
Central European Journal of Biology Enhanced production of valerenic acid in hairy root culture of...
11
Central European Journal of Biology * E-mail: [email protected] Research Article 1 Department of Biology, Faculty of Science, University of Urmia, P.O. Box 165, Urmia, Iran 2 Department of Plant Breeding and Biotechnology, Faculty of Agriculture, University of Urmia, P.O. Box 165, Urmia, Iran 3 Department of Agricultural Biotechnology, Institute of Biotechnology, University of Urmia, P.O. Box 165, Urmia, Iran 4 Department of Plant Pathology, Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran Mohammad Reza Dini Torkamani 1 , Morad Jafari 2,3 *, Nasser Abbaspour 1 , Reza Heidary 1 , Naser Safaie 4 Enhanced production of valerenic acid in hairy root culture of Valeriana officinalis by elicitation 1. Introduction Valerian (Valeriana officinalis L.), a perennial herb from the Valerianaceae family, is native to Europe and Asia. It has been used as a medicinal herb worldwide [1,2]. Pharmaceutical application of valerian is due to its sedative, anticonvulsant, hypnotic effects, and anxiolytic activity. Lately, the potential cytoprotective effect of an aqueous extract of valerian on human neuroblastoma cells has also been demonstrated [3]. The compounds accumulated mainly in the roots and rhizomes of valerian include valepotriates (iridoids), the components of the volatile oil, sesquiterpenes as valerenic acid (VA) and its derivatives, monoterpenes as well as a number of other constituents [4]. Although the identity of the active components responsible for the beneficial health effects in valerian is still uncertain, in recent findings the main pharmacological effects have been attributed to valerenic acid and its derivatives including acetoxyvalerenic acid and hydroxyvalerenic acid [5,6]. Also, VA along with its derivatives is often used to indicate medicinal quality [4,7]. The production of medicinal compounds from extraction of wild or cultivated plants can be limited by various problems: plants difficult to cultivate, risk of extinction for overexploited plants, and geopolitical problems, among other causes [8]. In vitro culture of plants can overcome these problems, since the Cent. Eur. J. Biol. • 9(9) • 2014 • 853-863 DOI: 10.2478/s11535-014-0320-3 853 Received 29 November 2013; Accepted 03 April 2014 Keywords: Elicitation • Hairy root culture • Valerenic acid production • Valeriana officinalis Abstract: Valerenic acid (VA) is a pharmacologically-active sesquiterpene found in valerian (Valeriana officinalis L., Valerianaceae) roots and rhizomes. The plant produces only small amounts of this metabolite naturally. So, induction of hairy roots as well as elicitation can be useful to increase its commercial production. In this study, Wild-type strain ‘A13’ of Agrobacterium rhizogenes was used to induce hairy roots in valerian. The influence of three different elicitors including Fusarium graminearum extract (FE), methyl jasmonate (MJ) and salicylic acid (SA) on VA production in the selected hairy root line ‘LeVa-C4’ was also investigated. The 23-day-old cultures were treated with different concentrations of the elicitors at exposure time of 3 and 7 days. FE (1%) and MJ (100 µM L -1 ) highly promoted VA production at 7 days after elicitation, to a level of 12.31- and 6-fold higher than that of non-elicited controls, respectively, and FE did not exert any negative effects on biomass yield of hairy root. SA did not significantly increase the production of VA. This is the first time study to assess the elicitation of hairy root cultures to promote VA biosynthesis in valerian and the resulting experiments demonstrated that F. graminearum extract and MJ were indeed a potent inducer of VA biosynthesis. © Versita Sp. z o.o.
Transcript of Central European Journal of Biology Enhanced production of valerenic acid in hairy root culture of...

Central European Journal of Biology
* E-mail: [email protected]
Research Article
1Department of Biology, Faculty of Science, University of Urmia, P.O. Box 165, Urmia, Iran2 Department of Plant Breeding and Biotechnology, Faculty of Agriculture, University of Urmia, P.O. Box 165, Urmia, Iran3Department of Agricultural Biotechnology, Institute of Biotechnology, University of Urmia, P.O. Box 165, Urmia, Iran4Department of Plant Pathology, Faculty of Agriculture, Tarbiat Modares University, Tehran, Iran
Mohammad Reza Dini Torkamani1, Morad Jafari2,3*, Nasser Abbaspour1, Reza Heidary1, Naser Safaie4
Enhanced production of valerenic acid in hairy root culture of Valeriana officinalis by elicitation
1. IntroductionValerian (Valeriana officinalis L.), a perennial herb from the Valerianaceae family, is native to Europe and Asia. It has been used as a medicinal herb worldwide [1,2]. Pharmaceutical application of valerian is due to its sedative, anticonvulsant, hypnotic effects, and anxiolytic activity. Lately, the potential cytoprotective effect of an aqueous extract of valerian on human neuroblastoma cells has also been demonstrated [3]. The compounds accumulated mainly in the roots and rhizomes of valerian include valepotriates (iridoids), the components of the volatile oil, sesquiterpenes as valerenic acid (VA) and its derivatives, monoterpenes
as well as a number of other constituents [4]. Although the identity of the active components responsible for the beneficial health effects in valerian is still uncertain, in recent findings the main pharmacological effects have been attributed to valerenic acid and its derivatives including acetoxyvalerenic acid and hydroxyvalerenic acid [5,6]. Also, VA along with its derivatives is often used to indicate medicinal quality [4,7].
The production of medicinal compounds from extraction of wild or cultivated plants can be limited by various problems: plants difficult to cultivate, risk of extinction for overexploited plants, and geopolitical problems, among other causes [8]. In vitro culture of plants can overcome these problems, since the
Cent. Eur. J. Biol. • 9(9) • 2014 • 853-863DOI: 10.2478/s11535-014-0320-3
853
Received 29 November 2013; Accepted 03 April 2014
Keywords: Elicitation • Hairy root culture • Valerenic acid production • Valeriana officinalis
Abstract: Valerenic acid (VA) is a pharmacologically-active sesquiterpene found in valerian (Valeriana officinalis L., Valerianaceae) roots and rhizomes. The plant produces only small amounts of this metabolite naturally. So, induction of hairy roots as well as elicitation can be useful to increase its commercial production. In this study, Wild-type strain ‘A13’ of Agrobacterium rhizogenes was used to induce hairy roots in valerian. The influence of three different elicitors including Fusarium graminearum extract (FE), methyl jasmonate (MJ) and salicylic acid (SA) on VA production in the selected hairy root line ‘LeVa-C4’ was also investigated. The 23-day-old cultures were treated with different concentrations of the elicitors at exposure time of 3 and 7 days. FE (1%) and MJ (100 µM L-1) highly promoted VA production at 7 days after elicitation, to a level of 12.31- and 6-fold higher than that of non-elicited controls, respectively, and FE did not exert any negative effects on biomass yield of hairy root. SA did not significantly increase the production of VA. This is the first time study to assess the elicitation of hairy root cultures to promote VA biosynthesis in valerian and the resulting experiments demonstrated that F. graminearum extract and MJ were indeed a potent inducer of VA biosynthesis.
© Versita Sp. z o.o.

Valerenic acid production in hairy roots of valerian by elicitation
environmental conditions that affect plant metabolism can then be strictly controlled. Working with plant cells drastically reduces preparation time, handling and storage costs compared with traditional whole plant approaches [9]. Although the plant cell is considered to be totipotent, the biosynthesis of many secondary metabolites requires a certain level of differentiation of the tissues. Hairy roots produced by Agrobacterium rhizogenes infection are characterized by fast growth, biosynthetic stability and ability to grow in hormone–free medium, all advantages that make transformed roots efficient sources for secondary metabolite production [10]. The greatest advantage of hairy roots is the ability to produce higher levels of secondary metabolites as compared to intact plants [11]. Medicinal plants have been widely explored for potential hairy root culture and production of their bioactive compounds [12].
In vitro plant secondary metabolite production is enhanced through elicitation in plant cell or tissue culture systems [13]. Elicitors as compounds or treatments have been proved to be an effective method to increase secondary metabolite production. A number of elicitors such as methyl jasmonate (MJ) [14-17] and salicylic acid (SA) [14,18] have been used successfully for enhanced production of secondary metabolites in hairy root cultures of many plant species. In addition, fungal elicitors are significant in triggering phytoalexin production in cultured cells and have been used for enhanced production of secondary metabolites in some medicinal plants [19-22]. High biosynthetic capability of valepotriate production has been reported in hairy root cultures of Valeriana wallichii DC [23], Valerianella discoidea L. Loisel [24], Valeriana officinalis L. var. sambucifolia Mikan [25] and Valerianella locusta [26] and in adventitious root cultures of Valeriana glechomifolia [27] and V. officinalis L. [28].
To our knowledge, there is no published study on elicitor-induced production of secondary metabolites in hairy root cultures of Valerian. In the present study, for the first time, the effects of three different elicitors i.e. MJ, SA and Fusarium graminearum extract with different time exposure were investigated on growth rate and VA content of valerian hairy root cultures. This paper suggests the optimal concentration of elicitors for high-level VA production in hairy root cultures of valerian.
2. Experimental procedures2.1 Hairy root production2.1.1. Plant materialsSeeds of valerian were provided by Pakan Bazr Co. (Isfahan, Iran) at late August 2011. To increase
germination rate, seeds were pre-treated with commercial grade sulfuric acid (10 %) for 15 min. After complete washing with sterile distilled water, the seeds were surface-sterilized in 70% (v/v) ethanol for 1 min, rinsed with sterile distilled water for 3 min and immersed in 2% (v/v) solution of sodium hypochlorite (commercial bleach) for 10 min, followed by three rinses with sterile distilled water. The seeds were blotted dry with sterilized filter paper and cultured on MS [29] basal medium containing 3% sucrose and 0.7% plant agar (Duchefa, Netherlands). The cultures were maintained in a growth chamber at 22±2°C and 70% humidity and a 16/8-h light/dark photoperiod with a photon flux density (PFD) of 60 μmol/m2s. After seed germination, seedlings were transferred to fresh medium for further growth. The pH of the medium was adjusted to 5.9 with KOH (0.1 N) or HCl (1 N) prior to autoclaving at 121°C temperature for 20 min.
2.1.2. Infection of leaf explants and hairy root inductionAmong the different explants tested (data not shown), 5-week-old leaf explant was used for hairy root induction because of its capacity for higher transformation efficiency. So, leaf explants obtained from 5-week-old in vitro grown seedlings were cut into small segments (5–10 mm) and were dipped in the suspension of Agrobacterium rhizogenes A13 strain for 30 min and the explants were then blotted with sterile filter paper to remove excess liquid. Then explants were transferred onto 100×15-mm sterile petri dishes (15 explants per dish) containing agar-solidified MS co–cultivation medium and maintained in the dark at 24±2°C for 2 days. As a control, a few explants were immersed in sterile distilled water and were cultured the same way. After 2 days, the explants were rinsed with sterile distilled water and placed on solid hormone-free MS medium containing 500 mg/l cefotaxime. After the appearance of hairy roots at the wound sites of the explants, they were subcultured at intervals of 2 weeks on a fresh medium and the concentration of cefotaxime was gradually reduced and then omitted after the third subculture.
2.1.3. Establishment of hairy rootsHairy roots with more than four lateral branches were cut and transferred to 100 ml Erlenmeyer flaks containing 25 ml of hormone–free MS liquid medium supplemented with 3% (w/v) sucrose. The root cultures were incubated on an orbital shaker (100 rpm) in darkness at 22°C and subcultured on fresh liquid medium at 2 week-intervals. To measure hairy root growth rate in terms of fresh weight (FW, mg/25 ml culture medium), they were harvested after 4 subcultures and blotted dry on sterile filter paper to remove excess moisture.
854

M.R.D. Torkamani et al.
2.2 Polymerase chain reaction (PCR) analysis for the rolB and virD2 genes
The extraction of DNA from both hairy roots and untransformed roots (control) was carried out using the CTAB method [30]. For molecular confirmation of the transgenic nature of hairy roots, the presence of the rol genes located on the T-DNA as the main determinants of hairy root development were examined by PCR analysis using corresponding gene-specific primer pairs. The Ri plasmid of A. rhizogenes strains ‘A13’ was used as a positive control. The primer sequences to amplify a 780-bp portion of the rolB gene were 5ʹ-TTAGGCTTCTTTCTTCAGGTTTACTGCAGC-3ʹ (forward primer) and 5 ʹ -TGGATCCCAAATTGCTATTCCTTCCACGA-3ʹ (reverse primer). The primers (5ʹ-ATGCCCGATCGAGCTCAAGT-3ʹ and 5ʹ-CCTGACCCAAACATCTCGGCTGCCCA-3ʹ as forward and reverse primers, respectively) specific to the virD2 gene present in the non-transferred virulence region of the A. rhizogenes Ri plasmid were also used in PCR analysis to ensure transformed root tissue was free of bacterial cells. PCR reaction profiles involved: initial denaturation at 94°C for 5 min, followed by 35 cycles of 94°C for 1 min, 58°C for 1 min, 72°C for 1 min, and a final extension at 72°C for 10 min. The amplified DNAs were analyzed on 0.8% agarose gel at 90 V for 1 h and stained with ethidium bromide.
2.3 Elicitor preparation and treatmentA stock solution of MJ was obtained by dissolving it in 96% ethanol after filter sterilization using 0.22 µm Sartorius filters. A stock solution of SA was prepared in methanol, and then filter-sterilized. F. graminearum [Teleomorph: Gibberella zeae (Schwenitz) Petch] was also used as fungal elicitor. Fungal extract (FE) was prepared as described by Esmaeilzadeh et al. [19]. Briefly, one-week-old mycelia of F. graminearum were harvested and rinsed with sterile distilled water. The collected mycelia were ground under liquid nitrogen and suspended with water to a final concentration of 250 mg L-1. The suspension was centrifuged at 10,000 gfor 10 min and then the supernatant was autoclaved for 10 min.
For the elicitation treatment, 23-day-old hairy root cultures of the LeVa-C4 line were inoculated into 100-ml Erlenmeyer flasks containing 25 ml of liquid MS medium supplemented with various concentrations of MJ (50, 100, 200 µM L-1), SA (50, 100, 150 µM L-1) or FE (0.5, 1, 2% v/v) for different exposure times (3 and 7 days). The control cultures received the same amount of 96% ethanol or water. All cultures were incubated on an orbital shaker at 100 rpm in darkness at 25°C.
At the end of the exposure times, the root cultures were harvested and subjected to VA content analysis. Growth rate of elicitor-treated hairy root cultures was also determined in term of dry weight (DW, mg/25 ml culture medium) and for DW measurement, the samples were lyophilized.
2.4 VA quantificationFor VA extraction, hairy roots and untransformed roots were lyophilized and ground to a fine powder. The root powder (approx. 200 mg) was extracted three times with 5 ml of 70% MeOH, sonicated for 10 min and then diluted to a final volume of 30 ml with MeOH. All samples were filtered through a 0.45 µm micro-filter (Merck, Germany) and this solution was used for chemical analysis.
High-performance liquid chromatography (HPLC) analysis was carried out using a Knauer HPLC system (Germany) equipped with a UV detector k-2501. Separation was achieved by reverse-phase isocratic C18 column (250×3 mm), with a pre-column (25×4.6 mm, particle size 5 µm). The mobile phase A was 0.5% (v/v) phosphoric acid and the mobile phase B was a 0.5% phosphoric acid:methanol mixture (27:73). The flow rate was 1.5 ml min−1 and the injection volume was 10 µl. Detection was by UV at wavelength 225 nm. VA was obtained from Sigma Aldrich Co. (USA). Standard solution was prepared by dissolving VA in methanol to obtain solutions with 7 different concentrations in the range of 0.01562-1 µg L-1. The calibration curves were established based on measuring their respective peak areas. All analyses were repeated three times employing three replicates each. The results were expressed as mean values ± SE. The metabolite contents were calculated as mg of the compound per gram of root DW.
2.5 Statistical analysisAll experiments were repeated three times and for each set of experiments, four replicates (each replication consisted of one Erlenmeyer flask) were included. The data are expressed as the average of results of triplicate experiments and obtained data were subjected to analysis of variance (ANOVA) using the general linear models (PROC GLM) procedure in the Statistical Analysis System (SAS 9.1) programme package. The means were compared using Fisher’s least significant difference (FLSD) test at the 5% probability level.
3. Results and Discussion3.1 Establishment of hairy root culturesLeaf explants were infected by strain A13 of A. rhizogenes. Hairy root formation was observed
855
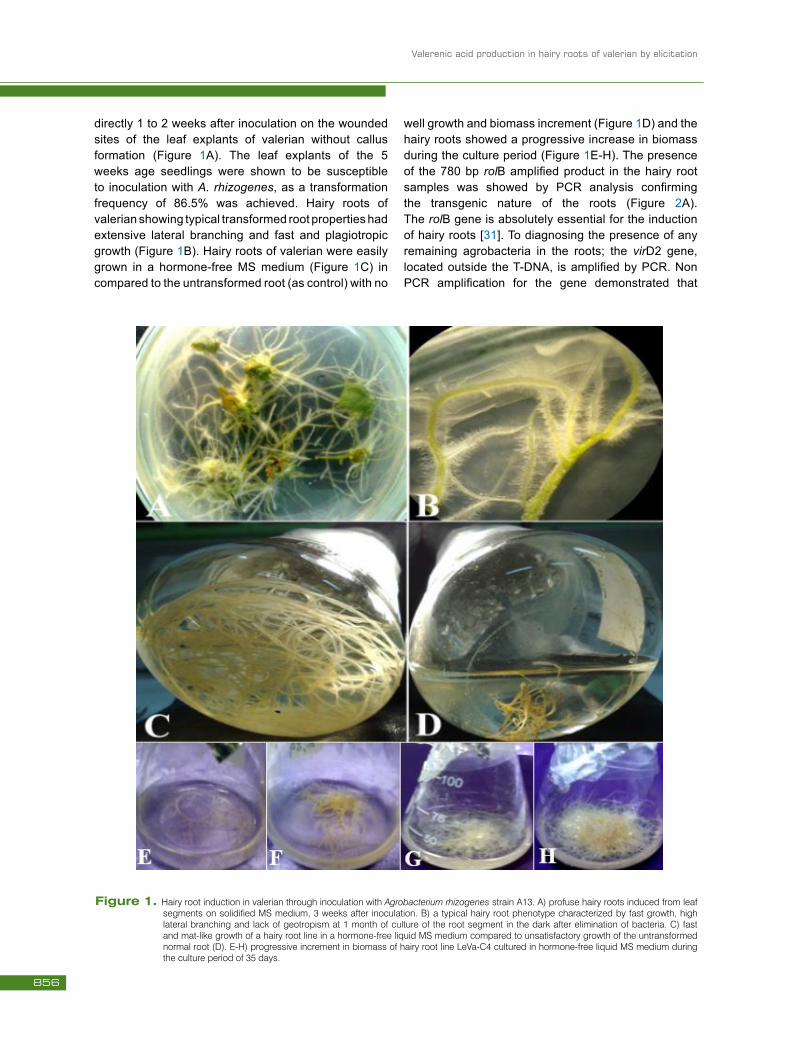
Valerenic acid production in hairy roots of valerian by elicitation
directly 1 to 2 weeks after inoculation on the wounded sites of the leaf explants of valerian without callus formation (Figure 1A). The leaf explants of the 5 weeks age seedlings were shown to be susceptible to inoculation with A. rhizogenes, as a transformation frequency of 86.5% was achieved. Hairy roots of valerian showing typical transformed root properties had extensive lateral branching and fast and plagiotropic growth (Figure 1B). Hairy roots of valerian were easily grown in a hormone-free MS medium (Figure 1C) in compared to the untransformed root (as control) with no
well growth and biomass increment (Figure 1D) and the hairy roots showed a progressive increase in biomass during the culture period (Figure 1E-H). The presence of the 780 bp rolB amplified product in the hairy root samples was showed by PCR analysis confirming the transgenic nature of the roots (Figure 2A). The rolB gene is absolutely essential for the induction of hairy roots [31]. To diagnosing the presence of any remaining agrobacteria in the roots; the virD2 gene, located outside the T-DNA, is amplified by PCR. Non PCR amplification for the gene demonstrated that
Figure 1. Hairy root induction in valerian through inoculation with Agrobacterium rhizogenes strain A13. A) profuse hairy roots induced from leaf segments on solidified MS medium, 3 weeks after inoculation. B) a typical hairy root phenotype characterized by fast growth, high lateral branching and lack of geotropism at 1 month of culture of the root segment in the dark after elimination of bacteria. C) fast and mat-like growth of a hairy root line in a hormone-free liquid MS medium compared to unsatisfactory growth of the untransformed normal root (D). E-H) progressive increment in biomass of hairy root line LeVa-C4 cultured in hormone-free liquid MS medium during the culture period of 35 days.
856

M.R.D. Torkamani et al.
there was no bacterial contamination of the hairy root tissues (Figure 2B).
Measuring the fresh biomass of transformed (hairy) roots and the normal root under the same condition after 4 subcultures showed that the transformed samples produced more biomass (maximum biomass of 4133 mg FW/25 ml culture medium) in comparison with the untransformed samples (132 mg FW/25 ml culture medium). The strain of Agrobacterium influences the development, growth rate and secondary metabolite production in transformed root cultures [32]. The A13 strain of A. rhizogenes is one of the aggressiveness strains whose superiority for hairy root induction and bioactive compound production have been reported in several other medicinal plants [33,34].
3.2 Effects of elicitors on biomass production of hairy roots
As a result of the transformation with A. rhizogenes and hairy root production, the LeVa-C4 hairy root line was selected for elicitation studies, because roots of the line were a light yellow, giving fast and stable growth without a tendency to develop callus. 23-day-old hairy root cultures of the LeVa-C4 line was treated with various elicitors and all the elicitors were shown to influence the growth of hairy roots in a dose-dependent and point time manner (Figure 3). FE did not inhibit growth at a time exposure of 3 days; but 7 days after treatment the fungal extract at a concentration of 1% significantly increased biomass by 13% compared to the non-elicited control, whereas increasing of FE concentration
to 1.5% decreased growth (Figure 3A). Decrease (50%) in growth in suspension cultures of Linum album using F. graminearum extract has been reported [19].
MJ treatment caused a slight decline in biomass throughout the exposure period and maximum inhibition of growth (decrease by 12.7%) was found after 7th days of exposure time at 200 µM MJ (Figure 3B). The addition of MJ to the cultures also changed the color of the roots from white to dark brown. This negative effect may be due to a direct toxic effect of MJ and loss of viability of the culture [38]. These results are in agreement with earlier reports [19,32,35]. SA decreased biomass 3 days after treatment but did not affect the growth rate at exposure time of 7 days, and DW of elicited roots at the end of the culture period was similar to the control (Figure 3C). This is in accordance with previous studies reporting that SA treatment has no significant negative effects on growth of Fagopyrum esculentum suspension cultures [35] and root cultures of Angelica gigas [13]. However, a negative effect of SA on growth of plant cell culture has also been reported by several studies [36,37]. In the present study, to enhance overall productivity and limit negative effects of elicitors (especially MJ) on growth of roots, elicitation in valerian hairy roots were tested when the roots were in the late linear or early stationary growth phase (23rd day of the culture period).
3.3 Effects of elicitors on VA productionAll concentrations of F. graminearum extract led to a significant increase in VA level at both exposure times compared to the control, however the highest content of
Figure 2. PCR analysis for hairy roots of valerian using the rolB gene (A) and virD2 gene (B) specific primers. M– 1 Kb DNA Ladder (Fermantas), Ri plasmid– the plasmid from A. rhizogenes strain A13 as a positive control, LeVa-A6, LeVa-C4 and LeVa-G7– Transgenic hairy root lines raised from leaf explants, UTR– Untransformed normal root as first negetive control, NDT– Non-DNA template PCR reaction as second negative control.
857

Valerenic acid production in hairy roots of valerian by elicitation
VA (3.02 mg/g DW) was observed after treatment with 1% FE for 7 days (Figures 4, 5A). The levels of VA were 11.06- and 12.31 times higher than that of non-elicited control (0.24 mg g-1 DW) at exposure times of 3 and 7 days, respectively. This substantial effect of FE has also been reported for podophyllotoxin production in suspension culture of Linum album [19]. Accumulation of JA and its derivatives, such as methyl jasmonate (MJ) is one of the common responses of plant cells to fungal elicitors and
as intermediate signals in elicitor-induced secondary metabolite accumulation in plant cells [39]. Few studies refer to the plant secondary metabolism mediated by the fungal elicitor. It has been speculated that recognition of the fungi elicitor by host plants triggers a cascade of signal transduction, which gives rise to a series of plant defense responses similar to plant pathogen interaction, thus leading to a noticeable change in plant metabolic state [40].
Figure 3. Effect of Fusarium graminearum extract (FE) (A), methyl jasmonate (MJ) (B) and salicylic acid (SA) (C) on growth of hairy root line LeVa-C4 of valerian. 23-day-old hairy root cultures were exposed to various concentrations of FE, SA and MJ for 3 and 7 days. Error bars represent the standard error of the mean of three repeated experiments. Means with the same letter are not significantly different according FLSD test (P < 0.05).
26
858

M.R.D. Torkamani et al.
Jasmonic acid and its analogues are known to induce the transcription of genes involved in plant defense metabolism [41]. The effects of MJ concentrations at two exposure times on VA production in hairy root cultures
of valerian is shown in Figure 5B. MJ had positive effect on VA accumulation at both timepoints examined. However, increasing MJ concentration from 100 to 150 µM L-1 significantly decreased VA content
Figure 4. Chromatograms of standard valerenic aicd (A), non-elicited control (B) and hairy root cultures exposed with 1% (v/v) Fusarium graminearum extract (for a 7 days) which was the best treatment of elicitation (C).
859

Valerenic acid production in hairy roots of valerian by elicitation
even below (0.16 mg g-1 DW) that of the control cultures (0.246 mg g-1 DW) after 7 days of exposure. The highest increase in VA accumulation (2.296 mg g-1 DW) was obtained with the addition of 100 µM L-1 MJ for a time period of 7 days, which was about 6-fold higher than that of non-elicited control hairy
roots (0.246 mg g-1 DW). MJ has also been reported to promote production of valtrate in the Valeriana amurensis adventitious root cultures [42].
It is known that secondary metabolites increase through MJ elicitation, but to reach the highest level the time required is different for each biosynthetic
28
Figure 5. Effect of Fusarium graminearum extract (FE) (A), methyl jasmonate (MJ) (B) and salicylic acid (SA) (C) on valerenic acid (VA) production in hairy root line LeVa-C4 of valerian. 23-day-old hairy root cultures were exposed to various concentrations of FE, SA and MJ for 3 and 7 days. Error bars represent the standard error of the mean of three repeated experiments. Means with the same letter are not significantly different according FLSD test (P < 0.05).
860

M.R.D. Torkamani et al.
pathway and even each compound type. For example the diterpenoid content in Salvia sclarea hairy root cultures reached a maximum after 7 days of exposure to 125 µM MJ [38]. But a longer period (10 days) was required for reaching the highest level of valepotriate production in hairy roots of Valerianella locusta [26]. In the present study, exposure to MJ for 7 days enhanced VA accumulation two-fold higher than 3 days of exposure time (1.266 mg g-1 DW).
SA slightly induced VA accumulation in the hairy root cultures in both exposure times compared to controls (Figure 5C). VA content at the highest concentration of SA used (150 mg L-1) was significantly higher (by 50%) than other concentrations at both exposure times, however SA did not improve VA contents of hairy roots compared with FE and MJ (Figures 5A, B). The inefficiency of SA in promoting valtrate production in adventitious roots of Valeriana amurensis has also been reported [42]. However, several studies have shown the effectiveness of SA in improving target metabolites in the plant cells [35,43]. SA and Jasmonic acid (JA)/MJ are messengers in the signal transduction chains involved in different pathways of biosynthesis and in activation of secondary metabolites and enzymes that participate in the plant stress response [44]; and exogenous application of MJ and SA may also induce the expression of many defense genes in plants [45]. JA signaling might be reinforced as a substitute for SA signaling to mediate elicitor-induced secondary
metabolite production when SA accumulation is impaired and it has been suggested that an unusual complementary relationship exists between JA and SA in mediating plant secondary metabolite accumulation, however SA does not have similar effects on JA signaling in mediating the fungal elicitor-induced secondary metabolite biosynthesis [46].
In conclusion, the results obtained in this study demonstrated that F. graminearum extract at 1% (v/v) and MJ at 100 µM L-1 significantly enhanced the biosynthesis of valerenic acid from hairy root cultures. Furthermore, SA does not appear to be an effective elicitor to increase VA content in this study. This work may help further investigate the optimal concentrations and exposure times of fungal elicitor and MJ for the possible large-scale production of valerenic acid from hairy root cultures of valerian in a bioreactor system.
Conflict of interestsThe authors have no conflict of interest to disclose.
AcknowledgementsThe authors would like to acknowledge the Biotechnology Research Center of Urmia University for providing the necessary laboratory facilities.
References
[1] Hazelhoff B., Malingre T.M., Meijer D.K.F., Antispasmodic effects of valeriana compounds: an in vivo and in vitro study on the guinea-pig ileum, Arch. Int. Pharmacodyn., 1982, 257, 274-287
[2] Rucker G., Tautges J., Sieck A., Wenzl H., Graf E., Isolation and pharmacodynamic activity of the sesquiterpene valeranone from Nardostachys jatamansi DC [in German], Arzneimittelforschung, 1987, 28, 7-13
[3] Oliveria D.M., Barreto G., De Andrade G.V.D., Saraceno E., Bertolino L.A., Capani F., Dos Santos El Bacha R., Giraldez L.D., Cytoprotective Effect of Valeriana officinalis Extract on an in vitro Experimental Model of Parkinson Disease, Neurochem. Res., 2009, 34, 215-220
[4] Bos R., Woerdenbag H.J., De Smet P.A.G.M., Scheffer J.J.C., Valeriana Species, In: De Smet P.A.G.M., Keller K., Chandler R.F. (Eds.), Adverse effects of herbal drugs, Springer-Verlag, 1997
[5] Khom S., Baburin I., Timin N., Hohaus A., Trauner G., Kopp B., Hering S., Valerenic acid potentiates and
inhibits GABAA receptors: molecular mechanism and subunit specificity, Neuropharmacol., 2007, 53, 178-187
[6] Trauner G., Khom S., Baburin I., Benedek B., Hering S., Kopp B., Modulation of GABAA receptors by valerian extracts is related to the content of valerenic acid. Planta Med., 2008, 74, 19-24
[7] Hansel R., Schulz J., Valerens auren und Valerenal als Leitstoffe des offizinellen Baldrians. Bestimmung mittels HPLC-Technik, Z. Phytother, 1982, 3, 333-340
[8] Verpoorte R., Contin A., Memelink J., Biotechnology for the production of plant secondary metabolites, Phytochem. Rev., 2002 1, 13-25
[9] Gerth A., Schmidt D., Wilken D., The production of plant secondary metabolites using bioreactors. The 27th International Horticultural Congress, Symposium on Plant Biotechnology: From Bench to Commercialization (13-19 August 2006, Seoul, Korea), Seoul, 2006
861

Valerenic acid production in hairy roots of valerian by elicitation
[10] Srivastava S., Srivastava A.K., Hairy root culture for mass-production of high-value secondary metabolites, Crit. Rev. Biotech., 2007, 27, 29-43
[11] Kim C.H., Park J.H., Chung I.S., Kim S.R., Lee S.W., Enhanced anthocyanin production in hairy root culture of daucus carota by fungal elicitor, Hort. Sci. J., 1992, 27, 694
[12] Guillon S., Tremouillaux-Guiller J., Pati P.K., Rideau M., Gantet P., Harnessing the potential of hairy roots: dawn of a new era, Trends Biotechnol, 2006, 24, 403–409
[13] Rhee H.S., Cho H.Y., Son S.Y., Yoon S.Y.H., Park J.M., Enhanced accumulation of decursin and decursinol angelate in root cultures and intact roots of Angelica gigas Nakai following elicitation, Plant Cell Tiss. Org. Cult., 2010, 1, 295-302
[14] Wielanek M., Urbanek H., Glucotropaeolin and myrosinase production in hairy root cultures of Tropaeolum majus, Plant Cell Tiss. Org. Cult., 1999, 57, 39-45
[15] Bauer N., Kiseljak D., Jelaska S., The effect of yeast extract and methyl jasmonate on rosmarinic acid accumulation in Coleus blumei hairy roots, Biol. Plantarum, 2009, 53, 650-656
[16] Sun J., Xiao J., Wang X., Yuan X., Zhao B., Improved cardenolide production in Calotropis gigantea hairy roots using mechanical wounding and elicitation, Biotechnol. Lett., 2012, 34, 563-569
[17] Kim O.T., Yoo N.H., Kim G.S., kim Y.C., Bang K.H., Hyun D.Y., Kim S.H., Kim M.Y., Stimulation of Rg3 ginsenoside biosynthesis in ginseng hairy roots elicited by methyl jasmonate, Plant Cell Tiss. Org. Cult., 2013, 112, 87-93
[18] Chotikadachanarong K., Dheeranupattana S., Jatisatienr C., Wangkarn S., Mungkornasawakul P., Pyne S.G., Ung A.T., Sastraruji T., Influence of salicylic acid on alkaloid production by root cultures of Stemona curtisii Hook, Cur. Res. J. Bio. Sci., 2011, 3, 322-325
[19] Esmaeilzadeh S., Sharifi M., Safaie N., Murata J., Yamagaki T., Satake H., Increased lignin biosynthesis in the suspension cultures of Linum album by fungal extracts, Plant Biotechnol. Rep., 2011, 5, 367-373
[20] Harsh P.B., Dattatreya B.S., Ravishankar G.A., Production of volatile compounds by hairy root cultures of Cichorium intybus L under the influence of fungal elicitors and their analysis using solid-phase micro extraction gas chromatography–mass spectrometry, J. Sci. Food Agri., 2003, 83, 769-774
[21] Karwasara V.S., Tomar P., Dixit V.K., Influence of fungal elicitation on glycyrrhizin production in
transformed cell cultures of Abrus precatorius Linn., Phcog. Mag., 2011, 7, 307-313
[22] Kim Y.J., Weathers P.J., Wyslouzil B.E., The growth of Artemisia annua hairy roots in liquid and gas phase reactors, Biotechnol. Bioeng., 2002, 80, 454-464
[23] Banerjee S., Rahman L., Uniyal G.C., Ahuja P.S., Enhanced production of valepotriates by Agrobacterium rhizogenes induced hairy root cultures of Valeriana wallichii DC., Plant Sci., 1998, 131, 203-208
[24] Caetano L.C., Charlwood B.V., Gahan P.B., The localization and accumulation of valepotriates in hairy roots of Valerianella discoidea (L.) Loisel, Phytochem. Anal., 1999, 10, 181-186
[25] Gränicher F., Christen P., Kapetanidis I., High-yield production of valepotriates by hairy root cultures of Valeriana officnalis L. var. sambucifolia Mikan. Plant Cell. Rep., 1992, 11, 339-342
[26] Kittipongpatana N., Davis D.L., Porter J.R., Methyl jasmonate increases the production of valepotriates by transformed root cultures of Valerianella locusta, Plant Cell Tiss. Org. Cult., 2002, 71, 65-75
[27] Maurmann N., Carvalho C.M.B., Silva L.A., Fett-Neto A.G., von Poser G.L., Rech S.B., Valepotriates accumulation in callus, suspended cells and untransformed root cultures of Valeriana glechomifolia, In Vitro Cell Dev. Biol. Plant, 2006, 42, 50-53
[28] Tousi S.E., Radjabianb T., Ebrahimzadeha H., Niknama V., Enhanced production of valerenic acids and valepotriates by in vitro cultures of Valeriana officinalis L., Int. J. Plant Prod., 2010, 4, 209-222
[29] Murashige T., Skoog F., A revised medium for rapid growth and bioassays with tobacco tissue cultures, Physiol. Plant, 1962, 15, 473-476
[30] Doyle J.J., Doyle J.L., A rapid isolation procedure for small quantities of fresh leaf tissue, Phytochem. Bull., 1987, 19, 11-15
[31] Nilsson O., Olsson O., The role of the Agrobacterium rhizogenes rol genes in the formation of hairy roots, Physiol. Plantarum, 1997, 100, 463-473
[32] Lee S.Y., Kim S.G., Song W.S., Kim Y.K., Park N.I., Park S.U., Influence of different strains of Agrobacterium rhizogenes on hairy root induction and production of alizarin and purpurin in Rubia akane Nakai, Romanian Biotechnol. Lett., 2010, 15, 5405-5409
[33] Afsaneh S., Carapetian J., Heidari R., Jafari M., Hassanzadeh Gorttapeh A., Hairy Root Induction in Linum mucronatum ssp. mucronatum, an Anti-Tumor Lignans Producing Plant, Not. Bot. Horti. Agrobo., 2012, 40, 125-131
862

M.R.D. Torkamani et al.
[34] Ooi C.T., Syahida A., Stanslas J., Maziah M., Efficiency of different Agrobacterium rhizogenes strains on hairy roots induction in Solanum mammosum, World J. Microb. Biotechnol., 2013, 29, 421-430
[35] Hu Y.H., Yu Y.T., Piao C.H., Liu J.M., Yu H.S., Methyl jasmonate- and salicylic acid-induced D-chiro-inositol production in suspension cultures of buckwheat (Fagopyrum esculentum), Plant Cell Tiss. Org. Cult., 2011, 106, 419-424
[36] Lee K.T., Hirano H., Yamakawa T., Kodama T., Igarashi Y., Shimomura K., Responses of transformed root culture of Atropa belladonna to salicylic acid stress, J. Biosci. Bioeng., 2001, 91, 586-589
[37] Ali M.B., Hahn E.J., Paek K.Y., Methyl jasmonate and salicylic acid induced oxidative stress and accumulation of phenolics in panax ginseng bioreactor root suspension cultures. Molecules, 2007, 12, 607-621
[38] Lukasz K., Elzbieta B., Halina W., Methyl jasmonate effect diterpenoid accumulation in Salvia sclarea hairy root culture in shake flasks and sprinkle bioreactor, Enzyme Microb. Tech., 2009, 44, 406-410
[39] Xu M.J., Dong J.F., Zhu M.Y., Nitric oxide mediates the fungal elicitor-induced hypericin production of hypericum perforatum cell suspension cultures through a jasmonic-acid-dependent signal pathway, Plant Physiol., 2005, 139, 991-998
[40] Chen X.M., Guo S.X., Effects of four species of endophytic fungi on the growth and polysaccharide
and alkaloid contents of Dendrobium nobile, China J. Chin. Mat. Med., 2005, 30, 253-257
[41] Van der Fits L., Memelink J., ORCA3 a jasmonate–responsive transcriptional regulator of plant primary and secondary metabolism, Science, 2000, 289, 295-297
[42] Cui L., Wang Z.Y., Zhou X.H., Optimization of elicitors and precursors to enhance valtrate production in adventitious roots of Valeriana amurensis Smir ex Kom, Plant Cell Tiss. Org. Cult., 2012, 108, 411-420
[43] Yousefzadi M., Sharifi M., Behmanesh M., Ghasempour A., Moyano E., Palazon J., Salicylic acid improves podophyllotoxin production in cell cultures of Linum album by increasing the expression of genes related with its biosynthesis, Biotechnol. Lett., 2010, 32, 1739-1743
[44] Pieterse C.M.J., Ton J., Van Loon L.C., Cross-talk between plant defense signaling pathways: boost or burden? Ag. Biotech. Net., 2001, 3, 1-8
[45] Ding C.K., Wang C.Y., Gross K.C., Smith D.L., Jasmonate and salicylate induce the expression of pathogenesis-related-proteins genes and increase resistance to chilling injury in tomato fruit, Planta, 2002, 214, 895-901
[46] Xu M., Dong J., Wang H., Huang L., Complementary action of jasmonic acid on salicylic acid in mediating fungal elicitor-induced flavonol glycoside accumulation of Ginkgo biloba cells, Plant Cell Environ., 2009, 32, 960-967
863




















