
Characterization of an acquired dps -containing gene island in the lactic acid bacterium Oenococcus...
10
ORIGINAL ARTICLE Characterization of an acquired dps-containing gene island in the lactic acid bacterium Oenococcus oeni A. Athane ´ 1 , E. Bilhe ` re 1 , E. Bon 1,2 , G. Morel 1 , P. Lucas 1 , A. Lonvaud 1 and C. Le Marrec 1 1 UMR 1219, INRA-Universite ´ Victor Segalen Bordeaux 2 – Universite ´ Bordeaux 1, Talence Cedex, France 2 Universite ´ Victor Segalen Bordeaux 2 – LaBRI (Universite ´ de Bordeaux – CNRS) 351, Talence Cedex, France Introduction Bacteria have evolved a complex network of stress response pathways to promote their growth and ⁄ or survival during environmental challenges. One important mechanism is the DNA binding protein from starved cells (Dps), first described in Escherichia coli, where its expression was found to be activated when the bacterium finds itself in nutritionally limiting conditions or under oxidative stress, regardless of whether cells are actively growing or not (Almiron et al. 1992). Recent evidence expands the role of Dps in tolerance to acid and osmotic stresses (Choi et al. 2000; Weber et al. 2006), metals, UV-irradiation (Nair and Finkel 2004), and high pressure (Malone et al. 2006). The Dps protein from E. coli has a molecular mass of 19 kDa, and is known to be a member of the Fe-binding protein family. This family forms large (approx. 150 kDa), hexameric complexes in cells that reduce the intracellular level of iron (Fe 2+ ), thereby preventing the deleterious effect of the superoxide anion through the Fenton reaction (Almiron et al. 1992). This combined action of hydrogen peroxide and intracellular iron is of paramount importance in vivo since aerobic growth generates sufficient H 2 O 2 to create toxic levels of DNA damage in E. coli (Park et al. 2005). Other Dps proteins such as the cyanobacterial Synechococcus DpsA proteins also display catalase activity, and can therefore degrade hydrogen peroxyde (Pena and Bullerjahn 1995). The DNA binding properties of some members of the Dps family constitute another protective mechanism. No apparent sequence specificity has been attributed to them. By coating the DNA, Dps provides a physical shield from damaging agents, and prevents mutagenesis (Nair and Finkel 2004; Park et al. 2005). In addition, the reduction of the nucleoid surface by Dps-dependent-compaction was recently suggested as an explanation for the reduced challenge of reactive oxygen species (Morikawa et al. 2006; Ceci et al. 2007). By modulating DNA structure, Keywords Dps, horizontal transfer, malolactic fermentation, Oenococcus oeni, stress. Correspondence Claire Le He ´ naff-Le Marrec, UMR Œnologie, Universite ´ Victor Segalen Bordeaux 2 – Universite ´ Bordeaux 1 (ISTAB)- INRA, 351, Cours de la Libe ´ ration, 33405 Talence Cedex, France. E-mail: [email protected] 2007 ⁄ 1692: received 22 October 2007, revised 8 April 2008 and accepted 8 May 2008 doi:10.1111/j.1365-2672.2008.03967.x Abstract Aims: To identify novel actors responsible for the marked adaptation of the Oenococcus oeni species to its environment. Methods and Results: Genomic surveillance of the available genome sequences from O. oeni indicated the presence of a small ORF, encoding a protein named Dps A . The cloned gene complemented the dps ) mutant of Escherichia coli and conferred resistance to hydrogen peroxide, wine, and metals. The dps A gene was flanked by IS-related elements. The entire region was characterized by an anomalously high GC content compared to those reported for oenococcal genomes. The dps A gene was present in 15 of the 38 tested isolates. Positive strains originated from different geographical areas and sources. No change in tolerance to wine or to oxidative stress was observed between O. oeni strains harbouring dps A and those not harbouring this gene. Conclusions: Some O. oeni have acquired a functional homologue to the dps gene from E. coli as part of a mobile element. Significance and Impact of the Study: Dps A probably increases the bacterial fitness in response to environmental challenges. However, the physiological condition under which it adds a selective advantage to O. oeni during wine- making remains to be found. Journal of Applied Microbiology ISSN 1364-5072 1866 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 ª 2008 The Authors
-
Upload
enseirb-matmeca -
Category
Documents
-
view
2 -
download
0
Transcript of Characterization of an acquired dps -containing gene island in the lactic acid bacterium Oenococcus...

ORIGINAL ARTICLE
Characterization of an acquired dps-containing gene islandin the lactic acid bacterium Oenococcus oeniA. Athane1, E. Bilhere1, E. Bon1,2, G. Morel1, P. Lucas1, A. Lonvaud1 and C. Le Marrec1
1 UMR 1219, INRA-Universite Victor Segalen Bordeaux 2 – Universite Bordeaux 1, Talence Cedex, France
2 Universite Victor Segalen Bordeaux 2 – LaBRI (Universite de Bordeaux – CNRS) 351, Talence Cedex, France
Introduction
Bacteria have evolved a complex network of stress response
pathways to promote their growth and ⁄ or survival during
environmental challenges. One important mechanism is
the DNA binding protein from starved cells (Dps), first
described in Escherichia coli, where its expression was
found to be activated when the bacterium finds itself in
nutritionally limiting conditions or under oxidative stress,
regardless of whether cells are actively growing or not
(Almiron et al. 1992). Recent evidence expands the role of
Dps in tolerance to acid and osmotic stresses (Choi et al.
2000; Weber et al. 2006), metals, UV-irradiation (Nair and
Finkel 2004), and high pressure (Malone et al. 2006).
The Dps protein from E. coli has a molecular mass of
19 kDa, and is known to be a member of the Fe-binding
protein family. This family forms large (approx.
150 kDa), hexameric complexes in cells that reduce the
intracellular level of iron (Fe2+), thereby preventing the
deleterious effect of the superoxide anion through the
Fenton reaction (Almiron et al. 1992). This combined
action of hydrogen peroxide and intracellular iron is of
paramount importance in vivo since aerobic growth
generates sufficient H2O2 to create toxic levels of DNA
damage in E. coli (Park et al. 2005). Other Dps proteins
such as the cyanobacterial Synechococcus DpsA proteins
also display catalase activity, and can therefore degrade
hydrogen peroxyde (Pena and Bullerjahn 1995). The
DNA binding properties of some members of the Dps
family constitute another protective mechanism. No
apparent sequence specificity has been attributed to them.
By coating the DNA, Dps provides a physical shield from
damaging agents, and prevents mutagenesis (Nair and
Finkel 2004; Park et al. 2005). In addition, the reduction
of the nucleoid surface by Dps-dependent-compaction
was recently suggested as an explanation for the reduced
challenge of reactive oxygen species (Morikawa et al.
2006; Ceci et al. 2007). By modulating DNA structure,
Keywords
Dps, horizontal transfer, malolactic
fermentation, Oenococcus oeni, stress.
Correspondence
Claire Le Henaff-Le Marrec, UMR Œnologie,
Universite Victor Segalen Bordeaux 2 –
Universite Bordeaux 1 (ISTAB)- INRA, 351,
Cours de la Liberation, 33405 Talence Cedex,
France. E-mail: [email protected]
2007 ⁄ 1692: received 22 October 2007,
revised 8 April 2008 and accepted 8 May
2008
doi:10.1111/j.1365-2672.2008.03967.x
Abstract
Aims: To identify novel actors responsible for the marked adaptation of the
Oenococcus oeni species to its environment.
Methods and Results: Genomic surveillance of the available genome sequences
from O. oeni indicated the presence of a small ORF, encoding a protein named
DpsA. The cloned gene complemented the dps) mutant of Escherichia coli and
conferred resistance to hydrogen peroxide, wine, and metals. The dpsA gene
was flanked by IS-related elements. The entire region was characterized by an
anomalously high GC content compared to those reported for oenococcal
genomes. The dpsA gene was present in 15 of the 38 tested isolates. Positive
strains originated from different geographical areas and sources. No change in
tolerance to wine or to oxidative stress was observed between O. oeni strains
harbouring dpsA and those not harbouring this gene.
Conclusions: Some O. oeni have acquired a functional homologue to the dps
gene from E. coli as part of a mobile element.
Significance and Impact of the Study: DpsA probably increases the bacterial
fitness in response to environmental challenges. However, the physiological
condition under which it adds a selective advantage to O. oeni during wine-
making remains to be found.
Journal of Applied Microbiology ISSN 1364-5072
1866 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875
ª 2008 The Authors

Dps may also regulate gene expression (Nair and Finkel
2004).
Dps and homologous proteins have been identified in
distantly-related bacteria facing different challenges in their
environment, suggesting that this protein plays an essential
role in bacterial vitality. This is highlighted by the fact that
Dps is considered one of the protective mechanisms dis-
played by bacterial pathogens against oxidative damage
during phagocytosis (Hasley et al. 2004; Olsen et al. 2005).
Dps has been shown to play a role in bacteriophage toler-
ance (Lacqua et al. 2006) and has recently been proposed
as a candidate protein for adaptation to epiphytic life in
Methylobacterium extorquens (Gourion et al. 2006). Consis-
tent with these pivotal roles, protein levels are thought to
be tightly regulated, with regulatory modes that may differ
according to the bacteria. This is illustrated by the observa-
tion that various transcriptional factors (OxyR ⁄ IHF, Per)
are responsible for dps gene expression in Proteobacteria
and Firmicutes (Morikawa et al. 2006). Dps also undergoes
protease-dependent regulation involving ClpXP and ClpAP
in E. coli (Stephani et al. 2003; Weichart et al. 2003), and
post-translational modifications including phosphorylation
in wall-less E. coli cells (Freestone et al. 1998) and glycosyl-
ation in Salmonella enterica (Hanna et al. 2007). The signifi-
cance of such modifications in vivo remains unknown.
Oenococcus oeni is a lactic acid bacterium responsible for
the malolactic fermentation of wines (MLF). This bacte-
rium is able to cope with a hostile environment including
acid, ethanol, toxic phenolic compounds and nutrient lim-
itation. Activation of membrane-bound H+-ATPases,
modification of membrane fluidity, stress protein synthesis
and exclusion of different cytoplasmic stressors through
multi-drug resistance proteins have been described as
essential mechanisms helping the strain to cope with the
harsh environment (See for review Spano and Massa
2006). The genomes of two strains of O. oeni have been
recently completed (Mills et al. 2005; GenBank accession
number AAUV01000000) and a dps-like gene was identi-
fied in both strains. We present evidence that the encoded
DpsA protein from O. oeni is a functional homologue of
dps from E. coli protecting the cell from H2O2, and suggest
its involvement in the defence of the lactic acid bacterium
against wine and metal stresses. We also posit the role of
dpsA during plasmid establishment. We further show that
dpsA is a mobile gene acquired as part of a mobile element.
Materials and methods
Bacterial strains, plasmids and growth conditions
The 38 O. oeni isolates used in this study are presented in
Table 1. Species identification was carried out by PCR,
using primers OO1 and OO2 targeting the gene encoding
the malolactic enzyme which are specific to O. oeni (Divol
et al. 2003). All strains were cultured in MRS broth pH 4Æ8.
The collection includes 27 wild strains, isolated from vari-
ous types of wines undergoing malolactic fermentation,
each isolate representing a specific vineyard of France
(Delaherche et al. 2006). These strains have been characte-
rized regarding their tolerance to wine, corresponding to
the ability to induce malolactic fermentation after inocula-
tion to wine (oenological potential). Briefly, precultures
grown in modified acidic grape medium were used to ino-
culate three different wines. Microvinifications were done
in duplicate using independent precultures. Assays were
conducted at 20�C and monitored for a three-week period
for their residual malic acid content. Consumption of malic
acid was checked weekly using a thin layer chromatography
method (Roche, Meylan, France). The strains which com-
pleted MLF in the three tested wines (n = 8) were further
considered as isolates of high tolerance (Table 1). Isolates
which did not perform MLF in any assay formed the low
performing group (n = 19). The group of highly perform-
ing strains was enriched with 11 commercial starters (see
Table 1). To avoid redundancy in our collection, the 38 iso-
lates were analysed by pulsed-field gel electrophoresis
(PFGE) using NotI, as previously described (Gindreau et al.
1997). All isolates exhibited distinct PFGE banding pattern
types.
Plasmid pBBR1 Kan carries a replicon from Bordetella
bronchiseptica, and replicates at a low-copy number in
E. coli (Kovach et al. 1995). It was used with the XL1-
Blue strain of E. coli for cloning purposes. The E. coli
dps) ZK1058 (ZK126 dps::kan) and SF2043 (ZK1058
pBR322), and the E. coli dps+ SF2042 (ZK1058 pJE106)
were kindly provided by Dr S. Finkel (Nair and Finkel
2004). They were grown in LB medium and cultures were
incubated at 37�C in rotary shakers for 16 h. When nec-
essary, the following antibiotic concentrations were used:
ampicillin 150 lg ml)1, kanamycin 50 lg ml)1.
Extraction of genomic DNA and subtractive
hybridization
Genomic DNA from O. oeni was isolated as previously
described (Le Marrec et al. 2007). Subtractive hybridiza-
tion was conducted using an adapted protocol of the PCR-
Select Bacterial Genome Subtraction Kit (Clontech), with
DNA from strain IOEB-SARCO 444 as the driver, and an
equimolar mixture of DNAs from strains IOEB-SARCO
277, IOEB-SARCO 384, IOEB-SARCO 450 and VP41 as
the tester. All DNAs were digested with RsaI. Restricted
tester DNA was marked by ligation with oligonucleotide
adapters and hybridized at 63�C with restricted driver
DNA in excess. Self-hybrid tester fragments were amplified
by PCR and nested PCR, using adapter-specific primers.
A. Athane et al. Acquired dps-containing gene island in lactic acid bacterium O. oeni
ª 2008 The Authors
Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 1867
Amplicons were cloned in the pGEM-T Easy Vector (Pro-
mega) and transformed into competent DH5 a cells. Posi-
tive clones were screened on LB medium supplemented
with ampicillin, X-Gal and IPTG. Inserted fragments were
sequenced. The specificity of the selected sequences was
assessed through a PCR assay in driver and tester DNAs.
Analysis of the distribution of dpsA by PCR
Primer design was achieved using the eprimer3 and Oligo
Analyser 1Æ0Æ3 softwares, using data from the genomes of
O. oeni PSU-1 and ATCC BAA-1163. Oligonucleotides
were purchased from Sigma-Aldrich. They are listed in
Table 2. PCR amplifications were carried out using the
primers AX1 and AX2, which are internal to the dpsA
gene from O. oeni PSU-1 (OEOE_1750) and ATCC BAA-
1163 (OENOO_64063). Genetic organization of the dpsA
region was first assessed using the primers AX3 and AX4,
whose sequences are part of the flanking OEOE_1749 and
OEOE_1751 genes, respectively, in O. oeni PSU-1. These
primers are 100% identical to the corresponding regions
in strain ATCC BAA-1163. The following additional sets
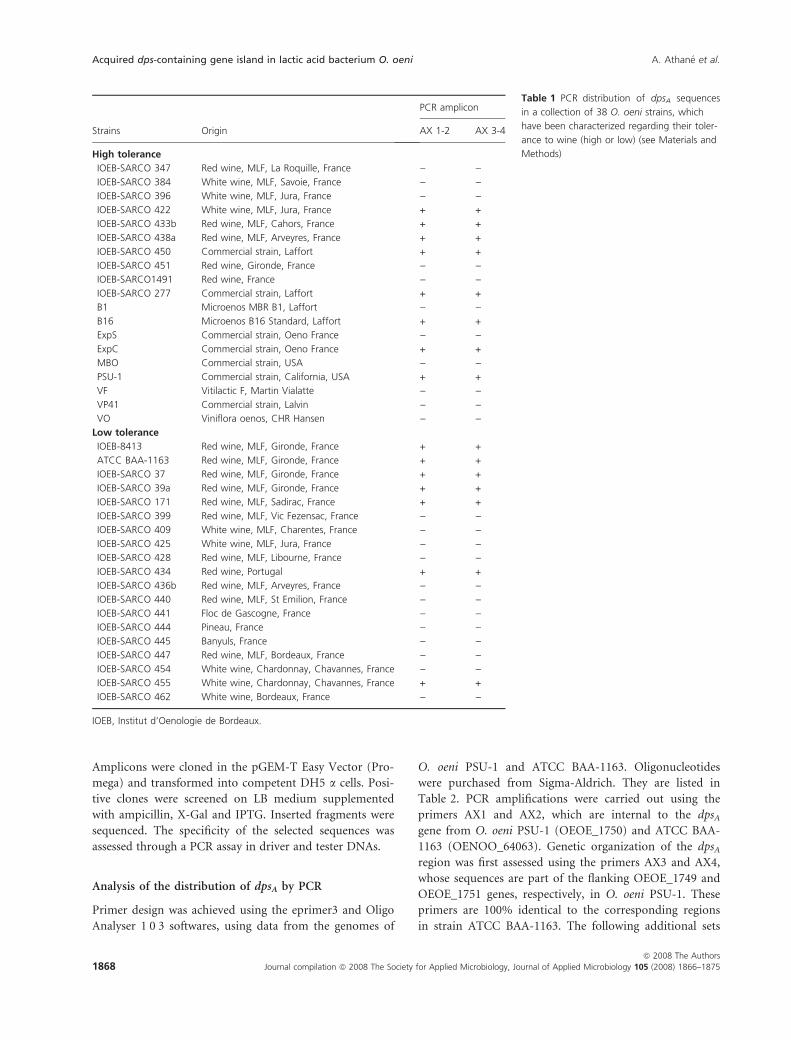
Table 1 PCR distribution of dpsA sequences
in a collection of 38 O. oeni strains, which
have been characterized regarding their toler-
ance to wine (high or low) (see Materials and
Methods)
Strains Origin
PCR amplicon
AX 1-2 AX 3-4
High tolerance
IOEB-SARCO 347 Red wine, MLF, La Roquille, France ) )IOEB-SARCO 384 White wine, MLF, Savoie, France ) )IOEB-SARCO 396 White wine, MLF, Jura, France ) )IOEB-SARCO 422 White wine, MLF, Jura, France + +
IOEB-SARCO 433b Red wine, MLF, Cahors, France + +
IOEB-SARCO 438a Red wine, MLF, Arveyres, France + +
IOEB-SARCO 450 Commercial strain, Laffort + +
IOEB-SARCO 451 Red wine, Gironde, France ) )IOEB-SARCO1491 Red wine, France ) )IOEB-SARCO 277 Commercial strain, Laffort + +
B1 Microenos MBR B1, Laffort ) )B16 Microenos B16 Standard, Laffort + +
ExpS Commercial strain, Oeno France ) )ExpC Commercial strain, Oeno France + +
MBO Commercial strain, USA ) )PSU-1 Commercial strain, California, USA + +
VF Vitilactic F, Martin Vialatte ) )VP41 Commercial strain, Lalvin ) )VO Viniflora oenos, CHR Hansen ) )
Low tolerance
IOEB-8413 Red wine, MLF, Gironde, France + +
ATCC BAA-1163 Red wine, MLF, Gironde, France + +
IOEB-SARCO 37 Red wine, MLF, Gironde, France + +
IOEB-SARCO 39a Red wine, MLF, Gironde, France + +
IOEB-SARCO 171 Red wine, MLF, Sadirac, France + +
IOEB-SARCO 399 Red wine, MLF, Vic Fezensac, France ) )IOEB-SARCO 409 White wine, MLF, Charentes, France ) )IOEB-SARCO 425 White wine, MLF, Jura, France ) )IOEB-SARCO 428 Red wine, MLF, Libourne, France ) )IOEB-SARCO 434 Red wine, Portugal + +
IOEB-SARCO 436b Red wine, MLF, Arveyres, France ) )IOEB-SARCO 440 Red wine, MLF, St Emilion, France ) )IOEB-SARCO 441 Floc de Gascogne, France ) )IOEB-SARCO 444 Pineau, France ) )IOEB-SARCO 445 Banyuls, France ) )IOEB-SARCO 447 Red wine, MLF, Bordeaux, France ) )IOEB-SARCO 454 White wine, Chardonnay, Chavannes, France ) )IOEB-SARCO 455 White wine, Chardonnay, Chavannes, France + +
IOEB-SARCO 462 White wine, Bordeaux, France ) )
IOEB, Institut d’Oenologie de Bordeaux.
Acquired dps-containing gene island in lactic acid bacterium O. oeni A. Athane et al.
1868 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875
ª 2008 The Authors

were also used: AX5-AX6, AX5-AX8, AX7-AX8 and AX7-
AX9. PCR with these primers amplified DNA fragments
of 6031-bp, 3600-bp, 8124-bp and 12000-bp, respectively
from O. oeni PSU-1 (Table 2 and Fig. 3). Amplifications
were carried out in a 20 ll reaction volume comprising
1 X iProof PCR Master Mix (Bio-Rad), 1 lmol l)1 for-
ward primer, 1 lmol l)1 reverse primer and 50 ng DNA
template using a iCycler thermal cycler (Bio-Rad).
Cycling parameters comprised an initial denaturation at
95�C for 3 min, 35 cycles of denaturation at 95�C for
30 s, annealing at x�C for 30 s and extension at 72�C for
y min (where annealing temperature x and extension time
y depend on the Tm of the primers and the expected
amplicon size, respectively), and a final extension of 72�C
for 10 min.
Cloning of dpsA and transformation in the E. coli dps)
mutant strain
The dpsA gene was PCR-amplified from the chromosome
of strain O. oeni PSU-1 using the primer pair AX3 and
AX4. The amplicon was cloned using the TOPO-XL
vector in TOP10 cells (Invitrogen). Three clones were
randomly selected and their plasmid DNA was extracted
using the Qiagen kit. The same sequence was obtained for
all three recombinant plasmids, and pAX1 was retained
for investigation. The 1-kb insert was subcloned as a
BamHI–XbaI fragment in the low copy AmpR vector
pBBR1-MCS4, yielding pAX19, which was introduced in
E. coli XL1-Blue using the classical transformation
method with calcium.
Plasmid DNAs from pAX19 and pBBR1-MCS4 were
also introduced in E. coli ZK1058 as described above. To
compare their transformation efficiencies in the dps)
mutant strain, pure plasmid DNAs were first isolated
from E. coli XL1-Blue and photometrically quantified.
Samples showing A260 ⁄ 280 ratio between 1Æ9 and 2 were
used for further investigations. The same quantities
(0Æ1 lg of DNA in 1–2 ll) were introduced in E. coli
XL1-blue and E. coli ZK1058. The ratio between the
transformation efficiencies (number of transformants per
lg DNA) of pAX19 and pBBR1-MCS4 were calculated in
each strain and compared. All assays were performed in
triplicate.
E. coli dps::kan mutant complementation assay and stress
tolerance assay in O. oeni
The parameters of the lethal shocks (duration and inten-
sity of the stressor) were defined using the two E. coli
ZK1058 derivatives containing pBR322 and pJE106 (dps
from E. coli). We selected stress conditions yielding differ-
ences in survival rates greater than 1 log unit after 15 to
30 min in the presence of the stressor. Sensitivity of the
strains containing pBBR1-MCS4 and pAX19 (dpsA from
O. oeni) were then assessed. Oxidative-stress assays were
conducted as follows. First, growth with a suboptimal
H2O2 concentration was tested. After overnight growth,
samples were inoculated in LB with or without H2O2
(4Æ85 mmol l)1) to an initial OD600 of 0Æ1, and were incu-
bated at 37�C for 6 h. Absorbance was recorded every
30 min. Survival to lethal oxidative stress was also
assessed. For this purpose, overnight LB cultures were
diluted 1 : 50 into 10 ml of fresh medium and incubated
at 37�C with shaking for 3h. The stationary growing
E. coli cells were exposed to a concentration of
40 mmol l)1 in H2O2 (obtained from Carlo Erba as a 9Æ7mol l)1 stock solution) in LB. After addition of the stres-
sor (time zero in the experiment), viable-cell counts were
periodically determined. Samples were recovered, serially
diluted in LB medium and immediately plated on LB
plates to determine the number of CFU and to calculate
survivors.
To assess sensitivity to wine, log phase cells grown in
LB broth were diluted to 1 ⁄ 10 in LB diluted with 50%
wine (Saint-Emilion, France). Stationary-phase cultures
were stressed similarly with pure wine. Metal stress assays
were conducted on stationary phase cells in LB containing
FeSO4 100 mmol l)1 or CuSO4 10 mmol l)1 as previously
reported (Nair and Finkel 2004). All assays were
performed in triplicate.
Survival of log phase cells of O. oeni to a lethal oxidative
stress was assessed in MRS containing H2O2 40 mmol l)1.
Survival was determined after 60 min as described above,
except that diluted samples were plates on MRS plates.
Results
DpsA from O. oeni is a functional homologue of Dps
from E. coli
Blast searches using the deduced amino acid sequence of
the Dps protein from E. coli revealed a significant identity
Table 2 Primers used for PCR amplification
Primer Sequence Location
AX1 5¢-TGCCTAGATAAGCACTGATC-3 ¢ OEOE_1750 (dpsA)
AX2 5¢-ATCAACTGATTGCCGATATC-3¢ OEOE_1750 (dpsA)
AX3 5¢-CGCCAGGTTCAAAATGTCTT-3¢ OEOE_1749 (IS1480)
AX4 5¢-TCAATTCGTATTCCCGAAGC-3¢ OEOE_1751 (IS1165)
AX5 5¢-ATCAATCCGGTCACGATAGTTC-3¢ OEOE_1748
AX6 5¢-TCTGGATACAGAAAGGGATCGT-3¢ OEOE_1755
AX7 5¢-CTGATAGTCTTCCGGATGATGC-3¢ OEOE_1745
AX8 5¢-CAGGCAGAGCTTACCTACACACT-3¢ OEOE_1753
AX9 5¢-GGTAGTATCAAGGACCGACTGG-3¢ OEOE_1759
A. Athane et al. Acquired dps-containing gene island in lactic acid bacterium O. oeni
ª 2008 The Authors
Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 1869
(29% identity, 48% similarity) to the OEOE_1750 and
OENOO_64063 gene products from O. oeni PSU-1 and
ATCC BAA-1163, respectively. Both products corre-
sponded to a putative polypeptide of 160 amino acids
(18Æ7 kDa, pI 4Æ56) henceforth named DpsA. This oeno-
coccal sequence displayed best BLAST matches (49% to
67% identity) with dps-like gene products from the lactic
acid bacteria Pediococcus pentosaceus, Lactobacillus fermen-
tum, Lactobacillus plantarum, Enterococcus faecalis, Lacto-
bacillus reuteri and Streptococcus thermophilus. Analysis
also revealed 46% identity to the Listeria monocytogenes
fri gene product, while a more remote homology was
found with MrgA from Bacillus subtilis (59% similarity,
33% identity).
Oenococcus oeni is not amenable to genetic transforma-
tions enabling dpsA gene inactivation. Therefore, to assess
the role of dpsA in-vivo, the gene was amplified from the
chromosome of O. oeni PSU-1, and cloned in the pBBR1-
MCS4 vector. The resulting plasmid was introduced into
the dps::kan mutant (ZK1058) of E. coli (Nair and Finkel
2004). The strain was also transformed with pBBR1-
MCS4 DNA to verify that the vector alone had no effect
on the growth ⁄ survival of the dps::kan mutant strain.
Surprisingly, the two plasmid derivatives (one harbouring
the dpsA gene and one that did not) differed in their
transformation efficiency of the E. coli ZK1058 strain. The
efficiency of the dpsA-containing plasmid was 750 (±50)-
fold higher than that observed for the empty vector,
although both values observed in E. coli XL1-Blue were
similar, yielding a ratio of 1Æ16 ± 0Æ2.
Given the role of Dps-proteins in resistance to oxida-
tive stress, strains ZK1058 (whether harbouring the dpsA
gene or not) were tested for their sensitivity to hydrogen
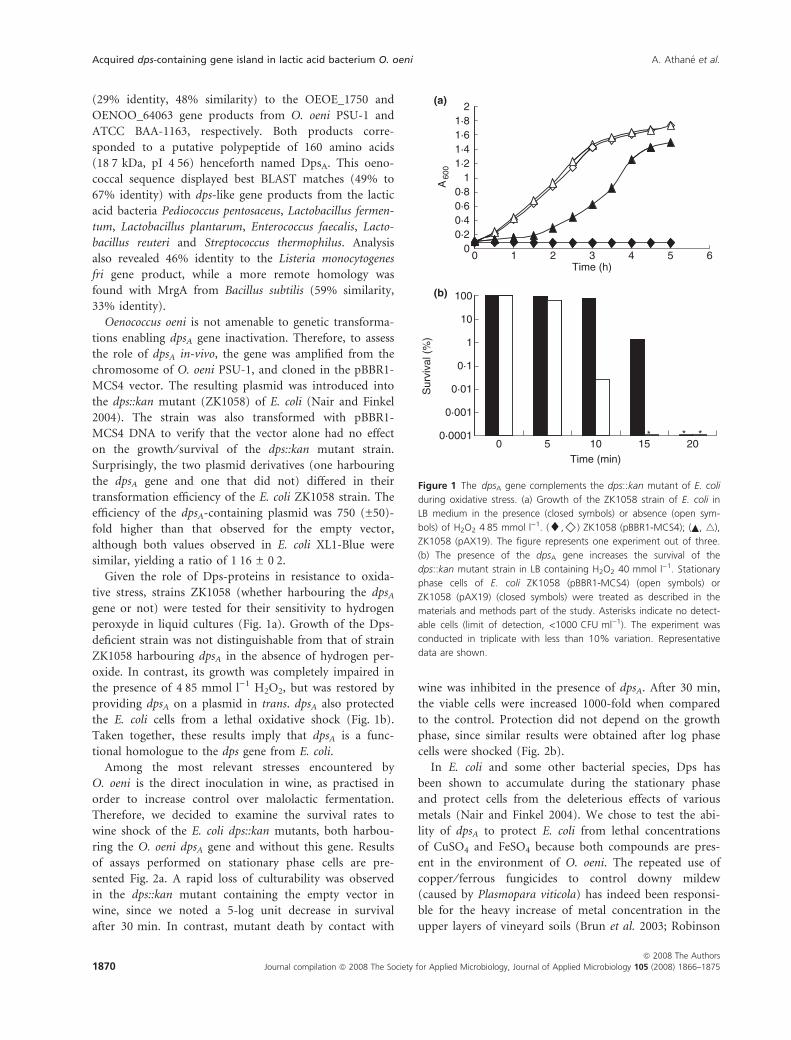
peroxyde in liquid cultures (Fig. 1a). Growth of the Dps-
deficient strain was not distinguishable from that of strain
ZK1058 harbouring dpsA in the absence of hydrogen per-
oxide. In contrast, its growth was completely impaired in
the presence of 4Æ85 mmol l)1 H2O2, but was restored by
providing dpsA on a plasmid in trans. dpsA also protected
the E. coli cells from a lethal oxidative shock (Fig. 1b).
Taken together, these results imply that dpsA is a func-
tional homologue to the dps gene from E. coli.
Among the most relevant stresses encountered by
O. oeni is the direct inoculation in wine, as practised in
order to increase control over malolactic fermentation.
Therefore, we decided to examine the survival rates to
wine shock of the E. coli dps::kan mutants, both harbou-
ring the O. oeni dpsA gene and without this gene. Results
of assays performed on stationary phase cells are pre-
sented Fig. 2a. A rapid loss of culturability was observed
in the dps::kan mutant containing the empty vector in
wine, since we noted a 5-log unit decrease in survival
after 30 min. In contrast, mutant death by contact with
wine was inhibited in the presence of dpsA. After 30 min,
the viable cells were increased 1000-fold when compared
to the control. Protection did not depend on the growth
phase, since similar results were obtained after log phase
cells were shocked (Fig. 2b).
In E. coli and some other bacterial species, Dps has
been shown to accumulate during the stationary phase
and protect cells from the deleterious effects of various
metals (Nair and Finkel 2004). We chose to test the abi-
lity of dpsA to protect E. coli from lethal concentrations
of CuSO4 and FeSO4 because both compounds are pres-
ent in the environment of O. oeni. The repeated use of
copper ⁄ ferrous fungicides to control downy mildew
(caused by Plasmopara viticola) has indeed been responsi-
ble for the heavy increase of metal concentration in the
upper layers of vineyard soils (Brun et al. 2003; Robinson
10
100(b)
(a)
1·61·8
2
0·001
0·01
0·1
1
Sur
viva
l (%
)
0·20·40·60·8
11·21·4
0·000120151050
06 5 4 3
Time (h)
Time (min)
2 1 0
* * *A
600
Figure 1 The dpsA gene complements the dps::kan mutant of E. coli
during oxidative stress. (a) Growth of the ZK1058 strain of E. coli in
LB medium in the presence (closed symbols) or absence (open sym-
bols) of H2O2 4Æ85 mmol l)1. (¤,)) ZK1058 (pBBR1-MCS4); ( , 4),
ZK1058 (pAX19). The figure represents one experiment out of three.
(b) The presence of the dpsA gene increases the survival of the
dps::kan mutant strain in LB containing H2O2 40 mmol l)1. Stationary
phase cells of E. coli ZK1058 (pBBR1-MCS4) (open symbols) or
ZK1058 (pAX19) (closed symbols) were treated as described in the
materials and methods part of the study. Asterisks indicate no detect-
able cells (limit of detection, <1000 CFU ml)1). The experiment was
conducted in triplicate with less than 10% variation. Representative
data are shown.
Acquired dps-containing gene island in lactic acid bacterium O. oeni A. Athane et al.
1870 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875
ª 2008 The Authors
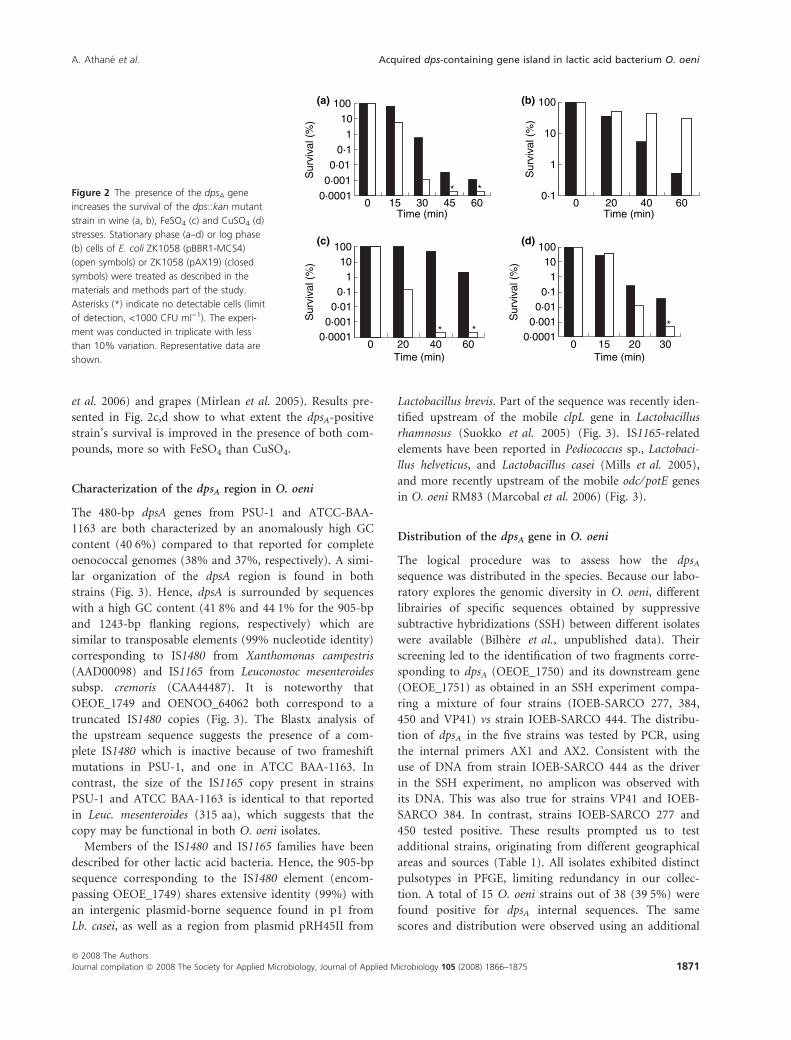
et al. 2006) and grapes (Mirlean et al. 2005). Results pre-
sented in Fig. 2c,d show to what extent the dpsA-positive
strain’s survival is improved in the presence of both com-
pounds, more so with FeSO4 than CuSO4.
Characterization of the dpsA region in O. oeni
The 480-bp dpsA genes from PSU-1 and ATCC-BAA-
1163 are both characterized by an anomalously high GC
content (40Æ6%) compared to that reported for complete
oenococcal genomes (38% and 37%, respectively). A simi-
lar organization of the dpsA region is found in both
strains (Fig. 3). Hence, dpsA is surrounded by sequences
with a high GC content (41Æ8% and 44Æ1% for the 905-bp
and 1243-bp flanking regions, respectively) which are
similar to transposable elements (99% nucleotide identity)
corresponding to IS1480 from Xanthomonas campestris
(AAD00098) and IS1165 from Leuconostoc mesenteroides
subsp. cremoris (CAA44487). It is noteworthy that
OEOE_1749 and OENOO_64062 both correspond to a
truncated IS1480 copies (Fig. 3). The Blastx analysis of
the upstream sequence suggests the presence of a com-
plete IS1480 which is inactive because of two frameshift
mutations in PSU-1, and one in ATCC BAA-1163. In
contrast, the size of the IS1165 copy present in strains
PSU-1 and ATCC BAA-1163 is identical to that reported
in Leuc. mesenteroides (315 aa), which suggests that the
copy may be functional in both O. oeni isolates.
Members of the IS1480 and IS1165 families have been
described for other lactic acid bacteria. Hence, the 905-bp
sequence corresponding to the IS1480 element (encom-
passing OEOE_1749) shares extensive identity (99%) with
an intergenic plasmid-borne sequence found in p1 from
Lb. casei, as well as a region from plasmid pRH45II from
Lactobacillus brevis. Part of the sequence was recently iden-
tified upstream of the mobile clpL gene in Lactobacillus
rhamnosus (Suokko et al. 2005) (Fig. 3). IS1165-related
elements have been reported in Pediococcus sp., Lactobaci-
llus helveticus, and Lactobacillus casei (Mills et al. 2005),
and more recently upstream of the mobile odc ⁄ potE genes
in O. oeni RM83 (Marcobal et al. 2006) (Fig. 3).
Distribution of the dpsA gene in O. oeni
The logical procedure was to assess how the dpsA
sequence was distributed in the species. Because our labo-
ratory explores the genomic diversity in O. oeni, different
librairies of specific sequences obtained by suppressive
subtractive hybridizations (SSH) between different isolates
were available (Bilhere et al., unpublished data). Their
screening led to the identification of two fragments corre-
sponding to dpsA (OEOE_1750) and its downstream gene
(OEOE_1751) as obtained in an SSH experiment compa-
ring a mixture of four strains (IOEB-SARCO 277, 384,
450 and VP41) vs strain IOEB-SARCO 444. The distribu-
tion of dpsA in the five strains was tested by PCR, using
the internal primers AX1 and AX2. Consistent with the
use of DNA from strain IOEB-SARCO 444 as the driver
in the SSH experiment, no amplicon was observed with
its DNA. This was also true for strains VP41 and IOEB-
SARCO 384. In contrast, strains IOEB-SARCO 277 and
450 tested positive. These results prompted us to test
additional strains, originating from different geographical
areas and sources (Table 1). All isolates exhibited distinct
pulsotypes in PFGE, limiting redundancy in our collec-
tion. A total of 15 O. oeni strains out of 38 (39Æ5%) were
found positive for dpsA internal sequences. The same
scores and distribution were observed using an additional
10
100 100
0·0001
0·001
0·01
0·1
1
0 15 30 45 60
Sur
viva
l (%
)S
urvi
val (
%)
Sur
viva
l (%
)
Sur
viva
l (%
)
* *0·1
1
10
0 20 40 60Time (min)
110
100(c) (d)
(a) (b)
Time (min)
110
100
0·00010·001
0·010·1
0 20 40 60Time (min)
* **
0·00010·001
0·010·1
0 15 20 30Time (min)
*
Figure 2 The presence of the dpsA gene
increases the survival of the dps::kan mutant
strain in wine (a, b), FeSO4 (c) and CuSO4 (d)
stresses. Stationary phase (a–d) or log phase
(b) cells of E. coli ZK1058 (pBBR1-MCS4)
(open symbols) or ZK1058 (pAX19) (closed
symbols) were treated as described in the
materials and methods part of the study.
Asterisks (*) indicate no detectable cells (limit
of detection, <1000 CFU ml)1). The experi-
ment was conducted in triplicate with less
than 10% variation. Representative data are
shown.
A. Athane et al. Acquired dps-containing gene island in lactic acid bacterium O. oeni
ª 2008 The Authors
Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 1871

primer pair (AX3 and AX4) designed in the upstream
and downstream ORFs in O. oeni PSU-1 and ATCC
BAA-1163 (Fig. 3). These results support the ideas (i) that
dpsA flanking sequences are conserved among strains
positive for this trait and (ii) that dpsA is part of a mobile
region encompassing at least OEOE_1749, OEOE_1750
and OEOE_1751. We next tried to assess the outer limits
of the dpsA containing gene island, using primers at
upstream and downstream of OEOE_1749 and
OEOE_1752 (see Fig. 3). Amplicons with the expected
sizes were obtained with all dpsA+ strains. In contrast,
dpsA) isolates yielded no products. These results suggest
that the dpsA variable region is larger than 12-kb.
The distribution of dpsA sequences in the collection
indicated no particular association with specific geograph-
ical areas, nor the type of samples used for bacterial isola-
tion. Lastly, we evaluated the possibility of an association
with enhanced oenological potential. Our collection con-
tained 19 strains previously known as highly tolerant to
wine (HP), including commercial starters, and 19 isolates
which failed at inducing malolactic fermentation after
direct inoculation in wines (LP) (Table 1). An equal dis-
tribution of the dpsA sequences in the HP and LP groups
was observed (8 and 7 strains were dpsA+, respectively).
This is suggesting that dpsA is not critical for survival in
wine. We could draw a similar conclusion regarding the
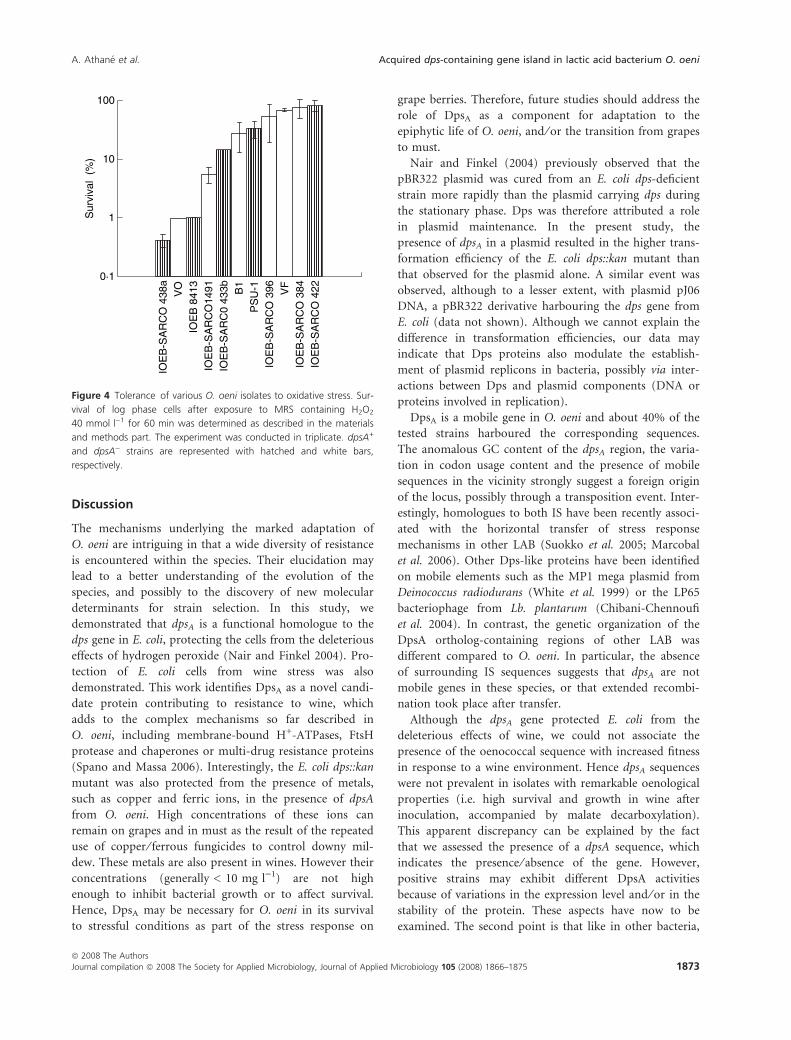
tolerance to oxidative conditions. Survival to H2O2 was
assessed for 11 O. oeni strains, five dpsA+ and six dpsA)
(Fig. 4). dpsA positive strains were not significantly more
resistant to an oxidative environment.
AX5AX3 AX1/2 AX4
AX6AX7 AX8 AX9
OE
OE
_174
8
OE
OE
_175
0
OE
OE
_174
9
OE
OE
_175
1
OE
OE
_175
2*
OE
OE
_175
3
OE
OE
_175
4
OE
OE
_175
5
OE
OE
_175
6
OE
OE
_175
7
OE
OE
_175
8
OE
OE
_175
9
OE
OE
_174
7
OE
OE
_174
6
OE
OE
_174
5
OE
OE
_174
4LPS synthesisglycosyltransferase
MethioninetRNAsynthase
Permease IS1165 mrr Cysteinesynthase
HP IS1480
dpsA Methyltransferase
Hydrolase Phosphoglyceratemutase
Cystathioninebeta-lyase
HP Pseudogene Serineacetyltransferase
+3
O. oeni ATCCBAA-1163
O. oeni PSU-1+2
+1
+1+3
O. oeni RM83
3128 4497 8866 11103 OE
NO
O_6
4063
OdcIS1165
Plasmid p1(Lb. casei)
Plasmid pRH45II(Lb. brevis)
IS1480
14335 15192
16703 17557 18466 19926ABC
transporterHP
IS1480
Lb. rhamnosusClpL2
167 462 839 2953
IS1480
Figure 3 Schematic overview of the dpsA chromosomal region in O. oeni PSU-1 and ATCC-BAA 1163. The dpsA gene is represented as a
hatched arrow, and flanking IS as grey arrows. Frameshifts in the IS1480 and IS1165 sequences are indicated. The sequence of the leftward
IS-related transposase in strain PSU-1 (OEOE_1752) has been incorrectly predicted, due to an error when locating the translation start site. The
correction of this error is indicated with an asterisk (OEOE_1752*). The positions of the nine primers are also indicated. Gene sequence conserva-
tion between PSU-1 genome and plasmid or cloned chromosomal regions from other lactic acid bacteria is represented by shaded areas. The
nucleotide position of the homologous regions in these sequences is also indicated.
Acquired dps-containing gene island in lactic acid bacterium O. oeni A. Athane et al.
1872 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875
ª 2008 The Authors
Discussion
The mechanisms underlying the marked adaptation of
O. oeni are intriguing in that a wide diversity of resistance
is encountered within the species. Their elucidation may
lead to a better understanding of the evolution of the
species, and possibly to the discovery of new molecular
determinants for strain selection. In this study, we
demonstrated that dpsA is a functional homologue to the
dps gene in E. coli, protecting the cells from the deleterious
effects of hydrogen peroxide (Nair and Finkel 2004). Pro-
tection of E. coli cells from wine stress was also
demonstrated. This work identifies DpsA as a novel candi-
date protein contributing to resistance to wine, which
adds to the complex mechanisms so far described in
O. oeni, including membrane-bound H+-ATPases, FtsH
protease and chaperones or multi-drug resistance proteins
(Spano and Massa 2006). Interestingly, the E. coli dps::kan
mutant was also protected from the presence of metals,
such as copper and ferric ions, in the presence of dpsA
from O. oeni. High concentrations of these ions can
remain on grapes and in must as the result of the repeated
use of copper ⁄ ferrous fungicides to control downy mil-
dew. These metals are also present in wines. However their
concentrations (generally < 10 mg l)1) are not high
enough to inhibit bacterial growth or to affect survival.
Hence, DpsA may be necessary for O. oeni in its survival
to stressful conditions as part of the stress response on
grape berries. Therefore, future studies should address the
role of DpsA as a component for adaptation to the
epiphytic life of O. oeni, and ⁄ or the transition from grapes
to must.
Nair and Finkel (2004) previously observed that the
pBR322 plasmid was cured from an E. coli dps-deficient
strain more rapidly than the plasmid carrying dps during
the stationary phase. Dps was therefore attributed a role
in plasmid maintenance. In the present study, the
presence of dpsA in a plasmid resulted in the higher trans-
formation efficiency of the E. coli dps::kan mutant than
that observed for the plasmid alone. A similar event was
observed, although to a lesser extent, with plasmid pJ06
DNA, a pBR322 derivative harbouring the dps gene from
E. coli (data not shown). Although we cannot explain the
difference in transformation efficiencies, our data may
indicate that Dps proteins also modulate the establish-
ment of plasmid replicons in bacteria, possibly via inter-
actions between Dps and plasmid components (DNA or
proteins involved in replication).
DpsA is a mobile gene in O. oeni and about 40% of the
tested strains harboured the corresponding sequences.
The anomalous GC content of the dpsA region, the varia-
tion in codon usage content and the presence of mobile
sequences in the vicinity strongly suggest a foreign origin
of the locus, possibly through a transposition event. Inter-
estingly, homologues to both IS have been recently associ-
ated with the horizontal transfer of stress response
mechanisms in other LAB (Suokko et al. 2005; Marcobal
et al. 2006). Other Dps-like proteins have been identified
on mobile elements such as the MP1 mega plasmid from
Deinococcus radiodurans (White et al. 1999) or the LP65
bacteriophage from Lb. plantarum (Chibani-Chennoufi
et al. 2004). In contrast, the genetic organization of the
DpsA ortholog-containing regions of other LAB was
different compared to O. oeni. In particular, the absence
of surrounding IS sequences suggests that dpsA are not
mobile genes in these species, or that extended recombi-
nation took place after transfer.
Although the dpsA gene protected E. coli from the
deleterious effects of wine, we could not associate the
presence of the oenococcal sequence with increased fitness
in response to a wine environment. Hence dpsA sequences
were not prevalent in isolates with remarkable oenological
properties (i.e. high survival and growth in wine after
inoculation, accompanied by malate decarboxylation).
This apparent discrepancy can be explained by the fact
that we assessed the presence of a dpsA sequence, which
indicates the presence ⁄ absence of the gene. However,
positive strains may exhibit different DpsA activities
because of variations in the expression level and ⁄ or in the
stability of the protein. These aspects have now to be
examined. The second point is that like in other bacteria,
0·1
1
10
100S
urvi
val
(%)
IOE
B-S
AR
CO
438
aV
OIO
EB
841
3IO
EB
-SA
RC
O14
91IO
EB
-SA
RC
0 43
3b B1
PS
U-1
IOE
B-S
AR
CO
396 VF
IOE
B-S
AR
CO
384
IOE
B-S
AR
CO
422
Figure 4 Tolerance of various O. oeni isolates to oxidative stress. Sur-
vival of log phase cells after exposure to MRS containing H2O2
40 mmol l)1 for 60 min was determined as described in the materials
and methods part. The experiment was conducted in triplicate. dpsA+
and dpsA) strains are represented with hatched and white bars,
respectively.
A. Athane et al. Acquired dps-containing gene island in lactic acid bacterium O. oeni
ª 2008 The Authors
Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 1873

the stress response system to wine or oxidative conditions
in O. oeni is probably complex and can be decomposed
into functional (and maybe compensating) modules. dpsA
may be one of them, even though its contribution to the
overall resistance can be expected to be weak, since it
does not appear critical to survival. Alternatively, a dpsA
deficiency could be complemented by the presence
and ⁄ or enhanced activity of other resistance protein(s), or
by a Dps-paralog. The latter possibility is supported by
the observation that the O. oeni PSU-1 genome appears
to encode a second Dps-like protein. Paired Dps proteins
have been found in Bacillus and Lactococcus lactis but are
rare in other bacteria (Liu et al. 2006). Although these
hypothesis require further investigation, our results stress
the need in the future to address the role of ferritin-like
proteins in helping O. oeni to cope with its environment.
Acknowledgements
E. Bilhere received a grant from the French Ministry of
Education.
References
Almiron, M., Link, A.J., Furlong, D. and Kolter, R. (1992)
A novel DNA-binding protein with regulatory and protec-
tive roles in starved Escherichia coli. Genes Dev 6, 2646–
2654.
Brun, L.A., Le Corff, J. and Maillet, J. (2003) Effects of ele-
vated soil copper on phenology, growth and reproduction
of five ruderal plant species. Environ Poll 122, 361–368.
Ceci, P., Mangiarotti, L., Rivetti, C. and Chiancone, E. (2007)
The neutrophil-activating Dps protein of Helicobacter
pylori HP-NAP adopts a mechanism different from
Escherichia coli Dps to bind and condense DNA. Nucleic
Acid Res 35, 2247–2256.
Chibani-Chennoufi, S., Dillmann, M.L., Marvin-Guy, L.,
Rami-Shojaei, S. and Brussow, H. (2004) Lactobacillus
plantarum bacteriophage LP65 : a new member of the
SPO1-like genus of the family Myoviridae. J Bacteriol 186,
7069–7083.
Choi, S.H., Baumler, D.J. and Kaspar, C.W. (2000) Contribu-
tion of dps to acid stress tolerance and oxidative stress
tolerance in Escherichia coli O157:H7. Appl Environ Micro-
biol 66, 3911–3916.
Delaherche, A., Bon, E., Dupe, A., Lucas, M., Arveiler, B., De
Daruvar, A. and Lonvaud-Funel, A. (2006) Intraspecific
Diversity of Oenococcus oeni Strains Determined by
Sequence Analysis of Target Genes. Appl Microbiol Technol
73, 394–403.
Divol, B., Tonon, T., Morichon, S., Gindreau, E. and Larvaud-
Funel, A. (2003) Molecular characterization of Oenococcus
oeni genes encoding proteins involved in arginine trans-
port. J Appl Microbiol 94, 738–746.
Freestone, P., Grant, S., Trinei, M., Onada, T. and Norris, V.
(1998) Protein phosphorylation in Escherichia L form
NC-7. Microbiology 144, 3289–3295.
Gindreau, E., Joyeux, A., De Revel, G., Claisse, O. and
Lonvaud-Funel, A. (1997) Evaluation of the settling of
malolactic starters within the indigenous microflora of
wines. J Inter des Sciences de la Vigne et du Vin 31, 197–202.
Gourion, B., Rossignol, M. and Vorholt, J.A. (2006) A proteo-
mic study of Methylobacterium extorquens reveals a
response regulator essential for epiphytic growth. Proc Natl
Acad Sci USA 103, 13186–13191.
Hanna, E.S., Roque-Barreira, M.C., Bernardes, E.S., Panunto-
Castelo, A., Sousa, M.V., Almeida, I.C. and Brocchi, M.
(2007) Evidence for glycosylation on a DNA-binding
protein of Salmonella enterica. Microb Cell Fact 6, 11.
Hasley, T., Vasquez-Torres, A., Gravdahl, D., Fang, F. and
Libby, S. (2004) The Ferritin-Like Dps Protein Is Required
For Salmonella enterica Serovar Typhimurium Oxidative
Stress Resistance and Virulence. Infect Immun 72, 1155–
1158.
Kovach, M.E., Elzer, P.H., Hill, D.S., Robertson, G.T., Farris,
M.A., Roop, R.M. and Peterson, K.M. (1995) Four new
derivatives of the broad-host-range cloning vector
pBBR1MCS, carrying different antibiotic-resistance
cassettes. Gene 166, 175–176.
Lacqua, A., Wanner, O., Colangelo, T., Martinotti, M.G. and
Landini, P. (2006) Emergence of biofilm-forming subpop-
ulations upon exposure of Escherichia coli to environmen-
tal bacteriophages. Appl Environ Microbiol 72, 956–959.
Le Marrec, C., Bon, E. and Lonvaud-Funel, A. (2007)
Tolerance to high osmolality of the lactic acid bacterium
Oenococcus oeni and identification of potential osmo-
protectants. Int J Food Microbiol 115, 335–342.
Liu, X., Kim, K., Leighton, T. and Theil, E.C. (2006) Paired
Bacillus anthracis Dps (mini-ferritin) have different reacti-
vities with peroxide. J Biol Chem 281, 27827–27835.
Malone, A.S., Chung, Y.K. and Yousef, A.E. (2006) Genes of
Escherichia coli O157:H7 that are involved in high-pressure
resistance. Appl Environ Microbiol 72, 2621–2671.
Marcobal, A., de Las rivas, B., Moreno-Arribas, M.V. and
Munoz, R. (2006) Evidence for horizontal gene transfer as
origin of putrescine production in Oenococcus oeni RM83.
Appl Environ Microbiol 72, 7954–7958.
Mills, D., Rawsthorne, H., Parker, C., Tamir, D. and Makar-
ova, K. (2005) Genomic Analysis of Oenococcus oeni PSU-1
and its relevance to winemaking. FEMS Microbiol Rev 29,
465–475.
Mirlean, N., Roisenberg, A. and Hies, J.O. (2005) Copper-
based fungicide contamination and metal distribution in
Brazilian grape products. Bull Environ Cont Tox 75,
968–974.
Morikawa, K., Ohniwa, R.L., Kim, J., Maruyama, A., Ohta, T.
and Takeyasu, K. (2006) Bacterial nucleoid dynamics:
oxidative stress response in Staphylococcus aureus. Genes
Cells 11, 409–423.
Acquired dps-containing gene island in lactic acid bacterium O. oeni A. Athane et al.
1874 Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875
ª 2008 The Authors

Nair, S. and Finkel, S. (2004) Dps Protects Cells Against
Multiple Stresses During Stationary Phase. J Bacteriol 186,
4192–4198.
Olsen, K.N., Larsen, M.H., Mac Gahan, C., Kallipolitis, B.,
Wolf, X.A., Rea, R., Hill, C. and Ingmer, H. (2005) The
Dps-like Fri of Listeria monocytogenes promotes stress
tolerance and intracellular multiplication in macrophage-
like cells. Microbiology 151, 925–933.
Park, S., You, X. and Imlay, J.A. (2005) Substantial DNA
damage from submicromolar intracellular hydrogen
peroxide detected in Hpx) mutants of Escherichia coli.
Proc Natl Acad Sci USA 102, 9317–9322.
Pena, M.M. and Bullerjahn, G.S. (1995) The DpsA protein of
Synechococcus sp. Strain PC C7942 is a DNA-binding
hemoprotein. Linkage of the Dps and bacterioferritin
protein families. J Biol Chem 38, 22478–22482.
Robinson, B., Greven, M., Green, S., Sivakumaran, S.,
Davidson, P. and Clothier, B. (2006) Leaching of copper,
chromium and arsenic from treated vineyard posts in
Marlborough, New Zealand. Sci Tot Environ 364, 113–123.
Spano, G. and Massa, S. (2006) Environmental stress response
in wine lactic acid bacteria: beyond Bacillus subtilis. Crit
Rev Microbiol 32, 77–86.
Stephani, K., Weichart, D. and Hengge, R. (2003) Dynamic
Control of Dps Protein Levels by ClpXP and ClpAP
Proteases in Escherichia coli. Mol Biol 49, 1605–1614.
Suokko, A., Savijoki, K., Malinen, E., Palva, A. and Varmanen,
P. (2005) Characterization of a mobile clpL gene from
Lactobacillus rhamnosus. Appl Environ Microbiol 71, 2061–
2069.
Weber, A., Kogl, S.A. and Jung, K. (2006) Time-dependent
proteome alterations under osmotic stress during aerobic
and anaerobic growth in Escherichia coli. J Bacteriol 188,
7165–7175.
Weichart, D., Ouerfurth, N., Dreger, M. and Hengge-Aronis,
R. (2003) Global control for ClpP-containing proteases in
stationary-phase adaptation of Escherichia coli. J Bacteriol
185, 115–215.
White, O., Eisen, J.A., Heidelberg, J.F., Hickey, E.K., Peterson,
J.D., Dodson, R.J. and Haft, D.H. et al. (1999) Genome
sequence of the radioresistant bacterium Deinococcus
radiodurans R1. Science 286, 1571–1577.
A. Athane et al. Acquired dps-containing gene island in lactic acid bacterium O. oeni
ª 2008 The Authors
Journal compilation ª 2008 The Society for Applied Microbiology, Journal of Applied Microbiology 105 (2008) 1866–1875 1875




















