
centro de investigación y de estudios - Repositorio CINVESTAV
51
CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS AVANZADOS DEL INSTITUTO POLITÉCNICO NACIONAL UNIDAD ZACATENCO DEPARTAMENTO DE INFECTÓMICA Y PATOGÉNESIS MOLECULAR “Evaluación de la actividad antiviral de los fármacos NDGA, vitamina D, lovastatina e ivermectina ante infección con Dengue en células HuH7” T E S I S Que presenta BIÓLOGA ARELY CRUZ PÉREZ Para obtener el grado de MAESTRA EN CIENCIAS EN INFECTÓMICA Y PATOGÉNESIS MOLECULAR Directora de la Tesis: Dra. Rosa María del Ángel Nuñez Ciudad de México Agosto, 2017
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of centro de investigación y de estudios - Repositorio CINVESTAV

CENTRO DE INVESTIGACIÓN Y DE ESTUDIOS
AVANZADOS DEL INSTITUTO POLITÉCNICO NACIONAL
UNIDAD ZACATENCO
DEPARTAMENTO DE INFECTÓMICA Y PATOGÉNESIS
MOLECULAR
“Evaluación de la actividad antiviral de los fármacos NDGA,
vitamina D, lovastatina e ivermectina ante infección con Dengue en
células HuH7”
T E S I S
Que presenta
BIÓLOGA ARELY CRUZ PÉREZ
Para obtener el grado de
MAESTRA EN CIENCIAS EN INFECTÓMICA Y PATOGÉNESIS
MOLECULAR
Directora de la Tesis:
Dra. Rosa María del Ángel Nuñez
Ciudad de México Agosto, 2017

Agradecimientos
Agradezco al Consejo Nacional de Ciencia y Tecnología por la beca No.
583442 brindada para la realización de este proyecto, el cual fue
realizado en el laboratorio de Virología del Departamento de
Infectómica y Patogénesis Molecular del Centro de Investigaciones y
Estudios Avanzados del Instituto Politécnico Nacional, bajo la tutoría de
la Dra. Rosa María del Ángel Núñez de Cáceres y la asesoría de la Dra.
Ana Lorena Gutiérrez Escolano y del Dr. Juan Santiago Salas Benito.

ÍNDICE
Resumen…………………..…………………………………………………..1
Abstract…………………...……………………………………………………2
Introducción
Generalidades…………………………….…………………………..3
Antecedentes
Patogenia…………..………………………………………………….3
DENV…………………………….…………………………………….4
Ciclo replicativo viral…………………….…………………………....6
Importancia de los lípidos en la infección de dengue…...…….....8
NDGA…………………………………...…………………………….10
Lovastatina……….…………………………………………………..12
Vitamina D3……..……………………………………………………13
Ivermectina…………....……………………………………………..15
Justificación…………...……………...……………………………………...18
Hipótesis……………………………………………….……………………..19
Objetivo general……………..………………………………………………19

Objetivos particulares……………………………………...…………….....19
Material y Métodos
Células HuH7…………………………………………………….…..20
Stock viral DENV2 y DENV4……………………………………….20
Infección y tratamiento……………………………………………...20
Evaluación de la infección………………………………………….21
Citometría de flujo…………………………………………………...21
Western blot……………………………………………………….....22
Ensayo de focos……………………………………………………..22
Ensayos de viabilidad…………………………………………….…23
Análisis estadísticos………………………………..........……...….23
Resultados
Efecto de los fármacos en el porcentaje de células
infectadas…………………………………………………………….23
Efecto de los fármacos en la cantidad de proteína
viral……………………………………………………………………26
Efecto de los fármacos en la producción de progenie
viral.………….………………………………..………………………29
Viabilidad Ivermectina-Lovastatina.……………………………….31
Tratamiento de ivermectina-lovastatina en
células infectadas con DENV………………………………………31
Discusión……………………………………………………………………..37
Conclusiones………………..……………………………………………….37
Perspectivas…………………………………………………………………43
Referencias…………………………………………..………………………44

1
RESUMEN
El virus del dengue (DENV), perteneciente a la familia Flaviviridae, es el
agente etiológico de la fiebre del dengue y de su principal complicación
el dengue hemorrágico. A pesar de que los brotes de dengue se han
reportado desde hace más de 60 años, aún no se cuenta con un
tratamiento antiviral específico, ni con una vacuna capaz de conferir una
alta protección contra la infección. Sin embargo, en años recientes se
han descrito algunos fármacos, aprobados por la FDA, con actividad
antiviral contra DENV. Tal es el caso del NDGA que inhibe la síntesis de
lípidos y a su vez la replicación viral; la lovastatina que es inhibidor de la
síntesis del colesterol reduciendo la replicación de virus; la vitamina D3,
que regula la repuesta inmune; y la ivermectina que impide el importe
nuclear de la proteína NS5. Dado que todos estos fármacos han sido
evaluados de manera independiente uno del otro, fue de nuestro interés
identificar cuál de estos fármacos ejerce una mayor actividad antiviral y
cuál es el efecto de la combinación ivermectina-lovastatina ante la
infección con DENV2 y DENV4 en células hepáticas HuH7. En este
estudio se observó que vitamina D3 no induce una reducción
significativa de la infección por DENV, mientras que ivermectina es el
fármaco con un mejor efecto antiviral. Finalmente la combinación
ivermectina-lovastatina puede ser una buena opción para tratar
infección por DENV.
Con formato: Español (México)

2
ABSTRACT
Dengue virus belongs to the Flaviviridae family and is the etiologic agent
of dengue fever and its main complication, dengue hemorrhagic fever.
Although dengue disease cases have been reported for over 60 years,
there is no specific antiviral treatment or high protective vaccine to avoid
infection. Nevertheless in recent years it has been described some
drugs approved by FDA, with antiviral activity against DENV. Some of
them are NDGA, which inhibit lipid synthesis and reduces viral
replication; lovastatin, an inhibitor of the cholesterol synthesis, inhibiting
viral replication; vitamin D3, which regulates the immune response; and
ivermectin which blocks the nuclear import of the viral protein NS5.
Since all these drugs have been evaluated independently, it was our
interest to identify which of these drugs has a better antiviral activity and
what is the effect of the combination of ivermectin- lovastatin in the
infection with DENV2 and DENV4 in hepatic cells HuH7. In this study we
observed that vitamin D does not induce any significant viral reduction,
while ivermectin is the drug with the highest antiviral activity. Finally, the
combination ivermectin-lovastatin can be an effective strategy against
DENV because it induced the highest anti-DENV activity.

3
INTRODUCCIÓN
Generalidades
El virus del dengue (DENV) es un virus perteneciente a la familia
Flaviviridae del género Flavivirus, causante de la fiebre del dengue y de
su principal complicación el dengue hemorrágico. Cada año se registran
390 millones de infecciones por este virus, de las cuales únicamente 96
millones son sintomáticas (Gubler et al., 2014). DENV es un virus
envuelto cuyo genoma es una molécula de RNA de cadena positiva y
es transmitido por mosquitos de las especies Aedes aegypti y A.
albopictus (Clyde et al., 2006)
A pesar de que el virus infecta al hombre desde hace más de 60 años,
aún no se cuenta con un tratamiento antiviral específico, así como
tampoco se cuenta con una vacuna capaz de conferir una alta
protección contra la infección. Resultados de nuestro grupo han
permitido identificar algunos fármacos como el ácido
nordihidroguaiarético (NDGA), lovastatina, vitamina D3 y recientemente
a la ivermectina con actividad antiviral contra DENV. Su uso solo o en
combinación constituye una opción prometedora para el tratamiento
viral.
Antecedentes
Patogenia
DENV es el agente causal del dengue clásico, dengue hemorrágico y
síndrome de shock por dengue. El dengue clásico se caracteriza por
síntomas como fiebre que dura de 2 a 5 días, dolor de cabeza, mialgia y
salpullido, mientras que las formas graves como dengue hemorrágico
provocan incremento de la permeabilidad vascular, trombocitopenia y
manifestaciones hemorrágicas. Por último, en el caso del síndrome de

4
choque por dengue el principal síntoma es la fuga de fluido de los
espacios intersticiales que pueden causar la muerte (Clyde et al., 2006).
Es importante mencionar que la infección con un serotipo de DENV no
provee inmunidad ante los otros serotipos, y de hecho, infecciones
subsecuentes con diferentes serotipos, pueden causar las formas
graves de la enfermedad por un proceso conocido como amplificación
de la infección dependiente de anticuerpos (ADE: del inglés antibody-
dependent enhancement) (Damonte et al., 2004; Acosta EG &
Bartenschlager R, 2016).
A pesar de que aún no se conoce exactamente la causa del por qué se
desarrollan las formas graves de la enfermedad, la teoría más aceptada
es la del ADE, donde se propone que en una segunda infección con
otro serotipo de DENV, ocurre una reacción cruzada de anticuerpos no
neutralizantes contra el nuevo serotipo de DENV, causando que el
complejo partícula viral-anticuerpo no neutralizante se una a los
macrófagos mediante la fracción Fc. Dado que los macrófagos son uno
de los principales blancos del virus, la infección se potencializa, así
como la respuesta inmune y la producción de citocinas proinflamatorias
tales como TNFα, IL6, IL8, IL10 e IL12 (Ahmed S et al., 2009; Guzman
MG & Harris E, 2015). Esta elevación de la respuesta inmune y
sobreproducción de citocinas es la causante del daño en el endotelio
vascular (Damonte EB et al., 2004; Guzman MG & Harris E, 2015).
DENV
DENV es un virus de la familia Flaviviridae del género Flavivirus. El
nombre de la familia Flaviviridae proviene del latín Flavis que significa
amarillo, por el virus de la fiebre amarilla, primer virus identificado
perteneciente a la familia. DENV es un arbovirus y es transmitido al
hombre por la picadura de mosquitos del género Aedes, los cuales se

5
encuentran ampliamente distribuidos en el mundo (Clyde et al., 2006).
Se han identificado cuatro diferentes serotipos (DENV1 a DENV4), y a
pesar de que los cuatro serotipos de DENV poseen una homología en
secuencia de aminoácidos de apenas 60-70% (Guzman & Harris, 205;
Acosta & Bartenchiager, 2016), en cuestiones de epidemiología, éstos
se comportan de manera similar. Los serotipos DENV2 y DENV3 en
general se han asociado con casos severos de la enfermedad
(Balsameda et al.,2006)
La partícula viral madura de DENV es esférica de 500 Å de diámetro y
está compuesta por una sola cadena de RNA de polaridad positiva. La
partícula se encuentra empaquetada por la proteína estructural C que
forma la nucleocápside y ésta a su vez está envuelta por una bicapa
lipídica en la que se encuentran 180 copias de dos glicoproteínas: E y
prM. El genoma de aproximadamente 10.8 kb tiene un único marco de
lectura abierto (ORF) que codifica para una poliproteína, de la cual, a
partir del extremo amino, se traducen las tres proteínas estructurales
que constituyen la partícula viral: cápside (C), membrana (M, expresada
como prM que es el precursor de M) y envoltura (E); y a continuación
las siete proteínas no estructurales que son esenciales para la
replicación viral: NS1, NS2A, NS2B, NS3, NS4A, NS4B y NS5
(Mukhopadhyay et al., 2005) (Figura 1).

6
Figura 1. Estructura del genoma de DENV. Tomada de Clyde et al., 2006.
DENV es capaz de infectar in vitro distintos tipos celulares como células
dendríticas, monocitos, macrófagos, células B, células T, células
endoteliales, hepatocitos y células neuronales; sin embargo, in vivo se
ha visto que las células fagocíticas mononucleares (monocitos,
macrófagos y células dendríticas) son las principales células infectadas
por DENV (Clyde et al., 2006)
Ciclo replicativo viral
DENV es capaz de entrar a las células de mamífero y mosquito por
endocitosis mediada por clatrina después de interaccionar con su
receptor celular. Hasta ahora, se han descrito numerosos receptores
que se unen a DENV en células de mamífero, entre estos se
encuentran el heparán sulfato, Hsp70 y Hsp90, GRP78, el receptor de
laminina, CLEC5B, el receptor de manosa, entre otros, sin embargo, la
interacción mejor estudiada es con el receptor DC-SIGN. DC-SIGN es
una lectina que interacciona con los residuos de manosa de la proteína
E de los cuatro serotipos de DENV. Una vez que el virus interacciona
con el receptor, éste es movilizado a sitios recubiertos por clatrina en
donde el virus es internalizado por medio de vesículas. Eventualmente

7
estas vesículas forman endosomas tempranos y tardíos, y es en los
endosomas tardíos donde la acidificación causa cambios
conformacionales de la proteína E que inducen la fusión de la
membrana viral con la membrana endosomal. Posteriormente, el RNA
de cadena positiva se disocia de la cápside y es liberado al citosol
funcionando como un RNA mensajero que es traducido en ribosomas
presentes en el retículo endoplásmico rugoso. La poliproteína
sintetizada se escinde de manera co y postraduccional en al menos 10
proteínas maduras. El extremo amino codifica a las proteínas
estructurales y el resto a las proteínas no estructurales. Una vez que las
proteínas virales son traducidas y procesadas, el virus induce hipertrofia
en las membranas intracelulares generando los llamados paquetes
vesiculares y complejos de membrana en donde se lleva a cabo la
síntesis del RNA viral que comienza con la síntesis del RNA de cadena
negativa complementaria a la del RNA viral que va a servir como molde
para la amplificación de cadenas positivas de RNA viral, reacción
catalizada por NS5 en asociación con NS3 (Gubler et al., 2014).
El nuevo material genético sintetizado puede seguir tres vías: traducirse
para la producción de proteína, servir de molde para la síntesis de RNA
de polaridad negativa o bien, asociarse a la cápside viral para la
formación de nuevas partículas virales. En la células de mamífero,
éstas partículas virales se distribuyen en el retículo endoplásmico,
núcleo y superficie de gotas lipídicas. Una vez formadas las
nucleocápsides en el lúmen del retículo endoplásmico, éstas adquieren
la bicapa lipídica y las proteínas E y prM de las membranas del retículo
endoplásmico, y son liberadas por la vía secretora. Para la maduración
de las partículas virales es importante la proteólisis de prM mediada por
furina en el trans-Golgi para el rearreglo y la homodimerización de
E(Gubler et al.,2014) (Figura 2).

8
Figura 2. Replicación de DENV. El virus se une a la membrana celular y
entra por endocitosis mediada por receptor. El virus es capaz de fusionar su
membrana con la del endosoma para liberar su genoma en el citoplasma y
poder llevar a cabo su replicación, traducción y ensamblaje en el RE.
Finalmente la maduración del virus se realiza en el trans-Golgi y las nuevas
partículas son liberadas por exocitosis (Tomada de Mukhopadhyay et al.,
2005).
Importancia de los lípidos en la infección de dengue
Los lípidos son moléculas importantes en diversos pasos del ciclo
replicativo de los virus de la familia Flaviviridae, debido a que participan
en la formación de la envoltura del virus y a que son componentes de la
plataforma celular a través de la cual el virus entra y lleva a cabo la

9
replicación, traducción y ensamble viral. En diversos tipos celulares se
ha demostrado que los receptores virales se encuentran en balsas
lipídicas (ricas en colesterol) o son translocadas a ellas para la
internalización viral (Martinez-Gutierrez et al., 2011). La presencia de
colesterol en las balsas lípidicas es indispensable para la entrada viral,
ya que se ha observado que la eliminación del colesterol con
compuestos como metilciclodextrina, inhibe la infección por DENV
(Reyes-del Valle et al, 2005; Puerta-Guardoet al., 2012). Posterior a la
unión y entrada, un paso importante en la internalización viral es la
fusión entre la membrana viral y endosomal, proceso que requiere que
la membrana celular contenga lípidos aniónicos como bis
(monoacilglicerol) fosfato y fosfatidil serina, resaltando nuevamente la
participación de los lípidos (Soto-Acosta et al., 2017). Una vez en el
citoplasma, el virus necesita una buena cantidad de lípidos y colesterol
para formar las invaginaciones del retículo endoplásmico (RE) y realizar
la remodelación del retículo endoplásmico rugoso (RER), eventos
necesarios para la replicación y síntesis de la proteína viral(Peña J
Harris, 2012). De hecho, es la proteína NS3 quien secuestra a la
sintasa de ácidos grasos (FAS) y la lleva al RE manteniéndola activa y
por lo tanto causando un aumento de ácidos grasos tras la infección por
DENV (Acosta EG & Bartenschlager R, 2016). En estas invaginaciones
formadas a partir del RE, se encuentran las proteínas necesarias para
la replicación como la NS5 que es la RNA polimerasa dependiente de
RNA, las proteínas NS2B, NS3, NS4A, NS4B y el RNA de doble cadena
(Villareal et al., 2015). Es importante resaltar que dichas vesículas están
compuestas principalmente de esfingolípidos y esteroles por lo que la
participación de ácidos grasos y colesterol es importante. Finalmente
para la morfogénesis se ha descrito que la proteína C se acumula
alrededor de las gotas de lípidos (LD), ricas en lípidos y esteres de

10
colesterol para la formación de la partícula viral (Samsa et al., 2009).
Así que la inhibición de la síntesis del colesterol mediante estatinas, o a
través del bloqueo de la vía de biosíntesis del colesterol usando siRNA
reduce entrada y replicación viral (Martinez-Gutierrez et al., 2011;
Rothwell et al., 2009). Acorde a la necesidad de colesterol, se ha
descrito que hay un aumento en la cantidad de colesterol en las células
infectadas con DENV. Este aumento se debe a un incremento en la
captación de partículas de lipoproteínas de baja densidad a través del
aumento del receptor de lipoproteínas de baja densidad (LDLr) y a un
aumento en la síntesis de colesterol debido a la activación de la
reductasa 3-hidroxi-3-metil-glutaril-CoA (HMGRC) (enzima limitante en
la síntesis de colesterol) por su defosforilación (Soto-Acosta et al.,
2014). Dada la importancia de los lípidos en la replicación viral, estos
constituyen un blanco interesante para el desarrollo de antivirales.
NDGA (Ácido nordihidroguaiarético)
El NDGA (Figura 3) es el lignano principal de la planta gobernadora
(Larrea tridentata) a la cual se le han atribuido una amplia variedad de
actividades farmacológicas, su principal y mayormente descrita función
es la de ser inhibidorade la 5-lipoxigenasa de ácido araquidónico, sin
embargo la propiedad antioxidante, hipolipemiante, anti-inflamatoria,
potenciadora del sistema inmune (Lü et al., 2010) e inhibitoria de
algunos virus como VIH, HPV, virus del Herpes simple (Hwu et al.,
2008), así como de DENV (Soto et al., 2014) también ha sido descrita
(Arteaga et al., 2005).

11
Figura3 Estructura ácido nordihidroguaiarético (NDGA). Tomada de
Arteaga et al., 2005.
La planta Larrea tridentata es usada de manera tradicional en
infusiones o en tabletas para una variedad de padecimientos, aunque la
mayoría de estos usos no son respaldados clínica o experimentalmente.
Lo que si se ha descrito es su toxicidad, en donde se indica que el alto
consumo de la planta, por sus altos contenidos de NDGA, puede causar
daño a riñón e hígado. Debido a que el NDGA es metabolizado por la
COMT (Catecol-O-metiltransferasa) y posteriormente oxidado, produce
como metabolitos secundarios o-quinonas y/o p-quinonas, que pueden
ser los responsables de la toxicidad del fármaco (Billinsky et al., 2007;
Grice et al., 1968; Jeong et al., 2017; Lambert et al., 2002). La
farmacocinética y farmacodinámica del NDGA no han sido muy
explorados, sin embargo Lambert y colaboradores (2001) describen que
en ratones inyectados de manera intravenosa con 50 mg/kg del
fármaco, éste alcanza un pico en la concentración plasmática de
14.7mg/ml, con una vida media de 135 min y una eliminación del
torrente sanguíneo de 201.9 ml/mi/kg.
El NDGA causa un efecto hipolipemiante primero, debido a la
importancia que el ácido araquidónico tiene en la regulación de la
homeostasis del colesterol (Demetz et al., 2014) y segundo por su
capacidad de aumentar la actividad de la AMPK y por lo tanto disminuir
la activación de SREBPs (proteína de unión a elementos de respuesta a
esterol). Las proteínas SREBPs son factores transcripcionales que

12
actúan sobre genes que están relacionados con la síntesis de ácidos
grasos y colesterol. Además NDGA actúa sobre la acetil-CoA
carboxilasa y la ácido graso sintasa, causando la expresión de genes
implicados en el catabolismo de ácidos grasos libres y el efecto
hipolipidémico in vivo e in vitro (Lee et al., 2010; Syed & Siddiqui.,
2011). Este efecto hipolipidémico es capaz de inhibir la infección por
DENV en células HuH7 a través de la reducción de la replicación del
genoma viral y la inhibición del ensamblaje del virión (Soto-Acosta et al.
2014).
Lovastatina
Otros compuestos inhibidores de la síntesis de colesterol son las
estatinas. La lovastatina es una estatina lipofílica perteneciente al grupo
de las estatinas naturales extraídas del hongo Aspergillus terreus, y es
un inhibidor competitivo reversible que actúa de forma dosis-
dependiente de la HMGR (3-hidroxi-3-metilglutaril-coenzima A
reductasa) la cual es la enzima limitante en la síntesis de colesterol
(Gazerro et al., 2012).
Sobre la farmacología del fármaco, la dosis empleada en pacientes es
de 40mg, su absorción es de 31%, tiene una biodisponibilidad menor a
5%, el tiempo en que alcanza su pico máximo de concentración
plasmática es de 2 a 4 horas, la unión a proteínas plasmáticas es de
96%, es metabolizado de manera hepática mediante CYP3A4, la Ic50 es
de 2.7-11.1 nM y la vida media es de 2.5- 3 h (Gazerro et al., 2012).
Las estatinas, además de reducir los niveles de colesterol, son capaces
de inhibir a las lipoproteínas de baja densidad (LDL) y triglicéridos
(Malhotra & Goa, 2001) y tienen un efecto en la función endotelial que
resulta en una reducción en la expresión de citocinas proinflamatorias
(Greenwood J & Mason JC, 2007). Esto último controla la migración

13
leucocitaria en áreas de inflamación endotelial. Igualmente, las
estatinas pueden mejorar la función endotelial modulando la producción
de especies reactivas del oxígeno (Greenwood J & Mason JC, 2007).
Así, el tratamiento con estatinas como lovastatina no solo sería capaz
de inhibir el ciclo replicativo viral debido a la reducción de los niveles de
colesterol (Whitehorn et al., 2016) sino que también podría modular la
respuesta inflamatoria endotelial tan importante durante las formas
graves de la infección por DENV.
Vitamina D3
Como se mencionó anteriormente, los casos de Dengue grave están
asociados a una segunda infección por DENV y a un incremento
exacerbado de la producción de citocinas proinflamatorias por células T,
macrófagos y células endoteliales causando disfunción endotelial, que
genera extravasación sanguínea (Arboleda & Urcuqui., 2016). Es por
eso, que en el combate de la infección por Dengue, una alternativa es
encontrar fármacos que controlen la respuesta inmune, y tal es el caso
de la vitamina D3 que además de ser importante en el metabolismo del
calcio y de fosfato, tiene una función inmunoreguladora (Di Rosa et al.,
2011).
La 1,25-dihidroxivitamina D3 [1,25(OH)2D3], es la forma
hormonalmente activa de la vitamina D que genera respuestas celulares
como diferenciación y proliferación celular, además de potenciamiento
de la respuesta inmune ante cáncer y enfermedades autoinmunes (Di
Rosa et al., 2011), Esta vitamina se forma de manera natural por
irradiación solar en la piel a partir de 7-dehidrocolesterol, forma de
síntesis más abundante en el cuerpo, aunque también se puede adquirir
mediante la dieta, donde en adultos sanos la concentración plasmática
puede alcanzar los 20ng/mL con 600-800 IU por día (Christakos et al.,

14
2016; Jetter et al., 2014). La vitamina D3 es producida principalmente
en el riñón por la acción de la enzima CYP27B1, pero también se puede
producir en células del sistema inmune, ya que el reconocimiento de
PAMPs por los TLRs presentes en la células del sistema inmune
estimula la activación de CYP27B1 y por tanto, la producción de 1, 25-
dihidroxivitamina D3 (Borella et al., 2014).
Dentro de las funciones inmunomoduladoras descritas para la vitamina
D3 están la supresión de la proliferación celular de tipo Th-1 que
conduce a la producción reducida de interferón gamma e interleucina 2;
conduciendo a una menor presentación de antígeno por parte de las
células dendríticas, menor reclutamiento de linfocitos T y proliferación.
De manera general, la vitamina D3 polariza el sistema inmune hacia
una respuesta Th2, por lo que las citocinas asociadas aumentan (Beard
et al., 2011; Chun et al., 2014). De igual manera existen muchos
estudios que reportan la importancia de la vitamina D3 en la inducción
de autofagia, como un mecanismo de protección ante patógenos (Chun
et al., 2014)
Estudios con vitamina D3 (metabolito activo de la vitamina D)
demuestran también que esta hormona es capaz de inhibir la respuesta
inmune adaptativa y la proliferación celular, y por otro lado, es capaz de
promover la diferenciación celular y la respuesta inmune innata
(Arboleda & Urcuqui., 2016). Tratamientos en cultivo con monocitos han
demostrado que la Vitamina D3 es capaz de disminuir la expresión de
algunos TLRs como lo son TLR2 y TLR4 de una manera dosis-
dependiente, además de ser capaces de disminuir la producción de
citocinas proinflamatorias ante una infección con DENV (Puerta-Guardo
et al., 2012)
A pesar de que una vez que entra DENV a la célula, los principales TLR
que lo reconocen son TLR3, TL7 y TLR8 (Green et al., 2014), se ha

15
descrito que la proteína NS1 de DENV, es reconocida por los
TLR2,TLR4 Y TLR6 promoviendo la producción de citocinas
proinflamatorias, por lo que vitamina D3, al disminuir la expresión de
TLR2 y TLR4 y por lo tanto de citocinas proinflamatorias, puede ser un
fármaco ideal para tratar la enfermedad de Dengue, en especial de las
formas graves (Arboleda& Urcuqui., 2016).
Ivermectina
DENV durante su ciclo replicativo emplea diferentes componentes y
proteínas celulares. Entre ellas, utiliza diversas proteínas nucleares, las
cuales en la mayoría de los casos son translocadas del núcleo al
citoplasma (Pryor et al., 2007). Este exporte/importe nuclear incluye
también a algunas proteínas virales como NS5 y C, las cuales en
distintos momentos de la infección pueden estar presentes en el núcleo
de la célula infectada (Bhuvanakantham et al;2009; Pryor et al., 2007).
Es difícil entender el motivo por el cual las proteínas virales son
importadas al núcleo, cuando el ciclo replicativo de DENV es
citoplásmático, sin embargo, este proceso es indispensable ya que se
ha demostrado que la infección por DENV se inhibe de manera drástica
al tratar con ivermectina, un fármaco capaz de evitar el importe nuclear
de proteínas como NS5 y NS3.
La ivermectina es un fármaco antihelmíntico de amplio espectro que
proviene del compuesto activo avermectina aislado en Japón en 1974 a
partir de la bacteria Streptomycetes avermetilis. Inicialmente, la
ivermectina fue lanzada al mercado en 1981 únicamente para uso en
animales, sin embargo, 6 años después se empezó a emplear también
en humanos. El fármaco es capaz de bloquear la neurotransmisión,
interfiriendo con la sinapsis neuromuscular, actúa en los canales de
cloro y entradas de glutamato causando parálisis en invertebrados. En

16
humanos, la ivermectina actúa estimulando la salida de GABA en las
neuronas, sin embargo, la barrera hemato-encefálica impide la entrada
del fármaco hacia el cerebro (Omura, 2008).
Sobre la farmacología de ivermectina, la única vía de administración
aprobada es la oral, y es efectivo a dosis bajas de 0.15 mg/kg
alcanzando su Cmax a las 4h y una concentración plasmática de
100ng/mL (Kitzman D et al., 2006). Dada la alta solubilidad lipídica de
ivermectina, ésta se distribuye ampliamente en el cuerpo y tiene una
afinidad a proteínas plasmáticas mayor a 90% (González Canga et al.,
2008; Klotz et al., 1990). El citocromo P-4503A4 es el encargado del
metabolismo de ivermectina en el hígado, convirtiéndolo en al menos 10
metabolitos, y el compuesto es excretado principalmente en las heces
(González Canga et al., 2008), siendo la vida media de absorción de
0.5-2.5h, mientras que su vida media en plasma es de
aproximadamente 18h (Chaccour et al., 2017).
Existen estudios que han demostrado que ivermectina tiene una alta
afinidad de unión a NS3, funcionando como inhibidor de la actividad
helicasa de la proteína NS3 de varios flavivirus, incluido DENV
(Mastrangelo et al., 2012). También se ha descrito que la ivermectina es
un inhibidor del transporte mediado por el heterodímero importina α/β
utilizado por DENV para una eficiente producción viral
(Bhuvanakantham et al., 2009) mediante el transporte nuclear de NS5
y NS3 (Wagstaff et al. 2012). En infecciones con DENV se ha
observado que en la replicación, la proteína NS5 se encuentra en el
núcleo en diferentes etapas del ciclo celular, haciendo notar que la
localización nuclear de NS5 es diferencial en los cuatro serotipos de
DENV, mientras que la proteína de DENV2 y DENV3 se encuentra
predominantemente en el núcleo, en los otros dos serotipos, NS3 se

17
encuentra fundamentalmente en el citoplasma (Hannemann et al.,
2012).
La proteína NS5 está compuesta por 900 a.a y tiene un peso molecular
de 105kDa. Es la más conservada de las proteínas virales entre los
cuatro serotipos de DENV con un 70% de identidad (El Sahili & Lescar
J., 2017, Pryor M et al., 2007) y tiene función de metiltransferasa y
deRNA polimerasa dependiente de RNA viral. Su transporte nuclear es
mediado por importina Impα/β1 que son capaces de unirse a NS5, a
partir de sus dos secuencias de localicación nuclear (NLS) la primera,
reconocida por IMPα/β y la segunda, una bNLS reconocida únicamente
por IMPβ (Fraser et al., 2014). Se ha observado que al mutar los
residuos de Impα/β1 que reconocen NLS, se ve afectada la
translocación a núcleo de NS5 y con ello la producción viral. Así, la
ivermectina, al poder inhibir la unión de la IMPa/b a su sustrato (Tay, et
al., 2013), puede ser usada para inhibir la infección por DENV (Wagstaff
et al. 2012).

18
Justificación
DENV es el arbovirus más importante a nivel mundial. El secuestro de
la maquinaria celular para la eficiente replicación del virus permite
identificar pasos críticos en el ciclo viral, entre los que se encuentra el
aumento en la síntesis de colesterol y ácidos grasos y el importe
nuclear de la proteína NS5. Debido a que no hay tratamiento específico
para DENVsepropuso evaluar de manera conjunta el efecto antiviral de
fármacos ya aprobados por FDA que inhibieran algunos de los pasos
críticos del ciclo replicativo de DENV. Los fármacos evaluados fueron
NDGA que inhibe la síntesis de lípidos, la vitamina D3 que regula la
repuesta inmune, las estatinas que son inhibidores de la síntesis del
colesterol y la Ivermectina que inhibe el transporte nuclear de la
proteína NS5. La evaluación del efecto antiviral se realizó en células
HuH7. De igual manera, se propuso evaluar el efecto sinérgico de la
combinación de ivermectina y lovastatina en la actividad antiviral contra
DENV.

19
Hipótesis
Los fármacos NDGA, lovastatina, ivermectina y vitamina D3,
disminuirán la infección por DENV2 y DENV4 en células HuH7. La
combinación ivermectina y lovastatina tendrá un efecto sinérgico en la
inhibición de la infección por DENV.
Objetivo general
Evaluar efecto antiviral de los fármacos NDGA, lovastatina, ivermectina,
vitamina D3 y de la combinación ivermectina-lovastatina ante infección
con DENV2 y DENV4 en células HuH7.
Objetivos particulares
Evaluar el efecto de los fármacos NDGA, lovastatina, ivermectina y
vitamina D3 en la infección de DENV2 y DENV4 en células HuH7.
Evaluar el efecto antiviral de la combinación ivermectina-lovastatina.

20
MATERIALES Y MÉTODOS
Células HuH7
La línea celular Huh7, proveniente de un carcinoma hepato celular se
obtuvo de un tumor de hígado de un paciente de 57 años de edad en
Japón. La línea celular fue establecida por Nakabayshi y Sato en 1982.
Las células se cultivaron en medio DMEM Advanced (Thermo Fisher
Scientific®) suplementado con 10% de suero fetal bovino (SBS), 2mM
de glutamina y 1% de penicilina/estreptomicina a 37ºC en una
atmósfera de 5% de CO2.
Stock viral DENV2 y DENV4
La propagación de los serotipos 2 y 4 de DENV se realizó a partir de la
inoculación en cerebro de ratón y los títulos virales se evaluaron a partir
de un ensayo de focos. Cerebros de ratones lactante no infectados se
usaron para extracción de extracto Mock.
Infección y tratamiento
Las células HuH7 se infectaron con DENV2 o DENV4 por dos horas a
una MOI de 3. A continuación, las células se lavaron con HBSS (1X) de
(Gibco® by life technologies TM) y se incubaron con los diferentes
fármacos por 24 y 48 horas.
Las concentraciones de los fármacos utilizados fueron: NDGA a 50 µM,
la lovastatina a 50 µM, la vitamina D3 a 10 µM y la ivermectina fue
utilizada primero como pretratamiento de 50 µM por 4 horas y posterior
a la infección se volvió a tratar con 20µM de acuerdo a los estudios de
viabilidad realizados previamente en el laboratorio.

21
Evaluación de la infección
Para evaluar el efecto de los distintos fármacos en la replicación viral se
determinó:
a) La cantidad de células infectadas mediante citometría de flujo.
b) La cantidad de proteína viral mediante Western blot.
c) La cantidad de virus producidos mediante ensayos de focos.
Citometría de flujo.
Para determinar el nivel de infección con las diferentes dosis de los
fármacos, se realizaron ensayos de citometría de flujo. Para ello, las
células previamente infectadas con DENV2 o DENV4 a una MOI de 3,
fueron despegadas con tripsina EDTA 0.1%, y posteriormente
centrifugadasa 800 rpm durante 8 min a 4ºC, para ser fijadas con
formaldehído al 1% por 20 minutos. Posteriormente, las células se
lavaron con PBS1X. Una vez lavadas, se les colocó solución
permeabilizadora (Saponina 0.2%, SBF 1% diluido en PBS1X), se
incubaron por 20 min a temperatura ambiente, se centrifugaron por 7
minutos a 2000 rpm y se inició el inmunomarcaje con el anticuerpo
primario anti prM a una dilución 1:100 incubando una hora a 37ºC.
Pasado el tiempo de incubación, las células se lavaron con PBS1X
para eliminar el resto de anticuerpo y se realizóla incubación con el
segundo anticuerpo secundario Alexa 488 a una dilución 1:200, por 1
hora a temperatura ambiente en oscuridad, y por último se realizaron
los lavados con PBS1X. Las células fueron suspendidas en PBS1X
para la lectura en el citómetro. Los datos fueron analizados mediante el
programa Cytobank software.

22
Western blot.
Las células HuH7 o U937 DC-SIGN fueron infectadas con DENV2 o
DENV4 y tratadas con los diferentes fármacos. A las 24 y 48 horas
post-infección las células fueron lisadas con buffer de lisis (1.5 mM
MgCl2, Tris– HCl pH 7.5 10 mM, 10 mM NaCl, 1% IGEPAL) y las
proteínas separadas mediante electroforesis en SDS-PAGE al 10%. Las
proteínas fueron transferidas a membranas de nitrocelulosa mediante
una cámara semiseca e incubadas con los anticuerpos anti NS3
(GENETEX) y anti-GAPDH como control de carga. Las bandas se
revelaron mediante quimioluminiscencia y se analizaron por
densitometría en el programa ImageJ.
Ensayo de focos.
La cantidad de partículas virales liberadas al medio se analizó después
de cada tratamiento, mediante ensayos de focos. Para ello, se
realizaron diluciones de los sobrenadantes recuperados y con ellos se
infectaron las células HuH7 sembradas en placas de 96 pozos a una
confluencia de 70%. La infección se mantuvo por 2 horas y
posteriormente se agregó medio DMEM Advanced. Las células fueron
incubadas por 24 horas, y terminando el tiempo de incubación se
procedió a fijar las células con formaldehído al 1% por 20 min. Una vez
fijadas, se lavaron con PBS1X y se colocó la solución permeabilizadora
por 20 min para después incubar con el anticuerpo primario anti prM a
una dilución de 1:100, incubando dos horas a temperatura ambiente.
Las células fueron lavadas con solución permeabilizadora para después
colocar el anticuerpo secundario Alexa 488 (1:200) por una hora. Las
células se lavaron PBS 1X. El conteo de focos se realizó mediante
microscopía de epifluorescencia.

23
Ensayos de viabilidad
Para analizar la viabilidad celular después del tratamiento con las
distintas dosis de la combinación de lovastatina con ivermectina se
realizaron ensayos de viabilidad de incorporación de yoduro de propidio
utilizando la técnica de citometría de flujo.
Análisis estadísticos
Los datos fueron analizados por una t test múltiple para analizar el
efecto de cada fármaco con respecto a su vehículo considerándose un
intervalo de confianza de 95%, además, se realizó una ANOVA de dos
vías con un test de comparación múltiple de Tukey, para verificar el
efecto antiviral entre cada tratamiento. El programa utilizado fue
GraphPad Software.
RESULTADOS
Efecto de los fármacos en el porcentaje de células infectadas
Las células Huh7 fueron infectadas con DENV2 y DENV4 y tratadas con
los fámacos en las dosis que se describen en la tabla 1.
Tabla 1 Estrategia experimental
De
manera
general,
las
células
fueron infectadas por dos horas y posteriormente fueron tratadas con
Tipo Celular Serotipo Fármacos Tiempo
HuH7 DENV2
DENV4
NDGA 50µM
Lovastatina 50 µM
Vitamina D3 10 µM
Ivermectina 50/20 µM
24 Horas
48 Horas

24
NDGA 50µM, lovastatina 50µM o vitamina D3 10µM. En el caso de
ivermectina se realizó un pretratamiento con 50µM del fármaco 4 horas
antes de la infección, y posterior a ésta se trató con 20 µM del fármaco.
El efecto del fármaco en la infección viral fue evaluado a las 24 y 48
horas.
Para analizar de qué manera afectaba cada tratamiento en el número
de células infectadas, se realizaron ensayos de citometría de flujo.
En DENV2, se observó que a 24h y 48h hubo una reducción
significativa en la cantidad de células infectadas y tratadas con los
fármacos con respecto a las células control incubadas solo con el
vehículo (Figura 4). Los niveles de inhibición fueron desde un 15% para
el caso de vitamina D3 hasta el 60% para ivermectina. La reducción en
la cantidad de células infectadas fue más evidente a las 48h en todos
los tratamientos, excepto para ivermectina donde el efecto es menor
(Figura 4). Al momento de comprar el efecto entre los fármacos,
ivermectina resultó ser el que disminuyó de manera más eficiente el
número de células infectadas (hasta 75% de inhibición).
Con respecto a DENV4, se observó un efecto similar al de DENV2 en
donde todos los tratamientos disminuyeron la cantidad de células
infectadas desde un 15% de inhibición (vitamina D3) hasta un 50%
(ivermectina). Al igual que con DENV2, el mayor efecto inhibitorio fue
con ivermectina (Figura 4).
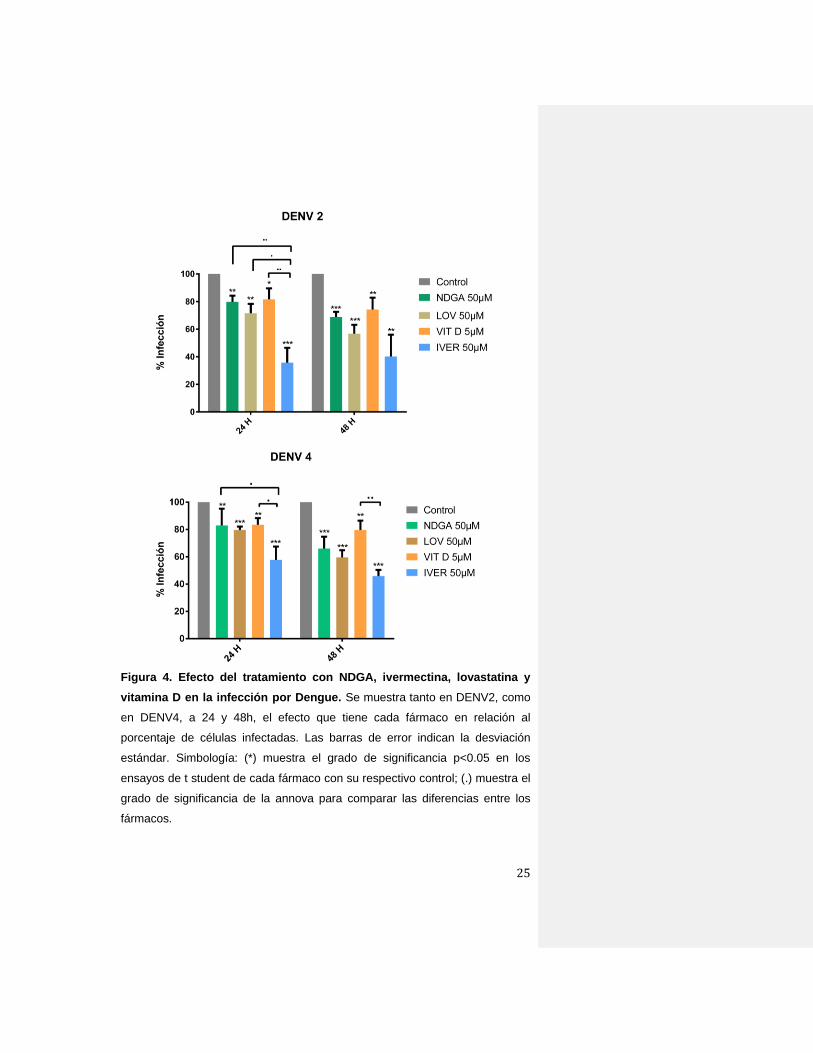
25
Figura 4. Efecto del tratamiento con NDGA, ivermectina, lovastatina y
vitamina D en la infección por Dengue. Se muestra tanto en DENV2, como
en DENV4, a 24 y 48h, el efecto que tiene cada fármaco en relación al
porcentaje de células infectadas. Las barras de error indican la desviación
estándar. Simbología: (*) muestra el grado de significancia p<0.05 en los
ensayos de t student de cada fármaco con su respectivo control; (.) muestra el
grado de significancia de la annova para comparar las diferencias entre los
fármacos.

26
Efecto de los fármacos en la cantidad de proteína viral
Otra manera de evaluar el efecto inhibitorio de la infección de los
distintos fármacos fue analizar la cantidad de proteína viral sintetizada
mediante ensayos de Western blot. A diferencia de lo observado en los
ensayos de citometría, en donde el efecto de los fármacos sobre la
inhibición de la infección se observó claramente, el método de Western-
blot fue menos sensible en detectar cambios en la cantidad de proteína
producida (Figura 5), sin embargo, para DENV2 fue evidente que hubo
una reducción significativa de la cantidad de la proteína viral después
del tratamiento con ivermectina a las 24 y 48 hpi. También pudimos
detectar una reducción en la cantidad de proteína viral en los
tratamientos con NDGA y lovastatina sin embargo no fueron
significativos a las 24 hpi. A las 48 hpi la inhibición en la cantidad de
proteína tanto con NDGA como con ivermectina fue significativa (Figura
5). Mediante esta metodología, no pudimos detectar una reducción
significativa de la cantidad de proteína viral después del tratamiento con
lovastatina ni con vitamina D3 (Figura 5).
Cuando se realizó la comparación entre tratamientos, se observó que a
24 H, ivermectina tiene un mejor efecto ante vitamina D3, mientras que
a 48 H Ivermectina tiene un mejor efecto ante vitamina D3 y lovastatina,
además también se muestra cómo NDGA tiene un mejor efecto antiviral
ante vitamina D3 (Figura 5).
En el caso de DENV4, se observó que a 24 horas sólo ivermectina
causó una reducción estadísticamente significativa en la cantidad de
proteína viral sintetizada (70% de reducción).Sin embargo, a las 48 hpi,
todos los fármacos probados, excepto vitamina D3, indujeron una
reducción significativa de la cantidad de proteína viral (40% de
inhibición para NDGA, 55% para lovastatina y 90% para ivermectina).
Como ocurrió con DENV2, el mayor efecto inhibitorio de la infección fue

27
causado por el tratamiento con ivermectina (de hasta 90% de inhibición
a las 48 hpi) (Figura 6).
Efecto de los fármacos en la producción de progenie viral.
Si bien los distintos tratamientos tenían un efecto inhibitorio en la
cantidad de células infectadas y en la cantidad de proteína viral
producida, decidimos evaluar el efecto de los fármacos en la cantidad
de virus producidos. La reducción de virus liberados de las células
infectadas y tratadas nos permite evaluar de manera directa la manera
en que los fármacos podrían reducir la diseminación viral, el cual es un
parámetro muy importante en la evaluación de antivirales. La cantidad
de virus liberado fue evaluado mediante ensayo de focos.

28
Figura 5. Evaluación de la expresión de la proteína viral NS3 en células
tratadas con NDGA, lovastatina, Vitamina D3 e ivermectina e infectadas
con DENV2. Las células Huh7 fueron infectadas con DENV2 y tratadas con
NDGA, LOV, VIT D o IVER por 24 o 48 h. La cantidad de la proteína NS3 fue
evaluada mediante Western-blot. Como control de carga se usó la proteína
GAPDH. Se muestra un Western blot representativo y el análisis
densitométrico de tres experimentos independientes realizados por duplicado.
Las barras de error indican la desviación estándar. Simbología: (*) muestra el
grado de significancia p<0.05 en los ensayos de t student de cada fármaco
con su respectivo control; (.) muestra el grado de significancia de la annova
para comparar las diferencias entre los fármacos.

29
Figura 6. Evaluación de la expresión de la proteína viral NS3 en células
HuH7 infectadas con DENV4 y tratadas con NDGA, lovastatina, vitamina
D3 e ivermectina. Las células Huh7 fueron infectadas con DENV4 y tratadas
con NDGA, LOV, VIT D o IVER por 24 o 48 h. La cantidad de la proteína NS3
fue evaluada mediante Western-blot. Como control de carga se usó la proteína
GAPDH. Se muestra un Western blot representativo y el análisis
densitométrico de tres experimentos independientes realizados por duplicado.
Las barras de error indican la desviación estándar. Simbología: (*) muestra el
grado de significancia p<0.05 en los ensayos de t student de cada fármaco
con su respectivo control; (.) muestra el grado de significancia de la annova
para comparar las diferencias entre los fármacos.
Con formato: Fuente: (Predeterminada) +Cuerpo
(Cambria), 12 pto, Sin Cursiva
Con formato: Normal, Izquierda, Interlineado: sencillo

30
Efecto de los fármacos en la producción de progenie viral.
Si bien los distintos tratamientos tenían un efecto inhibitorio en la
cantidad de células infectadas y en la cantidad de proteína viral
producida, decidimos evaluar el efecto de los fármacos en la cantidad
de virus producidos. La reducción de virus liberados de las células
infectadas y tratadas nos permite evaluar de manera directa la manera
en que los fármacos podrían reducir la diseminación viral, el cual es un
parámetro muy importante en la evaluación de antivirales. La cantidad
de virus liberado fue evaluado mediante ensayo de focos.
Con DENV2, se observó que todos los fármacos a 24 y 48 horas
afectaban el número de partículas virales liberadas (desde medio
logaritmo hasta tres logaritmos y medio), excepto vitamina D3 a las 48
horas donde no se observó diferencia con el vehículo (Figura 7).
Cuando se realizó el análisis comparativo entre tratamientos, se
observó que a 24 horas todos los tratamientos tienen un mayor efecto
antiviral y como en ensayos anteriores, el mayor efecto inhibitorio se
observó con ivermectina (Figura 7A).
En cuanto DENV4, a las 24 horas solo se mostró diferencia significativa
en reducción de virus producido con lovastatina e ivermectiva (un
logaritmo y medio y dos logaritmos y medio de inhibición
respectivamente). Sin embargo, a las 48 horas se observó una
diferencia significativa en el virus producido también con NDGA (un
logaritmo de inhibición). El tratamiento con vitamina D3 no indujo una
reducción de la cantidad de virus liberado (Figura 7B). Esto contrasta
con la inhibición observada con DENV2 (Figura 7A).
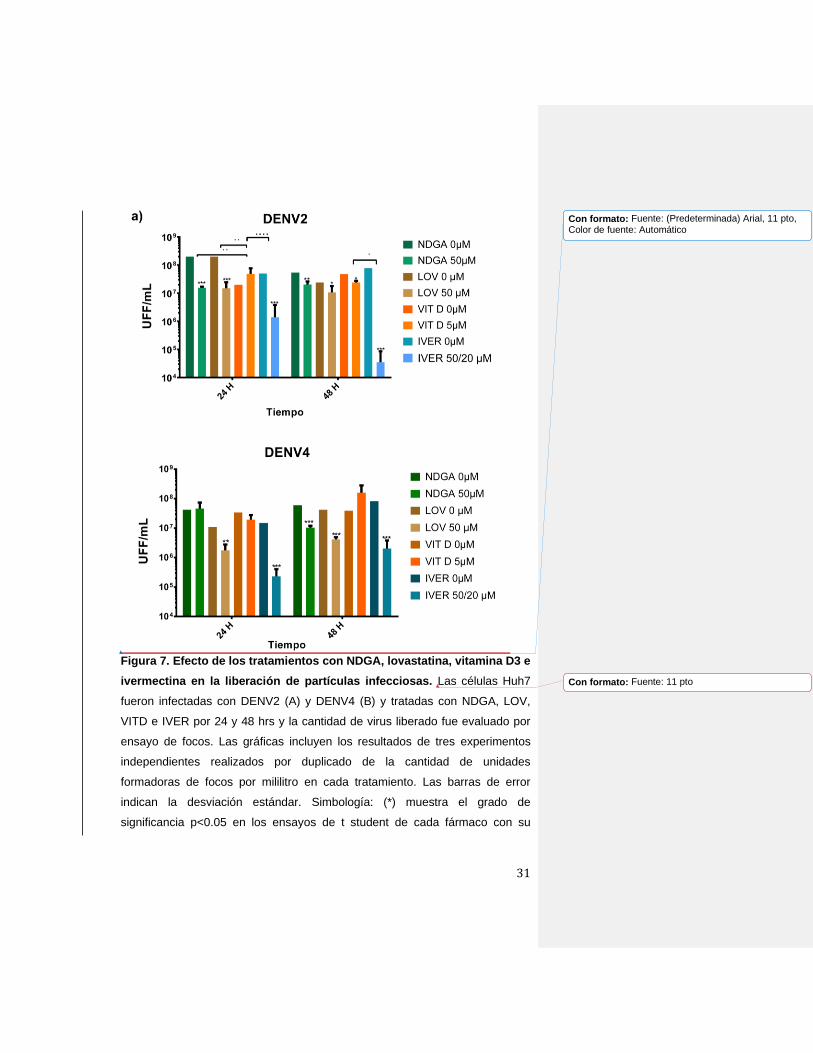
31
Figura 7. Efecto de los tratamientos con NDGA, lovastatina, vitamina D3 e
ivermectina en la liberación de partículas infecciosas. Las células Huh7
fueron infectadas con DENV2 (A) y DENV4 (B) y tratadas con NDGA, LOV,
VITD e IVER por 24 y 48 hrs y la cantidad de virus liberado fue evaluado por
ensayo de focos. Las gráficas incluyen los resultados de tres experimentos
independientes realizados por duplicado de la cantidad de unidades
formadoras de focos por mililitro en cada tratamiento. Las barras de error
indican la desviación estándar. Simbología: (*) muestra el grado de
significancia p<0.05 en los ensayos de t student de cada fármaco con su
Con formato: Fuente: (Predeterminada) Arial, 11 pto,
Color de fuente: Automático
Con formato: Fuente: 11 pto

32
respectivo control; (.) muestra el grado de significancia de la ANNOVA p<0.05
para comparar las diferencias entre los fármacos.
Viabilidad Ivermectina-Lovastatina.
Una vez evaluado el efecto independientemente de cada uno de los
fármacos, se decidió evaluar el efecto de algunos de ellos en
combinación. La vitamina D3 no fue contemplada pues fue el fármaco
con el menor efecto antiviral. Por otro lado, debido a que NDGA y
lovastatina actúan en la misma vía de reducción de colesterol, se
seleccionó solo uno de ellos que fue la lovastatina. Este fármaco se
combinó con ivermectina que actúa en la entrada de NS5 al núcleo, una
ruta diferente de inhibición de la infección por DENV.
Inicialmente, determinamos la viabilidad de las células con la
combinación de fármacos: 1) ivermectina 5µM-lovastatina 5µM, 2)
ivermectina 10µM-lovastatina 30µM, 3) ivermectina 10µM-lovastatina
40µM, y 4) ivermectina 15µM-lovastatina 35µM, y ninguna de ellas
afectó la viabilidad celular (Figura 8).
Tratamiento de ivermectina-lovastatina en células infectadas con DENV
Una vez corroborado que la viabilidad celular no era afectada con las
combinaciones de fármacos, evaluamos su efecto, para lo cual se
eligieron las dosis de ivermectina 10µM-lovastatina 40µM e ivermectina
15µM-lovastatina 35µM, ya que son las dosis a las cuáles cada fármaco
aún podría estar realizando su función antiviral. Para esto, las células
fueron infectadas con DENV2 o DENV4 y posteriormente fueron
tratadas con las dos diferentes dosis por 24-48 h. El medio se retiró y
en él se evaluó la cantidad de virus liberado por ensayo de focos.
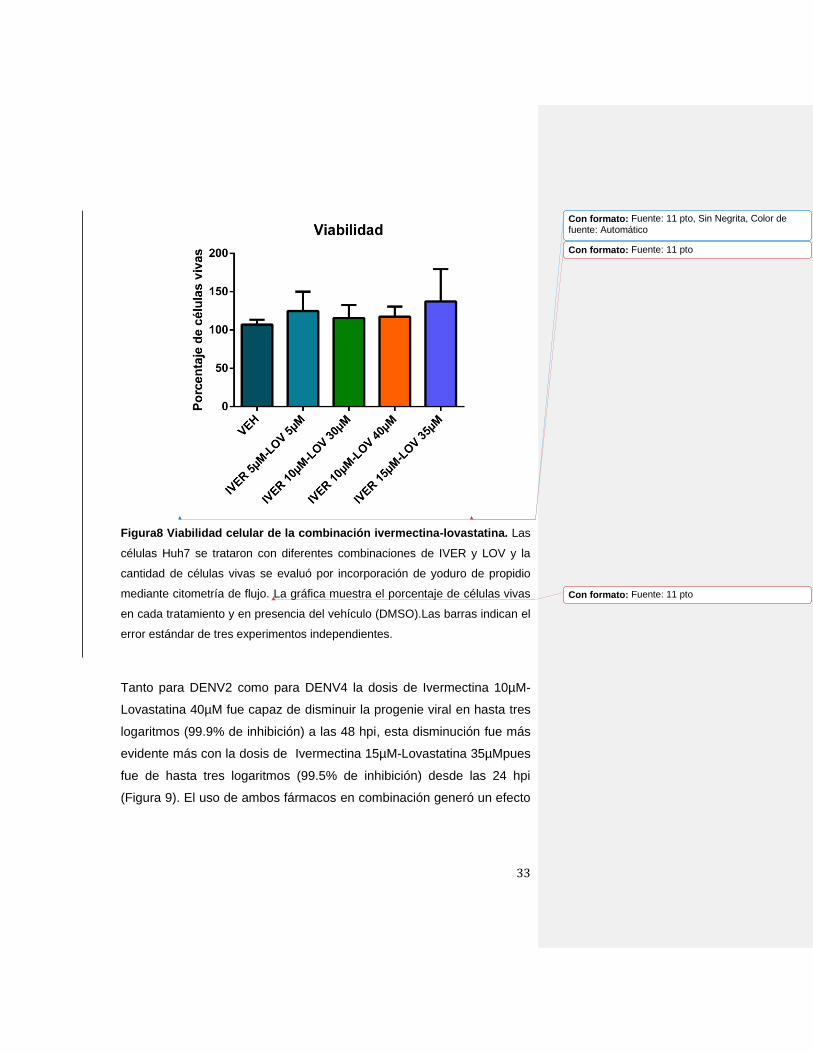
33
Figura8 Viabilidad celular de la combinación ivermectina-lovastatina. Las
células Huh7 se trataron con diferentes combinaciones de IVER y LOV y la
cantidad de células vivas se evaluó por incorporación de yoduro de propidio
mediante citometría de flujo. La gráfica muestra el porcentaje de células vivas
en cada tratamiento y en presencia del vehículo (DMSO).Las barras indican el
error estándar de tres experimentos independientes.
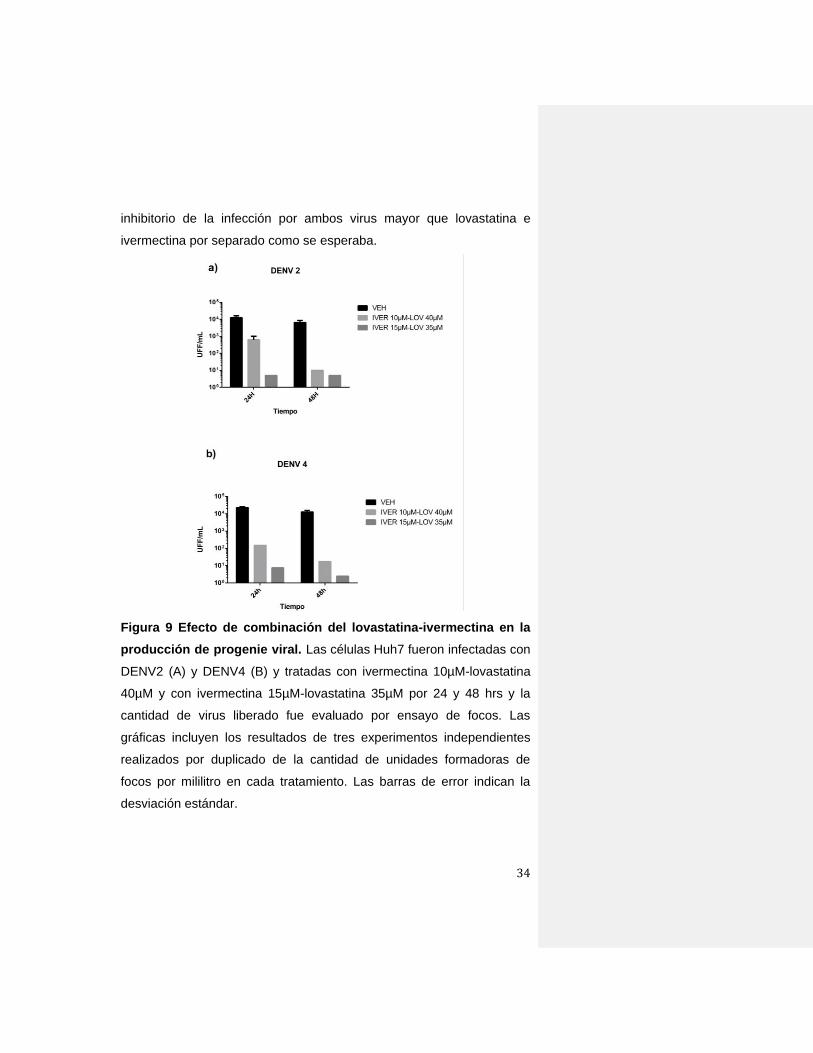
Tanto para DENV2 como para DENV4 la dosis de Ivermectina 10µM-
Lovastatina 40µM fue capaz de disminuir la progenie viral en hasta tres
logaritmos (99.9% de inhibición) a las 48 hpi, esta disminución fue más
evidente más con la dosis de Ivermectina 15µM-Lovastatina 35µMpues
fue de hasta tres logaritmos (99.5% de inhibición) desde las 24 hpi
(Figura 9). El uso de ambos fármacos en combinación generó un efecto
Con formato: Fuente: 11 pto, Sin Negrita, Color de
fuente: Automático
Con formato: Fuente: 11 pto
Con formato: Fuente: 11 pto
34
inhibitorio de la infección por ambos virus mayor que lovastatina e
ivermectina por separado como se esperaba.
Figura 9 Efecto de combinación del lovastatina-ivermectina en la
producción de progenie viral. Las células Huh7 fueron infectadas con
DENV2 (A) y DENV4 (B) y tratadas con ivermectina 10µM-lovastatina
40µM y con ivermectina 15µM-lovastatina 35µM por 24 y 48 hrs y la
cantidad de virus liberado fue evaluado por ensayo de focos. Las
gráficas incluyen los resultados de tres experimentos independientes
realizados por duplicado de la cantidad de unidades formadoras de
focos por mililitro en cada tratamiento. Las barras de error indican la
desviación estándar.

35
DISCUSIÓN
Existen diversos estudios en los que se ha evaluado la actividad anti-
DENV de diferentes fármacos. Esto se ha logrado empleando diferentes
ensayos, serotipos virales y a diferentes multiplicidades de infección
(MOI). En este estudio quisimos evaluar el efecto de algunos de los
fármacos ya caracterizados usando dos serotipos virales DENV2 y
DENV4 en las mismas condiciones experimentales. Decidimos evaluar
la capacidad inhibitoria de la infección de los fármacos empleando tres
parámetros. Por un lado, analizamos i) la cantidad de células infectadas
mediante citometría de flujo, ii) la eficiencia de replicación viral mediante
la cuantificación de la proteína viral producida mediante Western-blot y
iii) la cantidad de virus producido mediante ensayo de focos. Los dos
ensayos que mostraron mayor sensibilidad para poder evaluar el efecto
inhibitorio de los fármacos fueron la citometría de flujo y el ensayo de
focos. El Western-blot fue poco sensible para poder seguir la inhibición
de la infección. Sin embargo de manera global consideramos que la
cuantificación de virus liberado mediante ensayo de focos es un
excelente parámetro para evaluar el efecto de un fármaco en la
diseminación viral.
Los fármacos estudiados en este trabajo actúan en tres vías diferentes:
i) síntesis de lípidos y colesterol (NDGA y lovastatina), ii) respuesta
inmune (vitamina D3) y iii) transporte citoplasma-núcleo. Si bien, los
fármacos que actúan a nivel de síntesis de lípidos fueron capaces de
inhibir la infección, medida por los tres métodos diferentes, con ambos
serotipos de DENV, el fármaco con mejor actividad anti-DENV fue
ivermectina. Estos resultados son consistentes con los estudios
realizados por Tay y colaboradores (2013) donde se observa que
ivermectina disminuye la infección con DENV. Si bien la replicación de

36
DENV y de los flavivirus en general es citoplasmática, la presencia de
proteínas virales en el núcleo parece ser indispensable (Pryor et al.,
2007) así como la relocalización de proteínas celulares nucleares en el
citoplasma de las células infectadas (Wagstaff et al., 2012). Resultados
de nuestro laboratorio sugieren que el transporte núcleo-citoplasma
está siendo alterado por la infección por DENV y ZIKV. Esto abre la
posibilidad de que esta vía pueda ser usada como un blanco contra la
infección por flavivirus. En el caso específico de ivermectina, se ha
demostrado que éste fármaco tiene también actividad anti-ZIKV en
células en cultivo al igual que en cultivos primarios (Barrows et al.,
2016). Este hecho apoya la actividad anti-flavivirus de ivermectina.
A pesar de lo esperado, nuestros experimentos fueron consistentes con
el hecho de que vitamina D3 tuvo un pobre efecto anti-DENV. Estudios
previos realizados en el laboratorio, habían mostrado el efecto antiviral
en células HuH7 por citometría de flujo (Puerta Guardo et al., 2012).
Estos datos fueron consistentes con nuestros resultados en donde se
observó una reducción de la cantidad de células infectadas por
citometría de flujo, sin embargo, ésta reducción no se reflejó en una
inhibición significativa de la cantidad de proteína viral y de progenie
viral. El efecto inhibitorio más importante generado por vitamina D3 fue
reportado en células de sistema inmune como los monocitos U937 DC-
SIGN (Puerta Guardo et al., 2012). Este hecho es consistente con el
hecho de que muchas de las funciones que tiene vitamina D3 son
principalmente inmuno-reguladoras, realizadas en el caso de DENV por
macrófagos y células dendríticas. Existen diversos reportes que indican
que Vitamina D3 es capaz de inducir autofagia (Chun R et al., 2014). Si
bien la autofagia en células diferentes a macrófagos puede potenciar la
infección por DENV (Guzman MG & Harris E et al., 2015), la autofagia
inhibe la infección en macrófagos (Puerta-Guardo et al., 2012). Se ha

37
descrito que en células diferentes a macrófagos, DENV es capaz de
inducir autofagia con el fin de incrementar la degradación de las gotas
lipídicas (Heaton & Randal., 2011) y así aprovechar los ácidos grasos
en la remodelación del retículo endoplásmico, lugar donde se realiza la
replicación viral la inducción de éste proceso en macrófagos parece
actuar de manera contraria (Green et al., 2014).
Con base en nuestros resultados decidimos evaluar si el uso de una
combinación de dos fármacos podría darnos mejores resultados. Para
ello, decidimos emplear lovastatina e ivermectina. La elección se realizó
con base en el hecho de que las estatinas al igual que el NDGA tienen
como blanco los lípidos y el colesterol pero son menos tóxicas y mejor
aceptadas en el uso en humanos y que ivermectina había tenido el
mejor efecto inhibitorio. Usando la combinación logramos tener una
inhibición mayor que con cualquiera de los fármacos por separado y en
24 hpi (99.9% de inhibición). De la misma manera, no se requería de un
pretratamiento como se realizó con ivermectina sola. Uno de los
próximos pasos será probar la combinación en cultivos primarios, que
constituyen modelos más sensibles a los fármacos, además de
probarse contra otros flavivirus como ZIKV, YFV o WNV. El empleo de
un modelo animal sería un segundo paso en el proceso y podría
también dar pistas importantes de la relevancia del tratamiento
combinado en la actividad anti-flavivirus y de su posible aplicación en
humanos.

38
CONCLUSIONES
Ivermectina es el fármaco con el mejor efecto antiviral.
La combinación ivermectina-lovastatina puede constituir una estrategia
antiviral efectiva para DENV y tal vez para otros flavivirus.
PERSPECTIVAS
• Probar otras posibles combinaciones entre los fármacos
• Evaluar si a las concentraciones utilizadas en la combinación
ivermectina-lovastatina, los fármacos siguen teniendo el mismo
efecto propuesto.
• Emplear la combinación en cultivos primarios.
• Evaluar el efecto de los fármacos en la infección de células U937
DC-SIGN y en los niveles de citocinas proinflamatorias
producidas durante infección.
• Evaluar el efecto de los fármacos y sus posibles combinaciones
en otros flavivirus como ZIKV, YFV y WNV.

39
REFERENCIAS
• Acosta EG & Bartenschlager R (2016) The quest for host targets
to combat dengue virus infections. Current Opinion in Virology,
20:47–54
• Ahmed S, Finkelstein JL, Stewart AM, Kenneth J, Polhemus ME,
Endy TP, Cardenas W, Mehta S. (2009) Clinical response in
patients with dengue fever to oral calcium plus vitamin D
administration: study of 5 cases. Proc West Pharmacol
Soc;52:14-7.
• Arboleda JF, Urcuqui-Inchima S (2016) Vitamin D-Regulated
MicroRNAs: Are They Protective Factors against Dengue Virus
Infection? Adv Virol;2016:1016840. doi: 10.1155/2016/1016840.
• Arteaga S, Andrade-Cetto A, Cardenas R. (2005) Larrea
tridentata (Creosote bush), an abundant plant of Mexican and
US-American deserts and its metabolite nordihydroguaiaretic
acid. J Ethnopharmacol 2005;98:231–9. [PubMed: 15814253]
• Balsameda A, et al., (2006) Serotype-Specific Differences In
Clinical Manifestations Of Dengue. Am. J. Trop. Med. Hyg., 74(3),
2006, pp. 449–456
• Beard, J. A., Bearden, A., & Striker, R. (2011). Vitamin D and the
anti-viral state. Journal Of Clinical Virology: The Official
Publication Of The Pan American Society For Clinical Virology,
50(3), 194-200. doi:10.1016/j.jcv.2010.12.006

40
• Bhuvanakantham R, Chong MK, Ng ML (2009) Specific
interaction of capsid protein and importin-a/b influences West Nile
virus production. Biochemical and Biophysical Research
Communications 389 (2009) 63–69
• Billinsky JL, Marcoux MR, Krol ES. (2007) Oxidation of the lignan
nordihydroguaiaretic acid. Chem Res Toxicol 20: 1352–1358.
• Borella, E., Nesher, G. Israeli, E. & Shoenfeld, Y. (2014). Vitamin
D: A new anti-infective agent?. Blackwell Publishing Inc.
doi:10.1111/nyas.12321
• Chaccour, C., Hammann, F., & Rabinovich, N. R. (2017).
Ivermectin to reduce malaria transmission I. Pharmacokinetic and
pharmacodynamic considerations regarding efficacy and safety.
Malaria Journal, 16, 161. http://doi.org/10.1186/s12936-017-
1801-4
• Christakos, S., Dhawan, P., Verstuyf, A., Verlinden, L., &
Carmeliet, G. (2016). Vitamin D: Metabolism, Molecular
Mechanism of Action, and Pleiotropic Effects. Physiological
Reviews, 96(1), 365-408. doi:10.1152/physrev.00014.2015
• Chun R, Liu P, Modlin R, Adams J, Hewison M (2014) Impact of
vitamin D on immune function: Lessons learned from genome-
wide analysis. Frontiers in Physiology vol: 5
• Clyde K, Kyle J & Harria E. (2006) Recent Advances in
Deciphering Viral and Host Determinants of Dengue Virus
Replication and Pathogenesis. JOURNAL OF VIROLOGY, Dec.
2006, p. 11418–11431
• Damonte EB, Pujol CA & Coto CE (2004) Prospects for the
therapy and prevention of dengue virus infections. ADVANCES
IN VIRUS RESEARCH, VOL 63

41
• Demetz, E., Schroll, A., Auer, K., Heim, C., Patsch, J., Eller, P., &
... Tancevski, I. (2014). Article: The Arachidonic Acid Metabolome
Serves as a Conserved Regulator of Cholesterol Metabolism.
Cell Metabolism, 20787-798. doi:10.1016/j.cmet.2014.09.004
• Di Rosa M, Malaguarnera M, Nicoletti F & Malaguarnera L (2011)
Vitamin D3: a helpful immuno-modulator. Immunology, 134, 123–
139
• El Sahili, A., & Lescar, J. (2017). Dengue Virus Non-Structural
Protein 5. Viruses, 9(4), 91. http://doi.org/10.3390/v9040091
• Fraser JE, Rawlinson S, Wang C, Jans DA, Wagstaff (2014)
Investigating Dengue Virus Nonstructural Protein 5 (NS5) Nuclear
Import Radhakrishnan Padmanabhan and Subhash G.
Vasudevan (eds.), Dengue: Methods and Protocols, Methods in
Molecular Biology, vol. 1138, DOI 10.1007/978-1-4939-0348-
1_19, © Springer Science+Business Media, LLC 2014
• Gazzerro, P., Proto, M. C., Gangemi, G., Malfitano, A. M.,
Ciaglia, E., Pisanti, S., & ... Bifulco, M. (2012). Pharmacological
actions of statins: a critical appraisal in the management of
cancer. Pharmacological Reviews, 64(1), 102-146.
doi:10.1124/pr.111.004994
• González Canga, A., Sahagún Prieto, A. M., Diez Liébana, M. J.,
Fernández Martínez, N., Sierra Vega, M., & García Vieitez, J. J.
(2008). The Pharmacokinetics and Interactions of Ivermectin in
Humans—A Mini-review. The AAPS Journal, 10(1), 42–46.
http://doi.org/10.1208/s12248-007-9000-9
• Green A, Beatty P, Hadjilaou A, Harris E (2014) Innate Immunity
to Dengue Virus Infection and Subversion of Antiviral Responses.
J Mol Biol vol: 426 (6) pp: 1148-1160

42
• Greenwood J & Mason JC (2007) Statins and the vascular
endothelial inflammatory response. TRENDS in Immunology
Vol.28 No.2
• Grice HC, Becking G, Goodman T. (1968) Toxic properties of
nordihydroguaiaretic acid. Food Cosmet Toxicol 6: 155–161
• Gubler D, Ooi E, Vasudevan S & Farrar J. (2014). Dengue and
Dengue Hemorrhagic Fever. UK:CABI
• Guzman MG & Harris E (2015) Dengue Lancet 2015; 385: 453–
65
• Hannemann H1, Sung PY, Chiu HC, Yousuf A, Bird J, Lim SP,
Davidson AD. (2013) Serotype-specific Differences in Dengue
Virus Non-structural Protein 5 Nuclear Localization. The Journal
of Biological Chemestry VOL.288, NO.31 pp.22621–22635,
• Heaton, N. S., & Randall, G. (2011). Dengue Virus and
Autophagy. Viruses, 3(8),1332–1341.
• Hwu JR, Hsu MH, Huang RC. (2008) New nordihydroguaiaretic
acid derivatives as anti-HIV agents. Bioorg Med Chem Lett
2008;18:1884–8. [PubMed: 18321703
• Jeong, H., Kim, S., Lee, J., Park, J. Y., Zhou, W., Liu, X., Kim, S.
D., Song, Y. S., Jang, C. -Y., Oh, S. -R., Choi, S., and Chang, M.
(2017) Characterization of Phase I and Phase II Hepatic
Metabolism and Reactive Intermediates of Larrea nitida Cav. and
Its Lignan Compounds. Phytother. Res., 31: 140–151. doi:
10.1002/ptr.5742.
• Jetter A, Egli A, Dawson-Hughes B, Staehelin H, Stoecklin E
Goessl R, Henschkowski J, Bischoff-Ferrari H (2014)
Pharmacokinetics of oral vitamin D3 and calcifediol. Bone vol: 59
pp: 14-19

43
• Kitzman D, Wei SY, Fleckenstein L. Liquid chromatographic
assay of ivermectin in human plasma for application to clinical
pharmacokinetic studies. J Pharmaceut Biomed. 2006; 40(4):
1013–1020.
• Klotz U, J. E. Ogbuokiri, and P. O. Okonkwo. Ivermectin binds
avidly to plasma proteins. Eur. J. Clin. Pharmacol 39:607–608
(1990).
• Lambert JD, Zhao D, Meyers RO, Kuester RK, Timmermann BN,
Dorr RT. (2002). Nordihydroguaiaretic acid: hepatotoxicity and
detoxification in the mouse. Toxicon 40: 1701–1708.
• Lambert JD1, Meyers RO, Timmermann BN, Dorr RT. (2001)
Pharmacokinetic analysis by high-performance liquid
chromatography of intravenousnordihydroguaiaretic acid in the
mouse. J Chromatogr B Biomed Sci Appl. 2001 Apr 15;754(1):85-
90.
• Lee M Kim D Jo K Hwang J (2010) Nordihydroguaiaretic acid
protects against high-fat diet-induced fatty liver by activating
AMP-activated protein kinase in obese mice. Biochemical and
Biophysical Research Communications vol: 401 (1) pp: 92-97
• Lü, J.-M., Nurko, J., Weakley, S. M., Jiang, J., Kougias, P., Lin, P.
H., … Chen, C. (2010). Molecular mechanisms and clinical
applications of nordihydroguaiaretic acid (NDGA) and its
derivatives: An update. Medical Science Monitor : International
Medical Journal of Experimental and Clinical Research, 16(5),
RA93–R100.
• Malhotra, H. S., & Goa, K. L. (2001). Atorvastatin: an updated
review of its pharmacological properties and use in dyslipidaemia.
Drugs, 61(12), 1835-1881.
Con formato: Fuente: 12 pto, Sin Negrita, Color de
fuente: Automático, Inglés (Estados Unidos)
Con formato: Inglés (Estados Unidos)

44
• Martinez-Gutierrez M, Castellanos JE, Gallego-Gomez JC.
Statins reduce dengue virus production via decreased virion
assembly. Intervirology 2011; 54:202–16.
• Mastrangelo, E., Pezzullo, M., De Burghgraeve, T., Kaptein, S.,
Pastorino, B., Dallmeier, K., & ... Milani, M. (2012). Ivermectin is a
potent inhibitor of flavivirus replication specifically targeting NS3
helicase activity: new prospects for an old drug. The Journal Of
Antimicrobial Chemotherapy, 67(8), 1884-1894.
doi:10.1093/jac/dks147
• Mukhopadhyay, S., Kuhn, R. J., & Rossmann, M. G. (2005). A
structural perspective of the flavivirus life cycle. Nature Reviews.
Microbiology, 3(1), 13-22.
• Nicholas J. Barrows, Rafael K. Campos, Steven T. Powell,
K. Reddisiva Prasanth, Geraldine Schott-Lerner, Ruben Soto-
Acosta, Gaddiel Galarza-Muñoz, Erica L. McGrath, Rheanna
Urrabaz-Garza, Junling Gao, Ping Wu, Ramkumar Menon,
George Saade, Ildefonso Fernandez-Salas, Shannan L. Rossi,
Nikos Vasilakis, Andrew Routh, Shelton S. Bradrick, Mariano A.
Garcia-Blanco (2016) A Screen of FDA-Approved Drugs for
Inhibitors of Zika Virus InfectionCell Host & Microbe, Volume 20,
Issue 2, 10 August 2016, Pages 259-270
• Omura, S., 2008. Ivermectin: 25 years and still going strong. Int.
J. Antimicrob Agents 31 (2), 91–98.
• Peña J Harris E (2012) Early dengue virus protein synthesis
induces extensive rearrangement of the endoplasmic reticulum
independent of the UPR and SREBP-2 pathway. PLoS ONE
2012 vol: 7 (6)

45
• Pryor, M. J., Rawlinson, S. M., Butcher, R. E., Barton, C. L.,
Waterhouse, T. A., Vasudevan, S. G., Bardin, P. G., Wright, P. J.,
Jans, D. A. and Davidson, A. D. (2007), Nuclear Localization of
Dengue Virus Nonstructural Protein 5 Through Its Importin α/β–
Recognized Nuclear Localization Sequences is Integral to Viral
Infection. Traffic, 8: 795–807. doi:10.1111/j.1600-
0854.2007.00579.x
• Puerta-Guardo, H., De la Cruz Hernández, S. I., Rosales, V. H.,
Ludert, J. E., & del Angel, R. M. (2012). The 1α,25-dihydroxy-
vitamin D3 reduces dengue virus infection in human
myelomonocyte (U937) and hepatic (Huh-7) cell lines and
cytokine production in the infected monocytes. Antiviral
Research, 94(1), 57-61. doi:10.1016/j.antiviral.2012.02.006
• Reyes-Del Valle, J., Chávez-Salinas, S., Medina, F., & Del Angel,
R. (2005). Heat shock protein 90 and heat shock protein 70 are
components of dengue virus receptor complex in human cells.
Journal Of Virology, 79(8), 4557-4567.
doi:10.1128/JVI.79.8.4557-4567.2005
• Rothwell C, Lebreton A, Young Ng C, et al. Cholesterol
biosynthesis modulation regulates dengue viral replication.
Virology 2009; 389:8–19.
• Samsa, M.M., Mondotte, J.A., Iglesias, N.G., Assunção-Miranda,
I., Barbosa-Lima, G., Da Poian, A.T., Bozza, P.T., Gamarnik,
A.V., 2009. Dengue virus capsid protein usurps lipid droplets for
viral particle formation. PLoS Pathog. 5, e1000632.
• Soto-Acosta R, Bautista-Carbajal P, Cervantes-Salazar M, Angel-
Ambrocio AH, del Angel RM (2017) DENV up-regulates the
HMG- CoA reductase activity through the impairment of AMPK
Con formato: Fuente: 12 pto, Sin Negrita, Color de
fuente: Automático, Inglés (Estados Unidos)
Con formato: Inglés (Estados Unidos)
Con formato: Fuente: 12 pto, Sin Negrita, Color de
fuente: Automático, Inglés (Estados Unidos)

46
phosphorylation: A potential antiviral target. PLoS Pathog 13(4):
e1006257. doi:10.1371/ journal.ppat.
• Soto-Acosta, R., Bautista-Carbajal, P., Syed, G. H., Siddiqui, A.,
& Del Angel, R. M. (2014) Nordihydroguaiaretic acid (NDGA)
inhibits replication and viral morphogenesis of dengue virus.
Antiviral Research, 109132-140.
doi:10.1016/j.antiviral.2014.07.002
• Soto-Acosta, R., Mosso, C., Cervantes-Salazar, M., Puerta-
Guardo, H., Medina, F., Favari, L., & ... del Angel, R. M. (2013).
The increase in cholesterol levels at early stages after dengue
virus infection correlates with an augment in LDL particle uptake
and HMG-CoA reductase activity. Virology, 442(2), 132-147.
doi:10.1016/j.virol.2013.04.003
• Syed G Siddiqui A (2011) Effects of hypolipidemic agent
nordihydroguaiaretic acid on lipid droplets and hepatitis C virus.
Hepatology 2011 vol: 54 (6) pp: 1936-1946
• Tay M, Fraser J, Chan W, Moreland N, Rathore A, Wang C,
Vasudevan S, Jans D. (2013) Nuclear localization of dengue
virus (DENV) 1-4 non-structural protein 5; protection against all 4
DENV serotypes by the inhibitor Ivermectin. Antiviral Research
2013 vol: 99 (3) pp: 301-306
• Villareal, V. A., Rodgers, M. A., Costello, D. A., & Yang, P. L.
(2015). Targeting host lipid synthesis and metabolism to inhibit
dengue and hepatitis C viruses. Antiviral Research, 110.
doi:10.1016/j.antiviral.2015.10.013
• Wagner, A.H. et al. (2000) Improvement of nitric oxide-dependent
vasodilatation by HMG-CoA reductase inhibitors through
attenuation of endothelial superoxide anion formation.

47
Arterioscler. Thromb. Vasc. Biol. 20, 61–69
• Wagstaff, K., Heaton, S, Jans, D., Sivakumaran, H., & Harrich, D.
(2012). Ivermectin is a specific inhibitor of importin α/β-mediated
nuclear import able to inhibit replication of HIV-1 and dengue
virus. Biochemical Journal, 443(3), 851-856.
doi:10.1042/BJ20120150
• Whitehorn, J., Lam Phung, K., Duong Thi Hue, K., Nguyen Than
Ha, Q., Nguyen Thi Thanh, T., Chau Van Vinh, N., & ... Wills, B.
(2016). Lovastatin for the treatment of adult patients with dengue:
a randomized, double-blind, placebo-controlled trial. Clinical
Infectious Diseases, (4), 468.
Con formato: Fuente: 12 pto, Sin Negrita, Color de
fuente: Automático, Inglés (Estados Unidos)
Con formato: Inglés (Estados Unidos)




















