
Bowden, H., Steinhauer, K., Sanz, C. & Ullman, M. (2013) Native-like brain processing of syntax can...
20
Native-like brain processing of syntax can be attained by university foreign language learners Harriet Wood Bowden a,n , Karsten Steinhauer b , Cristina Sanz c , Michael T. Ullman d,nn a Department of Modern Foreign Languages & Literatures, University of Tennessee-Knoxville, 701 McClung Tower, Knoxville, TN 37996, USA b School of Communication Sciences and Disorders, McGill University, Beatty Hall, 1266 Pine Avenue West, Montreal, Quebec H3G-1A8, Canada c Department of Spanish & Portuguese, Georgetown University, Box 571039, Washington, DC 20057, USA d Department of Neuroscience, Georgetown University, Box 571464, Washington, DC 20057, USA article info Article history: Received 7 April 2012 Received in revised form 25 July 2013 Accepted 3 September 2013 Available online 17 September 2013 Keywords: ERP Event-related potentials Second language Syntax Semantics abstract Using event-related potentials (ERPs), we examined the neurocognition of late-learned second language (L2) Spanish in two groups of typical university foreign-language learners (as compared to native (L1) speakers): one group with only one year of college classroom experience, and low-intermediate proficiency (L2 Low), and another group with over three years of college classroom experience as well as 1–2 semesters of immersion experience abroad, and advanced proficiency (L2 Advanced). Semantic violations elicited N400s in all three groups, whereas syntactic word-order violations elicited LAN/P600 responses in the L1 and L2 Advanced groups, but not the L2 Low group. Indeed, the LAN and P600 responses were statistically indistinguishable between the L1 and L2 Advanced groups. The results support and extend previous findings. Consistent with previous research, the results suggest that L2 semantic processing always depends on L1-like neurocognitive mechanisms, whereas L2 syntactic processing initially differs from L1, but can shift to native-like processes with sufficient proficiency or exposure, and perhaps with immersion experience in particular. The findings further demonstrate that substantial native-like brain processing of syntax can be achieved even by typical university foreign- language learners. & 2013 Elsevier Ltd. All rights reserved. 1. Introduction Recent years have witnessed a blossoming of research on the neurocognitive mechanisms underlying late-acquired second lan- guage (L2) (for reviews, see Abutalebi, 2008; Clahsen, Felser, Neubauer, Sato, & Silva, 2010; de Groot, 2011; Doughty & Long, 2005; Green, 2003; Hernandez & Li, 2007; Indefrey, 2006; Kotz, 2009; Kroll & de Groot, 2005; Gass & Mackey, 2012; Morgan-Short & Ullman, 2011; Paradis, 2009; Schmidt & Roberts, 2009; Steinhauer, White, & Drury, 2009; Ullman, 2005). Much of this work has investigated whether the neurocognition of L2 differs from or is similar to that of first (native) language (L1). However, this research program has by no means yielded definitive conclu- sions, and many open questions remain. Three competing theoretical perspectives have addressed the neurocognitive relationship between L2 and L1. First, some researchers have hypothesized that the mechanisms underlying L2 are essentially the same as those subserving (fully-acquired) L1 (Abutalebi, 2008; Ellis, 2005; Hernandez, Li, & MacWhinney, 2005; Indefrey, 2006; MacWhinney, 2012). Second, others have proposed that the mechanisms underlying L2 are fundamentally different from those of L1 (Bley-Vroman, 1989). A third group of theories hypothesize that L2 learners initially depend heavily on different substrates than L1, but, with increas- ing experience or proficiency, gradually rely more on L1 neuro- cognitive mechanisms (Clahsen & Felser, 2006a; Paradis, 1994; Ullman, 2001). The nature of this shift differs among the particular theories. Paradis (1994, 2004, 2009) suggests that a shift between neurocognitive systems can take place both for rule-governed grammatical processes, and at least some lexical properties, specifically, grammatical properties of lexical items that are gen- erally implicit in L1. On the views of Ullman and Clahsen, lexical/ semantics relies on the same set of neurocognitive mechanisms in L2 and L1, whereas grammar shows a more complex relationship (Clahsen & Felser, 2006a, 2006b; Clahsen et al., 2010; Ullman, 2001, 2005, 2006, 2012). In particular, both Ullman and Clahsen hypothesize that aspects of rule-governed grammar are predicted to depend on different mechanisms at earlier versus later stages of L2 acquisition, corresponding to lower and higher levels of proficiency and/or experience. At lower levels, these aspects of Contents lists available at ScienceDirect journal homepage: www.elsevier.com/locate/neuropsychologia Neuropsychologia 0028-3932/$ - see front matter & 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.neuropsychologia.2013.09.004 n Corresponding author. Tel.: þ1 865 974 7099; fax: þ1 865 974 7096. nn Corresponding author. Tel.: þ1 202 687 6064; fax: þ1 202 687 6914. E-mail addresses: [email protected] (H.W. Bowden), [email protected] (K. Steinhauer), [email protected] (C. Sanz), [email protected] (M.T. Ullman). Neuropsychologia 51 (2013) 2492–2511
-
Upload
georgetown -
Category
Documents
-
view
2 -
download
0
Transcript of Bowden, H., Steinhauer, K., Sanz, C. & Ullman, M. (2013) Native-like brain processing of syntax can...

Native-like brain processing of syntax can be attained by universityforeign language learners
Harriet Wood Bowden a,n, Karsten Steinhauer b, Cristina Sanz c, Michael T. Ullman d,nn
a Department of Modern Foreign Languages & Literatures, University of Tennessee-Knoxville, 701 McClung Tower, Knoxville, TN 37996, USAb School of Communication Sciences and Disorders, McGill University, Beatty Hall, 1266 Pine Avenue West, Montreal, Quebec H3G-1A8, Canadac Department of Spanish & Portuguese, Georgetown University, Box 571039, Washington, DC 20057, USAd Department of Neuroscience, Georgetown University, Box 571464, Washington, DC 20057, USA
a r t i c l e i n f o
Article history:Received 7 April 2012Received in revised form25 July 2013Accepted 3 September 2013Available online 17 September 2013
Keywords:ERPEvent-related potentialsSecond languageSyntaxSemantics
a b s t r a c t
Using event-related potentials (ERPs), we examined the neurocognition of late-learned second language(L2) Spanish in two groups of typical university foreign-language learners (as compared to native (L1)speakers): one group with only one year of college classroom experience, and low-intermediateproficiency (L2 Low), and another group with over three years of college classroom experience as wellas 1–2 semesters of immersion experience abroad, and advanced proficiency (L2 Advanced). Semanticviolations elicited N400s in all three groups, whereas syntactic word-order violations elicited LAN/P600responses in the L1 and L2 Advanced groups, but not the L2 Low group. Indeed, the LAN and P600responses were statistically indistinguishable between the L1 and L2 Advanced groups. The resultssupport and extend previous findings. Consistent with previous research, the results suggest that L2semantic processing always depends on L1-like neurocognitive mechanisms, whereas L2 syntacticprocessing initially differs from L1, but can shift to native-like processes with sufficient proficiency orexposure, and perhaps with immersion experience in particular. The findings further demonstrate thatsubstantial native-like brain processing of syntax can be achieved even by typical university foreign-language learners.
& 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Recent years have witnessed a blossoming of research on theneurocognitive mechanisms underlying late-acquired second lan-guage (L2) (for reviews, see Abutalebi, 2008; Clahsen, Felser,Neubauer, Sato, & Silva, 2010; de Groot, 2011; Doughty & Long,2005; Green, 2003; Hernandez & Li, 2007; Indefrey, 2006; Kotz,2009; Kroll & de Groot, 2005; Gass & Mackey, 2012; Morgan-Short& Ullman, 2011; Paradis, 2009; Schmidt & Roberts, 2009;Steinhauer, White, & Drury, 2009; Ullman, 2005). Much of thiswork has investigated whether the neurocognition of L2 differsfrom or is similar to that of first (native) language (L1). However,this research program has by no means yielded definitive conclu-sions, and many open questions remain.
Three competing theoretical perspectives have addressed theneurocognitive relationship between L2 and L1. First, someresearchers have hypothesized that the mechanisms underlying
L2 are essentially the same as those subserving (fully-acquired) L1(Abutalebi, 2008; Ellis, 2005; Hernandez, Li, & MacWhinney, 2005;Indefrey, 2006; MacWhinney, 2012). Second, others have proposedthat the mechanisms underlying L2 are fundamentally differentfrom those of L1 (Bley-Vroman, 1989).
A third group of theories hypothesize that L2 learners initiallydepend heavily on different substrates than L1, but, with increas-ing experience or proficiency, gradually rely more on L1 neuro-cognitive mechanisms (Clahsen & Felser, 2006a; Paradis, 1994;Ullman, 2001). The nature of this shift differs among the particulartheories. Paradis (1994, 2004, 2009) suggests that a shift betweenneurocognitive systems can take place both for rule-governedgrammatical processes, and at least some lexical properties,specifically, grammatical properties of lexical items that are gen-erally implicit in L1. On the views of Ullman and Clahsen, lexical/semantics relies on the same set of neurocognitive mechanisms inL2 and L1, whereas grammar shows a more complex relationship(Clahsen & Felser, 2006a, 2006b; Clahsen et al., 2010; Ullman,2001, 2005, 2006, 2012). In particular, both Ullman and Clahsenhypothesize that aspects of rule-governed grammar are predictedto depend on different mechanisms at earlier versus later stagesof L2 acquisition, corresponding to lower and higher levels ofproficiency and/or experience. At lower levels, these aspects of
Contents lists available at ScienceDirect
journal homepage: www.elsevier.com/locate/neuropsychologia
Neuropsychologia
0028-3932/$ - see front matter & 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.neuropsychologia.2013.09.004
n Corresponding author. Tel.: þ1 865 974 7099; fax: þ1 865 974 7096.nn Corresponding author. Tel.: þ1 202 687 6064; fax: þ1 202 687 6914.E-mail addresses: [email protected] (H.W. Bowden),
[email protected] (K. Steinhauer), [email protected] (C. Sanz),[email protected] (M.T. Ullman).
Neuropsychologia 51 (2013) 2492–2511

grammar are thought to rely largely on lexical/semantic processesthat do not normally play a primary role in grammar in adult L1.The exact nature of these lexical/semantic processes is unclear(and differs somewhat between Clahsen and Ullman), but mayinclude chunking (memorizing complex forms, such as “walked”or the “the cat”), associative generalization in lexical memory,semantic-based parsing, and explicit rules. In addition, Ullmanspecifically ties these lexical/semantic processes to the declarativememory brain system. Crucially however, as proficiency and/orexperience increase, L2 grammar is expected to rely less and lesson these processes and more and more on mechanisms thatnormally underlie grammar in L1 (which Ullman ties to theprocedural memory brain system). For Ullman, most or all aspectsof grammar can potentially become native-like. For Clahsen, suchnative-like processing is typically restricted to local dependencies,and native-like processing of nonlocal dependencies and “complexsyntax” are not generally expected (Clahsen & Felser, 2006a,2006b). Thus, both Clahsen and Ullman predict that the neuro-cognition of lexical/semantics should be qualitatively the same inL2 and L1, at all proficiency and experience levels, whereas rule-governed aspects of grammar should rely largely on lexical/semantics at lower levels, but at least to some extent on native-like aspects of grammatical processing at higher levels – thoughthe exact time course of this shift and for what aspects of grammarthis shift may occur remains unclear, largely due to a lack ofrelevant empirical evidence. In this study, we investigate the timecourse and nature of this shift within the context of typicaluniversity-level language study in the U.S.
1.1. Event-related potentials
Event-related potentials (ERPs) may be one of the best methodsfor teasing apart these competing theories, and more generally forelucidating the neurocognitive relationship between L2 and L1.ERPs reflect real-time scalp-recorded electrophysiological brainactivity of cognitive processes that are time-locked to the pre-sentation of target stimuli. Unlike hemodynamic imaging methodslike fMRI, ERPs provide excellent temporal resolution, allowingone to examine the actual time course of processing. Additionally,ERP research has revealed a set of widely-studied language-relatedactivation patterns (“ERP components”) in L1, whose characteris-tics and underlying functions are reasonably well understood (seebelow). Moreover, lexical/semantic and (morpho)syntactic proces-sing in L1 are associated with largely distinct ERP components.These components thus provide a reasonably clear method ofcomparing the neurocognition of language processing between L2and L1, in particular for lexical/semantic and (morpho)syntacticprocessing. Importantly, since ERPs can be sensitive to effects notobserved with behavioral measures, including in language learn-ing studies (McLaughlin, Osterhout, & Kim, 2004; Tokowicz &MacWhinney, 2005), they can potentially reveal L2–L1 differencesand similarities that might not be found with purely behavioralapproaches.
In L1, ERP studies have shown that lexical/semantic difficulties(e.g., “That train has very large nmacaroni”, where n indicates aviolation or anomaly), which are examined in the present study,consistently elicit “N400” components. These are central/posteriorbilaterally distributed negativities that generally occur about 300–500 ms after the onset of the stimulus. They are associated withlexical/semantic processing (Kutas & Federmeier, 2010; Kutas &Hillyard, 1980; Steinhauer & Connolly, 2008), and have been linkedto the declarative memory brain system (Nobre, Allison, &McCarthy, 1994; Simos, Basile, & Papanicolaou, 1997; Ullman,2004). More specifically, it has been suggested that, at least inlanguage processing, the N400 may reflect aspects of lexical access
and the postlexical integration of word meanings into episodicmemory (Kutas & Federmeier, 2000).
In contrast, disruptions of rule-governed (morpho)syntacticprocessing in L1, such as word-order (phrase structure) violations(e.g., “The man hoped to nmeal the enjoy with friends”), which arealso examined in the present study, often produce three compo-nents. First, such difficulties sometimes, but not always (Hagoort &Brown, 1999; Osterhout, Bersick, & McLaughlin, 1997), elicit early(about 100–500 ms) left-distributed and/or anterior negativities(“LANs”; we emphasize that even in L1, such negativities may beleft-lateralized, anterior, or both; these distributional differencesare not yet well understood) (Friederici, Pfeifer, & Hahne, 1993;Neville, Nicol, Barss, Forster, & Garrett, 1991; Pakulak & Neville,2010). It is not yet clear whether the earlier-onset LAN (often calledan “ELAN”, which can begin as early as around 100 ms), and theLAN that occurs somewhat later (usually beginning around300 ms), reflect the same or different components. Indeed, it hasbeen argued that the presence of ELANs in the majority of previousstudies that report them may in fact be due to artifacts caused bybaseline problems from differences in the material preceding thetarget word (Steinhauer & Drury, 2012) – a problem that is avoidedin the design of the present study. For simplicity, here we use theterm LAN to refer to both earlier and later LAN effects. Although theexact nature of the processing mechanisms underlying LAN effectsremain to be elucidated, evidence suggests that they may reflectaspects of rule-governed automatic structure-building (Friederici &Kotz, 2003; Hahne & Friederici, 1999; Hasting & Kotz, 2008;Steinhauer & Connolly, 2008; Steinhauer & Drury, 2012; van denBrink & Hagoort, 2004) and they may depend on the proceduralbrain system (Hoen & Dominey, 2000; Ullman, 2004). In aninfluential account of language processing (Friederici, 1995, 2002;Friederici & Kotz, 2003) early LAN effects have been argued toreflect first-parse structure building, while the later LAN effectshave been interpreted as reflecting morpho-syntactic processing.
Second, (morpho)syntactic disruptions in L1 also usually elicit –and indeed, more reliably than LAN effects – late centro-parietalpositivities (P600s, often beginning around 600 ms). The P600 hasbeen linked to controlled (conscious) processing, syntactic integra-tion, and structural reanalysis (Friederici & Kotz, 2003; Hahne &Friederici, 1999; Kaan, Harris, Gibson, & Holcomb, 2000; Osterhout& Holcomb, 1992; Steinhauer & Connolly, 2008). It remains unclearwhether or not the P600 is language specific, and some evidencesuggests that this component may be part of the larger P300family, or might comprise P300 subcomponents (Steinhauer &Connolly, 2008).
Third, (morpho)syntactic disruptions have also often beenfound to elicit later sustained anterior negativities (“late anteriornegativities”; these may start as early as 500 or 600 ms, thoughthey are often more prevalent in later time windows) (Friedericiet al., 1993; Gillon Dowens, Vergara, Barber, & Carreiras, 2010;Martin-Loeches, Munoz, Casado, Melcon, & Fernandez-Frias, 2005;Pakulak & Neville, 2010). It has been suggested that such laternegativities, which have recently begun to receive more attention,may represent a continuation of the LAN (or similar processes tothe LAN) (Pakulak & Neville, 2010; Steinhauer & Drury, 2012), oralternatively, that they may reflect something other than auto-matic structure-building, such as processes related to increasedworking memory demands (Martin-Loeches et al., 2005; Sabourin& Stowe, 2008; Vos, Gunter, Kolk, & Mulder, 2001).
1.2. ERPs in second language
As in L1, in late-learned L2 (here we discuss studies in whichthe mean age of acquisition is at least 10 years) difficulties inlexical/semantic processing reliably yield an N400, though inmany cases this component is delayed or reduced in amplitude
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2493

as compared to L1 (Ardal, Donald, Meuter, Muldrew, & Luce, 1990;Hahne, 2001; Hahne & Friederici, 2001; Kutas & Kluender, 1994;Moreno & Kutas, 2005; Newman, Tremblay, Nichols, Neville, &Ullman, 2012; Ojima, Nakata, & Kakigi, 2005; Weber-Fox & Neville,1996). N400 effects have been found consistently across L2proficiency and experience levels – even at very low levels, forexample after one month of L2 classroom experience (McLaughlinet al., 2004; Osterhout et al., 2008). Interestingly, there is someevidence that the amplitude of the N400 violation effect may be atleast partly explained by proficiency, with the size of the N400(i.e., the difference in amplitude between anomalous and con-gruent words) increasing as proficiency increases, in both L1 andL2 (Newman et al., 2012).
L2 processing of (morpho)syntactic disruptions, on the otherhand, shows a more complex relationship as compared to L1. Herewe summarize the findings of the 20 studies of such disruptionsthat we are aware of (including in artificial language paradigms1)with regard to the LAN, P600, and late anterior negativity (for eachof these components, limiting ourselves to those studies thatdirectly test for the component in a relevant time window)(Chen, Shu, Liu, Zhao, & Li, 2007; Friederici, Steinhauer, & Pfeifer,2002; Gillon Dowens, Guo, Guo, Barber, & Carreiras, 2011; GillonDowens et al., 2010; Hahne, 2001; Hahne & Friederici, 2001;Hahne, Mueller, & Clahsen, 2006; Isel, 2007; McLaughlin et al.,2010; Morgan-Short, Finger, Grey, & Ullman, 2012; Morgan-Short,Sanz, Steinhauer, & Ullman, 2010; Morgan-Short, Steinhauer, Sanz,& Ullman, 2012; Mueller, Hahne, Fujii, & Friederici, 2005; Ojimaet al., 2005; Osterhout et al., 2008; Pakulak & Neville, 2011; Rossi,Gugler, Friederici, & Hahne, 2006; Steinhauer et al., 2009;Tokowicz & MacWhinney, 2005; Weber-Fox & Neville, 1996).
First, the presence of the LAN is quite variable. Although thiscomponent is reported in at least one experimental condition (i.e.,for at least one participant group or type of violation) in quite afew studies (Friederici, Steinhauer, et al., 2002; Gillon Dowenset al., 2010; Hahne et al., 2006; Isel, 2007; Morgan-Short, Finger,et al., 2012; Morgan-Short, Steinhauer, et al., 2012; Ojima et al.,2005; Rossi et al., 2006; Steinhauer et al., 2009; Weber-Fox &Neville, 1996), even more studies do not find a LAN in one or moreconditions (Chen et al., 2007; Gillon Dowens et al., 2011; GillonDowens et al., 2010; Hahne, 2001; Hahne & Friederici, 2001;Hahne et al., 2006; Morgan-Short, Finger, et al., 2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer, et al., 2012; Muelleret al., 2005; Ojima et al., 2005; Pakulak & Neville, 2011; Rossi et al.,2006; Steinhauer et al., 2009; Weber-Fox & Neville, 1996).
It is not yet clear what factors affect the presence or absence ofthe LAN in L2. Several studies have suggested that proficiency mayplay a role (e.g., Friederici, Steinhauer, et al., 2002; Gillon Dowenset al., 2010; Hahne, 2001; Hahne et al., 2006) (for reviews, see Kotz,2009; Steinhauer et al., 2009). Indeed, among studies that char-acterize their participants as having lower proficiency (e.g., “lowproficiency,” “less proficient,” “early stages of learning”), we areaware of only one that has reported a LAN (Rossi et al., 2006). Inthis study, the authors attributed this surprising finding to the lowsyntactic complexity of their stimuli (as compared to other studiesof L2 in which LANs were not found), which may have led to more
native-like processing. Additionally, it has been argued that theearly LAN reported in this study may be an artifact, since it seemsto occur prior to the auditory presentation of the suffix causing theviolation (Steinhauer & Drury, 2012). Instead of LANs, at lower L2proficiency (morpho)syntactic disruptions tend to produce eitherno effect in the LAN time window (Hahne & Friederici, 2001; Ojimaet al., 2005; Steinhauer et al., 2009), or an N400 or N400-likeresponse (McLaughlin et al., 2010; Morgan-Short, Finger, et al.,2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer, et al.,2012; Mueller et al., 2005; Osterhout et al., 2008), suggesting acompensatory role for lexical/semantic processes and declarativememory in at least some cases. In contrast, studies of participantscharacterized as having higher proficiency (e.g., “proficient,” “rela-tively high,” “high proficiency,” “high level of proficiency”) haveoften reported a LAN effect (that is, a left-lateralized and/oranterior negativity between about 100–500 ms) in at least oneexperimental condition (Friederici, Steinhauer, et al., 2002; GillonDowens et al., 2010; Hahne et al., 2006; Isel, 2007; Morgan-Short,Finger, et al., 2012; Morgan-Short, Steinhauer, et al., 2012; Ojimaet al., 2005; Rossi et al., 2006; Steinhauer et al., 2009).
Thus proficiency seems to correlate with the presence of a LAN.However, it is a far from perfect correlation, since a similar number ofstudies of higher proficiency groups have not found a LAN in at leastone condition (Chen et al., 2007; Gillon Dowens et al., 2011; GillonDowens et al., 2010; Hahne, 2001; Hahne et al., 2006; Morgan-Short,Finger, et al., 2012; Morgan-Short et al., 2010; Morgan-Short,Steinhauer, et al., 2012; Mueller et al., 2005; Pakulak & Neville,2011). Instead, no component was observed in most of these studiesin the LAN time window, while in one case an anterior positivity wasfound (Morgan-Short, Steinhauer, et al., 2012). The lack of a strongcorrelation could of course be due to the actual absence of anassociation between proficiency and LAN effects in L2, although thevirtual lack of a LAN at low proficiency suggests that there is indeedsome sort of relation. The weak correlation might also be partlyexplained by the fact that proficiency is inconsistently characterizedacross studies, and is assessed by widely different measures, and infact often only by self-ratings.
Additionally, factors other than proficiency might influence thedevelopment of LAN effects in L2. These include not only task-related factors such as syntactic complexity (see above), but alsofactors that are correlated with and thus confounded with profi-ciency. Indeed, even though L2 proficiency is evidently stronglyassociated with the amount of L2 experience, most studies focusonly on the factor of proficiency. Few even report estimates of thetotal amount of L2 experience, let alone of different types (i.e.,classroom as well as immersion experience), making it difficult todetermine whether the proficiency level and/or the amount ofexperience play important roles. Interestingly, recent evidencesuggests that the type of L2 experience may affect the presence ofthe LAN, even when holding proficiency level and amount ofexperience constant. Specifically, the results of some studies usingartificial languages suggest that immersion-like experience mightbe necessary for the emergence of the LAN even with higher levelsof proficiency and exposure (Morgan-Short, Finger, et al., 2012;Morgan-Short, Steinhauer, et al., 2012). Indeed, all of the studiesreviewed in which a LAN was found (see above) examinedparticipants with at least some immersion experience, includingthe one study that reported a LAN at low L2 proficiency (Rossiet al., 2006). Thus, it is possible that immersion may be necessaryfor the development of the LAN, or at least is an important factor.Finally, note that early LAN effects in many previous (L1 and L2)studies of word-order (phrase structure) violations may be pro-blematic because the stimulus paradigms unintentionally inducedcontext or baseline effects (Steinhauer & Drury, 2012).
Second, in addition to the LAN, (morpho)syntactic disruptionsalso often elicit a P600 in L2 learners – and as in L1, more reliably
1 Our review includes artificial languages and mini-languages (but not artificialgrammars), because they are natural-language like in that (a) their words andsentences have actual meanings, (b) their grammars follow those of naturallanguages, and (c) they elicit the same ERP components as natural languages. Weacknowledge that artificial language paradigms may be justifiably challenged onthe grounds that they lack native speaker controls, and that their trainingconditions do not fully reflect learning conditions in natural languages (e.g.,immersion-like training is not the same as true immersion). Nevertheless, webelieve that studies using these paradigms are informative, and we believe thatultimately what is needed is convergence between data from both artificial andnatural languages.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112494

than the LAN. Of the studies examined here, the great majorityreport a P600 in at least one condition (all but four; Chen et al.,2007; Hahne & Friederici, 2001; Isel, 2007; Ojima et al., 2005).Nevertheless, a fair number of studies have reported at least one(morpho)syntactic condition with no P600s (the four above andMcLaughlin et al., 2010; Morgan-Short et al., 2010; Osterhout et al.,2008; Tokowicz & MacWhinney, 2005). Like the LAN, the P600seems to be somewhat less prevalent at lower proficiency (P600swere found in five of eight studies of low L2 proficiency:McLaughlin et al., 2010; Osterhout et al., 2008; Rossi et al., 2006;Steinhauer et al., 2009; Tokowicz & MacWhinney, 2005) than athigher proficiency (found in 12 of 15 studies: Friederici,Steinhauer, et al., 2002; Gillon Dowens et al., 2011; GillonDowens et al., 2010; Hahne, 2001; Hahne et al., 2006; Morgan-Short, Finger, et al., 2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer, et al., 2012; Mueller et al., 2005; Pakulak &Neville, 2011; Rossi et al., 2006; Steinhauer et al., 2009). However,high proficiency does not seem to be sufficient for the attainmentof P600s, since of the 15 studies of high-proficiency participants,four report at least one condition for which no P600 is produced(Chen et al., 2007; Isel, 2007; Morgan-Short et al., 2010; Ojimaet al., 2005). Immersion may also affect the presence of P600effects. For example, of all studies reporting a P600 (see justabove), 75% examined participants with immersion experience(Friederici, Steinhauer, et al., 2002; Gillon Dowens et al., 2010;Hahne, 2001; Hahne et al., 2006; Morgan-Short, Finger, et al.,2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer, et al.,2012; Mueller et al., 2005; Pakulak & Neville, 2011; Rossi et al.,2006; Steinhauer et al., 2009; Weber-Fox & Neville, 1996). Thus,although proficiency appears to contribute to the emergence ofthe P600, it does not appear to tell the whole story, and immersionmay also play a role.
The picture for late anterior negativities is somewhat similar tothat for LANs and P600s, though somewhat less clear. Of the 20 L2(morpho)syntax studies reviewed here, 16 examined appropriatetime windows (beginning at 500 ms or later) and electrodes for alate anterior negativity (Chen et al., 2007; Gillon Dowens et al.,2011; Gillon Dowens et al., 2010; Hahne, 2001; Hahne & Friederici,2001; Hahne et al., 2006; Isel, 2007; Morgan-Short, Finger, et al.,2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer, et al.,2012; Mueller et al., 2005; Ojima et al., 2005; Pakulak & Neville,2011; Rossi et al., 2006; Tokowicz & MacWhinney, 2005; Weber-Fox & Neville, 1996). Of these 16 studies, only eight report whatappear to be a late anterior negativity (Chen et al., 2007; GillonDowens et al., 2011; Gillon Dowens et al., 2010; Isel, 2007;Morgan-Short, Finger, et al., 2012; Morgan-Short et al., 2010;Morgan-Short, Steinhauer, et al., 2012; Rossi et al., 2006). In allbut one of these eight studies (Rossi et al., 2006), this effect wasfound only for highly proficient learners, suggesting that profi-ciency or one of its correlates (e.g., amount of experience) mightplay a role. However, as with both LAN and P600 effects, highproficiency is clearly not sufficient for the presence of a lateanterior negativity, since only about 60% of the relevant studiesthat examined high-proficiency learners report this component inat least one condition (Chen et al., 2007; Gillon Dowens et al.,2011; Gillon Dowens et al., 2010; Isel, 2007; Morgan-Short, Finger,et al., 2012; Morgan-Short et al., 2010; Morgan-Short, Steinhauer,et al., 2012; Rossi et al., 2006). Immersion also appears to play arole: for example, of the eight studies reporting a late anteriornegativity, all but two examined participants with immersionexperience (all but Chen et al., 2007; Gillon Dowens et al., 2011).Thus, both high proficiency and immersion (as well as otherfactors) may influence the presence of this effect. In addition, itshould be noted that since in L1 a late anterior negativity oftenappears only after the P600 has peaked (Steinhauer & Drury,2012), and many L2 studies do not report such late time windows,
the proportions presented here may underestimate the actualprevalence of the late anterior negativity.
In sum, the following picture emerges from previous ERPstudies of later second language learners (note that although ageof acquisition also likely influences ERP outcomes, here we focusonly on later learners, and do not examine this factor). Lexical/semantic difficulties elicit an N400 in L2 as well as L1, though thiseffect may be delayed or smaller in L2, perhaps partly due to lowerproficiency and/or experience in L2 than L1. Thus the ERP evidencesuggests that L2 and L1 rely on the same set of neurocognitivemechanisms for lexical/semantics, though the relevant lexical/semantic knowledge may be better learned in L1. In contrast, theextent to which (morpho)syntactic processing in L2 is similar tothat of L1 seems to vary, apparently as a function of not onlyproficiency, but potentially other factors as well, including theamount and type of experience. At lower proficiency and/orexperience levels, LANs and late anterior negativities seem to bevirtually absent, though P600s are not as rare. In some cases anN400 or N400-like effect is seen in lieu of a LAN. In contrast, athigher proficiency/experience levels, LANs, P600s and late anteriornegativities are all reported in many studies. Nevertheless, highproficiency does not seem to be sufficient for these components,since LANs and late anterior negativities, and to a lesser extentP600s, are absent in a number of studies of high-proficiencylearners. Immersion experience may also play a role in theattainment of L1-like (morpho)syntactic processing, for LANs aswell as P600s and late anterior negativities. Overall, the previousevidence of L2 (morpho)syntactic disruptions suggests that L1-likeERP components may in fact be found in L2; these components aremore likely at higher than lower proficiency and/or exposure; andimmersion experience may be important, though other factorsmay also play a role.
1.3. Gaps and weaknesses
The picture painted by L2 ERP research is still far from clear,however, and various empirical gaps, as well as important meth-odological weaknesses in previous studies, suggest that it ispremature to draw any strong conclusions. In particular, we donot yet understand whether and at what levels proficiency and/orexperience might lead to the development of LAN, P600, and lateanterior negativity effects in L2 (morpho)syntax. This lack ofunderstanding may be attributed to a number of factors, includingthe following.
First, although a number of studies have focused on proficiency,the role of both proficiency and experience (including type ofexperience) remains unclear, for various reasons. Previous studieshave varied widely in their proficiency measures (and how theydefine low or high proficiency), as well as with respect to otherfactors that could affect ERP outcomes. For example, differentstudies have examined different L2s, in speakers with differentnative languages, using different (morpho)syntactic disruptionparadigms, in different modalities (visual vs. auditory presenta-tion). Thus outcomes that might be attributed to proficiency couldin fact be due to a host of other factors instead of or in combina-tion with proficiency. Indeed, we are aware of only a handful ofstudies that have directly compared low- and high-proficiencylearners with these factors held constant (Morgan-Short et al.,2010; Morgan-Short, Steinhauer, et al., 2012; Ojima et al., 2005;Rossi et al., 2006; Steinhauer et al., 2009). In addition, few studieshave considered the role of type of experience (Morgan-Short,Finger, et al., 2012; Morgan-Short et al., 2010; Morgan-Short,Steinhauer, et al., 2012), and many do not even report measuresof participants' classroom and immersion experience. Subject-level factors such as handedness (right vs. left) and sex (male vs.female) that can influence language processing (Ullman, Gelfand,
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2495

Walenski, Moffa, & Lee, in preparation; Ullman, Miranda, &Travers, 2008) are also not always reported or vary across studies,further reducing confidence in conclusions about the role ofproficiency. Along similar lines, even L2-related factors such asage of acquisition are inconsistently reported across (naturallanguage) studies, referring variably to age of arrival or age of firstclassroom exposure, while in some cases it is not clear what age ofacquisition refers to.
Second, a number of studies have not directly (statistically)compared L2 ERP patterns with those of L1 speakers using thesame paradigm, even for studies of natural language where this ispossible (Gillon Dowens et al., 2011; Gillon Dowens et al., 2010;McLaughlin et al., 2010; Osterhout et al., 2008; Tokowicz &MacWhinney, 2005). This suggests caution in drawing conclusionsin these studies about L1-like components in L2, and certainlyprecludes stronger statements of the sort that the L2 ERP patterndoes or does not differ from L1.
Third, a major empirical gap in previous L2 ERP research is thatmost studies of (morpho)syntax have examined immigrant groups(or immigrant-like groups, such as expatriates), and have largelyignored foreign-language learners of the sort that are often foundin the U.S. (and many other countries) – that is, whose experienceis primarily from classroom exposure (generally starting in middleschool or high school), with immersion experience limited largelyto study abroad, often towards the end of university. We are awareof only three L2 ERP studies of (morpho)syntax that have exam-ined such foreign-language learners (McLaughlin et al., 2010;Osterhout et al., 2008; Tokowicz & MacWhinney, 2005). However,all of these focused on relatively early stages of learning, withnone of their L2 groups having higher proficiency levels. Indeed,none had taken L2 classes beyond the third year of college, andnone had apparently had any study abroad or other immersionexperience. Additionally, none of these studies reported appro-priate electrodes and/or time windows for the examination of aLAN. Thus, it remains unclear whether, when, and how L1-likebrain processing may emerge in foreign-language learners.
Finally, as discussed above, most previous studies of word-order violations (both in L1 and L2) have had ERP baselineproblems due to differences in the material preceding the targetword, resulting in a lack of confidence in the presence or absenceof early ERP effects, in particular, early LAN effects (for discussion,see Steinhauer & Drury, 2012).
1.4. Motivations for the study
Here we attempt to address some of these gaps and weak-nesses. First, we focused on typical foreign-language learners inthe U.S. All learned the L2 first in classrooms (mean age of initialclassroom exposure: 12.9 years). We examined and directlycompared two such groups. The first had a mean of two semestersof L2 classroom training in college, with no immersion experience,and an expected proficiency of low intermediate (“L2 Low”; seeMethods section). The second group had about seven semesters ofuniversity-level classroom experience, plus about one to twosemesters of immersion experience abroad, with an expectedproficiency of mid advanced (“L2 Advanced”). This design allowedus to probe whether and when in the course of typical universityforeign-language learning native-like ERP components for (mor-pho)syntax might appear.
On the basis of the previous literature (see above), it wasunclear whether or not the L2 Low group would show a P600,whereas neither a LAN nor a late anterior negativity was expected.The absence of any of these components in this group wouldstrengthen the view that higher levels of proficiency and/orexperience (and possibly immersion experience) may be
important for their appearance, whereas their presence wouldsuggest that these factors are not critical.
The L2 Advanced group allowed us to test whether the fullpanoply of L1 ERP components (the LAN, P600, and possibly thelate anterior negativity), or perhaps a subset, can indeed emerge ina university foreign-language learning context, including studyabroad toward the end of this experience. The absence of a native-like pattern in these learners would suggest that this foreign-language learning experience is not sufficient to attain L1-likebrain processing. In contrast, the presence of these componentswould suggest that this experience is sufficient for attaining (atleast some) native-like brain processing. In turn, this findingwould allow further investigations to attempt to pinpoint exactlywhen native-like processing emerges, and under what conditions– e.g., whether it might already be present before study abroad, orperhaps with an equivalent amount of classroom exposure as thatexperienced during study abroad. In other words, the two groupsdelimit the beginning and end of the university foreign languagelearning experience available to many students, and elucidatewhich, if any, aspects of native-like processing are present at eachpoint, thus providing a foundation for future studies to determinewith greater precision when exactly within or after this period(or possibly before) various aspects of native-like processing mayemerge.
We selected Spanish as the L2 in part because it is the mostcommonly taught foreign language in the U.S. (Alonso, 2006;Rhodes & Pufahl, 2009). We examined word-order rather thanmorphosyntactic (e.g., agreement) violations because the latterseem to cause particular difficulties in L2 learning (Montrul,2004; White, 2003), and some previous studies have hinted thatnative-like ERP components might be more attainable for word-order than morphosyntactic violations (Morgan-Short et al., 2010;Morgan-Short, Steinhauer, et al., 2012). Thus, in the L2 Advancedgroup, we purposefully stacked the deck in favor of the emergenceof native-like processing, with the notion that if this group did notshow a fully native-like ERP pattern, we might reasonably con-clude that such a pattern may not be achievable in a typicalforeign-language learning context. Along the same lines, thefact that Spanish has a similar word-order structure to English(head-first phrase structure and canonical Subject–Verb–Objectword order, though word order is more flexible in Spanish thanEnglish) would favor the emergence of native-like brain processingif such structural similarity has an influence (Kotz, 2009;MacWhinney, 2007).
The study was designed to avoid many of the methodologicalweaknesses found in some previous research. The two L2 partici-pant groups did not differ in their age of initial classroomexperience (i.e., age of acquisition) or in the amount of classroomexperience prior to college. All L2 participants were given astandardized proficiency test of Spanish, in addition to self-ratings. The ERPs of the two L2 groups were compared directly(statistically) to each other and to a separate group of L1 (native)Spanish speakers (who moreover had English as their L2, and werethus matched to the L2 Spanish speakers in also having an L2). Allthree groups underwent the exact same experimental paradigm.All participants were right handed, and the three groups did notdiffer statistically on sex (distribution of males/females). Critically,the word-order violations were designed to avoid baseline pro-blems, and to have both the target words and preceding materialmatched across correct and violation sentences.
Finally, unlike some previous studies of L2 (morpho)syntax,lexical/semantic anomalies were also examined, for at least tworeasons. First, this condition is of course interesting in its ownright, allowing us to investigate the trajectory of lexical/semanticprocessing in foreign-language learners, and the influence ofproficiency and experience. Second, the condition serves as a
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112496

control for the word-order violations: if N400s are not found in L2in response to lexical/semantic anomalies, despite being stronglypredicted, then the absence of the expected syntactic ERP compo-nents would have to be treated with caution, as a null effect withpotentially little meaning.
2. Methods
2.1. Participants
We tested a total of 32 L2 Spanish speakers (with English as their L1), and 17 L1Spanish speakers (with English as their L2). All participants had at least somecollege education, and all were right-handed (Oldfield, 1971), had normal orcorrected hearing and vision, and had no known developmental, neurological, orpsychiatric disorders. They gave written informed consent and received monetarycompensation for their participation in the study. Four participants were excludedfrom analyses: two L2 participants who were outliers in their proficiency scores(see below), and two L1 participants due to the large number of artifacts in theirERP data. All statistics below are based on these 45 participants.
All of the L2 learners were native speakers of American English who had firstbeen exposed to Spanish in classroom environments. The two L2 groups did notdiffer in their initial age of classroom exposure to Spanish, or in their amount ofclassroom exposure to Spanish prior to age 17 (see Table 1). Additionally, none ofthe L2 learners had any significant immersion or immersion-like experience (otherthan the designated study abroad for the L2 Advanced group). Thus, none of the L2subjects had had family members or caretakers who spoke Spanish to them, noneattended a bilingual school (with Spanish or any other language), and none hadlived in a Spanish-speaking country or had visited one for more than a briefvacation.
The L2 Spanish participants were initially selected according to their specificlevel of L2 experience (see below), and were subsequently evaluated for proficiencyin Spanish with the Simulated Oral Proficiency Interview (SOPI; Center for AppliedLinguistics, Washington, DC).2 To maintain as much homogeneity as possible withineach of the L2 groups, outliers in each group (42 SDs from the group mean on theSOPI, with ratings converted to numerical scores following Henning 1992), wereexcluded from analysis. From the original participant pool, two were therebyexcluded (one from L2 Low and one from L2 Advanced).
See Table 1 for information on the three groups' age, education, and sex, and onthe L2 groups' classroom Spanish exposure and Spanish proficiency (as measuredby the SOPI and by self-ratings). Following are additional details for each of thethree groups.
2.1.1. L1 Spanish (L1)This group (n¼15) was selected to serve as a control group for the two L2
groups, and was tested only in Spanish, not in English. They had lived in Spanish-speaking countries (14 participants in Latin America and one in Spain) until at leastage 17 (except for one participant, who moved to the U.S. at age 15), with nosignificant immersion or immersion-like experience to English before this age.They were all living in the U.S. at the time of the study. Their average length ofresidence in English-speaking countries was 2.9 years (SD¼3.2), and all continuedto use Spanish on a regular basis.
2.1.2. Low experience L2 Spanish (L2 Low)Participants in this group (n¼16) were enrolled at the time of testing in 2nd–
4th semester Spanish language classes at Georgetown University (mean number ofsemesters of college Spanish¼2.0, SD¼1.1). As noted above, none of the partici-pants had any significant immersion or immersion-like Spanish experience. Theaverage level of proficiency for this group was Intermediate Low (that is, low-levelwithin Intermediate), as measured by the SOPI2.
2.1.3. Advanced experience L2 Spanish (L2 Advanced)This group (n¼14) had more classroom experience than the L2 Low group, as
well as immersion experience from study abroad. They had completed an averageof 6.9 semesters of university classroom Spanish (SD¼1.6), plus approximately oneto two semesters in a study abroad immersion situation in a Spanish-speakingcountry (mean number of months¼9.4, SD¼3.6). The average level of proficiencyfor this group was Advanced Mid (that is, mid-level within Advanced), as measuredby the SOPI.
2.2. Stimuli
The experimental stimuli consisted of 120 Spanish sentences, with 40 sen-tences in each of the two experimental conditions – that is, semantic (i.e., lexical/semantic) and word-order violations – as well as 40 correct control sentences. Thesentences were intermixed pseudorandomly (no more than three sentences of anycondition in a row), together with sentences of four additional conditions notreported here (352 additional sentences, with gender violations, number violations,or violations of regular or irregular morphology, and their corresponding correctsentences).3 To maximize the likelihood that the L2 Low participants were familiar
Table 1Participant background information.
L1 Spanish (n¼15) L2 Low Spanish (n¼16) L2 Advanced Spanish (n¼14)
Age (years) 26.9 (5.3) 19.1 (.8) 23.6 (3.5)Years of education 17.9 (3.1) 13.2 (.8) 16.5 (1.7)
Sex (females, males) 8 F, 7 M 9 F, 7 M 10 F, 4 MAge of initial classroom Spanish exposure (years) n/a 14.1 (3.6) 12.4 (2.4)Amount of classroom Spanish prior to age 17 (years) n/a 3.6 (3.6) 5.1 (2.3)L2 proficiency (SOPI) score (min¼ .1, max¼3.0) n/a 1.1 (.12) 2.4 (.33)
L2 self-rating (5-point scale)Listening n/a 2.5 (.7) 3.8 (.7)Reading n/a 2.8 (.6) 3.7 (.7)Speaking n/a 2.2 (.5) 3.6 (.9)Writing n/a 2.5 (.7) 3.4 (.9)
Note. The values represent means, with SDs in parentheses. Years of education is defined as the number of years of formal education. The L2 Low group had significantlylower ages and years of education than the other two groups (pso .05; see Discussion section), which in turn did not differ significantly from each other (ps4 .06). The threegroups did not differ in their distribution of females and males (χ2(2)¼1.13, p¼ .567). The two L2 groups did not differ either in the age of initial classroom exposure (t(28)¼1.51, p¼ .141) or in the amount of classroom exposure prior to age 17 (t(28)¼1.29, p¼ .206). As expected, proficiency scores as well as each of the four self-ratings differedbetween the L2 Low and L2 Advanced groups (pso .05).
2 The SOPI is based on the speaking proficiency guidelines of the AmericanCouncil on the Teaching of Foreign Languages (ACTFL). Designed for secondlanguage learners, it uses a variety of standardized, situation-based prompts toelicit oral production. The SOPI takes about 45 min to administer, and responses arerecorded for subsequent rating. Ratings range from “below novice high” to“superior”, with the following sublevels: Novice (two levels: ohigh, high; notethat the “ohigh” novice SOPI level corresponds to and does not distinguishbetween “low” and “mid” in the full ACTFL scale); Intermediate (three levels: low,mid, and high), Advanced (three levels: low, mid, high) and Superior (only onelevel). In this study, the SOPI was computer-administered and digitally recorded,and ratings were later assigned by trained SOPI raters. See http://www.cal.org/topics/ta/sopi.html for more information on the SOPI.
3 Sample sentences of other conditions not examined here (violations andcorrect controls): Gender: El lago es ntranquila por la mañana. (The lakems isntranquilfs in the morning.) Number: El lago es ntranquilos por la mañana. (Thelakems is ntranquilmp in the morning.) Correct: El lago es tranquilo por la mañana.(The lakems is tranquilms in the morning.) Regular morphology: Hoy en día yosiempre nbesar a mi esposa. (These days I always nto kiss my wife.) Correct: Hoy endía yo siempre beso a mi abuela. (These days I always kiss my grandmother.)Irregular morphology: Hoy en día yo siempre ncalentar el café. (These days I always
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2497

with the vocabulary, low-frequency words were avoided; additionally, 84% of thelexical items in the 120 sentences examined here were present in the first-yearGeorgetown University Spanish textbook.
The semantic and the word-order violation sentences were derived from thesame set of 40 correct sentences. All sentences contained 7–11 words. Thesentences were created following a paradigm which allowed ERPs of correct andviolation conditions to be compared while holding constant both the target wordsand the preceding (and following) sentence contexts (Steinhauer et al., 2009;Steinhauer, White, & Genesee, in preparation). This approach crucially eliminatesbaseline problems found in many ERP studies (for discussion, see Steinhauer &Drury, 2012; Steinhauer, White, & Genesee, in preparation).
Semantic violations were created by interchanging sentence-final nouns acrossthe entire set of 40 correct sentences (see Table 2 for examples). Thus, across the 40semantic violation sentences, the same sentence contexts and the same targetnouns were found as in the 40 correct sentences, but with different combinationsof sentence contexts and target nouns in the two sets of sentences. Sentence-finaltarget positions were chosen for several reasons, despite the potential disadvantageof sentence wrap-up effects (Hagoort, 2003; Osterhout, Holcomb, & Swinney, 1994;Osterhout & Nicol, 1999): given the structural constraints on the correct sentencesdue to the fact that they were also used for the creation of word-order violations,and sentence length constraints necessary to keep the ERP testing session to amanageable length, it turned out that only the final noun had sufficient contextualbuild up to generate high amplitude N400s. Importantly, the N400 is a robustcomponent, and many previous studies have successfully used sentence-finalstimuli (Ardal et al., 1990; Hagoort, 2003; Meuter, Donald, & Ardal, 1987; Moreno& Kutas, 2005).
Word-order violations were created by reversing the position of a noun and averb in the correct sentences, such that one was presented where the other wasexpected. As shown in Table 2, the correct sentences and word-order violationswere created in pairs, following the example of the English sentences created bySteinhauer et al. (2009, in preparation), such that both the target words and thepreceding sentence contexts were matched across the violation and correctsentence conditions, thus avoiding baseline problems. Violations occurred betweenthe third and the fifth word in the sentences.
All 120 sentences were presented to all participants. Six different pseudor-andomized presentation orders were created, and were approximately equallydistributed across participants within each of the three participant groups. For anyone base sentence, the three different versions (correct, semantic violation, word-order violation) were maximally separated in each presentation order. Finally,these 120 sentences were intermixed with the gender, number, regular andirregular morphological violation and correct control sentences mentioned above.
2.3. Procedure
At least two weeks prior to the ERP testing session, participants underwent abehavioral testing session in which background questionnaires (collecting demo-graphic, language-related, and medical information) and proficiency tests wereadministered, as well as regular/irregular inflectional morphology tasks reportedelsewhere (Bowden, Gelfand, Sanz, & Ullman, 2010). During the ERP session,4
participants were seated 70 cm from a 16-in. CRT monitor, in a dark, quiet testingroom. After application of the ERP cap, participants were instructed to minimizeeye and body movements during the presentation of each sentence, and were
instructed how and when to respond to the stimuli. The experimental sentenceswere divided into eight blocks lasting approximately 8–10 min each, with breaksbetween blocks. Prior to the first experimental block, participants performed apractice block with 10 items. In addition, six pre-items (not analyzed) werepresented at the beginning of the first experimental block. Participants wereinstructed to respond to each sentence by judging its acceptability in Spanish. Theacceptability judgment task was selected in order to provide behavioral measuresof accuracy.
Sentences were presented on the computer monitor one word at a time (RapidSerial Visual Presentation; RSVP), using the Presentation stimulus presentationprogram (Neurobehavioral Systems) on a Dell Inspiron 8600 laptop with WindowsXP. The participants saw the stimuli on a separate CRT monitor, thus avoiding thetiming inaccuracies inherent in LCD displays. Each trial began with a fixation crossin the center of the screen. After 1 s, the sentence was presented, one word at atime (duration¼300 ms; SOA¼500 ms) in the center of the screen. After eachsentence, the screen went blank for 1 s, after which an acceptability judgmentprompt appeared on the screen. This prompt remained on the screen for up to 3 s,or until a response was given (by pressing a mouse button with the right hand).A two-second intertrial (eye-blink) interval followed, after which the cross of thenext trial appeared.
2.4. EEG recording and pre-processing
Scalp EEG was continuously recorded at a sampling rate of 500 Hz from 64 tinelectrodes mounted in an elastic cap (Electro-Cap International), based on anextension of the 10–20 international system. Impedance at each scalp electrodewas reduced to 5 kΩ or below before recording. The EEG was amplified byNeuroscan SynAmps2 amplifiers (online band-pass filter: .05–100 Hz). Electrodeswere also placed above and below the right eye and on the outer canthi of botheyes to monitor vertical and horizontal eye movements and eye-blinks. Off-line,the EEG was re-referenced to the right mastoid and filtered with a band-passfilter (.16–30 Hz).
Trials with artifacts greater than 30 mV in the EOG channels or greater than75 mV in the EEG channels were removed from the raw data. One participant's datathat contained a relatively large number of eye movement artifacts in the semanticcondition were corrected with an EOG algorithm (EEProbe, ANT).5 Artifacts led torejections of 7.6% of the trials in the L1 group, 13.3% in the L2 Low group, and 10.0%in the L2 Advanced group. All remaining trials, regardless of behavioral responses,were included in the main analyses (all-trials analyses).6 Nevertheless, we alsoperformed all analyses only on those trials that were correctly responded to(response-contingent analyses), in order to ensure that we did not miss any effectsthat might be seen only in these correct trials; results from these analyses aresummarized below the main analyses, and are reported fully in the Supplementary
Table 2Sample control and violation sentences.
Violation condition Correct (control) sample sentences Violation sample sentences
Semantic Tengo muchas millas que correr esta semana. Tengo muchas millas que correr esta actriz.(I-have many miles to run this week.) (I-have many miles to run this actress.)Ellos van a las obras para ver a la actriz. La profesora espera ir en autobús a la semana.(They go to the plays to see the actress.) (The professor hopes to-go by bus to the week.)
Word-order Tengo que correr muchas millas esta semana. Tengo que millas muchas correr esta semana.(I-have to run many miles this week.) (I-have to miles many run this week.)Tengo muchas millas que correr esta semana. Tengo muchas correr que millas esta semana.(I-have many miles to run this week.) (I-have many run to miles this week.)
Note: English translations are given in parentheses. Critical words whose ERPs were compared in correct versus violation sentences are underlined.
(footnote continued)nto heat the coffee.) Correct: Hoy en día yo siempre caliento el agua. (These days Ialways heat the water.) Notes. ms: masculine singular; fs: feminine singular; mp:masculine plural. Critical words are underlined.
4 Pilot participants (five L1 speakers and six L2 Low speakers) included in thecurrent analyses performed the ERP experiment in two sessions rather than one.These two sessions corresponded exactly to the first and second halves of thesessions as presented to other participants.
5 The EEProbe EOG correction algorithm builds a prototype EOG event for asubject via manual selection of eyeblinks. It then calculates propagation factors thatdefine how EOG activity will be subtracted from each of the non-EOG channels inthe correction procedure.
6 Our main analyses (all-trials analyses) included all artifact-free trials, regard-less of their associated behavioral responses. This approach (as opposed toresponse-contingent analyses) was chosen for the primary analyses for severalreasons (also see Steinhauer et al., in preparation). First, this is a common approachin previous L2 studies (Chen et al., 2007; Foucart & Frenck-Mestre, 2012; Frenck-Mestre, Osterhout, McLaughlin, & Foucart, 2008; Mueller et al., 2005; Ojima et al.,2005; Weber-Fox & Neville, 1996). Second, previous research has revealed sig-nificant L2 ERP effects that reflect processing differences despite a lack ofbehavioral effects (McLaughlin et al., 2004; Tokowicz & MacWhinney, 2005).Furthermore, this approach (a) improves signal-to-noise ratio, (b) avoids eliminat-ing participants from analyses who may not have had a sufficient number of correcttrials in a given condition, and (c) avoids pseudo-effects due to asymmetric loss oftrials for a given condition (due to response bias).
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112498

Data. Note that for these analyses, two L2 Low subjects were excluded due tohaving an extremely low number of correct responses (o5) in at least onecondition.
2.5. Data analysis
2.5.1. Behavioral dataIn order to examine performance on the acceptability judgment task, partici-
pants' responses for each condition (see Table 3) were first transformed to d′ scores(see Table 4). Group differences in the ability to discriminate correct and violationsentences were then examined by submitting d′ scores for each participant toANOVAs (SPSS 19.0) with the between-subjects factor Group (L1, L2 Low, L2Advanced), separately for each condition (semantics, word order). Post-hoc Scheffétests were used to follow up on any significant differences across groups.Additionally, paired samples t-tests were used to compare performance onsemantic and word-order conditions within each group.
2.5.2. ERP dataERPs time-locked to the presentation of each target word were averaged by
condition type for each participant using a 200 ms pre-stimulus baseline, for asubset of 19 electrodes (16 lateral: F1/2, F5/6, FC1/2, FC5/6, CP1/2, CT5/6, P1/2, P5/6,and 3 midline: Fz, Cz, Pz). The subject averages were then entered into the grandaverage (group) ERP waveform for each set of target words (semantic violation,semantic control, word-order violation, word-order control). Mean amplitudeswere calculated for time-windows selected on the basis of the components ofinterest, together with both visual inspection of the waveforms and running t-testsof the these waveforms: for semantic violations, 300–500 ms and 500–800 ms; forword-order violations, 100–300 ms, 300–425 ms, 600–900 ms and 900–1200 ms,as well as 425–600 ms for completeness.
Mean amplitudes for each time-window were analyzed using global repeated-measures ANOVAs (SAS 9.3), separately for each condition (semantics, word order),first for the L1 group alone, as a baseline, and then across all three groups. Eachglobal ANOVA included, where applicable, the between-subjects factor Group (L1,L2 Low, L2 Advanced), as well as the within-subjects factor Violation (correct,violation), and the distributional factors Anterior/Posterior (anterior: F1/2, F5/6,centro-anterior: FC1/2, FC5/6, centro-posterior: CP1/2, CT5/6, posterior: P1/2, P5/6),Hemisphere (right, left), and Laterality (medial: F1/2, FC1/2, CP1/2, P1/2, lateral: F5/6, FC5/6, CT5/6, P5/6). Analogous analyses were conducted on the midlineelectrodes, without the factors Lat and Hemi, and with three levels of AP (anterior:Fz, central: Cz, posterior: Pz). When evaluating the AP factor, which included morethan one degree of freedom, the Greenhouse-Geisser correction was applied;corrected p values are reported here. For any global ANOVA that yielded asignificant interaction with the factor Viol, additional step-down ANOVAs wereperformed to clarify the nature of the interaction. When results from step-downanalyses of either lateral or midline electrodes across all three groups suggestedshared effects between only two of the groups, additional (lateral and midline)global ANOVAs (and step-down analyses for significant interactions) with only thetwo relevant groups were then performed, in order to elucidate similarities ordifferences in the effect between the two groups. We report all significant (pr .05)main effects and interactions with the factor Viol from each global ANOVA,as well as any group-specific or distributional Viol effects revealed at the lowestlevel by significant step-down analyses. The exact same set of analyses was run firston all artifact-free trials (all-trials analyses; reported as the main analyses) andthen on correctly-responded artifact-free trials (response-contingent analyses; seeSection 3.4).
3. Results
3.1. Behavioral data
The ANOVAs on the d′ scores for each condition (Table 4)yielded main effects of Group for both conditions (semantics: F(2,42)¼42.90, po .0001; word order: F(2,42)¼77.32, po .0001).Follow-up Scheffé tests revealed that for both conditions, the L1group outperformed the L2 Advanced group, who in turn out-performed the L2 Low group (pso .01). Additionally, the L2 Lowgroup performed significantly above chance (which would corre-spond to a d′ score of 0) in discriminating between correct andviolation sentences for both semantics and word order(pso .0001). Finally, paired t-tests showed that the L1 and L2Advanced groups both performed better at judging word orderthan semantics (pso .0001) whereas the L2 Low group performedsimilarly on the two conditions (p¼ .558).
3.2. ERP data: semantic violations
Grand average ERP waveforms and voltage maps for thesemantic violation condition (versus correct) are shown for eachgroup in Figs. 1–3, and results from the ANOVAs are reported inTables 5 and 6. Visual inspection and running t-tests suggestedthat, as predicted, N400s were elicited by all three groups. Inaddition, the L2 Advanced group, but not the other two groups,showed a late (600–800 ms) positivity.
3.2.1. Semantic violations in L1 SpanishIn the 300–500 ms time-window, the lateral ANOVA for the L1
group revealed a significant main effect of Viol (p¼ .003), reflectingthe N400, as well as several significant interactions qualifying thismain effect (see Fig. 1 and Table 5). First, there were significantViol�Hemi (p¼ .018) and Viol� Lat (p¼ .0004) interactions, whichwere in turn qualified by a Viol� Lat�Hemi interaction (p¼ .009).In addition, the ANOVA yielded a significant Viol�AP interaction(p¼ .001). Follow-up analyses on the three-way Viol� Lat�Hemiinteraction revealed significant left medial (F(1,14)¼11.71, p¼ .004)as well as right medial and right lateral negativities (F(1,14)¼21.80, p¼ .0004 and F(1,14)¼22.90, p¼ .0003, respectively). Thefollow-up analyses for the Viol�AP interaction pointed to anegativity that was most robust at centro-posterior and posteriorelectrodes (F(1,14)¼31.59, po .0001 and F(1,14)¼36.90, po .0001,respectively), but also extended, more weakly, to centro-anteriorelectrodes (F(1,14)¼5.09, p¼ .041). The midline ANOVA (also seeTable 5) yielded a significant main effect of Viol (p¼ .0009), as wellas a Viol�AP interaction (p¼ .004) that was due to significantnegativities restricted to the central (F(1,14)¼19.65, p¼ .0006) andposterior (F(1,14)¼45.39, po .0001) midline electrodes. Overall,the negativity in this time-window suggests a prototypical N400,with a posterior and slightly right-lateralized scalp distribution.
In the 500–800 ms time-window, the ANOVA for the L1 lateralelectrodes revealed several significant interactions: Viol�Hemi(p¼ .001), Viol� Lat�Hemi (p¼ .010), and Viol� Lat�AP (p¼.035). However, none of these resulted in significant follow-upanalyses. The midline ANOVA yielded no significant main effects orinteractions involving the factor Viol.
3.2.2. Semantic violations across groupsIn the 300–500 ms time-window, the global lateral ANOVA
across the three subject groups (L1, L2 Low, L2 Advanced; seeFigs. 1–3 and Table 6) yielded a significant main effect of Viol(p¼ .0006), reflecting an N400 in all three groups, though this wasqualified by several interactions involving topographical factors.First, there was a two-way Viol� Lat interaction (po .0001), which
Table 3Mean accuracy in percentage (and SDs) on the judgment task by group.
L1 Spanish(n¼15)
L2 LowSpanish (n¼16)
L2 AdvancedSpanish (n¼14)
Semantic violation 81.4 (14.0) 47.3 (22.9) 75.1 (9.9)Word-order violation 98.8 (2.3) 51.5 (22.9) 96.2 (8.8)Correct control 92.4 (7.5) 76.8 (8.8) 81.4 (10.1)
Table 4Mean d′ scores (and SDs) on the judgment task by group.
L1 Spanish(n¼16)
L2 LowSpanish (n¼16)
L2 AdvancedSpanish (n¼14)
Semantic condition 2.72 (.78) .72 (.52) 1.66 (.40)Word-order condition 4.41 (.98) .82 (.63) 3.39 (.85)
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2499
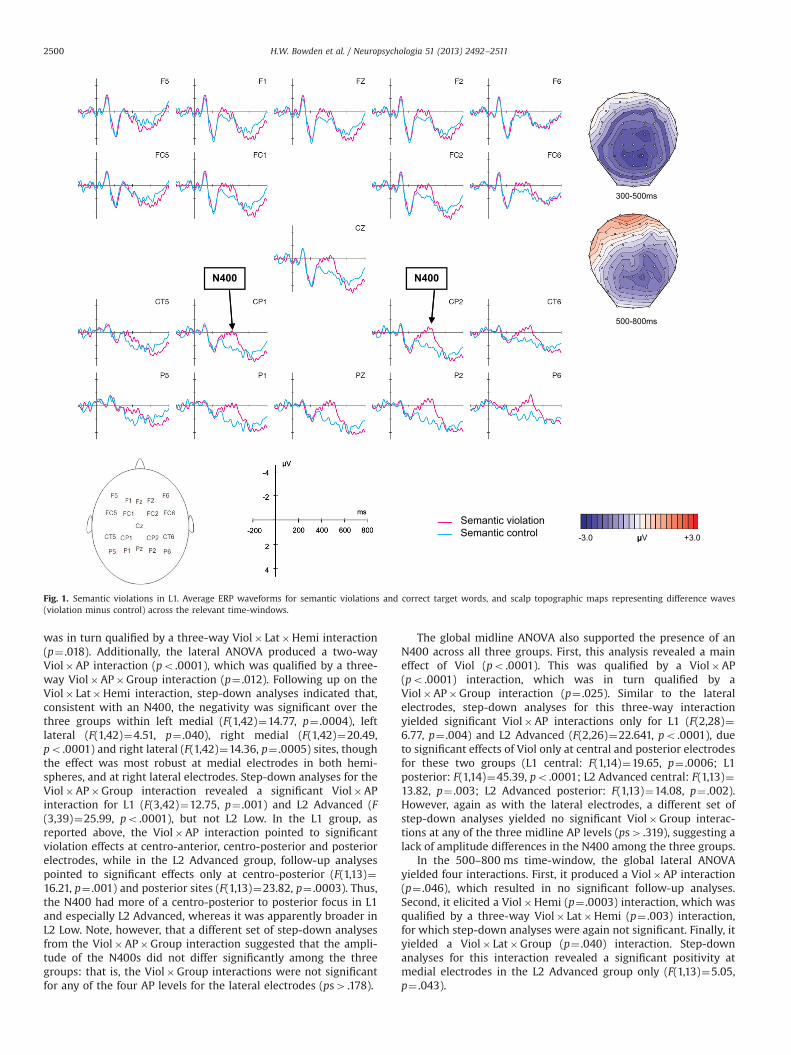
was in turn qualified by a three-way Viol� Lat�Hemi interaction(p¼ .018). Additionally, the lateral ANOVA produced a two-wayViol�AP interaction (po .0001), which was qualified by a three-way Viol�AP�Group interaction (p¼ .012). Following up on theViol� Lat�Hemi interaction, step-down analyses indicated that,consistent with an N400, the negativity was significant over thethree groups within left medial (F(1,42)¼14.77, p¼ .0004), leftlateral (F(1,42)¼4.51, p¼ .040), right medial (F(1,42)¼20.49,po .0001) and right lateral (F(1,42)¼14.36, p¼ .0005) sites, thoughthe effect was most robust at medial electrodes in both hemi-spheres, and at right lateral electrodes. Step-down analyses for theViol�AP�Group interaction revealed a significant Viol�APinteraction for L1 (F(3,42)¼12.75, p¼ .001) and L2 Advanced (F(3,39)¼25.99, po .0001), but not L2 Low. In the L1 group, asreported above, the Viol�AP interaction pointed to significantviolation effects at centro-anterior, centro-posterior and posteriorelectrodes, while in the L2 Advanced group, follow-up analysespointed to significant effects only at centro-posterior (F(1,13)¼16.21, p¼ .001) and posterior sites (F(1,13)¼23.82, p¼ .0003). Thus,the N400 had more of a centro-posterior to posterior focus in L1and especially L2 Advanced, whereas it was apparently broader inL2 Low. Note, however, that a different set of step-down analysesfrom the Viol�AP�Group interaction suggested that the ampli-tude of the N400s did not differ significantly among the threegroups: that is, the Viol�Group interactions were not significantfor any of the four AP levels for the lateral electrodes (ps4 .178).
The global midline ANOVA also supported the presence of anN400 across all three groups. First, this analysis revealed a maineffect of Viol (po .0001). This was qualified by a Viol�AP(po .0001) interaction, which was in turn qualified by aViol�AP�Group interaction (p¼ .025). Similar to the lateralelectrodes, step-down analyses for this three-way interactionyielded significant Viol�AP interactions only for L1 (F(2,28)¼6.77, p¼ .004) and L2 Advanced (F(2,26)¼22.641, po .0001), dueto significant effects of Viol only at central and posterior electrodesfor these two groups (L1 central: F(1,14)¼19.65, p¼ .0006; L1posterior: F(1,14)¼45.39, po .0001; L2 Advanced central: F(1,13)¼13.82, p¼ .003; L2 Advanced posterior: F(1,13)¼14.08, p¼ .002).However, again as with the lateral electrodes, a different set ofstep-down analyses yielded no significant Viol�Group interac-tions at any of the three midline AP levels (ps4 .319), suggesting alack of amplitude differences in the N400 among the three groups.
In the 500–800 ms time-window, the global lateral ANOVAyielded four interactions. First, it produced a Viol�AP interaction(p¼ .046), which resulted in no significant follow-up analyses.Second, it elicited a Viol�Hemi (p¼ .0003) interaction, which wasqualified by a three-way Viol� Lat�Hemi (p¼ .003) interaction,for which step-down analyses were again not significant. Finally, ityielded a Viol� Lat�Group (p¼ .040) interaction. Step-downanalyses for this interaction revealed a significant positivity atmedial electrodes in the L2 Advanced group only (F(1,13)¼5.05,p¼ .043).
300-500ms
N400 N400
500-800ms
Semantic violationSemantic control -3.0 μV +3.0μ
Fig. 1. Semantic violations in L1. Average ERP waveforms for semantic violations and correct target words, and scalp topographic maps representing difference waves(violation minus control) across the relevant time-windows.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112500

In sum, semantic violations elicited a classic N400 in all threegroups (L1, L2 Low, L2 Advanced), which did not differ betweenthe groups in amplitude, though the focus was more centro-posterior to posterior in the L1 and L2 Advanced groups ascompared to the L2 Low group. Additionally, a later medial(P600-like) positivity was found in the L2 Advanced group.
3.3. ERP data: word-order violations
Grand average ERP waveforms and voltage maps for theword-order violation condition (versus correct) are shown foreach group in Figs. 4–6, and results from the ANOVAs arereported in Tables 7–8. Visual inspection and running t-testssuggested that, consistent with our predictions, the word-orderviolations elicited a LAN-like effect in the 300–425 ms time-window and a centro-posterior P600 in the 600–900 ms time-window for both the L1 and L2 Advanced groups. In contrast, theL2 Low group appeared to show only a broad positivity in thesetime-windows, which was apparently strongest at anterior sites.In the 900–1200 ms time-window, the L1, and possibly L2Advanced group as well, appeared to show posterior positivitiesand late anterior negativities, whereas the L2 Low groupappeared to evidence only a positivity that seemed strongest atanterior sites.
3.3.1. Word-order violations in L1 SpanishIn the 100–300 ms time-window, the lateral ANOVA for the L1
Spanish group yielded a Viol�Hemi interaction (p¼ .040), thoughthe follow-up analyses were not significant. The midline ANOVAyielded no significant effects involving Viol. See Fig. 4 and Table 7.
In the 300–425 ms time-window, the lateral ANOVA yielded asignificant main effect of Viol (p¼ .032), and a significantViol�Hemi interaction (p¼ .006), both of which were qualifiedby a Viol� Lat�Hemi interaction (p¼ .044). This three-way inter-action was due to significant negativities at left medial and leftlateral electrodes (but not right medial or right lateral), with theeffect being more robust over left lateral (F(1,14¼9.73, p¼ .008))than left medial (F(1,14¼6.95, p¼ .020)), suggesting a LAN. At themidline, the ANOVA produced a significant main effect of Viol(p¼ .028), again due to the negativity in the L1 group.
In the 425–600 ms time-window, there were no significantmain effects or interactions involving Viol in either the lateral ormidline ANOVAs.
In the 600–900 ms time-window, there were two significantinteractions: a Viol� Lat�Hemi interaction (p¼ .0002) and aViol�AP interaction (p¼ .0007). The first of these did not lead tosignificant follow-ups. Step-down ANOVAs from the Viol�APinteraction pointed to significant positivities in centro-posteriorand posterior electrodes (F(1,14)¼5.59, p¼ .033 and F(1,14)¼12.92,p¼ .003, respectively), consistent with a P600. The midline ANOVA
300-500ms
N400 N400
500-800ms
Semantic violationSemantic control -3.0 μV +3.0μ
Fig. 2. Semantic violations in L2 Low. Average ERP waveforms for semantic violations and correct target words, and scalp topographic maps representing difference waves(violation minus control) across the relevant time-windows.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2501
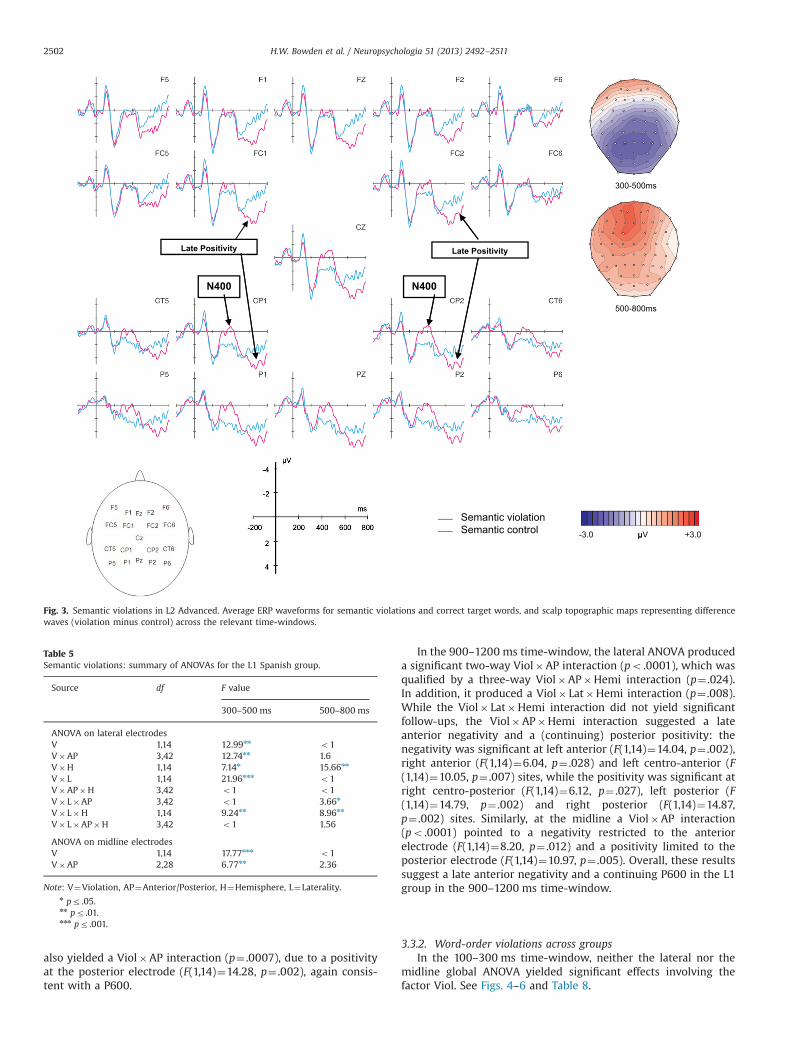
also yielded a Viol�AP interaction (p¼ .0007), due to a positivityat the posterior electrode (F(1,14)¼14.28, p¼ .002), again consis-tent with a P600.
In the 900–1200 ms time-window, the lateral ANOVA produceda significant two-way Viol�AP interaction (po .0001), which wasqualified by a three-way Viol�AP�Hemi interaction (p¼ .024).In addition, it produced a Viol� Lat�Hemi interaction (p¼ .008).While the Viol� Lat�Hemi interaction did not yield significantfollow-ups, the Viol�AP�Hemi interaction suggested a lateanterior negativity and a (continuing) posterior positivity: thenegativity was significant at left anterior (F(1,14)¼14.04, p¼ .002),right anterior (F(1,14)¼6.04, p¼ .028) and left centro-anterior (F(1,14)¼10.05, p¼ .007) sites, while the positivity was significant atright centro-posterior (F(1,14)¼6.12, p¼ .027), left posterior (F(1,14)¼14.79, p¼ .002) and right posterior (F(1,14)¼14.87,p¼ .002) sites. Similarly, at the midline a Viol�AP interaction(po .0001) pointed to a negativity restricted to the anteriorelectrode (F(1,14)¼8.20, p¼ .012) and a positivity limited to theposterior electrode (F(1,14)¼10.97, p¼ .005). Overall, these resultssuggest a late anterior negativity and a continuing P600 in the L1group in the 900–1200 ms time-window.
3.3.2. Word-order violations across groupsIn the 100–300 ms time-window, neither the lateral nor the
midline global ANOVA yielded significant effects involving thefactor Viol. See Figs. 4–6 and Table 8.
300-500ms
Late Positivity Late Positivity
N400 N400
500-800ms
Semantic violationSemantic control -3.0 μV +3.0μ
Fig. 3. Semantic violations in L2 Advanced. Average ERP waveforms for semantic violations and correct target words, and scalp topographic maps representing differencewaves (violation minus control) across the relevant time-windows.
Table 5Semantic violations: summary of ANOVAs for the L1 Spanish group.
Source df F value
300–500 ms 500–800 ms
ANOVA on lateral electrodesV 1,14 12.99nn o1V�AP 3,42 12.74nn 1.6V�H 1,14 7.14n 15.66nn
V� L 1,14 21.96nnn o1V�AP�H 3,42 o1 o1V� L�AP 3,42 o1 3.66n
V� L�H 1,14 9.24nn 8.96nn
V� L�AP�H 3,42 o1 1.56
ANOVA on midline electrodesV 1,14 17.77nnn o1V�AP 2,28 6.77nn 2.36
Note: V¼Violation, AP¼Anterior/Posterior, H¼Hemisphere, L¼Laterality.n pr .05.nn pr .01.nnn pr .001.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112502
In the 300–425 ms time-window, the global lateral ANOVAelicited a Viol�Group interaction (p¼ .0004) that was qualified bya four-way Viol�AP�Hemi�Group interaction (p¼ .032). Step-down analyses for this four-way interaction revealed negativitiesin the L1 and L2 Advanced groups, and a positivity in the L2 Lowgroup as follows. The negativity in the L1 group was significant atleft anterior (F(1,14)¼9.56, p¼ .008) and left centro-anterior (F(1,13)¼14.02, p¼ .002) sites. In the L2 Advanced group, thenegativity was also significant at left anterior (F(1,13)¼11.15,p¼ .005) and left centro-anterior (F(1,13)¼14.02, p¼ .002) sites,as well as, though less robustly, right centro-anterior (F(1,13)¼6.72, p¼ .022) and left centro-posterior electrodes (F(1,13)¼6.44,p¼ .025). Both of these negativities are consistent with a LAN,though apparently with a somewhat broader distribution for L2Advanced than for L1. The positivity in the L2 Low group wassignificant at left anterior (F(1,15)¼4.81, p¼ .044) and left centro-anterior (F(1,15)¼9.00, p¼ .009) sites. At the midline, a Viol�Group interaction (p¼ .004) pointed to a significant main effect ofViol within only L1 and L2 Advanced groups, reflecting thenegativity present in those groups (L1: F(1,14)¼6.01, p¼ .028; L2Advanced: F(1,13)¼5.083, p¼ .042).
Given the similar left anterior negativities in both L1 and L2Advanced in the 300–425 ms time-window, we performed globalANOVAs with only these two groups (see Methods section).Neither the lateral nor the midline global ANOVA revealed anysignificant interactions involving both Viol and Group, indicatingthe two groups did not differ in this time-window. Instead, therewas a main effect of Viol over the two groups in both the lateraland midline ANOVAs (lateral: F(1,27)¼14.80, p¼ .0007; midline: F(1,27)¼11.06, p¼ .003), which was qualified (only for the lateralANOVA) by three interactions: a Viol�Hemi interaction (F(1,27)¼13.29, p¼ .001), which was in turn qualified by both a Viol�AP�Hemi interaction (F(3,81)¼4.15, p¼ .016) and a Viol� Lat�Hemi interaction (F(1,27)¼6.20, p¼ .019). Step-down analyses forthe Viol� Lat�Hemi interaction revealed a shared negativity that
was significant at left medial (F(1,27)¼13.20, p¼ .001) and leftlateral (F(1,27)¼11.53, p¼ .002) regions, as well as right medial (F(1,27)¼8.73, p¼ .006) and right lateral (F(1,27)¼10.50, p¼ .003)sites. The Viol�AP�Hemi interaction indicated that the negativ-ity was significant primarily in left anterior (F(1,27)¼19.08,p¼ .0002) and left centro-anterior (F(1,27)¼28.21, po .0001)regions, extending more weakly (reflected by the interaction withHemi) to right anterior (F(1,27)¼7.05, p¼ .013) and right-centro-anterior (F(1,27)¼8.54, p¼ .007) sites. Overall, these findingsindicate a LAN in both the L1 and L2 Advanced groups, whichmoreover did not differ between them.
In the 425–600 ms time-window, the global lateral ANOVAproduced four significant interactions. First of all, it elicited aViol� Lat interaction (p¼ .007), which was qualified by a three-way Viol� Lat�Group interaction (p¼ .014), for which step-downanalyses yielded non-significant results. In addition, it produced aViol� Lat�AP interaction (p¼ .010), which pointed to a sharedpositivity across the three groups in centro-anterior medialelectrodes (F(1,42)¼4.25, p¼ .046). Finally, it produced a four-way Viol�AP�Hemi�Group interaction, which yielded no sig-nificant follow-ups. No significant effects including the factor Violwere produced by the global midline ANOVA. Thus, in this time-window, there was a shared centro-anterior medial positivity.
In the 600–900 ms time-window, the global lateral ANOVAyielded a significant main effect of Viol (p¼ .009), and severalinteractions. There were two two-way interactions: Viol� Lat(p¼ .0008) and Viol�AP (p¼ .0002), which were qualified bythree three-way interactions: Viol� Lat�AP (p¼ .028), Viol�Lat�Group (p¼ .022), Viol�AP�Group (p¼ .002). The last ofthese was in turn qualified by a four-way Viol�AP�Hemi�Group (p¼ .020) interaction. The first of the three unqualifiedinteractions, Viol� Lat�AP, pointed to shared positivities acrossthe three groups in the following regions: centro-posterior medial(F(1,42)¼17.66, p¼ .0001), centro-posterior lateral (F(1,42)¼14.50,p¼ .0005), posterior medial (F(1,42)¼30.28, po .0001) and poster-ior lateral (F(1,42)¼24.64, po .0001). Step-down analyses for thesecond unqualified interaction (Viol� Lat�Group) pointed to amedial positivity (F(1,13)¼5.71, p¼ .033) in L2 Advanced only,consistent with a P600. Step-down analyses for the third unqua-lified interaction, Viol�AP�Hemi�Group, revealed positivitiesin left and right centro-posterior and posterior regions for both L1(left centro-posterior: F(1,14)¼4.96, p¼ .043; right centro-poster-ior: F(1,14)¼5.79, p¼ .030; left posterior: F(1,14)¼11.33, p¼ .004;right posterior: F(1,14)¼13.36, p¼ .003) and L2 Advanced (leftcentro-posterior: F(1,13)¼6.50, p¼ .024; right centro-posterior: F(1,13)¼11.28, p¼ .005; left posterior: F(1,13)¼12.91, p¼ .003; rightposterior: F(1,13)¼17.53, p¼ .001), but not L2 Low, suggestive ofP600s only for the L1 and L2 Advanced groups. At the midlineelectrodes, there was a similar pattern of results: the globalANOVA yielded a main effect of Viol (p¼ .006), reflecting thepositivity, as well as a Viol�AP interaction (p¼ .0001), thoughthese were qualified by a Viol�AP�Group interaction (p¼ .0009).Step-down analyses for this interaction revealed a posteriorpositivity for L1 (F(1,14)¼14.28, p¼ .002), and a central (F(1,13)¼8.49 p¼ .012) to posterior (F(1,13)¼13.80, p¼ .003) positivity for L2Advanced. This pattern of results suggests that though a positivitywas shared in (medial and lateral) centro-posterior to posteriorregions across the three groups, the positivity had a centro-posterior to posterior focus (reflected in the interactions withthe AP factor) only in the L1 and L2 Advanced groups. Thus, onlythese groups appeared to show a classic P600, consistent withvisual inspection of Figs. 4–6, while the L2 Low group showed abroad positivity only.
Given evidence of a similar positivity in L1 and L2 Advanced,we also performed global ANOVAs with only these two groups.Neither the lateral nor midline global ANOVAs yielded any
Table 6Semantic violations: summary of ANOVAs across L1, L2 Low and L2 Advanced.
Source df F value
300–500 ms 500–800 ms
ANOVA on lateral electrodesV 1,42 13.63nnn o1V�G 2,42 o1 2.80þ
V�AP 3,126 31.54nnn 3.72n
V�H 1,42 3.04þ 15.20nnn
V� L 1,42 26.99nnn o1V�AP�G 6,126 4.20n o1V�H�G 2,42 2.27 1.11V� L�G 2,42 o1 3.47n
V�AP�H 3,126 o1 o1V� L�AP 3,126 1.44 2.89þ
V� L�H 1,43 6.02n 10.24nn
V�AP�H�G 6,126 1.16 2.06þ
V� L�AP�G 6,126 1.31 1.72V� L�H�G 2,43 1.65 o1V� L�AP�H 3,126 o1 o1V� L�AP�H�G 6,126 o1 o1
ANOVA on midline electrodesV 1,42 18.57nnn o1V�G 2,42 o1 2.90þ
V�AP 2,84 15.56nnn 5.33n
V�AP�G 4,84 3.46n o1
Note: V¼Violation, AP¼Anterior/Posterior, H¼Hemisphere, L¼Laterality.þ pr .10.n pr .05.nn pr .01.nnn pr .001.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2503

significant effects involving the factors Viol and Group, indicatingthat the observed positivities were statistically indistinguishablebetween the L1 and L2 Advanced groups. Instead, the lateralANOVA revealed a main effect of Viol (F(1,27)¼4.78, p¼ .038),reflecting the positivity, as well two two-way interactions(Viol� Lat: F(1,27)¼23.16, po .0001, and Viol�Hemi: F(1,27)¼9.71, p¼ .004) which were qualified by three higher-order inter-actions: Viol� Lat�Hemi (F(1,27)¼7.74, p¼ .010), Viol� Lat�AP(F(3,81)¼3.48, p¼ .032), and Viol�AP�Hemi (F(3,81)¼5.20,p¼ .011). Step-down analyses for the Viol� Lat�Hemi interactionwere not significant. Step-down analyses for the Viol� Lat�APinteraction revealed a shared centro-posterior to posterior posi-tivity significant at both medial and lateral sites (centro-posteriormedial: F(1,27)¼17.24, p¼ .0003; centro-posterior lateral: F(1,27)¼10.61, p¼ .003; posterior medial: F(1,27)¼31.10, po .0001; pos-terior lateral: F(1,27)¼24.36, po .0001), though (reflecting the inter-action with both Lat and AP) it was stronger at medial and posteriorelectrodes, consistent with a P600. Follow-up analyses for theViol�AP�Hemi interaction signaled a significant positivity atcentro-posterior and especially posterior electrodes in both the leftand right hemispheres (left centro-posterior: F(1,27)¼11.59, p¼ .002;right centro-posterior: F(1,27)¼16.88, po .0001; left posterior: F(1,27)¼24.42, po .0001; right posterior: F(1,27)¼30.86, po .0001).
Similarly, the global midline ANOVA including only the L1 and L2Advanced groups produced a main effect of Viol (F(1,27)¼6.78,p¼ .015) and one interaction: Viol�AP (F(2,54)¼24.87, po .0001).Step-down analyses from this interaction revealed a shared positivityat the central (F(1,27)¼7.94, p¼ .009) and posterior (F(1,27)¼28.06,po .0001) electrodes, which was again more robust at the posteriorelectrode, reflecting the P600.
In the 900–1200 ms time-window, the global lateral ANOVAyielded six significant interactions. Three of these (Viol�Hemi:p¼ .003; Viol� Lat: p¼ .043; and Viol�AP: po .0001) were qua-lified by three higher-level interactions: Viol� Lat�Hemi (p¼.019), Viol�AP�Hemi (p¼ .022), and Viol�AP�Group (po.0001). Step-down analyses from the Viol� Lat�Hemi interactiondid not yield significant results. Step-down analyses from theViol�AP�Hemi interaction revealed a right centro-posterior (F(1,42)¼5.38, p¼ .025) to right posterior (F(1,42)¼9.23, p¼ .004)positivity that was shared across the three groups. Step-downanalyses from the Viol�AP�Group interaction revealed, for theL1 group only, an anterior (F(1,14)¼10.04, p¼ .007) to centro-anterior (F(1,14)¼4.72, p¼ .048) negativity, as well as a centro-posterior (F(1,14)¼4.80, p¼ .046) to posterior (F(1,14)¼16.05,p¼ .001) positivity. The global midline ANOVA yielded a Viol�APinteraction (po .0001), which was qualified by a Viol�AP�Group
LAN
Late Anterior Negativity
100-300ms
300-425ms
P600
425-600ms
Word-order violationWord-order control
600-900ms
900-1200ms
-3.0 μV +3.0
Fig. 4. Word-order violations in L1. Average ERP waveforms for word-order violations and correct target words, and scalp topographic maps representing difference waves(violation minus control) across the relevant time-windows.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112504
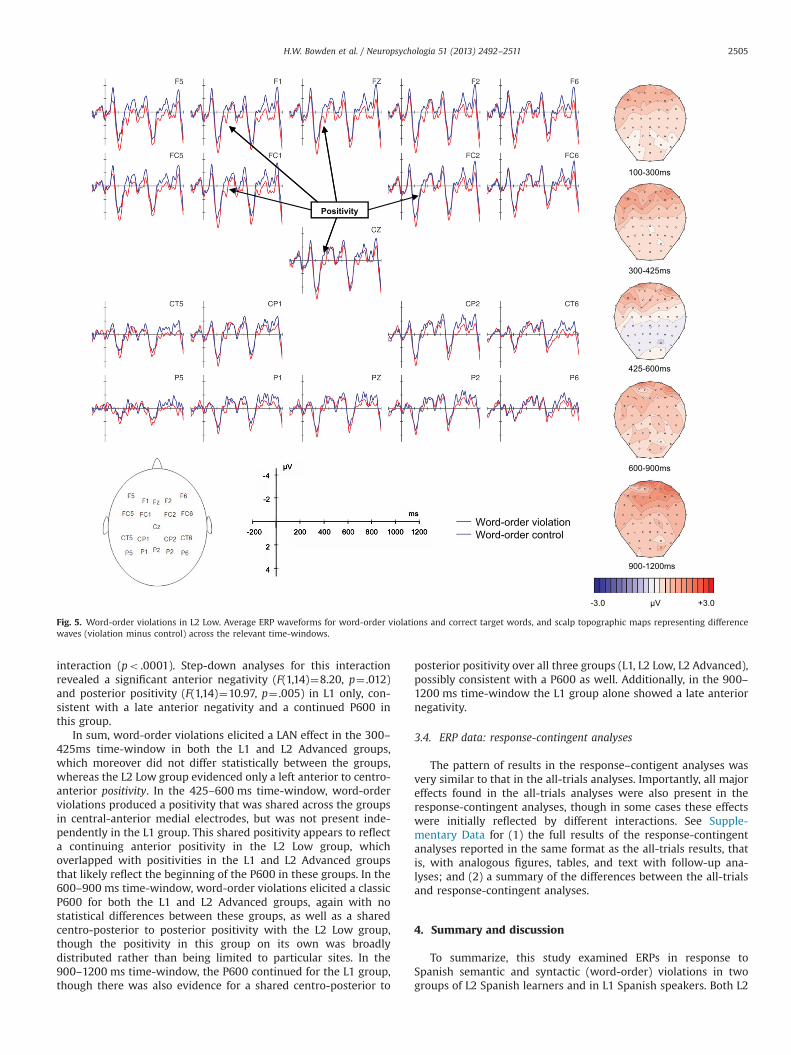
interaction (po .0001). Step-down analyses for this interactionrevealed a significant anterior negativity (F(1,14)¼8.20, p¼ .012)and posterior positivity (F(1,14)¼10.97, p¼ .005) in L1 only, con-sistent with a late anterior negativity and a continued P600 inthis group.
In sum, word-order violations elicited a LAN effect in the 300–425ms time-window in both the L1 and L2 Advanced groups,which moreover did not differ statistically between the groups,whereas the L2 Low group evidenced only a left anterior to centro-anterior positivity. In the 425–600 ms time-window, word-orderviolations produced a positivity that was shared across the groupsin central-anterior medial electrodes, but was not present inde-pendently in the L1 group. This shared positivity appears to reflecta continuing anterior positivity in the L2 Low group, whichoverlapped with positivities in the L1 and L2 Advanced groupsthat likely reflect the beginning of the P600 in these groups. In the600–900 ms time-window, word-order violations elicited a classicP600 for both the L1 and L2 Advanced groups, again with nostatistical differences between these groups, as well as a sharedcentro-posterior to posterior positivity with the L2 Low group,though the positivity in this group on its own was broadlydistributed rather than being limited to particular sites. In the900–1200 ms time-window, the P600 continued for the L1 group,though there was also evidence for a shared centro-posterior to
posterior positivity over all three groups (L1, L2 Low, L2 Advanced),possibly consistent with a P600 as well. Additionally, in the 900–1200 ms time-window the L1 group alone showed a late anteriornegativity.
3.4. ERP data: response-contingent analyses
The pattern of results in the response–contigent analyses wasvery similar to that in the all-trials analyses. Importantly, all majoreffects found in the all-trials analyses were also present in theresponse-contingent analyses, though in some cases these effectswere initially reflected by different interactions. See Supple-mentary Data for (1) the full results of the response-contingentanalyses reported in the same format as the all-trials results, thatis, with analogous figures, tables, and text with follow-up ana-lyses; and (2) a summary of the differences between the all-trialsand response-contingent analyses.
4. Summary and discussion
To summarize, this study examined ERPs in response toSpanish semantic and syntactic (word-order) violations in twogroups of L2 Spanish learners and in L1 Spanish speakers. Both L2
100-300ms
Positivity
300-425ms
425-600ms
Word-order violationWord-order control
600-900ms
900-1200ms
-3.0 μV +3.0
Fig. 5. Word-order violations in L2 Low. Average ERP waveforms for word-order violations and correct target words, and scalp topographic maps representing differencewaves (violation minus control) across the relevant time-windows.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2505
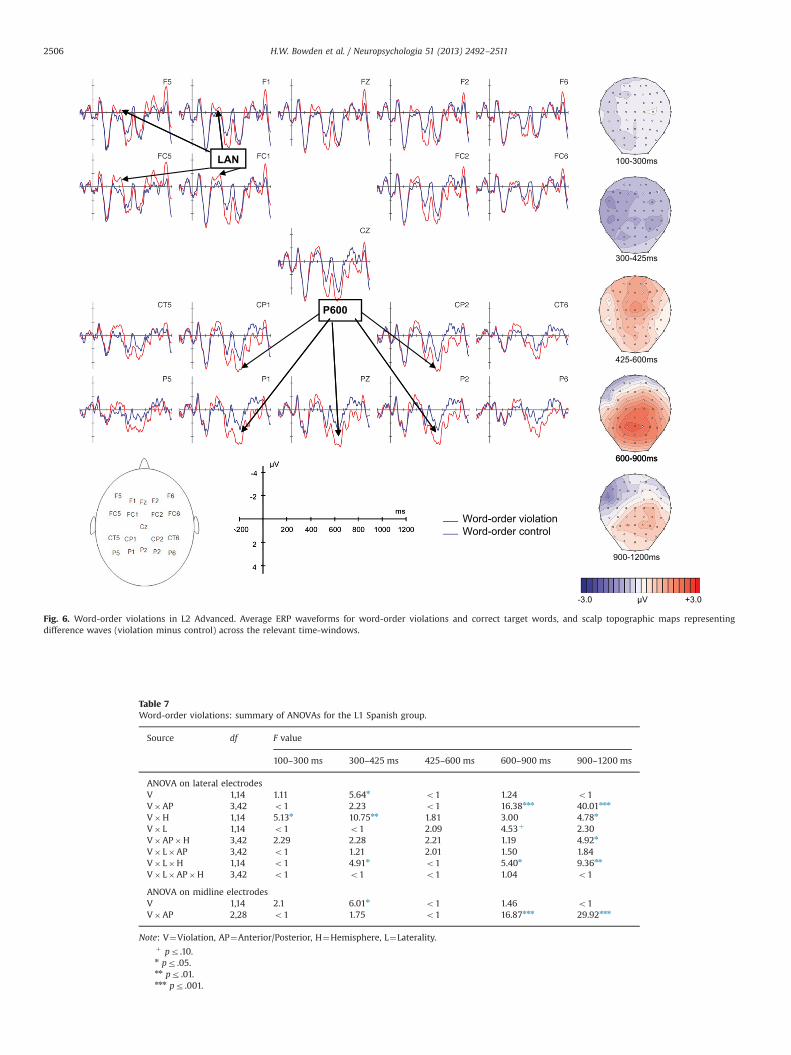
100-300msLAN
300-425ms
425-600ms
P600
600-900ms
Word-order violationWord-order control
600-900ms
900-1200ms
-3.0 μV +3.0
Fig. 6. Word-order violations in L2 Advanced. Average ERP waveforms for word-order violations and correct target words, and scalp topographic maps representingdifference waves (violation minus control) across the relevant time-windows.
Table 7Word-order violations: summary of ANOVAs for the L1 Spanish group.
Source df F value
100–300 ms 300–425 ms 425–600 ms 600–900 ms 900–1200 ms
ANOVA on lateral electrodesV 1,14 1.11 5.64n o1 1.24 o1V�AP 3,42 o1 2.23 o1 16.38nnn 40.01nnn
V�H 1,14 5.13n 10.75nn 1.81 3.00 4.78n
V� L 1,14 o1 o1 2.09 4.53þ 2.30V�AP�H 3,42 2.29 2.28 2.21 1.19 4.92n
V� L�AP 3,42 o1 1.21 2.01 1.50 1.84V� L�H 1,14 o1 4.91n o1 5.40n 9.36nn
V� L�AP�H 3,42 o1 o1 o1 1.04 o1
ANOVA on midline electrodesV 1,14 2.1 6.01n o1 1.46 o1V�AP 2,28 o1 1.75 o1 16.87nnn 29.92nnn
Note: V¼Violation, AP¼Anterior/Posterior, H¼Hemisphere, L¼Laterality.þ pr .10.n pr .05.nn pr .01.nnn pr .001.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112506
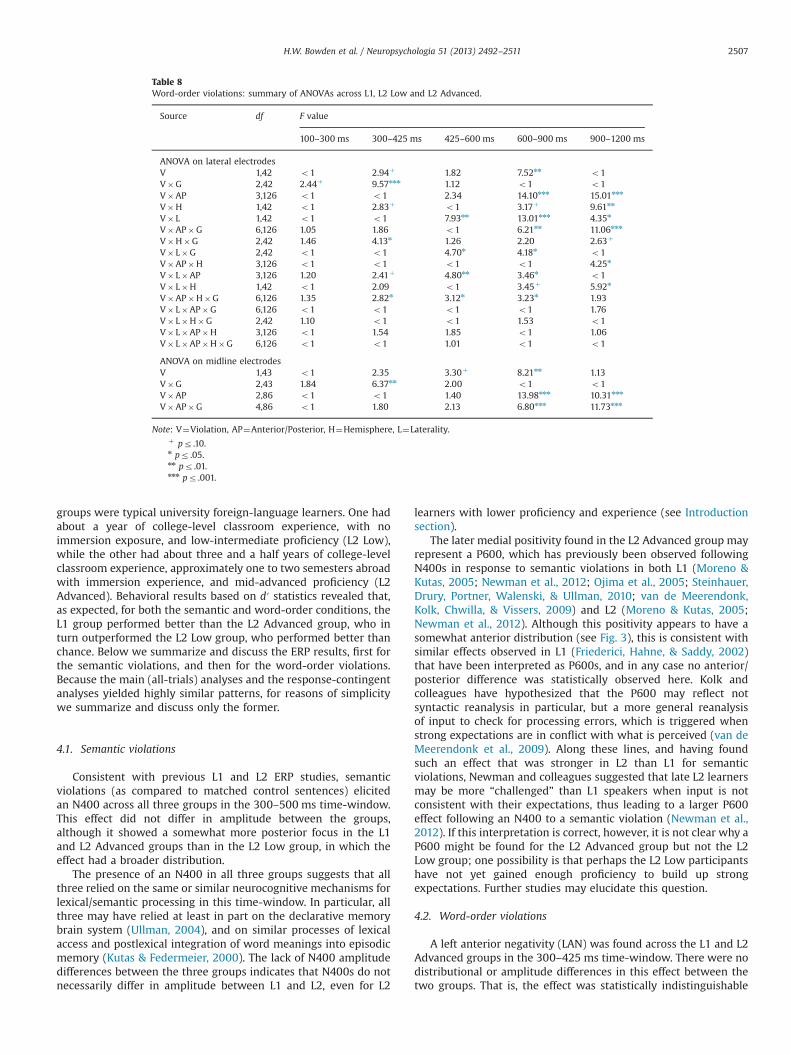
groups were typical university foreign-language learners. One hadabout a year of college-level classroom experience, with noimmersion exposure, and low-intermediate proficiency (L2 Low),while the other had about three and a half years of college-levelclassroom experience, approximately one to two semesters abroadwith immersion experience, and mid-advanced proficiency (L2Advanced). Behavioral results based on d′ statistics revealed that,as expected, for both the semantic and word-order conditions, theL1 group performed better than the L2 Advanced group, who inturn outperformed the L2 Low group, who performed better thanchance. Below we summarize and discuss the ERP results, first forthe semantic violations, and then for the word-order violations.Because the main (all-trials) analyses and the response-contingentanalyses yielded highly similar patterns, for reasons of simplicitywe summarize and discuss only the former.
4.1. Semantic violations
Consistent with previous L1 and L2 ERP studies, semanticviolations (as compared to matched control sentences) elicitedan N400 across all three groups in the 300–500 ms time-window.This effect did not differ in amplitude between the groups,although it showed a somewhat more posterior focus in the L1and L2 Advanced groups than in the L2 Low group, in which theeffect had a broader distribution.
The presence of an N400 in all three groups suggests that allthree relied on the same or similar neurocognitive mechanisms forlexical/semantic processing in this time-window. In particular, allthree may have relied at least in part on the declarative memorybrain system (Ullman, 2004), and on similar processes of lexicalaccess and postlexical integration of word meanings into episodicmemory (Kutas & Federmeier, 2000). The lack of N400 amplitudedifferences between the three groups indicates that N400s do notnecessarily differ in amplitude between L1 and L2, even for L2
learners with lower proficiency and experience (see Introductionsection).
The later medial positivity found in the L2 Advanced group mayrepresent a P600, which has previously been observed followingN400s in response to semantic violations in both L1 (Moreno &Kutas, 2005; Newman et al., 2012; Ojima et al., 2005; Steinhauer,Drury, Portner, Walenski, & Ullman, 2010; van de Meerendonk,Kolk, Chwilla, & Vissers, 2009) and L2 (Moreno & Kutas, 2005;Newman et al., 2012). Although this positivity appears to have asomewhat anterior distribution (see Fig. 3), this is consistent withsimilar effects observed in L1 (Friederici, Hahne, & Saddy, 2002)that have been interpreted as P600s, and in any case no anterior/posterior difference was statistically observed here. Kolk andcolleagues have hypothesized that the P600 may reflect notsyntactic reanalysis in particular, but a more general reanalysisof input to check for processing errors, which is triggered whenstrong expectations are in conflict with what is perceived (van deMeerendonk et al., 2009). Along these lines, and having foundsuch an effect that was stronger in L2 than L1 for semanticviolations, Newman and colleagues suggested that late L2 learnersmay be more “challenged” than L1 speakers when input is notconsistent with their expectations, thus leading to a larger P600effect following an N400 to a semantic violation (Newman et al.,2012). If this interpretation is correct, however, it is not clear why aP600 might be found for the L2 Advanced group but not the L2Low group; one possibility is that perhaps the L2 Low participantshave not yet gained enough proficiency to build up strongexpectations. Further studies may elucidate this question.
4.2. Word-order violations
A left anterior negativity (LAN) was found across the L1 and L2Advanced groups in the 300–425 ms time-window. There were nodistributional or amplitude differences in this effect between thetwo groups. That is, the effect was statistically indistinguishable
Table 8Word-order violations: summary of ANOVAs across L1, L2 Low and L2 Advanced.
Source df F value
100–300 ms 300–425 ms 425–600 ms 600–900 ms 900–1200 ms
ANOVA on lateral electrodesV 1,42 o1 2.94þ 1.82 7.52nn o1V�G 2,42 2.44þ 9.57nnn 1.12 o1 o1V�AP 3,126 o1 o1 2.34 14.10nnn 15.01nnn
V�H 1,42 o1 2.83þ o1 3.17þ 9.61nn
V� L 1,42 o1 o1 7.93nn 13.01nnn 4.35n
V�AP�G 6,126 1.05 1.86 o1 6.21nn 11.06nnn
V�H�G 2,42 1.46 4.13n 1.26 2.20 2.63þ
V� L�G 2,42 o1 o1 4.70n 4.18n o1V�AP�H 3,126 o1 o1 o1 o1 4.25n
V� L�AP 3,126 1.20 2.41þ 4.80nn 3.46n o1V� L�H 1,42 o1 2.09 o1 3.45þ 5.92n
V�AP�H�G 6,126 1.35 2.82n 3.12n 3.23n 1.93V� L�AP�G 6,126 o1 o1 o1 o1 1.76V� L�H�G 2,42 1.10 o1 o1 1.53 o1V� L�AP�H 3,126 o1 1.54 1.85 o1 1.06V� L�AP�H�G 6,126 o1 o1 1.01 o1 o1
ANOVA on midline electrodesV 1,43 o1 2.35 3.30þ 8.21nn 1.13V�G 2,43 1.84 6.37nn 2.00 o1 o1V�AP 2,86 o1 o1 1.40 13.98nnn 10.31nnn
V�AP�G 4,86 o1 1.80 2.13 6.80nnn 11.73nnn
Note: V¼Violation, AP¼Anterior/Posterior, H¼Hemisphere, L¼Laterality.þ pr .10.n pr .05.nn pr .01.nnn pr .001.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2507

between the L1 and L2 Advanced groups. This left and anteriordistribution was found not only across the two groups, but alsoindependently in both groups in follow-up analyses to the globalANOVA that was carried out across the three groups. The finding ofa left-lateralized but not anteriorly-distributed negativity in theanalyses carried out separately on the L1 group may either reflectthe actual distribution of this negativity (consistent with other L1LAN findings; see Introduction) or may be due to insufficientpower in this analysis (consistent with the voltage maps andwaveforms, which suggest that the LAN effect may indeed have ananterior distribution in the L1 group; see Fig. 4). Note that sincethe analyses across the L1 and L2 Advanced groups had morepower, yet did not reveal any group differences for the sharedeffect at left anterior electrodes, we believe the most appropriateinterpretation of this pattern is that, indeed, both groups elicited aleft-anterior LAN (consistent with their respective voltage maps).There was no evidence for an ELAN in any group in the earlier100–300 ms time-window.
The LAN in the L1 and L2 Advanced groups suggests that in thistime-window word-order violations may undergo similar or perhapseven identical neural processing in native speakers and reasonablyadvanced L2 learners – specifically, university foreign-languagelearners with about seven semesters of college Spanish, immersionduring one or two semesters abroad, and reasonably high profi-ciency. The implication of the LAN in automatic structure-building(see Introduction section) suggests that even such an automatic andimplicit process can be achieved by such learners.
The L2 Low group, in contrast, showed a left anterior to centro-anterior positivity during the 300–425ms time-window. It isunclear how this positivity should be interpreted. A similar effecthas also been observed at relatively high proficiency levels in L2learners (of an artificial language, albeit one that was actuallyspoken and comprehended) who had received only explicit, class-room-like, instruction (Morgan-Short, Finger, et al., 2012; Morgan-Short, Steinhauer, et al., 2012). In that study the effect wasinterpreted as a possible P3a, which is driven by attentionalmechanisms, since explicit, classroom-like, training conditionsmay be more effective than implicit training conditions in direct-ing learners' attention to L2 forms. This interpretation seemsconsistent with the finding in the present study, given that onlythe L2 Low group had just classroom experience. More generally, itmay be that such effects may tend to reflect explicit, instructedclassroom experience rather than uninstructed, naturalisticimmersion experience in L2 learners.
Word-order violations again elicited a common effect acrossthe L1 and L2 Advanced groups in the 600–900 ms time-window,in this case a P600. This effect was again statistically indistinguish-able between the two groups. The L2 Low group in thistime-window shared a positivity with these two groups incentro-posterior to posterior regions, though the follow-up ana-lyses suggested that in the L2 Low group it was broadly distributedrather than being restricted to particular regions.
In the 900–1200 ms time-window, the P600 continued for theL1 group, though there was also a shared centro-posterior toposterior positivity over all three groups. In the 900–1200 mstime-window the L1 group, but neither L2 group, also showed alate anterior negativity.
Thus, the L1 group showed a LAN, P600, and late anteriornegativity, while the L2 Advanced group showed only a LAN and aP600, and the L2 Low group showed evidence for only an earlyanterior positivity and a continued weak positivity. Several impor-tant observations are in order. First, the L2 Advanced group and L1group were statistically indistinguishable in all time-windows upto 900–1200 ms. This suggests that, during the initial second or sofollowing a target word in syntactic word-order processing, theneural mechanisms of native speakers and of the type of advanced
L2 learners examined here differ minimally if at all. Second,however, the L2 Advanced group was not identical to the L1 groupfor syntactic processing, in that the L1 group but not the L2Advanced group showed a late anterior negativity in the 900–1200 ms time-window. Third, as expected, the L2 Advanced groupshowed greater L1-like brain processing of syntax than the L2 Lowgroup, which showed only an early anterior positivity and acontinuing weak positivity.
Therefore, at least in the case of Spanish learned by nativeEnglish speakers, typical undergraduate foreign-language learnerswith only about a year of university exposure to the L2, noimmersion experience at all, and relatively low proficiency donot seem to show (on average) any native-like processing ofsyntactic word order. In contrast, on average, university foreign-language learners who take L2 classes through much of collegeand also study abroad for one or two semesters, and who haverelatively high (but not native-like) proficiency, show evidence ofmuch more native-like brain processing of syntax, including whatmay represent both early automatic structure-building and latercontrolled processing. Nevertheless, even these learners do notshow all aspects of native-like brain processing for syntax, asevidenced by the absence of late anterior negativities.
More generally, the findings here are both consistent with andextend previous findings. As we have seen in the introduction,previous studies suggest that proficiency, or one of its correlates,as well as possibly immersion experience, may be associated withmore native-like brain processing of (morpho)syntax, includingthe presence of LANs, P600s, and late anterior negativities. Con-sistent with these findings, the present study shows that higherlevels of proficiency and experience are associated with increasednative-like processing, in particular the presence of both LAN andP600 effects. Moreover, as in previous research, the LAN was onlyfound in the group with immersion experience, though theselearners also had significant classroom exposure, and the presentstudy does not tease apart these two factors.
Importantly, whereas previous studies have for the most partfocused on L2 learners with an immigrant-like experience, andalmost exclusively on such groups for higher proficiency levels, thepresent study extends the literature to the type of foreign-language learner typical in the U.S. and many other countries –
that is, who is exposed to the L2 exclusively through classroomexperience from middle school or high school through much ofcollege, with their only immersion experience from study abroad,generally towards the end of college. The findings reported hereshow that although at the end of their first year of L2 study incollege, such learners show no signs of native-like processing ofsyntactic word order, toward the end of their college L2 experi-ence, including study abroad, their brain processing of syntax ishighly similar to native speakers, and indeed is statisticallyindistinguishable in the initial second or so of processing followinga target word.
We should emphasize that attaining native-like brain proces-sing evidently does not necessarily entail native-like proficiency.Rather, in this study and many previous studies of L2, native-likeERP patterns of syntax (and semantics) have been observed in L2learners without native-like performance. Nevertheless, it is pos-sible that native-like processing might facilitate the eventualattainment of native-like proficiency, since presumably nativespeakers use the most efficient processing mechanisms for lan-guage. Further studies are needed to examine this issue; seeMorgan-Short et al. (2012) for additional discussion.
Finally, we underscore the fact that the balanced stimulusparadigm employed here for word-order violations providesassurance that the observed effects are not due to context orbaseline confounds, but rather should be explained by languageprocessing itself.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112508

4.3. Theoretical implications
The evidence from this study, together with findings fromprevious research, helps elucidate the theoretical landscape. Overall,the data do not seem in accordance with the hypothesis that L1 andL2 processing (in similarly aged individuals) always depend onessentially the same neurocognitive mechanisms (Abutalebi, 2008;Ellis, 2005; Hernandez et al., 2005; Indefrey, 2006; MacWhinney,2012), nor with the view that their mechanisms are entirely different(Bley-Vroman, 1989).
Instead, the findings seem to be consistent with the view thatat least for grammar, L2 learners initially rely on largely differentsubstrates than (similarly aged) L1 individuals, but, with increas-ing experience or proficiency, can gradually come to depend on L1neurocognitive mechanisms (Clahsen & Felser, 2006a; Paradis,1994; Ullman, 2001). In particular, the results specifically supportthe following (Clahsen & Felser, 2006a, 2006b; Clahsen et al., 2010;Ullman, 2001, 2005, 2012): for lexical/semantics, L1 and L2 speak-ers rely largely on a common neurocognitive system; in contrast,for grammatical processing, at lower proficiency and/or experi-ence L2 learners rely largely on lexical/semantic processing, andperhaps declarative memory in particular, whereas at higher L2levels learners can come to depend on L1-like mechanisms,including perhaps procedural memory. (Note that this view doesnot preclude the possibility that L2 learners follow the samedevelopmental trajectory as L1 learners, that is, children; indeed,it has been suggested that L1 children may follow a similar patternUllman, 2005)
The findings presented here also further elucidate and specifythis view. First, if indeed the early positivity found in the lowproficiency/exposure group in response to word-order violationsreflects a P3a related to explicit knowledge (which is still spec-ulative), this would further support the role of declarative memory(which is critical for explicit knowledge) in syntactic processingfollowing just classroom learning, prior to any immersion. It is notclear why such a P3a-like effect has been found in some studies,and an N400 in others. Second, taken together with previousresearch, the fact that the L2 group showing a LAN had not onlysignificant classroom instruction but also several months ofimmersion experience, suggests that immersion might be parti-cularly important for fully-native like processing of syntax, thoughit is important to emphasize that this study did not tease apartimmersion and additional classroom experience; further researchis clearly needed to examine this issue. Third, the finding of a LANas well as a P600 in the L2 Advanced group, both of which werestatistically indistinguishable from those of the native speakers,further constrains the theoretical landscape (Clahsen & Felser,2006b; Ullman, 2005) by revealing another type of syntacticstructure for which L1-like processing of grammar can beachieved. Moreover, it suggests that L1-like brain processing ofsyntax is possible even for typical university foreign-languagelearners who follow a purely classroom regimen plus just one ortwo semesters of study abroad.
4.4. Potential limitations and future directions
Before drawing any conclusions, we consider potential limita-tions and future directions. First, the participants in the L2 Lowgroup were somewhat younger and less educated than the othertwo groups (in part because they were college students in theirearlier years). It might be argued that this could explain thedifferences seen in their responses to the word-order violationsboth in earlier and later time-windows. However, this accountdoes not seem plausible, given that children younger than 19 aswell as adults have been shown to reliably elicit LAN and P600effects (Hahne, Eckstein, & Friederici, 2004). Second, following
Osterhout, McLaughlin, Kim, Greenwald, and Inoue (2004), it maybe possible that between-subject variability obscured any LAN-likeeffects in the L2 Low group. However, the presence of a positivityin the LAN time window argues against such an account. Third, thesignificance of both the later positivity found for semantic viola-tions in the L2 Advanced group, and the early anterior positivityfound for word-order violations in the L2 Low group, remainunclear, and should be further clarified in future studies. Fourth, asdiscussed above, a primary purpose of this study was to determinewhether the type of university L2 learner who is typical in the U.S.(and various other countries) could attain native-like brain pro-cessing of syntax toward the end of their foreign-language learn-ing experience in college. The answer seems to be yes, at least forthose processes reflected by the LAN and P600. Crucially, the studyis designed precisely to open doors to further investigations topinpoint exactly when during the foreign-language trajectory suchnative-like processing, that is, a LAN/P600 biphasic response,emerges.
There are several directions that future studies could take tofurther elucidate this question. In particular, they could askwhether and to what extent immersion and/or classroom experi-ence might most reliably lead to native-like processing (andproficiency). For example, might immersion experience be eithernecessary and/or sufficient for the emergence of a LAN/P600response? If so, how much immersion and when (e.g., is a monthor two sufficient, and might it be more beneficial after a certainamount of classroom experience (cf. DeKeyser, 2010; Llanes,2011))? Can a native-like response be found after an entirelyclassroom-based experience? If so, how much is necessary, and ofwhat type (e.g., more or less meta-linguistic vs. naturalistic)? Ordoes a combination of classroom exposure and immersion experi-ence most reliably yield native-like processing in universityforeign-language learners? Additionally, further studies shouldelucidate at what point, that is, after what further training, lateanterior negativities may emerge. Interestingly, visual inspectionof Fig. 6 suggests that this component is already beginning toappear in the L2 Advanced group. Thus perhaps a relativelymodest amount of further (classroom or immersion) trainingmight be sufficient for it to become robust. Future studies shouldelucidate whether the emergence of native-like processing ofgrammar may vary according to other factors not examined here,including linguistic structure (e.g., whether it emerges for wordorder prior to morphosyntax) and L1-L2 structural similarity.Indeed, as mentioned above, the present study stacked the deckto maximize the likelihood of the emergence of native-likeresponses for grammar; it seems plausible that under similar L2learning conditions, less native-like processing might be foundfor morphosyntax than word order, and in languages with lessstructural similarity to English.
5. Conclusion
In sum, the findings from the present study, together withprevious evidence, suggest the following. Lexical/semantic proces-sing in L2 always seems to rely on qualitatively similar neuralprocesses as in L1, independent of factors such as proficiency orexposure – though in some cases these can apparently quantita-tively affect these mechanisms. In contrast, the neurocognitivemechanisms underlying grammatical processing in L2 seem tochange qualitatively during the language learning process. Thisshift appears to depend on various factors, including (thoughprobably not limited to) proficiency and/or the amount of expo-sure, and possibly immersion experience in particular. Althoughthe exact role of these factors is still not well understood, theevidence suggests that higher proficiency and/or exposure, and
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2509

possibly some amount of immersion experience, may be critical(though possibly not sufficient) for fully native-like processing tobe evidenced.
The present experiment shows that at least the best-studiedERP components associated with native-like grammatical proces-sing, that is, the LAN and P600, can be attained even in the type offoreign-language learning environment that is often found in theU.S. as well as other countries: namely, solely classroom experi-ence from middle school or high school through much of college,plus one or two semesters of study abroad, and the attainment ofhigh but not native-like proficiency. Thus, the type of fullerimmersion experience that is typically found in immigrant-likecontexts, and has been the focus of most previous ERP studies ofL2, does not appear to be necessary to attain substantial native-like processing of at least some aspects of syntax.
Acknowledgments
We would like to thank David Birdsong, Matthew Gelfand, RonLeow, Christopher Maloof, Robbin Miranda, Kara Morgan-Short,and Aaron Newman for support in various aspects of the project.Any errors or omissions are ours alone. Support for this projectwas provided to MTU by the NIH under RO1 MH58189 and RO1HD049347, by the NSF under Doctoral Dissertation ImprovementGrant 0519133, to HWB by the NIH under Ruth L. KirschsteinNational Research Service Award 5 F31MH68143-04 from theNIMH, and to KS by the Canada Research Chair program and theCanada Foundation for Innovation (CRC/CFI; project # 201876) andthe Natural Sciences and Engineering Research Council of Canada(NSERC; # RGPGP 312835-05 and RGPIN 402678-11).
Appendix A. Supplementary Information
Supplementary data associated with this article can be found inthe online version at doi:10.1016/j.neuropsychologia.2013.09.004.
References
Abutalebi, J. (2008). Neural aspects of second language representation and languagecontrol. Acta Psychologica, 128(3), 466–478.
Alonso, C. J. (2006). Spanish: The foreign national language. ADFL Bulletin, 37(2–3),15–20.
Ardal, S., Donald, M. W., Meuter, R., Muldrew, S., & Luce, M. (1990). Brain responsesto semantic incongruity in bilinguals. Brain and Language, 39(2), 187–205.
Bley-Vroman, R. (1989). What is the logical problem of foreign language learning?.In: S. M. Gass, & J. Schacter (Eds.), Linguistic perspectives on second languageacquisition (pp. 41–68). Cambridge: Cambridge University Press.
Bowden, H. W., Gelfand, M. P., Sanz, C., & Ullman, M. T. (2010). Verbal inflectionalmorphology in L1 and L2 Spanish: A frequency effects study examining storagevs. composition. Language Learning, 60(1), 44–87.
Chen, L., Shu, H., Liu, Y., Zhao, J., & Li, P. (2007). ERP signatures of subject-verbagreement in L2 learning. Bilingualism: Language and Cognition, 10(2), 161–174.
Clahsen, H., & Felser, C. (2006a). Grammatical processing in language learners.Applied Psycholinguistics, 27(1), 3–42.
Clahsen, H., & Felser, C. (2006b). How native-like is non-native language procees-sing? Trends in Cognitive Sciences, 10(12), 564–570.
Clahsen, H., Felser, C., Neubauer, K., Sato, M., & Silva, R. (2010). Morphologicalstructure in native and nonnative language processing. Language Learning, 60(1), 21–43.
de Groot, A. M. B. (2011). Language and cognition in bilinguals and multilinguals: Anintroduction. New York: Psychology Press.
DeKeyser, R. (2010). Monitoring processes in Spanish as a second language during astudy abroad program. Foreign Language Annals, 43(1), 80–92.
Doughty, C. J., & Long, M. H. (2005). The handbook of second language acquisition.Oxford: Wiley-Blackwell Publishing Ltd.
Ellis, N. C. (2005). At the interface: Dynamic interactions of explicit and implicitlanguage knowledge. Studies in Second Language Acquisition, 27(2), 305–352.
Foucart, A., & Frenck-Mestre, C. (2012). Can late L2 learners acquire new gramma-tical features? Evidence from ERPs and eye-tracking. Journal of Memory andLanguage, 66, 226–248.
Frenck-Mestre, C., Osterhout, L., McLaughlin, J., & Foucart, A. (2008). The effect ofphonological realization of inflectional morphology on verbal agreement inFrench: Evidence from ERPs. Acta Psychologica, 128(3), 528–536.
Friederici, A., Hahne, A., & Saddy, D. (2002). Distinct neurophysiological patternsreflecting aspects of syntactic complexity and syntactic repair. Journal ofPsycholinguistic Research, 31(1), 45–63.
Friederici, A. D. (1995). The time course of syntactic activation during languageprocessing: A model based on neuropsychological and neurophysiological data.Brain and Language, 50, 259–281.
Friederici, A. D. (2002). Towards a neural basis of auditory sentence processing.Trends in Cognitive Sciences, 6(2), 78–84.
Friederici, A. D., & Kotz, S. A. (2003). The brain basis of syntactic processes:Functional imaging and lesion studies. NeuroImage, 20, S8–S17.
Friederici, A. D., Pfeifer, E., & Hahne, A. (1993). Event-related brain potentials duringnatural speech processing: Effects of semantic, morphological and syntacticviolations. Cognitive Brain Research, 1(3), 183–192.
Friederici, A. D., Steinhauer, K., & Pfeifer, E. (2002). Brain signatures of artificiallanguage processing: Evidence challenging the critical period hypothesis.Proceedings of the National Academy of Sciences, 99(1), 529–534.
Gillon Dowens, M., Guo, T., Guo, J., Barber, H., & Carreiras, M. (2011). Gender andnumber processing in Chinese learners of Spanish – Evidence from eventrelated potentials. Neuropsychologia, 49(7), 1651–1659.
Gillon Dowens, M., Vergara, M., Barber, H., & Carreiras, M. (2010). Morphosyntacticprocessing in late second-language learners. Journal of Cognitive Neuroscience,22(8), 1870–1887.
Green, D. W. (2003). Neural basis of the lexicon and the grammar in L2 acquisition.In: R. v. Hout, A. Hulk, F. Kuiken, & R. Towell (Eds.), The interface between syntaxand the lexicon in second language acquisition (pp. 197–208). Amsterdam: JohnBenjamins.
Hagoort, P. (2003). Interplay between syntax and semantics during sentencecomprehension: ERP effects of combining syntactic and semantic violations.Journal of Cognitive Neuroscience, 15(6), 883–899.
Hagoort, P., & Brown, C. M. (1999). Gender electrified: ERP evidence on thesyntactic nature of gender processing. Journal of Psycholinguist Research, 28(6), 715–728.
Hahne, A. (2001). What's different in second-language processing? Evidence fromevent-related brain potentials. Journal of Psycholinguistic Research, 30(3),251–266.
Hahne, A., Eckstein, K., & Friederici, A. D. (2004). Brain signatures of syntactic andsemantic processes during children's language development. Journal of Cogni-tive Neuroscience, 16(7), 1302–1318.
Hahne, A., & Friederici, A. D. (1999). Electrophysiological evidence for two steps insyntactic analysis: Early automatic and late controlled processes. Journal ofCognitive Neuroscience, 11(2), 194–205.
Hahne, A., & Friederici, A. D. (2001). Processing a second language: Late learners'comprehension mechanisms as revealed by event-related brain potentials.Bilingualism: Language and Cognition, 4, 123–141.
Hahne, A., Mueller, J. L., & Clahsen, H. (2006). Morphological processing in a secondlanguage: Behavioral and event-related brain potential evidence for storageand decomposition. Journal of Cognitive Neuroscience, 18(1), 121–134.
Hasting, A. S., & Kotz, S. A. (2008). Speeding up syntax: On the relative timing andautomaticity of local phrase structure and morphosyntactic processing asreflected in event-related brain potentials. Journal of Cognitive Neuroscience,20(7), 1207–1219.
Henning, G. (1992). The ACTFL oral proficiency interview: Validity evidence. System,20(3), 365–372.
Hernandez, A., Li, P., & MacWhinney, B. (2005). The emergence of competingmodules in bilingualism. Trends in Cognitive Sciences, 9(5), 220–225.
Hernandez, A. E., & Li, P. (2007). Age of acquisition: Its neural and computationalmechanisms. Psychological Bulletin, 133(4), 638–650.
Hoen, M., & Dominey, P. F. (2000). ERP analysis of cognitive sequencing: A leftanterior negativity related to structural transformation processing. NeuroRe-port, 11(14), 3187–3191.
Indefrey, P. (2006). A meta-analysis of hemodynamic studies on first and secondlanguage processing: Which suggested differences can we trust and what dothey mean? Language Learning, 56(Suppl. 1), S279–S304.
Isel, F. (2007). Syntactic and referential processes in second-language learners:Event-related brain potential evidence. NeuroReport, 18(18), 1885–1889.
Kaan, E., Harris, A., Gibson, E., & Holcomb, P. (2000). The P600 as an index ofsyntactic integration difficulty. Language and Cognitive Processes, 15(2),159–201.
Kotz, S. A. (2009). A critical review of ERP and fMRI evidence on L2 syntacticprocessing. Brain and Language, 109, 68–74.
Kroll, J. F., & de Groot, A. M.B. (Eds.). (2005). Handbook of bilingualism: Psycholin-guistic approaches. New York: Oxford University Press.
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology reveals semantic memoryuse in language comprehension. Trends in Cognitive Sciences, 4(12), 463–470.
Kutas, M., & Federmeier, K. D. (2010). Thirty years and counting: Finding meaningin the N400 component of the event-related brain potential (ERP). The AnnualReview of Psychology, 62, 14.11–14.27.
Kutas, M., & Hillyard, S. A. (1980). Reading senseless sentences: Brain potentialsreflect semantic incongruity. Science, 207(1), 203–205.
Kutas, M., & Kluender, R. (1994). What is who violating? A reconsideration oflinguistic violations in light of event-related brain potentials. In: H.-J. Heinze,T. F. Münte, & G. R. Mangun (Eds.), Cognitive electrophysiology: Basic and clinicalapplications (pp. 183–210). Boston: Birkhäuser.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–25112510

Llanes, A. (2011). The many faces of study abroad: An update on the research on L2gains emerged during a study abroad experience. International Journal ofMultilingualism, 8(3), 189–215.
Gass, S. M., & Mackey, A. (2012). Handbook of second language acquisition. New York:Routledge.
MacWhinney, B. (2007). A unified model. In: N. Ellis, & P. Robinson (Eds.), Handbookof cognitive linguistics and second language acquisition. New York: Routledge.
MacWhinney, B. (2012). The logic of the unified model. In: S. Gass, & A. Mackey(Eds.), Handbook of second language acquisition (pp. 211–227). New York:Routledge..
Martin-Loeches, M., Munoz, F., Casado, P., Melcon, A., & Fernandez-Frias, C. (2005).Are the anterior negativities to grammatical violations indexing workingmemory? Psychophysiology, 42, 508–519.
McLaughlin, J., Osterhout, L., & Kim, A. (2004). Neural correlates of second-languageword learning: Minimal instruction produces rapid change. Nature Neuroscience7(7), 703–704.
McLaughlin, J., Tanner, D., Pitkanen, I., Frenck-Mestre, C., Inoue, K., Valentine, G.,et al. (2010). Brain potentials reveal discrete stages of L2 grammatical learning.Language Learning, 60(1), 123–150.
Meuter, R., Donald, M. W., & Ardal, S. (1987). A comparison of first- and second-language ERPs in bilinguals. Electroencephalography and Clinical Neurophysiol-ogy, Suppl. 40, S412–S416.
Montrul, S. A. (2004). The acquisition of Spanish: Morphosyntactic development inmonolingual and bilingual L1 acquisition and adult L2 acquisition.
Moreno, E. M., & Kutas, M. (2005). Processing semantic anomalies in twolanguages: An electrophysiological exploration in both languages of Spanish–English bilinguals. Brain Research: Cognitive Brain Research, 22(2), 205–220.
Morgan-Short, K., Finger, I., Grey, S., & Ullman, M. T. (2012). Second languageprocessing shows increased native-like neural responses after months of noexposure. PLoS ONE, 7(3), e32974.
Morgan-Short, K., Steinhauer, K., Sanz, C., & Ullman, M. T. (2012). Explicit andimplicit second language training differentially affect the achievement ofnative-like brain activation patterns. Journal of Cognitive Neuroscience, 24(4),933–947.
Morgan-Short, K., Sanz, C., Steinhauer, K., & Ullman, M. T. (2010). Second languageacquisition of gender agreement in explicit and implicit training conditions: Anevent-related potential study. Language Learning, 60(1), 154–193.
Morgan-Short, K., & Ullman, M. T. (2011). The neurocognition of second language.In: S. M. Gass, & A. Mackey (Eds.), Handbook of second language acquisition (pp.282–299). New York: Routledge.
Mueller, J. L., Hahne, A., Fujii, Y., & Friederici, A. D. (2005). Native and nonnativespeakers' processing of a miniature version of Japanese as revealed by ERPs.Journal of Cognitive Neuroscience, 17(8), 1229–1244.
Neville, H. J., Nicol, J. L., Barss, A., Forster, K. I., & Garrett, M. F. (1991). Syntacticallybased sentence processing classes: Evidence from event-related brain poten-tials. Journal of Cognitive Neuroscience, 3(2), 151–165.
Newman, A. J., Tremblay, A., Nichols, E. S., Neville, H. J., & Ullman, M. T. (2012). Theinfluence of language proficiency on lexical-semantic processing in native andlate learners of English. Journal of Cognitive Neuroscience, 24(5), 1205–1223.
Nobre, A. C., Allison, T., & McCarthy, G. (1994). Word recognition in the humaninferior temporal lobe. Nature, 372, 260–263.
Ojima, S., Nakata, H., & Kakigi, R. (2005). An ERP study on second language learningafter childhood: Effects of proficiency. Journal of Cognitive Neuroscience, 17(8),1212–1228.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburghinventory. Neuropsychologia, 9(1), 97–113.
Osterhout, L., Bersick, M., & McLaughlin, J. (1997). Brain potentials reflect violationsof gender stereotypes. Memory & Cognition, 25(3), 273–285.
Osterhout, L., & Holcomb, P. J. (1992). Event-related brain potentials elicited bysyntactic anomaly. Journal of Memory and Language, 31, 785–806.
Osterhout, L., Holcomb, P. J., & Swinney, D. A. (1994). Brain potentials elicited bygarden-path sentences: Evidence of the application of verb information duringparsing. Journal of Experimental Psychology: Learning, Memory, and Cognition, 20(4), 786–803.
Osterhout, L., McLaughlin, J., Kim, A., Greenwald, R., & Inoue, K. (2004). Sentences inthe brain: Event related potentials as real time reflections of sentencecomprehension and language learning. In: M. Carreiras, & J. Clifton (Eds.), Theon-line study of sentence comprehension: Eye tracking, ERP and beyond (pp. 271–308). New York: Psychology Press.
Osterhout, L., & Nicol, J. (1999). On the distinctiveness, independence, and timecourse of the brain responses to syntactic and pragmatic anomalies. Languageand Cognitive Processes, 14(3), 283–317.
Osterhout, L., Poliakov, A., Inoue, K., McLaughlin, J., Valentine, G., Pitkanen, I., et al.(2008). Second-language learning and changes in the brain. Journal of Neuro-linguistics, 21(6), 509–521.
Pakulak, E., & Neville, H. J. (2010). Proficiency differences in syntactic processing ofmonolingual native speakers indexed by event-related potentials. Journal ofCognitive Neuroscience, 22(12), 2728–2744.
Pakulak, E., & Neville, H. J. (2011). Maturational constraints on the recruitment ofearly processes for syntactic processing. Journal of Cognitive Neuroscience, 23(10), 2752–2765.
Paradis, M. (1994). Neurolinguistic aspects of implicit and explicit memory:Implications for bilingualism and SLA. In: N. C. Ellis (Ed.), Implicit and explicitlearning of languages (pp. 393–419). London, UK: Academic Press.
Paradis, M. (2004). A neurolinguistic theory of bilingualism. Amsterdam, Nether-lands: John Benjamins.
Paradis, M. (2009). Declarative and procedural determinants of second languages, Vol.40. Amsterdam: John Benjamins Publishing Company.
Rhodes, N. C., & Pufahl, I. (2009). Foreign language teaching in U.S. schools: Resultsof a national survey. Washington, DC: Center for Applied Linguistics.
Rossi, S., Gugler, M. F., Friederici, A. D., & Hahne, A. (2006). The impact ofproficiency on syntactic second-language processing of German and Italian:Evidence from event-related potentials. Journal of Cognitive Neuroscience, 18(12), 2030–2048.
Sabourin, L., & Stowe, L. A. (2008). Second language processing: When are first andsecond languages processed similarly? Second Language Research, 24(3),397–430.
Schmidt, G. L., & Roberts, T. P. L. (2009). Second language research usingmagnetoencephalography: A review. Second Language Research, 25(1), 135–166.
Simos, P. G., Basile, L. F. H., & Papanicolaou, A. C. (1997). Source localization of theN400 response in a sentence-reading paradigm using evoked magnetic fieldsand magnetic resonance imaging. Brain Research, 762(1–2), 29–39.
Steinhauer, K., & Connolly, J. F. (2008). Event-related potentials in the study oflanguage. In: B. Stemmer, & H. A. Whitaker (Eds.), Handbook of the neuroscienceof language (pp. 91–104). Oxford, U.K.: Elsevier.
Steinhauer, K., & Drury, J. E. (2012). On the early left-anterior negativity (ELAN) insyntax studies. Brain and Language, 120(2), 135–162.
Steinhauer, K., Drury, J. E., Portner, P., Walenski, M., & Ullman, M. T. (2010). Syntax,concepts, and logic in the temporal dynamics of language comprehension:Evidence from event-related potentials. Neuropsychologia, 48(6), 1525–1542.
Steinhauer, K., White, E. J., & Drury, J. E. (2009). Temporal dynamics of late secondlanguage acquisition: Evidence from event-related brain potentials. SecondLanguage Research, 25(1), 13–41.
Steinhauer, K., White, E. J., & Genesee, F. (2013). Proficiency and first languagetransfer effects in a late acquired second language: ERP evidence from abalanced word-category violation paradigm (in preparation).
Tokowicz, N., & MacWhinney, B. (2005). Implicit and explicit measures ofsensitivity to violations in second language grammar: An event-relatedpotential investigation. Studies in Second Language Acquisition, 27(2), 173–204.
Ullman, M. T. (2001). The neural basis of lexicon and grammar in first and secondlanguage: The declarative/procedural model. Bilingualism: Language and Cogni-tion, 4(1), 105–122.
Ullman, M. T. (2004). Contributions of memory circuits to language: The declara-tive/procedural model. Cognition, 92(1–2), 231–270.
Ullman, M. T. (2005). A cognitive neuroscience perspective on second languageacquisition: The declarative/procedural model. In: C. Sanz (Ed.), Mind andcontext in adult second language acquisition: Methods, theory and practice (pp.141–178). Washington, DC: Georgetown University Press.
Ullman, M. T. (2006). The declarative/procedural model and the shallow-structurehypothesis. Applied Psycholinguistics, 27(1), 97–105.
Ullman, M. T. (2012). The declarative/procedural model. In: P. Robinson (Ed.),Routledge encyclopedia of second language acquisition (pp. 160–164). New York &London: Routledge.
Ullman, M. T., Gelfand, M. P., Walenski, M., Moffa, M., & Lee, S. (2013). The influenceof handedness on language: Storage versus composition differences in left- andright-handers (in preparation).
Ullman, M. T., Miranda, R. A., & Travers, M. L. (2008). Sex differences in theneurocognition of language. In: J. B. Becker, K. J. Berkley, N. Geary, E. Hampson,J. Herman, & E. Young (Eds.), Sex on the brain: From genes to behavior (pp. 291–309). New York: Oxford University Press.
van de Meerendonk, N., Kolk, H. H. J., Chwilla, D. J., & Vissers, C. T. W. M. (2009).Monitoring in language perception. Language and Linguistics Compass, 3(5),1211–1224.
van den Brink, D., & Hagoort, P. (2004). The influence of semantic and syntacticcontext constraints on lexical selection and integration in spoken-wordcomprehension as revealed by ERPs. Journal of Cognitive Neuroscience, 16(6),1068–1084.
Vos, S. H., Gunter, T. C., Kolk, H. H. J., & Mulder, G. (2001). Working memoryconstraints on syntactic processing: An electrophysiological investigation.Psychophysiology, 38(1), 41–63.
Weber-Fox, C. M., & Neville, H. J. (1996). Maturational constraints on functionalspecializations for language processing: ERP and behavioral evidence inbilingual speakers. Journal of Cognitive Neuroscience, 8(3), 231–256.
White, L. (2003). Fossilization in steady state L2 grammars: Persistent problemswith inflectional morphology. Bilingualism: Language and Cognition, 6, 129–141.
H.W. Bowden et al. / Neuropsychologia 51 (2013) 2492–2511 2511




















