor; LPS binding; Phagocytosis 1. Introduction Leucaena leucocephala is a tropical plant, which belongs to Leguminosae family and provides a useful source for fuel, protein, oil, and commercial gum [1–3]. It has a total carbohydrate content of approximately 35– 45%, reducing sugars about 5.2% and an average degree of polymerization of 150 [4]. The highly viscous solution of seed gum had the potential to be used as a laxative, in vegetable soups, and in other food commer- cial products [5]. L. leucocephala was reported to have few medicinal properties ranging from controlling of stomach diseases to contraception [2]. Galactomannans (Gs) are the second most abundant storage polysaccharide in Leucaena sp. They are mainly found in the endosperm cell wall of seeds from the Leguminosae family [5]. The structure of Gs is relatively simple, consisting of galactose (Gal) and mannose (Man) International Immunopharmacology 7 (2007) 871 – 878 www.elsevier.com/locate/intimp ⁎ Corresponding author. Biochemistry Department, Genetic Engi- neering and Biotechnology Division, National Research Centre, Dokki 12622, Cairo, Egypt. Tel.: +20 10 60 53 903; fax: +20 2 33 70 931. E-mail addresses: [email protected], [email protected] (A.M. Gamal-Eldeen). 1567-5769/$ - see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.intimp.2007.02.002
Transcript of Chemically-modified polysaccharide extract derived from Leucaena leucocephala alters Raw 264.7...

International Immunopharmacology 7 (2007) 871–878www.elsevier.com/locate/intimp
Preliminary report
Chemically-modified polysaccharide extract derived from Leucaenaleucocephala alters Raw 264.7 murine macrophage functions
Amira M. Gamal-Eldeen a,⁎, Hassan Amer b, Wafaa A. Helmy b,Roba M. Talaat c, Halla Ragab a
a Cancer Biology Laboratory, Nobel Project, Department of Biochemistry, National Research Centre, Dokki, Cairo, Egyptb Department of Natural and Microbial Products Chemistry, National Research Center, Dokki, Cairo, Egypt
c Department of Molecular Biology, Genetic Engineering and Biotechnology Research Institute, Minofyia University, Egypt
Received 8 October 2006; received in revised form 5 February 2007; accepted 8 February 2007
Abstract
In this study, a chemical modification of the polysaccharides extract (E) derived from Leucaena leucocephala seeds wasperformed to prepare C-glycosidic 2-propanol derivative (PE), and its sulphated derivative (SPE). This study aimed to characterizeimmunomodulatory activities of the original extract and its derivatives by exploring their effects on Raw macrophage 264.7functions and their antioxidant activity. Our results indicated that PE was an effective radical scavenger to hydroxyl, peroxyl, andsuperoxide anion radicals, and SPE was a peroxyl radical scavenger. PE and SPE were found to influence the macrophagefunctions. Both of PE and SPE enhanced the macrophage proliferation and phagocytosis of FITC-zymosan; PE inhibited nitricoxide (NO) generation and tumor necrosis factor-α (TNF-α) secretion in lipopolysaccharide (LPS)-stimulated Raw macrophage264.7. In contrast, SPE over-induced NO generation and TNF-α secretion. Moreover, PE strongly inhibited the binding affinity ofFITC-LPS to Raw 264.7, as indicated by flow cytometry analysis. These findings revealed that PE may act as a potent anti-inflammatory agent; however SPE may act as an inducer of macrophage functions against pathogens.© 2007 Elsevier B.V. All rights reserved.
Keywords: Leucaena leucocephala; Macrophage; Nitric oxide; Tumor necrosis factor; LPS binding; Phagocytosis
1. Introduction
Leucaena leucocephala is a tropical plant, whichbelongs to Leguminosae family and provides a usefulsource for fuel, protein, oil, and commercial gum [1–3].It has a total carbohydrate content of approximately 35–
⁎ Corresponding author. Biochemistry Department, Genetic Engi-neering and Biotechnology Division, National Research Centre, Dokki12622, Cairo, Egypt. Tel.: +20 10 60 53 903; fax: +20 2 33 70 931.
E-mail addresses: [email protected], [email protected](A.M. Gamal-Eldeen).
1567-5769/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.intimp.2007.02.002
45%, reducing sugars about 5.2% and an average degreeof polymerization of 150 [4]. The highly viscoussolution of seed gum had the potential to be used as alaxative, in vegetable soups, and in other food commer-cial products [5]. L. leucocephala was reported to havefew medicinal properties ranging from controlling ofstomach diseases to contraception [2].
Galactomannans (Gs) are the second most abundantstorage polysaccharide in Leucaena sp. They are mainlyfound in the endosperm cell wall of seeds from theLeguminosae family [5]. The structure of Gs is relativelysimple, consisting of galactose (Gal) and mannose (Man)

872 A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
with a β [1–4] Man backbone solubilized with a short α[1–6] linked Gal side chain [6]. The Man/Gal ratio isvariable among species, portions or fractions. Theproperties of Gs depend on their chemical structures i.e.chain length, Gal/Man ratio, cis-OH groups, sterichindrance, substitutions, and degree of polymerization[7,8]. They have biological activities including cancerchemopreventive, anti-cancer [9], immunostimmulation[9,10], anti-viral [11], anticoagulant and anti-thrombotic[12] activities. Sulphated polysaccharides were alsoreported to have in vitro anti-viral and anticoagulantactivities, which were attributed to the negatively chargedsulphate groups [13].
Macrophages are the first line of defense in innateimmunity against microbial infection. Professional pha-gocytes engulf and kill microorganisms and presentantigens for triggering adaptive immune responses [14].During phagocytosis, macrophages secrete preformedgranule constituents and newly synthesized products thatplay a critical role in inflammation and tissue repair[14]. Accordingly, phagocytic activity and macrophagereactivity against bacterial epitopes are crucial in theassessment of macrophage function. Macrophage defenseagainst pathogens includes cytokine secretions like tumornecrosis factor-α (TNF-α), and inflammation mediatorslike nitric oxide (NO). In phagocytes, NO is produced inlarge quantities by the action of inducible NO synthetase(iNOS), and cyclooxygenase-2 (COX-2) [15].
Phytochemicals are anticipated to be highly useful inproviding new immunomodulators as immunostimulatorsor immunosuppressors to help in the treatment of differentdiseases such as inflammatory diseases and immunode-ficiency disorders. In a trial to find new immunomodu-lators, we recently modified the guar gum structure, in away that improve and develop its immunomodulatory,anti-inflammatory, cancer chemopreventive properties byC-glycosylation and by an additional sulphation [16].Consequently, the present work is another attempt tomodify the structure of L. leucocephala polysaccharideextract, which is mainly Gs. This modification may alterthe macrophage functions and provide new immunomod-ulatory properties to the inactive crude extract. Theultimate goal is to use these derivatives as alternatives ofL. leucocephala polysaccharide extract (gum) in produc-tion of healthy foods.
2. Material and methods
2.1. Plant extraction and chemical modification
L. leucocephala gel complexwas extracted frommilled seed(5g),withdistilledwater (200ml×3 times)at 100 °Cunder reflux
for 1 h as previously described [17]. The combined extracts werecollected by centrifugation and most of the protein was removedby 20% trichloroacetic acid precipitation, and then dialyzedagainst distilled water for 2 days. The retentate was concentratedand then precipitated with 3 volumes of ethanol. The resultedprecipitatewas isolated by centrifugation,washedby acetone andthen lyophilized.
According to a previously reported methodology [18],200 mg of the lyophilized extract (E), NaHCO3 (200 mg) andpentane-2, 4-dione (300 μl) were stirred in 20mlwater at 100 °Cfor 20 h. The solution was diluted and neutralized with Dowex50 resin (H+ form). The resin was filtered off, and the retentatewas mixed with 100 mg NaBH4. The reaction mixture wasstirred for 24 h at room temperature, and the excess NaBH4 wasneutralized by glacial acetic acid. The solution was dialyzedagainst running distilled water for 48 h before lyophilized. Thesulphation of the dried 2, 4-pentanedione-treated extract (PE),to prepare SPE, was performed as follows: 0.3 g PE wassuspended in 2 ml of dry formamide and stirred at roomtemperature for 24 h. A solution was prepared by dropping 5 mlof HClSO3 on 20 ml of formamide in ice, and added to the PE-mixture. The reaction was neutralized by 30% NaOH solution,and dialyzed against running distilled water for 48 h beforelyophilized.
The total carbohydrates content of PE and SPE wasdetermined by phenol-sulphuric acid assay [19], and the totalprotein was assayed according to [20]. The sugar compositionwas determined after complete hydrolysis with H2SO4 (2 mol/l) at 100 °C for 8 h. The mixture was neutralized with BaCO3,centrifuged, filtered, neutralized with Dowex 50 resin(H+ form) and concentrated. The paper chromatography ofhydrolysate was carried out by the descending method onWhatman No.1 paper using butanol:acetone:water (4:5:1). Thesugars were detected by aniline phthalate [21], and thesulphate content was determined after hydrolysis with HCl[22] and liberated sulphate ions were determined by the BaCl2turbidimetric method [23].
2.2. Cell culture
Murine macrophage cell line (Raw 264.7, ATCC, USA)was routinely cultured in RPMI-1640 supplemented with 10%fetal bovine serum (FBS), 2 mM L-glutamine, 100 units/mlpenicillin sodium, 100 units/ml streptomycin sulphate, and250 ng/ml amphotericin B. Cells were maintained sub-confluent at 37 °C in humidified air containing 5% CO2.Raw 264.7 cells were harvested by gentle scraping. Cells wereused when confluence had reached 75%. Before dissolving inmedium, E, PE and SPE were tested for endotoxin usingPyrogent® Ultra gel clot assay, and they were found endotoxinfree. Optimum concentrations were selected to be used in therest of the study. All experiments were repeated four times,unless mentioned, and the data were represented as meanvalues. Cell culture material was obtained from CambrexBioScience (Copenhagen, Denmark). Chemicals were pur-chased from sigma (USA), except mentioned.
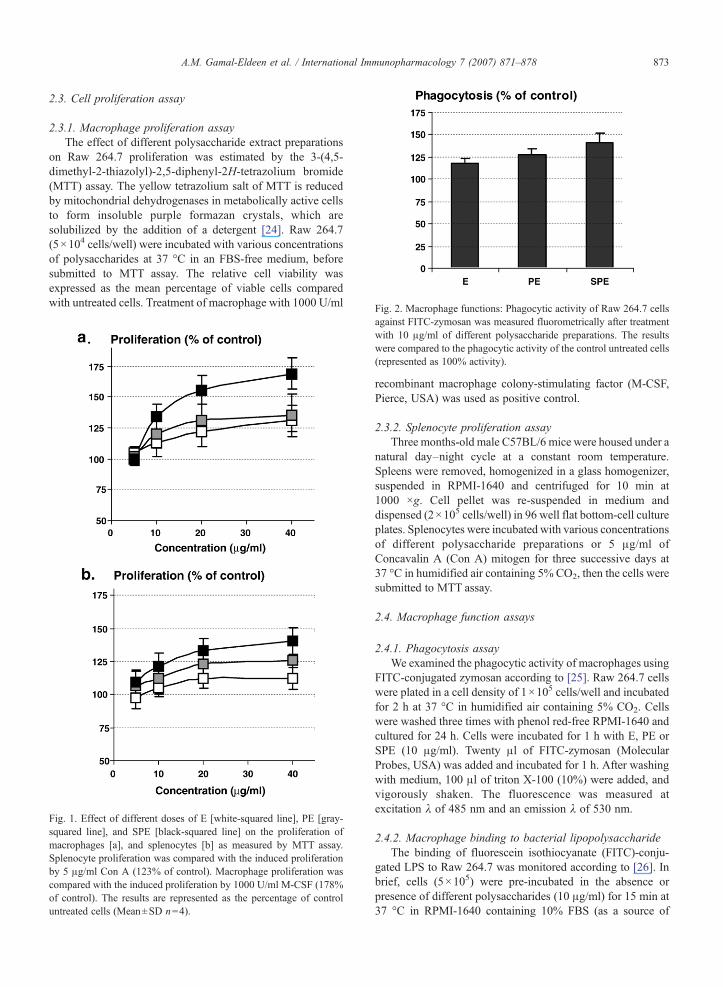
Fig. 2. Macrophage functions: Phagocytic activity of Raw 264.7 cells
873A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
2.3. Cell proliferation assay
2.3.1. Macrophage proliferation assayThe effect of different polysaccharide extract preparations
on Raw 264.7 proliferation was estimated by the 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide(MTT) assay. The yellow tetrazolium salt of MTT is reducedby mitochondrial dehydrogenases in metabolically active cellsto form insoluble purple formazan crystals, which aresolubilized by the addition of a detergent [24]. Raw 264.7(5×104 cells/well) were incubated with various concentrationsof polysaccharides at 37 °C in an FBS-free medium, beforesubmitted to MTT assay. The relative cell viability wasexpressed as the mean percentage of viable cells comparedwith untreated cells. Treatment of macrophage with 1000 U/ml
Fig. 1. Effect of different doses of E [white-squared line], PE [gray-squared line], and SPE [black-squared line] on the proliferation ofmacrophages [a], and splenocytes [b] as measured by MTT assay.Splenocyte proliferation was compared with the induced proliferationby 5 μg/ml Con A (123% of control). Macrophage proliferation wascompared with the induced proliferation by 1000 U/ml M-CSF (178%of control). The results are represented as the percentage of controluntreated cells (Mean±SD n=4).
against FITC-zymosan was measured fluorometrically after treatmentwith 10 μg/ml of different polysaccharide preparations. The resultswere compared to the phagocytic activity of the control untreated cells(represented as 100% activity).
recombinant macrophage colony-stimulating factor (M-CSF,Pierce, USA) was used as positive control.
2.3.2. Splenocyte proliferation assayThree months-old male C57BL/6 mice were housed under a
natural day–night cycle at a constant room temperature.Spleens were removed, homogenized in a glass homogenizer,suspended in RPMI-1640 and centrifuged for 10 min at1000 ×g. Cell pellet was re-suspended in medium anddispensed (2×105 cells/well) in 96 well flat bottom-cell cultureplates. Splenocytes were incubated with various concentrationsof different polysaccharide preparations or 5 μg/ml ofConcavalin A (Con A) mitogen for three successive days at37 °C in humidified air containing 5% CO2, then the cells weresubmitted to MTT assay.
2.4. Macrophage function assays
2.4.1. Phagocytosis assayWe examined the phagocytic activity of macrophages using
FITC-conjugated zymosan according to [25]. Raw 264.7 cellswere plated in a cell density of 1×105 cells/well and incubatedfor 2 h at 37 °C in humidified air containing 5% CO2. Cellswere washed three times with phenol red-free RPMI-1640 andcultured for 24 h. Cells were incubated for 1 h with E, PE orSPE (10 μg/ml). Twenty μl of FITC-zymosan (MolecularProbes, USA) was added and incubated for 1 h. After washingwith medium, 100 μl of triton X-100 (10%) were added, andvigorously shaken. The fluorescence was measured atexcitation λ of 485 nm and an emission λ of 530 nm.
2.4.2. Macrophage binding to bacterial lipopolysaccharideThe binding of fluorescein isothiocyanate (FITC)-conju-
gated LPS to Raw 264.7 was monitored according to [26]. Inbrief, cells (5×105) were pre-incubated in the absence orpresence of different polysaccharides (10 μg/ml) for 15 min at37 °C in RPMI-1640 containing 10% FBS (as a source of
874 A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
CD14 and LPS binding protein to induce binding) beforeincubated for 1 h with FITC-conjugated LPS (200 ng/ml).After washing with phosphate buffered saline (PBS), thebinding of FITC-LPS was analyzed by flow cytometry(MoFlo, DakoCytomation, Denmark), and median fluores-cence intensity was determined.
2.4.3. Nitrite assayThe accumulation of nitrite, an indicator of nitric oxide (NO)
synthesis, wasmeasured byGriess reagent [27]. Raw 264.7weregrown in phenol red-free RPMI-1640 containing 10% FBS.Cells were incubated for 24 h with LPS (1 μg/ml) in the presenceor absence of different polysaccharides (10 μg/ml). Fifty μl ofcell culture supernatant was mixed with 50 μl of Griess reagentand incubated for 10 min. The absorbance was measuredspectrophotometrically at 550 nm. A standard curve was blottedusing serial concentrations of sodium nitrite. The nitrite contentwas normalized to the cellular protein content as measured bybicinchoninic acid assay [28].
2.4.4. Determination of tumor necrosis factor-αCultured supernatants were harvested after 24 h of incubation
of LPS-stimulated Raw 264.7 with or without differentpolysaccharides (10 μg/ml), and then stored at −80 °Cuntil used. TNF-α was determined in the supernatants using
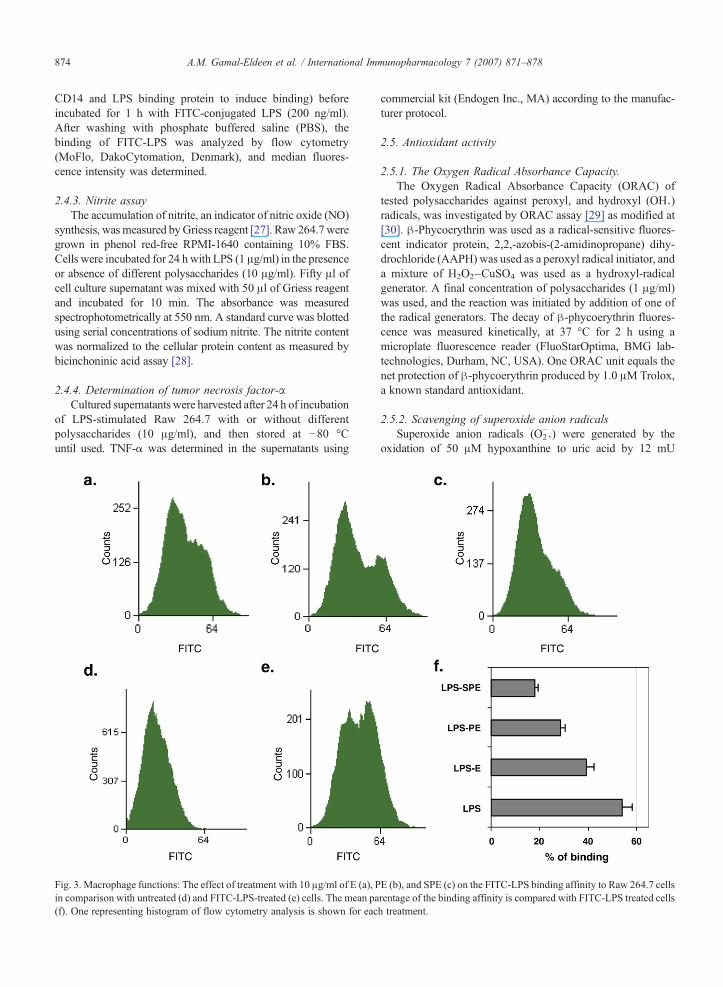
Fig. 3. Macrophage functions: The effect of treatment with 10 μg/ml of E (a), Pin comparison with untreated (d) and FITC-LPS-treated (e) cells. The mean pa(f). One representing histogram of flow cytometry analysis is shown for eac
commercial kit (Endogen Inc., MA) according to the manufac-turer protocol.
2.5. Antioxidant activity
2.5.1. The Oxygen Radical Absorbance Capacity.The Oxygen Radical Absorbance Capacity (ORAC) of
tested polysaccharides against peroxyl, and hydroxyl (OHU)radicals, was investigated by ORAC assay [29] as modified at[30]. β-Phycoerythrin was used as a radical-sensitive fluores-cent indicator protein, 2,2,-azobis-(2-amidinopropane) dihy-drochloride (AAPH) was used as a peroxyl radical initiator, anda mixture of H2O2–CuSO4 was used as a hydroxyl-radicalgenerator. A final concentration of polysaccharides (1 μg/ml)was used, and the reaction was initiated by addition of one ofthe radical generators. The decay of β-phycoerythrin fluores-cence was measured kinetically, at 37 °C for 2 h using amicroplate fluorescence reader (FluoStarOptima, BMG lab-technologies, Durham, NC, USA). One ORAC unit equals thenet protection of β-phycoerythrin produced by 1.0 μM Trolox,a known standard antioxidant.
2.5.2. Scavenging of superoxide anion radicalsSuperoxide anion radicals (O2
−U) were generated by theoxidation of 50 μM hypoxanthine to uric acid by 12 mU
E (b), and SPE (c) on the FITC-LPS binding affinity to Raw 264.7 cellsrentage of the binding affinity is compared with FITC-LPS treated cellsh treatment.
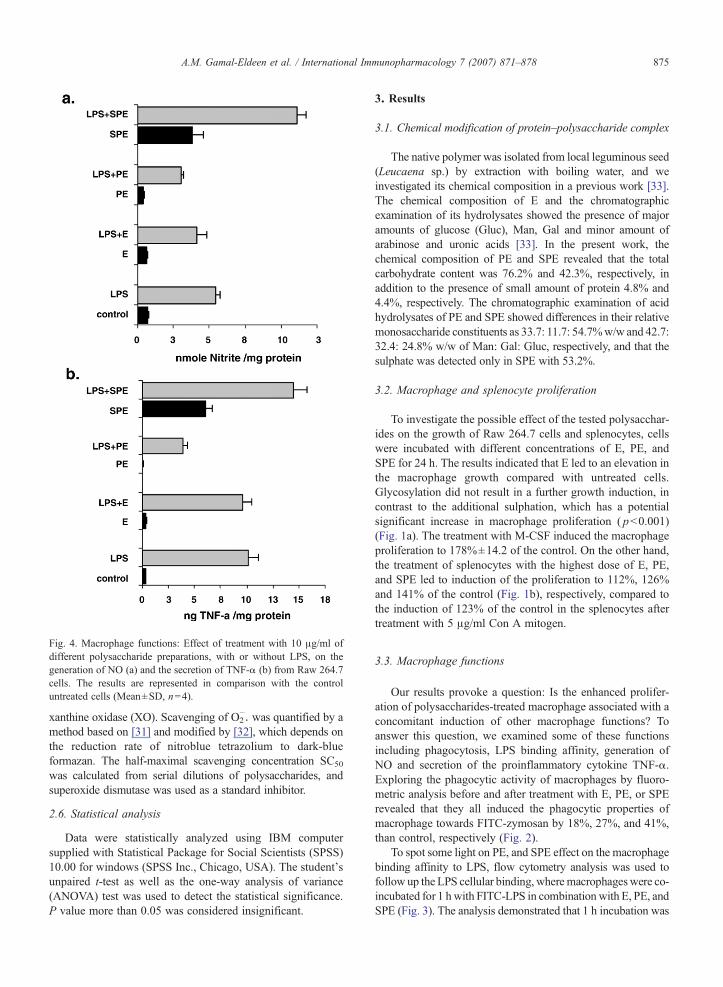
Fig. 4. Macrophage functions: Effect of treatment with 10 μg/ml ofdifferent polysaccharide preparations, with or without LPS, on thegeneration of NO (a) and the secretion of TNF-α (b) from Raw 264.7cells. The results are represented in comparison with the controluntreated cells (Mean±SD, n=4).
875A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
xanthine oxidase (XO). Scavenging of O2−U was quantified by a
method based on [31] and modified by [32], which depends onthe reduction rate of nitroblue tetrazolium to dark-blueformazan. The half-maximal scavenging concentration SC50
was calculated from serial dilutions of polysaccharides, andsuperoxide dismutase was used as a standard inhibitor.
2.6. Statistical analysis
Data were statistically analyzed using IBM computersupplied with Statistical Package for Social Scientists (SPSS)10.00 for windows (SPSS Inc., Chicago, USA). The student'sunpaired t-test as well as the one-way analysis of variance(ANOVA) test was used to detect the statistical significance.P value more than 0.05 was considered insignificant.
3. Results
3.1. Chemical modification of protein–polysaccharide complex
The native polymer was isolated from local leguminous seed(Leucaena sp.) by extraction with boiling water, and weinvestigated its chemical composition in a previous work [33].The chemical composition of E and the chromatographicexamination of its hydrolysates showed the presence of majoramounts of glucose (Gluc), Man, Gal and minor amount ofarabinose and uronic acids [33]. In the present work, thechemical composition of PE and SPE revealed that the totalcarbohydrate content was 76.2% and 42.3%, respectively, inaddition to the presence of small amount of protein 4.8% and4.4%, respectively. The chromatographic examination of acidhydrolysates of PE and SPE showed differences in their relativemonosaccharide constituents as 33.7: 11.7: 54.7%w/w and 42.7:32.4: 24.8% w/w of Man: Gal: Gluc, respectively, and that thesulphate was detected only in SPE with 53.2%.
3.2. Macrophage and splenocyte proliferation
To investigate the possible effect of the tested polysacchar-ides on the growth of Raw 264.7 cells and splenocytes, cellswere incubated with different concentrations of E, PE, andSPE for 24 h. The results indicated that E led to an elevation inthe macrophage growth compared with untreated cells.Glycosylation did not result in a further growth induction, incontrast to the additional sulphation, which has a potentialsignificant increase in macrophage proliferation ( pb0.001)(Fig. 1a). The treatment with M-CSF induced the macrophageproliferation to 178%±14.2 of the control. On the other hand,the treatment of splenocytes with the highest dose of E, PE,and SPE led to induction of the proliferation to 112%, 126%and 141% of the control (Fig. 1b), respectively, compared tothe induction of 123% of the control in the splenocytes aftertreatment with 5 μg/ml Con A mitogen.
3.3. Macrophage functions
Our results provoke a question: Is the enhanced prolifer-ation of polysaccharides-treated macrophage associated with aconcomitant induction of other macrophage functions? Toanswer this question, we examined some of these functionsincluding phagocytosis, LPS binding affinity, generation ofNO and secretion of the proinflammatory cytokine TNF-α.Exploring the phagocytic activity of macrophages by fluoro-metric analysis before and after treatment with E, PE, or SPErevealed that they all induced the phagocytic properties ofmacrophage towards FITC-zymosan by 18%, 27%, and 41%,than control, respectively (Fig. 2).
To spot some light on PE, and SPE effect on the macrophagebinding affinity to LPS, flow cytometry analysis was used tofollow up the LPS cellular binding, wheremacrophageswere co-incubated for 1 hwith FITC-LPS in combination with E, PE, andSPE (Fig. 3). The analysis demonstrated that 1 h incubation was

Table 1Radical-scavenging activity and antioxidant capacity of differentpolysaccharides
Sample ORAC−OHUa(units)
ORAC−ROOUa(units)
X/XO(O2−U) bSC50
(μg/ml)
E 0.8 0.6 146.2PE 2.6 1.5 74.3SPE 0.9 1.8 N250a 1.0 ORAC unit equals the net protection of β-phycoerythrin
produced by 1.0 μM Trolox.b SC50: Half-maximal scavenging concentration.
876 A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
enough for 54% of the macrophage population to bind FITC-LPS, however this binding was significantly inhibited 27.78%( pb0.05), 47.22 ( pb0.01), and 66.67% ( pb0.005) aftertreatment with E, PE, and SPE, respectively.
To enhance iNOS expression and NO production, Raw264.7 were stimulated with LPS and the nitrite level wasmeasured in cell culture supernatants. Although the treatmentwith PE resulted in a potent inhibition of the generated NO ashigh as 51.30% ( pb0.005), surprisingly SPE treatmentresulted in a strong significant over-induction of NO generation( pb0.001), to 2.21-fold of the LPS-stimulated NO generation(Fig. 4a). Similarly, omitting pretreatment with LPS, Thetreatment of Raw 264.7 with SPE alone induced the NOgeneration to an extent that is equivalent to 65% of thegenerated NO after treatment with LPS alone.
The effect of PE and SPE treatment on the secreted TNF-αfrom LPS-stimulated Raw 264.7 cells was evaluated usingenzyme-linked immunosorbent assay (ELISA). PE dramaticallyinhibited the induced-TNF-α concentration to 39.1% of LPS-stimulated TNF-α level ( pb0.001), while interestingly SPEover-enhanced the LPS-induced TNF-α concentration to145.25% of TNF-α concentration ( pb0.001) (Fig. 4b).Moreover, treatment with SPE alone without pretreatment withLPS lead to an enhanced TNF-α secretion from Raw 264.7,which was equivalent to 48% of the LPS-induced TNF-α.
3.4. Antioxidant activity
ORACassaywas performed to detect the scavenging activityof the polysaccharides to the physiological radicals (OHU andperoxyl). As shown in Table 1, PE was 2.6-fold and 1.5-foldmore potent than the reference compound Trolox (1 μM) inscavenging OHU and peroxyl, respectively. SPE was 1.8-foldmore active thanTrolox in scavenging peroxyl, and less effectivein scavenging OHU. In xanthine/xanthine oxidase system, PEdisplayed amoderate scavenging activity againstO2
−U, while SPEwas an inactive scavenger of O2
−U (Table 1). On the other hand, Ewas an inert scavenger for all the examined radicals.
4. Discussion
Laboratory studies revealed the critical role of poly-saccharides, especially Mans in the mechanism of im-munity [34]. We have focused our current work to extract
polysaccharides of L. leucocephala, and to modify thischeap, and widely-used polymer into an immunomodu-lator. Polysaccharides may have one terminal reducingglycosyl residue per linear polymer chain. This terminusprovides a selective and convenient site for direct covalentattachment of molecules with β-dicarbonyl compoundssuch as pentane-2, 4-dione. Knoevenagel-type conden-sation involves the reaction of the glycosyl residue withβ-dicarbonyl compounds in the presence of NaHCO3.Therefore, the reaction of L. leucocephala polysacchar-ides with pentane-2, 4-dione in the presence of NaHCO3
gave rise into C-glycosidic 2-propanone of the polysac-charide chain [18,35,36], which afforded C-glycosidic 2-propanol of corresponding polymer by NaBH4 as areducing agent.
Man-binding lectins present on macrophages can bindMan and activate the immunity via a non-self-recognitionmechanism [37]. Man receptor recognizes a repeating-unit associated with infectious agents and mediatesphagocytosis [38]. This recognition modulates the signaltransduction, cytokine production, and complementactivation. Zymosan particle, which is composed mainlyof b-glucan, Man, mannoproteins, and chitin, is recog-nized by dectin-1, Toll-like receptors (TLRs), and CD14.All are working together to enhance immune response tothe particles [39]. TLRs induce signaling through nucleartranscription factor-kappa B (NF-kB) results in thesecretion of inflammatory cytokines. The induction ofmacrophage proliferation and phagocytosis by any of thepolysaccharides indicated that the composition of galac-toglucomannan itself is responsible, to some extent, forthis induction, and that C-glycosylation and sulphationwere additive factors for both findings. The enhancedmacrophage proliferation may reflect a multiplication ofthe response to the treated polysaccharide preparations(PE and SPE) afforded by chemical modification of E.
LPS is a potent toxin released from gram negative cellwalls and it is notorious for initiation of inflammation [40].Macrophages are exclusively sensitive to the presence ofLPS and when activated they may orchestrate a multitudeof proinflammatory processes. LPS binding protein (LBP)is a glycoprotein present in sera [41] and is an acute phaseprotein, which increases in response to inflammatorystimuli. The resulting LPS-LBP complex binds to CD14receptor, which is a primary receptor for LPS that exists as asoluble form in sera and as a glycophosphatidylinositol(GPI)-linked molecule on the surface of mononuclearphagocytes [42]. Interaction of LBP and CD14 with LPSamplifies the response ofmacrophages to LPS and it resultsin the generation of proinflammatory cytokines. LPS-CD14 complex activates NF-kB as well as extracellularsignal-regulated kinase, c-Jun N-terminal kinase, and p38

877A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
mitogen-activated protein kinase, which mediate inflam-matory cytokine production [42].
Therefore, the strong inhibition of LPS bindingaffinity to macrophages after treatment with PE and SPEmay be attributed to a defect in the LPS activationconsequences. They may inhibit the interaction of LBPwith LPS, block the binding of LPS to CD14, and/orchange the configuration of the membrane surface ofmacrophage in a way that hinders the binding process.Man-rich polysaccharide may also collaborate withTLRs for the activation of immunity [39]. These factsmay explain the inhibited LPS binding due to thecompetitive recognition of receptors to the repeating-unit of Man of PE and SPE instead of that of LPS.However, the contradictory ability of PE, SPE to inducephagocytosis against zymosan and to inhibit binding toLPS, may suggest different binding mechanisms,especially that the binding mediators to LPS aredifferent from that of zymosan.
NO is a highly reactive free radical and it can form anumber of oxidation products such as NO2, NO2U,N2O3, and S-nitrosothiols [43]. Collectively, thesereactive nitrogen species have a spectrum of antimicro-bial activity mediated by their ability to react with keyprotein and lipid molecules in microbes [43]. Thus,inhibition of the generated NO by PE may be due to adirect scavenging capacity to NO, an inhibition of theiNOS pathway, and/or a modulation of other factors inthe NO cascade such as transcriptional factors.
The inhibition of TNF-α by PE may be also due to adepressed level of the extracellular signal-regulatedkinases resulted from the consequence inhibition ofLPS-CD14 complexes as predicted from the associatedPE-inhibition of LPS binding affinity to Raw 264.7cells. The effective ability of PE to inhibit the stimulatedNO and TNF-α may postulate a strong role in theinflammation cascade as anti-inflammatory agents. Onthe other hand the enhancing properties of SPE of NOgeneration, TNF-α secretion, phagocytosis, and prolif-eration indicated the immunomodulatory role of SPE asantimicrobial agent, which need to be explored againstliving microbes. In the presence of SPE, althoughmacrophage bound only one third of LPS, this lowbinding was leading to high activation of macrophage.This may be due to the remarkable activation ofmacrophage by SPE alone in the absence of LPS, andadditional indirect activation for the mediators in NOand TNF-α cascade.
Investigating the antioxidant activity resulted in avariable scavenging activity against different physio-logically relevant radicals. It seems that C-glycosylationpotentiates the affinity of the original extract against all
radicals. Sulphation seems to be responsible for theenhanced scavenging affinity only against peroxylradicals, while it depressed affinity against OHU andO2−U. Considering the known high molecular weights of
polysaccharides, the effective concentration values arerepresenting low effective molar concentrations in all ofthe experiments. The overall findings of SPE indicatedthat the sulphur content in the polysaccharide backbone,which represents the actual difference in structurerelationship activity between E and PE, evoked theinduction of the inflammatory mediators. While in theother activities, the increase seems accumulative from Eto additional C-glycosylation to additional sulphation,and not a specific advantage for the sulphur content.
Taken together, in a novel trial we successfully modifythe structure of the extracted L. leucocephala polysac-charide extract that leads to alteration of macrophagefunctions. C-glycosylation afforded anti-inflammatorypotentials, while additional sulphation afforded neweffective antimicrobial agent. Both modified polysacchar-ides could be used as an alternative of the originalextracted L. leucocephala polysaccharide in food indus-try. However the activity and the bioavailability of PE andSPE need to be explored in in vivo models.
Acknowledgements
We thank Prof. Mohammad A. Ali, Virology labora-tory, NRC for his support, cooperation and supply offacilities. This work was funded by the National ResearchCentre (NRC), Cairo, Egypt.
References
[1] Rushkin FR, Leucaena: promising forage and tree crops for thetropics. Washington, DC: National Research Council, NationalAcademy Press, 1984.
[2] Jagan Mohan Rao S, Azeemoddin G. Recovery of lecithin andrefining of subabul (Leucaena leucocephala) seed oil. J OilTechnol Assoc India 1988;20:16–7.
[3] Buckeridge MS, Dietrich SMC, Maluf AM. Galactomannan inseeds of different populations of Leucaena leucocephala. RevBras Bot 1987;10:25–7.
[4] Anderson DMW, Howlett JF, McNab CGA. The hydroxyprolinecontent of gum exudates from several plant genera. Phytochem-istry 1986;26:309–11.
[5] Reid JSG. Cell wall storage carbohydrates in seeds: biochemistry ofthe seed gums and hemicelluloses. Adv Bot Res 1985;11:125–55.
[6] Edwards ME, Marshall E, Gidley MJ, Reid GSJ. Transferspecificity of detergent-solubilized fenugreek galactomannangalactosyltransferase. Plant Physiol 2002;129:1391–7.
[7] Shcherbukhin VD. Galactomannans of native flora (review).Appl Biochem Microbiol 1993;29:599–606.
[8] Mercê ALR, Fernandes E, Mangrich AS, Sierakowski MR.Evaluation of the complexes of Galactomannan of Leucaena

878 A.M. Gamal-Eldeen et al. / International Immunopharmacology 7 (2007) 871–878
leucocephala and Co2+, Mn2+, Ni2+ and Zn2+. J Braz Chem Soc2000;11:224–31.
[9] Ingolfsdottir K, Jurcic K, Fischer B, Wagner H. Immunologicallyactive polysaccharide fromCetaria islandica. PlantaMed 1994;60:527–31.
[10] Ramesh HP, Yamaki K, Tsushida T. Effect of fenugreek(Trigonella foenum-graecum L.) galactomannan fractions onphagocytosis in rat macrophages and on proliferation and IgMsecretion in HB4C5 cells. Carbohydr Polym 2002;50:79–83.
[11] Herold BC, Gerber SI, Polonsky T, Belval BJ, Shaklee PN,Holme K. Identification of structural features of heparin requiredfor inhibition of herpes simplex virus type 1 binding. Virology1995;206:1108–16.
[12] Martinichen-Herrero JC, Carbonero ER, Sassaki GL, Gorin PA,Iacomini M. Anticoagulant and antithrombotic activities of achemically sulphated galactoglucomannanobtained from the lichenCladonia ibitipocae. Int J Biol Macromol 2005;35:97–102.
[13] Ono L, Wollinger W, Rocco IM, Coimbra TLM, Gorin PAJ,Sierakowski MR. In vitro and in vivo antiviral properties ofsulphated galactomannans against yellow fever virus (BeH111strain) and dengue 1 virus (Hawaii strain). Antivir Res2003;60:60201–8.
[14] Girotti M, Evans JH, Burke D, Leslie CC. Cytosolic phospholipaseA2 translocates to forming phagosomes during phagocytosis ofzymosan in macrophages. J Biol Chem 2004;279:19113–21.
[15] MacMicking J, Xie QW, Nathan C. Nitric oxide and macrophagefunction. Annu Rev Immunol 1997;15:323–50.
[16] Gamal-Eldeen AM, Amer H, Helmy WA. Cancer chemopreven-tive and anti-inflammatory activities of chemically modified guargum. Chem Biol Interact 2006;161:229–40.
[17] Whistler RL, Saarino J. Galactomannan from Soybean Hulls.J Am Chem Soc 1957;79:6055–7.
[18] Graziani A, Amer H, Zamyatina A, Hofinger A, Kosma P.Synthesis of C-glycosides related to glycero-β-D-manno-hep-toses. Tetrahedron Asymmetry 2005;16:167–75.
[19] Dubois M, Gillls KA, Hamilton JK, Rebers PA, Smith F.Colourimetric method for determination of sugars and relatedsubstances. Anal Chem 1956;28:350–6.
[20] Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ. Proteinmeasurement with the Folin-phenol reagent. J Biol Chem1952;193:265–75.
[21] Partridge SM. Aniline hydrogen phthalate as spraying reagent forchromatography of sugars. Nature 1949;164:443–7.
[22] Larsen B, Haug A, Painter JTJ. Sulphated polysaccharides inbrown algae I-isolation and preliminary characterization of threesulphated polysaccharides from Ascophylum nodosum. ActaChem Scand 1966;20:219–30.
[23] Garrido ML. Determination of sulphur in plant material. Analyst1962;89:61–6.
[24] Hansen MB, Nielsen SE, Berg K. Re-examination and furtherdevelopment of a precise and rapid dye method for measuringcell growth/cell kill. J Immunol Methods 1989;119:203–10.
[25] Kim KI, Shin KS, Jun WJ, Hong BS, Shin DH, Cho HY, et al.Effects of polysaccharides from Rhizomes of Curcuma zedoariaon macrophage function. Biosci Biotechnol Biochem 2001;65:2369–77.
[26] Carracedo J, Ramirez R, Martin-Malo A, Rodriguez M, AljamaP. The effect of LPS, uraemia, and haemodialysis membrane
exposure on CD14 _expression in mononuclear cells and itsrelation to apoptosis. Nephrol Dial Transplant 2002;17:428–34.
[27] Gerhäuser C, Klimo K, Heiss E, Neumann I, Gamal-Eldeen A,Knauft J, et al. Mechanism-based in vitro screening of potentialcancer chemopreventive agents. Mutat Res 2003;523–524:163–72.
[28] Smith PK, Krohn RI, Hermanson GT, Mallia AK, Gartner FH,Provenzano MD, et al. Measurement of protein usingbicinchoninic. Acid Anal Biochem 1985;150:76–85.
[29] CaoG, PriorRL.Measurement of oxygen radical absorbance capacityin biological samples. Methods Enzymol 1999;299:50–62.
[30] Gamal-Eldeen A, Kawashty S, Ibrahim L, Shabana M, NegoumyS. Evaluation of Antioxidant, anti-inflammatory, and antinoci-ceptive properties of aerial parts of Vicia sativa and its flavonoids.J Nat Rem 2004;4:81–96.
[31] Ukeda H, Maeda S, Ishii T, Sawamura M. Spectrophotometricassay for superoxide dismutase based on tetrazolium salt 3,1-(phenylamino(-carbonyl-3,4-tetrazolium)-bis(4-methoxy-6-nitro)benzenesulphonic acid hydrate reduction by xanthine/xanthine oxidase. Anal Biochem 1997;251:206–9.
[32] Gerhäuser C, Alt A, Heiss E, Gamal-Eldeen A, Klimo K, Knauft J,et al. Cancer chemopreventive activity of xanthohumol, a naturalproduct derived from hop. Mol Cancer Ther 2002;1:959–69.
[33] Hussein MM, Helmy WA, Salem HM. Biological activities ofsome galactomannans and their sulfated derivatives. Phytochem-istry 1998;48:479–84.
[34] Rudd PM,WormaldMR, Dwek RA. Mini Review. Glycosylationand the immune system. Trends Glycosci Glycotechnol 1999;11:1–21.
[35] Rodrigues F, Canac Y, Lubineau A. A convenient, one-step,synthesis of β-c-glycosidic ketones in aqueous media. ChemCommun 2000;42:2049–50.
[36] Wang Z, Shao H, Lacroix E, Wu SH, Jennings HJ, Zou W.Epimerization of 2′carboxyl-c-glycosides via enolation, beta-elimination and intramolecular cyclo-addition. J Org Chem2003;68:8097–105.
[37] Vasta GR, Quesenberry M, Ahmed H, O'Leary N. C-type lectinsand galectins mediate innate and adaptive immune functions:roles in the complement activation pathway. Dev Comp Immunol1999;23:401–20.
[38] Aderem A, Underhill DM. Mechanisms of phagocytosis inmacrophages. Annu Rev Immunol 1999;17:593–623.
[39] Gantner BN, Simmons RM, Canavera SJ, Akira S, UnderhillDM. Collaborative induction of inflammatory responses bydectin-1 and Toll-like receptor 2. J Exp Med 2003;197:1107–17.
[40] Ohlsson K, Bergenfeldt M, Bjork P. Functional studies of humansecretory leukocyte protease inhibitor. Adv Exp Med Biol1988;240:123–31.
[41] Schumann RR, Leong SR, Flaggs GW, Gray PW, Wright SD,Mathison JC, et al. Structure and function of lipopolysaccharidebinding protein. Science 1990;249:1429–31.
[42] Kitchens RL. Role of CD14 in cellular recognition of bacteriallipopolysaccharides. Chem Immunol 2000;74:61–7.
[43] Zhu L, Gunn C, Beckman J. Bactericidal activity of peroxynitrite.Arch Biochem Biophys 1992;298:452–7.










