
Contribution of D.R. Kasparyan to the knowledge of Mexican Ichneumonidae (Hymenoptera)

This article was downloaded by: [98.208.164.98]On: 14 January 2015, At: 10:15Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registeredoffice: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Click for updates
Journal of Natural HistoryPublication details, including instructions for authors andsubscription information:http://www.tandfonline.com/loi/tnah20
Similiclava nivea (Cnidaria: Hydrozoa:Similiclavidae): a new family, genusand species of athecate hydroid fromthe Pacific coast of North AmericaDale R. Caldera, Henry H. C. Choongab & Neil McDanielca Invertebrate Zoology Section, Department of Natural History,Royal Ontario Museum, Toronto, Canadab Palaeobiology Section, Department of Natural History, RoyalOntario Museum, Toronto, Canadac McDaniel Photography, Vancouver, CanadaPublished online: 12 Jan 2015.
To cite this article: Dale R. Calder, Henry H. C. Choong & Neil McDaniel (2015): Similiclava nivea(Cnidaria: Hydrozoa: Similiclavidae): a new family, genus and species of athecate hydroid from thePacific coast of North America, Journal of Natural History, DOI: 10.1080/00222933.2014.979261
To link to this article: http://dx.doi.org/10.1080/00222933.2014.979261
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the“Content”) contained in the publications on our platform. However, Taylor & Francis,our agents, and our licensors make no representations or warranties whatsoever as tothe accuracy, completeness, or suitability for any purpose of the Content. Any opinionsand views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Contentshould not be relied upon and should be independently verified with primary sourcesof information. Taylor and Francis shall not be liable for any losses, actions, claims,proceedings, demands, costs, expenses, damages, and other liabilities whatsoever orhowsoever caused arising directly or indirectly in connection with, in relation to or arisingout of the use of the Content.
This article may be used for research, teaching, and private study purposes. Anysubstantial or systematic reproduction, redistribution, reselling, loan, sub-licensing,systematic supply, or distribution in any form to anyone is expressly forbidden. Terms &

Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Similiclava nivea (Cnidaria: Hydrozoa: Similiclavidae): a new family,genus and species of athecate hydroid from the Pacific coast of NorthAmerica
Dale R. Caldera*, Henry H. C. Choonga,b† and Neil McDanielc
aInvertebrate Zoology Section, Department of Natural History, Royal Ontario Museum,Toronto, Canada; bPalaeobiology Section, Department of Natural History, Royal OntarioMuseum, Toronto, Canada; cMcDaniel Photography, Vancouver, Canada
(Received 3 July 2014; accepted 16 October 2014)
The hydroid Similiclava nivea, gen. nov., sp. nov., is described from coloniescollected in nearshore waters of southern British Columbia, Canada. It has beenobserved by divers, and recorded as Clava sp., several hundred times at locationsbetween southeast Alaska and southern Oregon, USA. While resembling thehydractiniid Clava multicornis, tentacles of S. nivea are arranged in two closewhorls in extended hydranths rather than being scattered over the distal end ofthe hydranth. Moreover, the hypostome is dome-shaped to flattened instead ofbeing proboscidiform. Similiclava nivea also resembles species of the hydractiniidgenera Stylactaria, Hydractinia and Podocoryna, but differs in being mono-morphic, in lacking spines on the hydrorhiza and in having numerous (as manyas 50 or more) sporosacs borne in a broad whorl below the tentacles on normalhydranths. Molecular analysis using the mitochondrial 16S gene sequences revealsthat S. nivea is closer to Clathrozoellidae than to Hydractiniidae, although it isclearly distinct from both. A new family, Similiclavidae, is established to accom-modate the species. Hydroids of S. nivea were observed at relatively shallowdepths (10–30 m) on rocks and epibiota attached to rocky substrates. Coloniesare conspicuous because of the relatively large surface area they occupy (up to20 cm across, and in aggregations often exceeding 100 or more hydranths) togetherwith the height (about 15 mm high when extended) and striking white colour ofliving hydranths. The cnidome consists of desmonemes, microbasic euryteles andmicrobasic mastigophores. A substrate generalist observed on rocks, algae,sponges, barnacles, bryozoans and other hydroids in cold-temperate waters,S. nivea is most prevalent in shallow subtidal areas swept by tidal currents.
http://zoobank.org/urn:lsid:zoobank.org:pub:2691E02E-7E14-4820-8D91-41D9E7E8BF62
Keywords: invertebrates; marine biology; natural history; taxonomy; zoology
Introduction
Current knowledge of the hydroid fauna of the Pacific coast of Canada and theUnited States is based largely on collections made during the late nineteenth and earlytwentieth centuries. Reviews of those sampling efforts, and a taxonomic account ofthe fauna as then known, have been given by Nutting (1901) and Fraser (1937). While
*Corresponding author. Email: [email protected]†Present address: Fairbanks Museum and Planetarium, 1302 Main Street, St. Johnsbury,Vermont 05819, USA
Journal of Natural History, 2015http://dx.doi.org/10.1080/00222933.2014.979261
© 2015 Taylor & Francis
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

hydroids of the region appear to be moderately well known from these two mono-graphs, recent evidence suggests that much taxonomic work remains to be done. Theexistence of knowledge gaps is revealed by discoveries of more than a dozen newspecies in the region within the past few decades (Brinckmann-Voss 1980, 1989, 2000;Cairns 1991; Brinckmann-Voss et al. 1993; Hewitt and Goddard 2001; Miglietta2006; Schuchert and Reiswig 2006; Brinckmann-Voss and Lindner 2008; Cairnsand Lindner 2011; Choong et al. 2012; Brinckmann-Voss and Calder 2013).
An account is provided here of an undescribed family, genus and species ofanthoathecate hydroid that has been observed by one of us (NM) over a period ofmore than four decades in waters of British Columbia (BC), Canada. The species wasfirst noticed in 1971, when photographs were taken of a colony in Howe Sound, nearthe city of Vancouver. It was photographed again in 1974 (Porlier Pass, Strait ofGeorgia), 1976 (Sutton Islands, Jervis Inlet, NW of Vancouver), 1978 (Howe Sound)and 2014 (Kuldekduma Point, Broughton Strait; Sakinaw Rock, Sechelt Inlet;Prominent Point, Knight Inlet; Ball Point, Jervis Inlet; Althorp Point, WellboreChannel). From the observations of a number of divers, it appears to be bothcommon and widespread in nearshore waters along the west coast of NorthAmerica from southeastern Alaska to southern Oregon.
Materials and methods
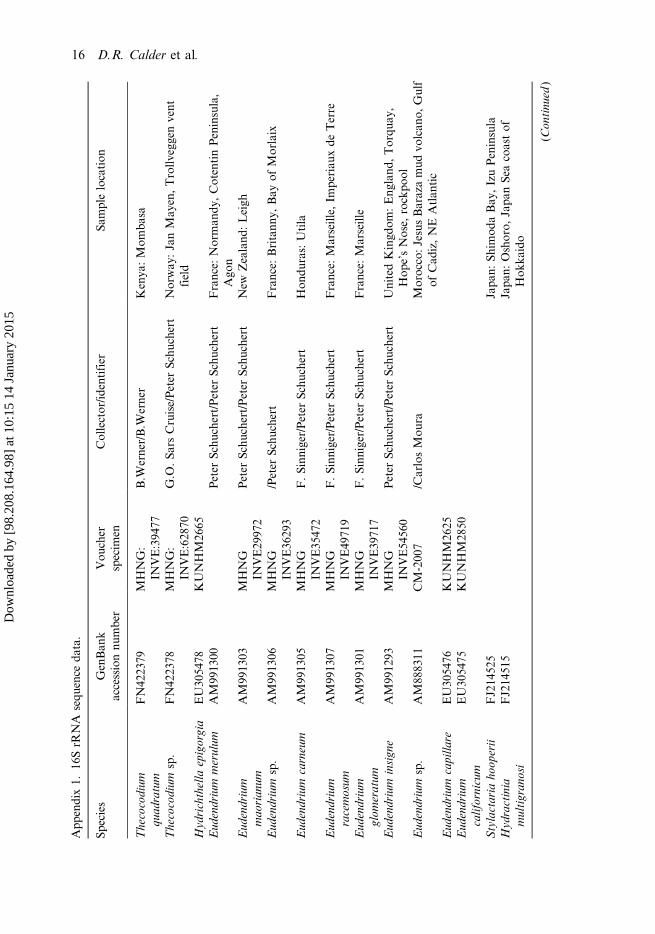
Specimens examined here were collected in southern BC during March, April andMay 2014. Materials for morphological study were fixed in formalin and latertransferred to 75% ethanol. Those intended for molecular analyses were fixed andpreserved in 95% ethanol. Type material has been deposited in collections of theInvertebrate Zoology Section, Department of Natural History, Royal OntarioMuseum (ROM). 16S rRNA sequence data are deposited in GenBank (accessionnumber KM099316).
DNA was extracted using 50:49:1 phenol:chloroform:isoamyl alcohol and precipi-tated with ethanol (Sambrook et al. 1989). Polymerase chain reaction (PCR) amplifica-tion and sequencing followed the protocol outlined in Moura et al. (2008). To amplifyc. 600 base pairs of the mitochondrial 16S gene, we used the primers SHA(ACGGAATGAACTCAAATCATGT) and SHB (TCGACTGTTTACCAAAAACATA) (Cunningham and Buss 1993; Moura et al. 2011). Forty additional 16Ssequences of hydroids, representing nine families, were obtained from GenBank(Appendix 1). Sequences were aligned using MAFFT version 7 (Katoh et al. 2002)with default alignment parameters. The maximum composite likelihood (MCL) methodinMEGA version 6.06 (Tamura et al. 2013) employing the general time reversible model(Nei andKumar 2000) was used to perform phylogenetic analyses.MEGAwas also usedto calculate pairwise distances for comparisons of sequence divergence between taxa.Sertularia unguiculata (family Sertulariidae) was used as the outgroup for the analyses. Inall, 1000 bootstrap replications were run. The tree with the highest log likelihood (–4760.4222) was used. The percentage of trees in which the associated taxa clusteredtogether is shown next to the branches. Initial tree(s) for the heuristic search wereobtained by applying the neighbour-joining method to a matrix of pairwise distancesestimated using the MCL approach. The analysis involved 41 nucleotide sequences. Allpositions containing gaps and missing data were eliminated. A total of 456 positionsexisted in the final dataset.
2 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Systematic account
Phylum CNIDARIA Verrill, 1865Subphylum MEDUSOZOA Petersen, 1979
Class HYDROZOA Owen, 1843Subclass HYDROIDOLINA Collins, 2000
Order ANTHOATHECATA Cornelius, 1992Suborder FILIFERA Kühn, 1913
Family SIMILICLAVIDAE fam. nov.
Diagnosis
Filiferan hydroids with strictly stolonal colonies. Hydrorhiza consisting of a creepingstolon with branches and anastomoses, becoming a dense network; coenosarcal tubesof stolon encased in a sheath of perisarc, not encrusting, or calcareous or coveredwith naked coenosarc. Hydrorhizal spines and nematophores absent. Zooids mono-morphic, consisting of sessile hydranths encased basally in a short, wrinkled sheath ofperisarc, otherwise naked. Hydranths of a colony tending to be separated rather thanclustered in a dense clump, sessile, highly contractile, elongate and nearly cylindricalwhen extended, with a slight to distinct bulbous area below a somewhat constrictedneck region, bulbous again distally at insertion of tentacles; without carbonategranules in the gastrodermis; tentacles filiform, in two very closely-placed whorls,appearing as a single whorl in contracted hydranths; hypostome dome-shaped.
Gonophores fixed and rounded sporosacs without traces of radial canals, ringcanal or tentacles, borne on unmodified hydranths, arising in grape-like clusters onhydranth column from a distal band beneath tentacular whorl.
Genus Similiclava gen. nov.
Diagnosis
With characters of the family.
Type species
Similiclava nivea sp. nov., designated herein.
Etymology
The name Similiclava is derived from a combination of the Latin word ‘similis’(resembling, like, similar) and the genus name Clava. The gender of the name isfeminine.
Remarks
The morphology of Similiclava conforms in many respects with diagnoses of thehydrozoan family Hydractiniidae L. Agassiz, 1862 (Bouillon et al. 2006; Schuchert
Journal of Natural History 3
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

2008; Calder 2010), and the hydroid examined here was initially thought to be ahydractiniid. Colonies are stolonal, with hydranths arising from a creeping hydro-rhiza that is enveloped in perisarc. Hydranths are naked except at their base, whereprotection is afforded by a cone of perisarc. Tentacles are filiform and amphicor-onate, and while arranged in two close whorls in extended hydranths, they mayappear to occur in a single whorl in contracted hydranths. Unlike in most hydracti-niids, zooids are monomorphic and spines of chitin or calcium carbonate are absent.There are no carbonate granules in the gastrodermis. Gonophores are fixed sporosacsthat arise in a broad band on normal hydranths below the tentacles.
Amongst genera currently assigned to Hydractiniidae, Similiclava most closelyresembles Clava Gmelin, 1790 in being monomorphic, in lacking chitinous or calcar-eous spines, in having numerous small rounded gonophores that arise in a bandaround the body of the hydranth below the tentacles and in having a hydrorhiza thatis creeping and completely covered with perisarc. Unlike in Clava, the tentacles arearranged in two close whorls (these appearing to comprise a single whorl in con-tracted hydranths) rather than being scattered over the distal end of the hydranth,and the hypostome is dome-shaped rather than being proboscidiform. It differs mostimmediately from the widely overlooked Hydrodendrium Nutting, 1905, anothermonomorphic hydractiniid genus (Calder 2010), in lacking a massive, spongy, arbor-escent hydrocaulus. Colonies of Similiclava differ from those of the more widelyknown hydractiniid genera Hydractinia Van Beneden, 1844, Podocoryna M. Sars,1846, Clavactinia Thornely, 1904, Hydrissa Stechow, 1921a, Stylactaria Stechow,1921a, Hydrocorella Stechow, 1921b, Janaria Stechow, 1921b, BouillonactiniaMiglietta et al. 2010, and Schuchertinia Miglietta et al. 2010 in being monomorphicrather than polymorphic, in lacking hydrorhizal spines and in having numerous smallgonophores (as sporosacs) that are borne on normal hydranths. The hydroid alsolacks a calcareous skeleton, found in many species within several genera included inHydractiniidae. Molecular analysis using the mitochondrial 16S gene indicates thatSimiliclava and its type species S. nivea are closer to Clathrozoellidae than toHydractiniidae, although it is clearly distinct from both.
Similiclava nivea sp. nov.Figures 1–3, 5
Clava sp. Lamb and Hanby, 2005: 101, fig. CN74.
Material examined
Holotype. Sakinaw Rock, Sechelt Inlet, British Columbia, Canada (49°34.075’N,123°48.238’W), depth 20 m, 7 March 2014, hydroid colony overgrowing algae,sponge tissue, and hydroid stems from a rocky substrate, some hydranths with femalegonophores, coll. Neil McDaniel, preserved in 95% ethanol, ROMIZ B4021.
Paratypes. Sakinaw Rock, Sechelt Inlet, British Columbia, Canada (49°34.075’N,123°48.238’W), depth 20 m, 8 April 2014, on algae, sponge tissue, and hydroid stemsfrom a rocky substrate, some hydranths with male gonophores, coll. Neil McDaniel,preserved in 95% ethanol, ROMIZ B4022.—Prominent Point, Knight Inlet, BritishColumbia, Canada (50°40.052’N, 126°00.557’W), depth 15 m, 3 May 2014, on
4 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

sponge, barnacle shell and bryozoan colony, some hydranths with male gonophores,coll. Neil McDaniel, preserved initially in formalin, now in 75% ethanol, ROMIZB4023.—Ball Point, Jervis Inlet, British Columbia, Canada (49°45.086’N, 124°13.426’W), depth 15 m, 6 May 2014, on barnacle shell, sponge and hydroid stem,some hydranths with male gonophores, coll. Neil McDaniel, preserved initially informalin, now in 75% ethanol, ROMIZ B4024.—Sakinaw Rock, Sechelt Inlet, BritishColumbia, Canada (49°34.075’N, 123°48.238’W), depth 20 m, 8 May 2014, on algae,hydroids and sponge, some hydranths with female gonophores, coll. Neil McDaniel,preserved initially in formalin, now in 75% ethanol, ROMIZ B4025.
Etymology
The specific name is the Latin adjective nivea (snowy, white, snow-white) in referenceto the striking snowy colour of hydranths and colonies.
Description
Colonies stolonal, monomorphic, with hydranths arising from a hydrorhiza creepingover rocks and epibiota on rocky substrates. Aggregations often comprising 100 ormore hydranths, covering several square centimetres of substrate. Hydroids thusconspicuous in nature, especially given the rather large size and snowy white colourof extended hydranths. Stolons of hydrorhiza branching and anastomosing, forming a
Figure 1. Preserved hydranths of Similiclava nivea, holotype colony, ROMIZ B4021; (A)sterile hydranth; (B) fertile hydranth with female gonophores. Scale bars equal 0.5 mm.
Journal of Natural History 5
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

dense and tangled network in older colonies, completely covered with mostly smoothperisarc; spines absent; nematophores absent; naked coenosarc absent. Hydranthslarge, 2–4 mm high and 1 mm or more wide when contracted, to 15 mm high or morewhen extended, usually somewhat separated rather than clumped together, protectedbasally by a wrinkled and creased collar of perisarc; perisarcal collar of varied lengthbut usually rather short, 1–2 mm or less, covering only basalmost part of hydranth,
Figure 2. Photographs of living colonies of Similiclava nivea: (A) diffuse colony, with mosthydranths sterile, Sakinaw Rock, Sechelt Inlet, BC, 7 March 2014; (B) small colony, with a fewfertile hydranths, Sakinaw Rock, Sechelt Inlet, BC, 7 March 2014, holotype, ROMIZ B4021;(C) dense colony, with some fertile hydranths, Sutton Islands, Jervis Inlet, BC, January 1976;(D) close-up of hydranths, some with well-developed gonophores, Prominent Point, KnightInlet, BC, 3 May 2014; (E) regressing hydranths, most with gonophores, Sakinaw Rock,Sechelt Inlet, BC, 13 July 2014; (F) renewed growth of colony, with small, sterile, juvenilehydranths, Sakinaw Rock, Sechelt Inlet, BC, 20 August 2014; note stolons of hydrorhiza. Allphotographs by Neil McDaniel.
6 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

tapering from its expanded distal end to a more slender base at insertion with stolon,but even there usually still much broader than diameter of stolon; hydranth columnhighly contractile, lacking wart-like batteries of nematocysts, sac-shaped to bulbousto pyriform when fully contracted, mostly cylindrical when extended but with a slightto sometimes distinct bulge on distal half of column followed by a constricted neckregion below tentacular whorl, then becoming bulbous again at level of tentacularring; hypostome dome-shaped to almost flattened, lacking a refringent band ofnematocysts around mouth; tentacles filiform, about 15–20 in number in fully devel-oped hydranths, tapering gradually from base to tip, alternating in two very closewhorls in extended hydranths, with those of upper whorl usually reaching upwardsand those of lower whorl most commonly directed outwards or somewhat down-wards, appearing to be arranged in a single whorl in contracted hydranths.
Gonophores fixed sporosacs, borne on normal hydranths just beneath tentacles,translucent white to pale peach in colour; colonies unisexual. Sporosacs sessile, small,nearly spherical to oviform to pyriform to reniform, with a spadix but lacking radialcanals, tentacles and ring canal. Fertile hydranths initially with one or a few smallgonophores, these later becoming numerous (as many as 50 or more) and aggregatedinto a dense contiguous band of sporosacs of various sizes and degrees of maturity,resembling clusters of grapes or coconuts; smaller and younger gonophores usuallyarising proximal to larger and more fully developed ones; as gonophores develop andincrease in number, hydranths bearing them tend to remain more contracted in life
Figure 3. Nematocysts of the hydranth of Similiclava nivea, holotype colony, ROMIZ B4021:(A) two desmonemes; (B) microbasic eurytele; (C) microbasic mastigophore; (D) two desmo-nemes, discharged; (E) microbasic eurytele, discharged; (F) microbasic mastigophore, dis-charged; only the shaft of the thread is visible. For nematocyst sizes, see text.
Journal of Natural History 7
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

than sterile ones. Female sporosacs producing a single egg; hydroids larviparous, withegg developing into a planula within the gonophore. Male sporosacs during develop-ment with a digitate central spadix surrounded by a spermatic mass.
Nematocyst complement (Figure 3)Desmonemes (n = 12): 5.5–7.0 μm long × 4.0–4.6 μm wide (undischarged)Microbasic euryteles (n = 15): 9.2–10.6 μm long × 4.4–5.7 μm wide (undischarged)Microbasic mastigophores (n = 12): 17.5–20.1 μm long × 7.2–8.8 μm wide(undischarged)
Remarks
Based on morphology, Similiclava nivea is assigned to order AnthoathecataCornelius, 1992, and to suborder Filifera Kühn, 1913, and it most closely resemblesClava multicornis (Forsskål 1775) [see Schuchert (2008) for a recent description]. Inparticular, hydranths with dense clusters of sporosacs occurring below the tentacularwhorl appear much alike in the two species. However, hydranths of S. nivea caneasily be distinguished in having tentacles in no more than two close whorls instead ofbeing scattered. Also different is the appearance of the hypostome, which is dome-shaped to flattened rather than being proboscis-shaped. Other differences exist inhabitat, with C. multicornis being mostly intertidal and S. nivea being predominantlysubtidal, and in colour, with C. multicornis being variously cream to pink to reddishto salmon-coloured and S. nivea being snowy white. Given its colour, an appropriatecommon name applied to S. nivea is the ‘white hydroid’ (Lamb and Hanby 2005,p. 101, as Clava sp.). As noted by Lamb and Hanby, ‘Often this hydroid grows sodensely that it mimics a “dusting of snow”’.
Hydranths with well-developed gonophores appear little different morphologi-cally than those without, although photographs of living colonies reveal that they arefrequently more contracted (Figure 2B–D). Greater protection may thereby beafforded the reproductive structures beneath a canopy provided by adjacent andmore fully extended sterile hydranths.
Of three kinds of nematocysts observed on hydranths of the species, microbasiceuryteles were most abundant, followed by desmonemes and finally by microbasicmastigophores (Figure 3A–F). The cnidome of Similiclava nivea is thus typical of thesuborder Filifera.Microbasic mastigophores were larger than those seen in hydractiniids,and they were distinctive in shape. An oval to nearly spherical bulb was consistentlypresent just beneath the operculum in the undischarged nematocyst (Figure 3C), and thisdilation was apparent at the base of the shaft in discharged ones (Figure 3F). Thedischarged thread somewhat resembles that seen in pseudostenoteles, but these largenematocysts are nevertheless regarded here as microbasic mastigophores.
No granules of calcium carbonate were observed in hydranths of Similiclavanivea. This was to be expected, inasmuch as there is no calcification of the skeletonanywhere in this species.
Phylogenetic analysis
As with its morphological characters, molecular analyses are consistent in placingSimiliclava nivea within the component subgroup Filifera of the order Anthoathecata.
8 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Nodes within the major clades exhibit strong bootstrap values and are consistent withprevious analyses (Cartwright et al. 2008; Peña Cantero et al. 2010). Species groupedin distinct clusters with high support in the 16S tree. 16S has been shown to be apowerful phylogenetic marker in delineating species boundaries for marine hydroids,and has also been demonstrated to be more successful than the mitochondriallyencoded cytochrome c oxidase I (COI) gene in extrapolating phylogenetic relation-ships of hydroids at the generic level, and in some cases at the family level (Migliettaet al. 2007, 2009; Moura et al. 2008, 2011; Peña Cantero et al. 2010; Zheng et al.2014). Our findings are also consistent with previous work supporting the non-monophyly of the Anthoathecata and Filifera (Cartwright et al. 2008). The topologyof the 16S rRNA tree (Figure 4) reveals that the phylogenetic position of S. nivea isnot within Hydractiniidae. There is strong bootstrap support for situating the specieswithin a separate clade (Similiclavidae fam. nov.) as a sister group to ClathrozoellidaePeña Cantero et al. 2003, within the monophyletic clade formed by Eudendriidae L.Agassiz, 1862 (maximum likelihood (ML) bootstrap values = 88 and 77). Between-group distances indicate considerable divergence between Similiclavidae and othertaxa (Figure 4; Appendix 2). Morphologically, this new family is distinct fromClathrozoellidae in having stolonal rather than erect colonies, in lacking an exoske-leton of adherent perisarcal stolons with a core of coenosarc, in lacking nematothecaeand in having gonophores directly on the hydranth rather than next to it at the baseof a pseudohydrotheca. Our molecular results support the inclusion of ClathrozoellaStechow, 1921a (cf. Peña Cantero et al. 2003) within Anthoathecata, given thepresence in both groups of unprotected gonophores and a cnidome comprised ofdesmonemes, microbasic euryteles and microbasic mastigophores.
Distribution
Specimens or photographs, or both, of Similiclava nivea examined during this studydocument the occurrence of the species in waters of BC from Kuldekduma Point,Broughton Strait, southwards to Porlier Pass, Strait of Georgia, at depths from 10–30 m. A white anthoathecate hydroid identified by Lamb and Hanby (2005) as Clavasp., regarded by us as conspecific with S. nivea, was reported to occur from BC tosouthern California. Meanwhile, its bathymetric distribution was given in their accountas ‘intertidal to subtidal’. Reports of this hydroid (Andy Lamb and Marc Chamberlain,personal communication, May 2014) now extend northwards to the Inian Islands (58°14’N, 136°21’W), Icy Strait, southeast Alaska, and southwards to Island Rock nearPort Orford, southern Oregon. Records from locations further south, including thosefrom southern California mentioned in Lamb and Hanby (2005), are now taken to beerroneous (Andy Lamb, personal communication, 3 June 2014).
Similiclava nivea is of surprisingly frequent occurrence within its known range. Atotal of 691 records of hydroids identified as Clava sp. (=Similiclava nivea) exist in anunpublished database provided to us by Andy Lamb (personal communication, 28 June2014), based onmore than 40 years of diving in the region. Among them are reports (withnumbers of observations in parentheses) from the following locations in BritishColumbia: Burrard Inlet (four), Central Coast (seven), Discovery Passage (44), GulfIslands (257), Haro Strait (one), Howe Sound (94), Johnstone Strait (65), QueenCharlotte Islands (Haida Gwaii) (three), Queen Charlotte Strait (10), Seymour Inlet(six), Strait of Georgia (36), Strait of Juan de Fuca (one), Sunshine Coast (98) and the
Journal of Natural History 9
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Figure 4. Condensed results of the maximum likelihood (ML) analysis (general time reversiblemodel and 1000 pseudoreplicates). Bootstrap values higher than 50 are shown.
10 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

outer coast of Vancouver Island (four). Lamb’s database also records the species atlocations in southern Alaska (14), Washington (40), and Oregon (one), USA.
Hydroids identified as Clava multiformis (a misspelling of C. multicornis) fromAlaska (Port Dick, Kenai Peninsula, 59°13.3’N, 151°10.0’W, 22 May 1975, intertidal,coll. J. Gnagy, California Academy of Sciences, Invertebrate Zoology, CAS:IZ:13160) were examined to determine whether they too might be Similiclava nivea.While the specimens were fragmentary and in poor condition, no large microbasicmastigophore nematocysts like those of S. nivea were observed. We conclude thatthey are referable to another species.
Ecology
A focused search for Similiclava nivea at three locations in coastal BC was under-taken by one of us (NM) during May 2014. Observations were made on 3 May atProminent Point, Knight Inlet, BC. Knight Inlet is a fjord extending inland some120 km from its entrance in Queen Charlotte Strait, and Prominent Point is adjacentto its sill. Conspicuous colonies of large hydranths (up to 15 mm high) were abundantthere at depths of about 10–25 m along a steep rocky dropoff. Irregular patches up to20 cm in diameter were noted on bedrock. Prominent Point is swept by modest tomoderate tidal currents, and the species appears to thrive under such conditions. AtBall Point, Jervis Inlet, BC, on 6 May, Similiclava nivea was found to be relativelyabundant on a steep rocky dropoff at depths of 10–20 m. This site is also impacted bymodest tidal currents. Finally, specimens were collected on 8 May at a depth of 20 mfrom Sakinaw Rock, Sechelt Inlet, BC, a site with a bottom of sloping rock that isswept by modest tidal currents. Pauline Ridings (personal communication, 14 May2014) reports having observed the species at most of the high-current locales whereshe has dived in BC, including Work Channel, Watts Narrows in Grenville Channel,Petrel Channel, Beaver Passage (close to Kitkatla), around Port Hardy, DoddNarrows, Steep Island (Campbell River) and the Stubbs Island area.
Similiclava nivea is a substrate generalist. Collected material has been observed onalgae, sponges, barnacle shells, a bryozoan colony and stems of other hydroids. It hasalso been observed to grow on bedrock and boulders.
In terms of seasonality, active colonies have been observed and photographed inwaters of BC during the months of January (1974, 1976, 2014), March (2014), April(1978, 2014), May (2014), June (2014), July (2014), August (2014), September (2013,2014), and November (1971). During each of those months, gonophores were dis-cernible on at least some of the hydranths. Of particular note, colonies monitored atSakinaw Rock, Sechelt Inlet, declined significantly in size and vigour during earlysummer (13 July 2014) at water temperatures above 11°C, with the few remaininghydranths fertile but much regressed (Figure 2E). Later in summer (20 August 2014)at the same location, at a water temperature of 13.1°C, mostly small, sterile, juvenilehydranths were seen (Figure 2F). By contrast, colonies observed during the samemonth (30 August 2014) under colder conditions (8.2°C) at Prominent Point inKnight Inlet appeared normal in size and overall appearance, with both fertile andinfertile hydranths (observations and photographs by NM). Meanwhile, the hydroididentified as Clava sp. (= Similiclava nivea) has been recorded throughout the year inthe northeast North Pacific marine life database of Andy Lamb (personal commu-nication, 28 June 2014). From its known distribution, and from the phenology of the
Journal of Natural History 11
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

species as observed at Sakinaw Rock during 2014, we conclude that S. nivea is aspecies restricted to cold-temperate waters.
The nudibranch Flabellina japonica has been identified as a predator ofSimiliclava nivea in nature. Specimens of that species were observed on colonies ofS. nivea in two photographs from the Sutton Islands, Skookumchuck Narrows, JervisInlet, BC, on 7 January 1976 (Figure 5A, B), and others have been observed amongstcolonies of the hydroid at three other locations in the province. Our suspicion thatF. japonica preys on the hydroid heretofore known as Clava sp. was confirmed bynudibranch specialist Sandra V. Millen (personal communication, 8 July 2014). Inanother photograph of S. nivea, a small nudibranch (Doto sp.) and a stenothoidamphipod were seen amongst its hydranths on 3 May 2014 at Prominent Point,Knight Inlet, BC, but there was no evidence that either was grazing on the species.On 1 June 2014, a specimen of Flabellina trophina, a nudibranch that preys on otherspecies of hydroids, was placed (by NM) on a colony of S. nivea at Sakinaw Rock,Sechelt Inlet, BC (Figure 5C) to observe its behaviour. Its reaction was stronglynegative, and the nudibranch moved rapidly away from the hydroid. The extent towhich S. nivea is venomous remains to be determined, although the large microbasicmastigophore nematocysts observed in the species (Figure 3C, F) may afford protec-tion from many potential predators.
Geolocation informationSimiliclava nivea. ROMIZ B4021 (point): 49°34’4”N, 123°48’14”W; ROMIZ B4022 (point):49°34’4”N, 123°48’14”W; ROMIZ B4023 (point): 50°40’3”N, 126°0’33”W; ROMIZ B4024(point): 49°45’5”N, 124°13’25”W; ROMIZ B4025 (point): 49°34’4”N, 123°48’14”W.
Acknowledgements
We are grateful to Andy Lamb, and to Donna and Charlie Gibbs, for providing access to theirlarge database on the white hydroid (as Clava sp.). Marc Chamberlain provided the northern-most sighting of the species, at the Inian Islands, southeast Alaska. Rick Harbo, Jan Kocian,Janna Nichols and Pauline Ridings also provided sighting records. Particular thanks are due to
Figure 5. Nudibranchs on the hydroid Similiclava nivea, from nature: (A) Flabellina japonica,observed naturally on a hydroid colony, Sutton Islands, Jervis Inlet, BC, 7 January 1976;(B) Flabellina japonica, observed naturally on a hydroid colony, Sutton Islands, Jervis Inlet,BC, 7 January 1976; (C) Flabellina trophina, placed intentionally (by NM) on a hydroid colony,Sakinaw Rock, Sechelt Inlet, BC, 1 June 2014; the nudibranch reacted negatively (see text). Allphotographs by Neil McDaniel.
12 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Oliver Haddrath of the ROM for help with the molecular analysis. Thanks are also due toKristen Choffe and Maureen Zubowski of the ROM, for assistance in the Laboratory ofMolecular Systematics and in the Invertebrate Zoology Section, respectively. ChristinaPiotrowski and Elizabeth Kools of the California Academy of Sciences provided a loan of ahydroid species from Alaska (CAS:IZ:13160) that we compared with our material. SandraMillen confirmed that the nudibranch Flabellina japonica preys on Similiclava nivea.
References
Agassiz L. 1862. Contributions to the natural history of the United States of America. Vol. IV.Boston (MA): Little, Brown.
Bouillon J, Gravili C, Pagès F, Gili J-M, Boero F. 2006. An introduction to Hydrozoa. MemMus Nat Hist Nat Paris. 194:1–591.
Brinckmann-Voss A. 1980. A new species of the genus Sarsia (Hydrozoa, Corynidae) fromVancouver Island and Puget Sound. Roy Ont Mus Life Sci Occ Pap. 34:1–4.
Brinckmann-Voss A. 1989. Sarsia cliffordi n.sp. (Cnidaria, Hydrozoa, Anthomedusae) fromBritish Columbia, with distribution records and evaluation of related species. Can J Zool.67:685–691. doi:10.1139/z89-099
Brinckmann-Voss A. 2000. The hydroid and medusa of Sarsia bella sp. nov. (Hydrozoa,Anthoathecatae, Corynidae), with a correction of the “life cycle” of Polyorchis penicillatus(Eschscholtz). Sci Mar. 64:189–195.
Brinckmann-Voss A, Calder DR. 2013. Zyzzyzus rubusidaeus (Cnidaria, Hydrozoa,Tubulariidae), a new species of anthoathecate hydroid from the coast of BritishColumbia, Canada. Zootaxa. 3666:389–397. doi:10.11646/zootaxa.3666.3.9
Brinckmann-Voss A, Lickey ADM, Mills CE. 1993. Rhysia fletcheri (Cnidaria, Hydrozoa,Rhysiidae), a new species of colonial hydroid from Vancouver Island (British Columbia,Canada) and the San Juan Archipelago (Washington, U.S.A). Can J Zool. 71:401–406.
Brinckmann-Voss A, Lindner A. 2008. Monocoryne colonialis sp. nov., a colonial candelabridhydroid (Cnidaria: Hydrozoa: Candelabridae) from the North Pacific. J Mar Biol AssocUK. 88:1631–1635. doi:10.1017/S002531540800180X
Cairns SD. 1991. Cyclohelia lamellata, new genus and species of Stylasteridae (Cnidaria:Hydrozoa) from the Bering Sea. Pac Sci. 45:383–388.
Cairns SD, Lindner A. 2011. A revision of the Stylasteridae (Cnidaria, Hydrozoa, Filifera)from Alaska and adjacent waters. ZooKeys. 158:1–88.
Calder DR. 2010. Some anthoathecate hydroids and limnopolyps (Cnidaria, Hydrozoa) fromthe Hawaiian archipelago. Zootaxa. 2590:1–91.
Cartwright P, Evans NM, Dunn CW, Marques AC, Miglietta MP, Schuchert P, Collins AG.2008. Phylogenetics of Hydroidolina (Hydrozoa: Cnidaria). J Mar Biol Assoc UK.88:1663–1672. doi:10.1017/S0025315408002257
Choong HHC, Calder DR, Brinckmann-Voss A. 2012. Sertularella maureenae, a new species ofhydroid (Cnidaria: Hydrozoa: Sertulariidae) from the Pacific coast of Canada. Zootaxa.3297:57–63.
Collins AG. 2000. Towards understanding the phylogenetic history of Hydrozoa: hypothesistesting with 18S gene sequence data. Sci Mar. 64:5–22.
Cornelius PFS. 1992. Medusa loss in leptolid Hydrozoa (Cnidaria), hydroid rafting, andabbreviated life-cycles among their remote-island faunae: an interim review. Sci Mar.56:245–261.
Cunningham CW, Buss LW. 1993. Molecular evidence for multiple episodes of paedomorpho-sis in the family Hydractiniidae. Biochem Syst Ecol. 21:57–69. doi:10.1016/0305-1978(93)90009-G
Journal of Natural History 13
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Forsskål P. 1775. Descriptiones animalium avium, amphibiorum, piscium, insectorum, ver-mium; quae in itinere orientali observavit Petrus Forskål. Prof. Haun. Post mortemauctoris edidit Carsten Niebuhr. Hauniae: Mölleri.
Fraser CM. 1937. Hydroids of the Pacific coast of Canada and the United States. Toronto(ON): University of Toronto Press.
Gmelin JF. 1790. Caroli a Linné, systema naturae per regna tria naturae, secundum classes,ordines, genera, species, cum characteribus, differentiis, synonymis, locis. Editio decimatertia, aucta reformata. Tomus 1, Pars 6. Lipsiae: GE Beer.
Hewitt CL, Goddard JHR. 2001. A new species of large and highly contractile hydroid in thegenus Candelabrum (Hydrozoa: Anthoathecatae) from southern Oregon, U.S.A. Can JZool. 79:2280–2888. doi:10.1139/cjz-79-12-2280
Katoh K, Misawa K, Kuma K, Miyata T. 2002. MAFFT: a novel method for rapid multiplesequence alignment based on fast Fourier transform. Nucleic Acid Res. 30:3059–3066.doi:10.1093/nar/gkf436
Kühn A. 1913. Entwicklungsgeschichte und Verwandtschaftsbeziehungen der Hydrozoen. I.Teil: Die Hydroiden. Erg Fortschr Zool. 4:1–284.
Lamb A, Hanby BP. 2005. Marine life of the Pacific Northwest. A photographic encyclopediaof invertebrates, seaweeds and selected fishes. Madeira Park, BC: Harbour Publishing.
Miglietta MP. 2006. Hydractinia antonii sp. nov.: a new, partially calcified hydractiniid(Cnidaria: Hydrozoa: Hydractiniidae) from Alaska. J Mar Biol Assoc UK. 86:993–996.doi:10.1017/S0025315406013968
Miglietta MP, McNally L, Cunningham CW. 2010. Evolution of calcium-carbonate skeletonsin the Hydractiniidae. Integr Comp Biol. 50:428–435. doi:10.1093/icb/icq102
Miglietta MP, Piraino S, Kubota S, Schuchert P. 2007. Species in the genus Turritopsis(Cnidaria, Hydrozoa): a molecular evaluation. J Zool Syst Evol Res. 45:11–19.doi:10.1111/j.1439-0469.2006.00379.x
Miglietta MP, Schuchert P, Cunningham CW. 2009. Reconciling genealogical and morpholo-gical species in a worldwide study of the family Hydractiniidae (Cnidaria, Hydrozoa).Zool Scripta. 38:403–430. doi:10.1111/j.1463-6409.2008.00376.x
Moura CJ, Cunha MR, Porteiro FM, Rogers AD. 2011. The use of the DNA barcode gene16S mRNA for the clarification of taxonomic problems within the family Sertulariidae(Cnidaria, Hydrozoa). Zool Scripta. 40:520–537. doi:10.1111/j.1463-6409.2011.00489.x
Moura CJ, Harris DJ, Cunha MR, Rogers AD. 2008. DNA barcoding reveals cryptic diversityin marine hydroids (Cnidaria, Hydrozoa) from coastal and deep-sea environments. ZoolScripta. 37:93–108.
Nei M, Kumar S. 2000. Molecular evolution and phylogenetics. New York (NY): OxfordUniversity Press.
Nutting CC. 1901. Papers from the Harriman Alaska Expedition. XXI. The hydroids. ProcWash Acad Sci. 3:157–216.
Nutting CC. 1905. Hydroids of the Hawaiian Islands collected by the steamer Albatross in1902. Bull US Fish Comm. 1903:931–959.
Owen R. 1843. Lectures on the comparative anatomy and physiology of the invertebrateanimals: delivered at the Royal College of Surgeons, in 1843. London: Longman,Brown, Green, & Longmans.
Peña Cantero AL, Sentandreu V, Latorre A. 2010. Phylogenetic relationships of the endemicAntarctic benthic hydroids (Cnidaria, Hydrozoa): what does the mitochondrial 16S rRNAtell us about it? Polar Biol. 33:41–57. doi:10.1007/s00300-009-0683-5
Peña Cantero AL, Vervoort W, Watson J. 2003. On Clathrozoellidae (Cnidaria, Hydrozoa,Anthoathecatae), a new family of rare deep-water leptolids, with the description of threenew species. Zool Verh. 345:281–296.
14 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

Petersen KW. 1979. Development of coloniality in Hydrozoa. In: Larwood G, Rosen BR,editors. Biology and systematics of colonial animals. New York (NY): Academic Press; p.105–139.
Sambrook J, Fritsch EF, Maniatis T. 1989. Molecular cloning: a laboratory manual. 2nd edn.New York (NY): Cold Spring Harbor Laboratory.
Sars M. 1846. Fauna littoralis Norvegiae oder Beschreibung und Abbildungen neuer oderwenig bekannten Seethiere, nebst Beobachtungen über die Organisation, Lebensweise undEntwickelung derselben. Heft I. Christiania: Johann Dahl.
Schuchert P. 2008. The European athecate hydroids and their medusae (Hydrozoa, Cnidaria).Filifera Part 3. Rev Suisse Zool. 115:221–302.
Schuchert P, Reiswig HM. 2006. Brinckmannia hexactinellidophila, n. gen., n. sp.: a hydroidliving in tissues of glass sponges of the reefs, fjords, and seamounts of Pacific Canada andAlaska. Can J Zool. 84:564–572. doi:10.1139/z06-031
Stechow E. 1921a. Neue Genera und Species von Hydrozoen und anderen Evertebraten. ArchNaturgesch. 87:248–265.
Stechow E. 1921b. Neue Gruppen skelettbildender Hydrozoen und Verwandtschaftsbeziehungenrezenter und fossiler Formen. Verh Deut Zool Ges. 26:29–31.
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. 2013. MEGA6: Molecular evolu-tionary genetics analysis version 6.0. Mol Biol Evol. 30:2725–2729. doi:10.1093/molbev/mst197
Thornely LR. 1904. Report on the Hydroida collected by Professor Herdman, at Ceylon, in1902. Report to the Government of Ceylon on the pearl oyster fisheries of the Gulf ofManaar, with supplementary reports upon the marine biology of Ceylon. Part II:107–126.
Van Beneden P-J. 1844. Sur les genres Eleuthérie et Synhydre. Bull Acad Roy Sci Belles-LettBruxelles. 11:305–314.
Verrill AE. 1865. Classification of polyps: (extract condensed from a synopsis of the Polypi ofthe North Pacific Exploring Expedition, under Captains Ringgold and Rodgers, U.S.N.Proc Essex Inst. 4:145–152.
Zheng L, He J, Lin Y, Cao W, Zhang W. 2014. 16S rRNA is a better choice than COI forDNA barcoding hydrozoans in the coastal waters of China. Acta Oceanol Sin. 33:55–76.doi:10.1007/s13131-014-0415-8
Journal of Natural History 15
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15
App
endix1.
16SrR
NA
sequ
ence
data.
Species
GenBan
kaccessionnu
mber
Vou
cher
specim
enCollector/id
entifier
Samplelocation
Thecocodium
quad
ratum
FN42
2379
MHNG:
INVE:394
77B.W
erner/B.W
erner
Kenya
:Mom
basa
Thecocodium
sp.
FN42
2378
MHNG:
INVE:628
70G.O
.Sa
rsCruise/Peter
Schu
chert
Norway
:JanMay
en,Trollv
eggenvent
field
Hyd
richthella
epigorgia
EU30
5478
KUNHM26
65Eud
endrium
merulum
AM99
1300
Peter
Schu
chert/Peter
Schu
chert
France:
Norman
dy,CotentinPeninsula,
Ago
nEud
endrium
mao
rian
umAM99
1303
MHNG
INVE29
972
Peter
Schu
chert/Peter
Schu
chert
New
Zealand
:Leigh
Eud
endrium
sp.
AM99
1306
MHNG
INVE36
293
/Peter
Schu
chert
France:
Britann
y,Bay
ofMorlaix
Eud
endrium
carneum
AM99
1305
MHNG
INVE35
472
F.Sinn
iger/Peter
Schu
chert
Hon
duras:Utila
Eud
endrium
racemosum
AM99
1307
MHNG
INVE49
719
F.Sinn
iger/Peter
Schu
chert
France:
Marseille,
Imperiau
xde
Terre
Eud
endrium
glom
eratum
AM99
1301
MHNG
INVE39
717
F.Sinn
iger/Peter
Schu
chert
France:
Marseille
Eud
endrium
insign
eAM99
1293
MHNG
INVE54
560
Peter
Schu
chert/Peter
Schu
chert
UnitedKingd
om:Eng
land
,Torqu
ay,
Hop
e’sNose,
rockpo
olEud
endrium
sp.
AM88
8311
CM-200
7/CarlosMou
raMorocco:Jesus
Barazamud
volcan
o,Gulf
ofCad
iz,NEAtlan
tic
Eud
endrium
capilla
reEU30
5476
KUNHM26
25Eud
endrium
californicum
EU30
5475
KUNHM28
50
Stylactaria
hoop
erii
FJ214
525
Japa
n:Sh
imod
aBay
,IzuPeninsula
Hyd
ractinia
multigran
osi
FJ214
515
Japa
n:Oshoro,
Japa
nSeacoastof
Hok
kaido
(Con
tinu
ed)
16 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

App
endix1.
(Con
tinu
ed).
Stylactaria
inermis
FJ214
546
Italy:
Otran
to,Apu
liaStylactaria
sp.
FJ214
494
Japa
n:Kom
inato,
BosoPeninsula
Clava
multicornis
FJ214
439
Icelan
d:Sa
ngerdi
vicinity
Clava
multicornis
EU30
5471
France:
Roscoff
Hyd
ractinia
fucicola
FJ214
437
Italy:
Torre
delSerpe,
Otran
to,Apu
liaPod
ocoryn
aam
erican
aFJ214
449
USA
:Woo
dsHole,
MA
Pod
ocoryn
aareolata
AM93
9651
MHNG
INVE48
749
Peter
Schu
chert/Peter
Schu
chert
Norway
:Fan
afjord
Hyd
ractinia
conchicola
FJ214
435
Japa
n:Oshoro,
Japa
nSeacoastof
Hok
kaido
Hyd
ractinia
sp.
FJ214
379
Hyd
ractinia
sp.
EU30
5477
KUNHM28
76Hyd
ractinia
serrata
FJ214
597
USA
:Frida
yHarbo
r,WA
Errinop
orana
nneca
EU64
5358
USN
M10
2782
0Errinop
oraun
dulata
JN57
2434
USN
M11
2352
7ho
lotype
Errinop
oradisticha
JN57
2438
USN
M11
2352
4ho
lotype
Hyd
richthys
boycei
EU44
8102
MHNG
INVE37
417
Lafoeadu
mosa
FN42
4137
/AlvaroL.PeñaCan
tero
Antarctica:
Antarctic
Peninsula
Lafoeasp.
AM88
8327
/CarlosMou
raMorocco:Gulfof
Cad
iz,NEAtlan
tic
Filellu
mcf
serratum
JN71
4660
MercatorMV,Stn.
B09
-01W
out,35
4m,
Gulfof
Cad
izSertulariaun
guiculata
AY78
7904
MHNG
INVE29
969
New
Zealand
:Leigh
Sym
plectoscyp
hus
exochu
sFN42
4147
/AlvaroL.PeñaCan
tero
Antarctica:
Low
Island
(Con
tinu
ed)
Journal of Natural History 17
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

App
endix1.
(Con
tinu
ed).
Species
GenBan
kaccessionnu
mber
Vou
cher
specim
enCollector/id
entifier
Samplelocation
Sym
plectoscyp
hus
plectilis
FN42
4150
/AlvaroL.PeñaCan
tero
Antarctica:
Deception
Island
Antarctoscyph
usgran
dis
FN42
4139
/AlvaroL.PeñaCan
tero
Antarctica:
RossSea
Halecium
incertum
FN42
4124
/AlvaroL.PeñaCan
tero
Antarctica:
RossSea
Halecium
muricatum
AM88
8318
/CarlosMou
raUnitedKingd
om:Lun
dyIsland
,Devon
Clathrozoella
dryg
alskii
FN42
4154
/AlvaroL.PeñaCan
tero
Antarctica:
RossSea
Sim
iliclavanivea
KM09
9316
ROMIZ
B40
22pa
ratype
NeilMcD
aniel/D
aleR.Calder,
Henry
H.C.Cho
ong
Can
ada:
British
Colum
bia,
Sakina
wRock,
SecheltInlet
18 D.R. Calder et al.
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15

App
endix
2.Pairw
isedistan
cesbetw
een
grou
psusing
themax
imum
compo
site
likelihoo
dmod
el.Stan
dard
errorestimate(s)
show
nab
ovethe
diag
onal.
12
34
56
78
910
1.Eud
endriid
ae0.01
60.01
90.01
50.01
90.01
80.01
60.01
70.01
60.01
52.
Hyd
ractiniid
ae0.16
40.01
40.01
50.01
70.01
80.01
70.01
80.01
70.01
93.
Stylasterida
e0.170
0.118
0.017
0.021
0.020
0.019
0.020
0.020
0.021
4.Ptilocodiidae
0.15
00.14
20.15
40.01
80.01
70.01
60.01
70.01
60.01
65.
Pan
deidae
0.19
00.14
40.16
30.16
10.01
90.02
00.02
10.02
10.02
16.
Lafoeidae
0.18
30.16
30.17
50.15
80.16
30.01
70.01
90.02
00.02
07.
Sertulariid
ae0.16
30.15
90.16
40.15
50.18
60.14
90.01
50.01
80.01
88.
Haleciid
ae0.16
80.15
70.16
80.15
50.17
80.17
20.12
50.02
10.02
19.Clathrozoellid
ae0.14
20.14
30.16
10.14
80.17
80.18
00.16
20.17
70.01
410
.Similiclav
idae
0.14
00.16
90.16
60.13
60.17
50.16
80.16
00.17
00.09
1
Journal of Natural History 19
Dow
nloa
ded
by [
98.2
08.1
64.9
8] a
t 10:
15 1
4 Ja
nuar
y 20
15
Copyright © 2022 FDOKUMEN














